
A genetically modulated, intrinsic cingulate circuit supports human nicotine addiction
Upload
independentCategory
view
3download
0
ttidobMdGodthacacpcwmmBgMmCcmalmtsnab
Experimental Neurology 156, 149–164 (1999)Article ID exnr.1998.7009, available online at http://www.idealibrary.com on
Impaired Inflammatory Response to Glial Cell Death in GeneticallyMetallothionein-I- and -II-Deficient Mice
Milena Penkowa,* Mercedes Giralt,† Torben Moos,* Pernille S. Thomsen,* Joaquın Hernandez,†and Juan Hidalgo†,1
*Institute of Medical Anatomy, Section C, The Panum Institute, University of Copenhagen, DK-2200 Copenhagen, Denmark;and †Departamento de Biologıa Celular y Fisiologıa, Unidad de Fisiologıa Animal, Facultad de Ciencias,
Universidad Autonoma de Barcelona, Bellaterra, Barcelona, Spain 08193
Received July 23, 1998; accepted November 24, 1998
fid
tl
rrdmcTag(mt(acpCe
anaMgrviicr
Metallothionein I1II (MT-I1II) are acute-phase pro-eins which are upregulated during pathological condi-ions in the brain. To elucidate the neuropathologicalmportance of MT-I1II, we have examined MT-I1II-eficient mice following ip injection with 6-aminonic-tinamide (6-AN). 6-AN is antimetabolic and toxic forone marrow cells and grey matter astrocytes. InT1/1 mice, injection with 6-AN resulted in break-
own of the blood–brain barrier (BBB) and absence ofFAP-positive astrocytes in specific grey matter areasf the brain stem. Reactive astrocytosis encircled theamaged grey matter areas, which were heavily infil-rated by microglia/macrophages. The recruitment ofematogenous macrophages was accompanied by leak-ge of the BBB. The immunoreactivity (ir) of granulo-yte–macrophage-colony-stimulating factor (GM-CSF)nd the receptor for GM-CSF (GM-CSFrec) was signifi-antly upregulated in astrocytes and microglia/macro-hages, respectively. MT-I1IIir was also clearly in-reased in astrocytes surrounding the damaged areas,hile that of the CNS-specific MT isoform, MT-III, wasildly increased in both astrocytes and microglia/acrophages. In MT2/2 mice injected with 6-AN, theBB remained almost intact. The damage to specificrey matter areas was similar to that observed inT1/1 mice, but reactive astrocytosis, microglia/acrophages infiltration, and GM-CSFir and GM-SFrecir were clearly reduced in MT2/2 mice. Inontrast, MT-IIIir was dramatically increased in MT2/2ice. Total zinc decreased and histochemically detect-
ble zinc increased in the brain stem after 6-AN simi-arly in MT1/1 and MT2/2 mice. Bone marrow myeloid
onocytes and macrophages were increased as a reac-ion to 6-AN only in MT1/1 mice. The results demon-trate that the capability of MT2/2 mice to mount aormal inflammatory response in the brain is severelyttenuated, at least in part because of 6-AN-inducedone marrow affectation, involving MT-I1II for the
1
aTo whom correspondence should be addressed.149
rst time as major factors during CNS tissueamage. r 1999 Academic Press
Key Words: Metallothionein-I, -II, and -III; inflamma-ory response; GM-CSF; GM-CSFrec; astrocytes; microg-ia; 6-aminonicotinamide; zinc.
INTRODUCTION
Injury to the CNS elicits an acute inflammatoryesponse involving astrocytes, microglia, and bone mar-ow-derived monocytes (52, 54, 59). Reactive astrocytesisplay hyperplasia and hypertrophy (1, 54), whileicroglia and recruited bone marrow-derived mono-
ytes transform into amoeboid phagocytes (1–3, 52).he regulation of the inflammatory response is medi-ted by several proinflammatory cytokines, includingranulocyte–macrophage-colony-stimulating factorGM-CSF), which is expressed by both astrocytes andicroglia/macrophages (24, 38, 41). GM-CSF is a po-
ent mitogen for cells of the myelomonocytic lineage21, 23), and infusion of GM-CSF leads to recruitmentnd activation of microglia/macrophages and astro-ytes (23, 26, 61). Astrocytes and microglia/macro-hages also express the receptor for GM-CSF (GM-SFrec) (56), and during pathological conditions thisxpression is upregulated (38, 53).Astrocytes and microglia/macrophages also express
ntioxidant stress-related proteins, such as metallothio-eins (MTs) (see Refs. 5, 31, and 55 for review). MTs arefamily of proteins which in CNS occur in isoformsT-I, MT-II, and MT-III, the latter also referred to as
rowth inhibitory factor, because it inhibits survival ofat neurons in vitro (49, 63). MT-I1II are detected inirtually all tissues, including the brain, while MT-IIIs primarily confined to the brain and has been detectedn neurons and astrocytes (9, 30, 36, 44). The physiologi-al roles of the different MT isoforms in the brain stillemain to be established, but their high affinity for zinc
nd copper suggest that these proteins are involved in0014-4886/99 $30.00Copyright r 1999 by Academic Press
All rights of reproduction in any form reserved.

taasoucAatiPtsgdbscf
Mms6abaOtsh
A
erm
E
numwcfdmtnwr
podmasfmmPisdowawcicts(Ndkidcm
I
(isSopmp1m(pammpmzasa
150 PENKOWA ET AL.
he metabolism of these essential heavy metals. Inddition, MT-I1II have been shown to have significantntioxidant functions in vitro (62), and a number oftudies suggest that they could also function againstxidative stress in vivo (55). In the brain, MT-I1II arepregulated during pathological conditions where in-reased oxidative stress is likely to occur, such aslzheimer’s disease (15), brain trauma (51), kainiccid-induced seizures (14), intraperitoneal administra-ion of 6-aminonicotinamide (6-AN) (50), stress (30), orn transgenic overexpression of interleukin-6 (12, 29).resumably, the functions of MT-III will differ fromhose of MT-I1II, as suggested by in vitro studieshowing that MT-III but not MT-I1II affects the out-rowth of neurites (17, 63). MT-III has also beenemonstrated to be increased by animal models ofrain damage (4, 33–35, 64, 65). Taken together, thesetudies suggest that MTs are important proteins foroping with the brain damage elicited by a wide array ofactors, injuries, and diseases.
To elucidate the neuropathological importance ofT-I1II, we have examined mice carrying a nullutation in the MT-I1II genes (MT2/2 mice) (43)
ubjected to an ip injection with the niacin antagonist-AN. 6-AN induces systemic antimetabolic effects andstroglial cell death in certain grey matter areas ofrain stem and medulla spinalis, whereas other brainreas like the forebrain are unaffected (27, 39, 40, 50).ur results suggest that MT-I1II deficiency impairs
he capability to resist systemic treatment with 6-AN,ince MT2/2 mice are left with a severe depression ofemopoietic and inflammatory activity.
MATERIAL AND METHODS
nimals
Homozygous MT-I1II knockout (KO) mice were gen-rated as previously described (43). The KO mice wereaised on the 129/Sv genetic background; therefore,ice from this strain were used as controls.
xperimental Procedures
To induce CNS injury, mice were injected intraperito-eally with 6-AN, an antimetabolite with inhibitionpon rapidly dividing cells, such as those of the bonearrow. 6-AN inhibits the hexose monophosphate path-ay, which is used preferentially by protoplasmic astro-
ytes, whereby these cells undergo cytotoxic edemaollowed by cell death (27, 39, 40, 50). However, theamaging effect of 6-AN is confined to certain greyatter areas, whether or not such neurons have projec-
ions outside the CNS (50). Thus, only astrocytes of theuclei of brain stem and medulla spinalis are injured,hile other CNS regions, such as the forebrain, appear
esistant to 6-AN (39, 50). e
Adult MT2/2 mice and MT1/1 mice weighing ap-roximately 35 g received an intraperitoneal injectionf 10 mg/kg body weight of 6-AN (Sigma), which wasissolved in physiological saline in a concentration of 5g/ml. Three days after injections the mice were deeply
naesthetized with Brietal and fixed by vascular perfu-ion with 0.1% Na2S in 0.1 M PBS, pH 7.4, for 5 min,ollowed by perfusion Zamboni’s fixative, pH 7.4, for 10in, in one experiment. In another separate experi-ent the mice were fixed with 4% paraformaldehyde inBS, pH 7.4, for 10 min. 4% paraformaldehyde in PBS
n one experiment and with Zamboni’s fixative in aecond, separate experiment. Brains were afterwardissected and processed for cryosectioning as previ-usly described (46). Brains fixed in Zamboni’s fixativeere used for immunohistochemistry, histochemistry,nd Neo–Timm staining (sulfide silver staining). Na2Sas chosen because this concentration of sulfide ions
helates specifically zinc ions in the tissue. Brains fixedn 4% paraformaldehyde were used for immunohisto-hemistry and histochemistry. Thin 2-µm-thick consecu-ive sections were made for colocalizing antibodies. Theections were incubated in 1.0% H2O2 in TBS/NonidetTBS: 0.05 M Tris, pH 7.4, 0.15 M NaCl; with 0.01%onidet P-40 (Sigma)) to quench endogenous peroxi-ase. In yet a third separate experiment, the mice wereilled by cervical dislocations and the brains dissected
nto brain stem, cerebellum, and remaining brain andigested in nitric/perchloric acid for total zinc quantifi-ation. In each of the three experiments, three to sixice per group were used.
mmunohistochemistry
The sections were preincubated with pronase Eprotease type XIV, Sigma No. P5147, 0.025 g dissolvedn 50 ml TBS for 10 min, pH 7.4, at 37°C) and 10% goaterum in TBS/Nonidet (TBS with 0.01% Nonidet P-40,igma) for 15 min at room temperature, and incubatedvernight with one of the following primary antibodies:olyclonal rabbit anti-human NSE (as a neuronalarker) diluted 1:1000 (Dakopatts, DK; Code A589),
olyclonal rabbit anti-rat MT-I1II diluted 1:500 (18,9, 50), polyclonal rabbit anti-mouse albumin (as aarker for the blood–brain barrier (BBB) diluted 1:5000
Nordic Immunology, The Netherlands; Code 3291),olyclonal rabbit anti-cow GFAP (as a marker forstrocytes) diluted 1:250 (Dakopatts, DK; Code Z 334);onoclonal rat anti-mouse F4/80 (as a marker of allicroglia and bone marrow-derived monocytes/macro-
hages) diluted 1:15 (Serotec, UK; Code MCA 497),onoclonal rat anti-mouse GM-CSF diluted 1:50 (Gen-
yme Diagnostics, UK; Code 1723-01), polyclonal rabbitnti-mouse GM-CSFrec a-chain diluted 1:500 (Re-earch Diagnostics, Inc., USA), and polyclonal rabbitnti-rat MT-III diluted 1:1000 (to be described in full
lsewhere).
n1sRwpKtt(rs
itipAd(rw(
L
t1mgbpkrrip
N
vc
T
l6ze
s
dMageFen
atdestwtstemni
B
auaosMteAdbititisassda
Bts(6
151INFLAMMATORY RESPONSE IN MT-I1II-DEFICIENT MICE
The primary antibodies were detected using monoclo-al anti-rabbit IgG absorbed with rat serum proteins:400 (Sigma, USA; Code B3275) or biotinylated species-pecific sheep anti-rat IgG 1:1000 (Amersham, CodePN1002). The binding of these secondary antibodiesas verified using streptavidin–biotin–peroxidase com-lex (StreptABComplex/HRP, Dakopatts, DK; Code377) prepared at manufacturer’s recommended dilu-
ions for 30 min at room temperature. The immunoreac-ion was visualized using 0.015% H2O2 in DAB/TBSTBS: 0.05 M Tris, pH 7.4, 0.15 M NaCl), for 10 min atoom temperature. Consecutive sections were immuno-tained for MT-III and F4/80 and for MT-III and GFAP.In order to evaluate the extent of nonspecific binding
n the immunohistochemical experiments, control sec-ions of normal and MT-I1II null mice were incubatedn (i) the DAB medium alone (to examine endogenouseroxidase activity), (ii) the DAB medium and theBComplex prepared at manufacturer’s recommendedilutions (to examine endogenous biotin activity), andiii) the absence of primary antibody (to examine cross-eaction among IgGs of the different species). Resultsere considered only if these controls were negative
11).
ectin Histochemistry
Biotinylated tomato lectin from Lycopersicon esculen-um (Sigma, USA; Code L9389) was diluted 1:500 in0% goat serum and used as a marker for cells of theyelomonocytic cell lineage, such as ramified micro-
lial cells and amoeboid brain macrophages. Labelingy lectin was verified using streptavidin–biotin–eroxidase complex (StreptABComplex/HRP, Da-opatts, DK; Code K377) prepared at manufacturer’secommended dilution and incubated for 30 min atoom temperature. The immunoreaction was visual-zed using 0.015% H2O2 in DAB/TBS (TBS: 0.05 M Tris,H 7.4, 0.15 M NaCl), for 10 min at room temperature.
eo–Timm Staining
Sections were physically (autometallographically) de-eloped for 60 min at room temperature in solutionontaining silver lactate (45, 51).
otal Zinc Levels
For measuring total zinc levels of brain stem, cerebel-um, and remaining brain, the tissue was digested at0°C with a mixture of nitric and perchloric acid andinc measured by atomic absorption spectrophotom-try.
RESULTS
Essentially the same results were obtained in the two
eparate experiments carried out. Except from minor sepigmentation of the tales in MT2/2 mice, uninjectedT1/1 and MT2/2 mice were clinically indistinguish-
ble. Brain stem sections from uninjected mice of eitherroup did not reveal significantly histological differ-nces as verified by the immunoreactivity of GFAP,4/80, NSE, GM-CSF, GM-CSFrec, and MT-III. Asxpected, only MT1/1 mice displayed MT-I1II immu-oreactivity.Both MT1/1 and MT2/2 mice became hypoactive
nd developed weakness and motor impairments atheir limbs on the first day after 6-AN injection. By theay of sacrifice, animals were clearly paralyzed in theirxtremities, but they gained body weight and only atelightly less than uninjected mice. By gross examina-ion of the CNS and internal organs, no abnormalitiesere observed between MT1/1 and MT2/2 mice. In
oluidine blue-stained and NSE-immunostained braintem sections, no significant differences between 6-AN-reated MT1/1 and MT2/2 mice were observed. How-ver, in all mice specific grey matter areas in pons andedulla oblongata (for instance, the oliva inferior
ucleus) displayed tissue degeneration following 6-ANnjection.
BB
In uninjected mice of both genotypes, the BBB tolbumin was intact, and albumin immunoreaction prod-ct was only seen over the entire pial surface and inreas devoid of a BBB, such as the circumventricularrgans (e.g., area postrema, median eminence, andubfornical organ). In the brain stem of 6-AN-injectedT1/1 mice, the BBB to albumin was compromised, in
hat albumin immunoreaction product was observedxtra- and intracellularly in the brain stem (Fig. 1A).lbumin immunoreactivity at 3 days postinjection (3pi) was most pronounced intracellularly, which coulde due to the number of days passing after the 6-ANnjection combined with the regenerative capability ofhe BBB. This is supported by examination of 6-AN-njected rats at 1 dpi, in that brain stem sections ofhese animals display significant extracellular and lessntracellular albumin immunoreactivity (data nothown). The observed intracellular albumin immunore-ction staining product was never seen in controlections, in which we excluded the primary and/orecondary antibody and/or the ABC complex. In cortex,iencephalon, and cerebellum, the BBB to albuminppeared similar to that of uninjected mice.In the brain stem of 6-AN-injected MT2/2 mice, theBB to albumin was barely affected, when compared to
hat of uninjected mice. Only a few cells in the braintem displayed albumin immunoreactivity at 3 dpiFig. 1B). In cortex, diencephalon, and cerebellum of-AN-injected MT2/2 mice, the BBB to albumin was
imilar to that of uninjected mice.
A
tsarcplr
tse6gHcM62lc
M
nsiwgnattsG
ip
tbt(prlrt
G
Gjctinipg
achc4pCma
G
Gjchd
bAs
152 PENKOWA ET AL.
strocytes
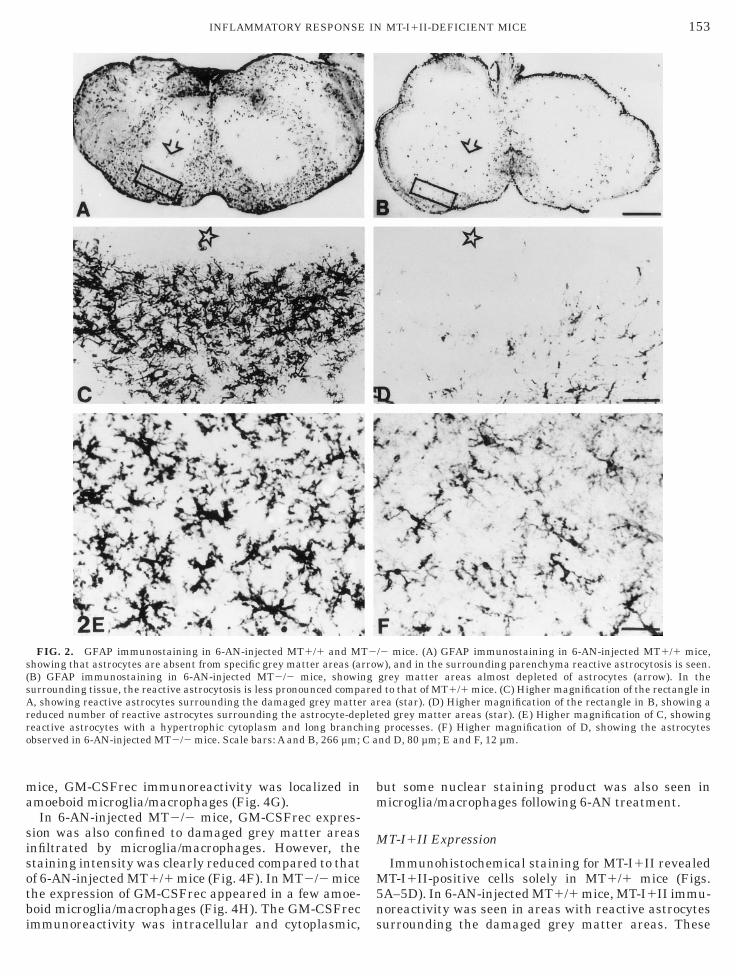
In 6-AN-treated MT1/1 mice, GFAP-expressing as-rocytes were absent in grey matter regions of the braintem, including cranial nerve nuclei. Surrounding thesereas reactive astrogliosis was observed (Fig. 2A). Theeactive astrocytes exhibited hypertrophy with swollenell bodies, and they displayed long elongated cellrocesses (Figs. 2C and 2E). In the cortex, diencepha-on, and cerebellum of 6-AN-treated MT1/1 mice, noeactive astrocytes were identified.In 6-AN-treated MT2/2 mice, GFAP-expressing as-
rocytes were absent in grey matter regions of the braintem, including cranial nerve nuclei (Fig. 2B). Thextent of astrocytic degeneration was similar to that of-AN-injected MT1/1 mice. Surrounding the damagedrey matter areas, reactive astrogliosis was observed.owever, the number of reactive astrocytes was signifi-
antly reduced compared to that of 6-AN-injectedT1/1 mice, and only some of the astrocytes in
-AN-treated MT2/2 mice displayed hypertrophy (Figs.D and 2F). In the cortex, diencephalon, and cerebel-um of 6-AN-treated MT2/2 mice, no reactive astro-ytes were identified.
icroglia/Macrophages
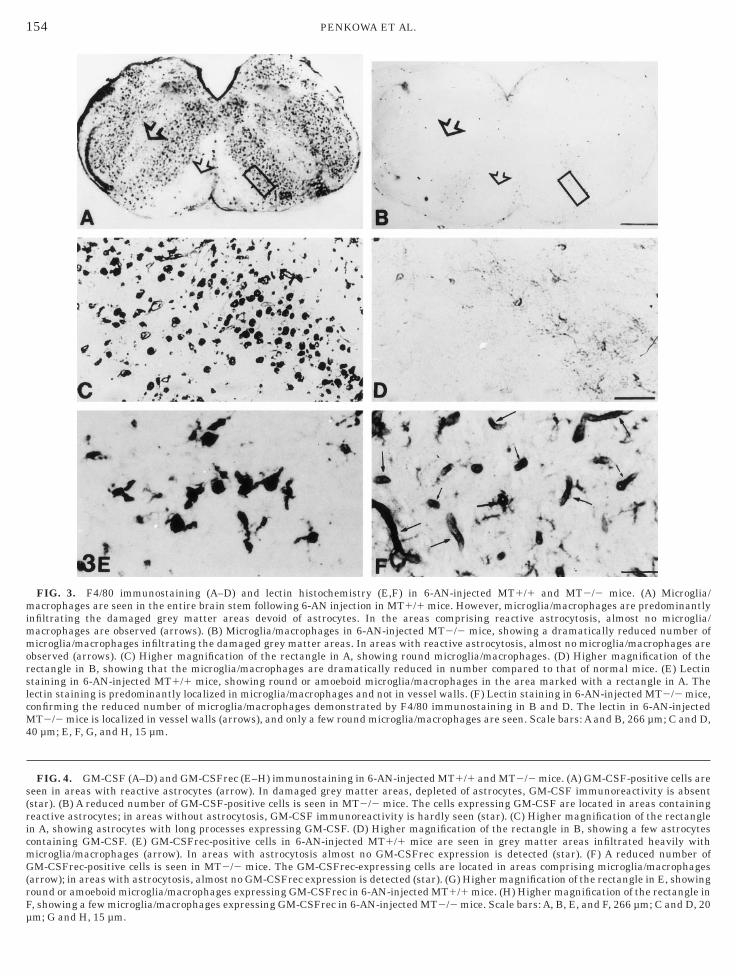
In 6-AN-treated MT1/1 mice, an increase in theumber of amoeboid microglia/macrophages was ob-erved in the entire brain stem as verified from F4/80mmunostaining and lectin staining. The brain stemas mostly infiltrated with microglia/macrophages inrey matter areas as the oliva inferior nuclei, pontineuclei, and vestibular nuclei (Fig. 3A). The meningesnd area postrema also contained numerous inflamma-ory microglia/macrophages. The highest levels of infil-rating inflammatory microglia/macrophages were ob-erved in the damaged grey matter areas depleted ofFAP-positive astrocytes (Figs. 3A, 3C, and 3E).In 6-AN-treated MT2/2 mice, the number of F4/80-
mmunoreacting and lectin-stained microglia/macro-
FIG. 1. Albumin immunostaining in 6-AN-injected MT1/1 andrain stem of 6-AN-injected MT1/1 mice. Both damaged grey mattelbumin immunostaining in 6-AN-injected MT2/2 mice, showing anome albumin is detected. Scale bars: A and B, 266 µm.
hages was dramatically decreased when compared to a
hat of 6-AN-treated MT1/1 mice. Only a few amoe-oid microglia/macrophages were observed infiltratinghe damaged grey matter areas depleted of astrocytesFigs. 3B, 3D, and 3F). Even in the meninges and areaostrema the number of microglia/macrophages waseduced compared to that of MT1/1 mice. The microg-ia/macrophages appeared round or amoeboid, whileamified or resting microglia was hardly seen in any ofhe mice following 6-AN injection (Figs. 3A–3F).
M-CSF Expression
In 6-AN-injected MT1/1 mice, the expression ofM-CSF was upregulated compared to that of unin-
ected MT1/1 mice. GM-CSF immunoreactivity wasonfined to areas with reactive astrocytosis encirclinghe damaged grey matter areas (Fig. 4A). In the areasnfiltrated with microglia/macrophages, GM-CSF immu-oreactivity was mostly absent. The GM-CSF-display-
ng cells had a phenotype similar to that of GFAPositive astrocytes with abundant cytoplasm and elon-ated processes (Fig. 4C).In 6-AN-injected MT2/2 mice, GM-CSF immunore-
ctivity was also confined to areas with reactive astro-ytosis encircling the damaged grey matter areas;owever, the staining intensity was clearly reducedompared to that of 6-AN-injected MT1/1 mice (Fig.B). In MT2/2 mice the expression of GM-CSF ap-eared in some stellate astrocytes (Fig. 4D). The GM-SF immunoreactivity was intracellular and cytoplas-ic, but some nuclear staining product was also seen in
strocytes following 6-AN treatment.
M-CSFrec Expression
In 6-AN-injected MT1/1 mice, the expression ofM-CSFrec was upregulated compared to that of unin-
ected MT1/1 mice. GM-CSFrec immunoreactivity wasonfined to damaged grey matter areas infiltratedeavily with microglia/macrophages (Fig. 4E). In areasisplaying reactive astrocytosis GM-CSFrec immunore-
2/2 mice. (A) The BBB to albumin was compromised in the entirereas and the surrounding are containing extravasated albumin. (B)ost intact BBB, and only in the center of damaged grey matter areas
MTr aalm
ctivity was merely absent. In 6-AN-injected MT1/1
ma
sisotbi
bm
M
M5n
s(sArro
153INFLAMMATORY RESPONSE IN MT-I1II-DEFICIENT MICE
ice, GM-CSFrec immunoreactivity was localized inmoeboid microglia/macrophages (Fig. 4G).In 6-AN-injected MT2/2 mice, GM-CSFrec expres-
ion was also confined to damaged grey matter areasnfiltrated by microglia/macrophages. However, thetaining intensity was clearly reduced compared to thatf 6-AN-injected MT1/1 mice (Fig. 4F). In MT2/2 micehe expression of GM-CSFrec appeared in a few amoe-oid microglia/macrophages (Fig. 4H). The GM-CSFrec
FIG. 2. GFAP immunostaining in 6-AN-injected MT1/1 and Mhowing that astrocytes are absent from specific grey matter areas (aB) GFAP immunostaining in 6-AN-injected MT2/2 mice, showinurrounding tissue, the reactive astrocytosis is less pronounced comp, showing reactive astrocytes surrounding the damaged grey matte
educed number of reactive astrocytes surrounding the astrocyte-depeactive astrocytes with a hypertrophic cytoplasm and long branchbserved in 6-AN-injected MT2/2 mice. Scale bars: A and B, 266 µm;
mmunoreactivity was intracellular and cytoplasmic, s
ut some nuclear staining product was also seen inicroglia/macrophages following 6-AN treatment.
T-I1II Expression
Immunohistochemical staining for MT-I1II revealedT-I1II-positive cells solely in MT1/1 mice (Figs.
A–5D). In 6-AN-injected MT1/1 mice, MT-I1II immu-oreactivity was seen in areas with reactive astrocytes
/2 mice. (A) GFAP immunostaining in 6-AN-injected MT1/1 mice,), and in the surrounding parenchyma reactive astrocytosis is seen.
grey matter areas almost depleted of astrocytes (arrow). In thed to that of MT1/1 mice. (C) Higher magnification of the rectangle inrea (star). (D) Higher magnification of the rectangle in B, showing aed grey matter areas (star). (E) Higher magnification of C, showingprocesses. (F) Higher magnification of D, showing the astrocytes
nd D, 80 µm; E and F, 12 µm.
T2rrowg
arer aletingC a
urrounding the damaged grey matter areas. These
mimmorslcM4
s(ricmG(rFµ
154 PENKOWA ET AL.
FIG. 3. F4/80 immunostaining (A–D) and lectin histochemistry (E,F) in 6-AN-injected MT1/1 and MT2/2 mice. (A) Microglia/acrophages are seen in the entire brain stem following 6-AN injection in MT1/1 mice. However, microglia/macrophages are predominantly
nfiltrating the damaged grey matter areas devoid of astrocytes. In the areas comprising reactive astrocytosis, almost no microglia/acrophages are observed (arrows). (B) Microglia/macrophages in 6-AN-injected MT2/2 mice, showing a dramatically reduced number oficroglia/macrophages infiltrating the damaged grey matter areas. In areas with reactive astrocytosis, almost no microglia/macrophages are
bserved (arrows). (C) Higher magnification of the rectangle in A, showing round microglia/macrophages. (D) Higher magnification of theectangle in B, showing that the microglia/macrophages are dramatically reduced in number compared to that of normal mice. (E) Lectintaining in 6-AN-injected MT1/1 mice, showing round or amoeboid microglia/macrophages in the area marked with a rectangle in A. Theectin staining is predominantly localized in microglia/macrophages and not in vessel walls. (F) Lectin staining in 6-AN-injected MT2/2 mice,onfirming the reduced number of microglia/macrophages demonstrated by F4/80 immunostaining in B and D. The lectin in 6-AN-injectedT2/2 mice is localized in vessel walls (arrows), and only a few round microglia/macrophages are seen. Scale bars: A and B, 266 µm; C and D,
0 µm; E, F, G, and H, 15 µm.
FIG. 4. GM-CSF (A–D) and GM-CSFrec (E–H) immunostaining in 6-AN-injected MT1/1 and MT2/2 mice. (A) GM-CSF-positive cells areeen in areas with reactive astrocytes (arrow). In damaged grey matter areas, depleted of astrocytes, GM-CSF immunoreactivity is absentstar). (B) A reduced number of GM-CSF-positive cells is seen in MT2/2 mice. The cells expressing GM-CSF are located in areas containingeactive astrocytes; in areas without astrocytosis, GM-CSF immunoreactivity is hardly seen (star). (C) Higher magnification of the rectanglen A, showing astrocytes with long processes expressing GM-CSF. (D) Higher magnification of the rectangle in B, showing a few astrocytesontaining GM-CSF. (E) GM-CSFrec-positive cells in 6-AN-injected MT1/1 mice are seen in grey matter areas infiltrated heavily withicroglia/macrophages (arrow). In areas with astrocytosis almost no GM-CSFrec expression is detected (star). (F) A reduced number ofM-CSFrec-positive cells is seen in MT2/2 mice. The GM-CSFrec-expressing cells are located in areas comprising microglia/macrophages
arrow); in areas with astrocytosis, almost no GM-CSFrec expression is detected (star). (G) Higher magnification of the rectangle in E, showingound or amoeboid microglia/macrophages expressing GM-CSFrec in 6-AN-injected MT1/1 mice. (H) Higher magnification of the rectangle in, showing a few microglia/macrophages expressing GM-CSFrec in 6-AN-injected MT2/2 mice. Scale bars: A, B, E, and F, 266 µm; C and D, 20
m; G and H, 15 µm.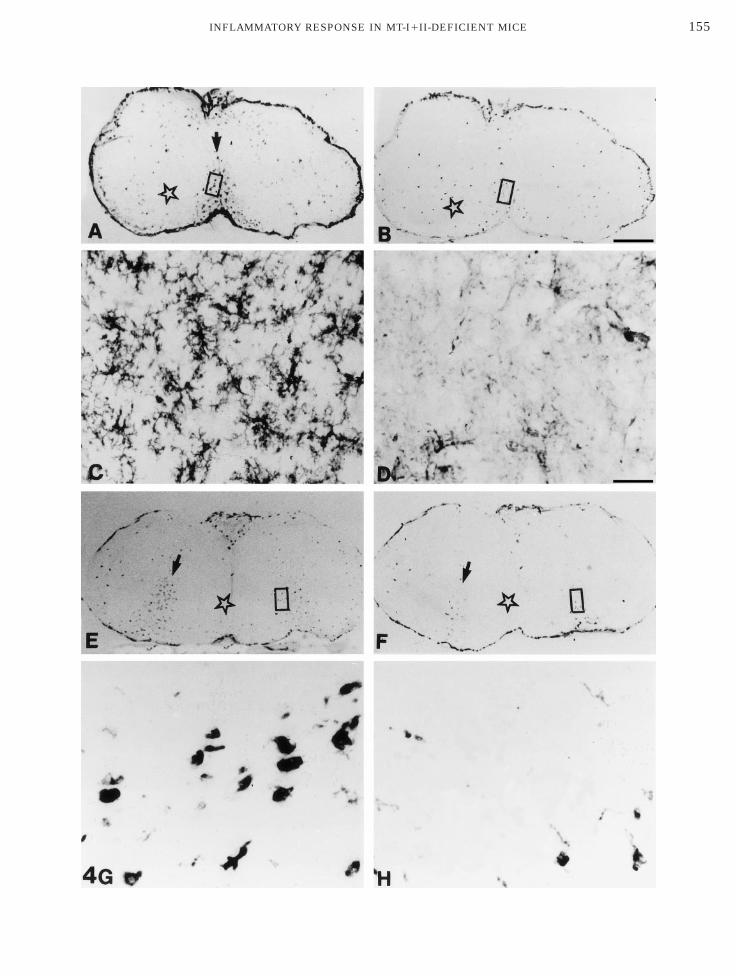
155INFLAMMATORY RESPONSE IN MT-I1II-DEFICIENT MICE

cG(om
M
tiMailmt
MgprwaMtp
(iaid
appwM(
B
iaisitrr(M
mimi
156 PENKOWA ET AL.
ells displayed a phenotype similar to that of GFAP andM-CSF positive astrocytes with elongated processes
Fig. 5C). MT-I1II expression was absent from the centerf damaged grey matter areas infiltrated with amoeboidicroglia/macrophages following the 6-AN injection.
T-III Expression
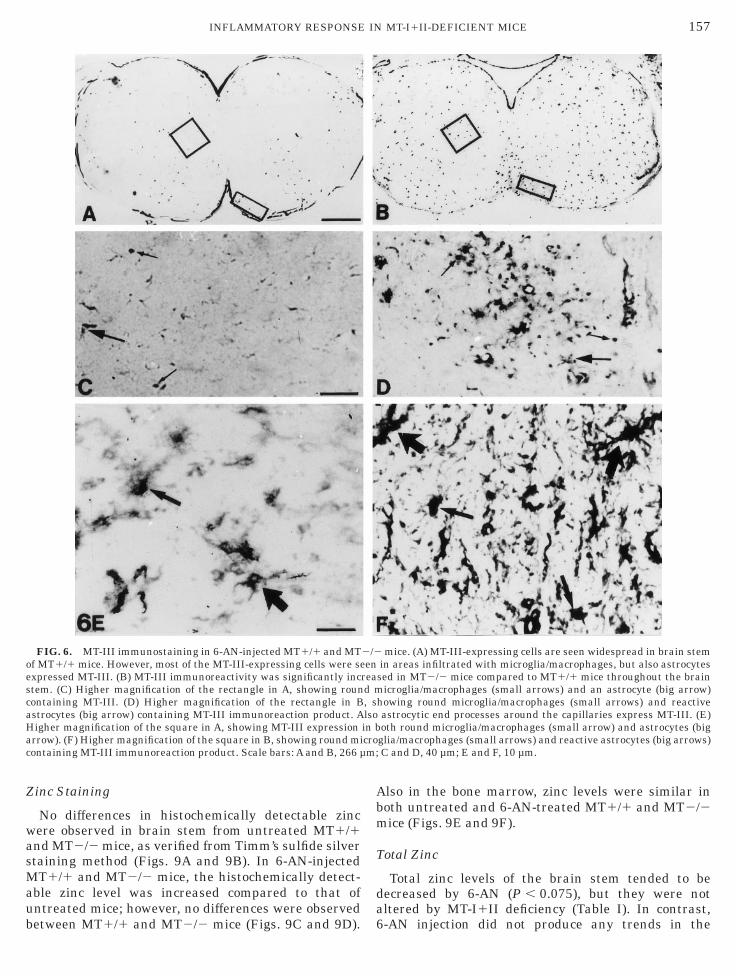
In 6-AN-injected MT1/1 mice, MT-III immunoreac-ivity was mildly upregulated in the entire brain stemn both grey and white matter areas (Fig. 6A), with
T-III expression confined to reactive astrocytes andmoeboid microglia/macrophages. The MT-III-express-ng astrocytes showed cytoplasmic hypertrophy andong cell processes, while MT-III expressing microglia/
acrophages were round or amoeboid without ramifica-ion (Figs. 6C and 6E).
In 6-AN-injected MT2/2 mice, the expression ofT-III was markedly increased in both the damaged
rey matter areas infiltrated with microglia/macro-hages and in the surrounding parenchyma comprisingeactive astrocytes (Fig. 6B). MT-III immunoreactivityas observed in virtually all of the astrocytes andmoeboid microglia/macrophages present in these mice.T-III expressing astrocytes displayed hypertrophy of
he cell body and long processes. Microglia/macro-
FIG. 5. MT-I1II immunostaining in 6-AN-injected MT1/1 andice, showing MT-I1II expression in areas with reactive astrocytosis
nfiltrated with microglia/macrophages (star). (B1C) MT-I1II immunatter areas infiltrated with microglia/macrophages (star) almost no
s absent from MT2/2 mice. Scale bars: A, 266 µm; B and D, 40 µm; C
hages were amoeboid or round without ramifications c
Figs. 6D and 6F). MT-III immunoreactivity was primar-ly observed intracellularly with labeling of both nucleusnd the paranuclear cytoplasm. MT-III immunoreactiv-ty was upregulated in the brain stem, while in cortex,iencephalon, and cerebellum no upregulation was seen.Consecutive sections were immunostained for MT-III
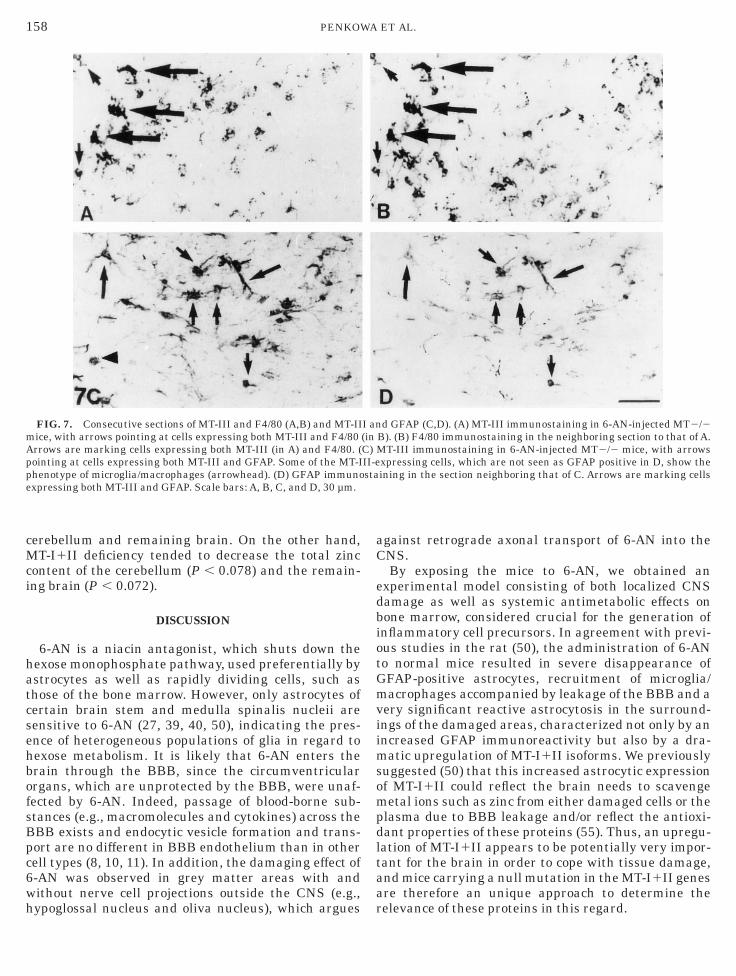
nd F4/80 to show that MT-III-expressing cells dis-layed the marker for amoeboid microglia/macro-hages (Figs. 7A and 7B). Also, consecutive sectionsere immunostained for MT-III and GFAP to show thatT-III expression was localized in astrocytes as well
Figs. 7C and 7D).
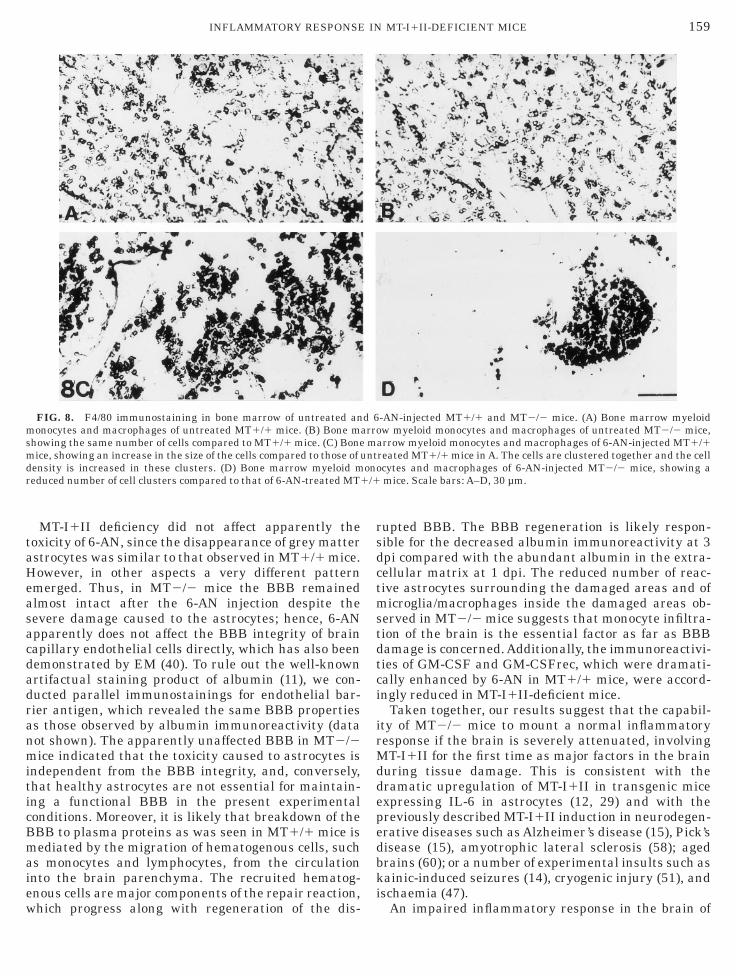
one Marrow Myelomonocytes
The number of myeloid monocytes and macrophagesn the bone marrow was the same in untreated MT1/1nd MT2/2 mice as verified from F4/80 immunoreactiv-ty (Figs. 8A and 8B). Following 6-AN injection, theizes of myeloid monocytes and macrophages werencreased in MT1/1 mice and the cells clusteredogether (Fig. 8C). In 6-AN-injected MT2/2 mice, theeactive response of the bone marrow was clearlyeduced compared to that of 6-AN-treated MT1/1 miceFig. 8D). Following 6-AN injection, the bone marrow of
T2/2 mice showed a reduced number of cell clusters
2/2 mice. (A) MT-I1II immunoreactivity in 6-AN-injected MT1/1row). MT-I1II expression is less pronounced in the grey matter areasactivity is seen in reactive astrocytes with long processes. In the grey-I1II immunoreactivity was detected. (D) MT-I1II immunoreactivityµm.
MT(aroreMT, 30
ompared to that of 6-AN-treated MT1/1 mice.
Z
wasMaub
Abm
T
da
oescaHac m;
157INFLAMMATORY RESPONSE IN MT-I1II-DEFICIENT MICE
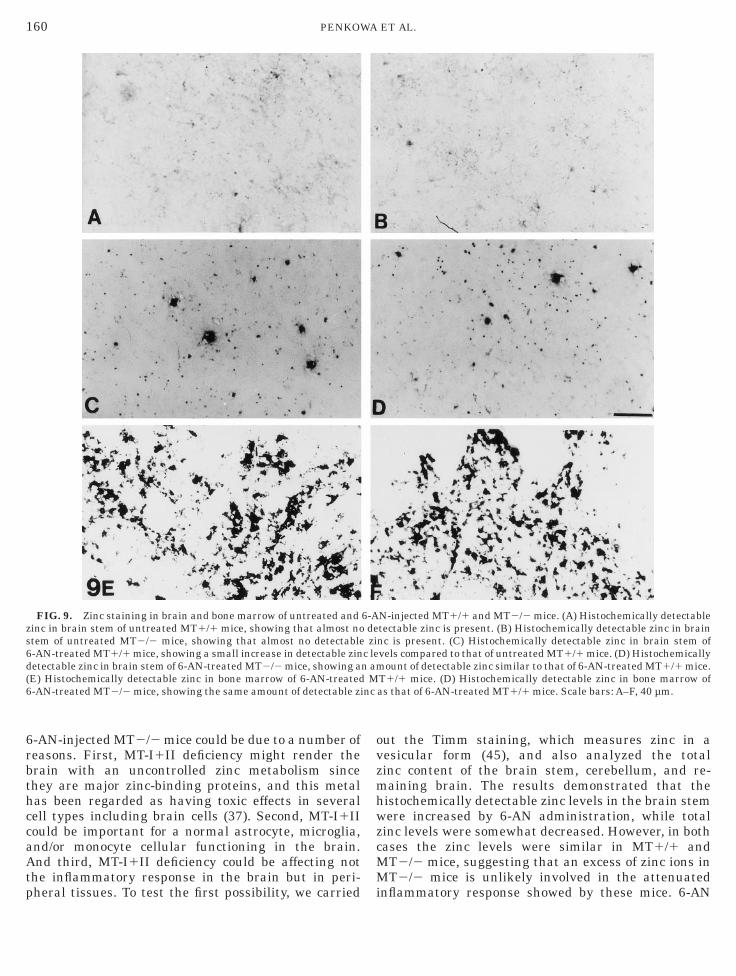
inc Staining
No differences in histochemically detectable zincere observed in brain stem from untreated MT1/1nd MT2/2 mice, as verified from Timm’s sulfide silvertaining method (Figs. 9A and 9B). In 6-AN-injectedT1/1 and MT2/2 mice, the histochemically detect-
ble zinc level was increased compared to that ofntreated mice; however, no differences were observed
FIG. 6. MT-III immunostaining in 6-AN-injected MT1/1 and MTf MT1/1 mice. However, most of the MT-III-expressing cells were sexpressed MT-III. (B) MT-III immunoreactivity was significantly inctem. (C) Higher magnification of the rectangle in A, showing rounontaining MT-III. (D) Higher magnification of the rectangle in Bstrocytes (big arrow) containing MT-III immunoreaction product. Aigher magnification of the square in A, showing MT-III expressionrrow). (F) Higher magnification of the square in B, showing round miontaining MT-III immunoreaction product. Scale bars: A and B, 266 µ
etween MT1/1 and MT2/2 mice (Figs. 9C and 9D). 6
lso in the bone marrow, zinc levels were similar inoth untreated and 6-AN-treated MT1/1 and MT2/2ice (Figs. 9E and 9F).
otal Zinc
Total zinc levels of the brain stem tended to beecreased by 6-AN (P , 0.075), but they were notltered by MT-I1II deficiency (Table I). In contrast,
mice. (A) MT-III-expressing cells are seen widespread in brain stemin areas infiltrated with microglia/macrophages, but also astrocytessed in MT2/2 mice compared to MT1/1 mice throughout the brain
icroglia/macrophages (small arrows) and an astrocyte (big arrow)howing round microglia/macrophages (small arrows) and reactiveastrocytic end processes around the capillaries express MT-III. (E)oth round microglia/macrophages (small arrow) and astrocytes (big
glia/macrophages (small arrows) and reactive astrocytes (big arrows)C and D, 40 µm; E and F, 10 µm.
2/2en
read m, slsoin bcro
-AN injection did not produce any trends in the
cMci
hatcsehbofsBpc6wh
aC
edbiotGmviimsompdltaa
mAppe
158 PENKOWA ET AL.
erebellum and remaining brain. On the other hand,T-I1II deficiency tended to decrease the total zinc
ontent of the cerebellum (P , 0.078) and the remain-ng brain (P , 0.072).
DISCUSSION
6-AN is a niacin antagonist, which shuts down theexose monophosphate pathway, used preferentially bystrocytes as well as rapidly dividing cells, such ashose of the bone marrow. However, only astrocytes ofertain brain stem and medulla spinalis nucleii areensitive to 6-AN (27, 39, 40, 50), indicating the pres-nce of heterogeneous populations of glia in regard toexose metabolism. It is likely that 6-AN enters therain through the BBB, since the circumventricularrgans, which are unprotected by the BBB, were unaf-ected by 6-AN. Indeed, passage of blood-borne sub-tances (e.g., macromolecules and cytokines) across theBB exists and endocytic vesicle formation and trans-ort are no different in BBB endothelium than in otherell types (8, 10, 11). In addition, the damaging effect of-AN was observed in grey matter areas with andithout nerve cell projections outside the CNS (e.g.,
FIG. 7. Consecutive sections of MT-III and F4/80 (A,B) and MT-IIice, with arrows pointing at cells expressing both MT-III and F4/80rrows are marking cells expressing both MT-III (in A) and F4/80. (ointing at cells expressing both MT-III and GFAP. Some of the MT-Ihenotype of microglia/macrophages (arrowhead). (D) GFAP immunoxpressing both MT-III and GFAP. Scale bars: A, B, C, and D, 30 µm.
ypoglossal nucleus and oliva nucleus), which argues r
gainst retrograde axonal transport of 6-AN into theNS.By exposing the mice to 6-AN, we obtained an
xperimental model consisting of both localized CNSamage as well as systemic antimetabolic effects onone marrow, considered crucial for the generation ofnflammatory cell precursors. In agreement with previ-us studies in the rat (50), the administration of 6-ANo normal mice resulted in severe disappearance ofFAP-positive astrocytes, recruitment of microglia/acrophages accompanied by leakage of the BBB and a
ery significant reactive astrocytosis in the surround-ngs of the damaged areas, characterized not only by anncreased GFAP immunoreactivity but also by a dra-
atic upregulation of MT-I1II isoforms. We previouslyuggested (50) that this increased astrocytic expressionf MT-I1II could reflect the brain needs to scavengeetal ions such as zinc from either damaged cells or the
lasma due to BBB leakage and/or reflect the antioxi-ant properties of these proteins (55). Thus, an upregu-ation of MT-I1II appears to be potentially very impor-ant for the brain in order to cope with tissue damage,nd mice carrying a null mutation in the MT-I1II genesre therefore an unique approach to determine the
d GFAP (C,D). (A) MT-III immunostaining in 6-AN-injected MT2/2B). (B) F4/80 immunostaining in the neighboring section to that of A.MT-III immunostaining in 6-AN-injected MT2/2 mice, with arrowsxpressing cells, which are not seen as GFAP positive in D, show theining in the section neighboring that of C. Arrows are marking cells
I an(inC)II-esta
elevance of these proteins in this regard.
taHeasacdadranmiticBmaiew
rsdctmstdtci
irMddepedbki
msmdr
159INFLAMMATORY RESPONSE IN MT-I1II-DEFICIENT MICE
MT-I1II deficiency did not affect apparently theoxicity of 6-AN, since the disappearance of grey matterstrocytes was similar to that observed in MT1/1 mice.owever, in other aspects a very different pattern
merged. Thus, in MT2/2 mice the BBB remainedlmost intact after the 6-AN injection despite theevere damage caused to the astrocytes; hence, 6-ANpparently does not affect the BBB integrity of brainapillary endothelial cells directly, which has also beenemonstrated by EM (40). To rule out the well-knownrtifactual staining product of albumin (11), we con-ucted parallel immunostainings for endothelial bar-ier antigen, which revealed the same BBB propertiess those observed by albumin immunoreactivity (dataot shown). The apparently unaffected BBB in MT2/2ice indicated that the toxicity caused to astrocytes is
ndependent from the BBB integrity, and, conversely,hat healthy astrocytes are not essential for maintain-ng a functional BBB in the present experimentalonditions. Moreover, it is likely that breakdown of theBB to plasma proteins as was seen in MT1/1 mice isediated by the migration of hematogenous cells, such
s monocytes and lymphocytes, from the circulationnto the brain parenchyma. The recruited hematog-nous cells are major components of the repair reaction,
FIG. 8. F4/80 immunostaining in bone marrow of untreated anonocytes and macrophages of untreated MT1/1 mice. (B) Bone m
howing the same number of cells compared to MT1/1 mice. (C) Boneice, showing an increase in the size of the cells compared to those of
ensity is increased in these clusters. (D) Bone marrow myeloid meduced number of cell clusters compared to that of 6-AN-treated MT1
hich progress along with regeneration of the dis-
upted BBB. The BBB regeneration is likely respon-ible for the decreased albumin immunoreactivity at 3pi compared with the abundant albumin in the extra-ellular matrix at 1 dpi. The reduced number of reac-ive astrocytes surrounding the damaged areas and oficroglia/macrophages inside the damaged areas ob-
erved in MT2/2 mice suggests that monocyte infiltra-ion of the brain is the essential factor as far as BBBamage is concerned. Additionally, the immunoreactivi-ies of GM-CSF and GM-CSFrec, which were dramati-ally enhanced by 6-AN in MT1/1 mice, were accord-ngly reduced in MT-I1II-deficient mice.
Taken together, our results suggest that the capabil-ty of MT2/2 mice to mount a normal inflammatoryesponse if the brain is severely attenuated, involvingT-I1II for the first time as major factors in the brain
uring tissue damage. This is consistent with theramatic upregulation of MT-I1II in transgenic micexpressing IL-6 in astrocytes (12, 29) and with thereviously described MT-I1II induction in neurodegen-rative diseases such as Alzheimer’s disease (15), Pick’sisease (15), amyotrophic lateral sclerosis (58); agedrains (60); or a number of experimental insults such asainic-induced seizures (14), cryogenic injury (51), and
schaemia (47).
-AN-injected MT1/1 and MT2/2 mice. (A) Bone marrow myeloidw myeloid monocytes and macrophages of untreated MT2/2 mice,
arrow myeloid monocytes and macrophages of 6-AN-injected MT1/1reated MT1/1 mice in A. The cells are clustered together and the cellcytes and macrophages of 6-AN-injected MT2/2 mice, showing amice. Scale bars: A–D, 30 µm.
d 6arro
muntono/1
An impaired inflammatory response in the brain of
6rbthccaAtp
ovzmhwzcMM
zs6d(6
160 PENKOWA ET AL.
-AN-injected MT2/2 mice could be due to a number ofeasons. First, MT-I1II deficiency might render therain with an uncontrolled zinc metabolism sincehey are major zinc-binding proteins, and this metalas been regarded as having toxic effects in severalell types including brain cells (37). Second, MT-I1IIould be important for a normal astrocyte, microglia,nd/or monocyte cellular functioning in the brain.nd third, MT-I1II deficiency could be affecting not
he inflammatory response in the brain but in peri-
FIG. 9. Zinc staining in brain and bone marrow of untreated andinc in brain stem of untreated MT1/1 mice, showing that almost notem of untreated MT2/2 mice, showing that almost no detectable-AN-treated MT1/1 mice, showing a small increase in detectable zinetectable zinc in brain stem of 6-AN-treated MT2/2 mice, showing aE) Histochemically detectable zinc in bone marrow of 6-AN-treated-AN-treated MT2/2 mice, showing the same amount of detectable zi
heral tissues. To test the first possibility, we carried i
ut the Timm staining, which measures zinc in aesicular form (45), and also analyzed the totalinc content of the brain stem, cerebellum, and re-aining brain. The results demonstrated that the
istochemically detectable zinc levels in the brain stemere increased by 6-AN administration, while total
inc levels were somewhat decreased. However, in bothases the zinc levels were similar in MT1/1 andT2/2 mice, suggesting that an excess of zinc ions inT2/2 mice is unlikely involved in the attenuated
N-injected MT1/1 and MT2/2 mice. (A) Histochemically detectabletectable zinc is present. (B) Histochemically detectable zinc in brainnc is present. (C) Histochemically detectable zinc in brain stem ofvels compared to that of untreated MT1/1 mice. (D) Histochemically
ount of detectable zinc similar to that of 6-AN-treated MT1/1 mice.T1/1 mice. (D) Histochemically detectable zinc in bone marrow ofas that of 6-AN-treated MT1/1 mice. Scale bars: A–F, 40 µm.
6-Adezi
c len am
Mnc
nflammatory response showed by these mice. 6-AN

wdmM
dslmaMiam
bmmasmprmmcmiomMmcrmpbipp
mrflowti(aw2ms
MmobmdsIacsacmacaodoitnfr
aso
BCR
am
161INFLAMMATORY RESPONSE IN MT-I1II-DEFICIENT MICE
as without effect in other brain areas, and MT-I1IIeficiency tended to decrease total zinc content, veryuch in agreement with what has been observed inT-III deficient mice (16).Regarding the second possibility, the available evi-
ence does not support an essential role of MT-I1II,ince no difference in the number of astrocytes, microg-ia, microglia/macrophages, or bone marrow-derived
yelomonocytes was seen between untreated MT1/1nd MT2/2 mice. Therefore, it seems more likely thatT-I1II deficiency in the present experiments is caus-
ng an abnormal brain inflammatory response throughperipheral mechanism, such as that seen in the bonearrow.We suspected that brain inflammatory response could
e attenuated in MT-deficient mice because the bonearrow could be severely affected in 6-AN-injectedice. MT1/1 mice injected with 6-AN show an intense
strogliosis and microgliosis (see above), but they alsohowed a significant increase of the number of bonearrow myeloid monocytes and macrophages com-
ared to that of untreated MT1/1 mice. This is likelyeflecting an increased proliferation of bone marrowyeloid cells in response to 6-AN toxicity. These bone-arrow-derived cells, together with resident microglial
ells, are considered the precursors of microglia/acrophages needed during inflammatory conditions
n CNS (3, 59). Not unexpectedly, the reactive responsef myeloid monocytes and macrophages in the bonearrow was significantly reduced in 6-AN-treatedT2/2 mice compared to that of 6-AN-injected MT1/1ice. Hence, our results suggest that MT-I1II defi-
iency dramatically impairs the brain inflammatoryesponse to the antimetabolite 6-AN, because the bone-arrow-derived monocytes and macrophages are not
roduced properly. This, in turn, severely impairs therain phagocyte response to astrocyte death, suggest-ng a major role of bone marrow-derived cells as therecursors of amoeboid brain macrophages and in
TABLE 1
Effect of 6-AN on Total Zinc Levels of MT1/1and MT2/2 Mice
MT1/1 MT2/2
Control 6-AN Control 6-AN
rain stem 14.95 6 0.73 13.77 6 0.76 15.47 6 1.23 13.92 6 0.47erebellum 20.71 6 1.90 19.67 6 0.56 17.19 6 1.42 16.95 6 1.85emaining
brain 20.38 6 0.58 21.12 6 0.88 19.34 6 0.50 19.74 6 0.52
Note. The tissues were acid-digested and zinc levels measured bytomic absorption spectrophotometry. Results are µg Zn/g wet tissue,eans 6 SE (n 5 5 or 6).
hagocytic reactivity of the CNS in this particular
odel of brain damage. That astrocytosis also has aole is obvious. Microglia/macrophages secrete proin-ammatory cytokines, such as interleukins, interfer-ns, tumor necrosis factors, and other growth factors,hich are instrumental in astrocyte activation, induc-
ion of cell adhesion molecule expression, neovascular-zation, wound repair, and recruitment of lymphocytes6, 20, 22, 24, 32). The reduced number of reactivestrocytes may lead to the reduced activity of GM-CSF,hich is a microglial and macrophage mitogen (21, 23,8). Thus, the 6-AN-induced supression of microglia/acrophages may be further enhanced by the supres-
ion of reactive astrocytes.The bone marrow suppression seen in 6-AN-injectedT2/2 mice could be mediated by at least two possibleechanisms when considering the putative functions
f MT-I1II. First, the absence of important zinc-inding proteins as MT-I1II, could render the bonearrow with an uncontrolled zinc metabolism, and, as
iscussed above, this metal has been shown to causeome toxicity to cultured cells (37). Additionally, MT-1II are significant antioxidant proteins, and thereforen increased oxidative stress and oxidative damageould be responsible for the bone marrow impairmenteen in 6-AN-injected MT2/2 mice. 6-AN is a niacinntagonist which induces death of rapidly dividingells, such as those of the bone marrow (27). Dying cellsay release substantial amounts of both heavy metals
nd reactive oxygen intermediates, which in excess areapable of inducing bystander tissue damage (55). Inddition, activated macrophages release cytotoxic super-xide and reactive oxygen intermediates (7, 13, 25). Noifferences in histochemically detectable zinc werebserved in the bone marrow of untreated and 6-AN-njected MT1/1 and MT2/2 mice, suggesting that it ishe antioxidant role of MT-I1II which is the predomi-ant factor responsible for the impaired bone marrow
unction and, in turn, impaired brain inflammatoryesponse.The CNS expresses MT-III in addition to MT-I1II, inrather specific fashion (36). Considerable uncertainty
till exists about the regulation and putative functionsf this CNS isoform (5, 31), and the MT-I1II- or
MT-III-deficient mice provide an essential tool to under-stand the CNS needs of the different MT isoforms. Asstated above, in normal MT1/1 mice 6-AN injectionproduced a very significant upregulation of MT-I1II inastrocytes surrounding the lesioned gray matter areasof the brain stem. In contrast, MT-III was more gener-ally upregulated, since increased immunoreactivitywas observed in areas comprising both astrocytes andmicroglia/macrophages. However, the increase wassmall compared to that of the MT-I1II isoforms. Such amild upregulation of MT-III has also been shown in a
number of animal models of brain damage (5, 31). In
cm6fiwarIbsmvtdpmnnlnsdab
Mttrdecre
DT(S
1
1
1
1
1
1
1
1
1
1
2
2
2
2
162 PENKOWA ET AL.
ontrast to the modest response observed in MT1/1ice, a very robust upregulation was observed in
-AN-injected MT2/2 mice, which indicates for therst time a sort of compensatory response of MT-IIIhen its normal counterparts MT-I1II are not avail-ble. This is in principle consistent with the compa-able metal binding properties of MT-III versus MT-1II (57), should the role of these MT isoforms beasically related to cellular zinc metabolism as recentlyuggested (42). Thus, one might speculate about com-on regulatory mechanisms. However, the factors in-
olved in MT-III regulation are mostly unknown, andhe available evidence suggests that they are ratherifferent from those of MT-I1II (5, 31). Moreover, areliminary experiment indicates that MT-III-deficientice do not express more MT-I1II protein than that oformal mice following a focal freeze lesion to theeocortex (unpublished observations).Also, MT-I mRNA
evels are not affected in MT-III-deficient mice underormal conditions (16). Thus, it seems as if the compen-atory upregulation of MT-III in 6-AN-injected MT-I1II-eficient mice is not a bidirectional mechanism. Inddition, MT-III has been suggested to have differentiological roles from those of MT-I1II (48, 63).In summary, the present results demonstrate thatT-I1II are essential for mounting a normal inflamma-
ory response in the brain following 6-AN administra-ion, presumably because of an abnormal bone marrowesponse. The overexpression of MT-III in MT-I1II-eficient mice suggest that MT-III is compensating forither the zinc homeostasis and/or the antioxidantapacity of the brain during MT-I1II deficiency, but ouresults also indicate that different regulatory factorsxist for the three MT isoforms.
ACKNOWLEDGMENTS
These studies were supported by Warwara Larsens Fond (M.P.),ir. Leo Nielsens Fond (M.P.) and Novo Nordisk Fonden (M.P. and.M.), and CICYT SAF96-0189 and Fundacion ‘‘La Caixa’’ 97/102-00J.H.). The excellent technical assistance of Hanne Hadberg and Keldtub is gratefully acknowledged.
REFERENCES
1. Amat, J., H. Ishiguro, K. Nakamura, and W. Norton. 1996.Pheotypic diversity and kinetics of proliferating microglia andastrocytes following cortical stab wounds. Glia 16: 368–382.
2. Andersson, P., V. Perry, and S. Gordon. 1992. The acute inflam-matory response to lipopolysaccharide in CNS parenchymadiffers from that in other body tissues. Neuroscience 48: 169–186.
3. Andersson, P., V. Perry, and S. Gordon. 1991. The kinetics andmorphological characteristics of the macrophage–microglial re-sponse to kaininc acid-induced neuronal degeneration. Neurosci-ence 42: 201–214.
4. Anezaki, T., H. Ishiguro, I. Hozumi, T. Inuzuka, M. Hiraiwa, H. 2
Kobayashi, T. Yuguchi, A. Wanaka, Y. Uda, T. Miyatake, K.Yamada, M. Tohyama, and S. Tsuji. 1995. Expression of growthinhibitory factor (GIF) in normal and injured rat brains. Neuro-chem. Int. 27: 89–94.
5. Aschner, M. 1996. The functional significance of brain metallo-thioneins. Faseb J 10: 1129–1136.
6. Balasingam, V., T. Tejada-Berges, E. Wright, R. Bouckova, andV. Yong. 1994. Reactive astrogliosis in the neonatal mouse brainand its modulation by cytokines. J. Neurosci. 14: 846–856.
7. Banati, R., J. Gehrmann, P. Schubert, and G. Kreutzberg. 1993.Cytotoxicity of microglia. Glia 7: 111–118.
8. Banks, W. A., A. J. Kastin, and R. D. Broadwell. 1995. Passage ofcytokines across the blood-brain barrier. Neuroimmunomodula-tion 2: 241–248.
9. Belloso, E., J. Hernandez, M. Giralt, P. Kille, and J. Hidalgo.1996. Effect of stress on mouse and rat brain metallothionein Iand III mRNA levels. Neuroendocrinology 64: 430–439.
0. Broadwell, R. D. 1992. Pathways into, through, and around thefluid-brain barriers. NIDA Res. Monogr. 120: 230–258.
1. Broadwell, R. D., and M. V. Sofroniew. 1993. Serum proteinsbypass the blood-brain fluid barriers for extracellular entry tothe central nervous system. Exp. Neurol. 120: 245–263.
2. Carrasco, J., J. Hernandez, B. Gonzalez, I. Campbell, and J.Hidalgo. 1998. Localization of metallothionein-I and -III expres-sion in the CNS of transgenic mice with astrocyte-targetedexpression of interleukin 6. Exp. Neurol., 153: 184–194.
3. Colton, A., and D. Gilbert. 1987. Production of superoxide by aCNS macrophage, the microglia. FEBS Lett. 223: 284–288.
4. Dalton, T., T. L. Pazdernik, J. Wagner, F. Samson, and G. K.Andrews. 1995. Temporalspatial patterns of expression of metal-lothionein-I and -III and other stress related genes in rat brainafter kainic acid-induced seizures. Neurochem. Int. 27: 59–71.
5. Duguid, J. R., C. W. Bohmont, N. G. Liu, and W. W. Tourtellotte.1989. Changes in brain gene expression shared by scrapie andAlzheimer disease. Proc. Natl. Acad. Sci. USA 86: 7260–7264.
6. Erickson, J. C., G. Hollopeter, S. A. Thomas, G. J. Froelick, andR. D. Palmiter. 1997. Disruption of the metallothionein-III genein mice: Analysis of brain zinc, behavior, and neuron vulnerabil-ity to metals, aging, and seizures. J. Neurosci. 17: 1271–1281.
7. Erickson, J. C., A. K. Sewell, L. T. Jensen, D. R. Winge, and R. D.Palmiter. 1994. Enhanced neurotrophic activity in Alzheimer’sdisease cortex is not associated with down-regulation of metallo-thionein-III (GIF). Brain. Res. 649: 297–304.
8. Gasull, T., M. Giralt, J. Hernandez, P. Martinez, I. Bremner, andJ. Hidalgo. 1994. Regulation of metallothionein concentrationsin rat brain: Effect of glucocorticoids, zinc, copper, and endo-toxin. Am. J. Physiol. 266: E760–E767.
9. Gasull, T., D. V. Rebollo, B. Romero, and J. Hidalgo. 1993.Development of a competitive double antibody radioimmunoas-say for rat metallothionein. J. Immunoassay 14: 209–225.
0. Gehrman, J., Y. Matsumoto, and G. Kreutzberg. 1995. Microg-lia: Intrinsec immunoeffector cell of the brain. Brain Res. Rev.20: 269–287.
1. Gehrmann, J. 1995. Colony-stimulating factors regulate pro-grammed cell death of rat microglia/brain macrophages in vitro.J. Neuroimmunol. 63: 55–61.
2. Giulian, D., J. Chen, J. Ingemann, J. George, and M. Noponen.1989. The role of mononuclear phagocytes in wound healingafter traumatic injury to adult mammalian brain. J. Neurosci.12: 4416–4429.
3. Giulian, D., and J. Ingemann. 1988. Colony-stimulating factorsas promoters of ameboid microglia. J. Neurosci. 8: 4707–4717.
4. Giulian, D., J. Li, X. Li, J. George, and P. Rutecki. 1994. The

2
2
2
2
2
3
3
3
3
3
3
3
3
3
3
4
4
4
4
4
4
4
4
4
4
5
5
5
5
5
5
5
5
5
5
6
6
6
163INFLAMMATORY RESPONSE IN MT-I1II-DEFICIENT MICE
impact of microglia-derived cytokines upon gliosis in the CNS.Dev. Neurosci. 16: 128–136.
5. Giulian, D., and C. Robertson. 1990. Inhibition of mononuclearphagocytes reduces ischemic injury in the spinal cord. Ann.Neurol. 27: 33–42.
6. Guillemin, G., F. D. Boussin, R. L. G. R., J. Croitoru, H. Coffigny,and D. Dormont. 1996. Granulocyte macrophage colony stimu-lates in vitro proliferation of astrocytes derived from simianmature brains. Glia 16: 71–80.
7. Haghignat, N., and D. McCandless. 1997. Effect of 6-aminonic-otinamide on metabolism of astrocytes and C6-glioma cells.Metabol. Brain Dis. 12: 29–45.
8. Hansen, P. B., M. Penkowa, and H. Joensen. 1998. Hemopoieticgrowth factors, G-CSF, GM-CSF, Epo and IL-3 for the treatmentof myelodysplastic syndromes. Leukemia Lymphoma 28.
9. Hernandez, J., A. Molinero, I. L. Campbell, and J. Hidalgo.1997. Transgenic expression of interleukin 6 in the centralnervous system regulates brain metallothionein-I and -III ex-pression in mice. Mol. Brain Res. 48: 125–131.
0. Hidalgo, J., M. Borras, J. S. Garvey, and A. Armario. 1990. Liver,brain, and heart metallothionein induction by stress. J. Neuro-chem. 55: 651–654.
1. Hidalgo, J., B. Castellano, and I. L. Campbell. 1997. Regulationof brain metallothioneins. Curr. Topics Neurochem. 1: 1–26.
2. Hopkins, S., and N. Rothwell. 1995. Cytokines and the nervoussystem. I. Expression and recognition. Trends Neurosci. 18:83–88.
3. Hozumi, I., T. Inuzuka, M. Hiraiwa, Y. Uchida, T. Anezaki, H.Ishiguro, H. Kobayashi, Y. Uda, T. Miyatake, and S. Tsuji. 1995.Changes of growth inhibitory factor after stab wounds in ratbrain. Brain Res. 688: 143–148.
4. Hozumi, I., T. Inuzuka, H. Ishiguro, M. Hiraiwa, Y. Uchida, andS. Tsuji. 1996. Immunoreactivity of growth inhibitory factor innormal rat brain and after stab wounds—An immunocytochemi-cal study using confocal laser scan microscope. Brain Res. 741:197–204.
5. Inuzuka, T., I. Hozumi, A. Tamura, M. Hiraiwa, and S. Tsuji.1996. Patterns of growth inhibitory factor (GIF) and glialfibrillary acidic protein relative level changes differ followingleft middle cerebral artery occlusion in rats. Brain Res. 709:151–153.
6. Kobayashi, H., Y. Uchida, Y. Ihara, K. Nakajima, S. Kohsaka, T.Miyatake, and S. Tsuji. 1993. Molecular cloning of rat growthinhibitory factor cDNA and the expression in the central ner-vous system. Mol. Brain Res. 19: 188–194.
7. Koh, J., and D. Choi. 1994. Zinc toxicity in cultured corticalneurons: Involvement of N-methyl-D-aspartate receptors. Neuro-science 60: 1049–1057.
8. Kreutzberg. 1996. Microglia: A sensor for pathological events inthe CNS. Trends Neurosci. 19: 312–318.
9. Krum, J. 1995. Age-dependent susceptibility of CNS glial popu-lations in situ to the antimetabolite 6-aminonicotinamide. Mol.Chem. Neuropathol. 26: 79–94.
0. Krum, J. M. 1994. Experimental gliopathy in adult rat CNS:Effect on the blood–spinal cord barrier. Glia 11: 354–366.
1. Malipiero, U., K. Frei, and A. Fontana. 1990. Production ofhemopoietic colony-stimulating factors by astrocytes. J. Immu-nol. 144: 3816–3821.
2. Maret, W., and B. Vallee. 1998. Thiolate ligands in metallothio-nein confer redox activity on zinc clusters. Proc. Natl. Acad. Sci.USA 95: 3478–3482.
3. Masters, B. A., E. J. Kelly, C. J. Quaife, R. L. Brinster, and R. D.
Palmiter. 1994. Targeted disruption of metallothionein I and IIgenes increases sensitivity to cadmium. Proc. Natl. Acad. Sci.USA 91: 584–588.
4. Masters, B. A., C. J. Quaife, J. C. Erickson, E. J. Kelly, G. J.Froelick, B. P. Zambrowicz, R. L. Brinster, and R. D. Palmiter.1994. Metallothionein III is expressed in neurons that sequesterzinc in synaptic vesicles. J. Neurosci. 14: 5844–5857.
5. Moos, T. 1993. Simultaneous application of Timm’s sulphidesilver method and immunofluorescence histochemistry. J. Neu-rosci. Methods 48: 149–156.
6. Moos, T., and P. Hoyer. 1996. Detection of plasma proteins inCNS: Conspicuous influence of tissue-processing parametersand the utilization of serum for blocking non-specific reactions.J. Histochem. Cytochem. 44: 591–603.
7. Neal, J. W., S. K. Singhrao, B. Jasani, and G. R. Newman. 1996.Immunocytochemically detectable metallothionein is expressedby astrocytes in the ischaemic human brain. Neuropathol. Appl.Neurobiol. 22: 243–247.
8. Palmiter, R. D. 1995. Constitutive expression of metallothionein-III (MT-III), but not MT-I, inhibits growth when cells becomezinc deficient. Toxicol. Appl. Pharmacol. 135: 139–146.
9. Palmiter, R. D., S. D. Findley, T. E. Whitmore, and D. M.Durnam. 1992. MT-III, a brain-specific member of the metallo-thionein gene family. Proc. Natl. Acad. Sci. USA 89: 6333–6337.
0. Penkowa, M., J. Hidalgo, and T. Moos. 1997. Increased astro-cytic expression of metallothioneins I 1 II in brain stem of adultrats treated with 6-aminonicotinamide. Brain Res. 774: 256–259.
1. Penkowa, M., and T. Moos. 1995. Disruption of the blood–braininterface in neonatal rat neocortex induces a transient expres-sion of metallothionein in reactive astrocytes. Glia 13: 217–227.
2. Perry, V., M. Bell, H. Brown, and M. Matyszak. 1995. Inflamma-tion in the nervous system. Curr. Opinion Neurobiol. 5: 636–641.
3. Raivich, G., J. Gehrmann, and G. W. Kreutzberg. 1991. Increaseof macrophage colony-stimulating factor and granulocyte–macrophage colony-stimulating factor receptors in the regener-ating rat facial nucleus. J. Neurosci. Res. 30: 682–686.
4. Ridet, J. L., A. Malhotra, and F. Gage. 1997. Reactive astrocytes:Cellular and molecular cues to biological function. TrendsNeurosci. 20: 570–577.
5. Sato, M., and I. Bremner. 1993. Oxygen free radicals andmetallothionein. Free Radic. Biol. Med. 14: 325–337.
6. Sawada, M., Y. Itoh, A. Suzumura, and T. Marunouchi. 1993.Expression of cytokine receptors in cultured neuronal and glialcells. Neurosci. Lett. 160: 131–134.
7. Sewell, A. K., L. T. Jensen, J. C. Erickson, R. D. Palmiter, andD. R. Winge. 1995. Bioactivity of metallothionein-3 correlateswith its novel beta domain sequence rather than metal bindingproperties. Biochemistry 34: 4740–4747.
8. Sillevis Smitt, P. A., H. G. Blaauwgeers, D. Troost, and J. M. deJong. 1992. Metallothionein immunoreactivity is increased inthe spinal cord of patients with amyotrophic lateral sclerosis.Neurosci. Lett. 144: 107–110.
9. Stevens, A., and M. Bahr. 1993. Origin of macrophages incentral nervous tissue. J. Neurol. Sci. 118: 117–122.
0. Suzuki, K., K. Nakajima, U. Kawaharada, K. Uehara, F. Hara,N. Otaki, M. Kimura, and Y. Tamura. 1992. Metallothionein inthe human brain. Acta Histochem. Cytochem. 25: 617–622.
1. Suzumura, A., M. Sawada, and T. Marunouchi. 1996. Selectiveinduction of interleukin-6 in mouse microglia by granulocyte-macrophage colony-stimulating factor. Brain Res. 713: 192–198.
2. Thornalley, P. J., and M. Vasak. 1985. Possible role for metallo-
thionein in protection against radiation-induced oxidative stress:
6
6
6
164 PENKOWA ET AL.
Kinetics and mechanism of its reaction with superoxide andhydroxyl radicals. Biochim. Biophys. Acta 827: 36–44.
3. Uchida, Y., K. Takio, K. Titani, Y. Ihara, and M. Tomonaga. 1991.The growth inhibitory factor that is deficient in the Alzheimer’sdisease brain is a 68 amino acid metallothionein-like protein.Neuron 7: 337–347.
4. Yuguchi, T., E. Kohmura, K. Yamada, T. Sakaki, T. Yamashita,
H. Otsuki, K. Kataoka, S. Tsuji, and T. Hayakawa. 1995.Expression of growth inhibitory factor mRNA following corticalinjury. J. Neurotrauma 12: 299–306.
5. Yuguchi, T., E. Kohmura, K. Yamada, T. Sakaki, T. Yama-shita, H. Otsuki, A. Wanaka, M. Tohyama, S. Tsuji, and T.Hayakawa. 1995. Changes in growth inhibitory factormRNA expression compared with those in c-jun mRNA expres-sion following facial nerve transection. Mol. Brain Res. 28:
181–185.Copyright © 2022 FDOKUMEN




















