
I.3 - The Evidence for Evolution
12
I.3 The Evidence for Evolution Gregory C. Mayer OUTLINE 1. The fossil record 2. Comparative biology 3. Biogeography 4. Evolution in action 5. Evolution as fact and theory The evidence for evolution was first comprehensively as- sembled by Charles Darwin, who succeeded in convincing essentially all his scientific contemporaries of the fact of descent with modification. A signal factor in Darwin’s achievement was that he was able to weave together nu- merous strands of natural history—paleontology, sys- tematics, embryology, morphology, biogeography—into a coherent framework. Since Darwin, genetics has joined this synthesis, and, in a development that might have surprised Darwin, evolution in natural populations has proven to occur sufficiently rapidly that it can be observed on human timescales. The most direct evidence of evo- lution comes from the fossil record, in which the dynamic changes of life over time are recorded, including many transitions between major taxa. A host of phenomena in comparative biology (e.g., systematics, morphology, em- bryology, genomics) and biogeography that otherwise appear inexplicable or anomalous are readily explained under the hypothesis of descent with modification. In addition, direct observation of natural and artificial pop- ulations shows the process of evolutionary change in ac- tion. Together, these sources of evidence lead to a “con- silience of inductions” that makes the fact of evolution one of the most securely established generalizations in science. GLOSSARY Adaptation. A feature of an organism that fits it to its con- ditions of existence, giving rise to similarity among organisms leading the same or similar ways of life. Homology. The correspondence, determined by their rel- ative positions and connections, of organs in different organisms, which is indicative of affinity; the cause of this correspondence is inheritance from a common ancestor. Oceanic Island. An island that has never been connected to a mainland and thus has received its fauna and flora over water by occasional means of transport. Phylogenetic Tree. A representation of the history of life, with branching indicating the splitting of lineages, and the connection of the branches indicating the pas- sage of genetic information and materials from one generation to the next. Progression. The pattern in the fossil record in which earlier forms of life differ from later forms, with major groups first appearing in the record in a generalized form and later as more diversified members of the same group. Some of the earlier forms may become extinct, and later forms may more closely resemble modern forms. Not to be confused with progress, a different concept, according to which evolution pro- ceeds toward some externally defined goal. Speciation. The splitting of a lineage into two or more daughter lineages reproductively isolated and evo- lutionarily independent from other lineages. Tetrapods. The group of four-limbed vertebrates com- prising amphibians, reptiles, mammals, and birds; includes species that have secondarily lost their limbs, such as snakes. Unity of Type. Similarities among organisms leading differ- ent ways of life under diverse conditions of existence, going beyond any functional need for similarity. The evidence for evolution, it has been remarked, is not the result of some crucial experiment but something more like the contents of the American Museum of Natural History. And so it is: the evidence for evolution comes from a plethora of biological and geological subdisciplines—systematics, paleontology, stratigraphy,
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of I.3 - The Evidence for Evolution

I.3The Evidence for EvolutionGregory C. Mayer
OUTLINE
1. The fossil record2. Comparative biology3. Biogeography4. Evolution in action5. Evolution as fact and theory
The evidence for evolution was first comprehensively as-sembledbyCharlesDarwin,whosucceeded inconvincingessentially all his scientific contemporaries of the fact ofdescent with modification. A signal factor in Darwin’sachievement was that he was able to weave together nu-merous strands of natural history—paleontology, sys-tematics, embryology, morphology, biogeography—intoa coherent framework. Since Darwin, genetics has joinedthis synthesis, and, in a development that might havesurprised Darwin, evolution in natural populations hasproven tooccur sufficiently rapidly that it canbe observedon human timescales. The most direct evidence of evo-lution comes from the fossil record, in which the dynamicchanges of life over time are recorded, including manytransitions between major taxa. A host of phenomena incomparative biology (e.g., systematics, morphology, em-bryology, genomics) and biogeography that otherwiseappear inexplicable or anomalous are readily explainedunder the hypothesis of descent with modification. Inaddition, direct observation of natural and artificial pop-ulations shows the process of evolutionary change in ac-tion. Together, these sources of evidence lead to a “con-silience of inductions” that makes the fact of evolutionone of the most securely established generalizations inscience.
GLOSSARY
Adaptation. Afeature of anorganism thatfits it to its con-ditions of existence, giving rise to similarity amongorganisms leading the same or similar ways of life.
Homology. The correspondence, determined by their rel-ative positions and connections, of organs in differentorganisms, which is indicative of affinity; the cause ofthis correspondence is inheritance from a commonancestor.
Oceanic Island. An island that has never been connectedto a mainland and thus has received its fauna andflora over water by occasional means of transport.
Phylogenetic Tree. A representation of the history of life,with branching indicating the splitting of lineages,and the connection of the branches indicating the pas-sage of genetic information and materials from onegeneration to the next.
Progression. The pattern in the fossil record in whichearlier formsof life differ fromlater forms,withmajorgroups first appearing in the record in a generalizedform and later as more diversified members of thesame group. Some of the earlier forms may becomeextinct, and later forms may more closely resemblemodern forms. Not to be confused with progress, adifferent concept, according to which evolution pro-ceeds toward some externally defined goal.
Speciation. The splitting of a lineage into two or moredaughter lineages reproductively isolated and evo-lutionarily independent from other lineages.
Tetrapods. The group of four-limbed vertebrates com-prising amphibians, reptiles, mammals, and birds;includes species that have secondarily lost their limbs,such as snakes.
Unity of Type. Similaritiesamongorganisms leadingdiffer-ent ways of life under diverse conditions of existence,going beyond any functional need for similarity.
The evidence for evolution, it has been remarked, is notthe result of some crucial experiment but somethingmore like the contents of the American Museum ofNatural History. And so it is: the evidence for evolutioncomes from a plethora of biological and geologicalsubdisciplines—systematics, paleontology, stratigraphy,

geochronology, biogeography, morphology, botany, zo-ology, embryology, genetics—many of which find theirobjects of study in the vast and varied collections of nat-ural history museums.
It is the diversity of these sources of evidence, allleading to the conclusion that life on earth has under-gone a long history of descent with modification, that isthe great strength, and the most striking aspect, of theevidence for evolution. The varied sources of evidenceare all brought into the unified explanatory scheme ofevolution, forming what the philosopher WilliamWhewell (1794–1866) called a “consilience of induc-tions,” each piece of evidence reinforcing the whole.Charles Darwin (1809–1882) in On the Origin ofSpecies used precisely such a form of argumentation,marshaling the disparate facts of geology, systemat-ics, morphology, embryology, and biogeography tosupport his theory of descent with modification.
Progression, Unity of Type, and Adaptation
By the time Darwin began his scientific career, it wasalready well established that the earth was old and thatthe fossil record was progressive, that is, that earlierforms of life differed from later forms, that some of theearlier forms had become extinct, and that later formsmore closely resembled modern forms. It was also be-coming clear that in this progression not only were olderforms replaced by newer ones but later forms were insome way related to earlier ones. Thus, a major groupwould appear in the fossil record in a generalized formand would be succeeded by more diversified members ofthe same group.
In addition to progression, two other great, but un-explained, classes of phenomena occupied biologists atthis time: unity of type and adaptation. Unity of typerefers to the similarities among organisms living differ-ent ways of life, similarities that extend far beyond anyfunctional needs. The same basic skeletal plan of theforelimb—a humerus, then radius and ulna, then car-pals, then metacarpals, then phalanges—occurs in alltetrapods, even though the limbs might appear quitedifferent externally and be used for very different func-tions (figure 1). Such similarities extend to embryolog-ical features as well: all tetrapod embryos, for example,have four limb buds, even if the adults (e.g., whales,snakes) lack one or two sets of limbs.
Adaptation refers to those features of organisms thatsuit them to their conditions of existence (see chapterIII.1; for a nuanced discussion of terminology, seechapter II.6), which may be shared by organisms withsimilar ways of life. Thus, sharks and whales share flat-tened tail flukes and dorsal fins, both features being ofobvious functional importance for aquatic organisms.
Despite these adaptive similarities, sharks and whalesdiffer in many features, such as their respiratory, circu-latory, and reproductive systems, that mark them asbelonging to different major groups of organisms—fishand mammals, respectively.
Darwin provided in his theory of descent with mod-ification a unified explanation of the geological (progres-sion), structural (unity of type), and functional (adapta-tion) phenomena. Unity of type reflects inheritance offeatures from common ancestors, and the changes fromthe common ancestor are due to modification. The mostimportant means of modification, natural selection,leads to adaptation. And when played out over geolog-ical time, descent with modification leads to progres-sion. Once admitted, descent with modification andcommon ancestry would also account for the hier-archical relationships revealed by systematics and forthe distributions of organisms across the surface of theglobe. It was Darwin’s triumph to combine geological,morphological, embryological, systematic, and biogeo-graphic evidence into a single explanatory theory.
This chapter considers the classes of evidence ad-duced by Darwin to support his theory of descent withmodification, including examples from post-Darwiniandisciplines such as genetics, and adds as an additionalclass of evidence observations of evolution in action, aclass that was largely unavailable to Darwin.
1. THE FOSSIL RECORD
Progression
The earliest fossils are of simple (prokaryotic), single-celled, photosynthetic bacteria that lived in mats calledstromatolites 3.5 billion years ago (see chapter II.11).One and a half to 2 billion years ago, more complex,organelle-bearing (eukaryotic) but still single-celledforms appear, and then multicellular forms. The originsof eukaryotes and multicellularity are not well docu-mented in the record, and molecular data suggest thatthey may have occurred considerably earlier than therecord shows (see chapter II.12). In the last part of thePrecambrian, about 600million years ago, diverse formsofmarine invertebrates appear, and then in theCambrianmany more invertebrate groups arise, as well as the firstvertebrates (see chapter II.15).
The earliest vertebrates, such as the recently dis-covered Myllokunmingia and Haikouichthys, whichlook remarkably like previously hypothesized general-ized vertebrates, were soft-bodied jawless forms thatlived about 525 million years ago. Diverse jawless fisheswith mineralized hard tissues followed. The first jawedfishes appear in the Late Ordovician, about 445 millionyears ago. The bony fishes, or Osteichthyes, the most
Evidence for Evolution 29

diverse living group of jawed fishes, are first found inthe Late Silurian, about 420 million years ago. The firstfour-legged vertebrates (tetrapods) are the approxi-mately 365 million-year-old amphibians Acanthostegaand Ichthyostega from theDevonian (see chapter II.17).Reptiles, the first tetrapods to be independent of water(amniotes),make their appearance in thePennsylvanian,about315million years ago. In theMesozoic era, the lasttwo major tetrapod groups arise: the first mammals areknown from the Triassic/Jurassic boundary (about 200million years ago), and the first bird is from the LateJurassic, about 150 million years ago.
The fossil record of vertebrates, then, exemplifies thesequential origin and diversification of themajor groups
of organisms through geological time.Moreover, as onemoves toward the present, many taxa go extinct andothers dwindle in diversity, and the array of majorgroups comes to progressively resemble that of today.
Transitions
Darwin did not have closely spaced transitional formsthat could demonstrate evolutionary continuity betweenmajor groups; he attributed their absence to the im-perfections of the geological record. The record is indeedimperfect: only hard parts of organisms are readily fos-silized, only certain sedimentary environments are con-ducive to fossilization, fossils must survive erosion and
Bird
Pterosaur
Bat
Humpback whale
Penguin
Lizard
Seal
Wrist bonesand digits
RadiusUlna
Human arm
Humerus
Theropod dinosaur
Figure 1. Unity of type and adaptation, illustrated by the forelimbsof tetrapods. The pattern of one bone (humerus), two bones (radiusand ulna), many bones (wrist and digits) is present in all, even
though the size, shape, and relative proportions of the elementshave been modified for varied ways of life, including walking, flying,grasping, and swimming. (Copyright Kalliopi Monoyios.)
30 Introduction

metamorphism and, finally, must be exposed on thesurface and discovered. As a consequence, fewer than1 percent of all species that ever lived are represented inthe fossil record, and the record is spotty temporally,geographically, and taxonomically (see chapter II.9). Forexample, only about 30 localities have yielded importantfossil faunas of tetrapods from the 60million years of theCarboniferous, when the first major radiation of am-phibians occurred.
Despite the imperfections, the transitional forms thatDarwin hoped might be found soon started turning up.In 1861,Archaeopteryxwas discovered and became thefirst of the proverbial “missing links” to be found.Clothed in the feathers of a bird with broad wings, itnonetheless had the long bony tail, toothed jaws, andfree, clawed fingers of a reptile. T. H. Huxley (1825–1895) saw it clearly as intermediate between reptiles andbirds and suggested a relationship to dinosaurs, a sug-gestion now well documented. Extraordinarily well-preserved fossils show not only the close skeletal resem-blance of Archaeopteryx and early birds to theropoddinosaurs but also that feathers are not peculiar to birds;they were present on quite a few nonflying dinosaurs aswell.
A few years later, Richard Owen (1804–1892) andE. D. Cope (1840–1897) both recognized that certainancient reptiles (synapsids) bore an affinity to mammals.The originofmammals from the synapsids is nowknownin exquisite detail (figure 2). Reptiles have several bonesin the lower jaw, and one of the more posterior ones, thearticular, forms the jaw joint with the quadrate bone ofthe skull. Mammals have a single bone in the lower jaw,the dentary (also present in reptiles), which articulateswith the squamosal bone of the skull. A long series offossils, beginning with fully reptilian forms in the Penn-sylvanian, lead gradually to themammals at the Triassic/Jurassic boundary. The dentary enlarges, becoming thelargest bone of the lower jaw. The quadrate and articularbecome smaller. Eventually, the dentary contacts thesquamosal, leading to several forms, such as Probainog-nathus, that have a dual jaw joint—both the old reptilianone and the new mammalian one. The transition is sogradual that it becomes a matter of convention to decidewhich form is the first “mammal”: Morganucodon isoften so regarded, but it, too, has a dual jaw joint. In laterforms the quadrate and articular detach from the jawandbecome two of the three mammalian ear bones. Manyother features that change during this transition—forexample, the dentition becomes cusped, a secondarypalate forms, the ilium becomes rod shaped—are like-wise documented in the fossil record.
Another well-documented transition is that betweenthe lobe-finned osteolepiform fishes and tetrapods
(figure 3). Osteolepiformswere “typical” fish, with dor-sal and anal fins, rounded heads with short snouts, andtheir shoulder girdles connected to their heads. But theirpectoral and pelvic fins extended from the body in fleshylobes, andwithin the lobewas a skeleton that, starting atthe base (the end near the body), had a pattern of “one
d an
d
d
d
an
ar
q
ar
q
ar
an
an
q
q
an
ar
Early synapsid
Advanced synapsid
Morganucodon
Advanced mammal
Figure 2. Homology of jaw bones of reptiles and ear bones of mam‐mals. In early synapsids the lower jaw comprises the tooth-bearingdentary (d) and several postdentary bones, including the angular (an)and articular (ar). The latter articulates with the quadrate (q), form‐ing the reptilian jaw joint. In advanced synapsids the latter bonesare reduced, while the dentary enlarges. In the earliest mammals(Morganucodon) these bones are reduced further, and the dentarymakes contact with the upper jaw, forming the mammalian jaw joint.In advanced mammals the angular, articular, and quadrate detachfrom the jaw entirely, becoming the tympanic, malleus, and incusof the mammalian ear. (After D. Davis 1991, K. Kardong 2012, andR. Carroll 1988.)
Evidence for Evolution 31

bone, two bones, many bones.” This is the same patternas in tetrapods: theplan that exemplifies theunity of typeof the tetrapod forelimb extends to osteolepiform fishes.In Panderichthys, from about 380million years ago, thehead and body are flattened, the snout is elongated, andthe eyes are on top of the head; the anal and dorsal finshave been lost. In the remarkable Tiktaalik from 375million years ago, the shoulder girdle (equivalent to ourcollarbone and shoulder blades) has been freed from theskull—Tiktaalik had a neck; and there is a joint withinthe “many bones” of the forelimb—it also had a wrist.Tenmillion years laterwe have the first actual tetrapods,Acanthostega and Ichthyostega, which have legs withtoes but retain some of the gill-cover bones and thecaudal fin of their fish ancestors.
The origins of birds, mammals, and tetrapods rep-resent fairly large changes inmorphology andwayof life,and the intermediate forms bridge the differences be-tween these major taxa. The much smaller transitionsthat occur as one species evolves into another are alsodocumented in the fossil record, but continuous sedi-mentary deposition is necessary to record such fine-scaletemporal events. Such conditions are not common butoccur most often in the fossil record of shelled marine
planktonic protists, such as foraminifera and radiolar-ians, that can be recovered by extracting cores from theseabed. The shells of dead individuals rain downonto theocean floor, forming an essentially continuous sedimen-tary record. In these organisms, such as the foraminif-erans in the genera Globorotalia and Contusotruncana,and the radiolarian genusEucyrtidium, such fine-scalechanges can be observed, and in the latter even thesplit into two species from an original one has beenrecorded.
There are many other examples of transitional formsin the fossil record—such as ancestral whales and snakeswith hindlegs—and more are being discovered everyyear.
2. COMPARATIVE BIOLOGY
Unity of Type
While the fossil recordprovides direct evidenceof changeof life over time, comparisons among organisms, eitherliving or extinct, give evidence that the link between thedifferent forms at different times is genealogical. Primaryamong these comparisons are the observed similarities infundamental structure among organisms referred to as“unity of type.” These similarities, which extend frommorphology to development to the genome, are homol-ogies, that is, similarities due to inheritance from acommon ancestor (see chapter II.6).
Famous homologies are the “one bone, two bones,many bones” pattern of the tetrapod limb (figure 1), andthe jaw and ear bones of reptiles and mammals, bothmentioned previously (figure 2). In the case of the limbs,the strong similarity of the same bone among the varioustetrapods allows the homologous bones to be easily rec-ognized: for example, humerus, radius, ulna. The sim-ilarity in plan is not accounted for by the functionalrequirements of the limbs but by inheritance from acommon ancestor. In the jaw and ear, the homologies aretraced with the aid of fossil and embryological (see thesection Markers of History) evidence. The angular andarticular bones of the reptilian lower jaw are homologuesof the tympanic andmalleus bones of themammalian ear,while the quadrate of the reptilian upper jaw is the incusof the mammalian middle ear.
These patterns can be seen not only in the skeletonbut in the genome itself at the cellular level. The humangenome contains 23 pairs of chromosomes, whereas thegenome of great apes has 24 pairs. The difference of onepair can readily be accounted for: human chromosome 2is homologous to two ape chromosomes, which havebecome fused in the human lineage. The homology has
Acanthostega gunnari
Tiktaalik roseae
Eusthenopteron foordi
Figure 3. Transition from lobe-finned fish to tetrapods. The lobe-finned osteolepiform fish Eusthenopteron has the tetrapod-like “onebone, two bones, many bones” pattern in its fin. Tiktaalik, the “fish-apod,”has lost thedorsal andanalfins, thehead is freeof theshouldergirdle, the snout is elongatedwith the eyesdirectedupward, and thereis awrist joint in the forelimb. Acanthostega, one of thefirst tetrapods,has digits but retains the caudal fin of its fish ancestors. The bones ofthe left forelimb shown in gray are, from darkest to lightest, the hu-merus, the radius, and the ulna, respectively; more distal elementsare unshaded. (Copyright Kalliopi Monoyios.)
32 Introduction

been confirmedby thediscoveryof remnantsof the central(centromere) and end (telomere) portions of the apechromosomes within the human chromosome, showingthat the latter arose from what were originally twochromosomes.
At perhaps the most basic level, the near universalityof the genetic code is another homology that argues forthe common ancestry of all life. The genetic code as-sociates each amino acid with a codon of three nucleo-tides, which carries the information for the making ofproteins. But the identity of the three nucleotides is ar-bitrary, so the code’s universality cannot be due tofunctional constraint but arises as a legacy of the codeestablished in distant progenitors.
Common Ancestry
Although the common ancestry indicated by the geneticcode embraces all (or nearly all) living beings, homol-ogies do not generally have such a wide distribution.Rather,homologies characterize smaller groups, and thesegroups are nested within larger homology-characterizedgroups, which in turn are nested within yet larger suchgroups, and soon.This nestingof homologies results from
the branching history of life, represented by the phyloge-netic tree (see chapter II.1).When a lineage divides, givingrise to anewbranch in the tree of life, the characteristics ofthe splitting lineage are passed on to its descendants.Modifications may occur in a descendant lineage, whichwill in turn be passed on to its, and only its, descendants.Each homology is the origin of a new feature, sharedwithdescendants but notwith collateral relatives. These nestedsets of homologies, which are exactly what one wouldexpect from a process of descent with modification, arepowerful evidence for evolution.
For example, possession of a toepad, composed oflaterally expanded scales at the end of the digit, char-acterizes lizards of the genus Anolis (figure 4). Anolesalso have a hemipenis (oddly named, as this means thatthey have two penises, rather than half a penis, as theword might imply) as the male intromittent organ, butthis organ characterizes a larger group, the squamates(lizardsplus snakes),withinwhichanoles are nested.Thesquamates, in turn, are nested within a yet-larger groupcharacterized by the presence of an amniotic membranearound their embryos. The amniotes (reptiles, birds, andmammals) also have four legs, a trait shared with am-phibians as well, which together make up the tetrapods.
Tetrapods Amniotes
Hemipenis
Amniotic membrane
Four legs
Toepads
Squamates Anoles
Figure 4. Phylogenetic tree of anole lizards showing nestedhomologies. The common ancestor of all tetrapods possessed fourlegs, a trait that, with modifications, was passed on to all of itsdescendants. One of these descendants evolved the amnioticmembrane, thus nesting the amniotes (reptiles, birds, and mam-mals) within the tetrapods. Squamates (snakes and lizards), which
evolved hemipenes, in turn are nested within amniotes. And finally,anoles are characterized by the possession of expanded toepads.Anoles thus possess toepads, hemipenes, amniotic membranes,and four legs, each trait inherited from a successively more distantancestor, and each marking a more inclusive group. The nestedboxes in the figure indicate the named clades.
Evidence for Evolution 33

The tetrapods, also in turn, share features of the limbwith certainfishes, formingagainamore inclusivegroup.Continuing in this manner, the bony fishes, vertebrates,chordates, and deuterostomes constitute successivelylarger groups within which anoles are nested, and thisnesting is indicative of the history of common ancestry.
Eventually all, or almost all, life can be subsumedwithin the nested tree. For organisms as divergent asinsects and vertebrates it is hard to recognize morpho-logical homologies, but genetic data show that homol-ogies of gene and genome structure can be recognizedin, for example, the presence of homologous sets of de-velopmental regulatory genes (Hox genes) in both ar-thropods and vertebrates. While nestedness holds truefor eukaryotes, there is some question whether at thebase of the tree of life transfer of genetic material be-tween prokaryotes may be so common as to obscure orefface the nested pattern (see chapter II.11).
Markers of History
Among the most striking evidences of evolution are thefeatures of organisms that appear to reflect a constraint:organisms inherit from their ancestors a developmentalsystem, and evolutionary modifications must take thatinherited system as a starting point. Indeed, organ-ismal features give every indication of having arisen by“tinkering”with this inherited preexisting developmen-tal system, and make sense only as a result of descentwith modification.
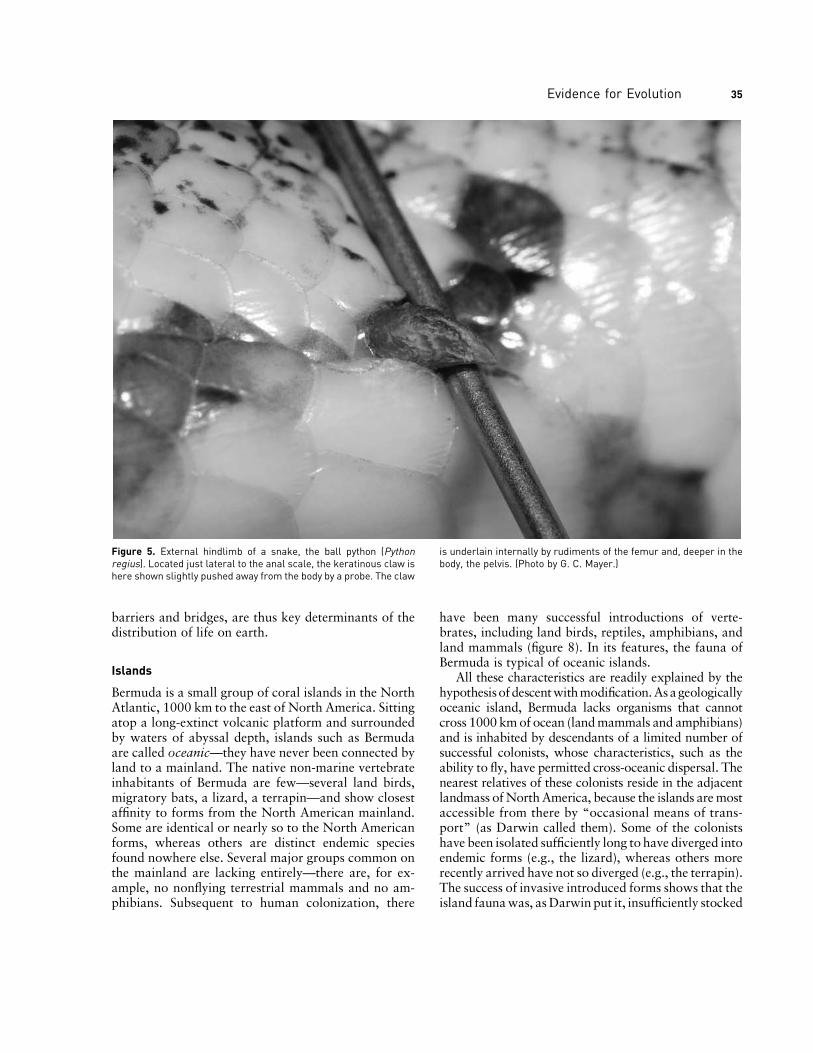
This constraint is evident in the similarities amongthe embryos of jawed vertebrates. The embryos gothrough a stage in which they resemble one another inthepossessionofpharyngeal arches, limbbuds, tails, andother traits. After this stage, embryos diverge, develop-ing into the varied forms they will become as adults.What perhaps is most striking is that structures presentin the embryo are not always present in the eventualadult. Thus, humans and apes have an embryonic tailthat largely disappears in the adult; only a bony vestigeremains internally. Humans develop a coat of fine fur,the lanugo,which is lost just before or shortly after birth.All four limb buds develop in tetrapods, such as whalesand snakes, that in the adult lack one or both pairs oflimbs. In some snakes, a vestigial leg is still visible ex-ternally (figure 5). In each case, the eventual adult formdevelops from a shared state and subsequently passesthrough stages shared with smaller nested groups untilfinally arriving at its own specific state.
Many structures show the traces of ancestry. In fetalmammals, the bones that eventually become the bonesof the middle ear begin their development in positions
along the jaw corresponding to those occupied by thehomologous jaw bones of reptiles. The “thumb” of pan-das (figure 6) is not homologous to othermammals’ innerdigits, nor is it even a digit: it is amodified radial sesamoidbone, pressed into service as a makeshift digit, in an ex-quisite example of tinkering—that is, the modification ofavailable structures, rather than an engineering ideal. Inthe Australian lungfish (Neoceratodus), there is a singledorsal lung, unlike in other lungfish (and tetrapods), inwhich the lungs are paired (figure 7). In all lungfish (andtetrapods), the lung attaches to the ventral part of theesophagus. In Australian lungfish, the pneumatic ducttravels up alongside the right side of the esophagus to thedorsally positioned lung, seeming to trace the course of itsmorphological modification. Confirming this movement,the left pulmonary artery, instead of going directly to thedorsal lung, travels down the left side of the esophagus,curls under the esophagus, and follows the pneumaticduct up to the lung.
Such examples are not limited to morphology: pseu-dogenes, nonfunctional versions of genes that are func-tional in related species or even in other copies in thesameorganism, occur commonly in organism’s genomes(see chapters V.3 and V.5). For example, in mostmammals, vitamin C is synthesized by a battery of en-zymes. In primates, which get vitamin C in their diet, agene for one of the necessary enzymes is present as anonfunctional pseudogene, so that the vitamin is notsynthesized. The broken gene is a relict from earliermammals that do use it in their synthesis of the vitamin.In primates, the gene has been disabled by a mutation,but the now-inactive gene remains as a marker of pri-mates’ forebears.
3. BIOGEOGRAPHY
Historically, the biogeographic evidence for evolutionwas crucial, because it was the evidence that convincedDarwin himself. Many features of the distribution oforganisms that are anomalous or merely curious undera hypothesis of special creation became explicable andexpected under the hypothesis of descent with mod-ification. Under the latter hypothesis, related speciesshould occur in geographically connected areas or inareas that could have been reached by a common an-cestor of the related species. The connectedness and“reachability” of areas are determined chiefly by thedistance and geographic barriers between them, andthe ability of organisms to cross such barriers. Geo-graphic conditions that either present barriers to dis-persal (e.g., the ocean around islands) or facilitate it(e.g., the continuous land of continents), and organ-isms’ abilities to overcome or utilize these geographic
34 Introduction
barriers and bridges, are thus key determinants of thedistribution of life on earth.
Islands
Bermuda is a small group of coral islands in the NorthAtlantic, 1000 km to the east of North America. Sittingatop a long-extinct volcanic platform and surroundedby waters of abyssal depth, islands such as Bermudaare called oceanic—they have never been connected byland to a mainland. The native non-marine vertebrateinhabitants of Bermuda are few—several land birds,migratory bats, a lizard, a terrapin—and show closestaffinity to forms from the North American mainland.Some are identical or nearly so to the North Americanforms, whereas others are distinct endemic speciesfound nowhere else. Several major groups common onthe mainland are lacking entirely—there are, for ex-ample, no nonflying terrestrial mammals and no am-phibians. Subsequent to human colonization, there
have been many successful introductions of verte-brates, including land birds, reptiles, amphibians, andland mammals (figure 8). In its features, the fauna ofBermuda is typical of oceanic islands.
All these characteristics are readily explained by thehypothesisofdescentwithmodification.Asageologicallyoceanic island, Bermuda lacks organisms that cannotcross 1000 kmof ocean (landmammals and amphibians)and is inhabited by descendants of a limited number ofsuccessful colonists, whose characteristics, such as theability to fly, have permitted cross-oceanic dispersal. Thenearest relatives of these colonists reside in the adjacentlandmass ofNorthAmerica, because the islands aremostaccessible from there by “occasional means of trans-port” (as Darwin called them). Some of the colonistshave been isolated sufficiently long to have diverged intoendemic forms (e.g., the lizard), whereas others morerecently arrived have not so diverged (e.g., the terrapin).The success of invasive introduced forms shows that theisland faunawas, asDarwin put it, insufficiently stocked
Figure 5. External hindlimb of a snake, the ball python (Pythonregius). Located just lateral to the anal scale, the keratinous claw ishere shown slightly pushed away from the body by a probe. The claw
is underlain internally by rudiments of the femur and, deeper in thebody, the pelvis. (Photo by G. C. Mayer.)
Evidence for Evolution 35

by the “creative force”: it was difficulties of dispersal,not habitat suitability, that caused the fauna to be re-stricted in species richness and taxonomic diversity.
Continents
Oncontinents, the accessibilityof adjacent areas leads, ingeneral, to related forms being found throughout. Dar-win noted this for the mammals of South America. Thecurrent mammals of the varied South American habitats—tropical, temperate, alpine—are all related to one an-other and not to mammals of similar habitats in distantplaces. Although organisms can sometimes disperse overlarge distances and across barriers (as in the case ofmanyoceanic islands), dispersal occurs more readily betweennearby areas, so that the biota of nearby areas, such asthe different habitats of South America, have similarcompositions. Similarly, the fossil SouthAmericanmam-malsDarwin foundwere related to the ones nowpresent.Thus, related organisms live in places they can reach bydispersal, and descend from the previous inhabitants.
This pattern is also well illustrated by the mammalfauna of Australia, a continent that became isolated fromthe other continents tens ofmillions of years ago.Most ofthe mammals of Australia are marsupials, a group now
found elsewhere only in the Americas, where they aremuch less diverse. In Australia, the marsupials occupy allthemajorhabitats andhavea rich fossil recordgoingbacktens of millions of years. Having been isolated in Aus-tralia, the marsupials have prevailed and diversified intomost ecological roles: herbivores (kangaroos), carnivores(thylacine), burrowers (wombats), climbers (koala), gli-ders (phalangers), and many more. The only native pla-cental mammals are bats andmurid rodents. Bats can fly,and rodents are able to colonize across water barriers(although not ones aswide as those that can be crossedbybats).
Geographic distributions thatmight seemanomalousunder the hypothesis of descent with modification areexplicablewhen themovement of the continents is takeninto account. For example, Cynognathus, a terrestrialsynapsid of the Triassic, about 240 million years ago,hasbeen found inSouthAmerica,Africa, andAntarctica,all now widely separated by seas unlikely to be crossedby a stout, meter-long animal like Cynoganthus. But inthe Triassic all these continents were connected in a sin-gle landmass, Gondwana. Far from being a problematiccase, Cynognathus shows again the importance of his-torical connectedness, both genealogical and geological,in the distribution of organisms.
4. EVOLUTION IN ACTION
Changes within Populations
Although fossils record descent with modification overbillions of years, evolution often occurs quickly enough
Figure 6. The right manus in the brown bear (Ursus arctos; left) andthe giant panda (Ailuropodamelanoleuca; right). The panda’s thumb isnot a true digit but a modified radial sesamoid bone. (After D. D. Davis1964.)
Lung
Gut
Rightpulmonary
artery
Left pulmonary artery
Figure 7. The lung and its arterial blood supply in the Australianlungfish (Neoceratodus). Note that the pneumatic duct swings aroundthe gut to make the lung dorsal, and the left pulmonary artery, afterbranching fromthedorsal aorta, follows it downandaround (insteadofgoing straight to the lung). (After E. S. Goodrich 1909.)
36 Introduction

to be observed within one or a few human lifetimes,and such cases allow the full panoply of evolutionarymechanisms—mutation, gene flow, genetic drift, andnatural selection—to be seen in action.
Darwin’s chief examples of observed evolution in-volved the work of animal and plant breeders in pro-ducing and elaborating the features of domesticatedorganisms (see chapter VIII.5). Studies of such speciesshow the importance of selection in establishing andfixing desired characteristics. The many varieties of do-mestic dog, differing so much in size, shape, and be-havior, have all been produced from the wolf in the lastfew thousands of years. Corn, one of the most highlymodified and economically important organisms evercreated by humans, was developed by selective breedingfrom a wild species of grass.
Rapid evolution has also unintentionally been causedby humans, who changed the environment in ways thatprompted evolutionary responses from affected organ-isms (seechaptersVIII.2andVIII.3). Industrialmelanism,the evolution of darker coloration in animals living inenvironments darkened by pollution, is widespread in avariety of insects in industrialized areas of both Europeand North America. When pollution controls have beenenacted, the evolutionary change has been reversed, andthe lighter-colored forms have again increased in fre-quency. Both the spreadofmelanismand its reversal havebeenobserved inBritain in themothBistonbetulariaovera period of about 150 years.
The use of pesticides and antibiotics has frequently ledto the undesired evolution of resistance in the targetedorganisms, including rodents, insects, bacteria, andvirus-es. Multiple-antibiotic-resistant strains of bacteria havebecome a major health problem, and one of the greatestdifficulties in treating acquired immune deficiency syn-drome (AIDS) has been the rapid evolution of resistanceby the human immunodeficiency virus (HIV). In the evo-lution of HIV, high mutation rates, short generationtime, large population sizes, and strong selection have allcombined to make the virus adapt extremely rapidly todrugs. Drug mixtures, which attack the metabolism ofthe virus in different ways simultaneously, have provenmore effective, as the multiple mutations required forresistance to all the drugs are less likely to occur.
Species introduced intonewgeographicareas,whetherby humans or by natural means, are likely to find them-selves in new environments and thus are more likely toundergo evolutionary changes. Such introductions haveindeed produced many examples of divergence in fea-tures such as size, shape, and coloration. One exampleis the house sparrow (Passer domesticus), introducedinto North America from Europe about 1851. By themiddle of the twentieth century, the sparrow hadgeographically differentiated in coloration, size, shape,and physiology in a manner parallel to that of nativespecies.
Natural populations have also been observed evolv-ing in response to natural environmental changes (see
Figure 8. Endemic and invasive oceanic island species. Left: Theendemic Bermuda skink (Eumeces [Plestiodon] longirostris) is theonly extant native terrestrial reptile of Bermuda. Its nearest rel‐atives are from the nearest mainland, North America, 1000 km tothe west. Right: A Bermudian specimen of Bufo (Rhinella) marinus,
native to Middle and South America, and introduced to Bermuda in1885. It is one of several exotic amphibian and reptile species thathave thrived and even become pests there, after the barrier to dis-persal was overcome by human agency. (Skink photo by RichardGround; toad photo by G. C. Mayer.)
Evidence for Evolution 37

chapter III.7). Perhaps the best-studied case is that ofDarwin’s finches (Geospiza) in the Galapagos, wherepopulations have been carefully tracked over decades.During this time span, repeated episodes of morpholog-ical evolution have occurred in response to climaticallyinduced variations in the food supply. Further, the ge-netic basis of these changes has been demonstrated byobservations in nature. These studies show that naturalpopulations are not evolutionarily static but can adap-tively track changes in the environment.
Speciation
Speciation, the origin of new lineages reproductively iso-lated and evolutionarily independent from other line-ages, is a key part of evolution (see Section VI: Specia-tion and Macroevolution). Without it, evolution mightoccurwithin a lineage, but therewould be no increases inbiodiversity. Divergence of lineages is enhanced by theirgeographic separation (since it reduces or eliminates thehomogenizing force of gene flow). Isolated populationsmay differentiate independently—including in character-istics affecting reproductive compatibility—to the pointthat they can no longer exchange genes if brought againinto contact. The insensible gradation over space of pop-ulations varying from mere geographic isolates tohighly differentiated populations approaching genus-level distinction provides evidence for speciation by geo-graphic isolation, and there aremany examples, suchas inkingfishers of the generaTanysiptera andHalcyon on theislands of the southwest Pacific. In most cases though,divergence takes too long to be observed within a humanlifetime. Laboratory studies have shown that incipientreproductive isolation can arise in separated populationsundergoing differential adaptation, and that reproductiveisolation can be enhanced by selection against individualswho hybridize with the “wrong” population.
In some cases, though, speciation occurs sufficientlyquickly to be observed in the wild within a human life-time. Speciation by polyploidy (i.e., the duplication ofchromosome sets) is an important mode of splitting inplants (see chapterVI.9).Awell-studied case is the recentorigin of the British salt marsh grass Spartina anglica, apolyploid derived from the natural crossing of the in-troduced American species S. alterniflora (diploid chro-mosomenumber 2n=60)with thenative S.maritima (2n= 62). Spartina alterniflora was accidentally introducedin 1829. In 1870, sterile hybrids with S. maritima werefirst recorded. Sterility of the hybrids, due to mismatch-ing of the parental chromosomes, was overcome bychromosome doubling, and the fertile S. anglica (2n =122) was first recorded in 1892. The new species hassince spread along coastlines throughout Britain.
5. EVOLUTION AS FACT AND THEORY
It is sometimes noted pejoratively that evolution is a“theory.” This pejorative usage confuses a vernacularnotion of “theory” as something uncertain or conjec-tural,with theword’s scientificusage. In science, a theoryis not a mere conjecture but a connected series of prop-ositions supported by, and explanatory of, many andvaried lines of evidence. We refer to the “germ theory ofdisease” not because we are unsure that microbes causedisease but because the theory is a set of high-level andpowerful generalizations that account for and are, inturn, supported by a huge amount of data.
Regarding evolution, it was Darwin who first con-vincingly assembled the many and varied lines of evi-dence that could be accounted for by, and provide evi-dence of, descentwithmodification.Hismany successorshave carried on his work, and Darwin would have beenboth pleased and astonished by the further lines of evi-dence that have been brought tobear. In his own lifetime,transitional fossils began to be found, and we now havethem in abundance. Darwin’s own attempts at for-mulating principles of inheritance failed, so he wouldhave been gratified by the explosive growth of geneticsand now genomics filling in what he did not know.Crucially, the facts of genetics couldhave turnedout tobeincompatible with Darwin’s evolutionary views but, in-stead, his ideas have passed this important test.Hewouldperhaps have been most astonished by the evidence forthe rapidity with which evolution can occur, includingthe formation of new species within a human lifetime.
As briefly reviewedhere, all these lines of evidence, allleading to the same conclusion, serve to make descentwith modification one of the most securely establishedhigh-level generalizations in science and allow us tospeak confidently of the fact of evolution.
FURTHER READING
Carroll, R. 2009. The Rise of Amphibians: 365Million Yearsof Evolution. Baltimore: Johns Hopkins University Press.Includes an account of the origins of vertebrates andreptiles, as well as of the origin of amphibians.
Coyne, J. A. 2009. Why Evolution Is True. New York: VikingPenguin. An account of the evidence for evolution for ageneral audience.
Dawkins, R. 2009. The Greatest Show on Earth. New York:Free Press. Another account of the evidence for evolutionfor a general audience.
Futuyma, D. J. 2013. Evolution. 3rd ed. Sunderland, MA:Sinauer. The leading textbook of evolutionary biology.
Grant, P. R., and Grant, B. R. 2008. How and Why SpeciesMultiply: The Radiation of Darwin’s Finches. Princeton,NJ: Princeton University Press. An account of the detailedevolutionary studies conducted by the authors and their
38 Introduction

associates over four decades, including several episodes ofclosely observed evolutionary changes.
Mayr, E. 2001.What Evolution Is. NewYork: Basic Books.Asummary of the evidence for evolution and its mechanismsby the “Darwin of the twentieth century.”
Prothero, D. R. 2007. Evolution: What the Fossils Say andWhy It Matters.New York: Columbia University Press. Arichly illustrated account of the fossil record emphasizingtransitions between major groups.
Shubin, N. 2008. Your Inner Fish. New York: PantheonBooks. A popular account of the discovery of the fish-
amphibian transitional form Tiktaalik, and of the tracesof common ancestry within vertebrates (including man),and of the common ancestry of vertebrates with otheranimals.
Young, D. 2007. The Discovery of Evolution. 2nd ed. Cam-bridge: Cambridge University Press. Intended for a generalaudience, this superb and richly illustrated history servesas a text for evolutionary biology itself, because it in-troduces and explicates not just the ideas and historicalfigures but the evidence on which the major discoveries ofevolutionary biology are based.
Evidence for Evolution 39


















![1+1=3 farm-making new rural [r]evolution](https://static.fdokumen.com/doc/165x107/63215f82f2b35f3bd10fdafa/113-farm-making-new-rural-revolution.jpg)

