
Immunohistochemical evidence for coexistence of glycine and GABA in nerve terminals on cat spinal...
10
Journal of Chemical Neuroanatomy, Vol. 5:1 l0 (1992) Immunohistochemical Evidence for Coexistence of Methionine-Enkephalin and Tyrosine Hydroxylase in Neurons of the Locus Coeruleus Complex Projecting to the Spinal Cord of the Cat Huang Zhuo, Simon J. Fung, 11. Krishna Reddy and Charles D. Barnes Department of Veterinary and Comparative Anatomy, Pharmacology and Physiology, College of Veterinary Medicine, Washington State University, Pullman, WA 99164-6520, USA ABSTRACT Previous studies have revealed the presence of pontospinal neurons with either methionine-enkephalin- or tyrosine hydroxylase-like immunoreactivity in the dorsolateral pontine tegmentum of the cat. Using a combined fast blue retrograde transport technique and simultaneous immunofluorescence histo- chemistry, the present study was designed to reveal the coexistence of enkephalin and tyrosine hydroxy- lase in cat coerulospinal neurons and to determine if and to what extent the coerulospinal pathway is heterogeneous. Fast blue-labelled neurons with tyrosine hydroxylase- and enkephalin-like immunoreactivities were found in the nucleus locus coeruleus, nucleus subcoeruleus, K611iker-Fuse nucleus, and the medial and lateral parabrachial nuclei. Approximately 87% of tyrosine hydroxylase-like immunoreactive neurons had enkephalin-like immunoreactivity, whereas about 76% of the enkephalin-like immunoreactive neurons had tyrosine hydroxylase-like immunoreactivity. About 71% of all coerulospinal neurons exhibited both tyrosine hydroxylase- and enkephalin-like immunoreactivities. These findings indicate that coerulospinal activity may lead to spinal cord effects reflecting both norepinephrine and enkephalin activity in most cases but do not rule out each transmitter's isolated functions. KEYWORDS: Immunofluorescence Catecholamines Opioid peptides Fast blue INTRODUCTION The dorsolateral pontine tegmentum of mammals is characterized by the presence of a large population of catecholaminergic neurons (Dahlstrom and Fuxe, 1964; Fuxe et al., 1970; Holets et at., 1988; Jones and Moore. 1974; Jones and Friedman, 1983; Leger et al., 1983; Stevens et al., 1982; Wiklund et al., 1981). In cats, this catecholaminergic neuronal population, estimated at about 9000 cells unilaterally (Wiklund et al., 1981), is scattered over a large area of the dorsolateral pons, namely, in the nuclei locus coeruleus (LC), subcoeruleus (SC), parabrachialis medialis (NPM) and lateralis (NPL), and the K611iker-Fuse nucleus (KF) (Leger et al., 1983; Maeda et al., 1973). These nuclear groups are often referred to as the LC complex, and are the primary source o f spinal norepinephrine (NE) (Commissiong et al., 1978; Nygren and Olson, 1977; Reddy et al., 1989; Satoh et al., 1977; Stevens et al., 1982, 1985; Addressee for correspondence: Dr C. D. Barnes, Department of VCAPP, Washington State University, Pullman, WA 99164-6520, USA. 0891 0618/92/010001 10505.00 © 1992 by John Wiley and Sons Ltd Westlund and Coulter, 1980; Westlund et at., 1981, 1983). Since the discovery of the two opiate pentapeptides leucine-enkaphalin and methionine-enkephalin in the brain (Hughes et al., 1975) several investi- gators have explored the distributions of these two endogenous opioid peptides in specific areas of the central nervous system. Accordingly, enkephalin (ENK)-like immunoreactive neurons have been described in the LC complex of humans (Bouras et al., 1984), rats (Finley et al., 1981 ; Johnson et al., 1979; Khachaturian et al., 1983, Murakami et al., 1989; Sar et al., 1978, Zamir et al., 1985) and cats (Charnay et al., 1982; Leger et al., 1983, Reddy et al., 1990). In addition, ENK-like immunoreactive terminals have also been localized in the spinal cord of the cat (Conrath-Verrier et al., 1983: Glazer and Basbaum, 1981), suggesting an opioid component in the pontospinal system. The existence of ENK in the catecholaminergic neurons in the cat LC com- plex has been demonstrated indirectly on adjacent sections (Charnay et al., 1982). Similarly, an earlier report from this laboratory alluded to the presence
Transcript of Immunohistochemical evidence for coexistence of glycine and GABA in nerve terminals on cat spinal...

Journal of Chemical Neuroanatomy, Vol. 5:1 l0 (1992)
Immunohistochemical Evidence for Coexistence of Methionine-Enkephalin and Tyrosine Hydroxylase in Neurons of the Locus Coeruleus Complex Projecting to the Spinal Cord of the Cat
Huang Zhuo, Simon J. Fung, 11. Krishna Reddy and Charles D. Barnes
Department of Veterinary and Comparative Anatomy, Pharmacology and Physiology, College of Veterinary Medicine, Washington State University, Pullman, WA 99164-6520, USA
ABSTRACT
Previous studies have revealed the presence of pontospinal neurons with either methionine-enkephalin- or tyrosine hydroxylase-like immunoreactivity in the dorsolateral pontine tegmentum of the cat. Using a combined fast blue retrograde transport technique and simultaneous immunofluorescence histo- chemistry, the present study was designed to reveal the coexistence of enkephalin and tyrosine hydroxy- lase in cat coerulospinal neurons and to determine if and to what extent the coerulospinal pathway is heterogeneous.
Fast blue-labelled neurons with tyrosine hydroxylase- and enkephalin-like immunoreactivities were found in the nucleus locus coeruleus, nucleus subcoeruleus, K611iker-Fuse nucleus, and the medial and lateral parabrachial nuclei. Approximately 87% of tyrosine hydroxylase-like immunoreactive neurons had enkephalin-like immunoreactivity, whereas about 76% of the enkephalin-like immunoreactive neurons had tyrosine hydroxylase-like immunoreactivity. About 71% of all coerulospinal neurons exhibited both tyrosine hydroxylase- and enkephalin-like immunoreactivities. These findings indicate that coerulospinal activity may lead to spinal cord effects reflecting both norepinephrine and enkephalin activity in most cases but do not rule out each transmitter's isolated functions.
KEY WORDS: Immunofluorescence Catecholamines Opioid peptides Fast blue
I N T R O D U C T I O N
The dorsolateral pontine tegmentum of mammals is characterized by the presence of a large population of catecholaminergic neurons (Dahlstrom and Fuxe, 1964; Fuxe et al., 1970; Holets et at. , 1988; Jones and Moore. 1974; Jones and Friedman, 1983; Leger et al. , 1983; Stevens et al., 1982; Wiklund et al. , 1981). In cats, this catecholaminergic neuronal population, estimated at about 9000 cells unilaterally (Wiklund et al. , 1981), is scattered over a large area of the dorsolateral pons, namely, in the nuclei locus coeruleus (LC), subcoeruleus (SC), parabrachialis medialis (NPM) and lateralis (NPL), and the K611iker-Fuse nucleus (KF) (Leger e t al. , 1983; Maeda et al. , 1973). These nuclear groups are often referred to as the LC complex, and are the primary source o f spinal norepinephrine (NE) (Commissiong et al. , 1978; Nygren and Olson, 1977; Reddy et al. , 1989; Satoh et al. , 1977; Stevens e t al. , 1982, 1985;
Addressee for correspondence: Dr C. D. Barnes, Department of VCAPP, Washington State University, Pullman, WA 99164-6520, USA.
0891 0618/92/010001 10505.00 © 1992 by John Wiley and Sons Ltd
Westlund and Coulter, 1980; Westlund et at., 1981, 1983).
Since the discovery of the two opiate pentapeptides leucine-enkaphalin and methionine-enkephalin in the brain (Hughes et al., 1975) several investi- gators have explored the distributions of these two endogenous opioid peptides in specific areas of the central nervous system. Accordingly, enkephalin (ENK)-like immunoreactive neurons have been described in the LC complex of humans (Bouras et al. , 1984), rats (Finley et al. , 1981 ; Johnson et al., 1979; Khachaturian et al. , 1983, Murakami et al., 1989; Sar et al., 1978, Zamir et al., 1985) and cats (Charnay et al. , 1982; Leger et al., 1983, Reddy et al. , 1990). In addition, ENK-like immunoreactive terminals have also been localized in the spinal cord of the cat (Conrath-Verrier et al. , 1983: Glazer and Basbaum, 1981), suggesting an opioid component in the pontospinal system. The existence of ENK in the catecholaminergic neurons in the cat LC com- plex has been demonstrated indirectly on adjacent sections (Charnay et al. , 1982). Similarly, an earlier report from this laboratory alluded to the presence

2 H. Zhuoetal .
of ENK in many of the catecholaminergic neurons that project to the spinal cord (Reddy et al., 1990). As part of a long-term physiological study on the stimulation-produced effects of these cells on the spinal cord of the cat, we attempted to determine if any of the coerulospinal neurons co-localize catecholamine and ENK. To achieve this we used the technique of Staines et al. (1988) to immunostain for ENK and tyrosine hydroxylase (TH) in neurons that were retrogradely labelled by injection of fast blue (FB) into the lumbar segments of the spinal cord.
MATERIALS AND METHODS
Six adult cats (weighing 2.5~1.4 kg), under sodium pentobarbital anesthesia (35mg/kg, i.p.), had laminectomies performed at the appropriate level of the lumbar spinal cord. Eight microlitres of 5% FB in saline was injected bilaterally into the sixth and seventh (Lr-L7) lumbar segments of the spinal cord, with eight penetrations, using a glass rnicropipette attached to a Hamilton syringe. The injections were made to include both the dorsal and ventral horns of the grey matter of the spinal cord. Following a 2- week survival period, the animals were reanesthe- tized, and colchicine (300-400 ltg in saline) was administered intracerebroventricularly. Twenty- four hours later, the animals were anesthetized again and perfused intracardially with physiological saline followed by ice-cold 3.8% paraformaldehyde in 0.1 M-phosphate buffer (pH 7.4). The brainstem and the spinal cord segments injected with FB were removed and soaked overnight in 30% sucrose in 0.1 M-phosphate buffer (pH 7.4) at 4°C.
The appropriate brainstem level was frozen sectioned at 20 lam thickness with a cryostat and the tissue sections were collected in 0.1 m-phosphate- buffered saline (PBS). Free-floating brainstem sec- tions were processed by the method of Staines et al. (1988) as follows. The sections were first incubated in a mixture of primary antisera containing mouse anti-TH (Boehringer Mannheim Biochem. USA) and rabbit anti-methionine-ENK (Incstar Corp., USA) in 0.1 M-PBS (dilution l:l:1000) for 48 h at 4°C, and then incubated in a mixture of secondary antisera containing fluorescein-labelled, goat anti- mouse IgG and L-rhodamine-labelled goat anti- rabbit IgG (Boehringer Mannheim Biochem.) in 0.1 M-PBS (1:1:50) for 1 h at room temperature. The sections were thoroughly washed (3 x 10min) in PBS after each reaction. The processed sections were wet-mounted onto glass slides and cover- slipped with a mixture containing glycerol and PBS (3:1).
The brainstem sections were examined and photographed with an Olympus (BH2) fluorescence photomicroscope. The filter systems used to visual- ize the three fluorescent markers were as follows: rhodamine, excitation 546 nm G, dichroic mirror G, barrier filter 590 nm; fluorescein, excitation 405-
435 nm B + EY455, dichroic mirror B, barrier filter 515nm; FB, excitation 334-365nm U, dichroic mirror U, barrier filter 435 nm. In three cats, photo- montages of one representative section in every 30 sections were made using these three filter systems. The different cell types were counted from the photomontages.
The spinal cord segments (injection sites) were sectioned at 40 lam thickness. The sections were collected in 0.1 M-PBS and wet-mounted on glass slides. The injection sites in the lumbar segments of the spinal cord were identified and drawn using camera lucida.
The terminology used for the nuclei of the cat brainstem was adapted from Leger and Hernandez- Nicaise (1980) and the stereotaxic levels referred to in this paper conformed to the description of Berman (1968).
To ascertain the immunostaining specificity, five control experiments were performed on representa- tive sections from one animal. These experiments are summarized below.
Experiment 1 The sections were incubated separately with the mouse anti-TH or rabbit anti-ENK (1:1000) anti- serum. The sections were then reacted with specific homologous secondary antibody (1:50); the sections incubated with mouse anti-TH were reacted with fluorescein-lahelled, goat anti-mouse IgG and those incubated in rabbit anti-ENK were reacted with L- rhodamine-labelled, goat anti-rabbit IgG.
Experiment 2 Sections were separately incubated with primary antibodies as in experiment 1 and then reacted in a mixture of two secondary antibodies containing fluorescein-labelled, goat anti-mouse IgG and L- rhodamine-labelled, goat anti-rabbit IgG in PBS (1:1:50).
Experiment 3 After the tissue sections were first incubated in the primary antibodies similar to those described in experiment 1, they were reacted with the non- specific heterologous secondary antiserai After microscopic examination and cofirmation of a negative immunoreactivity, these sections were washed in PBS and re-incubated in the specific homologous antibodies.
Experiments 4 and 5 In these experiments the primary (experiment 4) or the specific homologous secondary (experiment 5) antibodies were omitted.
The control experiments resulted in: (I) immuno- staining characteristic of the corresponding primary antibodies (Fig. 1); (2) no immunoreactivity, when treated with heterologous secondary antibodies or when primary or homologous secondary anti-

E N K
A
C
ENK and TH in Cat Coerulospinal Neurons
TH
B
O m
Fig. I. Results of control experiment 2. Top: photomicrographs of neurons from nucleus subcoeruleus. Section incubated with mouse anti-TH and then reacted in a mixture containing fluorescein-labelled, goat anti-mouse IgG and L-rhodamine-labelled, goat anti-rabbit IgG shows lack of affinity for the heterologous secondary antibody (A). Bottom: photomicrographs of neurons from nucleus locus coeruleus. Section processed by rabbit anti-ENK and reacted in a mixture of secondary antibodies (same as above) also exhibits a lack of affinity for the heterologous secondary antibody (D). Calibration bar = 50 lam.
bodies were omitted; (3) emission of fluorescence by fluorescein and L-rhodamine only when appropriate filter systems were used.
RESULTS
Bilateral injection of FB into the sixth and seventh lumbar segments of the spinal cord caused the retro- grade marker to spread into the dorsal and ventral grey matter and into the adjacent white matter.
Following injection of FB and subsequent pro- cessing for simultaneous immunofluorescence stain- ing for TH- and ENK-like immunoreactivities, several cell types were found in the LC complex, i.e., nuclei LC, SC, NPM and NPL, and the KF. Neurons showing either TH-or ENK-like immunoreactivity were found in all pontine areas described previously (Charnay et al., 1982; Hunt and Lovick, 1982; Leger and Hernandez-Nicaise, 1980; Leger et al., 1983; Maeda et al., 1973; Reddy et al., 1989, 1990; Wiklund

H. Zhuo et al.
P1.5
°° 7+
5 4 3
o*ao* ;-----, ***---~--
o e
-1
. . . . - 3
L i - 4 1
H _ _ d
0
o
P3.0 0
!
L_4 3 2 1
i ~ J
o
P2.0 \
+"+ .++'i ¢.. " "I-+?
:+"* I " 3 2
L
-1
- 2 H
q
- 3
- - - - 4
1 + 4 ,+
i P4,0
i
4 3 2
L
- - - 1
- 2
. . . . I _ 4
Fig. 2. Representative plottings of two kinds of TH- and ENK-Iik¢ immunoreactive neurons in the dorsolatera! pontine tegmentum (P 1.5- P4 stereotaxic planes) o f one animal (cat no. 170). FB-labelled neurons immunostained with TH- and ENK,like immunoreactivities ( m); neurons with TH- and ENK-like immunoreactivities but not labelled with FB ( o ).
et al., 1981); thus, they will not be described here in detail. Neurons containing both TH- and ENK-like immunoreactivities were present throughout the rostrocaudal extent of the LC complex from coronal levels P1.5 to P4 (Fig. 2). Neurons that were labelled with FB and contained evidence for both TH- and ENK-like immunoreactivities (triple-labelled neurons) were observed mainly in the caudoventral portion of the LC complex (Figs 2 and 3). Triple- labelled neurons were rarely found rostral to level PI.5. In the LC, triple-labelled neurons were commonly seen in its caudoventral portion (P3-P4). Additionally, such neurons were consistently observed in the SC (P1.5-P3), NPM (P2-P3), NPL (P1.5-P4), and the KF (P2-P3).
Figure 3 is a plot of all spinally projecting neurons in the dorsolateral pontine tegmentum of a repre- sentative animal (cat no. 170). Included are FB- labelled neurons that are immunoreactive to either
TH or ENK, those that are immunoreactive to both TH and EN K (same as those plotted in Fig. 2) and those that are immunoreactive to neither TH nor ENK. These neurons extend ventrolaterally from the ventral half of the mesencephalic tract of the tn- geminal nerve to the ventral margin of the rostral cerebeUar peduncle and rostrocaudally from coronal levels P1.5 to P4. Samples of neurons with both TH- and ENK-like immunoreactivities and retrogradely labelled with FB are illustrated in Fig. 4. In the LC complex, triple-labelled neurons are either oval (Fig. 4), fusiform (Fig. 4) or polygonal in shape (picture not shown) with the latter cell type being distributed mainly in the KF.
Table 1 presents the distribution of variously labelled cell types at different levels (P1.5-P4) of the LC complex (cat no. 170). About 87% of all catecholaminergic neurons in the LC complex were found to have ENK-like immunoreactivity, while

-.: J
e ) $
5 4 3
P1.5
- 1 -1
-2 H
- 3
- 4 1
ENK and TH in Cat Coerulospinal Neurons 5
P3.0
t l
5 4 3 2
- 3
- - - 2
- 4
-1
- 2
- 3
- 4
P2.0
5 4 3 2
L
- 2
- 3
- 4 1
H
P4.0
e m m x
x •
5 4 3
L Fig. 3. A plotting of four kinds of spinally projecting neurons in the dorsolateral pontine tegmentum of the cat (no. 170). TH-ENK-FB- labelled neurons ( ~-); TH-FB-labelled neurons (--~); ENK-FB-labelled neurons ( x ); FB-only-labelled neurons (O).
close to 76% of all enkephalinergic neurons showed TH-like immunoreactivity. In addition, approxi- mately 71% of the coerulospinal neurons exhibited both TH- and ENK-like immunoreactivities, 7.7% exhibited TH-like immunoreactivity alone and 7.5% contained ENK alone. In about 14% of the spinally projecting neurons there was no immuno- staining for either TH or ENK.
DISCUSSION
This study utilized a combined retrograde transport of FB and simultaneous immunofluorescence histo- chemistry. Several problems could arise when using this method; they include (1) the possibility of cross- reaction between the primary antibodies and the in- appropriate secondary antibodies (Erlandsen et al., 1975; Wessendorf and Elde, 1985); (2) potential for
cross-reaction between the secondary antibodies themselves (Grimmelikhuijzen, 1983); or (3) a tend- ency to overestimate the immunoreactive cells due to inappropriate selection of secondary antisera (Pilowsky et al., 1986). To avoid these pitfalls, certain control studies are believed to be absolutely necessary (Staines et al., 1988). The potential inter- actions between the various antisera used in this study were rigorously tested and no inappropriate interactions among the primary antibodies and the secondary antisera were found. Additionally, fluorescein is known to fluoresce red, simulating the fluorescence ofrhodamine; conversely, tetramethyl- rhodamine fluoresces green (Wessendorf and Elde, 1985), leading to erroneous cell counts. We employed fluorescein and lissamine rhodamine- conjugated secondary antibodies and found the two fluorochromes to fluoresce only when appropriate filter systems were used.

6 H. Z h u o e t a l .
Fig. 4. Photomicrographs of neurons from nucleus locus coeruleus (LC) (A~C), nucleus subcoeruleus (D-F) and K611iker-Fuse nucleus (KF) (G-I). The left column (A, D and G), middle column (B, E and H) and right column (C, F and I) depict FB-labelled, ENK-like and TH-like immunoreactive neurons, respectively. Arrowheads indicate triple-labelled neurons. Calibration bar = 30 lam.
The present s tudy demons t r a t e s the presence o f LC-complex neurons tha t show evidence for bo th
T H - and E N K - l i k e immunoreac t iv i t ies . The results o f this s tudy are consis tent with ear l ier repor ts
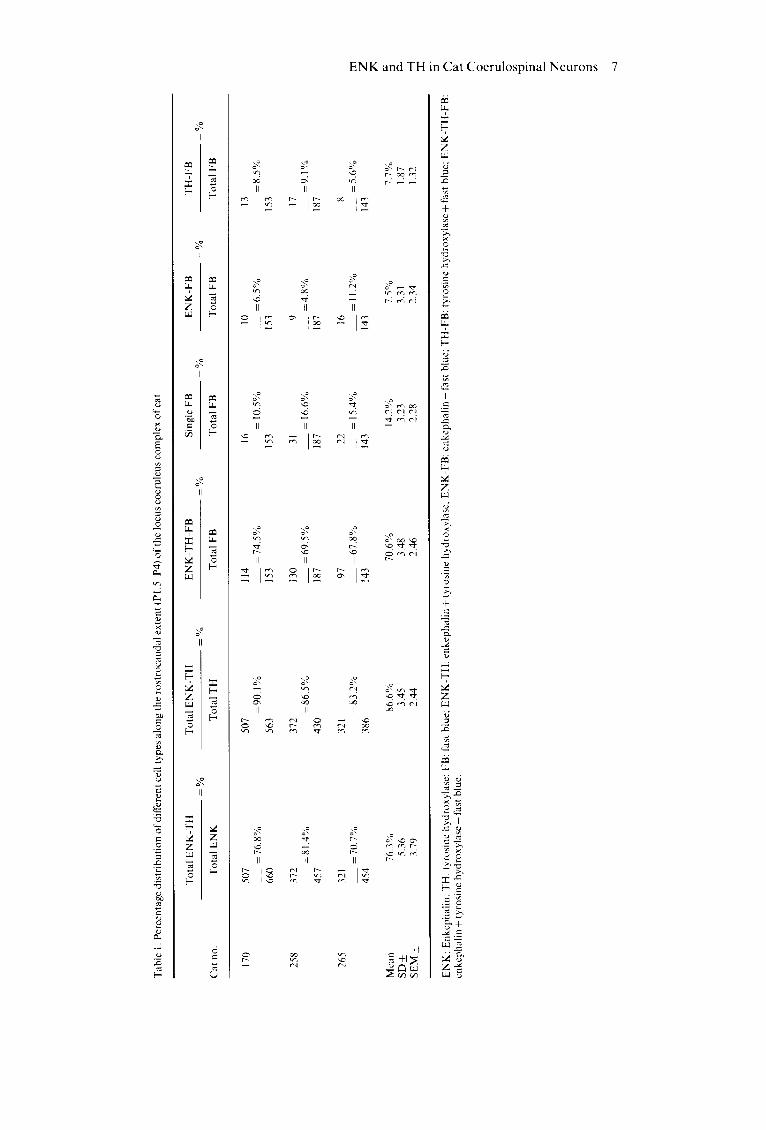
Tab
le 1
. Per
cent
age
dist
ribu
tion
of
diff
eren
t ce
ll ty
pes
alo
ng
the
ro
stro
cau
dal
ext
ent
(P1.
5 P
4) o
f th
e lo
cus
coer
uleu
s co
mp
lex
of
cat
To
tal
EN
K-T
H
Tot
al E
NK
-TH
E
NK
-TH
-FB
S
ingl
e F
B
EN
K-F
B
TH
-FB
=
%
=%
=
%
%
=%
=
%
Cat
no.
T
otal
EN
K
Tot
al T
H
Tot
al F
B
To
tal
FB
T
ota
l F
B
To
tal
FB
170
258
265
507
507
114
16
10
13
=7
6.8
%
=9
0.1
%
=7
4.5
%
= 1
0.5%
=
6.5
%
=8
.5%
66
0 56
3 15
3 15
3 15
3 15
3
372
372
130
31
9 17
=
81.
4%
= 8
6.5%
=
69
.5%
=
16.
6%
=4
.8%
=
9.1
%
457
430
187
187
187
187
321
321
97
22
16
8 =
70
.7%
=
83
.2%
=
67
.8%
=
15.
4°/.
= 1
1.2%
=
5.6
%
454
386
143
143
143
143
Mea
n 76
.3%
86
.6%
70
.6%
14
.2%
7.
5%
7.7%
SD
_+
5.36
3.
45
3.48
3.
23
3.31
1.
87
SE
M +
3.
79
2.44
2.
46
2.28
2.
34
1.32
z 7~
.-q :Z
EN
K:
Enk
cpha
lin;
TH
: ty
rosi
ne h
ydro
xyla
se,
FB
: fa
st b
lue;
EN
K-T
H:
enke
phal
in +
tyr
osin
e hy
drox
ylas
e; E
NK
-FB
: en
keph
alin
+ f
ast
blue
: T
H-F
B:
tyro
sine
hyd
roxy
lase
+ f
ast
blue
; E
NK
-Tt
I-F
B:
enke
phal
in +
tyr
osin
e hy
drox
ylas
e +
fas
t bl
ue.
¢3
© '-"1
'z3 Z
-,-,3

8 H. Zhuoetal.
(Charnay et al., 1982; Leger et al., 1983) in which pairs of adjacent sections were used to examine the coexistence of ENK and TH in the LC. Quantitative analysis of these data suggests that approximately 87% of all catecholaminergic neurons counted exhibited ENK-like immunoreactivity and 76% of the enkephalinergic neurons had TH-like immunoreactivity.
The neurons that project to the spinal cord and show evidence for both TH- and ENK-like immunoreactivities (triple-labelled) were limited to the caudoventral part of the LC complex; this is in agreement with previous studies using retrograde markers (Hancock and Fougerousse, 1976; Hayes and Rustioni, 1981; Nakazato, 1987). These data also revealed that a majority (71%) of all spinally projecting neurons of the dorsolateral pontine teg- mentum co-localize TH and ENK, and the present findings compliment those earlier reports suggesting that, in the cat, ENK is co-localized in many of the catecholaminergic neurons in the LC complex projecting to the spinal cord (Reddy et al., 1989, 1990).
Our finding of a dual presence of TH- and ENK- like immunoreactivities within individual coerulo- spinal neurons accords well with data from other indirect studies. There are biochemical data suggest- ing that NE and ENK are co-stored in the dense- cored vesicles of axonal terminals in cat stellate ganglion (Bastiaensen et al., 1988). Earlier studies by Costa and co-workers have shown that 8-Br- cyclic AMP triggered an increase in cultured bovine adrenal medullary chromaffin cell contents of proENK mRNA (Quach et al., 1984) as well as TH activity (Kumakura et al., 1979), suggesting a common mode of cellular induction of ENK and NE synthesis. Other pharmacological studies have also provided evidence of co-release of these two transmitters from the chromaffin cells in vitro (Livett et al., 1981). Although these findings tend to support the dual presence of NE and ENK in the soma and axon terminals ofcoerulospinal neurons, it may be possible that in triple-labelled cells, only one antigen might be contained in the spinal trajec- tory while the other contributed to local, recurrent axon collaterals or other efferent pathways.
The functional role of NE and ENK co-trans- mission in spinally projecting systems is not yet known. Anatomical studies, however, have estab- lished independently that NE- (Mizukawa, 1980) and ENK-containing (Atsumi and Sakamoto, 1987; Bouras et al., 1984; Honda and Lee, 1985; Tashiro et al., 1989) axon terminals contact spinal cord cells including motoneurons. Electrophysiological studies have indicated that both LC stimulation (Barnes et aL, 1989) and exogenously applied NE (Connell and Wallis, 1989; Fung et al., 1989) excite spinal motoneurons. When applied iontophoreti- cally, ENK is reported to produce no effect on the motoneuron resting membrane potential (Duggan and Zhao, 1984; Zieglgansberger and Tulloch,
1979), although an excitatory action is observed on Renshaw cells (Davies and Dray, 1976). Through the latter action, the ENK transmitter could reduce the segmental motor outflow indirectly. While the NE coerulospinal action is excitatory on the moto- neuron, the ENK co-transmitter may act antagonis- tically to limit any excessive firing of motoneurons. A recent study also points to the inhibitory ~t-opioid receptor regulation of NE release from various ter- minal trajectories of LC neurons (Werling et al., 1987). Taken together, these results suggest that the co-localized classic (NE) and peptide (ENK) trans- mitters may interact pre- and postsynaptically upon release to maintain a coordinated motoneuronal output.
The spinal dorsal horn contains an abundance of ENK- (Glazer and Basbaum, 1981) and NE-like immunoreactive (Nygren and Olson, 1977) vari- cosities, with the latter being reduced following bilateral lesioning of the LC in rats (Commissiong et al., 1978). Physiological evidences indicate that ENK (Randic and Miletic, 1978) and noradrenergic LC neurons (Mokha et aL, 1985) exert an inhibitory action on pain-evoked activity of dorsal horn inter- neurons. It appears that both ENK and NE trans- mitters may act in concert to produce the inhibitory response of pain-activated interneuronal activity: In corollary fashion, the co-transmitter (ENK) may co-operate or amplify the postsynaptic effects of the classic (NE) transmitter towards its target neurons.
In conclusion, our findings suggest that feline coerulospinal neurons may affect the spinal sensory and motor systems in myriad, interactive ways, through their classic neurotransmitter (NE) and peptide (ENK) co-transmitter.
ACKNOWLEDGEMENTS
We wish to express our appreciation to Catherine Smith, Paula Perron-Bates and Rebecca Thompson for help with editing and word processing. This work was supported by NIH grant NS24388.
REFERENCES
Atsumi, S. and Sakamoto, H. (1987). Enkephalin-like immunoreactive axon terminals make synap~s with a- motoneurons in the chicken. Brain Res. 409, 187-192.
Barnes, C. D., Fung, S. J. and Pompeiano, O. (1989). Descending catecholaminergic modulation of spinal cord reflexes in cat and rat. Modulation of Defined Vertebrate Neural Circuits. Ann. N.Y. Acad. Sci. 563, 45-58.
Bastiaensen, E., Miserez, B. and Potter, W. D. (1988). Subcellular fractionation of bovine ganglion stollatum: Co-storage of noradrenaline, Met-enkaphalin and neuropeptide Y in large 'dense-cored' vesicles. Brain Res. 442, 124-130.
Berman, A. L. (1968). The Brain Stem of the Cat: A Cytoarchitectonic Atlas with Stereotaxic Coordinates. The University of Wisconsin Press, Madison, Milwaukee and London.

Bouras, C., Taban, C. H. and Constantinidis, J. (1984). Mapping of enkephalins in human brain. An immuno- histofluorescence study on brains from patients with senile and presenile dementia. Neuroscience 12, 179 190.
Charnay, Y., Leger, L., Dray, F., Berod, A., Jouvet, M., Pujol, J. F. and Dubois, P. M. (1982). Evidence for the presence ofenkephalin ofcatecholaminergic neurons in cat locus coeruleus. Neurosci. Lett. 30, 147 151.
Commissiong, J. W., Hellstrom, S. O. and Neff, N. H. (1978). A new projection from locus coeruleus to the spinal ventral columns: Histochemical and biochemical evidence. Brain Res. 148, 207-213.
Connell, L. A. and Wallis, D. I. (1989). 5-Hydroxytrypta- mine depolarizes neonatal rat motoneurons through a receptor unrelated to an identified binding site. Neuropharmacology 28, 625-634.
Conrath-Verrier, M., Dietl, M., Arluison, M., Cesselin, F., Bourgoin, S. and Hamon, M. (1983). Localization of met-enkephalin-like immunoreactivity within pain- related nuclei of cervical spinal cord, brainstem and midbrain in the cat. Brain Res. Bull. 11,587 604.
Dahlstrom, A. and Fuxe, K. (1964). Evidence for the existence of monoamine-containing neurons in the cen- tral nervous system. I. Demonstration ofmonoamines in the cell bodies of brainstem neurons. Acta Physiol. Scand. 62, Suppl. 232, 1-55.
Davies, J. and Dray, A. (1976). Effects of enkephalin and morphine on Renshaw cells in feline spinal cord. Nature 262, 603-604.
Duggan, A. W. and Zhao, Z. Q. (1984). Microelectro- phoretic administration ofnaloxone near motoneurons fails to reproduce the effects of systemic naloxone in anesthetized cats. Neurosci. Lett. 45, 305-310.
Erlandsen, S. L., Parsons, J. A., Burke, J. P., Redick, J. A., Van Orden, D. E. and Van Order, L. S. (1975). A modification of the unlabelled antibody enzyme method using heterologous antisera for the light micro- scopic and ultrastructural localization of insulin, gluca- gon and growth hormone. J. Histochem. Cytochem. 23, 666 675.
Finley, J. C. W., Maderdrut, J. L. and Petrusz, P. (1981). The immunocytochemical localization ofenkephalin in the central nervous system of the rat. J. Comp. Neurol. 198, 541-565.
Fung, S. J., Manzoni, D., Chan, J. Y. H. and Barnes, C. D. (1989). Concordant depolarizing responses pro- duced by locus coeruleus stimulation and norepineph- rine in individual spinal motoneurons. Soc. Neurosci. Abstr. 15, 920.
Fuxe, K., Goldstein, M., H6kfelt, T. and Joh, T. H. (1970). Immunohistochemical localization of dopamine-B- hydroxylase in the peripheral and central nervous system. Res. Comm. Chem. Path. Pharm. 1(5), 627-636.
Glazer, E. Land Basbaum, A. I. (1981). Immunohisto- chemical localization of leucine-enkephalin in the spinal cord of the cat: Enkephalin-containing marginal neurons and pain modulation. J. Comp. Neurol. 196, 377-389.
Grimmelikhuijzen, C. J. P. (1983). Coexistence of neuro- peptides in hydra. Neuroscience 9(4), 837-845.
Hancock, M. B. and Fougerousse, C. L. (1976). Spinal projections from the nucleus locus coeruleus and nucleus subcoeruleus in the cat and monkey as demon- stratcd by the retrograde transport of horseradish peroxidase. Brain Res. Bull. 1,229 232.
E N K and T H in Cat Coerulospinal Neurons 9
Hayes, N. L. and Rustioni, A. (1981). Descending pro- jections from the brainstem and sensorimotor cortex to spinal enlargements in the cat. Exp. Brain Res. 41, 89-107.
Holets, V. R., H6kfelt, T., Rokaeus, A., Terenis, L. and Goldstein, M. (1988). Locus coeruleus neurons in the rat containing neuropeptide Y, tyrosine hydroxylase or galanin and their efferent projections to the spinal cord, cerebral cortex and hypothalamus. Neuroseience 24(3), 893-906.
Honda, C. N. and Lee, C. L. (1985). lmmunohisto- chemistry of synaptic input and functional character- izations of neuron near the central canal. Brain Res. 343, 120 128.
Hughes, J., Smith, T. W., Kosterlitz, H. W., Fothergill, L. A., Morgan, B. A. and Morris, H. R. (1975). Identifi- cation of two related pentapeptides from the brain with potent opiate agonist activity. Nature 258, 577 579.
Hunt, S. P. and Lovick, T. A. (1982). The distribution of serotonin, met-enkephalin and B-lipoproteinqike immunoreactivity in neuronal perikarya of the cat brainstem. Neurosci. Lett. 30, 139-145.
Johnson, R. P., Sar, M. and Stumpf, W. E. (19793. Relationships between the catecholaminergic and enkephalinergic systems demonstrated by a combined technique of formaldehyde-induced fluorescence and immunocytochemistry. Neurosci. Lett. 14, 321 326.
Jones, B. E. and Moore, R. Y. (1974). Catecholamine- containing neurons of the nucleus locus coeruleus in the cat. J. Comp. Neurol. 157, 43-52.
Jones, B. E. and Friedman, L. (1983). Atlas of catechol- amine perikarya, varicosities and pathways in the brainstem of the cat. J. Comp. Neurol. 215, 382-396.
Khachaturian, H., Lewis, M. E. and Watson. S. J. (19833. Enkephalin systems in diencephalon and brainstem of the rat. J. Comp. Neurol. 220, 310 320.
Kumakura, K., Guidotti, A. and Costa, E. (1979). Primary cultures of chromaffin cells: Molecular mechanisms for the induction of tyrosine hydroxylase mediated by 8-Br-cyclic AMP. Mol. Pharmacol. 16, 865 876.
Leger, L. and Hernandez-Nicaise, M. (1980). The cat locus coeruleus: Light and electron microscopic study of the neuronal somata. Anat. Embryol. (Berl.) 159, 181-198.
Leger, L., Charnay, Y., Chayvialle, J. A., Berod, A., Dray, F., Pujol, J. F., Jouvet, M. and Dubois, P. M. (1983). Localization of substance P- and enkephalin- like immunoreactivity in relation to catecholamine- containing cell bodies in the cat dorsolateral pontine tegmentum: An immunofluorescence study. Neuro- science 8, 525-546.
Livett, B. G., Dean, D. M., Whelan, L. G. Udenfriend, S. and Rossier, J. (1981). Co-release of enkephalin and catecholamines from cultured adrenal chromaffin cells. Nature 289, 317-319.
Maeda, T., Pin, C., Salvert, D., Ligier, M. and Jouvet, M. (1973). Les neurons contenant des catecholamines du tegmentum pontique et leurs voies de projection chez le chat. Brain Res. 57, 119 152.
Mizukawa, K. (1980). The segmental detailed topo- graphical distribution of monoaminergic terminals and their pathways in the spinal cord of the cat. Anat. Anz. 147, 125-144.
Mokha, S. S., McMillan, J. A. and Iggo, A. (1985). Descending control of spinal nociceptive transmission. Actions produced on spinal nociceptive neurons from

10 H. Zhuoe t al.
the nuclei locus coeruleus (LC) and raphe magnus (NRM). Exp. Brain Res. 58, 213-223.
Murakami, S., Okamura, H., Pelletier, G. and Ibata, Y. (1989). Differential colocalization of neuropeptide Y- and methionine-enkaphalin-Arg6-Gly7-Leu8-1ike immunoreactivity in catecholaminergic neurons in the rat brainstem. J. Comp. Neurol. 281,532-544.
Nakazato, T. (1987). Locus coeruleus neurons project- ing to the forebrain and the spinal cord in the cat. Neuroscience 23, 529-538.
Nygren, L. and Olson, L. (1977). A new major projection from locus coeruleus: The main source of noradrener- gic nerve terminals in the ventral and dorsal columns of the spinal cord. Brain Res. 132, 85-93.
Pilowsky, P., Minson, J., Hodgson, A., Howe, P., Chalmers, P. and Chalmers, J. (1986). Does substance P coexist with adrenaline in neurons of the rostral ventro- lateral medulla in the rat? Neurosci. Lett. 71,293-298.
Quach, T. T., Tang, F., Kageyama, H., Mocchetti, I., Guidotti, A., Meek, J. L., Costa, E. and Schwartz, J. P. (1984). Enkephalin biosynthesis in adrenal medulla, modulation of proenkephalin mRNA content of cul- tured chromaffin cells by 8-bromo-adenosine 3',5'- monophosphate. Mol. Pharmacol. 26, 255-260.
Randic, M. and Miletic, V. (1978). Depressant action of methionin-enkephalin and somatostatin in cat dorsal horn neurons activated by noxious stimuli. Brain Res. 152, 196-202.
Reddy, V. K., Fung, S. J., Zhuo, H. and Barnes, C. D. (1989). Spinally projecting noradrenergic neurons of the dorsolateral pontine tegmentum: A combined immunocytochemical and retrograde labelling study. Brain Res. 491,144-149.
Reddy, V. K., Fung, S. J., Zhuo, H. and Barnes, C. D. (1990). Localization of enkephalinergic neurons in the dorsolateral pontine tegmentum projecting to the spinal cord of the cat. J. Comp. Neurol. 29, 195-202.
Sar, M., Stumpf, W. E., Miller, R. J., Change, K. J. and Castrecasas, P. (1978). lmmunohistochemical localiz- ation of enkephalin in rat brain and spinal cord. J. Comp. Neurol. 182, 17-37.
Satoh, K., Tohyama, M., Yamamoto, K., Sakumoto, T. and Shimizu, N. (1977). Noradrenaline innervation of the spinal cord studied by the horseradish peroxidase method combined with monoamine oxidase staining. Exp. Brain Res. 30, 175-186.
Staines, W. A., Meister, B., Melander, T., Nagy, J. I. and Hrkfelt, T. (1988). Three-color immunofluorescence histochemistry allowing triple labeling within a single section. J. Histochem. Cytoehem. 36, 145-151.
Stevens, R. T., Hodge, C. J. and Apkarian, A. V. (1982). Krlliker-Fuse nucleus: The principal source of pontine catecholaminergic cells projecting to the lumbar spinal cord of cat. Brain Res. 239, 589-594.
Stevens, R., Apkarian, A. V. and Hodge, C. J. (1985). Funicular course of catecholamine fibers innervating the lumbar spinal cord of the cat. Brain Res. 336, 243-251.
Tashiro, T., Ruda, M. A., Satoda, T., Matsushima, R. and Mizuno, N. (1989). Convergence of serotonin-, enkephalin- and substance P-like immunoreactive afferent fibers onto cat medullary dorsal horn projec- tion neurons: A triple immunocytochemical staining technique combined with the retrograde HRP-tracing method. Brain Res. 491,360-365.
Werling, L. L., Brown, S. R. and Cox, B. M. (1987). Opioid receptor regulation of the release of norepi- nephrine in brain. Neuropharmacology 26, 987-996.
Wessendorf, M. R. and Elde, R. (1985). Characterization of immunofluorescence technique for the demon- stration of coexisting neurotransmitters within nerve fibers and terminals. J. Histochem. Cvtochem. 33, 984-994.
Westlund, K. N. and Coulter, J. D. (1980). Descending projections of the locus coeruleus and subcoeruleus/ medial parabrachial nuclei in monkey: Axon transport studies and dopamine-B-hydroxylase immunocyto- chemistry. Brain Res. Rev. 2, 235-264.
Westlund, K. N., Bowker, R. M., Ziegler, M. G. and Coulter, J. D. (1981). Origins of spinal noradrenergic pathways demonstrated by retrograde transport of antibody to dopamine-B-hydroxylase. Neurosei. Lett. 25, 243-249.
Westlund, K. N., Bowker, R. M., Ziegler, M. G. and Coulter, J. D. (1983). Noradrenergic projections to the spinal cord of the rat. Brain Res. 263, 15-31.
Wiklund, L., Leger, L. and Persson, M. (1981). Mono- amine cell distribution in the cat brainstem. A fluor- escence histochemical study with quantification of indoleaminergic and locus coeruleus cell groups. J. Comp. Neurol. 203, 617- 647.
Zamir, N., Palkovits, M., Brownstein, M. and Brownstein, M. (1985). Distribution of immunoreactive met- enkephalin-Arg6-Gly7-Leu8 and leu-enkephalin in dis- crete regions of the rat brain. Brain Res. 326, 1-8.
Zieglgansberger, W. and Tulloch, I. F. (1979). The effects of methionine- and leucine-enkephalin on spinal neurons of the cat. Brain Res. 167, 533-564.




















