
Hemisphere Specific Impairments in Reach-to-Grasp Control After Stroke: Effects of Object Size
13
1 Neurorehabilitation and Neural Repair Volume XX Number X Month XXXX xx-xx © 2009 The Author(s) 10.1177/1545968309332733 http://nnr.sagepub.com Hemisphere Specific Impairments in Reach-to-Grasp Control After Stroke: Effects of Object Size Jarugool Tretriluxana, PhD, PT, James Gordon, EdD, PT, FAPTA, Beth E. Fisher, PhD, PT, and Carolee J. Winstein, PhD, PT, FAPTA Background and objective. The authors investigated hemispheric specialization for the visuomotor transformation of grasp preshaping and the coordination between transport and grasp in individuals poststroke. Based on a bilateral model, the authors hypothesized that after uni- lateral stroke there would be hemisphere-specific deficits revealed by the ipsilesional limb. Methods. Right or left stroke and age- and limb- matched nondisabled participants performed rapid reach-to-grasp of 3 sized objects. The authors quantified grasp preshaping as the correlation between initial aperture velocity and peak aperture, and peak aperture and object diameter. A cross correlation analysis using transport velocity and aperture size was performed to quantify transport-grasp coordination. All statistical tests for hemisphere-specific deficits involved comparisons between each stroke group and the matched nondisabled group. Results. Overall, the right stroke group, but not left stroke group, demonstrated prolonged movement time. For grasp preshaping there was a higher correlation between initial aperture velocity and peak aperture for the right stroke group and a lower correlation between peak aperture and object diameter for the left stroke group. For transport-grasp coordination the correlation between transport velocity and aperture size was higher for the left stroke group and lower for the right stroke group, which also demonstrated a higher standard deviation of time lag. Conclusions. After left stroke, there was deficient scaling of grasp preshaping and stronger transport-grasp coordination. In contrast, after right stroke, grasp preshaping began earlier and transport-grasp coordination was weaker. Together, these hemisphere-specific deficits suggest a left hemisphere specialization for the visuomotor transformation of grasp preshaping and a right hemisphere specialization for transport-grasp coordination. Keywords: Hemispheric specialization; Ipsilesional hand; Kinematics; Dexterity T he notion that the left and right hemispheres are asymmet- ric for the control of skilled movements originally came from careful observations of human behavior. 1,2 In a previous study with nondisabled adults, we identified manual asymme- tries in grasp preshaping and transport-grasp coordination. 3 In particular, each hand performed reach-to-grasp to different sized objects with grasp preshaping that was scaled to object size. However, the process by which different objects were successfully procured was distinguishable for each hand. For example, the left hand exhibited anticipatory control of grasp aperture preshaping, whereas the right hand exhibited stronger transport-grasp coordination. These hand-specific effects pro- vide support for the existence of manual asymmetries in reach- to-grasp actions. However, extrapolation of these findings to hemispheric specialization can be confounded by hand domi- nance, such that the stronger transport-grasp coordination found for the right hand may simply reflect more practice or experience 4,5 with little relevance to hemispheric specializa- tion. Alternatively, if we assume that manual asymmetries are not solely a reflection of hand dominance, well-designed investigations using patients with unilateral brain damage can be used to provide insight into hemispheric specialization. Indeed, neuropsychological investigations of aiming move- ments in individuals with focal hemispheric stroke have pro- vided important data about hemispheric specialization. 6,7 Therefore, the purpose of this study was to investigate hemi- spheric specialization in reach-to-grasp actions with individu- als post focal hemispheric stroke using the same experimental protocol used in our previous study. 3 An important consideration in investigations of hemi- spheric specialization is that it is well known that both hemi- spheres participate in the control of unimanual goal-directed movements including reach-to-grasp actions. Imaging studies consistently show bilateral activation during both preparation and execution of unilateral finger movements. 8-10 Ipsilateral contributions to unilateral movements have also been demon- strated using transcranial magnetic stimulation (TMS). 11 For example, a recent TMS study demonstrated an inhibitory com- munication between primary motor areas of the 2 hemispheres prior to movement initiation. 12,13 This inhibition is likely medi- ated through excitatory transcallosal connections, which ter- minate onto inhibitory interneurons. 13-15 Kobayashi and From the Division of Biokinesiology and Physical Therapy, University of Southern California, Los Angeles (JT, JG, BEF, CJW); the Department of Neurology, Keck School of Medicine, University of Southern California, Los Angeles (BEF, CJW); and the Faculty of Physical Therapy and Applied Movement Science, Mahidol University, Nakhon Pathom, Thailand (JT). Address correspondence to Carolee J. Winstein, Division of Biokinesiology and Physical Therapy, University of Southern California, Los Angeles, CA 90033. E-mail: [email protected]. Neurorehabil Neural Repair OnlineFirst, published on May 1, 2009 as doi:10.1177/1545968309332733
Transcript of Hemisphere Specific Impairments in Reach-to-Grasp Control After Stroke: Effects of Object Size

1
Neurorehabilitation and Neural Repair
Volume XX Number XMonth XXXX xx-xx© 2009 The Author(s)
10.1177/1545968309332733http://nnr.sagepub.com
Hemisphere Specific Impairments in Reach-to-Grasp Control After Stroke: Effects of Object Size
Jarugool Tretriluxana, PhD, PT, James Gordon, EdD, PT, FAPTA, Beth E. Fisher, PhD, PT, and Carolee J. Winstein, PhD, PT, FAPTA
Background and objective. The authors investigated hemispheric specialization for the visuomotor transformation of grasp preshaping and the coordination between transport and grasp in individuals poststroke. Based on a bilateral model, the authors hypothesized that after uni-lateral stroke there would be hemisphere-specific deficits revealed by the ipsilesional limb. Methods. Right or left stroke and age- and limb-matched nondisabled participants performed rapid reach-to-grasp of 3 sized objects. The authors quantified grasp preshaping as the correlation between initial aperture velocity and peak aperture, and peak aperture and object diameter. A cross correlation analysis using transport velocity and aperture size was performed to quantify transport-grasp coordination. All statistical tests for hemisphere-specific deficits involved comparisons between each stroke group and the matched nondisabled group. Results. Overall, the right stroke group, but not left stroke group, demonstrated prolonged movement time. For grasp preshaping there was a higher correlation between initial aperture velocity and peak aperture for the right stroke group and a lower correlation between peak aperture and object diameter for the left stroke group. For transport-grasp coordination the correlation between transport velocity and aperture size was higher for the left stroke group and lower for the right stroke group, which also demonstrated a higher standard deviation of time lag. Conclusions. After left stroke, there was deficient scaling of grasp preshaping and stronger transport-grasp coordination. In contrast, after right stroke, grasp preshaping began earlier and transport-grasp coordination was weaker. Together, these hemisphere-specific deficits suggest a left hemisphere specialization for the visuomotor transformation of grasp preshaping and a right hemisphere specialization for transport-grasp coordination.
Keywords: Hemispheric specialization; Ipsilesional hand; Kinematics; Dexterity
The notion that the left and right hemispheres are asymmet-ric for the control of skilled movements originally came
from careful observations of human behavior.1,2 In a previous study with nondisabled adults, we identified manual asymme-tries in grasp preshaping and transport-grasp coordination.3 In particular, each hand performed reach-to-grasp to different sized objects with grasp preshaping that was scaled to object size. However, the process by which different objects were successfully procured was distinguishable for each hand. For example, the left hand exhibited anticipatory control of grasp aperture preshaping, whereas the right hand exhibited stronger transport-grasp coordination. These hand-specific effects pro-vide support for the existence of manual asymmetries in reach-to-grasp actions. However, extrapolation of these findings to hemispheric specialization can be confounded by hand domi-nance, such that the stronger transport-grasp coordination found for the right hand may simply reflect more practice or experience4,5 with little relevance to hemispheric specializa-tion. Alternatively, if we assume that manual asymmetries are not solely a reflection of hand dominance, well-designed investigations using patients with unilateral brain damage can
be used to provide insight into hemispheric specialization. Indeed, neuropsychological investigations of aiming move-ments in individuals with focal hemispheric stroke have pro-vided important data about hemispheric specialization.6,7 Therefore, the purpose of this study was to investigate hemi-spheric specialization in reach-to-grasp actions with individu-als post focal hemispheric stroke using the same experimental protocol used in our previous study.3
An important consideration in investigations of hemi-spheric specialization is that it is well known that both hemi-spheres participate in the control of unimanual goal-directed movements including reach-to-grasp actions. Imaging studies consistently show bilateral activation during both preparation and execution of unilateral finger movements.8-10 Ipsilateral contributions to unilateral movements have also been demon-strated using transcranial magnetic stimulation (TMS).11 For example, a recent TMS study demonstrated an inhibitory com-munication between primary motor areas of the 2 hemispheres prior to movement initiation.12,13 This inhibition is likely medi-ated through excitatory transcallosal connections, which ter-minate onto inhibitory interneurons.13-15 Kobayashi and
From the Division of Biokinesiology and Physical Therapy, University of Southern California, Los Angeles (JT, JG, BEF, CJW); the Department of Neurology, Keck School of Medicine, University of Southern California, Los Angeles (BEF, CJW); and the Faculty of Physical Therapy and Applied Movement Science, Mahidol University, Nakhon Pathom, Thailand (JT). Address correspondence to Carolee J. Winstein, Division of Biokinesiology and Physical Therapy, University of Southern California, Los Angeles, CA 90033. E-mail: [email protected].
Neurorehabil Neural Repair OnlineFirst, published on May 1, 2009 as doi:10.1177/1545968309332733

2 Neurorehabilitation and Neural Repair
All participants with stroke presented a clinical picture consistent with unilateral anterior circulation involvement.23 They all presented with some amount of hemiparesis contral-ateral to the lesion as measured by grip and pinch strength using Jamar dynamometer (Table 1). Specific information about lesion location was obtained from medical records and from MRI or a computerized tomography scan for 9 partici-pants with right lesion and 11 participants with left lesion. There was no significant difference in time poststroke between the 2 groups (Table 1).
Handedness was determined using the Edinburgh Handedness Inventory.24 For participants poststroke, it is based on their pre-stroke handedness. All were right hand dominant. The partici-pants were recruited from the local community. None of the nondisabled participants reported any neurological or muscu-loskeletal disorder at the time of the experiment. A written informed consent form approved by the University Institutional Review Board was read and signed prior to the experiment.
Experimental Setup and Task
The basic task was to reach forward from a designated start position, grasp a prepositioned cylinder, and lift it off the table as soon and as quickly using the thumb and index finger.
Each participant sat in a straight back chair with the torso secured to the chair back using a cross-shaped belt, the hand rested on a hand switch, and the thumb and index fingertips were positioned in opposition at the tips. A light emitting diode (LED) located 25 cm above the table surface was positioned behind the moveable target cylinder. The LED signal was triggered externally by the experimenter. Three different cylinders were used that measured 1.2 cm, 2.4 cm, and 5.6 cm in diameter, each were 10 cm in height and wrapped with tape at the center. Prior to each trial, the cylin-der was positioned on an object switch located 30 cm from the start switch (Figure 1, top view). Finally, vision of the reaching arm and hand was occluded by a wooden shield positioned to block the arm and hand at the beginning of the movement path but not during target approach (Figure 1, lateral view).
Behavioral kinematics data were acquired using the MotionMonitor (Innovative Sports Training Inc, Chicago, IL), an electromagnetic motion system with 6 degrees of freedom miniBIRD sensors (Ascension Technologies). Three sensors were attached with tape to the upper extremity, 1 on the forearm proximal to the styloid process of the radius and the other 2 on the nail bed of the thumb and index fingers. Position data were captured at 120 Hz. The LED hand and object lift switches were interfaced with the MotionMonitor through a timer and signaled the following 3 events: LED onset, hand lift onset, and movement termination (object lift switch). The entire data acquisition session was also recorded with a video camera positioned above and in front of the workspace.
coworkers provide further evidence for the inhibitory inter-hemispheric interaction in nondisabled adults with combined functional magnetic resonance imaging (MRI) and TMS.16 With evidence for bilateral activity during unimanual move-ments, a purely contralateral control model provides little explanatory power for hemisphere-specific contributions pre-viously demonstrated in nondisabled individuals.3 Instead, bilateral control, hereafter referred to as the bilateral model, provides greater explanatory power for these previous findings in nondisabled individuals.3 The bilateral model includes the unique contributions of each hemisphere and the associated interhemispheric communication.
To test the bilateral model predictions of hemispheric spe-cialization, we employed the same experimental protocol that we used previously but with a stroke-induced unilateral brain damage group. We reasoned that any deficits in the perfor-mance of the nonparetic, so-called “ipsilesional” upper extrem-ity would be expected to reflect the unique contributions of each hemisphere, one of which is impaired, and the effects of altered interhemispheric communication known to occur after stroke.17 After hemisphere damage, transcallosal communica-tion effectively downregulates the damaged hemisphere and upregulates the nondamaged hemisphere. This imbalance from the stroke lesion increases the probability for hemisphere-specific effects to emerge, compared to that in the nondisabled case. Because these deficits are apparent for the ipsilesional limb and are not evident in movements of matched nondis-abled participants they can provide a unique window into hemispheric specialization.
There is considerable evidence for hemisphere-specific pat-terns of deficits in goal-directed aiming movements performed by the ipsilesional hand of individuals with unilateral stroke.18-22 Our previous findings from right-handed nondisabled indi-viduals showed an earlier aperture preshaping response in the nondominant left hand and more coordinated control of hand reaching and finger grasping in the dominant right hand.3 If each hemisphere contributes uniquely to the control of reach-to-grasp movements, we expect that hemisphere-specific defi-cits will be observed in the performance of the ipsilesional hand. If the bilateral model is supported, our hypothesis is that participants with a dominant left hemisphere stroke will exhibit deficits in aperture preshaping in the left hand. In con-trast, participants with a nondominant right hemisphere stroke will exhibit deficits in the coordination of hand reaching and finger grasping in the right hand.
Method
Participants
A total of 36 right-handed volunteers participated in this study: 12 participants with right stroke (right hand mean [SD] age = 53 [13.8] years), 12 participants with left stroke (left hand mean [SD] age = 56 [16.7] years), and 12 age-matched nondisabled participants (mean [SD] age = 52 [14.9] years).

Tretriluxana et al / Hemisphere Specific Impairments in Reach-to-Grasp Control After Stroke 3
followed immediately by 3 test blocks in the fixed order of sizes 2.4 cm, 1.2 cm, and 5.6 cm. Each block consisted of 20 trials for each object diameter.
Reaction time (time from LED onset to hand lift onset) was displayed to the experimenter. If the latency exceeded 350 mil-liseconds, the trial was deemed invalid and another trial was collected. This assured that a full complement of 20 valid trials with each object diameter was used for analysis.
Data Reduction of Kinematic Variables
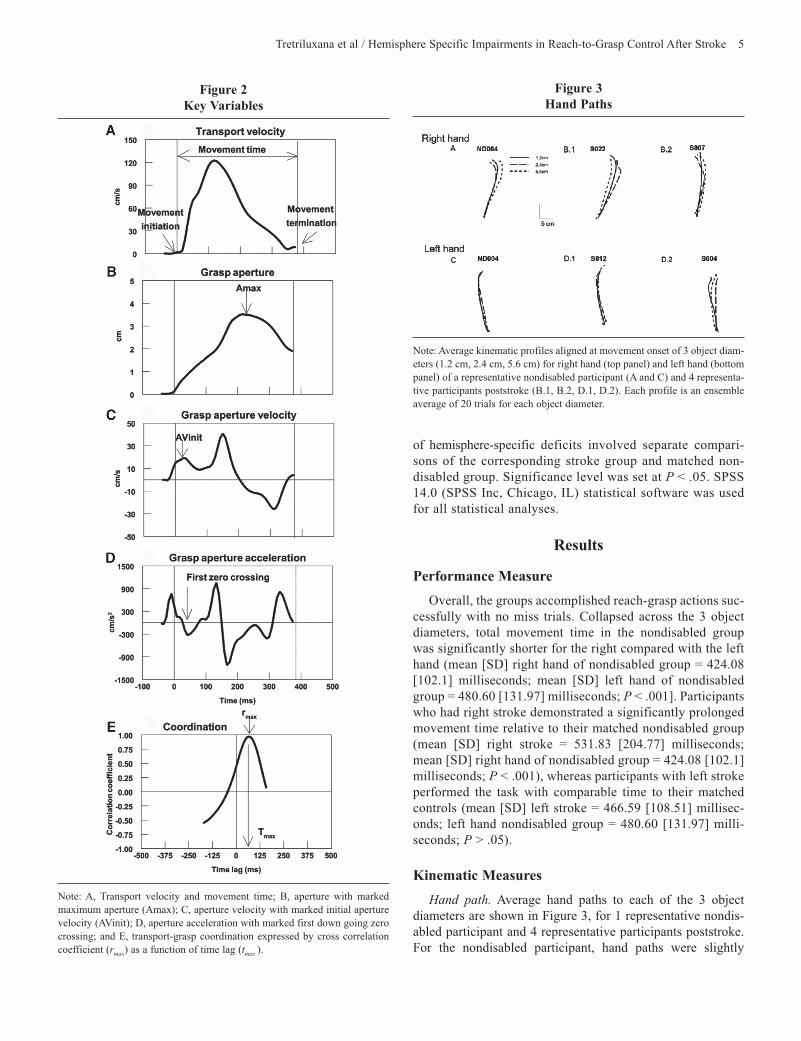
All kinematic data were filtered using a zero-lag Butterworth low-pass filter with a 20 Hz cut-off frequency. Hand path was derived from x-y-z coordinates of the wrist sensor. Tangential velocity of transport (Figure 2A) was calculated from the wrist sensor position using a finite-difference technique.27 Movement initiation was defined as the first bin of a continuous rise in transport velocity. Movement termination was defined as the
Experimental Procedures
For nondisabled participants, each hand was tested with the order counterbalanced across participants such that 6 partici-pants performed first with the right hand and the other 6 began with the left hand. The nonparetic so-called “ipsilesional” limb of individuals with stroke was tested.25,26 Therefore, the perfor-mance of the ipsilesional limb reflects the combined contribu-tion of the 2 hemispheres, one of which is damaged. Differences in deficits observed for the ipsilesional limb compared with the same limb of age-matched nondisabled individuals pro-vides the unique contribution of the lesioned hemisphere with-out the confounding effects of weakness and control deficits of the hemiparetic limb.7
After the LED signal, the participant reached forward from the start position, grasped the cylinder around the taped area, and lifted it off the table using only the thumb and index fin-ger. The experiments started with 3 to 5 practice trials using the middle-sized (2.4 cm) cylinder. These practice trials were
Table 1 Demographic of 12 Participants Post Right Stroke and 12 Post Left Stroke
GS PS Age Time Post L R L RGroup ID (Years) Gender Onset (Months) (% Normal) (% Normal) Locations/Types of Lesion
Right stroke S007 59 M 85 17.2 96.8 10.3 51.1 MCA occlusion (T-P, M1, and posterior IC)S008 68 M 30 51.6 72.4 48.8 56.5 CR and BG infarctionS010 60 F 131 4.8 99.1 0 96.6 Thalamus, posterior IC, and CR infarctionS011 68 M 83 17.2 64.0 0 51.4 N/AS014 63 M 5 24.3 78.5 38.9 75.7 Medial O (lingual gyrus and cuneus) infarctionS016 46 M 27 84 81.1 67.3 66.5 N/AS017 51 M 17 58.3 74.6 50.4 73.9 PV (tail of the caudate nucleus) and CR infarctionS021 70 F 20 53.0 88.7 62.9 64.9 PV (caudate nucleus) and part of CR infarctionS022 39 M 19 33.1 90.1 25.5 58.8 M1 and PM infarctionS024 52 F 22 24.3 107.0 9.6 18.4 SM1 and PM infarctionS025 30 F 16 82.5 89.5 63.8 76.9 Globus pallidus and part of posterior IC infarctionS026 39 M 41 75.8 90.4 50.7 74.6 N/A
Mean (SD) 53 8M/4F 41.3 43.8 86.0 35.7 63.8 counts (13.1) (38.0) (27.5) (12.3) (25.5) (19.2)Left stroke
S002 50 F 34 122.2 58.5 100.0 57.2 MCA occlusion (SM1 and small part of thalamus)S003 77 M 84 76.0 11.7 42.1 17.6 SM1 and T with posterior IC and lentiform nucleus
infarctionS004 58 M 8 138.8 48.4 98.5 20.9 Superior T and P hemorrhageS006 70 M 134 96.0 29.2 96.2 54.7 Deep white matter and thalamus infarction S009 57 F 34 130.2 55.7 64.3 41.3 MCA occlusion (insular cortex, caudate, putamen, and IC)S012 26 F 17 121.3 62.0 87.4 49.7 M1 infarctionS013 73 F 65 95.4 0 62.9 0 BG hemorrhageS015 56 M 2 95.2 6.5 82.6 27.8 N/AS018 75 F 15 85.1 22.3 52.2 20.8 PV (tail of caudate nucleus) and part of CR infarctionS019 27 F 22 114.3 63.4 80.9 41.0 M1 infarctionS020 59 M 16 116.3 53.2 77.5 25.5 F (including M1) and P hemorrhageS023 55 F 16 111.6 46.1 78.6 55.0 Posterior IC infarction
Mean (SD) 56 5M/7F 37.3 108.5 38.1 77.0 34.3 counts (16.7) (38.7) (19.0) (23.0) (18.4) (18.2)
Abbreviations: BG, basal ganglia; CR, corona radiata; F, frontal cortex; IC, internal capsule; M1, primary motor area; O, occipital cortex; P, parietal cortex; PM, premotor area; PV, periventricular area; SM1, primary sensorimotor area; T, temporal cortex; N/A, no information.Note: Grip (GS) and pinch (PS) values represent percentage strength of more affected and less affected limbs compared to limb, age, and gender matched norma-tive data.57

4 Neurorehabilitation and Neural Repair
(x-axis). Phase plane plots have been used previously to char-acterize coordination of functional behaviors such as locomo-tion.28 The coordination was quantified by cross-correlation analysis between transport velocity and aperture size. The highest cross correlation coefficient (rmax), the associated time lag for which the highest correlation coefficient was observed (tmax), and the standard deviation of the time lag across trials were calculated (Figure 2E). Using our convention, a positive lag represented a lead of peak transport velocity over peak aperture. It is noteworthy that both primary measures used here were aperture velocity of grasp preshaping and the cross-correlation analyses for transport-grasp coordination, which demonstrated good sensitivity to manual asymmetries in our previous study with nondisabled adults.3
Statistical Analysis
Z score (Fisher’s z) transformed correlations were used for comparisons across participants. Transformed z scores were converted back to equivalent r coefficients for reporting. Multivariate analysis of variance (ANOVA) was performed for group comparison on correlation coefficient values, time lag, and standard deviation of time lag. Two factors (groups X object diameters) ANOVA, repeating on object diameter, were used to compare total movement time, peak transport velocity, maximum aperture, and standard deviation of maximum aper-ture. The Bonferroni correction was applied for post hoc com-parisons between object diameters and hands. All statistical tests
time of object lift-off. Movement time (Figure 2A) was the time from movement initiation to movement termination. Aperture (Figure 2B) was derived from the distance between the thumb and index finger sensors, corrected for finger pad thickness. Grasp preshaping behavior was measured using maximum aper-ture (Amax) and initial maximum aperture velocity (AVinit). Amax (Figure 2B, arrow) was defined as the maximum value from the aperture displacement trajectory, It was observed most often between 250 and 400 milliseconds after movement initia-tion. AVinit (Figure 2C, arrow) was defined as the maximum aperture velocity that coincided with the first down going zero crossing of aperture acceleration (Figure 2D). This event was observed most often within 150 milliseconds after movement onset. All kinematic variables were extracted for each trial using customized automatic computer routines written in MatLab (The MathWorks Inc., Natick, MA). Each trial was visually inspected for veracity and manually corrected using transport acceleration profiles as a more sensitive measure to detect movement initiation, if necessary.
Dependent Measures
Visuomotor transformation of grasping was quantified using Pearson product moment correlation coefficients (r) calculated for the relationship between Amax and AVinit, and Amax and object diameter (20 trials for each object diameter).
Transport-grasp coordination was illustrated using phase plane plots of transport velocity (y-axis) and aperture size
Figure 1 Experimental Setup
Note: The wooden shield was used to occlude participants’ view of their reaching arm and hand at the beginning of the movement to maximize anticipatory control of aperture preshaping behavior (lateral view). Top view shows the relative position of the hand switch (home position), object, and light emitting diode (LED) signal.
Tretriluxana et al / Hemisphere Specific Impairments in Reach-to-Grasp Control After Stroke 5
of hemisphere-specific deficits involved separate compari-sons of the corresponding stroke group and matched non-disabled group. Significance level was set at P < .05. SPSS 14.0 (SPSS Inc, Chicago, IL) statistical software was used for all statistical analyses.
Results
Performance Measure
Overall, the groups accomplished reach-grasp actions suc-cessfully with no miss trials. Collapsed across the 3 object diameters, total movement time in the nondisabled group was significantly shorter for the right compared with the left hand (mean [SD] right hand of nondisabled group = 424.08 [102.1] milliseconds; mean [SD] left hand of nondisabled group = 480.60 [131.97] milliseconds; P < .001]. Participants who had right stroke demonstrated a significantly prolonged movement time relative to their matched nondisabled group (mean [SD] right stroke = 531.83 [204.77] milliseconds; mean [SD] right hand of nondisabled group = 424.08 [102.1] milliseconds; P < .001), whereas participants with left stroke performed the task with comparable time to their matched controls (mean [SD] left stroke = 466.59 [108.51] millisec-onds; left hand nondisabled group = 480.60 [131.97] milli-seconds; P > .05).
Kinematic Measures
Hand path. Average hand paths to each of the 3 object diameters are shown in Figure 3, for 1 representative nondis-abled participant and 4 representative participants poststroke. For the nondisabled participant, hand paths were slightly
Figure 2 Key Variables
Note: A, Transport velocity and movement time; B, aperture with marked maximum aperture (Amax); C, aperture velocity with marked initial aperture velocity (AVinit); D, aperture acceleration with marked first down going zero crossing; and E, transport-grasp coordination expressed by cross correlation coefficient (rmax) as a function of time lag (tmax ).
Figure 3 Hand Paths
Note: Average kinematic profiles aligned at movement onset of 3 object diam-eters (1.2 cm, 2.4 cm, 5.6 cm) for right hand (top panel) and left hand (bottom panel) of a representative nondisabled participant (A and C) and 4 representa-tive participants poststroke (B.1, B.2, D.1, D.2). Each profile is an ensemble average of 20 trials for each object diameter.

6 Neurorehabilitation and Neural Repair
both right (Figure 4A) and left hands (Figure 4C). Additionally, peak transport velocities were nearly identical across the 3 object diameters. In contrast to the matched control, the 2 participants with right stroke exhibited 2 different velocity profiles. Participant S022 demonstrated flatter peak velocities (Figure 4B.1) while S007 scaled peak velocities to object diameter (Figure 4B.2) corresponding to a pattern observed with hand paths. On the contrary, the 2 participants post left stroke were able to accomplish the task with velocity profiles comparable to those of matched nondisabled controls.
Collapsed across the 3 object diameters, significantly lower average peak velocity was observed for the right stroke group compared with the right nondisabled group. In contrast, aver-age peak velocity for the left stroke group was comparable to their nondisabled counterpart (mean [SD] right hand of nondisabled group = 107.95 [20.34] cm/s; mean [SD] right stroke = 95.42 [25.16] cm/s; P < .05; mean [SD] left hand of nondisabled group = 100.05 [22.23] cm/s; mean [SD] left stroke = 100.70 [23.05] cm/s; P > .05).
Grasp aperture. Average grasp aperture to each of the 3 object diameters is shown in Figure 5 for the same partici-pants. For all participants, there were 2 common features of the grasp aperture profiles across hands, distinct phases for grasp preshaping and scaled maximum aperture to object diameter.3
curved for all object diameters and similar for each hand (Figures 3A and 3C). The hand paths of the 3 objects over-lapped for most participants. In Figure 3A, the 3 paths for the right hand of this particular participant appear to diverge mid-way through the path. This exception was the result of variable wrist angles for each object. Overall, for the nondisabled group, there was no effect of object diameter or hand on hand path. In contrast to their matched nondisabled control, hand path varied with object size for the 2 representative partici-pants with right stroke. Participant S022, who had right pri-mary motor cortex lesion (M1), demonstrated a sharp deflection curve while approaching a 5.6-cm cylinder (Figure 3B.1). Additionally, S007, who had right temporoparietal and M1 lesions, scaled hand path distance to the 3 object diameters (Figure 3B.2) and demonstrated an atypical straight path to grasp a 1.2-cm cylinder (usually observed in reaching-only actions) and an atypical curved path deflecting approximately half way of the path during grasping a 2.4-cm cylinder. On the contrary, the 2 representatives post left stroke were able to accomplish the task with hand path profiles comparable to matched nondisabled control.
Transport velocity. Average transport velocity is shown in Figure 4 for 1 representative nondisabled participant and 4 representative participants poststroke. For the nondisabled participant, velocity profiles were single sharply peaked for
Figure 4 Tangential Transport Velocity
-100 0 100 200 300 400 500 600 700 800
ms
0
30
60
90
120
150
cm/s
ND004
1.2 cm2.4 cm5.6 cm
-100 0 100 200 300 400 500 600 700 800
ms
0
30
60
90
120
150
cm/s
S022
-100 0 100 200 300 400 500 600 700 800
ms
0
30
60
90
120
150
cm/s
S007
-100 0 100 200 300 400 500 600 700 800ms
0
30
60
90
120
150
cm/s
ND0004
-100 0 100 200 300 400 500 600 700 800
ms
0
30
60
90
120
150
cm/s
S012
-100 0 100 200 300 400 500 600 700 800ms
0
30
60
90
120
150
cm/s
S004
B.21.B.ARight hand
D.2C. D.1Left hand
Note: Average kinematic profiles aligned at movement onset of 3 object diameters (1.2 cm, 2.4 cm, 5.6 cm) for right hand (top panel) and left hand (bottom panel) of a representative nondisabled participant (A and C) and 4 representative participants poststroke (B.1, B.2, D.1, D.2). Each profile is an ensemble average of 20 trials for each object diameter.

Tretriluxana et al / Hemisphere Specific Impairments in Reach-to-Grasp Control After Stroke 7
control (right hand of nondisabled group). In contrast, the 2 rep-resentative participants post left stroke showed less anticipatory scaling of preshaping and appeared to overestimate maximum aperture compared to the matched nondisabled representative (left hand of nondisabled group). These hemisphere-specific deficits are shown in the bar plots of group data (Figures 6A and 6B). For early initial aperture, there was a significantly higher r between initial aperture velocity and peak aperture for the right stroke group compared to their matched control (r for right hand of nondisabled group = 0.10; r for right stroke = 0.61; P < .001) whereas there was lower r for the left stroke (r for left hand of nondisabled group = 0.6; r for left stroke = 0.55; P =.15). For maximum aperture, considered a relatively late measure of the visuomotor transformation, there was no difference in r between maximum aperture and object diame-ter of right stroke relative to right hand of disabled group, whereas there was a significantly lower r for left stroke than for their matched control (r for left hand of disabled group = 0.94; r for left stroke = 0.76; P < .05). A microanalysis was performed to identify the locus of the reduction in r. As expected, there was a significantly greater maximum aperture with smaller objects only for left stroke, not right stroke, com-pared to their control group (Figure 6C). Overall, grasp- prescaling capability for stroke groups was opposite of what was observed in their matched controls. This capability of the
For nondisabled participants, although no difference between hands in maximum aperture was observed (P > .05), the method of achieving maximum was different between hands. As shown for one representative participant, regardless of object diameter, the right hand reached maximum aperture through a similar initial aperture velocity (Figure 5A), whereas the left hand showed a proportional scaling of initial velocity (Figure 5C). This proportional scaling is suggestive of antici-patory control of visuomotor transformation.3 The nature of this feature is shown in the bar plots of group data (Figure 6A). Maximum aperture, considered a relatively late measure of the visuomotor transformation, for reach and grasp demonstrated no between-hand differences. In contrast, the earlier initial aperture was quite sensitive to hand difference (r for left hand of nondisabled group = 0.62, P < .001; r for right hand of nondisabled group = 0.04, P > .05). Overall, for the nondis-abled group, there was a robust manual asymmetry for initial grasp preshaping to object diameter.
Matched limb comparisons between the 2 stroke groups and their nondisabled counterparts revealed hemisphere- specific deficits. Both participants post right stroke showed an initial anticipatory scaling of aperture preshaping to object diameter (Figures 5B.1 and 5B.2), which was not observed in the matched nondisabled representative (Figure 5A), as well as achieved the comparable maximum aperture as their matched
-100 0 100 200 300 400 500 600 700 800ms
0
2
4
6
8
10
12
14
cm
ND004
1.2cm2.4cm5.6cm
-100 0 100 200 300 400 500 600 700 800ms
0
2
4
6
8
10
12
14
cm
S022
-100 0 100 200 300 400 500 600 700 800ms
0
2
4
6
8
10
12
14
cm
S007
-100 0 100 200 300 400 500 600 700 800ms
0
2
4
6
8
10
12
14
cm
ND004
-100 0 100 200 300 400 500 600 700 800ms
0
2
4
6
8
10
12
14
cm
S012
-100 0 100 200 300 400 500 600 700 800ms
0
2
4
6
8
10
12
14
cm
S004
B.21.B.A
D.2C. D.1
Right hand
Left hand
Figure 5 Aperture Displacement for Each Object Diameter
Note: Average kinematic profiles aligned at movement onset of 3 object diameters (1.2 cm, 2.4 cm, 5.6 cm) for right hand (top panel) and left hand (bottom panel) of a representative nondisabled participant (A and C) and 4 representative participants poststroke (B.1, B.2, D.1, D.2). Each profile is an ensemble average of 20 trials for each object diameter.

8 Neurorehabilitation and Neural Repair
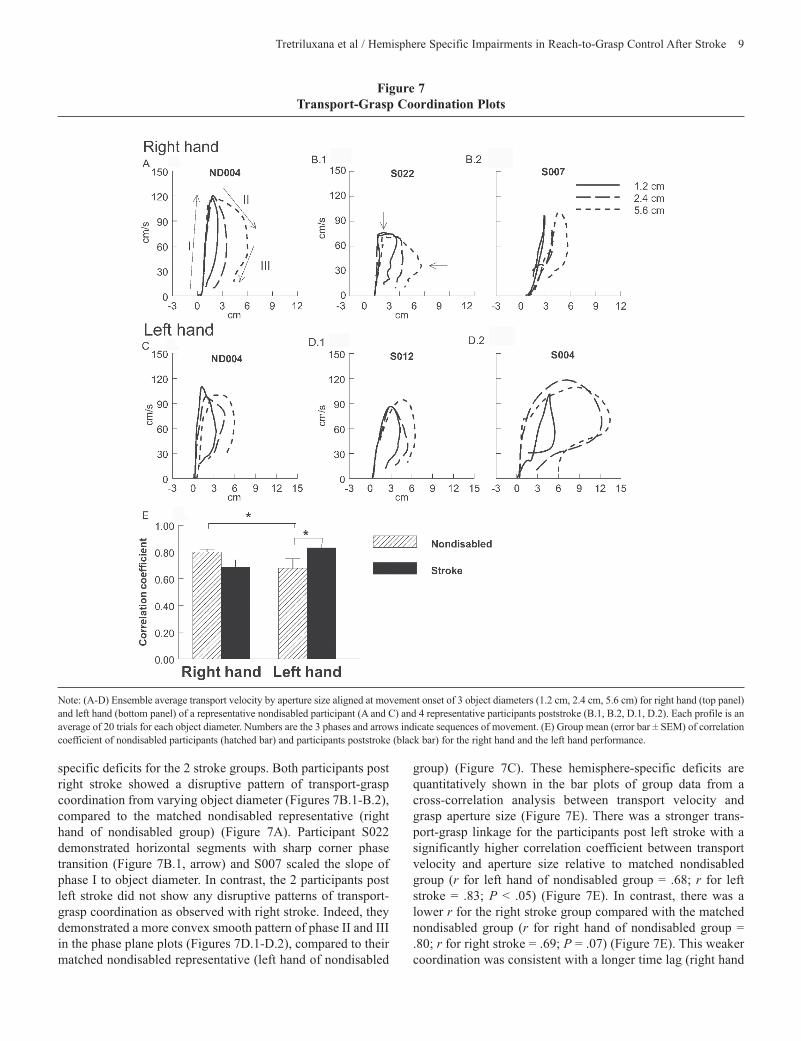
Visualization of the whole pattern, revealed a smooth con-vex segment for the right hand (Figure 7A), suggestive of parallel processing of transport and grasp.3 In contrast, the left hand demonstrated a relatively horizontal phase II trajectory with a sharp corner at the phase transition (Figure 7C.). This pattern is suggestive of sequential processing of transport and grasp.3 The nondisabled (ND) group demonstrated a stronger transport-grasp linkage for the right hand with a significantly higher correlation between transport velocity and aperture size (r for right hand of nondisabled group = 0.80; r for left hand of nondisabled group = 0.68; P < .05) (Figure 7E.). Overall, for the nondisabled group, the 2 hands accomplished the task of adapting to 3 object diameters using different coordinated patterns with a stronger transport-grasp relationship for the right hand.
Matched limb comparisons between the 2 stroke groups and their nondisabled counterparts revealed hemisphere
right hand system increased after the right hemispheric stroke, whereas those of the left hand system decreased after the left hemisphere was damaged.
Transport-grasp coordination. The phase plane plots for the 3 object diameters are shown in Figures 7A to 7D for the same participants. The plots for all participants reveal 3 funda-mental phases. Phase I consists of the acceleration phase of transport velocity with a preparatory preshaping. Phase II begins near the time of peak transport velocity and includes the later phase of preshaping that begins with rapid finger opening. Maximum aperture is achieved during transport deceleration. Phase III commences during the last half of the deceleration phase of transport velocity and includes aperture closure that terminates the reach-to-grasp action (Figure 7A, number indicates each phase and arrows are sequences of movement).
Figure 6 Visuomotor Transformation of Grasping Plots
Right hand Left hand0.00
0.20
0.40
0.60
0.80
1.00
Co
rrel
atio
n c
oef
fici
ent
A. AVinit and AmaxNondisabled
Stroke
Right hand Left hand0.00
0.20
0.40
0.60
0.80
1.00
Co
rrel
atio
n c
oef
fici
ent
B. Amax and object diameter
0 2 4 6 8 10
Object diameter (cm)
0
2
4
6
8
10
Max
imu
m a
per
ture
(cm
)
C.1 Right hand performance
RND RS
0 2 4 6 8 10
Object diameter (cm)
0
2
4
6
8
10
Max
imu
m a
per
ture
(cm
)
C.2 Left hand performanceLND LS
****
*
**
Note: Bar plot of group mean (error bar = SEM) correlation coefficient of nondisabled participants (hatched bar) and participants poststroke (black bar) for maximum aperture and object diameter (A), and maximum aperture and initial aperture velocity (B). (C) Group mean (error bar = SEM) of maximum aperture, the diagonal broken line indicates a 1:1 ratio between maximum aperture and object diameter (slope = 1) across trials for 3 object diameters of nondisabled participants (empty symbols) and participants poststroke (filled symbols) for the right hand performance (C.1) and the left hand performance (C.2).
Tretriluxana et al / Hemisphere Specific Impairments in Reach-to-Grasp Control After Stroke 9
group) (Figure 7C). These hemisphere-specific deficits are quantitatively shown in the bar plots of group data from a cross-correlation analysis between transport velocity and grasp aperture size (Figure 7E). There was a stronger trans-port-grasp linkage for the participants post left stroke with a significantly higher correlation coefficient between transport velocity and aperture size relative to matched nondisabled group (r for left hand of nondisabled group = .68; r for left stroke = .83; P < .05) (Figure 7E). In contrast, there was a lower r for the right stroke group compared with the matched nondisabled group (r for right hand of nondisabled group = .80; r for right stroke = .69; P = .07) (Figure 7E). This weaker coordination was consistent with a longer time lag (right hand
specific deficits for the 2 stroke groups. Both participants post right stroke showed a disruptive pattern of transport-grasp coordination from varying object diameter (Figures 7B.1-B.2), compared to the matched nondisabled representative (right hand of nondisabled group) (Figure 7A). Participant S022 demonstrated horizontal segments with sharp corner phase transition (Figure 7B.1, arrow) and S007 scaled the slope of phase I to object diameter. In contrast, the 2 participants post left stroke did not show any disruptive patterns of transport-grasp coordination as observed with right stroke. Indeed, they demonstrated a more convex smooth pattern of phase II and III in the phase plane plots (Figures 7D.1-D.2), compared to their matched nondisabled representative (left hand of nondisabled
Figure 7 Transport-Grasp Coordination Plots
Note: (A-D) Ensemble average transport velocity by aperture size aligned at movement onset of 3 object diameters (1.2 cm, 2.4 cm, 5.6 cm) for right hand (top panel) and left hand (bottom panel) of a representative nondisabled participant (A and C) and 4 representative participants poststroke (B.1, B.2, D.1, D.2). Each profile is an average of 20 trials for each object diameter. Numbers are the 3 phases and arrows indicate sequences of movement. (E) Group mean (error bar ± SEM) of correlation coefficient of nondisabled participants (hatched bar) and participants poststroke (black bar) for the right hand and the left hand performance.

10 Neurorehabilitation and Neural Repair
movements that involve distal muscles (eg, grasp preshaping) through contralateral reticulospinal and vestibulospinal neu-rons.33 Together, the evidence suggests that the ipsilateral CST projections may play only a minor role in the expression of hypofunctional deficits (Figure 8, thin dashed lines). Instead, it is likely that contralateral CST projection (Figure 8, solid diagonal lines) through transcallosal communication (Figure 8, horizontal arrows) plays a more substantial role in the expres-sion of ipsilesional hypofunction.
The hyperfunction deficit is thought to reflect the special-ized contribution of the nonlesioned hemisphere that is unmasked or disinhibited by the stroke-affected hemisphere (Figure 8, thinner horizontal arrows). This idea is supported by recent evidence of altered interhemispheric communication after experimentally induced (eg, virtual lesion) and naturally from stroke in humans.17,34,35 For example, hyperfunction of motor performance in the ipsilesional hand was identified after transient virtual cortical lesions using repetitive TMS in humans36 and after an ischemic lesion in a rodent model of stroke. After hemisphere damage, because of the altered tran-scallosal communication,17,34,35 there is a downregulation of the damaged hemisphere and an upregulation of the nondam-aged hemisphere. This effectively allows the expression of the upregulated hemisphere’s unique contributions to be unmasked more so than in the nondisabled condition. The following dis-cussion puts our hemisphere specific findings in the context of other work concerning praxis and the visual processing system.
The left hemisphere is specialized for grasp preshaping. The grasping deficits we observed illustrate the preferential role of the left hemisphere for grasp preshaping. This deficit that was unique to those with left hemisphere stroke has not been observed previously for natural grasping actions.22,37 This may be the result of 2 experimental manipulations used in our study: (1) vision of the arm and hand at the beginning of the movement path was occluded by a wooden shield and (2) par-ticipants were encouraged to complete the task as quickly as possible. These 2 factors may have magnified the left hemi-sphere deficits by minimizing the use of visual guidance and external cues in the environment. However, a recent study using the Mueller–Lyer illusion in those with either right or left hemisphere stroke found that only those with left hemi-sphere stroke were susceptible to the illusion when grasping the target with their left hand.38
Another interpretation of a left hemisphere advantage for grasping control may be related to the praxis system, that is, the representation of acquired manual skills including tool use39 and pantomimed prehension.40 In both right- and left-handed indi-viduals postcallosotomy, Frey and coworkers observed that the right hand more accurately pantomimed tool use such as ham-mering.39 Similarly, when participants post left stroke were asked to pantomime reach-to-grasp and drink actions, left hand aperture preshaping was either absent or significantly reduced relative to matched nondisabled controls.40 Though pantomime cannot easily be translated to actual task execution, like in the
of nondisabled group = 93.88 milliseconds; right stroke = 140.26 milliseconds; P = .17) and a significantly higher stan-dard deviation of the time lag (right hand of nondisabled group = 33.63 milliseconds; right stroke = 88.24 milliseconds; P < .001) observed in right stroke, but not left stroke, compared to matched controls. Overall, for stroke groups, the transport-grasp linkage for the left hand increased after the left hemi-spheric stroke, whereas this linkage for the right hand decreased after the right hemisphere was damaged.
Discussion
We observed hemisphere-specific deficits in reach-to-grasp movements of the ipsilesional limb in patients with unilateral stroke through experimental manipulation of object size. Specifically, compared with the matched-limb of the nondis-abled control group, we found that after left hemisphere stroke there was deficient scaling ability and a tendency for a delayed anticipatory grasp preshaping and a stronger transport-grasp linkage. In contrast, after right hemisphere stroke, there was earlier anticipatory grasp preshaping and a weaker transport-grasp coordination.
The comparison with nondisabled participants revealed 2 kinds of hemisphere-specific deficits: (1) an amplification of normal function or hyperfunction and (2) a reduction in func-tion or hypofunction. For the left hemisphere stroke group there was hypofunction for grasp preshaping, and hyperfunc-tion for transport-grasp coordination (Figure 8A). For the right hemisphere group the opposite pattern was observed; there was hyperfunction for grasp preshaping and hypofunction for transport-grasp coordination (Figure 8B). One plausible model for the expression of these 2 deficits in relation to control emerges from the bilateral model contribution that includes appropriately weighted projections from each hemisphere and the known interhemispheric communication illustrated in Figure 8. This bilateral model predicts that after unilateral stroke, there would be 2 types of performance deficits revealed by performance of the ipsilesional limb: (1) hypofunction of the specialized contribution from the lesioned hemisphere and (2) hyperfunction of the specialized contributions that are spe-cific to the nonlesioned hemisphere. Using this model, our findings suggest that the left hemisphere is specialized for grasp preshaping and the right hemisphere is specialized for transport-grasp coordination (Figure 8).
The hypofunctional ipsilesional deficits are consistent with previous studies that have used aiming and reaching tasks.7,18,25 Commonly, the hypofunctional deficit is attributed to the cognitive-motor requirements of the task.25,29,30 The reach-to-grasp task used in our experiment likely engages the reach and grasp pathways within each hemisphere, the 2 specialized dorsal stream channels of visuomotor transformation. It is estimated that 10% to 15% of the corticospinal tract (CST) axons project to ipsilateral spinal motoneurons that innervate primarily proximal muscles.31,32 A recent review suggested that the ipsilateral CST may contribute indirectly to ipsilateral

Tretriluxana et al / Hemisphere Specific Impairments in Reach-to-Grasp Control After Stroke 11
not the right temporoparietal area (S007), demonstrated the most profound grasp preshaping deficits. The temporoparietal area damaged by the stroke includes components of both ven-tral and dorsal streams and is adjacent to AIP. The fact that this deficit was observed after left, but not right temporoparietal area, leads us to speculate that the exaggeration of aperture estimation in the 2 participants may be a reflection of stroke-induced disruption of a left hemisphere putative intracortical network that includes AIP. However, more systematic investi-gations into the laterality issue are needed before any defini-tive conclusion can be made as to the specializations within the left and right AIP.
An alternative consideration for the exaggeration of aper-ture estimation concerns the speed instruction that was suc-cessfully implemented by participants with left hemisphere
present experiment, it is possible that the processing associated with praxis also mediates aperture preshaping.
A specific neuroanatomical locus of grasp perception and grasp formation has been identified both in lesioned popula-tions and imaging studies. Goodale and Milner proposed that the visual mechanisms of object perception and the action of grasping are mediated separately through ventral (temporooc-cipital cortex) and dorsal (parietooccipital cortex) streams.41 This idea was later supported by imaging studies.42,43 Although a distinct lateralization of these pathways has not been system-atically investigated, recent functional MRI (fMRI) and TMS studies have shown that the left anterior intraparietal (AIP) cortex, which is part of the dorsal stream, plays a key role in visually guided grasping.44,45 In our experiment, participants with a lesion in the left temporoparietal area (S004 and S020),
Figure 8 Bilateral Model for Hemispheric Specialization of Unimanual Control
Ipsilesional deficits
L R
Ipsilesional deficits
B. Right hemisphere stroke
L R
Hypofunction: weaker trans port-grasp coordination Hyperfunction: earlier antici patory grasp pre-shaping
Hypofunction: deficient scaling ability and later anticipatory grasp pre-shaping Hyperfunction: stronger transport-grasp coordination
A. Left hemisphere stroke
Note: (A) Right hemisphere stroke model. (B) Left hemisphere stroke model. Solid diagonal lines indicate contralateral projection, thin dash lines indicate ipsi-lateral projection, and horizontal arrows represent interhemispheric communications.

12 Neurorehabilitation and Neural Repair
more convincing verification of the role of the right hemi-sphere in transport-grasp coordination.
In summary, our results provide support for hemisphere-specific deficits in ipsilesional limb performance with deficient grasp scaling and stronger transport-grasp coordination after left hemisphere stroke, and weaker transport-grasp coordination with earlier anticipatory control of grasp preshaping after right hemisphere stroke. This pattern of findings is explained using a bilateral model in which the left hemisphere is specialized for grasp preshaping, whereas the right hemisphere may be more specialized for transport-grasp coordination.
Acknowledgments
We thank Dr Allan Wu for DataWizard programming con-sultation and Dr Michael A. Arbib for critical comments on an earlier version of this manuscript.
References
1. Eviatar Z, Hellige JB, Zaidel E. Individual differences in lateralization: effects of gender and handedness. Neuropsychology. 1997;11:562-576.
2. Hellige JB. Hemispheric Asymmetry: What’s Right and What’s Left. Cambridge, MA: Harvard University Press; 2001.
3. Tretriluxana J, Gordon J, Winstein CJ. Manual asymmetries in grasp preshap-ing and transport-grasp coordination. Exp Brain Res. 2008;188:305-315.
4. Haaland KY, Harrington D. The role of the hemispheres in closed loop movements. Brain Cogn. 1989;9:158-180.
5. Peters M. Prolonged practice of a simple motor task by preferred and nonpreferred hands. Percept Mot Skills. 1976;42:447-450.
6. Haaland KY, Harrington D. Hemispheric control of the initial and corrective components of aiming movements. Neuropsychologia. 1989;27:961-969.
7. Winstein CJ, Pohl PS. Effects of unilateral brain damage on the control of goal-directed hand movements. Exp Brain Res. 1995;105:163-174.
8. Cramer SC, Finklestein SP, Schaechter JD, Bush G, Rosen BR. Activation of distinct motor cortex regions during ipsilateral and contralateral finger movements. J Neurophysiol. 1999;81:383-387.
9. Kim S-G, Ashe J, Hendrich K, et al. Functional magnetic resonance imag-ing of motor cortex: Hemispheric asymmetry and handedness. Science. 1993;261:615-617.
10. Urbano A, Babiloni C, Onorati P, Babiloni F. Human cortical activity related to unilateral movements. A high resolution EEG study. Neuroreport. 1996;8:203-206.
11. Ziemann U, Ishii K, Borgheresi A, et al. Dissociation of the pathways mediating ipsilateral and contralateral motor-evoked potentials in human hand and arm muscles. J Physiol. 1999;518:895-906.
12. Di Lazzaro V, Rothwell JC, Oliviero A, et al. Intracortical origin of the short latency facilitation produced by pairs of threshold magnetic stimuli applied to human motor cortex. Exp Brain Res. 1999;129:494-499.
13. Ferbert A, Priori A, Rothwell JC, Day BL, Colebatch JG, Marsden CD. Interhemispheric inhibition of the human motor cortex. J Physiol. 1992; 453:525-546.
14. Harris-Love M, Cohen LG. High level bilateral talks. Focus on “effect of low-frequency repetitive transcranial magnetic stimulation on interhemi-spheric inhibition.” J Neurophysiol. 2005;94:1664-1665.
15. Daskalakis ZJ, Christensen BK, Fitzgerald PB, Roshan L, Chen R. The mechanisms of interhemispheric inhibition in the human motor cortex. J Physiol. 2002;543:317-326.
16. Kobayashi M, Hutchinson S, Schlaug G, Pascual-Leone A. Ipsilateral motor cortex activation on functional magnetic resonance imaging during unilateral hand movements is related to interhemispheric interactions. Neuroimage. 2003;20:2259-2270.
stroke, and possibly causing a speed-accuracy trade-off. To test this possibility, we ran a post hoc correlation analysis between total movement time and maximum grasp aperture of all trials within the left stroke group. There was no association between the time that the task was completed and maximum distance that the fingers were opened. This provides evidence against a speed-accuracy trade-off explanation, but instead supports an inherent left hemisphere specialization for grasp preshaping.
The right hemisphere is specialized for transport-grasp coordination. The weaker coordination after right hemisphere stroke, but not left, illustrates the preferential role of the right hemisphere for coordinated transport-grasp control. The lower transport velocities are consistent with previous findings in which a right stroke group performed reach, grasp, and drink actions using their ipsilesional arm.40
One explanation for weaker coordination after right hemi-sphere stroke pertains to its preferential role for processing global features of the workspace including hand position, the wooden shield, and object location. This contrasts with a left hemisphere specialization for processing local features of the object/target itself.2,46,47 This distinction in visual information processing has been attributed to a differential sensitivity of the hemispheres to extract spatial frequency information: the low spatial frequencies represent the global information, whereas high spatial frequencies highlight local information.47,48 Global processing might be used to allow the right hemisphere to effec-tively coordinate the speed of arm movement and the magnitude of finger opening and closing. Alternatively, compared to the left hemisphere, the right hemisphere might use a global infor-mation processing mode to direct spatial-attention over a larger portion of the visual field.49
Our interpretation of a right hemisphere specialization for transport-grasp coordination differs from that of studies that used a reaching-only task.7,19,20,50,51 This discrepancy may be because of the different task paradigm. The visuomotor prob-lem of coordinating transport and grasp through dynamic online communication between reach and grasp pathways is most likely a challenge to the nervous system, one that distin-guishes transport-grasp coordination from other forms of coordination, including intersegmental coordination in reach-only actions20,52 and spatial-segmental coordination between arm and trunk movements.19
One limitation of this study was that the effect of hypofunc-tion in transport-grasp coordination was not always reliable. One possible explanation for this was that we used object size as a task constraint, which in previous studies had been shown to be a more direct perturbation to grasping behavior than transport behavior.53,54 To more directly investigate the right hemisphere specialization for transport-grasp coordination, we conducted an experiment that used a barrier paradigm task.
The barrier task has been shown previously to perturb the transport path and thereby more likely to disrupt the linkage between transport and grasp components.55,56 Using this more direct perturbation to transport-grasp coordination allowed a

Tretriluxana et al / Hemisphere Specific Impairments in Reach-to-Grasp Control After Stroke 13
38. Radoeva PD, Cohen JD, Corballis PM, Lukovits TG, Koleva SG. Hemispheric asymmetry in a dissociation between the visuomotor and visuoperceptual streams. Neuropsychologia. 2005;43:1763-1773.
39. Frey SH, Funnell MG, Gerry VE, Gazzaniga MS. A dissociation between the representation of tool-use skills and hand dominance: insights from left- and right-handed callosotomy patients. J Cogn Neurosci. 2005;17:262-272.
40. Laimgruber K, Goldenberg G, Hermsdorfer J. Manual and hemispheric asymmetries in the execution of actual and pantomimed prehension. Neuropsychologia. 2005;43:682-692.
41. Goodale MA, Milner AD. Separate visual pathways for perception and action. Trends Neurosci. 1992;15:20-25.
42. Castiello U. The neuroscience of grasping. Nat Rev Neurosci. 2005;6: 726-736.
43. Culham JC, Kanwisher NG. Neuroimaging of cognitive functions in human parietal cortex. Curr Opin Neurobiol. 2001;11:157-163.
44. Frey SH, Vinton D, Norlund R, Grafton ST. Cortical topography of human anterior intraparietal cortex active during visually guided grasping. Brain Res Cogn Brain Res. 2005;23:397-405.
45. Tunik E, Frey SH, Grafton ST. Virtual lesions of the anterior intraparietal area disrupt goal-dependent on-line adjustments of grasp. Nat Neurosci. 2005;8:505-511.
46. Fink GR, Marshall JC, Halligan PW, Frith CD, Frackowiak RS, Dolan RJ. Hemispheric specialization for global and local processing: the effect of stimulus category. Proc Biol Sci. 1997;264:487-494.
47. Ivry RB, Robertson LC. The Two Sides of Perception. Cambridge, MA: MIT Press; 1997.
48. Serrien DJ, Ivry RB, Swinnen SP. Dynamics of hemispheric specialization and integration in the context of motor control. Nat Rev Neurosci. 2006;7:160-166.
49. Evert DL, McGlinchey-Berroth R, Verfaellie M, Milberg WP. Hemispheric asymmetries for selective attention apparent only with increased task demands in healthy participants. Brain Cogn. 2003;53:34-41.
50. Sainburg RL. Handedness: differential specializations for control of tra-jectory and position. Exerc Sport Sci Rev. 2005;33:206-213.
51. Sainburg RL, Duff SV. Does motor lateralization have implications for stroke rehabilitation? J Rehabil Res Dev. 2006;43:311-322.
52. Sainburg RL. Evidence for a dynamic-dominance hypothesis of handed-ness. Exp Brain Res. 2002;142:241-258.
53. Paulignan Y, Frak VG, Toni I, Jeannerod M. Influence of object position and size on human prehension movements. Exp Brain Res. 1997;114:226-234.
54. Marteniuk RG, Leavitt JL, MacKenzie CL, et al. Functional relationships between grasp and transport components in a prehension task. Hum Mov Sci. 1990;9:149-176.
55. Haggard P, Wing AM. Remote responses to perturbation in human prehen-sion. Neurosci Lett. 1991;122:103-108.
56. Saling M, Alberts J, Stelmach GE, Bloedel JR. Reach-to-grasp movements during obstacle avoidance. Exp Brain Res. 1998;118:251-258.
57. Mathiowetz V, Kashman N, Volland G, Weber K, Dowe M, Rogers S. Grip and pinch strength: normative data for adults. Arch Phys Med Rehabil. 1985;66:69-74.
17. Murase N, Duque J, Mazzocchio R, Cohen LG. Influence of interhemi-spheric interactions on motor function in chronic stroke. Ann Neurol. 2004;55:400-409.
18. Goodale MA. Hemispheric differences in motor control. Behav Brain Res. 1988;30:203-214.
19. Esparza DY, Archambault PS, Winstein CJ, Levin MF. Hemispheric spe-cialization in the coordination of arm and trunk movements during point-ing in patients with unilateral brain. Exp Brain Res. 2003;148:488-497.
20. Haaland KY, Prestopnik JL, Knight RT, Lee RR. Hemispheric asymmetries for kinematic and positional aspects of reaching. Brain. 2004;127:1145-1158.
21. Hermsdorfer J, Laimgruber K, Kerkhoff G, Mai N, Goldenberg G. Effects of unilateral brain damage on grip selection, coordination, and kinematics of ipsilesional prehension. Exp Brain Res. 1999;128:41-51.
22. Farne A, Roy AC, Paulignan Y, et al. Visuomotor control of the ipsilateral hand: evidence from right brain-damaged patients. Neuropsychologia. 2003;41:739-757.
23. Bamford J, Sandercock P, Dennis M, Burn J, Warlow C. Classification and natural history of clinically identifiable subtypes of cerebral infarction. Lancet. 1991;337:1521-1526.
24. Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 1971;9:97-113.
25. Haaland KY, Harrington DL. Hemispheric asymmetry of movement. Curr Opin Neurobiol. 1996;6:796-800.
26. Pohl P, Winstein CJ, Onla-or S. Sensory-motor control in the ipsilesional upper extremity after stroke. NeuroRehabilitation. 1997;9:57-69.
27. Winter DA. Biomechanics and Motor Control of Human Movement. New York: John Wiley & Sons; 2004.
28. Winstein CJ, Garfinkel A. Qualitative dynamics of disordered human locomotion: a preliminary investigation. J Mot Behav. 1989;21:373-391.
29. Haaland KY, Harrington D, Yeo R. The effects of task complexity in left and right CVA patients. Neuropsychologia. 1987;25:783-794.
30. Haaland KY, Harrington DL. Limb-sequencing deficits after left but not right hemisphere damage. Brain Cogn. 1994;24:104-122.
31. Colebatch JG, Deiber MP, Passingham RE, Friston KJ, Frackowiak RS. Regional cerebral blood flow during voluntary arm and hand movements in human subjects. J Neurophysiol. 1991;65:1392-1401.
32. Wassermann EM, Pascual-Leone A, Hallett M. Cortical motor representa-tion of the ipsilateral hand and arm. Exp Brain Res. 1994;100:121-132.
33. Jankowska E, Edgley SA. How can corticospinal tract neurons contribute to ipsilateral movements? A question with implications for recovery of motor functions. Neuroscientist. 2006;12:67-79.
34. Duque J, Hummel F, Celnik P, Murase N, Mazzocchio R, Cohen LG. Transcallosal inhibition in chronic subcortical stroke. Neuroimage. 2005;28:940-946.
35. Liepert J, Storch P, Fritsch A, Weiller C. Motor cortex disinhibition in acute stroke. Clin Neurophysiol. 2000;111:671-676.
36. Kobayashi M, Hutchinson S, Théoret H, Schlaug G, Pascual-Leone A. Repetitive TMS of the motor cortex improves ipsilateral sequential simple finger movements. Neurology. 2004;62:91-98.
37. Hermsdörfer J, Ulrich S, Marquardt C, Goldenberg G, Mai N. Prehension with the ipsilesional hand after unilateral brain damage. Cortex. 1999;35:139-161.
For reprints and permission queries, please visit SAGE’s Web site at http://www.sagepub.com/journalsPermissions.nav




















