
Patterns of diversity in microscopic animals: are they comparable to those in protists or in larger...
10
© 2006 Blackwell Publishing Ltd www.blackwellpublishing.com/geb DOI: 10.1111/j.1466-822x.2005.00193.x 153 Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2006) 15, 153–162 RESEARCH PAPER Blackwell Publishing, Ltd. Patterns of diversity in microscopic animals: are they comparable to those in protists or in larger animals? Diego Fontaneto*, Gentile Francesco Ficetola, Roberto Ambrosini and Claudia Ricci ABSTRACT Aim General patterns of biodiversity, such as latitudinal gradients and species-area relationships, are found consistently in a wide range of organisms, but recent results for protist diversity suggest that organisms shorter than 2 mm do not display such patterns. We tested this prediction in bdelloid rotifers, pluricellular metazoans smaller than 2 mm, but with size and ecology comparable to protists. Location A single valley in northern Italy was surveyed in detail and compared to all available faunistic data on bdelloids worldwide. Methods We analysed 171 local assemblages of bdelloid rotifers living in 5 systems of dry mosses and submerged mosses in running water and in lakes. We compared patterns of alpha, beta, and gamma diversity, and nestedness of metacommunities, with those known from protists and larger organisms. Results Bdelloid rotifers showed low local species richness (alpha diversity), with strong habitat selection, as observed in larger organisms. The number of species dif- fered among systems, with a higher number of species in dry than in aquatic mosses. There was no hierarchical structure or exclusion of species in the metacommunity pattern within each system. Local diversity for the entire valley was surprisingly high compared with worldwide bdelloid diversity, similar to observed patterns in protists. Main Conclusions Bdelloid rotifers have some of the peculiarities of protist biodiversity, although at slightly different spatial scales, thus confirming the idea of a major change in biodiversity patterns among organisms shorter than 2 mm. However, bdelloids show stronger habitat selection than protists. We suggest two possible explanations for the observed patterns: (1) dispersal is very rare, and not all bdelloid clones are arriving everywhere; and (2) dispersal is effective in displacing propagules, but environmental heterogeneity is very high and prevents many species from colonizing a given patch of moss. Keywords Bdelloidea, biodiversity, community, metacommunity, rotifera, species richness. *Correspondence: Diego Fontaneto, Dipartimento di Biologia, Università di Milano, via Celoria 26, I-20133 Milano, Italy. E-mail: [email protected] Dipartimento di Biologia, Università di Milano, via Celoria 26, I-20133 Milano, Italy. INTRODUCTION General patterns of biodiversity are found consistently across a wide range of organisms. Many such patterns have been described, for example latitudinal and altitudinal gradients, species-area relationships, and species-energy relationships (e.g. Rosenzweig, 1995; Lawton, 1999; Andrew et al ., 2003; Brehm et al ., 2003; Bonn et al ., 2004; Davies et al ., 2004; Dimitrakopoulos & Schmid, 2004; Gaston et al ., 2005; McAbendroth et al ., 2005; Rahbek, 2005). In addition, many organisms experience historical constraints on their distributions at different scales. For example, onycophorans and marsupials are distributed almost exclusively in the Southern Hemisphere (Monge-Najera & Hou, 2000; Sanmartin & Ronquist, 2004), and the local distribution of many species of plants and animals in the Northern Hemisphere can be explained by the location of glacial refugia (Hausdorf & Henning, 2004; Tribsch, 2004). Although debates continue on the processes that cause these patterns, and on the exact nature of patterns in different groups, the existence of gradients and structure in species distributions is clear.
Transcript of Patterns of diversity in microscopic animals: are they comparable to those in protists or in larger...

© 2006 Blackwell Publishing Ltd www.blackwellpublishing.com/geb DOI: 10.1111/j.1466-822x.2005.00193.x
153
Global Ecology and Biogeography, (Global Ecol. Biogeogr.)
(2006)
15
, 153–162
RESEARCHPAPER
Blackwell Publishing, Ltd.
Patterns of diversity in microscopic animals: are they comparable to those in protists or in larger animals?
Diego Fontaneto*, Gentile Francesco Ficetola, Roberto Ambrosini
and Claudia Ricci
ABSTRACT
Aim
General patterns of biodiversity, such as latitudinal gradients and species-arearelationships, are found consistently in a wide range of organisms, but recent resultsfor protist diversity suggest that organisms shorter than 2 mm do not display suchpatterns. We tested this prediction in bdelloid rotifers, pluricellular metazoanssmaller than 2 mm, but with size and ecology comparable to protists.
Location
A single valley in northern Italy was surveyed in detail and compared toall available faunistic data on bdelloids worldwide.
Methods
We analysed 171 local assemblages of bdelloid rotifers living in 5 systemsof dry mosses and submerged mosses in running water and in lakes. We comparedpatterns of alpha, beta, and gamma diversity, and nestedness of metacommunities,with those known from protists and larger organisms.
Results
Bdelloid rotifers showed low local species richness (alpha diversity), withstrong habitat selection, as observed in larger organisms. The number of species dif-fered among systems, with a higher number of species in dry than in aquatic mosses.There was no hierarchical structure or exclusion of species in the metacommunitypattern within each system. Local diversity for the entire valley was surprisingly highcompared with worldwide bdelloid diversity, similar to observed patterns in protists.
Main Conclusions
Bdelloid rotifers have some of the peculiarities of protistbiodiversity, although at slightly different spatial scales, thus confirming the idea ofa major change in biodiversity patterns among organisms shorter than 2 mm.However, bdelloids show stronger habitat selection than protists. We suggest twopossible explanations for the observed patterns: (1) dispersal is very rare, and not allbdelloid clones are arriving everywhere; and (2) dispersal is effective in displacingpropagules, but environmental heterogeneity is very high and prevents many speciesfrom colonizing a given patch of moss.
Keywords
Bdelloidea, biodiversity, community, metacommunity, rotifera, species richness.
*Correspondence: Diego Fontaneto, Dipartimento di Biologia, Università di Milano, via Celoria 26, I-20133 Milano, Italy. E-mail: [email protected]
Dipartimento di Biologia, Università di Milano,
via Celoria 26, I-20133 Milano, Italy.
INTRODUCTION
General patterns of biodiversity are found consistently across a
wide range of organisms. Many such patterns have been described,
for example latitudinal and altitudinal gradients, species-area
relationships, and species-energy relationships (e.g. Rosenzweig,
1995; Lawton, 1999; Andrew
et al
., 2003; Brehm
et al
., 2003;
Bonn
et al
., 2004; Davies
et al
., 2004; Dimitrakopoulos & Schmid,
2004; Gaston
et al
., 2005; McAbendroth
et al
., 2005; Rahbek,
2005). In addition, many organisms experience historical
constraints on their distributions at different scales. For example,
onycophorans and marsupials are distributed almost exclusively
in the Southern Hemisphere (Monge-Najera & Hou, 2000;
Sanmartin & Ronquist, 2004), and the local distribution of many
species of plants and animals in the Northern Hemisphere can
be explained by the location of glacial refugia (Hausdorf &
Henning, 2004; Tribsch, 2004). Although debates continue on
the processes that cause these patterns, and on the exact nature
of patterns in different groups, the existence of gradients and
structure in species distributions is clear.

D. Fontaneto
et al.
154
Global Ecology and Biogeography
,
15
, 153–162 © 2006 Blackwell Publishing Ltd
One limitation of these studies for establishing general diver-
sity patterns is that they all considered organisms longer than
2 mm. Recent results for microscopic organisms indicate a major
change in biodiversity patterns among organisms less than 2 mm
in body length (Finlay, 2002). For example, Fenchel
et al
. (1997)
counted the number of ciliates in the sediment beneath a
few square centimetres in a pond and found around 50% of all
species recorded during comprehensive sampling of that pond,
corresponding to about 8% of all named freshwater ciliates.
Similarly, Persson (2002) found roughly 25% of all known Swedish
sea-plankton dinoflagellates and diatoms in 100 ml of sediment.
Moreover, no latitudinal or species-area gradients in species
composition has ever been found. Hence, Fenchel & Finlay
(2004) concluded that, for microscopic organisms, ‘everything is
everywhere’ and that the protist species found in a given habitat
are a function only of habitat properties and not of historical
factors or restricted dispersal.
The degree of endemism is much higher in larger animals. For
example, in amphibians, even the richest countries, such as Brazil
and Colombia, possess only 8% and 6% of the global diversity
(Global Amphibian Assessment, 2005). Similarly, Europe has
17% of the species of ground beetles known worldwide (Audisio
& Vigna Taglianti, 2004) and each sampling locality presents less
than 0.1% of the global diversity (e.g. Allegro & Sciaky, 2003; Kotze
& O’Hara, 2003). This difference between organisms larger and
smaller than 2 mm seems to be related to size and not to environ-
ment, since many copepods, living in the same habitats as protists
but larger that 2 mm, share patterns of distribution with larger
animals (Jersabek
et al
., 2001; Suarez-Morales
et al
., 2004).
The difference in diversity patterns between small and large
organisms can be considered in terms of species-area curves. At
the local scale (alpha diversity), small organisms dominate in
species richness: many more species of small organisms than of
large ones can co-occur at finer scales. However, the species-area
curve increases less steeply among small organisms than among
large organisms. In macrofaunal groups, the curves increase
monotonically because larger areas contain different faunistic or
floristic domains and more endemic species are included. In
contrast, the worldwide richness (gamma diversity) of protists is
not much larger than local richness. No true endemism is known
in protists and most species are believed to have cosmopolitan
distributions (Finlay, 2002). For example, about 50% of all named
species of heterotrophic flagellates have been recorded in a small
Danish Bay (Fenchel & Finlay, 2004). Moreover, differences in
species composition among samples (beta diversity) appear to be
due to strong habitat differences and species preferences, and not
to biogeographical constraints to dispersal. For example, irrespective
of geographical location, moisture is the most important factor in
controlling the distribution of testate amoebae species (Mitchell
et al
., 1999). Of course unreliable taxonomy in protists could
mask true distributions (Coleman, 2002), but studies of closely
related forms from different areas demonstrate that isolates from
different areas can mate with each other, confirming true cosmo-
politism (Finlay & Fenchel, 2002).
The expectation for organisms smaller than 2 mm is therefore
to have very high local richness compared to a relatively low
worldwide richness, with differences among areas due only to
habitat heterogeneity. These expectations derive from studies on
small unicellular organisms, but do similar patterns apply to
small, multicellular organisms? We propose to test this hypothesis
using as a model microscopic but multicellular bdelloid rotifers
living in mosses. Bdelloid rotifers are a group of microscopic
animals (on average 200–600
µ
m in length) belonging to the so
called ‘meiofauna’. Meiofaunal organisms are the same size as
unicellular organisms, but they are true pluricellular animals, with
recognizable organs and apparatuses. Bdelloid rotifers, and other
terrestrial meiofaunal groups such as tardigrades, gastrotrichs
and some nematodes, share with protists size and other features:
(1) they live in the same aquatic interstitial habitats, where they
can be very abundant (Linhart
et al
., 2002; Wallace & Ricci,
2002); (2) they have amictic reproduction, allowing them to
increase rapidly in population size (Ricci, 2001); and (3) they are
able to resist drought or freezing by entering dormant stages,
which may also represent propagules for dispersal (Cáceres,
1997; Örstan, 1998). Do bdelloid rotifers share diversity features
with protists, because of their similar size, biology and ecology?
Or, if their distributions are similar to those of other larger
multicellular organisms, what might be the biological causes of
the differences from protist diversity? We are unaware of existing
studies on the community composition of a meiofaunal group in
continental habitats.
We focus on 171 local assemblages of bdelloid rotifers found in
moss cushions of similar size in a valley in northern Italy occupy-
ing three different habitats: terrestrial mosses, mosses in streams,
and mosses in lakes. The habitats differ in ecological features,
such as water availability and oxygenation, and also in the amount
of connectivity among moss patches: greater potential for dispersal
exists between local assemblages in a single stream than among
terrestrial moss patches separated by bare rock, or among lakes in
different hydrographical basins.
METHODS
Field procedures
All samples were collected haphazardly along a single valley in
northern Italy, Sesia Valley.
Each sample consisted of 5 cm
2
of mosses along with any
adhering soil particles. To avoid differences in growth stage
of mosses we collected our samples from the central, more
homogeneous part of each cushion. All moss cushions were
approximately 15–25 cm in diameter. Samples were taken directly
to the laboratory and bdelloids were isolated under a dissection
microscope. All the bdelloid rotifers present in the sample were
determined and counted. Species were identified under a
compound microscope at 100–1000x following Donner (1965).
We defined as a community the local assemblage of all species
encountered in each sample.
Almost no previous knowledge is available on bdelloid species
presence in different mosses, but there is no evidence of species-
specificity between bdelloids and moss or lichen species (Burger,
1948; Francez, 1980; Ricci, 1987). Therefore, as a measure of

Diversity patterns in bdelloid rotifers
Global Ecology and Biogeography
,
15
, 153–162 © 2006 Blackwell Publishing Ltd
155
habitat we used a similar classification of moisture content in
mosses as used for protist studies (Jung, 1936; Meisterfeld, 1977),
rather than discriminating moss species. We categorized
three different kind of mosses: (1) completely dry terrestrial
mosses, called ‘dry’, (2) submerged mosses in stagnant water
from oligotrophic lakes, called ‘lentic’ and (3) submerged mosses
in running water, called ‘lotic’. These three habitats are at the
extremes of the scale of moisture: completely submerged or
completely dry.
Dry, terrestrial moss cushions
We collected 24 samples of moss along an altitudinal transect from
800 m (municipality of Campertogno, Sesia Valley, Piedmont,
northern Italy, approximate Gauss-Boaga coordinates 1424750 E,
5072400 N) to about 1800 m (Becco della Guardia Mount) in
spring 2002. All sampled mosses had a northern exposure.
Mosses were collected both on rocks (gneiss in all cases) and tree
trunks (at 20 cm from the base of different trees).
Lentic, alpine lakes
We sampled submerged mosses in 16 alpine lakes above 1700 m
in the Sesia Valley during the summers of 2001 and 2002 (see
Fontaneto & Melone, 2003 for details) and in Argnaccia Lake
(1100 m a.s.l.), in the municipality of Campertogno. Four different
mosses were haphazardly sampled in each lake, and the diversity of
each lake is the cumulative diversity of these 4 samples. All lakes
were on acidic substrates and had similar pH and temperature.
Lotic mosses in streams
We sampled submerged mosses in three different streams during
summer 2003. We sampled 15 bdelloid local assemblages, named
R1 to R15 from upstream to downstream, along Res Stream
(about 900 m a.s.l.), and 15 local assemblages, A1 to A15, along
Argnaccia Stream (about 900 m a.s.l.) (see Fontaneto
et al
.,
2004b). One hundred local assemblages, named V00 to V99,
were analysed along Valnava Stream (about 400 m a.s.l.) (see
Fontaneto
et al
., 2005 for details). Res and Argnaccia streams
are on acidic rock substrates, while Valnava is partially on
calcareous rocks.
This structure allows us to propose possible relationships with
(1) moisture of the moss, as mosses in lakes (lentic) and streams
(lotic) are aquatic, permanently submerged habitats, while dry
mosses are usually dry and became wet only during rain, and
(2) connectivity of the system, as mosses inside each stream are
connected by water flow while terrestrial mosses and alpine lakes
have no direct connection between patches.
Community analyses
We treat the local assemblage of bdelloid species in each moss
cushion as a potentially isolated community: although bdelloids
can creep on a substratum and swim in water, they cannot move
actively for long distances (Örstan, 1998).
Species richness
Local richness (alpha diversity) was treated as the number of
species in each moss patch. To detect differences in local species
richness among the 3 habitats, and then among the 3 streams, we
used analysis of variance (
), without transforming our
original count data because residuals were normally distributed.
Tukey Post-Hoc Honestly Significant Difference (Crawley, 2002)
was then used to recognize significantly different habitats.
Overall richness (gamma diversity) was obtained by counting
all species found across all habitats, dry, lentic or lotic. Richness in
each habitat was compared through rarefaction curves (Colwell
& Coddington, 1994), obtained by plotting the average number of
species in the cumulative number of analysed local assemblages,
through resampling techniques, using
EstimateS
7 (Colwell,
2004). We also estimated the diversity in each system and overall
according to ICE, the Incidence-based Coverage Estimator,
which estimates the overall number of species given the observed
number of species and how many species are found only once or
twice (Chazdon
et al
., 1998).
Species replacement in space (beta diversity)
First, we evaluated differences in species composition among the
three habitats (dry, lentic and lotic). We used reciprocal averaging
to ordinate the data matrix, using the first axis only, as proposed
by Leibold and Mikkelson (2002), so that sites with the most
similar species list are close together and species with the most
similar distribution are close together. Then we tested the signi-
ficance of the differences in species composition of local assem-
blages in the three different habitats.
We used Jaccard’s distances as a dissimilarity index for all the
pairwise comparisons and tested for significant differences using
analysis of similarities (
), based on 1000 permutations,
using the software R 2.0.1, package vegan (R Development Core
Team, 2004). We repeated the analysis to test for differences in
species composition among the local assemblages of the three
different streams.
To analyse species replacements within each habitat in detail, we
performed the analysis of metacommunity structure proposed
by Leibold and Mikkelson (2002). We defined as a metacommunity
all the local assemblages in each system, so we obtained 5 different
metacommunities, one from dry mosses, one from lentic mosses
and one from each lotic system. As these metacommunities came
from different habitats with different ecological features and
different connectivity of the patches, we tested whether there were
different rates of species replacement among local assemblages.
The probability of obtaining the observed spatial turnover was
calculated using a Monte Carlo simulation. For each metacommu-
nity, we generated 200 random matrices following a ‘random 0
Null Model’, subject only to the constraint that the matrices have
the same number of presences of the analysed metacommunity,
and that no row or column have only absences (Leibold &
Mikkelson, 2002). We tested the significance of differences
between the observed and the simulated matrices using a two-
tailed
z
-test. Low rates of spatial replacements (low turnover)
D. Fontaneto
et al.
156
Global Ecology and Biogeography
,
15
, 153–162 © 2006 Blackwell Publishing Ltd
represent nested structures, with hierarchical structures among
communities. Higher rates than expected provide evidence of
groups of species that exclude each other.
A high rate of species replacement, that is high spatial turnover of
species, is the opposite of nestedness; therefore, as a confirmation of
the results obtained by the previous procedure, we used Atmar
and Patterson’s (1995) nestedness calculator. This program uses
a thermodynamic model and measures the extent of order in a
matrix (T
°
) by Monte Carlo simulations (Atmar & Patterson, 1993).
The null-model normally used for this kind of analysis (Null Model
0) is known to overestimate the degree of nestedness; therefore, we
generated random matrices using the Null Model 1 proposed by
Fischer and Lindenmayer (2002). We generated random matrices
based on Null-model 1 using the program Random Matrix
Generator (P. Colombo, 2003), and subsequently we loaded them
into the nestedness calculator. The T
°
values for each matrix were
recorded; then we calculated the mean and standard deviation of
the resulting distribution of T
°
values. We calculated the associated
P
-values using a two-tailed
z
-test, performing 50 runs of Monte Carlo
simulation, as suggested by Fischer and Lindenmayer (2002).
RESULTS
Species richness
A total of 73 species was identified, 45 species in 24 terrestrial
mosses, 20 species in 17 alpine lakes, and 17, 15 and 8 species in
local assemblages from Res, Valnava and Argnaccia streams,
respectively (Fig. 1). Undetermined species, named ‘sp.’ followed
by a number, refer to morphotaxa that are recognizable but are
not previously described.
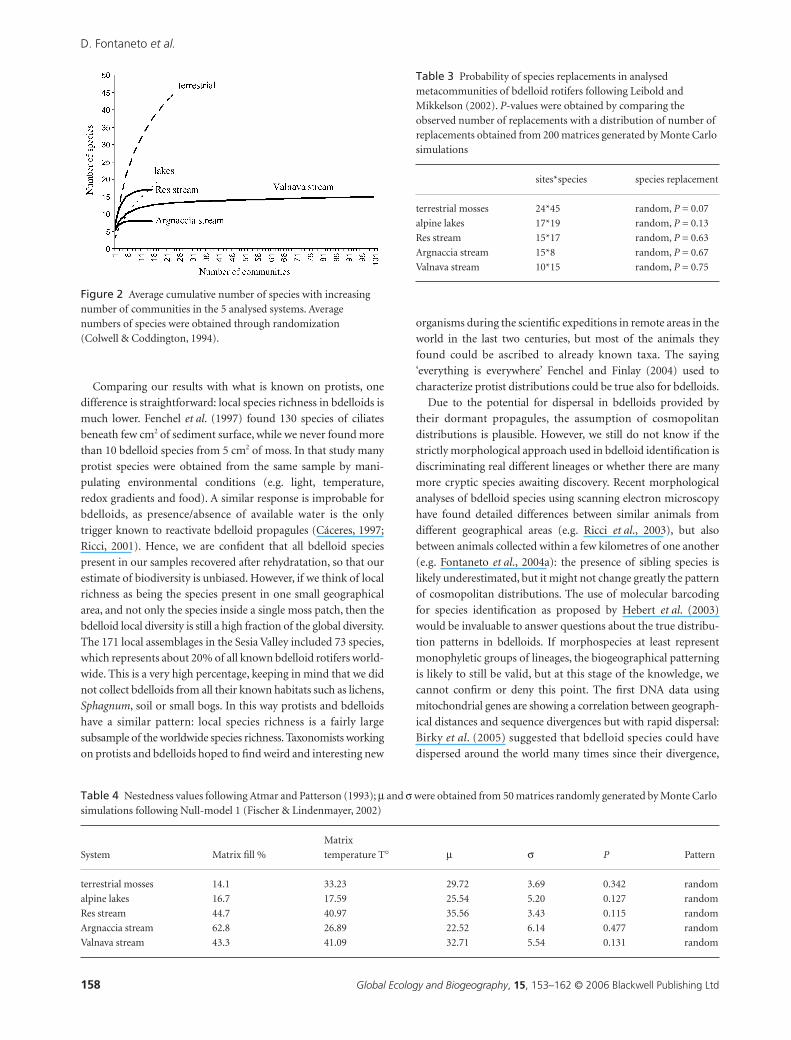
Species richness of samples differed significantly among the 5
systems (
test:
F
4,166
= 16.75,
P
< 0.0001): species richness
in alpine lakes was lower than in all the other systems, and species
richness in Res stream higher than in the other aquatic systems
(Table 1). These differences, although highly significant, did not
relate to any simple difference of aquatic versus terrestrial habi-
tats or connected versus unconnected systems.
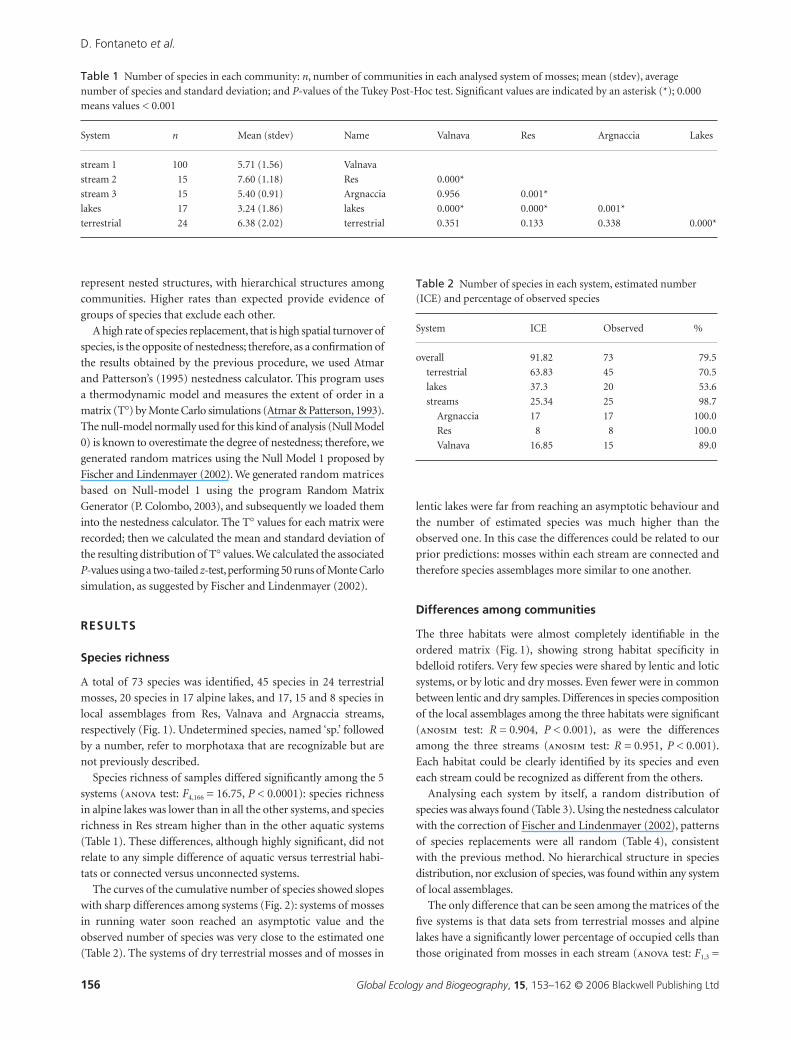
The curves of the cumulative number of species showed slopes
with sharp differences among systems (Fig. 2): systems of mosses
in running water soon reached an asymptotic value and the
observed number of species was very close to the estimated one
(Table 2). The systems of dry terrestrial mosses and of mosses in
lentic lakes were far from reaching an asymptotic behaviour and
the number of estimated species was much higher than the
observed one. In this case the differences could be related to our
prior predictions: mosses within each stream are connected and
therefore species assemblages more similar to one another.
Differences among communities
The three habitats were almost completely identifiable in the
ordered matrix (Fig. 1), showing strong habitat specificity in
bdelloid rotifers. Very few species were shared by lentic and lotic
systems, or by lotic and dry mosses. Even fewer were in common
between lentic and dry samples. Differences in species composition
of the local assemblages among the three habitats were significant
(
test:
R
= 0.904,
P
< 0.001), as were the differences
among the three streams (
test:
R
= 0.951,
P
< 0.001).
Each habitat could be clearly identified by its species and even
each stream could be recognized as different from the others.
Analysing each system by itself, a random distribution of
species was always found (Table 3). Using the nestedness calculator
with the correction of Fischer and Lindenmayer (2002), patterns
of species replacements were all random (Table 4), consistent
with the previous method. No hierarchical structure in species
distribution, nor exclusion of species, was found within any system
of local assemblages.
The only difference that can be seen among the matrices of the
five systems is that data sets from terrestrial mosses and alpine
lakes have a significantly lower percentage of occupied cells than
those originated from mosses in each stream (
test:
F
1,3
=
Table 1 Number of species in each community: n, number of communities in each analysed system of mosses; mean (stdev), average number of species and standard deviation; and P-values of the Tukey Post-Hoc test. Significant values are indicated by an asterisk (*); 0.000 means values < 0.001
System n Mean (stdev) Name Valnava Res Argnaccia Lakes
stream 1 100 5.71 (1.56) Valnava
stream 2 15 7.60 (1.18) Res 0.000*
stream 3 15 5.40 (0.91) Argnaccia 0.956 0.001*
lakes 17 3.24 (1.86) lakes 0.000* 0.000* 0.001*
terrestrial 24 6.38 (2.02) terrestrial 0.351 0.133 0.338 0.000*
Table 2 Number of species in each system, estimated number (ICE) and percentage of observed species
System ICE Observed %
overall 91.82 73 79.5
terrestrial 63.83 45 70.5
lakes 37.3 20 53.6
streams 25.34 25 98.7
Argnaccia 17 17 100.0
Res 8 8 100.0
Valnava 16.85 15 89.0

Diversity patterns in bdelloid rotifers
Global Ecology and Biogeography
,
15
, 153–162 © 2006 Blackwell Publishing Ltd
157
29.34,
P
= 0.012 on arcsin-squareroot transformed data). This is
because average similarity between local assemblages is higher in
stream systems than in the other two systems.
DISCUSSION
The analysis of distribution of bdelloid species in mosses from
different habitats revealed: (1) few species in each single assemblage
(low alpha diversity); (2) significant differences among systems, but
not in relation to prior expectations relating to presumed connect-
edness or moisture content; (3) that the overall number of species
(gamma diversity) differed among systems, with a higher number
of species in dry than in aquatic mosses; (4) different species compo-
sition of local assemblages from different systems, indicating strong
habitat selection; and (5) no hierarchical structure or clear exclusion
of species in the metacommunity pattern within each system.
Figure 1 Matrix of the presence of species in all the analysed communities, ordered with reciprocal averaging. Habitat type: ( ) terrestrial, ( ) lakes, (�) streams.
D. Fontaneto
et al.
158
Global Ecology and Biogeography
,
15
, 153–162 © 2006 Blackwell Publishing Ltd
Comparing our results with what is known on protists, one
difference is straightforward: local species richness in bdelloids is
much lower. Fenchel
et al
. (1997) found 130 species of ciliates
beneath few cm
2
of sediment surface, while we never found more
than 10 bdelloid species from 5 cm
2
of moss. In that study many
protist species were obtained from the same sample by mani-
pulating environmental conditions (e.g. light, temperature,
redox gradients and food). A similar response is improbable for
bdelloids, as presence/absence of available water is the only
trigger known to reactivate bdelloid propagules (Cáceres, 1997;
Ricci, 2001). Hence, we are confident that all bdelloid species
present in our samples recovered after rehydratation, so that our
estimate of biodiversity is unbiased. However, if we think of local
richness as being the species present in one small geographical
area, and not only the species inside a single moss patch, then the
bdelloid local diversity is still a high fraction of the global diversity.
The 171 local assemblages in the Sesia Valley included 73 species,
which represents about 20% of all known bdelloid rotifers world-
wide. This is a very high percentage, keeping in mind that we did
not collect bdelloids from all their known habitats such as lichens,
Sphagnum
, soil or small bogs. In this way protists and bdelloids
have a similar pattern: local species richness is a fairly large
subsample of the worldwide species richness. Taxonomists working
on protists and bdelloids hoped to find weird and interesting new
organisms during the scientific expeditions in remote areas in the
world in the last two centuries, but most of the animals they
found could be ascribed to already known taxa. The saying
‘everything is everywhere’ Fenchel and Finlay (2004) used to
characterize protist distributions could be true also for bdelloids.
Due to the potential for dispersal in bdelloids provided by
their dormant propagules, the assumption of cosmopolitan
distributions is plausible. However, we still do not know if the
strictly morphological approach used in bdelloid identification is
discriminating real different lineages or whether there are many
more cryptic species awaiting discovery. Recent morphological
analyses of bdelloid species using scanning electron microscopy
have found detailed differences between similar animals from
different geographical areas (e.g. Ricci
et al
., 2003), but also
between animals collected within a few kilometres of one another
(e.g. Fontaneto
et al
., 2004a): the presence of sibling species is
likely underestimated, but it might not change greatly the pattern
of cosmopolitan distributions. The use of molecular barcoding
for species identification as proposed by Hebert
et al
. (2003)
would be invaluable to answer questions about the true distribu-
tion patterns in bdelloids. If morphospecies at least represent
monophyletic groups of lineages, the biogeographical patterning
is likely to still be valid, but at this stage of the knowledge, we
cannot confirm or deny this point. The first DNA data using
mitochondrial genes are showing a correlation between geograph-
ical distances and sequence divergences but with rapid dispersal:
Birky
et al
. (2005) suggested that bdelloid species could have
dispersed around the world many times since their divergence,
Figure 2 Average cumulative number of species with increasing number of communities in the 5 analysed systems. Average numbers of species were obtained through randomization (Colwell & Coddington, 1994).
Table 3 Probability of species replacements in analysed metacommunities of bdelloid rotifers following Leibold and Mikkelson (2002). P-values were obtained by comparing the observed number of replacements with a distribution of number of replacements obtained from 200 matrices generated by Monte Carlo simulations
sites*species species replacement
terrestrial mosses 24*45 random, P = 0.07
alpine lakes 17*19 random, P = 0.13
Res stream 15*17 random, P = 0.63
Argnaccia stream 15*8 random, P = 0.67
Valnava stream 10*15 random, P = 0.75
Table 4 Nestedness values following Atmar and Patterson (1993); µ and σ were obtained from 50 matrices randomly generated by Monte Carlo simulations following Null-model 1 (Fischer & Lindenmayer, 2002)
System Matrix fill %
Matrix
temperature T° µ σ P Pattern
terrestrial mosses 14.1 33.23 29.72 3.69 0.342 random
alpine lakes 16.7 17.59 25.54 5.20 0.127 random
Res stream 44.7 40.97 35.56 3.43 0.115 random
Argnaccia stream 62.8 26.89 22.52 6.14 0.477 random
Valnava stream 43.3 41.09 32.71 5.54 0.131 random

Diversity patterns in bdelloid rotifers
Global Ecology and Biogeography
,
15
, 153–162 © 2006 Blackwell Publishing Ltd
159
consistent with cosmopolitan distributions. Monogonont
rotifers show a higher degree of geographical constraint than
previously thought (e.g. Segers, 2001; Gomez
et al
., 2002), but a
direct comparison with this taxon is not possible, as they repro-
duce by sex occasionally, and only hypotheses can be drawn on
the differences in speciation patterns without sex in bdelloids
(Barraclough
et al
., 2003).
Apart from cosmopolitanism, bdelloid species seem to be very
specific in their choice of substratum: almost no species overlap
was evident among the three analysed habitats. This means that
not all bdelloid propagules arriving in a place can survive, but that
at least three groups of moss-dwelling species can be described,
those able to live in dry terrestrial mosses, in submerged mosses
from running water and in submerged mosses from alpine lakes.
However, even within the riverine habitat, the three streams were
different in species composition. The two most similar streams,
Argnaccia and Res, were at higher elevation and on more acidic
substratum than the other stream, but further replicates
from different streams would be needed to correlate differences
with elevation or pH. Local assemblages in lakes were different
among different lakes, although lakes had similar pH, temperature,
rocky substratum, and surrounding and submerged vegetation
(Fontaneto & Melone, 2003). This strong selection for the habitat
is in contrast with the patterns found in protists; the generalization
that ‘most microorganisms can be found at the local seashore or
lake or, for that matter, in a garden pond’ (Fenchel & Finlay,
2004) might be true for protists but not for bdelloids. The above
generalization does not fit all protists, as a recent study on testate
amoeba (Protozoa, Rhizopoda) found significant differences in
species composition of communities from mosses with different
moisture (Vincke
et al
., 2004). The habitat specificity of some
bdelloid species can be confirmed by comparison with other
faunistic surveys; the only available recent data are from riverine
ecosystems (e.g. Zullini & Ricci, 1980; Schmid-Araya, 1998), and
their species lists are very close to the communities of our streams.
This does argue for a habitat-type explanation for rotifers not
being found everywhere. However, this explanation of habitat
differences between the streams is a posteriori, and we do not
have repeated habitat measures or samples to show that it is
habitat characteristics, rather than particular sites just being
separate, that determines composition in species.
An alternative explanation could be that colonization of each
habitat is rare and random from the global pool; therefore, there
are good chances of getting significant differences in species
composition. Once an area is colonized, subsequent colonization
events are harder, hence the partitioning among different areas
we sampled. More generally, bdelloids might have globally
cosmopolitan distributions but dispersal is rare enough that they
still have patchy distributions at narrower scales, unrelated to
habitat characteristics. This hypothesis could explain why species
replacements among local assemblages within each single system
revealed a random pattern, and why we could not identify groups
or gradients of species.
We suggest two possible explanations for the observed patterns:
(1) dispersal is very rare, and not all propagules are arriving
everywhere, so a species potentially able to live in a local assemblage
can be absent because no individual of this species ever reached
that particular moss; and (2) dispersal is effective in displacing
propagules, but environmental heterogeneity is much higher
than we measured: what is defined as a homogeneous system in
our analysis can be not homogeneous for bdelloids. These two
hypotheses, the former linked to stochastic events, and the latter
to ecological features, are not mutually exclusive and can both
contribute to the observed diversity pattern. Both hypotheses on
the effect of dispersal are supported by studies on other freshwater
organisms (e.g. Bohonak & Jenkins, 2003; Cohen & Shurin, 2003).
We found no differences in species occurrence in lakes
analysed in two subsequent years (Fontaneto & Melone, 2003),
suggesting that communities could be stable through time. This
observation is not enough to discriminate between the two
hypotheses, as in the case of effective dispersal, we do not know
whether different populations of the same species are replacing
one another through time, or if populations are stable. Ricci
et al
.
(1988) suggested an existing temporal turnover of different
clones of one bdelloid species in mosses, while De Meester
et al
.
(2002) suggested a monopolization hypothesis, with priority
effect, for other freshwater organisms.
Fenchel and Finlay (2004) argued that the composition and
diversity of a local biotic community in protists depend only on
the immigration and extinction of species populations, linked to
abiotic features, and not to interspecific interactions such as
competition and predation. This supports the general emphasis
given to dispersal and extinction as statistical phenomena more
important than species interactions in driving community
structure and diversity (Bell, 2001; Hubbell, 2001). The neutral
models apply only to guilds of species ecologically identical or
similar. This can be a reliable approximation for protists (Fenchel
& Finlay, 2004), and possibly for bdelloids, all being microfagous
filter-feeders or scrapers with very similar masticatory apparatuses
(Melone
et al
., 1998; Melone & Fontaneto, 2005). Moreover, the
random replacements of bdelloid species among local assem-
blages inside the same habitat could be easily explained by these
‘neutral’ models.
The differences in local species richness observed between
protists and bdelloids could be due to differences in reproductive
rates and therefore in effective population size: protists have
huge populations, and higher reproductive rates than bdelloids
(Ricci, 1983; Fenchel & Finlay, 2004). Therefore, a lower number
of individuals, and thus a lower number of propagules, can be
linked to low local richness, with higher differences among
patches, if each colonization event is less probable. Of course this
hypothesis has to be tested in further experiments.
In summary, it seems that bdelloid rotifers have some of the
peculiarities of protist biodiversity, but with much stronger
habitat selection. The most plausible explanation is that because
of their small size, bdelloids can be as easily dispersed as protists,
but then multicellular organisms are more complex in their
interactions with the environment and propagules cannot
survive everywhere.
Of course more work is needed in order to understand
meiofaunal biodiversity. No large data sets are available for other
small meiofaunal organisms. Data from mites, springtails and

D. Fontaneto
et al.
160
Global Ecology and Biogeography
,
15
, 153–162 © 2006 Blackwell Publishing Ltd
harpacticoid copepods indicate a higher degree of endemism and
historic biogeographical constraint (e.g. Jersabek
et al
., 2001;
Garrick
et al
., 2004; Niedbala, 2004), but these organisms,
although small, are larger than bdelloids and size matters for
dispersal mechanisms.
Regrettably, determining the global distribution of micro-
scopic organisms is a more difficult task than for large plants and
animals, because of undersampling in many parts of the world.
The vast majority of records of protists and meiofauna derive
from Europe and North America, with many fewer from most
other parts of the world. Intensive sampling is needed to clarify
biodiversity patterns of microscopic organisms. Even in the ‘well
known’ areas faunistic data are not available: only 54 bdelloid
species were known in Italy before this study (Braioni & Ricci,
1995; Fauna Europaea Service, 2004), and in our small valley we
found 73 species. Greater sampling and detailed molecular analyses
are needed to determine the true diversity of bdelloid rotifers.
Our results suggest that the geographical distribution of animals
and therefore biodiversity patterns may be strongly influenced
by animal size. This evidence of a gap between small and large
animals should be carefully taken into account in future studies
of biodiversity patterns.
ACKNOWLEDGEMENTS
We wish to thank Mathew A. Leibold for providing us the macro
for metacommunity analysis, P. Colombo for writing the
program Random Matrix Generator, Timothy G. Barraclough
for his suggestions on an earlier draft of the manuscript and for
revising the English text, and two anonymous reviewers for their
suggestions. Financial support came from a grant by the Italian
Space Agency.
REFERENCES
Allegro, G. & Sciaky, R. (2003) Assessing the potential role of
ground beetles (Coleoptera, Carabidae) as bioindicators in
poplar stands, with a newly proposed ecological index (FAI).
Forest Ecology and Management
,
175
, 275–284.
Andrew, N.R., Rodgerson, L. & Dunlop, M. (2003) Variation in
invertebrate–bryophyte community structure at different
spatial scales along altitudinal gradients.
Journal of Biogeography
,
30
, 731–746.
Atmar, W. & Patterson, B.D. (1993) The measure of order and
disorder in the distribution of species in fragmented habitat.
Oecologia
,
96
, 373–382.
Atmar, W. & Patterson, B.D. (1995
) The nestedness temperature
calculator: a visual basic program, including 294 presence-
absence matrices.
AICS Research Incorporate and the Field
Museum.
Audisio, P. & Vigna Taglianti, A. (2004)
Fauna Europaea: Coleoptera,
Carabidae
. Fauna Europaea Version 1.1, http://www.faunaeur.org
Barraclough, T.G., Birky, C.W. & Burt, A. (2003) Diversification
in sexual and asexual organisms.
Evolution
,
57
, 2166–
2172.
Bell, G. (2001) Neutral macroecology. Science, 293, 2413–2418.
Birky, C.W., Wolf, C., Maughan, H., Herbertson, L. & Henry,
E. (2005) Speciation and selection without sex. Hydrobiologia,
546, 29–45.
Bohonak, A.J. & Jenkins, D.G. (2003) Ecological and evolutionary
significance of dispersal by freshwater invertebrates. Ecology
Letters, 6, 783–796.
Bonn, A., Storch, D. & Gaston, K.J. (2004) Structure of the
species–energy relationship. Proceedings of the Royal Society of
London Series B, 271, 1685–1691.
Braioni, M.G. & Ricci, C. (1995) Rotifera. Vol. 8. Checklist delle
specie della fauna italiana (ed. by A. Minelli, S. Ruffo and S. La
Posta), pp. 1–11. Edagricole, Bologna.
Brehm, G., Sussenbach, D. & Fiedler, K. (2003) Unique eleva-
tional diversity patterns of geometrid moths in an Andean
montane rainforest. Ecography, 26, 456–466.
Burger, A. (1948) Studies on the moss dwelling bdelloids
(Rotifera) of Eastern Massachusetts. Transactions of the American
Microscopical Society, 67, 111–142.
Cáceres, C.E. (1997) Dormancy in invertebrates. Invertebrate
Biology, 116, 371–383.
Chazdon, R.L., Colwell, R.K., Denslow, J.S. & Guariguata, M.R.
(1998) Statistical methods for estimating species richness of
woody regeneration in primary and secondary rain forests of
NE Costa Rica. Forest biodiversity research, monitoring and
modeling: Conceptual background and Old World case studies
(ed. by F. Dallmeier and J.A. Comiskey), pp. 285–309. Parthe-
non Publishing, Paris.
Cohen, G.M. & Shurin, J.B. (2003) Scale-dependence and
mechanisms of dispersal in freshwater zooplankton. Oikos,
103, 603–617.
Coleman, A.W. (2002) Microbial eukaryote species. Science, 297,
337–337.
Colwell, R.K. (2004) EstimateS version 7. {http://viceroy.
eeb.uconn.edu/estimates}
Colwell, R.K. & Coddington, J.A. (1994) Estimating terrestrial
biodiversity through extrapolation. Philosophical Transactions
of the Royal Society of London Series B, 345, 101–118.
Crawley, M.J. (2002) Statistical computing. An introduction to
data analysis using S-plus. John Wiley & Sons, Chichester.
Davies, T.G., Savolainen, V., Chase, M.W., Moat, J. & Barraclough,
T.G. (2004) Environmental energy and evolutionary rates in
flowering plants. Proceedings of the Royal Society of London
Series B, 271, 2195–2200.
De Meester, L., Gómez, A., Okamura, B. & Schwenk, K. (2002)
The monopolization hypothesis and the dispersal-gene flow
paradox in aquatic organisms. Acta Oecologica, 23, 121–135.
Dimitrakopoulos, P.G. & Schmid, B. (2004) Biodiversity effects
increase linearly with biotope space. Ecology Letters, 7, 574–
583.
Donner, J. (1965) Ordnung Bdelloidea. Springer Verlag, Berlin.
Fauna Europaea Service (2004) {http://www.faunaeur.org}
Fenchel, T., Esteban, G.F. & Finlay, B.J. (1997) Local versus global
diversity of microorganisms: Cryptic diversity of ciliated
protozoa. Oikos, 80, 220–225.
Fenchel, T. & Finlay, B.J. (2004) The ubiquity of small species:
patterns of local and global diversity. BioScience, 54, 777–784.

Diversity patterns in bdelloid rotifers
Global Ecology and Biogeography, 15, 153–162 © 2006 Blackwell Publishing Ltd 161
Finlay, B.J. (2002) Global dispersal of free-living microbial
eukaryote species. Science, 296, 1061–1063.
Finlay, B.J. & Fenchel, T. (2002) Microbial eukaryote species:
response. Science, 297, 337–337.
Fischer, J. & Lindenmayer, D.B. (2002) Treating the nestedness
temperature calculator as a ‘black box’ can lead to false conclu-
sions. Oikos, 99, 193–199.
Fontaneto, D. & Melone, G. (2003) Bdelloid rotifers from
lakes above 1700 m in Western Italian Alps, with taxonomic
notes on Dissotrocha macrostyla. International Review of
Hydrobiology, 88, 594–601.
Fontaneto, D., Melone, G. & Cardini, A. (2004a) Shape diversity
in the trophi of different species of Rotaria (Rotifera, Bdelloidea):
a geometric morphometric study. Italian Journal of Zoology,
71, 63–72.
Fontaneto, D., Melone, G. & Ricci, C. (2004b) Structure of
bdelloid rotifer meta-communities in two alpine streams.
Acta Biologica, Studi Trentini di Scienze Naturali, 80, 23–
26.
Fontaneto, D., Melone, G. & Ricci, C. (2005) Connectivity
and nestedness of the meta-community structure of moss
dwelling bdelloid rotifers along a stream. Hydrobiologia,
542, 131–136.
Francez, A.J. (1980) Quelques Bdelloides (Rotiferes) des
tourbieres d’Auvergne. Revue du Science Naturelle d’Auvergne,
46, 21–26.
Garrick, R.C., Sands, C.J., Rowell, D.M., Tait, N.N., Greenslade, P.
& Sunnucks, P. (2004) Phylogeography recapitulates topography:
very fine-scale local endemism of a saproxylic ‘giant’ springtail
at Tallaganda in the Great Dividing Range of south-east
Australia. Molecular Ecology, 13, 3329–3344.
Gaston, K.J., Davies, R.G., Gascoigne, C.E. & Williamson, M.
(2005) The structure of global species-range size distribu-
tions: raptors & owls. Global Ecology and Biogeography, 14,
67–76.
Global Amphibian Assessment (2005) Global Amphibian Assess-
ment. http://www.globalamphibians.org
Gomez, A., Adcock, G.J., Lunt, D.H. & Carvalho, G.R. (2002)
The interplay between colonization history and gene flow in
passively dispersing zooplankton: microsatellite analysis of
rotifer resting egg banks. Journal of Evolutionary Biology, 15,
158–171.
Hausdorf, B. & Hennig, C. (2004) Does vicariance shape biotas?
Biogeographical tests of the vicariance model in the north-
west European land snail fauna. Journal of Biogeography, 31,
1751–1757.
Hebert, P.D.N., Ratnasingham, S. & deWaard, J.R. (2003)
Barcoding animal life: cytochrome c oxidase subunit 1 diver-
gences among closely related species. Proceedings of the Royal
Society of London Series B, 270, 96–99.
Hubbell, S.P. (2001) The unified neutral theory of biodiversity and
biogeography. Princeton University Press, Princeton (NJ).
Jersabek, C.D., Brancelj, A., Stoch, F. & Schabetsberger, R. (2001)
Distribution and ecology of copepods in mountainous regions
of the Eastern Alps. Hydrobiologia, 453, 309–324.
Jung, W. (1936) Thekamöben ursprünglicher, lebender deutscher
Hochmoore. Abhandlungen Landesmuseum Provinz Westfalen
Museum Naturkunde, 7, 1–87.
Kotze, D.J. & O’Hara, R.B. (2003) Species decline — but why?
Explanations of carabid beetle (Coleoptera, Carabidae)
declines in Europe. Oecologia, 135, 138–148.
Lawton, J.H. (1999) Are there general laws in ecology? Oikos, 84,
177–192.
Leibold, M.A. & Mikkelson, G.M. (2002) Coherence, species
turnover, and boundary clumping: elements of meta-
community structure. Oikos, 97, 237–250.
Linhart, J., Fiuraskova, M. & Uvira, V. (2002) Moss- and mineral
substrata-dwelling meiobenthos in two different low-order
streams. Archive für Hydrobiologie, 154, 543–560.
McAbendroth, L., Foggo, A., Rundle, S.D. & Bilton, D.T. (2005)
Unravelling nestedness and spatial pattern in pond assemblages.
Journal of Animal Ecology, 74, 41–49.
Meisterfeld, R. (1977) Die horizontale und vertikale Verteilung
der Testaceen (Rhizopoden, Testacea) in Sphagnum. Archive
für Hydrobiologie, 79, 319–356.
Melone, G., Ricci, C. & Segers, H. (1998) The trophi of Bdelloidea
(Rotifera): a comparative study across the class. Canadian
Journal of Zoology, 76, 1755–1765.
Melone, G. & Fontaneto, D. (2005) Trophi structure in bdel-
loid rotifers. Hydrobiologia, 546, 197–202.
Mitchell, E.A.D., Buttler, A.J., Warner, B.G. & Gobat, J.M. (1999)
Ecology of testate amoebae (Protozoa: Rhizopoda) in
Sphagnum peatlands in the Jura mountains, Switserland and
France. Ecoscience, 6, 565–576.
Monge-Najera, J. & Hou, X.G. (2000) Disparity, decimation and
the Cambrian ‘explosion’: comparison of early Cambrian
and Present faunal communities with emphasis on velvet worms
(Onychophora). Revue du Biologie Tropicale, 48, 333–351.
Niedbala, W. (2004) Ptyctimous mites (Acari, Oribatida) of the
neotropical region. Annales Zoologici, 54, 1–288.
Örstan, A. (1998) Microhabitats and dispersal routes of bdelloid
rotifers. Scientiae Naturae, 1, 27–36.
Persson, A. (2002) Proliferation of cryptic protists and germina-
tion of resting stages from untreated sediment samples with
emphasis on dinoflagellates. Ophelia, 55, 152–166.
R Development Core Team (2004) R: A language and environ-
ment for statistical computing. Vienna, Austria. {http://www.
R-project.org}
Rahbek, P. (2005) The role of spatial scale and the perception of
large-scale species-richness patterns. Ecology Letters, 8, 224–
239.
Ricci, C. (1983) Life histories of some species of Rotifera
Bdelloidea. Hydrobiologia, 104, 175–180.
Ricci, C. (1987) Ecology of bdelloids: how to be successful.
Hydrobiologia, 147, 117–127.
Ricci, C. (2001) Dormancy patterns in rotifers. Hydrobiologia,
446, 1–11.
Ricci, C., Pagani, M. & Bolzern, A.M. (1988) Temporal analysis
of clonal structure in a moss bdelloid population. Hydro-
biologia, 186/187, 145–152.
Ricci, C., Shiel, R.J., Fontaneto, D. & Melone, G. (2003) Bdelloid
rotifers recorded from Australia with description of

D. Fontaneto et al.
162 Global Ecology and Biogeography, 15, 153–162 © 2006 Blackwell Publishing Ltd
Philodinavus aussiensis n.sp. Zoologischer Anzeiger, 242,
241–248.
Rosenzweig, M.L. (1995) Species diversity in space and time.
Cambridge University Press, Cambridge.
Sanmartin, I. & Ronquist, F. (2004) Southern Hemisphere
biogeography inferred by event-based models: Plant versus
animal patterns. Systematic Biology, 53, 216–243.
Schmid-Araya, J.M. (1998) Small-sized invertebrates in a gravel
stream: community structure and variability of benthic rotifers.
Freshwater Biology, 39, 25–39.
Segers, H. (2001) Zoogeography of the Southeast Asian Rotifera.
Hydrobiologia, 446, 233–246.
Suarez-Morales, E., Reid, J.W., Fiers, F. & Iliffe, T.M. (2004)
Historical biogeography and distribution of the freshwater
cyclopine copepods (Copepoda, Cyclopoida, Cyclopinae) of
the Yucatan Peninsula, Mexico. Journal of Biogeography, 31,
1051–1063.
Tribsch, A. (2004) Areas of endemism of vascular plants in the
Eastern Alps in relation to Pleistocene glaciation. Journal of
Biogeography, 31, 747–760.
Vincke, S., Gremmen, N., Beyens, L. & Van de Vijver, B. (2004)
The moss dwelling testacean fauna of Île de la Possession. Polar
Biology, 27, 753–766.
Wallace, R.L. & Ricci, C. (2002) Rotifera. Freshwater meiofauna:
biology and ecology (ed. by S.D. Rundle, A.L. Robertson and
J.M. Schmid-Araya), pp. 15–44. Backhuys Publishers, Leiden,
The Netherlands.
Zullini, A. & Ricci, C. (1980) Bdelloid rotifers and nematodes in
a small Italian stream. Freshwater Biology, 10, 67–72.
Editor: Tim Blackburn
BIOSKETCHES
Diego Fontaneto has research interest in the fields of
systematics, phylogeny, and community ecology of
invertebrates, in particular of bdelloid rotifers.
G. Francesco Ficetola is interested in determinants of
biodiversity. He is currently researching conservation
biology and community ecology of amphibians and the
role of environmental factors at different scales.
Roberto Ambrosini is interested in behavioural ecology
and conservation biology of birds, in particular of the
barn swallow.
Claudia Ricci is a zoologist dealing with systematics and
ecology of bdelloid rotifers and thus with anhydrobiotic
responses, parthenogenetic reproduction and its
consequences on populations.




















