
Identification and characterization of visual pigments in caecilians (Amphibia: Gymnophiona), an...
7
3586 INTRODUCTION The Gymnophiona, commonly known as caecilians, are one of the three orders of extant Amphibia. They are readily distinguished from frogs (Anura) and salamanders (Urodela) by their annulated, limbless bodies, and are distinct in many other characters, including rudimentary eyes (e.g. Taylor, 1968; Wilkinson and Nussbaum, 2006). Reduction in the visual system of the Gymnophiona, associated with a mainly fossorial lifestyle, has long been recognized (see Norris and Hughes, 1918; Walls, 1942) but the extent to which the caecilian eye is functional remains uncertain (Wake, 1980; Wake, 1985; Himstedt, 1995; Himstedt, 1996). Adult caecilian eyes are relatively small, covered with skin and sometimes bone, lack features expected of well-developed vertebrate eyes and have several features apparently co-opted to novel sensory structures and functions (e.g. Himstedt and Simon, 1995). Relaxed molecular clock analysis suggests that the last common ancestor of extant caecilians occurred in the Triassic or Jurassic (Roelants et al., 2007; San Mauro, 2010) and that caecilians may therefore have possessed a reduced visual system for over 150 million years. It is widely assumed that variations in the peak spectral sensitivities (l max ) of the visual system represent adaptations to specific visual needs associated with particular habitats or lifestyles (Lythgoe, 1979; Peichl, 2005; Davies et al., 2009). Spectral sensitivity is determined either by changes in the tertiary structure of the opsin protein via amino acid substitutions and/or by the use of alternate chromophores derived from vitamin A1 or A2 (Crescitelli et al., 1972; Yokoyama, 2000; Bowmaker and Hunt, 2006; Hart and Hunt, 2007). Rods, which generally express the rh1 opsin gene and rod-specific components of the phototransduction cascade, are generally responsible for scotopic or dim light vision. By contrast, up to four classes of cone opsin genes exist in vertebrates that are associated with colour and photopic vision (Hunt et al., 2001a; Bowmaker and Hunt, 2006). The encoded proteins of these genes have l max values (based on a vitamin A1 chromophore) between about 360–440 nm (‘ultraviolet’ or ‘violet’, short wavelength-sensitive; the sws1 gene) 440–470 nm (‘blue’, short wavelength-sensitive; the sws2 gene), 470–520 nm (‘green’, middle wavelength-sensitive; the rh2 gene) and 500–570 nm (‘red’, long wavelength-sensitive; the lws gene). Electroretinography has detected a single class of visual pigment in the caecilian Ichthyophis cf. kohtaoensis (Himstedt, 1995), and this was presumed to be a rod pigment based on its l max value at 502 nm. Consistent with this, fragments of rh1 coding sequence have been isolated from four species of morphologically and phylogenetically diverse caecilians (Frost et al., 2006; Venkatesh et al., 2001), suggesting that this class of visual pigment may be present throughout the Gymnophiona. Immunohistochemistry has also been performed on a similarly diverse group of caecilians using antibodies raised to rod opsin, transducin, and cone opsin (Nguyen, 2003). Strong reactions were observed with anti-rod antibodies and weaker reactions with one anti-cone antibody, although the latter reaction is most probably the result of cross-reactivity with the rod The Journal of Experimental Biology 213, 3586-3592 © 2010. Published by The Company of Biologists Ltd doi:10.1242/jeb.045914 Identification and characterization of visual pigments in caecilians (Amphibia: Gymnophiona), an order of limbless vertebrates with rudimentary eyes S. M. Mohun 1,2 , W. L. Davies 1,3 , J. K. Bowmaker 1 , D. Pisani 4 , W. Himstedt 5 , D. J. Gower 2 , D. M. Hunt 1,6 and M. Wilkinson 2, * 1 UCL Institute of Ophthalmology, 11-43 Bath Street, London EC1V 9EL, UK, 2 Department of Zoology, Natural History Museum, London, Cromwell Road, London SW7 5BD, UK, 3 Nuffield Laboratory of Ophthalmology, University of Oxford, Levels 5-6, West Wing, John Radcliffe Hospital, Headley Way, Oxford OX3 9DU, UK, 4 Department of Biology, National University of Ireland Maynooth, County Kildare, Ireland, 5 Department of Biology, Darmstadt University of Technology, Schnittspahnstrasse 3-5, D-64287, Germany and 6 School of Animal Biology, University of Western Australia, 35 Stirling Highway, Perth, WA 6009, Australia *Author for correspondence ([email protected]) Accepted 13 July 2010 SUMMARY In comparison with the other amphibian orders, the Anura (frogs) and Urodela (salamanders), knowledge of the visual system of the snake-like Gymnophiona (caecilians) is relatively sparse. Most caecilians are fossorial with, as far as is known any surface activity occurring mainly at night. They have relatively small, poorly developed eyes and might be expected to possess detectable changes in the spectral sensitivity of their visual pigments. Microspectrophotometry was used to determine the spectral sensitivities of the photoreceptors in three species of caecilian, Rhinatrema bivittatum, Geotrypetes seraphini and Typhlonectes natans. Only rod opsin visual pigment, which may be associated with scotopic (dim light) vision when accompanied by other ‘rod- specific’ components of the phototransduction cascade, was found to be present. Opsin sequences were obtained from the eyes of two species of caecilian, Ichthyophis cf. kohtaoensis and T. natans. These rod opsins were regenerated in vitro with 11-cis retinal to give pigments with spectral sensitivity peaks close to 500 nm. No evidence for cone photoreception, associated with diurnal and colour vision, was detected using molecular and physiological methods. Additionally, visual pigments are short- wavelength shifted in terms of the maximum absorption of light when compared with other amphibian lineages. Supplementary material available online at http://jeb.biologists.org/cgi/content/full/213/20/3586/DC1 Key words: opsin, visual pigments, evolution, spectral tuning, caecilian, Gymnophiona. THE JOURNAL OF EXPERIMENTAL BIOLOGY
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Identification and characterization of visual pigments in caecilians (Amphibia: Gymnophiona), an...

3586
INTRODUCTIONThe Gymnophiona, commonly known as caecilians, are one of thethree orders of extant Amphibia. They are readily distinguished fromfrogs (Anura) and salamanders (Urodela) by their annulated,limbless bodies, and are distinct in many other characters, includingrudimentary eyes (e.g. Taylor, 1968; Wilkinson and Nussbaum,2006). Reduction in the visual system of the Gymnophiona,associated with a mainly fossorial lifestyle, has long been recognized(see Norris and Hughes, 1918; Walls, 1942) but the extent to whichthe caecilian eye is functional remains uncertain (Wake, 1980; Wake,1985; Himstedt, 1995; Himstedt, 1996). Adult caecilian eyes arerelatively small, covered with skin and sometimes bone, lackfeatures expected of well-developed vertebrate eyes and haveseveral features apparently co-opted to novel sensory structures andfunctions (e.g. Himstedt and Simon, 1995). Relaxed molecular clockanalysis suggests that the last common ancestor of extant caeciliansoccurred in the Triassic or Jurassic (Roelants et al., 2007; San Mauro,2010) and that caecilians may therefore have possessed a reducedvisual system for over 150 million years.
It is widely assumed that variations in the peak spectralsensitivities (lmax) of the visual system represent adaptations tospecific visual needs associated with particular habitats or lifestyles(Lythgoe, 1979; Peichl, 2005; Davies et al., 2009). Spectralsensitivity is determined either by changes in the tertiary structureof the opsin protein via amino acid substitutions and/or by the useof alternate chromophores derived from vitamin A1 or A2
(Crescitelli et al., 1972; Yokoyama, 2000; Bowmaker and Hunt,2006; Hart and Hunt, 2007). Rods, which generally express the rh1opsin gene and rod-specific components of the phototransductioncascade, are generally responsible for scotopic or dim light vision.By contrast, up to four classes of cone opsin genes exist invertebrates that are associated with colour and photopic vision (Huntet al., 2001a; Bowmaker and Hunt, 2006). The encoded proteins ofthese genes have lmax values (based on a vitamin A1 chromophore)between about 360–440nm (‘ultraviolet’ or ‘violet’, shortwavelength-sensitive; the sws1 gene) 440–470nm (‘blue’, shortwavelength-sensitive; the sws2 gene), 470–520nm (‘green’,middle wavelength-sensitive; the rh2 gene) and 500–570nm (‘red’,long wavelength-sensitive; the lws gene).
Electroretinography has detected a single class of visual pigmentin the caecilian Ichthyophis cf. kohtaoensis (Himstedt, 1995), andthis was presumed to be a rod pigment based on its lmax value at502nm. Consistent with this, fragments of rh1 coding sequence havebeen isolated from four species of morphologically andphylogenetically diverse caecilians (Frost et al., 2006; Venkateshet al., 2001), suggesting that this class of visual pigment may bepresent throughout the Gymnophiona. Immunohistochemistry hasalso been performed on a similarly diverse group of caecilians usingantibodies raised to rod opsin, transducin, and cone opsin (Nguyen,2003). Strong reactions were observed with anti-rod antibodies andweaker reactions with one anti-cone antibody, although the latterreaction is most probably the result of cross-reactivity with the rod
The Journal of Experimental Biology 213, 3586-3592© 2010. Published by The Company of Biologists Ltddoi:10.1242/jeb.045914
Identification and characterization of visual pigments in caecilians (Amphibia:Gymnophiona), an order of limbless vertebrates with rudimentary eyes
S. M. Mohun1,2, W. L. Davies1,3, J. K. Bowmaker1, D. Pisani4, W. Himstedt5, D. J. Gower2, D. M. Hunt1,6
and M. Wilkinson2,*1UCL Institute of Ophthalmology, 11-43 Bath Street, London EC1V 9EL, UK, 2Department of Zoology, Natural History Museum,London, Cromwell Road, London SW7 5BD, UK, 3Nuffield Laboratory of Ophthalmology, University of Oxford, Levels 5-6, West
Wing, John Radcliffe Hospital, Headley Way, Oxford OX3 9DU, UK, 4Department of Biology, National University of IrelandMaynooth, County Kildare, Ireland, 5Department of Biology, Darmstadt University of Technology, Schnittspahnstrasse 3-5, D-64287,
Germany and 6School of Animal Biology, University of Western Australia, 35 Stirling Highway, Perth, WA 6009, Australia*Author for correspondence ([email protected])
Accepted 13 July 2010
SUMMARYIn comparison with the other amphibian orders, the Anura (frogs) and Urodela (salamanders), knowledge of the visual system ofthe snake-like Gymnophiona (caecilians) is relatively sparse. Most caecilians are fossorial with, as far as is known any surfaceactivity occurring mainly at night. They have relatively small, poorly developed eyes and might be expected to possess detectablechanges in the spectral sensitivity of their visual pigments. Microspectrophotometry was used to determine the spectralsensitivities of the photoreceptors in three species of caecilian, Rhinatrema bivittatum, Geotrypetes seraphini and Typhlonectesnatans. Only rod opsin visual pigment, which may be associated with scotopic (dim light) vision when accompanied by other ‘rod-specific’ components of the phototransduction cascade, was found to be present. Opsin sequences were obtained from the eyesof two species of caecilian, Ichthyophis cf. kohtaoensis and T. natans. These rod opsins were regenerated in vitro with 11-cisretinal to give pigments with spectral sensitivity peaks close to 500nm. No evidence for cone photoreception, associated withdiurnal and colour vision, was detected using molecular and physiological methods. Additionally, visual pigments are short-wavelength shifted in terms of the maximum absorption of light when compared with other amphibian lineages.
Supplementary material available online at http://jeb.biologists.org/cgi/content/full/213/20/3586/DC1
Key words: opsin, visual pigments, evolution, spectral tuning, caecilian, Gymnophiona.
THE JOURNAL OF EXPERIMENTAL BIOLOGY

3587Visual pigments in caecilians
pigment (Nguyen, 2003; Foster et al., 1993). An anti-bovinetransducin antibody also reacted positively, suggesting that the rodphototransduction pathway is intact in the retina of caecilians. Thus,a functional scotopic visual pathway would appear to be present incaecilians, even in species with rudimentary eyes concealed underbone, without any evidence for the presence of photopic vision.
The aim of this study was to identify and characterize the visualpigments expressed in the retina within ecologically andphylogenetically diverse caecilian species and to determine the lmax
of such pigments using in vivo and in vitro methods.
MATERIALS AND METHODSTissue sources
Adult animals were used for all experiments as follows: onespecimen of Rhinatrema bivittatum (Guérin-Méneville 1829)(voucher number MW 2390) was used for microspectrophotometry(MSP); six specimens of Typhlonectes natans (Fischer 1880) (MW7346–MW 7351), with one specimen being used for MSP and fivespecimens for mRNA extraction; one specimen of Geotrypetesseraphini (Duméril 1859) (MW 3854) was used for MSP; twospecimens of Ichthyophis cf. kohtaoensis (Taylor, 1960) (MW 4764and MW 4765) for mRNA extraction. All caecilian species wereobtained by fieldwork or from licensed commercial sources asapproved by the local Animal Ethics Committee and were killedusing an approved procedure. All vouchers will be accessioned intothe collections of the Natural History Museum, London.
MicrospectrophotometryPhysiological data were determined using MSP. A modifiedLiebman dual beam microspectrophotometer under computer controlwas used to determine the spectral sensitivities of the photoreceptors.With the help of an infrared converter, the measuring beam(normally 2mm square cross section) was aligned to pass transverselythrough the photoreceptor outer segments or transparent pieces ofskin covering the eye, while the reference beam passed through aclear space adjacent to the object examined. Spectra were scannedfrom 750 to 350nm in 2nm increments and back from 251 to 749nmat the interleaved wavelengths. Only one absorption spectrum wasusually obtained from a given outer segment to minimize the effectsof bleaching. To verify the presence of a photolabile pigment,putative outer segments were bleached by exposure to white lightfrom the monochromator passing through measuring beams of themicrospectrophotometer. A standardized computer program wasused to estimate the lmax for each outer segment. The spectraobtained from the outer segments were averaged to obtain a meancurve. The curve was then fitted to a standard template curve inorder to obtain an estimate of lmax (Govardovskii et al., 2000). Thestandard template curve used was the Dartnall standard curve forrhodopsin placed with its lmax at 502nm and expressed on anabscissal scale of log frequency.
Messenger RNA extraction and complementary DNAsynthesis
Complementary DNA (cDNA) was generated from mRNA extractedfrom the eyes of two species of caecilian, I. cf. kohtaoensis and T.natans, using three and ten eyes, respectively. The QuickPrepTM
micro mRNA purification kit (GE Healthcare, Little Chalfont, UK)was used to purify polyadenylated mRNA. First strand cDNA wassynthesized in a reaction using 500ng of oligo(dT) or for 3�-RACEprimer from the 5�-/3�-RACE Kit (Roche, Burgess Hill, UK) and1–2mg of mRNA, 1i.u. Superscript III reverse transcriptase(Invitrogen, Paisley, UK), following the manufacturer’s instructions.
Polymerase chain reactionFragments of opsin coding sequence were amplified from cDNAusing degenerate primers designed from an alignment of vertebrateopsins (Davies et al., 2007a; Davies et al., 2007b). Polymerase chainreaction (PCR) amplifications using genomic DNA from the samecaecilian species and amphibian cone class-specific and degenerateprimers were also performed (supplementary material TableS1). Anested protocol was used to detect sequences from exon 1 to exon4. From the sequences obtained using standard nested PCR, specificprimers (supplementary material TableS1) were designed for 5�-and 3�-RACE to isolate the untranslated regions (UTRs) of the rodopsin mRNA transcript. The reaction mixture typically contained10mmoll–1 of each dNTP, 2mmoll–1 MgCl2, 0.1mmoll–1 of eachforward and reverse primer, 2.5i.u. Biotaq DNA polymerase(Bioline, London, UK) and 20–100 ng of cDNA. Cycling conditionswere, an initial denaturing step at 94°C for 5min, then 40 cycles ofdenaturing at 94°C for 30s, annealing at 55°C for 1min andextension at 72°C for 1.5min, with a final extension at 72°C for10min. Negative controls were run in parallel containing the samereagents but without the addition of cDNA. A second roundamplification used 1/10 dilution of the first round PCR products,under identical conditions to the first round except for an annealingtemperature of 60°C.
The 5� and 3� ends of the visual pigment cDNA were amplifiedusing the 5�/3�-RACE kit (Roche), following the manufacturer’sinstructions. PCR products were cloned into pGEM T-Easy vector(Promega, Southampton, UK) prior to sequencing using either T7or SP6 primers and the Big Dye Terminator v3.1 Cycle Sequencingkit on an ABI 3730 sequencer (Applied Biosystems, Warrington,UK).
In vitro regeneration of visual pigmentsThe full-length coding sequences of T. natans and I. cf. kohtaoensisrh1 were amplified using a proofreading KOD XL Taq polymerase(Novagen, Nottingham, UK) (supplementary material Fig. S1). Theprimer pairs were designed for the 5� and 3� ends with restrictionsites for EcoRI and SalI. The digested PCR products weredirectionally cloned into a derivative of the mammalian expressionvector pMT4 carrying the sequence of bovine rod opsin 1D4 epitope(including the stop codon) at the 3� end of the coding sequence(Franke et al., 1988). HEK293T cells were transfected using acalcium phosphate precipitation technique with vector containingthe opsin insert. Cells were harvested after 48h and washed with1� phosphate-buffered saline (PBS) buffer. Pigments werereconstituted by resuspending the harvested cells in PBS buffer andincubating with 20mmoll–1 11-cis retinal in the dark. The pigmentswere isolated by incubating with 1% (w/v) n-dodecyl-b-D-maltoside(DDM) and 20mgml–1 phenylmethylsulphonylfluoride (PMSF)before passage over a cyanogen-bromide-activated (CNBr)Sepharose binding column coupled to an anti-1D4 monoclonalantibody.
The dark and the bleached (photobleached with broad spectrumwhite light for at least 30min) absorption spectra were measuredusing a dual path spectrophotometer (Spectronic Unicam,Cambridge, UK). The lmax was determined from the differencespectra (dark minus light bleached) by fitting a Govardovskii A1template (Govardovskii et al., 2000) to the data using an Excelprogram.
Phylogenetic analysisAn alignment of coding region amino acids was done manually inMacClade 4.08 (Maddison and Maddison, 2000), using the
THE JOURNAL OF EXPERIMENTAL BIOLOGY

3588 S. M. Mohun and others
translated caecilian sequences and 68 vertebrate visual opsinsequences downloaded from the NCBI database (alignment andGenBank accession numbers are given in supplementary materialFig.S1), representing the five classes of visual opsins and includingall amphibian representatives (typically up to three where availableand including any with rudimentary eyes) of major vertebratelineages. Bayesian inferences used Markov Chain Monte Carlo(MCMC) implemented in MRBAYES 3.1.2 (Huelsenbeck andRonquist, 2001; Ronquist and Huelsenbeck, 2003) sampling every1000 generations from two parallel runs (each of four chains)of 1,000,000 generations. Convergence was established bycomparing the standard deviations of split frequencies and burninsdetermined by visual inspection of likelihoods using TRACER 1.4(Rambaut and Drummond, 2007) (Tracer v1.4, Available fromhttp://beast.bio.ed. ac.uk/Tracer). Model jumping was used withinMRBAYES 3.1.2 to estimate the fixed-rate model for amino aciddata during the tree-search, which chose the BLOSUM62 (BLOckSUbstitution Matrix) model of amino acid substitution that wasproduced from multiple alignments of evolutionarily divergentproteins (Henikoff and Henikoff, 1992). The analysis was rootedusing LWS opsin protein sequences as outgroups. The sequenceswere identified using BLASTP 2.2.20 and BLASTN (Altschul etal., 1997) against published opsin sequences from the NCBIdatabase.
RESULTSMicrospectrophotometry
Three species from ecologically diverse and phylogeneticallydistinct caecilian lineages were investigated. Rhinatrema bivittatum(a species that retains many ancestral features) and Geotrypetesseraphini, are terrestrial as adults, whereas Typhlonectes natans isaquatic. All have relatively well-developed eyes for caecilians, butthose of T. natans and G. seraphini are possibly secondarily welldeveloped. In all three species, only a single class of rodphotoreceptors was found, and microspectrophotometry (MSP)demonstrated the presence in each species of a single visual pigmentwith similar lmax values of 488.5±0.9nm for T. natans (N31),487.5±1.0nm for R. bivittatum (N13), and 486.6±2.2nm for G.seraphini (N8; Fig.1).
The light transmission of the skin immediately above the eye ofT. natans was determined by MSP. Fig.2 shows that this skin isalmost transparent with only a small amount of absorbanceattenuation of about 10% at wavelengths <500nm.
Opsin sequence analysis and phylogeneticsOf the three species used for the MSP analysis, only eye tissue samplesfrom T. natans were available for molecular studies. A fourth species,I. cf. kohtaoensis, was therefore added to the study at this stage. Opsinsequences were PCR amplified from cDNA; in all cases, the sequenceof the amplified fragments corresponded to the rh1 gene. This wasconfirmed by phylogenetic analysis, with both caecilian sequencesfalling, as expected, within the amphibian rh1 group as a sister groupof Batrachia (Fig.3). The aligned sequences used are shown insupplementary material Fig.S1. No cone opsin sequences wereobtained by PCR amplifications from genomic DNA or cDNA usingeither gene specific or degenerate primers.
The full rh1 coding and deduced amino acid sequences for T.natans and I. cf. kohtaoensis are shown in supplementary materialFigsS1 and S2. The sequences show an 80–82% nucleotide anda 87–89% amino acid identity for T. natans and a 79–81%nucleotide and a 86–90% amino acid identity for I. cf. kohtaoensisto the rh1 orthologues of other amphibian species. There is a 96%
amino acid identity between T. natans and I. cf. kohtaoensissequences.
Expression of visual pigmentsIn order to confirm that the cloned gene sequences encoded thepigments identified in situ in photoreceptors by MSP, the T. natanspigment was generated in vitro by isolation of recombinant opsin
400 500 600
Abs
orba
nce
Wavelength (nm)
Dark
Bleached
E
0
0.01
0.02
0.03
400 500 600
486.6 nmF
0
0.01
–0.01
0.02
0.03
Dark
Bleached
A
0.01
0
0.02
0.03
0.04 488.5 nmB
0
0.01
–0.01
0.02
0.03
Dark
Bleached
C
0
0.02
0.06
0.04
487.5 nmD
0
0.02
0.04
Fig.1. MSP absorbance spectra for three caecilian species, Typhlonectesnatans (A,B), Rhinatrema bivittatum (C, D) and Geotrypetes seraphini(E,F). (A,C,E) Dark and bleached spectra, (B,D,F) difference spectra. Foreach species, the absorbance peak shifts to around 380nm afterbleaching. This represents the oxime (stabilized form of the bleachedphotopigment chromophore) of the liberated retinal. The fitted spectra toboth the dark and difference spectra are shown as the solid lines.
100
75
50
25
400 500 600 700
% T
rans
mis
sion
Wavelength (nm)
Fig.2. MSP transmission spectrum through the skin covering the eye ofTyphlonectes natans.
THE JOURNAL OF EXPERIMENTAL BIOLOGY

3589Visual pigments in caecilians
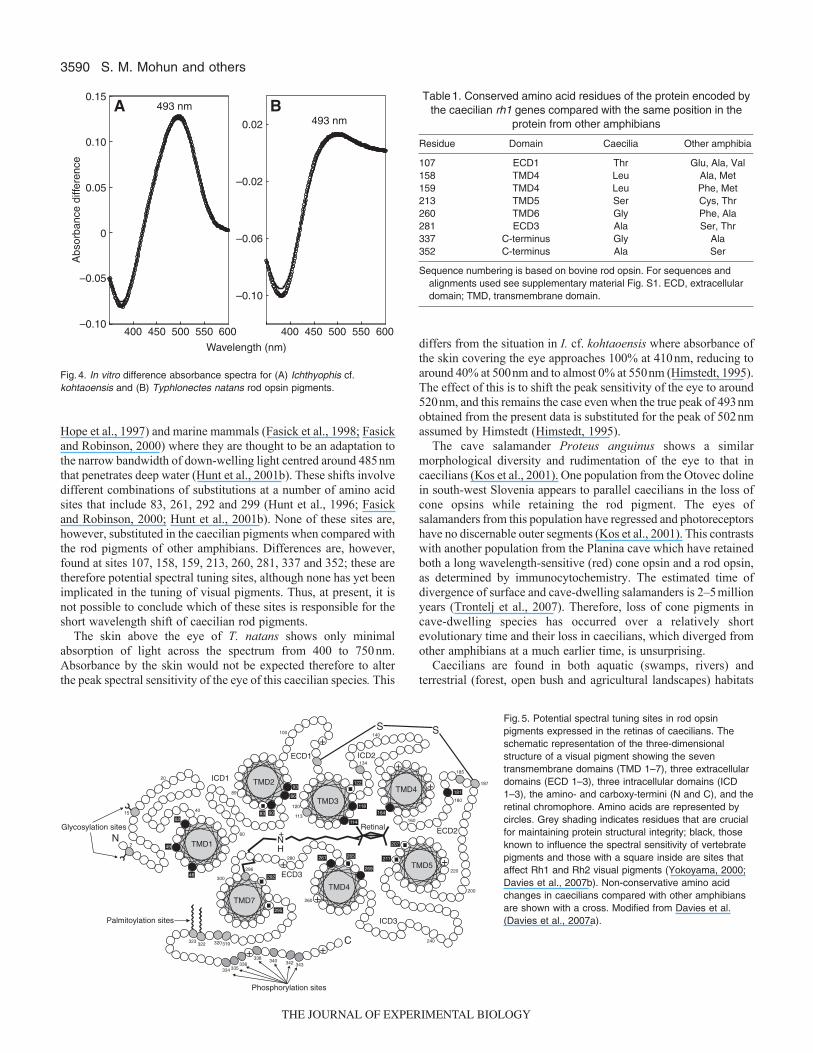
from mammalian cells transfected with an expression vectorcontaining the opsin coding sequence. After pigment reconstitutionby the addition of 11-cis-retinal, the dark and bleached absorptionspectra were measured by conventional UV–visiblespectrophotometry. The difference spectrum yielded a lmax at493nm (Fig.4), which is sufficiently similar to the photoreceptorpeak at 488.5±0.9nm obtained in situ by MSP to confirm the identityof the photoreceptor pigment as rod opsin. The opsin codingsequence of I. cf. kohtaoensis was also expressed and regeneratedin vitro to give an identical lmax to that of T. natans at 493nm.
Compared with the rod pigments of other Amphibia, which peakaround 502–506nm (Partridge et al., 1992; Chen et al., 1996;Fyhrquist et al., 1998; Ala-Laurila et al., 2002), the caecilianpigments measured in situ are short wavelength-shifted by between13 and 19nm. This shift must be the result of particular amino aciddifferences in the caecilian rh1 opsin pigments compared with otheramphibian rh1 opsins. Key amino acid sites 83, 122, 207, 211, 265,292, 295, which are known to be important in the spectral tuningof rh1 pigments (Davies et al., 2007a) are invariant across all theamphibian pigments (Fig.5). However, the two caecilian pigmentsshow consistent differences from other amphibians at sites 107, 158,159, 213, 260, 289, 337 and 352 (Table1) which may influence thespectral tuning of caecilian visual pigments.
DISCUSSIONOnly a single spectral class of photoreceptor was found in threecaecilian species, Rhinatrema bivittatum, Typhlonectes natans andGeotrypetes seraphini, with mean absorbance peaks between487–489nm. Consistent with this, the only expressed opsin detectedin the eye of two caecilian species, T. natans and I. cf. kohtaoensis,belongs to the rod or Rh1 class of opsins that is expressed in rodphotoreceptors throughout the vertebrate kingdom. No other classesof photoreceptors or expressed opsins were found and it is concludedthat these caecilian species possess a rod-only retina, which agreeswith previous research (Wake, 1985; Himstedt, 1995). Because coneopsins homologous with those of other vertebrates are found inmembers of the Urodela (salamanders) and Anura (frogs) (Rohlichand Szel, 2000; Takahashi et al., 2001; Sakikabara et al., 2002), itwould appear that the loss of cone opsins, and cone photoreceptors,is a synapomorphy of Gymnophiona.
The peak sensitivities of rod visual pigments in Amphibiatypically lie between 502–506nm (Liebman and Entine, 1968;Harosi, 1975; Partridge et al., 1992; Chen et al., 1996; Fyhrquist etal., 1998; Ala-Laurila et al., 2002); peak sensitivities of caecilianrod pigments are therefore short wavelength-shifted by between 13and 19nm. These shifts are comparable to those seen in a numberof deep-water vertebrates (Hunt et al., 2001b; Bowmaker et al., 1994;
Fig.3. Phylogenetic results of Bayesian analyses ofvertebrate amino acid sequence analysis using Blosum62.Numbers are posterior probabilities. Branches withoutnumbers are maximally supported and branches withposterior probability support values below 0.6 werecollapsed. See legend to supplementary material Fig.S2for opsin sequence accession numbers.
THE JOURNAL OF EXPERIMENTAL BIOLOGY
3590
Hope et al., 1997) and marine mammals (Fasick et al., 1998; Fasickand Robinson, 2000) where they are thought to be an adaptation tothe narrow bandwidth of down-welling light centred around 485nmthat penetrates deep water (Hunt et al., 2001b). These shifts involvedifferent combinations of substitutions at a number of amino acidsites that include 83, 261, 292 and 299 (Hunt et al., 1996; Fasickand Robinson, 2000; Hunt et al., 2001b). None of these sites are,however, substituted in the caecilian pigments when compared withthe rod pigments of other amphibians. Differences are, however,found at sites 107, 158, 159, 213, 260, 281, 337 and 352; these aretherefore potential spectral tuning sites, although none has yet beenimplicated in the tuning of visual pigments. Thus, at present, it isnot possible to conclude which of these sites is responsible for theshort wavelength shift of caecilian rod pigments.
The skin above the eye of T. natans shows only minimalabsorption of light across the spectrum from 400 to 750nm.Absorbance by the skin would not be expected therefore to alterthe peak spectral sensitivity of the eye of this caecilian species. This
S. M. Mohun and others
differs from the situation in I. cf. kohtaoensis where absorbance ofthe skin covering the eye approaches 100% at 410nm, reducing toaround 40% at 500nm and to almost 0% at 550nm (Himstedt, 1995).The effect of this is to shift the peak sensitivity of the eye to around520nm, and this remains the case even when the true peak of 493nmobtained from the present data is substituted for the peak of 502nmassumed by Himstedt (Himstedt, 1995).
The cave salamander Proteus anguinus shows a similarmorphological diversity and rudimentation of the eye to that incaecilians (Kos et al., 2001). One population from the Otovec dolinein south-west Slovenia appears to parallel caecilians in the loss ofcone opsins while retaining the rod pigment. The eyes ofsalamanders from this population have regressed and photoreceptorshave no discernable outer segments (Kos et al., 2001). This contrastswith another population from the Planina cave which have retainedboth a long wavelength-sensitive (red) cone opsin and a rod opsin,as determined by immunocytochemistry. The estimated time ofdivergence of surface and cave-dwelling salamanders is 2–5millionyears (Trontelj et al., 2007). Therefore, loss of cone pigments incave-dwelling species has occurred over a relatively shortevolutionary time and their loss in caecilians, which diverged fromother amphibians at a much earlier time, is unsurprising.
Caecilians are found in both aquatic (swamps, rivers) andterrestrial (forest, open bush and agricultural landscapes) habitats
400 450 500 550 600
–0.10
–0.06
–0.02
0.02
0
0.05
0.10
–0.05
–0.10
0.15
400 450 500 550600
493 nm493 nm
A B
Abs
orba
nce
diffe
renc
e
Wavelength (nm)
Fig.4. In vitro difference absorbance spectra for (A) Ichthyophis cf.kohtaoensis and (B) Typhlonectes natans rod opsin pigments.
334
340C
N
S S
NH
+
ECD1
ICD1
Palmitoylation sites
RetinalGlycosylation sites
ICD2
ICD3
ECD2
ECD3
TMD1
TMD2
TMD3
TMD4
T 4MD
TMD5
T 7MD
+
+
+
+
+
+
+
+335
336338 340 342 343
320322323 319
295
292
296
300
260
280
113
261 265
269
211
207
220
200
240
187
185
180
160
181
164
140
134
118
114
122
120
100
93
86
9083
89
40
60
20
15
2
52
49
46
Phosphorylation sites
Fig.5. Potential spectral tuning sites in rod opsinpigments expressed in the retinas of caecilians. Theschematic representation of the three-dimensionalstructure of a visual pigment showing the seventransmembrane domains (TMD 1–7), three extracellulardomains (ECD 1–3), three intracellular domains (ICD1–3), the amino- and carboxy-termini (N and C), and theretinal chromophore. Amino acids are represented bycircles. Grey shading indicates residues that are crucialfor maintaining protein structural integrity; black, thoseknown to influence the spectral sensitivity of vertebratepigments and those with a square inside are sites thataffect Rh1 and Rh2 visual pigments (Yokoyama, 2000;Davies et al., 2007b). Non-conservative amino acidchanges in caecilians compared with other amphibiansare shown with a cross. Modified from Davies et al.(Davies et al., 2007a).
Table1. Conserved amino acid residues of the protein encoded bythe caecilian rh1 genes compared with the same position in the
protein from other amphibians
Residue Domain Caecilia Other amphibia
107 ECD1 Thr Glu, Ala, Val158 TMD4 Leu Ala, Met159 TMD4 Leu Phe, Met213 TMD5 Ser Cys, Thr260 TMD6 Gly Phe, Ala281 ECD3 Ala Ser, Thr337 C-terminus Gly Ala352 C-terminus Ala Ser
Sequence numbering is based on bovine rod opsin. For sequences andalignments used see supplementary material Fig. S1. ECD, extracellulardomain; TMD, transmembrane domain.
THE JOURNAL OF EXPERIMENTAL BIOLOGY

3591Visual pigments in caecilians
in the wet tropics (e.g. Gower et al., 2004; Jones et al., 2006).Terrestrial species live mostly in soil but may also exploit epigeicmicrohabitats such as under leaf litter and rotting vegetation (e.g.Burger et al., 2007). Caecilians (at least those species that arepartially surface active) are mostly considered nocturnal (Himstedt,1995; Kupfer et al., 2004; Burger et al., 2007), so it is likely thatthe peak absorbance of caecilian rod photoreceptor pigments arespectrally tuned to maximize the absorbance of light and thewavelengths available in either a forest floor, soil or nightenvironment. Tropical forests are heterogeneous in the spectralcomposition of ambient light (Endler, 1993), where the light variesfrom white in large gaps between vegetation to yellowish-red insmall gaps. In forest shade, essentially all of the light has beentransmitted through or reflected from leaves to give a range ofwavelengths of 470–490 nm (Endler, 1993), similar therefore to themaximum absorbance of caecilian rod visual pigments. By contrast,at night the wavelengths of light are long wavelength-shifted, butthere is no evidence for a shift towards longer wavelengths in anynocturnal animals (Peichl, 2005), including caecilians. The light attwilight is predominately around 480nm, which is similar to thelmax of caecilian pigments, and after the sun sinks below the horizon,the loss of middle wavelengths becomes more pronounced (Endler,1993). Therefore, it is possible that the main role of the rod visualpigment in caecilians is to detect twilight, a critical period duringwhich diurnal and nocturnal species change behaviour patterns andlocations and when prey detection and predator avoidance becomeshighly significant (Munz and Mcfarland, 1977).
ACKNOWLEDGEMENTSWe are grateful to Dr Rosalie Crouch for the generous gift of 11-cis retinal. Wethank Alex Kupfer (Universität Jena), Jeannot and Odette (Camp Patawa), GuyTiego (Direction regionale de l’environment Guyane), Céline Dupuy (Direction desServices, Vétérinaires de la Guyane), and especially Philippe Gaucher (CentreNational de la Recherche Scientifique) for facilitating our collection of Rhinatremabivittatum in French Guiana. We would also like to thank Dr Livia Carvalho, DrSusan Wilkie and Dr Jill Cowing for technical assistence with the in vitroregeneration of caecilian visual pigments. This work was supported by a UKBiotechnology and Biological Sciences Research Council (BBSRC) studentship,awards from The Museum Research Fund and from the Zoology Research Fundof the Natural History Museum.
REFERENCESAla-Laurila, P., Saarinen, P., Albert, R., Koskelainen, A. and Donner. K. (2002).
Temperature effects on spectral properties of red and green rods in toad retina. Vis.Neurosci. 19, 781-792.
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W. andLipman, D. J. (1997). Gapped BLAST and PSI-BLAST: a new generation of proteindatabase search programs. Nucleic Acids Res. 25, 3389-3402.
Bowmaker, J. K. and Hunt, D. M. (2006). Evolution of vertebrate visual pigmentsCurr. Biol. 16, R484-R489.
Bowmaker, J. K., Govardovskii, V. I., Shukolyukov, S. A., Zueva, L. V., Hunt, D.M., Sideleva, V. G. and Smirnova, O. G. (1994). Visual pigments and the photicenvironment: the cottoid fish of Lake Baikal. Vis. Res. 34, 591-605.
Burger, R. M., Boylan, J. and Aucone, B. M. (2007). The effects of phototaxis andthigmotaxis on microhabitat selection by a caecilian amphibian (genus Ichthyophis)Herpetol. J. 17, 19-23.
Chen, N., Ma, J. X., Corson, D. W., Hazard, E. S. and Crouch, R. K. (1996).Molecular cloning of a rhodopsin gene from salamander rods. Invest. Ophthalmol.Vis. Sci. 37, 1907-1913.
Crescitelli, F. (1972). The visual cells and visual pigments of the vertebrate eye. InPhotochemistry of Vision: Handbook of Sensory Physiology (ed. H. J. A. Dartnall),pp. 245-363. Berlin: Springer.
Davies, W. L., Cowing, J. A., Carvalho, L. S., Potter, I. C., Tresize, A. E. O., Hunt,D. M. and Collin, S. P. (2007a). Functional characterization tuning and regulation ofvisual pigment gene expression in an anadromous lamprey. FASEB J. 21, 2713-2724.
Davies, W. L., Carvalho, L. S., Cowing, J. A., Beazley, L. D., Hunt, D. M. andArrese, C. A. (2007b). Visual pigments of the platypus: a novel route to mammaliancolour vision. Curr. Biol. 17, 161-162.
Davies, W. L., Carvalho, L. S., Tay, B., Brenner, S., Hunt, D. M. and Venkatesh, B.(2009). Into the blue: gene duplication and loss underlie color vision adaptations in adeep-sea chimaera, the elephant shark Callorhinchus milii. Gen. Res. 19, 415-426.
Endler, J. A. (1993). The color of light in forests and its implications. Ecol. Monogr. 63,1-27.
Fasick, J. I. and Robinson, P. R. (2000). Spectral-tuning mechanisms of marinemammal rhodopsins and correlations with foraging depth. Vis. Neurosci. 17, 781-788.
Fasick, J. I., Cronin, T. W., Hunt, D. M. and Robinson, P. R. (1998). The visualpigments of the bottlenose dolphin (Tursiops truncatus). Vis. Neurosci. 15, 643-651.
Foster, R. G., Garcia Fernandez, J. M., Provencio, I. and Degrip, W. J. (1993).Opsin localisation and chromophore retinoids identified within the basal brain of thelizard Anolis carolinensis. J. Comp. Physiol. A 172, 33-45.
Franke, R. R., Sakmar, T. P., Oprian, D. D. and Khorana, H. G. (1988). A singleamino acid substitution in rhodopsin (Lys248Leu) prevents activation of transducin.J. Biol. Chem. 263, 2119-2122.
Frost, D. R., Grant, T., Faivovich, J., Bain, R. H., Haas, A., Haddad, C. F. B., DeSa, R. O., Channing, A., Wilkinson, M., Donnellan, S. C. et al. (2006). Theamphibian tree of life. Bull. Am. Mus. Nat. Hist. 297, 1-370.
Fyhrquist, N., Donner, K., Hargrave, P. A., Mcdowell, J. H., Popp, M. P. andSmith, W. C. (1998). Rhodopsins from three frog and toad species: sequences andfunctional comparisons. Exp. Eye Res. 66, 295-305.
Govardovskii, V. I., Fyhrquist, N., Reuter, T., Kuzmin, D. G. and Donner, K. (2000).In search of the visual pigment template. Vis. Neurosci. 17, 509-528.
Gower, D. J., Loader, S. P., Wilkinson, M. and Moncreiff, C. B. (2004). Nicheseparation and comparative abundance of Boulengerula boulengeri andScolecomorphus vittatus (Amphibia: Gymnophiona) in East Usambara forest,Tanzania. Afr. J. Herpetol. 53, 183-190.
Harosi, F. (1975). Absorption spectra and linear dichroism of some amphibianphotoreceptors J. Gen. Physiol. 66, 357-382.
Hart, N. S. and Hunt, D. M. (2007). Avian visual pigments: characteristics, spectraltuning, and evolution. Am. Nat. 169, S7-S26.
Henikoff, S. and Henikoff, J. G. (1992). Amino acid substitution matrices from proteinblocks. Proc. Nalt. Acad. Sci. 89, 10915-10919.
Himstedt, W. (1995). Structure and function of the eyes in the caecilian Ichthyophiskohtaoensis (Amphibia, Gymnophiona) Zoo. Anal. Comp. Syst. 99, 81-94.
Himstedt, W. (1996). Die Blindwuhlen. Neue Brehm-Bucherei, Vol. 630.Magdeburg/Heidelberg: Westarp/Spektrum.
Himstedt, W. and Simon, D. (1995). Sensory basis of foraging behaviour in caecilians(Amphibia, Gymnophiona) Herp. J. 5, 266-270.
Hope, A. J., Partridge, J. C., Dulai, K. S. and Hunt, D. M. (1997). Mechanisms ofwavelength tuning in the rod opsins of deep-sea fishes Proc. R. Soc. Lond. B. Biol.Sci. 264, 155-163.
Huelsenbeck, J. P. and Ronquist, F. (2001). MRBAYES: Bayesian inference ofphylogeny. Bioinformatics 17, 754-755.
Hunt, D. M., Fitzgibbon, J., Slobodyanyuk, S. J. and Bowmaker, J. K. (1996).Spectral tuning and molecular evolution of rod visual pigments in the species flock ofcottoid fish in Lake Baikal. Vision Res. 36, 1217-1224.
Hunt, D. M., Wilkie, S. E., Bowmaker, J. K. and Poopalasundaram, S. (2001a).Vision in the ultraviolet Cell. Mol. Life Sci. 58, 1583-1598.
Hunt, D. M., Dulai, K. S., Partridge, J. C., Cottrill, P. and Bowmaker, J. K. (2001b).The molecular basis for spectral tuning of rod visual pigments in deep-sea fish. J.Exp. Biol. 204, 3333-3344.
Jones, D. T., Loader, S. P. and Gower, D. J. (2006). Trophic ecology of East Africancaecilians (Amphibia: Gymnophiona), and their impact on forest soil invertebrates J.Zool. 269, 117-126.
Kos, M., Bulog, B., Szel, A. and Rohlich, P. (2001). Immunocytochemicaldemonstration of visual pigments in the degenerate retinal and pinealphotoreceptors of the blind cave salamander (Proteus anguinus). Cell Tissue Res.303, 15-25.
Kupfer, A., Nabhitabhata, J. and Himstedt, W. (2004). Reproductive ecology offemale caecilian amphibians (genus Ichthyophis): a baseline study. Biol. J. Linn.Soc. Lond. 83, 207-217.
Liebman, P. A. and Entine, G. (1968). Visual pigments of frog and tadpole (Ranapipiens). Vis. Res. 8, 761-775.
Lythgoe, J. N. (1979). The Ecology of Vision. Oxford: Clarendon.Maddison, D. R. and Maddison, W. P. (2000). Macclade Version 4, Analysis of
Phylogeny and Character Evolution. Sunderland, MA: Sinauer Associates.Munz, F. W. and McFarland, W. N. (1977). Evolutionary adaptations of fishes to the
photic environment. In The Visual System of Vertebrates (ed. F. Crescitelli), pp. 194-274. New York: Springer-Verlag.
Nguyen, B. (2003). Sind Blindwuehlen blind? Histologische und immunhistochemischeUntersuchungen an den Augen von Gymnophionen. PhD thesis University ofTechnology, Darmstadt, Germany.
Norris, H. P. and Hughes, S. P. (1918). The cranial and anterior spinal nerves of thecaecilian amphibians. J. Morphol. 31, 490-557.
Partridge, J. C., Speare, P., Shand, J., Muntz, W. R. and Williams, D. M. (1992).Microspectrophotometric determinations of rod visual pigments in some adult andlarval Australian amphibians. Vis. Neurosci. 9, 137-142.
Peichl, L. (2005). Diversity of mammalian photoreceptor properties: adaptations tohabitat and lifestyle? Anat. Rec. A Discov.Mol.Cell. Evol. Biol. 287A, 1001-1012.
Roelants, K., Gower, D. J., Wilkinson, M., Loader, S. P., Biju, S. D., Guillaume, K.,Moriau, L. and Bossuyt, F. (2007). Global patterns of diversification in the history ofmodern amphibians. Proc. Natl. Acad. Sci. USA 104, 887-892.
Rohlich, P. and Szel, A. (2000). Photoreceptor cells in the Xenopus retina. Microsc.Res. Tech. 50, 327-337.
Ronquist, F. and Huelsenbeck, J. P. (2003). MRBAYES 3, Bayesian phylogeneticinference under mixed models. Bioinformatics 19, 1572-1574.
Sakakibara, S., Hiramatsu, H., Takahashi, Y., Hisatomi, O., Kobayashi, Y., Sakami,S., Saito, T. and Tokunaga, F. (2002). Opsin expression in adult, developing, andregenerating newt retinas Mol. Brain Res. 103, 28-35.
San Mauro, D. (2010). A multilocus timescale for the origin of extant amphibians.Mol.Phylogenet. Evol. 56, 554-561.
THE JOURNAL OF EXPERIMENTAL BIOLOGY

3592 S. M. Mohun and others
Takahashi, Y., Hisatomi, O., Sakakibara, S., Tsukahara, Y., Ebrey, T. G. andTokunaga, F. (2001). Molecular cloning of the blue-sensitive opsin expressed in theJapanese common newt (Cynops pyrrhogaster). Invest. Ophthalmol. Vis. Sci. 42, 984.
Taylor, E. H. (1968). The Caecilians of the World: a Taxonomic Analysis. Lawrence,KS: University of Kansas Press.
Trontelj, P., Goricki, S., Polak, S., Verovnik, R., ZakSek, V. and Sket, B. (2007).Age estimates for some subterranean taxa and lineages in the Dinaric Karst. ActaCarsologica 36, 183-189.
Venkatesh, B., Erdmann, M. V. and Brenner, S. (2001). Molecular synapomorphiesresolve evolutionary relationships of extant jawed vertebrates. Proc. Natl. Acad. Sci,USA 98, 11382-11387.
Wake, M. H. (1980). Morphological information on caecilian eye function. Am. Zool. 20,785.
Wake, M. H. (1985). The comparative morphology and evolution of the eyes ofcaecilians (Amphibia; Gymnophiona). Zoomorphology 105, 277-295.
Walls, G. (1942). The Vertebrate Eye and its Adaptive Radiation. Bloomfield Hills, MI:Cranbrook Institute of Science, Bull 19.
Wilkinson, M. and Nussbaum, R. A. (2006). Caecilian phylogeny and classification.In Reproductive Biology and Phylogeny of Gymnophiona (Caecilians) (ed. Exbrayet),pp. 39-78. Enfield: Science Publishers.
Yokoyama, S. (2000). Molecular evolution of vertebrate visual pigments. Prog. Retin.Eye. Res. 19, 385-419.
THE JOURNAL OF EXPERIMENTAL BIOLOGY




















