
High Fluidity and Soft Elasticity of the Inner Membrane of Escherichia coli Revealed by the Surface...
12
High Fluidity and Soft Elasticity of the Inner Membrane of Escherichia coli Revealed by the Surface Rheology of Model Langmuir Monolayers Iva ´n Lo ´pez-Montero, ²,‡ Laura R. Arriaga, ² Francisco Monroy,* ,² Germa ´n Rivas, § Pedro Tarazona, | and Marisela Ve ´lez ‡ Departamento de Quı ´mica Fı ´sica I, UniVersidad Complutense de Madrid, 28040 Madrid, Spain, Departamento de Proteı ´nas, Centro de InVestigaciones Biolo ´ gicas, CIB-CSIC, 28040 Madrid, Spain, Departamento de Fı ´sica Teo ´ rica de la Materia Condensada and Instituto de Ciencia de los Materiales Nicola ´ s Cabrera, UniVersidad Auto ´ noma de Madrid, 28049 Madrid, Spain ReceiVed October 26, 2007. In Final Form: December 19, 2007 We have studied the equilibrium and linear mechanical properties of model membranes of Escherichia coli built up as Langmuir monolayers of a native lipid extract using surface thermodynamics, fluorescence microscopy, and surface rheology measurements. The experimental study has been carried out at different temperatures across the physiological operative range 15-37 °C. Lipid phase coexistence has been revealed over a broad pressure range by fluorescence microscopy. The presence of ordered domains has been invoked to explain the emergence of shear elasticity accompanying the hydrostatic compression elasticity typical of fluid monolayers. The surface rheology measurements point out the soft character of E. coli membranes; i.e., upon deformation they react as a near-ideal compliant body with minimal energy dissipation, thus optimizing the effectiveness of external stresses in producing membrane deformations. These mechanical features appear to be independent of temperature, suggesting the existence of a passive thermoregulation mechanism. Introduction The fluid mosaic model suggested by Singer and Nicholson 1 in the 1970s attributed a passive role to the lipid component of the membrane, just forming a homogeneous bilayer where membrane proteins are embedded. This picture has been revised in the past few years. 2 Increasing experimental evidence has shown that the lipid composition and distribution on the membrane play an active role not only in modulating protein function and insertion on the membranes 3 but also in determining membrane structural parameters such as curvature and membrane fluidity in dynamic processes such as fusion, division, and motility. 4-6 Fluidity, understood as an inverse viscosity, is one of the principal attributes of biological membranes essential for maintaining the proper distribution and diffusion of membrane-embedded pro- teins. The physiological importance of regulating the viscosity of the membrane, the so-called homeoViscous adaptation, 7 has become evident from studies in which the fatty acid content is adapted to the growing temperature of the organisms. Nowadays, it is a matter of consensus that membrane viscosity is governed by the adequate lipid content. The elastic parameters describing the capacity of the membrane for bending or compressing laterally under the action of an external stimulus have been measured from a variety of biophysical studies carried out on native membranes as well as model membranes built up as artificial bilayers and monolayers. 8 Membrane mechanics is indeed so relevant to cell function that great efforts are being directed toward understanding the mechanical properties of eukaryotic lipid membranes. 8 Much less attention has been devoted to the effect of lipid composition on the mechanical properties of bacterial membranes. 7,9,10 In particular, despite great advances in the understanding of the lipid distribution in relation to membrane fluidity, 11 questions referring to membrane mechanics in prokaryotes remain insufficiently explored. 12 The cytoplasm inner lipid membrane surrounding Gram-negative bacteria does not undergo as many structural and dynamic changes as membranes surrounding eukaryotic cells, in part due to the presence of a second external membrane. However, cell division, which is an essential life process, requires the deformation of the inner membrane at the midcell region during a constriction process and, later, the fusion with the external membrane to finally form two individual daughter cells. 13 The constriction process implies dilation and curvature membrane deformations working together under the concerted action of a constriction force whose source and mechanism remain unknown. 14 Even thought the cell wall structure is complex, it can be conceived of as an elastic mechanical structure. This is how the division process is modeled from continuous mechanics approaches, 15 but precise values of the elastic parameters upon stretching and bending are needed to achieve appropriate conclusions. For a lipid bilayer, similar * Corresponding author. E-mail: [email protected]. ² Departamento de Quı ´mica Fı ´sica I, Universidad Complutense de Madrid. ‡ Instituto de Ciencia de los Materiales Nicola ´s Cabrera, Universidad Auto ´noma de Madrid. § Centro de Investigaciones Biolo ´gicas, CSIC. | Departamento de Fı ´sica Teo ´ rica de la Materia Condensada, Universidad Auto ´noma de Madrid. (1) Singer, S. J.; Nicholson, G. L. Science 1972, 175, 720. (2) Engelman, D. M. Nature 2005, 438, 578. (3) Simons, K.; Ikonen, E. Nature 1997, 387, 569. (4) Malinin, V. S.; Lentz, B. R. Biophys. J. 2004, 86, 2951. (5) Haque, M. E.; Lentz, B. R. Biochemistry 2004, 43, 3507. (6) McMahon, H. T.; Gallop, J. L. Nature 2005, 438, 590. (7) Sinensky, M. Proc. Natl. Acad. Sci. U.S.A. 1974, 71, 522. (8) Boal, D. Mechanics of the Cell; Cambridge Univ. Press: Cambridge, 2002. (9) Overath, P. T. H. Biochemistry 1973, 12, 2625. (10) Herring, F. G.; Krisman, A.; Sedgwick, E. G.; Bragg, P. D. Biochim. Biophys. Acta 1985, 819, 231. (11) Matsumoto, K. K. J.; Nishibori, A.; Hara, H. Mol. Microbiol. 2006, 61, 1110. (12) White, G. F.; Lipski, A.; Hallett, F. R.; Wood, J. M. Biochim. Biophys. Acta. 2000, 1468, 175. (13) Bi, E.; Lutkenhaus, J. Nature 1991, 354, 161. (14) Harry, E.; Monahan, L.; Thompson, L. Int. ReV. Cytol. 2006, 253, 27. (15) Lan, G.; Wolgemuth, C. W.; Sun, S. X. Proc. Natl. Acad. Sci. U.S.A. 2007, 104, 16110. 4065 Langmuir 2008, 24, 4065-4076 10.1021/la703350s CCC: $40.75 © 2008 American Chemical Society Published on Web 03/14/2008
Transcript of High Fluidity and Soft Elasticity of the Inner Membrane of Escherichia coli Revealed by the Surface...

High Fluidity and Soft Elasticity of the Inner Membrane ofEscherichia coliRevealed by the Surface Rheology of Model Langmuir
Monolayers
Ivan Lopez-Montero,†,‡ Laura R. Arriaga,† Francisco Monroy,*,† German Rivas,§Pedro Tarazona,| and Marisela Ve´lez‡
Departamento de Quı´mica Fısica I, UniVersidad Complutense de Madrid, 28040 Madrid, Spain,Departamento de Proteı´nas, Centro de InVestigaciones Biolo´gicas, CIB-CSIC, 28040 Madrid, Spain,
Departamento de Fı´sica Teo´rica de la Materia Condensada and Instituto de Ciencia de los MaterialesNicolas Cabrera, UniVersidad Auto´noma de Madrid, 28049 Madrid, Spain
ReceiVed October 26, 2007. In Final Form: December 19, 2007
We have studied the equilibrium and linear mechanical properties of model membranes ofEscherichia colibuiltup as Langmuir monolayers of a native lipid extract using surface thermodynamics, fluorescence microscopy, andsurface rheology measurements. The experimental study has been carried out at different temperatures across thephysiological operative range 15-37 °C. Lipid phase coexistence has been revealed over a broad pressure range byfluorescence microscopy. The presence of ordered domains has been invoked to explain the emergence of shearelasticity accompanying the hydrostatic compression elasticity typical of fluid monolayers. The surface rheologymeasurements point out the soft character ofE. coli membranes; i.e., upon deformation they react as a near-idealcompliant body with minimal energy dissipation, thus optimizing the effectiveness of external stresses in producingmembrane deformations. These mechanical features appear to be independent of temperature, suggesting the existenceof a passive thermoregulation mechanism.
Introduction
The fluid mosaic model suggested by Singer and Nicholson1
in the 1970s attributed a passive role to the lipid component ofthe membrane, just forming a homogeneous bilayer wheremembrane proteins are embedded. This picture has been revisedin the past few years.2 Increasing experimental evidence hasshown that the lipid composition and distribution on the membraneplay an active role not only in modulating protein function andinsertion on the membranes3 but also in determining membranestructural parameters such as curvature and membrane fluidityin dynamic processes such as fusion, division, and motility.4-6
Fluidity, understood as an inverse viscosity, is one of the principalattributes of biological membranes essential for maintaining theproper distribution and diffusion of membrane-embedded pro-teins. The physiological importance of regulating the viscosityof the membrane, the so-calledhomeoViscous adaptation,7 hasbecome evident from studies in which the fatty acid content isadapted to the growing temperature of the organisms. Nowadays,it is a matter of consensus that membrane viscosity is governedby the adequate lipid content.
The elastic parameters describing the capacity of the membranefor bending or compressing laterally under the action of an externalstimulus have been measured from a variety of biophysical studies
carried out on native membranes as well as model membranesbuilt up as artificial bilayers and monolayers.8 Membranemechanics is indeed so relevant to cell function that great effortsare being directed toward understanding the mechanical propertiesof eukaryotic lipid membranes.8 Much less attention has beendevoted to the effect of lipid composition on the mechanicalproperties of bacterial membranes.7,9,10In particular, despite greatadvances in the understanding of the lipid distribution in relationto membrane fluidity,11 questions referring to membranemechanics in prokaryotes remain insufficiently explored.12 Thecytoplasm inner lipid membrane surrounding Gram-negativebacteria does not undergo as many structural and dynamic changesas membranes surrounding eukaryotic cells, in part due to thepresence of a second external membrane. However, cell division,which is an essential life process, requires the deformation of theinner membrane at the midcell region during a constriction processand, later, the fusion with the external membrane to finally formtwo individual daughter cells.13The constriction process impliesdilation and curvature membrane deformations working togetherunder the concerted action of a constriction force whose sourceand mechanism remain unknown.14 Even thought the cell wallstructure is complex, it can be conceived of as an elasticmechanical structure. This is how the division process is modeledfrom continuous mechanics approaches,15 but precise values ofthe elastic parameters upon stretching and bending are neededto achieve appropriate conclusions. For a lipid bilayer, similar* Corresponding author. E-mail: [email protected].
† Departamento de Quı´mica Fısica I, Universidad Complutense de Madrid.‡ Instituto de Ciencia de los Materiales Nicola´s Cabrera, Universidad
Autonoma de Madrid.§ Centro de Investigaciones Biolo´gicas, CSIC.| Departamento de Fı´sica Teo´rica de la Materia Condensada, Universidad
Autonoma de Madrid.(1) Singer, S. J.; Nicholson, G. L.Science1972, 175, 720.(2) Engelman, D. M.Nature2005, 438, 578.(3) Simons, K.; Ikonen, E.Nature1997, 387, 569.(4) Malinin, V. S.; Lentz, B. R.Biophys. J.2004, 86, 2951.(5) Haque, M. E.; Lentz, B. R.Biochemistry2004, 43, 3507.(6) McMahon, H. T.; Gallop, J. L.Nature2005, 438, 590.(7) Sinensky, M.Proc. Natl. Acad. Sci. U.S.A.1974, 71, 522.
(8) Boal, D.Mechanics of the Cell; Cambridge Univ. Press: Cambridge, 2002.(9) Overath, P. T. H.Biochemistry1973, 12, 2625.(10) Herring, F. G.; Krisman, A.; Sedgwick, E. G.; Bragg, P. D.Biochim.
Biophys. Acta1985, 819, 231.(11) Matsumoto, K. K. J.; Nishibori, A.; Hara, H.Mol. Microbiol. 2006, 61,
1110.(12) White, G. F.; Lipski, A.; Hallett, F. R.; Wood, J. M.Biochim. Biophys.
Acta.2000, 1468, 175.(13) Bi, E.; Lutkenhaus, J.Nature1991, 354, 161.(14) Harry, E.; Monahan, L.; Thompson, L.Int. ReV. Cytol. 2006, 253, 27.(15) Lan, G.; Wolgemuth, C. W.; Sun, S. X.Proc. Natl. Acad. Sci. U.S.A.
2007, 104, 16110.
4065Langmuir2008,24, 4065-4076
10.1021/la703350s CCC: $40.75 © 2008 American Chemical SocietyPublished on Web 03/14/2008

mechanical work is required for dilation and bending,8,15 thusdifferent experimental efforts might be exerted to get insight onthe material relationships describing both modes of deformation.
Escherichia coliis an extensively studied prokaryote model.Its lipid formula at the inner membrane is known to be basedon three major phospholipids, phosphatidylethanolamine (PE),phosphatidylglycerol (PG), and cardiolipin (CL). Most workson E. coli membranes focus on the determination of thephysiological role of the different phospholipids and their locationin specific membrane sites.16 The use of selective stainingfluorescent dyes has allowed for the visualization in vivo of thelateral lipid organization across the division process; indeed, aspecialized lipid distribution based on CL-rich and PE-richdomains has been found in the midcell region ofE. colimembranesduring division.16,17
The present work aims to get insight on the dilationalviscoelastic behavior of the lipids extracted from the innermembranes ofE. coli, with the challenging perspective of betterunderstanding how the membrane deforms during the divisionprocess. We have used the commercially available polar lipidextract (E. coliPLE), which is devoid of the 18% of unspecifiednoncharged lipids present in the total lipid extract but enrichedin its relative content of PE, similar to the situation found at thedivision site. This enrichment is likely to reflect the geometricalrestrictions of accommodating cylindrical-like neutral lipids inhighly curved regions formed during constriction. Since we areinterested in the mechanical behavior of the lipid matrix at thedivision site, we usedE. coli PLE, the commercially availablelipid extract with higher PE relative content. We have takenadvantage of the surface mechanical rheology of model Langmuirmonolayers formed fromE. coli PLE spread on an aqueoussubphase mimicking the pH and ionic strength conditions foundinside the bacterial cell.
Surface Rheology of Langmuir Lipid Monolayers.Surfaceactive substances, most lipids among them, spontaneously adsorbas Langmuir monolayers at the air/water interface. The decreasein free energy upon the spontaneous adsorption of the monolayeris measured by the surface pressureΠ ()γ0 - γ, γ being thesurface tension of the monolayer-covered surface andγ0 its valuefor the bare aqueous surface).Π is indeed recognized to be themacroscopic descriptor for monolayer stability.18The mechanicalbehavior of these interfacial monolayers is characterized bysurface rheology as they possess large elasticity and viscosity.19
Two types of motions are possible in a planar monolayer:transversal, which are mainly restored by interfacial tension, andlongitudinal, which are driven by surface tension gradients orMarangoni gradients. The transverse motions, which could beeventually related to inter-monolayer coupling in bilayers, affectthe molecular distribution along the surface normal but are notefficient to dissipate energy in monolayers. Longitudinal motionsinvolve in-plane lipid motions, where membrane mosaicity andlateral lipid distribution are strongly concerned, and are morerelevant for membrane mechanics. If a Langmuir monolayer ismechanically strained along its plane (e.g., compressed betweenthe barriers of the Langmuir trough), the lateral distribution ofthe monolayer components is altered by the external stimulusand a proportional change in surface pressure is observed. Atdynamic conditions, a small surface dilation caused by aninfinitesimal change of the area accessible to the interfacial
monolayer,δA(t), causes the surface pressure to vary proportion-ally in a quantityδΠ(t), which evolves in time following thestrain functionu(t) ) δA(t)/A. If the monolayer is strained bythe barriers of the trough, the deformation ratio is actually auniaxial elongation-compression one. It must be noticed thatbecause of the uniaxial character of the compressive deformation,the viscoelastic response contains two terms, the hydrostaticcompression (xx) and in-plane shear (xy) response. This is whythe studied rheology is termeddilational.
For small enough amplitudes, the mechanical response of thesystem might be linear. In terms of a generalized stress function,σ, which is the actual change in surface pressure-δΠ producedupon straining the monolayer, the dilational causality functioncan be written as follows:20
whereε is the linear modulus of the dilational elasticity, whichcontains two coupled components together, hydrostatic (K) andshear (S).
The dilational stress acts as a longitudinal restoring force forrecovering the initial state of the monolayer when strain is at thedilating semicycle. Indeed, eq 1 is formally equivalent to Hooke’slaw for a pure elastic 2D-body. The presence of viscous frictionintroduces some delay in the response function, which can bethen written in terms of a viscoelastic modulus including adissipation operator:
The dissipative term in eq 2 accounts for viscous losses, whichare proportional to the dilation rate (Newton’s law). Theproportionality factors are respectively the elastic modulus (ε)and the dilational viscosity (κ), both including compression plusshear components. For small amplitude oscillatory motion,u(t)∼ u0eiωt, with frequencyω, the viscoelastic modulus is a complexquantity
which imposes some delay in the linear viscoelastic response,σ(t) ) εu0eiωt ) σ0ei(ωt+φ) (φ being the phase difference betweenthe applied strain and the observed stress).
The modulus of the complex viscoelasticity in eq 3 determinesthe amplitude of the stress response
and the phase difference (φ) is determined by the losses-to-storage ratio:
In a typical oscillatory barrier experiment performed in the linearviscoelastic regime at a frequencyω, the dilational parametersare calculated from experimental averaged parameters (σ0/u0
andφ) obtained by comparison between the strain-stress timetraces (see Materials and Methods for details). The dilationalelasticity modulus (ε) and the dilational viscosity (κ) are thencalculated as
(16) Dowhan, W.; Mileykovskaya, E.; Bogdanov, M.Biochim. Biophys. Acta.2004, 1666, 19.
(17) Fishov, I.; Woldringh, C. L.Mol. Microbiol. 1999, 32, 1166.(18) Gaines, H. E.Insoluble Monolayers at the Liquid-Gas Interface;
Interscience: New York, 1966.(19) Abraham, B. M.; Ketterson, J. K.Langmuir1985, 1, 708.
(20) Landau, L. D.; Lifshitz, E. M.Theory of Elasticity;Pergamon: Oxford,1986.
σ ) -∆Π ) εu ) (εK + εS)u (1)
σ(t) ) εu(t) ) (ε + κ∂
∂t)u(t) (2)
ε ) ε + iωκ (3)
σ0
u0) |ε| ) (ε2 + ω2
κ2)1/2 (4)
tgφ ) ωκ
ε(5)
4066 Langmuir, Vol. 24, No. 8, 2008 Lo´pez-Montero et al.
Materials and Methods
Lipids. E. coli PLE (PE 67.0 wt %, PG 23.2 wt %, CL 9.8 wt%) (E. coli PLE, >99% pure) and 1,2-dioleyl-sn-glycero-3-phosphoethanolamine-N-carboxylfluorescein (CF-DOPE) were sup-plied by Avanti Polar Lipids.E. coliPLE was dissolved in chloroformto achieve a final concentration of 0.1 or 1 mg/mL, depending onthe type of experiment (point-by-point or continuous compression,respectively). Stock chloroform solutions were stored at-20 °C.
Chemicals. Tris(hydroxymethyl)aminomethane hydrochloridebuffer was purchased from Fluka. KCl (>99.8%, Carlo Erba), MgCl2
(>99.8% Sigma-Aldrich), and chloroform (Chromasolv, Riedel-deHaen) was used as received. Ultrapure water from a Milli-Q system(Millipore; resistivity higher than 18 MΩ cm; organic residual lowerthan 1 ppb;γ ) 72.6 mN/m at 20°C) was used to prepare theaqueous subphase (50 mM Tris, 0.5 M KCl, 5 mM MgCl2) bufferedat pH ) 7.7.
Surface Pressure-Area Isotherms. The monolayer measure-ments were performed using a computer-controlled Langmuir balancewith a maximum area of 700 cm2 (702BAM, NIMA, UK). In orderto minimize the amount of subphase but to maintain a largecompression ratio, a volume reducer with an effective film area ofA0 ) 200 cm2 was used. This is a Teflon machined accessorycommercially available (NIMA). It is placed inside the conventional702BAM trough and preserves its rectangular geometry (650 mmlong, 30 mm width, 3 mm depth) but maintains a large compressionratio. The Teflon trough is equipped with two symmetrically movingbarriers made of hydrophilic Delrin. The surface pressure-areaisotherms (Π-A) were determined forE. coliPLE monolayers spreadon the buffered aqueous subphase at different temperatures. Thesurface pressure was measured using a paper Wilhelmy plate toensure a constant contact angle.21 This surface pressure sensor wasfaced parallel to the barriers. The temperature of the subphase wasmaintained constant within(0.1°C by recirculating water from anexternal thermostatic bath (Lauda) through a jacket placed at thebottom of the trough. The temperature of the subphase is measuredin situ by a Pt-100 sensor. In order to prevent surface perturbationsinduced by the air convection streams and to avoid undesirable dustcontamination, the whole setup is covered by a transparent Plexiglascase. Before each measurement, the subphase surface is cleaned bysweeping and suction. Lipids are dissolved in chloroform (0.1 mg/mL, typically) and then spread on the aqueous surface with a Hamiltonsyringe. By-addition isotherms were recorded after successiveaddition of small aliquots (10µL, typically) of the lipid stock. Elapsingtimes ranging between 2 min at very dilute states (Π ∼ 0) and 5 minat the highest pressures are left in order to allow for solventevaporation and to reach the equilibrium state of the monolayer,which is raised when theΠ is observed to fluctuate less than(0.1mN/m during a period longer than 5 min. Because the aliquot issmall, the present protocol ensures that (a) the lipid density is slightlyincreased between successive points and (b) the amount of chloroformis minimal. As a consequence, (a)Π increases along a quasicontinuouspath (less than 0.5 mN/m per aliquot), thus minimizing stressesagainst the pre-existing pressure, and (b) solvent flooding, typicalof standard protocols involving larger aliquots, is minimized. Nosolvent trapping was observed at the very dilute region of theisotherms, as revealed in control experiments performed by BAMmicroscopy (data not shown). Compression isotherms were recordedupon symmetric uniaxial compression of a diluted monolayer (14µL of 1 mg/mL lipid solution) at a constant compression rate of 10cm2/min. In both cases, the surface pressure is plotted against themean molecular area, obtained by dividing the total surface area bythe numerical amount of lipid spread on the surface (the meanmolecular weight of theE.coli PLE, Mw ) 971 g/mol, has beencalculated as an average from the single species and their relative
percentages in the mixture). Experiments were carried out attemperatures of 15, 25, and 37°C.
Oscillatory Dilational Rheology. This method has been thought-fully described elsewhere.22 Both, amplitude (u0) and period ()2π/ω) of the applied strain are controlled by the sinusoidal motion ofthe barriers of the Langmuir trough, which impose the available areato change in a fashion
whereA0 is the initial area,u0 ) (A - A0)/A0 and is the relativedeformation amplitude,ω is the frequency of the oscillatory motion,andφu accounts for a possible initial phase.
The stress response is measured by the pressure sensor andmonitored as a function of time taking a reading every 1 s. At smalldeformations,u0f0, the response is linear, and theΠ(t)-dependenceclosely follows a sinusoidal profile of frequencyω equal than thatof the strain wave
whereΠ0 is the initial pressure,σ0 the stress amplitude, andφπ aphase factor accounting for the delay imposed by viscous frictionwithin the response. This is characteristic for an elastic linear regime;thus, the linear dilational viscoelastic modulus is a strain-independentconstant that can be calculated from eq 6 by considering a stress-to-strain phase differenceφ ) φπ - φu.
Fluorescence Microscopy.Fluorescence images were taken witha Leica inverted microscope equipped with a Hamamatsu C2400-SIT camera. CF-DOPE was used as a fluorescence probe mixedwith the lipid at a proportion of 0.5 wt %. Because lipid probeaccommodates larger surface areas than a close-packed arrangementof the native lipid, the dye is excluded out of ordered regions, whichappear dark. The probe thus preferentially segregates to the disorderedfluid regions of the monolayer, which appear bright in the images.23
A given image represents an observed area of 700× 500 µm2.
Results
Π-A Isotherms: Aliquot Addition vs Continuous Com-pression. Figure 1 shows the surface pressure-area (Π-A)
(21) Grunfeld, F.ReV. Sci. Intsrum.1993, 64, 548.
(22) Hilles, H.; Monroy, F.; Bonales, L. J.; Ortega, F.; Rubio, R. G.AdV.Colloid. Interface Sci.2006, 122, 67.
(23) Knobler, C. M.Science1990, 249, 870.
ε ) (σ0/u0)cosφ
κ ) (σ0/ωu0)sinφ (6)
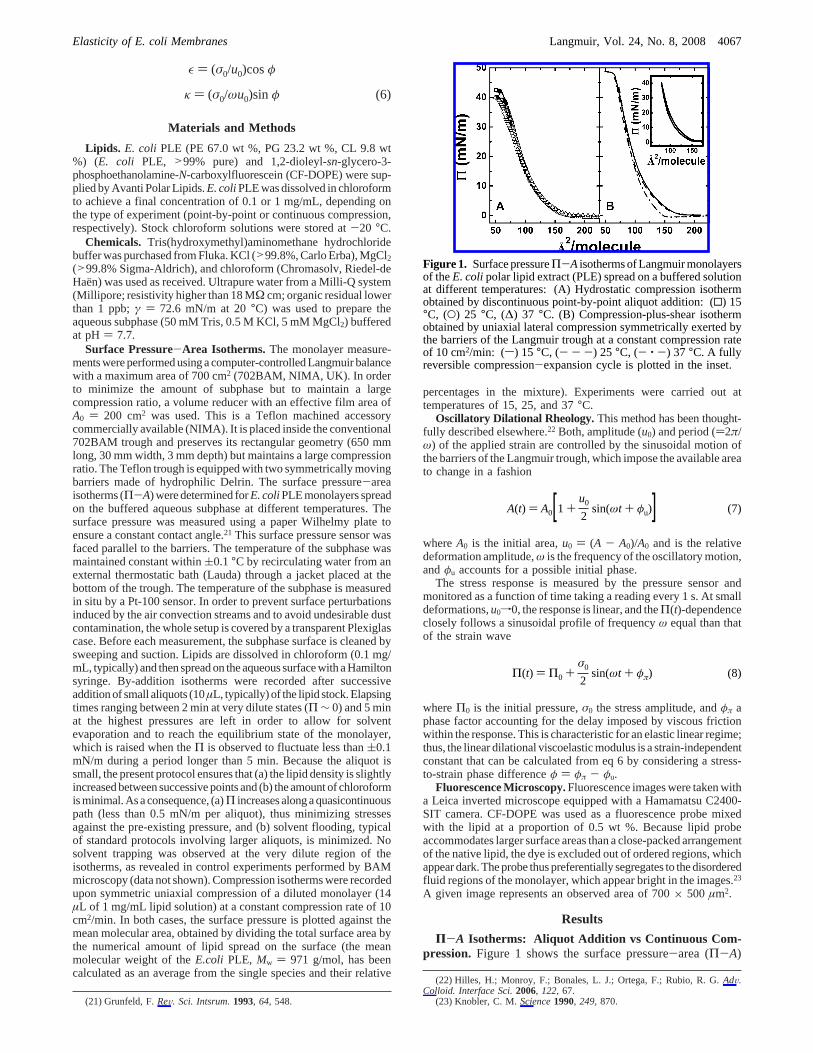
Figure 1. Surface pressureΠ-A isotherms of Langmuir monolayersof theE. colipolar lipid extract (PLE) spread on a buffered solutionat different temperatures: (A) Hydrostatic compression isothermobtained by discontinuous point-by-point aliquot addition: (0) 15°C, (O) 25 °C, (∆) 37 °C. (B) Compression-plus-shear isothermobtained by uniaxial lateral compression symmetrically exerted bythe barriers of the Langmuir trough at a constant compression rateof 10 cm2/min: (s) 15 °C, (- - -) 25 °C, (- ‚ -) 37 °C. A fullyreversible compression-expansion cycle is plotted in the inset.
A(t) ) A0[1 +u0
2sin(ωt + φu)] (7)
Π(t) ) Π0 +σ0
2sin(ωt + φπ) (8)
Elasticity of E. coli Membranes Langmuir, Vol. 24, No. 8, 2008 4067

isotherms of the Langmuir monolayers ofE. coli PLE obtainedby aliquot addition (Figure 1A) and by continuous compression(Figure 1B). Three different temperatures included in thephysiological operative range ofE. coli (15, 25, and 37°C) havebeen studied. The experimental isotherms exhibit a liquidexpanded (LE) shape, displaying very similar features to thoseof monounsaturated phospholipids24,25 and of native lipidmixtures, such as those extracted from erythrocyte membranes.26
The recordedΠ-A isotherms are continuous and differentiableover the whole pressure range up to the collapse, thus indicatingthe mutual miscibility of the lipid components. Although thisobservation could indicate single-phase behavior in simpler two-component monolayers, epifluorescence microscopy revealsphase coexistence persisting over a broadΠ range independentof the compression method (see below). Both, the quasiequi-librium aliquot addition isotherm and the continuous compressionone are almost identical up to lateral pressures near collapse. Formolecular areas below 175 Å2, a typical fluidlike isotherm isdisplayed as the available area is decreased down to ca. 100 Å.2
Uponcompressing further, the isothermentersanear-linearbranchfinishing at monolayer collapse. This is raised at a well-definedpressure plateau atΠc, recorded significantly higher for thecontinuous compression isotherm than for the aliquot-additionone. No hysteresis was observed in the reexpansion cyclesperformed after compression from different initial states up tohigh Π states below the collapse (see the inset in Figure 1B).This behavior points out that expansion recovery is fullyreversible, thus suggesting that no metastable states are reachedindependently of the initial condition, i.e., the studied states arewell at equilibrium. The compression isotherm ofE. coli PLEis found to be qualitatively similar to that previously reportedby Clausell et al. for the total lipid extract (TLE).27 A detailedanalysis reveals slightly larger molecular areas for PLE than forTLE, probably due to a larger electrostatic repulsion in PLE.
The experimental isotherms obtained from both methods werefound to be only weakly dependent on temperature up to highpressures below collapse. This near-athermal behavior of PLEmonolayers is rather general for expanded phases of lipidmonolayers.18This behavior implies that no net entropy changesoccur upon adsorption (∆Sads ) -dΠ/dT ∼ 0), indicating thatmonolayer stability mainly arises from adsorption interactions(Π ) -∆Gads∼-∆Hads). Significant differences emerge betweenthe two isotherms at surface areas smaller than 100 Å2. Whencompressing hydrostatically (aliquot method), the monolayercollapse is reached at a pressure of ca. 40 mN/m, but it is foundto be at least 5 mN/m higher when reached by the continuous(uniaxial) compression method. Collapse is reached at similarsurface area in both methods,Ac∼65 Å2. This value is compatiblewith the close-packing area of bichained charged lipids formingLE monolayers but is meaningfully larger than values found forsaturated hydrocarbon chains at the liquid-crystalline state.24,28
Differences described above could suggest either dynamicalmetastability or, alternatively, an equilibrium scenario whereuniaxial compression induces states with higher stability thanthose reached by the aliquot addition method. The respectiveplausibility of both scenarios will be discussed later in view ofthe coexisting microdomains.
Fluorescence Microscopy: Phase Behavior.Figure 2presents fluorescence micrographs of the lipid monolayer atdifferent surface pressures. The presence of domains is alreadyevident at surface pressures as low as 2-3 mN/m, but it has notbeen possible to determine the lower threshold for the coexistenceregion. At low pressures, lipid domains appear to be rather smalland essentially circular in shape. Domains appear as dark circlesfloating in the bright fluid phase. The fact that they exclude thefluorophore suggests their ordered character with respect to thecontinuous phase. Upon compressing further, the number of thesedomains increases but not their size (typical sizes are distributedaround ca. 2-5µm diameter). Some individual larger (ca. 10µmdiameter) domains are occasionally observed. AtΠ ∼ 35 mN/m,the small domains come into mutual contact and the monolayerenters a homogeneous state made of the dense,ordered, phase.Similar behavior and textures are found when lateral compressionis produced by the aliquot-addition discontinuous method andby the continuous one. This observation points toward the absenceof important dynamical effects on domain formation.
The observed phase coexistence seems thus to be mediatedby a progressive nucleation mechanism rather than the growingmechanism typical of liquid domains.29,30 The persistence ofdomains of small radius seems to support the idea of high orderinside, thus resulting in domains characterized by a high valueof the boundary line tension with the disordered fluid phase.29,30
The indirect evidence sketched above points to the orderedcharacter of the dense phase, but its precise structure (eitherliquid ordered or solid) remains however unknown. The fact that
(24) Mingotaud, A. F.; Mingotaud, C.; Patterson, L. K.Handbook ofMonolayers;Academic Press: San Diego: CA, 1993; Vol. 2.
(25) Cevc, G.Phospholipids Handbook; Marcel Dekker: New York, 1993.(26) Keller, S. L.; Pitcher, W. H., III; Huestis, W. H.; McConnell, H. M.Phys.
ReV. Lett. 1998, 81, 5019.(27) Clausell, A.; Busquets, M. A.; Pujol, M.; Alsin, A.; Cajal, Y.Biopolymers
2004, 75, 480.(28) Kjaer, K.; Als-Nielsen, J.; Helm, C. A.; Laxhuber, L. A.; Mo¨hwald, H.
Phys. ReV. Lett. 1987, 58, 2224.(29) McConnell, H. M.; Keller, D.; Gaub, H.J. Phys. Chem. 1986, 90, 1717.(30) McConnell, H. M.Annu. ReV. Phys. Chem.1991, 42, 171.
Figure 2. Fluorescence micrographs ofE. coli PLE monolayerscontaining 0.5 wt % fluorescent probe CF-DOPE (scale bar is 100µm). Images represent monolayer states raised along a continuouscompression isotherm at 25°C. At very low pressure,Π ) 1 mN/m,the monolayer exhibits a homogeneous fluorescent backgroundcorresponding to the homogeneous distribution of the fluorescentprobe in the fluid phase. (A) Upon further compression, small domains(2-5µm diameter, typically) start to nucleate above ca. 2-3 mN/m;because of the internal order within these domains, the fluorescentprobe is excluded out, thus appearing as dark spots. (B and C) Thenumber of domains, but not their size, increases as the monolayeris laterally compressed, thus suggesting the continuous nucleationof new domains instead of a growing mechanism (B,Π ) 15 mN/m;C, Π ) 17.5 mN/m, respectively). (D) Above 35 mN/m, imagesbecome again homogeneous, as it corresponds to the continuousordered phase. A typical image of the ordered phase is shown (D,Π ) 37 mN/m). Similar phase coexistence behavior was observedfor monolayers formed by the point-by-point isotherms (data notshown).
4068 Langmuir, Vol. 24, No. 8, 2008 Lo´pez-Montero et al.
the domains stay small, without ripening, suggests a solid phaseinstead of a liquid ordered one.
Equilibrium Elasticity: Dilational (Hydrostatic and Shear)Modulus. The equilibrium elasticity modulus is defined as thechange in monolayer pressure caused by an infinitesimal changein the molecular area
It can be easily calculated from the numerical derivative of theΠ-A isotherms in Figure 1. In the equilibrium isotherms obtainedby aliquot addition (Figure 1A), the increase in surface densityis isotropic; i.e., when adding lipid the hydrostatic change insurface area is directly related to the density increase,δAK/A )-δΓ/Γ; thus, the calculated values of the elasticity moduluscorrespond to the hydrostatic compression modulus,εK ) A(∂Π/∂AK)T ) -Γ(∂Π/∂Γ)T. Because the aliquot is small (10µL), thenumerical derivative of the discrete isotherm in Figure 1A canbe considered representative of an infinitesimal hydrostaticcompression path. On the other hand, a uniaxial compression isthe combination of a pure hydrostatic and a shear deformation.The elasticity modulus calculated from the derivative of theΠ-Aisotherms obtained by continuous uniaxial compression (Figure1B), εD ) (εK + εS) ) A(∂Π/∂AD)T, is therefore referred to asdilational and is the sum of the hydrostatic component (εK) plusthe shear one (εS). As a consequence, permanent shear-inducedredistributions of the structural elements of the monolayer couldcause extra energy storage when the compression is uniaxial.Thus,εS is finite only when order is present, in liquid orderedor solid phases. In disordered fluid phases it vanishes.31 Thus,the measured values of the dilational elasticity normally exceedthe hydrostatic modulus calculated from the aliquot method (εD
) εK + εS g εK) if shearing effects are present. For fluid phases,any structural redistribution is not permanent, and then thedilational modulus equals the hydrostatic modulus,εD ) εK.
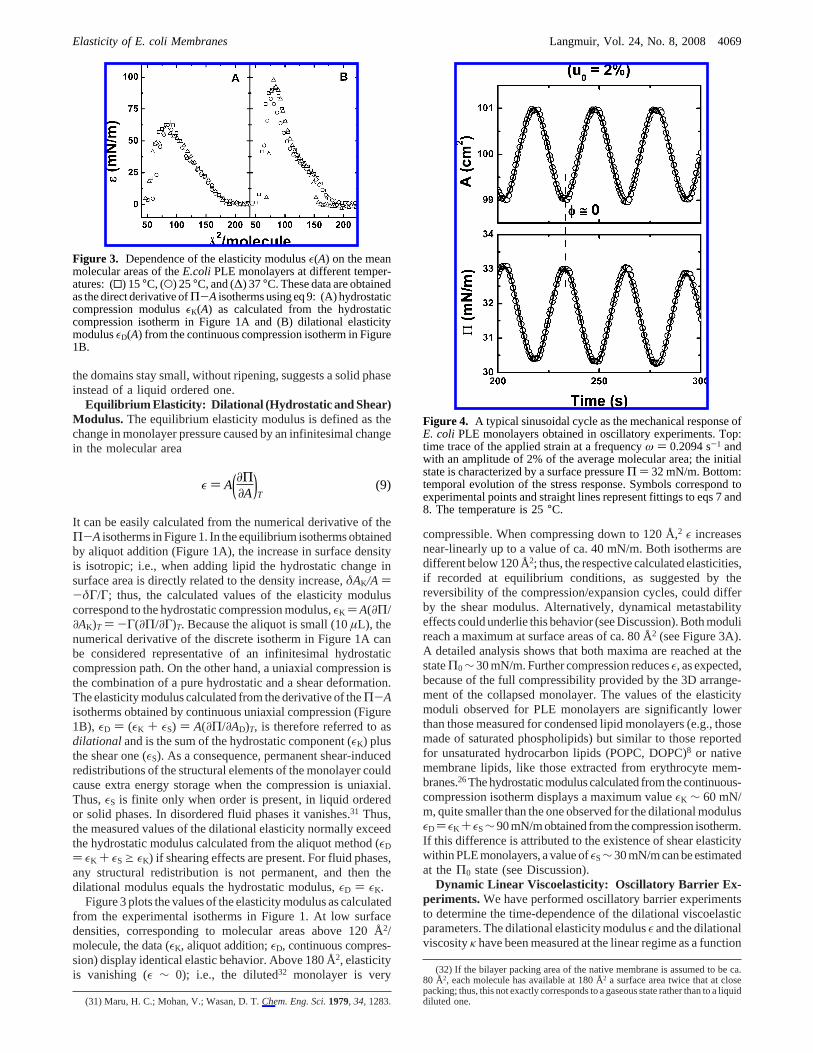
Figure 3 plots the values of the elasticity modulus as calculatedfrom the experimental isotherms in Figure 1. At low surfacedensities, corresponding to molecular areas above 120 Å2/molecule, the data (εK, aliquot addition;εD, continuous compres-sion) display identical elastic behavior. Above 180 Å2, elasticityis vanishing (ε ∼ 0); i.e., the diluted32 monolayer is very
compressible. When compressing down to 120 Å,2 ε increasesnear-linearly up to a value of ca. 40 mN/m. Both isotherms aredifferent below 120 Å2; thus, the respective calculated elasticities,if recorded at equilibrium conditions, as suggested by thereversibility of the compression/expansion cycles, could differby the shear modulus. Alternatively, dynamical metastabilityeffects could underlie this behavior (see Discussion). Both modulireach a maximum at surface areas of ca. 80 Å2 (see Figure 3A).A detailed analysis shows that both maxima are reached at thestateΠ0∼ 30 mN/m. Further compression reducesε, as expected,because of the full compressibility provided by the 3D arrange-ment of the collapsed monolayer. The values of the elasticitymoduli observed for PLE monolayers are significantly lowerthan those measured for condensed lipid monolayers (e.g., thosemade of saturated phospholipids) but similar to those reportedfor unsaturated hydrocarbon lipids (POPC, DOPC)8 or nativemembrane lipids, like those extracted from erythrocyte mem-branes.26The hydrostatic modulus calculated from the continuous-compression isotherm displays a maximum valueεK ∼ 60 mN/m, quite smaller than the one observed for the dilational modulusεD ) εK + εS∼90 mN/m obtained from the compression isotherm.If this difference is attributed to the existence of shear elasticitywithin PLE monolayers, a value ofεS∼30 mN/m can be estimatedat theΠ0 state (see Discussion).
Dynamic Linear Viscoelasticity: Oscillatory Barrier Ex-periments.We have performed oscillatory barrier experimentsto determine the time-dependence of the dilational viscoelasticparameters. The dilational elasticity modulusε and the dilationalviscosityκ have been measured at the linear regime as a function
(31) Maru, H. C.; Mohan, V.; Wasan, D. T.Chem. Eng. Sci.1979, 34, 1283.
(32) If the bilayer packing area of the native membrane is assumed to be ca.80 Å2, each molecule has available at 180 Å2 a surface area twice that at closepacking; thus, this not exactly corresponds to a gaseous state rather than to a liquiddiluted one.
Figure 3. Dependence of the elasticity modulusε(A) on the meanmolecular areas of theE.coli PLE monolayers at different temper-atures: (0) 15°C, (O) 25°C, and (∆) 37°C. These data are obtainedas the direct derivative ofΠ-A isotherms using eq 9: (A) hydrostaticcompression modulusεK(A) as calculated from the hydrostaticcompression isotherm in Figure 1A and (B) dilational elasticitymodulusεD(A) from the continuous compression isotherm in Figure1B.
ε ) A(∂Π∂A)T
(9)
Figure 4. A typical sinusoidal cycle as the mechanical response ofE. coli PLE monolayers obtained in oscillatory experiments. Top:time trace of the applied strain at a frequencyω ) 0.2094 s-1 andwith an amplitude of 2% of the average molecular area; the initialstate is characterized by a surface pressureΠ ) 32 mN/m. Bottom:temporal evolution of the stress response. Symbols correspond toexperimental points and straight lines represent fittings to eqs 7 and8. The temperature is 25°C.
Elasticity of E. coli Membranes Langmuir, Vol. 24, No. 8, 2008 4069
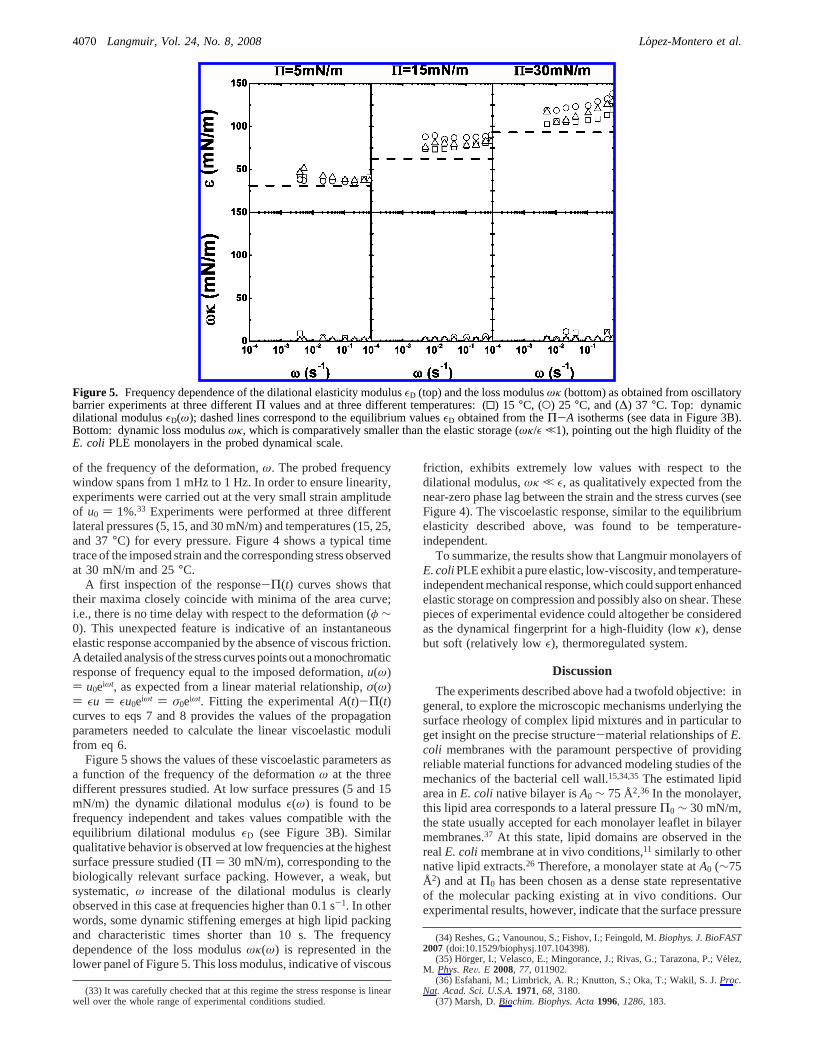
of the frequency of the deformation,ω. The probed frequencywindow spans from 1 mHz to 1 Hz. In order to ensure linearity,experiments were carried out at the very small strain amplitudeof u0 ) 1%.33 Experiments were performed at three differentlateral pressures (5, 15, and 30 mN/m) and temperatures (15, 25,and 37°C) for every pressure. Figure 4 shows a typical timetrace of the imposed strain and the corresponding stress observedat 30 mN/m and 25°C.
A first inspection of the response-Π(t) curves shows thattheir maxima closely coincide with minima of the area curve;i.e., there is no time delay with respect to the deformation (φ ∼0). This unexpected feature is indicative of an instantaneouselastic response accompanied by the absence of viscous friction.A detailed analysis of the stress curves points out a monochromaticresponse of frequency equal to the imposed deformation,u(ω)) u0eiωt, as expected from a linear material relationship,σ(ω)) εu ) εu0eiωt ) σ0eiωt. Fitting the experimentalA(t)-Π(t)curves to eqs 7 and 8 provides the values of the propagationparameters needed to calculate the linear viscoelastic modulifrom eq 6.
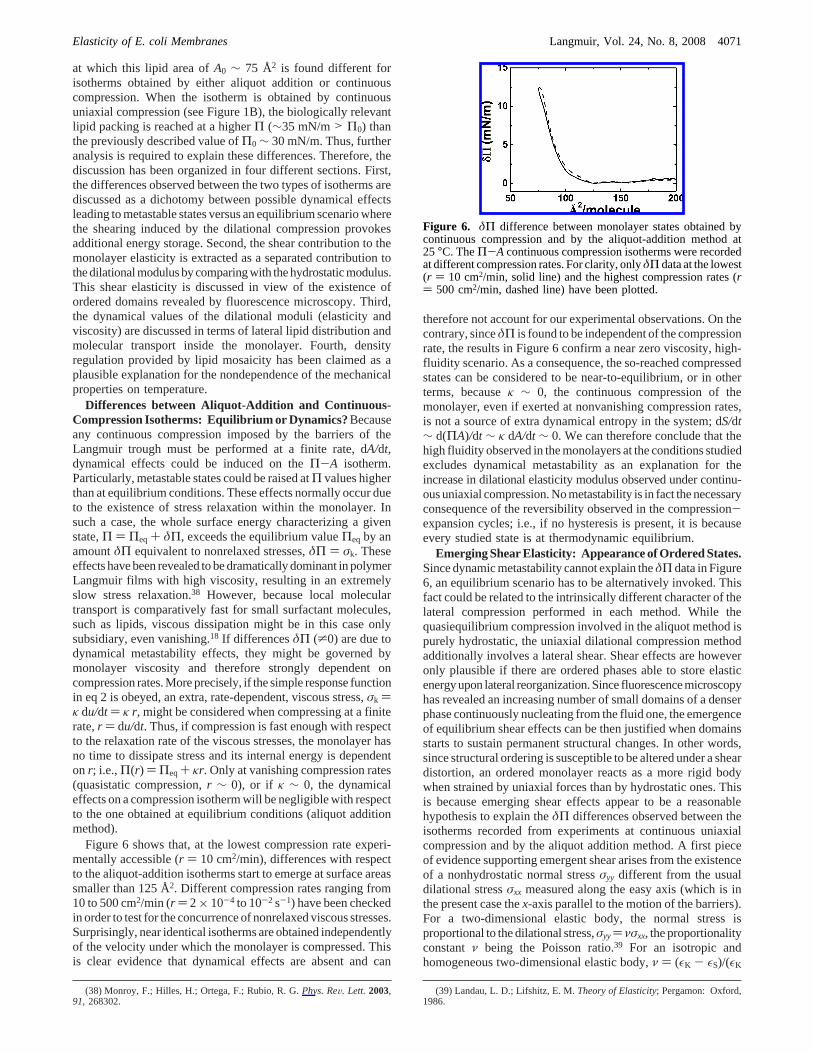
Figure 5 shows the values of these viscoelastic parameters asa function of the frequency of the deformationω at the threedifferent pressures studied. At low surface pressures (5 and 15mN/m) the dynamic dilational modulusε(ω) is found to befrequency independent and takes values compatible with theequilibrium dilational modulusεD (see Figure 3B). Similarqualitative behavior is observed at low frequencies at the highestsurface pressure studied (Π ) 30 mN/m), corresponding to thebiologically relevant surface packing. However, a weak, butsystematic,ω increase of the dilational modulus is clearlyobserved in this case at frequencies higher than 0.1 s-1. In otherwords, some dynamic stiffening emerges at high lipid packingand characteristic times shorter than 10 s. The frequencydependence of the loss modulusωκ(ω) is represented in thelower panel of Figure 5. This loss modulus, indicative of viscous
friction, exhibits extremely low values with respect to thedilational modulus,ωκ , ε, as qualitatively expected from thenear-zero phase lag between the strain and the stress curves (seeFigure 4). The viscoelastic response, similar to the equilibriumelasticity described above, was found to be temperature-independent.
To summarize, the results show that Langmuir monolayers ofE. coliPLE exhibit a pure elastic, low-viscosity, and temperature-independent mechanical response, which could support enhancedelastic storage on compression and possibly also on shear. Thesepieces of experimental evidence could altogether be consideredas the dynamical fingerprint for a high-fluidity (lowκ), densebut soft (relatively lowε), thermoregulated system.
Discussion
The experiments described above had a twofold objective: ingeneral, to explore the microscopic mechanisms underlying thesurface rheology of complex lipid mixtures and in particular toget insight on the precise structure-material relationships ofE.coli membranes with the paramount perspective of providingreliable material functions for advanced modeling studies of themechanics of the bacterial cell wall.15,34,35The estimated lipidarea inE. coli native bilayer isA0 ∼ 75 Å2.36 In the monolayer,this lipid area corresponds to a lateral pressureΠ0 ∼ 30 mN/m,the state usually accepted for each monolayer leaflet in bilayermembranes.37 At this state, lipid domains are observed in therealE. coli membrane at in vivo conditions,11 similarly to othernative lipid extracts.26 Therefore, a monolayer state atA0 (∼75Å2) and atΠ0 has been chosen as a dense state representativeof the molecular packing existing at in vivo conditions. Ourexperimental results, however, indicate that the surface pressure
(33) It was carefully checked that at this regime the stress response is linearwell over the whole range of experimental conditions studied.
(34) Reshes, G.; Vanounou, S.; Fishov, I.; Feingold, M.Biophys. J. BioFAST2007 (doi:10.1529/biophysj.107.104398).
(35) Horger, I.; Velasco, E.; Mingorance, J.; Rivas, G.; Tarazona, P.; Ve´lez,M. Phys. ReV. E 2008, 77, 011902.
(36) Esfahani, M.; Limbrick, A. R.; Knutton, S.; Oka, T.; Wakil, S. J.Proc.Nat. Acad. Sci. U.S.A.1971, 68, 3180.
(37) Marsh, D.Biochim. Biophys. Acta1996, 1286, 183.
Figure 5. Frequency dependence of the dilational elasticity modulusεD (top) and the loss modulusωκ (bottom) as obtained from oscillatorybarrier experiments at three differentΠ values and at three different temperatures: (0) 15 °C, (O) 25 °C, and (∆) 37 °C. Top: dynamicdilational modulusεD(ω); dashed lines correspond to the equilibrium valuesεD obtained from theΠ-A isotherms (see data in Figure 3B).Bottom: dynamic loss modulusωκ, which is comparatively smaller than the elastic storage (ωκ/ε ,1), pointing out the high fluidity of theE. coli PLE monolayers in the probed dynamical scale.
4070 Langmuir, Vol. 24, No. 8, 2008 Lo´pez-Montero et al.
at which this lipid area ofA0 ∼ 75 Å2 is found different forisotherms obtained by either aliquot addition or continuouscompression. When the isotherm is obtained by continuousuniaxial compression (see Figure 1B), the biologically relevantlipid packing is reached at a higherΠ (∼35 mN/m> Π0) thanthe previously described value ofΠ0 ∼ 30 mN/m. Thus, furtheranalysis is required to explain these differences. Therefore, thediscussion has been organized in four different sections. First,the differences observed between the two types of isotherms arediscussed as a dichotomy between possible dynamical effectsleading to metastable states versus an equilibrium scenario wherethe shearing induced by the dilational compression provokesadditional energy storage. Second, the shear contribution to themonolayer elasticity is extracted as a separated contribution tothedilationalmodulusbycomparingwith thehydrostaticmodulus.This shear elasticity is discussed in view of the existence ofordered domains revealed by fluorescence microscopy. Third,the dynamical values of the dilational moduli (elasticity andviscosity) are discussed in terms of lateral lipid distribution andmolecular transport inside the monolayer. Fourth, densityregulation provided by lipid mosaicity has been claimed as aplausible explanation for the nondependence of the mechanicalproperties on temperature.
Differences between Aliquot-Addition and Continuous-Compression Isotherms: Equilibrium or Dynamics?Becauseany continuous compression imposed by the barriers of theLangmuir trough must be performed at a finite rate, dA/dt,dynamical effects could be induced on theΠ-A isotherm.Particularly, metastable states could be raised atΠ values higherthan at equilibrium conditions. These effects normally occur dueto the existence of stress relaxation within the monolayer. Insuch a case, the whole surface energy characterizing a givenstate,Π ) Πeq + δΠ, exceeds the equilibrium valueΠeq by anamountδΠ equivalent to nonrelaxed stresses,δΠ ) σk. Theseeffects have been revealed to be dramatically dominant in polymerLangmuir films with high viscosity, resulting in an extremelyslow stress relaxation.38 However, because local moleculartransport is comparatively fast for small surfactant molecules,such as lipids, viscous dissipation might be in this case onlysubsidiary, even vanishing.18 If differencesδΠ (*0) are due todynamical metastability effects, they might be governed bymonolayer viscosity and therefore strongly dependent oncompression rates. More precisely, if the simple response functionin eq 2 is obeyed, an extra, rate-dependent, viscous stress,σk )κ du/dt ) κ r, might be considered when compressing at a finiterate,r ) du/dt. Thus, if compression is fast enough with respectto the relaxation rate of the viscous stresses, the monolayer hasno time to dissipate stress and its internal energy is dependentonr; i.e.,Π(r) ) Πeq+ κr. Only at vanishing compression rates(quasistatic compression,r ∼ 0), or if κ ∼ 0, the dynamicaleffects on a compression isotherm will be negligible with respectto the one obtained at equilibrium conditions (aliquot additionmethod).
Figure 6 shows that, at the lowest compression rate experi-mentally accessible (r ) 10 cm2/min), differences with respectto the aliquot-addition isotherms start to emerge at surface areassmaller than 125 Å2. Different compression rates ranging from10 to 500 cm2/min (r ) 2× 10-4 to 10-2 s-1) have been checkedin order to test for the concurrence of nonrelaxed viscous stresses.Surprisingly, near identical isotherms are obtained independentlyof the velocity under which the monolayer is compressed. Thisis clear evidence that dynamical effects are absent and can
therefore not account for our experimental observations. On thecontrary, sinceδΠ is found to be independent of the compressionrate, the results in Figure 6 confirm a near zero viscosity, high-fluidity scenario. As a consequence, the so-reached compressedstates can be considered to be near-to-equilibrium, or in otherterms, becauseκ ∼ 0, the continuous compression of themonolayer, even if exerted at nonvanishing compression rates,is not a source of extra dynamical entropy in the system; dS/dt∼ d(ΠA)/dt ∼ κ dA/dt ∼ 0. We can therefore conclude that thehigh fluidity observed in the monolayers at the conditions studiedexcludes dynamical metastability as an explanation for theincrease in dilational elasticity modulus observed under continu-ous uniaxial compression. No metastability is in fact the necessaryconsequence of the reversibility observed in the compression-expansion cycles; i.e., if no hysteresis is present, it is becauseevery studied state is at thermodynamic equilibrium.
Emerging Shear Elasticity: Appearance of Ordered States.Since dynamic metastability cannot explain theδΠ data in Figure6, an equilibrium scenario has to be alternatively invoked. Thisfact could be related to the intrinsically different character of thelateral compression performed in each method. While thequasiequilibrium compression involved in the aliquot method ispurely hydrostatic, the uniaxial dilational compression methodadditionally involves a lateral shear. Shear effects are howeveronly plausible if there are ordered phases able to store elasticenergyupon lateral reorganization.Since fluorescencemicroscopyhas revealed an increasing number of small domains of a denserphase continuously nucleating from the fluid one, the emergenceof equilibrium shear effects can be then justified when domainsstarts to sustain permanent structural changes. In other words,since structural ordering is susceptible to be altered under a sheardistortion, an ordered monolayer reacts as a more rigid bodywhen strained by uniaxial forces than by hydrostatic ones. Thisis because emerging shear effects appear to be a reasonablehypothesis to explain theδΠ differences observed between theisotherms recorded from experiments at continuous uniaxialcompression and by the aliquot addition method. A first pieceof evidence supporting emergent shear arises from the existenceof a nonhydrostatic normal stressσyy different from the usualdilational stressσxx measured along the easy axis (which is inthe present case thex-axis parallel to the motion of the barriers).For a two-dimensional elastic body, the normal stress isproportional to the dilational stress,σyy) νσxx, the proportionalityconstantν being the Poisson ratio.39 For an isotropic andhomogeneous two-dimensional elastic body,ν ) (εK - εS)/(εK
(38) Monroy, F.; Hilles, H.; Ortega, F.; Rubio, R. G.Phys. ReV. Lett. 2003,91, 268302.
(39) Landau, L. D.; Lifshitz, E. M.Theory of Elasticity; Pergamon: Oxford,1986.
Figure 6. δΠ difference between monolayer states obtained bycontinuous compression and by the aliquot-addition method at25°C. TheΠ-A continuous compression isotherms were recordedat different compression rates. For clarity, onlyδΠ data at the lowest(r ) 10 cm2/min, solid line) and the highest compression rates (r) 500 cm2/min, dashed line) have been plotted.
Elasticity of E. coli Membranes Langmuir, Vol. 24, No. 8, 2008 4071
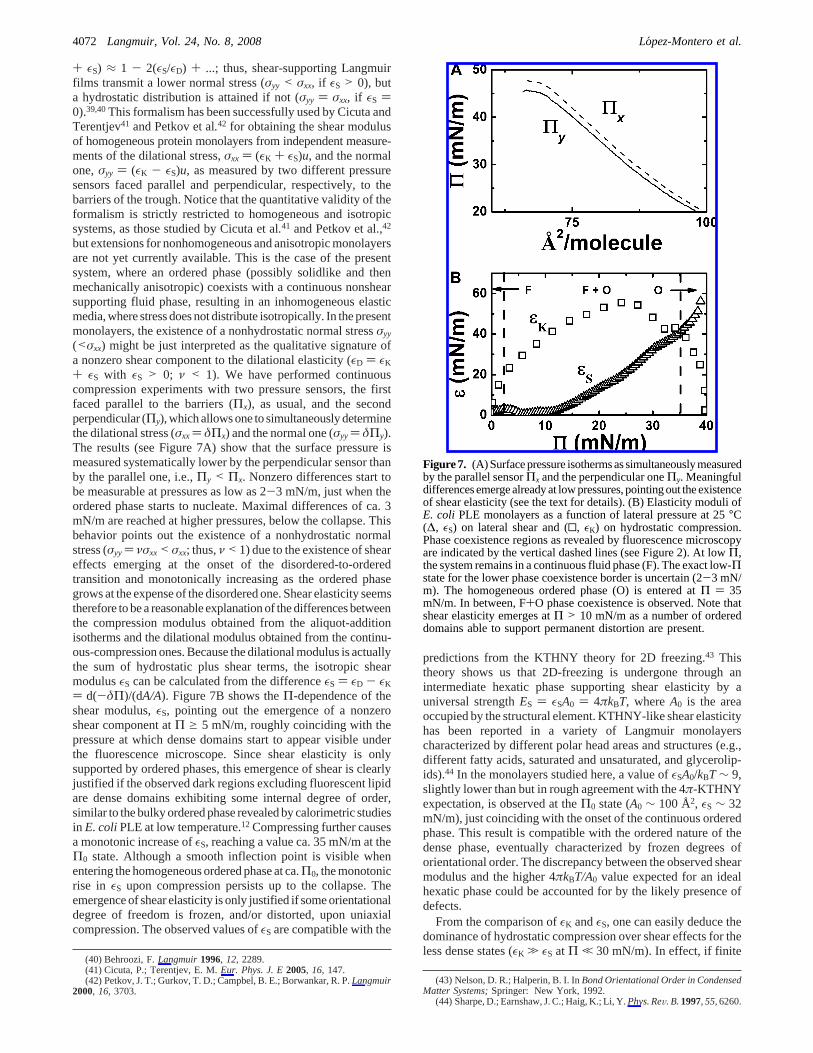
+ εS) ≈ 1 - 2(εS/εD) + ...; thus, shear-supporting Langmuirfilms transmit a lower normal stress (σyy < σxx, if εS > 0), buta hydrostatic distribution is attained if not (σyy ) σxx, if εS )0).39,40This formalism has been successfully used by Cicuta andTerentjev41 and Petkov et al.42 for obtaining the shear modulusof homogeneous protein monolayers from independent measure-ments of the dilational stress,σxx ) (εK + εS)u, and the normalone,σyy ) (εK - εS)u, as measured by two different pressuresensors faced parallel and perpendicular, respectively, to thebarriers of the trough. Notice that the quantitative validity of theformalism is strictly restricted to homogeneous and isotropicsystems, as those studied by Cicuta et al.41 and Petkov et al.,42
but extensions for nonhomogeneous and anisotropic monolayersare not yet currently available. This is the case of the presentsystem, where an ordered phase (possibly solidlike and thenmechanically anisotropic) coexists with a continuous nonshearsupporting fluid phase, resulting in an inhomogeneous elasticmedia, where stress does not distribute isotropically. In the presentmonolayers, the existence of a nonhydrostatic normal stressσyy
(<σxx) might be just interpreted as the qualitative signature ofa nonzero shear component to the dilational elasticity (εD ) εK
+ εS with εS > 0; ν < 1). We have performed continuouscompression experiments with two pressure sensors, the firstfaced parallel to the barriers (Πx), as usual, and the secondperpendicular (Πy), which allows one to simultaneously determinethe dilational stress (σxx ) δΠx) and the normal one (σyy ) δΠy).The results (see Figure 7A) show that the surface pressure ismeasured systematically lower by the perpendicular sensor thanby the parallel one, i.e.,Πy < Πx. Nonzero differences start tobe measurable at pressures as low as 2-3 mN/m, just when theordered phase starts to nucleate. Maximal differences of ca. 3mN/m are reached at higher pressures, below the collapse. Thisbehavior points out the existence of a nonhydrostatic normalstress (σyy) νσxx< σxx; thus,ν < 1) due to the existence of sheareffects emerging at the onset of the disordered-to-orderedtransition and monotonically increasing as the ordered phasegrows at the expense of the disordered one. Shear elasticity seemstherefore to be a reasonable explanation of the differences betweenthe compression modulus obtained from the aliquot-additionisotherms and the dilational modulus obtained from the continu-ous-compression ones. Because the dilational modulus is actuallythe sum of hydrostatic plus shear terms, the isotropic shearmodulusεS can be calculated from the differenceεS ) εD - εK
) d(-δΠ)/(dA/A). Figure 7B shows theΠ-dependence of theshear modulus,εS, pointing out the emergence of a nonzeroshear component atΠ g 5 mN/m, roughly coinciding with thepressure at which dense domains start to appear visible underthe fluorescence microscope. Since shear elasticity is onlysupported by ordered phases, this emergence of shear is clearlyjustified if the observed dark regions excluding fluorescent lipidare dense domains exhibiting some internal degree of order,similar to the bulky ordered phase revealed by calorimetric studiesin E. coliPLE at low temperature.12 Compressing further causesa monotonic increase ofεS, reaching a value ca. 35 mN/m at theΠ0 state. Although a smooth inflection point is visible whenentering the homogeneous ordered phase at ca.Π0, the monotonicrise in εS upon compression persists up to the collapse. Theemergence of shear elasticity is only justified if some orientationaldegree of freedom is frozen, and/or distorted, upon uniaxialcompression. The observed values ofεS are compatible with the
predictions from the KTHNY theory for 2D freezing.43 Thistheory shows us that 2D-freezing is undergone through anintermediate hexatic phase supporting shear elasticity by auniversal strengthES ) εSA0 ) 4πkBT, whereA0 is the areaoccupied by the structural element. KTHNY-like shear elasticityhas been reported in a variety of Langmuir monolayerscharacterized by different polar head areas and structures (e.g.,different fatty acids, saturated and unsaturated, and glycerolip-ids).44 In the monolayers studied here, a value ofεSA0/kBT ∼ 9,slightly lower than but in rough agreement with the 4π-KTHNYexpectation, is observed at theΠ0 state (A0 ∼ 100 Å2, εS ∼ 32mN/m), just coinciding with the onset of the continuous orderedphase. This result is compatible with the ordered nature of thedense phase, eventually characterized by frozen degrees oforientational order. The discrepancy between the observed shearmodulus and the higher 4πkBT/A0 value expected for an idealhexatic phase could be accounted for by the likely presence ofdefects.
From the comparison ofεK andεS, one can easily deduce thedominance of hydrostatic compression over shear effects for theless dense states (εK . εS at Π , 30 mN/m). In effect, if finite
(40) Behroozi, F.Langmuir1996, 12, 2289.(41) Cicuta, P.; Terentjev, E. M.Eur. Phys. J. E2005, 16, 147.(42) Petkov, J. T.; Gurkov, T. D.; Campbel, B. E.; Borwankar, R. P.Langmuir
2000, 16, 3703.(43) Nelson, D. R.; Halperin, B. I. InBond Orientational Order in Condensed
Matter Systems;Springer: New York, 1992.(44) Sharpe, D.; Earnshaw, J. C.; Haig, K.; Li, Y.Phys. ReV. B.1997, 55, 6260.
Figure7. (A)Surfacepressure isothermsassimultaneouslymeasuredby the parallel sensorΠx and the perpendicular oneΠy. Meaningfuldifferences emerge already at low pressures, pointing out the existenceof shear elasticity (see the text for details). (B) Elasticity moduli ofE. coli PLE monolayers as a function of lateral pressure at 25°C(∆, εS) on lateral shear and (0, εK) on hydrostatic compression.Phase coexistence regions as revealed by fluorescence microscopyare indicated by the vertical dashed lines (see Figure 2). At lowΠ,the system remains in a continuous fluid phase (F). The exact low-Πstate for the lower phase coexistence border is uncertain (2-3 mN/m). The homogeneous ordered phase (O) is entered atΠ ) 35mN/m. In between, F+O phase coexistence is observed. Note thatshear elasticity emerges atΠ > 10 mN/m as a number of ordereddomains able to support permanent distortion are present.
4072 Langmuir, Vol. 24, No. 8, 2008 Lo´pez-Montero et al.

values ofεS are assumed to exclusively arise from the presenceof ordered domains, the macroscopic response of the monolayermight be dominated by the high compliance of the continuousfluid phase but not by the stiffer grains. The absolute values ofthe compression modulusεK are lower than those characteristicof rigid monolayers at the liquid ordered or solid state, such uslipid mixtures including saturated lipids or cholesterol,24,45,46butsimilar to those of unsaturated phospholipids.24 The values ofboth moduli become comparable for the denser ordered phase,in particular,εK ∼ 1.5εS at Π ∼ Π0 ∼ 30 mN/m andεK ∼ εS
atΠ ∼ 35 mN/m, well inside the ordered phase. At the collapseof the monolayer,Π ∼ 40 mN/m, hydrostatic compression is notsupported anymore (εK ∼ 0), as expected, butεS remains finite(∼50 mN/m) since area is conserved upon shear. To conclude,one might expect a realE. colimembrane to behave as a relativelysoft (rather elastic) body able to support stress on lateralcompression but also on lateral shear, this consideration beingrelevant for further analysis of the mechanics of the constrictionprocess.
Membrane Dynamics: Lateral Lipid-Transport Drivenby Surface Diffusion.The experimental results in Figure 5 pointout a dynamical behavior characterized by a time-independentelastic storage of the fluid phase compatible with the equilibriumdilational compressibility (Figure 3A,B). However, higher valuesof the elasticity modulus are systematically observed for thedenser states as the deformation is stressed at higher dilationrates (see data atΠ ) 30 mN/m in Figure 5 at the higherωvalues). Hypothetically, thedynamical stiffeningobserved at theΠ0 state, mainly dominated by the presence of coexistencedomains, could be the elastic signature of the rigid grains of theordered phase. These domains could hinder relaxations at fastdeformation rates. In effect, composite materials made of rigidgrains dispersed in a compliant matrix behave stiffly under rapidshocks but elastic at quasistatic deformation conditions.47 Thisis because transport develops differently over different time scales,resulting in stress relaxation when the material is deformed.Getting insight on the possible relaxation effects requires a furtherdiscussion of lipid transport after a mechanical disturbance.Indeed, as lipid molecules can move inside the monolayer, thediffusive transport can modulate the viscoelastic response upona surface deformation strained at a given rate. In the simplestcase, if surface diffusion is considered the only mechanism forlipid transport operating under a sinusoidal deformation of themonolayer,u(t) ) ux
(0)eiωt, the frequency dependence of the linearviscoelastic parameters follows a Maxwell-like relaxation48
wherex ) ωτD, a reduced frequency.The viscoelastic relaxation in eq 10 is described by two
parameters: the instantaneous (high-frequency) limit of thedilational modulusε0 and a diffusive characteristic timeτD,determined by the ratio of the quadratic distance over whichdiffusion operates and the lateral diffusion coefficientD; i.e.,τD
∼ a2/D. Two extreme cases are easy to understand: when the
frequency of the sinusoidal dilatation is low, the monolayer hasenough time to transport the lipid molecules from the compressionsemiwave to the expanded zones; i.e., there is no elastic resistanceto compression and the injected energy is fully dissipated byviscous friction; thus,ε ) 0 andκ ) ε0τD. Conversely, when thefrequency is high (ω f ∞), the deformation is very fast and themonolayer has no time to respond, behaving as purely elasticand fluid: ε ) ε0 andκ ) 0. In other words, for fast deformationsoccurring at timest , τD diffusion is not able to operate and thesystem reacts as a pure elastic body. At the opposite regime,t. τD, a slow deformation allows for diffusion to work, resultingin a very compliant system under the external stimulus, dissipatingmost energy by viscous friction. Both regimes define the limitsof the stress relaxation driven by surface diffusion.
As stated above, the characteristic time of this relaxationdepends on the diffusion coefficient and on the spatial scaleaon which it operates, more preciselyτD ∼ a2/D.48 The spatialscale involved in an oscillatory barrier experiment is associatedwith the concentration longitudinal wave, or Lucassen wave,driven by surface tension gradients (Marangoni gradients) inducedby the external deformation. The propagation frequency of theso-created Lucassen wave disperses asω ) (x3/2)(ε2q4/ηF)1/3
(q being the wavevector of the longitudinal wave,F the density,andη the viscosity of the subphase on which the monolayer issupported).49 Thus, stiffer monolayers develop longer concentra-tion waves; at Langmuir monolayers (F ∼ 1 g/cm3; η ∼ 1 cP)the Lucassen wavelength isλ ()2π/q) ∼ 17.8 ε1/2/ω3/4. Moreprecisely, at the highly elastic diluted stateΠ ∼ 5 mN/m,εD ∼10 mN/m; thus,λmin ∼ 80 cm for the highest frequencies studied.At the more rigid state (Π ∼ 30 mN/m,εD ∼ 90 mN/m), a similarestimation,λmin ∼ 120 cm, is obtained for the highest probedfrequencyω ∼ 1 Hz, but it becomes much larger,λmax ∼ 6 ×104cm, for the slowest waves atω ∼ 1 mHz. In other words, theconcentration waves produced in our experiments exhibit largerwavelengths than the trough dimensions. These estimations havetwo important consequences: (a) The instantaneous densityprofile along thex-axis of the trough is near homogeneous; i.e.,the strain function distributes homogenously along the monolayer;thus, measurements of stress are independent of the relativeposition of theΠ sensor with respect to the barriers.22 (b) Theconcentration gradient created by the dilational wave has amacroscopic dimension. The first conclusion is important froma methodological point of view, but the last one is crucial forunderstanding the viscoelastic relaxation in Langmuir lipidmonolayers. Any concentration gradient relaxation due to laterallipid diffusion would take times longer than the characteristicdiffusive time, and the macroscopic concentration gradient createdby the dilational wave would take much longer time to relax thanthe characteristic periods of our experiments. For realistic valuesof D ∼ 10-8-10-7 cm2/s, the characteristic diffusive time fora macroscopic concentration gradient extending a distancea ∼λ ∼ 1 m is on the order ofτD ∼ a2/D ∼ 107-108s, longer byseveral orders of magnitude than the period of the deformation.In other words, the macroscopic concentration wave created bythe external deformation has no time to relax by lipid diffusionmechanism. The characteristic time of the observed relaxationis on the order ofτ ∼ 2π/ω ∼ 10s. If it were controlled bydiffusion (t ∼ a2/D), its characteristic spatial scale would be asshort asa ∼ (Dτ)1/2 ∼ 1-10µm. This estimation is surprisinglysimilar to the characteristic size of the lipid condensed domainsobserved by fluorescence microscopy. At higher frequencies (∼1Hz; see Figure 5,Π ) 30 mN/m, top) this relaxation mechanismmight apply to explain the observed increase in the dilational
(45) Li, J. B.; Kretzschmar, G.; Miller, R.; Mo¨hwald, H.Colloids Surf. A1999,149, 491.
(46) Stottrup, B.; Stevens, D. S.; Keller, S. L.Biophys. J.2005, 88, 269.(47) Ferry, J. D.Viscoelastic Properties of Polymers; Wiley: New York,
1980.(48) Monroy, F.; Khan, J.; Langevin, D.Colloids Surf. A1998, 143, 251. (49) Lucassen, J.; van den Tempel, M.Chem. Eng. Sci.1972, 27, 1283.
ε(ω) ) ε0x2
1 + x2
ωκ(ω) ) ε0x
1 + x2(10)
Elasticity of E. coli Membranes Langmuir, Vol. 24, No. 8, 2008 4073
modulus. We indeed suggest lipid domains as concentratedreservoirs of lipids actively contributing to the mechanicalresponse of the monolayer. Upon dilation, lipid domains workdispensing lipids to their surroundings, and conversely, newdomains can be nucleated from the continuous fluid phase oncompression. This domain-mediated mechanism allows for themacroscopic re-equilibration of the stressed monolayer. Mac-roscale lipid transport by diffusion is too slow to account for therapid lipid motions required to explain the fast equilibriumrestorations observed in our experiments.
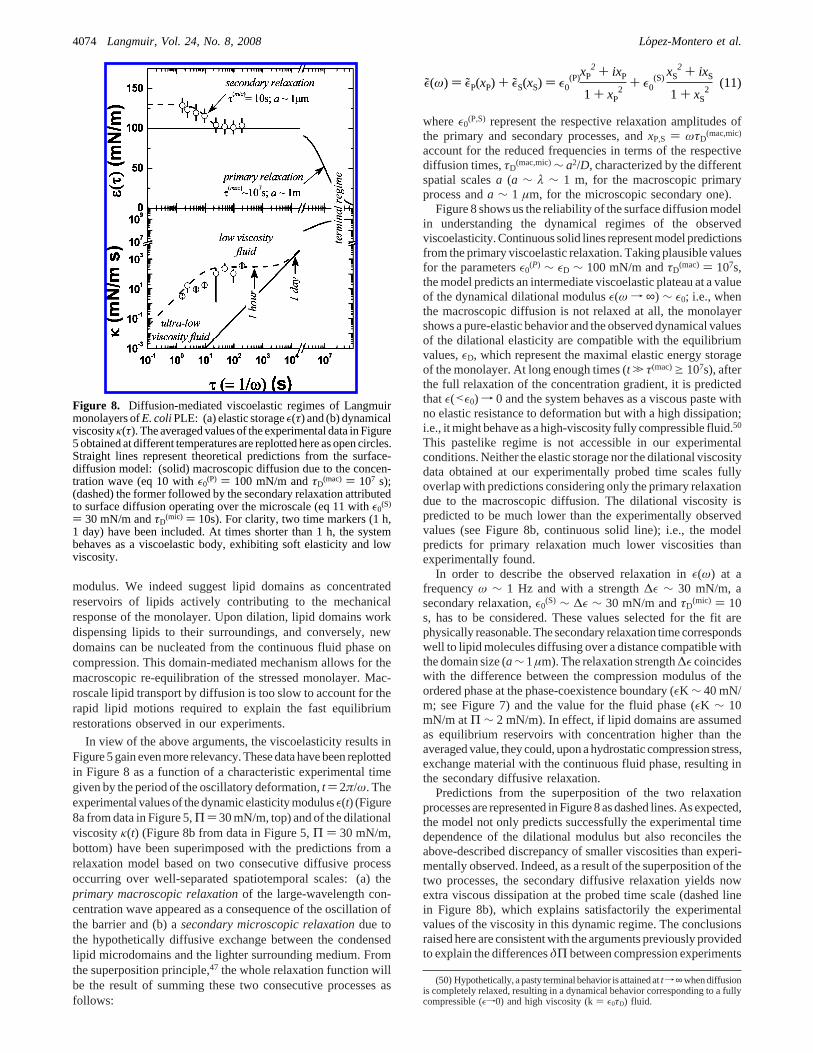
In view of the above arguments, the viscoelasticity results inFigure 5 gain even more relevancy. These data have been replottedin Figure 8 as a function of a characteristic experimental timegiven by the period of the oscillatory deformation,t ) 2π/ω. Theexperimental values of the dynamic elasticity modulusε(t) (Figure8a from data in Figure 5,Π ) 30 mN/m, top) and of the dilationalviscosityκ(t) (Figure 8b from data in Figure 5,Π ) 30 mN/m,bottom) have been superimposed with the predictions from arelaxation model based on two consecutive diffusive processoccurring over well-separated spatiotemporal scales: (a) theprimary macroscopic relaxationof the large-wavelength con-centration wave appeared as a consequence of the oscillation ofthe barrier and (b) asecondary microscopic relaxationdue tothe hypothetically diffusive exchange between the condensedlipid microdomains and the lighter surrounding medium. Fromthe superposition principle,47 the whole relaxation function willbe the result of summing these two consecutive processes asfollows:
whereε0(P,S) represent the respective relaxation amplitudes of
the primary and secondary processes, andxP,S ) ωτD(mac,mic)
account for the reduced frequencies in terms of the respectivediffusion times,τD
(mac,mic)∼ a2/D, characterized by the differentspatial scalesa (a ∼ λ ∼ 1 m, for the macroscopic primaryprocess anda ∼ 1 µm, for the microscopic secondary one).
Figure 8 shows us the reliability of the surface diffusion modelin understanding the dynamical regimes of the observedviscoelasticity. Continuous solid lines represent model predictionsfrom the primary viscoelastic relaxation. Taking plausible valuesfor the parametersε0
(P) ∼ εD ∼ 100 mN/m andτD(mac) ) 107s,
the model predicts an intermediate viscoelastic plateau at a valueof the dynamical dilational modulusε(ω f ∞) ∼ ε0; i.e., whenthe macroscopic diffusion is not relaxed at all, the monolayershows a pure-elastic behavior and the observed dynamical valuesof the dilational elasticity are compatible with the equilibriumvalues,εD, which represent the maximal elastic energy storageof the monolayer. At long enough times (t . τ(mac)g 107s), afterthe full relaxation of the concentration gradient, it is predictedthatε(<ε0) f 0 and the system behaves as a viscous paste withno elastic resistance to deformation but with a high dissipation;i.e., it might behave as a high-viscosity fully compressible fluid.50
This pastelike regime is not accessible in our experimentalconditions. Neither the elastic storage nor the dilational viscositydata obtained at our experimentally probed time scales fullyoverlap with predictions considering only the primary relaxationdue to the macroscopic diffusion. The dilational viscosity ispredicted to be much lower than the experimentally observedvalues (see Figure 8b, continuous solid line); i.e., the modelpredicts for primary relaxation much lower viscosities thanexperimentally found.
In order to describe the observed relaxation inε(ω) at afrequencyω ∼ 1 Hz and with a strength∆ε ∼ 30 mN/m, asecondary relaxation,ε0
(S) ∼ ∆ε ∼ 30 mN/m andτD(mic) ) 10
s, has to be considered. These values selected for the fit arephysically reasonable. The secondary relaxation time correspondswell to lipid molecules diffusing over a distance compatible withthe domain size (a∼ 1µm). The relaxation strength∆ε coincideswith the difference between the compression modulus of theordered phase at the phase-coexistence boundary (εK ∼ 40 mN/m; see Figure 7) and the value for the fluid phase (εK ∼ 10mN/m atΠ ∼ 2 mN/m). In effect, if lipid domains are assumedas equilibrium reservoirs with concentration higher than theaveraged value, they could, upon a hydrostatic compression stress,exchange material with the continuous fluid phase, resulting inthe secondary diffusive relaxation.
Predictions from the superposition of the two relaxationprocesses are represented in Figure 8 as dashed lines. As expected,the model not only predicts successfully the experimental timedependence of the dilational modulus but also reconciles theabove-described discrepancy of smaller viscosities than experi-mentally observed. Indeed, as a result of the superposition of thetwo processes, the secondary diffusive relaxation yields nowextra viscous dissipation at the probed time scale (dashed linein Figure 8b), which explains satisfactorily the experimentalvalues of the viscosity in this dynamic regime. The conclusionsraised here are consistent with the arguments previously providedto explain the differencesδΠ between compression experiments
(50) Hypothetically, a pasty terminal behavior is attained att f ∞ when diffusionis completely relaxed, resulting in a dynamical behavior corresponding to a fullycompressible (εf0) and high viscosity (k) ε0τD) fluid.
Figure 8. Diffusion-mediated viscoelastic regimes of Langmuirmonolayers ofE. coliPLE: (a) elastic storageε(τ) and (b) dynamicalviscosityκ(τ). The averaged values of the experimental data in Figure5 obtained at different temperatures are replotted here as open circles.Straight lines represent theoretical predictions from the surface-diffusion model: (solid) macroscopic diffusion due to the concen-tration wave (eq 10 withε0
(P) ) 100 mN/m andτD(mac) ) 107 s);
(dashed) the former followed by the secondary relaxation attributedto surface diffusion operating over the microscale (eq 11 withε0
(S)
) 30 mN/m andτD(mic) ) 10s). For clarity, two time markers (1 h,
1 day) have been included. At times shorter than 1 h, the systembehaves as a viscoelastic body, exhibiting soft elasticity and lowviscosity.
ε(ω) ) εP(xP) + εS(xS) ) ε0(P)xP
2 + ixP
1 + xP2
+ ε0(S)xS
2 + ixS
1 + xS2
(11)
4074 Langmuir, Vol. 24, No. 8, 2008 Lo´pez-Montero et al.

performed at different rates (see Figure 6). In effect, the (low)dilational viscosities in Figure 8b are compatible with a weaklydissipative scenario. If values ofδΠ ∼ 5 mN/m observed at theΠ ) 30 mN/m state were attributed to viscous stresses inducedby the lateral compressionδΠ ∼ κ du/dt, the dilational lossmodulus would be required to assume unphysical high valuesof ωκ ∼ ωδΠ/(du/dt) ∼ 500 mN/m at the minimum, in completedisagreement with the small values observed in the oscillatorybarrier experiments (ωκ , ε; see Figure 5B). It is thus reasonableto consider that it is the emergence of shear effects that underliesthe observedδΠ data.
As a general conclusion, Figure 8b shows us that the viscosityof the monolayer is strongly dependent on the time length of thedeformation. In the bacteria, the constriction process is acontinuous deformation taking less than 1 h, thusE. colimembranes work in vivo in afluid regimesimilar to that probedin our experiments; thus, constriction is exerted at an efficientregime characterized by small viscous losses.
Equilibrium and Dynamical Athermalicity: A PassiveThermal Regulation Mechanism?The near-athermal behaviorof the experimentalΠ-A isotherms teaches us that monolayerstability is largely dominated by strong adsorption interactions(∆Hads∼ -ΠA). However, the whole entropy remains essentiallyequal to that of the bare aqueous surface (∆Sads∼ -dΠ/dT ∼0), even iforderedstates are present, which is a consequenceof the counterbalance between the losses of conformationalentropy intrinsically related to organization of the monolayerand the disruption of the surface structure of water at the levelof the polar heads.18 In the present case, adsorption forces arelarge compared to thermal energy,-∆Hads ) ΠA . kBT, andessentially independent ofT. If the weakT-dependence of theΠ-A isotherms is assumed to be exclusively entropic,Π(T) ∼kBT, typical changes in surface pressureδΠ/Π ) δT/T areexpected to be on the order of a 0.3% per degree. Thus, theexpected entropy-driven changes in surface pressure along thestudiedT range (δT ∼ 22 °C) will be on the order ofδΠ ∼ 0.1mN/m atΠ ∼ 1 mN/m and as weak asδΠ ∼ 0.002 mN/m ata pressureΠ ∼ 30 mN/m, which are below the experimentaluncertainty and in close agreement with the experimentalobservation. In effect, because dΠ/dT ∼ 0, the surface entropyof the monolayer,s) s0 - dΠ/dT, equals that of the bare surface,s0 ) dγ0/dT (∼0.2 erg/cm2 K for water); thus, its absolute valueat the coexistence region might take a typical value,S) As)As0 ∼ kB, as supposed. Therefore, one could conclude thatadsorption is only driven by lipid-water and lipid-lipidinteractions but not by a decrease in entropy,∆Gads≡ - ΠA )∆Hads- T∆Sads∼ ∆Hads; otherwise,Π might display a strongT-dependence.
This equilibrium athermalicity is rather universal in lipidsystems, being of a transcendental importance for understandingthe ability of most lipids for self-assembling lipid bilayer structuresin aqueous media.51 When organized at hydrophobic interfaces,as the air/water interface of Langmuir monolayers or the oil/water interface typical of bilayers, an entropic penalty might bepaid against the energetic stability achieved by cohesiveinteractions, which are indeed the main driving forces for colloidalself-assembling.51 In the present case, thermodynamics showsus that cohesive contributions are largely dominating over entropy,which remains essentially unchanged (∆Hads . T∆Sads ∼ 0).These cohesion properties ofE. coli lipids yield the adequatethermodynamic scenario when discussed in terms of self-assembling of the lipid membrane. In effect, since∆Gads∼ ∆Hads
, 0, lipid self-assembling will be strongly spontaneous and thestructures formed, the lipid bilayer for instance, will result in alarge robustness because of the extremely low critical aggregationconcentration (log cac∼ ∆Gads/kBT). As a consequence, and thisis an essential requirement for biofunctionality,51the membranesremain intact under thermal fluctuations or even when the lipidswere strongly depleted from the medium.
Viscoelastic properties have been also found to be essentiallyindependent of temperature. This dynamic athermalicity contrastswith simpler lipid systems,52 where the temperature dependenceof the transport properties is governed by high activation energiesattributed to the high energy required to hop a molecule into anavailable free volume.53 In the present case, the formula of thelipid cocktail seems to have been optimized to allow a tightregulation of the mutual interactions in a way such that thisenergy penalty is reduced, resulting in transport propertiesessentially independent of the temperature. This unresponsive-to-temperature behavior could be related to the ability ofE. colimembranes to maintain fluidity and functionality across a broadtemperature range. The presence of domains could be plausiblyhypothesized to play a key role in the thermoregulationmechanism. In effect, relatively large density or concentrationdisturbances produced by a temperature change can be easilyassimilated and later resumed if domains are assumed to workas local reservoirs of condensed lipid material able to receiveinside or to deliver outside lipids upon request. As a consequenceof this thermal delivering mechanism, both equilibrium anddynamical properties could remain unaltered when macroscopi-cally averaged, resulting in a reliable mechanism for thermalregulation. The existence of a viscous plateau in Figure 8 (probablyrelated to the diffusive exchange with the microdomains) appearsto be essential to understand a dynamics unresponsive totemperature. In effect, time-temperature superposition principleteaches that despite the dependence of the molecular diffusionon temperatureE. colimembranes could maintain a temperature-invariant viscoelastic behavior if working at the plateau regimedescribed in Figure 8. However, much more work is needed inorder to verify the likelihood of this hypothesis.
ConclusionsThe monolayer model ofE. colistudied here displays dilational
elasticity that, at the biologically relevant packing representedby theΠ0 ∼ 30 mN/m state, is comparable to bilayer models ofsingle fluidlike lipid membranes, e.g., DMPC, POPC, EPC.54-56
Monolayers of theE. coli PLE behave like a very soft materialas compared to those based on eukaryote lipids.57,58 This softelasticity would support not only compression but also sheardeformations. Lipid domains of the ordered phase had beenconceived as the structural element to support elastic shear. Thedilational viscosity ofE. coli PLE monolayers has been foundlower than in other pasty materials, such as polymers.22 Moresignificantly, the viscous losses of this material are negligibleas compared to their own elasticity (ωκ , ε). Since the studiedPLE contains the main lipid components ofE. coli membranes,the present results could be relevant to understand the mechanicalbehavior of the real membrane under deformation. In theperspective of the division process ofE. coli, the presentmonolayer model retains those mechanical properties that one
(51) Israelachvili, J.Intermolecular and Surface Forces, 2nd ed; AcademicPress: London, 1992.
(52) Cicuta, P.; Keller, S. L.; Veatch, S. L.J. Phys. Chem. B2007, 111, 3328.(53) Almeida, P. F. F.; Vaz, W. L. C.; Thompson, T. E.Biochemistry1992,
31, 6739.(54) Needham, D.; McIntosh, T. J.; Evans, E.Biochemistry1988, 27, 4668.(55) McIntosh, T. J.; Magid, A. D.; Simon, S. A.Biochemistry 1989, 28, 17.(56) Kwok, R. E. E.Biophys. J.1981, 35, 637.(57) Evans, E. A.; Waugh, R.; Melnik, L.Biophys. J. 1976, 16, 5855.(58) Evans, E. A.; Waugh, R.Biophys. J.1977, 20, 307.
Elasticity of E. coli Membranes Langmuir, Vol. 24, No. 8, 2008 4075

would expect to be selected: it is essentially fluid and easilystretchable. These features are likely to renderE. colimembranesa soft mechanical behavior adequate for easy deformation underthe action of the division machinery and well-optimized todissipate a minimum amount of energy. The linear viscoelasticitydata reported here could be used in reliable material relationshipsfor the mechanical behavior ofE. colimembranes, and we hopethat such data will help studies of the division process fromcontinuous mechanics approaches.
Acknowledgment. This research strongly benefited from thecollaborative networking action from NANOBIO-M (http://
www.nanobiom.org) and COMBACT (www.cnb.csic.es/∼com-bact/combact). I.L.-M. thanks CAM for financial support fromNANOBIO-M. This work was financed by MEC under grantsFIS2006-01305 and BFU2005-04087-C02 and by CAM S-0505/MAT/0283. We thank R.G. Rubio and M. Vicente for their fruitfulcomments and J.E.F. Rubio and F. Ortega for technical assistancefrom the “Unidad IR-Raman-Correlacio´n” of the UCM-facilityCAI-Espectroscopia. F.M. gratefully acknowledges CAI-UCM“Centro de Microscopı´a y Citometrı´a” for allocating theinfrastructure for the experimental techniques.
LA703350S
4076 Langmuir, Vol. 24, No. 8, 2008 Lo´pez-Montero et al.




















