
Hepatitis B virus mutations in the pre-S genome before and after liver transplantatio
7
Hepatitis B Virus Mutations in the Pre-S Genome Before and After Liver Transplantation CHRISTIAN TRAUTWEIN, 1 HARALD SCHREM, 1 HANS L. TILLMANN, 1 STEFAN KUBICKA, 1 DIANA WALKER, 1 KLAUS H. W. BO ¨ KER, HANS-JO ¨ RG MASCHEK, 2 RUDOLPH PICHLMAYR, 3 AND MICHAEL P. MANNS 1 laxis with hepatitis B surface antigen antibody (anti-HBs) Mutational changes in the pre-S region of hepatitis B (hepatitis B immune globulin [HBIg]). This is most effective virus (HBV) were analyzed in 20 patients who experi- if serum HBV is below 1 pg/mL before OLT. 5-7 HBIg consists enced HBV reinfection after orthotopic liver trans- of polyclonal antibodies directed against the viral envelope plantation (OLT). HBV DNA was extracted from patient and is obtained from anti-HBs – positive donors. 6 Interest- sera before and after OLT. The pre-S sequence was am- ingly, in some patients, HBV reinfection is observed while plified via polymerase chain reaction, subcloned, se- receiving HBIg. One of the reasons for HBV reinfection dur- quenced, and analyzed. In 18 of 20 patients, mutational ing HBIg immunoprophylaxis could be escape mutations se- changes were found in the pre-S region pre- or post- lected by HBIg. Mutations are frequently found and selected OLT; 11 showed point mutations (1-10) and 7 cases major in the HBV genome because the virus replicates via an inter- changes (insertions/deletions). For the point mutations, mediate RNA step that lacks proofreading capacity. 8,9 there was no trend in the selection of wild-type (wt) HBV The pre-S region is believed to mediate hepatocyte attach- before or after OLT in the pre-S region. Additional HBV ment, contains B- and T-cell epitopes, and consists of regula- reinfection during hepatitis B surface antigen antibody tory elements that control env gene transcription. 10-17 Dele- (anti-HBs) administration had no influence on selection tions and insertions of the pre-S region were found in chronic pressure in the pre-S region. In contrast, insertions/dele- carriers, and they may have an impact on the progression of tions were more frequently found before OLT. In the 7 chronic HBV infection. 18,19 It is unclear whether these muta- patients with deletions/insertions, changes in the hepa- tions of the pre-S region might be involved in the poor out- tocyte attachment site were not seen after OLT. Interest- come after OLT and HBV reinfection. Alternatively, deletions ingly, the only patient with changes in a major virus and insertions of the pre-S region might interfere with HBV population after OLT had changes in the CCAAT-box reinfection of the graft, because the pre-S region is believed to of the S-promoter. As shown by gel shift analysis, this be involved in hepatocyte attachment. Additionally, selection mutation was associated with loss of specific binding to pressure of HBIg could create new mutants in the pre-S re- this element and thus probably led to dysregulation of gion after OLT. Thus, studies of the pre-S region before and S-gene transcription. Major changes in the pre-S genome after OLT could provide new clues for the prevention of HBV are mainly seen before OLT, and HBV reinfection does reinfection after OLT and new insights into the pathogenesis occur with the intact hepatocyte attachment sites after of chronic HBV infection. Therefore, 20 patients with HBV OLT. Anti-HBs (hepatitis B immune globulin [HBIg]) cre- reinfection of the graft after OLT were included in this study. ates no selection pressure on the pre-S region. The muta- All patients were transplanted for HBV-related end-stage tion in the CCAAT-box of the S-promoter potentially liver disease. Mutational changes in the pre-S genome were leads to its dysregulation and may be associated with investigated before and after OLT. the occurrence of fibrosing cholestatic hepatitis after OLT. (HEPATOLOGY 1996; 24:482-488.) PATIENTS AND METHODS Patients. Twenty patients experiencing HBV reinfection of the The last treatment option for patients with end-stage liver graft after OLT for HBV-related end-stage liver disease were in- disease or fulminant hepatic failure associated with hepatitis cluded in this study (Table 1). OLT of these patients was performed B virus (HBV) infection is orthotopic liver transplantation between 1985 and 1992. HBV reinfection was defined by the reap- (OLT). 1 HBV reinfection after OLT leads to graft dysfunction, pearance of the hepatitis B surface antigen, which was always associ- ated with positive polymerase chain reaction results for the pre-S retransplantation, acute or chronic hepatitis, and cirrhosis and C region of the HBV genome. Except for patients 2 and 14, all as early as 1 year after OLT. 2-4 The only effective treatment patients obtained HBIg prophylaxis after OLT. Routinely, 2 1 10,000 to reduce the rate of reinfection is long-term immunoprophy- IU were administered in the anhepatic phase, and 10,000 IU was administered each day for up to 1 week after surgery. 6 At later time points, anti-HBs titers were tested and HBIg was given only when titers fell below 100 IU/L. HBIg was administered for at least 3 Abbreviations: HBV, hepatitis B virus; OLT, orthotopic liver transplantation; anti-HBs, months (Fig. 1) or until patients became reinfected. hepatitis B surface antigen antibodies; HBIg, hepatitis B immune globulin; wt, wild-type; The patients had an average age of 45.35 years at the time of aa, amino acids; mRNA, messenger RNA. their first OLT. Sixteen of 20 patients (80%) were men. Two patients From the 1 Department of Gastroenterology and Hepatology, 2 Department of Pathology, and 3 Department of Surgery, Medizinische Hochschule Hannover, Hannover, Germany. (patients 6 and 7) were also infected with the hepatitis C virus before Received November 21, 1995; accepted April 29, 1996. and after OLT. A third patient (patient 18) was coinfected with the Presented in part at the 46th Annual Meeting of the American Association for the Study hepatitis D virus before OLT and experienced hepatitis D virus rein- of Liver Diseases, Chicago, IL. fection before HBV reinfection after OLT. In 7 of 20 patients (35%) Supported by BMFT-grant 01 Kl 9554 and SFB 265-C5. (Table 1) included in this study, hepatocellular carcinoma was found Address reprint requests to: M. P. Manns, M.D., Department of Gastroenterology and in the explanted liver. Patient 13 was retransplanted 7 years after Hepatology, Medizinische Hochschule Hannover, Konstanty-Gutschow-Str. 8, D-30625 his first transplantation because of the recurrence of hepatocellular Hannover, Germany. carcinoma in the first graft. This patient is still alive 13 years after Copyright q 1996 by the American Association for the Study of Liver Diseases. 0270-9139/96/2403-0003$3.00/0 his first OLT and works as a teacher (Fig. 1 and Table 1). 20 Patient 482 AID Hepa 0016 / 5p12$$$301 08-07-96 09:22:07 hptas WBS: Hepatology
-
Upload
mh-hannover -
Category
Documents
-
view
2 -
download
0
Transcript of Hepatitis B virus mutations in the pre-S genome before and after liver transplantatio

Hepatitis B Virus Mutations in the Pre-S Genome Before andAfter Liver Transplantation
CHRISTIAN TRAUTWEIN,1 HARALD SCHREM,1 HANS L. TILLMANN,1 STEFAN KUBICKA,1 DIANA WALKER,1 KLAUS H. W. BOKER,HANS-JORG MASCHEK,2 RUDOLPH PICHLMAYR,3 AND MICHAEL P. MANNS1
laxis with hepatitis B surface antigen antibody (anti-HBs)Mutational changes in the pre-S region of hepatitis B(hepatitis B immune globulin [HBIg]). This is most effectivevirus (HBV) were analyzed in 20 patients who experi-if serum HBV is below 1 pg/mL before OLT.5-7 HBIg consistsenced HBV reinfection after orthotopic liver trans-of polyclonal antibodies directed against the viral envelopeplantation (OLT). HBV DNA was extracted from patientand is obtained from anti-HBs–positive donors.6 Interest-sera before and after OLT. The pre-S sequence was am-ingly, in some patients, HBV reinfection is observed whileplified via polymerase chain reaction, subcloned, se-receiving HBIg. One of the reasons for HBV reinfection dur-quenced, and analyzed. In 18 of 20 patients, mutationaling HBIg immunoprophylaxis could be escape mutations se-changes were found in the pre-S region pre- or post-lected by HBIg. Mutations are frequently found and selectedOLT; 11 showed point mutations (1-10) and 7 cases majorin the HBV genome because the virus replicates via an inter-changes (insertions/deletions). For the point mutations,mediate RNA step that lacks proofreading capacity.8,9there was no trend in the selection of wild-type (wt) HBV
The pre-S region is believed to mediate hepatocyte attach-before or after OLT in the pre-S region. Additional HBVment, contains B- and T-cell epitopes, and consists of regula-reinfection during hepatitis B surface antigen antibodytory elements that control env gene transcription.10-17 Dele-(anti-HBs) administration had no influence on selectiontions and insertions of the pre-S region were found in chronicpressure in the pre-S region. In contrast, insertions/dele-carriers, and they may have an impact on the progression oftions were more frequently found before OLT. In the 7chronic HBV infection.18,19 It is unclear whether these muta-patients with deletions/insertions, changes in the hepa-tions of the pre-S region might be involved in the poor out-tocyte attachment site were not seen after OLT. Interest-come after OLT and HBV reinfection. Alternatively, deletionsingly, the only patient with changes in a major virusand insertions of the pre-S region might interfere with HBVpopulation after OLT had changes in the CCAAT-boxreinfection of the graft, because the pre-S region is believed toof the S-promoter. As shown by gel shift analysis, thisbe involved in hepatocyte attachment. Additionally, selectionmutation was associated with loss of specific binding topressure of HBIg could create new mutants in the pre-S re-this element and thus probably led to dysregulation ofgion after OLT. Thus, studies of the pre-S region before andS-gene transcription. Major changes in the pre-S genomeafter OLT could provide new clues for the prevention of HBVare mainly seen before OLT, and HBV reinfection doesreinfection after OLT and new insights into the pathogenesisoccur with the intact hepatocyte attachment sites afterof chronic HBV infection. Therefore, 20 patients with HBVOLT. Anti-HBs (hepatitis B immune globulin [HBIg]) cre-reinfection of the graft after OLT were included in this study.ates no selection pressure on the pre-S region. The muta-All patients were transplanted for HBV-related end-stagetion in the CCAAT-box of the S-promoter potentiallyliver disease. Mutational changes in the pre-S genome wereleads to its dysregulation and may be associated withinvestigated before and after OLT.the occurrence of fibrosing cholestatic hepatitis after
OLT. (HEPATOLOGY 1996;24:482-488.)PATIENTS AND METHODS
Patients. Twenty patients experiencing HBV reinfection of theThe last treatment option for patients with end-stage liver graft after OLT for HBV-related end-stage liver disease were in-disease or fulminant hepatic failure associated with hepatitis cluded in this study (Table 1). OLT of these patients was performedB virus (HBV) infection is orthotopic liver transplantation between 1985 and 1992. HBV reinfection was defined by the reap-(OLT).1 HBV reinfection after OLT leads to graft dysfunction, pearance of the hepatitis B surface antigen, which was always associ-
ated with positive polymerase chain reaction results for the pre-Sretransplantation, acute or chronic hepatitis, and cirrhosisand C region of the HBV genome. Except for patients 2 and 14, allas early as 1 year after OLT.2-4 The only effective treatmentpatients obtained HBIg prophylaxis after OLT. Routinely, 21 10,000to reduce the rate of reinfection is long-term immunoprophy-IU were administered in the anhepatic phase, and 10,000 IU wasadministered each day for up to 1 week after surgery.6 At later timepoints, anti-HBs titers were tested and HBIg was given only whentiters fell below 100 IU/L. HBIg was administered for at least 3
Abbreviations: HBV, hepatitis B virus; OLT, orthotopic liver transplantation; anti-HBs,months (Fig. 1) or until patients became reinfected.hepatitis B surface antigen antibodies; HBIg, hepatitis B immune globulin; wt, wild-type;
The patients had an average age of 45.35 years at the time ofaa, amino acids; mRNA, messenger RNA.their first OLT. Sixteen of 20 patients (80%) were men. Two patientsFrom the 1Department of Gastroenterology and Hepatology, 2Department of Pathology,
and 3Department of Surgery, Medizinische Hochschule Hannover, Hannover, Germany. (patients 6 and 7) were also infected with the hepatitis C virus beforeReceived November 21, 1995; accepted April 29, 1996. and after OLT. A third patient (patient 18) was coinfected with thePresented in part at the 46th Annual Meeting of the American Association for the Study hepatitis D virus before OLT and experienced hepatitis D virus rein-
of Liver Diseases, Chicago, IL. fection before HBV reinfection after OLT. In 7 of 20 patients (35%)Supported by BMFT-grant 01 Kl 9554 and SFB 265-C5. (Table 1) included in this study, hepatocellular carcinoma was foundAddress reprint requests to: M. P. Manns, M.D., Department of Gastroenterology and
in the explanted liver. Patient 13 was retransplanted 7 years afterHepatology, Medizinische Hochschule Hannover, Konstanty-Gutschow-Str. 8, D-30625his first transplantation because of the recurrence of hepatocellularHannover, Germany.carcinoma in the first graft. This patient is still alive 13 years afterCopyright q 1996 by the American Association for the Study of Liver Diseases.
0270-9139/96/2403-0003$3.00/0 his first OLT and works as a teacher (Fig. 1 and Table 1).20 Patient
482
AID Hepa 0016 / 5p12$$$301 08-07-96 09:22:07 hptas WBS: Hepatology

HEPATOLOGY Vol. 24, No. 3, 1996 TRAUTWEIN ET AL. 483
TABLE 1. Characteristics of Patients Included Into the Study
Patient No: 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Time between first HBV-diagnosisand OLT (y) ? 13 5 6 3 14 7 5 ? ? 2 11 7 ? 12 0.5 6 8 ? 3
Age at the time of OLT (y) 60 48 34 60 49 56 45 47 59 47 47 32 41 49 53 24 51 50 42 13Number of re-OLTs 1 2 1Hepatocellular carcinoma in the
explanted liver No No No No Yes No No No Yes Yes No Yes Yes No No No No Yes Yes NoHBsAg pre-OLT / / / / / / / / / / / / / / / / / / / /HBeAg pre-OLT 0 0 / / / 0 / / 0 / / 0 / 0 / / / 0 / /Anti-HBe pre-OLT / / 0 0 0 / 0 0 / 0 0 / 0 0 0 0 0 / / 0HBV-DNA dot-blot pre-OLT / / / / 0 0 0 0 0 / 0 0 0 0 0 / 0 0 / /HBsAg post-OLT, at reinfection / / / / / / / / / / / / / / / / / / / /HBeAg post-OLT, at reinfection 0 0 / / / 0 0 / 0 / / / / ND / / / 0 / /HBeAg post-OLT, 2.sequence 0 0 / / / / / / 0 / / / / / / / / 0 / /HBV-DNA PCR pre-OLT / / / / / / / / / / / / / / / / / / / /Anti-HCV pre-OLT 0 0 0 0 0 ? ? 0 0 0 0 0 0 0 0 0 0 0 0 0Anti-HCV post-OLT 0 0 0 0 0 / / 0 0 0 0 0 0 0 0 0 0 0 0 0Anti-HDV pre-OLT 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 / 0 0Anti-HDV post-OLT 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 / 0 0
NOTE: Twenty patients were included in the study. Some of the patients had characteristics besides end-stage liver disease for chronic hepatitis B infection.The different characteristics are listed.
Abbreviations: OLT, orthotopic liver transplantation; HBV, hepatitis Be virus; HBsAg, hepatitis Be surface antigen; HBeAg, hepatitis B antigen; HCV,hepatitis C virus; ND, not done.
20 was also retransplanted once, and patient 19 was retransplanted according to the manufacturer’s instructions. Insert size was deter-mined after EcoR1 digestion in a 1% agarose-electrophoresis gel.twice. Patient 16 was treated by OLT after orthotopic heart trans-
plantation. Sequencing of the clones was performed dependent on the migra-tion pattern in the gel. Four clones were sequenced in a equimolarSerological Assays. Serum samples were aliquoted and stored at
0207C until defrosted for testing. Hepatitis B surface antigen, anti- mix if their insert had the same molecular weight. All clones withaberrant insert size were sequenced. As well, single-clone sequencingHBs, antibodies against the hepatitis B core antigen, hepatitis C
virus antibodies, and antibodies against hepatitis D virus were tested was performed, if double bands appeared in the sequence of theequimolar mix as described before.23 Sequencing was performed us-by commercial enzyme immunoassays (Abbott Laboratories, Chi-
cago, IL). Samples positive for hepatitis C virus antibodies were ing the sequenase sequencing kit (United States Biochemicals;Braunschweig, Germany) according to the manufacturer’s instruc-tested for HCV RNA by polymerase chain reaction, as described be-
fore.21 Quantitative HBV-DNA assays were performed with the Ab- tions.The following primers were used for sequencing: 5*-GGG TCA CCAbott Hepatitis B viral DNA kit (Abbott Laboratories).
Extraction of DNA From Serum and Polymerase Chain Reaction of TAT TCT TG-3*, 5*-CCT GAT GTG ATG TTC TCC ATG-3*, 5*-CTACCT TGT TGG CGT CTG GCC-3*, 5*-CAT GTT CAG CGC AGG GTCthe Pre-S Genome. DNA was extracted from sera22 collected both be-
fore OLT and shortly after evidence for reinfection of the graft. CCC-3*, and 5*-TCC TGA CTG GCG ATT GGT GG-3*.Preparation of Nuclear Extracts. Six- to 9-week-old Sprague-Daw-Ninety microliters of serum was treated with 10 mL of proteinase K
and 300 mL of DNA-lysis buffer (0.67% sodium dodecyl sulfate, 13.33 ley rats were obtained from the animal facility of the MedizinischeHochschule Hannover. The animals were maintained on a 12-hourmmol/L Tris [pH 8.0], 6.6 mmol/L ethylenediaminetetraacetic acid
[pH 8.0], and 13.3 mg/mL transfer RNA). The mixture was incubated night and 12-hour light schedule. Twelve hours before surgery, foodwas withdrawn from the animals’ cages. Surgery was performed be-at 567C for 2 hours. Four hundred microliters of phenol:chloro-
form:isoamylalcohol (25:24:1) was added and briefly vortexed. The tween 8 and 10 AM. Anesthesia was performed by an intraperitonealinjection of a combination of rompun/ketamine. After a small subxy-mixture was centrifuged at 13,000 rpm for 30 minutes at 47C. The
DNA in 300 mL of the upper phase was precipitated by adding 660 phoid incision, the liver was removed and rinsed in ice-cold phos-phate-buffered saline. Four rats were used in parallel, and the liversmL of 100% ethanol and 30 mL of 3 mol/L NaOAc, pH 5.2, and was
centrifuged at 13,000 rpm for 30 minutes at 47C. After briefly wash- were pooled. Liver nuclear extracts were prepared essentially asdescribed by Descombes et al.24 All steps were performed at 47C.ing the pellet with 70% ethanol, the pellet was dried and resuspended
in 25 mL TE-buffer. Gel Retardation Assays. For the gel retardation assays, nuclearextracts from Sprague-Dawley rats were used as indicated. The bind-Polymerase chain reaction was performed using the following
primers: pre-S2: 5*-GGG TCA CCA TAT TCT TG-3*; pre-S4: 5*-CCT ing reaction was performed for 20 minutes on ice. For binding assays,an oligonucleotide spanning the region around the wt CCAAT-motifGAT GTG ATG TTC TCC ATG-3*. Five microliters of the resus-
pended DNA pellet was added to 95 mL of the following reaction mix: or the mutation found in patient 17, as indicated in Fig. 3A, wereused as a P32-labeled probe. Free DNA and DNA-protein complexes0.60 mmol/L primer pre-S2; 0.55 mmol/L primer pre-S4; 100 mmol/L
for each dNTP; 1.5 mmol/L MgCL2; 20 mmol/L Tris-HCl (pH 8.4); were resolved on a 6% polyacrylamide gel as described previously.25
Gels were dried and exposed for autoradiography.50 mmol/L KCl; and 25 mU Taq-polymerase. DNA amplification wasperformed using a thermocycler (Crocodile; Appligene) (Illkirch Ger-
RESULTSmany) using the following program: 2 minutes at 947C for one cycle;1 minute at 947C, 1 minute at 507C, and 1 minute 30 seconds at 727C Prevalence of Mutations. At least four clones were se-for 40 cycles; and 3 minutes at 727C for a final extension step. Ten quenced from each patient before and after OLT. The nucleo-microliters of the reaction mix was analyzed in a 1% agarose gel for tide sequence was translated into amino acids. A wild-typePCR products. For each sample, a negative control was treated in (wt) HBV consensus protein sequence for the pre-S genomeparallel, starting at the extraction step to lower the probability of was determined by combining sequence information of fivecross-contamination. Additionally, a positive control was added. In published wt HBV strains.8,26-29 According to Mimms,30 thecase of detected cross-contamination, all samples were discarded.
pre-S sequence was used to determine the subtype of theSubcloning and Sequencing of PCR Products. Subcloning of PCRHBV genome. Seventy percent of all 20 patients included inproducts was performed by using the TA cloning kit (Invitrogen, Santhe study had the subtype, adw, while 6 of 20 (30%) had theDiego, CA). White/blue colony selection was used to detect vectors
containing PCR products. All reaction conditions were performed subtype, ayw.
AID Hepa 0016 / 5p12$$$302 08-07-96 09:22:07 hptas WBS: Hepatology

484 TRAUTWEIN ET AL. HEPATOLOGY September 1996
HBV-DNA levels in patient sera were determined beforeand after OLT at the time of sequencing. Before OLT, theHBV-DNA level of all patients included was 69 { 235 pg/mL;after OLT, it was 180 { 306 pg/mL. The changes betweenthe pre- and post-OLT DNA levels were not significant.
Mutations were compared in each patient pre- to post-OLTand with the wt sequence. Eighteen of 20 patients showedmutations in the pre-S genome. In 11 patients, only pointmutations (between 1 and 10) were found, and, in 7 patients,major changes (deletions/insertions) were found. In the 11patients with point mutations, there was no trend versusthe selection of wt pre-S HBV either pre- or post-OLT. Fourpatients predominantly selected the wt sequence after OLT,4 before OLT and 3 patients selected the same numbers ofwt sequences before and after OLT.
Figure 2 shows the patients with insertions and/or dele-tions before and after OLT. These findings occurred in majorvirus populations—defined as ¢75% of the clones—in fivepatients. In four patients, these changes were found pre-OLT,but were found only in one patient post-OLT (Table 2), whichindicates that these changes occur more frequently in chroniccarriers than after HBV reinfection of the graft. This observa-
FIG. 2. Insertions and deletions found in the pre-S genome. In 7 out of 20patients, insertions and/or deletions were found in the pre-S genome. Changesare shown for each patient. The DNA sequence was translated into the aminoacid sequence, and the serotype was determined. The corresponding wt aminoacid sequence was compared with the sequence found in each clone. Deletionsand insertions are marked. The frequency of specific mutations is shown inthe virus population before and after OLT.
tion is strengthened by the fact that most of the insertions/deletions were found in regions that have been shown to me-diate the interplay with the immune system, and thereforeare probably the result of immune escape mechanisms (Fig.2).
The same deletion pre- and post-OLT was only found inpatient 1. Before OLT, in all the clones, a deletion of six aminoacids was detected between aa135 and 140. Two human B-cell epitopes overlap in this region.12,13,15,31 After OLT, thedeletion was found in less than 20% of the clones. In addition,new mutations occurred after OLT. At least five differentHBV virus populations, determined by the pre-S region, werefound in this patient. As the sole population found beforesurgery became a minority, this mutation was clearly notessential for virus survival after OLT.
HBIg is given to prevent HBV reinfection of the graft. Es-cape mutations in the S-gene could play a role in circum-FIG. 1. Date of reinfection after OLT. The time course of all 20 patientsventing effective therapy. Therefore, patients who becameafter OLT. The horizontal bar corresponds to the time course of each patient
after OLT. The change of color from dark to bright depicts the time point of reinfected during HBIg therapy were analyzed for the occur-HBV reinfection. Bright color corresponds to the detection of HBV DNA, and rence of escape mutations after OLT. Nine patients becamedark color to the time when no HBV DNA is present in serum of the patient. reinfected while receiving HBIg (Fig. 1). Three of these pa-The follow-up of each patient after OLT is at least 5 years, except for the
tients—patients 9, 10, and 12—belonged to the insertion/patients who died at earlier time points after OLT. The number of each patientis circled, indicating when reinfection occurred during therapy with HBIg. deletion group; 2 patients—patients 5 and 13—had no
AID Hepa 0016 / 5p12$$$302 08-07-96 09:22:07 hptas WBS: Hepatology
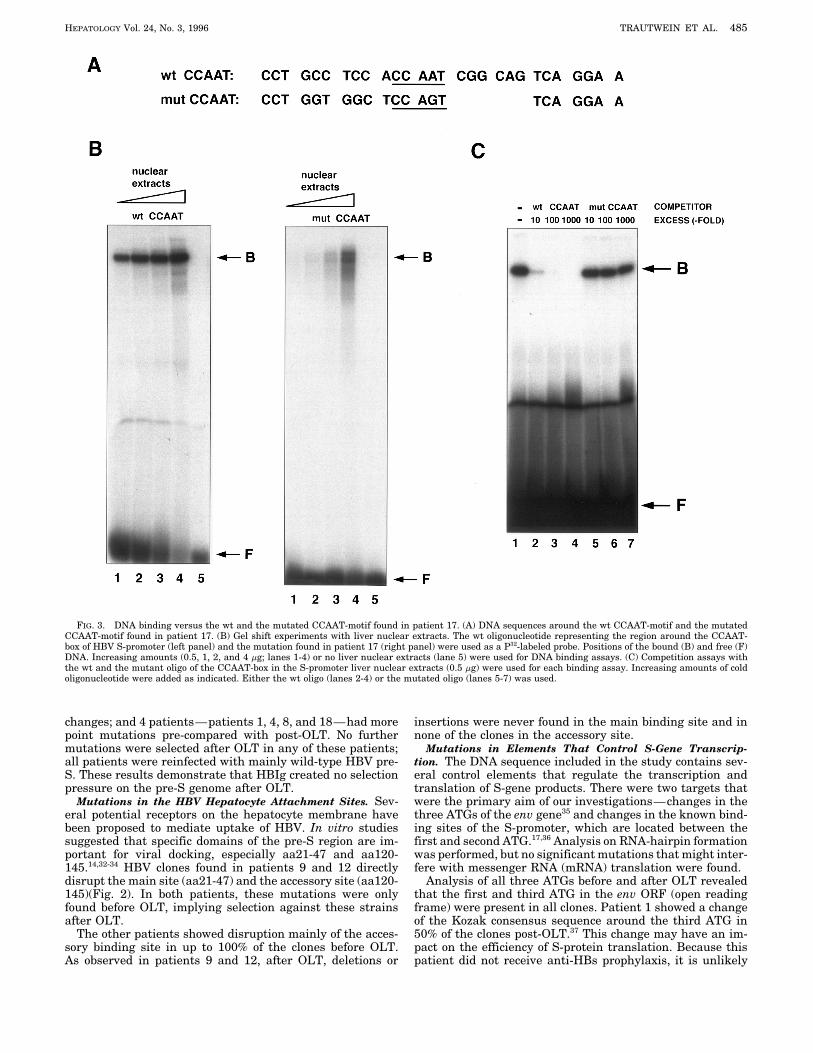
HEPATOLOGY Vol. 24, No. 3, 1996 TRAUTWEIN ET AL. 485
FIG. 3. DNA binding versus the wt and the mutated CCAAT-motif found in patient 17. (A) DNA sequences around the wt CCAAT-motif and the mutatedCCAAT-motif found in patient 17. (B) Gel shift experiments with liver nuclear extracts. The wt oligonucleotide representing the region around the CCAAT-box of HBV S-promoter (left panel) and the mutation found in patient 17 (right panel) were used as a P32-labeled probe. Positions of the bound (B) and free (F)DNA. Increasing amounts (0.5, 1, 2, and 4 mg; lanes 1-4) or no liver nuclear extracts (lane 5) were used for DNA binding assays. (C) Competition assays withthe wt and the mutant oligo of the CCAAT-box in the S-promoter liver nuclear extracts (0.5 mg) were used for each binding assay. Increasing amounts of coldoligonucleotide were added as indicated. Either the wt oligo (lanes 2-4) or the mutated oligo (lanes 5-7) was used.
changes; and 4 patients—patients 1, 4, 8, and 18—had more insertions were never found in the main binding site and innone of the clones in the accessory site.point mutations pre-compared with post-OLT. No further
mutations were selected after OLT in any of these patients; Mutations in Elements That Control S-Gene Transcrip-tion. The DNA sequence included in the study contains sev-all patients were reinfected with mainly wild-type HBV pre-
S. These results demonstrate that HBIg created no selection eral control elements that regulate the transcription andtranslation of S-gene products. There were two targets thatpressure on the pre-S genome after OLT.
Mutations in the HBV Hepatocyte Attachment Sites. Sev- were the primary aim of our investigations—changes in thethree ATGs of the env gene35 and changes in the known bind-eral potential receptors on the hepatocyte membrane have
been proposed to mediate uptake of HBV. In vitro studies ing sites of the S-promoter, which are located between thefirst and second ATG.17,36 Analysis on RNA-hairpin formationsuggested that specific domains of the pre-S region are im-
portant for viral docking, especially aa21-47 and aa120- was performed, but no significant mutations that might inter-fere with messenger RNA (mRNA) translation were found.145.14,32-34 HBV clones found in patients 9 and 12 directly
disrupt the main site (aa21-47) and the accessory site (aa120- Analysis of all three ATGs before and after OLT revealedthat the first and third ATG in the env ORF (open reading145)(Fig. 2). In both patients, these mutations were only
found before OLT, implying selection against these strains frame) were present in all clones. Patient 1 showed a changeof the Kozak consensus sequence around the third ATG inafter OLT.
The other patients showed disruption mainly of the acces- 50% of the clones post-OLT.37 This change may have an im-pact on the efficiency of S-protein translation. Because thissory binding site in up to 100% of the clones before OLT.
As observed in patients 9 and 12, after OLT, deletions or patient did not receive anti-HBs prophylaxis, it is unlikely
AID Hepa 0016 / 5p12$$$302 08-07-96 09:22:07 hptas WBS: Hepatology

486 TRAUTWEIN ET AL. HEPATOLOGY September 1996
TABLE 2. Percentage of Deletions/Insertions Found Before the wt sequence compared with the sequence found in patientand After OLT 17. To investigate the specificity of the binding versus the wt
CCAAT-box, competition assays were performed with the wt% of Clones With Insertions orDeletions and the mutated CCAAT-box oligo. While the cold wt oligo
very effectively competed for specific DNA binding, no compe-Patient Pre-Olt (%) Post-Olt (%)
tition was seen with the mutant oligo (Fig. 3C). These resultsindicate that specific DNA binding in liver nuclear extracts1 100 66
9 100 0 occurred only to the wt CCAAT sequence in the S-promoter.10 100 0 Therefore, these changes may lead to a similar dysregulation11 25 0 as shown by Lu et al.17 Thus, the liver histology of patient12 75 0 17 was investigated. Interestingly, patient 17 experienced14 0 25 HBV reinfection of the graft 4 months after OLT and died17 0 75 8.5 months after OLT. In the grafted liver, he showed the
characteristics of fibrosing cholestatic hepatitis (Fig. 4).NOTE: The relative percentage of clones in patients with deletions and/orinsertions in the HBV pre-S genome before and after OLT are shown.
DISCUSSION
The pre-S1 and pre-S2 domains of the viral envelope havebeen called immunodominant.10-13 The regions p21-47, p120-that the mutation was selected by an immune escape mecha-
nism. Loss of the second ATG was found in 8 of 20 (40%) of 141, p120-145, and p133-150 in the pre-S1 and pre-S2 do-mains confer protective immunity in chimpanzees.14-16,31 Ad-the patients, and was found in fewer patients before than
after OLT (3 vs. 6; in one patient before and after OLT). ditionally, multiple human B-cell epitopes and murine T-cellepitopes were mapped and described in the pre-S1 and pre-Comparison with wt HBV revealed several mutations in
the S-promoter before and after OLT. Single and double point S2 domains.12,13,38
Our results confirm that chronic HBV infection is associ-mutations occurred in known binding sites of transcriptionfactors. The ultimate role of these mutations on S-gene tran- ated with substantial deletions and insertions in the pre-
S genome.18,19 These mechanisms might be critical for viralscription must be analyzed by transfection and DNA bindingexperiments because the functional role of these cis-acting immune escape and could be important for the survival of
the virus. During chronic HBV infection, the liver is alreadyelements is poorly characterized. In patient 17, specific muta-tions were found in and around a cis-acting element called infected, and hepatocyte uptake might be less important for
HBV. Therefore, deletions and insertions of the pre-S regionthe CCAAT-box only after OLT (Fig. 3A). The regulatoryfunction of this element in the S-promoter was described in are frequently found and have no implication for the viral
life cycle. In contrast after OLT, when HBV reinfection of thedetail by Lu et al.17 An intact CCAAT-box promotes transcrip-tion of the S-promoter and suppresses pre-S transcription, new liver occurs, the situation changes. In vitro studies of
Neurath et al. showed that two domains, aa21-47 and aa120-leading to high small protein and low large protein expres-sion.17 These mechanisms were probably impaired in patient 145, are essential for viral uptake. aa21-47 has been called
the main hepatocyte binding site. Interestingly, this domain17 as the sequence in and around the CCAAT motif wasmutated. To investigate that these alterations indeed lead to also seems to be relevant in vivo, because mutations in this
region are only found before, but not after, OLT. In addition,different binding of transcription factors versus this element,gel shift experiments were performed. Liver nuclear extracts these results provide evidence that the deletions and inser-
tions found in chronic HBV carriers before OLT might beof Sprague-Dawley rats were prepared. Gel shift experimentswere performed with oligos representing the wt sequence in defective, unable to infect hepatocytes.
HBV reinfection of the graft correlates with poor prognosisand around the CCAAT motif and the corresponding se-quence of patient 17 (Fig. 3A). As demonstrated in Fig. 3B, after OLT.3,39,40 One-year survival after HBV reinfection has
been found to be as low as 50%.3 When there is high replica-there was a dramatic change in the binding pattern versus
FIG. 4. Liver histology of patient17. The liver histology of patient 17shows the typical signs of fibrosingcholestatic hepatitis with prolifera-tion of bile ductules interdigitatingbetween liver cell plates (pt, portaltract) (hematoxylin-eosin; originalmagnification 140.)
AID Hepa 0016 / 5p12$$$302 08-07-96 09:22:07 hptas WBS: Hepatology

HEPATOLOGY Vol. 24, No. 3, 1996 TRAUTWEIN ET AL. 487
2. Benner KG, Lee RG, Keeffe EB, Lopez RR, Sasaki AW, Pinson CW. Fibros-tion directly before OLT, the risk of HBV reinfection is high.ing cytolytic liver failure secondary to recurrent hepatitis B after liverTherefore, because there is a shortage of organs, the consen- transplantation. Gastroenterology 1990;103:1307-1312.
sus is not to transplant patients with high-replicative HBV 3. Todo S, Demetris AJ, Van Thiel D, Teperman L, Fung JJ, Sarzl TE. Or-infection.41 Besides promising experimental trials with nucle- thotopic liver transplantation for patients with hepatitis B virus–related
liver disease. HEPATOLOGY 1991;13:619-626.oside analogues,20,42,43 the treatment of choice to prevent HBV4. Shah G, Demetris AJ, Gavaler JS, Lewis JH, Todo S, Starzl TE, van Thielreinfection after OLT is HBIg prophylaxis. HBIg is obtained
DH. Incidence, prevalence, and clinical course of hepatitis B following liverfrom serum of anti-HBs–positive persons. Therefore, pa- transplantation. Gastroenterology 1992;103:323-329.tients after natural HBV infection and after HBV vaccination 5. Lauchert W, Muller R, Pichlmayr R. Immunoprophylaxis of hepatitis B
virus reinfection in recipients of human liver allografts. Transplant Procare included (Ebers G, Biotest, personal communication, De-1987;19:2387-2389.cember 1995).
6. Muller R, Gubernatis G, Farle M, Niehoff G, Klein H, Wittekind C, TuschHBIg therapy puts selection pressure on the coding region G, et al. Liver transplantation in Hbs antigen (HBsAg) carriers. Preventionof the small S-protein.44 Mutations were selected when HBV of hepatitis B virus (HBV) recurrence by passive immunisation. J Hepatol
1991;13:90-96.reinfection occurred while the patients were receiving HBIg7. Samuel D, Bismuth A, Mathieu D, Arulnaden JL, Reynes M, Benhamoutherapy. In contrast, no mutations were selected in the pre-
JP, Brechot C, et al. Passive immunoprophylaxis after liver transplanta-S genome. These results indicate that either HBIg contains tion in HBsAg positive patients. Lancet 1991;337:813-815.no protective anti–pre-S or these antibodies are not able to 8. Okamoto H, Tsuda F, Sakugawa H, Sastrosoewignjo RI, Imai M, Miya-
kawa Y, Mayumi M. Typing hepatitis B virus by homology in nucleotideneutralize the virus and prevent HBV reinfection. Becausesequence: comparison of surface antigen subtypes. J Gen Virol 1988;69:the pre-S region has been shown to be immunodominant,10-13
2575-2583.it is more likely that HBIg contains only minor anti–pre-9. Carman WF, Thomas H, Esteban D. Viral genetic variation: hepatitis B
S. Therefore, these results indicate that further research is as a clinical example. Lancet 1993;341:349-352.needed to investigate the protective effect of antibodies di- 10. Milich DR, Thornton GB, Neurath A, Kent S, Michel M, Tiollais P, Chisari
FV. Enhanced immunogenicity of the pre-S region of hepatitis B surfacerected against the main hepatocyte attachment site to pre-antigen. Science 1985;228:1195-1199.vent HBV reinfection after OLT.
11. Milich DR, McNamara MK, McLachlan A, Thornton GB, Chisari FV. Dis-The HBV env gene consists of three in-frame ATGs and tinct H-2–linked regulation of T-cell responses to the PreS and S regionsa common stop codon.35 Two regulatory elements affect the of the same hepatitis B surface antigen polypeptide allows circumvention
of nonresponsiveness to the S region. Proc Natl Acad Sci U S A 1985;82:transcription of the gene, the pre-S and the S-promoters. The8168-8172.pre-S promoter is located 5* of the first ATG, and the S-
12. Milich DR, McLachlan A, Chisari FV, Nakamura T, Thornton GB. Twopromoter is located between the first and second ATG.36,45,46distinct but overlapping antibody binding sites in the PreS(2) region of
Each promoter regulates a major transcript (2.4-kb and 2.1- HBsAg localized within 11 continuous residues. J Immunol 1986;137:2703-2710.kb mRNA). These two major transcripts lead to the transla-
13. Milich DR, McLachlan A, Chisari FV, Kent SBH, Thornton GB. Immunetion of the three proteins of the envelope: large, middle, andresponse to the PreS(1) region of the hepatitis B surface antigen (HBsAg):small S-protein.35 The second ATG, and thus the middle S- a preS(1)-specific T cell response can bypass nonresponsiveness to the
protein, was often deleted before and after OLT, and, as preS(2) and S regions of HBsAg. J Immunol 1986;137:315-322.shown previously by Gerken et al. and Santantonio et al.,18,19 14. Neurath AR, Kent SBH, Strick N, Parker K. Identification and chemical
synthesis of a host cell receptor binding site on hepatitis B virus. Cellthese data suggest that the middle S-protein is not important1986;46:429-436.for the normal HBV life cycle.
15. Neurath AR, Kent SBH, Parker K, Prince AM, Strick N, Brotman B,In patient 17, we found a mutation in the so-called CCAAT Sproul P. Antibodies to a synthetic peptide from the preS 120-145 region
motif, which was characterized by Lu et al.17 Lu et al. showed of the hepatitis B virus envelope are virus neutralizing. Vaccine 1986;4:35-37.that mutations in this motif lead to high transcription of the
16. Neurath AR, Strick N, Sproul P. Search for hepatitis B virus cell receptors2.4-kb S mRNA and low transcription of the 2.1-kb S mRNA.reveals binding sites for interleukin 6 on the virus envelope protein. J ExpThis is in contrast to the wt HBV genome with an intact Med 1992;175:461-469.
CCAAT motif, where the opposite regulation of transcription 17. Lu CC, Chen M, Ou JH, Yen TSB. Key role of a CCAAT element in regulat-ing hepatitis B virus surface protein expression. Virology 1995;206:1155-is evident. High amounts of the 2.4-kb S mRNA result in a1158.higher translation rate of the large S-protein, which has been
18. Gerken G, Kremsdorf D, Capel F, Dauguet C, Manns M, Meyer zumshown to have a cytopathic effect on hepatocytes.17 We show Buschenfelde K-H, Brechot C. Hepatitis B defective virus with rearrange-that the mutated S-promoter region in patient 17 results in ments in the preS gene during chronic HBV-infection. Virology 1991;183:
555-565.loss of binding of a specific transcription factor compared19. Santantonio T, Jung MC, Schneider R, Fernholz D, Milella M, Monno M,with the wt sequence. Therefore, these experiments indicate
Pastore G, et al. Hepatitis B virus genomes that cannot synthesize pre-that because of the mutation found in patient 17, S-gene S2 proteins occus frequently and as a dominant virus population in chronictranscription is altered compared with the wt sequence. Be- carriers in Italy. Virology 1992;188:948-952.cause this element has been characterized to have a dual role 20. Boker KH, Ringe B, Kruger M, Pichlmayr R, Manns MP. Prostaglandin
E plus famciclovir—a new concept for the treatment of severe hepatitison the transcription of the 2.4- and 2.1-kb S mRNAs, it isB after liver transplantation. Transplantation 1994;57:1706-1708.very likely that, in patient 17, the transcription of the 2.4-
21. Michitaka K, Durrazzo M, Tillmann HL, Walker D, Philipp T, Mannskb mRNA is enhanced; thus, this mutation may be relevant MP. Analysis of hepatitis C virus genome in patients with autoimmunefor the occurrence of fibrosing cholestatic hepatitis found in hepatitis type 2. Gastroenterology 1994;106:1603-1610.
22. Tillmann HL, Trautwein C, Walker D, Kubicka S, Boker K, Manns M.this patient after OLT.Prevalence of the hepatitis B e-minus variant in northern Europe. GutIn summary, our results show 1) Deletions and insertions1995;37:568-573.in the main hepatocyte binding site of chronic HBV carriers 23. Kubicka S, Trautwein C, Schrem H, Tillmann H, Manns MP. Hepatocellu-
are not found following HBV reinfection after OLT, 2) HBIg lar carcinoma and P53 mutation in northern Europe. J Hepatol 1995;23:412-419.has no selection pressure on the pre-S region, and 3) Muta-
24. Descombes P, Chojkier M, Lichtsteiner S, Falvey E, Schibler U. LAP, ations in the CCAAT-box of the S-promoter result in decreasednovel member of the C/EBP gene family, encodes a liver-enriched tran-binding versus this element, and may be one of the mecha-scriptional activator protein. Genes Dev 1990;4:1541-1551.
nisms leading to fibrosing cholestatic hepatitis after OLT. 25. Trautwein C, Walker D, Plumpe J, Manns MP. Transactivation of LAP/NF-IL6 is mediated by an acidic domain in the N-terminal part of theAcknowledgment: We thank F. W. Carman for critically protein. J Biol Chem 1995;270:15130-15136.
reviewing the manuscript for helpful comments. 26. Galibert F, Mandart E, Fitoussi F, Tiollais P, Charny P. Nucleotide se-quence of the hepatitis B virus genome (subtype ayw) cloned in E. coli.
REFERENCES Nature 1979;281:646-650.27. Valenzuela P, Quiroga M, Zalvidar J, Gray P, Rutter WJ. The nucleotide1. Samuel D, Bismuth H. Liver transplantation for hepatitis B. Gastroenterol
Clin North Am 1993;22:271-283. sequence of the hepatitis B viral genome and the identification of the
AID Hepa 0016 / 5p12$$$303 08-07-96 09:22:07 hptas WBS: Hepatology

488 TRAUTWEIN ET AL. HEPATOLOGY September 1996
major viral genes. In: Fields BN, Jaenisch R, Fox CF, eds. Animal virus 38. Milich DR, Jones JE, McLachlan A, Bitter G, Moriarty A, Hughes JL.genetics: New York: Academic Press, 1980:57-70. Importance of subtype in the immune response to the preS(2) region of
28. Kobayashi M, Koike K. Complete nucleotide sequence of hepatitis B virus the hepatitis B surface antigen II. Synthetic PreS(2) immunogen. J Immu-DNA of subtype adr and its conserved gene organization. Gene 1984;30: nol 1990;144:3544-3551.227-232. 39. Lau J, Bain VG, Davies SE, O’Grady JG, Alberti A, Alexander GJM, Wil-
29. Okamoto H, Imai M, Shimozaki M, Hoshi Y, Iizuka H, Gotanda T, Tsuda liams R. High level expression of hepatitis B viral antigens in fibrosingF, et al. Nucleotide sequence of a cloned hepatitis B virus genome, subtype cholestatic hepatitis. Gastroenterology 1992;102:956-962.ayr: comparison with genomes of the other three subtypes. J Gen Virol 40. Samuel D, Muller R, Alexander G, Fassati L, Ducot B, Benhamou J-P,1986;67:2305-2314. Bismuth H, and the investigators of the European Concerted Action
30. Mimms LT, Floreani M, Tyner J, Whitters E, Rosenlof R, Wray L, Goetze on Viral Hepatitis study. Liver transplantation in European patientsA, et al. Discrimination of hepatitis B virus (HBV) subtypes using mono- with the hepatitis B surface antigen. N Engl J Med 1993;329:1842-clonal antibodies to the preS1 and preS2 domains of the viral envelope. 1847.Virology 1990;176:604-619. 41. Indications for liver transplantation. Text of a consensus conference. Ann
31. Itoh Y, Takai E, Ohnuma H, Kitajima K, Tsuda F, Machida A, Mishiro S, Gastroenterol Hepatol (Paris) 1994;30:101-104, 109-112.et al. A synthetic peptide vaccine involving the product of the pre-S(2) 42. Grellier L, Brown D, Berenek P, Rolles K, Burroughs A, Dusheiko G.region of hepatitis B virus DNA: protective efficacy in chimpanzees. Proc Lamivudine prophylaxis: a new strategy for prevention of hepatitis B rein-Natl Acad Sci U S A 1986;83:9174-9178. fection in liver transplantation for hepatitis B DNA positive cirrhosis.
32. Neurath AR, Thanavala Y. Hepadnaviruses. In: Van Regenmortel MHV, Single topic symposium on liver transplantation for chronic viral hepatitis.Neurath AR, eds. Immunochemistry of Viruses II. Amsterdam: Elsevier, Reston, VA, 1995.1990:403-458. 43. Kruger M, Tillmann HL, Trautwein C, Bode U, Oldhafer K, Boker KHW,33. Petit MA, Dubanchet S, Capel F, Voet P, Dauguet C, Hauser P. HepG2 Pichlmayr R, et al. Treatment of hepatitis B virus reinfection after livercell binding activities of different hepatitis B isolates: inhibitory effect of transplantation with famciclovir. HEPATOLOGY 1994;20:130A.anti-HBs and anti-preS (21-47). Virology 1991;180:483-491.
44. Carman FW, Trautwein C, van Deursen FJ, Colman K, McIntyre G, Kliem34. Petit MA, Strick N, Dubanchet S, Capel F, Neurath AR. Inhibitory activityV, Waters J, et al. Hepatitis B virus envelope variation after transplanta-of monoclonal antibody F35.25 on the interaction between hepatocytestion with and without polyclonal antibody pressure. HEPATOLOGY 1994;(HepG2 cells) and preS1-specific ligands. Mol Immunol 1991;28:517-521.20:138A.35. Heermann KH, Goldmann U, Schwartz W, Seyffarth T, Baumgarten H,
45. Robinson WS. Hepadnaviridae and their replication. In: Fundamental vi-Gerlich WH. Large surface proteins of hepatitis B virus containing therology, 2nd ed. Fields-BN, Knipe-DM, eds. New York: Raven Press, 1991:pre-S sequence. J Virol 1994;52:396-402.989-1021.36. Zhou D-X, Yen TSB. The hepatitis B virus S promoter comprises a CCAAT
46. Raney AK, Millich DR, McLachlan A. Characterization of hepatitis B virusmotif and two initiation regions. J Biol Chem 1991;266:23416-23421.major surface antigen gene transcriptional regulatory elements in differ-37. Kozak M. The scanning model for translation: an update. J Cell Biol 1989;
108:229-241. entiated hepatoma cell lines. J Virol 1989;63:3919-3925.
AID Hepa 0016 / 5p12$$$303 08-07-96 09:22:07 hptas WBS: Hepatology




















