
In situ assay with the midge Kiefferulus calligaster for contamination evaluation in aquatic...
11
In situ assay with the midge Kiefferulus calligaster for contamination evaluation in aquatic agro-systems in central Thailand Ine ˆs Domingues a, * , Kriengkrai Satapornvanit b,c , Amararatne Yakupitiyage b , Amadeu M.V.M. Soares a , Anto ´ nio J.A. Nogueira a a CESAM & Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugal b Aquaculture & Aquatic Resources Management – School of Environment, Resources and Development, Asian Institute of Technology, P.O. Box 4, Klong Luang, Pathumthani 12120, Thailand c Kasetsart University, Paholyothin Road, P.O. Box 1054, 10900 Bangkok, Thailand Received 10 July 2007; received in revised form 4 January 2008; accepted 17 January 2008 Available online 10 March 2008 Abstract The aims of this study were to verify the suitability of in situ tests using the tropical midge Kiefferulus calligaster and to evaluate the most sensitive endpoint for the assessment of aquatic pesticide contamination. In situ tests were carried out in freshwater drainage chan- nels (farm channels) that supply vegetable crops and receive considerable pesticide spray drift, and at channels outside farms (main chan- nels). Moreover a pesticide-free farm was used as reference site. The endpoints analysed were: survival of the larvae, body length increment, capsule width increment, cholinesterase activity and glutathione S-transferase activity. Seasonal change was investigated as rainy season and dry season. Deleterious effects were observed at some farms especially during the rainy season when farmers apply hea- vier doses of pesticides. However, high mortality rates observed in main channels suggest that these water bodies are also affected by other impacts besides pesticide use. This work shows the potential of the in situ assay with K. calligaster as a tool for the environmental quality assessment of tropical aquatic ecosystems. Ó 2008 Elsevier Ltd. All rights reserved. Keywords: Tropical ecotoxicology; Pesticides; In situ assays; Biomarkers; Chironomids 1. Introduction Few studies have been done in tropical climates in terms of environmental risk assessment. Moreover, the registra- tion of new pesticides in developing countries typically relies on toxicity studies carried out on temperate species. However, information about the distribution, fate and mobility of pesticides and the factors that affect their pres- ence in temperate environments may not be applicable in tropical countries where climatic features, e.g. high temper- atures and rainfall, and agricultural practices differ (Abdul- lah et al., 1997). In situ bioassays are important tools for monitoring environment quality as it is difficult to represent in a labo- ratory experiment all the physical, chemical and biological stressors specific for a given exposure site (Den Besten et al., 2003). On the other hand, the simple detection and/or quantification of pesticides in both physical and biological compartments of an ecosystem has limitations in terms of costs and value of the information concerning the impact to communities. Thus, risk assessment studies should include a biological approach in which an evalua- tion of exposure of living organisms and subsequent effects at individual, population and/or community levels is done (Lagadic, 2002). Several methods have been developed for invertebrates in situ exposures and in situ chambers have been successfully designed for some chironomid taxa, 0045-6535/$ - see front matter Ó 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.chemosphere.2008.01.041 * Corresponding author. Tel.: +351 234 370790; fax: +351 234 426408. E-mail address: [email protected] (I. Domingues). www.elsevier.com/locate/chemosphere Available online at www.sciencedirect.com Chemosphere 71 (2008) 1877–1887
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of In situ assay with the midge Kiefferulus calligaster for contamination evaluation in aquatic...

Available online at www.sciencedirect.com
www.elsevier.com/locate/chemosphere
Chemosphere 71 (2008) 1877–1887
In situ assay with the midge Kiefferulus calligasterfor contamination evaluation in aquatic agro-systems
in central Thailand
Ines Domingues a,*, Kriengkrai Satapornvanit b,c, Amararatne Yakupitiyage b,Amadeu M.V.M. Soares a, Antonio J.A. Nogueira a
a CESAM & Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugalb Aquaculture & Aquatic Resources Management – School of Environment, Resources and Development, Asian Institute of Technology,
P.O. Box 4, Klong Luang, Pathumthani 12120, Thailandc Kasetsart University, Paholyothin Road, P.O. Box 1054, 10900 Bangkok, Thailand
Received 10 July 2007; received in revised form 4 January 2008; accepted 17 January 2008Available online 10 March 2008
Abstract
The aims of this study were to verify the suitability of in situ tests using the tropical midge Kiefferulus calligaster and to evaluate themost sensitive endpoint for the assessment of aquatic pesticide contamination. In situ tests were carried out in freshwater drainage chan-nels (farm channels) that supply vegetable crops and receive considerable pesticide spray drift, and at channels outside farms (main chan-nels). Moreover a pesticide-free farm was used as reference site. The endpoints analysed were: survival of the larvae, body lengthincrement, capsule width increment, cholinesterase activity and glutathione S-transferase activity. Seasonal change was investigated asrainy season and dry season. Deleterious effects were observed at some farms especially during the rainy season when farmers apply hea-vier doses of pesticides. However, high mortality rates observed in main channels suggest that these water bodies are also affected byother impacts besides pesticide use. This work shows the potential of the in situ assay with K. calligaster as a tool for the environmentalquality assessment of tropical aquatic ecosystems.� 2008 Elsevier Ltd. All rights reserved.
Keywords: Tropical ecotoxicology; Pesticides; In situ assays; Biomarkers; Chironomids
1. Introduction
Few studies have been done in tropical climates in termsof environmental risk assessment. Moreover, the registra-tion of new pesticides in developing countries typicallyrelies on toxicity studies carried out on temperate species.However, information about the distribution, fate andmobility of pesticides and the factors that affect their pres-ence in temperate environments may not be applicable intropical countries where climatic features, e.g. high temper-atures and rainfall, and agricultural practices differ (Abdul-lah et al., 1997).
0045-6535/$ - see front matter � 2008 Elsevier Ltd. All rights reserved.
doi:10.1016/j.chemosphere.2008.01.041
* Corresponding author. Tel.: +351 234 370790; fax: +351 234 426408.E-mail address: [email protected] (I. Domingues).
In situ bioassays are important tools for monitoringenvironment quality as it is difficult to represent in a labo-ratory experiment all the physical, chemical and biologicalstressors specific for a given exposure site (Den Bestenet al., 2003). On the other hand, the simple detectionand/or quantification of pesticides in both physical andbiological compartments of an ecosystem has limitationsin terms of costs and value of the information concerningthe impact to communities. Thus, risk assessment studiesshould include a biological approach in which an evalua-tion of exposure of living organisms and subsequent effectsat individual, population and/or community levels is done(Lagadic, 2002). Several methods have been developed forinvertebrates in situ exposures and in situ chambers havebeen successfully designed for some chironomid taxa,

1878 I. Domingues et al. / Chemosphere 71 (2008) 1877–1887
namely Chironomus riparius (Crane et al., 2000; Castroet al., 2003; Den Besten et al., 2003) and Chironomus ten-
tans (Chappie and Burton, 1997; Sibley et al., 1999). Chi-ronomid in situ approaches include ecological riskassessment in agricultural areas and estuaries using Chiron-
omus riparius (Crane et al., 1995a; Soares et al., 2005b);assessment of metal bioavailability with Chironomus crass-
iforceps (Peck et al., 2002), and assessment of urban andagricultural runoff with Chironomus tentans (Tucker andBurton, 1999).
In the aquatic environment the concentration of pollu-tants may not always be high enough to cause mortality,thus it should be useful to include other functional end-points in ecological risk assessment. The biomarkersapproach has been widely used as it is based on theassumption that low toxicant concentrations cause a bio-chemical response within individuals before the manifesta-tion of those effects can be observed at higher levels ofbiological organization (e.g. Crane et al., 1995b; Escartinand Porte, 1996; McLoughlin et al., 2000; Robillardet al., 2003; Chang et al., 2006). As such, a biomarkercan be generally defined as a cellular or biochemical com-ponent, process, structure or function that can be xenobiot-ically induced and measured in a biological system orsample (Kendall et al., 2001). Two enzymes are of particu-lar interest as biochemical biomarkers for risk assessmentof pesticide use: acetylcholinesterases (AChE) and glutathi-one transferases (GST’s).
AChE has a key role in the maintenance of normal nervefunction and is inhibited by neurotoxic compounds such asphosphate and carbamate esters (commonly used as insec-ticides) that bind to the enzyme and lead to the accumula-tion of acetylcholine in the synapse resulting in thedisruption of the normal nervous system function (Olsenet al., 2001). In consequence, AChE inhibition has usedas a specific biomarker for organophosphate and carba-mate pesticides.
GSTs are a family of enzymes catalyzing the conjuga-tion of reduced glutathione (GSH) in the detoxificationof compounds having electrophilic centers (nitrocom-pounds, organophosphates, organochlorines). The GSHconjugation products become less toxic and more solublein water so that they can be easily excreted (Olsen et al.,2001). The increase in GST activity has been used as a bio-marker of organic contamination.
Biomarkers have already been successfully used as toolsfor in situ assessment of pesticides in both invertebrates(Radenac et al., 1998; Forget et al., 2003; Moreira andGuilhermino, 2005) and vertebrates (Sturm et al., 1999;Sanchez-Hernandez and Walker, 2000). Although thereare some constraints in the use of biomarkers relating totheir specificity, ecological relevance and the influence ofadditional non-contaminant factors on enzyme response,they have represented an important contribution to ecolog-ical pesticide risk assessment (Lagadic, 2002).
This study was part of a European project (MAMAS-managing agrochemical in multiuse aquatic systems) that
aimed to develop a battery of in situ tests using indigenousorganisms to assess pesticide contamination in aquaticenvironments that surround intensive multi-crop farms incentral Thailand. The objectives of this specific study areto verify the suitability of in situ tests with the tropicalmidge Kiefferulus calligaster and to evaluate the most sen-sitive endpoints for the assessment of aquatic pesticide con-tamination. Endpoints span a wide range of sensitivitiesfrom the molecular (biomarkers) to the individual (growthand mortality) levels.
2. Materials and methods
2.1. Test animals and culture conditions
Organisms deployed in field tests were based on theregionally common midge, K. calligaster. Individuals werecollected from the field, in an agricultural area of centralThailand (Nakhon Chaisi district, June 2002), and main-tained in the laboratory for several generations beforebeing deployed in the field. Laboratory cultures weremaintained in the aquaculture and aquatic resources man-agement laboratory (Asian Institute of Technology, Thai-land) following standard procedures (EPA, 2000). Theculture was kept under conditions of temperature and pho-toperiod identical to the observed in the exterior (28 ± 2 �Cand 12:12 h light:dark cycle) and larvae were fed every twodays with a suspension of Tetramin�, Tetra GmbH D-49304 totalizing 0.5 mg of Tetramin/day/larva (Peryet al., 2002). ASTM hard water (ASTM, 1980) was usedas culture medium in plastic culture vessels (base dimen-sions 20 � 10 cm) with a thin (1 cm) layer of sediment(holding approximately 80 larvae), gentle aeration wasprovided.
Third instar larvae were used in the field deploymentssince at this stage larvae are big enough to be handled eas-ily and will undergo moulding during the four days of fieldexposure (as they grow to the fourth instar) representing aperiod of higher sensitivity to pollutants.
2.2. Study sites
The study sites were located in the village of Kokpraja-dee, Pathum Thani district, central Thailand in farmingsystems characterized by long beds (2–3 m wide), uponwhich crops are grown, separated by freshwater irrigationditches (0.5–1 m wide, 0.2–0.5 m depth) that are suppliedby an outside source of water through either a channel inletor motorised pump. Principal crops produced are Chinesecabbage and Chinese kale. Farmer residence is located inthe vicinity of the farming area and, very often, water fromthe farm channels is also used for bathing, cooking andwashing cloths and dishes. Large quantities of pesticidesare applied to crops to kill pests and also as a preventivemeasure with a high probability of spray drift enteringadjacent water channels. Various pesticides are applied;some belonging to classes Ib (Dicrotophos) and II (Profe-

I. Domingues et al. / Chemosphere 71 (2008) 1877–1887 1879
nofos, Paraquat and Chlorfenapyr) of the World HealthOrganization’s (WHO) classification system: ‘‘WHO rec-ommended classification of pesticides by Hazard”. A preli-minary ecological risk assessment has been made for thisarea based on an estimation of a PEC/NEC ratio (pre-dicted environmental concentration/no effect concentra-tion), considering spray drift as the only entry route ofpesticide in the water. For all situations studied a potentialrisk was found (ratio PEC/NEC > 1), indicating that sig-nificant effects of pesticide exposure could be expected onaquatic life (Van den Brink et al., 2003; Satapornvanitet al., 2004). In this study, target sites were the channelssurrounding cultivation areas (farm channels – FC) andthe channels outside the farms (main channel – MC) thatserve as the water source to the farm (Fig. 1). An organicfarm (OF) was chosen as reference site since no pesticidesare applied.
2.3. In situ bioassay chamber and general protocol
The assay method was based on the exposure setdescribed by Soares et al. (2005a) specifically for chirono-mids with various endpoints: survival (S), head capsulewidth increment (CW), body length increment (BL), ChEactivity (ChE) and GST activity (GST). Test chambersconsisted of a 20 cm opaque PVC tube (6 cm diameter)with two lateral windows (10 � 4 cm) covered with200 lm nylon mesh. Both ends of the chamber are coveredwith mesh but on the upper one the mesh is removable(fixed with a rubber band) to allow introduction andremoving of sediment. The mesh was sealed to the chamberwith white thermal glue (Elis-Tawain, Tawain, TN122/WS). Four farms were selected as sites for in situ deploy-ment, namely Surat farm (S1), Chanchai farm (S2), SomJit farm (S3) and Chuchat farm (S4). Pesticide sprayingwas carried out between one and seven days before eachin situ deployment. Pesticides applied were: dicrotophos,chlorfenapyr and abamectin at S1; diafenthiuron + fenoxy-
Fig. 1. Diagram of a typical vegetable far
carb, dicrotophos, senep and chlorfenapyr at S2 and prof-enophos at S3 and S4. Samples of water and sediment weretaken for pesticide analysis. However, results of pesticideanalysis can not be presented since a problem with the ana-lytical equipment resulted in inaccurate values and sampleloss. In each site one deployment set was placed in the farmchannel and another in the main channel. Six replicatedchambers with 10 organisms each (the maximum numberof larvae attending to the dimensions of test chamber) con-stituted each set, placed in a holding structure consisting ofmetal wire basket. In this way, the total number of animalsexposed at each place was 60. One additional set wasdeployed in the organic farm. The same experimentaldesign was conducted in the rainy season and the dryseason.
2.3.1. Day 1
The day before starting the assay, the structure holdingthe chambers and the chambers were placed in the studyand reference sites, attached to the margins by a thin rope(no current was observed in these channels). A 2 m longtransparent plastic tube was connected to each chamberto allow the introduction of chironomids from the bankat day 0. Placement of deployment sets did not interferewith farming activities or boat passage. Artificial sediment(sand washed and sieved through 1 mm sieve) was usedsince preliminary trials showed that the local sedimentwas not suitable for bioassays as it is constituted by mudwith a dark colouration causing difficulties for chironomidretrieval (recovering rate of chironomids on mud was12.5% against 79.2% in clean sediment). Farm canals canbe drained, rebuilt and refilled several times a year, thusinterfering with fine particles accumulation at the bottomof the canals.
2.3.2. Day 0
Third instar chironomid larvae were transported to thefield in small glass vials with ASTM hard water. Ten larvae
m in Kokprajadee, central Thailand.

1880 I. Domingues et al. / Chemosphere 71 (2008) 1877–1887
were introduced in each chamber through the plastic tubewith the help of a Pasteur pipette. ASTM culture mediumwas also poured through the tube with the help of a syringeto propel the larvae to the chamber. An excess of food(1.5 ml of a Tetramin suspension (20 mg/ml)) was intro-duced in the same way to feed the animals and avoidstarvation.
2.3.3. Day 4Chambers were removed from the holding structure and
placed in the bucket filled with water from the channels andtransported directly to the laboratory.
In the laboratory sediment was removed from the cham-bers, inspecting chambers for chironomids that may havebecome stuck to the walls or mesh. Chironomids and sed-iments were sieved to separate organisms from particulatematter before calculating larval survival rate. Live chiron-omids from each set were divided into three groups: (i) ani-mals were preserved in ethanol 70% solution for bodylength and head capsule width measurements; (ii) and (iii)the entire animals were snap-frozen individually in eppen-dorfs with 1 ml phosphate buffer, 0.1 M, pH = 7.2 for cho-linesterase (ChE) and 6.5 for glutathione-S-transferase(GST) analysis (Guilhermino et al., 1996). Samples werefrozen at �20 �C for ChE determinations and at �80 �Cfor GST determinations for a maximum of two weeks.
2.4. Physicochemical analysis of water
Water samples were collected in dark glass bottles at thesites at the same time as bioassay deployment was initiatedfor chemical analysis. Analysis followed standard methods(Clesceri et al., 1989) and included the determination ofnitrite, ammonia, chlorophyll a, TVS (total volatile solids)and TSS (total suspended solids). Dissolved oxygen con-centration (DO), pH, water temperature and conductivitywere measured in situ with portable meters (pH meter-WTW pH 330i/SET,WTW, Weilheim Germany; conduc-tivity meter-WTW COND 330i/SET,WTW, Weilheim,Germany; Oximeter-WTW OXI 330/SET,WTW, Weil-heim, Germany).
2.5. Biomarkers analysis
Samples were defrosted in ice, homogenized in a YstralGmbH D-7801 tissue homogenizer and activity determina-tions performed on the supernatants obtained by centrifu-gation of the homogenates in a high speed brushlesscentrifuge (MPW med instruments, MPW-350R) (4 �C,3348g, 3 min for ChE and 4 �C, 8931g, 30 min for GST)(Moreira and Guilhermino, 2005).
The ChE assay was performed at 414 nm according tothe method of Ellman et al. (1961) adapted to microplate(Guilhermino et al., 1996), using 0.05 ml of homogenateand 0.250 ml of the reaction solution (1 ml of 5.50-dithio-bis-2-nitrobenzoic acid (DTNB) 10 mM solution, 0.2 ml
of 0.075 M acetylcholine solution and 30 ml of 0.1 M phos-phate buffer).
GST activity was determined at 340 nm by the methodof Habig et al. (1974) adapted to microplate (Boothet al., 2000), using 0.1 ml of homogenate and 0.200 ml ofthe reaction mixture (reduced glutathione (GSH) at10 mM and 1-chloro-2.4-dinitrobenzene (CDNB) at60 mM).
All enzymatic activities were determined in quadrupli-cate and expressed as nanomol of substrate hydrolyzedper minute per mg of protein. Protein concentrations weredetermined in quadruplicate by the Bradford method(Bradford, 1976) at 595 nm using c-globulin as standard.A Labsystem Multiskan EX microplate reader was usedfor all determinations.
2.6. Statistical analysis
For each season normality of data was tested (Kol-mogorov Smirnov normality test) and the homogeneityof variance was verified (Barlett’s test). Following transfor-mations were performed to normalise data: body lengthincrement and GST: log10; capsule width increment andChE: square root; survival: arcsin square root. One-wayanalysis of variance (ANOVA), followed by the Dunnet(post hoc) test was used to assess differences between studysites and references. The hypothesis that activities werelower than the control was tested in the case of ChE, whilea (greater than, less than) difference from the control wastested for the other endpoints. For sites S2 and S3, whereno mortality was observed and data for farm channeland main channel was available for all endpoints, athree-way ANOVA was performed to assess influence ofseason, type of channel and study site (farm). SigmaStat(SPSS, 2004) was used to perform all statistical analysis.The significance level used in all statistical testing was0.05. A redundancy analysis (RDA) was performed on end-point-datasets (as species data) including physicochemicalwater parameters (as environmental data) following thelog-transformation of data. Multivariate analysis was per-formed using CANOCO 4.5 (Ter Braak and Smilauer,2002).
3. Results
3.1. Multivariate analysis
Results from RDA are summarised in Fig. 2 with astrong correlation between toxicity endpoints and waterproperties on the first two axes (0.964 and 0.934, respec-tively). The first two axes explained 71.3% of the variationin toxicity endpoints, while the interaction between toxicityparameters and water physicochemical parameters accountfor an additional 20.1% of the total variability. Seasonswere identified by a gradient where higher levels of GSTactivity and lower survival rates are associated with therainy season. Body length and head capsule width incre-

Fig. 2. Species-environment triplot diagram from the RDA analysis. Endpoint-datasets as species data: S (survival), BL (body length increment), CW(capsule width increment), ChE (cholinesterase activity) and GST (glutathione S-transferase activity) and physicochemical water parameters asenvironmental variables: nitrite, ammonia, chloroph (chlorophyll a), TVS (total volatile solids) and TSS (total suspended solids), pH, conduct(conductivity), water T (water temperature) and DO (dissolved oxygen concentration).
resp
onse
(per
cent
age
of la
bora
tory
leve
ls)
EndpointSeason
GSTAChECWBLSurvivalDry RainyDry RainyDry RainyDry RainyDry Rainy
200
150
100
50
0
100
Fig. 3. Variation of endpoints measured at the reference site compared tolaboratory values measured under control conditions (BL-body lengthincrement; CW-capsule width increment, AChE-cholinesterase activity,GST-glutathione S-transferase activity).
I. Domingues et al. / Chemosphere 71 (2008) 1877–1887 1881
ments were more weakly associated with season, increasingtowards dry season. Monte Carlo forward selection identi-fied the effects of nitrite and total suspended solids as sig-nificant (see Table 1).
3.2. Reference site
The organic farm showed some variability between sea-sons for all endpoints (BL: F1,15 = 24.61, p < 0.001; CW:F1,15 = 50.33, p < 0.001; ChE: F1,11 = 34.70, p < 0.001;GST: F1,9 = 5.14, p = 0.05) except for survival (F1,19 =2.63; p = 0.121). Variation in parameters measured at theorganic farm relative to laboratory values are representedin Fig. 3. Mean values measured during rainy season com-pared to lab values ranged between 35% and 64% for bodylength increment; 37% and 96% for capsule width incre-ment; 35% and 60% for ChE activity and 79% and 160%for GST activity. Survival at reference site ranged between75% and 100%. Values for the parameters measured at theorganic farm during dry season compared to lab valuesranged between 17% and 30% for body length increment;12% and 18% for capsule width increment; 76% and 92%for ChE activity and 67% and 75% for GST activity. Sur-vival at reference site ranged between 73% and 100%.
Table 1Significant conditional effects obtained using Monte Carlo forwardselection
Variable Var N Lambda A p F
Nitrite 5 0.26 0.006 6.64TSS (total suspended solids) 8 0.26 0.012 4.5Conductivity 3 0.07 0.098 2.16
3.3. Survival
Two sites, S1 and S4, had recurrently high mortality orno survival in the main channels regardless of season. Infarm channels one site showed significantly lower survivalthan the reference (S2 in the rainy season; Fig. 4). Three-way ANOVA analysis of S2 and S3 sites showed that therewere significant differences between seasons (lower survivalduring the rainy season) and between sites with a stronginteraction between sites and channels (Table 2).
3.4. Developmental endpoints
During the rainy season, three farms (S1, S2 and S3) hadlower increments on body length compared to reference,although differences were only statistically significant for
R S1 S2 S3 S4
GS
T a
ctiv
ity
(n
mo
l/ m
in/ m
g p
rot)
0
10
20
30
40
R S1 S2 S3 S40
10
20
30
40R S1 S2 S3 S4
Ch
E a
ctiv
ity
(n
mo
l/ m
in/ m
g p
rot)
0
20
40
60
80
100
R S1 S2 S3 S40
20
40
60
80
100R S1 S2 S3 S4
Cap
sule
wid
thin
crem
ent
(mm
)
0.0
0.1
0.2
0.3
0.4
0.5
R S1 S2 S3 S40.0
0.1
0.2
0.3
0.4
0.5R S1 S2 S3 S4
Bo
dy
len
gth
in
crem
ent
(mm
)
0
1
2
3
4
5
R S1 S2 S3 S4
0
1
2
3
4
5R S1 S2 S3 S4
Su
rviv
al (
%)
0
20
40
60
80
100
120
140
R S1 S2 S3 S4
0
20
40
60
80
100
120
140 Farm channelMain channelReference
Rain Season Dry season
study sites
* *
**
* *
*
* *
*
***
*
©
©
©
©
©
©
©
©
@ © ©
*
@
@
@
@
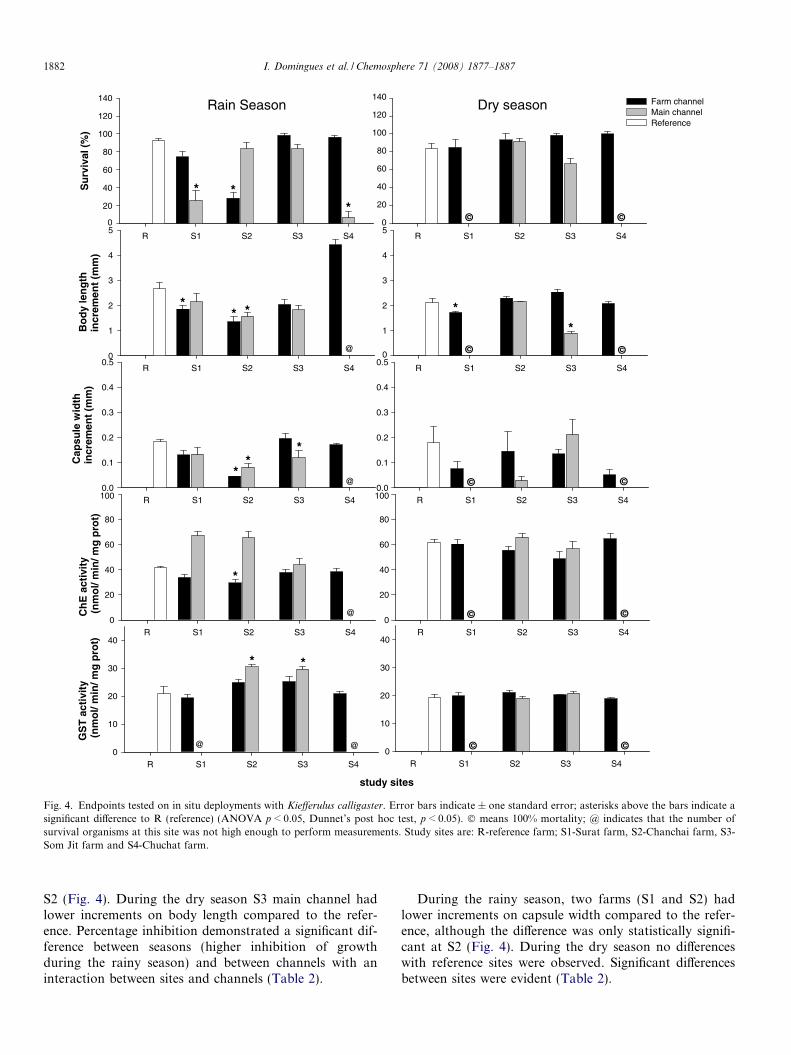
Fig. 4. Endpoints tested on in situ deployments with Kiefferulus calligaster. Error bars indicate ± one standard error; asterisks above the bars indicate asignificant difference to R (reference) (ANOVA p < 0.05, Dunnet’s post hoc test, p < 0.05). � means 100% mortality; @ indicates that the number ofsurvival organisms at this site was not high enough to perform measurements. Study sites are: R-reference farm; S1-Surat farm, S2-Chanchai farm, S3-Som Jit farm and S4-Chuchat farm.
1882 I. Domingues et al. / Chemosphere 71 (2008) 1877–1887
S2 (Fig. 4). During the dry season S3 main channel hadlower increments on body length compared to the refer-ence. Percentage inhibition demonstrated a significant dif-ference between seasons (higher inhibition of growthduring the rainy season) and between channels with aninteraction between sites and channels (Table 2).
During the rainy season, two farms (S1 and S2) hadlower increments on capsule width compared to the refer-ence, although the difference was only statistically signifi-cant at S2 (Fig. 4). During the dry season no differenceswith reference sites were observed. Significant differencesbetween sites were evident (Table 2).

Table 2Three-way ANOVA for endpoints measured in the field
Factors S BL CW ChE GST
Season F1,40 = 12.82; p = 0.001 F1,32 = 18.58; p = 0.000 F1,32 = 0.64; p = 0.428 F1,31 = 2.78; p = 0.105 F1,30 = 30.63; p = 0.000Site F1,40 = 6.2; p = 0.017 F1,32 = 0.000; p = 0.994 F1,32 = 11.37; p = 0.002 F1,31 = 3.72; p = 0.063 F1,30 = 0.01; p = 0.934Channel F1, 40 = 0.94; P = 0.338 F1,32 = 6.82; p = 0.014 F1,32 = 0.45; p = 0.505 F1,31 = 20.96; p = 0.000 F1,30 = 7.7; p = 0.009Site*channel F1,40 = 20.4; p = 0.000 F1,32 = 8.46; p = 0.007 F1,32 = 0.18; p = 0.675 F1,31 = 7.96; p = 0.008 F1,30 = 0.000; p = 0.975
Factors are season; site (farm) and type of channel (S-survival; BL-body length increment; CW-capsule width increment; ChE-cholinesterase activity;GST-Glutathione S-transferase activity).
I. Domingues et al. / Chemosphere 71 (2008) 1877–1887 1883
3.5. Biomarkers
In the rainy season two sites (S1 and S2) show an inhi-bition of cholinesterase activity on farm channels com-pared to the reference, although the difference was onlystatistically significant at S2 farm channel (Fig. 4). No dif-ferences compared to reference sites were evident duringthe dry season. Three-way ANOVA revealed differencesbetween channels but this is probably due to the abnor-mal values obtained for S1 and S2 main channels activities.An interaction between sites and channels was evident(Table 2).
In the rainy season two sites (S2 and S3) showed a slightincrease on GST activity compared to the reference but itwas only statistically significant in main channels. Duringthe dry season no differences were observed. Three-wayANOVA revealed differences between seasons with anincreased activity in the rainy season but not during thedry season.
4. Discussion
The bioassays performed proved to be suitable for thiskind of aquatic physical system although some further con-siderations are appropriate. Since deployments areperformed in farms with several kinds of activities co-occurring, precaution should be taken in order to not dis-turb farm activities like irrigation, harvesting, seeding orspraying. Moreover, attention should be given to watermanagement inside the farm that can lead to suddenchanges in the water level of test channels. Other con-straints relate to the structure of the water bodies such assteep margins and difficult access (in particular in the mainchannel).
4.1. Reference site
Differences were observed in the parameters measuredat the reference site between the two deployments (rainyseason and dry season). Survival levels were similarbetween seasons, while developmental endpoints (bodylength and head capsule width) were depressed duringthe dry season which may have been caused by a degra-dation of overall water quality associated with waterstagnation (water renewal did not occur). Evidence forthis is supported by RDA analysis which associated
low levels of dissolved oxygen with the reference site inthe dry season. Moreover, high chlorophyll a contentand low nitrite levels were associated with reference siteduring the rainy season presenting favourable conditionsfor organism growth. Nevertheless biochemical parame-ters (ChE and GST) presented values closer to laboratorylevels during the dry season and seemed to be alteredduring the rainy season. The chosen reference farm wasthe best candidate in the area since it is a closed systemwhere the inputs from outside are negligible although noguarantees can given regarding the total absence of exter-nal influences. In this way, the alteration of biomarkerslevels during the rain season can also be due to the run-off of chemicals not monitored. For instance, it is knownthat some heavy metals and detergents are susceptible ofinterfering with ChE activities (Guilhermino et al., 1998).The choice of a good reference site is crucial to resultsvalidation and unlike in situ testing in rivers where sitesupstream source of pollution are often suitable as refer-ence sites, reference sites in tropical agro-systems canbe more difficult to define because water bodies incorpo-rate a complex network of channels where pollutionsources can be difficult to identify. A sound approachfor future similar works would include the choice ofmore than one reference sites or the undertaking of pre-liminary work screening several possible reference sites toidentify the most suitable.
4.2. Survival
Between the four study sites, differences were observedfor several of the endpoints tested. Two farms (S1 andS4), independently of the season, presented either verylow levels of survival or no survival in main channelswhich could only be explained by other sources of con-tamination affecting these channels besides pesticide use.Factors such as the use of the water for domestic pur-poses, circulation of motorized boats and the potentialeffects of aquatic plants (water eutrophication) may causefurther alterations to water quality, potentially increasingobserved mortality. Survival levels were always higherthan 70% in farm channels except for S2 in the rainy sea-son. Unlike the main channels, no heavy impact wasobserved in these channels besides pesticide drift, sinceits use is restrained to watering the plots. Eventual spraydrift on farms do not seem to have a direct effect on thesurvival of test organisms.

1884 I. Domingues et al. / Chemosphere 71 (2008) 1877–1887
4.3. Developmental endpoints
During the rainy season, a depression on developmentalendpoints was observed for two farms although for bothparameters the difference was only significant at S2. Duringthe dry season no depression of these parameters wasobserved. Differences between seasons may be explainedby different agricultural practices since farmers applyhigher doses of pesticides during rainy season (MAMAS-project, 2003). Farmers have the perception that rain‘‘washes out” pesticides from soils and that higher dosesmust be applied to achieve the same effect on pest elimina-tion. This can eventually lead to an increase of pesticideresidues in water due to spray drift and run off.
Differences between farm channels and main channelswere expected since the former were located in the sprayingareas and consequently were more prone to receive spraydrift and accumulate residues while the main channels werelocated outside the farm and thus, outside the sprayedareas. The absence of a difference may be explained bythe dynamics of water management according to seasonalneeds; during the rainy season potentially contaminatedwater from farm channels might be pumped out into themain channels (to avoid overflow of farm channels)increasing their contamination. Moreover, potential effectsof pesticides on these main channels might be difficult todistinguish from contamination due to other use of thesewater bodies (see above). The lack of chemical analysisconstrains a more accurate interpretation of data andtherefore it should be a point to improve in future works.Although qualitative data regarding pesticide use is avail-able (amount and type of pesticides used, moment of spray-ing) no information is available concerning fate andmovements of pesticides in the water bodies which is essen-tial, in concert with the results obtained, to understand theoverall risk of pesticides use.
4.4. Biomarkers
The depression of ChE activity has been used as diag-nostic of carbamate or organophosphorous pesticides con-tamination. In our work, although it is known that OPpesticides were widely applied in the studied area, a slightdepression on ChE activity was only observed in S1 andS2 farm channels during the rainy season (only statisticallysignificant for S2), corresponding to the results for growthrate and head capsule increment. No inhibition of ChEactivity was observed in the main channels for any of theseasons suggesting that contamination of these channelsis probably not due to OP or carbamate pesticides. SinceChE activity in the reference site during the rainy seasonis slightly depressed it is first of all to consider that anunderestimation of risk can be occurring. Moreover, thehigh temperatures in tropical environments may, amongother factors, explain the lack of response observed in farmchannels. Some authors have suggested that elevated tem-perature might reduce toxicity of OP pesticides (Viswana-
than and Murti, 1989) and consequently reducinginhibition of cholinesterase. Moreover, temperature mayincrease ChE levels, rates of reactivation of inhibitedChE and general metabolic rates, affecting the toxicity ofsome cholinergic pollutants (Moulton et al., 1996). Fewin situ studies have investigated the cholinergic effects ofOP and carbamates pesticides in invertebrates. A clearresponse of ChE has been only obtained on events morelikely to constitute acute pulses of pesticide contamination,as is the case for the depression on ChE activity associatedwith the die-off of the mussel Elliptio complanata reportedby Fleming et al. (1995); the high AChE inhibition (55%)in the crayfish Procambarus clarkii after application of feni-trothion on Ebro Delta irrigation channels (Escartin andPorte, 1996) and the inhibition of AChE associated withsignificant levels of mortality in Gammarus pulex after theuse of malathion on watercress beds (Crane et al.,1995b). Many factors including differences in individualorganisms, age, diurnal and seasonal changes (Thompson,1999) which are impossible to control in a field situationmay contribute to a decrease of ChE sensitivity and repre-sent a constraint for its use as early warning signal. Never-theless, previous research has described in situ exposures inwhich ChE inhibition was successfully used as diagnostic ofpesticide contamination, e.g. Chironomus xanthus exposedto deltamethrin in mesocosms studies (Moreira-Santoset al., 2005).
In our study the low sensitivity of ChE may be attrib-uted to the two aspects: biotic and abiotic factors confer-ring high variability to the results and/or temperatureleading to a decrease in the toxicity of cholinergic pesti-cides. Moreover, possibly depressed ChE at reference sitecan lead to an underestimation of risk especially in therainy season.
GST activity increase has been associated with organicpollutants exposure in numerous insect species, however,according to Olsen et al. (2001) it remains unclear to whatextent GST induction can be expected in the chironomidChironomus riparius exposed in situ to xenobiotics. In ourwork with the chironomid K. calligaster, a slight increaseon GST activity on S2 and S3 sites during the rainy seasonwas observed. Absence of ChE inhibition in the main chan-nels of the same farms suggests that contamination at thesesites was not due to OP or carbamate pesticides but due toother environmental impacts previously discussed. Theincreased GST activity on farm channels on these samefarms might have been due to OP pesticide contamination(since a certain degree of ChE inhibition was also observed)but since GST is implied in non-specific detoxificationmechanisms conclusions are difficult to take.
The majority of published work on the in situ assessmentof organic pollutants have used mussels as test organismwith differing results. Some authors have demonstratedincreased GST activity as a tool for contaminant assess-ment (Kaaya et al., 1999; Gowland et al., 2002; Moreiraand Guilhermino, 2005) while others reported no increasein enzyme levels (Porte et al., 2000; Bebianno et al., 2004).

I. Domingues et al. / Chemosphere 71 (2008) 1877–1887 1885
Although no strong relationship was found betweenGST activity and any of the physicochemical parametersstudied, chironomid GST responds to some environmentalfactors such as temperature and dissolved oxygen (Choiet al., 2000; Callaghan et al., 2002). Moreover, some studieshave suggested that GST may not respond as rapidly asChE (Steevens and Benson, 1999) and the exposure in thefield may not have been long enough to elicit a response.All these factors may interfere with the interpretation ofthe results making a specific inference on the presenceand effects of pollutants difficult.
4.5. Sensitivity of endpoints
With the exception of site S2 during the rainy seasonwhere a general and consistent depression on the endpointsstudied (including survival) was observed, the responses ofthe selected endpoints in this study were weak and gener-ally not statistically significant. Moreover, biochemicalendpoints did not show higher sensitivity than develop-mental endpoints. This suggests that: (i) either the testorganisms and endpoints chosen in this work were not sen-sitive enough, or (ii) the studied aquatic environment wasless impacted than expected given the large amounts of pes-ticides applied in the area. The test species belongs to thechironomids family which are known to be ubiquitousand resistant organisms. This could be invoked as a reasonto question the sensitivity of this assay. However, using amore sensitive species (the freshwater prawn Macrobrachi-
um rosenbergii) similar results were obtained based on thepostexposure feeding rates of this prawn (Satapornvanit,2006). The endpoints studied represented a range of param-eters with different sensitivities. Survival is typically associ-ated with acute effects of contaminants; developmentalendpoints (growth and capsule width increment) are, incomparison, moderately sensitive while biochemicalparameters are, in theory, expected to be the most sensitive,responding to low levels of pollutants. The use of severalendpoints, spanning a wide range of sensitivities is gener-ally recommended since information provided by eachparameter presents complementary data. Nevertheless,the increase of the sensitivity of endpoints in relation totoxicants is likely to be coupled with an increase of theirsensitivity towards other biotic and abiotic factors makingit difficult to separate effects from contamination alone.
Tropical environments are characterized by high tem-peratures and humidity which affect the nature and courseof chemical reactions and the interactions between the bio-tic and abiotic components of the ecosystem (Murti, 1989).These represent critical factors for the fate of pesticides,leading to an accelerated process of pesticide degradation(Sethunathan, 1989), because under these conditions ratesof physicochemical and biological degradation are higher.Biological processes include higher rates of uptake, detox-ification and elimination, while physicochemical processesinclude higher volatilization and hydrolysis, both leadingto degradation of pesticides in a short period of time (Cas-
tillo et al., 1997; Henriques et al., 1997). Moreover hightemperatures may decrease the bioavailability of the con-taminants through indirect ways such as oxygen depletion,impact of nutrients and suspended solids. Agro-systemsstudied in this work were characterized by high levels ofsuspended solids due to water pumping in and out of thechannels. This may be especially important in explainingthe results obtained since pesticides attached to suspendedsolids may not be able to penetrate chironomid cages.Although it is difficult to draw specific conclusions toexplain the results obtained in this work, data on tropicaltoxicity support the idea that pesticide residues on aquaticenvironments are prone to a quick degradation after spay-ing and/or a quick drop in their bioavailability such thatthe bioavailable fraction may not be high enough to elicitresponses on the parameters studied. Further works shouldconsider the environmental fate of pesticides in these trop-ical aquatic systems to facilitate a greater understanding ofthe ecological effects of pesticides in the tropics.
5. Conclusion
The in situ bioassay developed in this study using K. cal-
ligaster permitted the detection of deleterious effects, dur-ing the rainy season, on some farm channels receivingspray drift from pesticides application. However, effectswere not always statistically significant. Main channelsappear to be affected by various impacts other than pesti-cides. All endpoints tested were shown to be suitable forthe bioassay and inclusion of biomarkers measurementsseemed to provide information related to the type of pollu-tants present in the aquatic environment. Further studiesshould assure technical means to quantify pesticides resi-dues in the water, since the absence of this data limits theinterpretation of the results.
This study demonstrated that the in situ bioassaydeveloped is a useful tool for pesticide monitoring andemphasises the need to adapt methodologies applied intemperate countries before their use in tropical climates.
Acknowledgments
The authors would like to thank Prof. P. Cranston whokindly identified the tropical chironomid species; Dr. Kier-an Monaghan for his comments and revision on this man-uscript; Alexandra Augusto for drawing the scheme of theThai farm and the MAMAS-project team for fieldassistance. This research was partly funded by the EC(MAMAS project, contract ICA4-2000-10247) and by adoctoral grant to Ines Domingues from FCT (Fundac�aopara a Ciencia e Tecnologia, Portugal).
References
Abdullah, A.R., Bajet, C.M., Matin, M.A., Nhan, D.D., Sulaiman, A.H.,1997. Ecotoxicology of pesticides in the tropical paddy field system.Environ. Toxicol. Chem. 16, 59–70.

1886 I. Domingues et al. / Chemosphere 71 (2008) 1877–1887
ASTM, 1980. Standard practice for conducting acute toxicity tests withfishes, macroinvertebrates and amphibians. E-729-80. American Stan-dard for Testing and Materials, Philadelphia, PA.
Bebianno, M.J., Geret, F., Hoarau, P., Serafim, M.A., Coelho, M.R.,Gnassia-Barelli, M., Romeo, M., 2004. Biomarkers in Ruditapes
decussatus: a potential bioindicator species. Biomarkers 9.Booth, L.H., Heppelthwaite, V., McGlinchy, A., 2000. The effect of
environmental parameters on growth, cholinesterase activity andglutathione S-transferase activity in the earthworm (Apporectodea
caliginosa). Biomarkers 5, 46–55.Bradford, M., 1976. A rapid and sensitive method for the quantification of
microgram quantities of protein utilizing the principle of proteindyebinding. Anal. Biochem. 72, 248–254.
Callaghan, A., Fisher, T.C., Grosso, A., Holloway, G.J., Crane, M., 2002.Effect of temperature and pirimiphos methyl on biochemical biomark-ers in Chironomus riparius meigen. Ecotoxicol. Environ. Safe. 52, 128–133.
Castillo, L.E., Cruz, E.d.l., Ruepert, C., 1997. Ecotoxicology andpesticides in tropical aquatic ecosystems of central america. Environ.Toxicol. Chem. 16, 41–51.
Castro, B.B., Guilhermino, L., Ribeiro, R., 2003. In situ bioassaychambers and procedures for assessment of sediment toxicity withChironomus riparius. Environ. Pollut. 125, 325–335.
Chang, C.-C., Lee, P.-P., Hsu, J.-P., Yeh, S.-P., Cheng, W., 2006.Survival, and biochemical, physiological, and histopathologicalresponses of the giant freshwater prawn, Macrobrachium rosenbergii,to short-term trichlorfon exposure. Aquaculture 253, 653–666.
Chappie, D.J., Burton, J.G.A., 1997. Optimization of in situ bioassayswith Hyalella azteca and Chironomus tentans. Environ. Toxicol. Chem.16, 559–564.
Choi, J., Roche, H., Caquet, T., 2000. Effects of physical (hypoxia,hyperoxia) and chemical (potassium dichromate, fenitrothion) stresson antioxidant enzyme activities in Chironomus riparius mg. (diptera,chironomidae) larvae: potential biomarkers. Environ. Toxicol. Chem.19, 495–500.
Clesceri, L.S., Greenberg, A.E., Trussel, R.R., 1989. Standard Methodsfor the Examination of Water and Wastewater. American PublicHealth Association, American Water Works Association, Washington.
Crane, M., Delaney, P., Mainstone, C., Clarke, S., 1995a. Measurementby in situ bioassay of water quality in an agricultural catchment. WaterRes. 29, 2441–2448.
Crane, M., Delaney, P., Watson, S., Parker, P., Walker, C., 1995b. Theeffect of malathion 60 on Gammarus pulex (L.) below watercress beds.Environ. Toxicol. Chem. 14, 1181–1188.
Crane, M., Higman, M., Olsen, T., Simpson, P., Callaghan, A.,Fisher, T., Kheir, R., 2000. An in situ system for exposing aquaticinvertebrates to contaminated sediments. Environ. Toxicol. Chem.19, 2715–2719.
Den Besten, P.J., Naber, A., Grootelaar, E.M.M., van de Guchte, C.,2003. In situ bioassays with Chironomus riparius: laboratory-fieldcomparisons of sediment toxicity and effects during wintering. Aquat.Ecosyst. Health Manage. 6, 217–228.
Ellman, G.L., Courtney, K.D., Andreas, V.J., Featherstone, R.M., 1961.A new and rapid colorimetric determination of acetylcholinesteraseactivity. Biochem. Pharmacol. 7, 88–95.
EPA, 2000. Methods for Measuring the Toxicity and Bioaccumulation ofSediment-Associated Contaminants with Freshwater Invertebrates.United States Environmental Protection Agency.
Escartin, E., Porte, C., 1996. Acetylcholinesterase inhibition in the crayfishProcambarus clarkii exposed to fenitrothion. Ecotoxicol. Environ.Safe. 34, 160–164.
Fleming, W.J., Augspurger, T.P., Alderman, J.A., 1995. Fresh-watermussel die-off attributed to anticholinesterase poisoning. Environ.Toxicol. Chem. 14, 877–879.
Forget, J., Beliaeff, B., Bocquene, G., 2003. Acetylcholinesterase activityin copepods (Tigriopus brevicornis) from the Vilaine river estuary,France, as a biomarker of neurotoxic contaminants. Aquat. Toxicol.62, 195–204.
Gowland, B.T.G., McIntosh, A.D., Davies, I.M., Moffat, C.F., Webster,L., 2002. Implications from a field study regarding the relationshipbetween polycyclic aromatic hydrocarbons and glutathione S-trans-ferase activity in mussels. Mar. Environ. Res. 54, 231–235.
Guilhermino, L., Lopes, M.C., Carvalho, A.P., Soares, A.M.V.M., 1996.Inhibition of acetylcholinesterase activity as effect criterion in acutetest with juvenile Daphnia magna. Chemosphere 32, 727–738.
Guilhermino, L., Barros, P., Silva, M.C., Soares, A.M.V.M., 1998. Shouldthe use of inhibition of cholinesterases as a specific biomarker fororganophosphate and carbamate pesticides be questioned? Biomarkers3, 157–163.
Habig, W.H., Pabst, M.J., Jakoby, W.B., 1974. Glutathione S-transfer-ases. J. Biol. Chem. 249, 7130–7137.
Henriques, W., Jeffers, R.D., Lacher, T.E., Kendall, R.J., 1997. Agro-chemical use on banana plantations in Latin America: perspectives onecological risk. Environ. Toxicol. Chem. 16, 91–99.
Kaaya, A., Najimi, S., Ribera, D., Narbonne, J.F., Moukrim, A., 1999.Characterization of glutathione S-transferases (GST) activities inPerna perna and Mytilus galloprovincialis used as a biomarker ofpollution in the Agadir marine bay (south of Morocco). Bull. Environ.Contam. Toxicol. 62, 623–629.
Kendall, R.J., Anderson, T.A., Baker, R.J., Bens, C.M., Carr, J.A.,Chiodo, L.A., Cobb III, G.P., Dickerson, R.L., Dixon, K.R., Fram,L.T., Hooper, M.J., Martin, C.F., McMurry, S.T., Patino, R., Smith,E.E., Theodorakis, C.W., 2001. Ecotoxicology. In: Klaassen, C.D.(Ed.), Casarett and Doull’s Toxicology: The Basic Science of Poisons.McGraw-Hill, pp. 1013–1045.
Lagadic, L., 2002. Biomarkers: useful tools for the monitoring of aquaticenvironments. Rev. Med. Vet. 153, 581–588.
MAMAS-project, 2003. State-of-the-System Workshop. Asian Institute ofTechnology, Thailand.
McLoughlin, N., Yin, D., Maltby, L., Wood, R.M., Yu, H., 2000.Evaluation of sensitivity and specificity of two crustacean biochemicalbiomarkers. Environ. Toxicol. Chem. 19, 2085–2092.
Moreira, S.M., Guilhermino, L., 2005. The Use of Mytilus Galloprovin-
cialis acetylcholinesterase and glutathione S-transferases activities asbiomarkers of environmental contamination along the northwestportuguese coast. Environ. Monit. Assess. 105, 309–325.
Moreira-Santos, M., Fonseca, A.L., Moreira, S.M., Osten, J.R.-v., Silva,E.M.d., Soares, A.M.V.M., Guilhermino, L., Ribeiro, R., 2005. Short-term sublethal (sediment and aquatic roots of floating macrophytes)assays with a tropical chironomid based on postexposure feeding andbiomarkers. Environ. Toxicol. Chem. 24, 2234–2242.
Moulton, C.A., Fleming, W.J., Purnell, C.E., 1996. Effects of twocholinesterase-inhibiting pesticides on freshwater mussels. Environ.Toxicol. Chem. 15, 131–137.
Murti, C.R.K., 1989. 3.4 chemicals in tropical and arid regions.Ecotoxicol. Climate, 97–136.
Olsen, T., Ellerbeck, L., Fisher, T., Callaghan, A., Crane, M., 2001.Variability in acetylcholinesterase and glutathione S-transferase activ-ities in Chironomus riparius meigen deployed in situ at uncontaminatedfield sites. Environ. Toxicol. Chem. 20, 1725–1732.
Peck, M.R., Klessa, D.A., Baird, D.J., 2002. A tropical sediment toxicitytest using the dipteran Chironomus crassiforceps to test metalbioavailability with sediment pH change in tropical acid-sulfatesediments. Environ. Toxicol. Chem. 21, 720–728.
Pery, A.R.R., Mons, R., Flammarion, P., Lagadic, L., Garric, J., 2002. Amodeling approach to link food availability, growth, emergence, andreproduction for the midge Chironomus riparius. Environ. Toxicol.Chem. 21, 2507–2513.
Porte, C., Biosca, X., Sole, M., Albaiges, J., 2000. The Aegean Sea oil spillon the Galician Coast (NW Spain). III: the assessment of long-termsublethal effects on mussels. Biomarkers 5, 436–446.
Radenac, G., Bocquene, G., Fichet, D., Miramand, P., 1998. Contami-nation of a dredged-material disposal site (La Rochelle Bay, France).The use of the acetylcholinesterase activity of Mytilus edulis (L.) as abiomarker of pesticides: the need for a critical approach. Biomarkers 3,305–315.

I. Domingues et al. / Chemosphere 71 (2008) 1877–1887 1887
Robillard, S., Beauchamp, G., Laulier, M., 2003. The role of abioticfactors and pesticide levels on enzymatic activity in the freshwatermussel Anodonta cygnea at three different exposure sites. Comp.Biochem. Phys. C 135, 49–59.
Sanchez-Hernandez, J.C., Walker, C.H., 2000. In vitro and in vivocholinesterase inhibition in lacertides by phosphonate- and phospho-rothioate-type organophosphates. Pestic. Biochem. Phys. 67, 1–12.
Satapornvanit, K., 2006. Feeding Behaviour of the Prawn Macrobrachium
Rosenbergii as an Indicator of Pesticide Contamination in TropicalFreshwater. University of Stirling.
Satapornvanit, K., Baird, D.J., Little, D.C., Milwain, G.K., Brink,P.J.V.D., Beltman, W.H.J., Nogueira, A.J.A., Daam, M.A., Domin-gues, I., Kodithuwakku, S.S., Perera, W., Yakupitiyage, A., Sures-hkumar, S.N., Taylor, M.G., 2004. Risks of pesticide use in aquaticecosystems adjacent to mixed vegetable and monocrop fruit growingareas in Thailand. Aust. J. Ecotoxicol. 10, 85–95.
Sethunathan, N., 1989. 5.2 Biodegradation of pesticides in tropical riceecosystems. Ecotoxicol. Climate, 247–264.
Sibley, P.K., Benoit, D.A., Balcer, M.D., Phipps, G.L., West, C.W., Hoke,R.A., Ankley, G.T., 1999. In situ bioassay chamber for assessment ofsediment toxicity and bioaccumulation using benthic invertebrates.Environ. Toxicol. Chem. 18, 2325–2336.
Soares, S., Cativa, I., Moreira-Santos, M., Soares, A., Ribeiro, R., 2005a.A Short-term sublethal in situ sediment assay with Chironomus riparius
based on postexposure feeding. Arch. Environ. Contam. Toxicol. 49,163–172.
Soares, S., Moreira, S., Andrade, S., Guilhermino, L., Ribeiro, R., 2005b.Freshwater-saltwater interface and estuarine sediment in situ assays
based on postexposure feeding of chironomids and polychaetes.Estuaries 28, 314–319.
SPSS, 2004. Sigma Stat for Windows (version 3.10). SPSS Inc., Chicago.Steevens, J.A., Benson, W.H., 1999. Toxicological interactions of chlor-
pyrifos and methyl mercury in the amphipod, Hyalella azteca. Toxicol.Sci. 52, 168–177.
Sturm, A., Wogram, J., Hansen, P.D., Liess, M., 1999. Potential use ofcholinesterase in monitoring low levels of organophosphates in smallstreams: natural variability in three-spined stickleback (Gasterosteus
aculeatus) and relation to pollution. Environ. Toxicol. Chem. 18, 194–200.
Ter Braak, C.J.F., Smilauer, P., 2002. CANOCO Reference Manual andCanoDraw for Windows User’s Guide: Software for CanonicalCommunity Ordination (Version 4.5). Microcomputer Power, Ithaca,USA.
Thompson, H.M., 1999. Esterases as markers of exposure to organo-phosphates and carbamates. Ecotoxicology 8, 369–384.
Tucker, K.A., Burton, G.A., 1999. Assessment of non-point-source runoffin a stream using in situ and laboratory approaches. Environ. Toxicol.18, 2797–2803.
Van den Brink, P.J., Sureshkumar, S.N., Daam, M.A., Domingues, I.,Milwain, G.K., Beltman, W.H.J., Perera, W., Satapornvanit, K., 2003.Environmental and human risks of pesticide use in Thailand and SriLanka. Results of a preliminar risk assessment. In: Alterra Report,789, A.r. (Ed.).
Viswanathan, P.N., Murti, C.R.K., 1989. 4.1 Effects of temperature andhumidity on ecotoxicology of chemicals. Ecotoxicol. Climate,139–154.




















