
Human Ecology of Shellfish Exploitation at a Prehistoric Fishing-Farming Village on the Pacific...
20
Journal of Island & Coastal Archaeology, 9:183–202, 2014 Copyright © 2014 Taylor & Francis Group, LLC ISSN: 1556-4894 print / 1556-1828 online DOI: 10.1080/15564894.2014.881935 Human Ecology of Shellfish Exploitation at a Prehistoric Fishing-Farming Village on the Pacific Coast of Mexico Carley B. Smith, 1 Claire E. Ebert, 2 and Douglas J. Kennett 2 1 Department of Anthropology, University of Oregon, Eugene, Oregon, USA 2 Department of Anthropology, The Pennsylvania State University, University Park, Pennsylvania, USA ABSTRACT Shellfish remains excavated from an early agricultural village on the Pacific Coast of Mexico (Guerrero) indicate a dietary shift from lo- cally obtained estuarine shellfish (1400–1100 BC) to a greater diversity of mollusks collected from more distant marine environments (900– 500 BC). The timing of this shift suggests that it occurred as human populations increased and impacted the availability of local estuarine resources. We argue that this prompted the incorporation of a more diverse array of shellfish species harvested at greater distances or ob- tained via trade, possibly with the use of boats to transport shellfish and other resources back to the village. Keywords behavioral ecology, formative period, historical ecology, maritime adaptations, Mesoamerica, mollusks, optimal foraging theory INTRODUCTION Ancient shell middens along the coasts of the world’s oceans indicate the importance of shellfish exploitation for anatomically Received 19 June 2013; accepted 10 December 2013. Address correspondence to Douglas J. Kennett, Department of Anthropology, The Pennsylvania State University, 409 Carpenter Building, University Park, PA 16802, USA. E-mail: [email protected] Color versions of one or more of the figures in the article can be found online at http://www.tandfonline.com/uica. modern humans as they expanded out of Africa over 100,000 years ago (Erlandson 2001). Most shellfish are sessile, distributed in aggregated clumps, and available year round. Little or no specialized technology is 183 Downloaded by [Penn State University] at 07:59 08 February 2016
Transcript of Human Ecology of Shellfish Exploitation at a Prehistoric Fishing-Farming Village on the Pacific...

Journal of Island & Coastal Archaeology, 9:183–202, 2014Copyright © 2014 Taylor & Francis Group, LLCISSN: 1556-4894 print / 1556-1828 onlineDOI: 10.1080/15564894.2014.881935
Human Ecologyof Shellfish Exploitationat a PrehistoricFishing-Farming Villageon the Pacific Coastof MexicoCarley B. Smith,1 Claire E. Ebert,2 and Douglas J. Kennett21Department of Anthropology, University of Oregon, Eugene, Oregon, USA2Department of Anthropology, The Pennsylvania State University, University
Park, Pennsylvania, USA
ABSTRACT
Shellfish remains excavated from an early agricultural village on thePacific Coast of Mexico (Guerrero) indicate a dietary shift from lo-cally obtained estuarine shellfish (1400–1100 BC) to a greater diversityof mollusks collected from more distant marine environments (900–500 BC). The timing of this shift suggests that it occurred as humanpopulations increased and impacted the availability of local estuarineresources. We argue that this prompted the incorporation of a morediverse array of shellfish species harvested at greater distances or ob-tained via trade, possibly with the use of boats to transport shellfish andother resources back to the village.
Keywords behavioral ecology, formative period, historical ecology, maritime adaptations,Mesoamerica, mollusks, optimal foraging theory
INTRODUCTION
Ancient shell middens along the coasts ofthe world’s oceans indicate the importanceof shellfish exploitation for anatomically
Received 19 June 2013; accepted 10 December 2013.Address correspondence to Douglas J. Kennett, Department of Anthropology, The Pennsylvania StateUniversity, 409 Carpenter Building, University Park, PA 16802, USA. E-mail: [email protected] versions of one or more of the figures in the article can be found online athttp://www.tandfonline.com/uica.
modern humans as they expanded out ofAfrica over 100,000 years ago (Erlandson2001). Most shellfish are sessile, distributedin aggregated clumps, and available yearround. Little or no specialized technology is
183
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Carley B. Smith et al.
required to collect mollusks and people ofall ages (men and women, young and old)can collect them easily (Bird et al. 2002; Er-landson 1988; Glassow and Wilcoxon 1988;Rick and Erlandson 2009; Yesner 1980). Be-cause of these characteristics, shellfish are aconvenient and plentiful source of proteinand often a dietary staple in coastal envi-ronments. During the Holocene shellfish andothermarineresourcesprovidedastablesub-sistence base for growing populations andthe emergence of complex hunter-gatherersocieties in some regions (e.g., California,Kennett 2005; Rick et al. 2005; NorthwestCoast, Ames and Maschner 1999). People liv-ing in areas of high productivity like coastaland estuarine regions were often slowerto implement and commit to agriculture asthey had access to a wide variety of aquaticprey such as shellfish and fish (Piperno2006).
The characteristics that make shellfishan attractive economic resource also makethem particularly susceptible to predation(Erlandson and Rick 2008). The differen-tial impacts of human predation on shell-fish species depend on species’ habitats, ge-ographic distribution, available refugia, andlife history characteristics as well as hu-man preference for particular species andtechnologies used (Claassen 1986; Erland-son et al. 2011; Mannino and Thomas 2002;Thacker 2011). Heavy predation and long-term human selection of individuals cancausepopulationstructures tochangesothatthe average size of available individuals in anarea decreases over time along with alter-ations in species composition (Braje et al.2007; Erlandson et al. 2011; Mannino andThomas 2002; Milner et al. 2007; Rick andErlandson 2009). Because of the ease of pro-curement, clumped shellfish that are foundin shell-beds (like oysters and mussels) orthose found on the surface (gastropods, oys-ters, mussels) are more prone to populationdecline due to predation compared to shell-fish with more elusive behavioral attributes(e.g., burrowing taxa; de Boer et al. 2000).Larger bodied solitary species are also proneto decline in number. An increase in di-versity in species preyed upon, changes inage and size profiles, as well as shifts to
focusonalternative shellfishspecies throughtime can signify human-caused resource de-pression sometimes influenced by naturalprocesses (de Boer et al. 2000; Erlandsonand Rick 2008; Erlandson et al. 2004;Klein et al. 2004; Mannino and Thomas2002).
Here we explore the effects of humanpredationonshellfishpopulationswithin thevicinity of the early agricultural village of LaZanja, located in the Acapulco Bay region ofGuerrero, Mexico. Estuaries, sandy beaches,and rocky shorelines supported a remark-able diversity of molluskan species duringthe Middle and Late Holocene (Kennettet al. 2008). Shellfish populations have beenstrongly and negatively impacted during thelast century with the expansion of Acapulcoas a major urban center and tourist destina-tion. Our study provides an environmentalbaseline for comparison with the contem-porary distribution and diversity of shellfishtaxa in this region of Mexico. This workalso provides a glimpse at changing shellfishharvesting strategies as expanding agricul-tural populations impacted the more pristinecoastal environments of Mexico. Using opti-mality models from behavioral ecology (opti-mal foraging theory[OFT])wepredictedthatresource depression at this settlement loca-tion would lead to expanding diet breadthand the acquisition of resources from moredistant locations via long-range forays ortrade. One of the unintended consequencesof an increasing commitment to agricultureis population expansion and a host of asso-ciated environmental impacts (Hooke 2000)that effect the distribution and abundance ofnon-agricultural subsistence resources. Thedeposits at La Zanja span this critical intervalof early agricultural commitment and expan-sion. Changing shellfish harvesting strategieswithin this context that are generally consis-tent with OFT predictions.
BACKGROUND
La Zanja is located 2 km from the PacificCoast, south of Acapulco, at the confluenceof the Sabana River and the Tres Palos La-goon (see Figure 1). The site is located
184 VOLUME 9 • ISSUE 2 • 2014
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Human Ecology of Shellfish Exploitation in Mexico
Figure 1. Map of coastal Guerrero, Mexico showing the location of Las Zanja relative to other sitesin the region.
at the edge of the northeastern end of theestuary, on a narrow coastal plain (2.5–13 km wide) between the Sierra Madredel Sur and the Pacific Ocean. Stratified ar-chaeological deposits in this low earthenmound site span the Early (∼1400–1100 BC)and Middle Formative (∼900–500 BC) pe-riods of Mesoamerican prehistory (see be-low). The Formative Period (∼1800 BC–AD300) followed a trajectory established duringthe Archaic Period (∼8000–1800 BC; Ken-nett 2012) of increasing reliance upon do-mesticated plants complemented by manywild foods (Flannery 1973, 1986; Kennettand Voorhies 1996; Kennett et al. 2006;Piperno 2006; Smith 2001; Voorhies 1996).Greater commitment to agricultural produc-tion required more intensive use of land sur-rounding central places, which altered habi-tats and gradually changed the relationship
between human and local coastal andterrestrial environments (Doolittle 1990;Lentz 2000; Piperno 2006). New rela-tionships were characterized by popula-tion growth, greater sedentism, environ-mental and social circumscription, and theemergence of social hierarchies that ulti-mately led to the first state institutionsin Mesoamerica (Clark and Blake 1994; Ken-nett et al. 2006; Rosenswig 2009; Webster2002).
Early excavations in the Acapulco regionby Charles Brush between 1959 and 1960identified a number of sites along the Sa-bana River and around Acapulco Bay (Brush1969). Manzanilla Lopez et al. (1991; Man-zanilla Lopez 2000) have greatly expandedour working knowledge of the prehistory inthis region and reassessed the regional ce-ramic typology/chronology based on new
JOURNAL OF ISLAND & COASTAL ARCHAEOLOGY 185
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Carley B. Smith et al.
excavations and Brush’s original collections.Puerto Marques is the earliest site in the re-gion, dating to the Late Archaic (∼3500 BC)and was occupied through the Classic Period(∼AD 800; Brush 1969; Kennett et al. 2008;Manzanilla Lopez 2000; Manzanilla Lopezet al. 1991). Puerto Marques is located di-rectly on the coast and the faunal assemblageof this site is dominated by species frommarine ecosystems in all periods. A deeplyburied shell mound at Puerto Marques wasdated to the Late Archaic (3500–2000 BC);this mound shows the importance of coastalresources to early foragers in this area (Brush1969; Kennett et al. 2008). La Zanja was es-tablished within a lagoonal/mangrove set-ting farther from the coast during the laterpart of the Early Formative (∼1400 BC). Theproximity of the two sites (∼5 km) and theearlier founding of Puerto Marques suggestthat La Zanja was originally settled by a groupof people from Puerto Marques.
Laguna Tres Palos and the bay at PuertoMarques were probably linked by a man-grove swamp during the early and middleHolocene but are now separated by a bar-rier beach (Kennett et al. 2004, 2008). Con-ditions in the modern lagoon change sea-sonally with the discharge of the La SabanaRiver and changing tidal influences as theinlet opens and closes to the Pacific Ocean(Kennett et al. 2004). The coastal lagoonsand rivers provided access by watercraftto many areas along the coastline and sea-sonal inundation of the lower reaches of thecoastal plain offered a form of natural irri-gation that could have facilitated agricultureduring the Formative Period. In this way LaZanja’s location allowed for strategic use ofa wide variety of resource patches with dif-ferent prey within a 5-km radius of the site.The coastal plain and piedmont areas sur-rounding the site provided access to terres-trial mammals and the adjacent marine habi-tat supplied fish, shellfish, and sea turtles.A mangrove estuary along which the siteis located offered additional local resourcessuch as birds, lagoon fish, gastropods, andbivalves that required little time and effortto acquire. Aquatic invertebrates were alsoavailable from the intertidal mudflat near thesite. A decrease in overall energetic returns
from wild fauna over time at La Zanja proba-bly coincided with a greater dependence ondomestic plant foods, which would be com-plemented by protein from shellfish. Agricul-ture and high populations would have had aperceptible impact on local resources thatmay have necessitated forager-fisher-farmersto travel farther into different habitats to pro-cure resources. Lagoon systems around thesite could have provided efficient travel tomore distant resource patches and the trans-port of these resources back to La Zanja viawatercraft.
THEORETICAL FRAMEWORK
Human behavioral ecology (HBE) providesa set of models that researchers can useto explore complex ecological interactionswithin changing social and natural environ-ments (Broughton and O’Connell 1999; Ka-plan and Hill 1992; Kennett 2005; Kennettet al. 2009; Winterhalder and Kennett 2006;Winterhalder et al. 2010; Winterhalder andSmith 2000). Within the broader HBE frame-work, OFT provides models to evaluate andexamine prehistoric subsistence strategies.Costs and benefits of different foraging be-haviors are evaluated given a set of environ-mental parameters and the models assumethat humans tend to make economically ra-tional decisions that optimize energetic re-turns while minimizing investment costs andrisk (Broughton and O’Connell 1999; Kaplanand Hill 1992; Smith 1983; Stephens andKrebs 1986). In OFT, benefits are estimatedby caloric energetic returns of different preyitems with costs including time spent inmanufacturing tools, search, pursuit, andprocessing.
We use the prey choice model andthe Marginal Value Theorem here to ex-plore shellfish harvesting strategies at LaZanja through the Early and Middle For-mative Periods and the expansion of dietbreadth through time. With long-term inten-sive use of a relatively homogenous local en-vironment, usually associated with increas-ing populations and targeting of high-rankedspecies, the prey choice model predictspredation pressure that results in declining
186 VOLUME 9 • ISSUE 2 • 2014
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Human Ecology of Shellfish Exploitation in Mexico
abundance of higher ranked large species(Broughton and O’Connell 1999; Kaplan andHill 1992; Smith 1983). With resource de-pression, encounter rates with high-rankedfoods decrease until foraging return rates de-crease enough that foragers expand their di-ets to incorporate lower ranked prey itemsand the optimal diet becomes more diverse(Broughton and O’Connell 1999; Hawkesand O’Connell 1992; Kennett et al. 2006).The Marginal Value Theorem makes predic-tions about how long an individual will stayin a given resource patch before moving onto exploit another (Charnov 1967). Individ-uals will widen their foraging range as localfood sources are impacted, and may focus onhigher rankedprey tocompensate forenergyexpended in travel until a diet that optimizesenergetic returns is reached.
The prey choice model assumes homo-geneity of prey distribution within the en-vironment, but a division of the foragingarea into resource patches allows applica-tion of measures of evenness and diver-sity within patches that are sensitive to re-sourcedepressionandwideningdietbreadth(Nagaoka 2002). A decrease in encounterratescouldprompt individuals towidentheirsearch area or to target different patches.However, at a certain point, if high-rankeditems become so depleted that the increasein search time is not met with sufficient re-turns, more low-ranked local resources maybeaddedto increasedietbreadth(Broughtonand O’Connell 1999; Hawkes and O’Connell1992; Hawkes et al. 1997). In this way, anincrease in diversity indices can also signala decrease in foraging returns. Hawkes andO’Connell (1992) point out that where thediet is relatively narrow, foragers will typi-cally target high-ranked prey to justify costs,with high search times relative to handlingtime. They go on to say, “Conversely, wherediet isbroadandhandlingrepresents thebulkof foraging effort, improvements in handlingefficiencywouldhave large effects” (Hawkesand O’Connell 1992:64). As targeted preyspecies decline and high populations pre-vent the expansion of sedentary groups intoless impacted environments, reliance on in-tensive food production with high handlingcosts may increase.
Within the La Zanja faunal assemblage,the highest ranked prey were marine tur-tles that were hunted locally while nestingon beaches and in the estuarine zone (Smithet al. 2007). Other high-ranked mammalsfound in the assemblage include deer andpeccary; the energetic returns for thesespecies may have been moderated by in-creased travel and search time in interiorlocations. Estuarine fish and shellfish mayhave individually represented a lower pro-tein/energycontributionthanlargeranimals,but were the easiest prey to acquire asthey were present in abundance in the la-goon near the settlement. There is a trade-off between energy capture and subsistencerisk and the reliability of shellfish may haveserved the foraging goals of men, women,and children differently (Zeanah 2004). Thehabitats of marine fish and shellfish wouldrequire more time spent in travel to acquirethan estuarine varieties, but were probablya more reliable and consistent food sourcethan many of the larger animals. Madsen andSchmitt (1998) emphasize that prey itemssuch as insects and shellfish can be collectedin mass and that their dietary ranking is den-sity dependent. They point out that whensmall animals are available in large amountsthey become a high-ranked item and can dis-place larger prey in optimal diets. Many ofthe shellfish species in the La Zanja assem-blage occur in clumps and were likely cap-tured with mass harvesting techniques (e.g.,rakes) and thus contributed to larger ener-getic returns compared with solitary animalsof the same size.
METHODS
The La Zanja site is an earthen mound 3.2 mhigh with an area of 40,000 m2. In 2001we excavated a 2 × 2 m unit at the high-est point in the mound down to a depth of3.6 m in order to gain a complete view of For-mative period occupation (20 cm levels; seeFigure 2). Previous excavations at the site byBrush (1969), along with our own system-atic auger tests, indicate that the Early andMiddle Formative Period deposits are onlyrepresented in the center of the site. The
JOURNAL OF ISLAND & COASTAL ARCHAEOLOGY 187
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Carley B. Smith et al.
Figure 2. North profile of excavated unit at La Zanja showing Early and Middle Formative Periodstratigraphic units and associated AMS 14C dates.
188 VOLUME 9 • ISSUE 2 • 2014
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Human Ecology of Shellfish Exploitation in Mexico
upper Middle Formative Period deposit ex-tends more widely and is indicative of pop-ulation expansion after the settlement wasinitially established in the Early Formative.The 2 × 2 m unit was placed in the centerof the site to sample the stratigraphic su-perposition of Early and Middle FormativePeriod deposits. The study was exploratorydue to budgetary constraints and additionalwork will be required to determine thedegree of spatial heterogeneity of thesedeposits.
Pumps were used to excavate below thewater table (∼108 cm beneath the surface;Kennett et al. 2004). Stratigraphy was essen-tially horizontal and there were two majorcultural components represented by uniqueartifact assemblages dating to the Early (360–240 cmbd) and Middle Formative (200–80cmbd) Periods. Seven AMS 14C dates werecalibrated and modeled stratigraphically inOxCal 4.2 to establish the chronology for thesite (Figure 3; Bronk Ramsey 2013; Kennettet al. 2011). Table 1 provides the 2-σ cali-brated ranges for each date and the modeledage for each phase (or period). Dates wereplaced in an ordered sequence and modeledwithin two phases, Early and Middle Forma-tive, based on stratigraphic association. Thespan of time representedbyeachdatedeventfor the Early Formative phase is estimatedto be between 24 and 344 years (∼1400–1100 BC), with a total maximum durationfor the phase of 846 years. The span of thedated events for the Middle Formative phaseis between 53 and 354 years (∼900–550 BC),with the total maximum duration estimatedat 796 years. While the maximum durationof occupation at the site for the Early Forma-tive is approximately 120 years longer thanthe Middle Formative, average durations foreach phase (423 and 420 years, respectively)suggest comparable occupations of compa-rable duration. The Formative period occu-pation at La Zanja is contemporary with thegrowth of settlements on the Gulf Coast (SanLorenzo; Cyphers 1996; Cyphers et al. 2007–2008), Oaxaca (San Jose Mogote; Flanneryand Marcus 2000), and the Soconusco (Pasode la Amada; Clark and Blake 1994).
Sediments from each 20-cm level of theunitwere screened through5-mmmeshwith
anadditional104-liter sample fromeach levelpassed through a 3-mm screen (Kennett et al.2004). Shells in the assemblage were frag-mented from processing, but large enoughthat the 5-mm sample considered here is rep-resentative. Field assessment of the 3-mmresidue indicate that shellfish species smallerthan 5 mm are rare in the assemblage. Lev-els analyzed are of equal volume (5.6 m3)for both periods and the duration of occupa-tion is similar so we argue that the samplesare directly comparable. Poteate and Fitz-patrick (2013) have suggested that samplingless than 16% of zooarchaeological remainsof a study population may produce resultsthatarenotstatisticallysignificantandarenotrepresentative. Though the assemblage ana-lyzed represents a subsample of the greaterpopulation at the site, analysis of smallerunits (e.g., 50 × 50 cm) are not uncommonfor coastal settings. Shellfish are abundant inthe excavation at La Zanja, and the overallzooarchaeological assemblage is closely mir-rored by the diverse assemblage excavated atthe nearby site of Puerto Marques. Previousresearch from both sites shows a trend of hu-man predation that resulted in the reducedavailability of high-ranked sea turtles fromthe Early to Middle Formative periods (Smithet al. 2007). Similar trends observed in theshellfish assemblage at La Zanja are likely tobe representative of long-term changes in hu-man foraging patterns that led to an increas-ing diet breadth. Modern trash was mixedinto the upper 80 cm of the deposit and therewas a single mixed level (200–220 cm) be-tween the Early and Middle Formative Periodcomponents. These mixed deposits were ex-cluded from our analysis.
Shellfish remains were identified us-ing a comparative collection developedby Barbara Voorhies and Douglas Kennett(Table 2). We used the Number of Identi-fiedSpecimens(NISP)andMinimumnumberof Individuals (MNI) to quantify taxonomicabundances for mollusk species. MNI andNISP are common zooarchaeological meth-ods that are often highly correlated. MNI pro-vides researchers with a close approxima-tion of the number of individuals belongingto a study population (Classen 1998; Reitzand Wing 1999).
JOURNAL OF ISLAND & COASTAL ARCHAEOLOGY 189
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Carley B. Smith et al.
#OS-40961 - RC 20
#OS-40963 - RC 5
#OS-40962 - RC 1
#OS-40865 - RC 2
#OS-40964 - RC 7
#OS-40963 - RC 4
#OS-41342 - RC 15
Modelled date (BC/AD)
0 500 1000 1500Calendar Years
Duration of Middle Formative Occupation
Duration of Early Formative Occupation
2500 2000 1500 1000 500 1BC/1AD 500
Earliest La Zanja Boundary
Latest La Zanja BoundaryM
iddl
e Fo
rmat
ive
Phas
eEa
rly F
orm
ativ
e Ph
ase
Boundary - Transition
Figure 3. Modeled calibrations for AMS 14C dates at La Zanja.
190 VOLUME 9 • ISSUE 2 • 2014
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Human Ecology of Shellfish Exploitation in Mexico
Table 1. AMS 14C dates from La Zanja used in Bayesian chronological model.
Sequence/phase OS # Sample 14C age (BP) Modelled 2-σ cal range
Boundary 784–266 BC
Middle Formative period
41343 RC 15 2510 ± 30 793–551 BC
40963 RC 4 2520 ± 35 797–552 BC
40964 RC 7 2720 ± 40 936–803 BC
40865 RC 2 2540 ± 40 805–551 BC
Boundary 1134–818 BC
Early Formative period
40962 RC 1 2890 ± 30 1208–1001 BC
41342 RC 5 3010 ± 40 1382–1116 BC
40961 RC 20 3030 ± 40 1386–1126 BC
Boundary 1737–1134 BC
Indices were calculated for richness, di-versity and evenness in the Early and Mid-dle Formative Period shellfish assemblages.Richness was determined as a raw countof taxa present in the Early and Middle For-mative assemblages. It is simply a measureof the number of shellfish taxa representedin the sub-assemblages being compared andis a product of the overall species diversity,prehistoric subsistence and processing deci-sions, and variations in sample size and shellpreservation. Diversity was calculated usingthe Shannon-Weaver Index and takes into ac-count number of species present and howevenly the numbers (MNI) are distributedamong taxa (Reitz and Wing 1999:104–106).This accounts for the overall dietary contri-bution of each species rather than simplepresence or absence. Evenness or equitabil-ity was calculated by dividing the Shannon-Weaver function by the number of species inthecommunity. Itprovidesameasureofhowevenly distributed the abundance of taxa arein the assemblage with 1 indicating evendistribution and lower values indicating thedominanceof1ormorespecies.This index istherefore sensitive to dietary shifts from sin-gle high-ranked prey to a greater diversity oflow-ranked prey, or vice versa, through time.
RESULTS
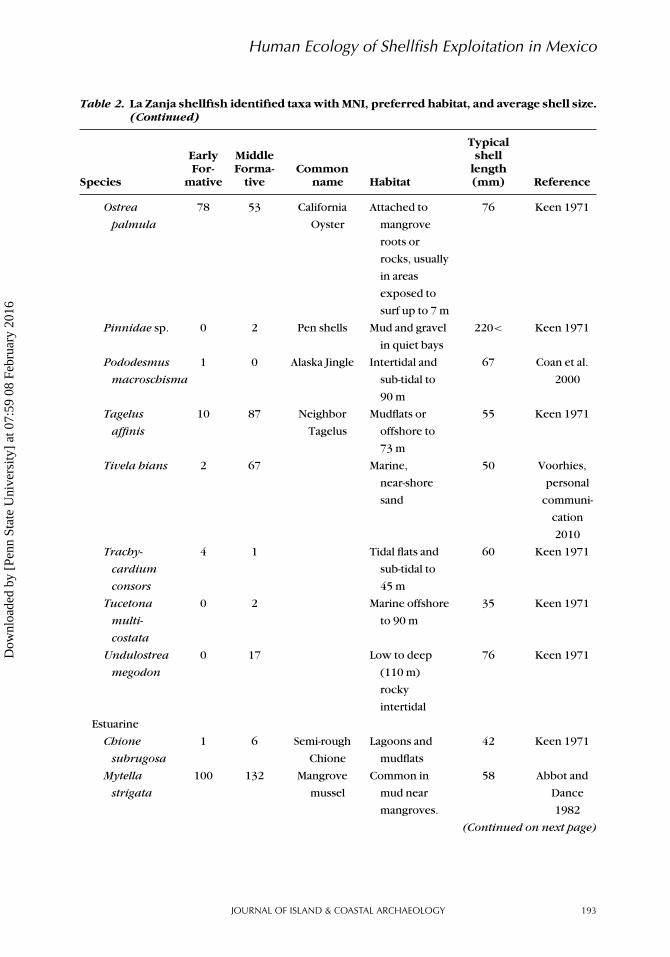
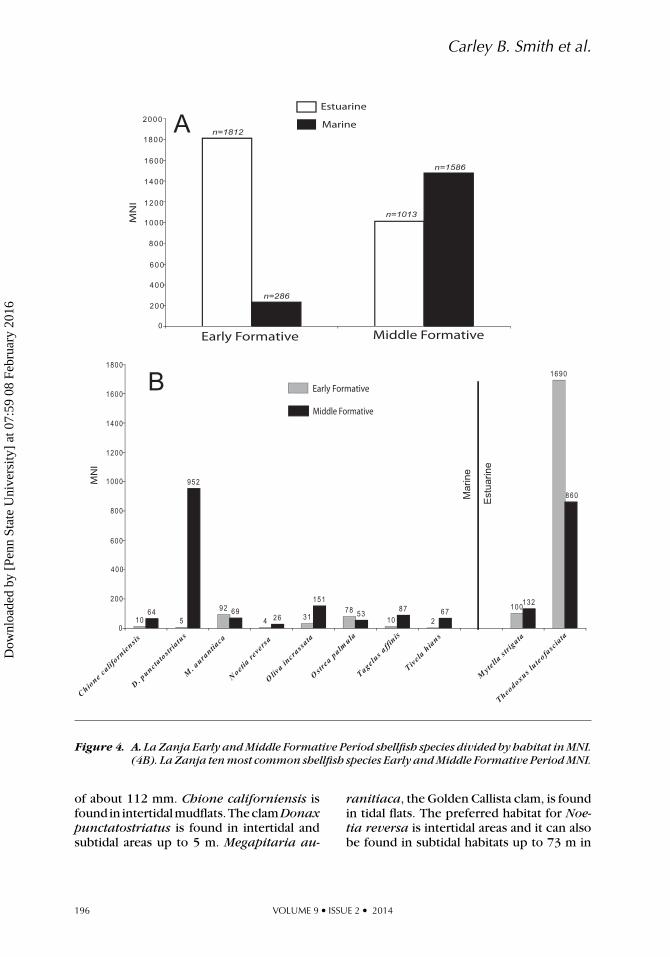
Shellfish taxa were compared using MNIwith most species divided into habitats(Table 2, Figure 4A). A total of 4702 shellfish(MNI) from La Zanja were identified fromboth Early and Middle Formative strata. Inthe Early Formative, 2,102 total shellfish(MNI) were recovered and 2,091 wereidentified to the species level (23 distinctspecies). Another 11 MNI were identifiedto the genus level (two additional generaidentified). In the Middle Formative, shell-fish remains increase to 2,600 MNI andrepresent a much more diverse collectionof taxa with 2,562 MNI identified to 31different species, 35 MNI identified to 5genera, 2 MNI identified to family, and 1MNI identified only as an unknown taxa ofbarnacle. Ten species dominate the La Zanjashellfish assemblage: Chione californiensis,Donax punctatostriatus, Megapitariaauranitiaca, Noetia reversa, Ostrea pal-mula, Tagelus affinis, Tivela hians, Mytellastrigata, Oliva incrassata, and Theodoxusluteofasciata. Together these species con-stitute 96% of the Early Formative shellfishassemblage and 95% of shellfish remains inthe Middle Formative (Figure 4B).
JOURNAL OF ISLAND & COASTAL ARCHAEOLOGY 191
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016
Carley B. Smith et al.
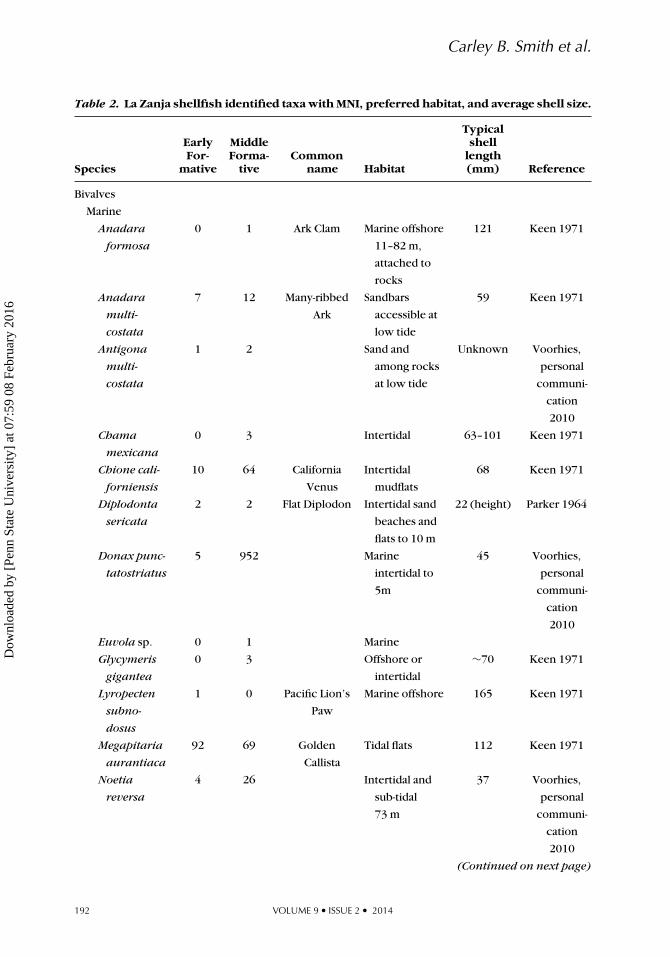
Table 2. La Zanja shellfish identified taxa with MNI, preferred habitat, and average shell size.
Species
EarlyFor-
mative
MiddleForma-
tiveCommon
name Habitat
Typicalshell
length(mm) Reference
Bivalves
Marine
Anadara
formosa
0 1 Ark Clam Marine offshore
11–82 m,
attached to
rocks
121 Keen 1971
Anadara
multi-
costata
7 12 Many-ribbed
Ark
Sandbars
accessible at
low tide
59 Keen 1971
Antigona
multi-
costata
1 2 Sand and
among rocks
at low tide
Unknown Voorhies,
personal
communi-
cation
2010
Chama
mexicana
0 3 Intertidal 63–101 Keen 1971
Chione cali-
forniensis
10 64 California
Venus
Intertidal
mudflats
68 Keen 1971
Diplodonta
sericata
2 2 Flat Diplodon Intertidal sand
beaches and
flats to 10 m
22 (height) Parker 1964
Donax punc-
tatostriatus
5 952 Marine
intertidal to
5m
45 Voorhies,
personal
communi-
cation
2010
Euvola sp. 0 1 Marine
Glycymeris
gigantea
0 3 Offshore or
intertidal
∼70 Keen 1971
Lyropecten
subno-
dosus
1 0 Pacific Lion’s
Paw
Marine offshore 165 Keen 1971
Megapitaria
aurantiaca
92 69 Golden
Callista
Tidal flats 112 Keen 1971
Noetia
reversa
4 26 Intertidal and
sub-tidal
73 m
37 Voorhies,
personal
communi-
cation
2010
(Continued on next page)
192 VOLUME 9 • ISSUE 2 • 2014
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016
Human Ecology of Shellfish Exploitation in Mexico
Table 2. La Zanja shellfish identified taxa with MNI, preferred habitat, and average shell size.(Continued)
Species
EarlyFor-
mative
MiddleForma-
tiveCommon
name Habitat
Typicalshell
length(mm) Reference
Ostrea
palmula
78 53 California
Oyster
Attached to
mangrove
roots or
rocks, usually
in areas
exposed to
surf up to 7 m
76 Keen 1971
Pinnidae sp. 0 2 Pen shells Mud and gravel
in quiet bays
220< Keen 1971
Pododesmus
macroschisma
1 0 Alaska Jingle Intertidal and
sub-tidal to
90 m
67 Coan et al.
2000
Tagelus
affinis
10 87 Neighbor
Tagelus
Mudflats or
offshore to
73 m
55 Keen 1971
Tivela hians 2 67 Marine,
near-shore
sand
50 Voorhies,
personal
communi-
cation
2010
Trachy-
cardium
consors
4 1 Tidal flats and
sub-tidal to
45 m
60 Keen 1971
Tucetona
multi-
costata
0 2 Marine offshore
to 90 m
35 Keen 1971
Undulostrea
megodon
0 17 Low to deep
(110 m)
rocky
intertidal
76 Keen 1971
Estuarine
Chione
subrugosa
1 6 Semi-rough
Chione
Lagoons and
mudflats
42 Keen 1971
Mytella
strigata
100 132 Mangrove
mussel
Common in
mud near
mangroves.
58 Abbot and
Dance
1982
(Continued on next page)
JOURNAL OF ISLAND & COASTAL ARCHAEOLOGY 193
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Carley B. Smith et al.
Table 2. La Zanja shellfish identified taxa with MNI, preferred habitat, and average shell size.(Continued)
Species
EarlyFor-
mative
MiddleForma-
tiveCommon
name Habitat
Typicalshell
length(mm) Reference
Polymesoda
sp.
4 6 Brackish to
freshwater,
not sandy
beach
30–60 Keen 1971
Gastropods
Marine
Cerithium
stercusmus-
carum
20 18 Cerith snails Intertidal rocky
shores
25 Parker 1964
Cymatium
wiegmanni
5 2 Deep water on
rocks; used
by hermit
crabs
76
Hexaplex
erythrosto-
mus
3 8 Pink Murex Intertidal; feed
on
Megaptaria
squalida
101 Keen 1971
Natica
chemnitzii
0 7 Intertidal
mudflats and
sand
33 Keen 1971
Northia
northiae
0 1 Offshore 45 Voorhies,
personal
communi-
cation
2010
Oliva
incrassata
31 151 Olivella Snail Outer edge of
sandspits at
very low
tides
55 Keen 1971
Semicassis
sp.
0 1 Sand at low tide 50 Keen 1971
Stramonita
biserialis
3 2 Rocky intertidal 75 Keen 1971
Strombus
galeatus
0 2 Cortez Conch Rocky areas at
low tide and
close
offshore
190 Keen 1971
(Continued on next page)
194 VOLUME 9 • ISSUE 2 • 2014
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Human Ecology of Shellfish Exploitation in Mexico
Table 2. La Zanja shellfish identified taxa with MNI, preferred habitat, and average shell size.(Continued)
Species
EarlyFor-
mative
MiddleForma-
tiveCommon
name Habitat
Typicalshell
length(mm) Reference
Tripsycha sp. 0 4 Worm Snails Marine. 80 (length
of coil); 10
(diam. of
opening)
Keen 1971
Turritella
leucostoma
0 2 Marine, up to
depths of
40 m mud
and sand flats
115
(length);
20 (diam.)
Keen 1971
Estuarine
Cerithidea
mazatlanica
17 10 Incidental?
Tidal flats,
mudflats, and
marshes
27 Keen 1971
Theodoxus
luteofasciata
1,690 860 Margins of
mangrove
swamps and
mudflats
12 Keen 1971
Marine/Estuarine
Melongena
patula
4 1 Pacific Crown
Conch
Sand, mudflats,
intertidal,
and estuaries
117 Keen 1971
Arthropods
Marine
Megabalanus
sp.
7 23 Barnacle Intertidal
marine on
rocks
Unknown
barnacle
0 1 Barnacle Intertidal
marine on
rocks
Total MNI 2,102 2,600
Total taxa 25 38
We identified the habitat for each mol-lusk taxa based on Keen’s (1971) designa-tions for western American tropical shellfish(Table 2, Figure 4A). Seven of the ten mostcommon species are marine clams that are
found in intertidal sandy beach or mudflathabitats. Most of the clam species in the as-semblage are small, with shell sizes that av-erage less than 70 mm. Megapitaria aurani-tiaca is larger with an average shell width
JOURNAL OF ISLAND & COASTAL ARCHAEOLOGY 195
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016
Carley B. Smith et al.
n=1812
n=286
0
200
400
600
800
1000
1200
1400
1600
1800
2000
MN
I
Early Formative Middle Formative
n=1013
n=1586
Estuarine
Marine
10 592
4 3178
10 2
100
1690
64
952
6926
151
53 87 67132
860
0
200
400
600
800
1000
1200
1400
1600
1800
Chione californ
iensis
D . puncta
tostriatu
s
M. a
urantia
ca
Noetia re
vers
a
O liva in
crass
ata
Ostrea palm
ula
Tagelus a
ffinis
Tivela hians
Mytella
strig
ata
Theodoxus lu teofa
sciata
Early Formative
Middle Formative
MN
I
Est
uarin
e
Mar
ine
A
B
Figure 4. A. La Zanja Early and Middle Formative Period shellfish species divided by habitat in MNI.(4B). La Zanja ten most common shellfish species Early and Middle Formative Period MNI.
of about 112 mm. Chione californiensis isfound in intertidalmudflats.TheclamDonaxpunctatostriatus is found in intertidal andsubtidal areas up to 5 m. Megapitaria au-
ranitiaca, the Golden Callista clam, is foundin tidal flats. The preferred habitat for Noe-tia reversa is intertidal areas and it can alsobe found in subtidal habitats up to 73 m in
196 VOLUME 9 • ISSUE 2 • 2014
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016
Human Ecology of Shellfish Exploitation in Mexico
depth. Tagelus affinis, the Neighbor Tagelusis found in mudflats or offshore sand to 73 m.The clam Tivela hians, is found in near-shoresand. Another bivalve species that is com-mon at La Zanja is the California oyster, Os-trea palmula, which attaches to mangroveroots and rocks typically in areas of tidalreach up to depths of 7 m. The Olivella snail,Oliva incrassata is also typically found inmarine, intertidal sandy beaches, commonlyfound on the surface at low tides. These ma-rine taxa were harvested either to the westor southwest of La Zanja along the barrierbeach or northwest in Puerto Marques. Bothof these areas were easily accessed by water-craft from La Zanja. The dominant estuarineshellfish taxa found in the assemblage (Theo-doxus luteofasciata and Mytella strigata)are both found in muddy lagoon sedimentsadjacent to the site.
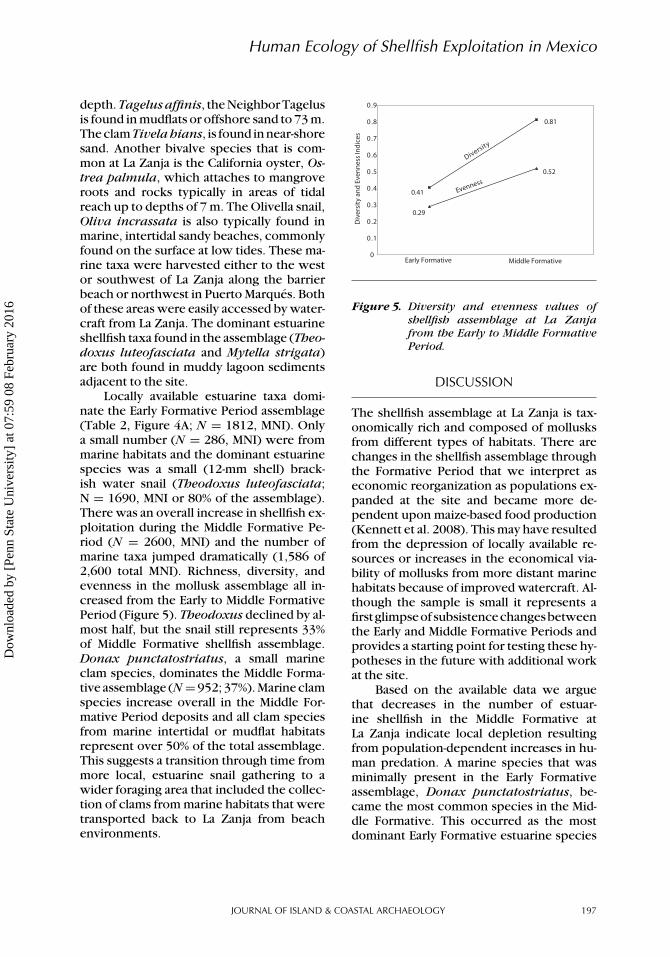
Locally available estuarine taxa domi-nate the Early Formative Period assemblage(Table 2, Figure 4A; N = 1812, MNI). Onlya small number (N = 286, MNI) were frommarine habitats and the dominant estuarinespecies was a small (12-mm shell) brack-ish water snail (Theodoxus luteofasciata;N = 1690, MNI or 80% of the assemblage).There was an overall increase in shellfish ex-ploitation during the Middle Formative Pe-riod (N = 2600, MNI) and the number ofmarine taxa jumped dramatically (1,586 of2,600 total MNI). Richness, diversity, andevenness in the mollusk assemblage all in-creased from the Early to Middle FormativePeriod (Figure 5). Theodoxus declined by al-most half, but the snail still represents 33%of Middle Formative shellfish assemblage.Donax punctatostriatus, a small marineclam species, dominates the Middle Forma-tive assemblage (N =952; 37%). Marine clamspecies increase overall in the Middle For-mative Period deposits and all clam speciesfrom marine intertidal or mudflat habitatsrepresent over 50% of the total assemblage.This suggests a transition through time frommore local, estuarine snail gathering to awider foraging area that included the collec-tion of clams from marine habitats that weretransported back to La Zanja from beachenvironments.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Early Formative Middle Formative
0.41
0.81
0.29
0.52
Diversity
Evenness
Div
ersi
ty a
nd
Eve
nn
ess
Ind
ices
Figure 5. Diversity and evenness values ofshellfish assemblage at La Zanjafrom the Early to Middle FormativePeriod.
DISCUSSION
The shellfish assemblage at La Zanja is tax-onomically rich and composed of mollusksfrom different types of habitats. There arechanges in the shellfish assemblage throughthe Formative Period that we interpret aseconomic reorganization as populations ex-panded at the site and became more de-pendent upon maize-based food production(Kennett et al. 2008). This may have resultedfrom the depression of locally available re-sources or increases in the economical via-bility of mollusks from more distant marinehabitats because of improved watercraft. Al-though the sample is small it represents afirstglimpseof subsistencechangesbetweenthe Early and Middle Formative Periods andprovides a starting point for testing these hy-potheses in the future with additional workat the site.
Based on the available data we arguethat decreases in the number of estuar-ine shellfish in the Middle Formative atLa Zanja indicate local depletion resultingfrom population-dependent increases in hu-man predation. A marine species that wasminimally present in the Early Formativeassemblage, Donax punctatostriatus, be-came the most common species in the Mid-dle Formative. This occurred as the mostdominant Early Formative estuarine species
JOURNAL OF ISLAND & COASTAL ARCHAEOLOGY 197
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Carley B. Smith et al.
(Theodoxus luteofasciata) was reduced toover half its former abundance. The col-lection of this marine species at more dis-tant locations may have been enhanced bythe availability of watercraft. There was anincreased focus in the Middle Formativeon bivalve clam species from sandy andmudflat intertidal zones. The two mostcommon shellfish species in the Early For-mative, the estuarine gastropod Theodoxusluteofasciata and the brackish water musselMytella strigata, are both perching speciesthat would be easily located and collectedin large numbers from the surface of sand,rocks, or mangrove roots. These specieswould require little skill or time to find andgather compared with the burrowing clamspecies that make up over 50% of the MiddleFormative shellfish assemblage.
Small species such as aquatic gastropodsindividually represent a small energeticreturn, but require little skill to collect inlarge quantities. These gastropods were alsoeasy to process by crushing in large quanti-ties or boiling, as demonstrated ethnograph-ically and archaeologically in other areas(Raab 1992; Voorhies 2004). Although themollusk assemblage is well preserved thesegastropods all show evidence for crackingfor meat extraction. Most of the marine shell-fish species evident in the Middle Forma-tive deposits were mudflat or intertidal clamsand have larger meat packages than the es-tuarine snail. Large tidal flat bivalves requiremore expertise and time to locate and har-vest because they burrow in the sand (Raab1992), although technologies such as rakesmay have been used to reduce time andeffort. Clam meat can be dried and storedfor long periods of time. The handling andtransportationcostsof acquiringmarinemol-lusks from more distant locations places lim-its on the taxa pursued and how they wereprocessed (Bird and Bliege Bird 1997). Ifwatercraft were more limited in the EarlyFormative Period, then extensive field pro-cessing (e.g., removal of shells, drying; Birdand Bliege Bird 1997) would have beennecessary to generate viable load sizes ex-ceeding the energy expended in transportbiasing the assemblage towards underrep-resentation of shellfish remains (Bird et al.
2002). The marine mollusks in the MiddleFormativeassemblage,ontheotherhand,arethe largestavailablealongthecoastandlargerthan locally available snails. This pattern isconsistent with the prey choice model pre-diction that larger species will be targeted astransport costs increase. However, the dom-inance of marine mollusks at La Zanja frommore distant locations during the Middle For-mative points to changes in transportationcosts consistent with the availability of moreviable watercraft (see below).
The overall quantity of shellfish in-creased in the assemblage from the EarlyFormative (MNI = 2098) to the Middle For-mative (MNI = 2599), perhaps indicating ahigher level of predation pressure as humanpopulations rose in the village (Table 2, Fig-ure 4A). Shellfish increased in diversity andevenness during the Middle Formative (Fig-ure 5) suggesting expanding diet breath inthe face of population-dependent resourcedepression locally. Larger shellfish were alsocollected at more distant locations and trans-ported back to La Zanja. This suggests thatpeople were searching farther and longer forprey during the Middle Formative. If water-craft were used more in the Middle Forma-tive, this could increase foraging efficiency,as these early farmers could cover moreground and easily return with materials frommore distant resource patches (Gomez et al.2011;Thomas2008).Arise in theuseofboatsmight negate the importance of removinglow-utility parts (shell) prior to transport ofshellfish back to central places (Ames 2002).More marine shell in the Middle Formativeassemblage is expected if boats were usedfor the transport of goods more frequentlythan in the Early Formative. However, if ma-rine shellfish were transported primarily onfoot in both periods they may be underrepre-sented in both assemblages and more impor-tant dietary constituents than faunal remainsindicate (Bird et al. 2002).
There isevidence for increasedmaritimetrade along the Pacific Coast during the lat-est Early Formative and earliest Middle For-mative Periods (Gomez et al. 2011). There isalso evidence for the development of moreintensive trade networks with the Gulf CoastOlmec either over land or across the Isthmus
198 VOLUME 9 • ISSUE 2 • 2014
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Human Ecology of Shellfish Exploitation in Mexico
of Tehuantepec and up and down the Pa-cific Coast. Lagoons and coastal river systemsserved as important flat-water conduits forboats carrying pottery, obsidian, and othervaluable resources. White slipped and in-cised pottery became more prevalent in theassemblage at La Zanja during the MiddleFormative along with other status items sug-gesting linkages/contact with the Gulf CoastOlmec (Blomster et al. 2005; Neff 2006). It islikely that watercraft were commonly usedat this time both for the transport of long-distance trade goods and for local travel. Iftransportation via watercraft became morecommon in the Middle Formative, there mayhave been less incentive to process mollusksbefore transport, consistent with the higherincidence of low-utility shell from marine in-vertebrates in the La Zanja Middle Formativecomponent. As populations increased dur-ing the Middle Formative, the use of water-craft might have served as a cost-reductionstrategy in the face of resource depression,since travel costs to more distant patcheswould be reduced. It is also possible that ma-rine mollusks were obtained via trade withcoastal communities. Agricultural productiv-ity was higher on the more expansive allu-vial portions of the coast compared with bar-rier beach locations and maize, beans, andsquash could have been exchanged for ma-rine foods including shellfish. Trade and theproduction of trade items to exchange forfood provides another mechanism for ex-panding dietary breadth (Kennett 2005). Re-gardless, changes in the La Zanja shellfish as-semblage through time are generally consis-tentwiththepreychoicemodelandMarginalValue Theorem for dietary expansion in theface of population-dependent resource de-pression.
CONCLUSION
The Formative Period occupants of La Zanjacombined the cultivation of domesticatedplants with a heavy reliance upon coastaland estuarine resources, largely shellfish andfish. Foraging strategies changed and dietbreadth expanded to incorporate a larger ar-ray of shellfish taxa as people became more
reliant on maize-based food production andvillage populations increased during the Mid-dle Formative Period. Mollusks were also col-lected from more distant beach habitats andtransported back to La Zanja. This parallelsan increased reliance upon boats for tradeevident along with coasts of Mesoamericaduring the Formative Period (Gomez et al.2011). The predictions of OFT models areconsistent with many of the patterns seenin the shellfish assemblage at La Zanja wherethe increase inshellfishcatchmentarea in theMiddle Formative seems to occur at a time ofpopulation expansion and increasing humanenvironmental impacts with a growing com-mitment to maize agriculture.
While our examination of shellfish ex-ploitation and expanding diet breadth at LaZanja predict a more generalized diet anda movement of foragers into more distanthabitats, with decreasing encounter rateswith local high-ranked prey (Broughton andBayham 2003; Broughton and O’Connell1999; Hawkes and O’Connell 1992), parallelshifts in diet breadth might also be exploredthrough changes in other aquatic taxa (fish,turtles, etc.). Our previous work suggests thelocal depression of nesting sea turtle popu-lations at this and other locations throughtime (Smith et al. 2007) that is consistentwith expanding human populations region-ally. Future work will focus on analysis offish taxa and other faunal classes (e.g., largemammals) in order to rank prey species andexamine changes in foraging patterns thatmay have existed at La Zanja. Through an ex-amination of a larger zooarchaeological as-semblage, optimal foraging models from be-havioral ecology provide an effective tool forexploring the ecological effects of popula-tion growth and human predation withincoastal settings.
ACKNOWLEDGEMENTS
We thank Madonna Moss, Jon Erlandson,Scott Fitzpatrick, and two anonymous re-viewers for thoughtful comments and sug-gestions that improved the article greatly.
JOURNAL OF ISLAND & COASTAL ARCHAEOLOGY 199
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Carley B. Smith et al.
FUNDING
This work was funded by the National Sci-ence Foundation (BCS- 0211215, Kennett).
REFERENCES
Abbott, R. T. and S. P. Dance. 1982. Compendiumof Seashells: A Color Guide to More than 4,200of the World’s Marine Shells. New York: E. P.Dutton.
Ames, K. M. 2002. Going by boat: The forager-collector continuum at sea. In Beyond For-aging and Collecting: Evolutionary Changein Hunter-Gatherer Settlement Systems (B.Fitzhugh B. and J. Habu, eds.):17–50. NewYork: Kluwer/Plenum Press.
Ames, K. M. and H. D. G. Maschner. 1999. Peoplesof theNorthwestCoast:TheirArchaeologyandPrehistory. New York: Thames and Hudson.
Bird, D. W. and R. L. Bliege Bird. 1997. Contem-porary shellfish gathering strategies among theMeriam of the Torres Strait Islands, Australia:Testing predictions of a central place forag-ing model. Journal of Archaeological Science24:39–63.
Bird, D. W., J. L. Richardson, P. M. Veth, A. J.Barham. 2002. Explaining shellfish variability inMiddens on the Meriam Islands, Torres Strait,Australia. Journal of Archaeological Science29:457–469.
Blomster, J. P., H. Neff, and M. D. Glascock. 2005.Olmec pottery production and export in an-cient Mexico determined through elementalanalysis. Science 307(5712):1068–1072.
Braje, T. J., D. J. Kennett, J. M. Erlandson, andB. J. Culleton. 2007. Trans-Holocene subsis-tence changes and human impacts on a ma-rine ecosystem on Santa Rosa Island, California.American Antiquity 72(4):735–756.
Bronk Ramsey, C. 2013. OxCal v. 4.2. Re-trieved from https://c14.arch.ox.ac.uk/login/login.php?Location=/oxcal/OxCal.html.
Broughton, J. M. and F. E. Bayham. 2003.Showing off, foraging models, and the as-cendance of large game hunting in theMiddle Archaic. American Antiquity 68(4):783–789.
Broughton, J. M. and J. F. O’Connell. 1999. Onevolutionary ecology, selectionist archaeology,and behavioral ecology. American Antiquity64(1):153–165.
Brush, C. F.1969. A contribution to the archaeol-ogy of coastal Guerrero, Mexico. Ph.D. Disser-tation. New York: Columbia University.
Charnov, E. L. 1967. Optimal foraging, theMarginal Value Theorem. Theoretical Popula-tion Biology 9:129–136.
Claassen, C. L. 1986. Temporal patterns in marineshellfish-species use along the Atlantic coast inthe southeastern United States. SoutheasternArchaeology 5:120–137.
Clark, J.E. andM.Blake.1994.Powerandprestige:Competitive generosity and the emergence ofrank in lowland Mesoamerica. In FactionalCompetition and Political Development inthe New World (E. M. Brumfiel and J. W. Fox,eds.):17–30. Cambridge: Cambridge UniversityPress.
Coan, E. V., P. V. Scott, F. R. Bernard. 2000. Bi-valve seashells of western North America: Ma-rine bivalve mollusks from Arctic Alaska toBaja California. Santa Barbara Museum of Nat-ural History Monographs, Studies in Biodiver-sity: no. 2. Santa Barbara: Museum of NaturalHistory.
Cyphers, A. 1996. Reconstructing Olmec life atSan Lorenzo. In Olmec Art of Ancient Mexico,edited by (E. P. B. Benson, B. de la Fuente, andM. Castro-Leal, eds.):61–71. Washington, DC:National Gallery of Art.
Cyphers, A., T. Murtha, J. Borstein, J. Zurita-Noguera, R. Lunagomez, S. Symonds, G.Jimenez, M. Arturo Ortiz, and J. M. Figueroa.2007–2008. Arqueologıa digital en la primeracapital olmeca, San Lorenzo. Thule 22–25:121–144.
de Boer, W. F., T. Pereira, and A. Guissamulo.2000. Comparing recent and abandoned shellmiddens to detect the impact of human ex-ploitation on the intertidal ecosystem. AquaticEcology 34:287–297.
Doolittle, W. E. 1990. Canal Irrigation in Prehis-toric Mexico: The Sequence of TechnologicalChange. Austin: University of Texas Press.
Erlandson, J. M. 1988. The role of shellfish inprehistoric economies: A protein perspective.American Antiquity 53:102–109.
Erlandson, J. M. 2001. The archaeology of aquaticadaptations: Paradigms for a new millennium.Journal of Archaeological Research 9:287–350
Erlandson, J. M., T. J. Braje, T. C. Rick, N. P.Jew, D. J. Kennett, N. Dwyer, and J. Watts.2011. 10,000 years of human predation andsize changes in the owl limpet (Lottia gigan-tea) on San Miguel Island, California. Journalof Archaeological Science 38:1127–1134.
Erlandson, J. M. and T. C. Rick. 2008. Archaeol-ogy, marine ecology, and human impacts onmarine environments. In Human Impacts onAncient Marine Ecosystems (T. C. Rick and J.
200 VOLUME 9 • ISSUE 2 • 2014
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Human Ecology of Shellfish Exploitation in Mexico
M. Erlandson, eds.):1–20. Berkeley: Universityof California Press.
Erlandson, J. M., T. C. Rick, and R. Vellanoweth.2004. Human impacts on ancient environ-ments: A case study from California’s North-ern Channel Islands. In Voyages of Discovery:the Archaeology of Island (S. M. Fitzpatrick,ed.):51–84. Westport: Praeger.
Flannery, K. V. 1973. The origins of agriculture.Annual Review of Anthropology 2:271–310.
Flannery, K. V. 1986. Guila Naquitz: Archaic For-aging and Early Agriculture in Oaxaca, Mex-ico. Orlando: Academic Press Inc.
Flannery, K. V. and J. Marcus. 2000. Forma-tive Mexican chiefdoms and the myth of the“Mother Culture”. Journal of AnthropologicalArchaeology 19:1–37.
Glassow, M. A. and L. R. Wilcoxon. 1988. Coastaladaptations near Point Conception California,with particular regard to shellfish exploitation.American Antiquity 53:36–51.
Gomez, J., D. J. Kennett, H. Neff, M. D. Glascock,and B. Voorhies. 2011. Early formative potteryproduction, mobility, and exchange on the Pa-cific Coast of Southern Mexico. Journal of Is-land and Coastal Archaeology 6:333–350.
Hawkes, K. and J. F. O’Connell. 1992. On op-timal foraging models and subsistence transi-tions. Current Anthropology 33(1):63–66.
Hawkes, K., J. F. O’Connell, and L. Rogers. 1997.The behavioral ecology of modern hunter gath-erers, and human evolution. Trends in Ecologyand Evolution 12:29–32.
Hooke, R. L., 2000. On the history of humans asgeomorphic agents. Geology 28:843–846.
Kaplan, H. and K. Hill. 1992. The evolutionaryecology of food acquisition. In EvolutionaryEcology and Human Behavior (E. A. SmithandB.Winterhalder, eds.):167–201.NewYork:Walter de Gruyter.
Keen, A. M. 1971. Seashells of Tropical WestAmerica: Marine Mollusks from Lower Cali-fornia to Peru. Stanford: Stanford UniversityPress.
Kennett, D. J. 2005. The Island Chumash: Behav-ioral Ecology of a Maritime Society. Berkeley:University of California Press.
Kennett, D. J. 2012. Archaic Period Foragers andFarmers in Mesoamerica. In Oxford Handbookof Mesoamerican Archaeology (D. L. Nicholsand C. A. Pool eds.):141–150. Oxford: OxfordUniversity Press.
Kennett, D. J., B. J. Culleton, B. Voorhies, and J. R.Southon. 2011. Bayesian analysis of high preci-sion AMS 14C dates from a Prehistoric Mexicanshellmound. Radiocarbon 53(2):101–116.
Kennett D. J. and B. Voorhies. 1996. Oxygen iso-topic analysis of archaeological shells to detect
seasonaluseofwetlandsonthesouthernPacificcoast of Mexico. Journal of Archaeological Sci-ence 23:689–704.
Kennett, D. J., B. Voorhies, J. Iriarte, J. G. Jones,D. Piperno, M. T. Ramırez Herrera, and T. A.Wake. 2004. Avances en el Proyecto Arcaico-Formativo: Costa de Guerrero. Mexico City:Estregado al Instituto Nacional de AntropologıaeHistoria.Manuscript inpossessionofD. J.Ken-nett, Guerrero, Mexico.
Kennett, D. J., B. Voorhies, and D. Martorana.2006. An ecological model for the origins offood production on the Pacific coast of south-ern Mexico. In Behavioral Ecology and theTransition to Agriculture (D. J. Kennett andB. Winterhalder, eds.):103–136. Berkeley andLos Angeles: University of California Press.
Kennett, D. J., B. Voorhies, T. A. Wake, andN. Martınez. 2008. Human impacts on marineecosystems in Guerrero, Mexico. In HumanImpacts on Marine Ecosystems (T. C. Rick andJ. M. Erlandson, eds.):103–124. Berkeley: Uni-versity of California Press.
Kennett, D. J., B. Winterhalder, J. Bartruff, andJ. M. Erlandson. 2009. An ecological model forthe emergence of institutionalized social hierar-chies on California’s Northern Channel Islands.In Pattern and Process in Cultural Evolution(S. Shennan, ed.):69–80. Berkeley: Universityof California Press.
Klein, R. G., G. Avery, K. Cruz-Uribe, D. Halkett,J. E. Parkington, T. Steele, T. P. Volman, andR. Yates. 2004. The Ysterfontein 1 MiddleStone Age site, South Africa, and early hu-man exploitation of coastal resources. Proceed-ings of the National Academy of the Sciences101:5708–5715.
Lentz, D. L. 2000. Anthropocentric food webs inthe precolumbian Americas. In Imperfect Bal-ance: Landscape Transformations in the Pre-columbian Americas (D.L. Lentz, ed.):89–119.New York: Columbia University Press.
Madsen, D. B. and D. N. Schmitt. 1998. Mass col-lecting and the diet breadth model: A GreatBasin example. Journal of Archaeological Sci-ence 25:445–455.
Mannino, M. A. and K. D. Thomas. 2002. De-pletion of a resource? The impact of prehis-torichumanforagingon intertidalmollusccom-munities and its significance for human settle-ment, mobility and dispersal. World Archaeol-ogy 33(3):452–474.
Manzanilla Lopez, R. A. 2000. La Region Ar-queologica de la Costa Grande de Guerrero:Su Definicion a traves de la OrganizacionSocial y Territorialidad Prehispanicas. Ph.D.Dissertation. Mexico City: Escuela Nacional deAntropologıa e Historia.
JOURNAL OF ISLAND & COASTAL ARCHAEOLOGY 201
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016

Carley B. Smith et al.
Manzanilla Lopez, R. A., A. Talavera Gonzalez, andE. Rogriguez Sanchez. 1991. Informe Tecnicode Campo de la Primavera Etapa del Proyectode Investigacion y Salvamento Arqueologicoen Puerto Marquez, Estado Guerrero. MexicoCity: Entregado al INAH Mexicanos y Cen-troamericanos, Gobierno del Estado de Guer-rero and Instituto Nacional de Antropologıa eHistoria.
Milner, N., Barrett, J., and Welsh, J. 2007. Marineresource intensification in Viking Age Europe:the molluscan evidence from Quoygrew,Orkney. Journal of Archaeological Science34:1461–1472.
Nagaoka, L. 2002. Explaining subsistence changein southern New Zealand using foraging theorymodels. World Archaeology 34(1):84–102.
Neff, H., J. Blomster, M. D. Glascock, R. L. Bishop,M. J. Blackman, M. D. Coe, G. L. Cowgill,R. A. Diehl, S. Houston, et al. 2006. Methodolog-ical issues in the provenance investigation ofearly formative Mesoamerican ceramics. LatinAmerican Antiquity 17(1):54–76.
Parker, R. H. 1964. Zoogeography and ecologyof macro-invertebrates of the Gulf of Californiaand continental slope of Western Mexico. InMarine Geology of the Gulf of California: ASymposium (T. H. van Andel and G. G. Shor,Jr., eds.):331–376. The American Associationof Petroleum Geologists, Memoir 3.
Piperno, D. R. 2006. The origins of plant cul-tivation and domestication in the neotropics.In Behavioral Ecology and the Transition toAgriculture (D. J. Kennett and B. Winterhalder,eds.):137–166. Berkeley and Los Angeles: Uni-versity of California Press.
Poteate, A. F., and S. M. Fitzpatrick. 2013. Testingthe efficacy and reliability of common zooar-chaeological sampling strategies: A case studyfrom the Caribbean. Journal of ArchaeologicalScience 40:3693–3705.
Raab, M. 1992. An optimal foraging analysis of pre-historic shellfish collecting on San ClementeIsland, California. Journal of Ethnobiology12:63–80.
Reitz, E. J. and E. S. Wing, 1999. Zooarchaeology.Cambridge: Cambridge University Press.
Rick, T. C. and J. M. Erlandson. 2009. Coastal ex-ploitation. Science 325:952–953.
Rick, T. C., J. M. Erlandson, R. L. Vellanoweth,and T. J. Braje. 2005. From Pleistocene marinersto complex hunter-gatherers: The archaeologyof the California Channel Islands. Journal ofWorld Prehistory 19:169–228.
Rosenswig, R. M. 2009. The Beginnings ofMesoamerican Civilization: Inter-RegionalInteraction and the Olmec. Cambridge: Cam-bridge University Press.
Smith, B. D. 2001. Low-level food production.Journal of Archaeological Research 9(1):1–43.
Smith, C. B., D. J. Kennett, T. A. Wake, andB. Voorhies. 2007. Prehistoric sea turtle hunt-ing on the Pacific coast of Mexico. Journalof Island & Coastal Archaeology 2(2):231–235.
Smith, E. A. 1983. Anthropological applicationsof optimal foraging theory: A critical review.Current Anthropology 24(5):625–640.
Stephens, D. W. and J. R. Krebs. 1986. Forag-ing Theory. Princeton: Princeton UniversityPress.
Thacker, H. B. 2011. Intensification of shellfishexploitation: evidence of species-specificdeviation from traditional expectations.Journal of Archaeological Science 38:2596–2605.
Thomas, D. H. 2008. Native American Land-scapes of St. Catherines Island, Georgia: Theo-retical Framework. New York: American Mu-seum of Natural History, Anthropologicial Pa-pers, Number 88.
Voorhies, B. 1996. The transformation fromforaging to farming in lowland Mesoamer-ica. In The Managed Mosaic: Ancient MayaAgriculture and Resource Use (S. L. Fedick,ed.):17–29. Salt Lake City: University of UtahPress.
Voorhies, B. 2004. Coastal Collectors in theHolocene: The Chantuto People of SouthwestMexico. Gainesville: University Press of Florida.
Webster, D. 2002. The Fall of the Ancient Maya.London: Thames and Hudson.
Winterhalder, B. and D. J. Kennett. 2006. Behav-ioral ecology and the transition from huntingand gathering to agriculture. In BehavioralEcology and the Transition to Agriculture(D. J. Kennett and B. Winterhalder, eds.):1–21.Berkeley and Los Angeles: University of Califor-nia Press.
Winterhalder, B., D. J. Kennett, M. Grote, andJ. Bartruff 2010. A Bayesian approach to idealfree settlement on California’s Northern Chan-nel Islands. Journal of Anthropological Ar-chaeology 29:469–490.
Winterhalder, B. and E. A. Smith. 2000. Analyz-ing adaptive strategies: Human behavioral ecol-ogyat twenty-five.EvolutionaryAnthropology9:51–72.
Yesner, D. R. 1980. Maritime hunter-gathererecology and prehistory. Current Anthropology21(6):727–750.
Zeanah, D. W. 2004. Sexual division of labor andcentral place foraging: A model for the CarsonDesert of western Nevada. Journal of Anthro-pological Archaeology 23:1–32.
202 VOLUME 9 • ISSUE 2 • 2014
Dow
nloa
ded
by [
Penn
Sta
te U
nive
rsity
] at
07:
59 0
8 Fe
brua
ry 2
016



![[Prehistoric America] - ScienceViews.com](https://static.fdokumen.com/doc/165x107/6333b0f7a6138719eb0abae5/prehistoric-america-scienceviewscom.jpg)
















