
Identification of Neospora caninum proteins regulated during the differentiation process from...
11
RESEARCH ARTICLE Identification of Neospora caninum proteins regulated during the differentiation process from tachyzoite to bradyzoite stage by DIGE Virginia Maruga ´n-Herna ´ndez, Gema A ´ lvarez-Garcı´a, Vero ´nica Risco-Castillo , Javier Regidor-Cerrillo and Luis Miguel Ortega-Mora SALUVET, Animal Health Department, Faculty of Veterinary Sciences, Complutense University of Madrid, Ciudad Universitaria s/n, Madrid, Spain Received: September 21, 2009 Revised: January 20, 2010 Accepted: January 25, 2010 Identification of differentially expressed proteins during Neospora caninum tachyzoite–bra- dyzoite conversion processes may lead to a better knowledge of the pathogenic mechanisms developed by this important parasite of cattle. In the present work, a differential expression proteomic study of tachyzoite and bradyzoite stages was accomplished for the first time by applying DIGE technology coupled with MS analysis. Up to 72 differentially expressed spots were visualized (1.5-fold in relative abundance, po0.05, t-test). A total of 53 spots were more abundant in bradyzoites and 19 spots in tachyzoites. MS analysis identified 26 proteins; 20 of them overexpressed in the bradyzoite stage and 6 in the tachyzoite stage. Among the novel proteins, enolase and glyceraldehyde-3-phosphate dehydrogenase (involved in glycolysis), HSP70 and HSP90 (related to stress response) as well as the dense granule protein GRA9, which showed higher abundance in the bradyzoite stage, might be highlighted. On the other hand, isocitrate dehydrogenase 2, involved in the Krebs cycle, was found to be more abundant in tachyzoites extract. Biological functions from most novel proteins were correlated with previously reported processes during the differentiation process in Toxoplasma gondii. Thus, DIGE technology arises as a suitable tool to study mechanisms involved in the N. caninum tachyzoite to bradyzoite conversion. Keywords: Bradyzoite / Developmentally expressed proteins / DIGE / Microbiology Neospora caninum / Tachyzoite 1 Introduction Neospora caninum is an obligate cyst-forming intracellular protozoan parasite initially misdiagnosed as the closely related Toxoplasma gondii [1]. N. caninum causes abortions or stillbirths, as well as the birth of weak or healthy but congenitally infected calves in cattle, which involves important economic losses in the bovine industry. The life cycle of N. caninum comprises three distinct invasive stages: tachyzoites, bradyzoites located inside tissue cysts and sporozoites contained in oocysts. N. caninum can persist in brain and skeletal muscle through the tissue cyst, forming the bradyzoite stage responsible for chronic infec- tion for many years without the manifestation of clinical signs [2]. However, the reactivation of a chronic infection in a pregnant cow may lead to the switch of quiescent brady- zoites into metabolically active tachyzoites, responsible for Abbreviations: PV, parasitophorous vacuole; PVM, PV membrane Current address: Dr. Vero ´nica Risco-Castillo, Institut National de la Sante ´ et de la Recherche Me ´dicale (INSERM), Unite Mixte de Recherche S945, Paris F-75013, France Correspondence: Dr. Gema A ´ lvarez-Garcı ´a, SALUVET, Animal Health Department, Faculty of Veterinary Sciences, Complutense University of Madrid, Ciudad Universitaria s/n, 28040-Madrid, Spain E-mail: [email protected] Fax: 134-913944095 & 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com 1740 Proteomics 2010, 10, 1740–1750 DOI 10.1002/pmic.200900664
Transcript of Identification of Neospora caninum proteins regulated during the differentiation process from...

RESEARCH ARTICLE
Identification of Neospora caninum proteins regulated
during the differentiation process from tachyzoite
to bradyzoite stage by DIGE
Virginia Marugan-Hernandez, Gema Alvarez-Garcıa, Veronica Risco-Castillo�,Javier Regidor-Cerrillo and Luis Miguel Ortega-Mora
SALUVET, Animal Health Department, Faculty of Veterinary Sciences, Complutense University of Madrid, CiudadUniversitaria s/n, Madrid, Spain
Received: September 21, 2009
Revised: January 20, 2010
Accepted: January 25, 2010
Identification of differentially expressed proteins during Neospora caninum tachyzoite–bra-
dyzoite conversion processes may lead to a better knowledge of the pathogenic mechanisms
developed by this important parasite of cattle. In the present work, a differential expression
proteomic study of tachyzoite and bradyzoite stages was accomplished for the first time by
applying DIGE technology coupled with MS analysis. Up to 72 differentially expressed spots
were visualized (1.5-fold in relative abundance, po0.05, t-test). A total of 53 spots were more
abundant in bradyzoites and 19 spots in tachyzoites. MS analysis identified 26 proteins; 20 of
them overexpressed in the bradyzoite stage and 6 in the tachyzoite stage. Among the novel
proteins, enolase and glyceraldehyde-3-phosphate dehydrogenase (involved in glycolysis),
HSP70 and HSP90 (related to stress response) as well as the dense granule protein GRA9,
which showed higher abundance in the bradyzoite stage, might be highlighted. On the other
hand, isocitrate dehydrogenase 2, involved in the Krebs cycle, was found to be more abundant
in tachyzoites extract. Biological functions from most novel proteins were correlated with
previously reported processes during the differentiation process in Toxoplasma gondii. Thus,
DIGE technology arises as a suitable tool to study mechanisms involved in the N. caninumtachyzoite to bradyzoite conversion.
Keywords:
Bradyzoite / Developmentally expressed proteins / DIGE / Microbiology
Neospora caninum / Tachyzoite
1 Introduction
Neospora caninum is an obligate cyst-forming intracellular
protozoan parasite initially misdiagnosed as the closely
related Toxoplasma gondii [1]. N. caninum causes abortions or
stillbirths, as well as the birth of weak or healthy but
congenitally infected calves in cattle, which involves
important economic losses in the bovine industry.
The life cycle of N. caninum comprises three distinct
invasive stages: tachyzoites, bradyzoites located inside tissue
cysts and sporozoites contained in oocysts. N. caninum can
persist in brain and skeletal muscle through the tissue cyst,
forming the bradyzoite stage responsible for chronic infec-
tion for many years without the manifestation of clinical
signs [2]. However, the reactivation of a chronic infection in
a pregnant cow may lead to the switch of quiescent brady-
zoites into metabolically active tachyzoites, responsible for
Abbreviations: PV, parasitophorous vacuole; PVM, PV
membrane
�Current address: Dr. Veronica Risco-Castillo, Institut National de la
Sante et de la Recherche Medicale (INSERM), Unite Mixte de
Recherche S945, Paris F-75013, France
Correspondence: Dr. Gema Alvarez-Garcıa, SALUVET, Animal
Health Department, Faculty of Veterinary Sciences, Complutense
University of Madrid, Ciudad Universitaria s/n, 28040-Madrid,
Spain
E-mail: [email protected]
Fax: 134-913944095
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
1740 Proteomics 2010, 10, 1740–1750DOI 10.1002/pmic.200900664

the acute phase of infection. Tachyzoites disseminate
quickly, are able to cross the placenta and their replication in
the fetus may cause abortion or birth of congenitally infec-
ted calves [3]. Mechanisms triggering tachyzoite–bradyzoite
stage conversion and vice versa allow Neospora to be a highly
successful intracellular pathogen; however, these mechan-
isms are still unknown. It has been suggested that the
immune status of the host might play an important role in
these events, as described for T. gondii [4].
The expression of developmentally expressed antigens
has been suggested to be involved in immune system
evasion, as well as in the establishment of chronic infection
[2]. Thus, its use in vaccine formulations has been suggested
as a promising alternative, according to the results obtained
in a T. gondii vaccination assay [5]. Moreover, serological and
molecular assays based on antigenic stage-specific proteins
may offer additional information on the determination of
the phase of infection [6, 7]. Until now, Neospora stage-
specific proteins have been identified by genomic approa-
ches. In this sense, only two N. caninum bradyzoite-specific
antigens, NcSAG4 and NcBSR4, have been reported [8, 9],
but their role in parasite persistence remains unknown. On
the other hand, several tachyzoite-specific proteins have
been described, including surface antigens [10, 11] and
organelle components, such as micronemes and
dense granules [12]. In contrast, proteomic techniques
allow large-scale identification of proteins, which may
further lead to the identification of antigens with diagnostic
and vaccine value. Currently, several studies have been done
and focused on the N. caninum tachyzoite stage proteome
[13, 14].
Thus, a proteomic comparative study of N. caninumtachyzoite and bradyzoite stages was accomplished for the
first time. DIGE was coupled with MS to identify proteins
involved in the stage conversion.
2 Materials and methods
2.1 Tachyzoites and bradyzoites
N. caninum tachyzoites from Nc-Liv isolate were grown by
continuous passage in MARC-145 cell culture following
standard procedures. Tachyzoites were harvested 4 days
post-infection. In vitro stage conversion for bradyzoites was
carried out in sodium nitroprusside (Sigma, St. Louis, MO,
USA)-treated cultures [15]. Both zoite productions were
purified by disposable PD-10 desalting columns (GE
Healthcare, Buckinghamshire, UK), and microscope obser-
vations (Nikon Eclipse TS 100) were carefully carried out to
discard parasite batches with host cell contamination. Then,
parasites were counted, pelleted and stored at �801C until
use. Extracts from tachyzoites and bradyzoites were grouped
into four different batches of production of approximately
2� 108 zoites each, which is the required number of repli-
cates to carry out DIGE technology.
2.2 SDS-PAGE and immunoblotting analysis of
tachyzoite and bradyzoite extracts
Homogeneity between parasite batches and bradyzoite
conversions were checked by immunoblotting analysis. A
sample from each batch was resolved by 15% SDS-PAGE
under reducing conditions and transferred to nitrocellulose
membranes following a previously described procedure [16].
Membranes were incubated with both a polyclonal rabbit
antiserum developed against the recombinant NcSAG4
protein [8] at a 1:6000 dilution and a monoclonal mouse
antiserum raised against the intracytoplasmic bradyzoite
antigen TgBAG1 [17, 18] at a 1:2000 dilution. An anti-rabbit
(Sigma) or anti-mouse (GE Healthcare) IgG conjugated with
peroxidase was employed as the secondary antibody
(1:15 000 or 1:3000, respectively). The expression of NcSAG4
and TgBAG1 proteins was visualized by Immobilon
chemiluminescence (Millipore, Bedford, MA, USA).
2.3 Immunofluorescence analysis of tachyzoite and
bradyzoite extracts
The tachyzoite–bradyzoite conversion rate was assessed with
a double-immunofluorescence assay [15]. Coverslips with
infected monolayers were labeled with a monoclonal mouse
antibody directed against the tachyzoite surface antigen
NcSAG1 (1:1000) and a polyclonal rabbit antiserum raised
against the bradyzoite antigen TgBAG1 (1:500) [15] followed
by the appropriate goat IgG coupled with Alexa Fluor 488 or
Alexa Fluor 594 (1:1000) (Invitrogen, Carlsbad, CA, USA).
Coverslips were mounted on a glass slide with Dabco
(Sigma), and photographs were taken using a digital camera
(Nikon Digital Sight DS-L1) connected to an inverted
fluorescence microscope (model TE200 Nikon, 100� oil
immersion objective).
2.4 Preparation and labeling of tachyzoite and
bradyzoite extracts for DIGE
Parasite pellets were suspended in lysis buffer (6 M urea,
2 M thiourea, 4% CHAPS, 1 mM PMSF in 30 mM Tris-HCl,
pH 8.5) and disrupted by sonication cycles. Next, samples
were precipitated with the 2D-Clean up Kit (GE Healthcare)
and resuspended in 50mL of DIGE solution (30 mM Tris,
7 M urea, 2 M thiourea, 4% CHAPS). Protein concentration
was quantified by Bradford assay employing BSA as the
calibration standard. CyDye labeling was performed
following the manufacturer’s protocols (GE Healthcare).
Briefly, 50 mg protein per sample was labeled with 400 pmol
of Cy2, Cy3 or Cy5 fluorochromes dissolved in DMF (99.8%)
for 30 min at 41C in the dark. Then, reactions were quen-
ched with 1mL of lysine (10 mM/50mg protein) for 15 min at
41C in the dark. All DIGE gels included the internal stan-
dard, which was prepared by pooling equal amounts of
Proteomics 2010, 10, 1740–1750 1741
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

protein from each biological sample in the experiment and
labeling with Cy2 dye, so that all proteins from all samples
were represented in the internal standard [19].
2.5 DIGE
A total of 150 mg of protein containing the internal standard
(Cy2-labeled) and tachyzoite (Cy3-labeled for gels 1 and 2,
Cy5-labeled for gels 3 and 4) and bradyzoite extracts (Cy5-
labeled for gels 1 and 2, Cy3-labelled for gels 3 and 4) were
mixed, and an equivalent volume of loading buffer was
added (8 M urea, 4% CHAPS, 130 mM DTT and 2% IPG
buffer, pH 3–10).
Samples were loaded into 24 cm non-linear, pH 3–11,
IPG strips (GE Healthcare) by anodic cup loading and
placed on a manifold. Strips were previously hydrated
overnight with 0.45 mL hydration buffer (7 M urea, 2 M
thiourea, 2% CHAPS w/v, 10 mM DTE, IPG buffer, pH
3–10, and blue bromophenol traces). IEF was performed
with an IPGphor II unit (GE Healthcare) up to a total of
50–55 kVh.
Before the second dimension, strips were equilibrated
twice in 10 mL equilibration buffer (6 M urea, 30% glycerol
v/v, 2% SDS, 100 mM Tris-HCl, pH 6.8), first for 12 min
adding DTE (0.5%) and second for 5 min by adding iodo-
acetamide (4.5%). Strips were sealed with 1% agarose to
12% polyacrylamide gels and proteins were separated (17 h,
2 W/gel) in an Ettan Dalt Six unit (GE Healthcare). To
estimate pI and Mr of spots, a gel containing 2-D SDS-PAGE
Standards (Biorad) was run in parallel to DIGE gels and spot
matching was performed with the DeCyder 6.0 package (GE
Healthcare).
When insufficient amount of differentially expressed
spots was present in DIGE gels for MS, preparative 2-D gels
were run with a total of 300 mg protein and visualized with
an MS-compatible Coomassie blue staining. Prior to MS
analysis, spots from the preparative gel were matched with
those visualized in DIGE gels with DeCyder software for
accurate spot correspondence [20].
2.6 Image analysis and statistics
Image visualization of fluorochrome-labeled proteins was
generated with laser excitation at 488, 532 and 633 nm and
emission filters of 520, 580 and 670 nm for Cy2, Cy3 and
Cy5 fluorochromes, respectively, using a Typhoon 9400
fluorescence scanner (GE Healthcare). Image cropping and
filtering were carried out with Image Quant v. 5.2 software
(GE Healthcare), and image analyses for detection of
different abundance between spots from different stages
were performed with the DIA (Differential In gel Analysis)
module of the DeCyder 6.0 package (GE Healthcare). The
relative protein abundance of a spot was defined as the
normalized spot volume observed in the Cy3 or Cy5 channel
(protein from specific stage) divided by the normalized spot
volume of the same spot measured in the Cy2 channel
(protein reference pool) on the same gel [21]. This value was
used in the t-test statistical analysis. Spots exhibiting over
1.5-fold in their relative abundance with a p-value less than
0.05 in t-test between both stages were considered as
differentially expressed spots.
2.7 MS analysis (MS-MS/MS)
Differentially expressed spots were excised from gels and
proteins selected were in-gel reduced, alkylated and digested
with trypsin [22]. Briefly, spots were washed twice with
water, shrunk for 15 min with 100% ACN and dried in a
Savant SpeedVac for 30 min. Samples were then reduced
with 10 mM DTE in 25 mM ammonium bicarbonate (561C,
30 min) and alkylated with 55 mM iodoacetamide in 25 mM
ammonium bicarbonate for 20 min in the dark. Finally,
samples were digested overnight with 12.5 ng/mL sequen-
cing-grade trypsin (Roche) in 25 mM ammonium bicarbo-
nate (pH 8.5) at 371C.
After digestion, the supernatant was collected, and 1 mL
was spotted onto a MALDI target plate and allowed to air-dry
at room temperature. Matrix (0.4 mL of a 3 mg/mL solution
of CHCA (Sigma) in 50% ACN) was added to the dried
peptide digest spots and allowed again to air-dry at
room temperature. MALDI-TOF MS fingerprinting was
performed in a MALDI-TOF/TOF mass spectrometer (4700
Proteomics Analyzer; PerSeptive Biosystems) operating in
reflector mode with an accelerating voltage of 20 000 V. All
mass spectra were calibrated externally using a standard
peptide mixture (Sigma).
For protein identification monoisotopic peptide masses
were compared with NCBI non-redundant (NCBI nr),
Swiss-PROT/TrEMBL and ToxoDB 5.0 databases using
the MASCOT algorithm v2.1 (Matrix Science) through the
Global Protein Server v3.5 from Applied Biosystems. The
apicomplexan-specific ToxoDB database contains T. gondiiand N. caninum genome sequences. For MS/MS sequencing
analyses, suitable precursors were selected and fragmenta-
tion was carried out using CID. MASCOT search para-
meters were as follows: carbamidomethyl cysteine as fixed
modification and oxidized methionine as variable modifi-
cation; peptide mass tolerance 50–100 ppm; one missed
trypsin cleavage site; MS/MS fragments tolerance 0.3 Da.
The parameters for the combined search (peptide mass
fingerprint and MS/MS spectra) were as described above. In
all protein identifications, the probability scores were greater
than the score fixed by MASCOT as significant with a
p-value less than 0.05.
De novo sequencing from fragmentation spectra of
peptides was performed using the DeNovo software tool
(Applied Biosystems), and database homology searches of
the sequences were carried out by BLAST (http://
www.ncbi.nlm.nih.gov/BLAST).
1742 V. Marugan-Hernandez et al. Proteomics 2010, 10, 1740–1750
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

3 Results
3.1 Analysis of tachyzoite and bradyzoite extracts
First, double-immunofluorescence assays in cell cultures
raised around 60% of bradyzoite-positive parasitophorous
vacuoles (PVs) (Fig. 1A). Second, positive reactions with
anti-TgBAG1 and anti-NcSAG4 antibodies directed against
specific bradyzoite proteins of 30 and 18 kDa, respectively,
were obtained by immunoblotting (Fig. 1B). Neither anti-
body showed any reaction with pure tachyzoite extracts (data
not shown).
3.2 Differentially expressed proteins detected by
DIGE analysis
More than 2000 spots were detected for each gel, and spots
with a significant increase (or decrease) in their relative
abundance were considered as developmentally expressed
proteins. After bioinformatics analysis, a total of up to 72
differentially expressed proteins were obtained: 53 were
more abundant in the bradyzoite stage and 19 in the
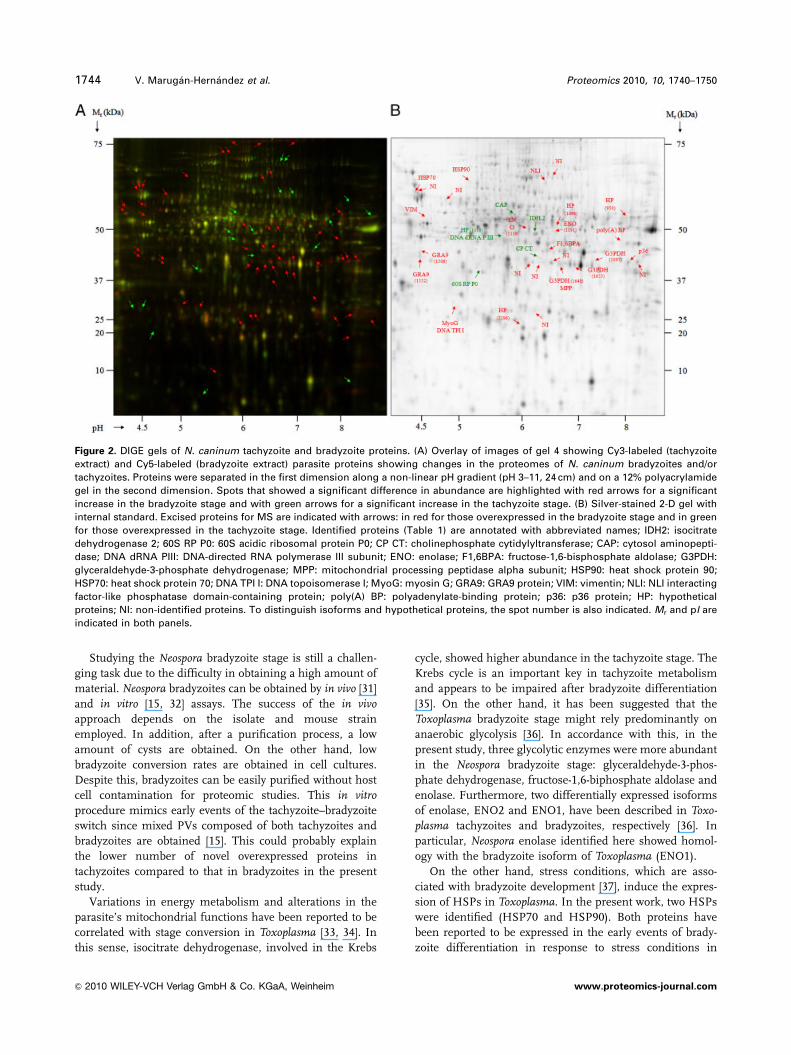
tachyzoite stage (Fig. 2A).
3.3 Protein identification
A total of 31 spots were excised for MS analysis (Fig. 2B).
Only four proteins were identified as Toxoplasma proteins
when spectra data were compared against GenBank or
SwissProt non-redundant databases (data not shown).
Identifications notably increased when the ToxoDB 5.0
database was employed, and 23 spots were identified. Mass
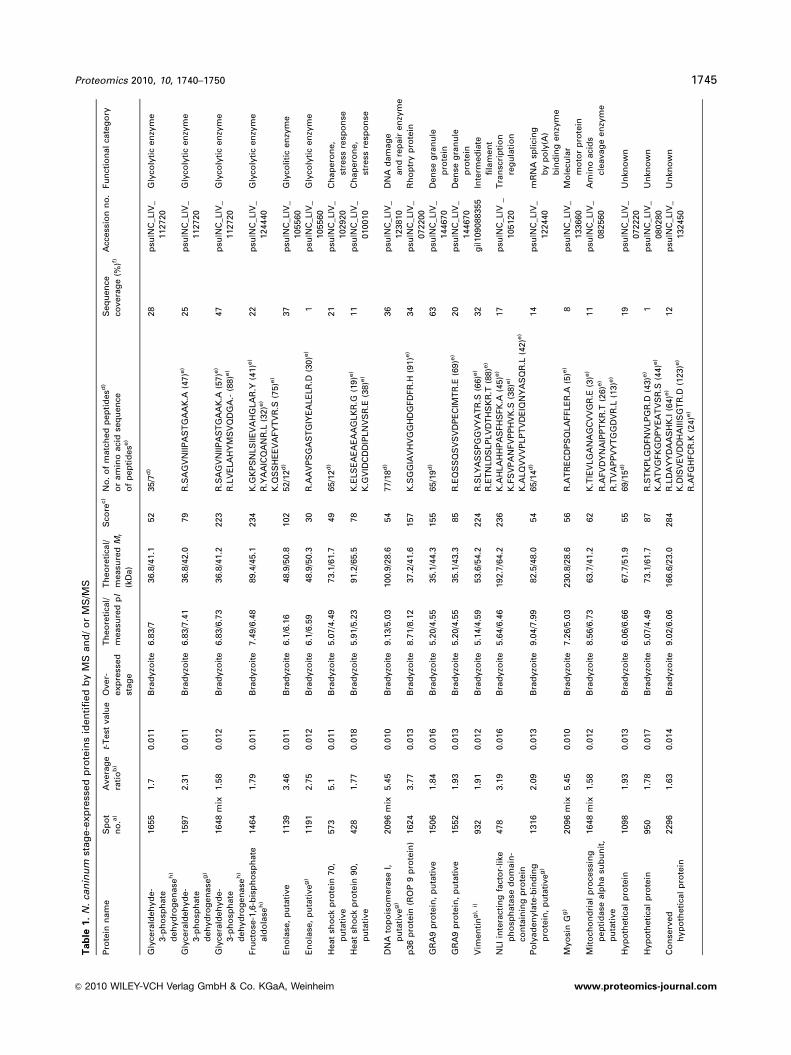
fingerprint identified 11, and MS/MS combined with MS
identified 15 (Table 1). Of these spots, three showed double
protein identification so that 25 different Neospora proteins
were identified. In addition, a host cell protein named
vimentin was also identified, and spectra from eight addi-
tional spots did not correspond to any annotated protein.
In total, 20 out of 26 novel proteins were more abundant
in the bradyzoite stage, whereas six proteins were detected
as over-expressed in tachyzoites (Figs. 2B; Table 1).
The three double identifications were as follows: glycer-
aldehyde-3-phosphate dehydrogenase – mitochondrial
processing peptidase a subunit, DNA topoisomerase I –
myosin G in bradyzoites, and a hypothetical protein – DNA-
directed RNA polymerase III subunit in tachyzoites.
Different isoforms were detected for enolase, glycer-
aldehyde-3-phosphate dehydrogenase and GRA9 (Table 1).
4 Discussion
Tissue cyst formation containing bradyzoites is thought to
be a key factor in immune evasion by N. caninum, which
ensures its persistence in the host and makes possible
further transmission to newborn animals. During the
development of an acute infection, tachyzoites are partially
cleared by the immune response developed by the host. The
surviving parasites switch into metabolically quiescent and
low-dividing bradyzoites gathered in tissue cysts. In this
way, the parasite maintains a balance between multi-
plication and survival in the host and succeeds in persisting
[23].
Mechanisms triggering N. caninum tachyzoite–brady-
zoite conversion in apicomplexa are still unknown. In the
present study, N. caninum developmentally expressed
proteins in both tachyzoite and bradyzoite stages have been
identified for the first time by DIGE technology coupled to
MS analysis, a powerful tool to quantitatively detect devel-
opmentally expressed proteins [24].
Currently, few proteomic studies have been developed in
N. caninum, and these were only focussed on the tachyzoite
stage [13, 14, 25]. In addition, no attempt has ever been
made to identify bradyzoite-specific proteins by means of
proteomic tools in cyst-forming coccidia. This proteomic
approach has been successfully employed in other apicom-
plexan parasites to study parasite–host cell interactions
[26, 27]. Furthermore, large-scale studies on tachyzoite–
bradyzoite conversion have only been accomplished in
T. gondii by means of genomic tools [28–30].
Figure 1. Analysis of bradyzoite extracts. (A) Immuno-
fluorescence of N. caninum bradyzoite-positive parasitophorous
vacuoles in cell culture at day 7 post-treatment with 70 mM of
sodium nitroprusside. Panel 1: cell nuclear material detected by
DAPI staining. Panel 2: tachyzoite recognition by polyclonal anti-
NcSAG1 (red). Panel 3: bradyzoite recognition by monoclonal
anti-TgBAG1 (green). Panel 4: merge. (B) Immunoblotting
analysis of bradyzoite extracts with antibodies directed against
the bradyzoite stage-specific proteins NcSAG4 (18 kDa) and
TgBAG1 (30 kDa). Lane number indicates the bradyzoite batch
number for each DIGE gel replicate.
Proteomics 2010, 10, 1740–1750 1743
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
Studying the Neospora bradyzoite stage is still a challen-
ging task due to the difficulty in obtaining a high amount of
material. Neospora bradyzoites can be obtained by in vivo [31]
and in vitro [15, 32] assays. The success of the in vivoapproach depends on the isolate and mouse strain
employed. In addition, after a purification process, a low
amount of cysts are obtained. On the other hand, low
bradyzoite conversion rates are obtained in cell cultures.
Despite this, bradyzoites can be easily purified without host
cell contamination for proteomic studies. This in vitroprocedure mimics early events of the tachyzoite–bradyzoite
switch since mixed PVs composed of both tachyzoites and
bradyzoites are obtained [15]. This could probably explain
the lower number of novel overexpressed proteins in
tachyzoites compared to that in bradyzoites in the present
study.
Variations in energy metabolism and alterations in the
parasite’s mitochondrial functions have been reported to be
correlated with stage conversion in Toxoplasma [33, 34]. In
this sense, isocitrate dehydrogenase, involved in the Krebs
cycle, showed higher abundance in the tachyzoite stage. The
Krebs cycle is an important key in tachyzoite metabolism
and appears to be impaired after bradyzoite differentiation
[35]. On the other hand, it has been suggested that the
Toxoplasma bradyzoite stage might rely predominantly on
anaerobic glycolysis [36]. In accordance with this, in the
present study, three glycolytic enzymes were more abundant
in the Neospora bradyzoite stage: glyceraldehyde-3-phos-
phate dehydrogenase, fructose-1,6-biphosphate aldolase and
enolase. Furthermore, two differentially expressed isoforms
of enolase, ENO2 and ENO1, have been described in Toxo-plasma tachyzoites and bradyzoites, respectively [36]. In
particular, Neospora enolase identified here showed homol-
ogy with the bradyzoite isoform of Toxoplasma (ENO1).
On the other hand, stress conditions, which are asso-
ciated with bradyzoite development [37], induce the expres-
sion of HSPs in Toxoplasma. In the present work, two HSPs
were identified (HSP70 and HSP90). Both proteins have
been reported to be expressed in the early events of brady-
zoite differentiation in response to stress conditions in
Figure 2. DIGE gels of N. caninum tachyzoite and bradyzoite proteins. (A) Overlay of images of gel 4 showing Cy3-labeled (tachyzoite
extract) and Cy5-labeled (bradyzoite extract) parasite proteins showing changes in the proteomes of N. caninum bradyzoites and/or
tachyzoites. Proteins were separated in the first dimension along a non-linear pH gradient (pH 3–11, 24 cm) and on a 12% polyacrylamide
gel in the second dimension. Spots that showed a significant difference in abundance are highlighted with red arrows for a significant
increase in the bradyzoite stage and with green arrows for a significant increase in the tachyzoite stage. (B) Silver-stained 2-D gel with
internal standard. Excised proteins for MS are indicated with arrows: in red for those overexpressed in the bradyzoite stage and in green
for those overexpressed in the tachyzoite stage. Identified proteins (Table 1) are annotated with abbreviated names; IDH2: isocitrate
dehydrogenase 2; 60S RP P0: 60S acidic ribosomal protein P0; CP CT: cholinephosphate cytidylyltransferase; CAP: cytosol aminopepti-
dase; DNA dRNA PIII: DNA-directed RNA polymerase III subunit; ENO: enolase; F1,6BPA: fructose-1,6-bisphosphate aldolase; G3PDH:
glyceraldehyde-3-phosphate dehydrogenase; MPP: mitochondrial processing peptidase alpha subunit; HSP90: heat shock protein 90;
HSP70: heat shock protein 70; DNA TPI I: DNA topoisomerase I; MyoG: myosin G; GRA9: GRA9 protein; VIM: vimentin; NLI: NLI interacting
factor-like phosphatase domain-containing protein; poly(A) BP: polyadenylate-binding protein; p36: p36 protein; HP: hypothetical
proteins; NI: non-identified proteins. To distinguish isoforms and hypothetical proteins, the spot number is also indicated. Mr and pI are
indicated in both panels.
1744 V. Marugan-Hernandez et al. Proteomics 2010, 10, 1740–1750
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
Tab
le1.
N.
can
inu
mst
ag
e-e
xp
ress
ed
pro
tein
sid
en
tifi
ed
by
MS
an
d/
or
MS
/MS
Pro
tein
nam
eS
po
t
no
.a)
Avera
ge
rati
ob
)
t-T
est
valu
eO
ver-
exp
ress
ed
stag
e
Th
eo
reti
cal/
measu
red
pI
Th
eo
reti
cal/
measu
red
Mr
(kD
a)
Sco
rec)
No
.o
fm
atc
hed
pep
tid
esd
)
or
am
ino
aci
dse
qu
en
ce
of
pep
tid
ese
)
Seq
uen
ce
covera
ge
(%)f)
Acc
ess
ion
no
.Fu
nct
ion
al
cate
go
ry
Gly
cera
ldeh
yd
e-
3-p
ho
sph
ate
deh
yd
rog
en
ase
h)
1655
1.7
0.0
11
Bra
dyzo
ite
6.8
3/7
36.8
/41.1
52
35/7
d)
28
psu
|NC
_LIV
_
112720
Gly
coly
tic
en
zym
e
Gly
cera
ldeh
yd
e-
3-p
ho
sph
ate
deh
yd
rog
en
ase
g)
1597
2.3
10.0
11
Bra
dyzo
ite
6.8
3/7
.41
36.8
/42.0
79
R.S
AG
VN
IIP
AS
TG
AA
K.A
(47)e
)25
psu
|NC
_LIV
_
112720
Gly
coly
tic
en
zym
e
Gly
cera
ldeh
yd
e-
3-p
ho
sph
ate
deh
yd
rog
en
ase
h)
1648
mix
1.5
80.0
12
Bra
dyzo
ite
6.8
3/6
.73
36.8
/41.2
223
R.S
AG
VN
IIP
AS
TG
AA
K.A
(57)e
)
R.L
VE
LA
HY
MS
VQ
DG
A.-
(88)e
)
47
psu
|NC
_LIV
_
112720
Gly
coly
tic
en
zym
e
Fru
cto
se-1
,6-b
isp
ho
sph
ate
ald
ola
seh
)
1464
1.7
90.0
11
Bra
dyzo
ite
7.4
9/6
.48
89.4
/45.1
234
K.G
KP
SN
LS
IIE
VA
HG
LA
R.Y
(41)e
)
R.Y
AA
ICQ
AN
R.L
(32)e
)
22
psu
|NC
_LIV
_
124440
Gly
coly
tic
en
zym
e
K.Q
SS
HE
EV
AFY
TV
R.S
(75)e
)
En
ola
se,
pu
tati
ve
1139
3.4
60.0
11
Bra
dyzo
ite
6.1
/6.1
648.9
/50.8
102
52/1
2d
)37
psu
|NC
_LIV
_
105560
Gly
coli
tic
en
zym
e
En
ola
se,
pu
tati
ve
g)
1191
2.7
50.0
12
Bra
dyzo
ite
6.1
/6.5
948.9
/50.3
30
R.A
AV
PS
GA
ST
GIY
EA
LE
LR
.D(3
0)e
)1
psu
|NC
_LIV
_
105560
Gly
coly
tic
en
zym
e
Heat
sho
ckp
rote
in70,
pu
tati
ve
573
5.1
0.0
11
Bra
dyzo
ite
5.0
7/4
.49
73.1
/61.7
49
65/1
2d
)21
psu
|NC
_LIV
_
102920
Ch
ap
ero
ne,
stre
ssre
spo
nse
Heat
sho
ckp
rote
in90,
pu
tati
ve
428
1.7
70.0
18
Bra
dyzo
ite
5.9
1/5
.23
91.2
/65.5
78
K.E
LS
EA
EA
EA
AG
LK
R.G
(19)e
)
K.G
VID
CD
DIP
LN
VS
R.E
(38)e
)
11
psu
|NC
_LIV
_
010010
Ch
ap
ero
ne,
stre
ssre
spo
nse
DN
Ato
po
iso
mera
seI,
pu
tati
ve
g)
2096
mix
5.4
50.0
10
Bra
dyzo
ite
9.1
3/5
.03
100.9
/28.6
54
77/1
8d
)36
psu
|NC
_LIV
_
123810
DN
Ad
am
ag
e
an
dre
pair
en
zym
e
p36
pro
tein
(RO
P9
pro
tein
)1624
3.7
70.0
13
Bra
dyzo
ite
8.7
1/8
.12
37.2
/41.6
157
K.S
GG
IAV
HV
GG
HD
GFD
FR
.H(9
1)e
)34
psu
|NC
_LIV
_
072200
Rh
op
try
pro
tein
GR
A9
pro
tein
,p
uta
tive
1506
1.8
40.0
16
Bra
dyzo
ite
5.2
0/4
.55
35.1
/44.3
155
65/1
9d
)63
psu
|NC
_LIV
_
144670
Den
seg
ran
ule
pro
tein
GR
A9
pro
tein
,p
uta
tive
1552
1.9
30.0
13
Bra
dyzo
ite
5.2
0/4
.55
35.1
/43.3
85
R.E
QS
SQ
SV
SV
DP
EC
IMT
R.E
(69)e
)20
psu
|NC
_LIV
_
144670
Den
seg
ran
ule
pro
tein
Vim
en
tin
g),
i)932
1.9
10.0
12
Bra
dyzo
ite
5.1
4/4
.59
53.6
/54.2
224
R.S
LY
AS
SP
GG
VY
AT
R.S
(66)e
)32
gi|109088355
Inte
rmed
iate
fila
men
tR
.ET
NLD
SLP
LV
DT
HS
KR
.T(8
8)e
)
NLI
inte
ract
ing
fact
or-
like
ph
osp
hata
sed
om
ain
-
con
tain
ing
pro
tein
478
3.1
90.0
16
Bra
dyzo
ite
5.6
4/6
.46
192.7
/64.2
236
K.A
HLA
HH
PA
SFH
SFK
.A(4
5)e
)17
psu
|NC
_LIV
_
105120
Tra
nsc
rip
tio
n
reg
ula
tio
nK
.FS
VP
AN
FV
PP
HV
K.S
(38)e
)
K.A
LQ
VV
VP
LP
TV
DE
IQN
YA
SQ
R.L
(42)e
)
Po
lyad
en
yla
te-b
ind
ing
pro
tein
,p
uta
tive
g)
1316
2.0
90.0
13
Bra
dyzo
ite
9.0
4/7
.99
82.5
/48.0
54
65/1
4d
)14
psu
|NC
_LIV
_
122440
mR
NA
spli
cin
g
by
po
ly(A
)
bin
din
gen
zym
e
Myo
sin
Gg
)2096
mix
5.4
50.0
10
Bra
dyzo
ite
7.2
6/5
.03
230.8
/28.6
56
R.A
TR
EC
DP
SQ
LA
FFLE
R.A
(5)e
)8
psu
|NC
_LIV
_
133660
Mo
lecu
lar
mo
tor
pro
tein
Mit
och
on
dri
al
pro
cess
ing
pep
tid
ase
alp
ha
sub
un
it,
pu
tati
ve
1648
mix
1.5
80.0
12
Bra
dyzo
ite
8.5
6/6
.73
63.7
/41.2
62
K.T
IEV
LG
AN
AG
CV
VG
R.E
(3)e
)11
psu
|NC
_LIV
_
082560
Am
ino
aci
ds
cleavag
een
zym
eR
.AFV
DY
NA
IPP
TK
R.T
(26)e
)
R.T
VA
PP
VY
TG
GD
VR
.L(1
3)e
)
Hyp
oth
eti
cal
pro
tein
1098
1.9
30.0
13
Bra
dyzo
ite
6.0
6/6
.66
67.7
/51.9
55
69/1
5d
)19
psu
|NC
_LIV
_
072220
Un
kno
wn
Hyp
oth
eti
cal
pro
tein
950
1.7
80.0
17
Bra
dyzo
ite
5.0
7/4
.49
73.1
/61.7
87
R.S
TK
PLG
DFN
VLP
GR
.D(4
3)e
)1
psu
|NC
_LIV
_
080280
Un
kno
wn
K.A
TV
GFK
GD
PY
EA
TV
SR
.S(4
4)e
)
Co
nse
rved
hyp
oth
eti
cal
pro
tein
2296
1.6
30.0
14
Bra
dyzo
ite
9.0
2/6
.06
166.6
/23.0
284
R.L
DA
YY
DA
AS
HK
.I(6
4)e
)12
psu
|NC
_LIV
_
132450
Un
kno
wn
K.D
ISV
EV
DD
HA
IIIS
GT
R.D
(123)e
)
R.A
FG
HFC
R.K
(24)e
)
Proteomics 2010, 10, 1740–1750 1745
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Tab
le1.
Co
nti
nu
ed
Pro
tein
nam
eS
po
t
no
.a)
Avera
ge
rati
ob
)
t-T
est
valu
eO
ver-
exp
ress
ed
stag
e
Th
eo
reti
cal/
measu
red
pI
Th
eo
reti
cal/
measu
red
Mr
(kD
a)
Sco
rec)
No
.o
fm
atc
hed
pep
tid
esd
)
or
am
ino
aci
dse
qu
en
ce
of
pep
tid
ese
)
Seq
uen
ce
covera
ge
(%)f)
Acc
ess
ion
no
.Fu
nct
ion
al
cate
go
ry
Iso
citr
ate
deh
yd
rog
en
ase
2
1225
�2.5
10.0
11
Tach
yzo
ite
5.5
2/6
.26
112.5
/49.6
222
K.A
ES
LV
CE
GP
GD
FT
ISFT
PA
GG
AR
.V(6
1)e
)19
psu
|NC
_LIV
_
132780
TC
Aen
zym
eK
.QG
IWY
EH
R.L
(12)e
)
K.T
ST
NP
IAS
IFA
WT
R.G
(43)e
)
R.L
GQ
FC
LA
LE
R.A
(28)e
)
60S
aci
dic
rib
oso
mal
pro
tein
P0
h)
1738
�2.1
30.0
11
Tach
yzo
ite
5.4
4/5
.42
33.9
/38.8
54
47/7
d)
30
psu
|NC
_LIV
_
141370
Mem
bra
ne
pro
tein
Ch
oli
nep
ho
sph
ate
cyti
dyly
ltra
nsf
era
se,
pu
tati
ve
1569
�1.7
60.0
11
Tach
yzo
ite
7.8
1/6
.32
33.1
/43.0
57
R.I
LQ
NY
ED
YV
DR
.S(4
2)e
)15
psu
|NC
_LIV
_
135050
Mem
bra
ne
ph
osp
ho
lip
id
syn
thesi
sen
zym
e
Cyto
sol
am
ino
pep
tid
ase
902
�1.5
80.0
16
Tach
yzo
ite
6.0
2/5
.92
60.4
/55.2
100
72/1
6d
)34
psu
|NC
_LIV
_
113250
Am
ino
aci
ds
cleavag
een
zym
e
DN
A-d
irect
ed
RN
A
Po
lym
era
seII
Isu
bu
nit
,
pu
tati
ve
1318
mix�
2.1
10.0
14
Tach
yzo
ite
8.7
8/5
.99
151.8
/47.9
57
65/1
9d
)11
psu
|NC
_LIV
_
103390
Rib
on
ucl
eo
tid
e
po
lym
eri
zati
on
en
zym
e
Hyp
oth
eti
cal
pro
tein
1318
mix�
2.1
10.0
14
Tach
yzo
ite
8.9
3/5
.99
50.9
/47.9
51
67/1
0d
)23
psu
|NC
_LIV
_
094430
Un
kno
wn
a)
Sp
ots
that
con
tain
ed
mo
reth
an
on
ep
rote
inare
lab
ell
ed
as
‘‘m
ix.’
’b
)A
vera
ge
vo
lum
era
tio
of
the
spo
ts(b
rad
yzo
ites
vers
us
tach
yzo
ites)
qu
an
tifi
ed
by
DeC
yd
er
bio
log
ical
vari
ati
on
an
aly
sis
mo
du
le.
On
lysp
ots
exh
ibit
ing
over
1.5
-fo
ldin
their
rela
tive
ab
un
dan
cew
ith
apo
0.0
5in
t-te
stb
etw
een
bo
thst
ag
es
were
an
aly
zed
by
MS
for
iden
tifi
cati
on
.c)
MA
SC
OT
MS
pro
tein
sco
re,
ob
tain
ed
fro
mM
ALD
IT
OF/T
OF
spect
ra.
Inall
case
s,th
ep
rob
ab
ilit
ysc
ore
waso
0.0
5.
d)N
um
ber
of
pep
tid
em
ass
es
valu
es
searc
h/m
atc
hed
.e)
Am
ino
aci
dse
qu
en
ceid
en
tifi
ed
by
MS
/MS
;th
eio
nsc
ore
isin
dic
ate
din
pare
nth
ese
s.f)
Am
ino
aci
dse
qu
en
ceco
vera
ge
for
the
iden
tifi
ed
pro
tein
sb
yM
San
dM
S/M
S.
g)A
pp
are
ntl
yo
nly
vis
uali
zed
inb
rad
yzo
ite
extr
act
s.h
)Als
oid
en
tifi
ed
inN
CB
In
rd
ata
base
as
the
sam
ep
rote
in,
bu
tfr
om
T.
go
nd
ii.
i)V
imen
tin
was
iden
tifi
ed
inG
en
Ban
kas
aM
aca
cca
mu
latt
a(r
hesu
sm
aca
qu
es)
pro
tein
,co
rresp
on
din
gto
the
ho
stce
ll(M
AR
C-1
45
cell
lin
e).
1746 V. Marugan-Hernandez et al. Proteomics 2010, 10, 1740–1750
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Toxoplasma [37, 38]. Moreover, one bradyzoite overexpressed
hypothetical protein showed high homology (87% identity)
with a T. gondii bradyzoite-specific HSP (HSP21) when
BLAST was performed. Even more, it exhibited similar Mr
in a DIGE gel to TgHSP21 (data not shown). These stress
conditions could result in DNA damage in the slow repli-
cating bradyzoite stage [36], which may inhibit cell replica-
tion and delay a single cell’s entry into mitosis. Different
enzymes involved in DNA damage and repair have been
detected overexpressed in encysted bradyzoites in Toxo-plasma [39, 40]. Concerning this issue, a DNA topoisome-
rase I was found to be more abundant in the Neosporabradyzoite stage.
Other events associated with stage conversion in Toxo-plasma are changes in the main surface antigens [41]. In this
sense, the ribosomal protein P0 was found overexpressed in
tachyzoites. This protein is detected on the surface of N.caninum tachyzoites, though no assays have been performed
in bradyzoites, and seems to be directly implicated in host
cell invasion [42].
Moreover, several changes take place in the PV
membrane (PVM) after parasite invasion, with the subse-
quent formation of a thicker structure to protect bradyzoites
from environmental conditions and communicate with host
cells. Within the vacuolar compartment, Toxoplasma rhoptry
proteins are involved in PV formation, and some of them are
targeted into the host cell nucleus. On the other hand, dense
granule proteins associate with the tubular network deli-
miting membrane [43]. In our study, we report the over-
expression of ROP9 and GRA9 in the bradyzoite stage.
Conversely, in T. gondii, ROP9 has been reported as the only
tachyzoite-specific rhoptry protein [44], and GRA9 has been
described in both stages [45]. According to this finding, the
higher level of GRA9 expression in bradyzoites may give
evidence for its secretion in the PV and its implication in the
network of membranous tubules in the latest moment of
vacuole formation.
On the other hand, it is also known that tissue cysts are
wrapped into host cell filaments [46]. Interestingly, one host
cell intermediate filament, vimentin, was also more abun-
dant in bradyzoites. An explanation for this finding is the
association between vimentin and bradyzoite proteins
involved in the matrix conformation of PVs. Indeed,
vimentin is an intermediate filament that arises from the
host cell nuclear surface and progressively rearranges
around the enlarging PV compartment. In this way, the host
filament network probably serves to dock the parasite
compartment to the host cell nuclear surface [46].
Two additional identifications might be associated with
the tachyzoite–bradyzoite switch, although no previous data
have been reported in any apicomplexan parasite. First, NLI
interacting factor-like phosphatase domain-containing
protein may be involved in differentiation processes since it
plays an essential role in the development of neuronal
lineages and embryonic development [47]. Second, poly(A)
tail metabolism driven by a putative polyadenylate-binding
protein might be one of numerous cellular processes that
normally protect cells from damaging effects of high stress
conditions [48]. No biological function has been associated
with the mitochondrial processing peptidase a subunit in
apicomplexan parasites.
The myosin G heavy chain, belonging to class XXIII, has
been previously described in Toxoplasma and has an ortho-
logue in Eimeria tenella [49]. It remains unclear why myosin
G heavy chain is more abundant in the bradyzoite stage, due
to the low replication and motility found when the differ-
entiation process occurs. The only feasible explanation is its
involvement in organelle redistribution.
Our findings in tachyzoites were mainly related with the
replication process. In particular, cholinephosphate cytidy-
lyltransferase is necessary for the synthesis of phosphati-
dylcholine, which is essential for rapid parasite replication
when significant biogenesis of parasite membranes and the
concomitant enlargement of the PVM occur in T. gondii [50].
On the other hand, the overexpression of cytosolic amino-
peptidase in tachyzoites indicates that this enzymatic activity
increases during the stage when the parasite is most meta-
bolically active. Finally, DNA-dependent RNA polymerase
was also more abundant in the tachyzoite stage, probably
associated with a higher multiplication rate.
Several novel proteins showed differences between their
theoretical and measured molecular weight in DIGE gels. In
the case of isocitrate dehydrogenase 2, NLI interacting
factor-like phosphatase domain-containing protein and a
conserved hypothetical protein, these differences could be
explained by the annotation of Neospora sequences in the
database. When BLAST was performed against Toxoplasmadatabase, the theoretical molecular weight of the Neosporaprotein corresponded with two different and consecutive
Toxoplasma proteins. However, Neospora protein exhibited
homology with just one of the Toxoplasma proteins, whose
molecular weight was similar to that one exhibited by the
Neospora protein in DIGE gels. On the contrary, the theo-
retical molecular weight of myosin G, DNA topoisomerase I
and DNA-directed RNA Polymerase III subunit were iden-
tical to their Toxoplasma homologues. Thus, differences
between theoretical and measured molecular weights could
be explained by a protein processing.
Two SAG1 related sequence-family proteins, NcSAG4
and NcBSR4, have been previously identified as devel-
opmentally expressed in the Neospora bradyzoite stage [8, 9].
However, they were not found in the present study, probably
due to two possible explanations: the poor resolution of
hydrophobic proteins in 2-D PAGE and, in particular,
NcBSR4 seems to be expressed late during the tachyzoite to
bradyzoite differentiation process [9].
Furthermore, our data show that most spots are shared
between both developmental stages and do not show
differential expression, indicating that most visualized
proteins belong to housekeeping processes. As DIGE
analysis does not allow the identification of stage-specific
proteins, it is likely that several proteins detected
Proteomics 2010, 10, 1740–1750 1747
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

in the present study, such as one enolase and
glyceraldehyde-3-phosphate dehydrogenase isoforms, poly-
adenylate-binding protein, DNA topoisomerase I, myosin G
and vimentin, seem to be bradyzoite-specific proteins
according to DeCyder analysis (data not shown).
It is worthwhile mentioning that several of novel proteins
identified in the present study have been proposed as ther-
apeutic targets for diseases caused by apicomplexan parasites.
Apicomplexan glycolytic enzymes, which are plant homo-
logues, have been suggested as novel chemotherapeutic
targets [51]. Similarly, disruption of phospholipid metabolic
pathways was suggested to combat parasite growth in Toxo-plasma [50] and Plasmodium [52]. Inhibitors against amino-
peptidases [53] and type I topoisomerases [54] seem to be
potential drugs for the treatment of human parasitic diseases,
such as malaria or leishmaniasis, together with drugs block-
ing HSP90 functions that affect invasion and conversion of
the parasite [38]. Concerning vaccine development, antigens
that are known to be important for parasite survival should be
considered for inclusion in a N. caninum vaccine [55].
Consequently, the novel proteins involved in membrane
surface (ribosomal protein P0) and PVM modification (ROP9,
GRA9) might be candidates for vaccine development.
Recently, several authors have suggested the importance
of proteome and genome integration data, since surprising
discrepancies between protein abundances and transcript
expression data have been shown in Toxoplasma [56]. Our
novel proteins differ from previously described ones in those
genomic approaches, but they are certainly involved in the
same processes. In this way, determining both absolute
protein expression and post-translational events will be a key
factor in gaining a more complete understanding of the
biology of these pathogenic organisms.
In conclusion, the present results have evidenced for the
first time in Neospora the previously reported developmental
strategies in Toxoplasma during stage conversion. This study
opens the gate to further studies on tachyzoite–bradyzoite
conversion mechanisms and sets the basis for further studies
in potential targets for drug and vaccine development.
We thank Louis M. Weiss (Albert Einstein College of Medi-cine, New York) for the monoclonal anti-recombinant TgBAG1serum, Andrew Hemphill (Institute for Parasitology, Universityof Berne, Switzerland) for providing the polyclonal rabbit anti-recombinant NcSAG1 serum, and Diana Williams (LiverpoolSchool of Tropical Medicine, Liverpool, UK) for the N. caninumNc-Liv isolate. We also thank Aida Pitarch and MonserratMartınez (Pharmacy Faculty, Complutense University ofMadrid, Spain) for carefully reading the manuscript. Theproteomics work was done at the Proteomics Facility UCM-PCM, a member of ProteoRed network, funded by GenomaEspana. VMH was supported by a fellowship from the Ministryof Science and Innovation of Spain. This work was financiallysupported by AGL 2007–60132/GAN.
These authors have declared no conflict of interest.
5 References
[1] Bjerkas, I., Dubey, J. P., Evidence that Neospora caninum is
identical to the Toxoplasma-like parasite of Norwegian
dogs. Acta Vet. Scand. 1991, 32, 407–410.
[2] Buxton, D., McAllister, M. M., Dubey, J. P., The comparative
pathogenesis of neosporosis. Trends Parasitol. 2002, 18,
546–552.
[3] Innes, E. A., Wright, S. E., Maley, S., Rae, A. et al., Protection
against vertical transmission in bovine neosporosis. Int. J.
Parasitol. 2001, 31, 1523–1534.
[4] Lyons, R. E., McLeod, R., Roberts, C. W., Toxoplasma gondii
tachyzoite-bradyzoite interconversion. Trends Parasitol.
2002, 18, 198–201.
[5] Di Cristina, M., Del Porto, P., Buffolano, W., Beghetto, E.
et al., The Toxoplasma gondii bradyzoite antigens
BAG1 and MAG1 induce early humoral and cell-
mediated immune responses upon human infection.
Microbes Infect. 2004, 6, 164–171.
[6] Aguado-Martinez, A., Ortega-Mora, L. M., Alvarez-Garcia,
G., Rodriguez-Marco, S. et al., Stage-specific expression of
Nc SAG4 as a marker of chronic Neospora caninum infec-
tion in a mouse model. Parasitology 2009, 136, 757–764.
[7] Aguado-Martinez, A., Alvarez-Garcia, G., Fernandez-Garcia,
A., Risco-Castillo, V. et al., Usefulness of rNcGRA7- and
rNcSAG4-based ELISA tests for distinguishing primo-
infection, recrudescence, and chronic bovine neosporosis.
Vet. Parasitol. 2008, 157, 182–195.
[8] Fernandez-Garcia, A., Risco-Castillo, V., Zaballos, A.,
Alvarez-Garcia, G., Ortega-Mora, L. M., Identification and
molecular cloning of the Neospora caninum SAG4 gene
specifically expressed at bradyzoite stage. Mol. Biochem.
Parasitol. 2006, 146, 89–97.
[9] Risco-Castillo, V., Fernandez-Garcia, A., Zaballos, A.,
Aguado-Martinez, A. et al., Molecular characterisation of
BSR4, a novel bradyzoite-specific gene from Neospora
caninum. Int. J. Parasitol. 2007, 37, 887–896.
[10] Hemphill, A., Fuchs, N., Sonda, S., Gottstein, B., Hentrich,
B., Identification and partial characterization of a 36 kDa
surface protein on Neospora caninum tachyzoites. Para-
sitology 1997, 115, 371–380.
[11] Howe, D. K., Crawford, A. C., Lindsay, D., Sibley, L. D., The
p29 and p35 immunodominant antigens of Neospora cani-
num tachyzoites are homologous to the family of surface
antigens of Toxoplasma gondii. Infect. Immun. 1998, 66,
5322–5328.
[12] Fuchs, N., Sonda, S., Gottstein, B., Hemphill, A., Differential
expression of cell surface- and dense granule-associated
Neospora caninum proteins in tachyzoites and bradyzoites.
J. Parasitol. 1998, 84, 753–758.
[13] Shin, Y. S., Shin, G. W., Kim, Y. R., Lee, E. Y. et al.,
Comparison of proteome and antigenic proteome between
two Neospora caninum isolates. Vet. Parasitol. 2005, 134,
41–52.
[14] Lee, E. G., Kim, J. H., Shin, Y. S., Shin, G. W. et al., Appli-
cation of proteomics for comparison of proteome of
Neospora caninum and Toxoplasma gondii tachyzoites.
1748 V. Marugan-Hernandez et al. Proteomics 2010, 10, 1740–1750
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

J. Chromatogr. B Analyt. Technol. Biomed. Life. Sci. 2005,
815, 305–314.
[15] Risco-Castillo, V., Fernandez-Garcia, A., Ortega-Mora, L. M.,
Comparative analysis of stress agents in a simplified in vitro
system of Neospora caninum bradyzoite production.
J. Parasitol. 2004, 90, 466–470.
[16] Alvarez-Garcia, G., Pereira-Bueno, J., Gomez-Bautista, M.,
Ortega-Mora, L. M., Pattern of recognition of Neospora
caninum tachyzoite antigens by naturally infected
pregnant cattle and aborted foetuses. Vet. Parasitol. 2002,
107, 15–27.
[17] Bohne, W., Gross, U., Ferguson, D. J., Heesemann, J.,
Cloning and characterization of a bradyzoite-specifically
expressed gene (hsp30/bag1) of Toxoplasma gondii, related
to genes encoding small heat-shock proteins of plants. Mol.
Microbiol. 1995, 16, 1221–1230.
[18] Parmley, S. F., Weiss, L. M., Yang, S., Cloning of a brady-
zoite-specific gene of Toxoplasma gondii encoding a cyto-
plasmic antigen. Mol. Biochem. Parasitol. 1995, 73,
253–257.
[19] Alban, A., David, S. O., Bjorkesten, L., Andersson, C. et al.,
A novel experimental design for comparative two-dimen-
sional gel analysis: two-dimensional difference gel electro-
phoresis incorporating a pooled internal standard.
Proteomics 2003, 3, 36–44.
[20] Gorg, A., Obermaier, C., Boguth, G., Harder, A. et al., The
current state of two-dimensional electrophoresis with
immobilized pH gradients. Electrophoresis 2000, 21,
1037–1053.
[21] Fodor, I. K., Nelson, D. O., Alegria-Hartman, M., Robbins, K.
et al., Statistical challenges in the analysis of two-dimen-
sional difference gel electrophoresis experiments using
DeCyder. Bioinformatics 2005, 21, 3733–3740.
[22] Sechi, S., Chait, B. T., Modification of cysteine residues by
alkylation. A tool in peptide mapping and protein identifi-
cation. Anal. Chem. 1998, 70, 5150–5158.
[23] Innes, E. A., The host-parasite relationship in pregnant
cattle infected with Neospora caninum. Parasitology 2007,
134, 1903–1910.
[24] Marouga, R., David, S., Hawkins, E., The development of the
DIGE system: 2D fluorescence difference gel analysis tech-
nology. Anal. Bioanal. Chem. 2005, 382, 669–678.
[25] Lee, E. G., Kim, J. H., Shin, Y. S., Shin, G. W. et al., Estab-
lishment of a two-dimensional electrophoresis map for
Neospora caninum tachyzoites by proteomics. Proteomics
2003, 3, 2339-2350.
[26] Lefevre, T., Thomas, F., Schwartz, A., Levashina, E. et al.,
Malaria Plasmodium agent induces alteration in the head
proteome of their Anopheles mosquito host. Proteomics
2007, 7, 1908–1915.
[27] Nelson, M. M., Jones, A. R., Carmen, J. C., Sinai, A. P. et al.,
Modulation of the host cell proteome by the intracellular
apicomplexan parasite Toxoplasma gondii. Infect. Immun.
2008, 76, 828–844.
[28] Cleary, M. D., Singh, U., Blader, I. J., Brewer, J. L., Boot-
hroyd, J. C., Toxoplasma gondii asexual development:
identification of developmentally regulated genes and
distinct patterns of gene expression. Eukaryot. Cell 2002, 1,
329–340.
[29] Radke, J. R., Behnke, M. S., Mackey, A. J., Radke, J. B. et al.,
The transcriptome of Toxoplasma gondii. BMC Biol. 2005, 3,
26.
[30] Boyle, J. P., Saeij, J. P., Harada, S. Y., Ajioka, J. W., Boot-
hroyd, J. C., Expression quantitative trait locus mapping of
Toxoplasma genes reveals multiple mechanisms for strain-
specific differences in gene expression. Eukaryot. Cell. 2008,
7, 1403–1414.
[31] McGuire, A. M., McAllister, M. M., Jolley, W. R., Anderson-
Sprecher, R. C., A protocol for the production of
Neospora caninum tissue cysts in mice. J. Parasitol. 1997,
83, 647–651.
[32] Vonlaufen, N., Guetg, N., Naguleswaran, A., Muller, N.
et al., In vitro induction of Neospora caninum bradyzoites in
vero cells reveals differential antigen expression, localiza-
tion, and host-cell recognition of tachyzoites and brady-
zoites. Infect. Immun. 2004, 72, 576–583.
[33] Bohne, W., Heesemann, J., Gross, U., Reduced replication
of Toxoplasma gondii is necessary for induction of brady-
zoite-specific antigens: a possible role for nitric oxide in
triggering stage conversion. Infect. Immun. 1994, 62,
1761–1767.
[34] Tomavo, S., Boothroyd, J. C., Interconnection between
organellar functions, development and drug resistance in
the protozoan parasite, Toxoplasma gondii. Int. J. Parasitol.
1995, 25, 1293–1299.
[35] Denton, H., Roberts, C. W., Alexander, J., Thong, K. W.,
Coombs, G. H., Enzymes of energy metabolism in the
bradyzoites and tachyzoites of Toxoplasma gondii. FEMS
Microbiol. Lett. 1996, 137, 103–108.
[36] Tomavo, S., The differential expression of multiple isoen-
zyme forms during stage conversion of Toxoplasma gondii:
an adaptive developmental strategy. Int. J. Parasitol. 2001,
31, 1023–1031.
[37] Weiss, L. M., Ma, Y. F., Takvorian, P. M., Tanowitz, H. B.,
Wittner, M., Bradyzoite development in Toxoplasma gondii
and the hsp70 stress response. Infect. Immun. 1998, 66,
3295–3302.
[38] Echeverria, P. C., Matrajt, M., Harb, O. S., Zappia, M. P.
et al., Toxoplasma gondii Hsp90 is a potential drug target
whose expression and subcellular localization are devel-
opmentally regulated. J. Mol. Biol. 2005, 350, 723–734.
[39] Yahiaoui, B., Dzierszinski, F., Bernigaud, A., Slomianny, C.
et al., Isolation and characterization of a subtractive library
enriched for developmentally regulated transcripts
expressed during encystation of Toxoplasma gondii. Mol.
Biochem. Parasitol. 1999, 99, 223–235.
[40] Manger, I. D., Hehl, A., Parmley, S., Sibley, L. D. et al.,
Expressed sequence tag analysis of the bradyzoite stage of
Toxoplasma gondii: identification of developmentally
regulated genes. Infect. Immun. 1998, 66, 1632–1637.
[41] Boothroyd, J. C., Hehl, A., Knoll, L. J., Manger, I. D., The
surface of Toxoplasma: more and less. Int. J. Parasitol.
1998, 28, 3–9.
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
Proteomics 2010, 10, 1740–1750 1749

[42] Zhang, H., Lee, E. G., Liao, M., Compaore, M. K. et al.,
Identification of ribosomal phosphoprotein P0 of Neospora
caninum as a potential common vaccine candidate for the
control of both neosporosis and toxoplasmosis. Mol.
Biochem. Parasitol. 2007, 153, 141–148.
[43] Carruthers, V. B., Sibley, L. D., Sequential protein secretion
from three distinct organelles of Toxoplasma gondii
accompanies invasion of human fibroblasts. Eur. J. Cell
Biol. 1997, 73, 114–123.
[44] Reichmann, G., Dlugonska, H., Fischer, H. G., Characteriza-
tion of TgROP9 (p36), a novel rhoptry protein of Toxo-
plasma gondii tachyzoites identified by T cell clone. Mol.
Biochem. Parasitol. 2002, 119, 43–54.
[45] Adjogble, K. D., Mercier, C., Dubremetz, J. F., Hucke, C.
et al., GRA9, a new Toxoplasma gondii dense granule
protein associated with the intravacuolar network of tubular
membranes. Int. J. Parasitol. 2004, 34, 1255–1264.
[46] Halonen, S. K., Weidner, E., Overcoating of Toxoplasma
parasitophorous vacuoles with host cell vimentin type inter-
mediate filaments. J. Eukaryot. Microbiol. 1994, 41, 65–71.
[47] Warren, A. J., Colledge, W. H., Carlton, M. B., Evans, M. J.
et al., The oncogenic cysteine-rich LIM domain protein rbtn2 is
essential for erythroid development. Cell 1994, 78, 45–57.
[48] Osteryoung, K. W., Sundberg, H., Vierling, E., Poly(A) tail
length of a heat shock protein RNA is increased by severe
heat stress, but intron splicing is unaffected. Mol. Gen.
Genet. 1993, 239, 323–333.
[49] Foth, B. J., Goedecke, M. C., Soldati, D., New insights into
myosin evolution and classification. Proc. Natl. Acad. Sci.
USA 2006, 103, 3681–3686.
[50] Gupta, N., Zahn, M. M., Coppens, I., Joiner, K. A., Voelker,
D. R., Selective disruption of phosphatidylcholine metabo-
lism of the intracellular parasite Toxoplasma gondii arrests
its growth. J. Biol. Chem. 2005, 280, 16345–16353.
[51] Dzierszinski, F., Popescu, O., Toursel, C., Slomianny, C.
et al., The protozoan parasite Toxoplasma gondii expresses
two functional plant-like glycolytic enzymes. Implications
for evolutionary origin of apicomplexans. J. Biol. Chem.
1999, 274, 24888–24895.
[52] Vial, H., Ancelin, M. L., Research for new antimalarial
molecules: an emergency, a hope. Pathol. Biol. (Paris) 1994,
42, 138–144.
[53] Berry, C., Humphreys, M. J., Matharu, P., Granger, R. et al.,
A distinct member of the aspartic proteinase gene family
from the human malaria parasite Plasmodium falciparum.
FEBS Lett. 1999, 447, 149–154.
[54] Malik, M., Nitiss, J. L., DNA repair functions that control
sensitivity to topoisomerase-targeting drugs. Eukaryot. Cell.
2004, 3, 82–90.
[55] Innes, E. A., Andrianarivo, A. G., Bjorkman, C., Williams,
D. J., Conrad, P. A., Immune responses to Neospora cani-
num and prospects for vaccination. Trends Parasitol. 2002,
18, 497–504.
[56] Xia, D., Sanderson, S. J., Jones, A. R., Prieto, J. H. et al., The
proteome of Toxoplasma gondii: integration with the
genome provides novel insights into gene expression and
annotation. Genome Biol. 2008, 9, R116.
1750 V. Marugan-Hernandez et al. Proteomics 2010, 10, 1740–1750
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com












