
Habitat selective factors influencing the distribution of larger benthic foraminiferal assemblages...
9
INTERNAL ARCHITECTURE OF MIOCENE PSEUDOTABERINA AND ITS RELATION TO CARIBBEAN ARCHAIASINS by WILLEM RENEMA Nationaal Natuurhistorisch Museum Naturalis, PO Box 9517, 2300 RA Leiden, the Netherlands; e-mail: [email protected], and Centre for Marine Studies, University of Queensland, Brisbane, Qld 4072, Australia Typescript received 31 January 2006: accepted in revised form 8 January 2007 Abstract: Well-preserved, isolated specimens of Pseudotabe- rina were found at three localities in Indonesia, thus enabling the study of the relationships between this genus and Carib- bean Archaiasinae. The internal architecture of the chambers of Pseudotaberina consists of four layers of stolon planes, each of which is divided by superposed septula into chamb- erlets. The chamberlets are connected to the next chambers by foramina opposing the septula. The stolons are Y-shaped. All other Archaiasinae have radial stolon axes combined with an initial involute growth stage, whereas the Soritinae have cross-wise oblique stolon axes, but with evolute growth in the early stages. Pseudotaberina shows a combination of the characters of these two subfamilies, confirming the need to study more Tethyan material to elucidate phylogenetic rela- tionships within this group of large benthic foraminifera. Key words: Archaiasinae, Soritinae, large benthic foramini- fera, Indonesia, Burdigalian. Phylogenetic analyses of the Soritidae have revealed ambiguous results with regard to the monophyly of the subfamily Archaiasinae. Analyses based on morphological characters show a monophyletic grouping of all Archaiasi- nae as a sister group to the Soritinae (Gudmundsson 1994; Richarson 2001). The analysis carried out by Gudmundsson (1994) only involved extant taxa, but Richarson (2001) included fossil taxa as well. In the latter analysis both Nemophora floridana (=Miarchaias florid- anus) and N. malabarica (= Pseudotaberina malabarica) were included and they grouped as sister taxa in a clade ancestral to the Recent species of Archaias. This would imply a palaeobiogeographical connection between the Middle and Far East and the Caribbean for which hardly any other evidence is available. However, Holzman et al. (2001) found in a molecular phylogeny of the Soritacea, obviously restricted to living taxa only, that taxa without an endoskeleton form a paraphyletic group within the sub- clade Archaiasinae. Taxa with an endoskeleton are derived independently from basal taxa without endoskeletons. To understand the phylogenetic relationships in fossil and Recent large benthic foraminifera (LBF) an analysis of the complex internal structure is necessary. This com- plicates the study of fossil taxa, since only exceptionally well preserved, isolated specimens allow for this kind of detailed investigation. Well-preserved, isolated specimens from shallow, in particular reef-associated, habitats are even rarer. One of the groups in which phylogenetic rela- tionships are unclear is the soritids, and especially the archaiasins. At least six species of the Archaiasinae occur in the Caribbean province, whereas only one genus occurs in the Indo-West Pacific (Hallock 1988; Hallock and Peebles 1993; Holzman et al. 2001). All species harbour chlorophytes as symbionts and occur in shallow environments. Three genera of Archaiasins have been described from the Upper Miocene of the Dominican Republic (Hottin- ger 2001). The preservation of this material was sufficient to allow a full description of the microstructure of these species, as well as of some co-occuring soritins. Hottinger concluded that these species are unlikely to be candidates for a direct ancestry of the species living today in the Caribbean province, and he described several new species and genera from this material. The taxa reflect differences in the apertural face and the respective arrangement of radial partitions (Hottinger 2001). One of them, Miarcha- ias meander Hottinger, 2001, develops cyclical agamonts with a diameter up to 1.5 cm and with meandering struc- tures covering the lateral surface of the disc, while smaller species do not develop cyclical growth. The Tethyan genus Pseudotaberina Eames, in Davies 1971 (emended by Banner and Highton 1989) has the same basic architec- ture. These descriptions are based on material in thin sec- tions. In the present study, isolated specimens of Pseudotaberina, allowing study of the internal architec- ture, are described from the Miocene of Java. These turn out to have a different architecture, and thus the genera are not synonymous. [Palaeontology, Vol. 51, Part 1, 2008, pp. 71–79] ª The Palaeontological Association doi: 10.1111/j.1475-4983.2007.00731.x 71
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Habitat selective factors influencing the distribution of larger benthic foraminiferal assemblages...

INTERNAL ARCHITECTURE OF MIOCENE
PSEUDOTABERINA AND ITS RELATION TO
CARIBBEAN ARCHAIASINS
by WILLEM RENEMANationaal Natuurhistorisch Museum Naturalis, PO Box 9517, 2300 RA Leiden, the Netherlands; e-mail: [email protected], and Centre for Marine Studies,
University of Queensland, Brisbane, Qld 4072, Australia
Typescript received 31 January 2006: accepted in revised form 8 January 2007
Abstract: Well-preserved, isolated specimens of Pseudotabe-
rina were found at three localities in Indonesia, thus enabling
the study of the relationships between this genus and Carib-
bean Archaiasinae. The internal architecture of the chambers
of Pseudotaberina consists of four layers of stolon planes,
each of which is divided by superposed septula into chamb-
erlets. The chamberlets are connected to the next chambers
by foramina opposing the septula. The stolons are Y-shaped.
All other Archaiasinae have radial stolon axes combined with
an initial involute growth stage, whereas the Soritinae have
cross-wise oblique stolon axes, but with evolute growth in
the early stages. Pseudotaberina shows a combination of the
characters of these two subfamilies, confirming the need to
study more Tethyan material to elucidate phylogenetic rela-
tionships within this group of large benthic foraminifera.
Key words: Archaiasinae, Soritinae, large benthic foramini-
fera, Indonesia, Burdigalian.
Phylogenetic analyses of the Soritidae have revealed
ambiguous results with regard to the monophyly of the
subfamily Archaiasinae. Analyses based on morphological
characters show a monophyletic grouping of all Archaiasi-
nae as a sister group to the Soritinae (Gudmundsson
1994; Richarson 2001). The analysis carried out by
Gudmundsson (1994) only involved extant taxa, but
Richarson (2001) included fossil taxa as well. In the latter
analysis both Nemophora floridana (=Miarchaias florid-
anus) and N. malabarica (= Pseudotaberina malabarica)
were included and they grouped as sister taxa in a clade
ancestral to the Recent species of Archaias. This would
imply a palaeobiogeographical connection between the
Middle and Far East and the Caribbean for which hardly
any other evidence is available. However, Holzman et al.
(2001) found in a molecular phylogeny of the Soritacea,
obviously restricted to living taxa only, that taxa without
an endoskeleton form a paraphyletic group within the sub-
clade Archaiasinae. Taxa with an endoskeleton are derived
independently from basal taxa without endoskeletons.
To understand the phylogenetic relationships in fossil
and Recent large benthic foraminifera (LBF) an analysis
of the complex internal structure is necessary. This com-
plicates the study of fossil taxa, since only exceptionally
well preserved, isolated specimens allow for this kind of
detailed investigation. Well-preserved, isolated specimens
from shallow, in particular reef-associated, habitats are
even rarer. One of the groups in which phylogenetic rela-
tionships are unclear is the soritids, and especially the
archaiasins. At least six species of the Archaiasinae occur
in the Caribbean province, whereas only one genus occurs
in the Indo-West Pacific (Hallock 1988; Hallock and
Peebles 1993; Holzman et al. 2001). All species harbour
chlorophytes as symbionts and occur in shallow
environments.
Three genera of Archaiasins have been described from
the Upper Miocene of the Dominican Republic (Hottin-
ger 2001). The preservation of this material was sufficient
to allow a full description of the microstructure of these
species, as well as of some co-occuring soritins. Hottinger
concluded that these species are unlikely to be candidates
for a direct ancestry of the species living today in the
Caribbean province, and he described several new species
and genera from this material. The taxa reflect differences
in the apertural face and the respective arrangement of
radial partitions (Hottinger 2001). One of them, Miarcha-
ias meander Hottinger, 2001, develops cyclical agamonts
with a diameter up to 1.5 cm and with meandering struc-
tures covering the lateral surface of the disc, while smaller
species do not develop cyclical growth. The Tethyan
genus Pseudotaberina Eames, in Davies 1971 (emended by
Banner and Highton 1989) has the same basic architec-
ture. These descriptions are based on material in thin sec-
tions. In the present study, isolated specimens of
Pseudotaberina, allowing study of the internal architec-
ture, are described from the Miocene of Java. These turn
out to have a different architecture, and thus the genera
are not synonymous.
[Palaeontology, Vol. 51, Part 1, 2008, pp. 71–79]
ª The Palaeontological Association doi: 10.1111/j.1475-4983.2007.00731.x 71

AGE AND ORIGIN OF THE MATERIAL
In the collection of the Nationaal Natuurhistorisch
Museum, Leiden, samples collected by Martin in
1910 ⁄ 1911 contain isolated, sediment-free specimens of
Pseudotaberina malabarica. They were labelled and stored
under the name Orbiculina cf. adunca Fichtel and Moll,
1798, which is also the name under which they were pub-
lished by Rutten (1917).
The samples were collected at Kembang Sokkoh and
Gunung Spolong (Yogyakarta, Indonesia; Text-fig. 1), two
localities separated by less than 4 km. The lower part of
the Jonggrangang Formation comprises a thick succession
of Oligocene–earliest Miocene andesitic volcaniclastics,
overlying the Middle–Late Eocene Nanggulan marls. The
upper part of this formation consists of an alternation of
marls and limestones, and grades into a bedded lime-
stone. A detailed description of the sample sites and their
relative position was published by Martin (1911). In total,
at least 103 species of molluscs (Martin 1917), 15–17 spe-
cies of corals (Gerth 1921), and nine species of benthic
foraminifera (Rutten in Martin 1917; Tan Sin Hok in van
Bemmelen 1949) have been described from the Jonggran-
gang Formation. The setting was interpreted as reefal,
with Kembang Sokkoh situated near a (small) river-
mouth (van Bemmelen 1949). Accompanying LBF were,
among others, Lepidosemicyclina thecidaeformis, Austrotril-
lina howchini, Flosculinella globulosa and Planogypsina.
This is a typical LBF zone Tf1 fauna, approximately
equivalent to early Burdigalian.
Though rare, Pseudotaberina malabarica has also been
found in some other localities. Van der Vlerk (1924) fig-
ured a specimen from the Njalindung beds at Ci Talahab
(West Java, Indonesia; Text-fig. 1). The accompanying
fauna at this locality indicates a slightly younger age: Tf2
or late Burdigalian. An assemblage very similar to that on
Java was described by de Neve (1947) from the Sangkuli-
ran area in East Kalimantan (Text-fig. 1). The age of this
locality is Tf1. Just as in the Jonggrangan Formation, the
specimens of Flosculinella globulosa were very small and
spherical. De Neve (1947) described his specimens as a
new species, Archaias vandervlerki.
In East Kalimantan, P. malabarica was also found at a
locality near Bontang (Renema, unpublished data) where
it is present in dark marls containing numerous Flosculi-
nella bontangensis (Rutten) and Lepidosemicyclina bifida
(Rutten), as well as a limited number of Nephrolepidina
angulosa (Rutten). Flosculinella bontangensis is a fusiform
alveolinid species (length approximately twice height).
The specimens found at Bontang (the type locality) show
very limited variation in elongation. In this friable marl,
very rare specimens of Pseudotaberina malabarica were
found in one sample. The age of these samples is Tf2, lat-
est Burdigalian.
Wonders and Adams (1991) reported another occur-
rence of Pseudotaberina from the Darai Limestone (Papua
New Guinea: PNG). They determined the stratigraphical
position of their samples as basal Tf1, earliest Burdigalian.
Other localities outside Indonesia include Saipan (Mari-
ana Islands) in the West Pacific (Cole, 1957) where
Pseudotaberina was found in cores in association with
Austrotrillina howchini, Flosculinella globulosa and
Nephrolepidina spp., suggesting an age very similar to that
of the Javanese specimens. The illustrations are not of
sufficient quality to allow identification to species level
but they suggest Pseudotaberina vandervlerki.
In Flosculinella, a general trend from spherical to fusi-
form is observed, with intermediate variable populations
120° E
0 200 400
km
10° S
10° N
0°
140° E
1
2
34
N
TEXT-F IG . 1 . Map showing the
localities mentioned in the text. 1,
Menoreh Mountains, Yogyakarta; 2,
Bontang, East Kalimantan; 3, Ci Talahab,
West Java; 4, Darai Limestone, Papua
New Guinea.
72 P A L A E O N T O L O G Y , V O L U M E 5 1

(Reichel 1936–37; Wonders and Adams 1991). The associ-
ation with both basal spherical and derived fusiform pop-
ulations shows that Pseudotaberina occurred over a longer
period. The faunal associations fit well within Tf1–Tf2 of
Burdigalian age. Thus Pseudotaberina has a very wide geo-
graphical distribution but is restricted to a short strati-
graphic range through most of the Burdigalian.
TERMINOLOGY
A complete description of the architectural elements in
archaiasin tests has been provided by Hottinger (2001,
2005). Here the elements used in the description of Pseu-
dotaberina are repeated. A schematic overview of the
structural elements is given in Text-figure 2. Archaiasins
are porcellaneous, imperforate foraminifera. In this group,
subdivisions of the chambers by structural elements are
produced by localized thickening of the chamber wall.
Apertures, openings in the chamber wall through which
the foraminiferan communicates with the outside envi-
ronment, develop on the apertural face, the area of the
chamber wall where single or multiple apertures are posi-
tioned. During the formation of the next chamber, the
apertural face is covered and the apertures transform into
intercameral foramina through which there is communi-
cation between successive chambers. The axes of aligned
apertures in successive chambers are called apertural axes.
These can be parallel to the radius of the test: in this case
the apertural axis does not cross the previous or the next
apertural axes. This pattern is called radial. In a cross-
wise-oblique pattern the apertural axes are oblique to the
a
a
pi
su
ap
f
f
ap su
s
ma
lw
A
B
af
so
TEXT -F IG . 2 . The structure of archaiasinid frontal and lateral
chamber margins. Over part of the chamber, the frontal and
lateral walls are cut off to show the septal face and partitions
below the lateral chamber wall. Stereographs, schematic, not to
scale. A, Pseudotaberina sp.; B, Miarchaias sp. (after Hottinger,
2001). Abbreviations: a, aperture; af, apertural face; ap, annular
passage; f, foramen; lw, lateral chamberwall; ma, marginal
aperture; pi, pillar; s, septum; so, socculus; su, septula.
so
bts
su
su
A
B
f
f
s
s
s s
lw
TEXT -F IG . 3 . A, scanning electron microscope (SEM)
micrograph of the ultimate chambers of Pseudotaberna
vandervlerki, Kembang Sokkoh, Menoreh Mountains,
Yogyakarta, RGM 508454; white hatched lines represent the
direction of the passages through the foramina; solid white lines
represent the test radius. B, schematic interpretation of the
structural elements in Text-figure 2. Abbreviations: as for Text-
figure 2 and bts, basal thickening of the septula. Scale bar
represents 0.1 mm.
R E N E M A : I N T E R N A L A R C H I T E C T U R E O F M I O C E N E P S E U D O T A B E R I N A 73

radius of the test, and cross each other regularly (Text-
fig. 3). In archaiasins the chambers can be subdivided by
septula (continuous walls) or free-standing pillars. Septula
and pillars tend to thicken at their proximal contact with
the basal layer. These structures have been called socculi
(Seiglie et al. 1976; Hottinger 2001). The annular passage
through the septula connects chamberlets of the same
chamber.
During growth, planispiral, involute archaiasinids tend
to extend their apertural face by peripheral extension of
their spiral chambers during ontogeny: a first spiral,
peneropline growth stage ends with the first chamber over-
growing the spiral periphery of the previous chamber. This
growth stage ends where a chamber produces a complete
annulus for the first time (Hottinger 2005). To increase
further the extension of the apertural face, two structures
can be formed: (1) forward growing: meandrine chambers
growing from the apertures of the last involute chambers
form lateral supplementary chambers overgrowing the
annular growth stage (Text-fig 4A); or (2) backward
growing: the marginally placed apertures produce lateral
supplementary chambers overgrowing the central part of
the test at the point where the distal part of the apertural
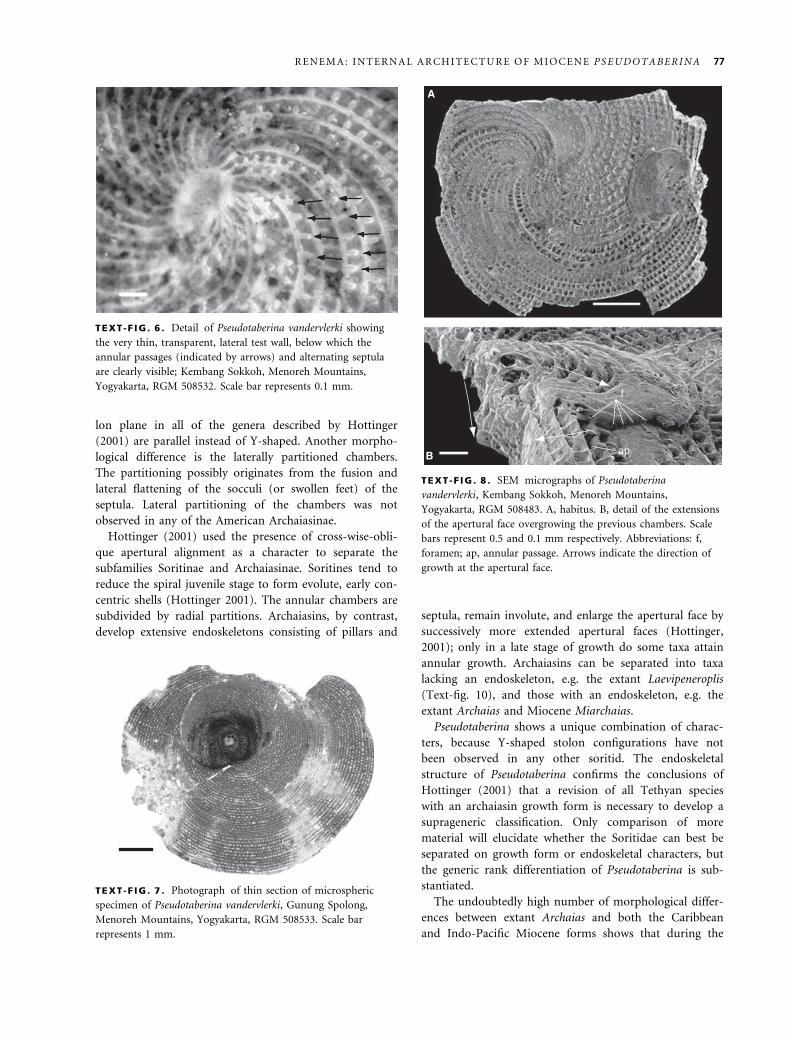
face touches the more proximal part (Text-figs 4B, 8).
SYSTEMATIC PALAEONTOLOGY
Superfamily SORITACEA Ehrenberg, 1839
Family SORITIDAE Ehrenberg, 1839
Genus PSEUDOTABERINA Eames, in Davies 1971, emend.
Banner and Highton, 1989
Type species. Orbitolites malabarica Carter, 1853a.
Pseudotaberina malabarica (Carter, 1853a)
* 1853a Orbitolites malabarica Carter, pp. 142–144, pl. 2A.
1853b Orbitolites malabarica Carter; Carter, pp. 425–427,
pl. 16B, figs 1–4.
1989 Pseudotaberina malabarica (Carter); Banner and
Highton, pp. 113–129, pl. 1, figs 6–8.
? 1923 Orbiculina malabarica (Carter); Wayland and
Davies, pp. 591–592, pl. 28, figs 1–4.
Description. Banner and Highton (1989) formally erected the
genus Pseudotaberina, which up to then had been a nomen nu-
dum, and emended the type species P. malabarica based on the
specimens used by Carter (1853a). The size of the topotype
specimens (Banner and Highton 1989, pl. 1, figs 6–8) varies
from 2.9 to 6 mm. The figured apertural face of the 6 mm speci-
men shows multiple, 5–7, rows of apertures. The alternating,
rimmed apertures are positioned in depressions. At a diameter
of 2.9 mm the alar prolongations are 0.6 mm. The chamber
height averages 0.04 mm.
In the macrospheric generation, specimens reach a diameter
of up to 6.5 mm and a thickness of 0.7 mm. Microspheric speci-
mens can reach a diameter of up to 15 mm, and attain annular
chambers at a diameter of c. 8 mm. Larger specimens develop
forward-growing meandrine chambers from the apertures of the
ultimate involute chamber.
The internal architecture, as shown in the oblique sections, is
very similar to that of the Indonesian specimens. Both have alter-
nating septula that reach approximately half-way to the chamber
just below the lateral wall, and increase in length and width, cut-
ting deeper into the test. Laterally flattened socculi subdividing
the chamberlets are also present in the Indian specimens.
Remarks. Wayland and Davies (1923) recorded a larger
maximum diameter (up to 21 mm) than that for the
specimens described by Banner and Highton (1989). The
specimens attain cyclical growth when they reach a diam-
eter of 3–5 mm, which is slightly smaller than in the
specimens figured in Banner and Highton (1989). Way-
land and Davies (1923) were the first to discuss the differ-
ence between the Indian and Indonesian specimens. They
also observed that at the same size, the Indonesian speci-
mens do not attain cyclical growth. Interestingly, they
mentioned (but did not figure) a single specimen of
P. vandervlerki from India. By contrast, Tobler (1927)
mentioned a Meandropsina from Kalimantan, a genus that
externally resembles Pseudotaberina in attaining cyclical
growth, but the internal structure is different. So far, it is
known only from the Cretaceous (L. Hottinger, pers.
comm. 2005). The dimensions of the Kalimantan speci-
mens equal those of P. malabarica from India, although
the largest specimens are larger than those mentioned by
Wayland and Davies (1923). In the samples from Kali-
mantan, the small, probable macrospheric, specimens
measure 5–7 mm, the largest, probable microspheric,
specimens 25 mm (Tobler 1927).
A B
TEXT -F IG . 4 . Schematic drawing of the outside of the test. A,
forward-growing Miarchaias, after reaching annular growth;
lateral extensions of the apertural face of the alar prolongation
of the ultimate chamber before reaching annular growth. B,
backward-growing Pseudotaberina vandervlerki, extensions of the
apertural face grow over the previous chamber starting at the
point where the distal part of the apertural face touches the
more distal part at the onset of evolute growth (see also Text-
fig. 8); thin lines indicate sutures, thick black line indicates
apertural face where growth occurs. Not to scale.
74 P A L A E O N T O L O G Y , V O L U M E 5 1

Pseudotaberina vandervlerki (de Neve, 1947)
Text-figures 2A, 3–9.
1917 Orbiculina cf. adunca Fichtel and Moll, 1798;
Rutten, pp. 276–285, pl. 5, fig. 142.
1924 Orbiculina? sp.; van der Vlerk 1924, pp. 27–28,
fig. 25.
* 1947 Archaias vandervlerki de Neve 1947, pp. 14–15,
text-figs 1–4.
? 1957 Archaias vandervlerki De Neve; Cole, p. 335,
pl. 103, figs 5–9.
Material. More than 100 isolated specimens, of which only two
are microspheric, from Kembang Sokkoh and Gunung Spolong,
Yogyakarta, Indonesia (RGM 3267, 3481, 47264, 47265, 508454–
508456, 508479, 508481, 508483–508484). Type specimens
(unnumbered) of Archaias vandervlerki de Neve, 1947, deposited
in the Geological Museum in Bandung, Indonesia. Unnumbered
single specimen labelled Orbiculina sp. from the Njalindung
beds, figured by van der Vlerk (1924), also deposited in the Geo-
logical Museum in Bandung.
Description. Lenticular test, planispiral involute throughout
ontogeny. In the macrospheric generation, specimens can reach
a diameter of 5 mm and a thickness of 0.7 mm. Microspheric
specimens can reach a diameter of 12 mm. Annular stages have
not been observed. The chambers proximally embrace the previ-
ous whorls but distally they are evolute. They are low but long,
with a strong posterior curvature so that they cover at least half,
but usually more, of the previously formed test, and cover more
as growth continues. Chamber height increases gradually from
58–71 lm in the first whorl to 65–86 lm in the last whorl. The
last chambers in the microspheric generations become 90–
120 lm in height. The alar prolongations extend 4 mm in a
specimen of 12 mm (microsphere), and 1.2–1.4 mm in macro-
spheric specimens of 3.1–3.3 mm diameter. The embracing com-
ponent (length of alar prolongation ⁄ radius) in specimens with a
diameter of 3–4 mm is 0.76–0.84. Apertures are round and
rimmed. They are placed in four alternating superposed rows;
the shape of those in the marginal rows is the same as those in
the equatorial layers (Text-fig. 5A). The lateral chamber wall is
thin and slightly pitted.
The length of the chambers increases with growth. In the later
chambers test growth occurs along the entire circumference, but
without becoming annular as in Miarchaias. At the place where
the distal part of the apertural face meets the more proximal
part, the marginal apertures of the proximal part form back-
ward-growing meandrine lateral chambers (Text-figs 4, 8, 9E).
The occurrence of this structure at exactly the transition from
involute to evolute growth, and never at other positions along
the apertural face, indicates that this backward-growing is not
the result of shell repair. No forward-growing lateral extensions
of the alar prolongations have been observed.
The megalospheric embryo consists of an approximately
spherical megalosphere 0.25–0.30 mm in diameter followed by a
wide flexostyle of c. 0.1 mm and about one-quarter volution in
length. The flexostyle has two apertures at the proximal part: the
first spiral chamber does not show a subdivision in all thin sec-
tions, but the second and later chambers always have subdivi-
sions. There are 3–3.5 volutions in macrospheric specimens. The
lateral test wall is very thin, and the septula are visible from the
outside.
The chambers consist of four stolon planes and are radially
divided into chamberlets by septula. An annular passage, of
which the diameter is about one-half to one-third of the cham-
ber height, is positioned distally in the septula directly below the
test wall (Text-fig. 2A). The septula are somewhat irregularly
placed and are thicker proximally than distally. The laterally flat-
tened bases, which can possibly be referred to as socculi, physi-
cally separate the stolon planes. The chamberlets are laterally
divided into four stolon planes by the flattened bases of the sep-
tula (soculi; Text-fig. 2A).
Between the septula are round, rimmed foramina in the sep-
tum, with a diameter of c. 0.025–0.031 mm. The foramina are
placed opposite the septula in the previous chamber, resulting in
Y-shaped stolon passages (Text-fig. 3). Owing to this pattern,
every chamberlet is connected to two chamberlets in the more
distal chamber. This results in the absence of stolon axes, pro-
viding a direct line of transport from the central part of the test
to the apertural face.
Remarks. The specimens described here differ from those
described by Banner and Highton (1989) in that the
embracing component of the alar prolongations is much
larger. Other differences relate to the apertural face, which
does not show the rimmed apertures positioned in depres-
sions, and the smaller diameter of full-grown tests. Also,
annular chambers and forward-growing lateral extensions
of the alar prolongation as described for P. malabarica
(Banner and Highton 1989) have not been observed in
Indonesian specimens.
Stratigraphic distribution. Burdigalian of Java, Borneo, PNG and
India.
DISCUSSION AND CONCLUSIONS
Henson (1950) found Taberina [Archaias, Orbiculina, Or-
bitolites] malabarica Carter, 1853a in Iraq. However, no
isolated specimens are available and his material could
not be assigned with certainty to either of the two species
described here.
The type material of Archaias vandervlerki de Neve, 1947
is deposited at the Geological Museum in Bandung and
was available for comparison. The specimens concerned
are identical to those from Java; thus the name P. vander-
vlerki (de Neve, 1947) is available for this species.
Based on external morphology, Pseudotaberina resem-
bles Miarchaias, described by Hottinger (2001) from the
Miocene of the Dominican Republic. However, the stolon
axes in Miarchaias are radial instead of Y-shaped as in
Pseudotaberina (Text-fig. 3). The stolon axes in each sto-
R E N E M A : I N T E R N A L A R C H I T E C T U R E O F M I O C E N E P S E U D O T A B E R I N A 75

lw
af
a
a f
f
s
su
su
bts
bts
so
C
B
1
2 3 4
A
D
TEXT-F IG . 5 . SEM micrographs of details of the test of
Pseudotaberina vandervlerki, RGM 508484. A, oblique view
of the apertural face showing four rows of irregularly
superposed apertures. B, oblique view of chamberlets in the
ultimate and penultimate whorl; white arrows indicate two
superposed foramina, the top one with a stolon passage to
the left, the lower one to the right; black arrow indicates the
annular passage. C, schematic interpretation of B;
abbreviations as for Text-figures 2 and 3. D, axial view of
broken specimens showing four stolon planes. Scale bars
represent 50 lm in A–B, 0.1 mm in D.
76 P A L A E O N T O L O G Y , V O L U M E 5 1
lon plane in all of the genera described by Hottinger
(2001) are parallel instead of Y-shaped. Another morpho-
logical difference is the laterally partitioned chambers.
The partitioning possibly originates from the fusion and
lateral flattening of the socculi (or swollen feet) of the
septula. Lateral partitioning of the chambers was not
observed in any of the American Archaiasinae.
Hottinger (2001) used the presence of cross-wise-obli-
que apertural alignment as a character to separate the
subfamilies Soritinae and Archaiasinae. Soritines tend to
reduce the spiral juvenile stage to form evolute, early con-
centric shells (Hottinger 2001). The annular chambers are
subdivided by radial partitions. Archaiasins, by contrast,
develop extensive endoskeletons consisting of pillars and
septula, remain involute, and enlarge the apertural face by
successively more extended apertural faces (Hottinger,
2001); only in a late stage of growth do some taxa attain
annular growth. Archaiasins can be separated into taxa
lacking an endoskeleton, e.g. the extant Laevipeneroplis
(Text-fig. 10), and those with an endoskeleton, e.g. the
extant Archaias and Miocene Miarchaias.
Pseudotaberina shows a unique combination of charac-
ters, because Y-shaped stolon configurations have not
been observed in any other soritid. The endoskeletal
structure of Pseudotaberina confirms the conclusions of
Hottinger (2001) that a revision of all Tethyan species
with an archaiasin growth form is necessary to develop a
suprageneric classification. Only comparison of more
material will elucidate whether the Soritidae can best be
separated on growth form or endoskeletal characters, but
the generic rank differentiation of Pseudotaberina is sub-
stantiated.
The undoubtedly high number of morphological differ-
ences between extant Archaias and both the Caribbean
and Indo-Pacific Miocene forms shows that during the
TEXT -F IG . 7 . Photograph of thin section of microspheric
specimen of Pseudotaberina vandervlerki, Gunung Spolong,
Menoreh Mountains, Yogyakarta, RGM 508533. Scale bar
represents 1 mm.
f
apB
A
TEXT -F IG . 8 . SEM micrographs of Pseudotaberina
vandervlerki, Kembang Sokkoh, Menoreh Mountains,
Yogyakarta, RGM 508483. A, habitus. B, detail of the extensions
of the apertural face overgrowing the previous chambers. Scale
bars represent 0.5 and 0.1 mm respectively. Abbreviations: f,
foramen; ap, annular passage. Arrows indicate the direction of
growth at the apertural face.
TEXT -F IG . 6 . Detail of Pseudotaberina vandervlerki showing
the very thin, transparent, lateral test wall, below which the
annular passages (indicated by arrows) and alternating septula
are clearly visible; Kembang Sokkoh, Menoreh Mountains,
Yogyakarta, RGM 508532. Scale bar represents 0.1 mm.
R E N E M A : I N T E R N A L A R C H I T E C T U R E O F M I O C E N E P S E U D O T A B E R I N A 77
Miocene this group was highly diverse. At present, no
extant Archaiasinae with an endoskeleton are known from
the Indo-West Pacific. Pseudotaberina and the Middle
Eastern taxa described by Henson (1950) became extinct
at the end of the Early Miocene. Two separate radiations
can be hypothesized, one during the Oligocene–Miocene
in the Tethyan realm and another in the Middle–Late
Miocene in the Caribbean. The stratigraphical use of the
occurrence of Pseudotaberina has long been recognised in
the Indo-West Pacific. The Indonesian specimens
reported here are restricted to the Burdigalian. The
assemblages in which Pseudotaberina was found are typi-
cal of very shallow, epiphytic depositional environments.
Acknowledgements. Frank Wesselingh and Steve Donovan read
earlier drafts of this manuscript and their constructive remarks
improved the paper. Comments by Lukas Hottinger and an
anonymous reviewer further improved its quality.
REFERENCES
B A N N E R , F. T. and H I G H T ON , J. 1989. On Pseudotaberina
malabarica (Carter) (Foraminiferida). Journal of Micropalaeon-
tology, 8, 113–129.
B E M M E L E N , R. W. van 1949. Geology of Indonesia. Govern-
ment Printing Office, The Hague, 732 pp.
C A R T E R , H. J. 1853a. A description of Orbitolites malabarica,
illustrative of the spiral and not concentric arrangement of
A B
C D E
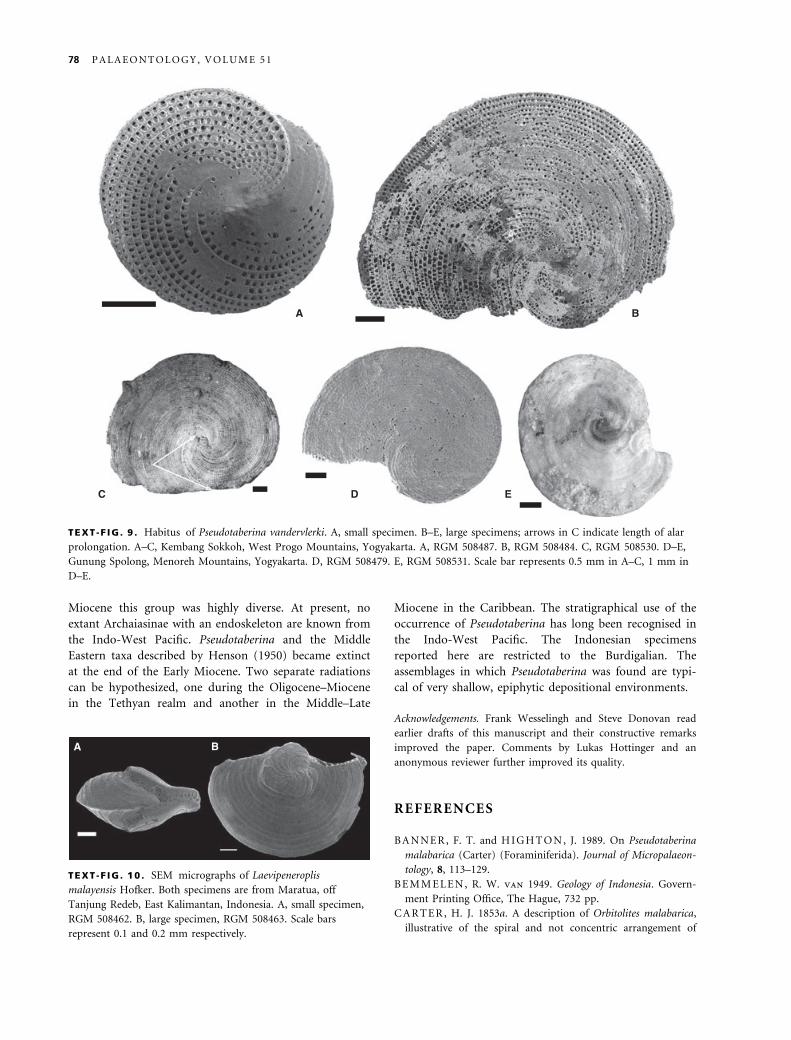
TEXT -F IG . 9 . Habitus of Pseudotaberina vandervlerki. A, small specimen. B–E, large specimens; arrows in C indicate length of alar
prolongation. A–C, Kembang Sokkoh, West Progo Mountains, Yogyakarta. A, RGM 508487. B, RGM 508484. C, RGM 508530. D–E,
Gunung Spolong, Menoreh Mountains, Yogyakarta. D, RGM 508479. E, RGM 508531. Scale bar represents 0.5 mm in A–C, 1 mm in
D–E.
B A
TEXT -F IG . 10 . SEM micrographs of Laevipeneroplis
malayensis Hofker. Both specimens are from Maratua, off
Tanjung Redeb, East Kalimantan, Indonesia. A, small specimen,
RGM 508462. B, large specimen, RGM 508463. Scale bars
represent 0.1 and 0.2 mm respectively.
78 P A L A E O N T O L O G Y , V O L U M E 5 1

chambers in d’Orbigny’s order Cyclostegues. Journal of the
Bombay Branch, Royal Asiatic Society, 5, 142–144.
—— 1853b. A description of Orbitolites malabarica (H.J.C.),
illustrative of the spiral and not concentric arrangement of
chambers in d’Orbigny’s order Cyclostegues. Annals and Mag-
azine of Natural History (2), 11, 425–427.
C OL E , W. S. 1957. Larger foraminifera. In: Geology of Saipan,
Mariana Islands. United States Geological Survey, Professional
Paper, 280-I, 319–360.
D A V I E S , A. M. 1971. Tertiary faunas. A text-book for oilfield
palaeontologists and students of geology. Volume 1. Second edi-
tion. George Allen and Unwin, London, 571 pp.
E H R E N B E R G , C. G. 1839. Uber die Bildung der Kreidefelfsen
und des Kreidemergels durch ubsichtbare Organismen. Physi-
kalische Abhandlungen der Konglichen Akademie der Wissen-
schaften, Jahrgang, 1838, 59–147.
G E R T H , H. 1921. Coelenterata. Sammlungen Geologisches
Reichsmuseums, Neue Folge, 1 (2), 387–445.
G U DM UN D S S O N , G. 1994. Phylogeny, ontogeny and sys-
tematics of Recent Soritacea Ehrenberg 1839 (Foraminiferida).
Micropaleontology, 40, 101–155.
F I C H T E L , L. and M O L L , J. P. C. 1798. Testacea microscopia
aliaque minuta ex generibus Argonauta et Nautilus ad naturam
delineata et descripta. Anton Pichler, Wien, 123 pp.
H A L L O CK , P. 1988. Diversification in algal symbiont-bearing
foraminifera; a response to oligotrophy? Revue de Paleo-
biologie, Special Issue, 2, 789–797.
—— and PE E B L E S , M. W. 1993. Foraminifera with chloro-
phyte endosymbionts: habitats of six species in the Florida
Keys. Marine Micropaleontology, 20, 277–292.
H E N S O N , F. R. S. 1950. Middle Eastern Tertiary Peneroplidae
(foraminifera) with remarks on the phylogeny and taxonomy of
the family. West Yorkshire Printing Company, Wakefield, 70
pp.
H O L ZM A N , M., HO H E N E G G E R , J., H A L L O CK , P.,
PI L L E R , W. E. and PA W L O W S K I , J. 2001. Molecular
phylogeny of large miliolid foraminifera (Soritacea Ehrenberg
1839). Marine Micropaleontology, 43, 57–74.
H O TT I N G E R , L. 2001. Archaisinids and related porcelaneous
larger foraminifera from the late Miocene of the Dominican
Republic. Journal of Paleontology, 75, 475–512.
—— 2005. Geometrical constraints in foraminiferal architecture:
consequences of change from planispiral to annular growth.
Studia Geologica Polonica, 124, 99–115.
M A R TI N , K. 1911. Vorlaufiger Bericht uber geologische
Forschungen auf Java. Erster Teil. Sammlungen Geologisches
Reichsmuseums Leiden, 9, 1–76.
—— 1917. Die altmiocane Fauna des West-Progo Gebirges auf
Java. Sammlungen Geologisches Reichsmuseums Leiden, Neue
Folge, 2, 1–276.
N E V E , G. A. de 1947. A new Archaias species from east Bor-
neo. Dutch East Indies. Bulletin de Bureau des Mines and the
Geological Survey of Indonesia, 1, 13–16.
R I CH A R S O N , S. 2001. Endosymbiont change as a key inno-
vation in the adaptive radiation of Soritida (Foraminifera).
Paleobiology, 27, 262–280.
R E I CH E L , M. 1936–37. Etude sur les Alveolines. Schweizerische
Palaeontologische Abhandlungen, 57 (4), 1–93; 59 (3), 95–147.
R U T TE N , L. M. R. 1917. Rhizopoda. Sammlungen Geologisches
Reichsmuseums Leiden, Neue Folge, 2, 276–277.
S E I G L I E , A. K., G R O V E , A. K. and R I V E R A , J. A. 1976.
Revision of some Caribbean Archaiasinae, new genera,
species and subspecies. Eclogae Geologicae Helvetiae, 70, 855–
883.
T O BL E R , A. 1927. Meandropsina im Tertiar von Ostborneo.
Eclogae Geologicae Helvetiae, 20, 321–323.
V L E R K , I. M. van der 1924. Foraminiferen uit het Tertiair
van Java I. Wetenschappelijke Mededeelingen van de Dienst
Mijnbouw, 1, 1–35.
W A Y L A N D, E. J. and D A V I E S , A. M. 1923. The Miocene of
Ceylon. Quarterly Journal of the Geological Society of London,
79, 577–602.
W O N D E R S , A. A. H. and A D A M S , C. G. 1991. The
biostratigraphical and evolutionary significance of Alveolinella
praequoyii sp. nov. from Papua New Guinea. Bulletin of the
British Museum (Natural History), Geology, 47, 169–175.
R E N E M A : I N T E R N A L A R C H I T E C T U R E O F M I O C E N E P S E U D O T A B E R I N A 79




















