
How environment and grazing influence floristic composition of dry Puna in the southern Peruvian...
20
How environment and grazing influence floristic composition of dry Puna in the southern Peruvian Andes Andrea Catorci, Camerino, Italy, José Luis Velasquez, Arequipa, Peru, Sabrina Cesaretti, Luca Malatesta and Federico Maria Tardella, Camerino, Italy and Horacio Zeballos, Arequipa, Peru With 5 figures, 4 tables and 3 electronic appendices Abstract: High mountain environments located in the tropics account for as much as 10 % of the total surface area of high moun- tain ecosystems worldwide, yet their ecology has been studied much less thoroughly than outside the tropics. The dry Puna is the largest ecosystem of the high tropical Andes and one of the leading biodiversity hotspots worldwide. In this high mountain envi- ronment, the main stress factors are the long period of water shortage, high degree of potential evapotranspiration, wind erosion, low soil nutrient content, and extreme thermal variation. The aim of this study was to deepen our understanding of the relation between the dry Puna species composition and diversity, and environmental/anthropogenic variables. Species cover was deter- mined and soil samples were analysed from 121 plots, along altitudinal transects located between 3,900 and 4,900 m a.s.l. in South Peru. The data were statistically analysed by means of Multivariate regression tree analysis, Indicator Species analysis and Redun- dancy analysis. The hypothesis that in the dry Puna, altitudinal gradient, aspect and rockiness are key factors that mainly determine the floristic composition of the vegetation was confirmed. The local soil features emerged as a secondary driver in the plant com- munity composition. Moreover, disturbance intensity appeared to be a main factor in determining changes in the plant community diversity, also modifying the site ecology (nutrient content, pH) and the structure of ecosystems (from grass dominated systems to dwarf shrub-dominated communities). High grazing intensities caused the spread of species with avoidance strategies such as thorny dwarf shrubs (Tetraglochin cristatum), annual and prostrate plants. Harsh conditions (thin and coarse soil, severe solar ra- diation, low temperatures and thermal fluctuations) promoted the spread of cushion plants (Pycnophyllum sp. pl.), while aspect and soil features (pH, silt % and potassium content) shaped the species composition of Festuca orthophylla tall grassland, the dominant plant community in the dry Puna. Keywords: arid environments, facilitation processes, grazing disturbance, nurse species, soil features, South Peru Abbreviations: CVRE – Cross-Validation Relative Error; ISA – Indicator Species Analysis; IV – Indicator Value; MRT – Multi- variate Regression Trees; tb-RDA – transformation-based Redundancy Analysis. © 2014 Gebrüder Borntraeger, 70176 Stuttgart, Germany DOI: 10.1127/0340-269X/2014/0044-0577 www.borntraeger-cramer.de 0340-269X/2014/0044-0577 $ 8.10 Introduction Although high mountain environments in the tropics account for as much as 10 % of the total surface area of the high mountain ecosystems worldwide (Körner 2003), the plant community ecology of tropical alpine environments has been studied much less thoroughly than that of alpine ecosystems outside the tropics (An- thelme & Dangles 2012). The tropical Andes are in- cluded among the leading biodiversity hotspots world- wide (Myers et al. 2000) and one of their largest pastoral ecosystems is the “dry Puna”, which has a unique as- semblage of shrubs, grass tussocks, cushion, and mat- forming species (SMITH 1994). This pastoral landscape is found in northern Chile and Argentina, southern Peru and western Bolivia (Reese & Liu 2005, Galán de Mera et al. 2012). Dry Puna differs from the other types of Puna because of its lower annual rainfall (less than 400 mm yr -1 ) that leads to an 8/9-month long dry sea- son. The region lies at an elevation of 3,500–5,000 m a.s.l. and the vegetation supports herds of wild and do- mesticated camelids, which form the social and eco- nomic core of pastoral societies here (Bowman 1990, Caro 1992). Moreover, locals intensively harvest many of the woody species for fuel and medicinal use (Hodge 1960, Wickens 1995). Another traditional management practice is the burning of pastures to increase forage productivity and quality (Monteiro et al. 2011). Thomas et al. (2010) demonstrated that the species di- versity of dry Puna in the Bolivian Andes is predomi- nantly determined by altitude and that the abundance of species is associated with anthropogenic pressure con- nected to grazing and controlled fires. Arroyo et al. (1988) found a positive relation between the dominance of woody growth forms and increasing aridity and be- tween the dominance of longer lived and more deeply rooted plants and decreasing temperatures. Moreover, Lambrinos et al. (2006) stated that in the Puna of Chile both species richness and diversity in growth form in- creased with the degree of rockiness and on more xeric northerly and easterly slopes. Xeric, rocky sites sup- ported the greatest overall abundance of cushion forms, Phytocoenologia Vol. 44 (2014), Issue 1-2, 103–119 Article Published online February 2014 eschweizerbart_xxx
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of How environment and grazing influence floristic composition of dry Puna in the southern Peruvian...

How environment and grazing influence floristic composition of dry Puna in the southern Peruvian Andes
Andrea Catorci Camerino Italy Joseacute Luis Velasquez Arequipa Peru Sabrina Cesaretti Luca Malatesta and Federico Maria Tardella Camerino Italy and Horacio Zeballos Arequipa Peru
With 5 figures 4 tables and 3 electronic appendices
Abstract High mountain environments located in the tropics account for as much as 10 of the total surface area of high moun-tain ecosystems worldwide yet their ecology has been studied much less thoroughly than outside the tropics The dry Puna is the largest ecosystem of the high tropical Andes and one of the leading biodiversity hotspots worldwide In this high mountain envi-ronment the main stress factors are the long period of water shortage high degree of potential evapotranspiration wind erosion low soil nutrient content and extreme thermal variation The aim of this study was to deepen our understanding of the relation between the dry Puna species composition and diversity and environmentalanthropogenic variables Species cover was deter-mined and soil samples were analysed from 121 plots along altitudinal transects located between 3900 and 4900 m asl in South Peru The data were statistically analysed by means of Multivariate regression tree analysis Indicator Species analysis and Redun-dancy analysis The hypothesis that in the dry Puna altitudinal gradient aspect and rockiness are key factors that mainly determine the floristic composition of the vegetation was confirmed The local soil features emerged as a secondary driver in the plant com-munity composition Moreover disturbance intensity appeared to be a main factor in determining changes in the plant community diversity also modifying the site ecology (nutrient content pH) and the structure of ecosystems (from grass dominated systems to dwarf shrub-dominated communities) High grazing intensities caused the spread of species with avoidance strategies such as thorny dwarf shrubs (Tetraglochin cristatum) annual and prostrate plants Harsh conditions (thin and coarse soil severe solar ra-diation low temperatures and thermal fluctuations) promoted the spread of cushion plants (Pycnophyllum sp pl) while aspect and soil features (pH silt and potassium content) shaped the species composition of Festuca orthophylla tall grassland the dominant plant community in the dry Puna
Keywords arid environments facilitation processes grazing disturbance nurse species soil features South Peru
Abbreviations CVRE ndash Cross-Validation Relative Error ISA ndash Indicator Species Analysis IV ndash Indicator Value MRT ndash Multi-variate Regression Trees tb-RDA ndash transformation-based Redundancy Analysis
copy 2014 Gebruumlder Borntraeger 70176 Stuttgart GermanyDOI 1011270340-269X20140044-0577
wwwborntraeger-cramerde0340-269X20140044-0577 $ 810
Introduction
Although high mountain environments in the tropics account for as much as 10 of the total surface area of the high mountain ecosystems worldwide (Koumlrner 2003) the plant community ecology of tropical alpine environments has been studied much less thoroughly than that of alpine ecosystems outside the tropics (An-thelme amp Dangles 2012) The tropical Andes are in-cluded among the leading biodiversity hotspots world-wide (Myers et al 2000) and one of their largest pastoral ecosystems is the ldquodry Punardquo which has a unique as-semblage of shrubs grass tussocks cushion and mat-forming species (SMITH 1994) This pastoral landscape is found in northern Chile and Argentina southern Peru and western Bolivia (Reese amp Liu 2005 Galaacuten de Mera et al 2012) Dry Puna differs from the other types of Puna because of its lower annual rainfall (less than 400 mm yr-1) that leads to an 89-month long dry sea-son The region lies at an elevation of 3500ndash5000 m asl and the vegetation supports herds of wild and do-
mesticated camelids which form the social and eco-nomic core of pastoral societies here (Bowman 1990 Caro 1992) Moreover locals intensively harvest many of the woody species for fuel and medicinal use (Hodge 1960 Wickens 1995) Another traditional management practice is the burning of pastures to increase forage productivity and quality (Monteiro et al 2011)
Thomas et al (2010) demonstrated that the species di-versity of dry Puna in the Bolivian Andes is predomi-nantly determined by altitude and that the abundance of species is associated with anthropogenic pressure con-nected to grazing and controlled fires Arroyo et al (1988) found a positive relation between the dominance of woody growth forms and increasing aridity and be-tween the dominance of longer lived and more deeply rooted plants and decreasing temperatures Moreover Lambrinos et al (2006) stated that in the Puna of Chile both species richness and diversity in growth form in-creased with the degree of rockiness and on more xeric northerly and easterly slopes Xeric rocky sites sup-ported the greatest overall abundance of cushion forms
Phytocoenologia Vol 44 (2014) Issue 1-2 103ndash119 ArticlePublished online February 2014
phyto_44_1_2_103_119_Catorciindd 103phyto_44_1_2_103_119_Catorciindd 103 120314 0713120314 0713
eschweizerbart_xxx
104 A Catorci et al
while less rocky sites with deeper soil supported a greater abundance of grasses and shrubs
Notwithstanding the previous investigations more in-sight in the relation between dry Puna species composi-tion and environmentalanthropogenic variables and more information about the soil conditions of dry grass-lands are needed
Moreover in order to effectively address biodiversity conservation issues it is vital to identify the driving forces that affect plant community composition dynam-ics and spatial distribution (Grime 2001 2006 Biondi 2011 Catorci et al 2011a) Indeed stress and disturbance intensities influence species co-existence (Grime 2001) affecting the competitive ability of species and thus the diversity of plant communities (Lauenroth amp Aguilera 1998) and their distribution (Catorci amp Gatti 2010 Blasi et al 2012)
Given these findings we hypothesised that topo-graphic factors (altitude slope aspect and slope angle) degree of rockiness and soil physicalchemical features as well as the disturbance intensity due to grazing pres-sure and fire affect the dry Puna diversity and composi-tion as well as the abundance of plant growth forms and of potential nurse species (also known as facilitators or benefactors) ie plants which create more favorable mi-cro-habitats than those available in their surroundings for the germination establishment and survival of other plants also known as beneficiaries target or facilitated species (Valiente-Banuet et al 1991 Tewksbury amp Lloyd 2001) In fact it was stated that in harsh environments facilitation plays a key role in shaping the richness and diversity of plant communities (Pugnaire amp Luque 2001 Armas amp Pugnaire 2005 Brooker 2006 Catorci et al 2011b Kikvidze et al 2011)
Study area
The research was carried out in the Salinas and Aguada Blanca National Reserve (Arequipa and Moquegua De-partments ndash Fig 1) in the southern Peruvian Andes (cen-tral coordinates 16deg 04rsquo 59rdquo S 71deg 24rsquo 15rdquo W) This area is part of the Andean altiplano Although the majority of the Altiplano is a relatively flat region some areas have an extremely complex topography Numerous mountain ranges characterise the landscape with peaks reaching 6000 m asl (Mount Chachani in the study area)
The research area is characterised by volcanic struc-tures covered by deposits of volcanic ash and saline soils in depressed areas Precipitation is markedly seasonal with over 80 of the annual rainfall (350ndash400 mm) fall-ing between December and March (Vuille et al 1998) soil moisture ranges from 0 (dry season) to below 10 in the rainy season (Montenegro et al 2010) The mean annual temperature is 3ndash4deg C with sharp diurnal and annual var-iations from freezing to temperatures of more than
30deg C From a biogeographic point of view this area is included in the Neotropical kingdom Andean region Puna region Oruro-Arequipenian province Huaran-cante-Imata sector while for a bioclimatic characterisa-tion we can include it in the orotropical belt with a semi-arid-dry humidity interval (Rivas-Martiacutenez amp Tovar 1982 Galaacuten de Mera et al 2009 2010) In the Salinas and Aguada Blanca National Reserve the Puna ecosystem commonly occurs between 38003900 and 48005000 m asl (Montenegro et al 2010)
The study area comprises the typical dry Puna land-scape of Latin America mostly dominated by bunch grasses like Festuca orthophylla or Jarava ichu and also composed of resinous shrubs (eg Parastrephia quad-rangularis and P lucida) and cushion plants (eg Pycno-phyllum molle P weberbaueri) From a phytosociologi-cal viewpoint according to Galaacuten de Mera et al (2002 2003 2009) the vegetation of the study area is included in the Parastrephietalia lepidophyllae Navarro 1993 order and in the Calamagrostietea vicunarum Rivas-Martiacutenez amp Tovar 1982 class
The human settlements are limited to small villages and isolated farms around which the economic activities of inhabitants (mainly breeding of domestic camelids) are concentrated These areas are mainly located on the least steep slopes of the altiplano while the surrounding high mountain slopes are not utilised by humans and host wild camelids
Materials and methods
Data collection
Data were collected at altitudes ranging from 3900 to 4900 m asl avoiding wetland (ldquobofedalesrdquo) since these areas lie outside the aim of our study (focused on dry grasslands) even if they represent an important habitat for alpaca breeding in the Reserve Rocky cliffs and streams were avoided as well We laid 160 sampling plots of 100 m2 each (10 m x 10 m) along 20 a priori chosen 2 km long transects The distance between consecutive plots was 250 m Transects were oriented along the direc-tion of maximum slope angle and selected to sample the heterogeneity of landforms and substrates 39 plots were not sampled because they laid on roads hamlets open quarries and wetlands or because they included latrine areas (small areas used by camelids for defecation and urination) In each of the remaining 121 plots the cover percentage values of species were estimated (based on a visual evaluation of vertically projected cover of species on the ground) using the Braun-Blanquet (1964) scale (+ lt 1 1 2ndash5 2 6ndash25 3 26ndash50 4 51ndash75 5 76ndash100 ) The collected specimens are stored at ldquoBie-nestar ndash Centro de Investigacioacuten para la Promocioacuten de los Pueblosrdquo of Arequipa (Peru)
phyto_44_1_2_103_119_Catorciindd 104phyto_44_1_2_103_119_Catorciindd 104 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 105
Fig 1 The study area (Salinas and Aguada Blanca National Reserve) located within the Arequipa and Moquegua Departments in the southern Peruvian Andes
phyto_44_1_2_103_119_Catorciindd 105phyto_44_1_2_103_119_Catorciindd 105 120314 0713120314 0713
eschweizerbart_xxx
106 A Catorci et al
For each releveacute we collected soil samples and data on altitude (m asl) slope aspect (azimuth degrees) slope angle (vertical degrees) and rockiness Within each plot 5 separate soil samples were taken at a depth of 2 to 20 cm below the soil surface and combined to one bulked sam-ple Grazing intensity and occurrence of fire were deter-mined by more than ten interviews with local villagers and the rangers of the Salinas and Aguada Blanca Na-tional Reserve Releveacutes were carried out at the end of the rainy season (March 2010) when soil moisture is highest and the highest number of species can be observed
Data on the following plant traits were collected by field observations and through consultation of the Her-barium of the Universidad Nacional de San Agustin (HUSA) of Arequipa (Peru) and of the Herbarium at the Universidad Nacional Mayor de San Marcos (USM) of Lima (Peru) annual and perennial life span graminoid species non-graminoid species upright forb (plant with leaves distributed on upright stems or branches) hemiro-sulate forb (plant with leaves distributed both at ground level and raised from the ground) prostrate forb (plant with leaves distributed only at the ground level) tall shrub (gt 15 cm) dwarf shrub (lt 15 cm) cushion and succulent plants
As it has been demonstrated that in harsh environ-ments positive plant-plant spatial interaction between nurse and beneficiary species play a key role in shaping the community richness and diversity (Armas amp Pug-naire 2005 Catorci et al 2011b Kikvidze et al 2011) fol-lowing several authors (Rebollo et al 2002 Maestre amp Cortina 2005 Padilla amp Pugnaire 2006 Arredondo-Nu-ntildeez et al 2009 Monteiro et al 2011) we chose tall grass tussocks tall shrubs and cushions as potential nurse plants considering the potential nurse role as an addi-tional functional trait
Species nomenclature mainly follows Brako amp Zarucchi (1993) Later taxonomic changes were checked by consult-ing IPNI (httpwwwipniorgindexhtml) Tropicos (httpwwwtropicosorg) and other botanical studies from close geographic areas (eg Araraki amp Cano 2003 Schwarzer et al 2010 Montesinos 2011 Montesinos et al 2012)
Data analysis
Soil samples were analysed by the Instituto Nacional de Innovacion Agraria (INIA) soil analysis laboratory of Arequipa (Ministerio de Agricultura y Riego del Peru) The following soil parameters were determined (the methods applied are indicated in parentheses) texture ie sand loam and clay (Buoyuocosrsquo method) or-ganic matter (modified Walkley Blackrsquos method) ni-trogen (Micro Kjeldahlrsquos method) phosphorus in ppm (modified Olsenrsquos method 05 M NaHCO3 extrac-tion at pH 88) potassium in ppm (flame photometry
by ammonium acetate at pH 70) CaCO3 (Wesemaelrsquos method) pH (by potentiometer in a 125 soilwater sus-pension)
To assess floristic composition the species cover val-ues were transformed into percentages using the central values of the Braun-Blanquet scale (Podani 2007) Fol-lowing Warren (2008) aspect azimuth was converted from the 0ndash360 compass scale to a linear (0ndash180) scale giving northerly aspect a value approaching 0 and south-erly aspect a value approaching 180 Disturbance inten-sity was assessed using a 4-level scale as follows 1) ab-sence of grazing (absent) 2) presence of wild camelids and sporadically small herds of domestic camelids (low) 3) grazing of large herds of domestic camelids (moder-ate) 4) grazing of large herds of domestic camelids and periodic fires (strong)
In order to assess the influence of environmental vari-ables and disturbance intensity on the species assemblage and identify homogeneous groups of releveacutes a con-strained clustering using Multivariate regression tree (MRT) analysis (Dersquoath 2002) with 100 iterations and 29 cross-validation groups was executed on two matrices namely ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo Prior to MRT analy-sis species data have been transformed using chord trans-formation (Legendre amp Gallagher 2001) which expresses the data as relative abundances per sites This removes from the data the total abundance per site which is the response of the species to the total productivity of the sites (Borcard et al 2011) The result of MRT analysis is a tree whose ldquoleavesrdquo (terminal groups of releveacutes) are com-posed of subsets of releveacutes chosen to minimize the within-group sum of squares and each partition of the tree is defined by a threshold value of one explanatory variable (Borcard et al 2011) Cross-validation is a re-sampling analysis repeated a number of times for succes-sive and independent divisions of the data (in this case species cover) into random test groups of objects (rel-eveacutes) (Legendre amp Legendre 2012) An overall error sta-tistic (cross-validation relative error CVRE) was com-puted for each test group and partition size (number of groups) to choose the optimal size of the tree CVRE varies from zero for a perfect set of predictors chosen for the splits of a tree to close to one for poor predictors For each partition size the mean and standard error of all CVRE estimates was computed Generally the smallest tree (ie a tree with fewer splits) within one standard er-ror of the smallest CVRE value is selected (Breiman et al 1984) but one may select the tree that has the smallest CVRE value as well (Legendre amp Legendre 2012)
To provide a general overview of the ecological charac-teristics of the whole releveacute data set we calculated the 1st and 3rd quartiles for each environmental variable The mean value and the standard deviation of the same varia-bles were calculated for each releveacute group that emerged from the MRT analysis
phyto_44_1_2_103_119_Catorciindd 106phyto_44_1_2_103_119_Catorciindd 106 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 107
The species diversity of each group was assessed by calculating the Shannon-Wiener (Hrsquo) and Gini-Simpson (D) diversity indices while the dissimilarity between the groups was computed through the Jaccard distance index (Magurran 1988) In order to investigate the species com-position of the groups we performed the Indicator Spe-cies Analysis (ISA) a non-parametric method for identi-fying those items (speciestrait) that show significantly preferential distribution (frequency and abundance) compared to an a priori treatment group An indicator value is calculated by multiplying the relative abundance of each item in a particular group and the relative fre-quency of the item occurrence in the sample of that group (Dufrecircne amp Legendre 1997) The number of randomized indicator values higher than the observed ones is used to calculate the probability value (McCune amp Grace 2002) The statistical significance (P lt 005) of the observed maximum Indicator Values (IV) was tested using permu-tation tests with 4999 iterations Only Indicator Values higher than 30 were considered of interest
To assess the influence of the ecological factors on spe-cies composition and abundance we performed a transfor-mation-based Redundancy Analysis (tb-RDA ndash Legendre amp Gallagher 2001) on the same matrices processed for MRT analysis Disturbance was considered as a factor var-iable with the above-mentioned four levels (absent low moderate strong) thus each of these levels was repre-sented by its centroid instead of an arrow in the RDA plot
To assess the influence of the ecological factors on the trait composition of the plant communities we first transformed trait binary data (presenceabsence) into quantitative data (ie aggregated cover values of each trait) The ldquoreleveacutes x species coverrdquo matrix was multiplied by the ldquospecies x traitrdquo matrix to provide a ldquoreleveacutes x trait coverrdquo matrix (Pakeman et al 2009) which formed the basis for the following analyses Then we ran a tb-RDA on the ldquoreleveacutes x trait coverrdquo matrix using the above-mentioned parameters (environmental factors and distur-bance intensity) as constraining variables
Since the disturbance gradient emerged as the main factor affecting trait composition in dry Puna communi-ties in order to better understand its effect on trait com-position we performed an ISA on the above matrix using MRT groups as grouping variable As the first partition of the regression tree segregated releveacutes characterised by strong disturbance intensity we re-ran ISA comparing releveacutes of this group against all the other ones The statis-tical significance (P lt 005) of the observed maximum In-dicator Values (IV) was tested using permutation tests with 4999 iterations Only Indicator Values higher than 30 were considered of interest
We used the R software (version 2140 ndash R Core Team 2012 ndash httpwwwR-projectorg) and its mvpart labdsv and vegan packages (Oksanen et al 2013) to perform MRT analysis ISA tb-RDA and to calculate diversity and dis-tance indices besides basic statistics
Results
MRT analysis generated two trees with the same CVRE (0641) composed of 4 and 14 leaves respectively (Fig 2) The 4-leaf tree had a smaller standard error (0051) than the 14-leaf tree (0062) and was selected as the best solution more frequently during the cross-validation it-erations Nevertheless to avoid losing information po-tentially useful for the ecological interpretation of the distribution of plant communities as the CVRE of the 14-leaf tree fell within one standard error of the smallest CVRE value we decided to consider this tree as well We recorded 136 species (belonging to 69 genera and 28 fam-ilies) The species composition of each group highlighted by MRT is shown in Fig 3 while the related distance in-dices are given in Appendix A The values of the Shan-non-Wiener and the Gini-Simpson diversity indices are reported in Table 1 The indicator species identified by ISA for groups highlighted by MRT as well as their mean abundances are shown in Table 2 Table 3 indicates the mean the median and the 1st and 3rd quartiles for the eco-logical variables in each group Table 2 shows the output of ISA concerning the assessment of the indicator traits of the four relevegrave groups of the 4-leaf and 14-leaf trees The variable that discriminated between the two branches in the first node of the 4-leaves MRT was grazing distur-bance thus the first leaf (T41) was composed of the re-leveacutes characterised by a high level of disturbance The related plant communities were dominated by the dwarf shrub Tetraglochin cristatum they presented the lowest species diversity (Hrsquo = 163 D = 065) as well as the high-est distances compared to the other leaves Other envi-ronmental features of the T cristatum ndash dominated plant community were (Table 3) high sand percentage and low soil values of organic matter nitrogen phosphorus and potassium The pH value was the highest one compared to the other groups of releveacutes Dwarf shrubs non-grami-noid and annual species were the indicator traits of this group compared with all the other releveacutes jointly consid-ered (Table 4)
The second partition was defined by the percentage of outcropping rocks The first branch of this partition characterised by the lower values of outcropping rocks is in turn divided into two leaves the first (T42) grouping releveacutes located below 4000 m of altitude is dominated by Jarava ichu while Calamagrostis breviaristata domi-nates the second one (T43) grouping the releveacutes placed above 4000 m Both these groups were subjected to in-termediate level of disturbance and were characterised by flat or semi-flat slopes The Jarava ichu ndash dominated plant community was distinguished by low sand percent-age and relatively high organic matter phosphorus and potassium content in soil (Table 3) The pH value was similar to that of groups T43 and T44 groups Grami-noid and hemirosulate forbs as well as succulent and po-
phyto_44_1_2_103_119_Catorciindd 107phyto_44_1_2_103_119_Catorciindd 107 120314 0713120314 0713
eschweizerbart_xxx
108 A Catorci et al
Fig 2 The 4-leaf (a) and 14-leaf (b) Multivariate regression trees for the species data set constrained by their explanatory variables The threshold values shown for each partition of the tree correspond to the mean of the two limit values of the considered variables at the break between the branches The relative abundances of the species are shown in histograms at the tips of the branches with the species in the same order as in the input file the species names and abun-dances are shown in Figure 3 Under each histogram the sum of squared errors for the group and the number of releveacutes in the leaf (n) are indicated Below each tree the residual error (Error) the cross-validation error (CV Error) and standard error (SE) are indi-cated Alt ndash Altitude Asp_deg ndash aspect Dist_L ndash Disturbance level Hum_p ndash organic matter K_ppm ndash Potassium in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt
tential nurse plants were the indicator traits for the T42 group (Table 4)
The C breviaristata ndash dominated plant community was characterized by high sand percentage and low or-ganic matter and potassium content in the soil (Table 3) The pH value was similar to that of groups T43 and T44 Cushion and perennial plants were the indicator traits for group T43 (Table 4)
The releveacutes grouped in the fourth leaf (T44) are char-acterised by a percentage of outcropping rocks greater than 1 (mean cover value 178 ) In this group the dominant species is the graminoid Festuca orthophylla other indicator species being Deyeuxia brevifolia Trise-tum spicatum Nassella depauperata and Deyeuxia rigida (all grass species) Group T44 includes the greatest num-ber of releveacutes (69) and presents the highest diversity (Hrsquo = 282 D = 083) in the 4-leaved tree Other environmen-
tal features of the Festuca orthophylla ndash dominated plant community were (Table 3) the highest slopes altitude and angles among the four groups highlighted by MRT low sand percentage and high values of organic matter nitro-gen and phosphorus in the soil The pH value was similar to that of groups T42 and T43 These pastures were characterised by low disturbance intensity Within the high richness and internal variability of these grasslands particular environmental conditions or treatments caused minor differences in species composition highlighting different plant communities as shown by the 14-leaved MRT (Fig 2)
Despite the high number of groups yielded the val-ues of community diversity indices computed from the pairwise comparison of the 14 leaves were still high (never below 45 ) (Appendix A) The first partition of the Festuca orthophylla ndash dominated grassland was de-
phyto_44_1_2_103_119_Catorciindd 108phyto_44_1_2_103_119_Catorciindd 108 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 109
fined by aspect with a threshold value that can be inter-preted as the boundary between north-east-facing slopes (arid more stressful ndash groups T148-T1412) and south-west-facing ones (cool less stressful ndash groups T1413 and T1414) From a functional point of view the south-facing plots were characterized by a high number of indicator traits (perennial species graminoids hemi-rosulate forbs cushion plants and potential nurses) while no traits emerged as indicator of the plant com-munities growing on the north-facing slopes In addi-tion north-facing groups were characterised by a higher degree of dominance with the most widespread species often representing more than 50 of the total vegeta-tion cover
Altitude gradients and soil parameters defined further partitions for a more detailed differentiation of these two main clusters Among north-facing groups T148 was dominated by Nassella depauperata and had coarser soil as well as higher pH and organic matter values than T149 Groups T1410 T1411 and T1412 were charac-terised by higher altitudes (about 400 m above T148 and T149 ndash Table 3) T1410 was characterised by low distur-bance intensity and showed a more even distribution of species (particularly if compared with T1411) among which several palatable grasses are present (eg Deyeuxia brevifolia) Group T1412 dominated by the shrub spe-cies Chersodoma jodopappa showed a higher percentage of silt and organic matter in soil and steeper slopes with
Fig 3 Pie charts reporting the species composition of the groups defined in the 4-leaf (a) and in the 14-leaf (b) Multivariate regres-sion trees Species abbreviations are explained below Ast_die ndash As-tragalus dielsii Bac_tri ndash Baccharis tricuneata Bel_lon ndash Belloa longifolia Cal_bra ndash Calamagrostis breviaristata Can_aca ndash Calan-drina acaulis Chs_jod ndash Chersodoma jodopappa Cro_ful ndash Crocop-sis fulgens Des_myr ndash Descurainia myriophylla Dey_brf ndash De-yeuxia brevifolia Dey_rig ndash Deyeuxia rigida Dip_mey ndash Diplostephium meyenii Ech_pam ndash Echinopsis pampana Fes_ort ndash Festuca ortophylla Gna_lac ndash Gnaphalium lacteum Hyp_ech ndash Hypochaeris echegarayi Jar_ich ndash Jarava ichu Lpp_qua ndash Lepi-dopyllum quadrangulare Lup_sax ndash Lupinus saxatilis Muh_per ndash Muhlenbergia peruviana Nas_dep ndash Nassella depauperata Nas_nar ndash Nassella nardoides Ntt_tur ndash Nototriche turritella Oen_sp ndash Oenothera sp Par_luc ndash Parastrephia lucida Par_qua ndash Parastre-phia quadrangularis Pla_ser ndash Plantago sericea var lanuginosa Poa_gym ndash Poa gymnantha Poa_pea ndash Poa pearsonii Pyc_mol ndash Pycnophyllum molle Pyc_web ndash Pycnophyllum weberbaueri Sen_cha ndash Senecio chachaniensis Sen_nut ndash Senecio nutans Sti_han ndash Stipa hans-meyeri Tet_cri ndash Tetraglochin cristatum Tri_spi ndash Trise-tum spicatum Wer_pyg ndash Werneria pygmaea
phyto_44_1_2_103_119_Catorciindd 109phyto_44_1_2_103_119_Catorciindd 109 120314 0713120314 0713
eschweizerbart_xxx
110 A Catorci et al
groups yielded by the 4-leaf tree are clearly recognisable in the RDA three-dimensional scatterplot (as highlighted in Fig 4) The RDA performed on functional traits data with the environmental variables as constraints yielded similar results but explained a notably smaller fraction of the total variance (92 adjusted R2 = 056) The varia-bles showing the strongest correlation with the con-strained components coincided with those observed in the first RDA but for most environmental variables the observed correlation is weaker this is the case of altitude aspect percentage of outcropping rocks sand and silt percentage in the soil (Fig 5) Instead for disturbance in-tensity the observed correlation was slightly stronger The functional traits whose variance was best explained by the constrained components were nurse ability and various types of plant forms dwarf shrubs upright forbs non-graminoids hemirosulate forbs and graminoids (Fig 5) Non-graminoid species dwarf shrubs and up-right forbs were related to relatively higher soil pH and strong disturbance intensity nurse species were linked to higher altitudes South-facing slopes relatively lower sand percentages in the soil graminoid species and hemi-rosulate forbs were particularly tied to more fertile soils (higher phosphorus and nitrogen concentrations) and low disturbance intensity (Fig 5) RDA biplot scores are reported in the Appendix C
Discussion
Our results demonstrate that the dry Puna landscape is anything but homogeneous as observed for other semi-arid environments where local factors were detected as key drivers of species assemblages (eg Schlesinger et al 1990 Perelman et al 1997 Diacuteaz et al 1999) The variables showing the strongest correlations with the constrained RDA components were disturbance intensity slope as-pect altitude percentage of outcropping rocks and to a lesser degree soil features (organic matter content pH and potassium concentration) High disturbance inten-sity (which had the best correlation with the first con-strained component) emerged as the main factor shaping the species composition of the dry Puna pastures and was positively correlated with pH dwarf shrubs and non-graminoid plant forms while it was negatively cor-related with organic matternitrogen soil content nurse plant abundance hemirosulate forbs and graminoids Consistently with previous research carried out in semi-arid environments (eg Ares et al 1990 Carilla et al 2012) high grazing intensity promotes the spread of spe-cies with avoidance strategies (Table 4) such as thorny dwarf shrubs (Tetraglochin cristatum) annual species (eg Muhlenbergia peruviana) and small or prostrate plants (eg Nassella nardoides Astragalus dielsii Hy-pochaeris echegarayi) Thorny dwarf shrubs are fostered by herbivory given that the tough and spiny stems of this
Table 1 Values of Shannon and Simpson diversity indices for the releveacute groups of the 4-leaf and 14-leaf Multivariate regression tree
Groups Diversity
Shannon (Hlsquo) Simpson (D)
T41 1628 0653
T42 1831 0626
T43 2176 0765
T44 2822 0829
T141 1511 0582
T142 1611 0688
T143 1831 0626
T144 1714 0633
T145 2366 0822
T146 1913 0763
T147 2348 0824
T148 1316 0598
T149 1540 0574
T1410 2214 0792
T1411 2283 0675
T1412 2149 0714
T1413 2100 0741
T1414 2685 0863
high outcropping rock cover South-facing groups were in turn separated by potassium content in the soil group T1413 with higher concentration of potassium shows a higher coverage of graminoid species (eg Poa pearsonii)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grass Festuca or-thophylla the cushion plants Pycnophyllum molle and P weberbaueri the shrub species Parastrephia lucida and P quadrangularis) This plant community was located near the altitudinal limit of vegetation and characterised by very low disturbance
The Redundancy Analysis executed on species and en-vironmental data explained 638 of the total variance (adjusted R2 = 047) The results of the RDA are in line with the MRT analysis the variables showing the strong-est correlation with the constrained components were disturbance intensity altitude percentage of outcropping rocks slope aspect soil organic matter content pH and potassium concentration (Fig 4) RDA biplot scores are reported in Appendix B It is worth noting that this result coincides with those discriminating for the partitions of the 4-leaf and 14-leaf MRTs (in order of diminishing cor-relation with RDA axes) Another correspondence be-tween MRT and RDA analyses lies in the placement of site points in the RDA graph as a matter of fact the
phyto_44_1_2_103_119_Catorciindd 110phyto_44_1_2_103_119_Catorciindd 110 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 111
Table 2 Indicator species of the releveacute groups highlighted by Indicator Species Analysis for the 4ndashleaf and the 14ndashleaf Multivariate regres-sion tree (MRT) and their respective indicator values and mean cover percentages Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value)
Indicator species 4-leaf MRT 14-leaf MRTMax
GroupIndicator
ValueMean
cover Max
GroupIndicator
ValueMean
cover Nassella nardoides T41 663 199 ndash ndash ndash
Nothoscordum andicola T41 482 08 ndash ndash ndash
Lepidium meyenii T41 412 06 ndash ndash ndash
Astragalus peruvianus T41 315 07 ndash ndash ndash
Tetraglochin cristatum T41 826 544 T141 422 628
Muhlenbergia peruviana T41 675 44 T142 300 46
Cumulopuntia ignescens T42 405 07 ndash ndash ndash
Chenopodium petiolare T42 370 03 ndash ndash ndash
Urocarpidium echinatum T42 350 06 ndash ndash ndash
Nototriche turritella T42 345 24 ndash ndash ndash
Jarava ichu T42 979 598 T143 925 598
Plantago linearis T42 901 30 T143 493 30
Diplostephium meyenii T42 700 23 T143 634 23
Crocopsis fulgens T42 571 16 T143 571 16
Belloa piptolepis T42 592 11 T143 351 11
Senecio phylloleptus T42 490 10 T143 364 10
Hypochaeris echegarayi T43 380 08 ndash ndash ndash
Pycnophyllum weberbaueri T43 342 08 ndash ndash ndash
Junellia minima (Meyen) T43 300 08 ndash ndash ndash
Calamagrostis breviaristata T43 704 430 T144 398 581
Calceolaria inamoena ndash ndash ndash T145 315 08
Astragalus dielsii T43 460 11 T146 321 14
Nassella nardoides ndash ndash ndash T146 484 415
Calandrinia acaulis ndash ndash ndash T146 430 14
Oenothera sp ndash ndash ndash T146 420 14
Festuca orthophylla T44 522 389 ndash ndash ndash
Trisetum spicatum T44 406 08 ndash ndash ndash
Nassella depauperata T44 352 70 T148 860 600
Lupinus saxatilis ndash ndash ndash T148 657 80
Descurainia myriophylla ndash ndash ndash T148 437 133
Erigeron pazensis ndash ndash ndash T148 416 07
Deyeuxia brevifolia T44 464 53 T1410 414 193
Mniodes pulvinata ndash ndash ndash T1410 375 05
Lupinus chlorolepis ndash ndash ndash T1411 364 09
Calamagrostis vicunarum ndash ndash ndash T1412 500 07
Poa gymnantha ndash ndash ndash T1412 485 42
Dielsiochloa floribunda ndash ndash ndash T1412 450 07
Leucheria daucifolia ndash ndash ndash T1412 398 07
Echinopsis pampana ndash ndash ndash T1412 384 42
Perezia ciliosa ndash ndash ndash T1412 329 07
Poa pearsonii ndash ndash ndash T1413 670 237
Deyeuxia rigida T44 316 47 T1414 774 285
Poa aequigluma ndash ndash ndash T1414 453 06
Werneria spathulata ndash ndash ndash T1414 412 08
Luzula racemosa ndash ndash ndash T1414 409 09
Lucilia sp ndash ndash ndash T1414 401 12
Senecio scorzoneraefolius ndash ndash ndash T1414 376 05
Gnaphalium badium ndash ndash ndash T1414 338 10
Pycnophyllum molle ndash ndash ndash T1414 335 95
phyto_44_1_2_103_119_Catorciindd 111phyto_44_1_2_103_119_Catorciindd 111 120314 0713120314 0713
eschweizerbart_xxx
112 A Catorci et al
Table 3 Descriptive statistics of the environmental variables of the groups highlighted by the 4-leaf and 14-leaf Multivariate regression tree 1st and 3rd quartiles are indicated for the whole data set mean and standard deviation (SD) are reported for each group
Group Descriptive statistics
Constraining variables
Site characteristics Soil texture Soil composition
Altitude(m asl)
Aspect (deg)
Slope (deg)
Outcr rocks()
Sand()
Silt ()
Clay ()
Org matt()
N () P (ppm)
K (ppm)
CaCO3 ()
pH
1st quartile 407000 000 200 000 6280 1660 580 089 004 1136 11250 008 540 3rd quartile 447500 9000 2000 1500 7760 2780 910 174 009 2469 23250 075 623
T41 Mean 399700 2363 755 000 7867 1403 1051 071 003 1265 16381 049 652SD 1908 2777 894 000 295 180 1434 033 001 668 3318 035 028
T42 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T43 Mean 410500 2880 364 000 7265 2133 602 110 009 2173 17036 053 542SD 2933 4752 930 000 389 372 237 053 020 1234 6129 051 035
T44 Mean 440287 8739 2061 1787 6698 2444 849 171 008 2219 19384 045 572SD 22159 5242 1431 1192 976 866 304 083 004 1471 11137 039 045
T141 Mean 399833 3938 1208 000 7926 1374 1233 052 003 875 17452 049 666SD 2462 2561 901 000 333 203 1853 024 001 142 3181 032 028
T142 Mean 399500 000 075 000 7778 1445 778 099 004 1850 14775 049 631SD 535 000 104 000 214 141 213 022 001 725 3011 041 010
T143 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T144 Mean 411393 643 064 000 7373 1987 640 098 012 2552 18143 052 529SD 1712 2405 145 000 309 274 306 020 027 1421 5395 062 035
T145 Mean 412667 10500 2167 000 6777 2577 647 210 008 1611 18335 071 549SD 4619 1299 2082 000 224 145 093 017 001 516 13137 017 031
T146 Mean 409500 000 000 000 7385 2128 488 078 005 2370 18117 015 575SD 000 000 000 000 042 026 017 003 001 327 1019 019 018
T147 Mean 406750 7875 425 000 7135 2315 550 110 004 1073 11106 082 551SD 3663 5356 299 000 654 654 081 094 004 698 1577 019 031
T148 Mean 414800 7200 1900 1500 7368 2080 552 135 007 2044 9024 019 630SD 570 1006 224 000 618 567 064 037 002 1111 4791 021 015
T149 Mean 408833 9000 600 1833 6960 2093 947 096 005 1216 18803 054 566SD 5795 000 346 577 763 412 357 067 004 098 12521 057 071
T1410 Mean 458688 4922 2731 1813 7623 1591 787 136 006 2448 17087 042 547SD 22773 4043 1590 680 622 535 164 026 001 1461 9485 047 049
T1411 Mean 439614 5727 1114 2173 6501 2430 1038 147 007 1741 22177 052 583SD 16079 2840 551 1297 832 588 420 053 003 597 10086 035 049
T1412 Mean 445650 9000 4000 550 6025 3248 725 248 013 1653 25562 055 579SD 27365 000 2121 636 078 046 120 049 002 467 26427 040 013
T1413 Mean 421318 15341 3518 2409 5954 3227 819 285 014 2823 28413 025 564SD 11402 2814 1419 1338 1099 981 169 117 006 2824 11501 041 022
T1414 Mean 454300 14850 1600 590 6189 3104 707 183 009 2740 11102 065 571SD 6997 2846 615 1058 589 588 071 063 003 830 2759 027 026
life form discourage animals from biting it (Briske 1996) The observed spread of dwarf shrubs is in accordance with the findings of several studies carried out in semiarid environments (Perelman et al 1997 Swinton amp Quiroz 2003 Carilla et al 2012 Catorci et al 2012b 2013) and in pastoral ecosystems subjected to periodic fires (Liedloff
et al 2001) In semi-arid environments annual life span may be considered an avoidance strategy fostered by high grazing intensity (Diacuteaz et al 1999 McGregor 2002 Catorci et al 2012b) because it allows plants to dynami-cally face the grazing pressure by restarting the growth cycle every year (Diacuteaz et al 1999 2001 McGregor 2002)
phyto_44_1_2_103_119_Catorciindd 112phyto_44_1_2_103_119_Catorciindd 112 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

104 A Catorci et al
while less rocky sites with deeper soil supported a greater abundance of grasses and shrubs
Notwithstanding the previous investigations more in-sight in the relation between dry Puna species composi-tion and environmentalanthropogenic variables and more information about the soil conditions of dry grass-lands are needed
Moreover in order to effectively address biodiversity conservation issues it is vital to identify the driving forces that affect plant community composition dynam-ics and spatial distribution (Grime 2001 2006 Biondi 2011 Catorci et al 2011a) Indeed stress and disturbance intensities influence species co-existence (Grime 2001) affecting the competitive ability of species and thus the diversity of plant communities (Lauenroth amp Aguilera 1998) and their distribution (Catorci amp Gatti 2010 Blasi et al 2012)
Given these findings we hypothesised that topo-graphic factors (altitude slope aspect and slope angle) degree of rockiness and soil physicalchemical features as well as the disturbance intensity due to grazing pres-sure and fire affect the dry Puna diversity and composi-tion as well as the abundance of plant growth forms and of potential nurse species (also known as facilitators or benefactors) ie plants which create more favorable mi-cro-habitats than those available in their surroundings for the germination establishment and survival of other plants also known as beneficiaries target or facilitated species (Valiente-Banuet et al 1991 Tewksbury amp Lloyd 2001) In fact it was stated that in harsh environments facilitation plays a key role in shaping the richness and diversity of plant communities (Pugnaire amp Luque 2001 Armas amp Pugnaire 2005 Brooker 2006 Catorci et al 2011b Kikvidze et al 2011)
Study area
The research was carried out in the Salinas and Aguada Blanca National Reserve (Arequipa and Moquegua De-partments ndash Fig 1) in the southern Peruvian Andes (cen-tral coordinates 16deg 04rsquo 59rdquo S 71deg 24rsquo 15rdquo W) This area is part of the Andean altiplano Although the majority of the Altiplano is a relatively flat region some areas have an extremely complex topography Numerous mountain ranges characterise the landscape with peaks reaching 6000 m asl (Mount Chachani in the study area)
The research area is characterised by volcanic struc-tures covered by deposits of volcanic ash and saline soils in depressed areas Precipitation is markedly seasonal with over 80 of the annual rainfall (350ndash400 mm) fall-ing between December and March (Vuille et al 1998) soil moisture ranges from 0 (dry season) to below 10 in the rainy season (Montenegro et al 2010) The mean annual temperature is 3ndash4deg C with sharp diurnal and annual var-iations from freezing to temperatures of more than
30deg C From a biogeographic point of view this area is included in the Neotropical kingdom Andean region Puna region Oruro-Arequipenian province Huaran-cante-Imata sector while for a bioclimatic characterisa-tion we can include it in the orotropical belt with a semi-arid-dry humidity interval (Rivas-Martiacutenez amp Tovar 1982 Galaacuten de Mera et al 2009 2010) In the Salinas and Aguada Blanca National Reserve the Puna ecosystem commonly occurs between 38003900 and 48005000 m asl (Montenegro et al 2010)
The study area comprises the typical dry Puna land-scape of Latin America mostly dominated by bunch grasses like Festuca orthophylla or Jarava ichu and also composed of resinous shrubs (eg Parastrephia quad-rangularis and P lucida) and cushion plants (eg Pycno-phyllum molle P weberbaueri) From a phytosociologi-cal viewpoint according to Galaacuten de Mera et al (2002 2003 2009) the vegetation of the study area is included in the Parastrephietalia lepidophyllae Navarro 1993 order and in the Calamagrostietea vicunarum Rivas-Martiacutenez amp Tovar 1982 class
The human settlements are limited to small villages and isolated farms around which the economic activities of inhabitants (mainly breeding of domestic camelids) are concentrated These areas are mainly located on the least steep slopes of the altiplano while the surrounding high mountain slopes are not utilised by humans and host wild camelids
Materials and methods
Data collection
Data were collected at altitudes ranging from 3900 to 4900 m asl avoiding wetland (ldquobofedalesrdquo) since these areas lie outside the aim of our study (focused on dry grasslands) even if they represent an important habitat for alpaca breeding in the Reserve Rocky cliffs and streams were avoided as well We laid 160 sampling plots of 100 m2 each (10 m x 10 m) along 20 a priori chosen 2 km long transects The distance between consecutive plots was 250 m Transects were oriented along the direc-tion of maximum slope angle and selected to sample the heterogeneity of landforms and substrates 39 plots were not sampled because they laid on roads hamlets open quarries and wetlands or because they included latrine areas (small areas used by camelids for defecation and urination) In each of the remaining 121 plots the cover percentage values of species were estimated (based on a visual evaluation of vertically projected cover of species on the ground) using the Braun-Blanquet (1964) scale (+ lt 1 1 2ndash5 2 6ndash25 3 26ndash50 4 51ndash75 5 76ndash100 ) The collected specimens are stored at ldquoBie-nestar ndash Centro de Investigacioacuten para la Promocioacuten de los Pueblosrdquo of Arequipa (Peru)
phyto_44_1_2_103_119_Catorciindd 104phyto_44_1_2_103_119_Catorciindd 104 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 105
Fig 1 The study area (Salinas and Aguada Blanca National Reserve) located within the Arequipa and Moquegua Departments in the southern Peruvian Andes
phyto_44_1_2_103_119_Catorciindd 105phyto_44_1_2_103_119_Catorciindd 105 120314 0713120314 0713
eschweizerbart_xxx
106 A Catorci et al
For each releveacute we collected soil samples and data on altitude (m asl) slope aspect (azimuth degrees) slope angle (vertical degrees) and rockiness Within each plot 5 separate soil samples were taken at a depth of 2 to 20 cm below the soil surface and combined to one bulked sam-ple Grazing intensity and occurrence of fire were deter-mined by more than ten interviews with local villagers and the rangers of the Salinas and Aguada Blanca Na-tional Reserve Releveacutes were carried out at the end of the rainy season (March 2010) when soil moisture is highest and the highest number of species can be observed
Data on the following plant traits were collected by field observations and through consultation of the Her-barium of the Universidad Nacional de San Agustin (HUSA) of Arequipa (Peru) and of the Herbarium at the Universidad Nacional Mayor de San Marcos (USM) of Lima (Peru) annual and perennial life span graminoid species non-graminoid species upright forb (plant with leaves distributed on upright stems or branches) hemiro-sulate forb (plant with leaves distributed both at ground level and raised from the ground) prostrate forb (plant with leaves distributed only at the ground level) tall shrub (gt 15 cm) dwarf shrub (lt 15 cm) cushion and succulent plants
As it has been demonstrated that in harsh environ-ments positive plant-plant spatial interaction between nurse and beneficiary species play a key role in shaping the community richness and diversity (Armas amp Pug-naire 2005 Catorci et al 2011b Kikvidze et al 2011) fol-lowing several authors (Rebollo et al 2002 Maestre amp Cortina 2005 Padilla amp Pugnaire 2006 Arredondo-Nu-ntildeez et al 2009 Monteiro et al 2011) we chose tall grass tussocks tall shrubs and cushions as potential nurse plants considering the potential nurse role as an addi-tional functional trait
Species nomenclature mainly follows Brako amp Zarucchi (1993) Later taxonomic changes were checked by consult-ing IPNI (httpwwwipniorgindexhtml) Tropicos (httpwwwtropicosorg) and other botanical studies from close geographic areas (eg Araraki amp Cano 2003 Schwarzer et al 2010 Montesinos 2011 Montesinos et al 2012)
Data analysis
Soil samples were analysed by the Instituto Nacional de Innovacion Agraria (INIA) soil analysis laboratory of Arequipa (Ministerio de Agricultura y Riego del Peru) The following soil parameters were determined (the methods applied are indicated in parentheses) texture ie sand loam and clay (Buoyuocosrsquo method) or-ganic matter (modified Walkley Blackrsquos method) ni-trogen (Micro Kjeldahlrsquos method) phosphorus in ppm (modified Olsenrsquos method 05 M NaHCO3 extrac-tion at pH 88) potassium in ppm (flame photometry
by ammonium acetate at pH 70) CaCO3 (Wesemaelrsquos method) pH (by potentiometer in a 125 soilwater sus-pension)
To assess floristic composition the species cover val-ues were transformed into percentages using the central values of the Braun-Blanquet scale (Podani 2007) Fol-lowing Warren (2008) aspect azimuth was converted from the 0ndash360 compass scale to a linear (0ndash180) scale giving northerly aspect a value approaching 0 and south-erly aspect a value approaching 180 Disturbance inten-sity was assessed using a 4-level scale as follows 1) ab-sence of grazing (absent) 2) presence of wild camelids and sporadically small herds of domestic camelids (low) 3) grazing of large herds of domestic camelids (moder-ate) 4) grazing of large herds of domestic camelids and periodic fires (strong)
In order to assess the influence of environmental vari-ables and disturbance intensity on the species assemblage and identify homogeneous groups of releveacutes a con-strained clustering using Multivariate regression tree (MRT) analysis (Dersquoath 2002) with 100 iterations and 29 cross-validation groups was executed on two matrices namely ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo Prior to MRT analy-sis species data have been transformed using chord trans-formation (Legendre amp Gallagher 2001) which expresses the data as relative abundances per sites This removes from the data the total abundance per site which is the response of the species to the total productivity of the sites (Borcard et al 2011) The result of MRT analysis is a tree whose ldquoleavesrdquo (terminal groups of releveacutes) are com-posed of subsets of releveacutes chosen to minimize the within-group sum of squares and each partition of the tree is defined by a threshold value of one explanatory variable (Borcard et al 2011) Cross-validation is a re-sampling analysis repeated a number of times for succes-sive and independent divisions of the data (in this case species cover) into random test groups of objects (rel-eveacutes) (Legendre amp Legendre 2012) An overall error sta-tistic (cross-validation relative error CVRE) was com-puted for each test group and partition size (number of groups) to choose the optimal size of the tree CVRE varies from zero for a perfect set of predictors chosen for the splits of a tree to close to one for poor predictors For each partition size the mean and standard error of all CVRE estimates was computed Generally the smallest tree (ie a tree with fewer splits) within one standard er-ror of the smallest CVRE value is selected (Breiman et al 1984) but one may select the tree that has the smallest CVRE value as well (Legendre amp Legendre 2012)
To provide a general overview of the ecological charac-teristics of the whole releveacute data set we calculated the 1st and 3rd quartiles for each environmental variable The mean value and the standard deviation of the same varia-bles were calculated for each releveacute group that emerged from the MRT analysis
phyto_44_1_2_103_119_Catorciindd 106phyto_44_1_2_103_119_Catorciindd 106 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 107
The species diversity of each group was assessed by calculating the Shannon-Wiener (Hrsquo) and Gini-Simpson (D) diversity indices while the dissimilarity between the groups was computed through the Jaccard distance index (Magurran 1988) In order to investigate the species com-position of the groups we performed the Indicator Spe-cies Analysis (ISA) a non-parametric method for identi-fying those items (speciestrait) that show significantly preferential distribution (frequency and abundance) compared to an a priori treatment group An indicator value is calculated by multiplying the relative abundance of each item in a particular group and the relative fre-quency of the item occurrence in the sample of that group (Dufrecircne amp Legendre 1997) The number of randomized indicator values higher than the observed ones is used to calculate the probability value (McCune amp Grace 2002) The statistical significance (P lt 005) of the observed maximum Indicator Values (IV) was tested using permu-tation tests with 4999 iterations Only Indicator Values higher than 30 were considered of interest
To assess the influence of the ecological factors on spe-cies composition and abundance we performed a transfor-mation-based Redundancy Analysis (tb-RDA ndash Legendre amp Gallagher 2001) on the same matrices processed for MRT analysis Disturbance was considered as a factor var-iable with the above-mentioned four levels (absent low moderate strong) thus each of these levels was repre-sented by its centroid instead of an arrow in the RDA plot
To assess the influence of the ecological factors on the trait composition of the plant communities we first transformed trait binary data (presenceabsence) into quantitative data (ie aggregated cover values of each trait) The ldquoreleveacutes x species coverrdquo matrix was multiplied by the ldquospecies x traitrdquo matrix to provide a ldquoreleveacutes x trait coverrdquo matrix (Pakeman et al 2009) which formed the basis for the following analyses Then we ran a tb-RDA on the ldquoreleveacutes x trait coverrdquo matrix using the above-mentioned parameters (environmental factors and distur-bance intensity) as constraining variables
Since the disturbance gradient emerged as the main factor affecting trait composition in dry Puna communi-ties in order to better understand its effect on trait com-position we performed an ISA on the above matrix using MRT groups as grouping variable As the first partition of the regression tree segregated releveacutes characterised by strong disturbance intensity we re-ran ISA comparing releveacutes of this group against all the other ones The statis-tical significance (P lt 005) of the observed maximum In-dicator Values (IV) was tested using permutation tests with 4999 iterations Only Indicator Values higher than 30 were considered of interest
We used the R software (version 2140 ndash R Core Team 2012 ndash httpwwwR-projectorg) and its mvpart labdsv and vegan packages (Oksanen et al 2013) to perform MRT analysis ISA tb-RDA and to calculate diversity and dis-tance indices besides basic statistics
Results
MRT analysis generated two trees with the same CVRE (0641) composed of 4 and 14 leaves respectively (Fig 2) The 4-leaf tree had a smaller standard error (0051) than the 14-leaf tree (0062) and was selected as the best solution more frequently during the cross-validation it-erations Nevertheless to avoid losing information po-tentially useful for the ecological interpretation of the distribution of plant communities as the CVRE of the 14-leaf tree fell within one standard error of the smallest CVRE value we decided to consider this tree as well We recorded 136 species (belonging to 69 genera and 28 fam-ilies) The species composition of each group highlighted by MRT is shown in Fig 3 while the related distance in-dices are given in Appendix A The values of the Shan-non-Wiener and the Gini-Simpson diversity indices are reported in Table 1 The indicator species identified by ISA for groups highlighted by MRT as well as their mean abundances are shown in Table 2 Table 3 indicates the mean the median and the 1st and 3rd quartiles for the eco-logical variables in each group Table 2 shows the output of ISA concerning the assessment of the indicator traits of the four relevegrave groups of the 4-leaf and 14-leaf trees The variable that discriminated between the two branches in the first node of the 4-leaves MRT was grazing distur-bance thus the first leaf (T41) was composed of the re-leveacutes characterised by a high level of disturbance The related plant communities were dominated by the dwarf shrub Tetraglochin cristatum they presented the lowest species diversity (Hrsquo = 163 D = 065) as well as the high-est distances compared to the other leaves Other envi-ronmental features of the T cristatum ndash dominated plant community were (Table 3) high sand percentage and low soil values of organic matter nitrogen phosphorus and potassium The pH value was the highest one compared to the other groups of releveacutes Dwarf shrubs non-grami-noid and annual species were the indicator traits of this group compared with all the other releveacutes jointly consid-ered (Table 4)
The second partition was defined by the percentage of outcropping rocks The first branch of this partition characterised by the lower values of outcropping rocks is in turn divided into two leaves the first (T42) grouping releveacutes located below 4000 m of altitude is dominated by Jarava ichu while Calamagrostis breviaristata domi-nates the second one (T43) grouping the releveacutes placed above 4000 m Both these groups were subjected to in-termediate level of disturbance and were characterised by flat or semi-flat slopes The Jarava ichu ndash dominated plant community was distinguished by low sand percent-age and relatively high organic matter phosphorus and potassium content in soil (Table 3) The pH value was similar to that of groups T43 and T44 groups Grami-noid and hemirosulate forbs as well as succulent and po-
phyto_44_1_2_103_119_Catorciindd 107phyto_44_1_2_103_119_Catorciindd 107 120314 0713120314 0713
eschweizerbart_xxx
108 A Catorci et al
Fig 2 The 4-leaf (a) and 14-leaf (b) Multivariate regression trees for the species data set constrained by their explanatory variables The threshold values shown for each partition of the tree correspond to the mean of the two limit values of the considered variables at the break between the branches The relative abundances of the species are shown in histograms at the tips of the branches with the species in the same order as in the input file the species names and abun-dances are shown in Figure 3 Under each histogram the sum of squared errors for the group and the number of releveacutes in the leaf (n) are indicated Below each tree the residual error (Error) the cross-validation error (CV Error) and standard error (SE) are indi-cated Alt ndash Altitude Asp_deg ndash aspect Dist_L ndash Disturbance level Hum_p ndash organic matter K_ppm ndash Potassium in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt
tential nurse plants were the indicator traits for the T42 group (Table 4)
The C breviaristata ndash dominated plant community was characterized by high sand percentage and low or-ganic matter and potassium content in the soil (Table 3) The pH value was similar to that of groups T43 and T44 Cushion and perennial plants were the indicator traits for group T43 (Table 4)
The releveacutes grouped in the fourth leaf (T44) are char-acterised by a percentage of outcropping rocks greater than 1 (mean cover value 178 ) In this group the dominant species is the graminoid Festuca orthophylla other indicator species being Deyeuxia brevifolia Trise-tum spicatum Nassella depauperata and Deyeuxia rigida (all grass species) Group T44 includes the greatest num-ber of releveacutes (69) and presents the highest diversity (Hrsquo = 282 D = 083) in the 4-leaved tree Other environmen-
tal features of the Festuca orthophylla ndash dominated plant community were (Table 3) the highest slopes altitude and angles among the four groups highlighted by MRT low sand percentage and high values of organic matter nitro-gen and phosphorus in the soil The pH value was similar to that of groups T42 and T43 These pastures were characterised by low disturbance intensity Within the high richness and internal variability of these grasslands particular environmental conditions or treatments caused minor differences in species composition highlighting different plant communities as shown by the 14-leaved MRT (Fig 2)
Despite the high number of groups yielded the val-ues of community diversity indices computed from the pairwise comparison of the 14 leaves were still high (never below 45 ) (Appendix A) The first partition of the Festuca orthophylla ndash dominated grassland was de-
phyto_44_1_2_103_119_Catorciindd 108phyto_44_1_2_103_119_Catorciindd 108 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 109
fined by aspect with a threshold value that can be inter-preted as the boundary between north-east-facing slopes (arid more stressful ndash groups T148-T1412) and south-west-facing ones (cool less stressful ndash groups T1413 and T1414) From a functional point of view the south-facing plots were characterized by a high number of indicator traits (perennial species graminoids hemi-rosulate forbs cushion plants and potential nurses) while no traits emerged as indicator of the plant com-munities growing on the north-facing slopes In addi-tion north-facing groups were characterised by a higher degree of dominance with the most widespread species often representing more than 50 of the total vegeta-tion cover
Altitude gradients and soil parameters defined further partitions for a more detailed differentiation of these two main clusters Among north-facing groups T148 was dominated by Nassella depauperata and had coarser soil as well as higher pH and organic matter values than T149 Groups T1410 T1411 and T1412 were charac-terised by higher altitudes (about 400 m above T148 and T149 ndash Table 3) T1410 was characterised by low distur-bance intensity and showed a more even distribution of species (particularly if compared with T1411) among which several palatable grasses are present (eg Deyeuxia brevifolia) Group T1412 dominated by the shrub spe-cies Chersodoma jodopappa showed a higher percentage of silt and organic matter in soil and steeper slopes with
Fig 3 Pie charts reporting the species composition of the groups defined in the 4-leaf (a) and in the 14-leaf (b) Multivariate regres-sion trees Species abbreviations are explained below Ast_die ndash As-tragalus dielsii Bac_tri ndash Baccharis tricuneata Bel_lon ndash Belloa longifolia Cal_bra ndash Calamagrostis breviaristata Can_aca ndash Calan-drina acaulis Chs_jod ndash Chersodoma jodopappa Cro_ful ndash Crocop-sis fulgens Des_myr ndash Descurainia myriophylla Dey_brf ndash De-yeuxia brevifolia Dey_rig ndash Deyeuxia rigida Dip_mey ndash Diplostephium meyenii Ech_pam ndash Echinopsis pampana Fes_ort ndash Festuca ortophylla Gna_lac ndash Gnaphalium lacteum Hyp_ech ndash Hypochaeris echegarayi Jar_ich ndash Jarava ichu Lpp_qua ndash Lepi-dopyllum quadrangulare Lup_sax ndash Lupinus saxatilis Muh_per ndash Muhlenbergia peruviana Nas_dep ndash Nassella depauperata Nas_nar ndash Nassella nardoides Ntt_tur ndash Nototriche turritella Oen_sp ndash Oenothera sp Par_luc ndash Parastrephia lucida Par_qua ndash Parastre-phia quadrangularis Pla_ser ndash Plantago sericea var lanuginosa Poa_gym ndash Poa gymnantha Poa_pea ndash Poa pearsonii Pyc_mol ndash Pycnophyllum molle Pyc_web ndash Pycnophyllum weberbaueri Sen_cha ndash Senecio chachaniensis Sen_nut ndash Senecio nutans Sti_han ndash Stipa hans-meyeri Tet_cri ndash Tetraglochin cristatum Tri_spi ndash Trise-tum spicatum Wer_pyg ndash Werneria pygmaea
phyto_44_1_2_103_119_Catorciindd 109phyto_44_1_2_103_119_Catorciindd 109 120314 0713120314 0713
eschweizerbart_xxx
110 A Catorci et al
groups yielded by the 4-leaf tree are clearly recognisable in the RDA three-dimensional scatterplot (as highlighted in Fig 4) The RDA performed on functional traits data with the environmental variables as constraints yielded similar results but explained a notably smaller fraction of the total variance (92 adjusted R2 = 056) The varia-bles showing the strongest correlation with the con-strained components coincided with those observed in the first RDA but for most environmental variables the observed correlation is weaker this is the case of altitude aspect percentage of outcropping rocks sand and silt percentage in the soil (Fig 5) Instead for disturbance in-tensity the observed correlation was slightly stronger The functional traits whose variance was best explained by the constrained components were nurse ability and various types of plant forms dwarf shrubs upright forbs non-graminoids hemirosulate forbs and graminoids (Fig 5) Non-graminoid species dwarf shrubs and up-right forbs were related to relatively higher soil pH and strong disturbance intensity nurse species were linked to higher altitudes South-facing slopes relatively lower sand percentages in the soil graminoid species and hemi-rosulate forbs were particularly tied to more fertile soils (higher phosphorus and nitrogen concentrations) and low disturbance intensity (Fig 5) RDA biplot scores are reported in the Appendix C
Discussion
Our results demonstrate that the dry Puna landscape is anything but homogeneous as observed for other semi-arid environments where local factors were detected as key drivers of species assemblages (eg Schlesinger et al 1990 Perelman et al 1997 Diacuteaz et al 1999) The variables showing the strongest correlations with the constrained RDA components were disturbance intensity slope as-pect altitude percentage of outcropping rocks and to a lesser degree soil features (organic matter content pH and potassium concentration) High disturbance inten-sity (which had the best correlation with the first con-strained component) emerged as the main factor shaping the species composition of the dry Puna pastures and was positively correlated with pH dwarf shrubs and non-graminoid plant forms while it was negatively cor-related with organic matternitrogen soil content nurse plant abundance hemirosulate forbs and graminoids Consistently with previous research carried out in semi-arid environments (eg Ares et al 1990 Carilla et al 2012) high grazing intensity promotes the spread of spe-cies with avoidance strategies (Table 4) such as thorny dwarf shrubs (Tetraglochin cristatum) annual species (eg Muhlenbergia peruviana) and small or prostrate plants (eg Nassella nardoides Astragalus dielsii Hy-pochaeris echegarayi) Thorny dwarf shrubs are fostered by herbivory given that the tough and spiny stems of this
Table 1 Values of Shannon and Simpson diversity indices for the releveacute groups of the 4-leaf and 14-leaf Multivariate regression tree
Groups Diversity
Shannon (Hlsquo) Simpson (D)
T41 1628 0653
T42 1831 0626
T43 2176 0765
T44 2822 0829
T141 1511 0582
T142 1611 0688
T143 1831 0626
T144 1714 0633
T145 2366 0822
T146 1913 0763
T147 2348 0824
T148 1316 0598
T149 1540 0574
T1410 2214 0792
T1411 2283 0675
T1412 2149 0714
T1413 2100 0741
T1414 2685 0863
high outcropping rock cover South-facing groups were in turn separated by potassium content in the soil group T1413 with higher concentration of potassium shows a higher coverage of graminoid species (eg Poa pearsonii)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grass Festuca or-thophylla the cushion plants Pycnophyllum molle and P weberbaueri the shrub species Parastrephia lucida and P quadrangularis) This plant community was located near the altitudinal limit of vegetation and characterised by very low disturbance
The Redundancy Analysis executed on species and en-vironmental data explained 638 of the total variance (adjusted R2 = 047) The results of the RDA are in line with the MRT analysis the variables showing the strong-est correlation with the constrained components were disturbance intensity altitude percentage of outcropping rocks slope aspect soil organic matter content pH and potassium concentration (Fig 4) RDA biplot scores are reported in Appendix B It is worth noting that this result coincides with those discriminating for the partitions of the 4-leaf and 14-leaf MRTs (in order of diminishing cor-relation with RDA axes) Another correspondence be-tween MRT and RDA analyses lies in the placement of site points in the RDA graph as a matter of fact the
phyto_44_1_2_103_119_Catorciindd 110phyto_44_1_2_103_119_Catorciindd 110 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 111
Table 2 Indicator species of the releveacute groups highlighted by Indicator Species Analysis for the 4ndashleaf and the 14ndashleaf Multivariate regres-sion tree (MRT) and their respective indicator values and mean cover percentages Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value)
Indicator species 4-leaf MRT 14-leaf MRTMax
GroupIndicator
ValueMean
cover Max
GroupIndicator
ValueMean
cover Nassella nardoides T41 663 199 ndash ndash ndash
Nothoscordum andicola T41 482 08 ndash ndash ndash
Lepidium meyenii T41 412 06 ndash ndash ndash
Astragalus peruvianus T41 315 07 ndash ndash ndash
Tetraglochin cristatum T41 826 544 T141 422 628
Muhlenbergia peruviana T41 675 44 T142 300 46
Cumulopuntia ignescens T42 405 07 ndash ndash ndash
Chenopodium petiolare T42 370 03 ndash ndash ndash
Urocarpidium echinatum T42 350 06 ndash ndash ndash
Nototriche turritella T42 345 24 ndash ndash ndash
Jarava ichu T42 979 598 T143 925 598
Plantago linearis T42 901 30 T143 493 30
Diplostephium meyenii T42 700 23 T143 634 23
Crocopsis fulgens T42 571 16 T143 571 16
Belloa piptolepis T42 592 11 T143 351 11
Senecio phylloleptus T42 490 10 T143 364 10
Hypochaeris echegarayi T43 380 08 ndash ndash ndash
Pycnophyllum weberbaueri T43 342 08 ndash ndash ndash
Junellia minima (Meyen) T43 300 08 ndash ndash ndash
Calamagrostis breviaristata T43 704 430 T144 398 581
Calceolaria inamoena ndash ndash ndash T145 315 08
Astragalus dielsii T43 460 11 T146 321 14
Nassella nardoides ndash ndash ndash T146 484 415
Calandrinia acaulis ndash ndash ndash T146 430 14
Oenothera sp ndash ndash ndash T146 420 14
Festuca orthophylla T44 522 389 ndash ndash ndash
Trisetum spicatum T44 406 08 ndash ndash ndash
Nassella depauperata T44 352 70 T148 860 600
Lupinus saxatilis ndash ndash ndash T148 657 80
Descurainia myriophylla ndash ndash ndash T148 437 133
Erigeron pazensis ndash ndash ndash T148 416 07
Deyeuxia brevifolia T44 464 53 T1410 414 193
Mniodes pulvinata ndash ndash ndash T1410 375 05
Lupinus chlorolepis ndash ndash ndash T1411 364 09
Calamagrostis vicunarum ndash ndash ndash T1412 500 07
Poa gymnantha ndash ndash ndash T1412 485 42
Dielsiochloa floribunda ndash ndash ndash T1412 450 07
Leucheria daucifolia ndash ndash ndash T1412 398 07
Echinopsis pampana ndash ndash ndash T1412 384 42
Perezia ciliosa ndash ndash ndash T1412 329 07
Poa pearsonii ndash ndash ndash T1413 670 237
Deyeuxia rigida T44 316 47 T1414 774 285
Poa aequigluma ndash ndash ndash T1414 453 06
Werneria spathulata ndash ndash ndash T1414 412 08
Luzula racemosa ndash ndash ndash T1414 409 09
Lucilia sp ndash ndash ndash T1414 401 12
Senecio scorzoneraefolius ndash ndash ndash T1414 376 05
Gnaphalium badium ndash ndash ndash T1414 338 10
Pycnophyllum molle ndash ndash ndash T1414 335 95
phyto_44_1_2_103_119_Catorciindd 111phyto_44_1_2_103_119_Catorciindd 111 120314 0713120314 0713
eschweizerbart_xxx
112 A Catorci et al
Table 3 Descriptive statistics of the environmental variables of the groups highlighted by the 4-leaf and 14-leaf Multivariate regression tree 1st and 3rd quartiles are indicated for the whole data set mean and standard deviation (SD) are reported for each group
Group Descriptive statistics
Constraining variables
Site characteristics Soil texture Soil composition
Altitude(m asl)
Aspect (deg)
Slope (deg)
Outcr rocks()
Sand()
Silt ()
Clay ()
Org matt()
N () P (ppm)
K (ppm)
CaCO3 ()
pH
1st quartile 407000 000 200 000 6280 1660 580 089 004 1136 11250 008 540 3rd quartile 447500 9000 2000 1500 7760 2780 910 174 009 2469 23250 075 623
T41 Mean 399700 2363 755 000 7867 1403 1051 071 003 1265 16381 049 652SD 1908 2777 894 000 295 180 1434 033 001 668 3318 035 028
T42 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T43 Mean 410500 2880 364 000 7265 2133 602 110 009 2173 17036 053 542SD 2933 4752 930 000 389 372 237 053 020 1234 6129 051 035
T44 Mean 440287 8739 2061 1787 6698 2444 849 171 008 2219 19384 045 572SD 22159 5242 1431 1192 976 866 304 083 004 1471 11137 039 045
T141 Mean 399833 3938 1208 000 7926 1374 1233 052 003 875 17452 049 666SD 2462 2561 901 000 333 203 1853 024 001 142 3181 032 028
T142 Mean 399500 000 075 000 7778 1445 778 099 004 1850 14775 049 631SD 535 000 104 000 214 141 213 022 001 725 3011 041 010
T143 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T144 Mean 411393 643 064 000 7373 1987 640 098 012 2552 18143 052 529SD 1712 2405 145 000 309 274 306 020 027 1421 5395 062 035
T145 Mean 412667 10500 2167 000 6777 2577 647 210 008 1611 18335 071 549SD 4619 1299 2082 000 224 145 093 017 001 516 13137 017 031
T146 Mean 409500 000 000 000 7385 2128 488 078 005 2370 18117 015 575SD 000 000 000 000 042 026 017 003 001 327 1019 019 018
T147 Mean 406750 7875 425 000 7135 2315 550 110 004 1073 11106 082 551SD 3663 5356 299 000 654 654 081 094 004 698 1577 019 031
T148 Mean 414800 7200 1900 1500 7368 2080 552 135 007 2044 9024 019 630SD 570 1006 224 000 618 567 064 037 002 1111 4791 021 015
T149 Mean 408833 9000 600 1833 6960 2093 947 096 005 1216 18803 054 566SD 5795 000 346 577 763 412 357 067 004 098 12521 057 071
T1410 Mean 458688 4922 2731 1813 7623 1591 787 136 006 2448 17087 042 547SD 22773 4043 1590 680 622 535 164 026 001 1461 9485 047 049
T1411 Mean 439614 5727 1114 2173 6501 2430 1038 147 007 1741 22177 052 583SD 16079 2840 551 1297 832 588 420 053 003 597 10086 035 049
T1412 Mean 445650 9000 4000 550 6025 3248 725 248 013 1653 25562 055 579SD 27365 000 2121 636 078 046 120 049 002 467 26427 040 013
T1413 Mean 421318 15341 3518 2409 5954 3227 819 285 014 2823 28413 025 564SD 11402 2814 1419 1338 1099 981 169 117 006 2824 11501 041 022
T1414 Mean 454300 14850 1600 590 6189 3104 707 183 009 2740 11102 065 571SD 6997 2846 615 1058 589 588 071 063 003 830 2759 027 026
life form discourage animals from biting it (Briske 1996) The observed spread of dwarf shrubs is in accordance with the findings of several studies carried out in semiarid environments (Perelman et al 1997 Swinton amp Quiroz 2003 Carilla et al 2012 Catorci et al 2012b 2013) and in pastoral ecosystems subjected to periodic fires (Liedloff
et al 2001) In semi-arid environments annual life span may be considered an avoidance strategy fostered by high grazing intensity (Diacuteaz et al 1999 McGregor 2002 Catorci et al 2012b) because it allows plants to dynami-cally face the grazing pressure by restarting the growth cycle every year (Diacuteaz et al 1999 2001 McGregor 2002)
phyto_44_1_2_103_119_Catorciindd 112phyto_44_1_2_103_119_Catorciindd 112 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

Floristic composition of dry Puna in the southern Peruvian Andes 105
Fig 1 The study area (Salinas and Aguada Blanca National Reserve) located within the Arequipa and Moquegua Departments in the southern Peruvian Andes
phyto_44_1_2_103_119_Catorciindd 105phyto_44_1_2_103_119_Catorciindd 105 120314 0713120314 0713
eschweizerbart_xxx
106 A Catorci et al
For each releveacute we collected soil samples and data on altitude (m asl) slope aspect (azimuth degrees) slope angle (vertical degrees) and rockiness Within each plot 5 separate soil samples were taken at a depth of 2 to 20 cm below the soil surface and combined to one bulked sam-ple Grazing intensity and occurrence of fire were deter-mined by more than ten interviews with local villagers and the rangers of the Salinas and Aguada Blanca Na-tional Reserve Releveacutes were carried out at the end of the rainy season (March 2010) when soil moisture is highest and the highest number of species can be observed
Data on the following plant traits were collected by field observations and through consultation of the Her-barium of the Universidad Nacional de San Agustin (HUSA) of Arequipa (Peru) and of the Herbarium at the Universidad Nacional Mayor de San Marcos (USM) of Lima (Peru) annual and perennial life span graminoid species non-graminoid species upright forb (plant with leaves distributed on upright stems or branches) hemiro-sulate forb (plant with leaves distributed both at ground level and raised from the ground) prostrate forb (plant with leaves distributed only at the ground level) tall shrub (gt 15 cm) dwarf shrub (lt 15 cm) cushion and succulent plants
As it has been demonstrated that in harsh environ-ments positive plant-plant spatial interaction between nurse and beneficiary species play a key role in shaping the community richness and diversity (Armas amp Pug-naire 2005 Catorci et al 2011b Kikvidze et al 2011) fol-lowing several authors (Rebollo et al 2002 Maestre amp Cortina 2005 Padilla amp Pugnaire 2006 Arredondo-Nu-ntildeez et al 2009 Monteiro et al 2011) we chose tall grass tussocks tall shrubs and cushions as potential nurse plants considering the potential nurse role as an addi-tional functional trait
Species nomenclature mainly follows Brako amp Zarucchi (1993) Later taxonomic changes were checked by consult-ing IPNI (httpwwwipniorgindexhtml) Tropicos (httpwwwtropicosorg) and other botanical studies from close geographic areas (eg Araraki amp Cano 2003 Schwarzer et al 2010 Montesinos 2011 Montesinos et al 2012)
Data analysis
Soil samples were analysed by the Instituto Nacional de Innovacion Agraria (INIA) soil analysis laboratory of Arequipa (Ministerio de Agricultura y Riego del Peru) The following soil parameters were determined (the methods applied are indicated in parentheses) texture ie sand loam and clay (Buoyuocosrsquo method) or-ganic matter (modified Walkley Blackrsquos method) ni-trogen (Micro Kjeldahlrsquos method) phosphorus in ppm (modified Olsenrsquos method 05 M NaHCO3 extrac-tion at pH 88) potassium in ppm (flame photometry
by ammonium acetate at pH 70) CaCO3 (Wesemaelrsquos method) pH (by potentiometer in a 125 soilwater sus-pension)
To assess floristic composition the species cover val-ues were transformed into percentages using the central values of the Braun-Blanquet scale (Podani 2007) Fol-lowing Warren (2008) aspect azimuth was converted from the 0ndash360 compass scale to a linear (0ndash180) scale giving northerly aspect a value approaching 0 and south-erly aspect a value approaching 180 Disturbance inten-sity was assessed using a 4-level scale as follows 1) ab-sence of grazing (absent) 2) presence of wild camelids and sporadically small herds of domestic camelids (low) 3) grazing of large herds of domestic camelids (moder-ate) 4) grazing of large herds of domestic camelids and periodic fires (strong)
In order to assess the influence of environmental vari-ables and disturbance intensity on the species assemblage and identify homogeneous groups of releveacutes a con-strained clustering using Multivariate regression tree (MRT) analysis (Dersquoath 2002) with 100 iterations and 29 cross-validation groups was executed on two matrices namely ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo Prior to MRT analy-sis species data have been transformed using chord trans-formation (Legendre amp Gallagher 2001) which expresses the data as relative abundances per sites This removes from the data the total abundance per site which is the response of the species to the total productivity of the sites (Borcard et al 2011) The result of MRT analysis is a tree whose ldquoleavesrdquo (terminal groups of releveacutes) are com-posed of subsets of releveacutes chosen to minimize the within-group sum of squares and each partition of the tree is defined by a threshold value of one explanatory variable (Borcard et al 2011) Cross-validation is a re-sampling analysis repeated a number of times for succes-sive and independent divisions of the data (in this case species cover) into random test groups of objects (rel-eveacutes) (Legendre amp Legendre 2012) An overall error sta-tistic (cross-validation relative error CVRE) was com-puted for each test group and partition size (number of groups) to choose the optimal size of the tree CVRE varies from zero for a perfect set of predictors chosen for the splits of a tree to close to one for poor predictors For each partition size the mean and standard error of all CVRE estimates was computed Generally the smallest tree (ie a tree with fewer splits) within one standard er-ror of the smallest CVRE value is selected (Breiman et al 1984) but one may select the tree that has the smallest CVRE value as well (Legendre amp Legendre 2012)
To provide a general overview of the ecological charac-teristics of the whole releveacute data set we calculated the 1st and 3rd quartiles for each environmental variable The mean value and the standard deviation of the same varia-bles were calculated for each releveacute group that emerged from the MRT analysis
phyto_44_1_2_103_119_Catorciindd 106phyto_44_1_2_103_119_Catorciindd 106 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 107
The species diversity of each group was assessed by calculating the Shannon-Wiener (Hrsquo) and Gini-Simpson (D) diversity indices while the dissimilarity between the groups was computed through the Jaccard distance index (Magurran 1988) In order to investigate the species com-position of the groups we performed the Indicator Spe-cies Analysis (ISA) a non-parametric method for identi-fying those items (speciestrait) that show significantly preferential distribution (frequency and abundance) compared to an a priori treatment group An indicator value is calculated by multiplying the relative abundance of each item in a particular group and the relative fre-quency of the item occurrence in the sample of that group (Dufrecircne amp Legendre 1997) The number of randomized indicator values higher than the observed ones is used to calculate the probability value (McCune amp Grace 2002) The statistical significance (P lt 005) of the observed maximum Indicator Values (IV) was tested using permu-tation tests with 4999 iterations Only Indicator Values higher than 30 were considered of interest
To assess the influence of the ecological factors on spe-cies composition and abundance we performed a transfor-mation-based Redundancy Analysis (tb-RDA ndash Legendre amp Gallagher 2001) on the same matrices processed for MRT analysis Disturbance was considered as a factor var-iable with the above-mentioned four levels (absent low moderate strong) thus each of these levels was repre-sented by its centroid instead of an arrow in the RDA plot
To assess the influence of the ecological factors on the trait composition of the plant communities we first transformed trait binary data (presenceabsence) into quantitative data (ie aggregated cover values of each trait) The ldquoreleveacutes x species coverrdquo matrix was multiplied by the ldquospecies x traitrdquo matrix to provide a ldquoreleveacutes x trait coverrdquo matrix (Pakeman et al 2009) which formed the basis for the following analyses Then we ran a tb-RDA on the ldquoreleveacutes x trait coverrdquo matrix using the above-mentioned parameters (environmental factors and distur-bance intensity) as constraining variables
Since the disturbance gradient emerged as the main factor affecting trait composition in dry Puna communi-ties in order to better understand its effect on trait com-position we performed an ISA on the above matrix using MRT groups as grouping variable As the first partition of the regression tree segregated releveacutes characterised by strong disturbance intensity we re-ran ISA comparing releveacutes of this group against all the other ones The statis-tical significance (P lt 005) of the observed maximum In-dicator Values (IV) was tested using permutation tests with 4999 iterations Only Indicator Values higher than 30 were considered of interest
We used the R software (version 2140 ndash R Core Team 2012 ndash httpwwwR-projectorg) and its mvpart labdsv and vegan packages (Oksanen et al 2013) to perform MRT analysis ISA tb-RDA and to calculate diversity and dis-tance indices besides basic statistics
Results
MRT analysis generated two trees with the same CVRE (0641) composed of 4 and 14 leaves respectively (Fig 2) The 4-leaf tree had a smaller standard error (0051) than the 14-leaf tree (0062) and was selected as the best solution more frequently during the cross-validation it-erations Nevertheless to avoid losing information po-tentially useful for the ecological interpretation of the distribution of plant communities as the CVRE of the 14-leaf tree fell within one standard error of the smallest CVRE value we decided to consider this tree as well We recorded 136 species (belonging to 69 genera and 28 fam-ilies) The species composition of each group highlighted by MRT is shown in Fig 3 while the related distance in-dices are given in Appendix A The values of the Shan-non-Wiener and the Gini-Simpson diversity indices are reported in Table 1 The indicator species identified by ISA for groups highlighted by MRT as well as their mean abundances are shown in Table 2 Table 3 indicates the mean the median and the 1st and 3rd quartiles for the eco-logical variables in each group Table 2 shows the output of ISA concerning the assessment of the indicator traits of the four relevegrave groups of the 4-leaf and 14-leaf trees The variable that discriminated between the two branches in the first node of the 4-leaves MRT was grazing distur-bance thus the first leaf (T41) was composed of the re-leveacutes characterised by a high level of disturbance The related plant communities were dominated by the dwarf shrub Tetraglochin cristatum they presented the lowest species diversity (Hrsquo = 163 D = 065) as well as the high-est distances compared to the other leaves Other envi-ronmental features of the T cristatum ndash dominated plant community were (Table 3) high sand percentage and low soil values of organic matter nitrogen phosphorus and potassium The pH value was the highest one compared to the other groups of releveacutes Dwarf shrubs non-grami-noid and annual species were the indicator traits of this group compared with all the other releveacutes jointly consid-ered (Table 4)
The second partition was defined by the percentage of outcropping rocks The first branch of this partition characterised by the lower values of outcropping rocks is in turn divided into two leaves the first (T42) grouping releveacutes located below 4000 m of altitude is dominated by Jarava ichu while Calamagrostis breviaristata domi-nates the second one (T43) grouping the releveacutes placed above 4000 m Both these groups were subjected to in-termediate level of disturbance and were characterised by flat or semi-flat slopes The Jarava ichu ndash dominated plant community was distinguished by low sand percent-age and relatively high organic matter phosphorus and potassium content in soil (Table 3) The pH value was similar to that of groups T43 and T44 groups Grami-noid and hemirosulate forbs as well as succulent and po-
phyto_44_1_2_103_119_Catorciindd 107phyto_44_1_2_103_119_Catorciindd 107 120314 0713120314 0713
eschweizerbart_xxx
108 A Catorci et al
Fig 2 The 4-leaf (a) and 14-leaf (b) Multivariate regression trees for the species data set constrained by their explanatory variables The threshold values shown for each partition of the tree correspond to the mean of the two limit values of the considered variables at the break between the branches The relative abundances of the species are shown in histograms at the tips of the branches with the species in the same order as in the input file the species names and abun-dances are shown in Figure 3 Under each histogram the sum of squared errors for the group and the number of releveacutes in the leaf (n) are indicated Below each tree the residual error (Error) the cross-validation error (CV Error) and standard error (SE) are indi-cated Alt ndash Altitude Asp_deg ndash aspect Dist_L ndash Disturbance level Hum_p ndash organic matter K_ppm ndash Potassium in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt
tential nurse plants were the indicator traits for the T42 group (Table 4)
The C breviaristata ndash dominated plant community was characterized by high sand percentage and low or-ganic matter and potassium content in the soil (Table 3) The pH value was similar to that of groups T43 and T44 Cushion and perennial plants were the indicator traits for group T43 (Table 4)
The releveacutes grouped in the fourth leaf (T44) are char-acterised by a percentage of outcropping rocks greater than 1 (mean cover value 178 ) In this group the dominant species is the graminoid Festuca orthophylla other indicator species being Deyeuxia brevifolia Trise-tum spicatum Nassella depauperata and Deyeuxia rigida (all grass species) Group T44 includes the greatest num-ber of releveacutes (69) and presents the highest diversity (Hrsquo = 282 D = 083) in the 4-leaved tree Other environmen-
tal features of the Festuca orthophylla ndash dominated plant community were (Table 3) the highest slopes altitude and angles among the four groups highlighted by MRT low sand percentage and high values of organic matter nitro-gen and phosphorus in the soil The pH value was similar to that of groups T42 and T43 These pastures were characterised by low disturbance intensity Within the high richness and internal variability of these grasslands particular environmental conditions or treatments caused minor differences in species composition highlighting different plant communities as shown by the 14-leaved MRT (Fig 2)
Despite the high number of groups yielded the val-ues of community diversity indices computed from the pairwise comparison of the 14 leaves were still high (never below 45 ) (Appendix A) The first partition of the Festuca orthophylla ndash dominated grassland was de-
phyto_44_1_2_103_119_Catorciindd 108phyto_44_1_2_103_119_Catorciindd 108 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 109
fined by aspect with a threshold value that can be inter-preted as the boundary between north-east-facing slopes (arid more stressful ndash groups T148-T1412) and south-west-facing ones (cool less stressful ndash groups T1413 and T1414) From a functional point of view the south-facing plots were characterized by a high number of indicator traits (perennial species graminoids hemi-rosulate forbs cushion plants and potential nurses) while no traits emerged as indicator of the plant com-munities growing on the north-facing slopes In addi-tion north-facing groups were characterised by a higher degree of dominance with the most widespread species often representing more than 50 of the total vegeta-tion cover
Altitude gradients and soil parameters defined further partitions for a more detailed differentiation of these two main clusters Among north-facing groups T148 was dominated by Nassella depauperata and had coarser soil as well as higher pH and organic matter values than T149 Groups T1410 T1411 and T1412 were charac-terised by higher altitudes (about 400 m above T148 and T149 ndash Table 3) T1410 was characterised by low distur-bance intensity and showed a more even distribution of species (particularly if compared with T1411) among which several palatable grasses are present (eg Deyeuxia brevifolia) Group T1412 dominated by the shrub spe-cies Chersodoma jodopappa showed a higher percentage of silt and organic matter in soil and steeper slopes with
Fig 3 Pie charts reporting the species composition of the groups defined in the 4-leaf (a) and in the 14-leaf (b) Multivariate regres-sion trees Species abbreviations are explained below Ast_die ndash As-tragalus dielsii Bac_tri ndash Baccharis tricuneata Bel_lon ndash Belloa longifolia Cal_bra ndash Calamagrostis breviaristata Can_aca ndash Calan-drina acaulis Chs_jod ndash Chersodoma jodopappa Cro_ful ndash Crocop-sis fulgens Des_myr ndash Descurainia myriophylla Dey_brf ndash De-yeuxia brevifolia Dey_rig ndash Deyeuxia rigida Dip_mey ndash Diplostephium meyenii Ech_pam ndash Echinopsis pampana Fes_ort ndash Festuca ortophylla Gna_lac ndash Gnaphalium lacteum Hyp_ech ndash Hypochaeris echegarayi Jar_ich ndash Jarava ichu Lpp_qua ndash Lepi-dopyllum quadrangulare Lup_sax ndash Lupinus saxatilis Muh_per ndash Muhlenbergia peruviana Nas_dep ndash Nassella depauperata Nas_nar ndash Nassella nardoides Ntt_tur ndash Nototriche turritella Oen_sp ndash Oenothera sp Par_luc ndash Parastrephia lucida Par_qua ndash Parastre-phia quadrangularis Pla_ser ndash Plantago sericea var lanuginosa Poa_gym ndash Poa gymnantha Poa_pea ndash Poa pearsonii Pyc_mol ndash Pycnophyllum molle Pyc_web ndash Pycnophyllum weberbaueri Sen_cha ndash Senecio chachaniensis Sen_nut ndash Senecio nutans Sti_han ndash Stipa hans-meyeri Tet_cri ndash Tetraglochin cristatum Tri_spi ndash Trise-tum spicatum Wer_pyg ndash Werneria pygmaea
phyto_44_1_2_103_119_Catorciindd 109phyto_44_1_2_103_119_Catorciindd 109 120314 0713120314 0713
eschweizerbart_xxx
110 A Catorci et al
groups yielded by the 4-leaf tree are clearly recognisable in the RDA three-dimensional scatterplot (as highlighted in Fig 4) The RDA performed on functional traits data with the environmental variables as constraints yielded similar results but explained a notably smaller fraction of the total variance (92 adjusted R2 = 056) The varia-bles showing the strongest correlation with the con-strained components coincided with those observed in the first RDA but for most environmental variables the observed correlation is weaker this is the case of altitude aspect percentage of outcropping rocks sand and silt percentage in the soil (Fig 5) Instead for disturbance in-tensity the observed correlation was slightly stronger The functional traits whose variance was best explained by the constrained components were nurse ability and various types of plant forms dwarf shrubs upright forbs non-graminoids hemirosulate forbs and graminoids (Fig 5) Non-graminoid species dwarf shrubs and up-right forbs were related to relatively higher soil pH and strong disturbance intensity nurse species were linked to higher altitudes South-facing slopes relatively lower sand percentages in the soil graminoid species and hemi-rosulate forbs were particularly tied to more fertile soils (higher phosphorus and nitrogen concentrations) and low disturbance intensity (Fig 5) RDA biplot scores are reported in the Appendix C
Discussion
Our results demonstrate that the dry Puna landscape is anything but homogeneous as observed for other semi-arid environments where local factors were detected as key drivers of species assemblages (eg Schlesinger et al 1990 Perelman et al 1997 Diacuteaz et al 1999) The variables showing the strongest correlations with the constrained RDA components were disturbance intensity slope as-pect altitude percentage of outcropping rocks and to a lesser degree soil features (organic matter content pH and potassium concentration) High disturbance inten-sity (which had the best correlation with the first con-strained component) emerged as the main factor shaping the species composition of the dry Puna pastures and was positively correlated with pH dwarf shrubs and non-graminoid plant forms while it was negatively cor-related with organic matternitrogen soil content nurse plant abundance hemirosulate forbs and graminoids Consistently with previous research carried out in semi-arid environments (eg Ares et al 1990 Carilla et al 2012) high grazing intensity promotes the spread of spe-cies with avoidance strategies (Table 4) such as thorny dwarf shrubs (Tetraglochin cristatum) annual species (eg Muhlenbergia peruviana) and small or prostrate plants (eg Nassella nardoides Astragalus dielsii Hy-pochaeris echegarayi) Thorny dwarf shrubs are fostered by herbivory given that the tough and spiny stems of this
Table 1 Values of Shannon and Simpson diversity indices for the releveacute groups of the 4-leaf and 14-leaf Multivariate regression tree
Groups Diversity
Shannon (Hlsquo) Simpson (D)
T41 1628 0653
T42 1831 0626
T43 2176 0765
T44 2822 0829
T141 1511 0582
T142 1611 0688
T143 1831 0626
T144 1714 0633
T145 2366 0822
T146 1913 0763
T147 2348 0824
T148 1316 0598
T149 1540 0574
T1410 2214 0792
T1411 2283 0675
T1412 2149 0714
T1413 2100 0741
T1414 2685 0863
high outcropping rock cover South-facing groups were in turn separated by potassium content in the soil group T1413 with higher concentration of potassium shows a higher coverage of graminoid species (eg Poa pearsonii)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grass Festuca or-thophylla the cushion plants Pycnophyllum molle and P weberbaueri the shrub species Parastrephia lucida and P quadrangularis) This plant community was located near the altitudinal limit of vegetation and characterised by very low disturbance
The Redundancy Analysis executed on species and en-vironmental data explained 638 of the total variance (adjusted R2 = 047) The results of the RDA are in line with the MRT analysis the variables showing the strong-est correlation with the constrained components were disturbance intensity altitude percentage of outcropping rocks slope aspect soil organic matter content pH and potassium concentration (Fig 4) RDA biplot scores are reported in Appendix B It is worth noting that this result coincides with those discriminating for the partitions of the 4-leaf and 14-leaf MRTs (in order of diminishing cor-relation with RDA axes) Another correspondence be-tween MRT and RDA analyses lies in the placement of site points in the RDA graph as a matter of fact the
phyto_44_1_2_103_119_Catorciindd 110phyto_44_1_2_103_119_Catorciindd 110 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 111
Table 2 Indicator species of the releveacute groups highlighted by Indicator Species Analysis for the 4ndashleaf and the 14ndashleaf Multivariate regres-sion tree (MRT) and their respective indicator values and mean cover percentages Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value)
Indicator species 4-leaf MRT 14-leaf MRTMax
GroupIndicator
ValueMean
cover Max
GroupIndicator
ValueMean
cover Nassella nardoides T41 663 199 ndash ndash ndash
Nothoscordum andicola T41 482 08 ndash ndash ndash
Lepidium meyenii T41 412 06 ndash ndash ndash
Astragalus peruvianus T41 315 07 ndash ndash ndash
Tetraglochin cristatum T41 826 544 T141 422 628
Muhlenbergia peruviana T41 675 44 T142 300 46
Cumulopuntia ignescens T42 405 07 ndash ndash ndash
Chenopodium petiolare T42 370 03 ndash ndash ndash
Urocarpidium echinatum T42 350 06 ndash ndash ndash
Nototriche turritella T42 345 24 ndash ndash ndash
Jarava ichu T42 979 598 T143 925 598
Plantago linearis T42 901 30 T143 493 30
Diplostephium meyenii T42 700 23 T143 634 23
Crocopsis fulgens T42 571 16 T143 571 16
Belloa piptolepis T42 592 11 T143 351 11
Senecio phylloleptus T42 490 10 T143 364 10
Hypochaeris echegarayi T43 380 08 ndash ndash ndash
Pycnophyllum weberbaueri T43 342 08 ndash ndash ndash
Junellia minima (Meyen) T43 300 08 ndash ndash ndash
Calamagrostis breviaristata T43 704 430 T144 398 581
Calceolaria inamoena ndash ndash ndash T145 315 08
Astragalus dielsii T43 460 11 T146 321 14
Nassella nardoides ndash ndash ndash T146 484 415
Calandrinia acaulis ndash ndash ndash T146 430 14
Oenothera sp ndash ndash ndash T146 420 14
Festuca orthophylla T44 522 389 ndash ndash ndash
Trisetum spicatum T44 406 08 ndash ndash ndash
Nassella depauperata T44 352 70 T148 860 600
Lupinus saxatilis ndash ndash ndash T148 657 80
Descurainia myriophylla ndash ndash ndash T148 437 133
Erigeron pazensis ndash ndash ndash T148 416 07
Deyeuxia brevifolia T44 464 53 T1410 414 193
Mniodes pulvinata ndash ndash ndash T1410 375 05
Lupinus chlorolepis ndash ndash ndash T1411 364 09
Calamagrostis vicunarum ndash ndash ndash T1412 500 07
Poa gymnantha ndash ndash ndash T1412 485 42
Dielsiochloa floribunda ndash ndash ndash T1412 450 07
Leucheria daucifolia ndash ndash ndash T1412 398 07
Echinopsis pampana ndash ndash ndash T1412 384 42
Perezia ciliosa ndash ndash ndash T1412 329 07
Poa pearsonii ndash ndash ndash T1413 670 237
Deyeuxia rigida T44 316 47 T1414 774 285
Poa aequigluma ndash ndash ndash T1414 453 06
Werneria spathulata ndash ndash ndash T1414 412 08
Luzula racemosa ndash ndash ndash T1414 409 09
Lucilia sp ndash ndash ndash T1414 401 12
Senecio scorzoneraefolius ndash ndash ndash T1414 376 05
Gnaphalium badium ndash ndash ndash T1414 338 10
Pycnophyllum molle ndash ndash ndash T1414 335 95
phyto_44_1_2_103_119_Catorciindd 111phyto_44_1_2_103_119_Catorciindd 111 120314 0713120314 0713
eschweizerbart_xxx
112 A Catorci et al
Table 3 Descriptive statistics of the environmental variables of the groups highlighted by the 4-leaf and 14-leaf Multivariate regression tree 1st and 3rd quartiles are indicated for the whole data set mean and standard deviation (SD) are reported for each group
Group Descriptive statistics
Constraining variables
Site characteristics Soil texture Soil composition
Altitude(m asl)
Aspect (deg)
Slope (deg)
Outcr rocks()
Sand()
Silt ()
Clay ()
Org matt()
N () P (ppm)
K (ppm)
CaCO3 ()
pH
1st quartile 407000 000 200 000 6280 1660 580 089 004 1136 11250 008 540 3rd quartile 447500 9000 2000 1500 7760 2780 910 174 009 2469 23250 075 623
T41 Mean 399700 2363 755 000 7867 1403 1051 071 003 1265 16381 049 652SD 1908 2777 894 000 295 180 1434 033 001 668 3318 035 028
T42 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T43 Mean 410500 2880 364 000 7265 2133 602 110 009 2173 17036 053 542SD 2933 4752 930 000 389 372 237 053 020 1234 6129 051 035
T44 Mean 440287 8739 2061 1787 6698 2444 849 171 008 2219 19384 045 572SD 22159 5242 1431 1192 976 866 304 083 004 1471 11137 039 045
T141 Mean 399833 3938 1208 000 7926 1374 1233 052 003 875 17452 049 666SD 2462 2561 901 000 333 203 1853 024 001 142 3181 032 028
T142 Mean 399500 000 075 000 7778 1445 778 099 004 1850 14775 049 631SD 535 000 104 000 214 141 213 022 001 725 3011 041 010
T143 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T144 Mean 411393 643 064 000 7373 1987 640 098 012 2552 18143 052 529SD 1712 2405 145 000 309 274 306 020 027 1421 5395 062 035
T145 Mean 412667 10500 2167 000 6777 2577 647 210 008 1611 18335 071 549SD 4619 1299 2082 000 224 145 093 017 001 516 13137 017 031
T146 Mean 409500 000 000 000 7385 2128 488 078 005 2370 18117 015 575SD 000 000 000 000 042 026 017 003 001 327 1019 019 018
T147 Mean 406750 7875 425 000 7135 2315 550 110 004 1073 11106 082 551SD 3663 5356 299 000 654 654 081 094 004 698 1577 019 031
T148 Mean 414800 7200 1900 1500 7368 2080 552 135 007 2044 9024 019 630SD 570 1006 224 000 618 567 064 037 002 1111 4791 021 015
T149 Mean 408833 9000 600 1833 6960 2093 947 096 005 1216 18803 054 566SD 5795 000 346 577 763 412 357 067 004 098 12521 057 071
T1410 Mean 458688 4922 2731 1813 7623 1591 787 136 006 2448 17087 042 547SD 22773 4043 1590 680 622 535 164 026 001 1461 9485 047 049
T1411 Mean 439614 5727 1114 2173 6501 2430 1038 147 007 1741 22177 052 583SD 16079 2840 551 1297 832 588 420 053 003 597 10086 035 049
T1412 Mean 445650 9000 4000 550 6025 3248 725 248 013 1653 25562 055 579SD 27365 000 2121 636 078 046 120 049 002 467 26427 040 013
T1413 Mean 421318 15341 3518 2409 5954 3227 819 285 014 2823 28413 025 564SD 11402 2814 1419 1338 1099 981 169 117 006 2824 11501 041 022
T1414 Mean 454300 14850 1600 590 6189 3104 707 183 009 2740 11102 065 571SD 6997 2846 615 1058 589 588 071 063 003 830 2759 027 026
life form discourage animals from biting it (Briske 1996) The observed spread of dwarf shrubs is in accordance with the findings of several studies carried out in semiarid environments (Perelman et al 1997 Swinton amp Quiroz 2003 Carilla et al 2012 Catorci et al 2012b 2013) and in pastoral ecosystems subjected to periodic fires (Liedloff
et al 2001) In semi-arid environments annual life span may be considered an avoidance strategy fostered by high grazing intensity (Diacuteaz et al 1999 McGregor 2002 Catorci et al 2012b) because it allows plants to dynami-cally face the grazing pressure by restarting the growth cycle every year (Diacuteaz et al 1999 2001 McGregor 2002)
phyto_44_1_2_103_119_Catorciindd 112phyto_44_1_2_103_119_Catorciindd 112 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

106 A Catorci et al
For each releveacute we collected soil samples and data on altitude (m asl) slope aspect (azimuth degrees) slope angle (vertical degrees) and rockiness Within each plot 5 separate soil samples were taken at a depth of 2 to 20 cm below the soil surface and combined to one bulked sam-ple Grazing intensity and occurrence of fire were deter-mined by more than ten interviews with local villagers and the rangers of the Salinas and Aguada Blanca Na-tional Reserve Releveacutes were carried out at the end of the rainy season (March 2010) when soil moisture is highest and the highest number of species can be observed
Data on the following plant traits were collected by field observations and through consultation of the Her-barium of the Universidad Nacional de San Agustin (HUSA) of Arequipa (Peru) and of the Herbarium at the Universidad Nacional Mayor de San Marcos (USM) of Lima (Peru) annual and perennial life span graminoid species non-graminoid species upright forb (plant with leaves distributed on upright stems or branches) hemiro-sulate forb (plant with leaves distributed both at ground level and raised from the ground) prostrate forb (plant with leaves distributed only at the ground level) tall shrub (gt 15 cm) dwarf shrub (lt 15 cm) cushion and succulent plants
As it has been demonstrated that in harsh environ-ments positive plant-plant spatial interaction between nurse and beneficiary species play a key role in shaping the community richness and diversity (Armas amp Pug-naire 2005 Catorci et al 2011b Kikvidze et al 2011) fol-lowing several authors (Rebollo et al 2002 Maestre amp Cortina 2005 Padilla amp Pugnaire 2006 Arredondo-Nu-ntildeez et al 2009 Monteiro et al 2011) we chose tall grass tussocks tall shrubs and cushions as potential nurse plants considering the potential nurse role as an addi-tional functional trait
Species nomenclature mainly follows Brako amp Zarucchi (1993) Later taxonomic changes were checked by consult-ing IPNI (httpwwwipniorgindexhtml) Tropicos (httpwwwtropicosorg) and other botanical studies from close geographic areas (eg Araraki amp Cano 2003 Schwarzer et al 2010 Montesinos 2011 Montesinos et al 2012)
Data analysis
Soil samples were analysed by the Instituto Nacional de Innovacion Agraria (INIA) soil analysis laboratory of Arequipa (Ministerio de Agricultura y Riego del Peru) The following soil parameters were determined (the methods applied are indicated in parentheses) texture ie sand loam and clay (Buoyuocosrsquo method) or-ganic matter (modified Walkley Blackrsquos method) ni-trogen (Micro Kjeldahlrsquos method) phosphorus in ppm (modified Olsenrsquos method 05 M NaHCO3 extrac-tion at pH 88) potassium in ppm (flame photometry
by ammonium acetate at pH 70) CaCO3 (Wesemaelrsquos method) pH (by potentiometer in a 125 soilwater sus-pension)
To assess floristic composition the species cover val-ues were transformed into percentages using the central values of the Braun-Blanquet scale (Podani 2007) Fol-lowing Warren (2008) aspect azimuth was converted from the 0ndash360 compass scale to a linear (0ndash180) scale giving northerly aspect a value approaching 0 and south-erly aspect a value approaching 180 Disturbance inten-sity was assessed using a 4-level scale as follows 1) ab-sence of grazing (absent) 2) presence of wild camelids and sporadically small herds of domestic camelids (low) 3) grazing of large herds of domestic camelids (moder-ate) 4) grazing of large herds of domestic camelids and periodic fires (strong)
In order to assess the influence of environmental vari-ables and disturbance intensity on the species assemblage and identify homogeneous groups of releveacutes a con-strained clustering using Multivariate regression tree (MRT) analysis (Dersquoath 2002) with 100 iterations and 29 cross-validation groups was executed on two matrices namely ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo Prior to MRT analy-sis species data have been transformed using chord trans-formation (Legendre amp Gallagher 2001) which expresses the data as relative abundances per sites This removes from the data the total abundance per site which is the response of the species to the total productivity of the sites (Borcard et al 2011) The result of MRT analysis is a tree whose ldquoleavesrdquo (terminal groups of releveacutes) are com-posed of subsets of releveacutes chosen to minimize the within-group sum of squares and each partition of the tree is defined by a threshold value of one explanatory variable (Borcard et al 2011) Cross-validation is a re-sampling analysis repeated a number of times for succes-sive and independent divisions of the data (in this case species cover) into random test groups of objects (rel-eveacutes) (Legendre amp Legendre 2012) An overall error sta-tistic (cross-validation relative error CVRE) was com-puted for each test group and partition size (number of groups) to choose the optimal size of the tree CVRE varies from zero for a perfect set of predictors chosen for the splits of a tree to close to one for poor predictors For each partition size the mean and standard error of all CVRE estimates was computed Generally the smallest tree (ie a tree with fewer splits) within one standard er-ror of the smallest CVRE value is selected (Breiman et al 1984) but one may select the tree that has the smallest CVRE value as well (Legendre amp Legendre 2012)
To provide a general overview of the ecological charac-teristics of the whole releveacute data set we calculated the 1st and 3rd quartiles for each environmental variable The mean value and the standard deviation of the same varia-bles were calculated for each releveacute group that emerged from the MRT analysis
phyto_44_1_2_103_119_Catorciindd 106phyto_44_1_2_103_119_Catorciindd 106 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 107
The species diversity of each group was assessed by calculating the Shannon-Wiener (Hrsquo) and Gini-Simpson (D) diversity indices while the dissimilarity between the groups was computed through the Jaccard distance index (Magurran 1988) In order to investigate the species com-position of the groups we performed the Indicator Spe-cies Analysis (ISA) a non-parametric method for identi-fying those items (speciestrait) that show significantly preferential distribution (frequency and abundance) compared to an a priori treatment group An indicator value is calculated by multiplying the relative abundance of each item in a particular group and the relative fre-quency of the item occurrence in the sample of that group (Dufrecircne amp Legendre 1997) The number of randomized indicator values higher than the observed ones is used to calculate the probability value (McCune amp Grace 2002) The statistical significance (P lt 005) of the observed maximum Indicator Values (IV) was tested using permu-tation tests with 4999 iterations Only Indicator Values higher than 30 were considered of interest
To assess the influence of the ecological factors on spe-cies composition and abundance we performed a transfor-mation-based Redundancy Analysis (tb-RDA ndash Legendre amp Gallagher 2001) on the same matrices processed for MRT analysis Disturbance was considered as a factor var-iable with the above-mentioned four levels (absent low moderate strong) thus each of these levels was repre-sented by its centroid instead of an arrow in the RDA plot
To assess the influence of the ecological factors on the trait composition of the plant communities we first transformed trait binary data (presenceabsence) into quantitative data (ie aggregated cover values of each trait) The ldquoreleveacutes x species coverrdquo matrix was multiplied by the ldquospecies x traitrdquo matrix to provide a ldquoreleveacutes x trait coverrdquo matrix (Pakeman et al 2009) which formed the basis for the following analyses Then we ran a tb-RDA on the ldquoreleveacutes x trait coverrdquo matrix using the above-mentioned parameters (environmental factors and distur-bance intensity) as constraining variables
Since the disturbance gradient emerged as the main factor affecting trait composition in dry Puna communi-ties in order to better understand its effect on trait com-position we performed an ISA on the above matrix using MRT groups as grouping variable As the first partition of the regression tree segregated releveacutes characterised by strong disturbance intensity we re-ran ISA comparing releveacutes of this group against all the other ones The statis-tical significance (P lt 005) of the observed maximum In-dicator Values (IV) was tested using permutation tests with 4999 iterations Only Indicator Values higher than 30 were considered of interest
We used the R software (version 2140 ndash R Core Team 2012 ndash httpwwwR-projectorg) and its mvpart labdsv and vegan packages (Oksanen et al 2013) to perform MRT analysis ISA tb-RDA and to calculate diversity and dis-tance indices besides basic statistics
Results
MRT analysis generated two trees with the same CVRE (0641) composed of 4 and 14 leaves respectively (Fig 2) The 4-leaf tree had a smaller standard error (0051) than the 14-leaf tree (0062) and was selected as the best solution more frequently during the cross-validation it-erations Nevertheless to avoid losing information po-tentially useful for the ecological interpretation of the distribution of plant communities as the CVRE of the 14-leaf tree fell within one standard error of the smallest CVRE value we decided to consider this tree as well We recorded 136 species (belonging to 69 genera and 28 fam-ilies) The species composition of each group highlighted by MRT is shown in Fig 3 while the related distance in-dices are given in Appendix A The values of the Shan-non-Wiener and the Gini-Simpson diversity indices are reported in Table 1 The indicator species identified by ISA for groups highlighted by MRT as well as their mean abundances are shown in Table 2 Table 3 indicates the mean the median and the 1st and 3rd quartiles for the eco-logical variables in each group Table 2 shows the output of ISA concerning the assessment of the indicator traits of the four relevegrave groups of the 4-leaf and 14-leaf trees The variable that discriminated between the two branches in the first node of the 4-leaves MRT was grazing distur-bance thus the first leaf (T41) was composed of the re-leveacutes characterised by a high level of disturbance The related plant communities were dominated by the dwarf shrub Tetraglochin cristatum they presented the lowest species diversity (Hrsquo = 163 D = 065) as well as the high-est distances compared to the other leaves Other envi-ronmental features of the T cristatum ndash dominated plant community were (Table 3) high sand percentage and low soil values of organic matter nitrogen phosphorus and potassium The pH value was the highest one compared to the other groups of releveacutes Dwarf shrubs non-grami-noid and annual species were the indicator traits of this group compared with all the other releveacutes jointly consid-ered (Table 4)
The second partition was defined by the percentage of outcropping rocks The first branch of this partition characterised by the lower values of outcropping rocks is in turn divided into two leaves the first (T42) grouping releveacutes located below 4000 m of altitude is dominated by Jarava ichu while Calamagrostis breviaristata domi-nates the second one (T43) grouping the releveacutes placed above 4000 m Both these groups were subjected to in-termediate level of disturbance and were characterised by flat or semi-flat slopes The Jarava ichu ndash dominated plant community was distinguished by low sand percent-age and relatively high organic matter phosphorus and potassium content in soil (Table 3) The pH value was similar to that of groups T43 and T44 groups Grami-noid and hemirosulate forbs as well as succulent and po-
phyto_44_1_2_103_119_Catorciindd 107phyto_44_1_2_103_119_Catorciindd 107 120314 0713120314 0713
eschweizerbart_xxx
108 A Catorci et al
Fig 2 The 4-leaf (a) and 14-leaf (b) Multivariate regression trees for the species data set constrained by their explanatory variables The threshold values shown for each partition of the tree correspond to the mean of the two limit values of the considered variables at the break between the branches The relative abundances of the species are shown in histograms at the tips of the branches with the species in the same order as in the input file the species names and abun-dances are shown in Figure 3 Under each histogram the sum of squared errors for the group and the number of releveacutes in the leaf (n) are indicated Below each tree the residual error (Error) the cross-validation error (CV Error) and standard error (SE) are indi-cated Alt ndash Altitude Asp_deg ndash aspect Dist_L ndash Disturbance level Hum_p ndash organic matter K_ppm ndash Potassium in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt
tential nurse plants were the indicator traits for the T42 group (Table 4)
The C breviaristata ndash dominated plant community was characterized by high sand percentage and low or-ganic matter and potassium content in the soil (Table 3) The pH value was similar to that of groups T43 and T44 Cushion and perennial plants were the indicator traits for group T43 (Table 4)
The releveacutes grouped in the fourth leaf (T44) are char-acterised by a percentage of outcropping rocks greater than 1 (mean cover value 178 ) In this group the dominant species is the graminoid Festuca orthophylla other indicator species being Deyeuxia brevifolia Trise-tum spicatum Nassella depauperata and Deyeuxia rigida (all grass species) Group T44 includes the greatest num-ber of releveacutes (69) and presents the highest diversity (Hrsquo = 282 D = 083) in the 4-leaved tree Other environmen-
tal features of the Festuca orthophylla ndash dominated plant community were (Table 3) the highest slopes altitude and angles among the four groups highlighted by MRT low sand percentage and high values of organic matter nitro-gen and phosphorus in the soil The pH value was similar to that of groups T42 and T43 These pastures were characterised by low disturbance intensity Within the high richness and internal variability of these grasslands particular environmental conditions or treatments caused minor differences in species composition highlighting different plant communities as shown by the 14-leaved MRT (Fig 2)
Despite the high number of groups yielded the val-ues of community diversity indices computed from the pairwise comparison of the 14 leaves were still high (never below 45 ) (Appendix A) The first partition of the Festuca orthophylla ndash dominated grassland was de-
phyto_44_1_2_103_119_Catorciindd 108phyto_44_1_2_103_119_Catorciindd 108 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 109
fined by aspect with a threshold value that can be inter-preted as the boundary between north-east-facing slopes (arid more stressful ndash groups T148-T1412) and south-west-facing ones (cool less stressful ndash groups T1413 and T1414) From a functional point of view the south-facing plots were characterized by a high number of indicator traits (perennial species graminoids hemi-rosulate forbs cushion plants and potential nurses) while no traits emerged as indicator of the plant com-munities growing on the north-facing slopes In addi-tion north-facing groups were characterised by a higher degree of dominance with the most widespread species often representing more than 50 of the total vegeta-tion cover
Altitude gradients and soil parameters defined further partitions for a more detailed differentiation of these two main clusters Among north-facing groups T148 was dominated by Nassella depauperata and had coarser soil as well as higher pH and organic matter values than T149 Groups T1410 T1411 and T1412 were charac-terised by higher altitudes (about 400 m above T148 and T149 ndash Table 3) T1410 was characterised by low distur-bance intensity and showed a more even distribution of species (particularly if compared with T1411) among which several palatable grasses are present (eg Deyeuxia brevifolia) Group T1412 dominated by the shrub spe-cies Chersodoma jodopappa showed a higher percentage of silt and organic matter in soil and steeper slopes with
Fig 3 Pie charts reporting the species composition of the groups defined in the 4-leaf (a) and in the 14-leaf (b) Multivariate regres-sion trees Species abbreviations are explained below Ast_die ndash As-tragalus dielsii Bac_tri ndash Baccharis tricuneata Bel_lon ndash Belloa longifolia Cal_bra ndash Calamagrostis breviaristata Can_aca ndash Calan-drina acaulis Chs_jod ndash Chersodoma jodopappa Cro_ful ndash Crocop-sis fulgens Des_myr ndash Descurainia myriophylla Dey_brf ndash De-yeuxia brevifolia Dey_rig ndash Deyeuxia rigida Dip_mey ndash Diplostephium meyenii Ech_pam ndash Echinopsis pampana Fes_ort ndash Festuca ortophylla Gna_lac ndash Gnaphalium lacteum Hyp_ech ndash Hypochaeris echegarayi Jar_ich ndash Jarava ichu Lpp_qua ndash Lepi-dopyllum quadrangulare Lup_sax ndash Lupinus saxatilis Muh_per ndash Muhlenbergia peruviana Nas_dep ndash Nassella depauperata Nas_nar ndash Nassella nardoides Ntt_tur ndash Nototriche turritella Oen_sp ndash Oenothera sp Par_luc ndash Parastrephia lucida Par_qua ndash Parastre-phia quadrangularis Pla_ser ndash Plantago sericea var lanuginosa Poa_gym ndash Poa gymnantha Poa_pea ndash Poa pearsonii Pyc_mol ndash Pycnophyllum molle Pyc_web ndash Pycnophyllum weberbaueri Sen_cha ndash Senecio chachaniensis Sen_nut ndash Senecio nutans Sti_han ndash Stipa hans-meyeri Tet_cri ndash Tetraglochin cristatum Tri_spi ndash Trise-tum spicatum Wer_pyg ndash Werneria pygmaea
phyto_44_1_2_103_119_Catorciindd 109phyto_44_1_2_103_119_Catorciindd 109 120314 0713120314 0713
eschweizerbart_xxx
110 A Catorci et al
groups yielded by the 4-leaf tree are clearly recognisable in the RDA three-dimensional scatterplot (as highlighted in Fig 4) The RDA performed on functional traits data with the environmental variables as constraints yielded similar results but explained a notably smaller fraction of the total variance (92 adjusted R2 = 056) The varia-bles showing the strongest correlation with the con-strained components coincided with those observed in the first RDA but for most environmental variables the observed correlation is weaker this is the case of altitude aspect percentage of outcropping rocks sand and silt percentage in the soil (Fig 5) Instead for disturbance in-tensity the observed correlation was slightly stronger The functional traits whose variance was best explained by the constrained components were nurse ability and various types of plant forms dwarf shrubs upright forbs non-graminoids hemirosulate forbs and graminoids (Fig 5) Non-graminoid species dwarf shrubs and up-right forbs were related to relatively higher soil pH and strong disturbance intensity nurse species were linked to higher altitudes South-facing slopes relatively lower sand percentages in the soil graminoid species and hemi-rosulate forbs were particularly tied to more fertile soils (higher phosphorus and nitrogen concentrations) and low disturbance intensity (Fig 5) RDA biplot scores are reported in the Appendix C
Discussion
Our results demonstrate that the dry Puna landscape is anything but homogeneous as observed for other semi-arid environments where local factors were detected as key drivers of species assemblages (eg Schlesinger et al 1990 Perelman et al 1997 Diacuteaz et al 1999) The variables showing the strongest correlations with the constrained RDA components were disturbance intensity slope as-pect altitude percentage of outcropping rocks and to a lesser degree soil features (organic matter content pH and potassium concentration) High disturbance inten-sity (which had the best correlation with the first con-strained component) emerged as the main factor shaping the species composition of the dry Puna pastures and was positively correlated with pH dwarf shrubs and non-graminoid plant forms while it was negatively cor-related with organic matternitrogen soil content nurse plant abundance hemirosulate forbs and graminoids Consistently with previous research carried out in semi-arid environments (eg Ares et al 1990 Carilla et al 2012) high grazing intensity promotes the spread of spe-cies with avoidance strategies (Table 4) such as thorny dwarf shrubs (Tetraglochin cristatum) annual species (eg Muhlenbergia peruviana) and small or prostrate plants (eg Nassella nardoides Astragalus dielsii Hy-pochaeris echegarayi) Thorny dwarf shrubs are fostered by herbivory given that the tough and spiny stems of this
Table 1 Values of Shannon and Simpson diversity indices for the releveacute groups of the 4-leaf and 14-leaf Multivariate regression tree
Groups Diversity
Shannon (Hlsquo) Simpson (D)
T41 1628 0653
T42 1831 0626
T43 2176 0765
T44 2822 0829
T141 1511 0582
T142 1611 0688
T143 1831 0626
T144 1714 0633
T145 2366 0822
T146 1913 0763
T147 2348 0824
T148 1316 0598
T149 1540 0574
T1410 2214 0792
T1411 2283 0675
T1412 2149 0714
T1413 2100 0741
T1414 2685 0863
high outcropping rock cover South-facing groups were in turn separated by potassium content in the soil group T1413 with higher concentration of potassium shows a higher coverage of graminoid species (eg Poa pearsonii)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grass Festuca or-thophylla the cushion plants Pycnophyllum molle and P weberbaueri the shrub species Parastrephia lucida and P quadrangularis) This plant community was located near the altitudinal limit of vegetation and characterised by very low disturbance
The Redundancy Analysis executed on species and en-vironmental data explained 638 of the total variance (adjusted R2 = 047) The results of the RDA are in line with the MRT analysis the variables showing the strong-est correlation with the constrained components were disturbance intensity altitude percentage of outcropping rocks slope aspect soil organic matter content pH and potassium concentration (Fig 4) RDA biplot scores are reported in Appendix B It is worth noting that this result coincides with those discriminating for the partitions of the 4-leaf and 14-leaf MRTs (in order of diminishing cor-relation with RDA axes) Another correspondence be-tween MRT and RDA analyses lies in the placement of site points in the RDA graph as a matter of fact the
phyto_44_1_2_103_119_Catorciindd 110phyto_44_1_2_103_119_Catorciindd 110 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 111
Table 2 Indicator species of the releveacute groups highlighted by Indicator Species Analysis for the 4ndashleaf and the 14ndashleaf Multivariate regres-sion tree (MRT) and their respective indicator values and mean cover percentages Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value)
Indicator species 4-leaf MRT 14-leaf MRTMax
GroupIndicator
ValueMean
cover Max
GroupIndicator
ValueMean
cover Nassella nardoides T41 663 199 ndash ndash ndash
Nothoscordum andicola T41 482 08 ndash ndash ndash
Lepidium meyenii T41 412 06 ndash ndash ndash
Astragalus peruvianus T41 315 07 ndash ndash ndash
Tetraglochin cristatum T41 826 544 T141 422 628
Muhlenbergia peruviana T41 675 44 T142 300 46
Cumulopuntia ignescens T42 405 07 ndash ndash ndash
Chenopodium petiolare T42 370 03 ndash ndash ndash
Urocarpidium echinatum T42 350 06 ndash ndash ndash
Nototriche turritella T42 345 24 ndash ndash ndash
Jarava ichu T42 979 598 T143 925 598
Plantago linearis T42 901 30 T143 493 30
Diplostephium meyenii T42 700 23 T143 634 23
Crocopsis fulgens T42 571 16 T143 571 16
Belloa piptolepis T42 592 11 T143 351 11
Senecio phylloleptus T42 490 10 T143 364 10
Hypochaeris echegarayi T43 380 08 ndash ndash ndash
Pycnophyllum weberbaueri T43 342 08 ndash ndash ndash
Junellia minima (Meyen) T43 300 08 ndash ndash ndash
Calamagrostis breviaristata T43 704 430 T144 398 581
Calceolaria inamoena ndash ndash ndash T145 315 08
Astragalus dielsii T43 460 11 T146 321 14
Nassella nardoides ndash ndash ndash T146 484 415
Calandrinia acaulis ndash ndash ndash T146 430 14
Oenothera sp ndash ndash ndash T146 420 14
Festuca orthophylla T44 522 389 ndash ndash ndash
Trisetum spicatum T44 406 08 ndash ndash ndash
Nassella depauperata T44 352 70 T148 860 600
Lupinus saxatilis ndash ndash ndash T148 657 80
Descurainia myriophylla ndash ndash ndash T148 437 133
Erigeron pazensis ndash ndash ndash T148 416 07
Deyeuxia brevifolia T44 464 53 T1410 414 193
Mniodes pulvinata ndash ndash ndash T1410 375 05
Lupinus chlorolepis ndash ndash ndash T1411 364 09
Calamagrostis vicunarum ndash ndash ndash T1412 500 07
Poa gymnantha ndash ndash ndash T1412 485 42
Dielsiochloa floribunda ndash ndash ndash T1412 450 07
Leucheria daucifolia ndash ndash ndash T1412 398 07
Echinopsis pampana ndash ndash ndash T1412 384 42
Perezia ciliosa ndash ndash ndash T1412 329 07
Poa pearsonii ndash ndash ndash T1413 670 237
Deyeuxia rigida T44 316 47 T1414 774 285
Poa aequigluma ndash ndash ndash T1414 453 06
Werneria spathulata ndash ndash ndash T1414 412 08
Luzula racemosa ndash ndash ndash T1414 409 09
Lucilia sp ndash ndash ndash T1414 401 12
Senecio scorzoneraefolius ndash ndash ndash T1414 376 05
Gnaphalium badium ndash ndash ndash T1414 338 10
Pycnophyllum molle ndash ndash ndash T1414 335 95
phyto_44_1_2_103_119_Catorciindd 111phyto_44_1_2_103_119_Catorciindd 111 120314 0713120314 0713
eschweizerbart_xxx
112 A Catorci et al
Table 3 Descriptive statistics of the environmental variables of the groups highlighted by the 4-leaf and 14-leaf Multivariate regression tree 1st and 3rd quartiles are indicated for the whole data set mean and standard deviation (SD) are reported for each group
Group Descriptive statistics
Constraining variables
Site characteristics Soil texture Soil composition
Altitude(m asl)
Aspect (deg)
Slope (deg)
Outcr rocks()
Sand()
Silt ()
Clay ()
Org matt()
N () P (ppm)
K (ppm)
CaCO3 ()
pH
1st quartile 407000 000 200 000 6280 1660 580 089 004 1136 11250 008 540 3rd quartile 447500 9000 2000 1500 7760 2780 910 174 009 2469 23250 075 623
T41 Mean 399700 2363 755 000 7867 1403 1051 071 003 1265 16381 049 652SD 1908 2777 894 000 295 180 1434 033 001 668 3318 035 028
T42 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T43 Mean 410500 2880 364 000 7265 2133 602 110 009 2173 17036 053 542SD 2933 4752 930 000 389 372 237 053 020 1234 6129 051 035
T44 Mean 440287 8739 2061 1787 6698 2444 849 171 008 2219 19384 045 572SD 22159 5242 1431 1192 976 866 304 083 004 1471 11137 039 045
T141 Mean 399833 3938 1208 000 7926 1374 1233 052 003 875 17452 049 666SD 2462 2561 901 000 333 203 1853 024 001 142 3181 032 028
T142 Mean 399500 000 075 000 7778 1445 778 099 004 1850 14775 049 631SD 535 000 104 000 214 141 213 022 001 725 3011 041 010
T143 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T144 Mean 411393 643 064 000 7373 1987 640 098 012 2552 18143 052 529SD 1712 2405 145 000 309 274 306 020 027 1421 5395 062 035
T145 Mean 412667 10500 2167 000 6777 2577 647 210 008 1611 18335 071 549SD 4619 1299 2082 000 224 145 093 017 001 516 13137 017 031
T146 Mean 409500 000 000 000 7385 2128 488 078 005 2370 18117 015 575SD 000 000 000 000 042 026 017 003 001 327 1019 019 018
T147 Mean 406750 7875 425 000 7135 2315 550 110 004 1073 11106 082 551SD 3663 5356 299 000 654 654 081 094 004 698 1577 019 031
T148 Mean 414800 7200 1900 1500 7368 2080 552 135 007 2044 9024 019 630SD 570 1006 224 000 618 567 064 037 002 1111 4791 021 015
T149 Mean 408833 9000 600 1833 6960 2093 947 096 005 1216 18803 054 566SD 5795 000 346 577 763 412 357 067 004 098 12521 057 071
T1410 Mean 458688 4922 2731 1813 7623 1591 787 136 006 2448 17087 042 547SD 22773 4043 1590 680 622 535 164 026 001 1461 9485 047 049
T1411 Mean 439614 5727 1114 2173 6501 2430 1038 147 007 1741 22177 052 583SD 16079 2840 551 1297 832 588 420 053 003 597 10086 035 049
T1412 Mean 445650 9000 4000 550 6025 3248 725 248 013 1653 25562 055 579SD 27365 000 2121 636 078 046 120 049 002 467 26427 040 013
T1413 Mean 421318 15341 3518 2409 5954 3227 819 285 014 2823 28413 025 564SD 11402 2814 1419 1338 1099 981 169 117 006 2824 11501 041 022
T1414 Mean 454300 14850 1600 590 6189 3104 707 183 009 2740 11102 065 571SD 6997 2846 615 1058 589 588 071 063 003 830 2759 027 026
life form discourage animals from biting it (Briske 1996) The observed spread of dwarf shrubs is in accordance with the findings of several studies carried out in semiarid environments (Perelman et al 1997 Swinton amp Quiroz 2003 Carilla et al 2012 Catorci et al 2012b 2013) and in pastoral ecosystems subjected to periodic fires (Liedloff
et al 2001) In semi-arid environments annual life span may be considered an avoidance strategy fostered by high grazing intensity (Diacuteaz et al 1999 McGregor 2002 Catorci et al 2012b) because it allows plants to dynami-cally face the grazing pressure by restarting the growth cycle every year (Diacuteaz et al 1999 2001 McGregor 2002)
phyto_44_1_2_103_119_Catorciindd 112phyto_44_1_2_103_119_Catorciindd 112 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

Floristic composition of dry Puna in the southern Peruvian Andes 107
The species diversity of each group was assessed by calculating the Shannon-Wiener (Hrsquo) and Gini-Simpson (D) diversity indices while the dissimilarity between the groups was computed through the Jaccard distance index (Magurran 1988) In order to investigate the species com-position of the groups we performed the Indicator Spe-cies Analysis (ISA) a non-parametric method for identi-fying those items (speciestrait) that show significantly preferential distribution (frequency and abundance) compared to an a priori treatment group An indicator value is calculated by multiplying the relative abundance of each item in a particular group and the relative fre-quency of the item occurrence in the sample of that group (Dufrecircne amp Legendre 1997) The number of randomized indicator values higher than the observed ones is used to calculate the probability value (McCune amp Grace 2002) The statistical significance (P lt 005) of the observed maximum Indicator Values (IV) was tested using permu-tation tests with 4999 iterations Only Indicator Values higher than 30 were considered of interest
To assess the influence of the ecological factors on spe-cies composition and abundance we performed a transfor-mation-based Redundancy Analysis (tb-RDA ndash Legendre amp Gallagher 2001) on the same matrices processed for MRT analysis Disturbance was considered as a factor var-iable with the above-mentioned four levels (absent low moderate strong) thus each of these levels was repre-sented by its centroid instead of an arrow in the RDA plot
To assess the influence of the ecological factors on the trait composition of the plant communities we first transformed trait binary data (presenceabsence) into quantitative data (ie aggregated cover values of each trait) The ldquoreleveacutes x species coverrdquo matrix was multiplied by the ldquospecies x traitrdquo matrix to provide a ldquoreleveacutes x trait coverrdquo matrix (Pakeman et al 2009) which formed the basis for the following analyses Then we ran a tb-RDA on the ldquoreleveacutes x trait coverrdquo matrix using the above-mentioned parameters (environmental factors and distur-bance intensity) as constraining variables
Since the disturbance gradient emerged as the main factor affecting trait composition in dry Puna communi-ties in order to better understand its effect on trait com-position we performed an ISA on the above matrix using MRT groups as grouping variable As the first partition of the regression tree segregated releveacutes characterised by strong disturbance intensity we re-ran ISA comparing releveacutes of this group against all the other ones The statis-tical significance (P lt 005) of the observed maximum In-dicator Values (IV) was tested using permutation tests with 4999 iterations Only Indicator Values higher than 30 were considered of interest
We used the R software (version 2140 ndash R Core Team 2012 ndash httpwwwR-projectorg) and its mvpart labdsv and vegan packages (Oksanen et al 2013) to perform MRT analysis ISA tb-RDA and to calculate diversity and dis-tance indices besides basic statistics
Results
MRT analysis generated two trees with the same CVRE (0641) composed of 4 and 14 leaves respectively (Fig 2) The 4-leaf tree had a smaller standard error (0051) than the 14-leaf tree (0062) and was selected as the best solution more frequently during the cross-validation it-erations Nevertheless to avoid losing information po-tentially useful for the ecological interpretation of the distribution of plant communities as the CVRE of the 14-leaf tree fell within one standard error of the smallest CVRE value we decided to consider this tree as well We recorded 136 species (belonging to 69 genera and 28 fam-ilies) The species composition of each group highlighted by MRT is shown in Fig 3 while the related distance in-dices are given in Appendix A The values of the Shan-non-Wiener and the Gini-Simpson diversity indices are reported in Table 1 The indicator species identified by ISA for groups highlighted by MRT as well as their mean abundances are shown in Table 2 Table 3 indicates the mean the median and the 1st and 3rd quartiles for the eco-logical variables in each group Table 2 shows the output of ISA concerning the assessment of the indicator traits of the four relevegrave groups of the 4-leaf and 14-leaf trees The variable that discriminated between the two branches in the first node of the 4-leaves MRT was grazing distur-bance thus the first leaf (T41) was composed of the re-leveacutes characterised by a high level of disturbance The related plant communities were dominated by the dwarf shrub Tetraglochin cristatum they presented the lowest species diversity (Hrsquo = 163 D = 065) as well as the high-est distances compared to the other leaves Other envi-ronmental features of the T cristatum ndash dominated plant community were (Table 3) high sand percentage and low soil values of organic matter nitrogen phosphorus and potassium The pH value was the highest one compared to the other groups of releveacutes Dwarf shrubs non-grami-noid and annual species were the indicator traits of this group compared with all the other releveacutes jointly consid-ered (Table 4)
The second partition was defined by the percentage of outcropping rocks The first branch of this partition characterised by the lower values of outcropping rocks is in turn divided into two leaves the first (T42) grouping releveacutes located below 4000 m of altitude is dominated by Jarava ichu while Calamagrostis breviaristata domi-nates the second one (T43) grouping the releveacutes placed above 4000 m Both these groups were subjected to in-termediate level of disturbance and were characterised by flat or semi-flat slopes The Jarava ichu ndash dominated plant community was distinguished by low sand percent-age and relatively high organic matter phosphorus and potassium content in soil (Table 3) The pH value was similar to that of groups T43 and T44 groups Grami-noid and hemirosulate forbs as well as succulent and po-
phyto_44_1_2_103_119_Catorciindd 107phyto_44_1_2_103_119_Catorciindd 107 120314 0713120314 0713
eschweizerbart_xxx
108 A Catorci et al
Fig 2 The 4-leaf (a) and 14-leaf (b) Multivariate regression trees for the species data set constrained by their explanatory variables The threshold values shown for each partition of the tree correspond to the mean of the two limit values of the considered variables at the break between the branches The relative abundances of the species are shown in histograms at the tips of the branches with the species in the same order as in the input file the species names and abun-dances are shown in Figure 3 Under each histogram the sum of squared errors for the group and the number of releveacutes in the leaf (n) are indicated Below each tree the residual error (Error) the cross-validation error (CV Error) and standard error (SE) are indi-cated Alt ndash Altitude Asp_deg ndash aspect Dist_L ndash Disturbance level Hum_p ndash organic matter K_ppm ndash Potassium in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt
tential nurse plants were the indicator traits for the T42 group (Table 4)
The C breviaristata ndash dominated plant community was characterized by high sand percentage and low or-ganic matter and potassium content in the soil (Table 3) The pH value was similar to that of groups T43 and T44 Cushion and perennial plants were the indicator traits for group T43 (Table 4)
The releveacutes grouped in the fourth leaf (T44) are char-acterised by a percentage of outcropping rocks greater than 1 (mean cover value 178 ) In this group the dominant species is the graminoid Festuca orthophylla other indicator species being Deyeuxia brevifolia Trise-tum spicatum Nassella depauperata and Deyeuxia rigida (all grass species) Group T44 includes the greatest num-ber of releveacutes (69) and presents the highest diversity (Hrsquo = 282 D = 083) in the 4-leaved tree Other environmen-
tal features of the Festuca orthophylla ndash dominated plant community were (Table 3) the highest slopes altitude and angles among the four groups highlighted by MRT low sand percentage and high values of organic matter nitro-gen and phosphorus in the soil The pH value was similar to that of groups T42 and T43 These pastures were characterised by low disturbance intensity Within the high richness and internal variability of these grasslands particular environmental conditions or treatments caused minor differences in species composition highlighting different plant communities as shown by the 14-leaved MRT (Fig 2)
Despite the high number of groups yielded the val-ues of community diversity indices computed from the pairwise comparison of the 14 leaves were still high (never below 45 ) (Appendix A) The first partition of the Festuca orthophylla ndash dominated grassland was de-
phyto_44_1_2_103_119_Catorciindd 108phyto_44_1_2_103_119_Catorciindd 108 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 109
fined by aspect with a threshold value that can be inter-preted as the boundary between north-east-facing slopes (arid more stressful ndash groups T148-T1412) and south-west-facing ones (cool less stressful ndash groups T1413 and T1414) From a functional point of view the south-facing plots were characterized by a high number of indicator traits (perennial species graminoids hemi-rosulate forbs cushion plants and potential nurses) while no traits emerged as indicator of the plant com-munities growing on the north-facing slopes In addi-tion north-facing groups were characterised by a higher degree of dominance with the most widespread species often representing more than 50 of the total vegeta-tion cover
Altitude gradients and soil parameters defined further partitions for a more detailed differentiation of these two main clusters Among north-facing groups T148 was dominated by Nassella depauperata and had coarser soil as well as higher pH and organic matter values than T149 Groups T1410 T1411 and T1412 were charac-terised by higher altitudes (about 400 m above T148 and T149 ndash Table 3) T1410 was characterised by low distur-bance intensity and showed a more even distribution of species (particularly if compared with T1411) among which several palatable grasses are present (eg Deyeuxia brevifolia) Group T1412 dominated by the shrub spe-cies Chersodoma jodopappa showed a higher percentage of silt and organic matter in soil and steeper slopes with
Fig 3 Pie charts reporting the species composition of the groups defined in the 4-leaf (a) and in the 14-leaf (b) Multivariate regres-sion trees Species abbreviations are explained below Ast_die ndash As-tragalus dielsii Bac_tri ndash Baccharis tricuneata Bel_lon ndash Belloa longifolia Cal_bra ndash Calamagrostis breviaristata Can_aca ndash Calan-drina acaulis Chs_jod ndash Chersodoma jodopappa Cro_ful ndash Crocop-sis fulgens Des_myr ndash Descurainia myriophylla Dey_brf ndash De-yeuxia brevifolia Dey_rig ndash Deyeuxia rigida Dip_mey ndash Diplostephium meyenii Ech_pam ndash Echinopsis pampana Fes_ort ndash Festuca ortophylla Gna_lac ndash Gnaphalium lacteum Hyp_ech ndash Hypochaeris echegarayi Jar_ich ndash Jarava ichu Lpp_qua ndash Lepi-dopyllum quadrangulare Lup_sax ndash Lupinus saxatilis Muh_per ndash Muhlenbergia peruviana Nas_dep ndash Nassella depauperata Nas_nar ndash Nassella nardoides Ntt_tur ndash Nototriche turritella Oen_sp ndash Oenothera sp Par_luc ndash Parastrephia lucida Par_qua ndash Parastre-phia quadrangularis Pla_ser ndash Plantago sericea var lanuginosa Poa_gym ndash Poa gymnantha Poa_pea ndash Poa pearsonii Pyc_mol ndash Pycnophyllum molle Pyc_web ndash Pycnophyllum weberbaueri Sen_cha ndash Senecio chachaniensis Sen_nut ndash Senecio nutans Sti_han ndash Stipa hans-meyeri Tet_cri ndash Tetraglochin cristatum Tri_spi ndash Trise-tum spicatum Wer_pyg ndash Werneria pygmaea
phyto_44_1_2_103_119_Catorciindd 109phyto_44_1_2_103_119_Catorciindd 109 120314 0713120314 0713
eschweizerbart_xxx
110 A Catorci et al
groups yielded by the 4-leaf tree are clearly recognisable in the RDA three-dimensional scatterplot (as highlighted in Fig 4) The RDA performed on functional traits data with the environmental variables as constraints yielded similar results but explained a notably smaller fraction of the total variance (92 adjusted R2 = 056) The varia-bles showing the strongest correlation with the con-strained components coincided with those observed in the first RDA but for most environmental variables the observed correlation is weaker this is the case of altitude aspect percentage of outcropping rocks sand and silt percentage in the soil (Fig 5) Instead for disturbance in-tensity the observed correlation was slightly stronger The functional traits whose variance was best explained by the constrained components were nurse ability and various types of plant forms dwarf shrubs upright forbs non-graminoids hemirosulate forbs and graminoids (Fig 5) Non-graminoid species dwarf shrubs and up-right forbs were related to relatively higher soil pH and strong disturbance intensity nurse species were linked to higher altitudes South-facing slopes relatively lower sand percentages in the soil graminoid species and hemi-rosulate forbs were particularly tied to more fertile soils (higher phosphorus and nitrogen concentrations) and low disturbance intensity (Fig 5) RDA biplot scores are reported in the Appendix C
Discussion
Our results demonstrate that the dry Puna landscape is anything but homogeneous as observed for other semi-arid environments where local factors were detected as key drivers of species assemblages (eg Schlesinger et al 1990 Perelman et al 1997 Diacuteaz et al 1999) The variables showing the strongest correlations with the constrained RDA components were disturbance intensity slope as-pect altitude percentage of outcropping rocks and to a lesser degree soil features (organic matter content pH and potassium concentration) High disturbance inten-sity (which had the best correlation with the first con-strained component) emerged as the main factor shaping the species composition of the dry Puna pastures and was positively correlated with pH dwarf shrubs and non-graminoid plant forms while it was negatively cor-related with organic matternitrogen soil content nurse plant abundance hemirosulate forbs and graminoids Consistently with previous research carried out in semi-arid environments (eg Ares et al 1990 Carilla et al 2012) high grazing intensity promotes the spread of spe-cies with avoidance strategies (Table 4) such as thorny dwarf shrubs (Tetraglochin cristatum) annual species (eg Muhlenbergia peruviana) and small or prostrate plants (eg Nassella nardoides Astragalus dielsii Hy-pochaeris echegarayi) Thorny dwarf shrubs are fostered by herbivory given that the tough and spiny stems of this
Table 1 Values of Shannon and Simpson diversity indices for the releveacute groups of the 4-leaf and 14-leaf Multivariate regression tree
Groups Diversity
Shannon (Hlsquo) Simpson (D)
T41 1628 0653
T42 1831 0626
T43 2176 0765
T44 2822 0829
T141 1511 0582
T142 1611 0688
T143 1831 0626
T144 1714 0633
T145 2366 0822
T146 1913 0763
T147 2348 0824
T148 1316 0598
T149 1540 0574
T1410 2214 0792
T1411 2283 0675
T1412 2149 0714
T1413 2100 0741
T1414 2685 0863
high outcropping rock cover South-facing groups were in turn separated by potassium content in the soil group T1413 with higher concentration of potassium shows a higher coverage of graminoid species (eg Poa pearsonii)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grass Festuca or-thophylla the cushion plants Pycnophyllum molle and P weberbaueri the shrub species Parastrephia lucida and P quadrangularis) This plant community was located near the altitudinal limit of vegetation and characterised by very low disturbance
The Redundancy Analysis executed on species and en-vironmental data explained 638 of the total variance (adjusted R2 = 047) The results of the RDA are in line with the MRT analysis the variables showing the strong-est correlation with the constrained components were disturbance intensity altitude percentage of outcropping rocks slope aspect soil organic matter content pH and potassium concentration (Fig 4) RDA biplot scores are reported in Appendix B It is worth noting that this result coincides with those discriminating for the partitions of the 4-leaf and 14-leaf MRTs (in order of diminishing cor-relation with RDA axes) Another correspondence be-tween MRT and RDA analyses lies in the placement of site points in the RDA graph as a matter of fact the
phyto_44_1_2_103_119_Catorciindd 110phyto_44_1_2_103_119_Catorciindd 110 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 111
Table 2 Indicator species of the releveacute groups highlighted by Indicator Species Analysis for the 4ndashleaf and the 14ndashleaf Multivariate regres-sion tree (MRT) and their respective indicator values and mean cover percentages Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value)
Indicator species 4-leaf MRT 14-leaf MRTMax
GroupIndicator
ValueMean
cover Max
GroupIndicator
ValueMean
cover Nassella nardoides T41 663 199 ndash ndash ndash
Nothoscordum andicola T41 482 08 ndash ndash ndash
Lepidium meyenii T41 412 06 ndash ndash ndash
Astragalus peruvianus T41 315 07 ndash ndash ndash
Tetraglochin cristatum T41 826 544 T141 422 628
Muhlenbergia peruviana T41 675 44 T142 300 46
Cumulopuntia ignescens T42 405 07 ndash ndash ndash
Chenopodium petiolare T42 370 03 ndash ndash ndash
Urocarpidium echinatum T42 350 06 ndash ndash ndash
Nototriche turritella T42 345 24 ndash ndash ndash
Jarava ichu T42 979 598 T143 925 598
Plantago linearis T42 901 30 T143 493 30
Diplostephium meyenii T42 700 23 T143 634 23
Crocopsis fulgens T42 571 16 T143 571 16
Belloa piptolepis T42 592 11 T143 351 11
Senecio phylloleptus T42 490 10 T143 364 10
Hypochaeris echegarayi T43 380 08 ndash ndash ndash
Pycnophyllum weberbaueri T43 342 08 ndash ndash ndash
Junellia minima (Meyen) T43 300 08 ndash ndash ndash
Calamagrostis breviaristata T43 704 430 T144 398 581
Calceolaria inamoena ndash ndash ndash T145 315 08
Astragalus dielsii T43 460 11 T146 321 14
Nassella nardoides ndash ndash ndash T146 484 415
Calandrinia acaulis ndash ndash ndash T146 430 14
Oenothera sp ndash ndash ndash T146 420 14
Festuca orthophylla T44 522 389 ndash ndash ndash
Trisetum spicatum T44 406 08 ndash ndash ndash
Nassella depauperata T44 352 70 T148 860 600
Lupinus saxatilis ndash ndash ndash T148 657 80
Descurainia myriophylla ndash ndash ndash T148 437 133
Erigeron pazensis ndash ndash ndash T148 416 07
Deyeuxia brevifolia T44 464 53 T1410 414 193
Mniodes pulvinata ndash ndash ndash T1410 375 05
Lupinus chlorolepis ndash ndash ndash T1411 364 09
Calamagrostis vicunarum ndash ndash ndash T1412 500 07
Poa gymnantha ndash ndash ndash T1412 485 42
Dielsiochloa floribunda ndash ndash ndash T1412 450 07
Leucheria daucifolia ndash ndash ndash T1412 398 07
Echinopsis pampana ndash ndash ndash T1412 384 42
Perezia ciliosa ndash ndash ndash T1412 329 07
Poa pearsonii ndash ndash ndash T1413 670 237
Deyeuxia rigida T44 316 47 T1414 774 285
Poa aequigluma ndash ndash ndash T1414 453 06
Werneria spathulata ndash ndash ndash T1414 412 08
Luzula racemosa ndash ndash ndash T1414 409 09
Lucilia sp ndash ndash ndash T1414 401 12
Senecio scorzoneraefolius ndash ndash ndash T1414 376 05
Gnaphalium badium ndash ndash ndash T1414 338 10
Pycnophyllum molle ndash ndash ndash T1414 335 95
phyto_44_1_2_103_119_Catorciindd 111phyto_44_1_2_103_119_Catorciindd 111 120314 0713120314 0713
eschweizerbart_xxx
112 A Catorci et al
Table 3 Descriptive statistics of the environmental variables of the groups highlighted by the 4-leaf and 14-leaf Multivariate regression tree 1st and 3rd quartiles are indicated for the whole data set mean and standard deviation (SD) are reported for each group
Group Descriptive statistics
Constraining variables
Site characteristics Soil texture Soil composition
Altitude(m asl)
Aspect (deg)
Slope (deg)
Outcr rocks()
Sand()
Silt ()
Clay ()
Org matt()
N () P (ppm)
K (ppm)
CaCO3 ()
pH
1st quartile 407000 000 200 000 6280 1660 580 089 004 1136 11250 008 540 3rd quartile 447500 9000 2000 1500 7760 2780 910 174 009 2469 23250 075 623
T41 Mean 399700 2363 755 000 7867 1403 1051 071 003 1265 16381 049 652SD 1908 2777 894 000 295 180 1434 033 001 668 3318 035 028
T42 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T43 Mean 410500 2880 364 000 7265 2133 602 110 009 2173 17036 053 542SD 2933 4752 930 000 389 372 237 053 020 1234 6129 051 035
T44 Mean 440287 8739 2061 1787 6698 2444 849 171 008 2219 19384 045 572SD 22159 5242 1431 1192 976 866 304 083 004 1471 11137 039 045
T141 Mean 399833 3938 1208 000 7926 1374 1233 052 003 875 17452 049 666SD 2462 2561 901 000 333 203 1853 024 001 142 3181 032 028
T142 Mean 399500 000 075 000 7778 1445 778 099 004 1850 14775 049 631SD 535 000 104 000 214 141 213 022 001 725 3011 041 010
T143 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T144 Mean 411393 643 064 000 7373 1987 640 098 012 2552 18143 052 529SD 1712 2405 145 000 309 274 306 020 027 1421 5395 062 035
T145 Mean 412667 10500 2167 000 6777 2577 647 210 008 1611 18335 071 549SD 4619 1299 2082 000 224 145 093 017 001 516 13137 017 031
T146 Mean 409500 000 000 000 7385 2128 488 078 005 2370 18117 015 575SD 000 000 000 000 042 026 017 003 001 327 1019 019 018
T147 Mean 406750 7875 425 000 7135 2315 550 110 004 1073 11106 082 551SD 3663 5356 299 000 654 654 081 094 004 698 1577 019 031
T148 Mean 414800 7200 1900 1500 7368 2080 552 135 007 2044 9024 019 630SD 570 1006 224 000 618 567 064 037 002 1111 4791 021 015
T149 Mean 408833 9000 600 1833 6960 2093 947 096 005 1216 18803 054 566SD 5795 000 346 577 763 412 357 067 004 098 12521 057 071
T1410 Mean 458688 4922 2731 1813 7623 1591 787 136 006 2448 17087 042 547SD 22773 4043 1590 680 622 535 164 026 001 1461 9485 047 049
T1411 Mean 439614 5727 1114 2173 6501 2430 1038 147 007 1741 22177 052 583SD 16079 2840 551 1297 832 588 420 053 003 597 10086 035 049
T1412 Mean 445650 9000 4000 550 6025 3248 725 248 013 1653 25562 055 579SD 27365 000 2121 636 078 046 120 049 002 467 26427 040 013
T1413 Mean 421318 15341 3518 2409 5954 3227 819 285 014 2823 28413 025 564SD 11402 2814 1419 1338 1099 981 169 117 006 2824 11501 041 022
T1414 Mean 454300 14850 1600 590 6189 3104 707 183 009 2740 11102 065 571SD 6997 2846 615 1058 589 588 071 063 003 830 2759 027 026
life form discourage animals from biting it (Briske 1996) The observed spread of dwarf shrubs is in accordance with the findings of several studies carried out in semiarid environments (Perelman et al 1997 Swinton amp Quiroz 2003 Carilla et al 2012 Catorci et al 2012b 2013) and in pastoral ecosystems subjected to periodic fires (Liedloff
et al 2001) In semi-arid environments annual life span may be considered an avoidance strategy fostered by high grazing intensity (Diacuteaz et al 1999 McGregor 2002 Catorci et al 2012b) because it allows plants to dynami-cally face the grazing pressure by restarting the growth cycle every year (Diacuteaz et al 1999 2001 McGregor 2002)
phyto_44_1_2_103_119_Catorciindd 112phyto_44_1_2_103_119_Catorciindd 112 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

108 A Catorci et al
Fig 2 The 4-leaf (a) and 14-leaf (b) Multivariate regression trees for the species data set constrained by their explanatory variables The threshold values shown for each partition of the tree correspond to the mean of the two limit values of the considered variables at the break between the branches The relative abundances of the species are shown in histograms at the tips of the branches with the species in the same order as in the input file the species names and abun-dances are shown in Figure 3 Under each histogram the sum of squared errors for the group and the number of releveacutes in the leaf (n) are indicated Below each tree the residual error (Error) the cross-validation error (CV Error) and standard error (SE) are indi-cated Alt ndash Altitude Asp_deg ndash aspect Dist_L ndash Disturbance level Hum_p ndash organic matter K_ppm ndash Potassium in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt
tential nurse plants were the indicator traits for the T42 group (Table 4)
The C breviaristata ndash dominated plant community was characterized by high sand percentage and low or-ganic matter and potassium content in the soil (Table 3) The pH value was similar to that of groups T43 and T44 Cushion and perennial plants were the indicator traits for group T43 (Table 4)
The releveacutes grouped in the fourth leaf (T44) are char-acterised by a percentage of outcropping rocks greater than 1 (mean cover value 178 ) In this group the dominant species is the graminoid Festuca orthophylla other indicator species being Deyeuxia brevifolia Trise-tum spicatum Nassella depauperata and Deyeuxia rigida (all grass species) Group T44 includes the greatest num-ber of releveacutes (69) and presents the highest diversity (Hrsquo = 282 D = 083) in the 4-leaved tree Other environmen-
tal features of the Festuca orthophylla ndash dominated plant community were (Table 3) the highest slopes altitude and angles among the four groups highlighted by MRT low sand percentage and high values of organic matter nitro-gen and phosphorus in the soil The pH value was similar to that of groups T42 and T43 These pastures were characterised by low disturbance intensity Within the high richness and internal variability of these grasslands particular environmental conditions or treatments caused minor differences in species composition highlighting different plant communities as shown by the 14-leaved MRT (Fig 2)
Despite the high number of groups yielded the val-ues of community diversity indices computed from the pairwise comparison of the 14 leaves were still high (never below 45 ) (Appendix A) The first partition of the Festuca orthophylla ndash dominated grassland was de-
phyto_44_1_2_103_119_Catorciindd 108phyto_44_1_2_103_119_Catorciindd 108 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 109
fined by aspect with a threshold value that can be inter-preted as the boundary between north-east-facing slopes (arid more stressful ndash groups T148-T1412) and south-west-facing ones (cool less stressful ndash groups T1413 and T1414) From a functional point of view the south-facing plots were characterized by a high number of indicator traits (perennial species graminoids hemi-rosulate forbs cushion plants and potential nurses) while no traits emerged as indicator of the plant com-munities growing on the north-facing slopes In addi-tion north-facing groups were characterised by a higher degree of dominance with the most widespread species often representing more than 50 of the total vegeta-tion cover
Altitude gradients and soil parameters defined further partitions for a more detailed differentiation of these two main clusters Among north-facing groups T148 was dominated by Nassella depauperata and had coarser soil as well as higher pH and organic matter values than T149 Groups T1410 T1411 and T1412 were charac-terised by higher altitudes (about 400 m above T148 and T149 ndash Table 3) T1410 was characterised by low distur-bance intensity and showed a more even distribution of species (particularly if compared with T1411) among which several palatable grasses are present (eg Deyeuxia brevifolia) Group T1412 dominated by the shrub spe-cies Chersodoma jodopappa showed a higher percentage of silt and organic matter in soil and steeper slopes with
Fig 3 Pie charts reporting the species composition of the groups defined in the 4-leaf (a) and in the 14-leaf (b) Multivariate regres-sion trees Species abbreviations are explained below Ast_die ndash As-tragalus dielsii Bac_tri ndash Baccharis tricuneata Bel_lon ndash Belloa longifolia Cal_bra ndash Calamagrostis breviaristata Can_aca ndash Calan-drina acaulis Chs_jod ndash Chersodoma jodopappa Cro_ful ndash Crocop-sis fulgens Des_myr ndash Descurainia myriophylla Dey_brf ndash De-yeuxia brevifolia Dey_rig ndash Deyeuxia rigida Dip_mey ndash Diplostephium meyenii Ech_pam ndash Echinopsis pampana Fes_ort ndash Festuca ortophylla Gna_lac ndash Gnaphalium lacteum Hyp_ech ndash Hypochaeris echegarayi Jar_ich ndash Jarava ichu Lpp_qua ndash Lepi-dopyllum quadrangulare Lup_sax ndash Lupinus saxatilis Muh_per ndash Muhlenbergia peruviana Nas_dep ndash Nassella depauperata Nas_nar ndash Nassella nardoides Ntt_tur ndash Nototriche turritella Oen_sp ndash Oenothera sp Par_luc ndash Parastrephia lucida Par_qua ndash Parastre-phia quadrangularis Pla_ser ndash Plantago sericea var lanuginosa Poa_gym ndash Poa gymnantha Poa_pea ndash Poa pearsonii Pyc_mol ndash Pycnophyllum molle Pyc_web ndash Pycnophyllum weberbaueri Sen_cha ndash Senecio chachaniensis Sen_nut ndash Senecio nutans Sti_han ndash Stipa hans-meyeri Tet_cri ndash Tetraglochin cristatum Tri_spi ndash Trise-tum spicatum Wer_pyg ndash Werneria pygmaea
phyto_44_1_2_103_119_Catorciindd 109phyto_44_1_2_103_119_Catorciindd 109 120314 0713120314 0713
eschweizerbart_xxx
110 A Catorci et al
groups yielded by the 4-leaf tree are clearly recognisable in the RDA three-dimensional scatterplot (as highlighted in Fig 4) The RDA performed on functional traits data with the environmental variables as constraints yielded similar results but explained a notably smaller fraction of the total variance (92 adjusted R2 = 056) The varia-bles showing the strongest correlation with the con-strained components coincided with those observed in the first RDA but for most environmental variables the observed correlation is weaker this is the case of altitude aspect percentage of outcropping rocks sand and silt percentage in the soil (Fig 5) Instead for disturbance in-tensity the observed correlation was slightly stronger The functional traits whose variance was best explained by the constrained components were nurse ability and various types of plant forms dwarf shrubs upright forbs non-graminoids hemirosulate forbs and graminoids (Fig 5) Non-graminoid species dwarf shrubs and up-right forbs were related to relatively higher soil pH and strong disturbance intensity nurse species were linked to higher altitudes South-facing slopes relatively lower sand percentages in the soil graminoid species and hemi-rosulate forbs were particularly tied to more fertile soils (higher phosphorus and nitrogen concentrations) and low disturbance intensity (Fig 5) RDA biplot scores are reported in the Appendix C
Discussion
Our results demonstrate that the dry Puna landscape is anything but homogeneous as observed for other semi-arid environments where local factors were detected as key drivers of species assemblages (eg Schlesinger et al 1990 Perelman et al 1997 Diacuteaz et al 1999) The variables showing the strongest correlations with the constrained RDA components were disturbance intensity slope as-pect altitude percentage of outcropping rocks and to a lesser degree soil features (organic matter content pH and potassium concentration) High disturbance inten-sity (which had the best correlation with the first con-strained component) emerged as the main factor shaping the species composition of the dry Puna pastures and was positively correlated with pH dwarf shrubs and non-graminoid plant forms while it was negatively cor-related with organic matternitrogen soil content nurse plant abundance hemirosulate forbs and graminoids Consistently with previous research carried out in semi-arid environments (eg Ares et al 1990 Carilla et al 2012) high grazing intensity promotes the spread of spe-cies with avoidance strategies (Table 4) such as thorny dwarf shrubs (Tetraglochin cristatum) annual species (eg Muhlenbergia peruviana) and small or prostrate plants (eg Nassella nardoides Astragalus dielsii Hy-pochaeris echegarayi) Thorny dwarf shrubs are fostered by herbivory given that the tough and spiny stems of this
Table 1 Values of Shannon and Simpson diversity indices for the releveacute groups of the 4-leaf and 14-leaf Multivariate regression tree
Groups Diversity
Shannon (Hlsquo) Simpson (D)
T41 1628 0653
T42 1831 0626
T43 2176 0765
T44 2822 0829
T141 1511 0582
T142 1611 0688
T143 1831 0626
T144 1714 0633
T145 2366 0822
T146 1913 0763
T147 2348 0824
T148 1316 0598
T149 1540 0574
T1410 2214 0792
T1411 2283 0675
T1412 2149 0714
T1413 2100 0741
T1414 2685 0863
high outcropping rock cover South-facing groups were in turn separated by potassium content in the soil group T1413 with higher concentration of potassium shows a higher coverage of graminoid species (eg Poa pearsonii)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grass Festuca or-thophylla the cushion plants Pycnophyllum molle and P weberbaueri the shrub species Parastrephia lucida and P quadrangularis) This plant community was located near the altitudinal limit of vegetation and characterised by very low disturbance
The Redundancy Analysis executed on species and en-vironmental data explained 638 of the total variance (adjusted R2 = 047) The results of the RDA are in line with the MRT analysis the variables showing the strong-est correlation with the constrained components were disturbance intensity altitude percentage of outcropping rocks slope aspect soil organic matter content pH and potassium concentration (Fig 4) RDA biplot scores are reported in Appendix B It is worth noting that this result coincides with those discriminating for the partitions of the 4-leaf and 14-leaf MRTs (in order of diminishing cor-relation with RDA axes) Another correspondence be-tween MRT and RDA analyses lies in the placement of site points in the RDA graph as a matter of fact the
phyto_44_1_2_103_119_Catorciindd 110phyto_44_1_2_103_119_Catorciindd 110 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 111
Table 2 Indicator species of the releveacute groups highlighted by Indicator Species Analysis for the 4ndashleaf and the 14ndashleaf Multivariate regres-sion tree (MRT) and their respective indicator values and mean cover percentages Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value)
Indicator species 4-leaf MRT 14-leaf MRTMax
GroupIndicator
ValueMean
cover Max
GroupIndicator
ValueMean
cover Nassella nardoides T41 663 199 ndash ndash ndash
Nothoscordum andicola T41 482 08 ndash ndash ndash
Lepidium meyenii T41 412 06 ndash ndash ndash
Astragalus peruvianus T41 315 07 ndash ndash ndash
Tetraglochin cristatum T41 826 544 T141 422 628
Muhlenbergia peruviana T41 675 44 T142 300 46
Cumulopuntia ignescens T42 405 07 ndash ndash ndash
Chenopodium petiolare T42 370 03 ndash ndash ndash
Urocarpidium echinatum T42 350 06 ndash ndash ndash
Nototriche turritella T42 345 24 ndash ndash ndash
Jarava ichu T42 979 598 T143 925 598
Plantago linearis T42 901 30 T143 493 30
Diplostephium meyenii T42 700 23 T143 634 23
Crocopsis fulgens T42 571 16 T143 571 16
Belloa piptolepis T42 592 11 T143 351 11
Senecio phylloleptus T42 490 10 T143 364 10
Hypochaeris echegarayi T43 380 08 ndash ndash ndash
Pycnophyllum weberbaueri T43 342 08 ndash ndash ndash
Junellia minima (Meyen) T43 300 08 ndash ndash ndash
Calamagrostis breviaristata T43 704 430 T144 398 581
Calceolaria inamoena ndash ndash ndash T145 315 08
Astragalus dielsii T43 460 11 T146 321 14
Nassella nardoides ndash ndash ndash T146 484 415
Calandrinia acaulis ndash ndash ndash T146 430 14
Oenothera sp ndash ndash ndash T146 420 14
Festuca orthophylla T44 522 389 ndash ndash ndash
Trisetum spicatum T44 406 08 ndash ndash ndash
Nassella depauperata T44 352 70 T148 860 600
Lupinus saxatilis ndash ndash ndash T148 657 80
Descurainia myriophylla ndash ndash ndash T148 437 133
Erigeron pazensis ndash ndash ndash T148 416 07
Deyeuxia brevifolia T44 464 53 T1410 414 193
Mniodes pulvinata ndash ndash ndash T1410 375 05
Lupinus chlorolepis ndash ndash ndash T1411 364 09
Calamagrostis vicunarum ndash ndash ndash T1412 500 07
Poa gymnantha ndash ndash ndash T1412 485 42
Dielsiochloa floribunda ndash ndash ndash T1412 450 07
Leucheria daucifolia ndash ndash ndash T1412 398 07
Echinopsis pampana ndash ndash ndash T1412 384 42
Perezia ciliosa ndash ndash ndash T1412 329 07
Poa pearsonii ndash ndash ndash T1413 670 237
Deyeuxia rigida T44 316 47 T1414 774 285
Poa aequigluma ndash ndash ndash T1414 453 06
Werneria spathulata ndash ndash ndash T1414 412 08
Luzula racemosa ndash ndash ndash T1414 409 09
Lucilia sp ndash ndash ndash T1414 401 12
Senecio scorzoneraefolius ndash ndash ndash T1414 376 05
Gnaphalium badium ndash ndash ndash T1414 338 10
Pycnophyllum molle ndash ndash ndash T1414 335 95
phyto_44_1_2_103_119_Catorciindd 111phyto_44_1_2_103_119_Catorciindd 111 120314 0713120314 0713
eschweizerbart_xxx
112 A Catorci et al
Table 3 Descriptive statistics of the environmental variables of the groups highlighted by the 4-leaf and 14-leaf Multivariate regression tree 1st and 3rd quartiles are indicated for the whole data set mean and standard deviation (SD) are reported for each group
Group Descriptive statistics
Constraining variables
Site characteristics Soil texture Soil composition
Altitude(m asl)
Aspect (deg)
Slope (deg)
Outcr rocks()
Sand()
Silt ()
Clay ()
Org matt()
N () P (ppm)
K (ppm)
CaCO3 ()
pH
1st quartile 407000 000 200 000 6280 1660 580 089 004 1136 11250 008 540 3rd quartile 447500 9000 2000 1500 7760 2780 910 174 009 2469 23250 075 623
T41 Mean 399700 2363 755 000 7867 1403 1051 071 003 1265 16381 049 652SD 1908 2777 894 000 295 180 1434 033 001 668 3318 035 028
T42 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T43 Mean 410500 2880 364 000 7265 2133 602 110 009 2173 17036 053 542SD 2933 4752 930 000 389 372 237 053 020 1234 6129 051 035
T44 Mean 440287 8739 2061 1787 6698 2444 849 171 008 2219 19384 045 572SD 22159 5242 1431 1192 976 866 304 083 004 1471 11137 039 045
T141 Mean 399833 3938 1208 000 7926 1374 1233 052 003 875 17452 049 666SD 2462 2561 901 000 333 203 1853 024 001 142 3181 032 028
T142 Mean 399500 000 075 000 7778 1445 778 099 004 1850 14775 049 631SD 535 000 104 000 214 141 213 022 001 725 3011 041 010
T143 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T144 Mean 411393 643 064 000 7373 1987 640 098 012 2552 18143 052 529SD 1712 2405 145 000 309 274 306 020 027 1421 5395 062 035
T145 Mean 412667 10500 2167 000 6777 2577 647 210 008 1611 18335 071 549SD 4619 1299 2082 000 224 145 093 017 001 516 13137 017 031
T146 Mean 409500 000 000 000 7385 2128 488 078 005 2370 18117 015 575SD 000 000 000 000 042 026 017 003 001 327 1019 019 018
T147 Mean 406750 7875 425 000 7135 2315 550 110 004 1073 11106 082 551SD 3663 5356 299 000 654 654 081 094 004 698 1577 019 031
T148 Mean 414800 7200 1900 1500 7368 2080 552 135 007 2044 9024 019 630SD 570 1006 224 000 618 567 064 037 002 1111 4791 021 015
T149 Mean 408833 9000 600 1833 6960 2093 947 096 005 1216 18803 054 566SD 5795 000 346 577 763 412 357 067 004 098 12521 057 071
T1410 Mean 458688 4922 2731 1813 7623 1591 787 136 006 2448 17087 042 547SD 22773 4043 1590 680 622 535 164 026 001 1461 9485 047 049
T1411 Mean 439614 5727 1114 2173 6501 2430 1038 147 007 1741 22177 052 583SD 16079 2840 551 1297 832 588 420 053 003 597 10086 035 049
T1412 Mean 445650 9000 4000 550 6025 3248 725 248 013 1653 25562 055 579SD 27365 000 2121 636 078 046 120 049 002 467 26427 040 013
T1413 Mean 421318 15341 3518 2409 5954 3227 819 285 014 2823 28413 025 564SD 11402 2814 1419 1338 1099 981 169 117 006 2824 11501 041 022
T1414 Mean 454300 14850 1600 590 6189 3104 707 183 009 2740 11102 065 571SD 6997 2846 615 1058 589 588 071 063 003 830 2759 027 026
life form discourage animals from biting it (Briske 1996) The observed spread of dwarf shrubs is in accordance with the findings of several studies carried out in semiarid environments (Perelman et al 1997 Swinton amp Quiroz 2003 Carilla et al 2012 Catorci et al 2012b 2013) and in pastoral ecosystems subjected to periodic fires (Liedloff
et al 2001) In semi-arid environments annual life span may be considered an avoidance strategy fostered by high grazing intensity (Diacuteaz et al 1999 McGregor 2002 Catorci et al 2012b) because it allows plants to dynami-cally face the grazing pressure by restarting the growth cycle every year (Diacuteaz et al 1999 2001 McGregor 2002)
phyto_44_1_2_103_119_Catorciindd 112phyto_44_1_2_103_119_Catorciindd 112 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

Floristic composition of dry Puna in the southern Peruvian Andes 109
fined by aspect with a threshold value that can be inter-preted as the boundary between north-east-facing slopes (arid more stressful ndash groups T148-T1412) and south-west-facing ones (cool less stressful ndash groups T1413 and T1414) From a functional point of view the south-facing plots were characterized by a high number of indicator traits (perennial species graminoids hemi-rosulate forbs cushion plants and potential nurses) while no traits emerged as indicator of the plant com-munities growing on the north-facing slopes In addi-tion north-facing groups were characterised by a higher degree of dominance with the most widespread species often representing more than 50 of the total vegeta-tion cover
Altitude gradients and soil parameters defined further partitions for a more detailed differentiation of these two main clusters Among north-facing groups T148 was dominated by Nassella depauperata and had coarser soil as well as higher pH and organic matter values than T149 Groups T1410 T1411 and T1412 were charac-terised by higher altitudes (about 400 m above T148 and T149 ndash Table 3) T1410 was characterised by low distur-bance intensity and showed a more even distribution of species (particularly if compared with T1411) among which several palatable grasses are present (eg Deyeuxia brevifolia) Group T1412 dominated by the shrub spe-cies Chersodoma jodopappa showed a higher percentage of silt and organic matter in soil and steeper slopes with
Fig 3 Pie charts reporting the species composition of the groups defined in the 4-leaf (a) and in the 14-leaf (b) Multivariate regres-sion trees Species abbreviations are explained below Ast_die ndash As-tragalus dielsii Bac_tri ndash Baccharis tricuneata Bel_lon ndash Belloa longifolia Cal_bra ndash Calamagrostis breviaristata Can_aca ndash Calan-drina acaulis Chs_jod ndash Chersodoma jodopappa Cro_ful ndash Crocop-sis fulgens Des_myr ndash Descurainia myriophylla Dey_brf ndash De-yeuxia brevifolia Dey_rig ndash Deyeuxia rigida Dip_mey ndash Diplostephium meyenii Ech_pam ndash Echinopsis pampana Fes_ort ndash Festuca ortophylla Gna_lac ndash Gnaphalium lacteum Hyp_ech ndash Hypochaeris echegarayi Jar_ich ndash Jarava ichu Lpp_qua ndash Lepi-dopyllum quadrangulare Lup_sax ndash Lupinus saxatilis Muh_per ndash Muhlenbergia peruviana Nas_dep ndash Nassella depauperata Nas_nar ndash Nassella nardoides Ntt_tur ndash Nototriche turritella Oen_sp ndash Oenothera sp Par_luc ndash Parastrephia lucida Par_qua ndash Parastre-phia quadrangularis Pla_ser ndash Plantago sericea var lanuginosa Poa_gym ndash Poa gymnantha Poa_pea ndash Poa pearsonii Pyc_mol ndash Pycnophyllum molle Pyc_web ndash Pycnophyllum weberbaueri Sen_cha ndash Senecio chachaniensis Sen_nut ndash Senecio nutans Sti_han ndash Stipa hans-meyeri Tet_cri ndash Tetraglochin cristatum Tri_spi ndash Trise-tum spicatum Wer_pyg ndash Werneria pygmaea
phyto_44_1_2_103_119_Catorciindd 109phyto_44_1_2_103_119_Catorciindd 109 120314 0713120314 0713
eschweizerbart_xxx
110 A Catorci et al
groups yielded by the 4-leaf tree are clearly recognisable in the RDA three-dimensional scatterplot (as highlighted in Fig 4) The RDA performed on functional traits data with the environmental variables as constraints yielded similar results but explained a notably smaller fraction of the total variance (92 adjusted R2 = 056) The varia-bles showing the strongest correlation with the con-strained components coincided with those observed in the first RDA but for most environmental variables the observed correlation is weaker this is the case of altitude aspect percentage of outcropping rocks sand and silt percentage in the soil (Fig 5) Instead for disturbance in-tensity the observed correlation was slightly stronger The functional traits whose variance was best explained by the constrained components were nurse ability and various types of plant forms dwarf shrubs upright forbs non-graminoids hemirosulate forbs and graminoids (Fig 5) Non-graminoid species dwarf shrubs and up-right forbs were related to relatively higher soil pH and strong disturbance intensity nurse species were linked to higher altitudes South-facing slopes relatively lower sand percentages in the soil graminoid species and hemi-rosulate forbs were particularly tied to more fertile soils (higher phosphorus and nitrogen concentrations) and low disturbance intensity (Fig 5) RDA biplot scores are reported in the Appendix C
Discussion
Our results demonstrate that the dry Puna landscape is anything but homogeneous as observed for other semi-arid environments where local factors were detected as key drivers of species assemblages (eg Schlesinger et al 1990 Perelman et al 1997 Diacuteaz et al 1999) The variables showing the strongest correlations with the constrained RDA components were disturbance intensity slope as-pect altitude percentage of outcropping rocks and to a lesser degree soil features (organic matter content pH and potassium concentration) High disturbance inten-sity (which had the best correlation with the first con-strained component) emerged as the main factor shaping the species composition of the dry Puna pastures and was positively correlated with pH dwarf shrubs and non-graminoid plant forms while it was negatively cor-related with organic matternitrogen soil content nurse plant abundance hemirosulate forbs and graminoids Consistently with previous research carried out in semi-arid environments (eg Ares et al 1990 Carilla et al 2012) high grazing intensity promotes the spread of spe-cies with avoidance strategies (Table 4) such as thorny dwarf shrubs (Tetraglochin cristatum) annual species (eg Muhlenbergia peruviana) and small or prostrate plants (eg Nassella nardoides Astragalus dielsii Hy-pochaeris echegarayi) Thorny dwarf shrubs are fostered by herbivory given that the tough and spiny stems of this
Table 1 Values of Shannon and Simpson diversity indices for the releveacute groups of the 4-leaf and 14-leaf Multivariate regression tree
Groups Diversity
Shannon (Hlsquo) Simpson (D)
T41 1628 0653
T42 1831 0626
T43 2176 0765
T44 2822 0829
T141 1511 0582
T142 1611 0688
T143 1831 0626
T144 1714 0633
T145 2366 0822
T146 1913 0763
T147 2348 0824
T148 1316 0598
T149 1540 0574
T1410 2214 0792
T1411 2283 0675
T1412 2149 0714
T1413 2100 0741
T1414 2685 0863
high outcropping rock cover South-facing groups were in turn separated by potassium content in the soil group T1413 with higher concentration of potassium shows a higher coverage of graminoid species (eg Poa pearsonii)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grass Festuca or-thophylla the cushion plants Pycnophyllum molle and P weberbaueri the shrub species Parastrephia lucida and P quadrangularis) This plant community was located near the altitudinal limit of vegetation and characterised by very low disturbance
The Redundancy Analysis executed on species and en-vironmental data explained 638 of the total variance (adjusted R2 = 047) The results of the RDA are in line with the MRT analysis the variables showing the strong-est correlation with the constrained components were disturbance intensity altitude percentage of outcropping rocks slope aspect soil organic matter content pH and potassium concentration (Fig 4) RDA biplot scores are reported in Appendix B It is worth noting that this result coincides with those discriminating for the partitions of the 4-leaf and 14-leaf MRTs (in order of diminishing cor-relation with RDA axes) Another correspondence be-tween MRT and RDA analyses lies in the placement of site points in the RDA graph as a matter of fact the
phyto_44_1_2_103_119_Catorciindd 110phyto_44_1_2_103_119_Catorciindd 110 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 111
Table 2 Indicator species of the releveacute groups highlighted by Indicator Species Analysis for the 4ndashleaf and the 14ndashleaf Multivariate regres-sion tree (MRT) and their respective indicator values and mean cover percentages Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value)
Indicator species 4-leaf MRT 14-leaf MRTMax
GroupIndicator
ValueMean
cover Max
GroupIndicator
ValueMean
cover Nassella nardoides T41 663 199 ndash ndash ndash
Nothoscordum andicola T41 482 08 ndash ndash ndash
Lepidium meyenii T41 412 06 ndash ndash ndash
Astragalus peruvianus T41 315 07 ndash ndash ndash
Tetraglochin cristatum T41 826 544 T141 422 628
Muhlenbergia peruviana T41 675 44 T142 300 46
Cumulopuntia ignescens T42 405 07 ndash ndash ndash
Chenopodium petiolare T42 370 03 ndash ndash ndash
Urocarpidium echinatum T42 350 06 ndash ndash ndash
Nototriche turritella T42 345 24 ndash ndash ndash
Jarava ichu T42 979 598 T143 925 598
Plantago linearis T42 901 30 T143 493 30
Diplostephium meyenii T42 700 23 T143 634 23
Crocopsis fulgens T42 571 16 T143 571 16
Belloa piptolepis T42 592 11 T143 351 11
Senecio phylloleptus T42 490 10 T143 364 10
Hypochaeris echegarayi T43 380 08 ndash ndash ndash
Pycnophyllum weberbaueri T43 342 08 ndash ndash ndash
Junellia minima (Meyen) T43 300 08 ndash ndash ndash
Calamagrostis breviaristata T43 704 430 T144 398 581
Calceolaria inamoena ndash ndash ndash T145 315 08
Astragalus dielsii T43 460 11 T146 321 14
Nassella nardoides ndash ndash ndash T146 484 415
Calandrinia acaulis ndash ndash ndash T146 430 14
Oenothera sp ndash ndash ndash T146 420 14
Festuca orthophylla T44 522 389 ndash ndash ndash
Trisetum spicatum T44 406 08 ndash ndash ndash
Nassella depauperata T44 352 70 T148 860 600
Lupinus saxatilis ndash ndash ndash T148 657 80
Descurainia myriophylla ndash ndash ndash T148 437 133
Erigeron pazensis ndash ndash ndash T148 416 07
Deyeuxia brevifolia T44 464 53 T1410 414 193
Mniodes pulvinata ndash ndash ndash T1410 375 05
Lupinus chlorolepis ndash ndash ndash T1411 364 09
Calamagrostis vicunarum ndash ndash ndash T1412 500 07
Poa gymnantha ndash ndash ndash T1412 485 42
Dielsiochloa floribunda ndash ndash ndash T1412 450 07
Leucheria daucifolia ndash ndash ndash T1412 398 07
Echinopsis pampana ndash ndash ndash T1412 384 42
Perezia ciliosa ndash ndash ndash T1412 329 07
Poa pearsonii ndash ndash ndash T1413 670 237
Deyeuxia rigida T44 316 47 T1414 774 285
Poa aequigluma ndash ndash ndash T1414 453 06
Werneria spathulata ndash ndash ndash T1414 412 08
Luzula racemosa ndash ndash ndash T1414 409 09
Lucilia sp ndash ndash ndash T1414 401 12
Senecio scorzoneraefolius ndash ndash ndash T1414 376 05
Gnaphalium badium ndash ndash ndash T1414 338 10
Pycnophyllum molle ndash ndash ndash T1414 335 95
phyto_44_1_2_103_119_Catorciindd 111phyto_44_1_2_103_119_Catorciindd 111 120314 0713120314 0713
eschweizerbart_xxx
112 A Catorci et al
Table 3 Descriptive statistics of the environmental variables of the groups highlighted by the 4-leaf and 14-leaf Multivariate regression tree 1st and 3rd quartiles are indicated for the whole data set mean and standard deviation (SD) are reported for each group
Group Descriptive statistics
Constraining variables
Site characteristics Soil texture Soil composition
Altitude(m asl)
Aspect (deg)
Slope (deg)
Outcr rocks()
Sand()
Silt ()
Clay ()
Org matt()
N () P (ppm)
K (ppm)
CaCO3 ()
pH
1st quartile 407000 000 200 000 6280 1660 580 089 004 1136 11250 008 540 3rd quartile 447500 9000 2000 1500 7760 2780 910 174 009 2469 23250 075 623
T41 Mean 399700 2363 755 000 7867 1403 1051 071 003 1265 16381 049 652SD 1908 2777 894 000 295 180 1434 033 001 668 3318 035 028
T42 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T43 Mean 410500 2880 364 000 7265 2133 602 110 009 2173 17036 053 542SD 2933 4752 930 000 389 372 237 053 020 1234 6129 051 035
T44 Mean 440287 8739 2061 1787 6698 2444 849 171 008 2219 19384 045 572SD 22159 5242 1431 1192 976 866 304 083 004 1471 11137 039 045
T141 Mean 399833 3938 1208 000 7926 1374 1233 052 003 875 17452 049 666SD 2462 2561 901 000 333 203 1853 024 001 142 3181 032 028
T142 Mean 399500 000 075 000 7778 1445 778 099 004 1850 14775 049 631SD 535 000 104 000 214 141 213 022 001 725 3011 041 010
T143 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T144 Mean 411393 643 064 000 7373 1987 640 098 012 2552 18143 052 529SD 1712 2405 145 000 309 274 306 020 027 1421 5395 062 035
T145 Mean 412667 10500 2167 000 6777 2577 647 210 008 1611 18335 071 549SD 4619 1299 2082 000 224 145 093 017 001 516 13137 017 031
T146 Mean 409500 000 000 000 7385 2128 488 078 005 2370 18117 015 575SD 000 000 000 000 042 026 017 003 001 327 1019 019 018
T147 Mean 406750 7875 425 000 7135 2315 550 110 004 1073 11106 082 551SD 3663 5356 299 000 654 654 081 094 004 698 1577 019 031
T148 Mean 414800 7200 1900 1500 7368 2080 552 135 007 2044 9024 019 630SD 570 1006 224 000 618 567 064 037 002 1111 4791 021 015
T149 Mean 408833 9000 600 1833 6960 2093 947 096 005 1216 18803 054 566SD 5795 000 346 577 763 412 357 067 004 098 12521 057 071
T1410 Mean 458688 4922 2731 1813 7623 1591 787 136 006 2448 17087 042 547SD 22773 4043 1590 680 622 535 164 026 001 1461 9485 047 049
T1411 Mean 439614 5727 1114 2173 6501 2430 1038 147 007 1741 22177 052 583SD 16079 2840 551 1297 832 588 420 053 003 597 10086 035 049
T1412 Mean 445650 9000 4000 550 6025 3248 725 248 013 1653 25562 055 579SD 27365 000 2121 636 078 046 120 049 002 467 26427 040 013
T1413 Mean 421318 15341 3518 2409 5954 3227 819 285 014 2823 28413 025 564SD 11402 2814 1419 1338 1099 981 169 117 006 2824 11501 041 022
T1414 Mean 454300 14850 1600 590 6189 3104 707 183 009 2740 11102 065 571SD 6997 2846 615 1058 589 588 071 063 003 830 2759 027 026
life form discourage animals from biting it (Briske 1996) The observed spread of dwarf shrubs is in accordance with the findings of several studies carried out in semiarid environments (Perelman et al 1997 Swinton amp Quiroz 2003 Carilla et al 2012 Catorci et al 2012b 2013) and in pastoral ecosystems subjected to periodic fires (Liedloff
et al 2001) In semi-arid environments annual life span may be considered an avoidance strategy fostered by high grazing intensity (Diacuteaz et al 1999 McGregor 2002 Catorci et al 2012b) because it allows plants to dynami-cally face the grazing pressure by restarting the growth cycle every year (Diacuteaz et al 1999 2001 McGregor 2002)
phyto_44_1_2_103_119_Catorciindd 112phyto_44_1_2_103_119_Catorciindd 112 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

110 A Catorci et al
groups yielded by the 4-leaf tree are clearly recognisable in the RDA three-dimensional scatterplot (as highlighted in Fig 4) The RDA performed on functional traits data with the environmental variables as constraints yielded similar results but explained a notably smaller fraction of the total variance (92 adjusted R2 = 056) The varia-bles showing the strongest correlation with the con-strained components coincided with those observed in the first RDA but for most environmental variables the observed correlation is weaker this is the case of altitude aspect percentage of outcropping rocks sand and silt percentage in the soil (Fig 5) Instead for disturbance in-tensity the observed correlation was slightly stronger The functional traits whose variance was best explained by the constrained components were nurse ability and various types of plant forms dwarf shrubs upright forbs non-graminoids hemirosulate forbs and graminoids (Fig 5) Non-graminoid species dwarf shrubs and up-right forbs were related to relatively higher soil pH and strong disturbance intensity nurse species were linked to higher altitudes South-facing slopes relatively lower sand percentages in the soil graminoid species and hemi-rosulate forbs were particularly tied to more fertile soils (higher phosphorus and nitrogen concentrations) and low disturbance intensity (Fig 5) RDA biplot scores are reported in the Appendix C
Discussion
Our results demonstrate that the dry Puna landscape is anything but homogeneous as observed for other semi-arid environments where local factors were detected as key drivers of species assemblages (eg Schlesinger et al 1990 Perelman et al 1997 Diacuteaz et al 1999) The variables showing the strongest correlations with the constrained RDA components were disturbance intensity slope as-pect altitude percentage of outcropping rocks and to a lesser degree soil features (organic matter content pH and potassium concentration) High disturbance inten-sity (which had the best correlation with the first con-strained component) emerged as the main factor shaping the species composition of the dry Puna pastures and was positively correlated with pH dwarf shrubs and non-graminoid plant forms while it was negatively cor-related with organic matternitrogen soil content nurse plant abundance hemirosulate forbs and graminoids Consistently with previous research carried out in semi-arid environments (eg Ares et al 1990 Carilla et al 2012) high grazing intensity promotes the spread of spe-cies with avoidance strategies (Table 4) such as thorny dwarf shrubs (Tetraglochin cristatum) annual species (eg Muhlenbergia peruviana) and small or prostrate plants (eg Nassella nardoides Astragalus dielsii Hy-pochaeris echegarayi) Thorny dwarf shrubs are fostered by herbivory given that the tough and spiny stems of this
Table 1 Values of Shannon and Simpson diversity indices for the releveacute groups of the 4-leaf and 14-leaf Multivariate regression tree
Groups Diversity
Shannon (Hlsquo) Simpson (D)
T41 1628 0653
T42 1831 0626
T43 2176 0765
T44 2822 0829
T141 1511 0582
T142 1611 0688
T143 1831 0626
T144 1714 0633
T145 2366 0822
T146 1913 0763
T147 2348 0824
T148 1316 0598
T149 1540 0574
T1410 2214 0792
T1411 2283 0675
T1412 2149 0714
T1413 2100 0741
T1414 2685 0863
high outcropping rock cover South-facing groups were in turn separated by potassium content in the soil group T1413 with higher concentration of potassium shows a higher coverage of graminoid species (eg Poa pearsonii)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grass Festuca or-thophylla the cushion plants Pycnophyllum molle and P weberbaueri the shrub species Parastrephia lucida and P quadrangularis) This plant community was located near the altitudinal limit of vegetation and characterised by very low disturbance
The Redundancy Analysis executed on species and en-vironmental data explained 638 of the total variance (adjusted R2 = 047) The results of the RDA are in line with the MRT analysis the variables showing the strong-est correlation with the constrained components were disturbance intensity altitude percentage of outcropping rocks slope aspect soil organic matter content pH and potassium concentration (Fig 4) RDA biplot scores are reported in Appendix B It is worth noting that this result coincides with those discriminating for the partitions of the 4-leaf and 14-leaf MRTs (in order of diminishing cor-relation with RDA axes) Another correspondence be-tween MRT and RDA analyses lies in the placement of site points in the RDA graph as a matter of fact the
phyto_44_1_2_103_119_Catorciindd 110phyto_44_1_2_103_119_Catorciindd 110 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 111
Table 2 Indicator species of the releveacute groups highlighted by Indicator Species Analysis for the 4ndashleaf and the 14ndashleaf Multivariate regres-sion tree (MRT) and their respective indicator values and mean cover percentages Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value)
Indicator species 4-leaf MRT 14-leaf MRTMax
GroupIndicator
ValueMean
cover Max
GroupIndicator
ValueMean
cover Nassella nardoides T41 663 199 ndash ndash ndash
Nothoscordum andicola T41 482 08 ndash ndash ndash
Lepidium meyenii T41 412 06 ndash ndash ndash
Astragalus peruvianus T41 315 07 ndash ndash ndash
Tetraglochin cristatum T41 826 544 T141 422 628
Muhlenbergia peruviana T41 675 44 T142 300 46
Cumulopuntia ignescens T42 405 07 ndash ndash ndash
Chenopodium petiolare T42 370 03 ndash ndash ndash
Urocarpidium echinatum T42 350 06 ndash ndash ndash
Nototriche turritella T42 345 24 ndash ndash ndash
Jarava ichu T42 979 598 T143 925 598
Plantago linearis T42 901 30 T143 493 30
Diplostephium meyenii T42 700 23 T143 634 23
Crocopsis fulgens T42 571 16 T143 571 16
Belloa piptolepis T42 592 11 T143 351 11
Senecio phylloleptus T42 490 10 T143 364 10
Hypochaeris echegarayi T43 380 08 ndash ndash ndash
Pycnophyllum weberbaueri T43 342 08 ndash ndash ndash
Junellia minima (Meyen) T43 300 08 ndash ndash ndash
Calamagrostis breviaristata T43 704 430 T144 398 581
Calceolaria inamoena ndash ndash ndash T145 315 08
Astragalus dielsii T43 460 11 T146 321 14
Nassella nardoides ndash ndash ndash T146 484 415
Calandrinia acaulis ndash ndash ndash T146 430 14
Oenothera sp ndash ndash ndash T146 420 14
Festuca orthophylla T44 522 389 ndash ndash ndash
Trisetum spicatum T44 406 08 ndash ndash ndash
Nassella depauperata T44 352 70 T148 860 600
Lupinus saxatilis ndash ndash ndash T148 657 80
Descurainia myriophylla ndash ndash ndash T148 437 133
Erigeron pazensis ndash ndash ndash T148 416 07
Deyeuxia brevifolia T44 464 53 T1410 414 193
Mniodes pulvinata ndash ndash ndash T1410 375 05
Lupinus chlorolepis ndash ndash ndash T1411 364 09
Calamagrostis vicunarum ndash ndash ndash T1412 500 07
Poa gymnantha ndash ndash ndash T1412 485 42
Dielsiochloa floribunda ndash ndash ndash T1412 450 07
Leucheria daucifolia ndash ndash ndash T1412 398 07
Echinopsis pampana ndash ndash ndash T1412 384 42
Perezia ciliosa ndash ndash ndash T1412 329 07
Poa pearsonii ndash ndash ndash T1413 670 237
Deyeuxia rigida T44 316 47 T1414 774 285
Poa aequigluma ndash ndash ndash T1414 453 06
Werneria spathulata ndash ndash ndash T1414 412 08
Luzula racemosa ndash ndash ndash T1414 409 09
Lucilia sp ndash ndash ndash T1414 401 12
Senecio scorzoneraefolius ndash ndash ndash T1414 376 05
Gnaphalium badium ndash ndash ndash T1414 338 10
Pycnophyllum molle ndash ndash ndash T1414 335 95
phyto_44_1_2_103_119_Catorciindd 111phyto_44_1_2_103_119_Catorciindd 111 120314 0713120314 0713
eschweizerbart_xxx
112 A Catorci et al
Table 3 Descriptive statistics of the environmental variables of the groups highlighted by the 4-leaf and 14-leaf Multivariate regression tree 1st and 3rd quartiles are indicated for the whole data set mean and standard deviation (SD) are reported for each group
Group Descriptive statistics
Constraining variables
Site characteristics Soil texture Soil composition
Altitude(m asl)
Aspect (deg)
Slope (deg)
Outcr rocks()
Sand()
Silt ()
Clay ()
Org matt()
N () P (ppm)
K (ppm)
CaCO3 ()
pH
1st quartile 407000 000 200 000 6280 1660 580 089 004 1136 11250 008 540 3rd quartile 447500 9000 2000 1500 7760 2780 910 174 009 2469 23250 075 623
T41 Mean 399700 2363 755 000 7867 1403 1051 071 003 1265 16381 049 652SD 1908 2777 894 000 295 180 1434 033 001 668 3318 035 028
T42 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T43 Mean 410500 2880 364 000 7265 2133 602 110 009 2173 17036 053 542SD 2933 4752 930 000 389 372 237 053 020 1234 6129 051 035
T44 Mean 440287 8739 2061 1787 6698 2444 849 171 008 2219 19384 045 572SD 22159 5242 1431 1192 976 866 304 083 004 1471 11137 039 045
T141 Mean 399833 3938 1208 000 7926 1374 1233 052 003 875 17452 049 666SD 2462 2561 901 000 333 203 1853 024 001 142 3181 032 028
T142 Mean 399500 000 075 000 7778 1445 778 099 004 1850 14775 049 631SD 535 000 104 000 214 141 213 022 001 725 3011 041 010
T143 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T144 Mean 411393 643 064 000 7373 1987 640 098 012 2552 18143 052 529SD 1712 2405 145 000 309 274 306 020 027 1421 5395 062 035
T145 Mean 412667 10500 2167 000 6777 2577 647 210 008 1611 18335 071 549SD 4619 1299 2082 000 224 145 093 017 001 516 13137 017 031
T146 Mean 409500 000 000 000 7385 2128 488 078 005 2370 18117 015 575SD 000 000 000 000 042 026 017 003 001 327 1019 019 018
T147 Mean 406750 7875 425 000 7135 2315 550 110 004 1073 11106 082 551SD 3663 5356 299 000 654 654 081 094 004 698 1577 019 031
T148 Mean 414800 7200 1900 1500 7368 2080 552 135 007 2044 9024 019 630SD 570 1006 224 000 618 567 064 037 002 1111 4791 021 015
T149 Mean 408833 9000 600 1833 6960 2093 947 096 005 1216 18803 054 566SD 5795 000 346 577 763 412 357 067 004 098 12521 057 071
T1410 Mean 458688 4922 2731 1813 7623 1591 787 136 006 2448 17087 042 547SD 22773 4043 1590 680 622 535 164 026 001 1461 9485 047 049
T1411 Mean 439614 5727 1114 2173 6501 2430 1038 147 007 1741 22177 052 583SD 16079 2840 551 1297 832 588 420 053 003 597 10086 035 049
T1412 Mean 445650 9000 4000 550 6025 3248 725 248 013 1653 25562 055 579SD 27365 000 2121 636 078 046 120 049 002 467 26427 040 013
T1413 Mean 421318 15341 3518 2409 5954 3227 819 285 014 2823 28413 025 564SD 11402 2814 1419 1338 1099 981 169 117 006 2824 11501 041 022
T1414 Mean 454300 14850 1600 590 6189 3104 707 183 009 2740 11102 065 571SD 6997 2846 615 1058 589 588 071 063 003 830 2759 027 026
life form discourage animals from biting it (Briske 1996) The observed spread of dwarf shrubs is in accordance with the findings of several studies carried out in semiarid environments (Perelman et al 1997 Swinton amp Quiroz 2003 Carilla et al 2012 Catorci et al 2012b 2013) and in pastoral ecosystems subjected to periodic fires (Liedloff
et al 2001) In semi-arid environments annual life span may be considered an avoidance strategy fostered by high grazing intensity (Diacuteaz et al 1999 McGregor 2002 Catorci et al 2012b) because it allows plants to dynami-cally face the grazing pressure by restarting the growth cycle every year (Diacuteaz et al 1999 2001 McGregor 2002)
phyto_44_1_2_103_119_Catorciindd 112phyto_44_1_2_103_119_Catorciindd 112 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx
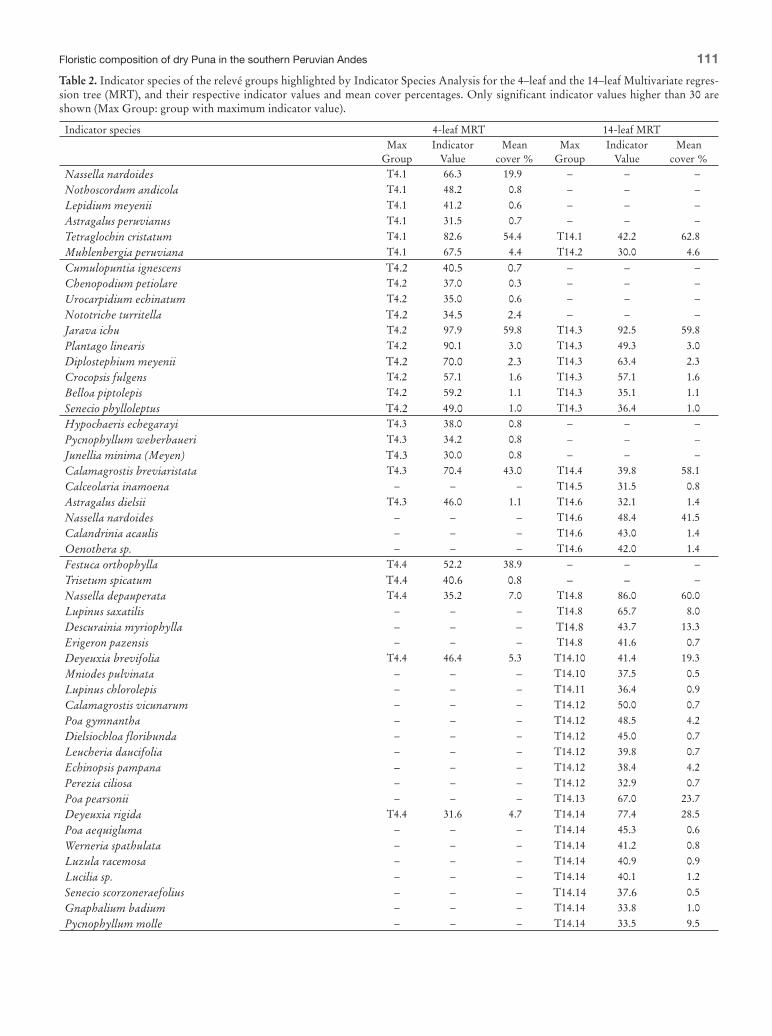
Floristic composition of dry Puna in the southern Peruvian Andes 111
Table 2 Indicator species of the releveacute groups highlighted by Indicator Species Analysis for the 4ndashleaf and the 14ndashleaf Multivariate regres-sion tree (MRT) and their respective indicator values and mean cover percentages Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value)
Indicator species 4-leaf MRT 14-leaf MRTMax
GroupIndicator
ValueMean
cover Max
GroupIndicator
ValueMean
cover Nassella nardoides T41 663 199 ndash ndash ndash
Nothoscordum andicola T41 482 08 ndash ndash ndash
Lepidium meyenii T41 412 06 ndash ndash ndash
Astragalus peruvianus T41 315 07 ndash ndash ndash
Tetraglochin cristatum T41 826 544 T141 422 628
Muhlenbergia peruviana T41 675 44 T142 300 46
Cumulopuntia ignescens T42 405 07 ndash ndash ndash
Chenopodium petiolare T42 370 03 ndash ndash ndash
Urocarpidium echinatum T42 350 06 ndash ndash ndash
Nototriche turritella T42 345 24 ndash ndash ndash
Jarava ichu T42 979 598 T143 925 598
Plantago linearis T42 901 30 T143 493 30
Diplostephium meyenii T42 700 23 T143 634 23
Crocopsis fulgens T42 571 16 T143 571 16
Belloa piptolepis T42 592 11 T143 351 11
Senecio phylloleptus T42 490 10 T143 364 10
Hypochaeris echegarayi T43 380 08 ndash ndash ndash
Pycnophyllum weberbaueri T43 342 08 ndash ndash ndash
Junellia minima (Meyen) T43 300 08 ndash ndash ndash
Calamagrostis breviaristata T43 704 430 T144 398 581
Calceolaria inamoena ndash ndash ndash T145 315 08
Astragalus dielsii T43 460 11 T146 321 14
Nassella nardoides ndash ndash ndash T146 484 415
Calandrinia acaulis ndash ndash ndash T146 430 14
Oenothera sp ndash ndash ndash T146 420 14
Festuca orthophylla T44 522 389 ndash ndash ndash
Trisetum spicatum T44 406 08 ndash ndash ndash
Nassella depauperata T44 352 70 T148 860 600
Lupinus saxatilis ndash ndash ndash T148 657 80
Descurainia myriophylla ndash ndash ndash T148 437 133
Erigeron pazensis ndash ndash ndash T148 416 07
Deyeuxia brevifolia T44 464 53 T1410 414 193
Mniodes pulvinata ndash ndash ndash T1410 375 05
Lupinus chlorolepis ndash ndash ndash T1411 364 09
Calamagrostis vicunarum ndash ndash ndash T1412 500 07
Poa gymnantha ndash ndash ndash T1412 485 42
Dielsiochloa floribunda ndash ndash ndash T1412 450 07
Leucheria daucifolia ndash ndash ndash T1412 398 07
Echinopsis pampana ndash ndash ndash T1412 384 42
Perezia ciliosa ndash ndash ndash T1412 329 07
Poa pearsonii ndash ndash ndash T1413 670 237
Deyeuxia rigida T44 316 47 T1414 774 285
Poa aequigluma ndash ndash ndash T1414 453 06
Werneria spathulata ndash ndash ndash T1414 412 08
Luzula racemosa ndash ndash ndash T1414 409 09
Lucilia sp ndash ndash ndash T1414 401 12
Senecio scorzoneraefolius ndash ndash ndash T1414 376 05
Gnaphalium badium ndash ndash ndash T1414 338 10
Pycnophyllum molle ndash ndash ndash T1414 335 95
phyto_44_1_2_103_119_Catorciindd 111phyto_44_1_2_103_119_Catorciindd 111 120314 0713120314 0713
eschweizerbart_xxx
112 A Catorci et al
Table 3 Descriptive statistics of the environmental variables of the groups highlighted by the 4-leaf and 14-leaf Multivariate regression tree 1st and 3rd quartiles are indicated for the whole data set mean and standard deviation (SD) are reported for each group
Group Descriptive statistics
Constraining variables
Site characteristics Soil texture Soil composition
Altitude(m asl)
Aspect (deg)
Slope (deg)
Outcr rocks()
Sand()
Silt ()
Clay ()
Org matt()
N () P (ppm)
K (ppm)
CaCO3 ()
pH
1st quartile 407000 000 200 000 6280 1660 580 089 004 1136 11250 008 540 3rd quartile 447500 9000 2000 1500 7760 2780 910 174 009 2469 23250 075 623
T41 Mean 399700 2363 755 000 7867 1403 1051 071 003 1265 16381 049 652SD 1908 2777 894 000 295 180 1434 033 001 668 3318 035 028
T42 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T43 Mean 410500 2880 364 000 7265 2133 602 110 009 2173 17036 053 542SD 2933 4752 930 000 389 372 237 053 020 1234 6129 051 035
T44 Mean 440287 8739 2061 1787 6698 2444 849 171 008 2219 19384 045 572SD 22159 5242 1431 1192 976 866 304 083 004 1471 11137 039 045
T141 Mean 399833 3938 1208 000 7926 1374 1233 052 003 875 17452 049 666SD 2462 2561 901 000 333 203 1853 024 001 142 3181 032 028
T142 Mean 399500 000 075 000 7778 1445 778 099 004 1850 14775 049 631SD 535 000 104 000 214 141 213 022 001 725 3011 041 010
T143 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T144 Mean 411393 643 064 000 7373 1987 640 098 012 2552 18143 052 529SD 1712 2405 145 000 309 274 306 020 027 1421 5395 062 035
T145 Mean 412667 10500 2167 000 6777 2577 647 210 008 1611 18335 071 549SD 4619 1299 2082 000 224 145 093 017 001 516 13137 017 031
T146 Mean 409500 000 000 000 7385 2128 488 078 005 2370 18117 015 575SD 000 000 000 000 042 026 017 003 001 327 1019 019 018
T147 Mean 406750 7875 425 000 7135 2315 550 110 004 1073 11106 082 551SD 3663 5356 299 000 654 654 081 094 004 698 1577 019 031
T148 Mean 414800 7200 1900 1500 7368 2080 552 135 007 2044 9024 019 630SD 570 1006 224 000 618 567 064 037 002 1111 4791 021 015
T149 Mean 408833 9000 600 1833 6960 2093 947 096 005 1216 18803 054 566SD 5795 000 346 577 763 412 357 067 004 098 12521 057 071
T1410 Mean 458688 4922 2731 1813 7623 1591 787 136 006 2448 17087 042 547SD 22773 4043 1590 680 622 535 164 026 001 1461 9485 047 049
T1411 Mean 439614 5727 1114 2173 6501 2430 1038 147 007 1741 22177 052 583SD 16079 2840 551 1297 832 588 420 053 003 597 10086 035 049
T1412 Mean 445650 9000 4000 550 6025 3248 725 248 013 1653 25562 055 579SD 27365 000 2121 636 078 046 120 049 002 467 26427 040 013
T1413 Mean 421318 15341 3518 2409 5954 3227 819 285 014 2823 28413 025 564SD 11402 2814 1419 1338 1099 981 169 117 006 2824 11501 041 022
T1414 Mean 454300 14850 1600 590 6189 3104 707 183 009 2740 11102 065 571SD 6997 2846 615 1058 589 588 071 063 003 830 2759 027 026
life form discourage animals from biting it (Briske 1996) The observed spread of dwarf shrubs is in accordance with the findings of several studies carried out in semiarid environments (Perelman et al 1997 Swinton amp Quiroz 2003 Carilla et al 2012 Catorci et al 2012b 2013) and in pastoral ecosystems subjected to periodic fires (Liedloff
et al 2001) In semi-arid environments annual life span may be considered an avoidance strategy fostered by high grazing intensity (Diacuteaz et al 1999 McGregor 2002 Catorci et al 2012b) because it allows plants to dynami-cally face the grazing pressure by restarting the growth cycle every year (Diacuteaz et al 1999 2001 McGregor 2002)
phyto_44_1_2_103_119_Catorciindd 112phyto_44_1_2_103_119_Catorciindd 112 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

112 A Catorci et al
Table 3 Descriptive statistics of the environmental variables of the groups highlighted by the 4-leaf and 14-leaf Multivariate regression tree 1st and 3rd quartiles are indicated for the whole data set mean and standard deviation (SD) are reported for each group
Group Descriptive statistics
Constraining variables
Site characteristics Soil texture Soil composition
Altitude(m asl)
Aspect (deg)
Slope (deg)
Outcr rocks()
Sand()
Silt ()
Clay ()
Org matt()
N () P (ppm)
K (ppm)
CaCO3 ()
pH
1st quartile 407000 000 200 000 6280 1660 580 089 004 1136 11250 008 540 3rd quartile 447500 9000 2000 1500 7760 2780 910 174 009 2469 23250 075 623
T41 Mean 399700 2363 755 000 7867 1403 1051 071 003 1265 16381 049 652SD 1908 2777 894 000 295 180 1434 033 001 668 3318 035 028
T42 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T43 Mean 410500 2880 364 000 7265 2133 602 110 009 2173 17036 053 542SD 2933 4752 930 000 389 372 237 053 020 1234 6129 051 035
T44 Mean 440287 8739 2061 1787 6698 2444 849 171 008 2219 19384 045 572SD 22159 5242 1431 1192 976 866 304 083 004 1471 11137 039 045
T141 Mean 399833 3938 1208 000 7926 1374 1233 052 003 875 17452 049 666SD 2462 2561 901 000 333 203 1853 024 001 142 3181 032 028
T142 Mean 399500 000 075 000 7778 1445 778 099 004 1850 14775 049 631SD 535 000 104 000 214 141 213 022 001 725 3011 041 010
T143 Mean 389929 9321 571 000 5883 3280 837 170 008 2378 21141 051 567SD 5541 4580 189 000 678 638 240 074 004 1608 4710 059 031
T144 Mean 411393 643 064 000 7373 1987 640 098 012 2552 18143 052 529SD 1712 2405 145 000 309 274 306 020 027 1421 5395 062 035
T145 Mean 412667 10500 2167 000 6777 2577 647 210 008 1611 18335 071 549SD 4619 1299 2082 000 224 145 093 017 001 516 13137 017 031
T146 Mean 409500 000 000 000 7385 2128 488 078 005 2370 18117 015 575SD 000 000 000 000 042 026 017 003 001 327 1019 019 018
T147 Mean 406750 7875 425 000 7135 2315 550 110 004 1073 11106 082 551SD 3663 5356 299 000 654 654 081 094 004 698 1577 019 031
T148 Mean 414800 7200 1900 1500 7368 2080 552 135 007 2044 9024 019 630SD 570 1006 224 000 618 567 064 037 002 1111 4791 021 015
T149 Mean 408833 9000 600 1833 6960 2093 947 096 005 1216 18803 054 566SD 5795 000 346 577 763 412 357 067 004 098 12521 057 071
T1410 Mean 458688 4922 2731 1813 7623 1591 787 136 006 2448 17087 042 547SD 22773 4043 1590 680 622 535 164 026 001 1461 9485 047 049
T1411 Mean 439614 5727 1114 2173 6501 2430 1038 147 007 1741 22177 052 583SD 16079 2840 551 1297 832 588 420 053 003 597 10086 035 049
T1412 Mean 445650 9000 4000 550 6025 3248 725 248 013 1653 25562 055 579SD 27365 000 2121 636 078 046 120 049 002 467 26427 040 013
T1413 Mean 421318 15341 3518 2409 5954 3227 819 285 014 2823 28413 025 564SD 11402 2814 1419 1338 1099 981 169 117 006 2824 11501 041 022
T1414 Mean 454300 14850 1600 590 6189 3104 707 183 009 2740 11102 065 571SD 6997 2846 615 1058 589 588 071 063 003 830 2759 027 026
life form discourage animals from biting it (Briske 1996) The observed spread of dwarf shrubs is in accordance with the findings of several studies carried out in semiarid environments (Perelman et al 1997 Swinton amp Quiroz 2003 Carilla et al 2012 Catorci et al 2012b 2013) and in pastoral ecosystems subjected to periodic fires (Liedloff
et al 2001) In semi-arid environments annual life span may be considered an avoidance strategy fostered by high grazing intensity (Diacuteaz et al 1999 McGregor 2002 Catorci et al 2012b) because it allows plants to dynami-cally face the grazing pressure by restarting the growth cycle every year (Diacuteaz et al 1999 2001 McGregor 2002)
phyto_44_1_2_103_119_Catorciindd 112phyto_44_1_2_103_119_Catorciindd 112 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

Floristic composition of dry Puna in the southern Peruvian Andes 113
Prostrate life form is considered an escape strategy in re-sponse to herbivory and studies worldwide have demon-strated that it is fostered by high grazing intensity (Ares et al 1990 Adler amp Morales 1999 Diacuteaz et al 2007 Ca-torci et al 2011a 2012a)
As mentioned before organic matter had a notably lower value in highly disturbed plots This could be due to a number of factors such as erosion treading poach-ing enhanced mineralization and in particular to the camelid behaviour of creating latrine areas namely small areas used by the herd for defecation and urination with a consequent transfer and accumulation of nutrients in specific areas (Naeth et al 1991 Zink et al 1995 Sheath amp Carlson 1998 McGregor 2002) Also there was a lower value of nitrogen percentage in highly disturbed plots This seems to be in contrast with the usual soil nu-trient dynamics (Bai et al 2012) However Pintildeeiro et al (2009) observed a similar trend in upland soils of arid and disturbed environments in Uruguay and Argentina which they hypothesised was due to the volatilization and leaching from urine and dung patches (nitrogen loss hypothesis) As a matter of fact the decrease in soil nutri-ent availability can actually contribute to lower the cover value of graminoids (non-graminoid is an indicator trait of the T41 cluster) (Bret-Harte et al 2004 Brooker 2006) because plants with tolerance strategies such as the graminoids need nutrient availability to restore photo-
synthetic tissues after the herbivores bite (Skarpe 2001) The high pH value observed in the Tetraglochin cristatum ndash dominated community (mean pH of 65 with all other groups below 57 ndash Table 3) might be ascribed to the pe-riodic fires In fact the combustion process releases bound nutrients mainly in elemental or radical form ie as cations When washed into the soil these ions ex-change with H+ ions and the resulting increase in H+ ions in solution increases soil pH (National Wildfire Coordi-nating Group 2001) Lastly the group T41 linked to highly disturbed conditions showed the lowest cover value of potential nurse plants This may contribute to explain the recorded lowest plant diversity within the 4-leaf tree probably due to the decrease in beneficiary species that in semiarid grazed systems are hosted inside or under the canopy of nurse plants (Padilla amp Pugnaire 2006 Arredondo-Nuntildeez et al 2009 Monteiro et al 2011 Catorci et al 2011b 2013)
The other three groups yielded by the 4-leaved MRT (Fig 2a) coincide with the plant communities of the dry Puna landscape undergoing moderate-low level of distur-bance intensity (grazing of large herds of domestic came-lids presence of wild camelids and sporadically small herds of domestic camelids) while altitude and the abun-dance of outcropping rocks are key environmental fac-tors discriminating among the clusters (Fig 2a) The Jar-ava ichu ndash dominated plant community (group T42) dif-
Table 4 Indicator trait states and respective indicator values and P values of the releveacute groups highlighted by Indicator Species Analysis for the 4-leaf Multivariate regression tree (T41 vs T42 vs T43 vs T44) The analysis was run also using the two groups which segregate at the first partition of the tree (T41 vs T42+T43+T44) Only significant indicator values higher than 30 are shown (Max Group group with maximum indicator value P proportion of randomized trials with an indicator value equal to or exceeding the observed indicator value)
Groups compared Trait Trait state Max group Indicator value P
T41vs
T42vs
T43vs
T44
Plant form graminoid T42 304 0037
non-graminoid T41 409 0000
hemirosulate forb T42 307 0005
dwarf shrub T41 625 0000
cushion plant T43 437 0009
succulent plant T42 392 0005
Nurse ability nurse T42 462 0000
T41vs
T42+T43+T44
Life span annual T41 577 0011
perennial T42+T43+T44 508 0017
Plant form graminoid T42+T43+T44 658 0000
non-graminoid T41 666 0000
upright forb T42+T43+T44 822 0000
hemirosulate forb T42+T43+T44 688 0000
tall shrub T42+T43+T44 636 0018
dwarf shrub T41 832 0000
cushion plant T42+T43+T44 463 0022
Nurse ability nurse T42+T43+T44 918 0000
phyto_44_1_2_103_119_Catorciindd 113phyto_44_1_2_103_119_Catorciindd 113 120314 0713120314 0713
eschweizerbart_xxx
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx
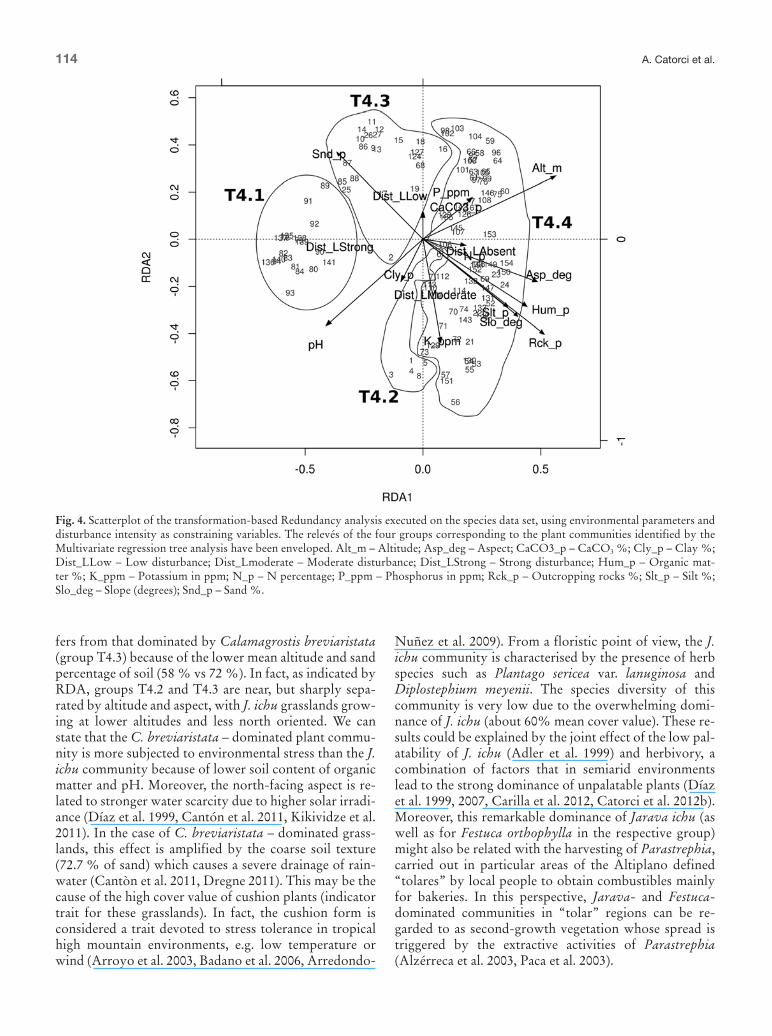
114 A Catorci et al
fers from that dominated by Calamagrostis breviaristata (group T43) because of the lower mean altitude and sand percentage of soil (58 vs 72 ) In fact as indicated by RDA groups T42 and T43 are near but sharply sepa-rated by altitude and aspect with J ichu grasslands grow-ing at lower altitudes and less north oriented We can state that the C breviaristata ndash dominated plant commu-nity is more subjected to environmental stress than the J ichu community because of lower soil content of organic matter and pH Moreover the north-facing aspect is re-lated to stronger water scarcity due to higher solar irradi-ance (Diacuteaz et al 1999 Cantoacuten et al 2011 Kikividze et al 2011) In the case of C breviaristata ndash dominated grass-lands this effect is amplified by the coarse soil texture (727 of sand) which causes a severe drainage of rain-water (Cantograven et al 2011 Dregne 2011) This may be the cause of the high cover value of cushion plants (indicator trait for these grasslands) In fact the cushion form is considered a trait devoted to stress tolerance in tropical high mountain environments eg low temperature or wind (Arroyo et al 2003 Badano et al 2006 Arredondo-
Nuntildeez et al 2009) From a floristic point of view the J ichu community is characterised by the presence of herb species such as Plantago sericea var lanuginosa and Diplostephium meyenii The species diversity of this community is very low due to the overwhelming domi-nance of J ichu (about 60 mean cover value) These re-sults could be explained by the joint effect of the low pal-atability of J ichu (Adler et al 1999) and herbivory a combination of factors that in semiarid environments lead to the strong dominance of unpalatable plants (Diacuteaz et al 1999 2007 Carilla et al 2012 Catorci et al 2012b) Moreover this remarkable dominance of Jarava ichu (as well as for Festuca orthophylla in the respective group) might also be related with the harvesting of Parastrephia carried out in particular areas of the Altiplano defined ldquotolaresrdquo by local people to obtain combustibles mainly for bakeries In this perspective Jarava- and Festuca-dominated communities in ldquotolarrdquo regions can be re-garded to as second-growth vegetation whose spread is triggered by the extractive activities of Parastrephia (Alzeacuterreca et al 2003 Paca et al 2003)
Fig 4 Scatterplot of the transformation-based Redundancy analysis executed on the species data set using environmental parameters and disturbance intensity as constraining variables The releveacutes of the four groups corresponding to the plant communities identified by the Multivariate regression tree analysis have been enveloped Alt_m ndash Altitude Asp_deg ndash Aspect CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic mat-ter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slt_p ndash Silt Slo_deg ndash Slope (degrees) Snd_p ndash Sand
phyto_44_1_2_103_119_Catorciindd 114phyto_44_1_2_103_119_Catorciindd 114 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx
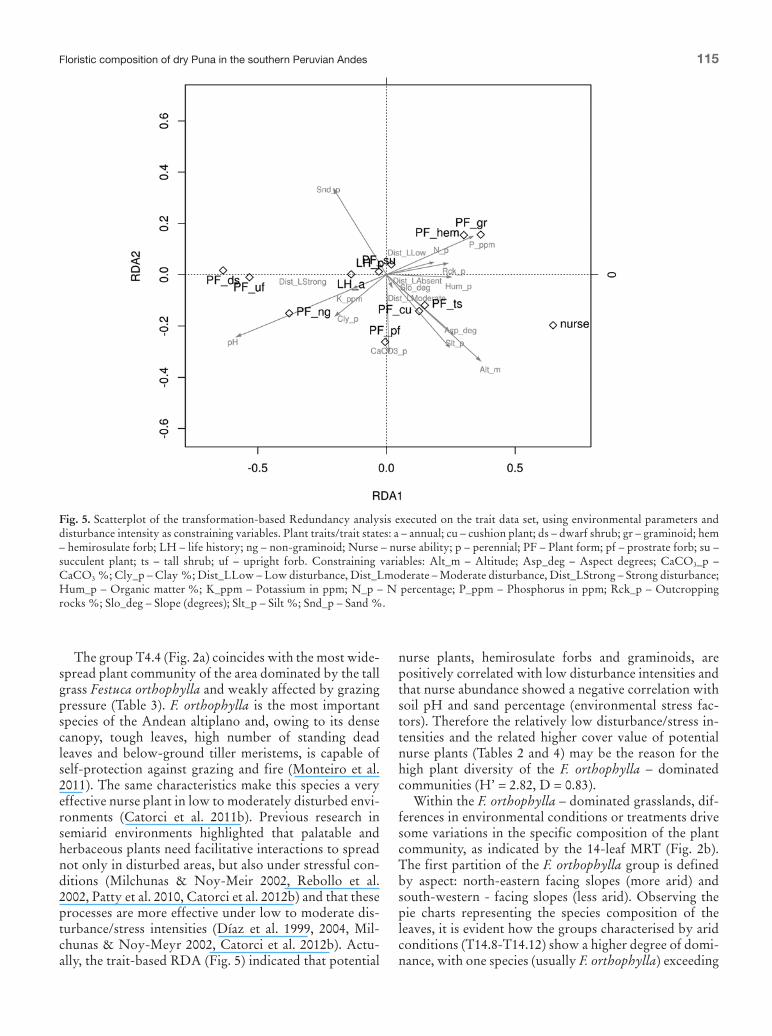
Floristic composition of dry Puna in the southern Peruvian Andes 115
Fig 5 Scatterplot of the transformation-based Redundancy analysis executed on the trait data set using environmental parameters and disturbance intensity as constraining variables Plant traitstrait states a ndash annual cu ndash cushion plant ds ndash dwarf shrub gr ndash graminoid hem ndash hemirosulate forb LH ndash life history ng ndash non-graminoid Nurse ndash nurse ability p ndash perennial PF ndash Plant form pf ndash prostrate forb su ndash succulent plant ts ndash tall shrub uf ndash upright forb Constraining variables Alt_m ndash Altitude Asp_deg ndash Aspect degrees CaCO3_p ndash CaCO3 Cly_p ndash Clay Dist_LLow ndash Low disturbance Dist_Lmoderate ndash Moderate disturbance Dist_LStrong ndash Strong disturbance Hum_p ndash Organic matter K_ppm ndash Potassium in ppm N_p ndash N percentage P_ppm ndash Phosphorus in ppm Rck_p ndash Outcropping rocks Slo_deg ndash Slope (degrees) Slt_p ndash Silt Snd_p ndash Sand
The group T44 (Fig 2a) coincides with the most wide-spread plant community of the area dominated by the tall grass Festuca orthophylla and weakly affected by grazing pressure (Table 3) F orthophylla is the most important species of the Andean altiplano and owing to its dense canopy tough leaves high number of standing dead leaves and below-ground tiller meristems is capable of self-protection against grazing and fire (Monteiro et al 2011) The same characteristics make this species a very effective nurse plant in low to moderately disturbed envi-ronments (Catorci et al 2011b) Previous research in semiarid environments highlighted that palatable and herbaceous plants need facilitative interactions to spread not only in disturbed areas but also under stressful con-ditions (Milchunas amp Noy-Meir 2002 Rebollo et al 2002 Patty et al 2010 Catorci et al 2012b) and that these processes are more effective under low to moderate dis-turbancestress intensities (Diacuteaz et al 1999 2004 Mil-chunas amp Noy-Meyr 2002 Catorci et al 2012b) Actu-ally the trait-based RDA (Fig 5) indicated that potential
nurse plants hemirosulate forbs and graminoids are positively correlated with low disturbance intensities and that nurse abundance showed a negative correlation with soil pH and sand percentage (environmental stress fac-tors) Therefore the relatively low disturbancestress in-tensities and the related higher cover value of potential nurse plants (Tables 2 and 4) may be the reason for the high plant diversity of the F orthophylla ndash dominated communities (Hrsquo = 282 D = 083)
Within the F orthophylla ndash dominated grasslands dif-ferences in environmental conditions or treatments drive some variations in the specific composition of the plant community as indicated by the 14-leaf MRT (Fig 2b) The first partition of the F orthophylla group is defined by aspect north-eastern facing slopes (more arid) and south-western - facing slopes (less arid) Observing the pie charts representing the species composition of the leaves it is evident how the groups characterised by arid conditions (T148-T1412) show a higher degree of domi-nance with one species (usually F orthophylla) exceeding
phyto_44_1_2_103_119_Catorciindd 115phyto_44_1_2_103_119_Catorciindd 115 120314 0713120314 0713
eschweizerbart_xxx
116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

116 A Catorci et al
the cover value of 50 Conversely plant species are more evenly distributed in relevegraves of groups character-ized by less arid conditions (T1413 and T1414) and also growth forms reach a higher diversity in these sites char-acterised by graminoids (Poa sp pl Calamagrostis sp pl Stipa hans-meyeri) hemirosulate forbs (Anthericum collinum Lucilia sp pl) cushion plants (Pycnophyllum sp pl Azorella sp pl) and above all a wide variety of potential nurse species (F orthophylla Deyeuxia rigida Parastrephia sp pl Pycnophyllum molle) These obser-vations are in contrast with the findings of Lambrinos et al (2006) in the Chilean Puna in which xeric aspects seem to promote a higher species richness and variability of plant growth forms and demonstrate how the Peru-vian Puna despite being shaped by similar environmental stress and disturbance factors has peculiar characteristics that deserve specific research and management efforts
Soil parameters define further partitions for a more de-tailed differentiation among south- and north-facing sub-clusters Among north-facing releveacute groups in cluster T148 characterised by lower soil acidity Nassella dep-auperata is the dominant species The lower coverage of F orthophylla in this cluster can be due to the heavier dis-turbance regime as also reflected in the reduced number of species and higher degree of dominance (Tables 2 and 3 Fig 3b) Conversely in group T1410 the absence of grazing disturbance (Table 3 Fig 3b) promotes a more even distribution of species with high coverage of palat-able plants such as Deyeuxia brevifolia South-facing groups are in turn separated by potassium content in soil Particularly group T1413 with a higher concentration of potassium showed a higher cover value of graminoid species (eg Poa pearsonii) Actually graminoids have been proven to be favoured by high potassium concen-trations in arid environments (Bell 1982 OrsquoConnor 2005)
The last group (T1414) represents a peculiar environ-ment characterised by a substantial dominance of differ-ent types of nurse species (the tussock grasses C brevia-ristata and F orthophylla the cushion plants Pycnophyl-lum molle and P weberbaueri and the shrub species Parastrephia lucida and P quadrangularis) This commu-nity located near the upper altitudinal limit of vegetation and characterised by cool aspects and very low distur-bance strongly depends on facilitation processes for its development due to the thin and coarse soil and the se-vere radiation and thermal fluctuations (Schlesinger 1990 Kikividze et al 2011) In particular it was demonstrated that seed sprouting and seedling survival of shrub and graminoid species are fostered by the favourable condi-tions (deeper soils seed trap and protection) provided by cushion plants (Cavieres et al 2007) that actually showed a very high cover value in this type of grassland
Conclusions
Our results confirm the hypothesis that in the dry Puna the floristic composition of the different plant communi-ties is mainly determined by altitude aspect and rocki-ness
Besides disturbance intensity appeared to be a main factor in changing the floristic diversity of the vegetation also modifying the site ecology (soil nutrient content and pH) and the structure of ecosystems (from grass domi-nated systems to dwarf shrub-dominated community) The local soil features emerged as a secondary driver in the plant community composition with sand percentage pH and amount of organic matter being the most impor-tant variables The cumulative effect of soil physical fea-tures (eg sand percentage) and slope aspect (eg north-facing slopes) also proved to be an important driver In turn high stress intensity (ie dryness or high altitude) emerged as the main driving force for the spread of cush-ion plants The greatest abundance of potential nurse plants as well as the highest species diversity was ob-served in intermediate stress and disturbance intensities
Acknowledgements The authors wish to thank the INIA soil analysis laboratory of Arequipa for soil analyses the Servicio Nacional de Aacutereas Naturales Protegidas por el Es-tado (SERNANP) for the authorization to investigate in the Salinas and Aguada Blanca National Reserve and Sheila Beatty for editing the English usage of the manuscript
References
Adler PB amp Morales JM (1999) Influence of environmental factors and sheep grazing on an Andean grassland ndash J Range Manage 52 471ndash480
Alzeacuterreca H Laura J Prieto G Ceacutespedes J Calle P Var-gas A Cardozo A (2003) Manual de manejo y uso sos-tenible de la tola y los tolares Proyecto Binacional Lago Titicaca La Paz Bolivia 59 pp
Anthelme F amp Dangles O (2012) Plant-plant interactions in tropical alpine environments ndash Perspect Plant Ecol Evol Syst 14 363ndash372
Arakaki M amp Cano A (2003) Composicioacuten floriacutestica de la cuenca del riacuteo Ilo-Moquegua y lomas de Ilo Moquegua Peruacute ndash Rev Per Biol 10 5ndash19
Ares J Beeskow AM Bertiller M Rostagno M Irisarri M Anchorena J Defosseacute G amp Merino C (1990) Struc-tural and dynamic characteristics of overgrazed lands of northern Patagonia Argentina ndash In A Breymeyer (ed) Managed Grasslands pp 149ndash175 ndash Elsevier Amsterdam NL
Armas C amp Pugnaire FI (2005) Plant interactions govern population dynamics in a semi-arid plant community ndash J Ecol 93 978ndash989
Arredondo-Nuntildeez A Badano EI amp Bustamante RO (2009) How beneficial are nurse plants A meta-analysis of the effects of cushion plants on high-Andean plant commu-nities ndash Comm Ecol 10 1ndash6
phyto_44_1_2_103_119_Catorciindd 116phyto_44_1_2_103_119_Catorciindd 116 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

Floristic composition of dry Puna in the southern Peruvian Andes 117
Arroyo MTK Squeo FA Armesto JJ amp Villagraacuten C (1988) Effects of aridity on plant diversity in the northern Chilean Andes results of a natural experiment ndash Ann Mis-souri Bot Gard 75 55-78
Arroyo MTK Cavieres LA Pentildealoza A amp Arroyo-Kalin MA (2003) Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes ndash Plant Ecol 169 121ndash129
Badano EI Jones CG Cavieres LA amp Wright JP (2006) Assessing impacts of ecosystem engineers on community or-ganization a general approach illustrated by effects of a high-Andean cushion plant ndash Oikos 115 369ndash385
Bai Y Wu J Clark CM Pan Q Zhang L Chen S Wang Q amp Han X (2012) Grazing alters ecosystem functioning and CNP stoichiometry of grasslands along a regional pre-cipitation gradient ndash J Appl Ecol 49 1204ndash1215
Bell RHV (1982) The effect of soil nutrient availability on community structure in African ecosystems ndash In Huntley BJ amp Walker BH (eds) Ecology of Tropical Savannas pp 193ndash216 ndash Springer-Verlag Berlin Germany
Biondi E (2011) Phytosociology today Methodological and conceptual evolution ndash Plant Biosyst 145 19-29
Blasi C Facioni L Burrascano S Del Vico E Tilia A amp Rosati L (2012) Submediterranean dry grasslands along the Tyrrhenian sector of central Italy Synecology syndynamics and syntaxonomy ndash Plant Biosyst 146 266ndash290
Borcard D Gillet F amp Legendre P (2011) Numerical eco-logy with R ndash Springer Sci + Business Media New York USA 306 pp
Bowman DL (1990) High altitude camelid pastoralism of the Andes ndash In Galaty JG amp Johnson D (eds) The world of pastoralism herding systems in comparative perspective pp 323ndash352 ndash Guildford Press New York USA
Brako L amp Zarucchi JL (eds 1993) Catalogue of the flowering plants and gymnosperms of Peru ndash Monogr Syst Bot Missouri Bot Gard 45 IndashXL 1ndash1286
Breiman L Friedman JH Olshen RA amp Stone CG (1984) Classification and regression trees ndash Wadsworth In-ternat Group Belmont CA USA 368 pp
Bret-Harte MS Garciacutea EA Sacreacute VM Whorley JR Wagner JL Lippert SC amp Chapin FS III (2004) Plant and soil responses to neighbour removal and fertilization in Alaskan tussock tundra ndash J Ecol 92 635ndash647
Brooker RW (2006) Plant-plant interactions and environ-mental change ndash New Phytologist 171 271ndash284
Briske DD (1996) Strategies of plant survival in grazed sys-tems a functional interpretation ndash In Hodgson Jamp Illius AW (eds) The Ecology and Management of Grazing Sys-tems pp 37ndash68 ndash CAB Internat Wallingford UK
Cantoacuten Y Soleacute-Benet A De Vente J Boix-Fayos C Calvo-Cases A Asensio C amp Puigdefaacutebregas J (2011) A review of runoff generation and soil erosion across scales in semi-arid south-eastern Spain ndash J Arid Environ 75 1254ndash1261
Carilla J Aragoacuten R amp Gurvich DE (2012) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands ndash Acta Oecol 37 337ndash345
Caro D (1992) The socioeconomic and cultural context of Andean pastoralism Constraints and potential for biological research and interventions ndash In Valdivia C (ed) Sustaina-ble crop livestock systems of the Bolivian highlands pp 71ndash92 ndash University of Missouri Columbia MO USA
Catorci A amp Gatti R (2010) Floristic composition and spatial distribution assessment of montane mesophilous grasslands in the central Apennines Italy A multi-scale and diachronic approach ndash Plant Biosyst 144 793ndash804
Catorci A Ottaviani G amp Cesaretti S (2011a) Functional and coenological changes under different long-term manage-ment conditions in Apennine meadows (central Italy) ndash Phytocoenologia 41 45ndash58
Catorci A Cesaretti S Velasquez JL amp Zeballos H (2011b) Plant-plant spatial interactions in the dry Puna (southern Pe-ruvian Andes) ndash Alpine Botany 121 113ndash121
Catorci A Ottaviani G Vitasović Kosić I amp Cesaretti S (2012a) Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland ndash Plant Biosyst 146 352ndash367
Catorci A Tardella FM Cesaretti S Bertellotti M amp San-tolini R (2012b) The interplay among grazing history plant-plant spatial interactions and species traits affects vege-tation recovery processes in Patagonian steppe ndash Comm Ecol 13 253ndash263
Catorci A Cesaretti S Velasquez JL Burrascano S amp Ze-ballos H (2013) Manafement type affects composition and facilitative processes in altoandine dry grassland ndash Acta Oecol 52 19ndash28
Cavieres LA Badano EI Sierra-Almeida A amp Molina-Montenegro MA (2007) Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native plants in the high-Andes of central Chile ndash Arctic Antarctic and Alpine Res 39 229ndash236
Dersquoath G (2002) Multivariate regression trees a new tech-nique for modeling species-environment relationships ndash Ecology 83 1105ndash1117
Diacuteaz S Cabido M Zak M Martinez Carretero E amp Araniacutebar J (1999) Plant functional traits ecosystem struc-ture and land-use history along a climatic gradient in central-western Argentina ndash J Vegetation Sci 10 651ndash660
Diacuteaz S Noy-Meir I amp Cabido M (2001) Can grazing re-sponse of herbaceous plants be predict from simple vegeta-tive traits ndash J Appl Ecol 38 497ndash508
Diacuteaz S Hodgson JG Thompson K Cabido M Cornelis-sen JHC et al (2004) The plant traits that drive ecosys-tems evidence from three continents ndash J Vegetation Sci 15 295ndash304
Diacuteaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas D G Skarpe C et al (2007) Plant trait re-sponses to grazing ndash a global synthesis ndash Global Change Biol 13 313ndash341
Dregne HE (2011) Soils of arid regions Vol 6 ndash Elsevier Amsterdam NL 237 pp
Dufrecircne M amp Legendre P (1997) Species assemblages and in-dicator species the need for a flexible asymmetrical ap-proach ndash Ecol Monogr 67 345ndash366
Galaacuten de Mera A Rosa MV amp Caacuteceres C (2002) Una aproximacioacuten sintaxonoacutemica sobre la vegetacioacuten del Perugrave Clases Oacuterdenes y Alianzas ndash Acta Bot Malacitana 27 75ndash103
Galaacuten de Mera A Caacuteceres C amp Gonzaacuteles A (2003) La vege-tacioacuten de la alta montantildea andina del sur del Perugrave ndash Acta Bot Malacitana 28 121ndash147
Galaacuten de Mera A Linares Perea E Campos de la Cruz J amp Vicente Orellana JA (2009) Nuovas observaciones sobre la vegetacioacuten del sur del Perugrave Del Desierto Pacifico al Altipi-ano ndash Acta Bot Malacitana 34 107ndash144
Galaacuten de Mera AG Perea EL Vera CT amp Benavidea FV (2010) Termoclima y humedad en el sur del Peruacute Bioclima-tologiacutea y bioindicadores en el departamento de Arequipa ndash Zonas Aacuteridas 14 71-83
Galaacuten de Mera A Perea EL de la Cruz JC amp Orellana JAV (2012) Las comunidades vegetales relacionadas con
phyto_44_1_2_103_119_Catorciindd 117phyto_44_1_2_103_119_Catorciindd 117 120314 0713120314 0713
eschweizerbart_xxx
118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

118 A Catorci et al
los ambientes humanos en el sur del Peruacute Plant communities linked with human environments in southern Peru ndash Phyto-coenologia 41 265ndash305
Grime JP (2001) Plant strategies vegetation processes and ecosystem properties (2nd ed) ndash Wiley Chichester UK 456 pp
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities Mechanisms and conse-quences ndash J Vegetation Sci 17 255ndash260
Hodge WH (1960) Yareta ndash fuel umbellifer of the Andean puna ndash Economic Bot 14 113ndash118
Kikvidze Z Michalet R Brooker RW Cavieres LA Lor-tie CJ Pugnaire FI amp Callaway RM (2011) Climatic drivers of plant-plant interactions and diversity in alpine communities ndash Alpine Bot 121 63ndash70
Koumlrner C (2003) Alpine plant life functional plant ecology of high mountain ecosystems (2nd ed) ndash Springer Berlin DE 344 pp
Lambrinos JG Kleier CC amp Rundel PW (2006) Plant community variation across a puna landscape in the Chilean Andes ndash Revista Chilena de Historia Natural 79 233ndash243
Lauenroth WK amp Aguilera MO (1998) Plant-plant interac-tions in grasses and grasslands ndash In Cheplick GP (ed) Population Biology of Grasses pp 209ndash230 ndash Cambridge University Press Cambridge UK
Legendre P amp Gallagher E (2001) Ecologically meaningful transformations for ordination of species data ndash Oecologia 129 271ndash280
Legendre P amp Legendre L (2012) Numerical Ecology (3rd ed) ndash Elsevier Amsterdam NL 853 pp
Liedloff AC Coughenour MB Ludwig JA amp Dyer R (2001) Modeling the trade off between fire and grazing in a tropical savanna landscape northern Australia ndash Environ Internat 27 173ndash180
Luebert F amp Gajardo R (2005) Vegetacioacuten alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacioacuten de la Puna meridional ndash Phytocoenologia 35 79ndash128
Maestre FT amp Cortina J (2005) Remnant shrubs in Mediter-ranean semi-arid steppes Effects of shrub size abiotic fac-tors and species identity on understorey richness and occur-rence ndash Acta Oecol 27 161ndash169
Magurran AE (1988) Ecological diversity and its measure-ment ndash Chapman and Hall London UK 179 pp
McCune B amp Grace JB (2002) Analysis of Ecological Com-munities ndash MjM Software Design Gleneden Beach Oregon 300 pp
McGregor BA (2002) Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures ndash Small Ruminant Res 44 219ndash232
Milchunas DG amp Noy-Meir I (2002) Grazing refuges ex-ternal avoidance of herbivory and plant diversity ndash Oikos 99 113ndash130
Monteiro JAF Hiltbrunner E amp Koumlrner C (2011) Func-tional morphology and microclimate of Festuca orthophylla the dominant tall tussock grass in the Andean Altiplano ndash Flora 206 387ndash396
Montenegro B Zuacutentildeiga S amp Zeballos H (2010) Climatologiacutea de la Reserva Nacional Salinas y Aguada Blanca suroeste del Peruacute ndash In Zeballos H Ochoa JA amp Loacutepez E (eds) Di-versidad bioloacutegica de la Reserva Nacional de Salinas y Aguada Blanca pp 261ndash273 ndash Litho amp arte sac Lima Perugrave
Montesinos D (2011) Diversidad floriacutestica de la cuenca alta del riacuteo Tambo-Ichuntildea (Moquegua Peru) ndash Rev Peru Biol 18 119ndash132
Montesinos DB Cleef AM amp Sykora KV (2012) Andean shrublands of Moquegua South Peru Prepuna plant com-munities ndash Phytocoenologia 42 29ndash55
Myers N Mittermeier RA Mittermeier CG Da Fonseca GAB amp Kent J (2000) Biodiversity hotspots for conser-vation priorities ndash Nature 403 853ndash858
Naeth MA Bailey AW Pluth DJ Chanasyk DS amp Hardin RT (1991) Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta ndash J Range Manage 44 7ndash12
National Wildfire Coordinating Group (2001) Fire Effects Guide ndash Available online at httpwwwnwcggovpmsRxFireFEGpdf (Accessed on Jan 10 2013)
OrsquoConnor TG (2005) Influence of land use on plant commu-nity composition and diversity in Highland Sourveld grass-land in the southern Drakensberg South Africa ndash J Appl Ecol 42 975ndash988
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR OrsquoHara RB Simpson GL Solymos P Stevens MHH amp Wagner H (2013) Vegan Community Ecology Package (v 20-7) ndash Available online at httpCRANR-projectorgpackage=vegan (Accessed on May 4th 2013)
Paca FR Palao A Canaza D Bustinza H Vaacutesquez G Chambilla R amp Chaacutevez M (2003) Repoblamiento de praderas del altiplano con tola en el aacutembito peruano del sis-tema TDPS ndash Projecto Binacional Lago Titicaca Puno Peru 31 pp
Padilla FM amp Pugnaire FI (2006) The role of nurse plants in the restoration of degraded environments ndash Frontiers Ecol Environ 4 196ndash202
Pakeman RJ Lepš J Kleyer M Lavorel S Garnier E amp the VISTA consortium (2009) Relative climatic edaphic and man- agement controls of plant functional trait signatures ndash J Veget Sci 20 148ndash159
Patty L Halloy SRP Hiltbrunner E Koumlrner C (2010) Biomass allocation in herbaceous plants under grazing im-pact in the high semi-arid Andes ndash Flora 205 695ndash703
Perelman S Leoacuten RJC amp Bussacca JP (1997) Floristic changes related to grazing intensity in a Patagonian shrub steppe ndash Ecography 20 400ndash406
Pintildeeiro G Paruelo JM Jobbagravegy EG Jackson RB amp Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temper-ate and subtropical grasslands of South America ndash Global Biogeochem Cycles 23
Podani J (2007) Analisi ed esplorazione multivariata dei dati in ecologia e biologia ndash Liguori Napoli Italy 544 pp
Pugnaire FI amp Luque MT (2001) Changes in plant interac-tions along a gradient of environmental stress ndash Oikos 93 42ndash49
R Core Team (2012) R A language and environment for statis-tical computing ndash R Foundation for Statistical Computing Wien 1730 pp
Rebollo S Milchunas DG Noy-Meir I amp Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities ndash Oikos 98 53ndash64
Reese CA amp Liu KB (2005) A modern pollen rain study from the central Andes region of South America ndash J Bio-geogr 32 709ndash718
Rivas-Martinez S amp Tovar O (1982) Vegetatio Andinae I Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Peru ndash Lazaroa 4 167ndash187
Schlesinger WH Reynolds JF Cunningham GL Huennke LF Jarrell WM Virginia RA amp Whitford WG (1990)
phyto_44_1_2_103_119_Catorciindd 118phyto_44_1_2_103_119_Catorciindd 118 120314 0713120314 0713
eschweizerbart_xxx
Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

Floristic composition of dry Puna in the southern Peruvian Andes 119
Biological feedbacks in global desertification ndash Science 247 1043ndash1048
Schwarzer C Caacuteceres F Cano A La Torre M I Weigend M (2010) 400 years for long-distance dispersal and diver-gence in the northern Atacama desert and insights from the Huaynaputina pumice slopes of Moquegua Peru ndash J Arid Environ 74 1540ndash1551
Sheath GW amp Carlson WT (1998) Impact of cattle treading on hill land 1 Soil damage patterns and pasture status ndash New Zealand J Agricult Res 41 271ndash278
Skarpe C (2001) Effects of Large Herbivores on Competition and Succession in Natural Savannah Rangelands ndash In Tow PG amp Lazenby A (eds) Competition and succession in pastures pp 175ndash192 ndash CABI Publishing Wallingford UK
Smith AP (1994) Introduction to Tropical Alpine Vegetation ndash In Rundel PW Smith AP amp Meinzer FC (eds) Tro-pical Alpine Environments Plant Form and Function pp 1ndash20 ndash Cambridge University Press Cambridge UK
Swinton MS amp Quiroz R (2003) Is Poverty to Blame for Soil Pasture and Forest Degradation in Perursquos Altiplano ndash World Development 31 1903ndash1919
Tewksbury JJ amp Lloyd JD (2001) Positive interactions un-der nurseplants spati al scale stress gradients and benefactor size ndash Oecologia 127 425ndash434
Thomas E Damme P amp Van Goetghebeur P (2010) Some factors determining species diversity of prepuna and puna vegetations in a Bolivian Andes region ndash Plant Ecol Evol 143 31ndash42
Valiente-Banuet A Vite F amp Zavala-Hurtado JA (1991) In-teraction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana ndash J Veget Sci 2 11ndash14
Vuille M Hardy DR Braun C Keimig F amp Bradley RS (1998) Atmospheric circulation anomalies associated with 19961997 summer precipitation events on Sajama Ice Cap Bolivia ndash J Geophys Res 103 11191ndash11204
Warren R (2008) Mechanisms driving understory evergreen herb distributions across slope aspects as derived from land-scape position ndash Plant Ecol 198 297ndash308
Wickens G E (1995) Azorella compacta Umbelliferae a re-view ndash Economic Bot 49 201-212
Zink TA Allen MF Heindl-Tenhunen B amp Allen EB (1995) Disturbance corridor on an ecological reserve ndash Res-toration Ecol 3 304ndash310
Zuloaga FO Morrone ON Belgrano MJ Marticorena C amp Marchesi E (eds 2008) Cataacutelogo de las plantas vascu-lares del Cono Sur ndash Monogr Syst Bot Missouri Bot Gard 107 3 Vols 3348 p
Addresses of the authorsAndrea Catorci Federico Maria Tardella Scuola di Scienze Ambientali Universitagrave di Camerino ItalyJoseacute Luis Velasquez Centro de Estudios y Promocioacuten del De-sarrollo (Desco) Arequipa PeruSabrina Cesaretti Luca Malatesta School of Advanced Studies PhD Course in Environmental Sciences and Public Health Universitagrave di Camerino ItalyHoracio Zeballos Centro Integral de Nutricioacuten e Investigacioacuten (BIENestar) Arequipa PeruCorresponding author lucamalatestaunicamit
Electronic Appendices
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory vari-ables are reported for the first six axesAppendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes
Appendix data associated with this article can be found in the online version atwwwschweizerbartdejournalsphyto
The publishers do not bear any liability for the lack of usability or correctness of supplementary material
phyto_44_1_2_103_119_Catorciindd 119phyto_44_1_2_103_119_Catorciindd 119 120314 0713120314 0713
eschweizerbart_xxx
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx
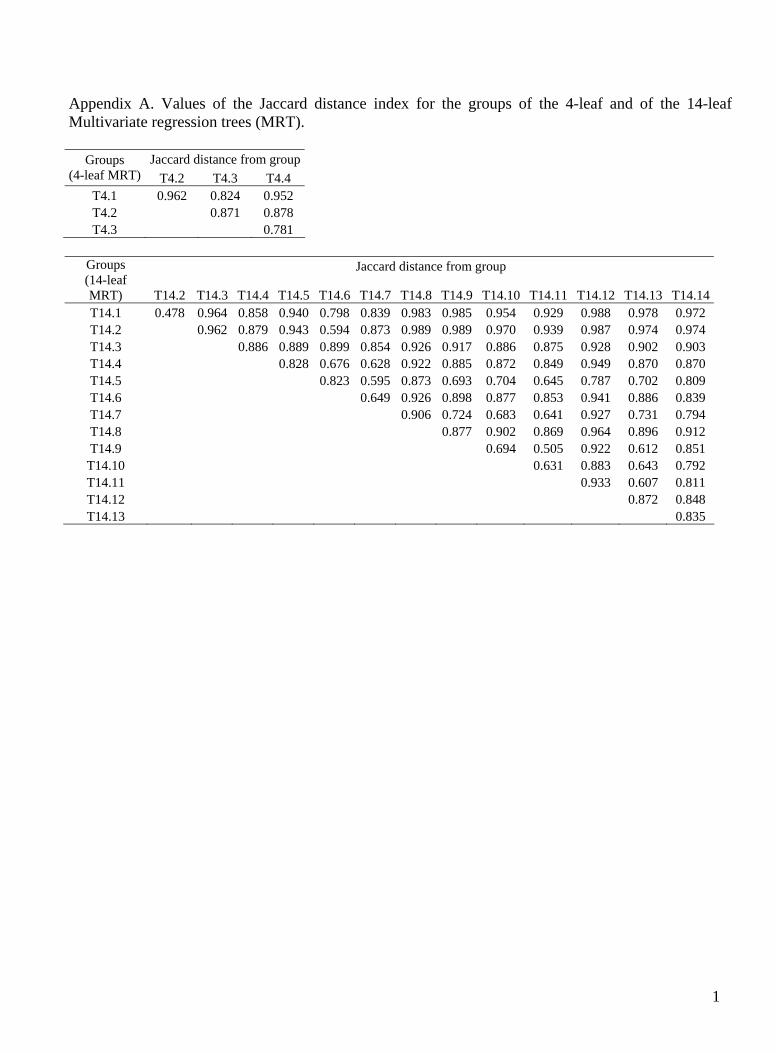
1
Appendix A Values of the Jaccard distance index for the groups of the 4-leaf and of the 14-leaf Multivariate regression trees (MRT)
Jaccard distance from group Groups (4-leaf MRT) T42 T43 T44
T41 0962 0824 0952 T42 0871 0878 T43 0781
Jaccard distance from group Groups
(14-leaf MRT) T142 T143 T144 T145 T146 T147 T148 T149 T1410 T1411 T1412 T1413 T1414T141 0478 0964 0858 0940 0798 0839 0983 0985 0954 0929 0988 0978 0972 T142 0962 0879 0943 0594 0873 0989 0989 0970 0939 0987 0974 0974 T143 0886 0889 0899 0854 0926 0917 0886 0875 0928 0902 0903 T144 0828 0676 0628 0922 0885 0872 0849 0949 0870 0870 T145 0823 0595 0873 0693 0704 0645 0787 0702 0809 T146 0649 0926 0898 0877 0853 0941 0886 0839 T147 0906 0724 0683 0641 0927 0731 0794 T148 0877 0902 0869 0964 0896 0912 T149 0694 0505 0922 0612 0851
T1410 0631 0883 0643 0792 T1411 0933 0607 0811 T1412 0872 0848 T1413 0835
eschweizerbart_xxx
2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

2
Appendix B Biplot scores (scaling 2) for the constraining variables resulting from a transformation-based Redundancy analysis run on the ldquospecies x releveacutesrdquo and ldquoenvironmental variables and disturbance intensity x releveacutesrdquo matrices Scores for explanatory variables are reported for the first six axes
Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6
Site characteristics Altitude 0660 0314 -0445 -0182 0039 -0301 Aspect 0569 -0212 -0368 0283 0045 0516 Slope in degrees 0472 -0383 -0332 -0170 0420 0263 Outcropping rocks 0604 -0473 -0108 -0426 0004 -0270 Soil texture Sand -0424 0429 0113 -0456 0153 -0178 Silt 0421 -0338 -0074 0536 -0127 0343 Clay -0109 -0212 -0141 -0051 -0016 -0196 Soil composition Organic matter 0516 -0334 -0140 0131 0124 0328 N 0214 -0032 0142 0055 0036 0144 Phosphorus in ppm 0247 0205 0059 0146 0028 0125 Potassium in ppm 0089 -0512 0148 0052 0314 -0005 CaCO3 -0001 0138 0013 0095 -0099 -0182 pH -0482 -0431 -0460 -0175 -0277 -0078 Disturbance level Low disturbance 0157 0532 0321 0050 -0540 0478 Moderate disturbance 0103 -0555 0040 0506 -0142 -0561 Strong disturbance -0871 -0066 -0373 -0216 0127 0040
eschweizerbart_xxx
3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx

3
Appendix C Results of a transformation-based Redundancy analysis of plant traits data constrained by environmental variables and disturbance intensity Scores (scaling 2) for traits and constraining variables are reported for the first six axes Constraining variables RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Site characteristics Altitude 0457 -0419 -0074 0206 -0056 0084 Aspect 0328 -0293 0554 -0259 -0086 0271 Slope in degrees 0029 -0064 0358 0056 -0386 0199 Outcropping rocks 0297 0055 0431 0521 -0001 -0175 Soil texture Sand -0255 0415 -0296 -0342 -0228 0262 Silt 0302 -0350 0212 0222 0227 -0171 Clay -0247 -0201 0216 0130 -0187 -0132 Soil composition Organic matter 0318 -0013 0248 0288 -0026 0051 N 0228 0061 -0030 0182 0063 0185 Phosphorus in ppm 0420 0188 -0150 0205 0138 0511 Potassium in ppm -0162 -0067 0194 0590 -0386 -0378 CaCO3 0012 -0385 -0149 0036 0037 -0139 pH -0730 -0304 0141 -0063 0182 -0233 Disturbance level Low disturbance 0392 0213 -0222 -0577 0354 -0059 Moderate disturbance -0070 -0306 0481 0575 0214 -0240 Strong disturbance -0874 0019 -0248 -0038 0013 0218 Traits RDA1 RDA2 RDA3 RDA4 RDA5 RDA6 Life span annual -0137 0001 0115 -0036 0187 -0009 perennial 0019 0039 -0059 0023 -0025 -0018 Plant form graminoid 0330 0159 -0071 0001 0054 -0023 non-graminoid -0378 -0150 0023 0022 0014 -0030 upright forbs -0532 -0010 0044 -0132 -0015 -0008 hemirosulate forbs 0366 0156 -0039 0022 0048 -0044 prostrate forbs -0005 -0262 -0198 0102 0053 -0008 tall shrubs 0149 -0119 -0010 -0083 -0038 -0101 dwarf shrubs -0635 0016 0086 0096 0011 -0018 cushion plants 0127 -0142 -0115 -0115 0036 0038 succulent plants -0030 0012 0073 0058 -0025 -0016 Nurse ability nurse 0647 -0197 0244 0022 -0007 0008
eschweizerbart_xxx




















