
In situ nitrogen enrichment experiments in two Idaho (U.S.A.) streams
13
IN SITU NITROGEN ENRICHMENT EXPERIMENTS IN TWO IDAHO (U.S.A,) STREAMS C. YVONNE MANUEL-FALER, G. WAYNE MINSHALL, RON W. DUNN, and DALE A. BRUNS Department of Biology, Idaho State University, Pocatello, Idaho (Received October 30, 1983) Abstract. We conducted a series of in situ experimental nutrient additions in two Idaho streams: Deep Creek, a desert stream located in Southeastern Idaho, and Big Wood River, situated in a mountainous region of Central Idaho. In both streams, a homogeneous reach was partitioned into almost identical channels. This allowed us to vary nutrient levels and measure algal response in experiments uncomplicated by differences in other factors. Ammonium nitrate was added to the treatment channels in Deep Creek. After nine days, Cladophora glomerata still predominated in both the control and treatment channels and algal biomass was not statistically different between the two. In the Big Wood River experiment significant differences in chloro- phyll a concentrations were not detected among three treatments of differing nitrogen enrichment and the control for both natural and artificial substrates. Also chlorophyll a levels were not correlated with nitrogen concentrations, and the composition of algal communities remained similar among all treatments. We conclude that macronutrient enrichment of streams does not necessarily result in an increase in algal density or a change in algal community structure so long as other factors, which may limit the use of nutrients by algae, remain unaltered. Our results suggest that light, current velocity, and macroinvertebrate grazing may account for the failure of the periphyton to respond to in situ additions of nitrogen (and phosphorus) in our study streams. 1. Introduction Algae in moderate abundance are natural and necessary components oflotic ecosystems. Frequently, however, excessive algal growth results in conditions that are detrimental to other stream biota and aesthetically displeasing to man. Therefore, management of algal levels in streams is a problem of common concern. Algal biomass levels'in streams have been positively correlated with concentrations of phosphorus and/or nitrogen (Tilley and Haushild, 1975; Stockner and Shortreed, 1976; Seki et al., 79; Marcus, 1980). The growth-stimulating effects of phosphorus and nitrogen enrichment have been demonstrated experimentally in laboratory algal bio- assay, chemostat, and other microcosm experiments (Greene et al., 1975; Rhee, 1978; Wilhm and Long, 1969), in outdoor artificial stream channels (Zimmerman, 1961; Stockner and Shortreed, 1978; Evans and Marcan, 1976), in experimental ponds (O'Brien and deNoyelles, 1974), in large enclosures in lakes (Goldman, 1962; Powers et al., 1972; Schindler, 1971)and in whole-lake studies (Schindler, 1971, 1975). These and other studies have provided a basis for the commonly held belief that nutrient increase in streams results in increased primary production, and have led to the establish- ment of State and Federal standards for nutrient concentrations in waters receiving industrial and domestic effluents. Environmental Monitoring and Assessment 4 (1984) 67-79. 0167-6369/84/0041-0067501.95. 1984 by D. Reidel Publishing Company.
Transcript of In situ nitrogen enrichment experiments in two Idaho (U.S.A.) streams

IN S I T U N I T R O G E N E N R I C H M E N T E X P E R I M E N T S IN TWO
I D A H O ( U . S . A , ) S T R E A M S
C. Y V O N N E M A N U E L - F A L E R , G. W A Y N E M I N S H A L L , RON W. D U N N , and DALE A. B R U N S
Department of Biology, Idaho State University, Pocatello, Idaho
(Received October 30, 1983)
Abstract. We conducted a series of in situ experimental nutrient additions in two Idaho streams: Deep Creek, a desert stream located in Southeastern Idaho, and Big Wood River, situated in a mountainous region of Central Idaho. In both streams, a homogeneous reach was partitioned into almost identical channels. This allowed us to vary nutrient levels and measure algal response in experiments uncomplicated by differences in other factors.
Ammonium nitrate was added to the treatment channels in Deep Creek. After nine days, Cladophora glomerata still predominated in both the control and treatment channels and algal biomass was not statistically different between the two. In the Big Wood River experiment significant differences in chloro- phyll a concentrations were not detected among three treatments of differing nitrogen enrichment and the control for both natural and artificial substrates. Also chlorophyll a levels were not correlated with nitrogen concentrations, and the composition of algal communities remained similar among all treatments.
We conclude that macronutrient enrichment of streams does not necessarily result in an increase in algal density or a change in algal community structure so long as other factors, which may limit the use of nutrients by algae, remain unaltered. Our results suggest that light, current velocity, and macroinvertebrate grazing may account for the failure of the periphyton to respond to in situ additions of nitrogen (and phosphorus) in our study streams.
1. Introduction
Algae in moderate abundance are natural and necessary components oflotic ecosystems. Frequently, however, excessive algal growth results in conditions that are detrimental to other stream biota and aesthetically displeasing to man. Therefore, management of algal levels in streams is a problem of common concern.
Algal biomass levels'in streams have been positively correlated with concentrations of phosphorus and/or nitrogen (Tilley and Haushild, 1975; Stockner and Shortreed, 1976; Seki et al., 79; Marcus, 1980). The growth-stimulating effects of phosphorus and nitrogen enrichment have been demonstrated experimentally in laboratory algal bio- assay, chemostat, and other microcosm experiments (Greene et al., 1975; Rhee, 1978; Wilhm and Long, 1969), in outdoor artificial stream channels (Zimmerman, 1961; Stockner and Shortreed, 1978; Evans and Marcan, 1976), in experimental ponds (O'Brien and deNoyelles, 1974), in large enclosures in lakes (Goldman, 1962; Powers et al., 1972; Schindler, 1971)and in whole-lake studies (Schindler, 1971, 1975). These and other studies have provided a basis for the commonly held belief that nutrient increase in streams results in increased primary production, and have led to the establish- ment of State and Federal standards for nutrient concentrations in waters receiving industrial and domestic effluents.
Environmental Monitoring and Assessment 4 (1984) 67-79. 0167-6369/84/0041-0067501.95. �9 1984 by D. Reidel Publishing Company.

68 C. Y. MANUEL-FALER ET AL.
However, Wuhrmann and Eichenberger (1975) found that the enrichment of groundwater with inorganic nitrogen and phosphorus had no significant effect on either algal biomass or algal community structure in semi-natural stream channels. Further- more, several field studies have shown that algal growth in streams is not necessarily correlated with or limited by nutrient concentrations (Kilkus et al., 1975; Campbell etal., 1976; Sumner and Fisher, 1979). Thus, to date, the evidence that nutrient enrichment in streams increases algal growth is inconclusive.
Greater predictive value, relative to the effect(s) of nutrient enrichment should derive from tests carried out in situ, where the natural complement of physical, chemical, and biological factors could be maintained. Studies of nutrient-rich streams, however, rarely include data from comparable non-enriched systems and studies involving the experi- mental manipulation of nutrient concentrations in natural streams are lacking.
In this paper we report results of in situ experimental nitrogen enrichment of two Idaho streams: Deep Creek, a cold-desert stream in Southern Idaho, and Big Wood River, a mountain stream of Central Idaho (Figure 1). Our approach was to partition a homogenous section of each stream into almost identical channels. This enabled us to vary nutrient levels and measure the algal response in experiments uncomplicated by differences in other factors.
BIG WOOD
IDAHO
eKetchum ~tello
P'" Holb ook DEEP CR.) e
;Great ? 'Salt L. I
Fig. 1. Site locations for experimental studies in Deep Creek and Big Wood River, Idaho.
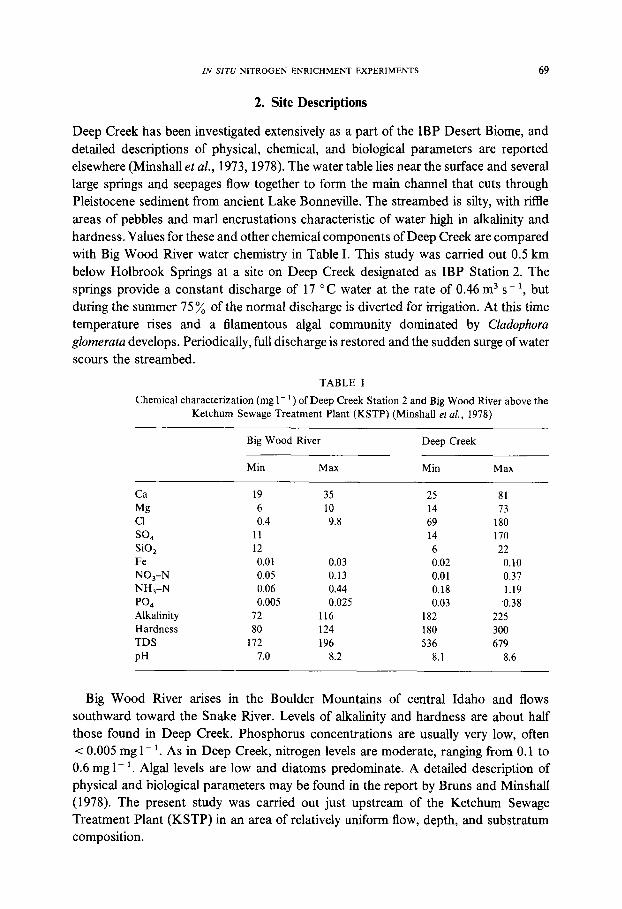
IN SITU NITROGEN ENRICHMENT EXPERIMENTS 69
2. Site Descriptions
Deep Creek has been investigated extensively as a part of the IBP Desert Biome, and detailed descriptions of physical, chemical, and biological parameters are reported elsewhere (Minshall et al., 1973, 1978). The water table lies near the surface and several large springs and seepages flow together to form the main channel that cuts through Pleistocene sediment from ancient Lake Bonneville. The streambed is silty, with riffle areas of pebbles and marl encrustations characteristic of water high in alkalinity and hardness. Values for these and other chemical components of Deep Creek are compared with Big Wood River water chemistry in Table I. This study was carried out 0.5 km below Holbrook Springs at a site on Deep Creek designated as IBP Station 2. The springs provide a constant discharge of 17 ~ water at the rate of 0.46 m 3 s- 1, but during the summer 75~o of the normal discharge is diverted for irrigation. At this time temperature rises and a filamentous algal community dominated by Cladophora glomerata develops. Periodically, full discharge is restored and the sudden surge of water scours the streambed.
TABLE I
Chemical characterization (mg 1 - 1 ) of Deep Creek Station 2 and Big Wood River above the Ketchum Sewage Treatment Plant (KSTP) (Minshall et aL, 1978)
Big Wood River Deep Creek
Min Max Min Max
Ca 19 35 25 81 Mg 6 10 14 73 C1 0.4 9.8 69 180 S O 4 11 14 170 SiO 2 12 6 22 Fe 0.01 0.03 0.02 0.10 NO3-N 0.05 0.13 0.01 0.37 NH3-N 0.06 0.44 0.18 1.19 PO4 0.005 0.025 0.03 0.38 Alkalinity 72 116 182 225 Hardness 80 124 180 300 TDS 172 196 536 679 pH 7.0 8.2 8.1 8.6
Big Wood River arises in the Boulder Mountains of central Idaho and flows southward toward the Snake River. Levels of alkalinity and hardness are about half those found in Deep Creek. Phosphorus concentrations are usually very low, often < 0.005 mg 1 - 1. As in Deep Creek, nitrogen levels are moderate, ranging from 0.1 to 0.6 mg 1- 1. Algal levels are low and diatoms predominate. A detailed description of physical and biological parameters may be found in the report by Bruns and Minshall (1978). The present study was carried out just upstream of the Ketchum Sewage Treatment Plant (KSTP) in an area of relatively uniform flow, depth, and substratum composition.

70 C. Y. MANUEL-FALER ET AL.
In addition to similar nitrogen levels, both streams are subject to the scouring action of high flow during snow-melt runoff and both have an open canopy. Big Wood River, however, receives more bank shading than Deep Creek. In Deep Creek the algae are predominantly filamentous and patchily distributed, whereas in Big Wood River the rocks support only a uniformly thin layer of diatoms. In both streams algal growth appears to be limited by some factor other than space.
3. Experimental Procedures
Our experiments in Deep Creek and Big Wood River were similar in basic design: nitrogen (ammonium nitrate) was added continuously to in situ channels and the response of algae was measured as changes in density (chlorophyll a or biomass) and/or community structure.
3.1. D E E P CREEK
In Deep Creek a homogenous riffle (26 m long x 2.8 m wide) was divided into two channels of equal area by constructing a partition using corrugated fiberglass panels and steel fence posts. Mean water velocity in the channels was 9 cm s-1. Ammonium nitrate, dissolved in stream was added to one channel (B). After mixing, the final concentration of nitrogen was 5.6 mgl-1 compared to 0.3 mgl-~ in the control channel (A). The duration of this experiment was nine days. Three 46-cm 2 samples were collected every other day from each channel by pushing a metal cylinder into the gravel and removing all enclosed algae. Algal biomass samples were then placed in tared crucibles, dried at 60 ~ for 3 d and weighed. On each sampling date a representative sample of algae from each channel was dried at 60 ~ C for 3 d, weighed, combusted at 450 ~ C for 4 hr and reweighed to determine the ratio of organic weight to dry weight. This ratio was then used to convert biomass from dry weight to organic weight (g ash free dry weight m - 2).
3.2. BIG WOOD RIVER
In Big Wood River four 0.5-m wide channels (A-D) were formed by partitioning a 2-m wide x 20-m long section of river. Partitions consisted of 0.6-m high 2-m long plywood panels painted white, fastened end-to-end, and secured in place by reinforcing steel rods driven into the riverbed.
River water was pumped into four polyethylene containers; anhydrous NH4NO 3 was added and the solution was delivered through PVC pipe to individual channels where flow was regulated by stopcocks. The volume of water flowing through the channels was monitored periodically and nutrient flow rates were adjusted to maintain nitrogen concentrations of approximately 0.5, 1.0, 2.0, and 4.0 mg 1 - 1 in channels A (control), B, C, and D, respectively.
Since the high N : P ratio in Big Wood River (normal range from 47 to 100 compared to 10 to 30 for Deep Creek) indicated that phosphorus was even more likely to be limiting than nitrogen, it seemed unlikely that the addition of nitrogen would increase algal

I N S I T U NITROGEN ENRICHMENT EXPERIMENTS 7l
growth if phosphorus levels remained low. Therefore, although our study was designed to test the effects of several levels of nitrogen enrichment, each of our experimental channels was enriched with phosphorus (equal amounts to each channel). Phosphorus was added by filling 390 ~tm mesh nylon bags (dimensions: 3 x 8 x 20 cm) with CaHPO 4 and suspending three such bags in the water at the head of each channel. All bags were refilled daily. During the day, bags were massaged hourly to clear pores in the mesh and one bag in each channel was refilled. Phosphorus levels ranged from 0.110 mg 1- 1 in D to 0.179 mg 1 - 1 in C. Total dissolved solids increased slightly (from 141 to 158 m g l - l ) with the addition of nutrients but alkalinity (113 mgl -~) and hardness (120 mgl -~) were practically unchanged.
Algal response to nutrient enrichment was evaluated by measuring chlorophyll a of periphyton on natural and artificial substrates and by comparing community structure of periphyton on artificial substrate. Direct measurements of biomass were not made because of the presence of large numbers of Simulium (Diptera: Simuliidae) larvae and pupae.
Natural substrate was sampled on day 17 and day 34 by covering the bottom of a 312 cm 2 tray with rocks (mostly cobbles) randomly selected from midway along the length of each channel. Rocks and attached periphyton were either frozen for later processing or chlorophyll extraction begun immediately. For processing, rocks were placed in a polyethylene container and covered with acetone. Extraction was done in the dark under refrigeration for > 24 hr followed by spectrophotometric determination with a Beckman DB-G spectrophotometer. Chlorophyll a was corrected for phaeophyton (Lorenzen, 1967) and results expressed as mg chl a m-2.
Artificial substrate consisted of sets of eight glass microscope slides (2.5 x 7.5 cm) inserted between plastic strips mounted on 20 x 25 cm plexiglass plates which were anchored horizontally on the riverbed. At the start of the experiment, sets of slides were placed in the upstream and downstream sections of each channel and in the river (outside the constructed channels) parallel to the upstream and downstream sites in channel A. Slides were removed from the water on day 34. A supplementary series of slides were incubated in the channels prior to the experiment reported here. Blackfly (Simulium) larvae and pupae were removed and periphyton from four slides of each set was scraped into polyethylene tubes (one tube per slide). Chlorophyll was extracted in 15 ml acetone and measured as described above. Community structure of algae was determined by counting individual cells in ten 4.8 x 103 gm 2 fields on one slide from each treament and expressing the results for each species as a percent of the total.
4. Results and Discussion
4.1. DEEP CREEK
Two days before the experiment was begun temporary restoration of normal flow dislodged and removed over half of the filamentous algae (mainly Cladophora glomerata). On day 1 algal standing crop was 36 g m- 2 in A (control) and 32 g m- 2 in B (Table II).

72 C. Y. MANUEL-FALER ET AL.
TABLE II Biomass of filamentous algae (g AFDW m- 2 + SE) in Deep
Creek
Day Channel
Control Nitrogen Student enriched t-test
1 36.4 + 6.1 32.5 + 4.1 NS 3 42.7 + 5.8 34.9 + 7.8 NS 5 63.8 + 2.9 57.8 + 9.1 NS 7 68.0 + 16.7 74.9 + 16.4 NS 9 70.8 + 8.3 76.6 + 7.6 NS
There were only minor changes in algal levels by day 3, but by day 5 mean biomass had increased to 63.8 g m - 2 in A. On the same day biomass in the enriched channel was 57.9 g m 2, an increase of 78~o compared to 75~o in A. Growth rate was lower after day 5 and there were no significant increases in either channel after day 7. Therefore, it is assumed that maximum levels had been reached. Mean biomass on day 9 was slightly higher in B (77 g m - 2) than in A (71 g m-2) , however, the difference was not
significant. A similar growth pattern and biomass level for filamentous algae also was observed in an experiment which tested the effects of phosphorus enrichment in Deep Creek (Minshall etaL, 1978). These studies indicate that values for biomass of filamentous algae in Deep Creek are lower than those for some other streams. Whitton
(1970) compiled data from several streams, all of which had a filamentous algal density of > 100gm -2.
Since algal biomass did not increase in response to nitrogen enrichment it is assumed that nitrogen was not the primary factor regulating algal levels in Deep Creek. Therefore, we have considered several other factors that might explain the failure of algal biomass to increase after day 7 or to attain levels comparable to those reported for other streams. These factors include phosphorus concentration, temperature, light, and the activities of invertebrates.
In another experiment (Minshall et al., 1978) we tested the effects of phosphorus enrichment with and without the addition of nitrogen. Algal biomass did not increase but there was a shift in community dominance from Cladophora to Spirogyra. The reason for this shift is unclear, particularly in view of the reports (cited by Whitton, 1970) that indicate that Cladophora has high phosphorus requirements and is favored by water with low N : P ratios. The experiment did indicate, however, that phosphorus probably is not limiting, and the results support the suggestion of Wong and Clark (1976) that a phosphorus concentration of 0.06 mg 1- ~ P is sufficient to promote maximum growth of Cladophora.
The experiments were carried out in midsummer, at a time when the spring growth of Cladophora typically disappears from most streams. This summer demise of Clado- phora has been reported by many investigators and is generally attributed to high water

IN SITU NITROGEN ENRICHMENT EXPERIMENTS 73
temperatures. However, temperatures can be ruled out as a limiting factor during the experiments in Deep Creek, since the 17 ~ source springs moderated temperature to limits well within those that normally promote growth of Cladophora (11 to 23 ~ according to Wong et aL (1978).
Deep Creek is a shallow, open-canopy stream with a summertime solar radiation mean of approximately 5500 kcal m- 2 day i. Normally, light is not considered to be a limiting factor, but the summer of 1975 was cooler than average, frequently cloudy, and insolution was estimated to be about 20?/0 lower than normal. Jaag (1938, cited by Whitton, 1970) observed that a series of cloudy days greatly reduced Cladophora production. Duffer and Dorris (1966) reported that algae in Blue River increased photosynthetic productivity with increases in solar radiation up to 72 cal m-2 h r - i (about 7200 kcal m -2 day-l , or around 50Yo higher than in Deep Creek in summer 1975). Thus, as algal mats developed in Deep Creek, it is probable that self-shading, coupled with the lower than normal insolation, resulted in light limitation of Cladophora production.
The effect of invertebrate activities on biomass of filamentous algae was not measured in this study. However, Koslucher and Minshall (1973) in a previous study in Deep Creek found only small amounts of Cladophora in the guts of invertebrates. Therefore, it is doubtful that direct consumption by invertebrates significantly affected the biomass of Cladophora. However, invertebrates may have regulated algal biomass by disruptive activities. Support for this hypothesis is provided by Eichenberger and Schlatter (1978) who found that in channels treated with insecticide, algal biomass (comprised of diatoms, filamentous green, and blue-green algae) reached levels greater than 200 g m - 2 whereas biomass in the untreated control channels was less than 50 g m -2. They attributed the difference partly to ingestion of food, but concluded that most of the losses were from the activities of the invertebrates that dislodged algae from the substrate in the process of feeding.
In summary, our data indicate that nitrogen was not limiting to the growth of filamentous algae in Deep Creek. Phosphorus concentration also did not appear to be limiting and indirect evidence suggests that water temperature was not critical. The effects of insufficient solar radiation and the activities of invertebrates may have combined to limit biomass to levels below those often encountered in streams of comparable nutrient enrichment.
4.2. WOOD RIVER
The concentrations of chlorphyll a on rocks collected on day 17 (August 28) of the experiment indicate that there were no differences in algal levels between the control channel (A) (58 mg-2) and the more enriched channels (Table III). However, all of the experimental channels including the control had higher concentrations of chlorophyll a than the river.
On day 34 estimates of mean chlorophyll a on rocks ranged from 41.1 mg m 2 in C to 65.0 mg m-2 in D. The more enriched channels, taken as a group did not have statistically higher concentrations of chlorophyll a than the control, which had a mean

74 C.Y. MANUEL-FALER ET AL.
TABLE III
Chlorophyll a of periphyton (mg m - 2) on rock and glass slides sampled from experimental channels and Big Wood River
A B C D River Analyses of variance
Among channels Between river and channels
Aug. 28 Rocks
Sept. 15 Rocks
Slides
Mean 57 .8 30 .5 54 .6 55 .0 3 5 . 3 ( A ) + ( C + D ) a ( R ) + ( A + C + D ) SD 3.5 6.0 12 .9 18.0 N 3 1 3 3 3 NS p < 0.05
Mean 46 .7 53 .9 41.1 65 .0 5 5 . 7 ( A ) + ( B + C + D ) ( R ) + ( A + B + C + D ) SD 22.4 29 .6 30 .9 12.2 20.1 N 3 3 3 3 3 NS NS
Mean 10.5 8.3 12.2 8.4 8.5 ( A ) + ( B + C + D ) ( R ) + ( A + B + C + D ) SD 7.4 2.9 2.3 3.8 3.8 N 7 7 7 7 7 NS NS
a A laboratory accident resulted in the loss of 2 samples from B, therefore this experimental channel is not included in ANOVA test.
concentration of 46.7 mg m -2. Moreover, on this second sampling date algal levels (based on rock samples) in the experimental channels were not significantly higher than
in the river. Levels of chlorophyll a of periphyton on glass slides (pooled upstream and down-
stream samples) were much lower than on rocks. However, the results of a comparison of slides from the four treatments were consistent with the data from the rock samples, i.e., there were no differences in concentrations of chlorophyll a that could be attributed to nitrogen enrichment. Also, as with the rock samples collected on the same day, the data show that algal levels in the experimental channels and in the river were not statistically different.
The thin layer of algae on the slides was mainly diatoms. Cor was the most abundant and together with Diatoma, occurred in all treatments. Other diatoms that were present in samples from one or more treatments were: Achnanthes, Cymbella cistula, Cymbella ventricosa, Fragilaria arcus, Gomphonema, and Synedra ulna. Six diatom species occurred on slides from the open river, 5 each from C and D, 4 from B, and 3 from A. A small, spherical (unidentified) blue-green alga was found in all samples but was never abundant.
Slides also had been placed in the river and in the experimental channels at the beginning of the two-week period prior to the experiment, when the nutrient delivery systems were being adjusted. During this time, they were exposed to nitrogen concen- trations ranging from 2 mg 1- 1 in A up to 16 mg 1 - 1 in D. Slides from each channel and from the river were removed and examined before the main experiment was begun.

IN S I T U N I T R O G E N E N R I C H M E N T E X P E R I M E N T S 7 5
Contrary to our expectations species richness at the end of this preliminary period was greater on slides from the channels (8-11 taxa) than from the fiver (6 taxa). Species richness was also greater on these slides than on any of the slides collected at the end
of the main experiment. Similarities in algal community structure among samples collected September 15 are
evident (Figure 2). The differences that exist were not related to nutrient concentration. Slides collected prior to the main experiment also had high among-treatment similarity in community structure but were, as a group, unlike the September set.
COCCONEIS SP, E'----'J FRAGILARIA SPP, L | CyMBELLA SPP, nTITrl DIATOMA SPP, I
~LCHNANTHES SPP, ~ ' ~ SYNEDRA SPP, k \ \ \ \ ~
NITZSCHIA SPP, ~iiii~ GOMPHONEMA SPP, ~ ' ~
AUGUST 8
i u �9 l n l l l [ I /
SE PTEi'IBE R 15
A B C D RIVER
CHANNEL
Fig. 2. Relative abundances of diatoms on glass slides in Big Wood River and in in situ experimental channels.
Failure of nutrient enrichment to modify algal densities or community structure in Big Wood River was even more surprising than in Deep Creek because of the much lower ambient nitrogen levels found in Big Wood River. There is some evidence, however, that the nutrient resources in the fiver may already be sufficient to support algae with higher biomass and a different community structure from that which occurred in the fiver at the time of our study. Concurrent with the in situ experiments we tested the effect of nutrient enrichment of Big Wood River in artificial stream experiments using water pumped continuously from the fiver (Manuel and Minshall, 1980). In all six artificial streams (including the control, with river water but no nutrients added) the predominant component of the periphyton was the filamentous alga Ulothrix, which grew in long strands on every available surface. Diatoms also were abundant, but grew mainly as

76 C. Y. MANUEL-FALER ET AL.
epiphytes on Ulothrix. Clorphyll a of periphyton on rocks collected from the control artificial stream September 7 (at the end ofa 16-d experiment)was 87 mg m - a compared to 56 mg m - : on rocks collected from the river September 15 (after a 34-day exposure). When either phosphorus or nitrogen was added, algal densities in the artificial streams more than doubled, and where both were added, chlorophyll a concentrations reached 693 mg m - z compared with 54 mg m - 2 in in situ channel A with similar nutrient con- centrations (approximately 0.1 mg 1 - 1 p and 1.0 mg 1- ~ N). Clearly, in the artificial streams some factor(s) differed from the river in such a way as to facilitate uptake and utilization of nutrients by algae and/or the retention of algae.
We were successful in simulating river conditions of temperature and light in the artificial stream experiment. However, two other important factors that affect algal densities, current velocity and invertebrate densities, were not duplicated. Quantitative evidence for the effect of current on periphyton is provided by Swanson and Bachmann (1976) who found that the daily export of benthic algae in Iowa streams ranged from 2.4~o to 8.9~ of the standing crop and Marker and Gunn (1977) who estimated downstream loss of benthic algae between March and May was over twice the standing crop. Tett et al. (1978) concluded that flow was the most important regulator of peri- phyton standing crops in the Mechums River. During our in situ study, mean current velocity (47 cm s - 1) was almost twice that of the artificial streams (26 cm s - 1). There- fore, we attribute some of the difference in algal densities between the artificial streams and the in situ channels to current velocity.
In the past, the effect of invertebrates on algal standing crop was largely ignored. However, Ball and Bahr (1975) found that summer suppression of periphyton produc- tivity coincided with the maximum demand by consumers in Red Cedar River. This is consistent with the results reported by Eichenberger and Schlatter (1978) (see above). In contrast, the observation by Sumner and Fisher (1979) that invertebrates consume less than 10~o of net periphyton production in Fort River suggests that, at least in some systems, invertebrates may not significantly affect algal density.
In the artificial stream experiment invertebrates were limited to those that could pass through the 1 mm mesh of the water intake, and that could adapt to the low current in the streams. Simulium, for example, could pass through the mesh (many were found attached to the plastic hose of the manifold that distributed river water to the streams) but none were observed in the streams themselves, presumably because of the low current velocity. In the river, however, the number of macroinvertebrates (in size classes > 390 lam) was estimated to be 6100 m - 2, based on samples collected 0.2 km upstream of the in situ channels (Bruns and Minshall, 1978). It is reasonable to assume that, at such high density, invertebrates contribute to the regulation of periphyton biomass.
Of particular relevance to the present study are the observations of Patrick (1948) of the superior substrate adhering properties of Cocconeis, and the resistance of Cocconeis to gazing by Physa heterostropha (Patrick, 1970). Hunter (1980), in agreement with Patrick, reported that snails were effective in removing filamentous algae and most diatoms from slides, but that Cocconeis and other small diatoms were not grazed and thus remained in unreduced numbers. Marcus (1980), however, discounted the effects

IN SITU NITROGEN ENRICHMENT EXPERIMENTS 77
of invertebrates (Simulium) and offered an alternate hypothesis to account for the predominance of Cocconeis observed on glass slides at 3 out of 4 sites downstream of a reservoir. At these sites nitrogen levels were lower than at a 4th site, where other diatoms were abundant. Although Simulium occurred mainly where Cocconeis predomi- nated, he concluded that Cocconeis was predominant because of a higher nutrient utilization efficiency which provided a competitive advantage in a nitrogen limiting environment. Our data do not support this hypothesis, since Cocconeis was the predomi- nant diatom on slides collected at the end of the main experiment. Simulium larva were present on all slides also. Although Simulium are primarily filter feeders, it has been observed that they spend part of their time grazing. In addition, their movement on the slides may tend to dislodge some diatoms and thereby favor others, such as Cocconeis, that attach more tenaciously to the substrate. Therefore, we attribute the predominance of Cocconeis, in part, to selective pressures associated with the presence of Simulium.
Cocconeis was not abundant in August, however, and this suggests that some factor associated with temporal changes was important in regulating a community that was apparently indifferent to nutrient changes.
3. Conclusions
Addition of nitrogen did not increase algal density in either Deep Creek or Big Wood River. In Big Wood River algal biomass remained sparse and continued to be dominated by diatoms even when nutrient levels were increased to 0.1 mg 1- l p and 3.3 mg 1- 1 N. In Deep Creek, filamentous algae was conspicuous but patchy, and biomass levels remained relatively low following the addition of nitrogen. These results indicate that some factors other than the concentration of macronutrients acted to regulate algal levels. The effects of grazing activities of insects, combined with low light intensity in Deep Creek, and with high current velocity in Big Wood River appear to be involved.
From our experiments in Deep Creek and Big Wood River we conclude that nutrient enrichment of streams does not necessarily result in an increase in algal density or a change in algal community structure so long as other factors, which may limit the utilization of nutrient by algae, remain unaltered. Conversely, a comparison of the in situ
experiments in Big Wood River with experiments carried out simultaneously in artificial streams indicates that when current velocity is reduced and/or invertebrates are excluded, algal levels may increase even in the absence of nutrient enrichment. Finally, these studies serve to emphasize that nuisance algal levels may not be due to high nutrient concentrations alone, but may be related to other factors which affect algal growth.
Acknowledgments
The research in Deep Creek was supported by the National Science Foundation IBP/Desert Biome and that in Big Wood River by a contract from the U.S. Army Corps of Engineers, Walla Walla, Washington. Special thanks are due to Dale McCullough

78 c.Y. MANUEL-FALER ET AL.
who developed the nutrient dispensing systems and to Tina Bruns who helped construct the in situ channels. Caryl Tickner Schmidt performed many of the laboratory analyses.
References
Ball, R. C. and Bahr, T. G.: 1975, 'Intensive Survey: Red Cedar River, Michigan', in River Ecology, Berkeley, University of California Press.
Bruns, D. A. and Minshall, G. W.: 1978, 'Effects of Drought on the Big Wood River', Idaho, U.S. Corps of Engineers, Walla Walla District.
Campbell, P. G., Couture, P., Lachance, M., and Talbot, L.: 1976, 'Etude integrre de la qualit6 des eaux des bassins versants des rivirres Saint-Francois et Yamaska', Vol. 2. Secteur des substances nntritives, INRS-Eau, Universit6 du Qurbec.
Duffer, W. R. and Dorris, T. C.: 1966, 'Primary Productivity in a Southern Great Plain Stream', Limnol. Oceanogr. 11, 143-151.
Eischenberger, E. and Schlatter, A.: 1978, 'Effect of Herbivorous Insects on the Production of Benthic Algal Vegetation in Out door Channels', Verh. Internat. Verein. Limnol. 20, 1806-1810.
Evans, G. H. and Marcan, E.: 1976, 'Diatom Community Response to Variations in Effluent Concentration', Environ. Pollut, 10, 115-126.
Goldman, C. R.: 1962, 'A Method of Studying Nutrient Limiting Factors in situ in Water Columns Isolated by Polyethylene Film', Limnol. Oceanogr. 7, 99-101.
Green, J. C., Miller, W. E., Shiroyama, T., and Maloney, T. E.: 1975, 'Utilization of Algal Assays to Assess the Effects of Municipal, Industrial, and Agricultural Wastewater Effluents upon Phytoplankton in the Snake River System', Water, Air, and Soil Pollut. 4, 415-434.
Hunter, R. D.: 1980, 'Effects of Grazing on the Quantity and Quality of Freshwater Aufwuchs', Hydrobiologia 69, 251-259.
Jaag, O.: 1938, 'Die Kryptogamenflora des Rheinfalls und des Hochrheins yon Stein bis Eglisau', Mitt. Naturf. Ges. SchlafJhausen 14, 1-158.
Kilkus, S. P., LaPerriere, J. D., and Bachmann, R. W.: 1975, 'Nutrients and Algae in some Iowa Streams', J. Water Pollut. Cont. 47, 1870-1879.
Koslucher, D. G. and Minshall, G. W.: 1973, 'Food Habits of some Benthic Invertebrates in a Northern Cold-Desert Stream (Deep Creek, Curlew Valley, Idaho-Utah)', Trans. Amer. Micros. Soc. 92, 441-452.
Lorenzen, C. J.: 1967, 'Determination of Chlorophyll and Pheo-Pigments: Spectrophotometric Equations', Limnol. Oceanogr. 12, 343-346.
Manuel, C. Y. and Minshall, G. W.: 1980, 'Limitations on the Use of Microcosms for Predicting Algal Response to Nutrient Enrichment in Lotic Systems', in J. P. Giesy, Jr. (ed.), Microcosms in Ecological Research, Aiken, S.C., Savannah River Ecology Lab. DOE Symposium Series No. 52, pp. 645-667.
Marcus, M. D.: 1980, 'Periphytic Community Response to Chronic Nutrient Enrichment by a Reservoir Discharge', Ecology 61, 387-399.
Marker, A. F. H. and Gunn, R. J. M.: 1977, 'The Benthic Algae in some Streams in Southern England. III. Seasonal Variations in Chlorophyll a in the Seston', J. Ecol. 65, 223-234.
Minshall, G. W., Andrews, D. A., Rose, F. L, Shaw, D. W., and Newell, R. L.: 1973, 'Validation Studies at Deep Creek, Curlew Valley', 1972 progress report, US/IBP Desert Biome.
Minshall, G. W., Manuel, C. Y., Dunn, R. W., Pace, R. K., McSorley, M. R., and Andrews, D. A.: 1978, 'Deep Creek Studies 1975-1976', US/IBP Desert Biome.
O'Brien, W. J. and DeNoyelles, F., Jr.: 1974, 'Relationship between Nutrient Concentration, Phytoplankton Diversity, and Zooplankton Diversity in Nutrient .Enriched Experimental Ponds', Hydrobiologia 44, 105-125.
Patrick, R.: 1948, 'Factors Affecting the Distribution of Diatoms', Bot. Rev. 14, 473-524. Patrick, R.: 1970, 'Benthic Stream Communities', Am. Sci. 58, 546-549. Powers, C. F., Schults, D. W., Malueg, K. W., Brice, R. M., and Schuldt, M. D.: 1972, 'Algal Responses
to Nutrient Additions in Natural Waters. II. Field Experiments', in Nutrients and Eutrophication: The Limiting Nutrient Controversy, American Society of Limnology and Oceanography, Special Volume I, pp. 141-154.
Rhee, G. Y.: 1978, 'Effects ofN : P Atomic Ratios and Nitrate Limitation on Algal Growth, Cell Composition, and Nitrate Uptake', Limnol. Oceanogr. 23, 10-25.

IN SITU NITROGEN ENRICHMENT EXPERIMENTS 79
Schindler, D. W.: 1971, 'Carbon, Nitrogen, and Phosphorus and the Eutrophication of Lakes', J. Phycol. 7, 321-329.
Schindler, D. W.: 1975, 'Whole-Lake Eutrophication Experiments with Phosphorus, Nitrogen, and Carbon', Verh. Internat. Verein. Limnol. 19, 3221-3231.
Seki, H., Hara, Y., and lchimura, S.: 1979, 'Impact of Nutrient Transfer into a Water System of River Sakuragawa and Lake Kasumigaura, Japan', Arch, Hydrobiol. 85, 291-301.
Stockner, J. C. and Shortreed, K. R. S.: 1976, 'Autotrophic Production in Carnation Creek, a Coastal Rainforest Stream on Vancouver Island, British Columbia', J. Fish. Res. Board Can. 33, 1553-1563.
Stockner, J. G. and Shortreed, K. R. S.: 1978, 'Enhancement of Autotrophic Production by Nutrient Addition in a Coastal Rainforest Stream on Vancouver Island', J. Fish. Res. Board Can. 35, 28-34.
Sumner, W. T. and Fisher, S. G.: 1979, 'Periphyton Production in Fort River, Massachusetts', Freshwater Biology 9, 205-212.
Swanson, C. D. and Bachmann, R. W.: 1976, 'A Model of Algal Export in some Iowa Streams', Ecology 57, 1076-1080.
Tett, P., Gallegos, C., Kelly, M. G., Hornberger, G. M., and Cosby, B. J.: 1978, 'Relationships among Substrate, Flow, and Benthic Microalgal Pigment Density in the Mechums River, Virginia', Limnol Oceanogr. 23, 785-797.
Tilley, L. J. and Haushild, W. L.: 1975, 'Use of Productivity of Periphyton to Estimate Water Quality',Journal WPCF 47, 2157-2171.
Whitton, B. A.: 1970, 'Biology of Cladophora in Freshwaters', War. Res. 4, 457-476. Wilhm, J. L. and Long, J.: 1969, 'Succession in Algal Mat Communities at Three Different Nutrient Levels',
Ecol. 50, 645-652. Wong, S. L. and Clark, B.: 1976, 'Field Determination of the Critical Nutrient Concentrations for Cladophora
in Streams', J. Fish Res. Board Can. 35, 85-92. Wong, S. L., Clark, B., Kirby, M., and Kosciuw, R. F.: 1978, 'Water Temperature Fluctuations and Seasonal
Periodicity of Cladophora and Potamogeton in Shallow Rivers', J. Fish Res. Board Can. 35, 866-870. Wuhrmann, K. and Eichenberger, E.: 1975, 'Experiments on the Effects of Inorganic Enrichment of Rivers
on Periphyton Primary Production', Verh. Internat. Verein. Limnol. 19, 2028-2034. Zimmerman, P.: 1961, 'Experimentelle Undersuchungen fiber die Oologische Wirkung der Str6mungsqesh
Windigkeit auf die Lebensgemeinschafter des fliessenden Wassers', Schweig. Z. Hydrol. 23, 1-81.




















