
Identification of the Endogenous Ligands for Chicken Growth Hormone-Releasing Hormone (GHRH)...
12
Identification of the Endogenous Ligands for Chicken Growth Hormone-Releasing Hormone (GHRH) Receptor: Evidence for a Separate Gene Encoding GHRH in Submammalian Vertebrates Yajun Wang, Juan Li, Crystal Ying Wang, Amy Ho Yan Kwok, and Frederick C. Leung Department of Zoology, The University of Hong Kong, Hong Kong, China It is generally believed that hypothalamic GHRH activates GHRH receptor (GHRHR) to stimulate GH synthesis and re- lease in the pituitary of mammals. However, the identity of the endogenous ligand of GHRHR is still unresolved in submam- malian vertebrates including birds. In this study, we have successfully identified the chicken GHRH (cGHRH) gene, which consists of seven exons including two exons (exons 4 and 5) coding for the predicted mature GHRH peptide of 47 amino acids. Interestingly, the differential usage of splice do- nor sites at exon 6 results in the generation of two prepro- GHRHs (172 and 188 amino acids in length) with different C-terminal tails. Similar to mammals, cGHRH was detected to be predominantly expressed in the hypothalamus by RT-PCR assay. Using the pGL3-CRE-luciferase reporter system, we further demonstrated that both the synthetic cGHRH pep- tides (cGHRH 1– 47 and cGHRH 1–31 ) and conditioned medium from CHO cells expressing cGHRH could strongly induce lu- ciferase activity via activation of cGHRHR, indicating that cGHRH could bind cGHRHR and activate downstream cAMP- protein kinase A signaling pathway. Using the same system, cGHRH-like peptide was also shown to be capable of activat- ing cGHRHR in vitro. As in chicken, a conserved GHRH gene was identified in the genomes of lower vertebrate species including zebrafish, fugu, tetraodon, and Xenopus by synteny analysis. Collectively, our data suggest that GHRH, perhaps together with GHRH-like peptide (chicken/carp-like), may function as the authentic endogenous ligands of GHRHR in chicken as well as in other lower vertebrate species. (Endo- crinology 148: 2405–2416, 2007) P ITUITARY GH SYNTHESIS and release are regulated by two main hypothalamic peptides: GHRH and soma- tostatin in mammals (1–3). GHRH belongs to the glucagon superfamily, which also includes pituitary adenylate cyclase- activating polypeptide (PACAP), secretin, and vasoactive intestinal polypeptide (VIP) (4). Mammalian GHRH is a polypeptide of 42– 44 amino acid residues and encoded by a gene of five exons (5–9). Compelling evidence demonstrated that GHRH could strongly stimulate pituitary GH synthesis and release both in vivo and in vitro (6, 10, 11). The biological actions of GHRH are mediated by GHRH receptor (GHRHR), a member of the G protein-coupled receptor family, whose activation could increase intracellular cAMP levels and ac- tivate the protein kinase A (PKA) signaling pathway (12–15). Mutations in either GHRH gene or GHRHR gene would cause pituitary hypoplasia and serum GH deficiency in mouse and human (12, 16 –21), further emphasizing the piv- otal role of GHRH in controlling GH synthesis and secretion, as well as somatotroph proliferation in the pituitary of mammals. In contrast to studies reported in mammals, the phys- iological role of GHRH in pituitary GH release is contro- versial in lower vertebrates (22). In teleost fish, human GHRH or fish GHRH-like peptide has only modest or weak effects on GH release in cultured pituitary cells of several species including rainbow trout (23), tilapia (24), and sockeye salmon (25). In European eels (Anguilla an- guilla), one of the most primitive teleost fish, human GHRH is totally devoid of an effect on GH secretion in cultured pituitary cells (26). In contrast to the minimal response to GHRH, PACAP has been demonstrated to act as a GH-releasing factor in pituitaries of grass carp (27, 28), sockeye salmon (25), and European eel (26, 29). Similar stimulatory effects of PACAP have also been reported in amphibians (30). All the evidence points to the possibility that PACAP, instead of GHRH, may play a dominant role in GH release in the pituitary of lower vertebrates, especially in teleost fishes (22). Using a cDNA cloning strategy, the se- quences of GHRH-like peptide have been determined in a number of lower vertebrate species including zebrafish (31– 33), sockeye salmon (25), catfish (34), and frog (35, 36). Interestingly, GHRH-like peptide and PACAP are encoded by a single gene, whereas GHRH and PACAP are encoded by two distinct genes in mammals (4, 22). The distinct or- ganization of GHRH and PACAP between mammals and lower vertebrates seems to partially explain the difference in pituitary responsiveness to GHRH and PACAP. First Published Online February 1, 2007 Abbreviations: a.a., Amino acid(s); c, chicken; CHO, Chinese hamster ovary; CRE, cAMP response element; GHRH-L, GHRH-like peptide; GHRHR, GHRH receptor; hpGHRH, human pancreatic GHRH; l, long form; oPACAP, ovine pituitary adenylate cyclase-activating polypep- tide; ORF, open reading frame; PACAP, pituitary adenylate cyclase- activating polypeptide; PRP, PACAP-related peptide; PAC 1 -R, PACAP type I receptor; PKA, protein kinase A; s, short form; VIP, vasoactive intestinal polypeptide. Endocrinology is published monthly by The Endocrine Society (http:// www.endo-society.org), the foremost professional society serving the endocrine community. 0013-7227/07/$15.00/0 Endocrinology 148(5):2405–2416 Printed in U.S.A. Copyright © 2007 by The Endocrine Society doi: 10.1210/en.2006-1013 2405
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Identification of the Endogenous Ligands for Chicken Growth Hormone-Releasing Hormone (GHRH)...

Identification of the Endogenous Ligands for ChickenGrowth Hormone-Releasing Hormone (GHRH) Receptor:Evidence for a Separate Gene Encoding GHRH inSubmammalian Vertebrates
Yajun Wang, Juan Li, Crystal Ying Wang, Amy Ho Yan Kwok, and Frederick C. Leung
Department of Zoology, The University of Hong Kong, Hong Kong, China
It is generally believed that hypothalamic GHRH activatesGHRH receptor (GHRHR) to stimulate GH synthesis and re-lease in the pituitary of mammals. However, the identity of theendogenous ligand of GHRHR is still unresolved in submam-malian vertebrates including birds. In this study, we havesuccessfully identified the chicken GHRH (cGHRH) gene,which consists of seven exons including two exons (exons 4and 5) coding for the predicted mature GHRH peptide of 47amino acids. Interestingly, the differential usage of splice do-nor sites at exon 6 results in the generation of two prepro-GHRHs (172 and 188 amino acids in length) with differentC-terminal tails. Similar to mammals, cGHRH was detected tobe predominantly expressed in the hypothalamus by RT-PCRassay. Using the pGL3-CRE-luciferase reporter system, wefurther demonstrated that both the synthetic cGHRH pep-
tides (cGHRH1–47 and cGHRH1–31) and conditioned mediumfrom CHO cells expressing cGHRH could strongly induce lu-ciferase activity via activation of cGHRHR, indicating thatcGHRH could bind cGHRHR and activate downstream cAMP-protein kinase A signaling pathway. Using the same system,cGHRH-like peptide was also shown to be capable of activat-ing cGHRHR in vitro. As in chicken, a conserved GHRH genewas identified in the genomes of lower vertebrate speciesincluding zebrafish, fugu, tetraodon, and Xenopus by syntenyanalysis. Collectively, our data suggest that GHRH, perhapstogether with GHRH-like peptide (chicken/carp-like), mayfunction as the authentic endogenous ligands of GHRHR inchicken as well as in other lower vertebrate species. (Endo-crinology 148: 2405–2416, 2007)
PITUITARY GH SYNTHESIS and release are regulated bytwo main hypothalamic peptides: GHRH and soma-
tostatin in mammals (1–3). GHRH belongs to the glucagonsuperfamily, which also includes pituitary adenylate cyclase-activating polypeptide (PACAP), secretin, and vasoactiveintestinal polypeptide (VIP) (4). Mammalian GHRH is apolypeptide of 42–44 amino acid residues and encoded by agene of five exons (5–9). Compelling evidence demonstratedthat GHRH could strongly stimulate pituitary GH synthesisand release both in vivo and in vitro (6, 10, 11). The biologicalactions of GHRH are mediated by GHRH receptor (GHRHR),a member of the G protein-coupled receptor family, whoseactivation could increase intracellular cAMP levels and ac-tivate the protein kinase A (PKA) signaling pathway (12–15).Mutations in either GHRH gene or GHRHR gene wouldcause pituitary hypoplasia and serum GH deficiency inmouse and human (12, 16–21), further emphasizing the piv-otal role of GHRH in controlling GH synthesis and secretion,
as well as somatotroph proliferation in the pituitary ofmammals.
In contrast to studies reported in mammals, the phys-iological role of GHRH in pituitary GH release is contro-versial in lower vertebrates (22). In teleost fish, humanGHRH or fish GHRH-like peptide has only modest orweak effects on GH release in cultured pituitary cells ofseveral species including rainbow trout (23), tilapia (24),and sockeye salmon (25). In European eels (Anguilla an-guilla), one of the most primitive teleost fish, humanGHRH is totally devoid of an effect on GH secretion incultured pituitary cells (26). In contrast to the minimalresponse to GHRH, PACAP has been demonstrated to actas a GH-releasing factor in pituitaries of grass carp (27, 28),sockeye salmon (25), and European eel (26, 29). Similarstimulatory effects of PACAP have also been reported inamphibians (30). All the evidence points to the possibilitythat PACAP, instead of GHRH, may play a dominant role inGH release in the pituitary of lower vertebrates, especially inteleost fishes (22). Using a cDNA cloning strategy, the se-quences of GHRH-like peptide have been determined in anumber of lower vertebrate species including zebrafish (31–33), sockeye salmon (25), catfish (34), and frog (35, 36).Interestingly, GHRH-like peptide and PACAP are encodedby a single gene, whereas GHRH and PACAP are encodedby two distinct genes in mammals (4, 22). The distinct or-ganization of GHRH and PACAP between mammals andlower vertebrates seems to partially explain the difference inpituitary responsiveness to GHRH and PACAP.
First Published Online February 1, 2007Abbreviations: a.a., Amino acid(s); c, chicken; CHO, Chinese hamster
ovary; CRE, cAMP response element; GHRH-L, GHRH-like peptide;GHRHR, GHRH receptor; hpGHRH, human pancreatic GHRH; l, longform; oPACAP, ovine pituitary adenylate cyclase-activating polypep-tide; ORF, open reading frame; PACAP, pituitary adenylate cyclase-activating polypeptide; PRP, PACAP-related peptide; PAC1-R, PACAPtype I receptor; PKA, protein kinase A; s, short form; VIP, vasoactiveintestinal polypeptide.Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
0013-7227/07/$15.00/0 Endocrinology 148(5):2405–2416Printed in U.S.A. Copyright © 2007 by The Endocrine Society
doi: 10.1210/en.2006-1013
2405

Although the same precursor encompasses GHRH-likepeptide and PACAP in chickens as reported in teleosts andamphibians (4, 37), we and others have clearly demonstratedthat human GHRH, and not PACAP, play a dominant rolein chicken GH release both in vivo and in vitro (38–46).Despite the fact that PACAP can increase intracellular cAMPlevel and GH release in cultured chicken pituitary cells, itseffect on GH release is much less potent than human GHRH(47). Surprisingly, iv administration of synthetic chicken (c)GHRH-like peptide has little effect on GH release even at ahigh dosage (44). Consistent with this finding, synthesizedcGHRH-like peptide also has little potency at cGHRHR ac-tivation (48). In view of these findings, there arises severalfundamental issues that need to be addressed: 1) whethercGHRH-like peptide represents the authentic ligand of cGH-RHR; 2) whether a new GHRH with high potency in stim-ulating pituitary GH secretion exists in the chicken and otherlower vertebrate species; and 3) how GHRH gene andGHRH-PACAP gene have evolved in the course of vertebrateevolution. Answering these questions would help us to re-evaluate the physiological roles of GHRH in the hypotha-lamic-pituitary axis of lower vertebrates.
In the present study, we isolated the full-length cDNAscoding for chicken GHRH. Using a luciferase reporter sys-tem, we further demonstrated that cGHRH could activatecGHRHR with high potency (EC50 � 0.1 nm). Together withthe information collected from genome database of modelvertebrate animals (http://www.ensembl.org/), our studiesfor the first time indicate that a conserved GHRH gene existsin submammalian vertebrates including birds, amphibians,and teleosts, and most likely, the identified GHRH is one ofthe endogenous ligands of GHRHR in submammalian ver-tebrate species.
Materials and MethodsChemicals and hormones
All chemicals were obtained from Sigma-Aldrich (St. Louis, MO), andrestriction enzymes were obtained from Amersham Biosciences (GEHealthcare Bio-Sciences Corp., Piscataway, NJ) unless stated otherwise.Human pancreatic GHRH (hpGHRH1–40) and ovine pituitary adenylatecyclase-activating polypeptide (oPACAP38) were purchased fromBachem (Torrance, CA). cGHRH1–31 and cGHRH1–47 with free carboxyltermini were synthesized using solid-phase Fmoc chemistry (GL Bio-chem, Shanghai, China). The purity of synthesized cGHRHs is greaterthan 95% (analyzed by HPLC), and their structure was verified by massspectrometry (GL Biochem). cGHRH1–47, cGHRH1–31, hpGHRH1–40, andoPACAP38 were first dissolved in distilled water and then diluted to thedesired concentrations with medium before use.
Animals
Adult chickens used in all experiments are of a local chicken strain(Shek-ki) of Hong Kong and was kindly provided by Kadoorie Agri-cultural Research Centre (Hong Kong). All experiments were performedunder license from the Government of the Hong Kong Special Admin-istrative Region and endorsed by the Animal Experimentation EthicsCommittee of The University of Hong Kong.
Total RNA isolation
Adult chickens were decapitated and different tissues including smallintestine, kidney, liver, lung, muscle, ovary, pituitary, spleen, testis, andbrain were collected for total RNA extraction. Total RNA was extractedfrom chicken tissues with Tri-reagent (Molecular Research Center, Cin-
cinnati, OH) according to the manufacturer’s instructions and dissolvedin diethylpyrocarbonate-treated H2O.
Cloning of the full-length cDNAs for GHRH fromchicken brain
Two gene-specific primers were designed based on an unannotatedcDNA sequence deposited in GenBank (accession no. BX929984,ChEST294a6) (Table 1). Two micrograms of total RNA from adultchicken brain were reverse transcribed into single-strand cDNA withMoloney murine leukemia virus reverse transcriptase (Promega Corp.,Madison, WI) followed by PCR amplification using high-fidelity TaqDNA polymerase (Roche Diagnostics, Basel, Switzerland). The PCRamplification (35 cycles) was performed using a cycle profile of 30 secat 95 C, 30 sec at 56 C, and 60 sec at 72 C, followed by a 15-min extensionat 72 C. The PCR products were cloned into pBluescript SK (�/�)(Stratagene, La Jolla, CA). The cloned GHRH cDNAs were sequencedwith the Bigdye terminator cycle sequencing, version 3.0, ready reactionkit (PerkinElmer, Foster City, CA) and analyzed on Prism 3100 geneticanalyzer (PerkinElmer). Each form (short and long form) of GHRHfull-length cDNAs was finally determined by sequencing three inde-pendent clones.
RT-PCR
Reverse transcription was performed at 42 C for 2 h in a total volumeof 10 �l consisting of 2 �g total RNA from different tissues, 1� single-strand buffer, 0.5 mm each deoxynucleotide triphosphate, 0.5 �g oli-godeoxythymide, and 100 U Moloney murine leukemia virus reversetranscriptase (Promega). All negative controls were carried out underthe same condition without reverse transcriptase added in the 10 �l ofreaction mix.
PCR was carried out in a total volume of 20 �l consisting of 1� PCRbuffer, 0.2 mm each deoxynucleotide triphosphate, 2.0 mm MgCl2, 0.2�m each primer, and 0.5 U of Taq DNA polymerase (Invitrogen, Carls-bad, CA) on the PTC-225 Peltier thermal cycler (MJ Research Inc.,Waltham, MA). To evaluate the relative mRNA levels of target genes, theoptimal cycle numbers were first determined according to our previ-ously established semiquantitative RT-PCR methods (49–52). For �-ac-tin gene, 23 cycles of 30 sec at 95 C, 30 sec at 58 C, and 60 sec at 72 Cwere used followed by 5 min extension at 72 C. For GHRH gene, 31cycles of 30 sec at 95 C, 30 sec at 58 C, and 60 sec at 72 C were usedfollowed by 5 min extension at 72 C. For GHRH-PACAP gene, 33 cycleswere used based on our previous report (53). The primers used forGHRH gene, GHRH-PACAP gene, and �-actin gene were listed in Table1. The PCR products were visualized on a UV-transilluminator (Bio-RadLaboratories, Inc., Hercules, CA) after electrophoresis on 2% agarose gelcontaining ethidium bromide.
Establishment of a system to monitor the biologicalactivities of cGHRH, cGHRH-like peptide, and cPACAP
Experiment 1: cloning of cGHRHR and chicken PACAP type I receptor(PAC1-R) into pcDNA3.1(�) expression vector. Based on the cDNA se-quences of cGHRHR (53) and PACAP type I receptor [cPAC1-R, shortform, 471 amino acids (a.a.)] (54), two pairs of gene-specific primersflanked by restriction enzyme recognition sites were designed to amplifythe complete open reading frame (ORF) regions of chicken GHRHR andPAC1-R with high-fidelity Taq DNA polymerase (Roche Diagnostics)(Table 1). The amplified PCR products were first cloned into pBluescriptSK (�/�) vector (Stratagene) and subjected to sequencing analysis(PerkinElmer). Then the inserts of interest were released by restrictionenzyme digestions and subcloned into pcDNA3.1(�) expression vector(Invitrogen).
Experiment 2: cell culture and cotransfection of cGHRHR (or cPAC1-R) ex-pression plasmid and pGL3-CRE-luciferase reporter construct into Chinesehamster ovary (CHO) cells. In this experiment, pGL3-CRE-luciferase re-porter construct, in which the expression of luciferase gene is driven bya promoter containing multiple cAMP-response elements (CRE), wasfirst constructed by cloning the promoter region of pCRE-SEAP vector(CLONTECH, Palo Alto, CA) into the promoterless pGL3-Basic vector(Promega). CHO cells were cultured in DMEM supplemented with 10%
2406 Endocrinology, May 2007, 148(5):2405–2416 Wang et al. • Identification of Chicken GHRHR Ligands

(vol/vol) fetal bovine serum (HyClone, Logan, UT), 100 U/ml penicillinG, and 100 �g/ml streptomycin (Life Technologies, Inc., Grand Island,NY) in a 90-cm culture dish (NUNC, Rochester, NY) and incubated at37 C with 5% CO2. Then CHO cells were plated in a 6-well plate at adensity of 3 � 105 cells/well 1 d before transfection. A mixture con-taining 700 ng of pGL3-CRE-luciferase reporter construct, 200 ng ofpcDNA3.1 construct containing either cGHRHR or cPAC1-R (or emptyvector), and 6 �l of DOSPER liposomal transfection reagent (RocheDiagnostics) was prepared in 50 �l of PBS solution and transfection wasperformed according to the manufacturer’s instructions when cells were70% confluent. After 24 h of culture, CHO cells were trypsinized andcultured in a 96-well plate at a density of 2 � 104 cells/well at 37 C for24 h before hormone treatment.
Experiment 3: examination of hormone specificity of hpGHRH1–40 andoPACAP38 on the activation of cGHRHR and cPAC1-R. The hpGHRH1–40and oPACAP38 were diluted by serum-free DMEM to the desired con-centrations, respectively. After removal of the DMEM from 96-wellplate, 50 or 60 �l of hormone-containing medium or hormone-free me-dium (used as control) were added and CHO cells were incubated foran additional 6 h at 37 C with 5% CO2 before being harvested forluciferase assay. After removal of the culture medium, CHO cells werelysed by adding 50 �l of 1� passive lysis buffer (Promega) per well, and
the luciferase activity of 15 �l of cellular lysates was determined usingluciferase assay reagent (Promega).
Transient expression of cGHRH(s), cGHRH(l), cGHRH(d),and cGHRH(c) in CHO cells and conditionedmedium collection
Five gene-specific primers flanked by restriction sites (KpnI or EcoRI)at their 5�-ends were designed to prepare four expression plasmids,named cGHRH(s) (encoding the short cGHRH precursor, 172 a.a.), cGH-RH(l) (encoding the long cGHRH precursor, 188 a.a.), cGHRH(d) (con-taining the signal peptide, cryptic peptide and putative GHRH peptide,121 a.a.), and cGHRH(c) (including signal peptide and cryptic peptideonly, 72 a.a.), respectively (see Table 1 and Fig. 6A). The PCR was firstperformed with high fidelity Taq DNA polymerase (Roche) by usingcloned GHRH plasmids as templates. Then the PCR products werecloned into pBluescript SK(�/�) vector (Stratagene) and sequenced byABI3100 genetic analyzer (PerkinElmer). The inserts with correct DNAsequences were released by KpnI and EcoRI restriction enzyme diges-tions and ligated into pcDNA3.1(�) expression vector (Invitrogen). Thetransfection procedure is slightly different from the above experiments.In brief, five mixtures containing 3 �g of different pcDNA3.1(�) con-
TABLE 1. Primers useda
Gene/construct Primer sequence (5�–3�) Size (bp)
Primers for amplifying full-length cDNAsghrh
Sense TACCTGAGTGGGAGCTGATC 704/667Antisense CATCAGTCTCCAGCTGGTCA
Primers for RT-PCR assayghrh
Sense TACCTGAGTGGGAGCTGATC 286Antisense CTGCATCCTTTTCTCAGTGG
ghrhSense CAGGATGTGAACGGCAGT 180/143Antisense CATCAGTCTCCAGCTGGTCA
ghrh-pacapSense GAGGTGTACGCGCTGTACTA 326Antisense GCTCATCGCTACAAATACGCTA
�-ActinSense CAATGGCTCCGGTATGTGCA 401Antisense AGGCATACAGGGACAGCACA
Primers for constructing the expression plasmidsb
cGHRH(s)Sense GGTACCACCATGCTGGATAAGGCCA 532Antisensec GAATTCACATGAACTTAGCCAG
cGHRH(l)Antisensec GAATTCATTGCTGTGAATGGC 580
cGHRH(d)Antisensec GAATTCATGTCAGGAGCTTGTGCA 379
cGHRH(c)Antisensec GAATTCTACATCCTTTTCTCAGTGG 233
cGHRH-PACAP(w)Sense GGTACCACCATGTGTAGCAAAGCGCT 538Antisensed GAATTCATCGCTACAAATACGC
cGHRH-PACAP(d)Antisensed GAATTCTAGCTGAGCGGTTCCGCCT 392
cGHRH-PACAP(c)Antisensed GAATTCTATCCCGGCGGGTAGTACA 248
cPAC1-RSense GGGTACCGCCGCCACCATGGCCATCTGTTGGAAG 1438Antisense GGAATTCTATGTCGCTAGATTCTCCG
cGHRHRSense GGATCCTAGCATGTCATACCA 1280Antisense TCTAGAGTCTTTAGCACACAGA
a All primers were synthesized by Invitrogen (Hong Kong).b The restriction sites added are underlined.c The same sense primer was used for constructing other GHRH expression plasmids.d The same sense primer was used for constructing other GHRH-PACAP expression plasmids.
Wang et al. • Identification of Chicken GHRHR Ligands Endocrinology, May 2007, 148(5):2405–2416 2407

structs and 6 �l of DOSPER liposomal transfection reagent (Roche) wereprepared in 50 �l of PBS solution, and transfection was carried outaccording to the manufacturer’s instruction when CHO cells were 70%confluent in a 6-well plate. Six hours after transfection, the medium wasreplaced by fresh medium supplemented with 10% fetal bovine serumand CHO cells were incubated at 37 C for an additional 18 h. Then themedium was replaced by 1 ml of serum-free DMEM. The serum-freeconditioned medium was collected after 36 h of culture at 37 C with 5%CO2. To avoid any contamination from cells or cell debris, the condi-tioned medium was centrifuged at 3000 rpm for 3 min and the super-natant was aliquoted and stored at �80 C before measurement of bio-logical activity.
Transient expression of cGHRH-PACAP(w), cGHRH-PACAP(d), and cGHRH-PACAP(c) in chicken DF-1 cellsand conditioned medium collection
Using the same approach as mentioned above, three sets of gene-specific primers were designed to prepare three pcDNA3.1 expressionplasmids: named cGHRH-PACAP(w) (encoding the GHRH-PACAPprecursor, 172 a.a.), cGHRH-PACAP(d) (containing signal peptide,cryptic peptide and GHRH-like peptide only, 125 a.a.), and cGHRH-PACAP(c) (containing signal peptide and cryptic peptide only, 77 a.a.)respectively (see Table 1 and Fig. 8A). An immortalized chicken embryofibroblast cell line DF-1 (cultured in high glucose DMEM) was chosenfor transient expression of various cGHRH-PACAP constructs becausethe conditioned medium collected from CHO cells had low activity(perhaps due to the low expression level of recombinant peptides inCHO cells) on the activation of cGHRHR (or cPAC1-R). Transfection andconditioned medium collection were performed in the same way men-tioned above.
Data analysis
The luciferase activities in each treatment group were expressed asrelative fold increase as compared with the control group (withouthormone treatment). The data were analyzed by one-way ANOVAfollowed by Dunnett’s test using GraphPad Prism 4 (GraphPad Soft-ware, San Diego, CA). To validate our results, all experiments wererepeated at least three times.
ResultsCloning of the full-length cDNAs encoding cGHRH
Based on an unannotated cDNA sequence deposited inGenBank (accession no. BX929984), we successfully clonedthe full-length cDNA coding for GHRH from adult chickenbrain. It is 704 bp in length and encodes a GHRH precursorof 172 amino acids (GenBank accession no. DQ465018). Com-parison of GHRH cDNA sequence with the chicken genomedatabase (http://www.ensembl.org/Gallus_gallus) re-vealed that cGHRH gene spans more than 6 kb on chromo-some 20 and consists of 7 exons (Fig. 1). The predicted matureGHRH peptide is 47 amino acid residues in length (encodedby exons 4 and 5) and shares high amino acid sequenceidentity with that of human (50%), rat (41%), and zebrafish(69%) (accession no. DQ832172) (Fig. 2A). Within GHRH1–47region, the N terminus (cGHRH1–27) is of the highest se-quence identity, whereas the carboxyl termini diversify sig-nificantly among species (Fig. 2A). Unlike GHRH precursorsof human, rat, and zebrafish, cGHRH precursor has a uniquelong carboxyl terminal flanking peptide (49 a.a.) and a crypticpeptide encoded by exon 3, which is absent in human, rat,and zebrafish GHRH precursors (Fig. 2A).
Although the GHRH gene may have evolved from a du-plicated GHRH-PACAP gene, the putative cGHRH1–47 isonly 46% identical with cGHRH-like peptide with the highest
sequence identity (70%) noted in the N-terminal 23 aminoacid residues (37, 53), and little or no homology was foundin the remaining parts of the two precursors (Fig. 2B).
Interestingly, a minor PCR band (667 bp) with faint in-tensity was also observed when RT-PCR assay was per-formed (Fig. 1A). Sequencing analysis revealed that it isresulted from the alternative usage of a splice donor sitelocated at 3� end of exon 6, which leads to a deletion of 37 bpincluding the stop codon on exon 6 (Fig. 1B). Thus, this shortcDNA fragment using a new stop codon on exon 7 encodesa larger prepro-GHRH (188 a.a.) with an altered carboxylterminal tail (GenBank accession no. DQ465017) (Fig. 2A).Consistent with our finding, the unannotated cDNA se-quence (BX929984) also encodes an identical precursor of 188amino acid residues although sequence variation was notedin the 5� untranslated region, confirming the presence ofalternative mRNA splicing in cGHRH gene. In this study, theshort and long GHRH precursors are designated as cGHRH-sand cGHRH-l, respectively.
Expression of GHRH gene and GHRH-PACAP gene in adultchicken brain
To demonstrate the hypothalamic origin of this novelcGHRH, spatial distribution of GHRH mRNA in adultchicken brain was examined by RT-PCR assay. As shown inFig. 3, GHRH mRNA was detected to be predominantlyexpressed in the hypothalamus and weakly expressed in themidbrain and hindbrain, clearly indicating that this polypep-tide is produced mainly by the hypothalamus (Fig. 3). More-over, when specific primers detecting both the long and shortGHRH precursor mRNAs were used, similar expression pat-tern was noted (Table 1 and Fig. 3). In contrast, cGHRH-PACAP mRNA is widely expressed in all regions of adult
FIG. 1. A, Amplification of cGHRH full-length cDNAs from brain byRT-PCR. �, RT without reverse transcriptase; �, RT with reversetranscriptase. B, Genomic structure of cGHRH gene. Seven exons arelabeled with e1, e2, e3, e4, e5, e6, and e7, respectively. The short andlong prepro-GHRHs are designated as cGHRH-s and cGHRH-l, re-spectively. SP, Signal peptide; CP, cryptic peptide; C-peptide, car-boxyl terminal flanking peptide. Arrow (B) indicates the location of analternative splice donor site at exon 6 (e6).
2408 Endocrinology, May 2007, 148(5):2405–2416 Wang et al. • Identification of Chicken GHRHR Ligands

chicken brain investigated, including telencephalon, cerebel-lum, midbrain, hindbrain, and hypothalamus (Fig. 3).
Tissue distribution of GHRH mRNA expression inchicken tissues
To examine the extrahypothalamic sources of cGHRH, wealso investigated the expression of cGHRH in other nineadult chicken tissues including small intestine, kidney, liver,lung, muscle, ovary, pituitary, spleen, and testis using RT-PCR assay. The expression of cGHRH was low but detectablein small intestine, kidney, ovary, pituitary, and spleen usinglower PCR cycle (31 cycles) (Fig. 4). However, using higherPCR cycle number (35 cycles), the expression of cGHRH,predominantly the cGHRH-s, was detected in all tissues ex-cept muscle (Fig. 4). Considering the low or no expression ofcGHRHR in extrapituitary tissues (48, 53, 55), the physio-
logical relevance of extrahypothalamic GHRH remains to bedetermined.
Validating the system for monitoring the biologicalactivities of cGHRH, cGHRH-like peptide, and cPACAP
To test the biological activities of cGHRH, cGHRH-likepeptide, and cPACAP from conditioned medium, a systemmonitoring hormone specificity on cGHRHR activation wasfirst established. In view of the fact that the GHRH gene islikely to be originated from a duplication of GHRH-PACAPgene (4, 22, 56), cPAC1-R was also introduced in the presentstudy to monitor whether peptides released from the shortor long cGHRH precursor could activate cPAC1-R. As shownin Fig. 5, human GHRH (10�12 to 10�6 m) could induceluciferase activities in a dose-dependent manner via activa-tion of cGHRHR (EC50 0.106 nm) but not cPAC1-R (Fig. 5A),whereas oPACAP38 could increase basal luciferase activities
FIG. 2. A, Comparison of GHRH precur-sors of chicken (cGHRH-s and cGHRH-l)with that of zebrafish (zfGHRH:DQ832172), rat (rGHRH: M73486), andhuman (hGHRH: X00094). The putativecryptic peptide is underlined and the de-duced or identified GHRH peptides areshaded. A putative amidation site (G28) isboxed. B, Comparison of the deducedamino acid sequence of cGHRH precursorwith that of cGHRH-PACAP precursor(AY956323). The predicted GHRH andC-peptide are boxed. GHRH-like peptideand PACAP38 are boxed and shaded. Dots(A and B) indicate the amino acid resi-dues identical with those of chicken pre-pro-GHRH (cGHRH-s). C-peptide, Car-boxyl terminal flanking peptide.
FIG. 3. Expression of GHRH gene and GHRH-PACAP gene in adultchicken brain regions including telencephalon (Te), cerebellum (Cb),midbrain (Mb), hindbrain (Hb), and hypothalamus (Hp). Gel picturesA and B show the specific amplification of N-terminal region andvariable C-terminal regions of GHRH precursors, respectively. Onlyvery faint minor band corresponding to cGHRH-l was observed in gelpicture B. Numbers in brackets indicate the PCR cycles used (allnegative controls are not shown).
FIG. 4. Expression of GHRH gene in chicken extrahypothalamic tis-sues including small intestine (In), kidney (Ki), liver (Li), lung (Lu),muscle (Mu), ovary (Ov), pituitary (Pi), spleen (Sp), and testis (Te).Gel images A and B indicate the specific amplification of cGHRH Nterminus, and gel image C shows the specific amplification of variablecGHRH C-termini. Numbers in brackets indicate the PCR cycle num-bers used (all negative controls are not shown).
Wang et al. • Identification of Chicken GHRHR Ligands Endocrinology, May 2007, 148(5):2405–2416 2409

via cPAC1-R, even at a low dosage (EC50 0.867 nm) (Fig. 5B).Interestingly, PACAP could also activate cGHRHR at higherdosages (�10�8 m, EC50 37.7 nm) (Fig. 5B), coincided with thereport by Toogood et al. (48). Most important of all, neitherhpGHRH1–40 nor oPACAP38 could increase basal luciferaseactivity in cultured CHO cells without transfected cGHRHRand cPAC1-R expression plasmids, indicating the bindingspecificity of hormone to receptor(s) (Fig. 5). Thus, this sys-tem was used to evaluate the biological activities of cGHRH,cGHRH-like peptide, and cPACAP from conditionedmedium.
Activation of cGHRHR by conditioned medium collectedfrom CHO cells with transient expression of cGHRH(s),cGHRH(l), and cGHRH(d) constructs
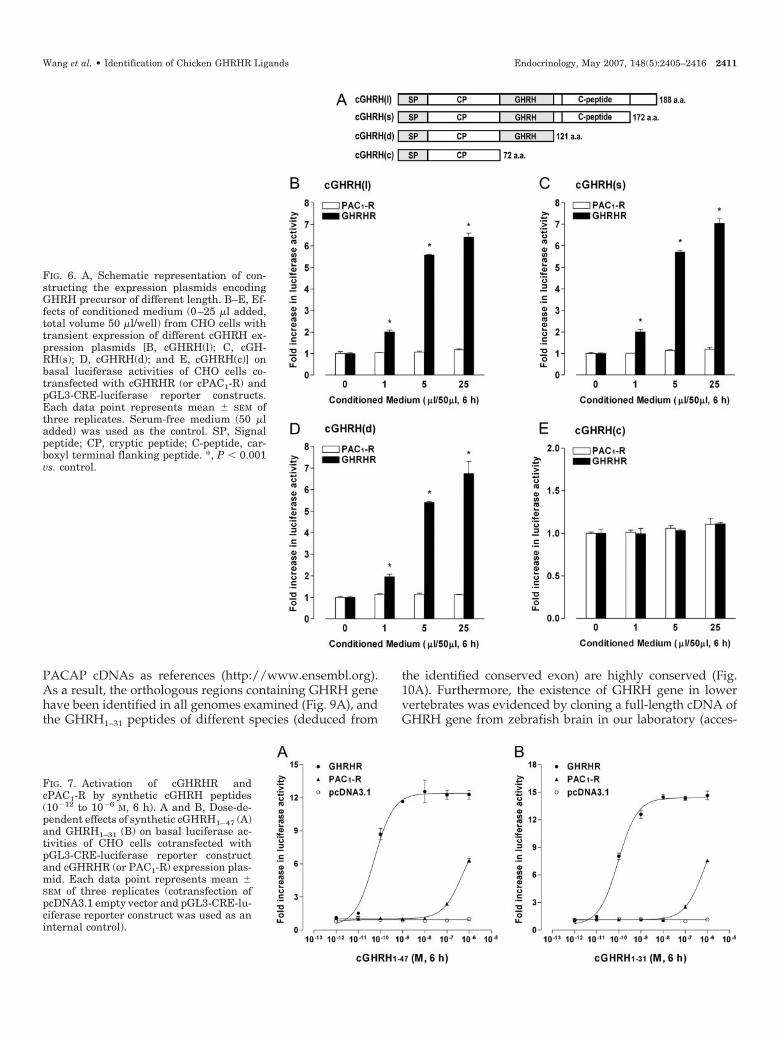
To examine whether cGHRH could activate cGHRHR spe-cifically, transient expression of four cGHRH constructs[cGHRH(s), cGHRH(l), cGHRH(d), and cGHRH(c)] in CHOcells was performed in this study (Fig. 6). As expected, onlythe conditioned medium collected from CHO cells with tran-sient expression of constructs containing cGHRH region[cGHRH(s), cGHRH(l), and cGHRH(d)] could increase lu-ciferase activity in a dose-dependent manner via activationof cGHRHR (not cPAC1-R) (Fig. 6, B–D). In contrast, cGH-RHR could not be activated by conditioned medium col-lected from CHO cells transfected with construct encodingonly the cryptic peptide [cGHRH(c)] under the same condi-tion (Fig. 6E). Similar results were obtained by using con-ditioned medium from cultured DF-1 cells (data not shown).These results suggest that cGHRH is a potential ligand ofcGHRHR.
Activation of cGHRHR by synthetic cGHRH1–31 andcGHRH1–47 peptides
To further evaluate the potency of cGHRH at cGHRHRactivation, two cGHRH peptides of different length weresynthesized. One is the predicted long cGHRH1–47 peptide.The other is a short cGHRH1–31 peptide containing the N-terminal 31 amino acid residues conserved between chickenand zebrafish (Fig. 2A). As shown in Fig. 7, both syntheticcGHRH1–47 and cGHRH1–31 could increase luciferase activityvia activation of cGHRHR with high potency (cGHRH1–47,EC50 0.053 nm; cGHRH1–31, EC50 0.087 nm) (Fig. 7). This
finding strongly suggests that cGHRH is a potent ligand ofcGHRHR.
In contrast to hpGHRH1–40, cGHRH peptide, either thelong or short form, could activate cPAC1-R at higher dosages(�10�7 m, 1000-fold less potent than oPACAP38) (Figs. 5 and7). This experiment was repeated more than three times, andsimilar patterns were noted, suggesting the low-affinitybinding of cGHRH to cPAC1-R and a close evolutionaryrelationship between GHRH and PACAP (4, 22).
Activation of cGHRHR by cGHRH-like peptide
It has been reported that the synthesized cGHRH-like pep-tide (or carp GHRH-like peptide) shows little detectablebinding to either cGHRHR or hGHRHR in vitro (48), sug-gesting that cGHRH-like peptide may not function as a li-gand of cGHRHR. To test this possibility, three GHRH-PACAP expression plasmids were used to transfect chickenDF-1 cells and conditioned medium was collected. As shownin Fig. 8, the conditioned medium from transient expressionof the construct cGHRH-PACAP(d) encoding GHRH-likepeptide could increase luciferase activity in a dose-depen-dent manner via specific activation of cGHRHR (Fig. 8C). Incontrast, the conditioned medium from transient expressionof construct cGHRH-PACAP(c) encoding the cryptic peptidehad no effect on cGHRHR activation (Fig. 8D), also support-ing the specific action of cGHRH-like peptide on cGHRHRactivation. Interestingly, the conditioned medium from tran-sient expression of the construct encoding complete ORFregion of GHRH-PACAP seemed to induce luciferase activ-ity via activation of cPAC1-R only (Fig. 8B). We repeatedthese experiments five times, and similar results wereobtained.
Evolutionary evidence for a separate gene encoding GHRHin submammalian vertebrate species
Two separate genes were shown to encode GHRH andGHRH-PACAP in chickens (37), leading us to speculate thatchicken GHRH gene and GHRH-PACAP gene may be or-thologous to mammalian GHRH gene and PACAP gene,respectively. To test this hypothesis, we blasted genome da-tabase of human (Homo sapiens), Xenopus (Xenopus tropicalis),zebrafish (Danio rerio), tetraodon (Tetraodon nigroviridis), andfugu (Takifugu rubripes) using chicken GHRH and GHRH-
FIG. 5. Hormone specificity of hpGHRH(10�12 to 10�6 M) (A) and oPACAP38 (10�12
to 10�7 M) (B) on the activation of cGHRHRand PACAP type I receptor (PAC1-R) mon-itored by a system of cotransfection ofpGL3-CRE-luciferase reporter constructand GHRHR (or PAC1-R) expression plas-mid in CHO cells. Each data point repre-sents mean � SEM of three replicates (co-transfection of pcDNA3.1 empty vector andpGL3-CRE-luciferase reporter constructwas used as an internal control).
2410 Endocrinology, May 2007, 148(5):2405–2416 Wang et al. • Identification of Chicken GHRHR Ligands
PACAP cDNAs as references (http://www.ensembl.org).As a result, the orthologous regions containing GHRH genehave been identified in all genomes examined (Fig. 9A), andthe GHRH1–31 peptides of different species (deduced from
the identified conserved exon) are highly conserved (Fig.10A). Furthermore, the existence of GHRH gene in lowervertebrates was evidenced by cloning a full-length cDNA ofGHRH gene from zebrafish brain in our laboratory (acces-
FIG. 6. A, Schematic representation of con-structing the expression plasmids encodingGHRH precursor of different length. B–E, Ef-fects of conditioned medium (0–25 �l added,total volume 50 �l/well) from CHO cells withtransient expression of different cGHRH ex-pression plasmids [B, cGHRH(l); C, cGH-RH(s); D, cGHRH(d); and E, cGHRH(c)] onbasal luciferase activities of CHO cells co-transfected with cGHRHR (or cPAC1-R) andpGL3-CRE-luciferase reporter constructs.Each data point represents mean � SEM ofthree replicates. Serum-free medium (50 �ladded) was used as the control. SP, Signalpeptide; CP, cryptic peptide; C-peptide, car-boxyl terminal flanking peptide. *, P � 0.001vs. control.
FIG. 7. Activation of cGHRHR andcPAC1-R by synthetic cGHRH peptides(10�12 to 10�6 M, 6 h). A and B, Dose-de-pendent effects of synthetic cGHRH1–47 (A)and GHRH1–31 (B) on basal luciferase ac-tivities of CHO cells cotransfected withpGL3-CRE-luciferase reporter constructand cGHRHR (or PAC1-R) expression plas-mid. Each data point represents mean �SEM of three replicates (cotransfection ofpcDNA3.1 empty vector and pGL3-CRE-lu-ciferase reporter construct was used as aninternal control).
Wang et al. • Identification of Chicken GHRHR Ligands Endocrinology, May 2007, 148(5):2405–2416 2411

sion no. DQ832172; 1017 bp, 112 a.a.) (Fig. 2A), confirmingthe expression of GHRH gene in lower vertebrate species(Figs. 2A and 9A). In contrast, cGHRH-PACAP gene andmammalian PACAP gene are localized on another conservedsynteny, which could also be found in teleosts and amphib-ians, indicating that they are of the same origin (Figs. 9B and11).
Discussion
It is well documented that GHRH plays a critical role instimulating pituitary GH synthesis and release both in vitroand in vivo in mammals (1). However, whether a GHRH geneexists in submammalian vertebrate species is still an openquestion (4, 22). In the present study, two chicken prepro-GHRH precursors with distinct C-terminal tails have beenidentified and both of them display biological activities oncGHRHR activation. Our studies also provide direct evi-dence that a functional GHRH is expressed predominantly inthe chicken hypothalamus.
cGHRH peptide shares high homology with mammalianGHRHs, especially in the bioactive core region (GHRH1–29)(Fig. 2A). However, the chicken GHRH gene differs from itsmammalian counterparts. In chickens, the GHRH gene con-
sists of seven exons, whereas five exons have been noted inmammals (5, 7), and the predicted chicken prepro-GHRHsare much longer (172 and 188 a.a.) than mammalian prepro-GHRHs (103–108 a.a.). Unlike that in mammals, a crypticpeptide is still present in chicken GHRH precursors (Fig. 2A)(4). In addition, no homology could be noted in the carboxylterminal flanking peptides between human, zebrafish, andchicken, suggesting that C-terminal flanking peptides maynot have a similar physiological role in chickens as proposedin mammals (57–60). In this study, two cGHRH mRNA spe-cies have been identified. The multiple splice variants coex-pressed in hypothalamus and extrahypothalamic tissueshave also been reported in mammals (5, 9, 61). However, thesplicing sites are not conserved between chicken and mam-mals. All the difference indicates that mammalian GHRHgene must have undergone an accelerated evolutionary pro-cess after its divergence from a common ancestor (Figs. 2 and11). Meanwhile, the obvious difference in GHRH gene be-tween chicken and mammals raises an interesting questionregarding whether cGHRH is a functional cGHRHR ligand.
The supporting evidence from our present study firstcomes from the restricted spatial expression pattern ofcGHRH in adult chicken brain. cGHRH mRNA is detected to
FIG. 8. A, Schematic representation ofconstructing the expression plasmidsencoding GHRH-PACAP precursor ofdifferent length. SP, Signal peptide; CP,cryptic peptide. B–D, Effects of condi-tioned medium (0–30 �l added, totalvolume 60 �l/well) from DF-1 cells withtransient expression of differentcGHRH-PACAP expression plasmids[B, cGHRH-PACAP(w); C, cGHRH-PACAP(d); and D, cGHRH-PACAP(c)]on basal luciferase activities of CHOcells cotransfected with GHRHR (orPAC1-R) expression plasmid and pGL3-CRE-luciferase reporter construct. Se-rum-free medium (60 �l added) wasused as the control. Each data point rep-resents mean � SEM of three replicates.# and *, P � 0.001 vs. respective con-trols.
2412 Endocrinology, May 2007, 148(5):2405–2416 Wang et al. • Identification of Chicken GHRHR Ligands

be expressed predominantly in the hypothalamus. This isconsistent with findings in mammals that GHRH is mainlylocalized in the arcuate nuclei of hypothalamus using im-munohistochemistry and in situ hybridization (62, 63). Incontrast, cGHRH-PACAP mRNA was detected to be ex-pressed in all regions of the brain examined including hy-pothalamus. The ubiquitous expression of cGHRH-PACAPmRNA in the brain also concurs with previous findings inchickens (64). The differential mRNA expression of the twogenes strongly hints that GHRH gene and GHRH-PACAPgene may have distinct functions in chicken brain. At least,the abundant expression of cGHRH mRNA in the hypothal-amus, together with the predominant expression of cGH-RHR in the pituitary (48, 53, 55), provides spatial evidencethat cGHRH is likely to be a hypophysiotropic factor in theregulation of pituitary functions.
Another proof to support the functionality of cGHRHcomes from the transient expression of cGHRH in CHO cells.Only the conditioned medium from CHO cells transfectedwith constructs encoding GHRH region could activate cGH-RHR (Fig. 6, B–D) and thus increase luciferase activity in adose-dependent manner, suggesting that cGHRH, as inmammals, could activate the intracellular cAMP-PKA sig-naling pathway (13). It also strongly suggests that cGHRHcould function as the endogenous ligand of cGHRHR (Fig. 6,B–D). At present, we do not know how many amino acidresidues are present in the mature chicken GHRH. But the
predicted GHRH peptide is flanked by monobasic (R74) anddibasic residues (R122R123) at its N and C terminus, respec-tively, which are the putative proteolytic cleavage sites formature peptide release (4). Thus, it is conceivable thatcGHRH may have 47 amino acid residues with a free car-boxyl terminus (named cGHRH1–47). This speculation is di-rectly supported by our study that the synthetic cGHRH1–47peptide is of the highest potency at cGHRHR activation (EC500.053 nm) (Fig. 7A). Because the carboxyl-terminally trun-cated cGHRH peptide (cGHRH1–31) still displays high po-tency at cGHRHR activation comparable with that ofcGHRH1–47 (Fig. 7), whether the short form of cGHRH existsin vivo remains to be determined. Interestingly, a conservedconsensus amidation site (G28), which also exists in the pre-cursors of other glucagon family members such as PACAPand VIP (4), has been identified within the cGHRH1–47 region(Figs. 2A and 10A). Therefore, whether an amidation wouldoccur at this specific location, thereby generating a shortamidated form of cGHRH (cGHRH1–27-NH2) in vivo, wouldbe of considerable interest to be investigated (Figs. 2A and10A).
Finally, the evolutionary evidence supports that chickenGHRH gene is orthologous to the mammalian GHRH gene.It was originally hypothesized that mammalian GHRH geneoriginated from a duplicated GHRH-PACAP gene just before
FIG. 9. Identification of two conserved syntenies containing GHRHgene and GHRH-PACAP (or PRP-PACAP) gene, respectively, in ver-tebrate species including chicken, human, Xenopus, and zebrafish. A,GHRH, RPN2, MANBAL, and Src genes are located on a conservedsynteny. B, GHRH-PACAP, clusterin-like 1 (CLUL1), thymidylatesynthetase (TYMS), and v-yes-1 Yamaguchi sarcoma viral oncogene(YES1) genes are located on another conserved synteny. A duplicationof GHRH-PACAP gene (GHRH-PACAP1 and GHRH-PACAP2) is iden-tified in zebrafish. Chr., Chromosome.
FIG. 10. A, Alignment of amino acid sequences of the partial GHRHpeptides from various species including chicken (cGHRH: DQ465018),Xenopus (xGHRH: CX961070), zebrafish (zGHRH: DQ832172), fugu(fGHRH: scaffold_79), tetraodon (tGHRH: CAF96902), and human(hGHRH: X00094). B, Alignment of amino acid sequences of GHRH-like peptides (or PRP in human) from different species includingchicken (cGHRH-L: AY956323), carp (cpGHRH-L), salmon(sGHRH-L: CAA51705), zebrafish (zGHRH-L1: AF217251; zGHRH-L2: AF329730), tetraodon (tGHRH-L1: CAG10213; tGHRH-L2:CAG12289), Xenopus (xGHRH-L: scaffold_420), and human (hPRP:NP_001108). Numbers in brackets indicate sequence identity ofchicken GHRH (A) or GHRH-Like peptide (B) with that of otherspecies. Dots represent the amino acid residues identical with thoseof cGHRH or cGHRH-like peptide. The predicted C-terminal amida-tion (G28) sites in A are boxed.
Wang et al. • Identification of Chicken GHRHR Ligands Endocrinology, May 2007, 148(5):2405–2416 2413

the emergence of mammals (4, 22). In this study, we havefound that GHRH gene is located on a conserved synteny ofall vertebrate species examined, including teleosts and am-phibians (Fig. 9A), revealing that a separate GHRH gene hadalready existed in the last common ancestor of tetrapods andteleosts (Figs. 9 and 11). Moreover, partial or whole GHRHregions, especially the bioactive core of GHRH, have beenidentified in zebrafish, tetraodon, fugu, and Xenopus. Inter-estingly, the cGHRH1–31 is 84% identical with the predictedteleost GHRH1–31 but is only 61% identical with humanGHRH1–31 (Fig. 10A). The high homology between chickenand fish strongly suggests that teleost and cGHRH may playa conserved role in the pituitary. Despite accumulated evi-dence suggesting that PACAP, instead of GHRH, appears toact as a major hypophysiotropic GH-releasing factor in te-leosts (22, 27, 28), it should be noted that cGHRHR could beactivated by oPACAP38 at higher dosages (�10�8 m, EC5037.7 nm) (Fig. 5B) (48), pointing out the possibility that theGH-releasing effect of PACAP, particularly in vitro, may bepartially mediated by pituitary GHRHR in the lower verte-brate species including chicken (Figs. 5B and 12) (48). Un-doubtedly, the identification of the conserved GHRH gene inlower vertebrate species provides us an opportunity to re-evaluate the roles of GHRH and PACAP and their receptorsin the pituitary.
In agreement with the concept of a separate gene encodingGHRH in lower vertebrates, the GHRH-PACAP gene is lo-cated on another conserved synteny of all vertebrate speciesinvestigated (Fig. 9B). Interestingly, two copies of the GHRH-PACAP gene have been identified in teleosts (32, 33), pro-viding a possibility that cGHRH might have evolved fromone of them. However, two copies of the GHRH-PACAPgene and its neighboring genes including clusterin-like 1,thymidylate synthetase, and v-yes-1 Yamaguchi sarcoma vi-ral oncogene were identified on two chromosomes of ze-brafish (chromosomes 2 and 7) (Fig. 9B), clearly showing thattwo copies of the GHRH-PACAP gene have resulted from awhole-genome duplication event that occurred in the teleostlineage (Figs. 9 and 11) (65), as proposed in our previousstudy (32).
The identification of GHRH gene in lower vertebrates alsoquestions the authentic identity of GHRH-like peptide insubmammalian species. Is it a GHRH-like peptide with
GHRH-like activity or a PACAP-related peptide (PRP) with-out known functions? In this study, we have demonstratedthat cGHRH-like peptide is likely to be a ligand of cGHRHR.However, the biological activity of cGHRH-like moleculedetected in conditioned medium largely depends on the typeof construct tested. The conditioned medium from culturedDF-1 cells with transient expression of GHRH-like peptidecould activate cGHRHR but not cPAC1-R (Fig. 8C). In con-trast, the conditioned medium from DF-1 cells with transientexpression of construct encoding complete ORF region ofGHRH-PACAP could activate cPAC1-R but not cGHRHR(Fig. 8B). This finding is quite interesting because bothcGHRH-like peptide and cPACAP are supposed to be re-leased from the same precursor at a theoretical ratio of 1:1and then activate both cGHRHR and cPAC1-R to a certainextent. Because the recombinant cGHRH-like peptide stilldisplays an effect on cGHRHR activation; thus, the inabilityof recombinant GHRH-PACAP(w) on cGHRHR activation isunlikely to be due to the instability of processed GHRH-like
FIG. 11. Proposed evolutionary history of GHRH gene and GHRH-PACAP gene in vertebrate species. The GHRH gene had already appearedin the last common ancestor of tetrapods and teleosts. After emergence of the mammalian lineage, GHRH gene and GHRH-PACAP gene haveundergone an accelerated evolutionary event. The whole-genome duplication (WGD) occurred in teleost lineage resulted in the duplicatedGHRH-PACAP genes (GHRH-PACAP1 and GHRH-PACAP2) in teleost species. Presence of an additional copy of GHRH gene in teleosts remainsquestionable. CP, Cryptic peptide; GHRH-L, GHRH-like peptide; C-peptide, carboxyl terminal flanking peptide.
FIG. 12. Proposed actions of GHRH, GHRH-like peptide (GHRH-L),and PACAP in chicken pituitary. Both GHRH and GHRH-L can ac-tivate GHRHR and thus are supposed to be the endogenous ligandsof GHRHR responsible for GH synthesis and release in somatotroph.The GH-releasing action of PACAP in lower vertebrate species in-cluding chicken is likely to be mediated by PAC1-R (or other PACAPreceptors) or GHRHR or both. The physiological relevance of GHRH-induced PAC1-R activation remains to be clarified.
2414 Endocrinology, May 2007, 148(5):2405–2416 Wang et al. • Identification of Chicken GHRHR Ligands

peptide under our experimental condition. One possible ex-planation is that tissue-specific posttranslational processingmay be involved. In mammals, the precursor of glucagongene, also a member of secretin/VIP/PACAP family, en-compasses three bioactive peptides (glucagon, glucagon-likepeptide-1, and glucagon-like peptide-2) (4, 13), and the list offunctional peptides produced depends on a tissue-specificprocessing event (13). If this is the case for cGHRH-PACAP,it would be of considerable importance to determine howcGHRH-PACAP precursor is processed in the hypothalamusand extrahypothalamic tissues, a question yet to be answeredin all lower vertebrates. Recently Toogood et al. (48) dem-onstrated that GHRH-like peptide has little potency in in-creasing intracellular cAMP level via activation of cGHRHR(48). In this study, we clearly demonstrated that cGHRH-likepeptide could activate cGHRHR with high reproducibility.The discrepancy between the two studies may be due todifferent approaches used (48). In agreement with our find-ings, chicken GHRH-like peptide is 76% identical with carpor salmon GHRH-like peptide (also named GHRH-L1 in thisstudy) of biological activity (66–68), but is only 47–67%identical with the other type of teleost GHRH-like peptide(named GHRH-L2 in this study) with no effect on GHRHRactivation (Fig. 11B) (68). The high degree of conservationbetween cGHRH-like peptide and carp GHRH-like peptide,together with evidence showing the biological activities ofchicken and carp GHRH-like peptides (66), suggests thatGHRH-like peptide (chicken/carp-like) may have importantbiological actions, at least in part, mediated by GHRHR inchicken and teleost fish (Fig. 12).
In summary, the full-length cDNAs encoding chickenGHRH have been cloned in the present study. RT-PCR assayrevealed that cGHRH mRNA is predominantly expressed inthe hypothalamus and functional studies confirmed thatcGHRH could activate cGHRHR with high potency (EC50 �0.1 nm). The activation of cGHRHR by cGHRH-like peptidewas also observed. Moreover, the conserved GHRH genewas identified in genomes of Xenopus, zebrafish, fugu, andtetraodon. Evidence presented here demonstrates thatGHRH, perhaps together with GHRH-like peptide (chicken/carp-like), function as the endogenous ligands of GHRHR inthe chicken, and may act similarly in other lower vertebratespecies (Fig. 12).
Acknowledgments
Received July 27, 2006. Accepted January 24, 2007.Address all correspondence and requests for reprints to: Dr. Frederick
C. Leung, Department of Zoology, The University of Hong Kong,Pokfulam Road, Hong Kong, China. E-mail: [email protected].
This work was supported by the Research Grant Council of the HongKong Government (HKU7345/03M).
The cDNA sequences encoding chicken GHRH and zebrafish GHRHhave been submitted to the DNA Data Base of Japan/European Mo-lecular Biology Laboratory/GenBank databases under accession no.DQ465018, DQ465017, and DQ832172.
Disclosure Statement: Y.W., J.L., C.Y.W., A.H.Y.K., and F.C.L. havenothing to disclose.
References
1. Muller EE, Locatelli V, Cocchi D 1999 Neuroendocrine control of growthhormone secretion. Physiol Rev 79:511–607
2. Frohman LA, Kineman RD 2002 Growth hormone-releasing hormone andpituitary development, hyperplasia and tumorigenesis. Trends EndocrinolMetab 13:299–303
3. Giustina A, Veldhuis JD 1998 Pathophysiology of the neuroregulation ofgrowth hormone secretion in experimental animals and the human. EndocrRev 19:717–797
4. Sherwood NM, Krueckl SL, McRory JE 2000 The origin and function of thepituitary adenylate cyclase-activating polypeptide (PACAP)/glucagon super-family. Endocr Rev 21:619–670
5. Mayo KE, Cerelli GM, Lebo RV, Bruce BD, Rosenfeld MG, Evans RM 1985Gene encoding human growth hormone-releasing factor precursor: structure,sequence, and chromosomal assignment. Proc Natl Acad Sci USA 82:63–67
6. Guillemin R, Brazeau P, Bohlen P, Esch F, Ling N, Wehrenberg WB 1982Growth hormone-releasing factor from a human pancreatic tumor that causedacromegaly. Science 218:585–587
7. Mayo KE, Cerelli GM, Rosenfeld MG, Evans RM 1985 Characterization ofcDNA and genomic clones encoding the precursor to rat hypothalamic growthhormone-releasing factor. Nature 314:464–467
8. Ling N, Esch F, Bohlen P, Brazeau P, Wehrenberg WB, Guillemin R 1984Isolation, primary structure, and synthesis of human hypothalamic somato-crinin: growth hormone-releasing factor. Proc Natl Acad Sci USA 81:4302–4306
9. Gubler U, Monahan JJ, Lomedico PT, Bhatt RS, Collier KJ, Hoffman BJ,Bohlen P, Esch F, Ling N, Zeytin F, Brazeau P, Poonian MS, Gage LP 1983Cloning and sequence analysis of cDNA for the precursor of human growthhormone-releasing factor, somatocrinin. Proc Natl Acad Sci USA 80:4311–4314
10. Rivier J, Spiess J, Thorner M, Vale W 1982 Characterization of a growthhormone-releasing factor from a human pancreatic islet tumour. Nature 300:276–278
11. Brazeau P, Ling N, Bohlen P, Esch F, Ying SY, Guillemin R 1982 Growthhormone releasing factor, somatocrinin, releases pituitary growth hormone invitro. Proc Natl Acad Sci USA 79:7909–7913
12. Lin SC, Lin CR, Gukovsky I, Lusis AJ, Sawchenko PE, Rosenfeld MG 1993Molecular basis of the little mouse phenotype and implications for cell type-specific growth. Nature 364:208–213
13. Mayo KE, Miller LJ, Bataille D, Dalle S, Goke B, Thorens B, Drucker DJ 2003International Union of Pharmacology. XXXV. The glucagon receptor family.Pharmacol Rev 55:167–194
14. Mayo KE 1992 Molecular cloning and expression of a pituitary-specific re-ceptor for growth hormone-releasing hormone. Mol Endocrinol 6:1734–1744
15. Gaylinn BD 2002 Growth hormone releasing hormone receptor. ReceptorsChannels 8:155–162
16. Alba M, Salvatori R 2004 A mouse with targeted ablation of the growthhormone-releasing hormone gene: a new model of isolated growth hormonedeficiency. Endocrinology 145:4134–4143
17. Gaylinn BD, Dealmeida VI, Lyons Jr CE, Wu KC, Mayo KE, Thorner MO1999 The mutant growth hormone-releasing hormone (GHRH) receptor of thelittle mouse does not bind GHRH. Endocrinology 140:5066–5074
18. Alba M, Schally AV, Salvatori R 2005 Partial reversibility of growth hormone(GH) deficiency in the GH-releasing hormone (GHRH) knockout mouse bypostnatal treatment with a GHRH analog. Endocrinology 146:1506–1513
19. Salvatori R, Fan X, Phillips 3rd JA, Espigares-Martin R, Martin De Lara I,Freeman KL, Plotnick L, Al-Ashwal A, Levine MA 2001 Three new mutationsin the gene for the growth hormone (gh)-releasing hormone receptor in familialisolated gh deficiency type ib. J Clin Endocrinol Metab 86:273–279
20. Salvatori R, Hayashida CY, Aguiar-Oliveira MH, Phillips 3rd JA, Souza AH,Gondo RG, Toledo SP, Conceicao MM, Prince M, Maheshwari HG, Bau-mann G, Levine MA 1999 Familial dwarfism due to a novel mutation of thegrowth hormone-releasing hormone receptor gene. J Clin Endocrinol Metab84:917–923
21. Wajnrajch MP, Gertner JM, Harbison MD, Chua Jr SC, Leibel RL 1996Nonsense mutation in the human growth hormone-releasing hormone recep-tor causes growth failure analogous to the little (lit) mouse. Nat Genet 12:88–90
22. Montero M, Yon L, Kikuyama S, Dufour S, Vaudry H 2000 Molecular evo-lution of the growth hormone-releasing hormone/pituitary adenylate cyclase-activating polypeptide gene family. Functional implication in the regulation ofgrowth hormone secretion. J Mol Endocrinol 25:157–168
23. Luo DS, McKeown BA, Rivier J, Vale W 1990 In vitro responses of rainbowtrout (Oncorhynchus mykiss) somatotrophs to carp growth hormone-releasingfactor (GRF) and somatostatin. Gen Comp Endocrinol 80:288–298
24. Melamed P, Eliahu N, Levavi-Sivan B, Ofir M, Farchi-Pisanty O, Rentier-Delrue F, Smal J, Yaron Z, Naor Z 1995 Hypothalamic and thyroidal regulationof growth hormone in tilapia. Gen Comp Endocrinol 97:13–30
25. Parker DB, Power ME, Swanson P, Rivier J, Sherwood NM 1997 Exon skip-ping in the gene encoding pituitary adenylate cyclase-activating polypeptidein salmon alters the expression of two hormones that stimulate growth hor-mone release. Endocrinology 138:414–423
26. Montero M, Yon L, Rousseau K, Arimura A, Fournier A, Dufour S, VaudryH 1998 Distribution, characterization, and growth hormone-releasing activityof pituitary adenylate cyclase-activating polypeptide in the European eel,Anguilla anguilla. Endocrinology 139:4300–4310
27. Wong AO, Leung MY, Shea WL, Tse LY, Chang JP, Chow BK 1998 Hy-pophysiotropic action of pituitary adenylate cyclase-activating polypeptide
Wang et al. • Identification of Chicken GHRHR Ligands Endocrinology, May 2007, 148(5):2405–2416 2415

(PACAP) in the goldfish: immunohistochemical demonstration of PACAP inthe pituitary, PACAP stimulation of growth hormone release from pituitarycells, and molecular cloning of pituitary type I PACAP receptor. Endocrinol-ogy 139:3465–3479
28. Wong AO, Li W, Leung CY, Huo L, Zhou H 2005 Pituitary adenylate cyclase-activating polypeptide (PACAP) as a growth hormone (GH)-releasing factorin grass carp. I. Functional coupling of cyclic adenosine 3�,5�-monophosphateand Ca2�/calmodulin-dependent signaling pathways in PACAP-induced GHsecretion and GH gene expression in grass carp pituitary cells. Endocrinology146:5407–5424
29. Montero M, Yon L, Rousseau K, Arimura A, Fournier A, Dufour S, VaudryH 1998 Localization of pituitary adenylate cyclase-activating polypeptide inthe central nervous system of the European eel Anguilla anguilla: stimulatoryeffect of PACAP on GH secretion. Ann NY Acad Sci 865:475–477
30. Martinez-Fuentes AJ, Gonzalez de Aguilar JL, Lacuisse S, Kikuyama S,Vaudry H, Gracia-Navarro F 1994 Effect of frog pituitary adenylate cyclase-activating polypeptide (PACAP) on amphibian pituitary cells. In: Rosselin G,eds. Vasoactive intestinal peptide, pituitary adenylate cyclase-activatingpolypeptide and related peptide. London: World Scientific; 376–380
31. Sherwood NM, Wu S 2005 Developmental role of GnRH and PACAP in azebrafish model. Gen Comp Endocrinol 142:74–80
32. Wang Y, Wong AO, Ge W 2003 Cloning, regulation of messenger ribonucleicacid expression, and function of a new isoform of pituitary adenylate cyclase-activating polypeptide in the zebrafish ovary. Endocrinology 144:4799–4810
33. Fradinger EA, Sherwood NM 2000 Characterization of the gene encoding bothgrowth hormone-releasing hormone (GRF) and pituitary adenylate cyclase-activating polypeptide (PACAP) in the zebrafish. Mol Cell Endocrinol 165:211–219
34. McRory JE, Parker DB, Ngamvongchon S, Sherwood NM 1995 Sequence andexpression of cDNA for pituitary adenylate cyclase activating polypeptide(PACAP) and growth hormone-releasing hormone (GHRH)-like peptide incatfish. Mol Cell Endocrinol 108:169–177
35. Hu Z, Lelievre V, Tam J, Cheng JW, Fuenzalida G, Zhou X, Waschek JA 2000Molecular cloning of growth hormone-releasing hormone/pituitary adenylylcyclase-activating polypeptide in the frog Xenopus laevis: brain distribution andregulation after castration. Endocrinology 141:3366–3376
36. Alexandre D, Vaudry H, Jegou S, Anouar Y 2000 Structure and distributionof the mRNAs encoding pituitary adenylate cyclase-activating polypeptideand growth hormone-releasing hormone-like peptide in the frog, Rana ridi-bunda. J Comp Neurol 421:234–246
37. McRory JE, Parker RL, Sherwood NM 1997 Expression and alternative pro-cessing of a chicken gene encoding both growth hormone-releasing hormoneand pituitary adenylate cyclase-activating polypeptide. DNA Cell Biol 16:95–102
38. Leung FC, Taylor JE 1983 In vivo and in vitro stimulation of growth hormonerelease in chickens by synthetic human pancreatic growth hormone releasingfactor (hpGRFs). Endocrinology 113:1913–1915
39. Leung FC, Taylor JE, van Iderstine A 1984 Thyrotropin-releasing hormonestimulates body weight gain and increases thyroid hormones and growthhormone in plasma of cockerels. Endocrinology 115:736–740
40. Leung FC, Taylor JE, Wien S, Van Iderstine A 1986 Purified chicken growthhormone (GH) and a human pancreatic GH-releasing hormone increase bodyweight gain in chickens. Endocrinology 118:1961–1965
41. Scanes CG, Harvey S 1984 Stimulation of growth hormone secretion by humanpancreatic growth-hormone-releasing factor and thyrotrophin-releasing hor-mone in anaesthetized chickens. Gen Comp Endocrinol 56:198–203
42. Scanes CG, Harvey S, Rivier J, Vale W 1986 Stimulation of in vivo growthhormone secretion in young chickens by rat hypothalamic growth hormone-releasing factor and synthetic analogues. J Endocrinol 108:413–416
43. Taylor JE, Ball CA, Leung FC 1986 Interaction of human pancreatic growthhormone-releasing factor, thyrotropin-releasing hormone, and somatostatinon growth hormone release in chickens. Proc Soc Exp Biol Med 183:363–367
44. Harvey S 1999 GHRH: a growth hormone-releasing factor in birds? In: PrasadaRao PD, Peter RE, eds. Neural regulation in the vertebrate endocrine system.New York: Kluwer Academic/Plenum Publishers; 69–83
45. Porter TE, Couger GS, Dean CE, Hargis BM 1995 Ontogeny of growth hor-mone (GH)-secreting cells during chicken embryonic development: initialsomatotrophs are responsive to GH-releasing hormone. Endocrinology 136:1850–1856
46. Dean CE, Piper M, Porter TE 1997 Differential responsiveness of somatotrophsto growth hormone-releasing hormone and thyrotropin-releasing hormoneduring chicken embryonic development. Mol Cell Endocrinol 132:33–41
47. Peeters K, Langouche L, Vandesande F, Darras VM, Berghman LR 1998Effects of pituitary adenylate cyclase-activating polypeptide (PACAP) oncAMP formation and growth hormone release from chicken anterior pituitarycells. Ann NY Acad Sci 865:471–474
48. Toogood AA, Harvey S, Thorner MO, Gaylinn BD 2006 Cloning of thechicken pituitary receptor for growth hormone-releasing hormone. Endocri-nology 147:1838–1846
49. Wang Y, Ge W 2003 Involvement of cyclic adenosine 3�,5�-monophosphate inthe differential regulation of activin ßA and ßB expression by gonadotropin inthe zebrafish ovarian follicle cells. Endocrinology 144:491–499
50. Wang Y, Ge W 2003 Gonadotropin regulation of follistatin expression in thecultured ovarian follicle cells of zebrafish, Danio rerio. Gen Comp Endocrinol134:308–315
51. Wang Y, Ge W 2004 Developmental profiles of activin ßA, ßB, and follistatinexpression in the zebrafish ovary: evidence for their differential roles duringsexual maturation and ovulatory cycle. Biol Reprod 71:2056–2064
52. Wang Y, Ge W 2004 Cloning of epidermal growth factor (EGF) and EGFreceptor from the zebrafish ovary: evidence for EGF as a potential paracrinefactor from the oocyte to regulate activin/follistatin system in the follicle cells.Biol Reprod 71:749–760
53. Wang CY, Wang Y, Li J, Leung FC 2006 Expression profiles of growth hor-mone-releasing hormone and growth hormone-releasing hormone receptorduring chicken embryonic pituitary development. Poult Sci 85:569–576
54. Peeters K, Gerets HH, Princen K, Vandesande F 1999 Molecular cloning andexpression of a chicken pituitary adenylate cyclase-activating polypeptidereceptor. Brain Res Mol Brain Res 71:244–255
55. Porter TE, Ellestad LE, Fay A, Stewart JL, Bossis I 2006 Identification of thechicken growth hormone-releasing hormone receptor (GHRH-R) mRNA andgene: regulation of anterior pituitary GHRH-R mRNA levels by homologousand heterologous hormones. Endocrinology 147:2535–2543
56. McRory J, Sherwood NM 1997 Two protochordate genes encode pituitaryadenylate cyclase-activating polypeptide and related family members. Endo-crinology 138:2380–2390
57. Nillni EA, Steinmetz R, Pescovitz OH 1999 Posttranslational processing ofprogrowth hormone-releasing hormone. Endocrinology 140:5817–5827
58. Bloch B, Baird A, Ling N, Guillemin R 1986 Immunohistochemical evidencethat growth hormone-releasing factor (GRF) neurons contain an amidatedpeptide derived from cleavage of the carboxyl-terminal end of the GRF pre-cursor. Endocrinology 118:156–162
59. Arase K, Sakaguchi T, Takahashi M, Bray GA, Ling N 1987 Effects of feedingbehavior of rats of a cryptic peptide from the C-terminal end on prepro-growthhormone-releasing factor. Endocrinology 121:1960–1965
60. Breyer PR, Rothrock JK, Beaudry N, Pescovitz OH 1996 A novel peptide fromthe growth hormone releasing hormone gene stimulates Sertoli cell activity.Endocrinology 137:2159–2162
61. Perez-Riba M, Gonzalez-Crespo S, Boronat A 1997 Differential splicing of thegrowth hormone-releasing hormone gene in rat placenta generates a novelpre-proGHRH mRNA that encodes a different C-terminal flanking peptide.FEBS Lett 402:273–276
62. Suhr ST, Rahal JO, Mayo KE 1989 Mouse growth-hormone-releasing hor-mone: precursor structure and expression in brain and placenta. Mol Endo-crinol 3:1693–1700
63. Bloch B, Brazeau P, Ling N, Bohlen P, Esch F, Wehrenberg WB, Benoit R,Bloom F, Guillemin R 1983 Immunohistochemical detection of growth hor-mone-releasing factor in brain. Nature 301:607–608
64. Peeters K, Gerets HH, Arckens L, Vandesande F 2000 Distribution of pituitaryadenylate cyclase-activating polypeptide and pituitary adenylate cyclase-ac-tivating polypeptide type I receptor mRNA in the chicken brain. J CompNeurol 423:66–82
65. Woods IG, Wilson C, Friedlander B, Chang P, Reyes DK, Nix R, Kelly PD,Chu F, Postlethwait JH, Talbot WS 2005 The zebrafish gene map definesancestral vertebrate chromosomes. Genome Res 15:1307–1314
66. Vaughan JM, Rivier J, Spiess J, Peng C, Chang JP, Peter RE, Vale W 1992Isolation and characterization of hypothalamic growth-hormone releasing fac-tor from common carp, Cyprinus carpio. Neuroendocrinology 56:539–549
67. Chan KW, Yu KL, Rivier J, Chow BK 1998 Identification and characterizationof a receptor from goldfish specific for a teleost growth hormone-releasinghormone-like peptide. Neuroendocrinology 68:44–56
68. Kee F, Ng SS, Vaudry H, Pang RT, Lau EH, Chan SM, Chow BK 2005 Asparticacid scanning mutation analysis of a goldfish growth hormone-releasing hor-mone (GHRH) receptor specific to the GHRH salmon-like peptide. Gen CompEndocrinol 140:41–51
Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
2416 Endocrinology, May 2007, 148(5):2405–2416 Wang et al. • Identification of Chicken GHRHR Ligands




















