
Efficient Identification of miRNAs for Classification of Tumor Origin

Experimental Gerontology 62 (2015) 7–13
Contents lists available at ScienceDirect
Experimental Gerontology
j ourna l homepage: www.e lsev ie r .com/ locate /expgero
Improved knee extensor strengthwith resistance training associateswithmuscle specific miRNAs in older adults
Tan Zhang a,b, Alexander Birbrair a, Zhong-Min Wang a, María L. Messi a, Anthony P. Marsh d, Iris Leng c,Barbara J. Nicklas a,b, Osvaldo Delbono a,b,⁎a Department of Internal Medicine, Section on Gerontology and Geriatric Medicine, Wake Forest School of Medicine, Winston-Salem, NC 27157, United Statesb J Paul Sticht Center on Aging, Wake Forest School of Medicine, Winston-Salem, NC 27157, United Statesc Department of Biostatistical Sciences, Wake Forest School of Medicine, Winston-Salem, NC 27157, United Statesd Department of Health and Exercise Science, Wake Forest University, Winston-Salem, NC 27109, United States
⁎ Corresponding author at: Department of InternalMedSchool of Medicine, Medical Center Boulevard, Winston-S
E-mail address: [email protected] (O. Delbon
http://dx.doi.org/10.1016/j.exger.2014.12.0140531-5565/© 2015 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 September 2014Received in revised form 22 December 2014Accepted 29 December 2014Available online 2 January 2015
Section Editor: Christiaan Leeuwenburgh
Keywords:MicroRNASkeletal muscleAgingResistance training
Regular exercise, particularly resistance training (RT), is the only therapy known to consistently improvemusclestrength and quality (force per unit of mass) in older persons, but there is considerable variability in responsive-ness to training. Identifying sensitive diagnostic biomarkers of responsiveness to RT may inform the design of amore efficient exercise regimen to improvemuscle strength in older adults. MicroRNAs (miRNAs) are small non-codingRNAs that regulate gene expression.Wequantified sixmuscle specificmiRNAs (miR-1, -133a, -133b, -206,-208b and -499) in both muscle tissue and blood plasma, and their relationship with knee extensor strength inseven older (age = 70.5 ± 2.5 years) adults before and after 5 months of RT. MiRNAs differentially respondedto RT; muscle miR-133b decreased, while all plasmamiRNAs tended to increase. Percent changes in knee exten-sor strength with RT showed strong positive correlations with percent changes in muscle miR-133a, -133b, and-206 and with percent changes in plasma and plasma/muscle miR-499 ratio. Baseline level of plasma or plasma/musclemiR-499 ratio further predictsmuscle response to RT,while changes inmusclemiR-133a, -133b, and -206may correlate with muscle TNNT1 gene alternative splicing in response to RT. Our results indicate that RT altersmuscle specific miRNAs in muscle and plasma, and that these changes account for some of the variation instrength responses to RT in older adults.
© 2015 Elsevier Inc. All rights reserved.
1. Introduction
Mammalian species, and humans in particular, undergo a progres-sive and gradual loss of skeletal muscle mass and strength with aging,which impairs performance of daily living activities and leads to majormedical and socioeconomic problems. Age-related decreases in musclemass only partially account for muscular weakening with age(Delbono, 2003, 2011; Goodpaster et al., 2006), and improvements inmuscle strengthwith resistance training (RT) are not entirely accountedfor by muscle hypertrophy with RT (Fiatarone et al., 1990; Leenderset al., 2013; Tracy et al., 1999). This suggests that strength is regulatedby cellular and molecular mechanisms that are independent of musclesize; yet these mechanisms have yet to be entirely elucidated. In addi-tion, there is considerable variability of strength responses to RT thatcan inform the discovery of these mechanisms. Recently, we reportedincreased specific force and power in single muscle fibers, but notmyofiber cross-sectional area, in the vastus lateralis muscle of older
icine, Gerontology,Wake Forestalem, NC 27157, United States.o).
adults subjected to a 5-month RT program (Zhang et al., 2014). Thisstudy indicated that RT altered the Troponin T1 (TNNT1) pre-mRNA al-ternative splicing (AS) profile in muscle and that muscle strength afterRT was related to these gene profiles. The next step is to determinethe signals induced by RT to trigger the alternative splicing of TNNT1.
MiRNAs, small (~20–30 nucleotides) noncoding RNAs that inhibitprotein translation or enhance mRNA degradation, have been implicat-ed in the response of various organs to a single bout of RT or endurancetraining (Davidsen et al., 2011; Drummond et al., 2008; Hamilton andBaulcombe, 1999; Reinhart et al., 2000; Tonevitsky et al., 2013;Zacharewicz et al., 2013). Muscle-specific miRNAs (myomiRs) are agroup of striated muscle (skeletal and/or cardiac) miRNAs, includingmiR-1, miR-133a, miR-133b, miR-206, miR-208, miR-208b, miR-486and miR-499 (Callis et al., 2008; McCarthy and Esser, 2007; Smallet al., 2010; van Rooij et al., 2008, 2009). Among these myomiRs, miR-1, miR-133a/b, and miR-206, make up nearly 25% of miRNA expressionin skeletal muscle in both humans and mice (McCarthy, 2008; Sempereet al., 2004). Epigenetic control of muscle performance in response tooverload, muscle innervation, specification of muscle fiber identity,and myoblast proliferation in response to tissue injury, are regulatedby specific miRNAs (Dey et al., 2011; Moresi et al., 2010; Nakasa et al.,

8 T. Zhang et al. / Experimental Gerontology 62 (2015) 7–13
2010; Small et al., 2010; van Rooij et al., 2009; Williams et al., 2009).Some muscle-enriched miRNAs, such as miR-1, miR-133a, miR-206and miR-499, can be detected in blood and respond to pathologies inhumans and animals (Deng et al., 2011; Donaldson et al., 2013;Eisenberg et al., 2007;McCarthy et al., 2007;Mizuno et al., 2011). As cir-culating miRNAs are stable (e.g., resistant to endogenous RNAses)(Mitchell et al., 2008), they are considered useful biomarkers(Alevizos and Illei, 2010) for disease incidence and prognosis. Interest-ingly, circulating miRNAs may be taken up by non-muscle cells andthus regulate protein expression, contributing to the pathogenesis ofdisease (Heneghan et al., 2010; Mitchell et al., 2008; Wang et al.,2009, 2010). Thus, miRNAs are promising diagnostic and therapeutictargets (Hoy and Buck, 2012; Uhlemann et al., 2014; Valadi et al.,2007; Weilner et al., 2013).
Only a few studies have analyzedwhether resistance exercise has aneffect onmiRNA levels in humans. A study in healthy youngmen under-taking a five day per week RT program for 12 weeks showed thatmiRNAs play a role in regulating muscle mass response to RT(Davidsen et al., 2011). Additionally, compared to young subjects,older subjects showed smaller decreases in miR-1 expression after anacute bout of resistance exercise (Drummond et al., 2008). However,whether chronic RT alters miRNAs in older adults and whether this ef-fect potentially contributes to the magnitude of strength gain with RTis not known. Therefore, the purpose of this study was to measure sixmuscle specific miRNAs (miR-1, -133a, -133b, -206, -208b and -499)in muscle tissue and blood plasma in older adults before and after fivemonths of RT and to examine the relationship between changes inknee extensor strength with changes in miRNAs or miRNAs at baselinelevel.
2. Methods and materials
2.1. Study participants
Participants in this study included older men (n = 3) and women(n = 4) who were recruited from the Piedmont Triad area of NorthCarolina. All participants were age 65–80 years, non-smoking, not onhormone replacement therapy, sedentary (b15 min of exercise,2 times/week) and weight stable (b5% weight change) for at least sixmonths prior to enrollment. All had normal liver, kidney, pulmonary,and thyroid function; no history of excessive alcohol intake; and nomajor chronic illness, anemia, or orthopedic impairment as described(Zhang et al., 2014). The study was approved by the Wake Forest Insti-tutional Review Board for Human Research and all participants signedinformed consent to participate in the study.
2.2. Resistance training intervention
The exercise intervention consisted of five months of progressive RTdesigned to elicit adaptations in skeletal muscle and increase strengthand power. To optimize the resistance stimulus for maximal functionalgain, the exercise prescription was based on a relative intensity leveland progressed with each individual's strength gain. Participantsexercised three days/week under the supervision of two exercise phys-iologists. Participants walked or cycled slowly for 5–10min to warm-upprior to RT. The resistance exercises were performed on machines(Cybex, Medway, MA and Nautilus, Vancouver, WA), on which theload could be adjusted in small increments. The participants completedfour lower extremity exercises, two of which targeted the quadricepsmuscles (leg press, knee extension), one that targeted the hamstrings(leg curl), and one that targeted the triceps surae (calf press). The pro-gram was based on American College of Sports Medicine guidelinesfor intensity, number of repetitions, number of sets, and number ofdays per week (American College of Sports Medicine, 2009). The onerepetition maximum (1RM), that is, the maximal weight that could belifted with correct form in a single repetition was used to prescribe
intensity. Intensity was progressively increased during the first monthtowards the training goal of three sets of 10 repetitions at 70% 1RMfor a given exercise. The 1RM strength testing was repeated every fourweeks, and the training loads adjusted to achieve the 70% 1RM goal.
2.3. Knee extensor strength
We assessed maximal isokinetic knee extensor strength using anisokinetic dynamometer (Biodex) at a fixed speed (60° per second)with the participant sitting and the hips and knee flexed at 90°. Thismethod is widely considered the gold standard of strength assessment(Martin et al., 2006). The 1-week test–retest reliability in our lab isvery high (intra-class correlation coefficient N 0.97). We set start andstop angles at 90° and 30° of knee flexion (range of motion of 60°).We used the peak torque observed during this range of motion as themeasure of strength for each knee extension movement.
2.4. Body composition measurements
Height andmassweremeasured to calculate BMI (kg/m2). All partic-ipants underwent a total-body scan using dual-energy X-ray absorpti-ometry (DXA, Model DPX-L, Lunar Corp., Madison, Wisconsin) todetermine lean muscle mass and percent body fat before and after RT.
2.5. Skeletal muscle biopsy and blood collection
Needle biopsies of the vastus lateralis were collected under localanesthesia with 1% lidocaine. All biopsies were performed in the earlymorning after an overnight fast. Subjects were asked to refrain fromtaking aspirin, prescription and over-the-counter non-steroidal anti-inflammatory drugs, or other compounds that may affect bleeding,platelets, or bruising for the week prior to the biopsy, and to refrainfrom any strenuous activity (including RT) for at least 36 h prior tothe biopsy. Visible blood and connective tissue were removed fromthemuscle specimen. Amuscle portion (30mg) used for RNAextractionwas snap-frozen in liquid nitrogen under RNAse free conditions, andstored at −80 °C until analysis. Participants completed their last boutof RT 36–72 h prior to the post-RT biopsies and blood sampling.Whole blood from participants was collected in sodium EDTA tubes be-fore and after RT and before muscle collection. Centrifugation was thenperformed at 4 °C with an initial spin at 2000 g for 10 min to separatethe plasma from the buffy coat and red blood cells and was stored at−80 °C until analysis. Plasma from two of the subjectswas not collectedbecause they did not provide consent to store their blood samples.
2.6. RNA isolation and quantitative real-time RT-PCR for miRNA detection
The miRNAs miR-1, miR-133a, miR-133b, miR-206, miR-208b, andmiR-499 are enriched in muscles, but typically found at low levels inother tissues (McCarthy, 2011). We examined the levels of muscle-specific miRNAs in both muscle and blood plasma using real-time RT-PCR. Total RNAwas extracted fromhuman vastus lateralismuscle tissuesusing themirVana™miRNA Isolation Kit (Applied Biosystems, Carlsbad,CA) or from 200 μl plasma using the phenol-free total RNA purificationkit (Amresco, Solon, OH) according to the manufacturer's instructions.RNA samples and the same volume of water (negative control) werereverse-transcribed using TaqMan® MicroRNA Reverse TranscriptionKit (Applied Biosystems). The TaqManmiRNA assaywas used accordingto the manufacturer's instructions to measure the expression of humanmiR-1, miR-133a, miR-133b, miR-206, miR-208b, and miR-499 usingpre-developed reagents from ABI (Applied Biosystems) run on anMx3000 (Stratagene, La Jolla, CA). The U6 small nuclear RNA (RNU6B),an endogenous control, was used to normalize blood plasma and skele-tal muscle miRNAs expression (Asaga et al., 2011; Ng et al., 2009; Songet al., 2012; Togliatto et al., 2013). Analysis was performed by the com-parative threshold cycle (Ct) method under the following cycling
9T. Zhang et al. / Experimental Gerontology 62 (2015) 7–13
conditions: 10min at 95 °C; 65 cycles of 15 s at 95 °C and 1min at 60 °C.Relative abundance of each miRNA was determined from the Ct valuesusing the 2−ΔΔCtmethod (Livak and Schmittgen, 2001) after normaliza-tion to RNU6B and all experiments were performed in triplicate. As thecut-off value for low-copy (b250 copies) circulation miRNAs is approx-imately 35 PCR cycles (Mestdagh et al., 2009), and plasma miR-208b'sCt value was more than 50 in all subjects, it was considered non-detectable or unreliable, and not reported further.
2.7. Statistical analysis
Data analysis was performed with SAS 9.4 (Cary, NC). Percentchange ((post-measure − pre-measure) / pre-measure × 100%) wascalculated for the outcomes of knee strength, body composition (BMI,%fat and lean mass) and miRNAs to take into account individual differ-ences at baseline. A single group t-test was used to determine interven-tion effect on each variable. Then we examined the age and gendereffect on the percent change of knee strength, body composition andmiRNAs after RT. We also used mixed effect models to control for ageand gender, where pre- and post-measures were treated as repeatedmeasures. Compound symmetry covariance structure was used to ac-count for within subject correlation. Pearson Product correlationswere calculated to determine the strength of the association betweenpairs of variables. For all tests, statistical significant level was set at0.05. In acknowledgement of our small sample size,we also report effectsizes (ES) of RT effect calculated with Cohen's D, where ES = (post-measure − pre-measure) / pooled standard deviation of pre-measureand post-measure.
3. Results
We noticed that the mixed effect model results did not differ mate-rially from the single group t-test results, sowe only report the t-test re-sults. It appeared that age did not affect percent change of kneestrength, body composition, or miRNAs, while gender only showed a
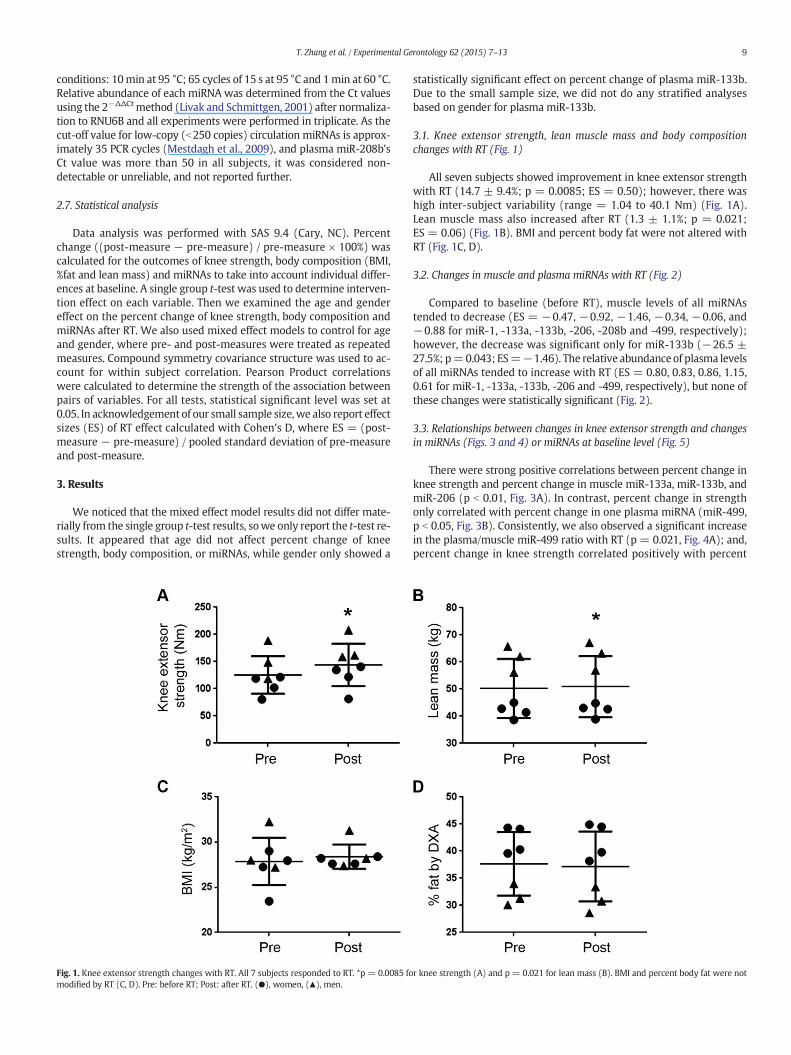
Fig. 1. Knee extensor strength changes with RT. All 7 subjects responded to RT. *p = 0.0085 fomodified by RT (C, D). Pre: before RT; Post: after RT. (●), women, (▲), men.
statistically significant effect on percent change of plasma miR-133b.Due to the small sample size, we did not do any stratified analysesbased on gender for plasma miR-133b.
3.1. Knee extensor strength, lean muscle mass and body compositionchanges with RT (Fig. 1)
All seven subjects showed improvement in knee extensor strengthwith RT (14.7 ± 9.4%; p = 0.0085; ES = 0.50); however, there washigh inter-subject variability (range = 1.04 to 40.1 Nm) (Fig. 1A).Lean muscle mass also increased after RT (1.3 ± 1.1%; p = 0.021;ES = 0.06) (Fig. 1B). BMI and percent body fat were not altered withRT (Fig. 1C, D).
3.2. Changes in muscle and plasma miRNAs with RT (Fig. 2)
Compared to baseline (before RT), muscle levels of all miRNAstended to decrease (ES = −0.47, −0.92, −1.46, −0.34, −0.06, and−0.88 for miR-1, -133a, -133b, -206, -208b and -499, respectively);however, the decrease was significant only for miR-133b (−26.5 ±27.5%; p=0.043; ES=−1.46). The relative abundance of plasma levelsof all miRNAs tended to increase with RT (ES = 0.80, 0.83, 0.86, 1.15,0.61 for miR-1, -133a, -133b, -206 and -499, respectively), but none ofthese changes were statistically significant (Fig. 2).
3.3. Relationships between changes in knee extensor strength and changesin miRNAs (Figs. 3 and 4) or miRNAs at baseline level (Fig. 5)
There were strong positive correlations between percent change inknee strength and percent change in muscle miR-133a, miR-133b, andmiR-206 (p b 0.01, Fig. 3A). In contrast, percent change in strengthonly correlated with percent change in one plasma miRNA (miR-499,p b 0.05, Fig. 3B). Consistently, we also observed a significant increasein the plasma/muscle miR-499 ratio with RT (p = 0.021, Fig. 4A); and,percent change in knee strength correlated positively with percent
r knee strength (A) and p = 0.021 for lean mass (B). BMI and percent body fat were not
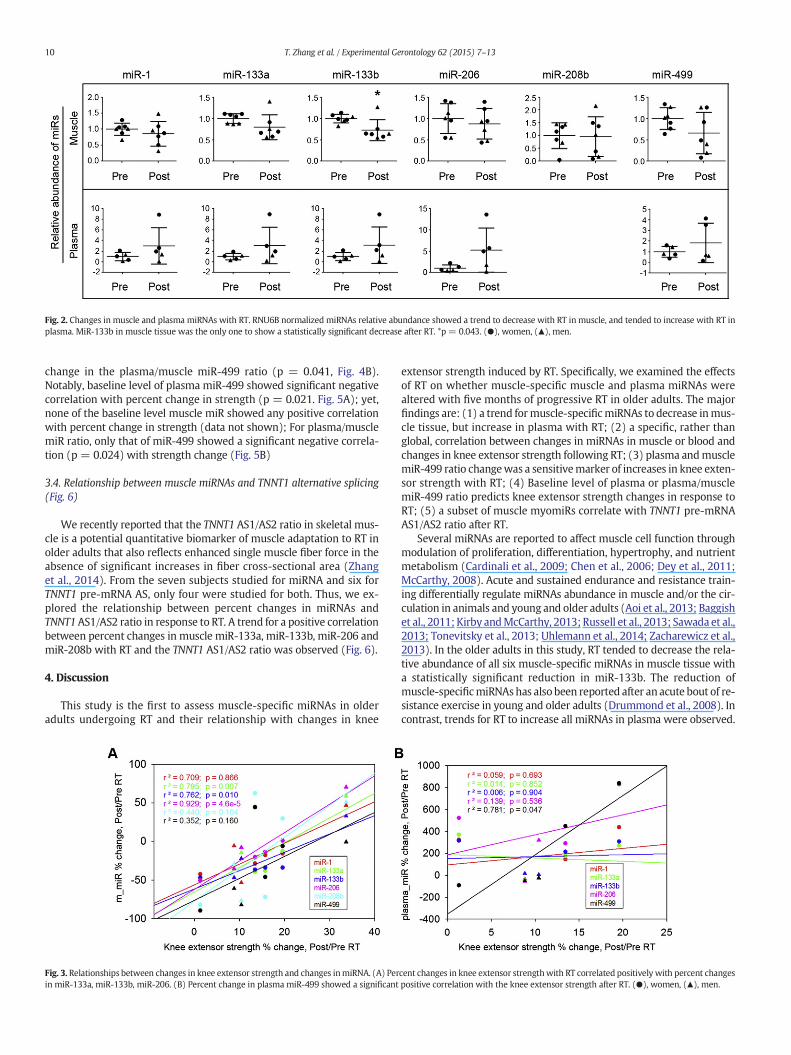
Fig. 2. Changes in muscle and plasma miRNAs with RT. RNU6B normalized miRNAs relative abundance showed a trend to decrease with RT in muscle, and tended to increase with RT inplasma. MiR-133b in muscle tissue was the only one to show a statistically significant decrease after RT. *p = 0.043. (●), women, (▲), men.
10 T. Zhang et al. / Experimental Gerontology 62 (2015) 7–13
change in the plasma/muscle miR-499 ratio (p = 0.041, Fig. 4B).Notably, baseline level of plasma miR-499 showed significant negativecorrelation with percent change in strength (p = 0.021. Fig. 5A); yet,none of the baseline level muscle miR showed any positive correlationwith percent change in strength (data not shown); For plasma/musclemiR ratio, only that of miR-499 showed a significant negative correla-tion (p = 0.024) with strength change (Fig. 5B)
3.4. Relationship between muscle miRNAs and TNNT1 alternative splicing(Fig. 6)
We recently reported that the TNNT1 AS1/AS2 ratio in skeletal mus-cle is a potential quantitative biomarker of muscle adaptation to RT inolder adults that also reflects enhanced single muscle fiber force in theabsence of significant increases in fiber cross-sectional area (Zhanget al., 2014). From the seven subjects studied for miRNA and six forTNNT1 pre-mRNA AS, only four were studied for both. Thus, we ex-plored the relationship between percent changes in miRNAs andTNNT1AS1/AS2 ratio in response to RT. A trend for a positive correlationbetween percent changes in muscle miR-133a, miR-133b, miR-206 andmiR-208b with RT and the TNNT1 AS1/AS2 ratio was observed (Fig. 6).
4. Discussion
This study is the first to assess muscle-specific miRNAs in olderadults undergoing RT and their relationship with changes in knee
Fig. 3. Relationships between changes in knee extensor strength and changes inmiRNA. (A) Perin miR-133a, miR-133b, miR-206. (B) Percent change in plasma miR-499 showed a significant
extensor strength induced by RT. Specifically, we examined the effectsof RT on whether muscle-specific muscle and plasma miRNAs werealtered with five months of progressive RT in older adults. The majorfindings are: (1) a trend formuscle-specificmiRNAs to decrease inmus-cle tissue, but increase in plasma with RT; (2) a specific, rather thanglobal, correlation between changes in miRNAs in muscle or blood andchanges in knee extensor strength following RT; (3) plasma andmusclemiR-499 ratio changewas a sensitivemarker of increases in knee exten-sor strength with RT; (4) Baseline level of plasma or plasma/musclemiR-499 ratio predicts knee extensor strength changes in response toRT; (5) a subset of muscle myomiRs correlate with TNNT1 pre-mRNAAS1/AS2 ratio after RT.
Several miRNAs are reported to affect muscle cell function throughmodulation of proliferation, differentiation, hypertrophy, and nutrientmetabolism (Cardinali et al., 2009; Chen et al., 2006; Dey et al., 2011;McCarthy, 2008). Acute and sustained endurance and resistance train-ing differentially regulate miRNAs abundance in muscle and/or the cir-culation in animals and young and older adults (Aoi et al., 2013; Baggishet al., 2011; Kirby andMcCarthy, 2013; Russell et al., 2013; Sawada et al.,2013; Tonevitsky et al., 2013; Uhlemann et al., 2014; Zacharewicz et al.,2013). In the older adults in this study, RT tended to decrease the rela-tive abundance of all six muscle-specific miRNAs in muscle tissue witha statistically significant reduction in miR-133b. The reduction ofmuscle-specificmiRNAshas also been reported after an acute bout of re-sistance exercise in young and older adults (Drummond et al., 2008). Incontrast, trends for RT to increase all miRNAs in plasma were observed.
cent changes in knee extensor strengthwith RT correlated positively with percent changespositive correlation with the knee extensor strength after RT. (●), women, (▲), men.
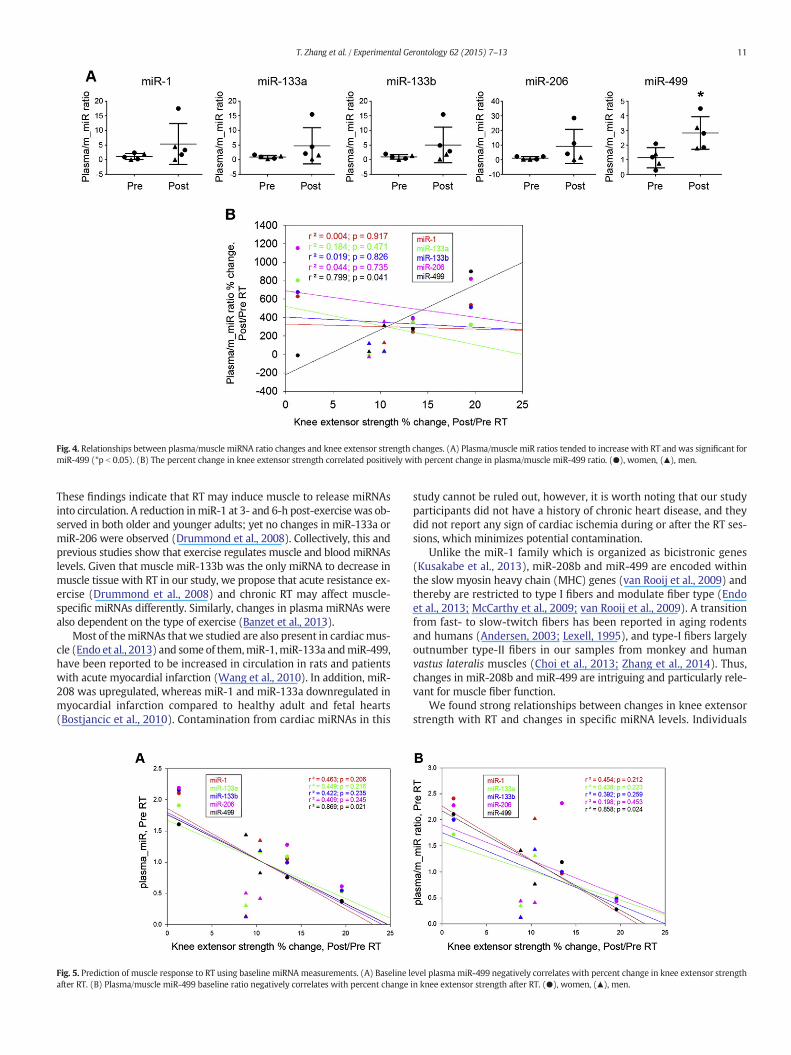
Fig. 4. Relationships between plasma/muscle miRNA ratio changes and knee extensor strength changes. (A) Plasma/muscle miR ratios tended to increase with RT and was significant formiR-499 (*p b 0.05). (B) The percent change in knee extensor strength correlated positively with percent change in plasma/muscle miR-499 ratio. (●), women, (▲), men.
11T. Zhang et al. / Experimental Gerontology 62 (2015) 7–13
These findings indicate that RT may induce muscle to release miRNAsinto circulation. A reduction inmiR-1 at 3- and 6-h post-exercise was ob-served in both older and younger adults; yet no changes in miR-133a ormiR-206 were observed (Drummond et al., 2008). Collectively, this andprevious studies show that exercise regulates muscle and blood miRNAslevels. Given that muscle miR-133b was the only miRNA to decrease inmuscle tissue with RT in our study, we propose that acute resistance ex-ercise (Drummond et al., 2008) and chronic RT may affect muscle-specific miRNAs differently. Similarly, changes in plasma miRNAs werealso dependent on the type of exercise (Banzet et al., 2013).
Most of themiRNAs thatwe studied are also present in cardiacmus-cle (Endo et al., 2013) and some of them,miR-1,miR-133a andmiR-499,have been reported to be increased in circulation in rats and patientswith acute myocardial infarction (Wang et al., 2010). In addition, miR-208 was upregulated, whereas miR-1 and miR-133a downregulated inmyocardial infarction compared to healthy adult and fetal hearts(Bostjancic et al., 2010). Contamination from cardiac miRNAs in this
Fig. 5. Prediction of muscle response to RT using baseline miRNA measurements. (A) Baseline lafter RT. (B) Plasma/muscle miR-499 baseline ratio negatively correlates with percent change
study cannot be ruled out, however, it is worth noting that our studyparticipants did not have a history of chronic heart disease, and theydid not report any sign of cardiac ischemia during or after the RT ses-sions, which minimizes potential contamination.
Unlike the miR-1 family which is organized as bicistronic genes(Kusakabe et al., 2013), miR-208b and miR-499 are encoded withinthe slow myosin heavy chain (MHC) genes (van Rooij et al., 2009) andthereby are restricted to type I fibers and modulate fiber type (Endoet al., 2013; McCarthy et al., 2009; van Rooij et al., 2009). A transitionfrom fast- to slow-twitch fibers has been reported in aging rodentsand humans (Andersen, 2003; Lexell, 1995), and type-I fibers largelyoutnumber type-II fibers in our samples from monkey and humanvastus lateralis muscles (Choi et al., 2013; Zhang et al., 2014). Thus,changes in miR-208b and miR-499 are intriguing and particularly rele-vant for muscle fiber function.
We found strong relationships between changes in knee extensorstrength with RT and changes in specific miRNA levels. Individuals
evel plasmamiR-499 negatively correlates with percent change in knee extensor strengthin knee extensor strength after RT. (●), women, (▲), men.
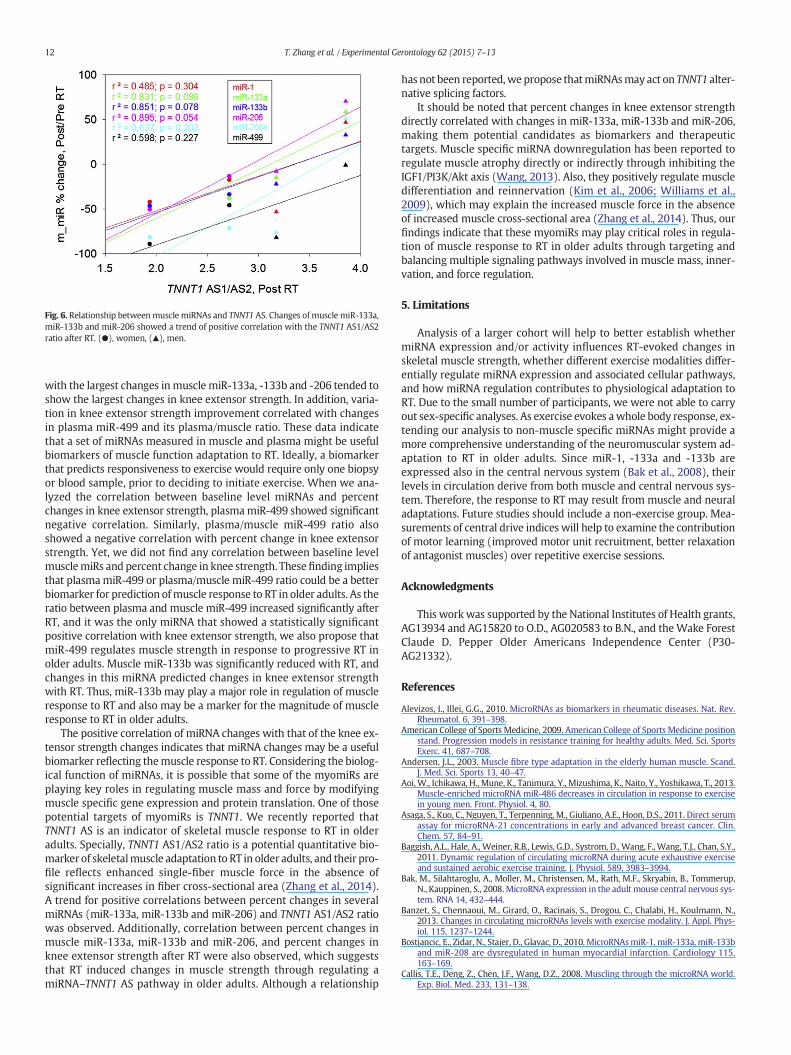
Fig. 6. Relationship betweenmuscle miRNAs and TNNT1 AS. Changes of muscle miR-133a,miR-133b and miR-206 showed a trend of positive correlation with the TNNT1 AS1/AS2ratio after RT. (●), women, (▲), men.
12 T. Zhang et al. / Experimental Gerontology 62 (2015) 7–13
with the largest changes inmuscle miR-133a, -133b and -206 tended toshow the largest changes in knee extensor strength. In addition, varia-tion in knee extensor strength improvement correlated with changesin plasma miR-499 and its plasma/muscle ratio. These data indicatethat a set of miRNAs measured in muscle and plasma might be usefulbiomarkers of muscle function adaptation to RT. Ideally, a biomarkerthat predicts responsiveness to exercise would require only one biopsyor blood sample, prior to deciding to initiate exercise. When we ana-lyzed the correlation between baseline level miRNAs and percentchanges in knee extensor strength, plasmamiR-499 showed significantnegative correlation. Similarly, plasma/muscle miR-499 ratio alsoshowed a negative correlation with percent change in knee extensorstrength. Yet, we did not find any correlation between baseline levelmusclemiRs and percent change in knee strength. Thesefinding impliesthat plasmamiR-499 or plasma/muscle miR-499 ratio could be a betterbiomarker for prediction ofmuscle response to RT in older adults. As theratio between plasma and muscle miR-499 increased significantly afterRT, and it was the only miRNA that showed a statistically significantpositive correlation with knee extensor strength, we also propose thatmiR-499 regulates muscle strength in response to progressive RT inolder adults. Muscle miR-133b was significantly reduced with RT, andchanges in this miRNA predicted changes in knee extensor strengthwith RT. Thus, miR-133b may play a major role in regulation of muscleresponse to RT and also may be a marker for the magnitude of muscleresponse to RT in older adults.
The positive correlation of miRNA changes with that of the knee ex-tensor strength changes indicates that miRNA changes may be a usefulbiomarker reflecting themuscle response to RT. Considering the biolog-ical function of miRNAs, it is possible that some of the myomiRs areplaying key roles in regulating muscle mass and force by modifyingmuscle specific gene expression and protein translation. One of thosepotential targets of myomiRs is TNNT1. We recently reported thatTNNT1 AS is an indicator of skeletal muscle response to RT in olderadults. Specially, TNNT1 AS1/AS2 ratio is a potential quantitative bio-marker of skeletalmuscle adaptation to RT in older adults, and their pro-file reflects enhanced single-fiber muscle force in the absence ofsignificant increases in fiber cross-sectional area (Zhang et al., 2014).A trend for positive correlations between percent changes in severalmiRNAs (miR-133a, miR-133b and miR-206) and TNNT1 AS1/AS2 ratiowas observed. Additionally, correlation between percent changes inmuscle miR-133a, miR-133b and miR-206, and percent changes inknee extensor strength after RT were also observed, which suggeststhat RT induced changes in muscle strength through regulating amiRNA–TNNT1 AS pathway in older adults. Although a relationship
has not been reported,we propose thatmiRNAsmay act on TNNT1 alter-native splicing factors.
It should be noted that percent changes in knee extensor strengthdirectly correlated with changes in miR-133a, miR-133b and miR-206,making them potential candidates as biomarkers and therapeutictargets. Muscle specific miRNA downregulation has been reported toregulate muscle atrophy directly or indirectly through inhibiting theIGF1/PI3K/Akt axis (Wang, 2013). Also, they positively regulate muscledifferentiation and reinnervation (Kim et al., 2006; Williams et al.,2009), which may explain the increased muscle force in the absenceof increased muscle cross-sectional area (Zhang et al., 2014). Thus, ourfindings indicate that these myomiRs may play critical roles in regula-tion of muscle response to RT in older adults through targeting andbalancing multiple signaling pathways involved in muscle mass, inner-vation, and force regulation.
5. Limitations
Analysis of a larger cohort will help to better establish whethermiRNA expression and/or activity influences RT-evoked changes inskeletal muscle strength, whether different exercise modalities differ-entially regulate miRNA expression and associated cellular pathways,and how miRNA regulation contributes to physiological adaptation toRT. Due to the small number of participants, we were not able to carryout sex-specific analyses. As exercise evokes awhole body response, ex-tending our analysis to non-muscle specific miRNAs might provide amore comprehensive understanding of the neuromuscular system ad-aptation to RT in older adults. Since miR-1, -133a and -133b areexpressed also in the central nervous system (Bak et al., 2008), theirlevels in circulation derive from both muscle and central nervous sys-tem. Therefore, the response to RT may result from muscle and neuraladaptations. Future studies should include a non-exercise group. Mea-surements of central drive indices will help to examine the contributionof motor learning (improved motor unit recruitment, better relaxationof antagonist muscles) over repetitive exercise sessions.
Acknowledgments
This work was supported by the National Institutes of Health grants,AG13934 and AG15820 to O.D., AG020583 to B.N., and theWake ForestClaude D. Pepper Older Americans Independence Center (P30-AG21332).
References
Alevizos, I., Illei, G.G., 2010. MicroRNAs as biomarkers in rheumatic diseases. Nat. Rev.Rheumatol. 6, 391–398.
American College of Sports Medicine, 2009. American College of Sports Medicine positionstand. Progression models in resistance training for healthy adults. Med. Sci. SportsExerc. 41, 687–708.
Andersen, J.L., 2003. Muscle fibre type adaptation in the elderly human muscle. Scand.J. Med. Sci. Sports 13, 40–47.
Aoi,W., Ichikawa, H., Mune, K., Tanimura, Y., Mizushima, K., Naito, Y., Yoshikawa, T., 2013.Muscle-enriched microRNA miR-486 decreases in circulation in response to exercisein young men. Front. Physiol. 4, 80.
Asaga, S., Kuo, C., Nguyen, T., Terpenning, M., Giuliano, A.E., Hoon, D.S., 2011. Direct serumassay for microRNA-21 concentrations in early and advanced breast cancer. Clin.Chem. 57, 84–91.
Baggish, A.L., Hale, A., Weiner, R.B., Lewis, G.D., Systrom, D., Wang, F., Wang, T.J., Chan, S.Y.,2011. Dynamic regulation of circulating microRNA during acute exhaustive exerciseand sustained aerobic exercise training. J. Physiol. 589, 3983–3994.
Bak, M., Silahtaroglu, A., Moller, M., Christensen, M., Rath, M.F., Skryabin, B., Tommerup,N., Kauppinen, S., 2008.MicroRNA expression in the adult mouse central nervous sys-tem. RNA 14, 432–444.
Banzet, S., Chennaoui, M., Girard, O., Racinais, S., Drogou, C., Chalabi, H., Koulmann, N.,2013. Changes in circulating microRNAs levels with exercise modality. J. Appl. Phys-iol. 115, 1237–1244.
Bostjancic, E., Zidar, N., Stajer, D., Glavac, D., 2010. MicroRNAsmiR-1, miR-133a, miR-133band miR-208 are dysregulated in human myocardial infarction. Cardiology 115,163–169.
Callis, T.E., Deng, Z., Chen, J.F., Wang, D.Z., 2008. Muscling through the microRNA world.Exp. Biol. Med. 233, 131–138.

13T. Zhang et al. / Experimental Gerontology 62 (2015) 7–13
Cardinali, B., Castellani, L., Fasanaro, P., Basso, A., Alema, S., Martelli, F., Falcone, G., 2009.MicroRNA-221 and microRNA-222 modulate differentiation and maturation of skel-etal muscle cells. PLoS One 4, e7607.
Chen, J.F., Mandel, E.M., Thomson, J.M., Wu, Q., Callis, T.E., Hammond, S.M., Conlon, F.L.,Wang, D.Z., 2006. The role of microRNA-1 and microRNA-133 in skeletal muscle pro-liferation and differentiation. Nat. Genet. 38, 228–233.
Choi, S.J., Shively, C.A., Register, T.C., Feng, X., Stehle, J., High, K., Ip, E., Kritchevsky, S.B.,Nicklas, B., Delbono, O., 2013. Force-generation capacity of single vastus lateralismuscle fibers and physical function decline with age in African green vervet mon-keys. J. Gerontol. A: Biol. Med. Sci. 68, 258–267.
Davidsen, P.K., Gallagher, I.J., Hartman, J.W., Tarnopolsky, M.A., Dela, F., Helge, J.W.,Timmons, J.A., Phillips, S.M., 2011. High responders to resistance exercise trainingdemonstrate differential regulation of skeletal muscle microRNA expression. J. Appl.Physiol. 110, 309–317.
Delbono, O., 2003. Neural control of aging skeletal muscle. Aging Cell 2, 21–29.Delbono, O., 2011. Expression and regulation of excitation–contraction coupling proteins
in aging skeletal muscle. Curr. Aging Sci. 4, 248–259.Deng, Z., Chen, J.F., Wang, D.Z., 2011. Transgenic overexpression of miR-133a in skeletal
muscle. BMC Musculoskelet. Disord. 12, 115.Dey, B.K., Gagan, J., Dutta, A., 2011. MiR-206 and -486 induce myoblast differentiation by
downregulating Pax7. Mol. Cell. Biol. 203–214.Donaldson, A., Natanek, S.A., Lewis, A., Man, W.D., Hopkinson, N.S., Polkey, M.I., Kemp,
P.R., 2013. Increased skeletal muscle-specific microRNA in the blood of patientswith COPD. Thorax 68, 1140–1149.
Drummond, M.J., McCarthy, J.J., Fry, C.S., Esser, K.A., Rasmussen, B.B., 2008. Aging differen-tially affects human skeletal muscle microRNA expression at rest and after an anabol-ic stimulus of resistance exercise and essential amino acids. Am. J. Physiol. Endocrinol.Metab. 295, E1333–E1340.
Eisenberg, I., Eran, A., Nishino, I., Moggio, M., Lamperti, C., Amato, A.A., Lidov, H.G., Kang,P.B., North, K.N., Mitrani-Rosenbaum, S., Flanigan, K.M., Neely, L.A., Whitney, D.,Beggs, A.H., Kohane, I.S., Kunkel, L.M., 2007. Distinctive patterns of microRNA expres-sion in primary muscular disorders. Proc. Natl. Acad. Sci. U. S. A. 104, 17016–17021.
Endo, K., Weng, H., Naito, Y., Sasaoka, T., Takahashi, A., Fukushima, Y., Iwai, N., 2013. Clas-sification of various muscular tissues using miRNA profiling. Biomed. Res. 34,289–299.
Fiatarone, M.A., Marks, E.C., Ryan, N.D., Meredith, C.N., Lipsitz, L.A., Evans, W.J., 1990.High-intensity strength training in nonagenarians. Effects on skeletal muscle. JAMA263, 3029–3034.
Goodpaster, B.H., Park, S.W., Harris, T.B., Kritchevsky, S.B., Nevitt, M., Schwartz, A.V.,Simonsick, E.M., Tylavsky, F.A., Visser, M., Newman, A.B., 2006. The loss of skeletalmuscle strength, mass, and quality in older adults: the health, aging and body com-position study. J. Gerontol. A: Biol. Med. Sci. 61, 1059–1064.
Hamilton, A.J., Baulcombe, D.C., 1999. A species of small antisense RNA in posttranscrip-tional gene silencing in plants. Science 286, 950–952.
Heneghan, H.M., Miller, N., Lowery, A.J., Sweeney, K.J., Newell, J., Kerin, M.J., 2010. Circu-lating microRNAs as novel minimally invasive biomarkers for breast cancer. Ann.Surg. 251, 499–505.
Hoy, A.M., Buck, A.H., 2012. Extracellular small RNAs: what, where, why? Biochem. Soc.Trans. 40, 886–890.
Kim, H.K., Lee, Y.S., Sivaprasad, U., Malhotra, A., Dutta, A., 2006. Muscle-specific microRNAmiR-206 promotes muscle differentiation. J. Cell Biol. 174, 677–687.
Kirby, T.J., McCarthy, J.J., 2013. MicroRNAs in skeletal muscle biology and exercise adapta-tion. Free Radic. Biol. Med. 64, 95–105.
Kusakabe, R., Tani, S., Nishitsuji, K., Shindo, M., Okamura, K., Miyamoto, Y., Nakai, K.,Suzuki, Y., Kusakabe, T.G., Inoue, K., 2013. Characterization of the compact bicistronicmicroRNA precursor, miR-1/miR-133, expressed specifically in Ciona muscle tissues.Gene Expr. Patterns 13, 43–50.
Leenders, M., Verdijk, L.B., van der Hoeven, L., van Kranenburg, J., Nilwik, R., van Loon, L.J.,2013. Elderly men and women benefit equally from prolonged resistance-type exer-cise training. J. Gerontol. A Biol. Sci. Med. Sci. 68, 769–779.
Lexell, J., 1995. Human aging, muscle mass, and fiber type composition. J. Gerontol. A Biol.Sci. Med. Sci. 50 (Spec No: 11–16).
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods 25, 402–408.
Martin, H.J., Yule, V., Syddall, H.E., Dennison, E.M., Cooper, C., Aihie Sayer, A., 2006. Ishand-held dynamometry useful for the measurement of quadriceps strength inolder people? A comparison with the gold standard Bodex dynamometry. Gerontol-ogy 52, 154–159.
McCarthy, J.J., 2008. MicroRNA-206: the skeletal muscle-specific myomiR. Biochim.Biophys. Acta 1779, 682–691.
McCarthy, J.J., 2011. The MyomiR network in skeletal muscle plasticity. Exerc. Sport Sci.Rev. 39, 150–154.
McCarthy, J.J., Esser, K.A., 2007. MicroRNA-1 and microRNA-133a expression are de-creased during skeletal muscle hypertrophy. J. Appl. Physiol. 102, 306–313.
McCarthy, J.J., Esser, K.A., Andrade, F.H., 2007. MicroRNA-206 is overexpressed in the dia-phragm but not the hindlimb muscle of mdx mouse. Am. J. Physiol. Cell Physiol. 293,C451–C457.
McCarthy, J.J., Esser, K.A., Peterson, C.A., Dupont-Versteegden, E.E., 2009. Evidence ofMyomiR network regulation of beta-myosin heavy chain gene expression duringskeletal muscle atrophy. Physiol. Genomics 39, 219–226.
Mestdagh, P., Van Vlierberghe, P., De Weer, A., Muth, D., Westermann, F., Speleman, F.,Vandesompele, J., 2009. A novel and universal method for microRNA RT-qPCR datanormalization. Genome Biol. 10, R64.
Mitchell, P.S., Parkin, R.K., Kroh, E.M., Fritz, B.R., Wyman, S.K., Pogosova-Agadjanyan, E.L.,Peterson, A., Noteboom, J., O'Briant, K.C., Allen, A., Lin, D.W., Urban, N., Drescher, C.W.,Knudsen, B.S., Stirewalt, D.L., Gentleman, R., Vessella, R.L., Nelson, P.S., Martin, D.B.,Tewari, M., 2008. CirculatingmicroRNAs as stable blood-basedmarkers for cancer de-tection. Proc. Natl. Acad. Sci. U. S. A. 105, 10513–10518.
Mizuno, H., Nakamura, A., Aoki, Y., Ito, N., Kishi, S., Yamamoto, K., Sekiguchi, M., Takeda, S.,Hashido, K., 2011. Identification of muscle-specific microRNAs in serum of musculardystrophy animal models: promising novel blood-based markers for muscular dys-trophy. PLoS One 6, e18388.
Moresi, V., Williams, A.H., Meadows, E., Flynn, J.M., Potthoff, M.J., McAnally, J., Shelton,J.M., Backs, J., Klein, W.H., Richardson, J.A., Bassel-Duby, R., Olson, E.N., 2010.Myogenin and class II HDACs control neurogenic muscle atrophy by inducing E3ubiquitin ligases. Cell 143, 35–45.
Nakasa, T., Ishikawa, M., Shi, M., Shibuya, H., Adachi, N., Ochi, M., 2010. Acceleration ofmuscle regeneration by local injection of muscle-specific microRNAs in rat skeletalmuscle injury model. J. Cell. Mol. Med. 14, 2495–2505.
Ng, E.K., Chong, W.W., Jin, H., Lam, E.K., Shin, V.Y., Yu, J., Poon, T.C., Ng, S.S., Sung, J.J., 2009.Differential expression of microRNAs in plasma of patients with colorectal cancer: apotential marker for colorectal cancer screening. Gut 58, 1375–1381.
Reinhart, B.J., Slack, F.J., Basson, M., Pasquinelli, A.E., Bettinger, J.C., Rougvie, A.E., Horvitz,H.R., Ruvkun, G., 2000. The 21-nucleotide let-7 RNA regulates developmental timingin Caenorhabditis elegans. Nature 403, 901–906.
Russell, A.P., Lamon, S., Boon, H., Wada, S., Guller, I., Brown, E.L., Chibalin, A.V., Zierath, J.R.,Snow, R.J., Stepto, N., Wadley, G.D., Akimoto, T., 2013. Regulation of miRNAs inhuman skeletal muscle following acute endurance exercise and short-term endur-ance training. J. Physiol. 591, 4637–4653.
Sawada, S., Kon, M., Wada, S., Ushida, T., Suzuki, K., Akimoto, T., 2013. Profiling of circulat-ing microRNAs after a bout of acute resistance exercise in humans. PLoS One 8,e70823.
Sempere, L.F., Freemantle, S., Pitha-Rowe, I., Moss, E., Dmitrovsky, E., Ambros, V., 2004. Ex-pression profiling of mammalian microRNAs uncovers a subset of brain-expressedmicroRNAs with possible roles in murine and human neuronal differentiation. Ge-nome Biol. 5, R13.
Small, E.M., O'Rourke, J.R., Moresi, V., Sutherland, L.B., McAnally, J., Gerard, R.D.,Richardson, J.A., Olson, E.N., 2010. Regulation of PI3-kinase/Akt signaling by muscle-enriched microRNA-486. Proc. Natl. Acad. Sci. U. S. A. 107, 4218–4223.
Song, J., Bai, Z., Han, W., Zhang, J., Meng, H., Bi, J., Ma, X., Han, S., Zhang, Z., 2012. Identifi-cation of suitable reference genes for qPCR analysis of serum microRNA in gastriccancer patients. Dig. Dis. Sci. 57, 897–904.
Togliatto, G., Trombetta, A., Dentelli, P., Cotogni, P., Rosso, A., Tschop, M.H., Granata, R.,Ghigo, E., Brizzi, M.F., 2013. Unacylated ghrelin promotes skeletal muscle regenera-tion following hindlimb ischemia via SOD-2-mediated miR-221/222 expression.J. Am. Heart Assoc. 2, e000376.
Tonevitsky, A.G., Maltseva, D.V., Abbasi, A., Samatov, T.R., Sakharov, D.A., Shkurnikov,M.U., Lebedev, A.E., Galatenko, V.V., Grigoriev, A.I., Northoff, H., 2013. Dynamicallyregulated miRNA–mRNA networks revealed by exercise. BMC Physiol. 13, 9.
Tracy, B.L., Ivey, F.M., Hurlbut, D., Martel, G.F., Lemmer, J.T., Siegel, E.L., Metter, E.J., Fozard,J.L., Fleg, J.L., Hurley, B.F., 1999. Muscle quality. II. Effects of strength training in 65- to75-yr-old men and women. J. Appl. Physiol. 86, 195–201.
Uhlemann, M., Mobius-Winkler, S., Fikenzer, S., Adam, J., Redlich, M., Mohlenkamp, S.,Hilberg, T., Schuler, G.C., Adams, V., 2014. Circulating microRNA-126 increases afterdifferent forms of endurance exercise in healthy adults. Eur. J. Prev. Cardiol. 21,484–491.
Valadi, H., Ekstrom, K., Bossios, A., Sjostrand, M., Lee, J.J., Lotvall, J.O., 2007. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic ex-change between cells. Nat. Cell Biol. 9, 654–659.
van Rooij, E., Liu, N., Olson, E.N., 2008. MicroRNAs flex their muscles. Trends Genet. 24,159–166.
van Rooij, E., Quiat, D., Johnson, B.A., Sutherland, L.B., Qi, X., Richardson, J.A., Kelm Jr., R.J.,Olson, E.N., 2009. A family of microRNAs encoded by myosin genes governs myosinexpression and muscle performance. Dev. Cell 17, 662–673.
Wang, X.H., 2013. MicroRNA in myogenesis and muscle atrophy. Curr. Opin. Clin. Nutr.Metab. Care 16, 258–266.
Wang, K., Zhang, S., Marzolf, B., Troisch, P., Brightman, A., Hu, Z., Hood, L.E., Galas, D.J.,2009. Circulating microRNAs, potential biomarkers for drug-induced liver injury.Proc. Natl. Acad. Sci. U. S. A. 106, 4402–4407.
Wang, G.K., Zhu, J.Q., Zhang, J.T., Li, Q., Li, Y., He, J., Qin, Y.W., Jing, Q., 2010. CirculatingmicroRNA: a novel potential biomarker for early diagnosis of acutemyocardial infarc-tion in humans. Eur. Heart J. 31, 659–666.
Weilner, S., Schraml, E., Redl, H., Grillari-Voglauer, R., Grillari, J., 2013. Secretion ofmicrovesicular miRNAs in cellular and organismal aging. Exp. Gerontol. 48, 626–633.
Williams, A.H., Valdez, G., Moresi, V., Qi, X., McAnally, J., Elliott, J.L., Bassel-Duby, R., Sanes,J.R., Olson, E.N., 2009. MicroRNA-206 delays ALS progression and promotes regener-ation of neuromuscular synapses in mice. Science 326, 1549–1554.
Zacharewicz, E., Lamon, S., Russell, A.P., 2013. MicroRNAs in skeletal muscle and their reg-ulation with exercise, ageing, and disease. Front. Physiol. 4, 266.
Zhang, T., Choi, S.J., Wang, Z.M., Birbrair, A., Messi, M.L., Jin, J.P., Marsh, A.P., Nicklas, B.,Delbono, O., 2014. Human slow troponin T (TNNT1) pre-mRNA alternative splicingis an indicator of skeletal muscle response to resistance exercise in older adults.J. Gerontol. A: Biol. Med. Sci. 69, 1437–1447.
Copyright © 2022 FDOKUMEN




















