
In vivo definition of parieto-motor connections involved in planning of grasping movements
13
In vivo definition of parieto-motor connections involved in planning of grasping movements Giacomo Koch a,b, ⁎, Mara Cercignani c , Cristiano Pecchioli a , Viviana Versace a , Massimiliano Oliveri a,d , Carlo Caltagirone a,e , John Rothwell f , Marco Bozzali c a Laboratory of Clinical and Behavioural Neurology, Santa Lucia Foundation IRCCS, Via Ardeatina 306, 00179 Rome, Italy b Stroke Unit, Policlinico Tor Vergata, University of Rome Tor Vergata, Viale Oxford, 00133 Rome, Italy c Neuroimaging Laboratory, Santa Lucia Foundation IRCCS, Via Ardeatina 306, 00179 Rome, Italy d Department of Psychology, University of Palermo, Palermo, Italy e Department of Neuroscience, University of Rome Tor Vergata, Via Montpellier 1, 00133 Rome, Italy f Sobell Department of Motor Neuroscience, Queen Square, UCL, London, UK abstract article info Article history: Received 19 November 2009 Revised 7 February 2010 Accepted 9 February 2010 Available online 13 February 2010 Keywords: TMS Connectivity Movement planning Superior longitudinal fasciculus Tractography Transcranial magnetic stimulation We combined bifocal transcranial magnetic stimulation (TMS) and diffusion tensor imaging (DTI) tractography to investigate in humans the contribution of connections originating from different parietal areas in planning of different reaching to grasp movements. TMS experiments revealed that in the left hemisphere functional connectivity between the primary motor cortex (M1) and a portion of the angular gyrus (AG) close to the caudal intraparietal sulcus was activated during early preparation of reaching and grasping movements only when the movement was made with a whole hand grasp (WHG) towards objects in contralateral space. In contrast, a different pathway, linking M1 with a part of the supramarginal gyrus (SMG) close to the anterior intraparietal sulcus, was sensitive only to the type of grasp required (precision grasping) but not to the position of the object in space. A triple coil experiment revealed that inactivation of the ventral premotor area (PMv) by continuous theta burst stimulation interfered with some of these interactions. Anatomical DTI tractography revealed that AG and SMG are strongly connected with PMv and with M1 by different bundles of the superior longitudinal fasciculus (SLF). These results demonstrate the existence of segregated parieto-premotor-motor pathways crucial for preparation of different grasping actions and indicate that these may process information relevant to both the position of the object and the hand shape required to use it. © 2010 Elsevier Inc. All rights reserved. Introduction Although reaching and grasping a cup to drink a coffee seems to be almost automatic, successful performance requires a complex inter- action of specific frontal and parietal brain networks to produce the optimum combination of arm and hand movement to manipulate the object most efficiently (Jeannerod et al., 1995). Classically, this type of action is supposed to involve cortical interconnected circuits including the parietal cortex, the premotor cortex and the primary motor cortex (Rizzolatti and Luppino, 2001). Experiments in monkeys have shown that the inferior parietal lobule (IPL) contains a rich variety of neurons that discharge in association with reaching and grasping movements directed to specific goals (Sakata et al., 1995; Murata et al., 2000; Fogassi et al., 2005; Rozzi et al., 2008). Motor responses are somatotopically organized in a rostro-caudal fashion. In regions of the anterior intraparietal area (AIP), such as the PF and PFG, the most commonly represented motor acts are grasping with the mouth and hand grasping. In contrast, the most commonly represented act in more caudal areas, such as area PG, is arm reaching, (e.g. Rozzi et al., 2008). Human imaging experiments have tended to confirm this, revealing that more anterior regions of the IPL, located in the supramarginal gyrus (SMG), seem crucially involved in precise grasping movements, so that this area has already been proposed as the homologue region of the AIP described in monkeys (hAIP) (Binkofski et al., 1998; Culham and Valyear, 2006; Olivier et al., 2007; Begliomini et al., 2007), while more posterior regions of the IPL, located in the angular gyrus (AG) close to the caudal intraparietal sulcus, have been associated with preparation of visually guided reaching movements (Jeannerod et al., 1995; Castiello, 2005; Karnath and Perenin, 2005). It has been postulated that the different subregions of the IPL supply PMv with visual information about the attributes of objects (e.g., size, orientation) that are needed to select appropriate grasp configurations (Murata et al., 1997; Rizzolatti and Luppino, 2001; Raos et al., 2006). Such information would eventually converge onto NeuroImage 51 (2010) 300–312 ⁎ Corresponding author. Fondazione S. Lucia IRCCS, Via Ardeatina 306, 00179 Rome, Italy. Fax: +39 6 51501388. E-mail addresses: [email protected], [email protected] (G. Koch). 1053-8119/$ – see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.neuroimage.2010.02.022 Contents lists available at ScienceDirect NeuroImage journal homepage: www.elsevier.com/locate/ynimg
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of In vivo definition of parieto-motor connections involved in planning of grasping movements

NeuroImage 51 (2010) 300–312
Contents lists available at ScienceDirect
NeuroImage
j ourna l homepage: www.e lsev ie r.com/ locate /yn img
In vivo definition of parieto-motor connections involved in planning ofgrasping movements
Giacomo Koch a,b,⁎, Mara Cercignani c, Cristiano Pecchioli a, Viviana Versace a, Massimiliano Oliveri a,d,Carlo Caltagirone a,e, John Rothwell f, Marco Bozzali c
a Laboratory of Clinical and Behavioural Neurology, Santa Lucia Foundation IRCCS, Via Ardeatina 306, 00179 Rome, Italyb Stroke Unit, Policlinico Tor Vergata, University of Rome Tor Vergata, Viale Oxford, 00133 Rome, Italyc Neuroimaging Laboratory, Santa Lucia Foundation IRCCS, Via Ardeatina 306, 00179 Rome, Italyd Department of Psychology, University of Palermo, Palermo, Italye Department of Neuroscience, University of Rome Tor Vergata, Via Montpellier 1, 00133 Rome, Italyf Sobell Department of Motor Neuroscience, Queen Square, UCL, London, UK
⁎ Corresponding author. Fondazione S. Lucia IRCCS, VItaly. Fax: +39 6 51501388.
E-mail addresses: [email protected], giakoch@lib
1053-8119/$ – see front matter © 2010 Elsevier Inc. Adoi:10.1016/j.neuroimage.2010.02.022
a b s t r a c t
a r t i c l e i n f oArticle history:Received 19 November 2009Revised 7 February 2010Accepted 9 February 2010Available online 13 February 2010
Keywords:TMSConnectivityMovement planningSuperior longitudinal fasciculusTractographyTranscranial magnetic stimulation
We combined bifocal transcranial magnetic stimulation (TMS) and diffusion tensor imaging (DTI)tractography to investigate in humans the contribution of connections originating from different parietalareas in planning of different reaching to grasp movements. TMS experiments revealed that in the lefthemisphere functional connectivity between the primary motor cortex (M1) and a portion of the angulargyrus (AG) close to the caudal intraparietal sulcus was activated during early preparation of reaching andgrasping movements only when the movement was made with a whole hand grasp (WHG) towards objectsin contralateral space. In contrast, a different pathway, linking M1 with a part of the supramarginal gyrus(SMG) close to the anterior intraparietal sulcus, was sensitive only to the type of grasp required (precisiongrasping) but not to the position of the object in space. A triple coil experiment revealed that inactivation ofthe ventral premotor area (PMv) by continuous theta burst stimulation interfered with some of theseinteractions. Anatomical DTI tractography revealed that AG and SMG are strongly connected with PMv andwith M1 by different bundles of the superior longitudinal fasciculus (SLF).These results demonstrate the existence of segregated parieto-premotor-motor pathways crucial forpreparation of different grasping actions and indicate that these may process information relevant to boththe position of the object and the hand shape required to use it.
ia Ardeatina 306, 00179 Rome,
ero.it (G. Koch).
ll rights reserved.
© 2010 Elsevier Inc. All rights reserved.
Introduction
Although reaching and grasping a cup to drink a coffee seems to bealmost automatic, successful performance requires a complex inter-action of specific frontal and parietal brain networks to produce theoptimum combination of arm and hand movement to manipulate theobject most efficiently (Jeannerod et al., 1995). Classically, this type ofaction is supposed to involve cortical interconnected circuits includingthe parietal cortex, the premotor cortex and the primarymotor cortex(Rizzolatti and Luppino, 2001).
Experiments in monkeys have shown that the inferior parietallobule (IPL) contains a rich variety of neurons that discharge inassociationwith reaching and graspingmovements directed to specificgoals (Sakata et al., 1995;Murata et al., 2000; Fogassi et al., 2005; Rozziet al., 2008). Motor responses are somatotopically organized in arostro-caudal fashion. In regions of the anterior intraparietal area
(AIP), such as the PF and PFG, the most commonly represented motoracts are grasping with the mouth and hand grasping. In contrast, themost commonly represented act inmore caudal areas, such as area PG,is arm reaching, (e.g. Rozzi et al., 2008).
Human imaging experiments have tended to confirm this,revealing that more anterior regions of the IPL, located in thesupramarginal gyrus (SMG), seem crucially involved in precisegrasping movements, so that this area has already been proposed asthe homologue region of the AIP described in monkeys (hAIP)(Binkofski et al., 1998; Culham and Valyear, 2006; Olivier et al.,2007; Begliomini et al., 2007), while more posterior regions of the IPL,located in the angular gyrus (AG) close to the caudal intraparietalsulcus, have been associated with preparation of visually guidedreaching movements (Jeannerod et al., 1995; Castiello, 2005; Karnathand Perenin, 2005).
It has been postulated that the different subregions of the IPLsupply PMv with visual information about the attributes of objects(e.g., size, orientation) that are needed to select appropriate graspconfigurations (Murata et al., 1997; Rizzolatti and Luppino, 2001;Raos et al., 2006). Such information would eventually converge onto

301G. Koch et al. / NeuroImage 51 (2010) 300–312
M1 for execution of the final motor plan (Rizzolatti and Luppino,2001; Shimazu et al., 2004; Prabhu et al., 2009).
Most of the previous studies focused on the role played by specificparietal regions in reaching and grasping. Nevertheless, importantand complementary information can be obtained by studying thephysiology of the interactions occurring between the brain regionsthat form part of this cortical network (Rizzolatti and Luppino, 2001;Koch and Rothwell, 2009a,b).
We recently developed (Koch et al., 2007) a new method forinvestigating functional connections between the posterior parietalcortex and ipsilateral motor cortex noninvasively in humans, using atwin-coil or bifocal transcranial magnetic stimulation (TMS) para-digm. In this method, a conditioning TMS pulse is applied over PPCshortly before a test pulse over the hand area of primary motor cortex(M1). At appropriate interstimulus intervals, the motor evokedresponse evoked by theM1 pulse is enhanced, indicating the presenceof functional interactions between the two sites. Using this approach,we have previously explored interactions between theAG andprimarymotor cortex (M1) during task in which subjects had to reach andpoint to two different targets. The results showed that the excitabilityof this connection changed during the reaction time of the task. Whenthe reaction signal indicated that subjects had to reach and point to atarget positioned in the contralateral (and not in the ipsilateral)hemispace, therewas a pronounced increase in excitability 50ms afterthe onset of the reaction signal. Therefore, we concluded that func-tional connectivity between human parietal and motor cortex isenhanced during early stages of planning a reach in the contralateraldirection (Koch et al., 2008a). It appeared that information about thelocation of the target was being sent to M1 very early in the reactionperiod of the task. In classical terms this could therefore be viewed ascontributing to the reaching component of the task.
The previous task involved subjects reaching and pointing to thetarget with their index finger. Here we used the same methods toinvestigate the contribution of different subregions of the IPL, such asthe more posterior AG and the more anterior SMG, in planning ofreaching and grasping actions. We studied whether the interactionbetween these regions of the IPL in the left hemisphere and theipsilateral M1 would change when subjects were planning to reachand grasp a single object in two different ways: a precision grip (PG)and whole hand grasp (WHG). We decided to test the left hemispheresince these movements in everyday life are performed with thedominant right hand. We also varied the position of the object inspace in order to test for specific reaching spatial components of thetask (Koch et al., 2008a).
We hypothesised that the AG-M1 connection would be involved inprogramming the upcoming grasp when the target object has to bereached by a lateral movement (Jeannerod et al., 1994; Castiello,2005; Karnath and Perenin, 2005; Koch et al., 2008a). On the otherhand we predicted that the SMG-M1 connection would be involved inspecifying specific parameters for grasp (i.e., WHG vs. PG) rather thanreach, given that the SMG can be considered part of the hAIP (Sakataet al., 1995; Murata et al., 2000; Fogassi et al., 2005; Rozzi et al., 2008;Binkofski et al., 1998; Culham and Valyear, 2006; Olivier et al., 2007;Begliomini et al., 2008).
Although this approach can provide reliablemeasures of the causalfunctional interactions ongoing between the two areas (Koch andRothwell, 2009b) it does not provide any information about the aboutthe possible anatomical pathways that might mediate the responses.Because of this, we used diffusion tensor magnetic resonance imaging(DTI) to obtain some insight into the white matter pathways thatconnect these areas in humans. DTI can reconstruct in vivo whitematter fibre bundles based on the assumption that the principaldirection of tissue water diffusion is parallel to the main fibredirection in every voxel. Other authors have found an associationbetween DTI values in certain white matter tracts and measures offunctional connectivity as measured by tcTMS. They suggest that this
increases the confidence that the physiological interaction observedwas produced by activity in that specific tract (Wahl et al., 2007;Boorman et al., 2007; Kloppel et al., 2008; Mars et al., 2009).
Materials and methods
Task-dependent parietal-motor connectivity assessed with TMS
Experiment 1
Grasping task. We used an auditory choice RT task similar to thatpreviously described by Koch et al. (2008a) but adapted to emphasizereaching to grasp movements according to our experimentalhypotheses about PPC–M1 influences.
The task was to reach, grasp and lift with the right hand a cuplocated on a central or right lateral position (targets visiblethroughout the experiment), as signaled by an auditory tone,indicating symbolically how the subject should have grasped onthat particular trial (Fig. 1A). Depending on the tone, subjects wererequired either to pinch the handle of the cup (precision grip-PG) orto grasp the whole cup from the top (whole hand grasp-WHG) (seeFig. 1B). We examined the excitability of the AG-M1 and SMG-M1interactions in the reaction interval between the onset of the auditorysignal and the start of the grasping movement. Since no arm or handmovement occurs in this period the TMS responses are notcontaminated by ongoing EMG activity in the hand muscles.
Right handed subjects (n=10; F/M=5/5, median age [range]=28 [22–34] years; mean laterality index: 74.5%; Oldfield, 1971) sat ona 45-cm-high straight-back chair facing a table, 120 cm wide×90 cmdeep, placed at a height of 70 cm. A headrest ensured a constant headposition throughout the experiment. During target presentation, theroom was illuminated with direct lighting from the ceiling. Subjectsplaced their right hand on an upraised bump (2.5-cm-diameter coin)on the table surface that could have been located by touch (startbutton). This point was in line with the subject's sagittal midline. Inthe central position, cups were positioned on a plastic panel in front ofthe start button at a reaching distance of 20 cm and a viewing distanceof 60 cm; in the right lateral position, targets were positioned 45°ahead and laterally, at a similar reaching distance. Reaction times andmovement timeswere recorded through custommade copper sensorspositioned over the start button, the cup and over the panel in whichcups were located.
Subjects were required to reach, grasp and lift to a height ofapproximately 5 cm the peripheral central or lateral right target assoon as they heard a cue sound. Each trial began with an auditorywarning (500 Hz, 40 ms).
Premovement EMG activity was measured and controlled throughelectrodes placed over the deltoid muscle. The imperative auditorysignal was given randomly 1–3 s later and consisted of either a high(800 Hz, 30 ms) or low (200 Hz, 30 ms) frequency tone pulse thatindicated how to reach and grasp the peripheral target, according tocounterbalanced instructions given to the subjects (high meaningprecision grip, low meaning whole hand grasp, or vice versa, fullycounterbalanced as described below). The intertrial interval was 6 s.After the warning sound, subjects had to fixate a fixation crosspositioned in the middle of the panel on which the cups were located.They were then instructed to direct their gaze towards the peripheralor central target as soon they heard the cue. Thus the target waslocated in the central visual field in both conditions. Eye position wasmeasured by DC electrooculogram (EOG) recorded by means of Ag/AgCl surface electrodes for horizontal and vertical EOG. The signal wasamplified and band pass filtered (5–2000 Hz) using SIGNAL software.EOG was used to assess initial saccadic latencies.
Before each test session, 10 practice trials were given until subjectsreported at least five continuous RTs within 150% of individual meanRT. For each site of PPC stimulation (AG and SMG), in every other

Fig. 1. (A) Schematic representation of bifocal TMS coils placement and recordings during the planning phase of the reaching to grasp task. In different experiments a conditioningstimulus was applied over subregions of the PPC (AG and SMG), preceding by 4ms in half of the trails the test stimulus delivered over M1. Motor evoked potentials were recorded bythe first dorsal interosseus (FDI) and the abductor digiti minimi (ADM) muscles. Depending on an acoustic cue, subjects were required to perform either a precision grip or a wholehand grasp (B) of a cup positioned in different blocks centrally or laterally on the side opposite of the stimulated hemisphere. During subsequent movement, grasping the handle ofthe cup (PG) involved significantly greater EMG activity in FDI, the muscle that abducts and flexes the index finger, compared with grasping the top of the cup (WHG).
302 G. Koch et al. / NeuroImage 51 (2010) 300–312
block the target was located in the central position, thus alternatingwith blocks where it was located in the right lateral position.
The order of different experimental blocks was counterbalancedacross subjects. Onset of hand movement (RT) was detected byrelease of a pushbutton. The reaching movement time (MT) wascalculated as the delay between the release of the start button and thetime as subjects touched the cup. Lift time (LT) was then calculated asthe delay between the release of the cup from the surface of the tableand the time it was dropped.
Transcranial magnetic stimulation. Electromyographic (EMG) traceswere recorded from the right first dorsal interosseous (FDI) andabductor digiti minimi (ADM) muscles of the right hand using 9-mmdiameter, Ag–AgCl surface cup electrodes. Although the transportcomponent of a reach movement depends on the activity of musclesacting at the shoulder joint, we choose to test only the hand musclessince thesemuscles were selectively activated in our reaching to grasptask and MEPs can be recorded more reliably from this muscle incomparison with proximal shoulder muscles. For FDI muscle, theactive electrode was placed over the muscle belly and the referenceelectrode over the metacarpophalangeal joint of the index finger. Forthe ADM muscle, the active electrode was placed over the musclebelly and the reference electrode over the metacarpophalangeal jointof the little finger. Responses were amplified with a Digitimer D360amplifier (Digitimer Ltd, Welwyn Garden City, Hertfordshire, UK)through filters set at 20 Hz and 2 kHz with a sampling rate of 5 kHz,then recorded by a computer using SIGNAL software (CambridgeElectronic Devices, Cambridge, UK).
We used a paired pulse stimulation technique with two high-power Magstim 200 machines (Magstim Co., Whitland, Dyfed, UK).The magnetic stimulus had a nearly monophasic pulse configura-tion with a rise time of about 100 μs, decaying back to zero overabout 0.8 ms.
First the intensity of TMS was adjusted to evoke an MEP ofapproximately 1 mV peak-to-peak in the relaxed right FDI and ADM.The hand motor area of left M1 was defined as the point wherestimulation evoked the largest MEP from both muscles. The teststimulator was connected to a small custom-made figure-of-eight-shaped coil (external diameter 50 mm). The coil over M1 was alwaysplaced tangentially to the scalp at 45 angle frommidline of the centralsulcus, inducing a posterior–anterior (PA) current flow. The condi-tioning stimulator was connected to another small custom-madefigure-of-eight-shaped coil (external diameter 50 mm).
We used a neuronavigation system (Softaxic, E.M.S., Bologna, Italy)to precisely position the coil over the stimulation sites, usingindividual anatomical magnetic resonance images; this techniquehas been described in detail previously (Koch et al., 2007). Theindividual coordinates of each stimulation site were normalized aposteriori into the Montreal Neurological Institute (MNI) coordinatesystem and averaged. To target AG, the coil was positioned in theposterior portion of the inferior parietal lobule (IPL), close to aposterior part of the adjoining intraparietal sulcus (cIPS) (Koch et al.,2007). The centre of the coil was positioned tangentially to the skullwith the handle pointing downward and slightly posteriorly (10) (Fig.1A). To target SMG, the coil was positioned in the anterior portion ofIPL, close to the intersection between the intraparietal sulcus andpostcentral sulcus. The centre of the coil was positioned tangentially tothe skull with the handle pointing downward and posteriorly (10)(Fig. 1A).
TMS was delivered over M1 of the left hemisphere at differentdelays (25, 50, 75, 100, 125, 150 ms) after the cue sound. On the basisof our recent published paired pulse TMS work with PPC and M1 sites(Koch et al., 2007) we used the following parameters to best activatePPC–M1 interplay. The intensity of the M1 pulse was adjusted toevoke an MEP of approximately 1 mV peak-to-peak in the relaxedright FDI. In half of the trials, M1 TMS was preceded by a conditioning

303G. Koch et al. / NeuroImage 51 (2010) 300–312
pulse delivered 4 ms earlier, over the ipsilateral AG or SMG at anintensity of 90% of the resting motor threshold (RMT). We definedRMT as the lowest intensity that evoked five small responses (about50 μV) in the contralateral FDI muscle in a series of ten pulses whenthe subject kept the FDI muscles relaxed in both hands.
In each block, 10 trials were performed for each subcondition inthe main experiment (PG or WHG movement planned, as instructedby the symbolic auditory imperative stimulus, fully crossed with M1pulse alone or paired PPC–M1 pulses), at each of the six delays. Intotal, four blocks of 240 trials were performed for every subject. Themean peak-to-peak amplitude of the conditioned MEP was expressedas a percentage of the mean peak-to-peak amplitude size of theunconditioned test pulse.
Experiment 2In this control experiment (n=6) we applied both pulses over M1
to verify whether the effects we observed in experiment 1 could beascribed to changes in the excitability of M1 or to spread of magneticpulses over M1 from the parietal regions. Both pulses were appliedover left M1. A subthreshold CS at 90% RMT was applied 12 ms beforethe test stimulus (TS). The intensity of the TS was adjusted to evoke anMEP of approximately 1mV peak-to-peak in the relaxed right FDI. Thisprotocol is known to activate intracortical facilitatory circuits (ICF)(Kujirai et al., 1993). TMSwas delivered overM1of the left hemisphereat delays of 50, 75, 100 and 125ms after the cue sound.We studied thecrucial delays of 50, 75, 100 and 125 ms in which significantmodulation of the AG and SMG activity was found in experiment 1.Ten trials were performed for each subcondition. The mean peak-to-peak amplitude of the conditionedMEPwas expressed as a percentageof the mean peak-to-peak amplitude size of the unconditioned testpulse.
Experiment 3Previous animal data as well as the data that arose from the DTI
investigations in the present experiments suggest that both AG andSMG have strong connections with PMv, where neurons with similarfunctions have also been identified (Petrides and Pandya, 1984;Luppino et al., 1999; Rozzi et al., 2006; Brochier and Umilta, 2007;Borra et al., 2008).
Since PMv has powerful connections with M1 (Shimazu et al.,2004; Prabhu et al., 2009; Davare et al., 2008), we therefore askedwhether the functional effects that we had observed in the twin coilTMS experiments would be affected by a temporary interference withthe function of PMv produced by a short period of repetitive TMS.
In six subjects, we applied the continuous theta burst stimulation(cTBS) protocol (Huang et al., 2005) over left PMv in order to induce along lasting inhibition of the stimulated area. A MagStim Super Rapidmagnetic stimulator (Magstim Company, Whitland, Wales, UK),connected with a figure-of-eight coil with a diameter of 70 mm wasused to deliver cTBS. Three-pulse bursts at 50 Hz repeated every200 ms for 40 s were delivered at 80% of the active motor threshold(AMT) over the left PMv (600 pulses) (Huang et al., 2005). In thisexperiment we used the neuronavigation system (Softaxic, E.M.S.,Bologna, Italy) to position the coil precisely over the stimulation siteusing individual anatomical magnetic resonance images. The centre ofthe coil was held tangentially to the skull with the handle pointingdownward and posteriorly (10°). The individual coordinates of eachstimulation site were normalized a posteriori into the MontrealNeurological Institute (MNI) coordinate system and averaged. Sub-jects performed the grasping task before and immediately after cTBS.We tested the interactions betweenAGand SMGwithM1at the crucialtimepoints of the planning phase previously observed in experiment 1.Therefore TMSwas delivered over M1 of the left hemisphere at delaysof 50 and 75ms after the cue sound. In one block, a subthreshold CS at90% RMT was applied 4 ms before the test stimulus (TS) over the leftAGwhen subjects were performing the taskwith the target situated in
the right lateral position. In another block, CS at 90% RMT was appliedover the left SMGwhen subjects were performing the task with targetlocated in the central position. The intensity of the TS was adjusted toevoke anMEPof approximately 1mVpeak-to-peak in the relaxed rightFDI. In each block, ten trialswere performed for each subcondition. Themean peak-to-peak amplitude of the conditioned MEP was expressedas a percentage of the mean peak-to-peak amplitude size of theunconditioned test pulse.
Experiment 4In this control experiment (n=6) we aimed to verify whether the
results observed in experiment 1 are specific to the graspingcomponent of the movement, since they may be related to differencesin the reach component in the two conditions. In fact in experiment 1,PG and WHG tasks differ not only for the hand shape but also in theorientation and position in space of the hand and arm. This was due tothe fact that the targets were approached with slightly differentreachingmovements,with the handbeingmore pronated for theWHGin comparison with PG. Therefore we performed another experimentin which the reach was held constant and only the grasp was altered.We instructed the subjects to either perform a PG or a WHG using thesamehand orientation and position in space. In this case, theWHGwasperformed not by grasping cup from the top, with the arm pronated by90°, but grasping the cup from the lateral surface, with the armadopting the same posture as for PG. We tested the interactionsbetween SMG and M1 at the crucial time points of the planning phasepreviously observed in experiment 1. Therefore TMS was deliveredover M1 of the left hemisphere at delays of 50 and 75 ms after the cuesound. CS at 90% RMT was applied over the left SMG when subjectswere performing the task with target located in the central position.The intensity of the TSwas adjusted to evoke anMEP of approximately1 mV peak-to-peak in the relaxed right FDI. In each block, 10 trialswere performed for each subcondition. The mean peak-to-peakamplitude of the conditioned MEP was expressed as a percentage ofthemean peak-to-peak amplitude size of the unconditioned test pulse.
MRI tractography
The same subjects that participated in experiment 1 had MRI withDTI acquisitions. All imaging was obtained using a head-only 3.0 Tscanner (Siemens Magnetom Allegra, Siemens Medical Solutions,Erlangen, Germany), equipped with a circularly polarized transmit–receive coil. The maximum gradient strength is 40 mT m−1, with amaximum slew rate of 400 mT m−1 μs−1.
The MRI session included, for every subject, a dual-echo turbo spinecho sequence (TEs=12/112 ms; TR=4500 ms; ETL=7, ma-trix=256×192; FOV=230×172.5 mm2; slice thickness=5 mm;total number of slices=24), to exclude the presence of any brainabnormality, and a pulsed-gradient twice refocused spin-echo EPI(TE/TR=90/8500 ms, bmax=1000 s mm−2, voxel size 2.3 mm3)with diffusion gradients applied in 81 noncollinear directions. Nineimages with no diffusion weighting (b0) were also acquired. Inaddition, a magnetization prepared rapid gradient echo (MPrage)sequence (TE=2.74 ms; TR=2500 ms; TI=900; flip angle=8°;matrix=256×208×176; FOV=256×208×176 mm3) was collectedfor every subject.
Image analysis
The analysis was performed off-line using tools from SPM8 (www.fil.ion.ucl.ac.uk/spm/), FSL (www.fmrib.ox.ac.uk/fsl/), and CAMINO(www.camino.org.uk).
PreprocessingDTI data were corrected for eddy current-induced distortions and
small head motions using affine registration (see eddycorrect, FMRIB

304 G. Koch et al. / NeuroImage 51 (2010) 300–312
Diffusion Toolbox (FDT) http://www.fmrib.ox.ac.uk/fsl/fdt/index.html). Next the diffusion tensor was estimated voxelwise (Basser etal., 1994) using CAMINO (www.camino.org.uk), and FA maps weregenerated. For each subject, the FA map was affine-registered to theMPrage, using FLIRT (Jenkinson et al., 2002), and the transformationmatrix was recorded. Each subject's MPrage was registered to the FSLT1-weighted template in MNI coordinates, again using FLIRT. Thetransformation matching the DTI data to MNI coordinates wasobtained by concatenating these two transformations.
Seed-point ROIs. We used probabilistic tractography (Parker et al.,2003) to investigate the possible pathways connecting the IPLwithM1and PMv. Tractography requires the definition of “seed-points” and“way-points”, i.e., the regions of interest (ROIs) from where the tractsare initiated, or through where they are constrained. For this study,four ROI masks (respectively encompassing SMG, AG, M1, and PMv)were created using as a reference each subject's MPrage transformedto MNI space. For SMG, AG and M1, three spherical ROIs, with a 4 mmradius and centred at the MNI coordinates corresponding to the site ofthe TMS stimulation, were obtained. Each subject's MPrage imageswere segmented into white and grey matter using SPM8 (Ashburnerand Friston, 2005). The position of each sphere was carefully checked,nonparenchymal voxels (those falling in the corticospinal fluid, oroutside the brain) were removed, and the location of the seed-pointswas compared with the result of tissue segmentation, to ensure theywere located at the boundary between grey and white matter.
The ROI masks were then transformed back into DTI native space,where tractographywas performed. Because of the process of “cleaningup” of nonparenchymal voxels described above, there was a slightvariability in ROI size: the median (across subjects) size (in voxels) inDTI space was: 17 [range 13–19] voxels for aIPS, 17 [range 15–19] forcIPS, 16 [range 12–19] for M1, and 17 [range 12–19] for PMv.
Q-ball and tractography. DTI can only resolve one fibre per voxel,while the white matter tracts object of this study are known to have acomplex architecture. In order to account for the presence of multiplefibre directions in a single voxel, we used Q-ball imaging (Tuch, 2004),implemented in Camino. Q-ball is a high angular resolution diffusionimaging method able to reconstruct the intra-voxel fibre orientationdistribution functions (ODFs), based on the Funk-Radon transform. Inregions containing multiple fibre directions, the ODF exhibits multiplepeaks. Given the relatively lowb-valueused in this study,we limited themaximum number of direction per voxel to 3. We used Tuch's originalradial-basis function formulation (Tuch, 2004), as implemented inCamino with the default parameter settings.
The tractography algorithm used for this studywas the probabiliticindex of connectivity (PICo) (Parker et al., 2003). PICo considersmultiple pathways emanating from a seed point and from each pointalong the reconstructed trajectories. The uncertainty due to thepresence of noise and associated with the determination of theprincipal direction of diffusion is accounted for by generating aprobability distribution function in every voxel, which is obtainedfrom the Q-ball reconstruction. This probability density functionprovides voxel-wise estimates of the confidence in fibre tractalignment. Streamline-based tracking is then performed and repeatedin aMonte Carlo fashion to produce amap that reflects, for every voxelin the brain, the probability of connection (ranging between 0 and 1)to the seed point. The analysis was restricted to the left hemisphere.As the neurophysiological interactions between the IPL and M1 couldresult either from the mediation of polysynaptic circuits involving thepremotor cortex (as already suggested by animal studies; Shimazu etal., 2004; Prabhu et al., 2009) or in alternative from direct projectionsto M1, we hypothesized two possible pathways connecting the intra-parietal sulcus and the motor cortex: either (a) the existence of directconnections between M1 and both SMG and AG , or (b) the existenceof a circuit which connects the IPL to PMv, and PMv with M1.
In order to assess which of these two hypotheses was more likelywe compared the reconstruction of the following putative tracts:
(a)the putative direct connection between SMG andM1, and betweenAG and M1, initiating the streamlines from SMG, and AG,respectively, but retaining only those passing through M1.
(b)the connection between SMG and PMv, the connection betweenAG and PMv, and the connection between PMv and M1. For theSMG-PMv (and AG-PMv) connection, the SMG (or AG) mask wasused as seed-point, but retaining only streamlines which passedthrough the PMv ROI. For the PMv-M1 connection, the PMv ROIwas used as seed-point, accepting only streamlines passingthrough M1. For all tracts, PICo was run with 5000 Monte Carloiterations. The resulting maps were thresholded at 0.1 (10%), thusrejecting voxels with a low probability (b0.1) of connection to theseed points, in order to remove noise.
Evaluation of tractography output. In order to estimate andcompare the likelihood of the hypothesised anatomical connectionsand their relationship with functional connectivity, we used thefollowing criteria: first, we carefully inspected the results obtainedfor each subject, and we counted the number of individuals where acontinuous tract was present for each of the given connections(Aron et al., 2007). Secondly, for a given seed point (either the AGor the SMG), we compared its probability of connection to twoalternative target ROIs (either M1 or PMv). This was achieved byrunning PICo from the seed ROI, and permitting only pathwaysconnecting to either target. Among these streamlines, the fractionthat reached a given target was counted. The streamline count wasnormalised by the total number of streamlines reaching eithertarget (Tomassini et al., 2007), thus giving a measure of theprobability of connecting to a specific target. For example, for agiven voxel in the seed point mask, if N streamlines connect toPMv, and M streamlines connect to M1, the probability ofconnection of the seed with PMv is given by N / (M+N), whilethe probability of connection with M1 is given by M / (M+N). Inorder to obtain a single value per subject, the percentages wereaveraged across those obtained from all the voxels in the seedmask. These fractions were expressed as percentages. We refer tothis quantity as “strength of connectivity” (SoC).
Investigation of the relationship between anatomy and function.According to the hypothesis that specific anatomo-functional path-ways connecting the IPL and M1 exist, we expect to observe somesort of association between neurophysiological measures of func-tional connectivity and measures of white matter structure such asSoC.
We then quantified the correlation between this quantity for eachreconstructed tract and the percentage increase in MEP recordingafter a conditioning TMS pulse applied over the SMG or AG.
We correlated SoC of different connections with changes in MEPsamplitude for the FDImuscles and ADMmuscles. Given the prominentphysiological role of the FDI in tasks examined here, we expected tofind more robust correlation only for this muscle.
Reconstruction of the cortico-spinal tract
In order to complete the reconstruction of the circuit involved inplanning and executing a hand motor task, we also reconstructed theleft cortico-spinal tract (CST) in every subject's brain. The seed pointwas manually positioned in the cerebral peduncles, according topublished guidelines (Wakana et al., 2007). This ROI (mediansize=17.5 voxels, range=12–19) was drawn directly on the colour-coded direction map obtained from DTI data (Pajevic and Pierpaoli,1999) to ensure the selection of voxels with primarily superior-
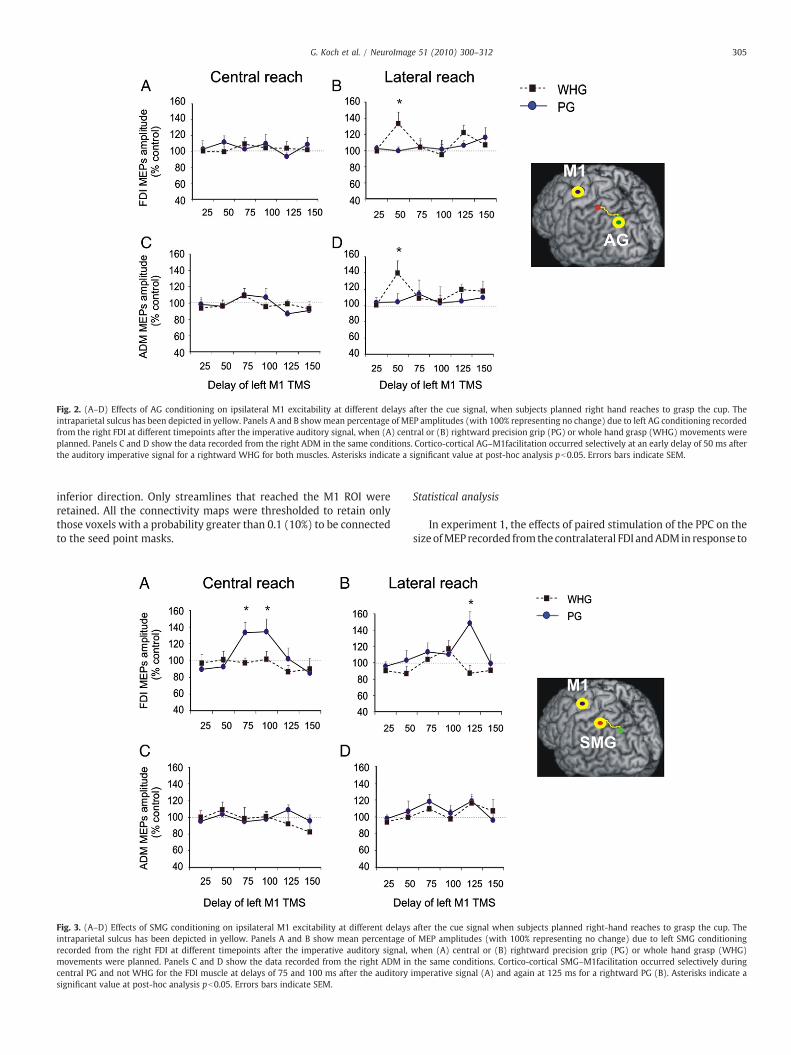
Fig. 2. (A–D) Effects of AG conditioning on ipsilateral M1 excitability at different delays after the cue signal, when subjects planned right hand reaches to grasp the cup. Theintraparietal sulcus has been depicted in yellow. Panels A and B showmean percentage of MEP amplitudes (with 100% representing no change) due to left AG conditioning recordedfrom the right FDI at different timepoints after the imperative auditory signal, when (A) central or (B) rightward precision grip (PG) or whole hand grasp (WHG) movements wereplanned. Panels C and D show the data recorded from the right ADM in the same conditions. Cortico-cortical AG–M1facilitation occurred selectively at an early delay of 50 ms afterthe auditory imperative signal for a rightward WHG for both muscles. Asterisks indicate a significant value at post-hoc analysis pb0.05. Errors bars indicate SEM.
305G. Koch et al. / NeuroImage 51 (2010) 300–312
inferior direction. Only streamlines that reached the M1 ROI wereretained. All the connectivity maps were thresholded to retain onlythose voxels with a probability greater than 0.1 (10%) to be connectedto the seed point masks.
Fig. 3. (A–D) Effects of SMG conditioning on ipsilateral M1 excitability at different delaysintraparietal sulcus has been depicted in yellow. Panels A and B show mean percentage orecorded from the right FDI at different timepoints after the imperative auditory signal,movements were planned. Panels C and D show the data recorded from the right ADM incentral PG and not WHG for the FDI muscle at delays of 75 and 100 ms after the auditorysignificant value at post-hoc analysis pb0.05. Errors bars indicate SEM.
Statistical analysis
In experiment 1, the effects of paired stimulation of the PPC on thesizeofMEP recorded from the contralateral FDI andADMin response to
after the cue signal when subjects planned right-hand reaches to grasp the cup. Thef MEP amplitudes (with 100% representing no change) due to left SMG conditioningwhen (A) central or (B) rightward precision grip (PG) or whole hand grasp (WHG)the same conditions. Cortico-cortical SMG–M1facilitation occurred selectively duringimperative signal (A) and again at 125 ms for a rightward PG (B). Asterisks indicate a

Fig. 4. In a control experiment when both pulses were applied over left M1 to test intracortical facilitatory circuits (ICF), no significant differences were observed during PG andWHGplanning for both central and lateral reaching movements. MEPs were recorded only from the right FDI. Errors bars indicate SEM.
Fig. 5. This figures illustrates the effects induced by cTBS applied on the left PMv on thesignificant interactions observed in experiment 1 for SMG stimulation during central PG(A, B) and for AG stimulation during lateral WHG (C, D). Asterisks indicate a significantvalue at post-hoc analysis pb0.05. Errors bars indicate SEM.
306 G. Koch et al. / NeuroImage 51 (2010) 300–312
M1 TMS were analyzed as the percentage of the mean peak-to-peakamplitude of the unconditioned test M1 pulse. For each site ofstimulation (SMG and AG), mean percentage values were analyzedwith a four-way repeated measures analyses of variance (ANOVA)with DIRECTION (central vs. rightward reaching), GRASP (PG vs.WHG),MUSCLE (FDI vs. ADM), and TIME (25, 50, 75, 100, 125, 150ms)as within-subject main factors. Repeated measures ANOVAs were alsoperformed on the mean RTs, MTs and LTs of all subjects in eachcondition for each site of stimulation with main factors of TIME, TMS(single vs. paired) or Type of hand movement (PG vs. WHG). A pvalueb0.05was considered as statistically significant. In experiment 2,when both pulses were applied overM1mean percentage values wereanalyzed with a three-way ANOVA with DIRECTION (central vs.rightward reaching), GRASP (PG vs. WHG), TIME (50, 75, 100,125 ms) as within-subject main factors. In experiment 3 separateANOVAs with TBS (pre vs. post) and TIME (50 vs. 75 ms) as withinsubjectsmain factorswere performed for each condition (central PG orlateral WHG) in which a significant change was observed inexperiment 1. In experiment 4 ANOVA was performed with GRASP(PG vs. WHG) and TIME (50 vs. 75 ms) as within-subject main factors.A significant main effect in the ANOVA was followed by Tukey's post-hoc analysis. The Greenhouse–Geisser correction was used fornonspherical data. The association between TMS data and SoC valueswas investigated using Pearson's correlation coefficient. A pvalueb0.05 was considered significant.
Results
Task dependent parietal-motor connectivity assessed with TMS
Experiment 1Mean normalized MNI coordinates of AG site were−38.4±7.1,−
72.2±4.8, and 50.0±5.5 mm (x, y, z, mean±SD). Mean normalizedMNI coordinates of SMG stimulation site were − 49.7±4.9, −41.8±8.3 and 56±4.2 mm (x, y, z, mean±SD) (Table S1). Measurements ofthe rectified EMG from each muscle integrated for the periodpreceding contact with the object, showed that grasping the handleof the cup (PG) involved significantly greater FDI EMG activity incomparison to that recorded during whole hand grip (WHG) and viceversa for ADM (all pb0.05 at t-test analysis) (Fig. 1C).
During the early planning phase of the task that we explored, theexcitability of the left motor cortex alone (i.e., unconditioned by leftAG or SMG stimulation) did not vary in any condition as shown byANOVA performed for each muscle on MEPs amplitude withDIRECTION, GRASP and TIME as within subjects main factors.
Figs. 2A–D plots the percentage change in MEPs caused by theconditioning stimulus to AG at intervals ranging from 25 to 150 msafter onset of the imperative auditory signal to perform a PG or aWHG. Therewas no effect of AG onMEPs evoked by the test pulse overM1 if the cup was placed in the midline whether imperative signalinstructed subjects to perform a PG or aWHG. However, when the cup
was placed to the right of the midline the excitability of theconnection to both the FDI and ADM areas of M1 increased, but onlywhen the imperative signal instructed subjects to perform a WHG butnot PG (as revealed by ANOVA showing significant effects of GRASP(F(1,9)=4.43; p=0.04) and DIRECTION (F(1,9)=4.11; p=0.04)main factors, and a significant DIRECTION×GRASP×TIME interaction(F(5,45)=3,46; p=0.03). Post-hoc analysis showed that the AG-M1grasp-specific facilitation arose selectively at an early delay of 50 msafter the auditory signal (WHG vs. PG; p=0.02 for both muscles; seeFig. 2B, D). In summary, the reach-specific effect (i.e., midline versusright) of the AG stimulus was modulated by the grasp that subjectswere about to perform.
A different pattern was found for the interactions of SMGwith M1.In this case, there was a grasp specific effect on the excitability of theinteraction, being present for PG but not WHG, but in this case it wasinsensitive to the direction of the reaching movement. This wasconfirmed in the ANOVA showing significant GRASP (F(1, 9)=6.51;p=0.03) and TIME (F(1,9)=3.46; p=0.04) main factors and a signi-ficant DIRECTION×MUSCLE×GRASP×TIME interaction (F(5,45)=4.82; p=0.002). Post-hoc analysis showed that for the midlinereach there was a grasp-specific SMG facilitation of the M1 projectionto FDI at delays of 75 and100ms after the auditory signal (WHGvs. PG;
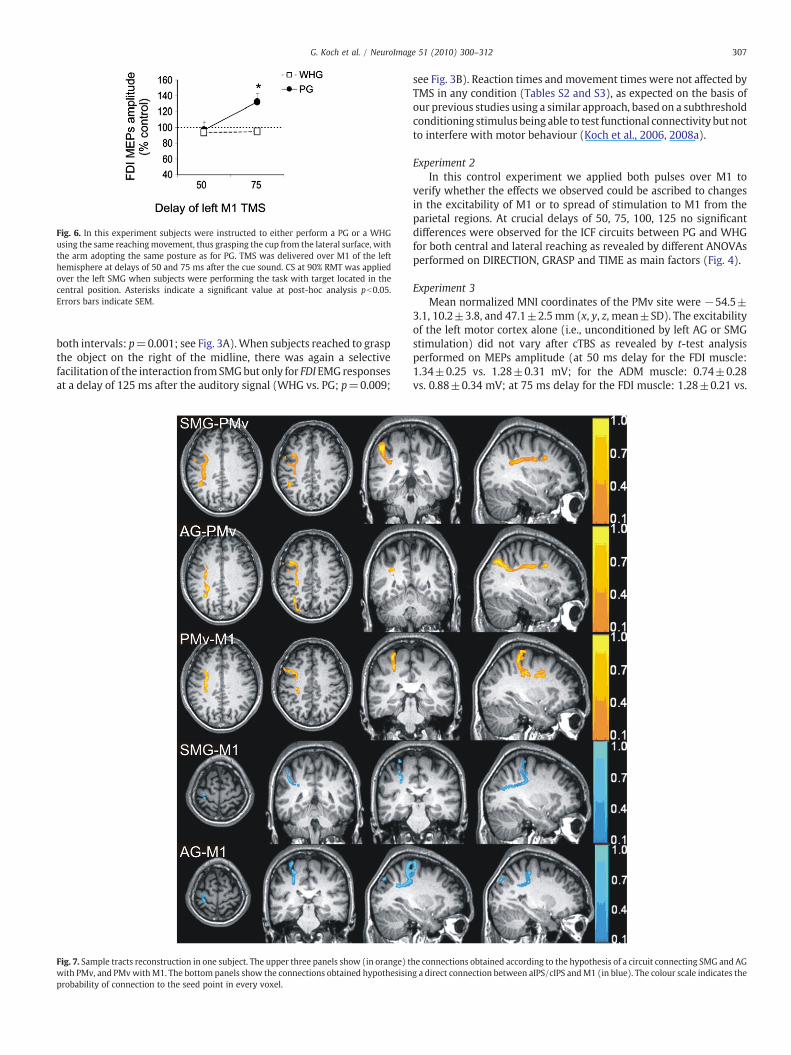
Fig. 6. In this experiment subjects were instructed to either perform a PG or a WHGusing the same reachingmovement, thus grasping the cup from the lateral surface, withthe arm adopting the same posture as for PG. TMS was delivered over M1 of the lefthemisphere at delays of 50 and 75 ms after the cue sound. CS at 90% RMT was appliedover the left SMG when subjects were performing the task with target located in thecentral position. Asterisks indicate a significant value at post-hoc analysis pb0.05.Errors bars indicate SEM.
307G. Koch et al. / NeuroImage 51 (2010) 300–312
both intervals: p=0.001; see Fig. 3A).When subjects reached to graspthe object on the right of the midline, there was again a selectivefacilitation of the interaction fromSMGbut only for FDI EMG responsesat a delay of 125 ms after the auditory signal (WHG vs. PG; p=0.009;
Fig. 7. Sample tracts reconstruction in one subject. The upper three panels show (in orange) twith PMv, and PMvwithM1. The bottom panels show the connections obtained hypothesisinprobability of connection to the seed point in every voxel.
see Fig. 3B). Reaction times and movement times were not affected byTMS in any condition (Tables S2 and S3), as expected on the basis ofour previous studies using a similar approach, based on a subthresholdconditioning stimulus being able to test functional connectivity but notto interfere with motor behaviour (Koch et al., 2006, 2008a).
Experiment 2In this control experiment we applied both pulses over M1 to
verify whether the effects we observed could be ascribed to changesin the excitability of M1 or to spread of stimulation to M1 from theparietal regions. At crucial delays of 50, 75, 100, 125 no significantdifferences were observed for the ICF circuits between PG and WHGfor both central and lateral reaching as revealed by different ANOVAsperformed on DIRECTION, GRASP and TIME as main factors (Fig. 4).
Experiment 3Mean normalized MNI coordinates of the PMv site were −54.5±
3.1, 10.2±3.8, and 47.1±2.5 mm (x, y, z, mean±SD). The excitabilityof the left motor cortex alone (i.e., unconditioned by left AG or SMGstimulation) did not vary after cTBS as revealed by t-test analysisperformed on MEPs amplitude (at 50 ms delay for the FDI muscle:1.34±0.25 vs. 1.28±0.31 mV; for the ADM muscle: 0.74±0.28vs. 0.88±0.34 mV; at 75 ms delay for the FDI muscle: 1.28±0.21 vs.
he connections obtained according to the hypothesis of a circuit connecting SMG and AGg a direct connection between aIPS/cIPS andM1 (in blue). The colour scale indicates the

Fig. 8. Intersubject variability. The three main pathways connecting the IPS, PMv and M1 are shown in the figure, by means of the overlap between subjects. The SMG–PMvconnection (top panel) the AG–PMv (middle panel) and the PMv–M1 connection were successfully reconstructed in all subjects and show a relatively reproducible shape acrossindividuals.
Table 1Mean strength of connectivity (across subjects).
Target seed PMv M1
SMG 62.4% 21.4%AG 26.0% 23.1%
The values indicate percentage strength of connectivity between either seed and thetargets, averaged across subjects. For each voxel in the seed region, these values wereobtained as the proportion of streamlines connecting to either target. Next, for everysubject, the results were averaged across all voxels in the seed. As there are voxelswhich do not connect to either target, please note that the percentages do not alwaysadd up to 100%.
308 G. Koch et al. / NeuroImage 51 (2010) 300–312
1.21±0.40 mV; for the ADM muscle: 0.83±0.38 vs. 0.81±0.31 mV;all p=n.s.). We found that the pronounced increase in the excitabilityof both the PPC–M1 connections were abolished following PMv cTBS.Thus, the interactions between AG and M1 during lateral WHG(ANOVA showing significant TBS main factor (F=7,47;p=0.03) andof TBS×TIME interaction (F=5,37; p=0.04) which were previouslymaximal at 50 ms were reduced following TBS (p=0.03). Similarlythe interactions between SMG and M1 during central PG changedfollowing PMv TBS, as revealed by ANOVA (showing a significant ISImain factor (F=8,41; p=0.03) and of TBS×TIME interaction(F=9,61; p=0.02). Post-hoc analysis showed that at 75 ms therewas a decrease in the excitability of the SMG–M1 connectionfollowing TBS (p=0.01) (Fig. 5).
Experiment 4In this experiment we found that for reaching to graspmovements
toward a central target there was a specific facilitation at 75ms duringthe planning phase of a PG movement as in experiment 1, even whenPG and WHG tasks were performed with the arm and the handapproaching the target object with the same posture. This borne outfrom ANOVA showing a significant GRASP TIME interaction (F=6,31;p=0.03). Post-hoc analysis showed that at 75 ms there was anincrease in the excitability of the SMG–M1 connection for PG incomparison with WHG (p=0.02) (Fig. 6).
DTI tractography
Anatomical evidence of continuous connectionsPICo was able to reconstruct a continuous tract in 10 out of 10
subjects for the following connections (Fig. 7):
• SMG to PMv: this tract descends from the SMG following the whitematter between the IPS and the angular gyrus and forms part of thesuperior longitudinal fasciculus (SLF); in particular, with referenceto the segmentation of the SLF (Makris et al., 2005; Croxson et al.,2005; Schmahmann et al., 2007), this tract forms part of SLF III;
• AG to PMv: this tract forms part of SLF II (Makris et al., 2005;Croxson et al., 2005; Schmahmann et al., 2007), it runs in directionposterior to anterior, dorsally and medially to the SMG to PMvconnection.
• PMv to M1: this subgyral short tract runs parallel to the previousone, in direction anterior-to-posterior, up to the coronal section
where the splenium of the corpus callosum appears, where itchanges direction to run inferiorly-to-superiorly.
For SMG to M1 connection, PICo was able to reconstruct acontinuous tract in 8 out of 10 subjects: this tract forms a subgyralshort connection between the pre- and the postcentral gyrus.
A tract connecting the AG to M1was reconstructed only in 5 out of10 subjects. This tract runs parallel to the AG to PMv in its first portion,then, at the level of the precentral gyrus, it changes abruptly directionto terminate in the primary motor cortex.
To provide an index of intersubject anatomical variability, a map ofthe between subject overlap is also provided (Fig. 8).
Strength of connectivity (SoC)Table 1 shows the mean strength of connectivity for both SMG and
AG towards M1 and PMv (averaged across subjects). Note that inmore than one subject there were some voxels in the seed, fromwhich none of the initiated streamlines reached a target, andtherefore the percentages do not add up to 100%. The values suggestthat the both SMG and AG are more likely to be directly connected toPMv then M1.
When the imperative signal instructed subjects to perform acentral PG, a positive correlation was found between the individualpeak facilitation of FDI MEPs induced by SMG stimulation (at 100 msafter the acoustic cue) and the SoC of the tract projecting from SMG toPMv (R=0.72, p=0.02). When subjects were instructed to perform alateral WHG, a positive correlation was found between the individualpeak FDI MEPs facilitation induced by AG stimulation (at 50 ms afterthe acoustic cue) with the SoC of the tract projecting from the AG to

Fig. 9. Correlation analyses between individual MEPs facilitation induced by PPCstimulation and tracts specific SoC provided further evidence of the linkage betweenmicrostructure and function of these parieto-motor pathways. Changes in MEPamplitude induced by SMG stimulation (individual values for peak of facilitation at100-ms delay during central PG preparation) correlated selectively with SoC of theSMG-PMv tract (A), but not with FA of other tracts; changes in connectivity due toAG stimulation (individual values for peak of facilitation at 50-ms delay duringlateral WHG preparation) correlated with SoC of the AG-PMv tract (C) but not withFA of the other tracts.
309G. Koch et al. / NeuroImage 51 (2010) 300–312
PMv (R=0.70, p=0.02) (Fig. 9), thus reinforcing the anatomo-functional relationship between SLF tracts and selective motor plans.
Discussion
These results demonstrate the existence of segregated anatomo-functional parieto-premotor-motor pathways that are involved inpreparation of different grasping actions and indicate that these mayprocess information relevant to both the position of the object and thehand shape required to use it.
Thus, the interaction between AG and M1 was modulated by boththe position of the object to be grasped as well as by the type of graspused to manipulate it (WHG); in contrast, the SMG interaction withM1 was specifically involved during preparation of a specific grasptype (PG) independently from the position of the object. Motor cortexexcitability did not vary over the same period, suggesting that themodifications in PPC–M1 connectivity reflect changes in the excit-ability of the stimulated regions (SMG or AG) rather than M1 itself.Temporary inactivation of PMv by means of cTBS abolished some of
the PPC–M1 interactions, suggesting that these circuits may include orat least be influenced by the ipsilateral PMv. Finally, DTI tractographyrevealed within the SLF the existence of connections between bothparietal areas and PMv, and to a lesser extent, M1. There was alsostrong anatomical connectivity between PMv–M1.
Functional interplay between AG and M1 during WHG
In a previous study (Koch et al., 2008a) we found that theexcitability of the interaction between AG and M1 was modulatedduring the reaction period when subjects had to point to a targetpositioned in contralateral space, but not if the target was in themidline. Here we show that this modulation is still present whensubjects have to pick up an object (cup), but only when they use awhole hand grip (WHG). If they are instructed to use a precision grip(PG) then there is no change in excitability in the reaction period. Weinterpret this change as reflecting an early biasing of movement plansin the contralateral direction that is specific for the WHG task. Ineffect, this connection appears to process information relevant to bothobject position and grasp type. Thus, the effect of object positionobserved for the AG–M1 connection may indicate that this pathway issensitive to the reach component of WHG task (Koch et al., 2008a).
The very early peak of facilitation observed at 50 ms would beconsistent with previous studies suggesting that PPC may be involvedin converting spatial specification of target location into motor plans(Andersen and Buneo, 2002; Snyder et al., 1997; Mattingley et al.,1998; Desmurget et al., 2009; Culham and Valyear, 2006; Prado et al.,2005) and that parietal regions may overrepresent the contralateralrelative to the ipsilateral workspace (Battaglia-Mayer et al., 2005).The novel aspect of the present results is that this spatial informationis affected by grasp type.
It has to be noted that the AG to M1 pathway was not activatedduring preparation of aWHG toward a target positioned in front of thesubject. This could imply that such movements could be coded byother nonprimary motor areas that were not tested in the currentstudy, such as the medial intraparietal sulcus (mIPS) or the caudalpart of the dorsal premotor cortex (PMd) as suggested by a recentfMRI investigation (Prado et al., 2005).
Functional interplay between SMG and M1 during PG
The results for the SMG–M1 interactionwere quite different: it wasmodulated by grasp type but not by position of the object. Thisoccurred at different delays from the acoustic cue in comparison withAG stimulation. In reaching to the central target, there was a specificpeak of facilitation for the M1 output to FDI some 75–100 ms after theimperative signal, but only if this instructed subjects to use a PG. Therewas no effect if a WHG was specified. The results were similar whensubjects were grasping the laterally placed object except that thefacilitation for PG was observed at 125 ms after the cue. We interpretthis change in excitability of SMG to M1 as reflecting a transfer ofinformation that is relevant to preparing activation of those handmuscles needed to perform precise grasping movements. Thisinterpretation is in line with studies in patients with circumscribedlesions of the anterior part of the PPC who have deficits in handpreshaping during visually guided reach-to-grasp movements (with arelative preservation of reaching) (Binkofski et al., 1998) and alsowiththe hypothesis that these regions may represent the humanhomologue of AIP (Culham and Valyear, 2006). Previous behaviouralinvestigations with TMS (using an interference approach rather thanthe bifocal TMS methodology introduced here) showing alteredmovement dynamics when magnetic pulses were applied duringvisually guided reach-to-grasp movements also support this role forSMG (Tunik et al., 2005; Rice et al., 2006). Our results are alsoconsistent with recent fMRI investigations, revealing that the left SMGis active when subjects perform a PG to reach and grasp a small object,

310 G. Koch et al. / NeuroImage 51 (2010) 300–312
with a negligible activity for WHG (Begliomini et al., 2007). They arealso in agreement with the idea that planning an accurate and precisemovement, such as PG, requires activity in a dorso-lateral circuit,which includes the SMG but not the AG (Grol et al., 2007).
Finally, our results may reflect previous findings from animalstudies, reporting that the neural coding of goal directed actions (i.e.,grasping for eating and grasping for placing) is mainly represented inthe AIP (Murata et al., 2000; Fogassi et al., 2005). As pointed out byRaos and colleagues (2003), in area AIP there are more visual-dominant neurons and visually responsive neurons than in PMv,while themotor neurons aremore represented in PMv than in AIP. AIPvisual-dominant neurons represent the shape, size, and/or orienta-tion of 3D objects, whereas AIP motor neurons probably represent theshape of the handgrip, grip size, or hand-orientation (Murata et al.,2000). These neurons described in AIP are thought to be involved inplanning different grasping movements by means of the reciprocalconnections linking AIP and F5 (PMv) (Rizzolatti and Luppino, 2001;Raos et al., 2003). Therefore the current findings of activation of SMG-M1 connectivity could represent the activation of such neural circuit,which is essential to recruit the appropriate “motor vocabulary”(Rizzolatti and Luppino, 2001).
There are several lines of evidence that suggest that the facilitationinduced by SMG and by AG stimulation may reflect the activation ofdistinct neural processes. First, the timing of the changes inexcitability was different. When subjects were planning a lateralWHG, a peak of interaction was observed as early as 50 ms for AGstimulation, while for SMG stimulation a peak occurred at 125 msafter the cue during a lateral PG. It is unlikely that two peakshappening with a delay of 75 ms could represent the same neuralprocess. Moreover, 50ms appears to be a very early timing after the gosignal to be interpreted as a phenomenon strictly related tovisuomotor transformation. In monkeys (although a different taskwas adopted) neural activity for a visuomotor transformation wasrecorded around 150 ms after the cue signal (Archambault et al.,2009). Since the position of the target was always known in advanceto subjects, we suggest that this early facilitation simply reflects therelease of preplanned information related to the spatial position of theobject. The fact that it was absent when the imperative signalinstructed a PG rather than a WHG suggests that this information hada direct input to the system and could cancel the prepared flow ofinformation. In contrast, it is possible that the later interactionsobserved between SMG andM1 during PG preparationmay reflect theneural activity coding the visuomotor transformations necessary toperform an accurate and precise movement.
We also measured whether the observed specific changes in PPC–M1 connectivitymay depend specifically onmodulation of excitabilityof the primary motor cortex itself. We did not find any change in MEPamplitude when measuring only the effects of TS alone in experiment1 (intervals ranging from 25 to 150 ms after onset of the imperativeauditory signal to perform a PG or a WHG). Moreover we failed todetect any change in intracortical facilitatory circuits at intervals of 50,75, 100, 125 ms (Experiment 2). This is consistent with severalprevious studies demonstrating that changes within M1 only arisejust before the onset of actual movements (Reynolds and Ashby,1999), not at early stages of movement planning (i.e., Koch et al.,2006) as tested in the current study.
Effects of cTBS over PMv
When cTBSwas applied over the left PMv, the task related PPC–M1connectivity changes were substantially reduced. In particular, thepeak of facilitation observed following AG stimulation duringplanning of lateral WHG at 50 ms SOA disappeared. Similarly, theincrease of excitability at 75 ms that occurred following AGstimulation during the reaction period of a central PG was abolished.Taken together, these results imply that the left PMv can be a crucial
node in the neural network linking AG and SMG with then ipsilateralM1. Therefore we may speculate that activation of cortico-corticalprojections originating from AG and SMG may synapse with neuronsof the ipsilateral PMv or at least be influenced by input from thatstructure. Further experiments are needed to fully elucidate the roleplayed by the connections with other regions of the network, such asfor instance PMd. A further limitation of the current experiment is thatwe did not explore the entire time course of activation of the SMG–M1connection following TBS and therefore we may have not detectedtemporal changes in the peaks of activation.
It might be argued that the interval that we used to test PPC–M1interactions (ISI=4 ms) is too short for an interposed synapticconnection in PMv. Indeed, previous TMS studies in resting subjectshave found that it takes a minimum of 2 ms for pulses applied to PMvto produce changes in excitability of M1 (Koch et al., 2007; Baumer etal., 2009). However the true latency of the interaction is complicatedby the fact that a single suprathreshold TMS pulse to M1 evokes aseries of I wave volleys in corticospinal neurones that can last 5 ms ormore. Since all of these contribute to the final amplitude of the MEP,inputs arriving as late as 9 ms after stimulation of PPC can affect thePPC–M1 interaction we measure. Thus, an ISI of 4 ms may besufficiently long to activate both direct and indirect (via PMv) circuitslinking PPC with M1.
Anatomical connections originating from AG and SMG
DTI tractography experiments revealed that AG and SMG areconnected with PMv by distinct fibres bundles of the SLF, likelycorresponding to the SLF II (fibres form AG) and SLF III (fibres fromSMG) subdivisions (Petrides and Pandya, 1984; Makris et al., 2005;Schmahmann et al., 2007). Analysis of the strength of connectivitysuggests that the SMG and AG are more likely to be directly connectedto PMv than M1. In fact, sparse direct connections between AG andSMG to M1 were found.
These findings are in agreement with previous anatomical studiesperformed in monkeys (Petrides and Pandya, 1984; Luppino et al.,1999; Rozzi et al., 2006; Brochier and Umilta, 2007; Borra et al., 2008),showing that most of the fibres originating from inferior parietallobule, that form part of the SLF (Schmahmann et al., 2007), terminatein PMv (F5). In contrast, direct prominent anatomical connectionsbetween the posterior parietal cortex and M1 have been describedonly for the superior parietal lobule (area PE) (Johnson et al., 1996;Matelli et al., 1998; Marconi et al., 2001).
Recent DTI investigations showed that the SLF III is prominentlylinkedwith PMv also in humans (Makris et al., 2005). Even though theSLF II was found to be highly connected with more dorsal prefrontalareas, reliable connections with PMv were observed also for this tract(Croxson et al., 2005). Our data are in line with this background,showing an higher probability of connection between SMG and PMvthan between AG and PMv (Table 1).
We also found correlations between individual TMS and SoCmeasures. There was a positive correlation between the SoC of theSMG-PMv tract and the amount of facilitation for the SMG-M1interaction during planning of PG; similarly there was a positivecorrelation between the SoC of the AG-PMv tract and the amount offacilitation for the AG-M1 interaction observed during preparation oflateral WHG. These data seem to suggest that subjects who havestronger anatomical representation of a certain white matter tractmay also have stronger neurophysiological interactions, supportingthe notion that the observed neurophysiological changes might bemediated by the anatomical pathways reconstructed by DTI analysis.
Nevertheless, it has to be noted that the current results, and thosein previous studies obtained linking other forms of DTI analysis withTMSmeasures (fractional anisotropy:Wahl et al., 2007; Boorman et al.,2007; Mars et al., 2009) were obtained in relatively small samples.Therefore, they should be takenwith caution and need to be replicated

311G. Koch et al. / NeuroImage 51 (2010) 300–312
in larger samples of subjects, since correlation analyses are highlysusceptible to interindividual variability.
In conclusion the current study reveals for the first time in humansthe existence of distinct functional and anatomical pathways linkingdifferent regions of the IPL with the ipsilateral motor cortex that areimportant during planning of grasping movements such as grasping asmall object precisely with the index and the thumb fingers or graspingwith the whole hand an object positioned in the contralateral space.Using combined noninvasive neurophysiological and neuroanatomicalmethods we show that these anatomo-functional connections becomeactive in a highly task-, condition- and time-dependent manner duringthe planning phase of different reaching to grasp actions. The currentmethod, if extended to other experimental conditions may providerelevant information to describe the function of other importantconnections. Moreover current information could also be important toinvestigate various neurological diseases inwhich a “miswiring” occurs,such as schizophrenia or stroke (Koch et al., 2008b,c) and to expandnew strategies of rehabilitation in patients with motor disabilities.
Acknowledgment
This work has been supported by grants of Ministero della Salute,Italy to G.K. (RF 06.60).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.neuroimage.2010.02.022.
References
Andersen, R.A., Buneo, C.A., 2002. Intentional maps in posterior parietal cortex. Annu.Rev. Neurosci. 25, 189–220.
Archambault, P.S., Caminiti, R., Battaglia-Mayer, A., 2009. Cortical mechanisms foronline control of hand movement trajectory: the role of the posterior parietalcortex. Cereb. Cortex 19, 2848–2864.
Aron, A.R., Behrens, T.E., Smith, S., Frank, M.J., Poldrack, R.A., 2007. Triangulating acognitive control network using diffusion-weighted magnetic resonance imaging(MRI) and functional MRI. J. Neurosci. 27, 3743–3752.
Ashburner, J., Friston, K.J., 2005. Unified segmentation. Neuroimage 26, 839–851.Basser, P.J., Mattiello, J., LeBihan, D., 1994. Estimation of the effective self-diffusion
tensor from the NMR spin echo. J. Magn. Reson. B 103, 247–254.Battaglia-Mayer, A., Mascaro, M., Brunamonti, E., Caminiti, R., 2005. The over-
representation of contralateral space in parietal cortex: a positive image ofdirectional motor components of neglect? Cereb. Cortex 15, 514–525.
Baumer, T., Schippling, S., Kroeger, J., Zittel, S., Koch, G., Thomalla, G., Rothwell, J.C.,Siebner, H.R., Orth, M., Munchau, A., 2009. Inhibitory and facilitatory connectivityfrom ventral premotor to primary motor cortex in healthy humans at rest – abifocal TMS study. Clin. Neurophysiol. 120, 1724–1731.
Begliomini, C., Wall, M.B., Smith, A.T., Castiello, U., 2007. Differential cortical activity forprecision and whole-hand visually guided grasping in humans. Eur. J. Neurosci. 25,1245–1252.
Binkofski, F., Dohle, C., Posse, S., Stephan, K.M., Hefter, H., Seitz, R.J., Freund, H.J., 1998.Human anterior intraparietal area subserves prehension: a combined lesion andfunctional MRI activation study. Neurology 50, 1253–1259.
Boorman, E.D., O'Shea, J., Sebastian, C., Rushworth, M.F., Johansen-Berg, H., 2007.Individual differences in white-matter microstructure reflect variation in func-tional connectivity during choice. Curr. Biol. 17, 1426–1431.
Borra, E., Belmalih, A., Calzavara, R., Gerbella, M., Murata, A., Rozzi, S., Luppino, G., 2008.Cortical connections of the macaque anterior intraparietal (AIP) area. Cereb. Cortex18, 1094–1111.
Brochier, T., Umilta, M.A., 2007. Cortical control of grasp in non-human primates. Curr.Opin. Neurobiol. 17, 637–643.
Castiello, U., 2005. The neuroscience of grasping. Nat. Rev. Neurosci. 6, 726–736.Croxson, P.L., Johansen-Berg, H., Behrens, T.E., Robson, M.D., Pinsk, M.A., Gross, C.G.,
Richter, W., Richter, M.C., Kastner, S., Rushworth, M.F., 2005. Quantitativeinvestigation of connections of the prefrontal cortex in the human and macaqueusing probabilistic diffusion tractography. J. Neurosci. 25, 8854–8866.
Culham, J.C., Valyear, K.F., 2006. Human parietal cortex in action. Curr. Opin. Neurobiol.16, 205–212.
Davare, M., Lemon, R., Olivier, E., 2008. Selective modulation of interactions betweenventral premotor cortex and primary motor cortex during precision grasping inhumans. J. Physiol. 586, 2735–2742.
Desmurget, M., Reilly, K.T., Richard, N., Szathmari, A., Mottolese, C., Sirigu, A., 2009.Movement intention after parietal cortex stimulation in humans. Science 324,811–813.
Fogassi, L., Ferrari, P.F., Gesierich, B., Rozzi, S., Chersi, F., Rizzolatti, G., 2005. Parietallobe: from action organization to intention understanding. Science 308, 662–667.
Grol, M.J., Majdandzic, J., Stephan, K.E., Verhagen, L., Dijkerman, H.C., Bekkering, H.,Verstraten, F.A., Toni, I., 2007. Parieto-frontal connectivity during visually guidedgrasping. J. Neurosci. 27, 11877–11887.
Huang, Y.Z., Edwards, M.J., Rounis, E., Bhatia, K.P., Rothwell, J.C., 2005. Theta burststimulation of the human motor cortex. Neuron 45, 201–206.
Jeannerod, M., Arbib, M.A., Rizzolatti, G., Sakata, H., 1995. Grasping objects: the corticalmechanisms of visuomotor transformation. Trends Neurosci. 18, 314–320.
Jenkinson, M., Bannister, P., Brady, M., Smith, S., 2002. Improved optimization for therobust and accurate linear registration and motion correction of brain images.Neuroimage 17, 825–841.
Johnson, P.B., Ferraina, S., Bianchi, L., Caminiti, R., 1996. Cortical networks for visualreaching: physiological and anatomical organization of frontal and parietal lobearm regions. Cereb. Cortex 6, 102–119.
Karnath, H.O., Perenin, M.T., 2005. Cortical control of visually guided reaching: evidencefrom patients with optic ataxia. Cereb. Cortex 15, 1561–1569.
Kloppel, S., Baumer, T., Kroeger, J., Koch, M.A., Buchel, C., Munchau, A., Siebner, H.R.,2008. The cortical motor threshold reflects microstructural properties of cerebralwhite matter. Neuroimage 40, 1782–1791.
Koch, G., Rothwell, J.C., 2009a. TMS investigations into the task-dependent functionalinterplay between human posterior parietal and motor cortex. Behav. Brain Res.202 (2), 147–152.
Koch, G., Rothwell, J.C., 2009b. Transcranial magnetic stimulation investigations ofreaching and grasping movements. In: Novak, D.A., Hermsdorfer, J (Eds.),Sensorimotor Control of Grasping: Physiology and Pathophysiology. CambridgeUniversity Press, pp. 72–83.
Koch, G., Franca, M., Fernandez Del Olmo, M., Cheeran, B., Milton, R., Alvarez Sauco, M.,Rothwell, J.C., 2006. Time course of functional connectivity between dorsalpremotor and contralateral motor cortex during movement selection. J. Neurosci.26, 7452–7459.
Koch, G., Fernandez Del Olmo, M., Cheeran, B., Ruge, D., Schippling, S., Caltagirone, C.,Rothwell, J.C., 2007. Focal stimulation of the posterior parietal cortex increases theexcitability of the ipsilateral motor cortex. J. Neurosci. 27, 6815–6822.
Koch, G., Fernandez Del Olmo, M., Cheeran, B., Schippling, S., Caltagirone, C., Driver, J.,Rothwell, J.C., 2008a. Functional interplay between posterior parietal and ipsilateralmotor cortex revealed by twin-coil transcranial magnetic stimulation during reachplanning toward contralateral space. J. Neurosci. 28, 5944–5953.
Koch, G., Ribolsi, M., Mori, F., Sacchetti, L., Codeca, C., Rubino, I.A., Siracusano, A.,Bernardi, G., Centonze, D., 2008b. Connectivity between posterior parietal cortexand ipsilateral motor cortex is altered in schizophrenia. Biol. Psychiatry 64,815–819.
Koch, G., Oliveri, M., Cheeran, B., Ruge, D., Lo Gerfo, E., Salerno, S., Torriero, S., Marconi,B., Mori, F., Driver, J., Rothwell, J.C., Caltagirone, C., 2008c. Hyperexcitability ofparietal-motor functional connections in the intact left-hemisphere of patientswith neglect. Brain 131, 3147–3155.
Kujirai, T., Caramia, M.D., Rothwell, J.C., Day, B.L., Thompson, P.D., Ferbert, A., Wroe, S.,Asselman, P., Marsden, C.D., 1993. Corticocortical inhibition in human motorcortex. J. Physiol. 471, 501–519.
Luppino, G., Murata, A., Govoni, P., Matelli, M., 1999. Largely segregated parietofrontalconnections linking rostral intraparietal cortex (areas AIP and VIP) and the ventralpremotor cortex (areas F5 and F4). Exp. Brain Res. 128, 181–187.
Makris, N., Kennedy, D.N., McInerney, S., Sorensen, A.G., Wang, R., Caviness Jr., V.S.,Pandya, D.N., 2005. Segmentation of subcomponents within the superiorlongitudinal fascicle in humans: a quantitative, in vivo, DT-MRI study. Cereb.Cortex 15, 854–869.
Marconi, B., Genovesio, A., Battaglia-Mayer, A., Ferraina, S., Squatrito, S., Molinari, M.,Lacquaniti, F., Caminiti, R., 2001. Eye-hand coordination during reaching. I.Anatomical relationships between parietal and frontal cortex. Cereb. Cortex 11,513–527.
Mars, R.B., Klein, M.C., Neubert, F.X., Olivier, E., Buch, E.R., Boorman, E.D., Rushworth,M.F., 2009. Short-latency influence of medial frontal cortex on primary motorcortex during action selection under conflict. J. Neurosci. 29, 6926–6931.
Matelli, M., Govoni, P., Galletti, C., Kutz, D.F., Luppino, G., 1998. Superior area 6 afferentsfrom the superior parietal lobule in the macaque monkey. J. Comp. Neurol. 402,327–352.
Mattingley, J.B., Husain, M., Rorden, C., Kennard, C., Driver, J., 1998. Motor role of humaninferior parietal lobe revealed in unilateral neglect patients. Nature 392, 179–182.
Murata, A., Fadiga, L., Fogassi, L., Gallese, V., Raos, V., Rizzolatti, G., 1997. Objectrepresentation in the ventral premotor cortex (area F5) of the monkey.J. Neurophysiol. 78, 2226–2230.
Murata, A., Gallese, V., Luppino, G., Kaseda, M., Sakata, H., 2000. Selectivity for the shape,size, and orientation of objects for grasping in neurons of monkey parietal area AIP.J. Neurophysiol. 83, 2580–2601.
Oldfield, R.C., 1971. The assessment and analysis of handedness: the Edinburghinventory. Neuropsychologia 9, 97–113.
Olivier, E., Davare, M., Andres, M., Fadiga, L., 2007. Precision grasping in humans: frommotor control to cognition. Curr. Opin. Neurobiol. 17, 644–648.
Pajevic, S., Pierpaoli, C., 1999. Color schemes to represent the orientation of anisotropictissues from diffusion tensor data: application to white matter fiber tract mappingin the human brain. Magn. Reson. Med. 42, 526–540.
Parker, G.J., Haroon, H.A., Wheeler-Kingshott, C.A., 2003. A framework for a streamline-based probabilistic index of connectivity (PICo) using a structural interpretation ofMRI diffusion measurements. J. Magn. Reson. Imaging 18, 242–254.
Petrides, M., Pandya, D.N., 1984. Projections to the frontal cortex from the posteriorparietal region in the rhesus monkey. J. Comp. Neurol. 228, 105–116.

312 G. Koch et al. / NeuroImage 51 (2010) 300–312
Prabhu, G., Shimazu, H., Cerri, G., Brochier, T., Spinks, R.L., Maier, M.A., Lemon, R.N.,2009. Modulation of primary motor cortex outputs from ventral premotorcortex during visually guided grasp in the macaque monkey. J. Physiol. 587,1057–1069.
Prado, J., Clavagnier, S., Otzenberger, H., Scheiber, C., Kennedy, H., Perenin, M.T., 2005.Two cortical systems for reaching in central and peripheral vision. Neuron 48,849–858.
Raos, V., Franchi, G., Gallese, V., Fogassi, L., 2003. Somatotopic organization of the lateralpart of area F2 (dorsal premotor cortex) of the macaque monkey. J. Neurophysiol.89, 1503–1518.
Raos, V., Umilta, M.A., Murata, A., Fogassi, L., Gallese, V., 2006. Functional properties ofgrasping-related neurons in the ventral premotor area F5 of the macaque monkey.J. Neurophysiol. 95, 709–729.
Reynolds, C., Ashby, P., 1999. Inhibition in the human motor cortex is reduced justbefore a voluntary contraction. Neurology 11, 730–735.
Rice, N.J., Tunik, E., Grafton, S.T., 2006. The anterior intraparietal sulcus mediates graspexecution, independent of requirement to update: new insights from transcranialmagnetic stimulation. J. Neurosci. 26, 8176–8182.
Rizzolatti, G., Luppino, G., 2001. The cortical motor system. Neuron 31, 889–901.Rozzi, S., Calzavara, R., Belmalih, A., Borra, E., Gregoriou, G.G., Matelli, M., Luppino, G.,
2006. Cortical connections of the inferior parietal cortical convexity of the macaquemonkey. Cereb. Cortex 16, 1389–1417.
Rozzi, S., Ferrari, P.F., Bonini, L., Rizzolatti, G., Fogassi, L., 2008. Functional organizationof inferior parietal lobule convexity in the macaque monkey: electrophysiologicalcharacterization of motor, sensory and mirror responses and their correlation withcytoarchitectonic areas. Eur. J. Neurosci. 28, 1569–1588.
Sakata, H., Taira, M., Murata, A., Mine, S., 1995. Neural mechanisms of visual guidance ofhand action in the parietal cortex of the monkey. Cereb. Cortex 5, 429–438.
Schmahmann, J.D., Pandya, D.N., Wang, R., Dai, G., D'Arceuil, H.E., de Crespigny, A.J.,Wedeen, V.J., 2007. Association fibre pathways of the brain: parallel observationsfrom diffusion spectrum imaging and autoradiography. Brain 130, 630–653.
Shimazu, H., Maier, M.A., Cerri, G., Kirkwood, P.A., Lemon, R.N., 2004. Macaque ventralpremotor cortex exerts powerful facilitation of motor cortex outputs to upper limbmotoneurons. J. Neurosci. 24, 1200–1211.
Snyder, L.H., Batista, A.P., Andersen, R.A., 1997. Coding of intention in the posteriorparietal cortex. Nature 386, 167–170.
Tomassini, V., Jbabdi, S., Klein, J.C., Behrens, T.E., Pozzilli, C., Matthews, P.M., Rushworth,M.F., Johansen-Berg, H., 2007. Diffusion-weighted imaging tractography-basedparcellation of the human lateral premotor cortex identifies dorsal and ventralsubregions with anatomical and functional specializations. J. Neurosci. 27,10259–10269.
Tuch, D.S., 2004. Q-ball imaging. Magn. Reson. Med. 52, 1358–1372.Tunik, E., Frey, S.H., Grafton, S.T., 2005. Virtual lesions of the anterior intraparietal area
disrupt goal-dependent on-line adjustments of grasp. Nat. Neurosci. 8, 505–511.Wahl, M., Lauterbach-Soon, B., Hattingen, E., Jung, P., Singer, O., Volz, S., Klein, J.C.,
Steinmetz, H., Ziemann, U., 2007. Human motor corpus callosum: topography,somatotopy, and link between microstructure and function. J. Neurosci. 27,12132–12138.
Wakana, S., Caprihan, A., Panzenboeck, M.M., Fallon, J.H., Perry, M., Gollub, R.L., Hua, K.,Zhang, J., Jiang, H., Dubey, P., Blitz, A., van Zijl, P., Mori, S., 2007. Reproducibility ofquantitative tractography methods applied to cerebral white matter. Neuroimage36, 630–644.




















