
High energy coralgal-stromatolite frameworks from Holocene reefs (Tahiti, French Polynesia
22
Srdimentology (1994) 41, 655-676 High energy coralgal-stromatolite frameworks from Holocene reefs (Tahiti, French Polynesia) GILBERT F. CAMOIN* and LUCIEN F. MONTAGGIONI*t * URA no. I208 du Centre National de la Recherche Scientifique, Centre de SPdimentologielPalPontolgie, Universitt! de Provence, 3 Place V. Hugo, I3331 Marseille cPdex 3, France t Centre de I’Environnement dOpunohu, Ecole Pratique des Hautes Etudes, Moorea, French Polynesia ABSTRACT Drill cores from Holocene reefs on Tahiti (French Polynesia) reveal a framework composed of massive branching acroporids encrusted by coralline algae associated with sessile vermetid gastropods and arborescent foraminifers. Laminated micritic crusts form coatings over coral branches or, more com- monly, over related encrusting organisms throughout the cored reef sections; these crusts appear as a major structural and volumetric component of the reef framework. The microbial nature of these micritic crusts is inferred from their typical organic growth forms and geometry, the occurrence of microbial remains and stable isotope measurements. The reef communities accumulated at depths less than 5 m below mean sea level in a high energy environment throughout vertical growth from 7140 f 170 yr BP to the present. The nature of the involved benthic communities, stable isotope data and high calcification rates of microbiaily encrusted corals strongly suggest that local environmental conditions have been optimal for reef development for the last 7000 years, The causes of the predominance of microbial communities over actual encrusters (red algae, foraminifers) remain problematic and could be related to short term fluctuations in ecological parameters. Microbial micritic crusts seemingly played a prominent role in protecting the coralgal colonies from bioeroders and grazers and, possibly, in strengthening the framework, due to rapid lithification. The record of similar microbial crusts in other Quaternary reef tracts suggests that microbial communities may have played a more prominent role in Quaternary reefs than presently recognized. INTRODUCTION Stromatolites are laminated organosedimentary structures, mainly built by microbes, through trap- ping, binding and precipitation processes. In Precambrian time, they formed prominent reef-like structures (Schopf, 1983), although a marked decline occurred in the late Proterozoic, coinciding with the appearance of skeletal organisms. In early Palaeozoic reef tracts, subtidal stromatolites were associated with sponges, metazoans and algal reefs. The propor- tion of stromatolites decreased sharply in late Palaeozoic and Mesozoic times with the rise of the newly evolving reef-forming organisms (sponges, corals and then rudists), which seemingly competed for substrate space in the subtidal habitat (Monty, 1973). However, their significance in Phanerozoic reefs has probably been underestimated (Pratt, 1982). Although it was thought that post-Cretaceous stro- matolites were restricted to marginal environments, especially lacustrine (Monty, 1973) and tidal flat (Riding, 1991) settings, abundant stromatolitic crusts overgrow corals in Miocene reefs (Pratt, 1982; Riding et a/., 1991a; G. F. Camoin, work in progress). Over the past three decades, modern stromatolites have been observed in various environments, ranging from hypersaline to freshwater, and from tropical to polar (Golubic, 1991), including high energy marine set- tings (Dravis, 1983; Dill et al., 1986; Shinn, 1987). Laminated Mg-calcite peloidal crusts have been described in Quaternary reef tracts, mainly from the walls of the deeper forereef from the Caribbean and the Red Sea (Ginsburg & James, 1973; Moore et al., 1976; James & Ginsburg, 1979; Land & Moore, 1980; 655
Transcript of High energy coralgal-stromatolite frameworks from Holocene reefs (Tahiti, French Polynesia

Srdimentology (1994) 41, 655-676
High energy coralgal-stromatolite frameworks from Holocene reefs (Tahiti, French Polynesia)
GILBERT F. CAMOIN* and LUCIEN F . MONTAGGIONI* t
* URA no. I208 du Centre National de la Recherche Scientifique, Centre de SPdimentologielPalPontolgie, Universitt! de Provence, 3 Place V. Hugo, I3331 Marseille cPdex 3, France
t Centre de I’Environnement dOpunohu, Ecole Pratique des Hautes Etudes, Moorea, French Polynesia
ABSTRACT
Drill cores from Holocene reefs on Tahiti (French Polynesia) reveal a framework composed of massive branching acroporids encrusted by coralline algae associated with sessile vermetid gastropods and arborescent foraminifers. Laminated micritic crusts form coatings over coral branches or, more com- monly, over related encrusting organisms throughout the cored reef sections; these crusts appear as a major structural and volumetric component of the reef framework. The microbial nature of these micritic crusts is inferred from their typical organic growth forms and geometry, the occurrence of microbial remains and stable isotope measurements. The reef communities accumulated at depths less than 5 m below mean sea level in a high energy environment throughout vertical growth from 7140 f 170 yr BP to the present. The nature of the involved benthic communities, stable isotope data and high calcification rates of microbiaily encrusted corals strongly suggest that local environmental conditions have been optimal for reef development for the last 7000 years, The causes of the predominance of microbial communities over actual encrusters (red algae, foraminifers) remain problematic and could be related to short term fluctuations in ecological parameters. Microbial micritic crusts seemingly played a prominent role in protecting the coralgal colonies from bioeroders and grazers and, possibly, in strengthening the framework, due to rapid lithification. The record of similar microbial crusts in other Quaternary reef tracts suggests that microbial communities may have played a more prominent role in Quaternary reefs than presently recognized.
INTRODUCTION
Stromatolites are laminated organosedimentary structures, mainly built by microbes, through trap- ping, binding and precipitation processes. In Precambrian time, they formed prominent reef-like structures (Schopf, 1983), although a marked decline occurred in the late Proterozoic, coinciding with the appearance of skeletal organisms. In early Palaeozoic reef tracts, subtidal stromatolites were associated with sponges, metazoans and algal reefs. The propor- tion of stromatolites decreased sharply in late Palaeozoic and Mesozoic times with the rise of the newly evolving reef-forming organisms (sponges, corals and then rudists), which seemingly competed for substrate space in the subtidal habitat (Monty, 1973). However, their significance in Phanerozoic reefs has probably been underestimated (Pratt, 1982).
Although it was thought that post-Cretaceous stro- matolites were restricted to marginal environments, especially lacustrine (Monty, 1973) and tidal flat (Riding, 1991) settings, abundant stromatolitic crusts overgrow corals in Miocene reefs (Pratt, 1982; Riding et a/., 1991a; G. F. Camoin, work in progress). Over the past three decades, modern stromatolites have been observed in various environments, ranging from hypersaline to freshwater, and from tropical to polar (Golubic, 1991), including high energy marine set- tings (Dravis, 1983; Dill et al., 1986; Shinn, 1987).
Laminated Mg-calcite peloidal crusts have been described in Quaternary reef tracts, mainly from the walls of the deeper forereef from the Caribbean and the Red Sea (Ginsburg & James, 1973; Moore et al., 1976; James & Ginsburg, 1979; Land & Moore, 1980;
655

656 G. F. Camoin and L. E Montaggioni
Brachert & Dullo, 1991), and in lagoonal to intertidal settings from the Caribbean (Jones & Hunter, 1991; Reid & Brown, 1991; Rasmussen et al., 1993). Marshall (1983) described lithified Mg-calcite crusts overgrowing corals and associated encrusting organ- isms in a Holocene section from the Great Barrier Reef.
In this paper we describe laminar to columnar micritic crusts capping or embedding high energy coralgal communities within a Holocene barrier reef from Tahiti (French Polynesia). These crusts are a major structural and volumetric constituent of the reef framework and seemingly played a significant sedimentological role in strengthening the reef (Montaggioni & Camoin, 1993).
METHODS
Three boreholes were drilled on the barrier reef complex facing Papeete City, one through the outer barrier reef flat (S,D,; depth 15,2m), about 30m behind the reef front, one on the south-western margin of the larger patch reef and one on the northern margin of the smaller patch reef (PA D,, PPT D,; depths 24 m and 20 m, respectively; Fig. la). None of the cores penetrated the entire reef sequence (Fig. 2). Cores were recovered using a porrable, hydraulically operated drilling rig which enables high recovery of 75 mm diameter cores in poorly consoli- dated sediments. Recovery was dependent upon framework type and on the size of internal cavities, but generally ranged from 50 to 95%; sections with poor recovery correspond to sandy beds. During drilling operations, the tube barrel was advanced every 1.5 m, so that we are confident that our core depths are accurate to & 0.3 m.
Microprobe analysis was performed at the Lab- oratoire de Microanalyse Electronique, Universite Paris-Sud Orsay. Polished thin sections of stroma- tolitic crusts were analysed by double wavelength dispersion spectrometry using a CAMEBAX microprobe analyser for quantitative determination of major, minor and trace elements and element mapping.
Radiocarbon ages were provided by the Centre des Faibles Radioactivities CNRS-CEA (Gif-sur- Yvette), the Laboratoire de Geologie du Quaternaire CNRS (Marseille) and the Laboratoire de Geochimie Isotopique (Universite Paris-sud-Orsay).
Stable isotope analyses (carbon and oxygen) on samples of corals and stromatolites were performed
I I
-REEFS
PREVAILING SWELLS
@
, 300 ~
4 4 PREVAILING WAVE DIRECTION
D R I L L I N G SITES
Fig. 1. Location map of the study area and drilling sites, on the outer barrier ~ reef flat (S,D,) and the two reef patches (PA D,; PPT D,) facing Papeete harbour, Tahiti.
at the Laboratoire de Geochimie Isotopique (Univer- site Paris-sud-Orsay) and at the Centre d’Analyses Isotopiques, Universite Claude Bernard (Lyon), using a FINNEGAN MAT 251 mass spectrometer coupled with a standard FINNEGAN ‘Bremen Carbonate Device’. This system allows automatic

Coralgal-stromatolite, Tahiti 657
s1 D1
8 7 0
PA D3
-I 6 950
PPT D5 - - REEF SURFACE
k =stromatol itic
crusts
detritus
-I C -14 ages
Fig. 2. Lithology and radiocarbon ages (yr BP) of drilled reef sequence. Depths are given to the reef top, situated at about 0.20 m below mean sea level. Reef framework is composed of scleractinian corals (mainly branching Acropora rohusta), associated with encrusting coralline algae, foraminifers and vermetid gastropods.
processing of the samples to be analysed with an individual acid bath. Reproducibility is 04)7%0 for 6"O and 0.05%0 for 6I3C. The 6"O and 6I3C isotopic compositions are expressed as:
6"O = [(180/'60) sample/ ('xO/160 standard- ')I x lo',
613C=[('3C/12C) ~ample/('~C/'*C standard- ')I x lo3.
The results are reported in the conventional delta notation, in per mil, relative to the Chicago PDB isotopic standard.
After collection, the branches of acroporid corals were cut in half in order to extract their growth axis. Each branch was split into subsamples; the relevant
isotopic composition therefore reflects at least a 3 week record. Such homogenization reduces the iso- topic variability that may result from pinpoint sub- sampling. After subsampling, the powder obtained from corals was treated for 20min in an oven at 400"C, under partial vacuum. In order to verify the isotopic effect of roasting, the composition of some subsamples was compared before and after heat- ing. Generally, there is no noticeable difference, indicating that organic matter was almost totally destroyed.
Stromatolites were sampled using a miniature lathe fitted with a 0.6 mm diameter dental drill; the drilling routine was observed under a binocular microscope.

658 G. F Camoin and L. F. Montaggioni
GENERAL SETTING
Study area and prevailing physical conditions The drilled reef is located on the north-west coast of 'Tahiti Island, off Papeete harbour (Fig. 1). Tahiti is a volcanic island situated at 17"SO'S and 149"20'W in the Society Archipelago (French Polynesia, central Pacific Ocean); it is surrounded by discontinuous fringing reefs that grade locally into barrier reefs.
The prevailing physical conditions at Tahiti are described in Delesalle et al. (1985). The climate is typically tropical, with two distinct seasons: from November to April (austral summer), the conditions are hot and rainy, with maximum sea surface tem- peratures of 28-29"C, while from May to October (austral winter), the climate is relatively less humid, with lower seawater temperatures (2425°C). Although the annual rainfall averages 2000 mm at Papeete, there are marked variations in rain intensity throughout the year: minimum monthly values are less than 50 mm in winter, while maximum values, up to 400 mm, occur in January and February.
The hydrological regime is primarily a response to the southeast and north-east trade winds. As the island is elongated along a NW-SE axis, the swells generated by these trade winds occur on the longest part of the coast. Surges ofthe north and west sectors originating with tropical depressions and cyclones are less frequent. Considering its location with respect to the prevailing conditions of swells, the studied reef is exposed to high energy conditions (Fig. 1); it pos- sesses reef flats which emerge at spring low tide water level and waves commonly break on the central areas of reef flats at high tide water level. During the winter, the winds, usually 5-8 m s - I (maximum of 15 m s - I) , may generate swells less than 2 m in amplitude in the outer shelves and plunging breakers on the reef front up to 3 m high. In summer there are only weak winds (less than 5 m s - ') and swells (maximal amplitude of 1 m), as well as occasional cyclones; the latter generate strong whirlwinds (100- 200 km h - I ) and ocean swells 8-10 m in amplitude. Calm winds, less than 1 m s - occur briefly during the interseasonal periods. The tides are semi-diurnal and their range rarely exceeds 0.4 m. the salinity of the open sea averages 36 g 1 - I , while variations in salinity are marked in the backreef water masses (3435 g 1 - I ) . Dissolved oxygen concentrations in proximal ocean waters are relatively low, ranging from 6.4 to 6.8 mg 1 - I . In contrast, the reef flats display higher concentrations (7-9 mg 1 - I ) , mark- edly above the oxygen saturation level, due to the
metabolic activity of coral zooxanthellid symbionts. Ocean waters off Papeete have very low nutrients concentrations (0.049.08 patoms-grams N . 1 - ': NO,; 0.060.2 patoms-grams N . 1 - I : NO,; 0.2- 0.45 patoms-grams P . 1 - PO,). Nutrient contents vary widely between the rainy and less humid seasons; maximum values are reached in the waters overlying the reef flat during the summer: 0-1-0.3 patoms-grams N . 1 - I : NO,; 0.3-0.6 patoms- grams N . 1 - NO, and 0 4 0 . 5 patoms- grams P . 1 - I : PO,. As indicated by direct field observations, such an enrichment in nitrogen and phosphorus is probably not high enough to initiate eutrophication or modify the balance between coral- gal and microbial communities.
Modern reef physiography and biozonation
The studied reef belongs to the barrier reef type. Based on morphology and biological patterns observed in the field, the following environments from the open sea landwards can be distinguished.
( 1 ) The outer reef slope comprises coral-built spurs alternating with grooves. The slope is relatively gentle (15-20°C) to a water depth of about 15 m. In the upper part of the spur and groove system, biological communities are typical of high energy environ- ments (Faure, 1982; Adey, 1986; Veron, 1986) and characterized by an abundance of branching and massive scleractinian corals (Acropora robusta, A. danai; Pocillopora verrucosa, Montipora erythraea, Montastraea curta, Porites lobata) and hydrocorals (Millepora platyphylla) with subordinate domal forms (Favia, Favites); associated framebuilders are coralline algae (Porolithon onkodes, Neogoniolithon, Hydrolithon, Lithothamnium and Lithophyllum) and vermetid gastropods (Dendropoma maximus, Serpulorhis annulatus).
(2 ) The ree f ja t zone is relatively narrow (130 m in maximum width) and consists of three distinct sub- zones. The outermost subzone (i.e. reef edge) is dominated by encrusting coralline algae (mostly Porolithon). The intermediate subzone (i.e. patchy reef flat) exhibits scattered coral heads including branching Acropora abrotanoides, A. cytherea, Pocil- lopora verrucosa) and massive (Porites australiensis. P. lutea) forms. In the innermost subzone (i.e rubble reef flat), the floor is chiefly covered by rubble and sand. Inhabitants are limited in diversity and abun- dance: only rare coral colonies occur (Acropora,
Coralgal-stromatolite, Tahiti 659
Porites). The reef flat is connected to the lagoonal floor through a gently dipping biodetrital talus apron
( 3 ) The back-reefzone corresponds to a 1 km wide bay, reaching a maximum depth of 20 m. Two large flat-topped patch reefs, respectively about 90 000 and 15 000 m2 in extent, occur in the central part of the lagoon. Their windward slope is steep with a nearly vertical drop-off along the northern and western margins; in contrast, the leeward slope is relatively gentle. In terms of zonation, the top of these patch reefs corresponds to an exposed reef flat zone domi- nated by branching corals (Acropora robusta, A. danai, Acropora abrotanoides, Pocillopora), associ- ated with scattered massive Porites and faviids; patchy algal pavements composed of chlorophytes (Halimeda) and rhodophytes (Amphiroa) are also present.
REEF DEVELOPMENT
Internal structure and biological composition of the reef
The cored framework belongs to one of the major biologicallsedimentological reef facies, i.e. branching Acropora facies (Fig. 2). This facies forms 0.1 to 1 m thick sections and comprises a cavernous framework of massive, branching or platy species of Acropora, dominated by A. robusta associated with A . danai and A. abrotanoides and a few small Porites heads. The major framework support is composed of veneers, up to several centimetres thick, dominated by coralline algae associated with sessile vermetid gastropods and arborescent foraminifers. The most promi- nent coraline algal contributors are Porolithon onkodes, Neogoniolithon absinde, N. fbsliei, and N. propinquum, intermingled with Hydrolithon cf. reinboldi, Mesophyllum cf. prolijer, Lithothamnium sp., Lithophyllum sp., Titanoderma sp., Heteroderma caribeum and Lithoporella sp.; the encrusting squa- mariaceae Polystratu dura and Peyssonnellia sp. occur occasionally. The vermetid assemblage is domi- nated by Serpulorbis annulatus, locally associated with Dendropoma maximus. Cryptic foraminifers include Homotrema rubrum, Carpenteria cf. monticu- laris and Acervulina inhaerens. Cavities, up to 10 cm in size, are common and are locally occupied by large fragments of the echinoid Heterocentrus trigonarius. Borings by sponges, bivalves and polychaetes are widespread.
AGES B.P. x 1000
0 1 2 3 4 5 6 7 8 0 '
5-
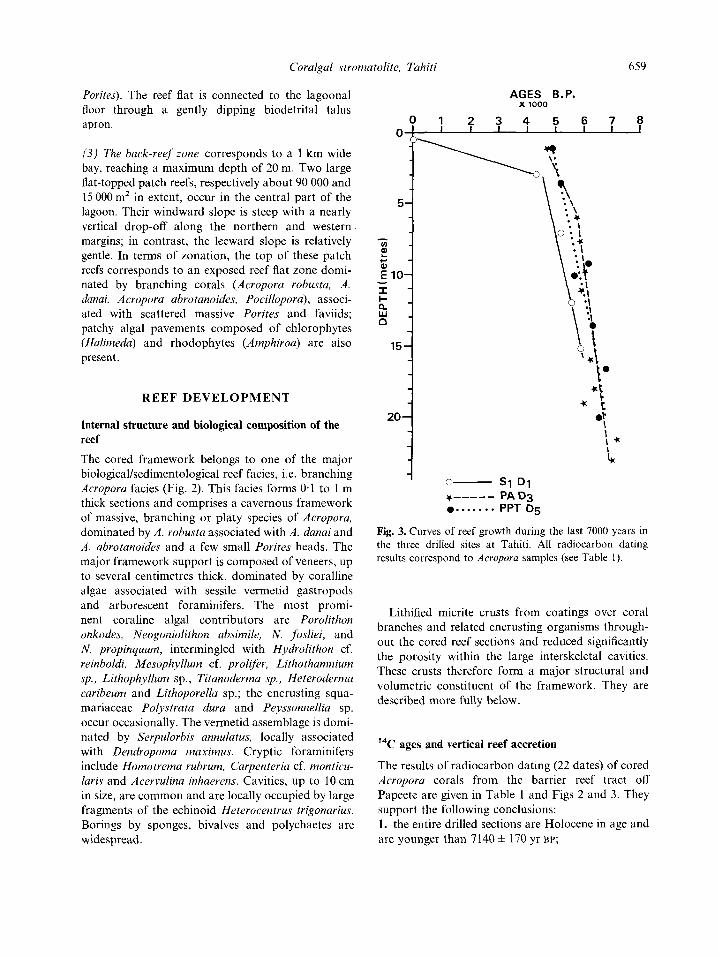
20 '1 Fig. 3. Curves of reef growth during the last 7000 years in the three drilled sites at Tahiti. All radiocarbon dating results correspond to Acropora samples (see Table I ) .
Lithified micrite crusts from coatings over coral branches and related encrusting organisms through- out the cored reef sections and reduced significantly the porosity within the large interskeletal cavities. These crusts therefore form a major structural and volumetric constituent of the framework. They are described more fully below.
I4C ages and vertical reef accretion
The results of radiocarbon dating (22 dates) of cored Acropora corals from the barrier reef tract off Papeete are given in Table 1 and Figs 2 and 3. They support the following conclusions: 1. the entire drilled sections are Holocene in age and are younger than 7 140 * 170 yr BP;
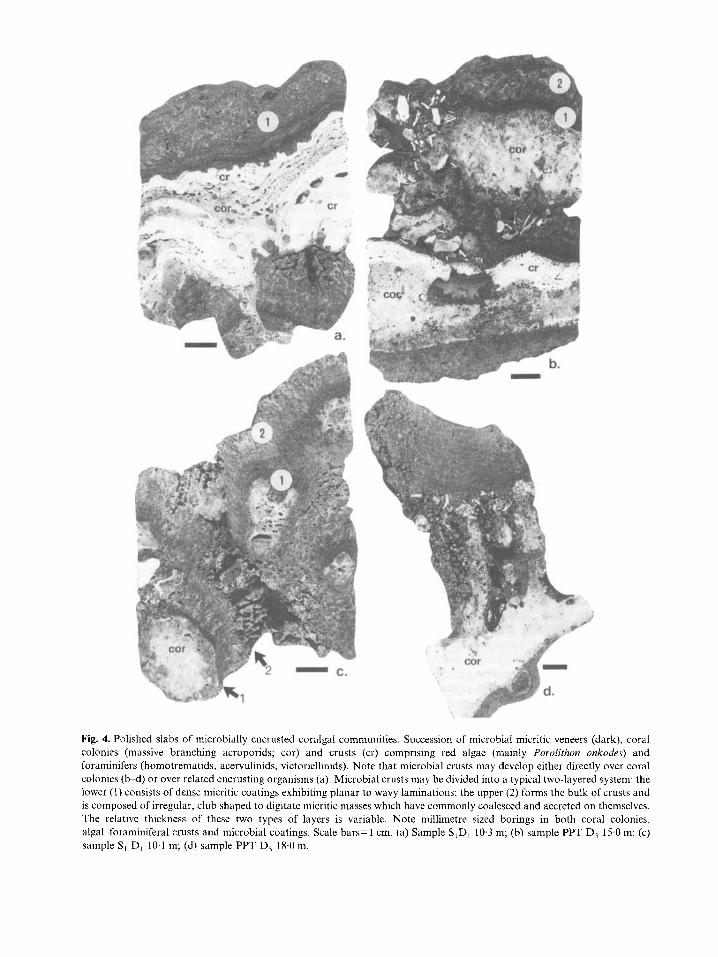
Fig. 4. Polished slabs of microbially encrusted coralgal communities. Succession of microbial micritic veneers (dark), coral colonies (inassive branching acroporids; cor) and crusts (cr) comprising red algae (mainly Porolithon onkodes) and foraminifers (homotrematids, acervulinids, victoriellinids). Note that microbial crusts may develop either directly over coral colonies (b-d) or over related encrusting organisms (a). Microbial crusts may be divided into a typical two-layered system: the lower ( I ) consists of dense micritic coatings exhibiting planar to wavy laminations; the upper (2) forms the bulk of crusts and is composed of irregular, club shaped to digitate micritic masses which have commonly coalesced and accreted on themselves. The relative thickness of these two types of layers is variable. Note millimetre sized borings in both coral colonies, algal foraminifera1 crusts and microbial coatings. Scale bars= 1 cm. (a) Sample S,D, 10.3 m; (b) sample PPT D, 15.0 m; (c) sample S , D, 10.1 m; (d) sample PPT D, 18.0 m.

Coralgal-stromatolite, Tahiti 66 1
Fig. 5. Thin section photomicrographs of microbial micritic crusts. Microbial micritic crusts exhibit a typical two-layered system including laminated coatings (1) and micritic masses (club shaped, digitate to shrubby, upward accreting growth forms) displaying a mottled to clotted thrombolite-like internal fabric (2). Note the occurrence of thin red algal crusts (white arrows) over coral branches before the development of microbial veneers. Scale bars ~ 0 . 5 cm. (a) Sample S,-D, 9.9 m; (b) sample PPT D, 9.3 m; (c) sample S,-D, 10.1 m.
2. various parts of the reef developed in different ways during growth to sea level (Montaggioni, 1988; Pirazzoli & Montaggioni, 1988). The barrier reef margin first tracked, then caught up sea level to reach an equilibrium position (‘catch-up strategy’), while the nearby patch reefs have been able to keep pace with rising sea level (‘keep-up’ growth strategy; Neumann & MacIntyre, 1985; Davies et al., 1985). The growth curves in Fig. 3 suggest that upward growth was largely uniforni; inflections in the curve represent variations in growth rates, which do not coincide with any change of facies. The total variation in the rate of vertical reef accretion is 0.6-14 mm yr -- I . The slowest vertical accretion rate recorded (0.6 mm yr I ) corresponds to the growth stabilization event (Davies el al., 1985), i.e.
the time when the reef top reached within 2 m of present day sea level (Fig. 3). According to Smith & Kinsey (1976) and Smith (1983), net calcifi- cation rates of reefs can be expressed as a function of net vertical accretion and framework porosity. Assuming that Acropora colonies yielded open framework structures with 75% primary porosity, net CaCO, production can be estimated for the main reef growth phases (Table 2). The outer barrier reef flat calcified at a rate of 6 kg CaCO, m - yr - from 15.2 to 3 m below present reef surface. The lagoonal patch reefs exhibited net calcification rates of 6.8-10.4 kg CaCO, m - * yr - I .
The slowest calcification rates recorded (i.e. 0.8 kg CaCO, mP2yr - ’ ) are related to the growth stabilization event.

662 G. F. Camoin and L. F. Montaggioni
Fig. 6. Thin section photomicrographs of microbial micritic crusts. (a, b) Club shaped to digitate microbial micritic accretions (arrows); they coalesced and accreted on themselves so that multiple generations are closely packed (microbial accretions are dark). The convex upward laminae resulted from intercalation of dense micritic films. Irregular shaped cavities between microbial accretions are partially filled with sediment (s). Sample S,-D, 10.1 m. Scale bars=500 pm. (c) Millimetre sized boring within a microbial micritic crust. Sample S,-D, 10.1 m. Scale bar=250 pm. (d) An irregular thrombolitic micritic mass (1) displaying primary cavities partially filled with sediment (s). Note the clotted to peloidal internal fabric of the micritic mass. Sample S,-D, 10.1 m. Scale bar=250 pm.
MICRITIC CRUSTS
Structure and composition of micritic crusts
Laminar to columnar Mg-calcite coatings, commonly 0.1-5.0 cm thick, develop over corals in the growth position or, more commonly, over associated encrusting red algae and foraminifers (Fig. 4). Thin crusts also occur in framework cavities or borings affecting corals and algal crusts, implying that the formation of these laminated coatings was not strictly related to light conditions. Crusts link
adjacent coral colonies and their development reduced the porosity of the entire framework by about 40%, thus restricting significantly the available space for subsequent internal sedimentation and cementation.
The outer limit to the crusts is sharp. Their internal microstructure ranges from dense laminated micrites to clotted, peloidal micrite; clots and peloids range in size from 5 to 50pm. No significant changes in the overall composition of laminated micritic crusts have been recorded through the sequence.
Corulgul~,~tromatolite. Tahiti 663
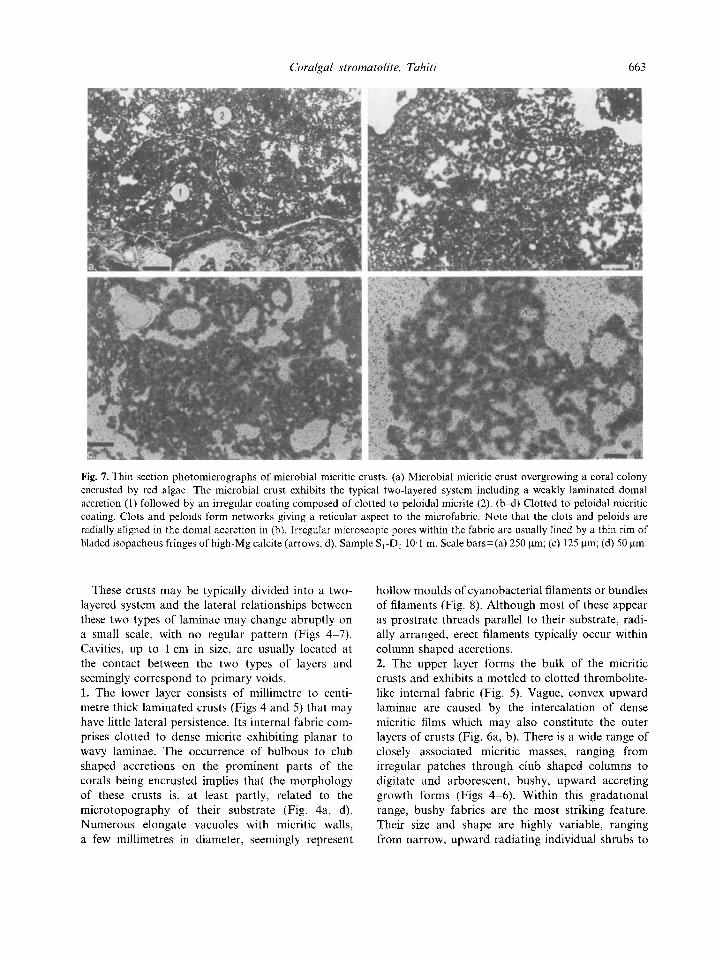
Fig. 7. Thin section photomicrographs of microbial micritic crusts. (a) Microbial micritic crust overgrowing a coral colony encrusted by red algae. The microbial crust exhibits the typical two-layered system including a weakly laminated domal accretion (1) followed by an irregular coating composed of clotted to peloidal micrite (2). (b-d) Clotted to peloidal micritic coating. Clots and peloids form networks giving a reticular aspect to the microfabric. Note that the clots and peloids are radially aligned in the domal accretion in (b). Irregular microscopic pores within the fabric are usually lined by a thin rim of bladed isopachous fringes of high-Mg calcite (arrows, d). Sample S,-D, 10.1 rn. Scale bars=(a) 250 pm; (c) 125 pm; (d) 50 pm.
These crusts may be typically divided into a two- layered system and the lateral relationships between these two types of laminae may change abruptly on a small scale, with no regular pattern (Figs 4-7). Cavities, up to 1 cm in size, are usually located at the contact between the two types of layers and seemingly correspond to primary voids. 1. The lower layer consists of millimetre to centi- metre thick laminated crusts (Figs 4 and 5 ) that may have little lateral persistence. Its internal fabric com- prises clotted to dense inicrite exhibiting planar to wavy laminae. The occurrence of bulbous to club shaped accretions on the prominent parts of the corals being encrusted implies that the morphology of these crusts is, at least partly, related to the microtopography of their substrate (Fig. 4a, d). Numerous elongate vacuoles with micritic walls, a few millimetres in diameter, seemingly represent
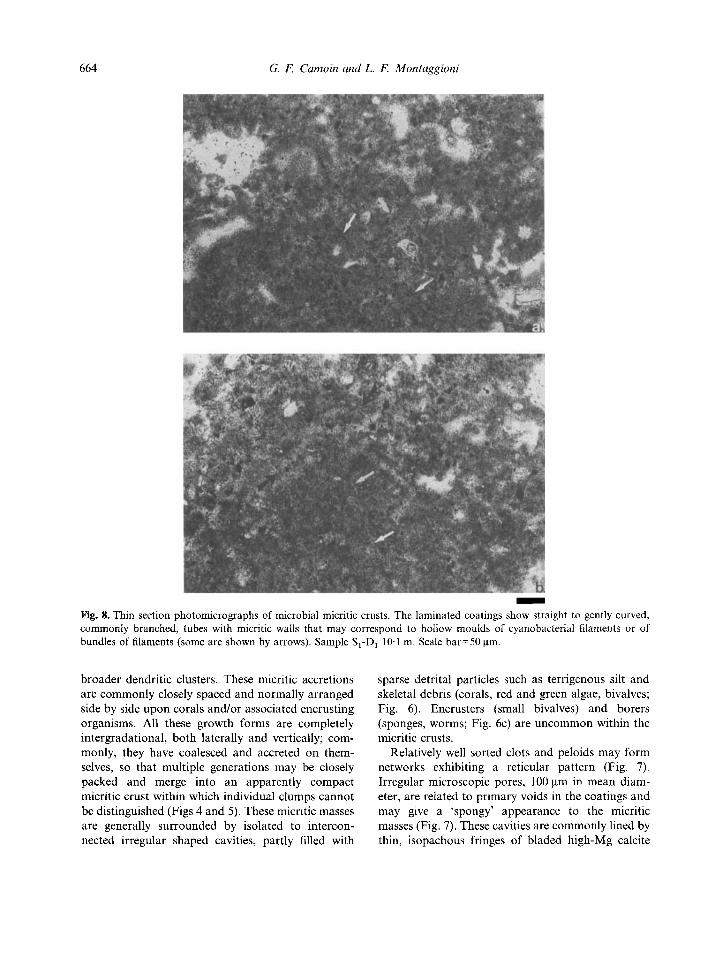
hollow moulds of cyanobacterial filaments or bundles of filaments (Fig. 8). Although most of these appear as prostrate threads parallel to their substrate, radi- ally arranged, erect filaments typically occur within column shaped accretions. 2. The upper layer forms the bulk of the micritic crusts and exhibits a mottled to clotted thrombolite- like internal fabric (Fig. 5). Vague, convex upward laminae are caused by the intercalation of dense micritic films which may also constitute the outer layers of crusts (Fig. 6a, b). There is a wide range of closely associated micritic masses, ranging from irregular patches through club shaped columns to digitate and arborescent, bushy, upward accreting growth forms (Figs 4-6). Within this gradational range, bushy fabrics are the most striking feature. Their size and shape are highly variable, ranging from narrow, upward radiating individual shrubs to
664 G. F. Curnoin and L. F. Montuggioni
- Fig. 8. Thin section photomicrographs of microbial micritic crusts. The laminated coatings show straight to gently curved, commonly branched, tubes with micritic walls that may correspond to hollow moulds of cyanobacterial filaments or of bundles of filaments (some are shown by arrows). Sample S,-D, 10.1 m. Scale bar=SO pm.
broader dendritic clusters. These micritic accretions are commonly closely spaced and normally arranged side by side upon corals andlor associated encrusting organisms. All these growth forms are completely intergradational, both laterally and vertically; com- monly, they have coalesced and accreted on them- selves, so that multiple generations may be closely packed and merge into an apparently compact micritic crust within which individual clumps cannot be distinguished (Figs 4 and 5 ) . These micritic masses are generally surrounded by isolated to intercon- nected irregular shaped cavities, partly filled with
sparse detrital particles such as terrigenous silt and skeletal debris (corals, red and green algae, bivalves; Fig. 6). Encrusters (small bivalves) and borers (sponges, worms; Fig. 6c) are uncommon within the micritic crusts.
Relatively well sorted clots and peloids may form networks exhibiting a reticular pattern (Fig. 7). Irregular microscopic pores, 100 pm in mean diam- eter, are related to primary voids in the coatings and may give a ‘spongy’ appearance to the micritic masses (Fig. 7). These cavities are commonly lined by thin, isopachous fringes of bladed high-Mg calcite
Coralgal-stromatolite, Tahiti 665
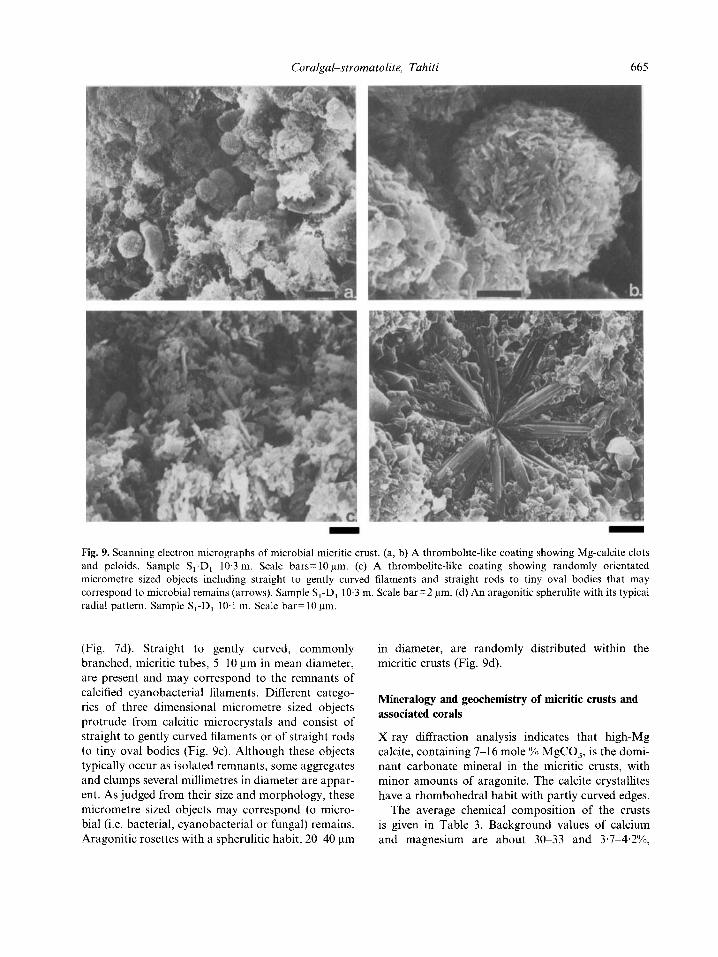
D - Fig. 9. Scanning electron micrographs of microbial micritic crust. (a, b) A thrombolite-like coating showing Mg-calcite clots and peloids. Sample S,-D, 10.3 m. Scale bars= 10 pm, (c) A thrombolite-like coating showing randomly orientated micrometre sized objects including straight to gently curved filaments and straight rods to tiny oval bodies that may correspond to microbial remains (arrows). Sample S,-D, 10.3 m. Scale bar=2 pm. (d) An aragonitic spherulite with its typical radial pattern. Sample S,-D, 10.1 m. Scale bar= 10 pm,
(Fig. 7d). Straight to gently curved, commonly branched, micritic tubes, 5-10 pm in mean diameter, are present and may correspond to the remnants of calcified cyanobacterial filaments. Different catego- ries of three dimensional micrometre sized objects protrude from calcitic microcrystals and consist of straight to gently curved filaments or of straight rods to tiny oval bodies (Fig. 9c). Although these objects typically occur as isolated remnants, some aggregates and clumps several millimetres in diameter are appar- ent. As judged from their size and morphology, these micrometre sized objects may correspond to micro- bial (i.e. bacterial, cyanobacterial or fungal) remains. Ardgonitic rosettes with a spherulitic habit, 2 0 4 0 pm
in diameter, are randomly distributed within the micritic crusts (Fig. 9d).
Mineralogy and geochemistry of micritic crusts and associated corals
X-ray diffraction analysis indicates that high-Mg calcite, containing 7-16 mole 'YO MgCO,, is the domi- nant carbonate mineral in the micritic crusts, with minor amounts of aragonite. The calcite crystallites have a rhombohedra1 habit with partly curved edges.
The average chemical composition of the crusts is given in Table 3. Background values of calcium and magnesium are about 30-33 and 3.7-4.2%,
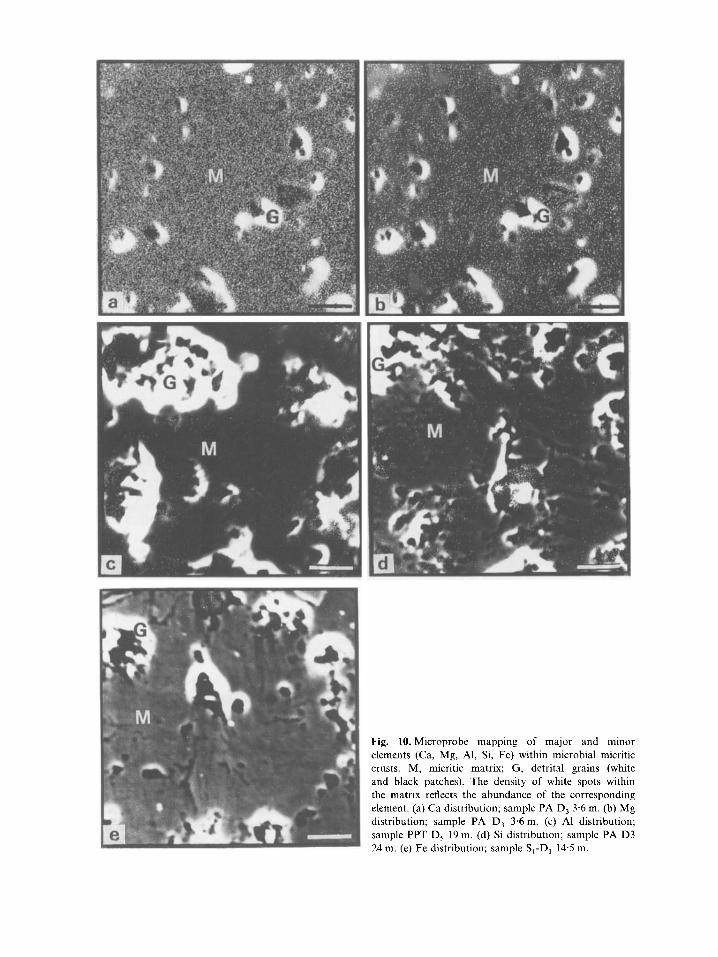
Fig. 10. Microprobe mapping of major and minor elements (Ca, Mg, Al, Si, Fe) within microbial micritic crusts. M, micritic matrix; G, detrital grains (white and black patches). The density of white spots within the matrix reflects the abundance of the corresponding element. (a) Ca distribution; sample PA D, 3.6 m. (b) Mg distribution; sample PA D, 3.6 m. (c) A1 distribution; sample PPT D, 19 m. (d) Si distribution; sample PA D3 24 m. (e) Fe distribution; sample S,-D, 14.5 m.

Coralgal-stromatolite, Tahiti 667
+5.0 -
+4.0 -
+3.0 -
a +2.0 -
n >o + l . O -
a 0
0 m r +o.o - m
-1.0 -
-2.0 -
-3.0 -
-4.0 -
STROMATOLITIC CRUSTS 1
Acropora corals
modern A S l D l 0
PPTD5 * Stromatolitic crusts
S1D1 0 PAD3 N PPTD5 0
-6.0 -5.0 -5.0 -4.0 -3.0 -2.0 -1.0 +1.0 +2.0
6 l 8 0 o/oo PDB Fig. 11. Oxygen and carbon isotope measurements (relative to PDB reference) of Acuopora branches and associated stroniatolitic crusts. Modern Acropora refers to samples taken from colonies on the present day reef front at depths of less than 5 m. Fossil Acropora and stromatolitic crusts refer to samples taken at various levels from drilled Holocene sequences (Sl-Dl, PA D3, PPT Ds).
respectively, the Mg/Ca ratio is almost constant and averages 0.12; this value fits with those given by Folk & Land (1975) for precipitation of calcite from interstitial normal marine waters.
Strontium concentrations range from 800 to 1600 ppm. The Sr/Ca ratio varies considerably and randomly from sample to sample (2.6-8.4); this may indicate a skeletal origin for strontium as an aragonite-incorporated element, rather than co-precipitation with calcite. The amounts of other cations (Mn, Ba, Al, Si, Fe) are generally lower than l000ppm. However, a sharp increase in the minor and trace element content occurs within the samples
poorer in Ca and Mg (those retaining less than 250/0 Ca and 3% Mg). The most striking variations dis- played by the crusts are peaks in concentrations of aluminium (10 000-15 000 ppm), silica (8200- 13 000 ppm) and iron (4100-6100 ppm) and, to a lesser extent, barium (less than 1600 to 2100 ppm).
Microprobe analysis of thin sections from micritic crusts reveals the spatial distribution of the most abundant elements (Fig. 10). All the elements con- sidered (Ca, Mg, Fe, Si, Al) are generally scattered throughout the sections examined; there is no evi- dence of metal-rich coatings on the crusts. The local occurrence of Si- and Fe-rich patches probably
668 G. F. Camoin and L. I? Montaggioni
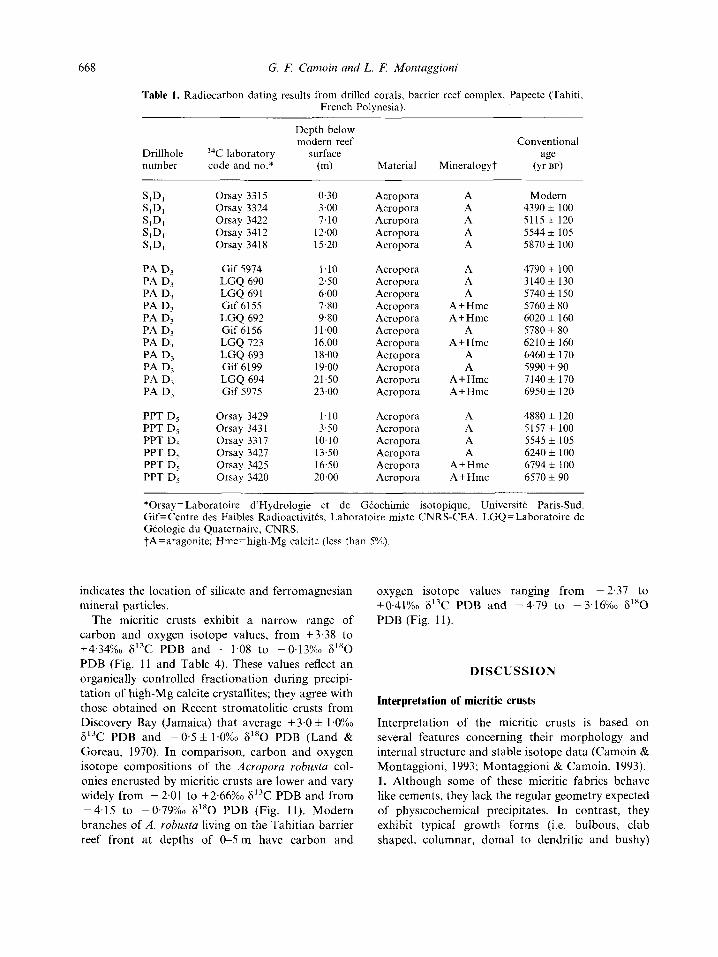
Table 1 . Radiocarbon dating results from drilled corals, barrier reef complex, Papeete (Tahiti, French Polynesia).
Depth below modern reef Conventional
Drillhole I4C laboratory surface age number code and no.* (m) Material Mineralogy? (Yr BPI
Orsay 3315 Orsay 3324 Orsay 3422 Orsay 3412 Orsay 3418
Gif 5974 LGQ 690 LGQ 691 Gif 6155 LGQ 692 Gif 6156 LGQ 123 LGQ 693 Gif 6199 LGQ 694 Gif 5975
Orsay 3429 Orsay 3431 Orsay 3317 Orsay 3427 Orsay 3425 Orsay 3420
0.30 3.00 7.10
12.00 15.20
1.10 2.50 6.00 7.80 9.80
11.00 16.00 18.00 19.00 21.50 23.00
1.10 3.50
10.10 13.50 16.50 20.00
Acropora Acropora Acropora Acropora Acropora
Acropora Acropora Acropora Acropora Acropora Acropora Acropora Acropora Acropora Acropora Acropora
Acropora Acropora Acropora Acropora Acropora Acropora
A A A A A
A A A
A+Hmc A+Hmc
A A+Hmc
A A
A + Hmc A + Hmc
A A A A
A+Hmc A+Hmc
Modern 4390 f 100 5115 f 120 5544 i 105 5870 f 100
4790 f 100 3140f 130 5740 f 150 5760 f 80 6020 f 160 5780 f 80 6210 f 160 6460 f 170 5990 f 90 7140 f 170 6950 + 120
4880 f 120 5157 f 100 5545 f 105 6240 f 100 6794 f 100 6570 f 90
*Orsay= Laboratoire d'Hydrologie et de Gtochimie isotopique, Universitt Paris-Sud. Gif= Centre des Faibles Radioactivitts, Laboratoire niixte CNRS-CEA. LGQ=Laboratoire de Geologie du Quaternaire, CNRS. ?A=aragonite; Hmczhigh-Mg calcite (less than 5%)).
indicates the location of silicate and ferromagnesian mineral particles.
The micritic crusts exhibit a narrow range of carbon and oxygen isotope values, from +3.38 to +4.34%0 6°C PDB and - 1.08 to -0.13%0 6 '*0 PDB (Fig. I I and Table 4). These values reflect an organically controlled fractionation during precipi- tation of high-Mg calcite crystallites; they agree with those obtained on Recent stromatolitic crusts from Discovery Bay (Jamaica) that average +3.0 k 1.0%0 613C PDB and - 0.5 f 1.0%0 6"O PDB (Land & Goreau, 1970). In comparison, carbon and oxygen isotope compositions of the Acropora robusta col- onies encrusted by micritic crusts are lower and vary widely from - 2.01 to +2.66%0 SI3C PDB and from - 4.15 to - 0.79%0 S1'O PDB (Fig. 11). Modern branches of A. robusta living on the Tahitian barrier reef front at depths of 0-5 m have carbon and
oxygen isotope values ranging from -2.37 to +0.410/00 6°C PDB and - 4.79 to - 3.16%0 6"O PDB (Fig. 11).
D I S C U S S I O N
Interpretation of micritic crusts
Interpretation of the micritic crusts is based on several features concerning their morphology and internal structure and stable isotope data (Camoin & Montaggioni, 1993; Montaggioni & Camoin, 1993). 1. Although some of these micritic fabrics behave like cements, they lack the regular geometry expected of physicochemical precipitates. In contrast, they exhibit typical growth forms (is. bulbous, club shaped, columnar, domal to dendritic and bushy)

Coralgal-stromatolite, Tahiti 669
Table 2. Estimates of net calcification rates during vertical reef growth of the barrier reef complex (Papeete, Tahiti).
Growth Growth Vertical growth Net calcification Drillhole thickness duration rate rate (*) number (m) (years) (mm yr ’) (kg CaCO, m-’yr-’)
S,D, S,Dl
PA D, PA D,
PPT D, PPT D,
12.20 1480 8.24 2.70 4390 0.61
17.00 1210 14.00 4.90 950 5.20
6.50 554 11.70 12.40 1360 9.10
6.1 0.8
10.4 4.0
8.6 6.8
*Assuming a framework porosity of 75% and aragonite mineral density of 2.9 g cm-3 for colonies of Arropora robustn and A . dnnai.
Table 3. Average chemical composition and Mg/Ca and SrKa molar ratios of stromatolitic crusts associated with Acropora reef framework of the barrier reef complex (Papeete).
S,D, S,D, S,D, PAD, PAD, PAD, PPTD, PPTD, Depth (m)t 7.6 12.0 14.5 3.6 16.0 23.0 12.0 19.0 n (6) ( 3 ) (3) (5) (6) (6) ( 5 ) (9)
Ca
Sr Mn Ba A1 Si Fe Mg/Ca Sr/Ca ( x 10,)
Mg 24.80
2.92 <2100 <2100 <2100 10 000
8200 6100 0.11 8.4
33.32 4.26 900
1100 <900
300 400
<600 0.12 2.7
32.70 4.04
< 1400 4 0 0 <900 <200 <200 <700 0.12 4.2
33.05 4.08 I400
<400 1700
100 300 700
0.12 4.2
30.63 3.77 800
<400 < 600 7700 2800 1700 0.12 2.6
33.59 4.19 1600
~ 5 0 0 <600 <300
< I300 500
0.12 4.7
21.23 2-88
<1600 <2800 < I600 15 200 13 300
4100 0.13 7.5
33.16 4.16 1300 <300 < 500
400 500
<400 0.12 3.9
Ca and Mg values are expressed in per cent, the values of the other elements are expressed in PPm. ?Below reef surface.
and laminations which allow them to be interpreted as bio-accretionary features. 2. The generalized occurrence of microbial remains (e.g. cyanobacterial filaments, traces and/or moulds of filaments, micrometre sized bacterial bodies pro- truding from calcitic microcrystals) suggest that these features result from the mineralization of cyanobac- terial, or more generally microbial, mucilaginous material. 3. The trapping of extraneous grains on the crusts, even on vertical surfaces, suggests that the crust exterior was at one stage composed of a sticky and soft mucilaginous material (see Monty, 1976; Riding et al., 1991a, for modern and Miocene counterparts, respectively).
4. Internally the crusts are predominantly com- posed of clotted to peloidal micrites (‘structure grumeleuse’, Cayeux, 1935; or ‘grumous’, Walter, 1972). Quite similar fabrics are common in micro- bial carbonates and are usually thought to be a result of in situ precipitation (Riding et al., 1991a). A similar peloidal fabric has been reported in lithi- fied Mg-calcite micritic crusts from a Holocene section of One Tree Reef (Marshall, 1983). 5. Crusts display great variations in thickness, lateral persistence and internal structure. 6. They exhibit abundant fenestrae which seemingly correspond to primary voids. 7. Carbon and oxygen isotope values indicate iso- topic enrichment with respect to the expected values

670 G. F. Camoin and L. F Montuggioni
Table 4. 6°C and 6"O &o, PDB) of stromatolitic crusts from the Holocene reef sequences of the barrier reef
complex (Papeete).
Drillhole number Depth (m)* 6°C 6I8O
S,D, 3.5 3.74 ~ 0.32 1.6 4.01 - 0.21 9.9 3.13 ~ 0.39
12.0 4.11 - 0.52 14.5 4-07 ~ 0.49
2.5 3-52 - 0.83 3.6 3.87 ~ 1.08 9.8 4-14 - 0.34
13.0 4.34 - 0.14 16.0 3-99 ~ 0.49 20.0 3.63 - 0.84 24.0 3.95 ~ 0.13
PPT D, 3.5 3.38 - 0.39 8.0 3-80 - 0.54
12.0 4-07 ~ 0.41 19.0 4-00 - 0.46 20.0 3.84 - 0.78
*Below reef surface. Samples analysed by M. Pachiaudi, University of Lyon, France.
for a cement precipitated at equilibrium with nor- mal seawater, thus suggesting organic fractionation during precipitation. These data strengthen the microbial interpretation of the relevant crusts.
As previously pointed out by Riding et al. (1991a) for Miocene micritic crusts, such a suite of fabrics has not been demonstrated to form through physico- chemical cementation.
Within the observed range of micritic encrusta- tions, arborescent dendritic fabrics are probably the most striking features and closely resemble calcified cyanobacterial sheath material, thus suggesting active microbial calcification processes. In modern oncoids, calcification of the thick mucilaginous sheath material of Oscillatoriaceaea, Schizothrix calcicola in particular, may produce irregular micritic tufted masses very reminiscent of the bushy fabrics reported here (Riding & Voronova, 1982). Quite similar fabrics are prominent in Cambrian and early Ordo- vician reefs (e.g. AngulocellularialEpiphyton group; Riding & Voronova, 1982), in early Cretaceous rocks (Camoin, 1983; G. F. Camoin et al., work in progress), in Miocene reefs (Riding et ul., 1991a; G. F. Camoin, work in progress) and in modern
sediments from Shark Bay (Monty, 1976). Recent analogues of these dendritic fabrics are also reported in various continental deposits (travertines, tufas and hot spring deposits) and were attributed to the activity of bacteria (Chafetz & Folk, 1984), lichens, algae or fungi (Jones & MacDonald, 1989). From the previous interpretations, it can be inferred that dis- tinct taxa of micro-organisms (bacteria, cyanobac- teria, fungi) may produce similar features, implying that the relevant similarities probably result from morphological convergence and are not indicative of systematic affinities. Nevertheless, it should be stressed that many stromatolites, even in modern environments, may retain little, if any, information concerning the organisms responsible for their for- mation (see also Riding, 1992). The prime evidence of microbial involvement may be obscured by rapid decay of microbial colonies (Monty, 1976) or by the calcification of a thick and diffuse, often degraded and homogenized, sheath material (Riding & Voronova, 1982). Thus, the apparent lack of micro- bial remains does not necessarily imply a pure physi- cochemical origin for the laminated crusts reported in other Quaternary reef tracts. A better knowledge of the role of microbial communities in Quaternary reef environments may also rely on the study of less distinctive, and often obscure, microbial features (cryptic microbial fabrics; Riding, 1991).
The predominantly clotted to peloidal nature of these micritic crusts raises the problem of the origin of clots and peloids. As suggested by Riding et al. (1991a), peloidal micritic crusts with more or less developed lamination may form in reefs as a result of distinct processes including (i) in situ precipitation (i.e. inorganic or microbially mediated) and (ii) trap- ping and binding of allochthonous grains by a micro- bial mat. In modern algal mats, clotted micrites may result from the replacement of coccoid cyanobac- terial communities, from the breakage of Scytonema sheaths or from erratic carbonate precipitation within cyanobacterial mucilage (Monty, 1976). Kennard & James (1986) attributed the clotted fabric of thrombolites, which is similar to that of the Tahiti crusts, to the calcification of coccoid microbial com- munities. The scarcity, or even the lack, of extraneous particles (e.g. terrigenous silt and sand, peloids, ooids and skeletal metazoan debris) as well as the abun- dance of microbial remains in our micritic crusts suggest that trapping and binding processes were of minor importance in comparison with calcification of microbial mucilaginous material and in situ, microbi- ally mediated, precipitation. The processes involved

Coralgal-stromatolite, Tahiti 67 1
in the formation of such predominantly prokaryotic peloidal micritic crusts contrast sharply with the trapping of sand material and the synsedimentary precipitation of carbonate cements involved in the formation of modern coarse grained columnar ‘eualgal-cyanobacterial’ stromatolites in shallow marine subtidal environments (Shark Bay, Western Australia, and bahamas; Riding et al., 1991b).
Current debate on the origin of spherulitic peloidal fabrics, particularly modern examples (MacIntyre, 1984, 1985; Chafetz, 1986), usually emphasized in situ precipitation, whether physicochemical, microbial, or both (MacIntyre & Marshall, 1989). MacIntyre (1984, 1985) and Lighty (1985) regarded spherulitic, well sorted (20-60mm) peloids in crusts and pro- tected cavities as marine cements, involving repeated nucleation around centres of growth. Conversely, Chafetz (1986) proposed that the formation of marine peloids may result from the microbially medi- ated precipitation of high-Mg calcite within and around active clumps of bacteria. Monty (1967) assumed that pellets and pellet-like structures found in algal mats and algal decay environments may be of bacterial origin. The occurrence of micrometre sized, probably bacterial, rods and cocci within micro- crystals constituting peloids and clots is another clue to the involvement of microbial organisms in precipitation processes.
Aragonitic rosettes with a spherulitic habit similar to those observed in the Tahiti micritic crusts have been reported within other stromatolitic crusts and were either regarded a calcified cyanobacteria (Buchbinder, 1981) or as by-products of the activity of bacteria, both in natural environments and in microbiological experiments (Krumbein, 1974; Krumbein & Cohen, 1977; Lighty, 1985; Chafetz, 1986; Brachert & Dullo, 1991).
The two-layered system of the Tahiti micritic crusts probably reflects slight differences in the com- position of the microbial communities; laminated crusts (i.e. stromatolites) should be related to epi- sodic sediment trapping and/or carbonate precipitat- ing activities of microbial communities dominated by filamentous microbes, whereas clotted micritic masses (i.e. thrombolites) should be interpreted as primary growth forms of calcified, internally poorly differentiated, coccoid dominated microbial commu- nities (see also Kennard & James, 1986). Thus, these microbial crusts provide an example of stromatolitic coatings grading upward and outward into throm- bolite caps. Furthermore, the poor development of internal laminations may be related to abundant in
situ production of carbonate muds, suggesting that the accretion of these microbial crusts was probably continuous and rapid.
Environmental conditions during reef growth
Benthic communities intermingled with micritic crusts in the Tahitian Holocene reef sequence provide information on water depth or wave energy at the time of framework formation. In modern environ- ments, the reef-building acroporids (Acropora rohusta, A . danai and A . abrotanoides) are restricted to depths less than 5 m at reef edges exposed to wave action (Faure, 1982; Veron, 1986). Massive coralline algal coatings dominated by Porolithon onkodes, Neogoniolithon josliei and Hydrolithon cf. reinholdi grow under rough water conditions at depths of less than 6 m (Adey, 1986). In French Polynesian reefs, the vermetid assemblage composed of Serpulorbis annulatus, with subordinate Dendropoma maximus is strictly limited to a depth interval ranging from near low tide level to about 3 m (Richard, 1982). The cavity dwelling echinoid Heterocentrus trigonavius occurs in high energy areas at depths of about 3 m (de Ridder, 1986). All these data imply that the reef frameworks have developed continuously in a high energy environment, at depths not greater than 5-6 m below the mean sea level. This is consistent with the reef growth curves presented in Fig. 3 .
The physicochemical environmental conditions during reef growth can be estimated from carbon and oxygen isotope data obtained from the acroporid corals. By comparison with their modern analogues, 6I3C and 6 l80 records from fossil acroporids display a positive shift of 2.1 and 2.30/00, respectively, and a wide overlap with the composition of modern corals (Fig. 11). As 6’’O values in corals are potentially temperature monitors (Weber & Woodhead, 1972; Smith & Kinsey, 1976; Swart & Coleman, 1980; Dunbar & Wellington, 1981; Patzold, 1984; Aharon, 1991) and, to a lesser extent, salinity gauges (Dunbar &Wellington, 1981; Aharon, 1991), it can be inferred that, as the Tahitian barrier reef grew upward, ambi- ent surface waters were at temperatures similar to or slightly lower than present and probably had normal open ocean salinities. The coral carbon isotope composition is mainly regarded as reflecting the biomass productivity of the reef community (Smith & Kroopnick, 198 1; Aharon, 1985), metabolic activity of coral symbionts (Weil et al., 1981; Swart, 1983; MacConnaughey, 1989) andlor seasonal light inten- sity (Patzold, 1984). In the study area, lower 6I3C

672 G. F. Camoin and L. F. Montaggioni
values in fossil Acropora (Fig. 11) may be due to higher light intensity or enhanced activity of the coral's endosymbiotic algae, thereby increasing growth rate of the colonies. This is in agreement with high calcification rates exhibited by the barrier reef during the period of vertical growth.
The chemical composition of the micritic crusts partially reflects the chemistry of ambient sea-water. In particular, the incorporation of metal and metal- loid elements into reefal deposits provides evidence for dissolved element concentration (Shen & Boyle, 1987; Shen et al., 1987, 1991; Lea et al., 1989; Brown et al., 1991). The elevated aluminium, silica and iron contents in micritic crusts from various stratigraphi- cal levels (Table 3) may indicate high terrigenous inputs to the reefs off Papeete, due to occasional freshwater discharge. Mg/Ca ratios (Table 3) indicate that the micritic crusts have precipitated from normal marine waters.
Another clue to water conditions during reef growth is provided by calcium carbonate accumula- tion rates (Table 2). As modern, normally function- ing coral reefs commonly calcify at an average rate of 4 kg m ~ yr ~ ' (Smith, 1981, 1983; Kinsey, 1983), with an extreme at 10 kg, it can be assumed that the Polynesian reefs studied, with calcification rates of 6.8-10.4 kg m ~ yr ~ ', grew under normal environ- mental conditions throughout the Holocene.
The nature and the high diversity of involved coralgal communities, the CaCO, accumulation rates and the stable isotope measurements indicate that ecological conditions for reef development have been optimal at Tahiti for the last 7000 years. Although these data imply that no significant long term environmental changes have occurred during reef growth, short term abnormal effects may be con- sidered in order to explain the abrupt changes in biota composition and the subsequent predominance of microbial communities over true encrusters (i.e. red algae and foraminifers). The scarcity or even the lack, of such encrusting organisms within microbial crusts suggests that these organisms were excluded during periods of microbial growth, either due to temporarily unfavourable ecological conditions or to chemical deterrents. Another possible explanation lies in subtle fluctuations of ecological parameters (e.g. salinity, temperature, nutrient availability). In particular, a temporary increase in nutrient availabil- ity would have favoured algal settlement (Nelson, 1985; Cuet et al., 1988; Montaggioni et al., 1989, 1993) and, finally, microbial communities at the expense of coral growth. The widespread occurrence
of microbial crusts throughout the cored sequences implies that such nutrient inputs would have occurred repeatedly during Holocene time, affecting both the barrier reef and the patch reefs. The only plausible mechanism could be related to temporary discharge of nutrient-enriched freshwater. Although freshwater influences are not recorded by mean iso- topic values reported here, they should be confirmed by high resolution isotopic analyses on stromatolitic crusts (G. F. Camoin et al., work in progress). However, the high reef accretion rates reported imply that such possible short term stressful periods did not significantly affect overall reef growth during Holocene time.
Sedimentary roles of micritic crusts in the reef framework
Microbial micritic crusts are a major structural and volumetric constituent of the reef framework throughout the Holocene sequence drilled at Tahiti. They usually form the last stage of encrustation since they develop over corals in growth position or, more commonly, over associated encrustations formed by coralline algae, vermetid gastropods and encrusting foraminifers. Alternations between coralgal colonies and microbial crusts are also present locally.
The occurrence of borings (clionids, worms and cyanobacteria) into red algal crusts developing between corals and microbial coatings indicates that some time elapsed prior to the formation of micritic crusts. Accordingly, the microbial crusts grew gener- ally over dead parts of coral branches, perhaps some way down within the dead reef framework, implying that there was generally no direct space competition between coralgal communities and microbial organ- isms, except for few alternations that are clues to local successive growths. Furthermore, the high accretion rates reported here imply that reef growth was not impeded by microbial crust formation which occurred primarily in interstices located between adjacent coral colonies. As a result, the microbial veneer thickly coated coralgal colonies and reduced significantly the porosity within large interskeletal cavities, subsequently filled with unbound detrital particles. Thus, the development of microbial micritic crusts seemingly acted upon porosity in two appar- ently opposite ways: (i) a reduction of the overall framework porosity (about 40%) and (ii) the preser- vation of this porosity as dense, early lithified, protective seals.

Coral~al-stroniutolite. Tahiti 673
Although probable, strengthening of the reef framework by micritic crusts is difficult to substanti- ate since such frameworks built by massive branching acroporids are strong structures, now restricted to reef zones exposed to rough water conditions. How- ever, rapid lithification of microbial micritic crusts may be deduced both from the presence of borings in them (Fig. 6c) and from their local interlayering with fast growing coralgal communities (Fig. 4) which require hard substrates for their development. It seems likely that these rapidly lithified micritic crusts may have played a prominent role in protecting the reef framework from biological eroders and grazers, thus reducing largely the production of particulate coral material. The calcification of modern microbial fabrics may take place during their growth (see Chafetz & Folk, I984), sometimes eilhancing their accretion rate (Riding, 1991). Such an early calcifica- tion is thought to result in a coarse, unlaminated or at best crudely laminated fabric (Riding, 1991), very reminiscent of the features observed in the Tahiti crusts.
Comparisons with Quaternary analogues
Lithified Mg-calcite crusts exhibiting a peloidal micritic fabric have been described in other Quater- nary reef tracts and may be compared with the crusts, or parts of crusts, reported here. The laminated parts of the Tahiti crusts are similar to the ‘smooth crusts’ of Land (1971), ‘cement crusts’ of MacIntyre (1977), ‘lithified crusts’ of Land & Moore (1980) and Marshall (1983) and ‘laminar micritic crusts’ of Brachert & Dullo (1991). The columnar crusts resemble the ‘knobby crusts’ of Land (1971), the columnar forms from the ‘lithified crusts’ of Marshall (1983) and the columnar microbialites reported by Jones & Hunter (1991) within Pleistocene shallow lagoons from Grand Cayman and San Salvador Island. Most of these examples report a very similar sequence of development, in which micritic crusts form the last stage of encrustation and generally overgrow red algal crusts. None of these examples report the interlayering between coralgal communi- ties and microbial crusts that we have observed in the Tahitian Holocene reefs.
Although some allusions have been made in the past to the resemblance between such micritic crusts and stromatolites (Land & Goreau, 1970; James & Ginsburg, 1979; Land & Moore, 1980), most of these studies remained inconclusive. Marshall (1983) assumed some form of organic influence during the
formation of micritic crusts in the Holocene reef sequence from the Great Barrier Reef, but indicated that the evidence of algal mats in the lithified crusts is circumstantial. Brachert & Dullo (1991) concluded that parts of the crusts they studied are probably biogenic in origin and referred to them as ‘cryptalgal fabrics’. Riding et al. (1991a) assumed that the peloidal micritic crusts reported in Recent reefs may be examples of microbial carbonates surviving to modern times in favourable habitats, but in a form which has tended to be overlooked.
None of the examples cited above report such widespread development of micritic crusts in a high energy reef environment as was observed in the Tahiti Holocene sequence, although Land & Goreau (1 970) and Marshall (1983) reported local micritic crust formation in zones of relatively high energy conditions (e.g. reef crest), immediately beneath the reef-water interface. In contrast, most of micritic crusts have been reported from the walls of the deeper forereef (Ginsburg & James, 1973; Moore et al., 1976; James & Ginsburg, 1979; Land & Moore, 1980; Brachert & Dullo, 1991) and in lagoonal settings in response to deteriorating environmental conditions (Jones & Hunter, I99 1).
The predominantly prokaryotic composition of the micritic crusts reported from Quaternary reef tracts contrasts sharply with that of coarse grained eualgal- cyanobacterial stromatolites, which include a signifi- cant eualgal component in their mats (Awramik & Riding, 1988) and can be expected to have formed in high energy environments (Riding et al., 1991b). This may suggest that, even if eualgal-cyanobacterial stromatolites reflect a change in stromatolitic mat components during the past 100 Ma (Riding et al., 1991 b), there is no clear link between the composition of microbial communities and environmental con- ditions, especially water turbulence. In particular, the tidal flat environment does not represent a refuge for fine grained cyanobacterial dominated stromatolites during the Cenozoic as was previously postulated (Awramik & Riding, 1988). In contrast, the data presented here and from other Quaternary reef tracts suggest that microbial communities may have played a more prominent role in Quaternary reefs than previously recognized.
CONCLUSIONS
Study of drill cores from Holocene barrier reefs from Tahiti (French Polynesia) affords the opportunity

674 G. F. Camoin and L. F. Montaggioni
to document the development of microbial micritic crusts in an open marine reef setting and in association with typical high energy coralgal communities. 1. The reef framework is predominantly composed of massive branching acroporids encrusted by coral- line algae associated with sessile vermetid gastropods and arborescent foraminifers. Microbial micritic crusts form coatings over coral branches or, more commonly, over related encrusting organisms, thus appearing as a major structural and volumetric con- stituent of the framework. 2. The reef communities developed throughout most of Holocene time, from 7140 & 170 yr BP to the present, at depths less than 5 m below mean sea level, in a high energy environment. 3. The nature of the benthic communities, stable isotope data and high calcification rates imply that, at least for the last 7000 years, the local environmen- tal conditions were optimal for reef development at Tahiti. In such conditions, the predominance of microbial communities over true encrusters (red algae, foraminifers) may be due to short term stress- ful periods related to temporary discharge of nutrient-enriched freshwater. 4. Microbial micritic crusts may have played a prominent role in protecting the coralgal framework from bioeroders and grazers and, possibly, in the strengthening of the framework due to their rapid lithification. 5. Since similar microbial crusts have been reported in other Quaternary reef tracts, we assume that microbial communities may have played a more prominent role in Quaternary reefs than presently recognized.
ACKNOWLEDGMENTS
The authors wish to thank B. Salvat and the French Polynesian Office of Works for financial support and drilling facilities. Isotope facilities were provided by J.-Ch. Fontes and M. Pachiaudi; 14C dating was performed by G. Delibrias and M. Lafon. G. Faure and M. Pichon identified the corals and G. Segonzac identified the red algae. We also thank I . G. MacIntyre and M. R. Talbot for constructive reviews and R. Riding for stimulating discussions. Technical assistance was provided by M. Lemoine for thin sections, M. Malea and D. Serrette for photographs, R. Notonier for S. E. M. analysis and G. Haufeurt for typing the tables.
REFERENCES ADEY, W.H. (1986) Coralline algae as indicators of sea level.
In: Sea-Level Research: A Manual for the Collection and Evaluation oj’Data (Ed. by 0. Van de Plassche), pp. 229- 280. Geo Books, Norwich, England.
AHARON, P. (1985) Carbon isotope record of Late Quater- nary coral reefs: possible index of sea surface paleopro- ductivity. In: The Carbon Cycle and Atmospheric CO,: Nutural Variations Archean to Present (Ed. by E. T. Sundquist & W. S. Broecker), pp. 343-356. American Geophysical Union.
AHARON, P. (1991) Recorders of reef environment histories: stable isotopes in corals, giant clams, and calcareous algae. In: Reefs as Recorders of Environmental Changes (Ed. by L. F. Montaggioni & I. G. MacIntyre). Spec. Issue Coral Reefs, 10, 71-90.
AWRAMIK, S.M. & RIDING, R. (1988) Role of algal eukary- otes in subtidal columnar stromatolite formation. Proc. Natl. Acad. Sci. USA, 85, 1321-1329.
BRACHERT, Y.C. & DULLO, W.C. (1991) Laminar micrite crusts and associated foreslope processes, Red Sea. J. sedim. Petrol., 61, 354-363.
BROWN, B.E., TUDHOPE, A.W., LE TISSIER, M.D.A. & SCOFFIN, T.P. (1991) A novel mechanism for iron incor- poration into coral skeletons. Coral Reefs, 10, 21 1-215.
BUCHBINDER, B. (1981) Morphology, microfabric and origin of stromatolites of the Pleistocene precursor of the Dead Sea, Israel. In: Phanerozoic Stromatolites (Ed. by C. L. V. Monty), pp. 181-186. Springer-Verlag, Berlin.
CAMOIN, G. (1983) Plates-formes carbonatees et recifs a rudistes du Cretace de Sicile. Trav. Lab. Giol. Hist. Pal., 13.
CAMOIN, G.F. & MONTAGGIONI L.F. (1993) Stromatolites associated with coralgal communities in Holocene high- energy reefs. Reply. Geology, 21, 956.
CAYEUX, L. (1935) Les Roches Sedimentaires de France. Masson, Paris.
CHAFETZ, H.S. (1986) Marine peloids: a product of bacteri- ally induced precipitation of calcite. J. sedim. Petrol., 56, 8 12-8 17.
CHAFETZ, H.S. & FOLK, R. L. (1984) Travertines: depo- sitional morphology and the bacterially constructed elements. J. sedim. Petrol., 54, 289-316.
CUET, P., NAIM, O., FAURE, G. & CONAN, J.Y. (1988) Nutrient-rich groundwater impact on benthic communi- ties of La Saline fringing reef (Reunion Island, Indian Ocean): preliminary results. Proc. Sixth Int. Coral Reef Symposium, Townsville, 2, 207-21 2.
DAVIES, P.J., MARSHALL, J.F. & HOPLEY, D. (1985) Rela- tionships between reef growth and sea level in the Great Barrier Reef. Proc. Fifth Int. CoralReef Cong., 3, 95-103.
DELESALLE, B., GALZIN, R. & SALVAT, B. (Eds) (1985) French Polynesian coral reefs. Proc. Fifth Int. Coral Reef Cong. 1, 1-554.
DE RIDDER, C. (1986) Echinides. In: Guide des Ctoiles de mer, oursins et autres Pehinodermes du lagon de Nouvelle Calidonie (Ed. by A. Guille et al.). Office de Recherche Scientifique et Technique d’Outre-Mer.
DILL, R.F., SHINN, E.A., JONES, A.T. KELLY K. & STEINEN, R.P. (1986) Giant subtidal stromatolites forming in normal salinity waters. Nature, 324, 55-58.
DRAVIS, J.J. (1983) Hardened subtidal stromatolites, Bahamas. Science, 219, 385-386.

Coralgal-stromatolite, Tahiti 675
DUNBAR, R.B. & WtLLlNGTON, G.M. (1981) Stable iso- topes in a branching coral monitor seasonal temperature variation. Nature, 293, 543-545.
FAURE, G., (1 982) Recherches sur les peuplemrnts de .sclPrac- tiniaires des rgclfs coralliens de l’archipel des Mascareignes (OcPan Indien occidental). DSc thesis, University of Marseille, France.
FOLK, R.N. & LAND, L.S. (1975) Mg/Ca ratio and salinity: two controls over crystallization of dolomite. Bull. Am. Ass. petrol. Geol.. 59, 60-68.
GINSBURG, R.N. & JAMES, N.P. (1973) British Honduras by Submarine. Geotimes, 18, 23-24.
GOLUBIC, S. (1991) Modern stromatolites: a review. In: Calcareous Algae and Stromutolites (Ed. by R. Riding), pp. 541 -561. Springer-Verlag, Berlin.
JAMES, N.P. & GINSBURG, R.N. (1979) The seaward margin of Belize barrier and atoll reefs. Spec. Puhl. int. Ass. Sediment. 3.
JONES, B. & HUNTER, I.G. (1991) Corals to rhodolites to microbialites - A community replacement sequence indicative of regressive conditions. Pulaios 6, 54-66.
JONES, B. MACDONALD, R.W. (1989) Micro-organisms and crystal fabrics in cave pisoliths from Grand Cayman, British West Indies. J. sedim. Petrol., 59, 387-396.
KENNARU, J.M. & JAMES, N.P. (1986) Thrombolites and stromatolites; two distinct types of microbial structures. Paluios. 1, 492-503.
KINSEY, D.W. (1983) Standards of performance in coral reef primary production and carbon turnover. I n : Perspectives on Coral Reefs (Ed. by D. J. Barnes), Austral. Inst. Mur. Sci. Contr., 200, 209-220.
KRUMBEIN, W.E. (1974) On the precipitation of aragonite on the surface of marine bacteria. Nuturwiss., 61, 167.
KRUMBEIN, W.E. & COHEN, Y . (1977) Primary production, mat formation and lithification chances of oxygenic and facultative anoxygenic cyanophytes (cyanobacteria). I n : Fos.sil Algae (Ed. by E. Flugel), pp. 37-56. Springer- Verlag, Berlin.
LAND, L.S. (1971) Submarine lithification of Jamaican reefs. In: Carbonate Cements (Ed. by 0. P. Bricker), pp. 59-62. John Hopkins University Press, Baltimore.
LAND, L.S. & GOREAU, T.F. (1970) Submarine lithification of Jamaican reefs. J. .sedim. Petrol., 40, 457462.
LAND, L.S. & MOORE, C.H. (1980) Lithification, micritiza- tion and syndepositional diagenesis of biolithites on the Jamaican island slope. J. sedim. Petrol., 50, 357 -370.
LEA, D.W., SHEN, G.T. & BOYLE, E.A. (1989) Coralline barium records temporal variability in equatorial Pacific upwelhng. Nature, 340, 373-376.
LIGHTY, R.G. (1985) Preservation of internal reef porosity and diagenetic sealing of submerged early Holocene bar- rier reef, Southeast Florida Shelf. In: Carbonate Cements (Ed. by N. Schneiderman & P. Harris), Spec. Puhl. Soc. Econ. Paleont. Miner., 36, 123-151.
MACCONNAUGHEY, T.A. (1989) I3C and ‘‘0 isotopic dis- equilibrium in biological carbonates: I-Patterns. Grochim. Cosmochim. Acta, 53, 15 1-162.
MACINTYRE, I.G. (1977) Distribution of submarine cements in a modern Caribbean fringing reef, Galeta Point, Panama. J. sedim. Petrol., 40, 503-516.
MACINTYRE,, I.G. (1984) Extensive submarille lithitication in a cave in the Belize Barrier Reef Platform. J. sedim. Petrol., 54. 221-235.
MACINTYW I.G. (1985) Submarine cements - the peloidal question. In: Carbonate Ccments (Ed. by N. Schneiderman & P. Harris), Spec. Publ. Soc. won. Paleont. Miner., 36, 109-1 16.
MACINTYRE, I.G. & MARSHALL, J.F. (1989) Submarine lithification in coral reefs: some facts and misconceptions. Proc. Sixth Int. Coral Reef Symposium, Townsville, 1, 263 272.
MARSHALL, J.F. (1983) Submarine cementation in a higher- energy platform reef: One Tree Reef, southern Great Barrier Reef. J. sedim. Petrol., 53, 1133-1 149.
MONTAGGIONI, L.F. (1988) Holocene reef growth history in midplate, high volcanic islands. Proc. Sixth Int. C’ol-a1 Reef Symposium, Torvnsville, 3, 455-460.
MONTAGGIONI, L.F. & CAMOIN, G.F. (1993) Stromatolites associated with coralgal communities in Holocene high- energy reefs. Geology, 21, 149-152.
MONTAGGIONI, L.F., CUET, P. & NAIM, 0. (1993) Effect of nutrient excess on a modern fringing reef (Reunion Island), Western Indian Ocean, geological implications. Global Aspects of Coral reefi, Case histories, Miami, N27TN33.
MONTAGGIONI, L.F., NAIM, 0. & CUET, P. (1989) Enregis- trement sedimentaire de I’eutrophisation sur les recifs coralliens actuels. Zime Congr. Fr. Skdim., Puris, 201.
MONTY, C.L.V. (1967) Distribution and structure of Recent stromatolitic algal mats, Eastern Andros Island, Bahamas. Ann. Soc. gPol. Belg., 90, 55-100.
MONTY, C.L.V. (1973) Precambrian background and Phanerozoic history of stromatolitic communities, an overview. Ann. Soc. gPol. Belg., 88, 269-276.
MONTY, C.L.V. (1976) The origin and development of cryptalgal fabrics. In: Stromutolites (Ed. by M. R. Walter), Derel. Sedimentol., 29, 193-249.
MOORE, C.H., GRAHAM, E.A. & LAND, L.S. (1976) Sediment transport and dispersal across the deep fore-reef and island slope ( - 55 m to - 305 m), Discovery Bay, Jamaica. J . .serlim. petrol., 46, 147-187.
NELSON, S.G. (1 985) Immediate enhancement of photosyn- thesis by coral reef macrophytes in response to ammonia enrichment. Proc. Fifth Coral Reef’ Cong., Tuhiti, 3, 105-1 10.
NEUMANN, A.C. & MACINTYRE, I.G. (1985) Reef response to sea level rise: keep-up, catch-up or give-up. Proc. Fifth Int. Coral re@/ Cong., 3, 105-1 10.
PATZOLD, J. (1984) Growth rhythms recorded in stable isotopes and density bands in the reef coral Porites lohata (Cebu, Philippines). Coral Reefy, 3, 87-90.
PIRAZZOLI, P.A. & MONTAGGIONI, L.F. (1988) The 7000 years sea level curve in French Polynesia: geodynamic implications for mid-plate volcanic islands. Proc. Sixth Int. Coral Reef Symposium, Toivnsville, 3, 467-472.
PRATT, B.R. (1982) Stromatolite decline - A reconsidera- tion. Geology, 10, 512-515.
RASMUSSEN, K.A., MACINTYKE, I.G. & PRUFERT, L. (1993) Modern stromatolite reefs fringing a brackish coastline, Chetumal Bay, Belize. Geology, 21, 199-202.
REID, R.P. & BROWN, K.M. (1991) Intertidal stromatolites in a fringing Holocene reef complex, Bahamas. Geology, 19, 15-18.
RICHARD, G. (1 982) Mollusques lagunuires et rPi,ifuu.r de PolynPsie Franpise: inventaire,faunistiyur. bionomie, hilan

676 G. l? Carnoin und L. l? Montuggioni
yuantitatq; croi.ssance, production. DSc thesis, University of Paris.
RIDING, R. (1991) Classification of microbial carbonates. In: Calcareous A l p e and Stromatolites (Ed. by R. Riding), pp. 21-51, Springer-Verlag, Berlin.
RIDING R. (1992) Temporal variation in calcification in marine cyanobacteria. J. geol. Soc., London, 149, 979- 989.
RIDING, R., AWRAMIK, S.M., WINSBOKOUGH, B.M., GRIPFIN, K.M. & DILL, R.F. (1991b) Bahamian giant stromatolites: microbial composition and surface mats. Geol. Mag., 128, 227-234.
RIDING, R., MAKTIN, J.M. & BKAGA, J.C. (1991a) Coral- stromatolite reef framework, Upper Miocene, Almeria, Spain. Sedimcvitology, 38, 799-8 18.
RIUING, R. & VORONOVA, L. (1982) Recent freshwater oscillatoriacean analogue of the Lower Paleozoic calcar- eous alga Angulocellularia. Lethaia, 15, 105-1 14.
SCHOPF, J.W. (1983) Earth’s Earliest Biosphere. Princeton University Press, New Jersey.
SHEN, G.T. & BOYLE E.A. (1987) Lead in corals: reconstruc- tion of historical fluxes to the surface ocean. Eurth planet. Sci. Lett., 82, 289-304.
SHEN, G.T, BOYLE, E.A. & LEA, D.W. (1987) Cadmium in corals as a tracer of historical upwelling and industrial fallout. Nuture, 328, 794-795.
SHEN, G.T., CAMPBELL, T.M., DUNBAK, R.B., WELLINGTON, G.M., COLGAN, M.W. & GLVNN, P.W. ( I 991) Paleochem- istry of manganese in corals from the Galagapos islands. In: Reefi as Recorders of Environmental Changes (Ed. by L. F. Montaggioni & I. G. MacIntyre). Spec. Issue Coral ReeJs, 10, 91 -100.
SHINN, E.A. (1987) Sand castles from the past: Bahamian stromatolites discovered. Sea Frontiers, 33, 334-343.
SMITH, S.V. (1981) The Houtman Abrolhos islands: carbon metabolism of coral reefs at high latitude. Limnol. Oceanogr., 26, 6 1 2-62 1 .
SMITH, S.V. (1983) Coral reef calcification. In: Perspectives in Coral Reefs (Ed. by D. J. Barnes) Austral. lnst. Mar. Sci. Contr. 200, 240-247.
SMITH, S.V. & KINSEY, D.W. (1976) Calcium carbonate production, coral reef growth, and sea level change. Science, 194, 937-939.
SMITH, S.V. & KROOPNICK, P. (1981) Carbon-13 isotopic fractionation as a measure of aquatic metabolism. Nature, 294, 252-253.
SWART, P.K. (1983) Carbon and oxygen isotope fraction- ation in scleractinian corals: A review. Earth Sci. Rev., 19, 51-80.
SWART, P.K. & COL~MAN, M.L. (1980) Isotopic data for scleractinian coral explain their paleotemperature uncertainties. Nalure. 283, 557-559.
VERON, J.E.N. (1986) Corals of’ Australia and the lndo- Pacijic. Angus & Robertson, London.
WALTER, M.R. (1972) Stromatolites and the biostratigraphy of the Australian Precambrian and Cambrian. Palaeon- tology Spec. Pup., 11.
WEBEK, J. & WOODHEAD, P.M.J. (1972) Temperature dependence of oxygen-I 8 concentration in reef coral carbonates. J geophys. Res., 77, 463473.
WEIL S.M., BUDDEMEIEK, R.W., SMITH, S.V. KROOPNICK, P.M. (1981) The stable isotopic composition of coral skeletons: control by environmental variables. Geochirn. Cosmorhim. Acta, 45, 1147-1 153.
(Manuscript received 3 February 1993; revision accepted 6 January 1994)




















