
Increase of Bradyrhizobium japonicum numbers in soils and enhanced nodulation of soybean (Glycine...
11
ELSEVIER FEMS Microbiology Ecology 20 ( 1996) 173- 183 Increase of Bradyrhizobium japonicum numbers in soils and enhanced nodulation of soybean (Glycine max CL) merr.) using granular inoculants amended with nutrients Georges Fouilleux I, Ckcile Revellin, Alain Hartmann, Gkrard Catroux * INRA-CMSE, Laboratoire de Microbiologic des Sols INRA, BV 1540. 21034 Dijon Ct!de.r, France Received 6 February 1996; accepted 22 March 1996 Abstract Mineral microgranules, amended with nutrients and inoculated with either peat or liquid Bradyrhizobium japonicum inoculants, increased the growth and recovery of the bacterium during laboratory incubation in unsterilized soil. Increases in the range of 1 log unit per g or ml inoculant used were observed in different soil types. B. japonicum showed better survival with nutrient-amended granules than in unamended ones, in soil undergoing desiccation. In a growth chamber experiment, the number of nodules per plant were significantly increased by nutrient-amendment of the granules, but only under suboptimal conditions for nodulation. Nutrient-amended granules significantly enhanced early nodulation of soybean and increased N content of the grain at harvest in four field trials. All these effects were obtained using an average of 10 kg granules amended with 1.14 kg glycerol and 0.16 kg sodium glutamate per hectare. The possible use of nutrient-amended granules to improve efficacy and reliability of microbial inoculation is discussed. Keyvords: Bradw%i:obium japonicum; Soybean; Inoculation; Inoculant 1. Introduction Inoculation of soybean (Glycine mar (L) Merr.) with Bradyrhizobium japonicum is a common prac- tice of significant agronomic benefit in areas where the crop is planted for the first time [1,2], or in soils where the number of B. japonicum is too low to allow efficient nodulation of the plant [3]. Increasing the number of B. japonicum in such situations in- creases nodule number and/or nodule weight [4] and may improve grain yield 121. The inoculation rate is thus important for the success of inoculation. The efficacy and ultimate economic benefit of soybean inoculation requires optimization of three factors: an effective inoculant population of B. japonicum in a stable formulation, a method for delivering the formulation with or near the seed with minimal losses of bacteria, and sufficient mainte- nance of a viable rhizobial population in the soil to nodulate roots emerging from the germinating seed. The first two factors have received considerable attention, mainly by increasing inoculation rates [ 1,2] and by attempts to decrease rhizobial losses during inoculation by the use of adhesives [5,6] or oils [7]. * Corresponding author. Tel: +33 80 63 32 91: Fax: t33 80 63 32 24; E-mail: [email protected] ’ Present address: Lvcee aaricole, 38200 Vienne-Sessuel. France. In contrast, the stability of inoculated populations 016%6496/96/$15.00 0 1996 Federation of European Microbiological Societies. Published by Elsevier Science B.V. PII SO 168.6496(96)00028- 1
Transcript of Increase of Bradyrhizobium japonicum numbers in soils and enhanced nodulation of soybean (Glycine...

ELSEVIER FEMS Microbiology Ecology 20 ( 1996) 173- 183
Increase of Bradyrhizobium japonicum numbers in soils and enhanced nodulation of soybean (Glycine max CL) merr.)
using granular inoculants amended with nutrients
Georges Fouilleux I, Ckcile Revellin, Alain Hartmann, Gkrard Catroux *
INRA-CMSE, Laboratoire de Microbiologic des Sols INRA, BV 1540. 21034 Dijon Ct!de.r, France
Received 6 February 1996; accepted 22 March 1996
Abstract
Mineral microgranules, amended with nutrients and inoculated with either peat or liquid Bradyrhizobium japonicum inoculants, increased the growth and recovery of the bacterium during laboratory incubation in unsterilized soil. Increases in the range of 1 log unit per g or ml inoculant used were observed in different soil types. B. japonicum showed better survival with nutrient-amended granules than in unamended ones, in soil undergoing desiccation. In a growth chamber experiment, the number of nodules per plant were significantly increased by nutrient-amendment of the granules, but only under suboptimal conditions for nodulation. Nutrient-amended granules significantly enhanced early nodulation of soybean and increased N content of the grain at harvest in four field trials. All these effects were obtained using an average of 10 kg granules amended with 1.14 kg glycerol and 0.16 kg sodium glutamate per hectare. The possible use of nutrient-amended granules to improve efficacy and reliability of microbial inoculation is discussed.
Keyvords: Bradw%i:obium japonicum; Soybean; Inoculation; Inoculant
1. Introduction
Inoculation of soybean (Glycine mar (L) Merr.) with Bradyrhizobium japonicum is a common prac- tice of significant agronomic benefit in areas where the crop is planted for the first time [1,2], or in soils where the number of B. japonicum is too low to allow efficient nodulation of the plant [3]. Increasing the number of B. japonicum in such situations in- creases nodule number and/or nodule weight [4] and
may improve grain yield 121. The inoculation rate is thus important for the success of inoculation.
The efficacy and ultimate economic benefit of soybean inoculation requires optimization of three factors: an effective inoculant population of B. japonicum in a stable formulation, a method for delivering the formulation with or near the seed with minimal losses of bacteria, and sufficient mainte- nance of a viable rhizobial population in the soil to nodulate roots emerging from the germinating seed. The first two factors have received considerable attention, mainly by increasing inoculation rates [ 1,2]
and by attempts to decrease rhizobial losses during
inoculation by the use of adhesives [5,6] or oils [7].
* Corresponding author. Tel: +33 80 63 32 91: Fax: t33 80 63 32 24; E-mail: [email protected]
’ Present address: Lvcee aaricole, 38200 Vienne-Sessuel. France. In contrast, the stability of inoculated populations
016%6496/96/$15.00 0 1996 Federation of European Microbiological Societies. Published by Elsevier Science B.V. PII SO 168.6496(96)00028- 1

174 G. Fouillru et al. / FEMS Microhiolo~q~ Ecology 20 (19961 173-183
and their possible growth in soils between the time of inoculation and root colonization has received little attention, except for the use of biocides and a corresponding resistant-strain of rhizobia [8], or the addition of microbial nutrients to seed pellets [9]. The latter method, which is supposed to encourage the multiplication of rhizobia in soils, gave no clear nodulation responses [9].
Significant increases in the number of rhizobia introduced into soils have been reported in labora- tory experiments following addition of a carbon source to the soil (0.1 to l.% dry wt soil) [lo-121, but, in an agricultural perspective, the introduction of large amounts of a carbon source into the soil matrix is unrealistic and uneconomical. We were conse- quently interested in methods for delivering small amounts of a concentrated carbon substrate along with the inoculant formulation, based on the use of granular carriers [ 13,141. Addition of a carbon-sub- strate to alginate pellets was previously reported to facilitate microbial growth in soils or improve inocu- lum survival of A;ospirillum brasiliense [ 151, Pseu- domonas Jluorescens [ 161, Trichoderma sp. [ 171. Gliocladium cirens [ 171, Talaromyces jlacus [ 181, and Fusarium solani sp. cucurbitae [19]. All these experiments were performed using large amount of inoculant processed with simultaneous inclusion of a carbon-substrate and the microorganism. However, from a practical point of view. such an inoculant containing an available carbon source may be more difficult to protect against contaminants which are known to drastically reduce the shelf life of rhizobial inoculants [20,2 11. In order to avoid this problem, we have used sterilized, nutrient-amended, mineral mi- crogranules inoculated at the time of sowing with peat-based or liquid inoculants. To the best of our knowledge, this is the first report of the use of B. japonicum inoculum on a microgranular carrier amended with a carbon-substrate which allows mul- tiplication of this bacterium in soils and improves nodulation of soybean.
2. Materials and methods
2.1. Bacterial strains and inoculants
B. juponicum strain G49 (originally strain SB16, Division of Microbiology. Indian Agricultural Re-
search Institute, New Delhi) was used for growth chamber and field experiments, in the form of com- mercial peat-based or liquid inoculants (provided by LIPHATEC, Lyon, France). Strain G49RfKm (CNCM No. l- 1264) a spontaneous G49 mutant resistant to rifampicin (100 mg l- ’ > and kanamycin (200 mg 1~ ’ >, was used for soil incubations. Prelimi- nary experiments indicated that the antibiotic-re- sistant and the wild-type strains had similar growth rates in Bergersen’s medium [22] as measured by the increase in optical density at 620 nm. G49RfKm inoculants were prepared by growing the bacteria in 200 ml of a malt extract medium [23] in a 1 1 conical flask agitated at 200 rpm for 6 days at 28°C. The culture broths (3.8 to 5.2 X lo9 cfu ml-‘) were stored as they were at 4°C for several months, with- out any change in bacterial counts, and used directly as liquid or peat-based inoculant. The latter was prepared by mixing the broth with irradiated sterile peat (provided by LIPHATEC, Lyon, France) to give approximately 4 X 10’ cfu B. japonicum per g of moist peat inoculant.
2.2. Granule amendment and inoculation
Three commercial mineral granules preparations were used, the physical characteristics of which are reported elsewhere [24]. M, are bentonite granules (LIPHATEC, Lyon, France), and M, are illite and smectite granules (MicroBio, Royston, UK). Both have a size of 0.2-0.8 mm with an apparent density of 1, and are currently used for soybean inoculation in France. M, are silica granules (Laporte S.A., Coppet, CH) with a size of 0.2-0.8 mm and an apparent density of 0.6. M, granules can absorb 30% of water (w/w> and maintain good gravity flow via a pesticide applicator. Granules were sterilized at 170°C for 1 h before nutrient amendment. The differ- ent amendments are given in Table 1. Concentrated, sterile aqueous solutions of nutrients were added to dry, sterile granules at rates of 12, 14 and 30% (v/w) for M,, M, and M, respectively. Nutrient- amended granules were dried at 60°C and stored dry at room temperature until inoculation.
Laboratory and growth chamber experiments were performed using only M, and M, granules. Peat inoculant (1 to 1.3 g> was mixed, prior to use, with 25 g dry M, and M, granules previously moistened with 3.2 and 7.5 ml sterile water, respectively. This
G. Fouilleux et al. / FEMS Microbiology Ecology 20 f 1996) 173-183 175
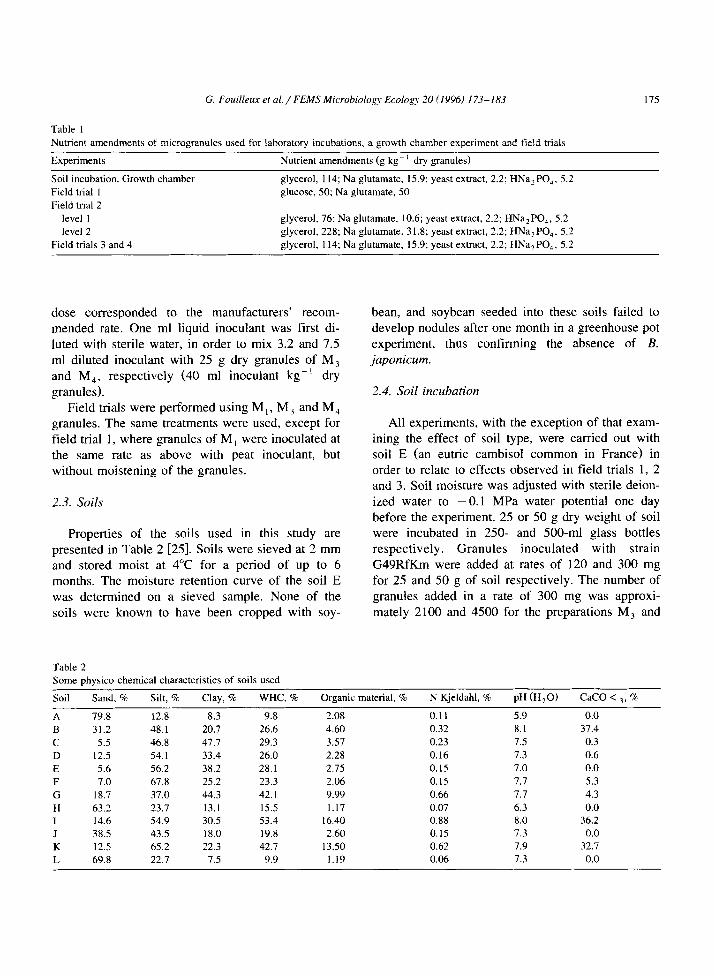
Table 1
Nutrient amendments of microgranules used for laboratory incubations, a growth chamber experiment and field trials
Experiments Nutrient amendments (g kg- ’ dry granules)
Soil incubation, Growth chamber glycerol, 114; Na glutamate, 15.9: yeast extract, 2.2; HNa,PO,, 5.2 Field trial 1 glucose, 50; Na glutamate, 50 Field trial 2
level 1 glycerol, 76: Na glutamate. 10.6; yeast extract, 2.2; HNaaPO,, 5.2 level 2 glycerol, 228; Na glutamate, 3 1.8; yeast extract, 2.2; HNa:PO,, 5.2
Field trials 3 and 4 glycerol, 114; Na glutamate, 15.9; yeast extract, 2.2; HNa,PO,, 5.2
dose corresponded to the manufacturers’ recom- mended rate. One ml liquid inoculant was first di- luted with sterile water, in order to mix 3.2 and 7.5 ml diluted inoculant with 25 g dry granules of M, and M,, respectively (40 ml inoculant kg-’ dry
granules). Field trials were performed using M,, M, and M,
granules. The same treatments were used, except for field trial 1, where granules of M, were inoculated at the same rate as above with peat inoculant, but without moistening of the granules.
2.3. Soils
Properties of the soils used in this study are presented in Table 2 [25]. Soils were sieved at 2 mm and stored moist at 4°C for a period of up to 6 months. The moisture retention curve of the soil E was determined on a sieved sample. None of the soils were known to have been cropped with soy-
bean, and soybean seeded into these soils failed to develop nodules after one month in a greenhouse pot experiment, thus confirming the absence of B. japonicum.
2.4. Soil incubation
All experiments, with the exception of that exam- ining the effect of soil type, were carried out with soil E (an eutric cambisol common in France) in order to relate to effects observed in field trials 1, 2 and 3. Soil moisture was adjusted with sterile deion- ized water to - 0.1 MPa water potential one day before the experiment. 25 or 50 g dry weight of soil were incubated in 250- and 500-ml glass bottles respectively. Granules inoculated with strain G49RfKm were added at rates of 120 and 300 mg for 25 and 50 g of soil respectively. The number of granules added in a rate of 300 mg was approxi- mately 2100 and 4500 for the preparations M, and
Table 2 Some physico-chemical characteristics of soils used
Soil Sand, % Silt, % Clay, % WHC, % Organic material, % N Kjeldahl, 9 pH (Ha01 CaCO < 3, %
A 79.8 12.8 8.3 9.8 2.08 0.11 5.9 0.0 B 31.2 48.1 20.7 26.6 4.60 0.32 8.1 37.4
C 5.5 46.8 47.7 29.3 3.57 0.23 7.5 0.3 D 12.5 54.1 33.4 26.0 2.28 0.16 7.3 0.6 E 5.6 56.2 38.2 28.1 2.75 0.15 7.0 0.0 F 7.0 67.8 25.2 23.3 2.06 0.15 7.7 5.3 G 18.7 37.0 44.3 42.1 9.99 0.66 7.7 4.3 H 63.2 23.7 13.1 15.5 1.17 0.07 6.3 0.0 I 14.6 54.9 30.5 53.4 16.40 0.88 8.0 36.2 J 38.5 43.5 18.0 19.8 2.60 0.15 7.3 0.0 K 12.5 65.2 22.3 42.7 13.50 0.62 7.9 32.7 L 69.8 22.7 7.5 9.9 1.19 0.06 7.3 0.0

176 G. Foui1leu.r et al. / FEMS Microbiology Ecology 20 C 19961 173- 183
M,, respectively. This inoculation rate was close to the manufacturers’ recommended rate, based on the assumptions that the seed-bed is a soil cylinder of 1 cm diameter, with a 30 cm space between plant rows, and inoculated with 10 kg granules per ha delivering a minimum of 4 X 10” viable B. jupon- icum (equivalent to lo6 bacteria per seed). After mixing, bottles were covered with semi-permeable plastic film and incubated at 20°C.
2.6. Growth chamber experiments
To evaluate the bacterial growth occurring during the process, chloramphenicol was added in two ex- periments, at 170 and 140 I_t,g g- ’ dry wt soil to half the treatments for granules inoculated with peat and liquid inoculants, respectively.
A two-step soil incubation experiment was per- formed in order to study the effect of granule amend- ment under conditions of soil desiccation. Incubation was performed with moist soil ( - 0.1 MPa) for one week for all treatments. For the following two weeks, half of the bottles were incubated at -0.1 MPa and the others were exposed to drying by opening the bottles.
All incubations were performed in triplicate. For each experiment, B. japonicum, in the inoculant and in the soil samples, was enumerated just after soil inoculation (t = 0) and after incubation (t = 7). Re- sults are expressed in log cfu related to the amount of inoculant added (per g of peat or ml of liquid inoculant).
For each treatment, five replicate 2.5-l plastic pots filled with soil E were sown with six soybean seeds (cv Maple Arrow), and thinned out to 4 plants after germination. A total of 200 mg granules were placed below the seeds in each pot; this amount is close to that used in agricultural practice for soybean inocula- tion with microgranule inoculant (10 kg microgran- ules for 400 to 600000 seeds ha- ’ ). Growth cham- ber day/night conditions were 13/ 11 h at 17,’ 12°C during the first two weeks, 13/ 1 I h at 19/ 14°C for the third week and 14/ 10 h at 20/ 16°C after three weeks with a lighting of 240 FE s-’ rn-‘. Pots were irrigated with deionized water and soil water poten- tial was maintained at - 5, - 0.1, or 0 MPa for the first ten days after sowing then at -0.1 MPa until the end of the experiment, by addition of deionized water to constant weight. The increase in plant weight was not taken into account for soil moisture correc- tion, as it produces only a maximum error of 5% in moisture content. M, and M, granules were inocu- lated with commercial peat and liquid inoculants in order to deliver lo6 (recommended rate) and 10” (I/ 10 recommended rate) B. juponicum per seed. Plate count numbers (lOh rate) were 5.6 X IO6 and 6.6 X lo6 for peat and liquid inoculant, respectively. Plants were harvested at the early growth stage V, [26] for nodulation assessment.
2.5. Enumeration of B. japonicum 2.7. Field experiments
Enumeration of B. juponicum was performed by plate counts on Bergersen’s medium [22], using aliquots of 1 ml of liquid inoculant or 1 g of peat inoculant or inoculated granules. Plates were inocu- lated with suspension-dilutions using the Spiral Sys- tem’” (Interscience, St. Nom la Breteche, France) and incubated for 6 days at 28°C. G49 RfKm popula- tions in soil were monitored by plate counts using a double layer technique, with the same medium sup- plemented with (mg 1~ ‘1 rifampicin (I 001, kanamycin (2001, cycloheximide (1001, benlate (251, and quintozene (25). Soil samples (25 or 50 g) treated with M, and M, granules were shaken in 100 ml of sterile sodium metaphosphate solution (2% w/v) or sterile deionized water, respectively, at 200 rpm for 30 min.
Field experiments were performed in soil E (trials 1. 2, 3) and in soil L (trial 41, using a randomized block design with four replicates per treatment. Plots were 2 X 10 m in size and each contained 6 plant rows. Soil preparation, weed control and irrigation were performed using methods commonly used in France for the commercial production of early matur- ing soybean. Cv Maple Arrow seeds were sown at a density of 600 000 per ha and inoculated with 10 kg inoculant granules per ha, delivered below the seeds. Nodule number was determined at stages V, and R, [26] on samples of 10 randomly chosen plants per plot. The nitrogen content of the plants and grains was determined by the Kjeldahl method [27]. For trial 1, M, granules inoculated with peat inoculant were used at a rate of 3.5, 1.0 and 8.7 X lo6 B.

G. Fouilieux et al. / FEMS Microbiology Ecology 20 (19%) 173-183 177
Table 3
Changes in the number of B. japonicum after incubation of peat and liquid inoculated M, and M, granules in soil E
Treatment Log ‘O number of B. japonicum ’ after time (t) in days
M 3 + Peat inoculant ‘.a
t=O t=7
M, + Liquid inoculant 2.3
t=o t=7
Unamended granules WAG)
Control
Chloramphenicol
8.27’ 8SXd’ 9.05c 9.08’
7.97” 7.58’ 9.05c 8.7fjd
Amended granules (AG)
Control 8.78b
Chloramphenicol 8.70b
’ Per g of peat or ml of liquid inoculant added.
9.25” 9.20b 10.11’
8.7 lb 9.15s’ 9.10c
’ Data pooled from the two columns (t = 0) and (t = 7) followed by the same letters, are not significantly different.
’ Initial amounts of B. japonicum added were 9.38 log units per g peat inoculant and 9.70 log units per ml liquid inoculant.
japonicum per seed, for granule, seed and seed bed liquid inoculation, respectively. Trial 2 was per- formed with M, granules inoculated with peat inocu- lant (60 g kg-l) at a rate corresponding to 2.6 X lo6 B. japonicum per seed. In trials 3 and 4, M, gran- ules prepared either with liquid inoculant or with peat inoculant at a rate of 4.5 or 4.3 X lo6 B. japonicum per seed were used, respectively.
2.8, Data processing
Statistical analyses were performed using STAT- ITCF software [28] for correlation, linear regression, variance analysis and Newman-Keuls tests, and sig- nificant differences were tested at the P = 0.05 level.
3. Results
3.1. B. japonicum recovery and growth after incuba- tion in soil E
Whatever the inoculant type (peat or liquid), the use of nutrient-amended granules (AG) significantly increased the number of viable B. japonicum recov- ered immediately after inoculation (to) and one week after incubation as compared to unamended granules &JAG). Numbers recovered immediately after inocu- lation (t,) were significantly higher for AG than for UAG for both peat and liquid inoculants (Table 3). After one week of incubation B. japonicum numbers
increased in AG treatments by 0.47 and 0.91 log cfu per g peat and per ml liquid inoculants respectively. No increase or a small decrease (peat) in B. japon- icum numbers was found in UAG-treated soils, and in all chloramphenicol-treated soils, incubated for one week. AG gave significantly higher viable counts
Table 4 Changes in the number of B. japonicnm after incubation of liquid
inoculated M, granules in different soils
Soil tested Log’0 number of B. japonicum after time (tl in days ‘Z
Unamended WAG) Amended (AG)
t=o t=7 t=o t=7
A 8.83Jk’m 8.96h” 9.1v’ 10.02b’
B 8.74Wm 8.881Jklm 9.18e’ 9.90c
C 8.86”k’m 9.12efg 9.WQ 10.18”
D 8.81Jklm 9.00ah1 9.15e@ 10.02bC
E 8.82Jklrn 9.0iYh’ 9. IS” 10.02bC
F 8.76k’m 8.55” 9.07’@ 9.49d
G 8.84’Jkim 8.70rn 9.12- 9.98’
H 8.82 Jklm 8.38“ 9.23cf 9.27e I 8.90iJk 8.81Jkirn 9.17e’ 10.W”
J 8.80Jk’“’ 8.88’Jk’ 9.16rf 10.08”b
K 8.711rn 8.70rn 9.13e’g lO.OOb’
Mean ’ 8.81’ 8.82’ 9.15b 9.92”
’ Per ml liquid inoculant added. a Data from all columns, followed by the same letters, are not significantly different.
’ Initial amount of B. japonicum added was 9.71 log units per ml
liquid inoculant.
A Means followed by the same letter are not significantly different.
178 G. Fouilleux et al. / FEMS Microbiology Ecology 20 (lYY61 173-183
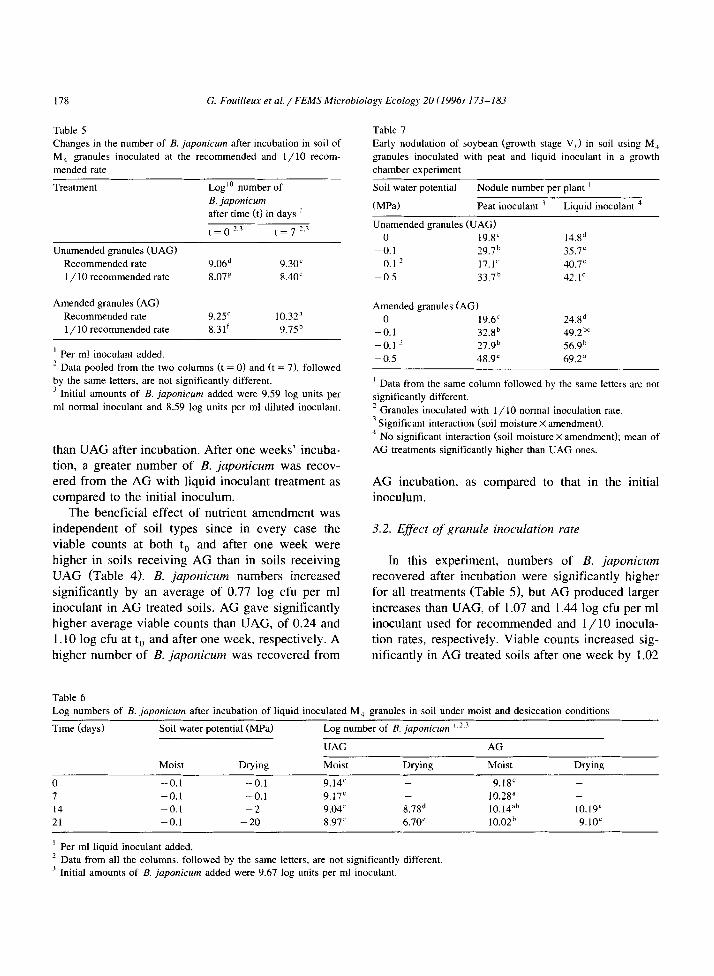
Table 5 Table 7
Changes in the number of B. japonicum after incubation in soil of
M, granules inoculated at the recommended and l/10 recom-
mended rate
Early nodulation of soybean (growth stage V,) in soil using M,
granules inoculated with peat and liquid inoculant in a growth
chamber experiment
Soil water potential Nodule number per plant ’
(MPa) Peat inoculant ’ Liquid inoculant ’
Unamended granules (UAG)
Treatment Log’0 number of
B. japonicum after time (tf in days ’
Unamended granules (IJAG)
Recommended rate
1 / 10 recommended rate
t = 0 2.3 t = 7 2.3
9.06d 9.30’
8.07’ 8.40’
Amended granules (AC)
Recommended rate
1 / 10 recommended rate
’ Per ml inoculant added.
9.25’ 10.32”
8.31’ 9.75b
’ Data pooled from the two columns (t = 0) and (t = 7), followed
by the same letters, are not significantly different.
’ Initial amounts of B. juponicum added were 9.59 log units per
ml normal inoculant and 8.59 log units per ml diluted inoculant.
than UAG after incubation. After one weeks’ incuba- tion, a greater number of B. japonicum was recov- ered from the AG with liquid inoculant treatment as compared to the initial inoculum.
The beneficial effect of nutrient amendment was independent of soil types since in every case the viable counts at both t, and after one week were higher in soils receiving AG than in soils receiving UAG (Table 4). B. japonicum numbers increased significantly by an average of 0.77 log cfu per ml inoculant in AG treated soils. AG gave significantly higher average viable counts than UAG, of 0.24 and 1.10 log cfu at t, and after one week, respectively. A higher number of B. japonicum was recovered from
0 19.8’ 14.gd -0.1 29.7b 35.7c -0.1 ? 17.1C 40.7’ - 0.5 33.7b 42.1’
Amended granules (AG)
0 19.6’ 24.gd - 0.1 32.gb 49.2b’ -0.1 z 27.9b 56.9b -0.5 48.9” 69.2”
’ Data from the same column followed by the same letters are not
significantly different.
’ Granules inoculated with l/10 normal inoculation rate.
’ Significant interaction (soil moisture X amendment).
’ No significant interaction (soil moisture X amendment); mean of
AG treatments significantly higher than UAG ones.
AG incubation, as compared to that in the initial inoculum.
3.2. Effect of granule inoculation rate
In this experiment, numbers of B. japonicum recovered after incubation were significantly higher for all treatments (Table 51, but AG produced larger increases than UAG, of 1.07 and 1.44 log cfu per ml inoculant used for recommended and l/10 inocula- tion rates, respectively. Viable counts increased sig- nificantly in AG treated soils after one week by 1.02
Table 6
Log numbers of B. juponicum after incubation of liquid inoculated M, granules in soil under moist and desiccation conditions
Time (days) Soil water potential (MPa) Log number of B. japonicum ‘A’
UAG AG
Moist Drying Moist Drying Moist Drying
0 -0.1 -0.1 9.14’ _ 9.18’ -
7 -0.1 -0.1 9.17< _ 10.28* -
14 -0.1 -2 9.04c 8.7gd 10.14”b 10.19”
21 -0.1 -20 8.97’ 6.70r 10.02b 9.10’
’ Per ml liquid inoculant added. a Data from all the columns. followed by the same letters, are not significantly different.
’ Initial amounts of B. japonicum added were 9.67 log units per ml inocuiant.
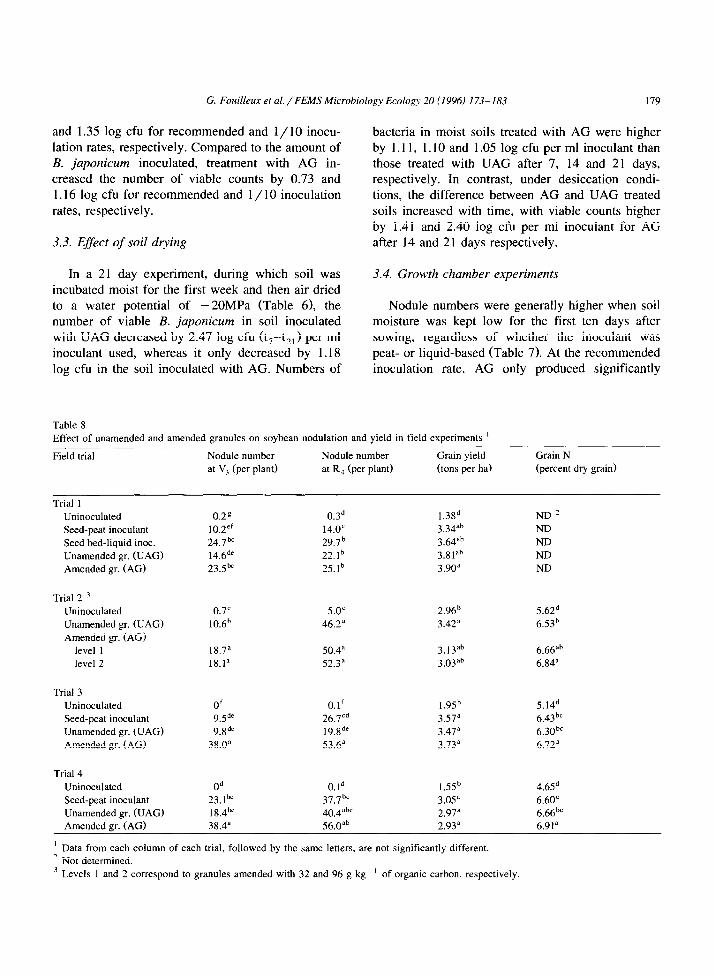
G. Fouilleux et al. /FEM.!? Microbiology Ecology 20 (I996) 173-183 179
and 1.35 log cfu for recommended and 1 / 10 inocu- lation rates, respectively. Compared to the amount of B. japonicum inoculated, treatment with AG in- creased the number of viable counts by 0.73 and 1.16 log cfu for recommended and 1 / 10 inoculation rates, respectively.
bacteria in moist soils treated with AG were higher by 1.11, 1.10 and 1.05 log cfu per ml inoculant than those treated with UAG after 7, 14 and 21 days, respectively. In contrast, under desiccation condi- tions, the difference between AG and UAG treated soils increased with time, with viable counts higher by 1.41 and 2.40 log cfu per ml inoculant for AG
3.3. Effect of soil dtying after 14 and 21 days respectively.
In a 21 day experiment, during which soil was incubated moist for the first week and then air dried to a water potential of - 20MPa (Table 6), the number of viable B. japonicum in soil inoculated with UAG decreased by 2.47 log cfu (t7-t?,) per ml inoculant used, whereas it only decreased by 1.18 log cfu in the soil inoculated with AG. Numbers of
3.4. Growth chamber experiments
Nodule numbers were generally higher when soil moisture was kept low for the first ten days after sowing, regardless of whether the inoculant was peat- or liquid-based (Table 7). At the recommended inoculation rate, AG only produced significantly
Table 8
Effect of unamended and amended granules on soybean nodulation and yield in field experiments ’
Field trial Nodule number
at V, (per plant)
Nodule number
at R, (per plant)
Grain yield
(tons per ha)
Grain N
(percent dry grain)
Trial 1 Uninoculated
Seed-peat inoculant
Seed bed-liquid inoc.
Unamended gr. &JAG)
Amended gr. (AG)
0.2 a 0.3d 1 .38d ND’
10.2ef 14.0’ 3.34ab ND
24.7k 29.1b 3.64ab ND
14.6de 22. lb 3.8 lab ND
23.+ 25.1b 3.90” ND
Trial 2 ’
Uninoculated
Unamended gr. (IJAG)
Amended gr. (AG)
level 1
level 2
o.7c 5.0’ 2.96b 5.62d
10.6b 46.2” 3.42” 6.53b
18.7a 50.4a 3.13as 6.66‘+
18.1” 52.3a 3.03as 6.84a
Trial 3 Uninoculated
Seed-peat inoculant
Unamended gr. WAG)
Amended gr. (AG)
0’ 0.1’ 1.95b 5.14d
9.5de 26.7cd 3.57a 6.43 bc
9.8d’ 19.8de 3.47a 6.30bc 38.0” 53.6= 3.73” 6.72a
Trial 4
Uninoculated Od 0.P 1.55b
Seed-peat inoculant 23. lbc 37.7sc 3.05a
Unamended gr. WAG) 18.4& 40.4abc 2.97a Amended gr. (AG) 38.4” 56.0ab 2.93a
’ Data from each column of each trial, followed by the same letters, are not significantly different.
’ Not determined.
’ Levels 1 and 2 correspond to granules amended with 32 and 96 g kg-’ of organic carbon, respectively
4.65d
6.60C
6.66bC 6.91a

180 G. Fouiileux et al. / FEMS Microbiology Ecology 20 (1996) 173-183
higher numbers of nodules than UAG when soil water potential was kept at - 0.5 MPa. At 1 /IO of the recommended inoculation rate, the number of nodules was higher in AG treated soils than in UAG treated soils when water potential was -0.1 MPa.
3.5. Field experiments
In four independent field trials, AG consistently yielded significantly higher numbers of nodules at the early growth stage V, than UAG (Table 8) but this effect tended to disappear at growth stage R, except in one trial. Nutrient amendment did not produce a significant increase in grain yield but the nitrogen content of grains was increased in two of the three trials where this parameter was measured.
4. Discussion
The use of nutrient-amended granules (AG) as carrier for soybean inoculant improved the immedi- ate recovery of B. juponicum after granule inocula- tion. Recovery from AG treated soils was generally higher than from UAG treated soils. This may be explained by a protection against desiccation pro- vided by the nutrient organic substrate added in the granules similar to that reported previously for car- bohydrates and rhizobial cultures [29,30]. Desicca- tion was reported recently to be a factor influencing the immediate recovery of B. juponicum after mix- ing with granules [24]. Thus. it may be supposed that amendment can fill a part of the granule porosity, decreasing the possibility of inoculant desiccation in dry granules. Nutrient-amendment promoted the sur- vival of B. japonicum introduced in soil undergoing desiccation. This resistance to desiccation may be the consequence of bacterial growth in the soil and a possible protection of cells in microcolonies, as pro- posed previously to explain the better survival of soil grown Bradyrhizobium sp. (lupini) and Rhizobium meliloti in soil undergoing drying [31].
Nutrient-amended granules significantly increased B. japonicum numbers after incubation in soils. One possible mechanism is cell division without biomass increase due to the introduction of the bacteria into an oligotrophic medium like soil, as reported for R.
leguminosarum bv. trifolii [32]. Some small in- creases observed without nutrient-amendment sup- port this observation, but bacterial growth probably remains the most important mechanism, as demon- strated by the effect of chloramphenicol on soil incubation (Table 3). An average increase of 0.77 log unit cfu per ml inoculant was found after 7 days incubation for amended granules (derived from Table 4). corresponding to 2.5 cell divisions. The relative contribution of the different nutrients added is un- known.
Increases in bacterial number after incubation were independent of soil type (Table 4) and could not be related to most of the soil characteristics (Table 2), as reported previously for the survival of introduced B. juponicum [33]. However in this work, the log cfu of bacteria recovered after one week in AG treated soils (Table 4) is correlated to the silt content (I = 0.646; r’ = 0.417; P = 0.030) and the difference in log cfu between AG and UAG treated soils after one week (Table 41, equivalent to the net microbial growth due to the amendment, is correlated with the soil organic matter content (r = 0.783; r’ = 0.613; P = 0.004).
In order to assess the percentage of carbon sub- strate utilization by the inoculated B. japonicum, we assumed that the bacteria have the same yield when mixed with the amended granules, as in liquid cul- ture. A standard culture medium [22] can produce lo9 bacteria ml-’ with 1 g 1-l of organic carbon corresponding to 10” bacteria per g of carbon. It can be deduced from Table 4 that the net increase in number of cells by amended granules is 0.9 X lo9 per ml liquid inoculant. As 1 kg of granules is amended with 48 g of organic carbon and inoculated with 40 ml inoculant, 1 ml inoculant corresponds to 1.2 g of carbon, which is able to sustain the produc- tion of 1.2 X 10” cells. Therefore, after inoculation onto granules. B. juponicum only utilizes approxi- mately 0.1% of the available substrate.
The results from soil incubations indicate that nutrient-amendment can promote the growth of a B. japonicum inoculant in soil, and its survival in soil undergoing drying. Thus, in practical terms, it may enable an increased inoculation rate, or recovery of the recommended inoculation rate, when using di- luted or poor quality inoculants. This may be of significant economic importance as increasing the

G. Fouilleux et al. / FEMS Microbiology Ecology 20 (1996) 173-183 181
number of B. japonicum inoculated per seed can increase the number of nodules per plant [4,34,35], the plant nitrogen content [4], the grain nitrogen content [34] and the grain yield [2,34-371.
Since the observed effect of nutrient-amendment was to increase the number of B. japonicum in soil, it may be expected that inoculation with amended granules would improve soybean nodulation. This hypothesis was confirmed by the results from a growth chamber experiment in suboptimal conditions of inoculation rate and soil moisture, and in four independent field trials.
Enhanced soybean nodulation may also be due to the direct action of the organic amendment, as re- ported previously [38]. However, the efficient con- centrations in the nutrient solution used in these experiments were 12 mM and 0.34 mM for glucose and glutamate, respectively. These values are IO- to 20-fold higher than the concentrations used in our work when extrapolated to field conditions. Thus,
our results are probably not due to a direct stimula- tion of nodulation by the amendment. Previous re- sults [ 121 have demonstrated the possibility of in- creasing the number of indigenous B. japonicum in soil by adding sugars to soil at a rate of 0. I % (w/w) every three days. After 12 and 36 days, increases in numbers of IO- and 1 OO-fold were observed with a total sugar supply of 0.4 and 1.2% of soil (w/w), respectively. Similar increases were observed for indigenous R. leguminosarum and R. meliloti when incorporating various organic substrates at 1% of soil (w/w> [IO]. Increases in numbers of introduced bac- teria were observed for B. juponicum [ 111 with a supply of 0.5% mannitol, and for Agrobacterium, Pseudomonas and Coyebacterium with a 1% glu- cose amendment [39]. In order to compare these results with our data, we assessed the volume of soil inoculated, i.e. the seed-bed. It may correspond to approximately 2.6 X IO6 cm3, or 3 900 kg of soil per ha (cylinder of 1 cm diameter per row, 30 cm space between plant rows). Amendment of the seed bed with 0.5 or 1% organic substrate will give amounts of 19.5 to 39 kg ha-‘, respectively. These amounts are probably uneconomical as they represent an un- acceptable increase in the cost of inoculation. On the other hand, our data have demonstrated that it is possible to increase the efficacy of inoculation with the supply of only 1 kg ha-’ of organic carbon
nutrients, providing that the substrate and the bacte- ria are localized on granules.
The proposed inoculation method involves the growth in soil of a microorganism that has been introduced on granules amended with nutrients. In order to avoid competition with the indigenous soil microflora, it may be possible to use a nutrient that is specifically assimilated by the inoculant microorgan- ism or use pesticides to control competitors, as re- ported previously [40]. The proposed method should improve efficacy and reliability of microbial inocula- tion of seeds and soils. Research efforts are currently applying this technology to other rhizobial strains and biocontrol agents.
Acknowledgements
This work was supported by the Conseil Regional de Bourgogne, France, and by LIPHATEC, Lyon, France. We thank Jean-Jacques Giraud and Georges Sommer for technical assistance and Vivienne Giani- nazzi-Pierson (INRA-CMSE, Laboratoire de Phy- toparasitologie, Dijon, France), Pauline Mele (De- partment of Agriculture, Rutherglen Research Insti- tute, Rutherglen, Victoria, Australia), and Nisha Parekh (NERC Institute of Terrestrial Ecology, Grange-over-Sands, Cumbria, UK) for helpful dis- cussion and advice.
References
111 Brockwell, J., Herridge, D.F., Morthorpe, L.J. and Roughley.
R.J. (1985) Numerical effects of Rhizobium population on
legume symbiosis. In: Nitrogen Fixation by Legumes in
Mediterranean Agriculture (Beck, D.P. and Materon, L.A..
Eds.), pp. 179-193. Martinus Nijhoff Publishers, Dordrecht.
The Netherlands.
[2] Hume, D.J. and Blair. D.H. (1992) Effect of numbers of
Br&rhkobium japonicum applied in commercial inoculants on soybean seed yield in Ontario. Can. J. Microbial. 38.
588-593.
[3] Singleton. P., Thies, J. and Bohlool. B.B. (1992) Useful models to predict response to legume inoculation. In: Biolog-
ical Nitrogen Fixation and Sustainability of Tropical Agricul-
ture, (Mulongoy, K., Gueye, M. and Spencer, S.D.C.. Eds.), pp. 245-256. John Wiley and Sons, Inc.. New York.
[4] Weaver, R.W. and Frederick, L.R. (1972) Effect of inoculum
size on nodulation of Gl@ze max (L.) Merril, variety Ford.
Agron. J. 64, 597-599. [5] Davidson, F. and Reuszer, H.W. (1978) Persistence of Rhi-

G. Fouilleux et al. / FEMS Microhiologx Ecology 20 f 19%) 173-183 182
t61
[71
[81
191
1101
[111
LIZI
1131
1141
1151
I161
iI71
1181
[I91
EOI
zobium japonicum on the soybean seed coat under controlled inoculants. In: A Treatise on Dinitrogen Fixation (Hardy.
temperature and humidity. Appl. Environ. Microbial. 35. R.W.F. and Gibson, A.H.A., Eds.). pp. 243-276. John Wiley
94-96. and Sons, Inc., New York.
Elegba. M.S. and Rennie. R.J. (1984) Effect of different
inoculant adhesive agents on rhizobial survival. nodulation,
and nitrogenase (acetylene-reducing) activity of soybeans
(G(vcine MAX (L.) Merrill). Can. J. Soil Sci. 64, 631-636.
Hoben. H.J.. Aung. N.N., Somasegaran, P. and Kang, U.G.
(1991) Oils as adhesives for seed inoculation and their
influence on the survival of Rhkobium spp. and Bradyhko-
bium spp. on inoculated seeds. World J. Microbial. Biot. 7.
324-330.
[2l] Rodriguez-Navarro. D.N., Temprano, F. and Orive. R. (1991)
Survival of Rhkobium sp. (Hedysarum coronurium L.) on
peat-based inoculants and inoculated seeds. Soil Biol.
Biochem. 23, 375-379.
Siddiqi, M.A. and Alexander, M. (1991) Use of aliette, a
basipetally translocated antimicrobial compound, to enhance
rhizophere colonization and nodulation by root-nodule bacte-
ria. Biol. Fert. Soils. 12. 141-146.
Brockwell, J. (1977) Application of legume seed inoculants,.
In: A Treatise on Dinitrogen Fixation (Hardy, R.W.F. and
Gibson. A.H.A.. Eds.). pp. 277-309. John Wiley and Sons,
Inc.. New York.
[22] Bergersen, F.J. (1961) The growth of Rhkobirtm in synthetic
media. Aust. J. Biol. Sci. 14, 349-360.
[23] Iswaran, V.. Graham, P.H. and Jauhri. KS. (1969) Modified
Burton’s medium for large-scale production of culture of
Rhizobium jnponicum. Curr. Sci. 38. 346.
[24] Fouilleux. G.. Revellin. C. and Catroux, G. (1994) Short
term recovery of Bradwhizobium japonicum during the inoc-
ulation process using mineral microgranules. Can. J. Micro-
biol. 40, 322-325.
[25] Klute. A. (1986) Methods of Soil Analysis. American Soci-
ety of Agronomy, Inc., Soil Science Society of America, Inc., Madison.
Germida, J.J. (1988) Growth of indigenous Rhi,-obium legu-
minosarum and Rhizobium meliloti in soils amended with
organic nutrients. Appl. Environ. Microbial. 54, 257-263.
Pena-Cabriales, J.J. and Alexander. M. (1983) Growth of
Rhkobium in soil amended with organic matter. Soil Sci.
Sot. Am. J. 47. 241-245.
[26] Fehr. W.R., Caviness. C.E., Burmood, D.T. and Pennington,
J.S. (1971) Stage of developmenr descriptions for soybean
G!\cine mar L. Merril. Crop Sci. 71. 929-931.
[27] Helrich, K. (1990) Official Methods of Analysis. Association
[281
Viteri. S.E. and Schmidt, E.L. (I 987) Ecology of indigenous
soil rhizobia: response of Bradyrhizobium japonicum to
readily available substrates. Appl. Environ. Microbial. 53.
l872- 1875.
E91
Brockwell, J.. Gault. R.R.. Chase. D.L.. Hely. F.W.. Zorin.
M. and Corbin, E.J. (1980) An appraisal of practical alterna-
tives to legume seed inoculation: field experiments on seed
bed inoculation with solid and liquid inoculants. Aust. J.
Agric. Res. 31. 47-60.
Smith, R.S. (1992) Legume inoculant formulation and appli-
cation. Can. J. Microbial. 38, 485-492.
Bashan. Y. (1986) Alginate beads as synthetic inoculant
carriers for slow release of bacteria that affect plant growth.
Appl. Environ. Microbial. 5 1, 1089-1098.
Van Elsas, J.D., Trevors, J.T., Jain, D., Wolters, A.C.,
Heijnen. C.E. and Van Overbeek. L.S. (1992) Survival of,
and root colonization by. alginate-encapsulated P.WU-
domonas j7uorescens cells following introduction into soil.
Biol. Fert. Soils. 14. 14-22.
1301
of Official Analytical Chemists, Inc., Arlington.
Institut Technique des C&ales et des Fourrages (1991)
STAT-ITCF. Manuel d’utilisation. Institut Technique des C&&ales et des Fourrages, Paris. France.
Bushby, H.V.A. and Marshall, K.C. (1977) Some factors
affecting the survival of root-nodules bacteria on desiccation.
Soil Biol. Biochem. 9. 143-147.
Salema. M.P.. Parker, C.A., Kidby, D.K. and Chatel. D.L.
(1982) Death of Rhizobia on inoculated seed. Soil Biol.
Biochem. 14. 13-14.
1311 Pena-Cabriales. J.J. and Alexander, M. (1979) Survival of
Rhizobium in soils undergoing drying. Soil Sci. Sot. Am. J.
43, 962-966.
1321
[331
Lewis, J.A. and Papavizas, G.C. (1985) Characteristics of
alginate pellets formulated with Trichoderma and Gliock-
dium and their effect on the proliferation of the fungi in soil. Plant Pathol. 34, 57 I-577.
Papavizas. G.C.. Fravel, D.R. and Lewis, J.A. (1987) Prolif-
eration of Talaromycesflarus in soil and survival in alginate pellets. Phytopathology 77, 13 1- 136. Weidemann, G.J. (1988) Effects of nutritional amendments
on conidial production of Fusarium soloni f. sp. cucurbitae
on sodium alginate granules and on control of Texas gourd. Plant Dis. 72, 757-759.
Date, R.A. and Roughley, R.J. (1977) Preparation of legume
[341
1351
[361
Postma. J., Van Elsas, J.D., Govaert, J.M. and Van Veen.
J.A. (I 988) The dynamics of Rhbobium /egumino.ww~r
biovar trifolii introduced into soil aa determined by im-
munofluorescence and selective plating techniques. FEMS
Microbial. Ecol. 53. 25 I-260.
Mahler. R.L. and Wollum, G. (1981) The influence of soil
water potential and soil texture on the survival of Rhkobium
juponicum and Rhizobiwn legurnir~osarurn isolates in the
soil. Soil Sci. Sot. Am. J. 45, 761-766.
Herridge. D.F., Roughley. R.J. and Brockwell. J. (1984)
Effect of rhizobia and soil nitrate on the establishment and
fonctioning of the soybean symbiosis in the field. Aust. J. Agric. Res. 35, 149-161.
Brockwell, .I., Gault. R.R., Chase. D.L.. Turner. G.L. and
Bergersen. F.J. (1985) Establishment and expression of soy- bean symbiosis in a soil previously free of Rhkobirtm japon-
icum. Aust. J. Agric. Res. 36, 397-409.
Obaton, M. and Rallier, M. (1970) L’inoculation du soja: influence de la qualitt de I’inoculum sur le rendement en grain et la richesse en proteine de la recolte. C.R. Sc6ances
Acad. Agric. Fr. 56, I l74- 1189.

G. Fouilleux er al. / FEMS Microbiology Ecokog.v 20 (19961 173-183 183
[37] Lagacherie, B. and Obaton, M. (1973) L’inoculation du soja.
R&hats d’essais et orientation du travail. C.R. ScCances
Acad. Agric. Fr. 59. 67-79.
[38] Kaneshiro, T. and Kwolek, W.F. (1985) Stimulated nodula-
tion of soybeans by Rhizobium japonicum mutant [B-14075)
that catabolizes the conversion of tryptophan to indol-3yl-
acetic acid. Plant Sci. 42, 141-146.
[39] Acea, M.J. and Alexander. M. (1988) Growth and survival of
bacteria introduced into carbon-amended soil. Soil Biol.
Biochem. 20, 703-709.
[40] Alexander. M. (1985) Enhancing nitrogen fixation by use of
pesticides: a review. Adv. Agron. 38, 267-282.




![Tolerance of Maize (Zea mays L.) and Soybean [Glycine max (L.) Merr.] to Late Applications of Postemergence Herbicides](https://static.fdokumen.com/doc/165x107/63438e5b1a2cfc44fc019bae/tolerance-of-maize-zea-mays-l-and-soybean-glycine-max-l-merr-to-late-applications.jpg)




![Purification, molecular cloning and ethylene-inducible expression of a soluble-type epoxide hydrolase from soybean (Glycine max [L.] Merr](https://static.fdokumen.com/doc/165x107/631a26e0b41f9c8c6e0a1180/purification-molecular-cloning-and-ethylene-inducible-expression-of-a-soluble-type.jpg)







![KARAKTERISTIKEKSTRAK AIR DAUN PUGUNTANO [Curanga fel-terrae (Lour.) Merr.]YANG BERPOTENSI SEBAGAI ANTELMINTIK](https://static.fdokumen.com/doc/165x107/633999c8a16e1672830dae6a/karakteristikekstrak-air-daun-puguntano-curanga-fel-terrae-lour-merryang-berpotensi.jpg)


