
2. Genetically modified food crops and their contribution to ...
19
2. Genetically modified food crops and their contribution to human nutrition and food quality Howarth E. Bouis a *, Bruce M. Chassy b and James O. Ochanda c a International Food Policy Research Institute, 2033 K Street, NW, Washington DC 20006 USA (fax: +1-202- 467-4439; e-mail: [email protected]) b Biotechnology Center, University of Illinois at Urbana-Champaign, 238 NSRC, 1101 West Peabody Drive, Urbana, IL 61801, USA c Deparment of Biochemistry, College of Biological & Physical Sciences, University of Nairobi, PO Box 30197 Nairobi, Kenya 2.1. Introduction This chapter discusses the potential of biotechnology to improve the health and nutrition of consumers in developing countries. In the relatively wealthy countries of Europe, North America and elsewhere, consumers spend perhaps 10% of their income on food. For the most part consumers in developed countries are free of classical nutrient deficiencies, although over-consump- tion is a problem for some. Also in relatively wealthy countries there is, in general, good access to affordable medical care to meet health needs and most consumers in rich countries have access to a relatively inexpensive supply of safe and healthy food. In these settings, the possibility that biotechnology might reduce the price of food or make food more beneficial to health is a rela- tively minor concern. Rather, public debate about genetically modified foods (GMFs) appears to have focused on the potential for harm to either the environ- ment or health without a clear definition of benefit to the consumer. The situation, of course, is quite different in poor countries where malnutrition and ill health are frequent. Poor consumers typically spend 70% of their incomes on food, and diets consist primarily of staple foods, which lack the vitamins, minerals and, very likely, other food components necessary to sustain good health and minimise the risk of adult onset diet-related chronic diseases. In addition, low incomes typically preclude the poor from access to adequate health care. There are three broad ways that biotechnology may benefit consumers in developing countries. First, bio- technology offers a powerful, new tool to improve crop productivity, both by making conventional breeding faster and more efficient and, more controversially, by the insertion of novel genes in a crop species, by use of transgenic methods. This includes the ability to bring new lands with unfavourable growing environments, such as those with high saline soils, under production. Second, pesticide applications may be reduced through adoption of Bt-containing crops (i.e. crops with insect resistance owing to the introduction of the Bacillus thuringiensis (Bt) gene encoding an insect-specific toxin). Reducing pesticide use can improve the health of farmers, in addition to lowering input costs. Third, transgenic methods may be used to improve the micro- nutrient content and/or bioavailability of commonly eaten foods in developing countries. The first pathway, that is improving crop productiv- ity, is associated with an often-asked question—Does feeding the world depend on the use of biotechnol- ogy?—The answer largely depends on perceptions of the 0924-2244/03/$ - see front matter # 2003 Published by Elsevier Ltd. doi:10.1016/S0924-2244(03)00073-6 Trends in Food Science & Technology 14 (2003) 191–209 * Corresponding author.
-
Upload
khangminh22 -
Category
Documents
-
view
5 -
download
0
Transcript of 2. Genetically modified food crops and their contribution to ...

2. Geneticallymodified food
crops and theircontribution to
human nutritionand food quality
Howarth E. Bouisa*, BruceM. Chassyb and James
O. Ochandac
aInternational Food Policy Research Institute, 2033 KStreet, NW, Washington DC 20006 USA (fax: +1-202-
467-4439; e-mail: [email protected])bBiotechnology Center, University of Illinois at
Urbana-Champaign, 238 NSRC, 1101 West PeabodyDrive, Urbana, IL 61801, USA
cDeparment of Biochemistry, College of Biological &Physical Sciences, University of Nairobi, PO Box
30197 Nairobi, Kenya
2.1. IntroductionThis chapter discusses the potential of biotechnology
to improve the health and nutrition of consumers indeveloping countries. In the relatively wealthy countriesof Europe, North America and elsewhere, consumersspend perhaps 10% of their income on food. For themost part consumers in developed countries are free of
classical nutrient deficiencies, although over-consump-tion is a problem for some. Also in relatively wealthycountries there is, in general, good access to affordablemedical care to meet health needs and most consumersin rich countries have access to a relatively inexpensivesupply of safe and healthy food. In these settings, thepossibility that biotechnology might reduce the price offood or make food more beneficial to health is a rela-tively minor concern. Rather, public debate aboutgenetically modified foods (GMFs) appears to havefocused on the potential for harm to either the environ-ment or health without a clear definition of benefit tothe consumer.The situation, of course, is quite different in poor
countries where malnutrition and ill health are frequent.Poor consumers typically spend 70% of their incomeson food, and diets consist primarily of staple foods,which lack the vitamins, minerals and, very likely, otherfood components necessary to sustain good health andminimise the risk of adult onset diet-related chronicdiseases. In addition, low incomes typically preclude thepoor from access to adequate health care.There are three broad ways that biotechnology may
benefit consumers in developing countries. First, bio-technology offers a powerful, new tool to improve cropproductivity, both by making conventional breedingfaster and more efficient and, more controversially, bythe insertion of novel genes in a crop species, by use oftransgenic methods. This includes the ability to bringnew lands with unfavourable growing environments,such as those with high saline soils, under production.Second, pesticide applications may be reduced throughadoption of Bt-containing crops (i.e. crops with insectresistance owing to the introduction of the Bacillusthuringiensis (Bt) gene encoding an insect-specifictoxin). Reducing pesticide use can improve the health offarmers, in addition to lowering input costs. Third,transgenic methods may be used to improve the micro-nutrient content and/or bioavailability of commonlyeaten foods in developing countries.The first pathway, that is improving crop productiv-
ity, is associated with an often-asked question—Doesfeeding the world depend on the use of biotechnol-ogy?—The answer largely depends on perceptions of the
0924-2244/03/$ - see front matter # 2003 Published by Elsevier Ltd.doi:10.1016/S0924-2244(03)00073-6
Trends in Food Science & Technology 14 (2003) 191–209
* Corresponding author.

current situation. Countries around the world arealready using most of the arable land available foragriculture. As populations continue to grow, increasingthe supply of food to meet future world food needs willdepend on increasing yields. This, in turn, requiresinvestments in agricultural research to increase the yieldpotentials of specific food crops as well as fish and live-stock production.Conventional breeding of cereals has been especially
successful over the past three decades. Through researchand extension efforts associated with the ‘Green Revo-lution’, the rate of increase of production of rice, wheat,and maize in developing countries has outpaced popu-lation growth and other factors associated withincreased demand (for example higher householdincomes), such that the inflation-adjusted prices of riceand wheat fell by 30–40% between 1970 and 1997(World Bank, 1990–1999).1
World supplies and prices of cereals are inter-con-nected through international trade. Advances in con-ventional breeding in Australia, Europe, and NorthAmerica have also contributed to falling cereal prices.Continued low prices of cereals are likely as long asinvestments in agricultural research continue, althougha worrisome trend is that funds for public research inagriculture have been declining (Rosegrant, Paisner,Meijer, & Witcover, 2001).It is often said that there is enough food in the world
to feed everyone. Except for some pockets of extremepoverty and dislocations owing to natural disasters andcivil strife, this is largely the case for cereals. Cereals(and roots and tubers) provide the cheapest sourcesof energy to meet requirements for poor populations.The availability of affordable staple foods preventsexplicit hunger, as defined narrowly in terms of energyrequirements.However, a range of non-staple foods, such as animal
and fish products, fruits, pulses, and vegetables, arefoods rich in bioavailable vitamins and minerals ormicronutrients and possibly other food components,which are necessary for good health and a productivelife. As discussed in more detail below, micronutrientundernutrition is widespread in poor countries, affectingmore than one-half of the population in the developingworld. This situation is due primarily to diets of lownutritional quality. The poor want to eat largeramounts of non-staple foods, and most often thesefoods are available in local markets. However, suchnon-staple foods are simply too expensive to be eaten bythe poor in large quantities owing to a combination ofhigh prices and low purchasing power. The inflation-adjusted prices of non-staple foods have not been
declining; in some cases they have been increasing overthe past 30 years.Because of poor dietary quality and consequent
micronutrient malnutrition, some people, especiallychildren and their mothers who have higher require-ments for vitamins and minerals, have higher mortality,become sick more often, have their cognitive abilitiescompromised for a lifetime, and are less productivemembers of the workforce than might otherwise havebeen the case. Their quality of life and aggregate eco-nomic growth are unnecessarily compromised. To makematters worse, the adverse synergy between under-nutrition and infection results in nutrition requirementsabove those that characterise the needs of well-nour-ished populations.In developing countries demand for non-staple foods
is growing faster than demand for cereals, as ever-richerhouseholds purchase more non-staple foods than cer-eals, with consequent pressure on non-staple food pricesto increase. In addition, increasing the productivity ofnon-staple foods through agricultural research (and solowering prices) is much more expensive than increasingthe productivity of staple foods, because of the far lar-ger number of non-staple foods involved.From this perspective, then, the world is already fail-
ing to provide an adequate supply of non-staple foodsfor a large proportion of its population, and prospectsare somewhat bleak for short- to medium-termimprovements. Biotechnology affords a powerful newtool to help increase the supply of food produced in theworld and redress, in part, situations characterised bypoor dietary quality.Chapter 3 discusses ways, including reduced use of
pesticides (i.e. the second pathway, above), in whichbiotechnology can provide breakthroughs in improvingthe productivity of crops. However, it is well beyond thescope of this special issue to speculate on the rate ofgrowth of world food supplies with and without the useof biotechnology.A third pathway through which biotechnology can
improve the nutrition and health of consumers indeveloping countries is through increasing the vitaminand mineral content and bioavailability of staples andother foods and/or reducing allergens, an approachwhich is focused on in this chapter. It must be empha-sised that none of the products discussed in this chapterhas been introduced commercially, though they are invarious stages of research and development. Thus,importantly, both the efficacy and effectiveness of thispathway requires documentation.Although it is obviously important to provide an
account of the range of specific efforts being developedto improve the nutritional content and health-enhancingproperties of particular foods, it is also necessary toplace these activities in the context of alternative inter-ventions (i.e. interventions that do not involve the use of
1 World Bank: Commodity Markets and the Developing Countries;Global Commodity Markets.
192 H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209

biotechnology) that might solve these malnutrition andhealth problems in other ways. This is accomplished by(i) examining the relative costs of nutrient supple-mentation and commercial nutrient fortification pro-grams that are already being implemented to reducemicronutrient malnutrition and (ii) examining the recenthistory of attempts to improve the micronutrient con-tent of staple food crops—efforts that have involveduse both of biotechnology and conventional plantbreeding. Recent developments in efforts to breed iron-and beta-carotene-dense rices are described in thischapter as a concrete example, illustrating several keygeneric issues associated with the use of biotechnologyto breed for characteristics that have direct benefits forconsumers.It is important to keep in mind that the final, sus-
tainable solution to micronutrient malnutrition indeveloping countries is a substantial improvement indietary quality by higher consumption of pulses, fruits,vegetables, fish, and animal products, which the poorneed and already desire, but cannot presently afford.Realisation of this goal of high quality diets will require(i) substantial investments by farmers, private busi-nesses and public agencies to build the infrastructure toproduce and bring to market the requisite supply ofnon-staple foods, (ii) sound government policies to sti-mulate agricultural and economic growth, (iii) con-siderable increases in the incomes of the poor during thecourse of economic development over many decades, or(iv) other strategies (e.g. subsidies) designed to enhancethe accessibility of these products to wider populations.Meanwhile breeding staple foods that are dense inminerals and vitamins can provide a low-cost, sustain-able, although as yet unproven strategy for reducingexisting levels of specific micronutrient undernutrition.Sections 2.2–2.5 (below) discuss, in some detail, iron-
and beta-carotene-dense rice, as an example of thedevelopment of a GMF crop of potential importance indeveloping countries. Section 2.6 describes the potentialof transgene technologies to enhance nutritional andquality factors of food crops in general; this is impor-tant in both developing and developed countries,though perhaps has more immediate relevance in thelatter.
2.2. Potential contribution of GM technology: acase study on breeding of iron- and beta-carotene-dense rices to help alleviate malnutrition
2.2.1. Background to the problem of micronutrientmalnutrition in poor countries, its economic costsand costs of present interventions to address theproblem
It is only relatively recently that nutritionists workingin developing countries have demonstrated conclusivelythat, during times of relative economic and politicalstability, many children and adults, particularly womenin their child-bearing years, suffer more from a lack ofessential vitamins and minerals in their diets than froma lack of calories. People, for the most part, are notaware that their diets lack such trace nutrients and so donot associate these deficiencies with listlessness, pooreyesight, impaired cognitive development and physicalgrowth, more frequent and severe bouts of illness, andhigh mortality. For this reason, the general problem ofpoor dietary quality has been dubbed ‘hidden hunger’.It is useful to summarise very briefly the scale and
consequences of micronutrient malnutrition in develop-ing countries. It is estimated that over 3 billion people indeveloping countries are iron-deficient (AdministrativeCommittee on Co-ordination, Subcommittee on Nutri-tion, & International Food Policy Research Institute,2000). The problem for women and children is moresevere because of their greater physiological need foriron. In poor countries, more than half of pregnantwomen and more than 40% of non-pregnant womenand preschool children are anaemic. Iron deficienciesduring childhood impair physical growth, mental devel-opment and learning capacity. In adults, iron deficiencyreduces the capacity to do physical labour. Severe irondeficiency is a leading contributor to death amongwomen during childbirth, owing to low haemoglobinlevels in the light of physiological blood loss.Globally, about 3 million children of preschool age
have visible eye damage owing to a vitamin A defi-ciency. Annually, an estimated 250 000–500 000 pre-school children go blind from this deficiency and abouttwo-thirds of these children die within months of goingblind. Estimates of the subclinical prevalence of vitaminA deficiency range from 100 to 250 million. A numberof clinical trials in developing countries have shown thatvitamin A supplementation, as capsules distributedbiannually in high doses, as weekly distributed lowdoses (consistent with levels attainable through food) oras a fortified product in diets can reduce mortality ratesamong preschool children on average by 23% (Beatonet al., 1993).Deficiencies in several other micronutrients, in parti-
cular zinc, may also be widespread, with equally seriousconsequences for health. However, because for some ofthese micronutrients, specific indicators are lacking toscreen for deficiencies, they have not received as muchattention as iron, vitamin A and iodine deficiencies.The World Bank (1994) estimates that at the levels of
micronutrient malnutrition existing in South Asia, 5%of gross national product is lost each year due to defi-ciencies in the intakes of just three nutrients, namelyiron, vitamin A and iodine. For each 50 million in
H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209 193

population, this translates into an economic loss of US$1 billion per year.Some general cost calculations can be made for inter-
ventions to relieve iron and vitamin A deficiencies bysupplementation and fortification in South Asia, whichhas a total population of roughly 1.25 billion people.The costs of vitamin A pills themselves are low: anoften-quoted cost of vitamin A supplementation, whichincludes the costs of delivery, is US $0.50 per person peryear (i.e. US $0.25 per capsule; World Bank, 1994). Ifone in every 12.5 people in South Asia were to receivesupplements (i.e. 100 million people in total), this wouldcost US $50 million per year, or US $500 million over10 years. An often-quoted cost of iron fortification isUS $0.12 per person per year (World Bank, 1994). If aparticular food vehicle fortified with iron were toreach 33% of the total (but untargeted) population inSouth Asia (i.e. 412 million people), the total costwould, again, be US $50 million annually, or US$500 million over 10 years. In absolute terms, thesenutrient-specific interventions may seem to be largeamounts of money, but they are very worthwhileinvestments and actually represent quite small percen-tages of the total economic activity of the South Asianeconomies.The Micronutrient Initiative (MI)2 estimates that 1
billion vitamin A pills have been distributed worldwidesince the inception of vitamin A distribution pro-grammes up to the end of 2000 and that 500 millionsupplements are now given out annually.A conservative estimate of the annual, recurrent cost
of global vitamin A supplementation programmes is,therefore, US $125 million. Assuming that 75% of theworld’s need for vitamin A is met, recurrent annualestimated costs to meet all worldwide needs would beabout US $165 million, or $1.65 billion over a 10-yearperiod.Although benefit–cost ratios are quite high, supple-
mentation and fortification programmes must be sus-tained at more or less the same level of funding yearafter year in any given country. If investments are notsustained, benefits of course disappear.Investments in plant breeding research and dis-
semination are far lower (see Section 2.3.2) andpotentially long lasting. Benefits of agriculturalresearch at a central location can be leveragedthroughout the world and across time. Breeding forstaple plants with high bioavailable micronutrientcontent in their seeds, hereinafter referred to as ‘bio-fortification’, treats the underlying cause of micro-nutrient undernutrition. Although plant breeding caninvolve relatively long lead times of 8–10 years before
nutritious varieties can be developed and their adop-tion by farmers can be initiated, such a strategy is sus-tainable once breeding has been completed, and seedshave been disseminated and adopted by farmers. Bio-fortification has the potential to provide coverage forremote rural populations, which supplementation andfortification programs may not reach, and it inherentlytargets the poor who consume high levels of staplefoods and little else.However, efficacy and effectiveness trials and other
studies have yet to be undertaken to determine both thedegree to which nutritionally dense varieties canimprove micronutrient status and whether there areunintended negative effects. For example, untargetedapproaches designed to increase bioavailable dietaryiron substantially must be considered with care. Thedocumented risks of iron deficiency must be weighedagainst the potential for genetically determinedincreased risks of toxicity from iron overload, given therelatively common occurrence of heterozygosity forhaemachromatosis. Thus ‘permanent’ changes in thefood supply, which result in long term ‘high’ intakes ofiron should be considered in the light of the incomple-tely described consequences of relevant polymorphismsin genes that appear to play roles in the regulation ofiron metabolism. Such risks have been studied mostadequately in developed countries (Merryweather-Clarke, Pointon, Sherman, & Robson, 1997).However, according to an expert group convened to
consider micronutrient supplementation and fortifica-tion with iron, iron overload disorders and haemochro-matosis are rare, even in those populations of Europeanorigin most susceptible to them; therefore, the use offortification and supplementation as public healthinterventions for preventing and controlling iron defi-ciency should not be constrained (UNICEF/UNU/WHO/MI, 1999). It is safe to assume that the sameconclusions would apply to biofortification. If bioforti-fication eventually were to replace supplementation insome areas, there is the possibility that biofortificationmight lessen risks of overdosing, such as that recentlyreported in India, where toxicity associated with a massdistribution of a vitamin A concentrate to children wasthought, possibly, to have been the result of a changeddelivery routine (Sharma, 2001).
2.2.2. Improving the nutrient content andbioavailability of minerals and vitamins in staple foodcrops through biofortification
Breeding strategies
There are three breeding sub-strategies that may beapplied individually or in various combinations. Theseare (i) reducing the level of antinutrients in food staples,
2 A Decade of Progress, a Lifetime of Hope: The MicronutrientInitiative, 1990–2000, available [July 2003] at http://www.micronu-trient.org/frame_HTML/decade.pdf.
194 H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209

which inhibit the bioavailability of minerals and vita-mins, (ii) increasing the levels of nutrients and com-pounds that promote the bioavailability of minerals andvitamins, and (iii) increasing the mineral and vitamincontent. Experience with increasing the mineral andvitamin content of rice is discussed in some detail inSection 2.2.3.
Reducing antinutrients
A breeding strategy of lowering the level of anti-nutrients, such as phytic acid, which binds metals ingrain, has often been suggested as a way to increase thebioavailability of minerals already consumed.Phytin is the primary storage form of phosphorus in
most mature seeds and grains and is an importantcompound required for early seed germination andseedling growth (Welch, 1986; Wise, 1995). Phytin playsan important role in determining mineral reserves ofseeds and thus contributes to the viability and vigour ofthe seedling produced (Welch, 1993).Graham and Welch (1996) argue that selecting for
seed and grain crops with substantially lower phytincontent could have an unacceptable effect on produc-tion, especially in regions of the world where there aresoils of low phosphorus status and/or poor micro-nutrient fertility. Such attempts to lower the anti-nutrient content of seeds and grains significantly requirea major shift in seed or grain composition. Becausemost of the antinutrients known to occur in seeds andgrains are major organic constituents of these organs,they may play beneficial roles in plant growth and mayhave other unidentified roles in human health. There-fore, Graham and Welch argue against a breedingstrategy that attempts to increase iron bioavailability byreducing antinutrient content. Antinutrients may alsoreduce absorption of potentially toxic trace mineralssuch as cadmium and other heavy metals (McLaughlin,Parker, & Clarke, 1999).However, Raboy (1996) has developed low phytic
acid (or lpa) mutant varieties of maize, rice, and barley.The phytic acid content of lpa seeds is reduced by 50–80% compared with non-mutant seeds. The totalamount of phosphorus remains the same—phytic acid isreplaced by inorganic phosphorus, which does not binda range of trace minerals.3 These mutations typicallyhave little observable effect on other seed or plantcharacteristics. These mutants are presently being testedfor agronomic performance and effects on micro-nutrient status in humans.
A related strategy is to increase the level of phytase.Phytase can break down phytates, thus increasing bioa-vailability of minerals. Phytases have been used suc-cessfully in animal feeds (Nelson, Shieh, Wodzinski, &Ware, 1971; Rimbach & Pallauf, 1993; Yi, Kornegay, &Veit, 1996). Phytase is present in the endosperm of rice,for example, but is destroyed when rice is boiled foreating. Heat-stable phytase may be introduced in foodstaples through genetic transformation (Holm, Kris-tiansen, & Pedersen, 2002).
Increasing levels of compounds that improveabsorption and utilisation of nutrients
Certain amino acids, such as cysteine, enhance ironand/or zinc bioavailability (Hallberg, 1981). Theseamino acids occur in many staple foods but their con-centrations are lower than those found in meat pro-ducts. A modest increase in the concentrations ofthese amino acids in plant foods may have positiveeffects on iron and/or zinc bioavailability in humans.Iron and zinc occur only in micromolar amounts inplant foods, so only micromolar increases in theamounts of these amino acids may be required tocompensate for the negative effects of antinutrients oniron and zinc bioavailability. These amino acids are nor-mal constituents of plants, so relatively small increases intheir concentrations in plant tissues should not haveadverse consequences on plant growth (Graham &Welch, 1996).However, when manipulating amino acid content,
the net effects on the resultant product’s proteinquality should also be considered. The three mostcommon measures of protein quality are protein effi-ciency ratios, biological value and net protein utilisa-tion. These measures are expressed as quotients ofretained nitrogen (a constituent of protein) divided bythe total intake of protein nitrogen. Among thedeterminants of protein quality are amino acid pattern,absolute levels of protein intake and physiologicalrequirements for essential amino acids. Essential aminoacids are those that must be provided pre-formedbecause of an inability for their de novo synthesis.The higher the quality of a specific protein, themore efficiently it is utilised and the less is neededto meet protein requirements. Thus changes in pro-tein content or amino acid pattern of the resultantprotein mix can have significant effects on the effi-ciency of protein utilisation to meet nutritionalrequirements.An additional possibility is that as intakes of beta-
carotene (converted to vitamin A after ingestion) areincreased, the utilisation of absorbed iron may be pro-moted and vice-versa. That is, there are possible syner-gies between increasing intakes of these two nutrients
3 When such mutants are used as animal feeds, this also avoidswhat has become a serious pollution problem—excretion of unu-tilised phytic acid.
H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209 195

(Garcia-Casal et al., 1998). There is already consider-able evidence for synergism between vitamin A and zincintakes (Smith, 1996).
Feasibility
Will famers adopt nutrient-rich varieties?. Impact on
plant nutrition is a particular concern of agriculturalists,
whose first priority may be to raise the yield potential of
plants rather than to address human nutrition pro-
blems. If yield and profitability are compromised by
breeding for nutrient content, farmers will not want to
adopt nutritionally enhanced cultivars. However, rela-
tively recent research has shown that mineral-dense
seeds assist plants in resisting disease and other envir-
onmental stresses (Graham, Welch, & Bouis, 2001;
Welch & Graham 1999). More seedlings survive and
initial growth is more rapid. Ultimately, yields are
higher, particularly in trace mineral ‘deficient’ soils in
arid regions.4
Will breeding for micronutrient-dense seeds change theprocessing or consumer characteristics of staple foods?.
Mineral micronutrients comprise a tiny fraction of the
physical mass of a seed; for example, 5–10 mg/kg (ppm)
in milled rice, and as much as 100 mg/kg in micro-
nutrient-dense bean seeds. It is not known whether such
small amounts will alter the appearance, taste, texture,
or cooking quality of foods. The dissemination strategy
for trace minerals, then, would be to include the
mineral-density trait in as many varietal releases as
possible, in order to benefit a high proportion of the
population, without having to rely on behavioural
change as a condition for success. This would be much
the same approach as adding fluoride to drinking water
in developed countries. As with the example of fluoride,
information on increased nutrient content would be
made publicly available.
However, increasing levels of beta-carotene will turnvarieties from white or light colours of yellow to darkyellow and orange. Often white varieties are much pre-ferred by consumers (e.g. milled rice, wheat flour, maize,cassava). Major nutrition education programs will prob-
ably be necessary to encourage consumers to switch tomore nutritious varieties. However, if these nutritioneducation programs are successful, the yellow-orangecolour will mark the more nutritious varieties from theless nutritious, and a disadvantage will have been turnedinto an advantage (Hagenimana & Low, 2000).
2.2.3. Increasing micronutrient content in rice
Conventional breedingIn a survey of traditional and modern varieties of rice,
Graham, Senadhira, Beebe, Iglesias, and Ortiz-Mon-asterio (1999) reported that iron density in unmilled ricevaried from 7 to 24 mg/kg (ppm) and zinc density from16 to 58 mg/kg. Nearly all the widely grown greenrevolution varieties were similar, at about 12 and 22 mg/kg for iron and zinc, respectively. The best lines dis-covered in the survey of the existing germplasm collec-tion were, therefore, twice as high in iron and 1.5 timesas high in zinc as the most widely grown varieties. Theseratios, therefore, are benchmarks for iron and zinc den-sity, which seem to be reasonably attainable using con-ventional breeding.High iron and to a lesser extent high zinc concentra-
tions were subsequently shown to be linked to the traitof aromaticity. Most aromatic rices, such as jasmineand basmati types, are high in both iron and zinc, andgenerally in most minerals (Graham, Senadhira, &Ortiz-Monasterio, 1997; Graham et al., 1999; Senadhira& Graham, 1999). A promising aromatic variety, desig-nated IR68-144, already being tested at the Interna-tional Rice Research Institute (IRRI) owing to itssuperior agronomic and consumer characteristics, wasfound to be high in iron (Gregorio, 2002).Expression of micronutrient-density traits has been
tested over a wide range of environments and, althoughthe environmental effect itself is strong, the genotypeeffect is consistent across environments (implying thatthe genotype-environment interaction is not serious), afinding that is sufficient to encourage a breeding effort.Environmental factors considered by one or more cropprograms include acid soils, alkaline soils, saline soils,acid-sulphate soils, iron-deficient soils, time of planting,field site, season, nitrogen fertilisation, phosphorus fer-tilisation, potassium status, elevation and drought stress(Graham et al., 2001).
Transgenic methods
Increasing iron. Ferritin is an iron-storage protein
found in animals, plants, and bacteria. The ferritin gene
has been isolated and sequenced in plants, including
soybean, French bean, pea, and maize. Recent studies
show that ferritin is used by both plants and animals as
the storage form of iron (Theil, Burton, & Beard, 1997);
4 Biofortification will not deplete soils of trace minerals. Sufficienttrace minerals are present in most soils for thousands of crops.Some soils are labelled ‘trace mineral-deficient’ because the prop-erties of these soils are such that the trace minerals are chemicallybound and so unavailable to the plant. Roots of trace mineralefficient lines release compounds into the soil, which chemicallyunbind the trace minerals in the soils and make them available tothe plant. Thus, a strategy of breeding for trace mineral efficientlines makes use of an untapped and abundant resource in the soil.This is a very different situation from depletion of macronutrientsin the soil, such as nitrogen and phosphorus (Graham & Welch,1996).
196 H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209

orally administered ferritin, in non-transgenic foods
(Beard, Burton, & Theil, 1996), and in transgenically-
derived ferritin-rich rice (Murray-Kolb et al., 2002)
can provide a source of iron for treatment of anaemia in
rats.
Human studies with extrinsically radiolabelled animalferritin have indicated that, when added to a meal, ironfrom ferritin is only about half as well absorbed as ironfrom vegetables (Martinez-Torres, Renzi, & Layrisse,1976; Taylor, Martinez-Torres, Romano, & Layrisse,1986) and iron as ferrous sulphate (Skikne, Fonzo,Lynch, & Cook, 1997). However, a recent study hasshown that intrinsically radiolabelled iron in soybeans ishighly bioavailable (27%) to iron-deficient women(Murray-Kolb, Welch, Theil, & Beard, 2003). About50% of the iron in the soybeans was ferritin; theremaining iron, ferric phytate, is not thought to behighly bioavailable.Goto, Yoshihara, Shigemoto, Toki, and Takaiwa
(1999) report improving the iron content of rice bytransferring the entire coding sequence of the soybeanferritin gene into a Japonica rice. The introduced ferri-tin gene was expressed under the control of a rice seed-storage protein glutelin promoter, to mediate the accu-mulation of iron specifically in the grain. The transgenicseeds stored up to three times more iron than normalseeds. Iron levels in the whole (unmilled) seeds ofthe transformants varied from 13 to 38 mg/kg (ppm);pooled mean values were 23 mg/kg for transformantsand 11 mg/kg for non-transformants. The averageiron content in the endosperm was 3.4 mg/kg in thetransformant and 1.6 mg/kg in the non-transformant.This underscores the importance of determining wherein endosperm the iron (and other trace minerals) aredeposited and how mineral levels are affected by milling.Trace minerals cannot be synthesised by plants andmust be transported from the soil, through the plantand to the seed. Several processes and genes may beinvolved. The authors speculate that the amount ofiron accumulation is restricted by transport of iron tothe ferritin molecule, rather than simply by levelsof ferritin protein. Thus, it may be possible to store lar-ger amounts of iron in the ferritin molecule by co-inte-grating the ferritin gene and the iron reductase-liketransporter gene. Robinson, Procter, Connolly, andGuerinot (1999) report a newly discovered ferric-chelatereductase gene, which allows plants to absorb moreiron from the soil, thus introducing the possibility ofwidening the scope of rice varieties with high ironuptake.A doubling of the iron content in rice using a ferritin
gene derived from Phaseolus vulgare has been reported(Gura, 1999). Metallothionine was also expressed in therice grain, increasing the cysteine content seven-fold.
Introducing a gene which codes for heat-stable phytase.
The level of phytase, which breaks down phytic acid, is
normally low in milled rice. Several studies have already
demonstrated the usefulness of adding phytase to the
rice diets of poultry (e.g. Adrizal & Sell, 1996; Farrell &
Martin, 1998; Martin, Nolan, Nitsan, & Farrell, 1998;
Munaro, Lopez, Lopez, & Rutz, 1996; Teichman, Lopez,
& Lopez, 1998). The phytase that does exist in rice seeds
will hydrolyse the phytic acid present at lower pH levels, if
seeds are left to soak until germination in water. However,
boiling destroys the phytases that occur naturally in rice.
In one study, an amino acid was changed in the pro-tein sequence to make the phytase heat stable (Pasa-montes, Haiker, Wyss, Tessier, & Van Loon, 1997). Thephytase was active under the pH conditions of digestionand degraded phytic acid in a very short time, duringmodel in vitro digestion.Lucca, Hurrell, and Potrykus (2001) reported the
introduction into rice of a transgene for this particularheat-stable phytase from Aspergillus fumigatus; thetransgene increased the level of phytase activity by 130-fold. Unfortunately after expressing the gene in thegrain, the phytase was no longer stable to heat and lostits activity on boiling.A new approach is now under investigation in which
the A. fumigatus phytase gene would break down phy-tate during seed maturation [when pH levels (6–7)would be conducive to phytase activity]. Fusing theglobulin promoter to the phytase gene, which tests haveshown is expressed only in the endosperm of rice, whichis consumed after milling, should prevent seed germina-tion being compromised (after harvesting and replanting)owing to low phosphorus content. The hope is that thephosphorus in the outer layers of the seed (which is milledaway for consumption but not for planting) would remainunaffected (Lucca, Hurrell, & Potrykus, 2002).
Increasing levels of compounds that improve absorptionand utilisation of nutrients. Levels of lysine, which is an
important, essential and limiting amino acid in rice and
which might promote the uptake of trace minerals, can
be improved by transgenic methods (Datta & Bouis,
2000). Introduction of two bacterial genes, expressing
DHSPS (dihydrodipicolinic acid synthase) and AK
(aspartokinase) enzymes, encoded by the Corynebacter-
ium dapA gene and a mutant Escherichia coli lysC gene,
has enhanced lysine levels about five-fold in canola and
soybean seeds (Falco et al., 1995).
Adding beta-carotene. Beta-carotene, a precursor of
vitamin A, does not occur naturally in the endosperm of
rice. Ye et al. (2000) have reported generating transgenic
plants that produce grain with yellow-coloured endo-
H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209 197

sperm. Biochemical analysis confirmed that the colour
represents beta-carotene (provitamin A). The reported
level of beta-carotene in one gram of the dry, uncooked
transformed rice (Golden Rice) is 1.6 mg. Research is pre-sently underway to increase the level of beta-carotene in
transformed rice, with promising ‘proof of concept’ results
in model systems (Beyer & Potrykus, 2001).
According to Ye et al. (2000), psy (cloned from Nar-cissus pseudonarcissus; Schledz et al., 1996), cryt1(cloned from Erwinia uredovora; Misawa et al., 1993),and the lyc gene have been introduced into the rice,driven by the endosperm specific glutelin promoter(Gt1). Crt1 was fused to the transit peptide (tp)sequence by the pea Rubisco small subunit (Misawa etal., 1993). The insertion of a multiple gene cassette is aremarkable accomplishment, considering that mosttraits engineered to date have only required the additionof a single gene (Guerinot, 2000).A discussion on the degree to which vitamin A intakes
may be improved by the use of Golden Rice in the ruralBangladesh context is provided below (Section 2.3.1).
2.3. Outcomes and impacts of breeding for iron-and beta-carotene-dense rice
2.3.1. Impact on iron- and beta-carotene intakesFor poor populations, food staple consumption so
dominates diets that primary food staples provide in therange of 40–55% of total iron intakes (Bouis, Graham,& Welch, 2000). If a single food staple provides 50% oftotal iron intakes for a poor population (e.g. rice inBangladesh), then a doubling of the iron density inthat food staple will result in a 50% increase in totaliron intakes. It would seem evident that a 50%increase in intakes of iron, if it were to be as bioa-vailable as iron already in the diet, would be ofconsiderable benefit to anaemic women with such lowiron intakes. Some empirical evidence is availablefrom a recently published study of a population ofBangladeshi rural women (Bhargava, Bouis, & Scrim-shaw, 2001). The study measured the relationshipbetween diets and blood haemoglobin. The estimatedrelationships suggested that a 50% increase in ironintakes from biofortified rice would reduce rates ofanaemia among these women by a minimum of 3% (e.g.from 53–50%).5 This is an estimate of the percentage of
women who would cross above the threshold of 12 g/dlhaemoglobin as a result of consuming biofortified rice.Women who remained below this threshold presumablywould also derive some benefit from a 50% increase iniron intakes.Biofortified varieties may reach undernourished
people for a cost of only US $0.02–0.03 per personper year (Hunt, 2002), compared with a cost of US$0.12 for commercial iron fortification (see Section2.2.1) and costs exceeding US $3.00 per person foriron supplements (World Bank, 1994). In the studyby Bhargava et al. (2001), the effect of iron supple-ments in reducing anaemia levels was estimated to bethree times as strong as that of biofortified rice.Given that, per person reached, the cost of iron sup-plements is more than 100 times the cost of biofortifiedrice (i.e. US $3.00 compared with <US 0.03 cents), thenthe cost per annual case of anaemia avoided is morethan 30 times higher for supplements than for bioforti-fied rice.For the same rural Bangladeshi population referred to
above, it is also possible to estimate the percentageincrease in total provitamin A intake provided byreplacement of presently eaten varieties of rice withGolden Rice (1.6 mg beta-carotene per gram of dry riceendosperm; Ye et al., 2000). For this poor population,animal and fish intakes provide only 3% of total energy(Bouis et al., 1998). Intakes of preformed vitamin A are,therefore, negligible and vegetables provide greater than90% of total provitamin A intakes. The US Institute ofMedicine6 recommends that a ratio of 12 to 1 be usedfor converting beta-carotene to retinol activity equiva-lents when evaluating mixed diets for healthy popula-tions (Institute of Medicine, Food and Nutrition Board,2001). Estimated average intakes for Bangladeshiwomen and for preschool children (2–5 years) are 260and 106 retinol equivalents, respectively.Recommended dietary allowances (RDAs) for heal-
thy women and children (4–8 years) are 700 and 400retinol equivalents, respectively (Institute of Medicine,Food and Nutrition Board, 2001). Vitamin A intakes inBangladesh are, therefore, currently well below recom-mended intakes.If Golden Rice were to replace the non-transgenic
rice, total provitamin A intakes would be increased by25% for adult women and preschool children; using aratio of 12 to 1 to convert provitamin A in both vege-tables and Golden Rice to retinol activity equivalents,total vitamin A intakes would still remain well belowrequirements.However, the conversion rate from beta-carotene to
retinol activity equivalents is a function of the foodmatrix in which the beta-carotene is present, the form in
,
5 Due to measurement error in a number of difficult-to-recordvariables, this estimate is likely to be a considerable underestimate.Roughly, it can be estimated that each additional 1mg of non-haeme iron added to rice (or to any other food in the diet) wouldreduce the anaemia prevalence by 1%. To see that this is a lowerbound estimate, it should be considered that, at this rate, anadditional 36 mg of iron, or twice the recommended daily allow-ance for women, would be required to lower prevalence by 36%e.g. from 50 to 14%.
6 IVACG (2002) Conversion Factors for Vitamin A and Carotenoidsavailable [July 2002] at http://www.ilsi.org/file/vitabookmark.pdf.
198 H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209

which the food is prepared, the presence of dietary fat,and other factors. For example, the conversion rate forsome leafy, green vegetables has been estimated to be 26to 1, whereas that from some orange fruits may be 13–15 to 1 (de Pee et al., 1998). It may be that the beta-carotene in the rice endosperm matrix would allow arelatively high conversion rate. Rice is a very simplefood matrix, which is totally digestible. Therefore, car-otenoids may be more easily released from rice thanfrom an undigested matrix of vegetable origin (Tyssan-dier, Lyan, & Borel, 2001).
It is also important to note that RDAs are set at levelsto maintain 4-month body stores. About one-half of theRDA is needed to prevent vitamin A deficiency (Insti-tute of Medicine, Food and Nutrition Board, 2001). Forthe Bangladeshi population described above, adultwomen would reach one-half of the RDA (deficiencyprevention) at an 8 to 1 conversion ratio and preschoolchildren at a 4 to 1 conversion ratio. However, such acalculation assumes that only Golden Rice would beconsumed (complete substitution with other rice vari-eties) and that no beta-carotene would be lost in pro-cessing and cooking. Research is currently beingundertaken to increase the beta-carotene content of newlines of Golden Rice (Beyer et al., 2002).However, the conversion factors and RDA, assume a
healthy population, which is unlikely for poorer seg-ments of the Bangladeshi population, or for poorerpopulations in other less developed countries. Under-nutrition is prevalent and enteric infections frequent;this erodes the integrity of the intestinal absorptionsurface where the mucosal beta-carotene cleavageenzyme normally resides. A vitamin A deficient statemay partially compensate by increasing both the con-version efficiency of the beta-carotene that is absorbedand its efficient utilisation. Data on how these numerousvariables interact to influence absorption efficiency fromGolden Rice are limited. Therefore, uncertainty sur-rounds estimates of the contribution to vitamin A statusthat high beta- carotene staple foods can make amongpoor populations in developing countries, whether suchfoods are selected from existing gene banks or devel-oped using classical breeding techniques or transgenicmethods. Efficacy trials will help to reduce some of thisuncertainty.
2.3.2. Costs and benefits of a biofortification strategyDevelopment of iron- and zinc- dense varieties of rice
or wheat using conventional breeding might cost asmuch as US $10 million each over 10 years, includingthe costs of nutrition safety and efficacy tests, the costsof dissemination in selected regions, and the costs of anevaluation of nutritional and economic impact (Hunt,2002). As indicated above (Section 2.2.1), such an
investment (of US $20 million in two crops, rice andwheat) is projected to have far reaching impacts if effi-cacy and effectiveness are achieved. Under such condi-tions, it would be reasonable to extrapolate benefits tocountries around the world, as countries would needonly to invest in adaptive breeding and disseminationcosts. Moreover, benefits should be sustainable at lowmaintenance costs. Benefits from breeding advancestypically do not disappear after initial investmentsand research have been successful, as long as an effec-tive domestic agricultural research infrastructure ismaintained.A simulation model, which was developed for Ban-
gladesh and India, based on development of iron- andzinc-dense varieties of rice and wheat, which wereassumed to be adopted on only 10% of approximately83 million hectares planted to rice and wheat, supportedthe expectation of enormous economic benefits from abiofortification strategy (Hunt, 2002). Total costs wereestimated to be US $42 million which, in addition to theUS $20 million in central development costs for rice andwheat, included costs of adaptive and maintenancebreeding and extension costs. These conservativeassumptions suggest that the returns that come on-stream during the second decade of research and devel-opment would be about $1.2 billion in benefits frombetter nutrition.A more formal economic evaluation of biofortifica-
tion in Bangladesh and India, in which the ratio of thepresent value of benefits to the present value of costswas discounted at a 3% rate (commonly used for eva-luation of social benefits), gave a benefit-cost ratio of19, for returns, simply for better iron nutrition inhumans; a similar ratio was found by Horton and Ross(1998) for fortification in South Asia. A different way ofexpressing the concept of discounting over time is theinternal rate of return, in which the interest rate is calcu-lated at which benefits would equal costs plus interest, ifthe funds were to be borrowed to make the investment. Inthis case the internal rate of return would be 29%, mea-sured as benefits to human nutrition (Hunt, 2002).In the longer term (years 11–25 of the simulation
model) it was estimated that a total of 44 million casesof anaemia would be prevented if nutritionallyimproved varieties were to be adopted on 10% of riceand wheat areas in Bangladesh and India. This wasbased on a conservative assumption of only a 3%reduction in anaemia among those consuming the high-iron rice. Thus, costs would be about US $1 per case ofanaemia per year prevented and US $0.03 per person peryear for those whose iron intakes would be increased by50% through consumption of iron-dense rice and wheat.These estimates have been made for conventional
breeding. It is not known whether transgenic methodswould be more or less costly than conventional breedingmethods, in part because the higher costs of meeting
H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209 199

stricter regulatory procedures for transgenic crops arenot yet known. These higher regulatory costs for trans-genic methods may be offset by the greater costs andtime of backcrossing involved with conventional tech-niques. However, the relative costs of transgenic andconventional methods are of secondary importance, inthe sense that, even if total costs for either methoddoubled the assumed costs for conventional breeding of$10 million over 10 years, biofortification would still bea very cost-effective proposition. For example, the veryconservative benefit calculations above included onlybenefits for India and Bangladesh, while the varietiesdeveloped under full development cost assumptionscould be used in other countries around the world aswell. It is very difficult to predict benefits and breedingcosts ex ante, so that it is important to make con-servative estimates of benefits.7 Extrapolations are madeon the assumption of specified levels of efficacy andeffectiveness for interventions characterised by uncer-tainties that result from incomplete knowledge.There is no a priori reason to believe that research
costs will be higher or lower for a widely grown crop(e.g. for a cereal such as rice) than for a ‘minor’ crop(e.g. pulses). Given constant costs, benefits in agri-cultural research will always be higher for the morewidely grown crop. Thus, there is a bias in researchinvestments toward those crops and, consequentlyminor crops lag behind in terms of productivity increa-ses. If the costs of developing transgenic crops are highowing to the need to meet regulatory procedures, thetendency to undertake research only on the more widelygrown crops will increase. On the other hand, if bio-technology can lower the costs of undertaking agri-cultural research by making the process more efficient,this tendency will be mitigated.
2.4. Evaluating outcomes and impacts of iron- andbeta-carotene-dense rice
2.4.1. Magnitudes of benefits and risksMethods for evaluating the outcome and impact of
nutritionally improved staple food crops fall into twocategories:
. simple calculations of probable increases intotal nutrient intakes from diets, such as thoseundertaken in section 2.3.1 in relation to RDAs;and
. assessment of the impact of the increased mineraland vitamin intakes on nutritional status.
The second category includes, in increasing order ofprecision, relevance, and cost:
. in vitro techniques (e.g. Glahn, Lee, & Miller,1999);
. animal models (e.g. Welch et al., 2000);
. test meals;
. efficacy trials (e.g. Haas, del Mundo, & Beard,2000); and
. effectiveness studies
2.4.2. Assessing likely beneficiaries of transgenictechnologiesIn vitro techniques and animal studies are relatively
inexpensive and quick to implement and are primarilyuseful for screening candidate lines in a breeding pro-gram for relative bioavailability; however, they providelittle information on the ultimate impact on micro-nutrient status of human populations of specific geno-types of crops being tested. For this assessment, severalfactors that affect the absorption and utilisation ofnutrients must be evaluated. Such factors include:
. the micronutrient and health status of subjects; forexample, those who are iron-deficient generallywill absorb a higher percentage of iron in the diet;
. the level of compounds and nutrients in the mealsbeing consumed that promote or inhibit theabsorption of nutrients; for example, phytatesinhibit the absorption of zinc; the presence ofparasites; and
. the absence of other significantly limiting nutrientsin the diets.
The relevance of results from test meals and efficacytrials, therefore, depend on how representative of theeconomically most disadvantaged population groups,are the nutritional status and diets of the test subjects.Efficacy trials, but not test meals, can allow for longer-term adaptations to the introduction of new foods andnutrients. Finally, effectiveness trials allow outcomeand impact evaluation under usual community livingsituations.An efficacy trial is currently being conducted with
iron-dense rice strains developed using classical breed-ing techniques. A specific line of Golden Rice withstable and relatively high levels of beta-carotene is cur-rently being selected for test meal evaluation of bioeffi-cacy in humans. Several months will be needed to growa sufficient quantity of rice with stable-isotope-labelledbeta-carotene to undertake the study. Meanwhile experi-ments continue to develop improved lines with higher
7 There are agronomic benefits associated with high zinc seeddensity which are not discussed here (Graham, Welch, & Bouis2001). Possible higher crop yields are not counted as possiblebenefits nor benefits to better zinc nutrition. During germplasmscreening, a high correlation between iron and zinc density in riceand wheat seeds has been found, therefore breeding for bothcharacteristics simultaneously is feasible.
200 H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209

levels of beta-carotene and even lines that produce retinol(Beyer & Potrykus, 2001; Beyer et al., 2002).
2.5. Knowledge gaps in breeding for iron- andbeta-carotene-dense rice
2.5.1. The genotype factors that optimally determinebioavailabilityDetermining the bioavailability to humans of micro-
nutrients in plant foods depends on a myriad of factors,which interact to determine the ultimate bioavailabilityof a particular micronutrient to an individual, eating amixed diet, within a given environment (Fairweather-Tait & Hurrell, 1996; Welch, 2002). These factorsinclude the combination of nutrients, antinutrients, andabsorption enhancers present in the nutrient enhancedrice and the meal in which it is eaten. Because of thiscomplexity, the data obtained using in vitro systems,animal models and test meals are always ambiguous(House, 1999; Van Campen & Glahn, 1999). Only dataobtained on reducing the prevalence of micronutrientdeficiencies, using feeding trials in test populationsunder free living conditions, can delineate the actualefficacy of using micronutrient-enriched varieties ofplant foods as an intervention tool. However, it isimpractical to test, in this way, the bioavailability ofselected micronutrients, in the numerous genotypes ofstaple plant foods that can be generated in plantbreeding programs (Graham et al., 2001; Graham &Welch, 1996). The ultimate proof of the biofortificationstrategy will lie in effectiveness studies undertaken inareas where nutritionally improved varieties will beintroduced.
2.5.2. Consumer acceptanceThere are various potential barriers to the adoption,
for example, of Golden Rice.
. It is yellow in colour, whereas Asian consumerswho stand to benefit the most from its introduc-tion are used to eating white rice.
. There may be consumer acceptance issues, poten-tially related to safety, ethics and localisation ofthe decision-making process in agricultural devel-opment and associated processes.
. There may be concerns that technological solu-tions used in isolation will exclude alternativeapproaches, including for example, failure to har-ness technology to social policy.
Whether public health education initiatives canaddress all of these concerns is open to question.Examples of successes in programs encouraging con-sumers to switch from white to yellow or orange vari-
eties of staple food crops provide some encouragement(Hagenimana & Low, 2000).
2.5.3. Relative efficiency of transgenic andconventional breeding methodologiesMuch experience has been accumulated with con-
ventional breeding and it continues to become moreefficient, in part owing to new techniques developedthrough the use of biotechnology. Yet, clearly thecapability to move genes across species holds greatpotential, as demonstrated by the examples presentedabove for rice, to accomplish goals that cannot beattained with conventional breeding. As experience isgained with biotechnology, development of transgenicvarieties will become more efficient and cost-effective.It is difficult if not impossible to determine howinvestments should best be allocated between con-ventional breeding, which will give benefits now, andtransgenic methods, which will generate future benefits.
2.5.4. Food and nutrient intakes stratified by country,urban-rural areas, and socioeconomic classA rigorous ex ante evaluation of the benefits of bio-
fortification requires the following:
. data on micronutrient status for various categoriesof individuals (e.g. by age, gender, income class,geographical location), matched with
. detailed knowledge of their food intakes and
. a reliably estimated empirical relationship, which‘explains’ observed micronutrient status as a func-tion of accurately measured levels of nutrients andother compounds in the diets.
A first attempt at such a study is described above(Section 2.3.2); however mismeasurement of foodintakes and nutrient content probably leads to anunderestimation of the benefit of dietary quality onmicronutrient status.
2.5.5. Unintended effectsWhen several nutrients are lacking in diets and a sin-
gle nutrient is added in significant quantities, it isunclear what the nutritional outcome will be, in the lightof important interactions among nutrients, which arenot fully understood. It will be important to check thatthere are no negative, unintended consequences of add-ing one or a few nutrients to the diet or of altering thelevels of other compounds in foods.
2.6. Other potential contributions of GM crops
Sections 2.2–2.5 have sought to place the effort tobreed for transgenic iron- and beta- carotene-dense rice
H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209 201
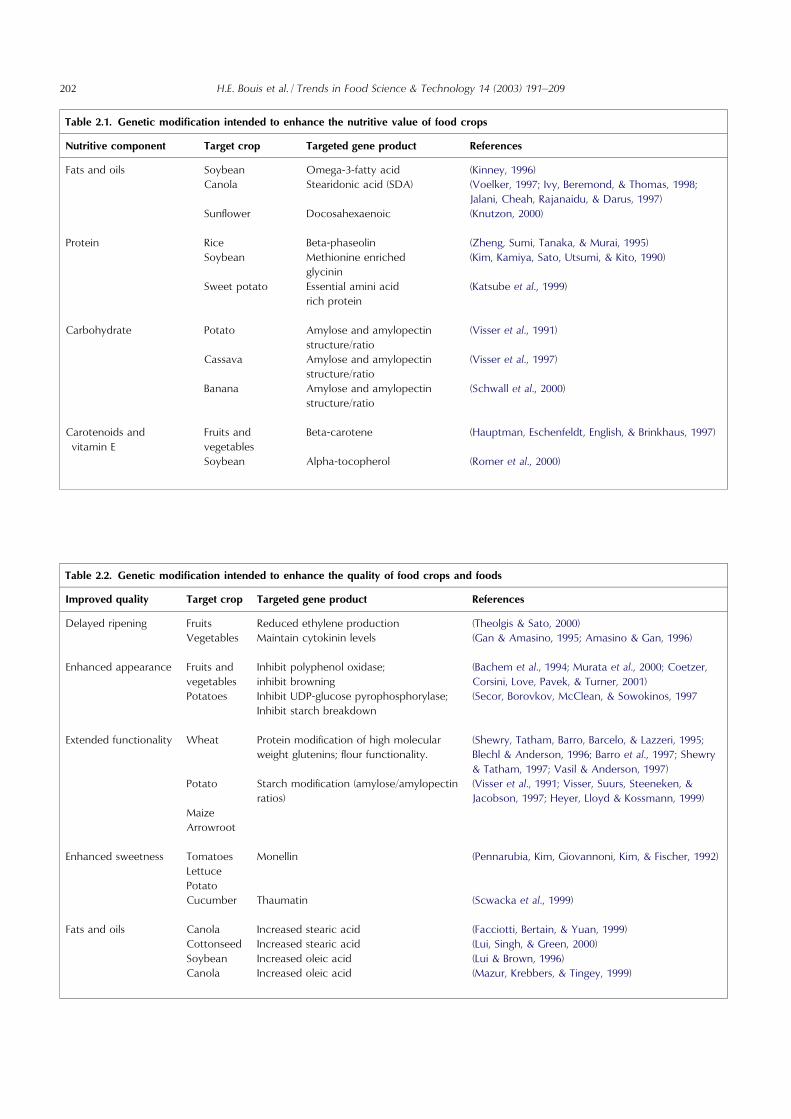
Table 2.1. Genetic modification intended to enhance the nutritive value of food crops
Nutritive component
Target crop Targeted gene product ReferencesFats and oils
Soybean Omega-3-fatty acid (Kinney, 1996) Canola Stearidonic acid (SDA) (Voelker, 1997; Ivy, Beremond, & Thomas, 1998;Jalani, Cheah, Rajanaidu, & Darus, 1997)
Sunflower Docosahexaenoic (Knutzon, 2000)Protein
Rice Beta-phaseolin (Zheng, Sumi, Tanaka, & Murai, 1995) Soybean Methionine enrichedglycinin
(Kim, Kamiya, Sato, Utsumi, & Kito, 1990)Sweet potato
Essential amini acidrich protein(Katsube et al., 1999)
Carbohydrate
Potato Amylose and amylopectinstructure/ratio(Visser et al., 1991)
Cassava
Amylose and amylopectinstructure/ratio(Visser et al., 1997)
Banana
Amylose and amylopectinstructure/ratio(Schwall et al., 2000)
Carotenoids andvitamin E
Fruits andvegetables
Beta-carotene
(Hauptman, Eschenfeldt, English, & Brinkhaus, 1997)Soybean
Alpha-tocopherol (Romer et al., 2000)Table 2.2. Genetic modification intended to enhance the quality of food crops and foods
Improved quality
Target crop Targeted gene product ReferencesDelayed ripening
Fruits Reduced ethylene production (Theolgis & Sato, 2000) Vegetables Maintain cytokinin levels (Gan & Amasino, 1995; Amasino & Gan, 1996)Enhanced appearance
Fruits andvegetablesInhibit polyphenol oxidase;inhibit browning
(Bachem et al., 1994; Murata et al., 2000; Coetzer,Corsini, Love, Pavek, & Turner, 2001)
Potatoes
Inhibit UDP-glucose pyrophosphorylase;Inhibit starch breakdown(Secor, Borovkov, McClean, & Sowokinos, 1997
Extended functionality
Wheat Protein modification of high molecularweight glutenins; flour functionality.(Shewry, Tatham, Barro, Barcelo, & Lazzeri, 1995;Blechl & Anderson, 1996; Barro et al., 1997; Shewry& Tatham, 1997; Vasil & Anderson, 1997)
Potato
Starch modification (amylose/amylopectinratios)(Visser et al., 1991; Visser, Suurs, Steeneken, &Jacobson, 1997; Heyer, Lloyd & Kossmann, 1999)
Maize
ArrowrootEnhanced sweetness
Tomatoes Monellin (Pennarubia, Kim, Giovannoni, Kim, & Fischer, 1992) Lettuce Potato Cucumber Thaumatin (Scwacka et al., 1999)Fats and oils
Canola Increased stearic acid (Facciotti, Bertain, & Yuan, 1999) Cottonseed Increased stearic acid (Lui, Singh, & Green, 2000) Soybean Increased oleic acid (Lui & Brown, 1996) Canola Increased oleic acid (Mazur, Krebbers, & Tingey, 1999)202 H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209

in the context of the need for such interventions indeveloping countries, the likelihood of success, alter-native interventions, and costs and benefits.Various attempts to enhance the nutritional or poten-
tial health benefits of crops or food ingredients, in otherways, have also been reported. A detailed analysis ofother biotechnology-related research to improve thenutritional and health enhancing effects of food of rele-vance to developed as well as developing countries is notgiven herein, although an overview of several of theseactivities is presented below. The requirements for thedevelopment and assessment of such GMFs will be simi-lar to those discussed above for GM rice. Possible geneticmodification interventions of relevance to nutritional andquality factors are outlined in Tables 2.1 and 2.2.
2.6.1. Enhancing nutritional valueStrategies similar to those described above for Golden
Rice have been employed to develop other crops withenhanced macro- or micronutrient content. Some ofthese modified crops are listed in Table 2.1; however,none of them has reached the stage of commercialisa-tion, so it is uncertain what drawbacks or risks might beassociated with each.
2.6.2. Non-nutritive active componentsThere is considerable private sector and consumer
interest in ‘functional foods’, that is foods with specialhealth benefits not associated with nutrients. This sec-tion gives a brief insight into how biotechnology may beused to produce healthier foods. It should be empha-sised, however, that these new varieties are the subjectof research that is in its earliest stages. Much funda-mental research needs to be done to substantiate theproposed health benefits and associated risks, prior tothe development of products with enhanced phyto-chemical content.It is now widely recognised that plants contain many
physiologically active components that have no classi-cally defined nutritional role, but may have positive oradverse effects on human health. Much as is the casewith carotenoids and antioxidants, consumers seemready to seek out and use new foods for which somehealth claim or expectation has been described. Many ofthese are viewed as a ‘natural’ means to ensure goodhealth. For the present, there are only a few food com-ponents that sound evidence links positively to healthoutcomes, as is the case, for example, for some phytos-terols (discussed below. Nonetheless, the pathways bywhich some of these compounds are synthesised arebeing elucidated so that ultimately their levels can bemanipulated. This has given rise to the new field called‘nutritional genomics’ or ‘metabolic engineering’ (Del-laPenna, 1999, 2001).Plant phytosterols, such as beta-sitosterol, were once
used as cholesterol-lowering drugs. Recently, functional
food products, such as margarines, containing phytos-terols or phytostanols have appeared on the market inEurope and the USA to help lower elevated cholesterollevels. The sources of these compounds include pine treeresin and soybeans. The content of phytosterols in oil-seeds has been enhanced by genetic modification (Ven-katramesh, 2000). Such an improvement could increasethe availability of this new functional component so itcould be included in more foods.It is easy to understand the consumer appeal of func-
tional foods that are claimed to prevent cancer. Forexample, glucoraphanin, a glucosinolate found in Bras-sica vegetables, is hypothesised to have cancer-prevent-ing effects in humans. It has been shown to inducedetoxification enzymes in the liver although it is unclearif that is the mechanism by which it could prevent can-cer. Glucoraphanin is not present in foods in high con-centrations and it is found in foods, such as broccoli,that are not particularly popular with some consumers.Research is now being directed at increasing levels ofglucoraphanin in certain vegetables. What effect thiswill have on the overall safety, flavour or acceptabilityof food will be the subject of much investigation if suchplants are developed.Flavonoids are a class of compounds, including iso-
flavones, anthocyanins, rutin, quercitin, and kaemp-ferol, that are synthesized from phenylalanine viachalcone synthase and chalcone isomerase. They arepresent in fruits, vegetables, nuts and seeds. Flavonoidshave strong antioxidant activity; both short- and long-term effects in humans of elevated consumption haveyet to be demonstrated. It is hypothesised, but has notyet been proved, that the health benefits associated withdiets rich in fruits and vegetables may derive, in part atleast, from intake of components such as flavonoids. Ithas been difficult to convince consumers in some cul-tures, particularly children, to eat recommended quan-tities of fruits and vegetables. An alternative may be tohelp increase the intake of components such as flavo-noids by enhancing their levels in fruits and vegetables,through biotechnology. Recently, the gene for soybeanisoflavone synthase has been expressed in the laboratorymodel plant, Arabidopsis thaliana; the transformantnon-leguminous plant produced the legume-derivedisoflavone, genestein (Jung et al., 2000). Also the geneencoding chalcone isomerase has been isolated frompetunia and inserted into tomato (Muir et al., 2001).Paste processed from the transformed tomatoes con-tained up to 1.9 mg flavonols per gram dry weight, a 21-fold increase compared with wild controls. Further-more, there was no negative effect on the plants and noundesirable flavours developed in the paste product.It is widely assumed that consumers will readily
accept functional foods, as people will perceive there is abenefit to health associated with such products. How-ever, some psychological factors are important determi-
H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209 203

nants of food choices, and there are inter-individual andcross-cultural differences in attitudes. For example,consumer research has demonstrated that, very gen-erally, older Americans are more positive towards‘functional foods’ than are younger Europeans. Con-sumers are more tolerant of the enrichment or fortifica-tion of foods with substances that they already containthan enrichment of foods with novel substances. Indivi-dual differences in barriers to dietary change have alsobeen identified. These include: ‘food neophobia’; theextent to which people are fatalistic about developingdisease and do not adopt dietary strategies to preventillness; and cultural factors that determine food choice.Other barriers to dietary change include factors like‘optimistic biases’, in which people rate their own risk ofdeveloping illnesses as less than people with whom theycompare themselves. Such psychological barriers havebeen shown to influence the extent to which individualsadopt healthy behaviours (such as healthy food choi-ces), in line with recommendations (Frewer, Scholderer,& Lambert, in press).In order to succeed, the development and use of
functional foods should be based on documented effi-cacy and should take account of diverse needs andrequirements of different populations and demographicgroups, as well as cultural factors that determinefood choices. In broad terms, simply providing infor-mation about health benefits is unlikely to be aninfluential strategy for changing well-established con-sumer behaviours.
2.6.3. Improving quality traitsBiotechnology offers the possibility to produce grains,
fruits and vegetables that retain their fresh appearancelonger and have a longer useful shelf life than non-modified products. Other targets for modificationinclude alterations in flavour, taste, texture and viscos-ity. Changes can be directed at alteration of the expres-sion of a key enzyme or introduction of a new proteinor enzymatic activity. For example, antisense RNA hasfrequently been employed to block the synthesis of keyenzymes. Some examples, representative of the scope ofthe modifications that have been reported, are presentedin Table 2.2.For example, fresh fruits and vegetables are relatively
perishable commodities that have limited shelf lives.Moreover, fresh produce and fruits must often bepicked in a green or unripened state in order to allowtime for shipping, marketing and, ultimately, ripeningprior to sale. Plant breeders have sought for manyyears, with only limited success, to develop varieties thathave delayed ripening and extended shelf lives. Theintroduction of molecular breeding techniques hasallowed the targeting and modification of pathwaysinvolved in ripening and senescence in certain crops. In
fact, the very first product of biotechnology to beapproved as a food was designed to enhance freshness.Softening of tomatoes can be delayed by insertion intotomatoes of an antisense mRNA that inhibits synthesisof an enzyme, polygalacturonase, that breaks downpectin. The resulting ‘Flavr SavrTM’ tomato was soldas whole fresh fruit in supermarkets while a similartomato was used in processed tomato products in theUK (Roller & Harlander, 1998). The products havebeen withdrawn from the market because they wereunprofitable.
2.6.4. Removing allergens and antinutrientsThe primary food allergens in a plant are frequently
major proteins. One approach to reducing allergenicityis to knock out the biosynthesis of the offending pro-tein. A second approach is to modify the structure ofthe offending protein so that it no longer contains theepitopes responsible for the undesirable immuneresponses that result in allergic signs and symptoms.The first approach has been applied to rice. The contentof the major allergenic protein in rice has been success-fully reduced; however, it is not known if this results indecreased allergenicity in humans (Tada et al., 1996).Peanut allergen (Bannon, Shin, Maleki, Kopper, &Burks, 1999) and potato allergen (Alibhai et al., 2000;Astwood, Alibhai, Lee, Fuchs, & Sampson, 2000) havebeen modified by protein engineering to remove epi-topes associated with allergenicty. A more generalmethod of disarming potential food allergens is alsobeing tested (Buchanan et al., 1997). Thioredoxin is aregulatory disulfide protein that is widely distributed innature. It has been found to reduce disulfide bonds inallergens, and the reduced allergens are less allergenic.Research is underway to determine if insertion of a genefor thioredoxin biosynthesis into wheat will reduce itsallergenicity without interfering with the functionalityof the wheat proteins.Members of the Brassica family, such as cabbage,
Brussels sprouts and broccoli, rapeseed and mustardcontain a family of compounds called glucosinolates.Glucosinolates can be hydrolysed to bitter tasting andpotentially toxic isothiocyanates. Extensive plantbreeding was required to produce varieties of rapeseedand mustard with greatly reduced levels of glucosino-lates. The meal remaining after oil extraction from thesevarieties can be used in animal feed. Glucosinolates arepart of the plant’s defence system against insect pestsand pathogens. Attempts are now being made todevelop plants that have high glucosinolate levels inleaves and seedpods but much reduced levels in the oil-seeds (Vageeshbabu & Chopra, 1997).Solanine and chaconine are naturally occurring gly-
coalkaloid toxins that are found in potatoes and othersolanaceous plants. While toxic to humans, the com-
204 H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209

pounds are destroyed by cooking. It has been reportedthat insertion of an invertase gene from yeast intopotato reduces the glycoalkaloid content (Engel, Blaas,Gabriel, & Beckman, 1996). The exact mechanism forthis inhibition is not known and further work seeks toestablish if the effect is caused by metabolic interferencewith the sugar moieties needed for biosynthesis of gly-coalkaloids. It is also possible to use antisense RNA toblock synthesis of a gene that codes for a key enzyme inglycoalkaloid synthesis.Fumonisin is a mycotoxin that is produced by the fun-
gus Fursarium. It accumulates in maize that has becomeinfected at the site of damage by insects such as the Eur-opean corn borer. Although the fumonisin content ofmaize is approximately proportional to the level of insectdamage, the temperature and humidity of storage canaffect the level of mycotoxin accumulation. Levels offumonisin in grain should be carefully controlled sincefumonisin B1 is a possible human carcinogen (IARC,2002). It has been linked to oesophageal cancer in humanswho consume high levels of fumonisin-contaminatedmaize. Fumonisin causes liver cancer in rats (Gelderblom,Abel, et al., 2001) and liver and kidney damage in non-human primates (Gelderbom, Seier, et al., 2001); it is alsoknown to be acutely toxic to livestock. De-esterase geneshave been transferred from yeast into maize for the pur-pose of detoxifying fumonisin (Duvick, 2001).As Bt-maize is resistant to attack by insect pests,
levels of fumonisin in Bt-maize are consistently greatlyreduced over levels observed in its conventional coun-terpart (Cahagnier & Melcion, 2000; Dowd, 2000;Munkvold, Hellmich, & Rice 1999). In a 3-year studyconducted in northern Italy, where fumonisin con-tamination of corn is a major problem, fumonisin levelsin Bt corn were greatly reduced compared with tradi-tional hybrids (Pietri & Piva, 2000). Reductions in thefumonisin content of maize represent a significantenhancement in food and feed safety and will alsoreduce post-harvest losses associated with the destruc-tion of contaminated grain
2.7. Standards for food useMany of the potential GM products that are descri-
bed herein have altered macro- or micronutrient com-position. Modifications are intended to enhancenutritional value or alter the quality characteristics ofthe food. These new plant varieties contain altered con-tents of macronutrients, micronutrients or non-nutritivecompounds of special interest that are ordinarily con-sumed in the human diet.These new crops are intended for distinct uses. Pro-
ducts such as Golden Rice are intended to replace orpartially replace a commodity crop, in regions wherepopulations consuming rice are at risk of vitamin Adeficiency. Many of the plant developments described in
this chapter will be segregated during cultivation andoffered for sale to consumers, based on their enhancednutritional or quality characteristics.Any type of crop whose nutritional composition has
been altered, whether by traditional or biotechnologicalmeans, must be shown to be stably maintained; fur-thermore the crop should not interfere with or nega-tively impact on other cropping systems or theenvironment. It will also be important to monitor thenutritional status of populations that have adopted newnutritionally enhanced crops.Also, any type of crop whose nutritional composition
has been altered, whether by traditional or biotechno-logical means, should pose no unique environmental orfood risk. The most significant challenge will be to pre-dict changes in the dietary intake of specific nutrientswhen these products are introduced into the market-place. It may be inadvisable to market products thatcontain elevated levels of nutrients that could be toxicto the 99th percentile of consumers. National food con-sumption surveys may be useful in monitoring changesin dietary intake.
2.8. ConclusionsThe following lessons may be drawn concerning the
potential usefulness of biotechnology in helping to pro-vide more nutritious foods in developing countries.
. To be of value plant breeding has to be demon-strated to be safe (see below) and more cost-effec-tive than alternative interventions already in placeto reduce micronutrient undernutrition. Such cost-effectiveness is anticipated, in large measurebecause of the far-reaching impacts of plantbreeding. A relatively small, fixed, initial invest-ment in research could benefit the health of mil-lions of poor people in developing countries allover the world, and at the same time couldimprove agricultural productivity on lands that arepresently among the least productive. Nutrient-dense varieties of crops can be developed for afraction of the recurrent estimated annual costs ofsupplementation programmes in developing coun-tries (for example US $1.65 billion for vitamin Asupplementation programmes) and can reach farmore people, as the benefits can be extended acrosscountries and time.
. There should be aspects of the breeding strategyfor which biotechnology is superior to conven-tional breeding techniques. For rice, this is the casefor adding beta-carotene-related and heat-stablephytase genes. In the long run, as more is under-stood about the factors driving translocation ofminerals in plants, biotechnology may also be amore efficient way to increase trace mineral density.
H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209 205

. Where biotechnology is superior to conventionalplant breeding, it must be recognised that the sameconditions as that apply to products developed viaconventional plant breeding, must also be met, asfollows:
� there should be no serious, negative agronomicconsequences associated with the characteristicbeing added;
� any noticeable changes in the colour, taste,texture, cooking qualities, and other featuresassociated with the characteristic being addedshould be acceptable to the consumer;
� the characteristic being added should result in ameasurable improvement in the nutritional sta-tus of the malnourished, target population; and
� there should be no unacceptable nutritionaleffects.
The conditions, relating to safety, acceptability andefficacy, in particular, have yet to be firmly established.Nutritionally improved transgenic foods have not beenfully developed and tested for their potential to improvemicronutrient status, even using in vitro techniques andanimal models, let alone in human subjects with micro-nutrient deficiencies.Definitive proof of the benefit of nutritionally
improved, transgenic foods obviously depends on fur-ther development, release and eventual impact evalua-tion, all of which are many years into the future.Nevertheless, in the shorter term, lessons learnedthrough initial experiences with conventional breedingefforts show just how cost-effective a biofortificationstrategy can be. Clearly, the nutritional quality of staplefoods can be substantially improved using transgenicmethods compared with what can be accomplishedusing conventional breeding. Knowledge in the area ofnutritional genomics should expand rapidly in thefuture and more efficient techniques for producingtransgenic plants will be developed. There are poten-tially substantial benefits to the poor in developingcountries in relation to costs.Characteristics of food, such as appearance, taste, and
colour, which are important to consumers in both devel-oped and developing countries, can also be improvedthrough biotechnology; however, such research currentlyprobably benefits wealthy consumers in developed coun-tries more than poor consumers in developing countries.Ultimately, good nutrition depends on adequate
intakes of a range of nutrients and other compounds, incombinations and levels that are not yet completelyunderstood. Thus, the best and final solution to elim-inating undernutrition as a public health problem indeveloping countries is to provide increased consump-tion of a range of non-staple foods. By reducing the costof producing food, biotechnology will, perhaps, make
its most important contribution to reducing malnutri-tion. However, this will require several decades to berealised, informed government policies, and a relativelylarge investment in agricultural research and otherpublic and on-farm infrastructure.In conceptualising solutions for a range of nutritional
deficiencies, interdisciplinary communication betweenplant scientists and human nutrition scientists holdsgreat potential. Human nutritionists need to beinformed, for example, about the extent to which thevitamin and mineral density of specific foods, as well ascompounds that promote and inhibit their bioavail-ability, can be modified through plant breeding. Plantbreeders need to be aware of both the major influencethat agricultural research may have had on nutrientutilisation in the past (e.g. the bioavailability of traceminerals in modern varieties versus bioavailability intraditional varieties), and the potential of plant breedingfor future improvements in nutrition and health.
2.9. References
Administrative Committee on Coordination, Subcommittee onNutrition, & International Food Policy Research Institute. (2000).Fourth report on the world nutrition situation. Geneva: UnitedNations.
Adrizal, P. E. P., & Sell, J. L. (1996). Utilization of defatted rice branby broiler chickens. Poultry Science, 75, 1012–1017.
Alibhai, M., Astwood, J., Joyce, E., Pershing, J., Sampson, H., & Pur-cell, J. (2000). Re-engineering patatin (Sol t 1) protein to eliminateIgE binding. Journal of Allergy and Clinical Immunology,105(Suppl.), S79.
Amasino, R., & Gan, S. (1996). Transgenic plants with altered senes-cence characteristics (International Patent Publication No. WO96/29858). Geneva: World Intellectual Property Organization.
Astwood, J., Alibhai, M., Lee, T., Fuchs, R., & Sampson, H. (2000).Identification and characterization of IgE binding epitopes ofpatatin, a major food allergen of potato. Journal of Allergy andClinical Immunology, 105(Suppl.), S184.
Bachem, C., Speckmann, G., van der Linde, P., Verheggen, F., Hunt,M., Steffens, J., & Zabeau, M. (1994). Antisense expression ofpolyphenol oxidase genes inhibits enzymatic browning in potatotubers. Bio-Technology, 12, 1101–1105.
Bannon, G., Shin, D., Maleki, S., Kopper, R., & Burks, A. W. (1999).Tertiary structure and biophysical properties of a major peanutallergen, implications for production of a hypoallergenic protein.International Archives of Allergy and Immunology, 118, 315–316.
Barro, F., Rooke, L., Bekes, F., Gras, P., Tatham, Fido, R., Lazzeri, P.,Shewry, P., & Barcelo, P. (1997). Transformation of wheat withhigh molecular weight subunit genes results in improved func-tional properties. Nature Biotechnology, 15, 1295–1299.
Beard, J. L., Burton, J. W., & Theil, E. C. (1996). Purified ferritin andsoybean meal can be sources of iron for treating iron deficiencyin rats. Journal of Nutrition, 126, 154–160.
Beaton, G. H., Martorell, R., Aronson, K. J., Edminston, B., McCabe,G., Ross, A. C., & Harvey, B. (1993). Effectiveness of vitamin Asupplementation in the control of young child morbidity andmortality in developing countries (ACC/SCN State of the ArtsSeries, Nutrition and Policy Discussion Paper No 13). Geneva,Switzerland: Administrative Committee on Coordination, Sub-
206 H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209

committee on Nutrition Available [Dec 2002] from; http://www.unsystem.org/scn/.
Beyer, P., & Potrykus, I. (2001). Golden rice: proof of concept andbeyond. Seeds of Health, 1, 2 and 10.
Beyer, P., Salim, A.-B., Xudong, Y., Lucca, P., Schaub, P., Welsch, R.,& Potrykus, I. (2002). ‘‘Golden Rice’’: introducing the B-carotenebiosynthesis pathway into rice endosperm by genetic engineer-ing to defeat vitamin A-deficiency. Journal of Nutrition,132(Suppl.), 506S–510S.
Bhargava, A., Bouis, H., & Scrimshaw, N. (2001). Dietary intakes andsocioeconomic factors are associated with the haemoglobinconcentration of Bangladeshi women. Journal of Nutrition, 131,758–764.
Blechl, A., & Anderson, O. (1996). Expression of a novel high-molecular-weight glutenin subunit gene in transgenic wheat.Nature Biotechnology, 14, 875–879.
Bouis, H., se la Briere, L., Guitierrez, K., Hallman, N., Hassan, O.,Hels, W., Quabili, A., Quisumbing, S., Thilsted, Z., Zihad, Z., &Zohir, S. (1998). Commercial vegetable and polyculture fish pro-duction in Bangladesh: impacts on income, household resourceallocation and nutrition. Washington DC: International FoodPolicy Research Institute.
Bouis, H., Graham, R., & Welch, R. (2000). The Consultative Groupon International Agricultural Research (CGIAR) MicronutrientsProject: justification and objectives. Food and Nutrition Bulletin,21, 374–381.
Buchanan, B., Adamidi, C., Lozano, R., Yee, B., Momma, M.,Kobrehel, K., Ermel, R. M., & Frick, O. (1997). Thioredoxin-linkedmitigation of allergic responses to wheat. Proceedings of theNational Academy of Sciences of the United States of America, 94,5372–5377.
Cahagnier, B., & Melcion, D. (2000). Mycotoxines de Fusarium dansles mais-grains a la recolte: relation entre la presence d’insects(pyrale, sesamie) et la teneur en mycotoxines. In Proceedings ofthe 6th international feed production conference, Piacenza, 27–28November 2000.
Coetzer, C., Corsini, D., Love, S., Pavek, J., & Turner, N. (2001).Control of enzymatic browning in potato (Solanum tuberosumL.) by sense and antisense RNA from tomato polyphenol oxi-dase. Journal of Agricultural and Food Chemistry, 49, 652–657.
Datta, S., & Bouis, H. E. (2000). Application of biotechnology toimproving the nutritional quality of rice. Food and NutritionBulletin, 21, 451–456.
de Pee, S., West, C. E., Permaesih, D., Martuti, S., Muhilal, ?., &Hautvast, J. G. A. J. (1998). Orange fruit is more effective than aredark-green, leafy vegetables in increasing serum concentrationsof retinol and beta-carotene in schoolchildren in Indonesia.American Journal of Clinical Nutrition, 68, 1058–1067.
DellaPenna, D. (1999). Nutritional genomics: manipulating plantmicronutrients to improve human health. Science, 285, 375–379.
DellaPenna, D. (2001). Plant metabolic engineering. Plant Physiol-ogy, 125, 160–163.
Dowd, P. (2000). Indirect reduction of ear moulds and associatedmycotoxins in Bacillus thuringiensis corn under controlled andopen field conditions: utility and limitations. Journal of EconomicEntomology, 93, 1669–1679.
Duvick, J. (2001). Prospects for reducing fumonism contaminationof maize through. Environmental Health Perspectives, 109(Suppl.2), 337–342.
Engel, K., Blaas, W., Gabriel, B., & Beckman, M. (1996). Modernbiotechnology in plant breeding: analysis of glycoalkaloids intransgenic potatoes. In G. R. Takeoka, R. Teranishi, P. J. Williams,& A. Kobayashi (Eds.), Biotechnology for improved foods andflavours (pp. 249–260). Washington: American ChemicalSociety.
Facciotti, M., Bertain, P., & Yuan, L. (1999). Improved stearate phe-notype in transgenic canola expressing a modified acylacyl car-rier protein thioesterase. Nature Biotechnology, 17, 593–597.
Fairweather-Tait, S., & Hurrell, R. F. (1996). Bioavailability of mineralsand trace elements. Nurition Research Reviews, 9, 295–324.
Falco, S. C., Guida, T., Locke, M., Mauvais, J., Sanders, C., Ward, R.T., & Webber, P. (1995). Transgenic canola and soybeans seedswith increased lysine. Bio-Technology, 13, 577–582.
Farrell, D. J., & Martin, E. A. (1998). Strategies to improve thenutritive value of rice bran in poultry diets. III. The addition ofinorganic phosphorus and a phytase to duck diets. British Poul-try Science, 39, 601–611.
Frewer, L., Scholderer, J., & Lambert, N. (in press). Consumeracceptance of functional foods: issue for the future. British FoodJournal.
Gan, S., & Amasino, R. (1995). Inhibition of leaf senescence byautoregaulation production of cytokinin. Science, 270, 1986–1988.
Garcia-Casal, M. N., Layrisse, M., Baron, M. A., Arguello, F., Llovera,D., Ramirez, J., Leets, I., & Tropper, E. (1998). Vitamin A and b-carotene can improve nonheme iron absorption from rice,wheat and corn by humans. Journal of Nutrition, 128, 646–650.
Gelderblom, W. C. A., Seier, J. V., Snijman, P. W., Van Schalkwyk,D. J., Shephard, G. S., & Marasas, W. F. O. (2001). Toxicity ofculture material of Fusarium verticillioides strain MRC 826 tononhuman primates. Environmental Health Perspectives,109(Suppl. 2), 267–276.
Gelderblom, W. C. A., Abel, S., Smuts, C. M., Marnewick, J., Mar-asas, W. F. O., Lemner, E. R., & Ramljak, D. (2001). Fumonisin-induced hepatocarcinogenesis: mechanisms related to cancerinitiation and promotion. Environmental Health Perspectives,109(Suppl. 2), 291–300.
Glahn, R. P., Lee, O. A., & Miller, D. D. (1999). In vitro digestion/Caco-2 cell culture model to determine optimal ascorbic acid toFe ratio in rice cereal. Journal of Food Science, 64, 925–928.
Goto, F. T., Yoshihara, T., Shigemoto, N., Toki, S., & Takaiwa, F.(1999). Iron fortification of rice seed by the soybean Ferritingene. Nature Biotechnology, 17, 282–286.
Graham, R. D., & Welch, R. M. (1996). Breeding for staple-food cropswith high micronutrient density: long-term sustainable agriculturalsolutions to hidden hunger in developing countries (AgriculturalStrategies for Micronutrients Working Paper 3). Washington,DC: International Food Policy Research Institute.
Graham, R. D., Senadhira, D., & Ortiz-Monasterio, I. (1997). Astrategy for breeding staple-food crops with high micronutrientdensity. Soil Science and Plant Nutrition, 43, 1153–1157.
Graham, R. D., Senadhira, D., Beebe, S. E., Iglesias, C., & Ortiz-Monasterio, I. (1999). Breeding for micronutrient density in edibleportions of staple food crops: conventional approaches. FieldCrop Research, 60, 57–80.
Graham, R. D., Welch, R. M., & Bouis, H. E. (2001). Addressingmicronutrient malnutrition through enhancing the nutritionalquality of staple foods: principles, perspectives and knowledgegaps. Advances in Agronomy, 70, 77–142.
Gregorio, G. B. (2002). Progress in breeding for trace minerals instaple crops (Abstract). J. Nutr., 132(special issue), 3.
Guerinot, M. L. (2000). The green revolution strikes Gold. Science,287, 241–243.
Gura, T. (1999). New genes boost rice nutrients. Science, 285, 994–995.
Haas, J., del Mundo, A. M., & Beard, J. (2000). A human feeding trialof iron-enhanced rice. Food and Nutrition Bulletin, 21, 440–444.
Hagenimana, V., & Low, J. (2000). Potential of orange-fleshed sweetpotatoes for raising vitamin A intake in Africa. Food and Nutri-tion Bulletin, 21, 414–418.
H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209 207

Hallberg, L. (1981). Bioavailability of dietary iron in man. AnnualReview of Nutrition, 1, 123–127.
Hauptmann, R., Eschenfeldt, W. H., English, J., & Brinkhaus, F. L.(1997). Enhanced carotenoid accumulation in storage organs ofgenetically engineered plants (United States Patent 5,618,988),Date Filed: 28/10/1994. Washington, DC: United States Patentsand Trademarks Office.
Heyer, A., Lloyd, J., & Kossmann, J. (1999). Production of modifiedpolymeric carbohydrates. Current Opinion in Biotechnology, 10,169–174.
Holm, P. B., Kristiansen, K. N., & Pedersen, H. B. (2002). Transgenicapproaches in commonly consumed cereals to improve ironand zinc content and bioavailability. Journal of Nutrition,132(Suppl.), 514–516.
Horton, S., & Ross, J. (1998). Economic consequences of iron effi-ciency. Ottawa, Canada: The Micronutrient Initiative.
House, W. A. (1999). Trace element bioavailability as exemplifiedby iron and zinc. Field Crop Research, 60, 115–141.
Hunt, J. M. (2002). Reversing productivity losses from iron defi-ciency: the economic case. Journal of Nutrition, 132(Suppl 4),794–801.
IARC. (2002). IARC Monographs on the evaluation of carcinogenicrisks to humans, vol 82, Some traditional herbal medicines, somemycotoxins, napthalene and styrene. Lyon, France: InternationalAgency for Research on Cancer.
Institute of Medicine, Food and Nutrition Board. (2001). Dietaryreference intakes for vitamin a, vitamin k, arsenic, chromium,copper, iodine, iron, manganese, molybdenum, nickel, silicon,vanadium and zinc. Washington, DC: National Academy Press.
Ivy, M. J., Beremond, P. D., & Thomas, T. L. (1998). Strategies formodifying fatty acid composition in transgenic plants. Bio-technology and Genetic Engineering Reviews, 15, 271–288.
Jalani, B. S., Cheah, S. C., Rajanaidu, N., & Darus, A. (1997).Improvement of palm oil through breeding and biotechnology.Journal of American Oil Chemists’ Society, 74, 1451–1455.
Jung, W., Yu, O., Lau, S., O’Keefe, D., Odell, J., Fader, G., & McGo-nigle, B. (2000). Identification and expression of isoflavone syn-thase, the key enzyme for biosynthesis of isoflavones inlegumes. Nature Biotechnology, 18, 208–212.
Katsube, T., Kurisaka, N., Ogawa, M., Maruyama, N., Ohtsuka, R.,Utsumi, S., & Takaiwa, F. (1999). Accumulation of soybean gly-cinin and its assembly with glutelins in rice. Plant Physiology,120, 1063–1073.
Kim, C. S., Kamiya, S., Sato, T., Utsumi, S., & Kito, M. (1990).Improvement of nutritional-value and functional-properties ofsoybean glycinin by protein engineering. Protein Engineering, 3,725–731.
Kinney, A. J. (1996). Designer oils for better nutrition. Nature Bio-technology, 14, 946.
Knutzon, D. (2000). Polyunsaturated fatty acids in plants (Interna-tional Patent Publication No. WO 99/64614, Date Filed: 6/10/1999). Geneva: World Intellectual Property Organization.
Lucca, P., Hurrell, R., & Potrykus, I. (2001). Genetic engineeringapproaches to improve the bioavailability and the level of ironin rice grains. Theoretical and Applied Genetics, 102, 392–397.
Lucca, P., Hurell, R., & Potrykus, I. (2002). Fighting iron deficiencyanemia with iron-rich rice. Journal of the Amercian College ofNutrition, 21, 184S–190S.
Lui, K. S., & Brown, E. A. (1996). Enhancing vegetable oil qualitythrough plant breeding and genetic engineering. Food Technol-ogy, 50, 67–71.
Lui, Q., Singh, S., & Green, A. (2000). Genetic modification of cottonseed oil using inverted-repeat gene-silencing techniques. In J.L. Harwood, & P. J. Quinn (Eds.), Recent advances in the bio-chemistry of plant lipids (pp. 927–929). London: Portland Press.
Martin, E. A., Nolan, J. C., Nitsan, Z., & Farrell, D. J. (1998). Strategiesto improve the nutritive value of rice bran in poultry diets. IV.Effects of addition of fish meal and a microbial phytase toduckling diets on bird performance and amino acid digestibility.British Poultry Science, 39, 612–621.
Martinez-Torres, C., Renzi, M., & Layrisse, M. (1976). Iron absorptionby humans for hemosiderin and ferritin, further studies. Journalof Nutrition, 106, 128–135.
Mazur, B., Krebbers, E., & Tingey, S. (1999). Gene discovery andproduct development for grain quality traits. Science, 285(5426),372–375.
McLaughlin, M. J., Parker, D. R., & Clarke, J. M. (1999). Metals andmacronutrients—food safety issues. Field Crop Research, 60,143–163.
Merryweather-Clarke, A. T., Pointon, J. J., Sherman, J. D., & Robson,K. J. (1997). Global prevalence of putative haemochromatosis.Journal of Medical Genetics, 34, 275–278.
Misawa, N., Yamona, H., Linden, H., de Felipe, M. R., Lucas, M.,Ikenga, H., & Sandmann, G. (1993). Functional expression of theErwinia uredovora biosynthesis gene crtl in transgenic plantsshowing an increase of b-carotene biosynthesis activity andresistance to the bleaching herbicide Norflurazon. Plant Journal,4, 833–840.
Muir, S., Collins, G., Robinson, S., Hughes, S., Bovy, A., De Vos, C.,van Tunen, A., & Verhoeyen, M. (2001). Overexpression of pet-unia chalcone isomerase in tomato results in fruit containingincreased levels of flavonols. Nature Biotechnology, 19, 470–474.
Munaro, F. A., Lopez, J., Lopez, S. E., & Rutz, F. (1996). Effect ofphytase on boiavailability of phosphorus in diets with defattaedrice bran for broiler chickens. Revista da Sociedade Brasileira deZootecnia, 25, 932–943.
Munkvold, G., Hellmich, R., & Rice, L. G. (1999). Comparison offumonisin concentrations in kernels of transgenic Bt maizehybrids and nontransgenic hybrids. Plant Disease, 83, 130–138.
Murata, M., Haruta, M., Murai, N., Tanikawa, N., Nishimura, M.,Homma, S., & Itoh, Y. (2000). Transgenic apple (Mal-us�domestica) shoot showing low browning potential. Journal ofAgricultural and Food Chemistry, 48, 5243–5248.
Murray-Kolb, L. E., Takaiwa, F., Goto, F., Yoshihara, T., Theil, E. C., &Beard, J. L. (2002). Transgenic rice as a source of iron for iron-depleted rats. Journal of Nutrition, 132, 957–960.
Murray-Kolb, L. E., Welch, R., Theil, E. C., & Beard, J. L. (2003).Women with low iron stores absorb iron form soybeans. Amer-ican Journal of Clinical Nutrition, 77, 180–184.
Nelson, T. S., Shieh, T. R., Wodzinski, R. J., & Ware, J. H. (1971).Effects of supplemental phytase on the utilization of phytatephosphorus by chicks. Journal of Nutrition, 101, 1289–1294.
Pasamontes, L., Haiker, M., Wyss, M., Tessier, M., & van Loon, A. P.(1997). Gene cloning, purification and characterization of a heat-stable phytase from the fungus Aspergillus fumigatus. Appliedand Environmental Microbiology, 63, 1696–1700.
Pennarubia, L., Kim, R., Giovannoni, J., Kim, S., & Fischer, R. (1992).Production of the sweet protein monoellin in transgenic plants.Bio-Technology, 10, 561–564.
Pietri, A., & Piva, G. (2000). Occurrence and control of mycotoxinsin maize grown in Italy. In Proceedings of the 6th internationalfeed conference, food safety: current situation and perspectives inthe European Community. Piacenza, Italy. 27–28 November, 2000,(pp. 226–236)
Raboy, V. (1996). Cereal low phytic mutants: a ‘‘global’’ approachto improving mineral nutritional quality. Micronutrients andAgriculture, 2, 15–16.
Rimbach, G., & Pallauf, J. (1993). Enhancment of zinc utilizationfrom phytate-rich soy protein isolate by microbial phytase.Zeitschrift fur Ernahrungswissenschaft, 32, 308–315.
208 H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209

Robinson, N. J., Procter, C. M., Connolly, M. L., & Guerinot, M. L.(1999). A ferric-chelate reductase for iron uptake from soils.Nature, 397, 694–697.
Roller, S., & Harlander, S. (1998). Modern food biotechnology:overview of key issues. In S. Roller, & S. Harlander (Eds.), Geneticmodification in the food industry—a strategy for food qualityimprovement. London, UK: Blackie Academic and Professional.
Romer, S., Fraser, J., Kiano, C., Shipton, C., Misawa, W., Schuch, W.,& Bramley, P. (2000). Elevation of the provitamin A content oftransgenic tomato plants. Nature Biotechnology, 18, 666–669.
Rosegrant, M. W., Paisner, M., Meijer, S., & Witcover, J. (2001).Global food projections to 2020: emerging trends and alternativefutures. Washington DC: International Food Policy ResearchInstitute.
Schledz, M., Al-Babili, S., Lintig, H., Haubruck, S., Rabbani, S., Klei-nig, H., & Beyer, P. (1996). Phytoene synthase from Narcissuspseudonarcissus: functional expression, galactolipid require-ment, topological distribution in chromoplasts and inductionduring flowering. Plant Journal, 10, 781–792.
Schwall, G., Safford, R., Westcott, R., Jeffcoat, A., Tayal, A., Shi, Y.-C., Gidley, M., & Jobling, S. (2000). Production of very-high-amy-lose potato starch by inhibition of SBE A and B. Nature Bio-technology, 18, 551–554.
Scwacka, M., et al. (1999). Transgenic cucumber plants expressingthe thaumatin gene. In Food biotechnology—proceedings of aninternational symposium, Zakopane, Poland. Amsterdam, Neth-erlands: Elsevier
Secor, G., Borovkov, A. Y., McClean, P. E., & Sowokinos, J. R. (1997).Modulation of sugar content in plants (United States Patent5,646,023) Date Filed: 8/7/1997). Washington DC: United StatesPatents and Trademarks Office.
Senadhira, D., & Graham, R. D. (1999). Genetic variation in iron and zincconcentrations in brown rice. Micronutrients in Agriculture, 3, 10–12.
Sharma, D. C. (2001). UN vitamin A campaign in India under fire.Lancet, 358(9295), 1791.
Shewry, P., Tatham, A., Barro, F., Barcelo, P., & Lazzeri, P. (1995).Biotechnology of breadmaking: unraveling and manipulating themulti-protein gluten complex. Bio-Technology, 13, 1185–1190.
Shewry, P., & Tatham, A. (1997). Biotechnology of wheat quality.Journal of the Science of Food and Agriculture, 73, 397–406.
Skikne, B., Fonzo, D., Lynch, S. R., & Cook, J. D. (1997). Bovine fer-ritin bioavailabilty in man. European Journal of Clinical Investiga-tions, 27, 228–233.
Smith, J.C. (1996). The vitamin A–Zinc connection. In Micro-nutrients and agriculture. Washington, DC: Federation of Amer-ican Scientists
Tada, Y., Nakase, M., Adachi, T., Nakamura, R., Shimada, H., Taka-hashi, M., Fujimura, T., & Matsuda, T. (1996). Reduction of 14-16kDa allergenic proteins in transgenic rice plants by antisensegene. FEBS Letters, 391, 341–345.
Taylor, P. G., Martinez-Torres, C., Romano, E. L., & Layrisse, M.(1986). The effect of cysteine-containing peptides released dur-ing meat digestion on iron absorption in humans. AmericanJournal of Clinical Nutrition, 43, 68–71.
Teichmann, H. F., Lopez, J., & Lopez, S. E. (1998). Effect of phytaseon the bioavailability of phosphorus in diets with rice bran forbroilers. Revista Braasileira de Zootechnia, 27, 338–344.
Theil, E. C., Burton, J. W., & Beard, J. L. (1997). A sustainable solutionfor dietary iron deficiency through plant biotechnology andbreeding to increase seed ferritin control. European Journal ofClinical Nutrition, 21(Suppl. 4), S28–S31.
Theologis, A., & Sato, T. (2000). Control of fruit ripening throughgenetic control of ACC synthase synthesis (United States Patent6,156,956, Date Filed: 2/3/1998). Washington DC: United StatesPatents and Trademark Office.
Tyssandier, V., Lyan, B., & Borel, P. (2001). Main factors governingthe transfer of caretenoids from emulsion lipid droplets tomi-celles. Biochimica and and Biophysica Acta, 1533, 285–292.
UNICEF/UNU/WHO/MI Technical Workshop (1999). Preventing irondeficiency in women and children: background and consensus onkey technical issues and resources for advocacy, planning andimplementing national programmes. Boston MA: InternationalNutrition Foundation: Ottawa, Canada: The MicronutrientInitiative
Vageeshbabu, H., & Chopra, V. (1997). Genetic and biotechnologi-cal approaches for reducing glucosinolates from rapeseed-mus-tard meal. Journal of Plant Biochemistry and Biotechnology, 6, 53–62.
Van Campen, D. R., & Glahn, R. P. (1999). Micronutrient bioavail-ability techniques: accuracy, problems and limitations. FieldCrop Research, 60, 93–113.
Vasil, I., & Anderson, O. (1997). Genetic engineering of wheat glu-ten. Trends in Plant Science, 2, 292–297.
Venkatramesh, M. (2000). Transgenic plants containing altered levelsof sterol compounds and tocopherols (International Patent Pub-lication No. WO 00/61771, Date Filed: 19/10/2000). Geneva:World Intellectual Property Organization.
Visser, R., Somhorst, I., Kuipers, G., Ruys, N., Feenstra, W., &Jacobsen, E. (1991). Inhibition of the expression of the gene forgranule-bound starch synthase in potato by antisense con-structs. Molecular and General Genetics, 225, 289–296.
Visser, R., Suurs, L., Steeneken, P., & Jacobsen, E. (1997). Somephysicochemical properties of amylose-free potato starch.Starch/Staerke, 49, 443–448.
Voelker, T. (1997). Transgenic manipulation of edible oilseeds. InT. Johns, & J. T. Romeo (Eds.), Functionality of Food Phytochem-icals (pp. 223–236). New York: Plenum Press.
Welch, R., & Graham, R. (1999). A new paradigm for world agri-culture: meeting human needs, productive, sustainable nutri-tious. Field Crop Research, 60, 1–10.
Welch, R. M. (1986). Effects of nutrient deficiencies on seed pro-duction and quality. Advances in Plant Nutrition, 2, 205–247.
Welch, R. M. (1993). Zinc concentrations and forms in plants forhumans and animals. In A. D. Robson (Ed.), Zinc in soils andplants (pp. 183–195). Kluwer Academic Publishers, Boston MA.
Welch, R. M., William, A., House, A. H., Beebe, S., Senadhira, D.,Gregoria, G., & Cheng, Z. (2000). Testing iron and zinc bioavai-labilty in genetically enriched bean (Phaselous Vulgaris L.) andrice (Oryza Sativa L.) using a rat model. Food and Nutrition Bul-letin, 21, 428–433.
Welch, R. M. (2002). Breeding strategies for biofortified staple plantfoods to reduce micronutrient malnutrition globally. Journal ofNutrition, 132(Suppl.), 495–499.
Wise, A. (1995). Phytate and zinc bioavailability. International Jour-nal of Food Science and Nutrition, 46, 53–63.
World Bank. (1994). Enriching lives: overcoming vitamin and mineralmalnutrition in developing countries (Development in PracticeSeries). Washington DC: The World Bank.
Ye, X., Al Babili, S., Kloeti, A., Zhang, J., Lucca, P., Beyer, P., &Potrykus, I. (2000). Engineering the provitamin A (b-carotene)biosynthetic pathway into (carotenoid-free) rice endosperm.Science, 287, 303–305.
Yi, Z., Kornegay, E. T., & Veit, H. P. (1996). Supplemental microbialphytase improves zinc utilization in broilers. Poultry Science, 75,540–546.
Zheng, A., Sumi, K., Tanaka, K., & Murai, N. (1995). The beanseed storage protein b-phaseolin is synthesized, processedand accumulated in the vacuolar type-II protein bodiesof transgenic rice endosperm. Plant Physiology, 109, 777–786.
H.E. Bouis et al. / Trends in Food Science & Technology 14 (2003) 191–209 209




















