
Gut-retention time in mycophagous mammals: a review and a study of truffle-like fungal spore...
12
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
edithcowan -
Category
Documents
-
view
2 -
download
0
Transcript of Gut-retention time in mycophagous mammals: a review and a study of truffle-like fungal spore...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Gut-retention time in mycophagous mammals: a review anda study of truffle-like fungal spore retention in the swampwallaby
Melissa A. DANKS*
Ecosystem Management, University of New England, Armidale, New South Wales 2351, Australia
a r t i c l e i n f o
Article history:
Received 28 February 2011
Revision received 11 July 2011
Accepted 13 July 2011
Available online 2 October 2011
Corresponding editor:
Jeff Powell
Keywords:
Forest ecology
Gut-passage
Marsupials
Mean retention time
Mycophagy
Mycorrhizas
Spore dispersal
Truffles
a b s t r a c t
Variation in the gut-retention time of macrofungal spores influences the distance to which
spores are dispersed by mycophagous (fungus-feeding) mammals and is of interest in
examining mammal-fungal interactions. In reviewing published studies of fluid and
particle (including macrofungal spore) digesta gut-retention times in ground-dwelling
mycophagous mammals, weighted mean retention times (MRT) were found to range
6.6e55.5 hr. Among macropodoid marsupials, fluid and small particle weighted MRT was
longer in mycophagous species than non-mycophagous species but statistical support for
this difference was weak (estimated mean difference 7.2 hr; 95 % CL [�0.8, 15.1] hr). Gut-
retention of truffle-like (below-ground fruiting) fungal spores was examined in the
swamp wallaby (Wallabia bicolor), a browsing macropodid marsupial that regularly eats
macrofungal fruit-bodies. Two wallabies of different body weights were examined in
a captive feeding trial. MRT of marker spores were 26.9 hr and 35.1 hr for the larger and
smaller animal respectively. A small number of marker spores were found in faecal pellets
up to 69 hr after ingestion, suggesting that there is potential for long distance dispersal of
fungal spores by swamp wallabies. The studied swamp wallabies probably carry fungal
spores for similar times to smaller mycophagous marsupials, including the strongly
mycophagous potoroids. Further studies of spore gut-passage, including MRT, in
mycophagous mammals would help clarify differences among species and groups of
species.
ª 2011 Elsevier Ltd and The British Mycological Society. All rights reserved.
Introduction
Interactions between mycorrhizal fungi, host plants, and
mammals influence forest ecosystem function. Trees and
other woody plants form biotrophic symbioses with ectomy-
corrhizal (EM) fungi (Brundrett 1991; Read 1991). Most EM fungi
produce fruit bodies (sporocarps) and these are an important
food resource for many mammals (Fogel & Trappe 1978;
Claridge & May 1994; Claridge et al. 1996; Maser et al. 2008).
Mammals are vital spore dispersal agents, particularly for
truffle-like (below-ground fruiting) sporocarpic fungi that do
not actively discharge their spores (Fogel & Trappe 1978;
Claridge & May 1994; Claridge et al. 1996; Johnson 1996;
Reddell et al. 1997; Bougher & Lebel 2001; Maser et al. 2008).
Spore dispersal is important for both maintenance of genetic
flow within and between fungal populations and for coloniza-
tion of new habitats (Bruns et al. 2009). For mycorrhizal host
plants and the plant-soil system, maintenance of a diverse
* Tel.: 61 424726667.E-mail address: [email protected]
available at www.sciencedirect.com
journal homepage: www.elsevier .com/locate/ funeco
1754-5048/$ e see front matter ª 2011 Elsevier Ltd and The British Mycological Society. All rights reserved.doi:10.1016/j.funeco.2011.08.005
f u n g a l e c o l o g y 5 ( 2 0 1 2 ) 2 0 0e2 1 0

Author's personal copy
mycorrhizal fungal community maintains resilience through
resource partitioning and competition amongst fungi (Perry
et al. 1989; Deacon & Fleming 1992; Bruns 1995). Rapid
seasonal turnover of plant root tips results in strong local-scale
competition among mycorrhizal fungi for this resource (Bruns
1995). By disseminating spores in their faecal pellets, mycoph-
agous (fungus-feeding) mammals help maintain fungal diver-
sity within their home range (Maser et al. 2008). While most
mycophagousmammalmovement occurswithin a frequently-
used area (home range), occasional long distance movements
occur and may be significant spore dispersal events.
Few mycophagous mammals remain on the New England
Tableland of north-eastern New South Wales; bettongs,
potoroos, and bandicoots have largely been extirpated from
the modified landscapes of this region in the period since
European occupation (Jarman & Vernes 2006). However, the
swamp wallaby, Wallabia bicolor, a plant browser and regular
mycophagist is resilient in these EM-forested landscapes.
Swamp wallabies consume a diversity of truffle-like fungi
year-round (M. Danks, unpublished data; Claridge et al. 2001;
Vernes & McGrath 2009; Vernes 2010) and may contribute to
the maintenance of vital mammal-truffle-plant relationships
in these simplified communities.
Gut-retention time of fungal spores in mycophagous
mammals is of interest in examining mammal-fungal inter-
actions as, along with movement patterns, variation in gut-
retention influences the distance to which spores may be
dispersed (Cork & Kenagy 1989b). Gut-passage is influenced by
body mass, gut morphology and diet (Cork & Kenagy 1989b;
Hume 1989). Most studies examining mammalian digesta
passage have held study animals in metabolism cages and
used chemical or physical markers to illustrate the passage of
fluid or particle digesta phases (e.g. Calaby 1958; Warner
1981a; Hume & Carlisle 1985; Sakaguchi & Hume 1990; Moyle
et al. 1995; McClelland et al. 1999; Gibson & Hume 2000; Pei
et al. 2001). Gut-passage has rarely been studied in free-
ranging mammals on a natural diet, making it problematic
to interpret the ecological meaning of reported gut-retention
times. Gut-retention time of fungal spores has also received
relatively little attention. The few studies that have directly
assessed gut-retention of fungal spores have examined
mycophagous rodents; for example, the giant white-tailed rat,
Uromys caudimaculatus (Comport & Hume 1998), the golden-
mantled ground squirrel, Spermophilus saturatus, and the
deer mouse, Peromyscus maniculatus (Cork & Kenagy 1989b).
The present study is the first to assess spore gut-passage in
a mycophagous mammal on a semi-natural diet, and the first
to measure spore gut-passage in a macropodoid marsupial.
In this paper I: (1) review published data on gut-retention
times in mycophagous mammals, including macropodids
(wallabies), potoroids (potoroos and bettongs), peramelids
(bandicoots), rodents (squirrels, voles, andmice), and possums;
and (2) examine the time taken for native truffle-like fungal
spores to pass through the gut of swamp wallabies. I use gut-
retention times to broadly examine the spore dispersal role of
these mammals rather than to compare their digestive effi-
ciency. Factors influencing digesta passage are discussed in
relation to fluid, small particle, and macrofungal spore gut-
retention time in the swamp wallaby and other mycophagous
mammals. Fluid and particulate digesta pass through the
digestive tract as different digesta phases (Faichney 1975).
Spores of EM fungi are thought, due to their small size of
generally <20 mm diameter, to move through the gut with the
fluid phase although some spores may remain attached to
sporocarp fragments and pass through the gut with the large
particlephase (Comport&Hume1998).Sporegut-retention time
in the swampwallaby is expected to bemost similar to fluidgut-
retention times reported for other browsing and grazing
wallabies as, despite their smaller size, these animals are most
similar to swampwallabies in termsofdietandgutmorphology.
Materials and methods
Published data on digesta gut-passage in mycophagousmammals
Published data on macrofungal spore, fluid, and particle
digesta mean retention times (MRT) in mycophagous
mammals were collated. Comparisons among macropodoid
marsupials (macropodids e kangaroos and wallabies; and
potoroids e potoroos, bettongs and rat-kangaroos), both
mycophagous and non-mycophagous, were also made. The
50 % excretion time (ET) measure, while not the same as MRT,
is considered similar enough for broad comparisons of gut-
retention times to be made (Stevens & Hume 1995), so 50 %
ET measures were included in this summary where MRT was
not reported. Both measures are referred to here as ‘gut-
retention time’. Several researchers provide information on
digesta passage in mycophagous mammals but do not report
MRT or 50 % ET (e.g. Calaby 1958; Hume & Carlisle 1985;
Richardson 1989), preventing comparison with the present
study, so these studies were not included in the summary.
In studies of large particle (size ranges between
300e1200 mm diameter) MRT, various chemical or radio-
isotope markers, including chromium mordanted onto cell-
wall constituents (Cr-CWC), dyed leaf particles, and 103Ru-
labelled tris-(1,10 phenanthroline)-ruthenium (11) chloride
(Ru-P) leaf particles were utilised. Fluid digesta MRT was
assessed using either cobalt-ethylenediamine tetra-acetic
acid (Co-EDTA), or the 51chromium complex of ethylenedi-
amine tetra-acetic acid (Cr-EDTA). The marker used can
influence the measured rate of passage of digesta. For
example, Ru-P migrates from large to small digesta particles
within the gut of sheep (Faichney & Griffiths 1978; Egan &
Doyle 1984) and macropods (Freudenberger & Hume 1992) so
that MRT of large particles may be underestimated using this
marker. Cr-EDTA may not completely associate with the fluid
phase (Faichney 1975), while Co-EDTA associates almost
exclusively with fluid (Ud�en et al. 1980). Additionally, the fibre
content and particle size distribution of the experimental diet,
frequency of feeding, and animal activity levels will also
influence digesta gut-passage and thereforemeasures of MRT.
Sakaguchi & Hume (1990) reported small (<75 mm diameter)
particle MRT and compared passage with that of fluids and
larger (300e600 mm) particles. The fine particles passed
through the gut with the fluid digesta phase. Therefore, I have
assumed measures of fluid digesta passage to be measures of
the passage of both solutes and small particulate digesta
(<75 mm).
Gut-retention time in mycophagous mammals 201

Author's personal copy
For the purpose of drawing broad comparisons among
mycophagous mammal species, weighted average MRT esti-
mates were calculated for each animal species (weighted by
original study sample size) and group of species (weighted by
the number of sources), based on mean MRT values reported
for those species on a range of experimental diets and varying
methodology. To compare average MRT between mycopha-
gous and non-mycophagous macropodoid marsupials, 95 %
confidence limits were calculated around the estimatedmean
difference between these groups. Sample size was generally
small (range 1e13 animals per experiment) and the number of
sources per species ranged between one and nine. Related
information including trophic classification, body mass,
and presence of selective gut-retention mechanisms was
summarised.
Swamp wallabies
Swamp wallabies were trapped in eucalypt woodland at
Newholme Field Station, northern New SouthWales, Australia
in Jun. 2009, and housed in large (1e1.2 ha) naturally vegetated
(eucalypt woodland) pens on site. Four purpose-built soft-
walled wallaby traps, 700 mm� 1200 mm� 700 mm, based on
the design of Di Stefano et al. (2005) were baited and set late
each afternoon, and checked early the following morning, for
three consecutive nights. Trapping to source animals for the
feeding trial was limited to three consecutive nights to mini-
mise the time for which animals would be held prior to
commencing the feeding trial, and only adults in good condi-
tion and without pouch young or young-at-foot were selected
for the feeding trial. Two wallabies were used in this study;
both were adult males and each weighed 21 kg and 10 kg
respectively. Lucerne hay, pelleted feed and apple pieces were
provided as supplementary feed throughout, although
wallabies were observed to browse on shrubs and grasses.
Duringanacclimationperiod of 3 d, fresh faecal pelletswere
regularly collected and samples examined microscopically for
the presence of macrofungal spores. A �1 cm3 portion was
taken from each pellet-group. Sub-samples (0.5 g) were rehy-
drated in 10 ml water for 1 hr, macerated, and washed through
a 50 mm� 50 mmsievewith a further 20 mlofwater. The sample
was then centrifuged and the supernatant removed by pipette
until 2 ml remained. The remaining sample was thoroughly
mixed and three drops placed on a slide with one drop of
Melzer’s Reagent. Melzer’s Reagent produces colour reactions
that aid identification of some fungi. Slideswere systematically
scanned at �400 magnification with an Olympus CX41 light
microscope (Fig 1A). Magnification of�1000 was used to clarify
spore morphology to aid identification. It was essential to
determine an appropriate fungal ‘marker’ taxon thatwas easily
found in the environment, but not currently in the diet of the
swamp wallabies in the pen area. Austrogautieria clelandii was
selected as amarker for this trial because (1) it was not found in
faecal pellets of the captured wallabies during the acclimation
period, (2) it is known to occur and be consumed by swamp
wallabies at nearby sites (M. Danks, unpublished data), (3) it is
easily identifiedby itsdistinctivesporemorphology (Fig 1B), and
(4) it was readily available in a quantity suitable for this study.
After theacclimationperiod, thewallabieswere recaptured, fed
(by use of a syringe to the back of themouth) a 10 ml pulse dose
of slurry ofA. clelandii sporocarps, and released back to the pen.
The dose was delivered as a slurry to ensure ingestion of
a standard quantity of spores (equivalent to approximately two
large sporocarps) at a known time. Swampwallabies were held
for a further 4 ½ d and then released. Wallaby capture and
handling protocol followed guidelines of the American Society
of Mammalogists (Gannon et al. 2007) and was approved by the
Animal Ethics Committee at the University of New England
(code AEC09-023).
Gut-passage of macrofungal spore digesta
Prior to dosing wallabies with the spore slurry, three transects
perpen (totalling5 %ofpenarea)weremarkedandclearedofall
Fig 1 e (A) EM of fungal spores in swamp wallaby faecal
pellet during acclimation period (pellet material stained
with Melzer’s Reagent to aid identification of some groups
of fungi). (B) Distinctive ridged spores of Austrogautieria sp.
in swamp wallaby faecal pellet collected after oral dosing
with Austrogautieria sp. sporocarp slurry. Inset:
Austrogautieria sp. spores from fresh sporocarp, showing
side and end view of spores. Micrographs taken at 4003
magnification (Inset micrograph taken at 10003
magnification). Bar scale for all images 10 mm.
202 M.A. Danks

Author's personal copy
faecal pellets. Faecal pelletswere collected fromthese transects
for 4 ½ d following dosing (4 hourly for 48 hr, then 6 hourly for
the next 36 hr, and then 12 hourly for a further 24 hr). Faecal
material was oven-dried (40 �C for 12e24 hr) and bulked by
collection period. Samples were processed for microscopic
examination as described earlier, omitting Melzer’s Reagent,
which was not necessary to distinguish marker spores. Slides
were examined at 400� magnification and the number of
marker spores in 30fields-of-view counted. Amounts ofmarker
excreted were expressed as proportions of the total number of
marker spores seen in faecal samples. ETs were estimated as
the mid-point of each collection period. Overall (dose to excre-
tion) mean retention time (MRT), the average time taken for
a marker particle to be excreted after a pulse dose, was calcu-
latedbyuseof the formulaP
(Miti)/P
MiwhereMi is theamount
of marker excreted in the ith defecation at time ti (Blaxter et al.
1956) as recommended byWarner (1981b).
Results
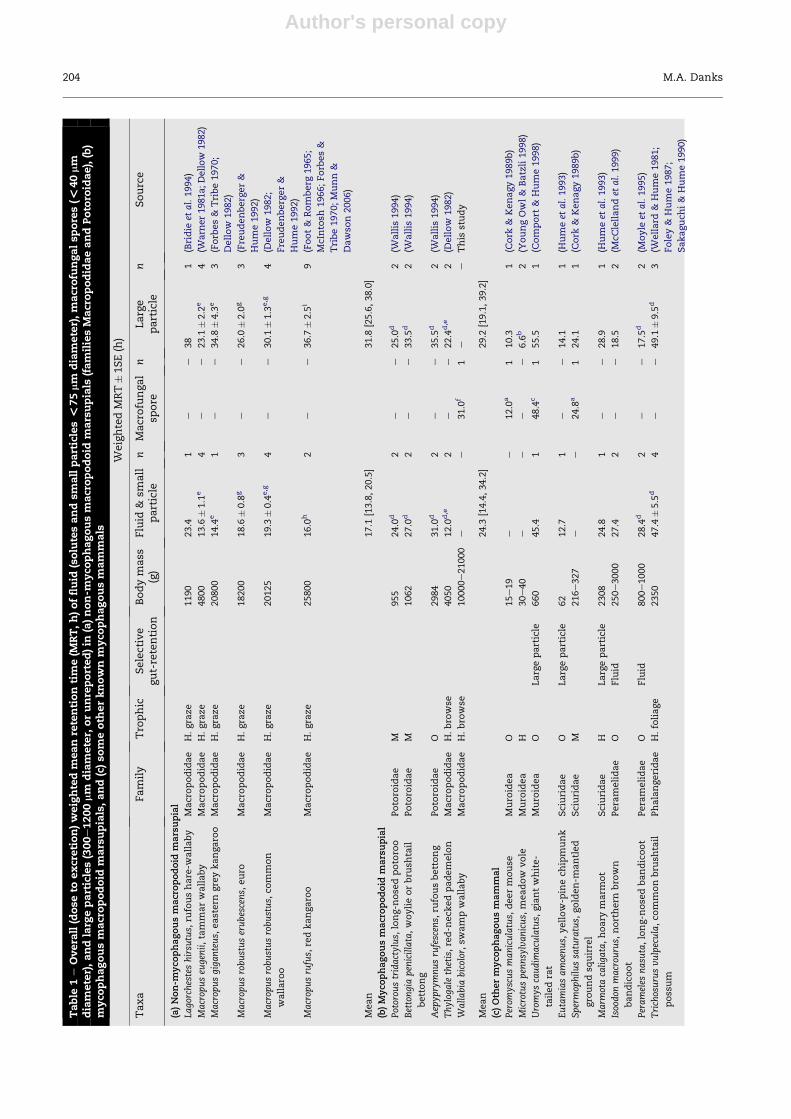
Digesta gut-passage in mycophagous mammals
Previous researchers specifically examining macrofungal
spore gut-passage examined mycophagous rodents with
mean body mass of 15e660 g and reported spore MRT of
12e48.4 hr. Studies of fluid (and small particle) and large
particle digesta marker passage in mycophagous macro-
podids, potoroids, peramelids, phalangerids, and rodents re-
ported fluid and particle marker MRT or 50 % ET of 6e71 hr.
WeightedMRT for fluid and small particles was shortest in the
red-necked pademelon Thylogale thetis (12 hr) and longest in
the brushtail possum Trichosurus vulpecula (47.4� 5.5 hr;
Table 1). Weighted MRT for large particles ranged from 6.6 hr
in the meadow vole Microtus pennsylvanicus to 49.1 (�9.5) hr in
the brushtail possum.
Digesta gut-passage in macropodoid marsupials
Among macropodoid marsupials, weighted MRT for fluid and
small particles (including macrofungal spores) ranged from
12 hr in the small browsing red-necked pademelon to 31 hr in
both the swamp wallaby and the small omnivorous rufous
bettong Aepyprymnus rufescens (Table 1). Fluids and small
particles including macrofungal spores were retained for
longer by mycophagous macropodoids (24.3 [14.4, 34.2] hr;
weighted mean [lower 95 % CL, upper 95 % CL]) than by non-
mycophagous macropodoids (17.6 [13.8, 20.5] hr), although
statistical support for a mean difference was weak (7.2 [�0.8,
15.1] hr: mean difference, [lower 95 % CL, upper 95 % CL]).
There was no support for a mean difference in large particle
MRT between the two groups (mycophagous 29.2 [19.1, 39.2]
hr; non-mycophagous 31.8 [25.6, 38] hr; mean difference 2.6
[�6.4, 11.6] hr).
Passage of fungal spore marker through swamp wallaby gut
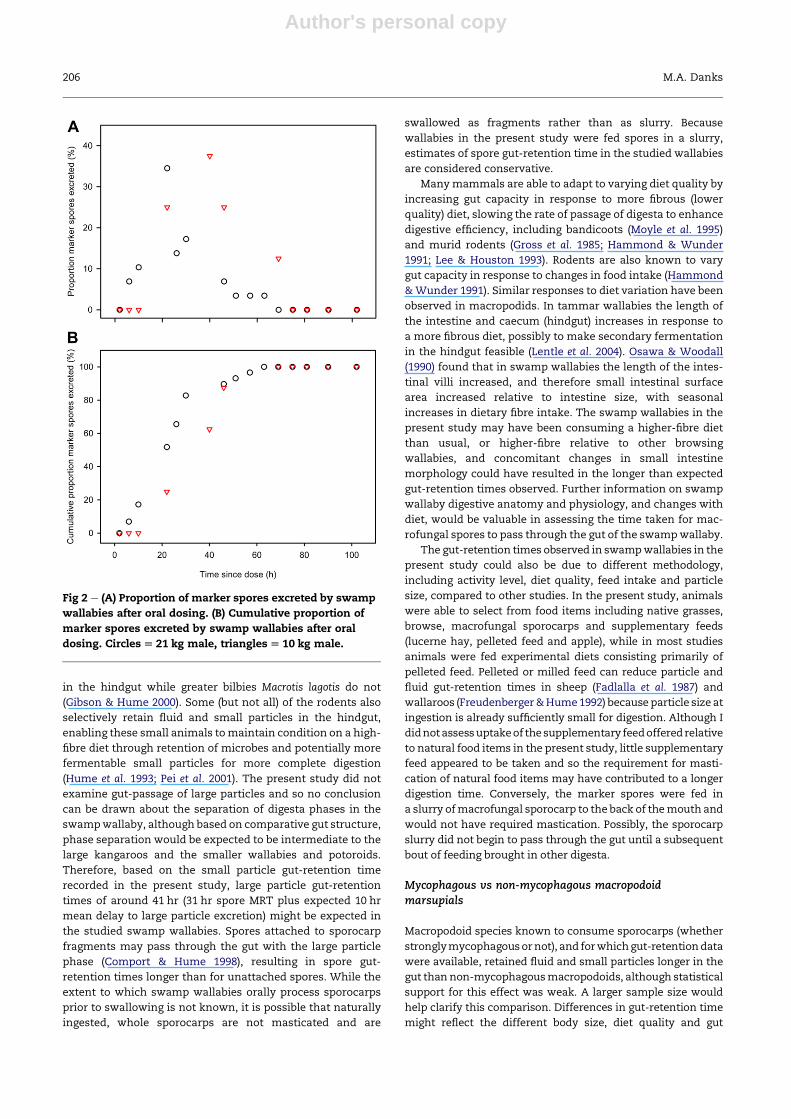
Patterns of excretion of marker spores over time were similar
but the timing of excretion differed slightly between the two
animals (Fig 2). The marker spores were first found in the
faecal pellets of the 21 kg and 10 kg animals at 6 hr and 22 hr,
andwere last seen at 63 hr and 69 hr respectively (Fig 2A). MRT
of spores was 27 hr and 35 hr, and modal retention time was
22 hr and 43 hr respectively. Fifty percent of themarker spores
were eliminated from the gut of both animals within 34 hr
(21 and 34 hr) and 90 % were eliminated within 51 hr (46 and
51 hr) after ingestion (Fig 2B).
Discussion
Fungal spore retention time in the swamp wallaby gut
This is the first study to examine digesta passage rate in the
swampwallaby, one of the few studies using a natural dietary
marker (macrofungal spores) to assess gut-retention time, and
one of the few studies to determine gut-retention time for
animals kept under semi-natural conditions. My results
suggest that the swamp wallabies sampled can carry ingested
spores for a mean 27e35 hr and some spores for up to 3 d. In
studies of smaller mycophagous mammals (potoroids and
rodents), spores remain viable after passage through the gut
(Kotter & Farentinos 1984; Lamont et al. 1985; Cork & Kenagy
1989a; Claridge et al. 1992; Caldwell et al. 2005). I therefore
assume that spores do not lose viability on dissemination in
swamp wallaby faeces and that swamp wallabies contribute
to effective dispersal of these spores. Swampwallabies occupy
home ranges 3e86 ha in size (M. Danks unpublished data;
Edwards & Ealey 1975; Troy & Coulson 1993; Di Stefano 2010;
Di Stefano et al. 2011) and so have the potential to disperse
spores across large areas. Occasional longer-distance move-
ments beyond the home range are also potentially significant
spore dispersal events.
Digesta gut-passage in macropodoid marsupials
Macropodoid marsupials are almost all foregut fermenters;
they have an enlarged forestomach, the principal site of
microbial digestion, made up of several regions, and a simple
hindgut (Langer 1980; Langer et al. 1980; Hume 1982). With
increasing body mass, the tubiform region of the forestomach
tends to increase in size and the sacciform region of the for-
estomach tends to decrease in size (Hume 1984). This trend is
explained by the ability of larger animals to utilise a more
fibrous (lower quality) diet; sincemetabolic requirements scale
with body size large animals require less energy relative tobody
mass thando small animals (Demment&Soest 1985). Anotable
exception to this trend is the rufous hare-wallaby, which
despite being small (1 kg; similar in size to the small potoroids),
has a high-fibre grazing diet and ‘kangaroo-like’ stomach
morphology (Bridie et al. 1994). The rufous hare-wallaby offsets
the disadvantages of small size on a high-fibre diet with low
food intake, discontinuous feeding, and a long distal colon for
storageof faeces, resulting in relatively longgut-retentiontimes
and thusmoreefficientdigestionofhigh-fibre food items (Bridie
et al. 1994). Digesta passes through the predominantly tubiform
stomach of the larger kangaroos relatively quickly (Hume 1984)
while in the potoroids the sacciform forestomach acts as
a storage chamber and fermentation site increasing the overall
rate of passage of digesta (Freudenberger et al. 1989).
Gut-retention time in mycophagous mammals 203
Author's personal copy
Table
1e
Overa
ll(dose
toexcretion)weightedm
eanretentiontim
e(M
RT,h)offluid
(solutesandsm
allparticles<
75mm
diam
eter),m
acrofu
ngalsp
ores(<
40mm
diam
eter),andlarg
eparticles(300e1200mm
diam
eter,
orunreported)in
(a)non-m
yco
phagousm
acropodoid
marsupials
(fam
iliesMacropodidaeandPotoro
idae),(b)
myco
phagousm
acropodoid
marsupials,and(c)so
meoth
erknownm
yco
phagousm
am
mals
WeightedMRT�1SE(h)
Taxa
Family
Tro
phic
Selective
gut-retention
Bodymass
(g)
Fluid
&sm
all
particle
nMacrofu
ngal
spore
nLarge
particle
nSource
(a)Non-m
yco
phagousmacropodoid
marsupial
Lago
rchesteshirsu
tus,
rufoushare-w
allaby
Macropodidae
H.graze
1190
23.4
1e
e38
1(Bridie
etal.1994)
Macrop
useu
genii,tammarwallaby
Macropodidae
H.graze
4800
13.6
�1.1
e4
ee
23.1
�2.2
e4
(Warn
er19
81a;Dellow
1982
)
Macrop
usgiga
nteus,
eastern
greykangaro
oMacropodidae
H.graze
20800
14.4
e1
ee
34.8
�4.3
e3
(Forb
es&
Tribe1970;
Dellow
1982)
Macrop
usrobustuserubescens,
euro
Macropodidae
H.graze
18200
18.6
�0.8
g3
ee
26.0
�2.0
g3
(Freudenberger&
Hume1992)
Macrop
usrobustusrobustus,
common
wallaro
o
Macropodidae
H.graze
20125
19.3
�0.4
e,g
4e
e30.1
�1.3
e,g
4(D
ellow
1982;
Freudenberger&
Hume1992)
Macrop
usrufus,
redkangaro
oMacropodidae
H.graze
25800
16.0
h2
ee
36.7
�2.5
i9
(Foot&
Romberg
1965;
McIntosh
1966;Forb
es&
Tribe1970;Munn&
Dawso
n2006)
Mean
17.1
[13.8,20.5]
31.8
[25.6,38.0]
(b)Myco
phagousmacropodoid
marsupial
Potorou
stridactylus,
long-n
ose
dpotoro
oPotoro
idae
M955
24.0
d2
ee
25.0
d2
(Wallis
1994)
Bettongiapen
icillata,woylieorbru
shtail
bettong
Potoro
idae
M1062
27.0
d2
ee
33.5
d2
(Wallis
1994)
Aep
yprymnusrufescen
s,ru
fousbettong
Potoro
idae
O2984
31.0
d2
ee
35.5
d2
(Wallis
1994)
Thylog
ale
thetis,red-n
eck
edpademelon
Macropodidae
H.bro
wse
4050
12.0
d,e
2e
e22.4
d,e
2(D
ellow
1982)
Wallabia
bicolor,sw
ampwallaby
Macropodidae
H.bro
wse
10000e21000
ee
31.0
f1
ee
This
study
Mean
24.3
[14.4,34.2]
29.2
[19.1,39.2]
(c)Oth
ermyco
phagousmammal
Perom
yscu
smaniculatus,
deermouse
Muro
idea
O15e19
ee
12.0
a1
10.3
1(C
ork
&Kenagy1989b)
Microtuspen
nsylvanicus,
meadow
vole
Muro
idea
H30e40
ee
ee
6.6
b2
(YoungOwl&
Batzli1998)
Uromys
caudim
acu
latus,
giantwhite-
tailedrat
Muro
idea
OLargeparticle
660
45.4
148.4
c1
55.5
1(C
omport
&Hume1998)
Eutamiasamoenus,
yellow-p
inech
ipmunk
Sciuridae
OLargeparticle
62
12.7
1e
e14.1
1(H
umeet
al.1993)
Spermop
hilussa
turatus,
golden-m
antled
gro
undsq
uirrel
Sciuridae
M216e327
ee
24.8
a1
24.1
1(C
ork
&Kenagy1989b)
Marm
otacaliga
ta,hoary
marm
ot
Sciuridae
HLargeparticle
2308
24.8
1e
e28.9
1(H
umeet
al.1993)
Isoodon
macrou
rus,
northern
bro
wn
bandicoot
Peramelidae
OFluid
250e3000
27.4
2e
e18.5
2(M
cClellandet
al.1999)
Perameles
nasu
ta,long-n
ose
dbandicoot
Peramelidae
OFluid
800e1000
28.4
d2
ee
17.5
d2
(Moyle
etal.1995)
Trichosurusvulpecula,co
mmonbru
shtail
possum
Phalangeridae
H.foliage
2350
47.4
�5.5
d4
ee
49.1
�9.5
d3
(Wellard
&Hume1981;
Foley&
Hume1987;
Sakaguch
i&
Hume1990)
204 M.A. Danks

Author's personal copy
Relatively little is known about swamp wallaby digestive
physiology, although some generalisations can bemade based
upon comparative gut morphology and diet habit. Gut
morphology is most similar to that of smaller wallabies; fore-
stomach and caecum (hindgut) structure are similar to that of
the tammar wallaby, a small (5 kg) grazer, and the red-necked
pademelon, a small (4 kg) browser (Dellow1979). The regionsof
the swamp wallaby gut are therefore expected to contribute
similarly to total stomach capacity (sacciform forestomach
30e51 %, tubiform forestomach 40e56 %, hindstomach
10e15 %; Dellow & Hume 1982) and the stomach to contribute
similarly to total gut capacity (76 %; Dellow&Hume 1982) as in
these two species and so account for similar proportions of
overall gut-retention times. The medium-sized, browsing
(low-fibre diet) swampwallaby has a forestomachmorphology
intermediate to the predominantly sacciform forestomach of
the small potoroids (low-fibre diets) and the predominantly
tubiform forestomach of the large grazing (high-fibre diets)
kangaroos (Stevens & Hume 1995). Osawa & Woodall (1992)
noted the similar gross anatomy of the swamp wallaby and
tammarwallaby intestinal tracts. The swampwallaby hindgut
(caecum) and large intestine are relatively small and simple
compared to the larger grazing kangaroos, most similar in
relative size and structure to that of the red-necked pademe-
lon, and consistent with a browsing, low-fibre diet (Osawa &
Woodall 1992). Osawa & Woodall (1992) also observed that
relative to body mass, the swamp wallaby small intestine is
longer while the large intestine is shorter than that of the red-
necked pademelon. Based on gut morphology and diet, reten-
tion timeoffluidsandsmall particles in the swampwallabygut
would be expected to be intermediate to the potoroids and the
large kangaroos, andmost similar to other browsingwallabies.
This prediction was not supported in the studied swamp
wallabies. The gut-retention time of truffle-like fungal spores
in the studied swamp wallabies is most similar to fluid gut-
retention times recorded for the smaller potoroids and per-
amelids despite great differences in body size and some
differences in gut morphology. The contribution of the small
intestine, although a small proportion of overall gut length,
might account for the longer than expected spore gut-
retention time in the study swamp wallabies.
Macropodoids exhibit differential digesta flow. Tubular
flow of digesta in the forestomach causes fluid and small
particle digesta to be expressed through the large particle
digesta, which remain longer in the forestomach formicrobial
fermentation resulting in longer overall gut-retention time of
large particles (Dellow 1982). The extent of digesta phase
separation is greater in the large kangaroos than in the small
wallabies and the potoroids (Dellow 1982; Hume 1989; Wallis
1994). In the larger kangaroos, differences between fluid and
large particle digesta range between 7 and 13 hr, compared to
10 hr in the small wallabies and 1e6 hr in the potoroids. In the
potoroid gut, fluid and particle digesta may pass through at
similar rates (Wallis 1994), although earlier radiographic
studies of digesta flow observed digesta phase separation
(Hume & Carlisle 1985; Richardson 1989). Of the peramelids
(hindgut fermenterswith a relatively simple forestomach), the
northern brown bandicoot Isoodon macrourus (McClelland et al.
1999) and the long-nosed bandicoot Perameles nasuta (Moyle
et al. 1995) selectively retain fluid and small particle digesta
Mean
33.4
[21.8,44.9]
26.9
[12.5,39.7]
Meanmyco
phagous
26.9
[20.0,33.7]
26.5
[17.7,35.3]
Meanmacropodoid
21.0
[16.4,25.5]
31.0
[26.5,35.3]
MeanMRT(�
1SE,w
here
n>3),weightedbysa
mple
size
,calculatedforsp
eciesacross
experimentald
iets.G
roupMRT[lower95%
confidence
limit,u
pper95%
confidence
limit],weightedbynumberof
sources.
MRTca
lculatedasreco
mmendedbyW
arn
er(1981b),exce
ptwhere
indicated.Bodymass:mean,orrangeifneithermeannorraw
valuesreported,ofstudyanim
als.
Tro
phic:H¼herb
ivore;O¼omnivore;M
¼primarily
myco
phagous.
n¼sa
mple
size
(numberofMRTestim
ates).
aMacrofu
ngalsp
ore
mark
er:
Elaphom
yces
granulatus.
bEstim
atedvisuallyfrom
graphedresu
lts.
cMacrofu
ngalsp
ore
mark
er:
Pisolithussp
.
dLargeparticle
mark
er:
Ru-P
(Dellow
1982;W
ellard
&Hume1981;Foley&
Hume1987;W
allis
1994).
eDellow
(1982):measu
redasmean50%
ET.MRTca
lculatedusingth
emeth
odofFaichney(1975)yieldedsimilarresu
lts.
fMacrofu
ngalsp
ore
mark
er:
Austroga
utieria
clelandii.
gCalculatedusingth
emeth
odofFaichney(1975).
hMunn&
Dawso
n(2006)only.
iFoot&
Romberg
(1965);McIntosh
(1966);Forb
es&
Tribe(1970):MRTca
lculatedusingth
emeth
odofCastle
(1956).
Gut-retention time in mycophagous mammals 205
Author's personal copy
in the hindgut while greater bilbies Macrotis lagotis do not
(Gibson & Hume 2000). Some (but not all) of the rodents also
selectively retain fluid and small particles in the hindgut,
enabling these small animals tomaintain condition on a high-
fibre diet through retention of microbes and potentially more
fermentable small particles for more complete digestion
(Hume et al. 1993; Pei et al. 2001). The present study did not
examine gut-passage of large particles and so no conclusion
can be drawn about the separation of digesta phases in the
swampwallaby, although based on comparative gut structure,
phase separation would be expected to be intermediate to the
large kangaroos and the smaller wallabies and potoroids.
Therefore, based on the small particle gut-retention time
recorded in the present study, large particle gut-retention
times of around 41 hr (31 hr spore MRT plus expected 10 hr
mean delay to large particle excretion) might be expected in
the studied swamp wallabies. Spores attached to sporocarp
fragments may pass through the gut with the large particle
phase (Comport & Hume 1998), resulting in spore gut-
retention times longer than for unattached spores. While the
extent to which swamp wallabies orally process sporocarps
prior to swallowing is not known, it is possible that naturally
ingested, whole sporocarps are not masticated and are
swallowed as fragments rather than as slurry. Because
wallabies in the present study were fed spores in a slurry,
estimates of spore gut-retention time in the studied wallabies
are considered conservative.
Manymammals are able to adapt to varying diet quality by
increasing gut capacity in response to more fibrous (lower
quality) diet, slowing the rate of passage of digesta to enhance
digestive efficiency, including bandicoots (Moyle et al. 1995)
and murid rodents (Gross et al. 1985; Hammond & Wunder
1991; Lee & Houston 1993). Rodents are also known to vary
gut capacity in response to changes in food intake (Hammond
&Wunder 1991). Similar responses to diet variation have been
observed in macropodids. In tammar wallabies the length of
the intestine and caecum (hindgut) increases in response to
a more fibrous diet, possibly to make secondary fermentation
in the hindgut feasible (Lentle et al. 2004). Osawa & Woodall
(1990) found that in swamp wallabies the length of the intes-
tinal villi increased, and therefore small intestinal surface
area increased relative to intestine size, with seasonal
increases in dietary fibre intake. The swamp wallabies in the
present study may have been consuming a higher-fibre diet
than usual, or higher-fibre relative to other browsing
wallabies, and concomitant changes in small intestine
morphology could have resulted in the longer than expected
gut-retention times observed. Further information on swamp
wallaby digestive anatomy and physiology, and changes with
diet, would be valuable in assessing the time taken for mac-
rofungal spores to pass through the gut of the swampwallaby.
The gut-retention times observed in swampwallabies in the
present study could also be due to different methodology,
including activity level, diet quality, feed intake and particle
size, compared to other studies. In the present study, animals
were able to select from food items including native grasses,
browse, macrofungal sporocarps and supplementary feeds
(lucerne hay, pelleted feed and apple), while in most studies
animals were fed experimental diets consisting primarily of
pelleted feed. Pelleted or milled feed can reduce particle and
fluid gut-retention times in sheep (Fadlalla et al. 1987) and
wallaroos (Freudenberger&Hume1992) because particle size at
ingestion is already sufficiently small for digestion. Although I
didnotassessuptakeof thesupplementary feedoffered relative
to natural food items in the present study, little supplementary
feed appeared to be taken and so the requirement for masti-
cation of natural food items may have contributed to a longer
digestion time. Conversely, the marker spores were fed in
a slurry ofmacrofungal sporocarp to the back of themouth and
would not have required mastication. Possibly, the sporocarp
slurry did not begin to pass through the gut until a subsequent
bout of feeding brought in other digesta.
Mycophagous vs non-mycophagous macropodoidmarsupials
Macropodoid species known to consume sporocarps (whether
stronglymycophagousornot), and forwhichgut-retentiondata
were available, retained fluid and small particles longer in the
gut than non-mycophagousmacropodoids, although statistical
support for this effect was weak. A larger sample size would
help clarify this comparison. Differences in gut-retention time
might reflect the different body size, diet quality and gut
Fig 2 e (A) Proportion of marker spores excreted by swamp
wallabies after oral dosing. (B) Cumulative proportion of
marker spores excreted by swamp wallabies after oral
dosing. Circles[ 21 kg male, triangles[ 10 kg male.
206 M.A. Danks

Author's personal copy
morphology ofmycophagousmacropodoids. Species known to
bemycophagous tend to be smaller, consume less fibrous diets
and have a sacciform forestomach-dominated gut structure, as
compared to non-mycophagous macropodoids which tend to
be larger, have a specialised grazing (fibrous) diet, and a tubi-
form forestomach-dominated gut structure (Freudenberger
et al. 1989). These features of mycophagous macropodoid
anatomy and feeding strategy enable utilisation of putative
low-quality food items such as macrofungal sporocarps. Addi-
tionally, these species occur predominantly in the mesic
eucalypt-dominated (EM) forests andwoodlands of Australia in
whichEMsporocarps, particularly truffle-like fungi, areahighly
diverse and abundant food resource (Claridge & May 1994;
Johnson 1994a; Allen et al. 1995; Bougher 1995; Claridge et al.
2000; Bougher & Lebel 2001). A diversity of fungi is probably
important for a balanced fungal diet (Claridge & Trappe 2005).
Macrofungal sporocarps have been considered a low-
quality food item because much of the nutrient content is in
indigestible forms or associated with cell walls (Cork & Foley
1990) requiring special gut adaptations, such as microbial
fermentation, for nutrient absorption. However, foregut
fermentation, as compared to hindgut fermentation, is
thought to improve the nutritional value of fungal sporocarps
for mammals by enabling better digestion of complex amino
acids through microbial fermentation in the forestomach
(Kinnear et al. 1979; Claridge&Cork 1994; Johnson 1994b). Most
foregut fermenting potoroids rely heavily on sporocarps, and
some macropodids, including wallabies and pademelons,
consistently consume sporocarps as part of amore varied diet.
The swamp wallaby, as a foregut fermenter, probably derives
nutrition from macrofungal sporocarps to supplement the
plant browse component of its diet, and retaining sporocarp
digesta in the gut for longer may enhance absorption of
nutrients from this relatively indigestible food item.
Spore gut-passage in other mycophagous mammals
Hindgut fermentingmammals also regularly utilise fungi, and
some, such as the northern flying squirrel (Glaucomys sabrinus)
(Maser et al. 1985; Vernes et al. 2004), rely heavily on macro-
fungal sporocarps, despite their apparently limited ability to
digest the ‘unavailable’ nutrients in these foods (Cork &
Kenagy 1989a; Bozinovic & Mu~noz-Pedreros 1995; D’Alva
et al. 2007). Among the Australian hindgut fermenters, all
weighing less than 3 kg, some (e.g. bush rat, Tory et al. 1997)
rely seasonally upon sporocarps and many, including other
rodents, peramelids and possums, consume sporocarps
regularly or occasionally (Claridge & May 1994; Claridge et al.
1996). Some of these animals have long gut-retention times,
for example the common brushtail possum and the giant
white-tailed rat. The giant white-tailed rat retains large
particulate digesta in an enlarged fundic diverticulum, slow-
ing the rate of passage of particulate digesta and increasing
absorption of nutrients (Comport & Hume 1998). Physiological
and morphological adaptations of the hindgut such as these
improve hindgut fermentation capabilities and consequently
enhance utilisation of otherwise low-quality food items such
as macrofungal sporocarps (Claridge et al. 1999).
Mycophagous mammals perform an important spore
dispersal role particularly for truffle-like fungi, which rely
upon such vectors for longer-distance spore dispersal.
Weighted average of available fluid and particle gut-retention
time estimates for mycophagous mammal species sympatric
with the swamp wallaby range from 12 hr (red-necked pade-
melon), to more than 2 d (brushtail possum). A community of
mammals with varying foraging habits, microhabitat use
patterns, home range size, and gut-retention times would
result in dispersal of spores of the truffle-like sporocarp
community to a variety of microhabitats and distances from
points of ingestion. Spore dispersal at multiple scales is likely
to be vital in maintaining macrofungal community diversity,
populations and individuals.
Mean gut-retention time of macrofungal spores alone does
not characterise an animal’s spore dispersal role but it is
a useful measure of the time after ingestion at which most
ingested spores are deposited in faeces; so it is a useful metric
when assessing spore dispersal by mycophagous mammals.
Variance in gut-retention time may also influence spore
dispersal patterns. Combining information on spore retention
with movement patterns, distance travelled, or area covered
within the average gut-retention time is also important, and
would enable relative rates of macrofungal spore dispersal by
mycophagous mammals to be examined. Further studies of
spore gut-passage in wild swamp wallabies and other
mycophagous mammals under semi-natural conditions are
necessary to obtain statistically robust and ecologically
meaningful measures of spore gut-retention time. Additional
information on the diversity and amounts of sporocarps
eaten, defecation patterns, and the viability of spores on
passage through the gut of mycophagous mammals with
varying digestive physiology would also be valuable in
building a picture of spore dispersal within a community of
mycophagous mammals.
Acknowledgements
This work was funded by a Hermon Slade Foundation grant
(HSF08-6, to K. Vernes, T. Lebel, and the author) and an Austra-
lian Research Council Discovery Grant (DP0557022) to K. Vernes.
TheauthorwassupportedbyanAustralianPostgraduateAward.
Research was conducted under University of New England
Animal Ethics Committee approval (AEC09-023), andNewSouth
Wales National Parks and Wildlife Service scientific license
(S12493). University of New England provided facilities and
access to Newholme Field Station. I thank S. Steinhart for
assistance inthefield,andK.Vernes,T. Lebel,N.Andrew, J. Price,
and three anonymous reviewers for critical feedback and
constructive comments that greatly improved this manuscript.
r e f e r e n c e s
Allen E, Allen M, Helm D, Trappe J, Molina R, Rincon E, 1995.Patterns and regulation of mycorrhizal plant and fungaldiversity. Plant and Soil 170: 47e62.
Blaxter KL, McGraham NM, Wainman FW, 1956. Someobservations on the digestibility of food by sheep and onrelated problems. British Journal of Nutrition 10: 69e91.
Gut-retention time in mycophagous mammals 207

Author's personal copy
Bougher NL, 1995. Diversity of ectomycorrhizal fungi associatedwith eucalypts in Australia. In: Brundrett M, Dell B,Malajczuk N, Mingqin G (eds),Mycorrhizas for Plantation Forestryin Asia. ACIAR, Canberra, pp. 8e14.
Bougher NL, Lebel T, 2001. Sequestrate (truffle-like) fungi ofAustralia and New Zealand. Australian Systematic Botany 14:439e484.
Bozinovic F, Mu~noz-Pedreros A, 1995. Nutritional ecology anddigestive responses of an omnivorous-insectivorous rodent(Abrothrix longipilis) feeding on fungus. Physiological Zoology 68:474e489.
Bridie A, Hume I, Hill D, 1994. Digestive-tract function andenergy-requirements of the rufous hare-wallaby, Lagorchesteshirsutus. Australian Journal of Zoology 42: 761e774.
Brundrett MC, 1991. Mycorrhizas in natural ecosystems. Advancesin Ecological Research 21: 171e315.
Bruns T, 1995. Thoughts on the processes that maintain localspecies diversity of ectomycorrhizal fungi. Plant and Soil 170:63e73.
Bruns TD, Peay KG, Boynton PJ, Grubisha LC, Hynson NA,Nguyen NH, Rosenstock NP, 2009. Inoculum potential ofRhizopogon spores increases with time over the first 4 yr ofa 99-yr spore burial experiment. New Phytologist 181: 463e470.
Calaby JH, 1958. Studies on marsupial nutrition II. The rate ofpassage of food residues and digestibility of crude fibre andprotein by the quokka, Setonix brachyurus (Quoy & Gaimard).Australian Journal of Biological Sciences 11: 571e580.
Caldwell IR, Vernes K, Baerlocher F, 2005. The northern flyingsquirrel (Glaucomys sabrinus) as a vector for inoculation of redspruce (Picea rubens) seedlings with ectomycorrhizal fungi.Sydowia 57: 166e178.
Castle EJ, 1956. The rate of passage of foodstuffs through thealimentary tract of the goat. British Journal of Nutrition 10:15e23.
Claridge AW, Barry SC, Cork SJ, Trappe JM, 2000. Diversity andhabitat relationships of hypogeous fungi. II. Factorsinfluencing the occurrence and number of taxa. Biodiversityand Conservation 9: 175e199.
Claridge AW, Castellano MA, Trappe JM, 1996. Fungi as a foodresource for mammals in Australia. In: Mallett K,Grgurinovic C (eds), The Fungi of Australia, vol. 1B. AustralianBiological Resource Study, Canberra, pp. 239e267.
Claridge A, Cork S, 1994. Nutritional value of hypogeal fungalsporocarps for the long-nosed potoroo (Potorous tridactylus),a forest-dwelling mycophagous marsupial. Australian Journal ofZoology 42: 701e710.
Claridge AW, May TW, 1994. Mycophagy among Australianmammals. Australian Journal of Ecology 19: 251e275.
Claridge AW, Tanton MT, Seebeck JH, Cork SJ, Cunningham RB,1992. Establishment of ectomycorrhizae on the roots of twospecies of Eucalyptus from fungal spores contained in thefaeces of the long-nosed potoroo (Potorous tridactylus).Australian Journal of Ecology 17: 207e217.
Claridge AW, Trappe JM, 2005. Sporocarp mycophagy: nutritional,behavioral, evolutionary, and physiological aspects. In:Dighton J, White JF, Oudemans P (eds), The Fungal Community:Its Organization and Role in the Ecosystem. CRC Press, Boca Raton,pp. 599e611.
Claridge AW, Trappe JM, Claridge DL, 2001. Mycophagy by theswamp wallaby (Wallabia bicolor). Wildlife Research 28: 643e645.
Claridge AW, Trappe JM, Cork SJ, Claridge DL, 1999. Mycophagy bysmall mammals in the coniferous forests of North America:nutritional value of sporocarps of Rhizopogon vinicolor,a common hypogeous fungus. Journal of Comparative PhysiologyB Biochemical Systemic and Environmental Physiology 169:172e178.
Comport SS, Hume ID, 1998. Gut morphology and rate of passageof fungal spores through the gut of a tropical rodent, the giant
white-tailed rat (Uromys caudimaculatus). Australian Journal ofZoology 46: 461e471.
Cork SJ, Foley WJ, 1990. Nutritional quality of hypogeous fungi forsmall mammals. Proceedings Nutritional Society Australia 15: 168.
Cork SJ, Kenagy GJ, 1989a. Nutritional value of hypogeousfungus for a forest-dwelling ground squirrel. Ecology 70:577e586.
Cork SJ, Kenagy GJ, 1989b. Rates of gut passage and retention ofhypogeous fungal spores in two forest-dwelling rodents.Journal of Mammalogy 70: 512e519.
D’Alva T, Lara C, Estrada-Torres A, Castillo-Guevara C, 2007.Digestive responses of two omnivorous rodents (Peromyscusmaniculatus and P. alstoni) feeding on epigeous fungus (Russulaoccidentalis). Journal of Comparative Physiology B BiochemicalSystemic and Environmental Physiology 177: 707e712.
Deacon JW, Fleming LV, 1992. Interactions of ectomycorrhizalfungi. In: Allen MF (ed), Mycorrhizal Functioning: An IntegrativePlant-Fungal Process. Routledge, Chapman & Hall, New York,pp. 249e295.
Dellow D, 1979. Physiology of Digestion in the MacropodineMarsupials. The Department of Biochemistry and Nutrition.University of New England, Armidale.
Dellow D, 1982. Studies on the nutrition of macropodinemarsupials. 3. The flow of digesta through the stomach andintestine of macropodines and sheep. Australian Journal ofZoology 30: 751e765.
Dellow D, Hume I, 1982. Studies on the nutrition of macropodinemarsupials. 4. Digestion in the stomach and the intestine ofMacropus giganteus, Thylogale thetis and Macropus eugenii.Australian Journal of Zoology 30: 767e777.
Demment MW, Soest PJV, 1985. A nutritional explanation forbody-size patterns of ruminant and nonruminant herbivores.The American Naturalist 125: 641e672.
Di Stefano J, 2010. Effect of habitat type, sex and time of day onspace use by the swamp wallaby. In: Coulson G, Eldridge M(eds), Macropods: The Biology of Kangaroos, Wallabies and Rat-kangaroos. CSIRO Publishing, Collingwood, Victoria, Australia,pp. 187e196.
Di Stefano J, Coulson G, Greenfield A, Swan M, 2011. Resourceheterogeneity influences home range area in the swampwallaby Wallabia bicolor. Ecography 34: 187e196.
Di Stefano J, Moyle R, Coulson G, 2005. A soft-walled double-layered trap for capture of swamp wallabies Wallabia bicolor.Australian Mammalogy 27: 235e238.
Edwards GP, Ealey EHM, 1975. Aspects of the ecology of theSwamp Wallaby, Wallabia bicolor (Marsupialia: Macropodidae).Australian Mammalogy 1: 307e317.
Egan J, Doyle P, 1984. A comparison of particulate markers for theestimation of digesta flow from the abomasum of sheepoffered chopped oaten hay. Australian Journal of AgriculturalResearch 35: 279e291.
Fadlalla B, Kay RNB, Goodall ED, 1987. Effects of particle size ondigestion of hay by sheep. The Journal of Agricultural Science 109:551e561.
Faichney GJ, 1975. The use of markers to partition digestionwithin the gastro-intestinal tract of ruminants. In: McDonaldIW, Warner ACI (eds), Digestion and Metabolism in the Ruminant.University of New England Publishing Unit, Armidale, pp.277e291.
Faichney GJ, Griffiths DA, 1978. Behaviour of solute and particlemarkers in the stomach of sheep given a concentrate diet.British Journal of Nutrition 40: 71e82.
Fogel R, Trappe JM, 1978. Fungus consumption (mycophagy) bysmall animals. Northwest Science 52: 1e31.
Forbes D, Tribe D, 1970. The utilization of roughages by sheep andkangaroos. Australian Journal of Zoology 18: 247e256.
Foley WJ, Hume ID, 1987. Passage of digesta markers in twospecies of arboreal folivorous marsupials: the greater glider
208 M.A. Danks

Author's personal copy
(Petauroides volans) and the brushtail possum (Trichosurusvulpecula). Physiological Zoology 60: 103e113.
Foot J, Romberg B, 1965. The utilization of roughage by sheep andthe red kangaroo, Macropus rufus (Desmarest). AustralianJournal of Agricultural Research 16: 429e435.
Freudenberger D, Hume I, 1992. Ingestive and digestive responsesto dietary fiber and nitrogen by 2 macropodid marsupials(Macropus robustus robustus andM. r. erubescens) and a ruminant(Capra hircus). Australian Journal of Zoology 40: 181e194.
Freudenberger DO, Wallis IR, Hume ID, 1989. Digestiveadaptations of kangaroos, wallabies and rat-kangaroos. In:Grigg G, Jarman P, Hume ID (eds), Kangaroos, Wallabies and Rat-kangaroos. Surrey Beatty & Sons, Chipping Norton, Australia,pp. 179e187.
Gannon WL, Sikes RS, Animal Care and Use Committee of theAmerican Society of Mammalogists, 2007. Guidelines of theAmerican Society of Mammalogists for the use of wildmammals in research. Journal of Mammalogy 88: 809e823.
Gibson LA, Hume ID, 2000. Digestive performance and digestapassage in the omnivorous greater bilby, Macrotis lagotis(Marsupialia: Peramelidae). Journal of Comparative Physiology BBiochemical Systemic and Environmental Physiology 170: 457e467.
Gross JE, Wang Z, Wunder BA, 1985. Effects of food quality andenergy needs: changes in gut morphology and capacity ofMicrotus ochrogaster. Journal of Mammalogy 66: 661e667.
Hammond KA, Wunder BA, 1991. The role of diet quality andenergy need in the nutritional ecology of a small herbivore,Microtus ochrogaster. Physiological Zoology 64: 541e567.
Hume ID, 1982. Digestive Physiology and Nutrition of Marsupials.University Press, Cambridge.
Hume ID, 1984. Microbial fermentation in herbivorousmarsupials. Bioscience 34: 435e440.
Hume ID, 1989. Optimal digestive strategies in mammalianherbivores. Physiological Zoology 62: 1145e1163.
Hume I, Carlisle C, 1985. Radiographic studies on the structureand function of the gastrointestinal tract of two species ofpotoroine marsupials. Australian Journal of Zoology 33: 641e654.
Hume ID, Morgan KR, Kenagy GJ, 1993. Digesta retention anddigestive performance in sciurid and microtine rodents:effects of hindgut morphology and body size. PhysiologicalZoology 666: 396e411.
Jarman P, Vernes K, 2006. Wildlife. In: Atkinson A, Ryan JS,Davidson I, Piper A (eds), High Lean Country. Land, People andMemory in New England. Allen & Unwin, Crows Nest, NSW, pp.44e56.
Johnson C, 1994a. Fruiting of hypogeous fungi in dry sclerophyllforest in Tasmania, Australia: seasonal variation and annualproduction. Mycological Research 98: 1173e1182.
Johnson CN, 1994b. Nutritional ecology of a mycophagousmarsupial in relation to production of hypogeous fungi.Ecology 75: 2015e2021.
Johnson CN, 1996. Interactions between mammals andectomycorrhizal fungi. Trends in Ecology & Evolution 11:503e507.
Kinnear JE, Cockson A, Christensen P, Main AR, 1979. Thenutritional biology of the ruminants and ruminant-likemammals e a new approach. Comparative Biochemistry andPhysiology Part A Comparative Physiology 64: 357e365.
Kotter MM, Farentinos RC, 1984. Formation of ponderosa pineectomycorrhizae after inoculation with feces of tassel-earedsquirrels. Mycologia 76: 758e760.
Lamont BB, Ralph CS, Christensen PES, 1985. Mycophagousmarsupials as dispersal agents for ectomycorrhizal fungi onEucalyptus calophylla and Gastrolobium bilobum. New Phytologist101: 651e656.
Langer P, 1980. Anatomy of the stomach in three species ofPotoroinae (Marsupialia: Macropodidae). Australian Journal ofZoology 28: 19e31.
Langer P, Dellow D, Hume I, 1980. Stomach structure and functionin three species of macropodine marsupials. Australian Journalof Zoology 28: 1e18.
Lee WB, Houston DC, 1993. The role of coprophagy in digestion involes (Microtus agrestis and Clethrionomys glareolus). FunctionalEcology 7: 427e432.
Lentle R, Stafford K, Hume I, 2004. A comparison of the grossgastrointestinal morphology of genetically similar tammarwallabies (Macropus eugenii) from different nutritionalenvironments. Australian Journal of Zoology 52: 437e446.
MaserC,ClaridgeAW,Trappe JM (eds), 2008. Trees, Truffles, andBeasts:How Forests Function. Rutgers University Press, New Brunswick.
Maser Z, Maser C, Trappe JM, 1985. Food habits of the northernflying squirrel (Glaucomys sabrinus) in Oregon. Canadian Journalof Zoology 63: 1084e1088.
McClelland KL, Hume ID, Soran N, 1999. Responses of thedigestive tract of the omnivorous northern brown bandicoot,Isoodon macrourus (Marsupialia: Peramelidae), to plant- andinsect-containing diets. Journal of Comparative Physiology BBiochemical Systemic and Environmental Physiology 169: 411e418.
McIntosh D, 1966. The digestibility of two roughages and the ratesof passages of their residues by the red kangaroo, Megaleia rufa(Desmarest), and the merino sheep. CSIRO Wildlife Research 11:125e135.
Moyle DI, Hume ID, Hill DM, 1995. Digestive performance andselective digesta retention in the long-nosed bandicoot,Perameles nasuta, a small omnivorous marsupial. Journal ofComparative Physiology B Biochemical Systemic and EnvironmentalPhysiology 164: 552e560.
Munn AJ, Dawson TJ, 2006. Forage fibre digestion, rates of feedpassage and gut fill in juvenile and adult red kangaroosMacropus rufus Desmarest: why body size matters. Journal ofExperimental Biology 209: 1535e1547.
Osawa R, Woodall P, 1990. Feeding strategies of the swampwallaby, Wallabia bicolor, on North Stradbroke Island,Queensland .2. Effects of seasonal changes in diet quality onintestinal morphology. Australian Wildlife Research 17: 623e632.
Osawa R, Woodall P, 1992. A comparative study of macroscopicand microscopic dimensions of the intestine in fivemacropods (Marsupialia, Macropodidae).2. Relationship withfeeding habits and fiber content of the diet. Australian Journalof Zoology 40: 99e113.
Pei YX, Wang DH, Hume ID, 2001. Selective digesta retention andcoprophagy in Brandt’s vole (Microtus brandti). Journal ofComparative Physiology B Biochemical Systemic and EnvironmentalPhysiology 171: 457e464.
Perry DA, Amaranthus MP, Borchers JG, Borchers SL, Brainerd RE,1989. Bootstrapping in ecosystems. Bioscience 39: 230e237.
Read DJ, 1991. Mycorrhizas in ecosystems. Experientia 47: 376e391.Reddell P, Spain AV, Hopkins M, 1997. Dispersal of spores of
mycorrhizal fungi in scats of native mammals in tropicalforests of northeastern Australia. Biotropica 29: 184e192.
Richardson KC, 1989. Radiographic studies on the form andfunction of the gastrointestinal tract of the Woylie (Bettongiapenicillata). In: Grigg G, Jarman P, Hume ID (eds), Kangaroos,Wallabies and Rat-Kangaroos. Surrey Beatty & Sons, ChippingNorton, Australia, pp. 205e215.
Sakaguchi E, Hume ID, 1990. Digesta retention and fibre digestionin brushtail possums, ringtail possums and rabbits.Comparative Biochemistry and Physiology, Part A: Molecular &Integrative Physiology 96A: 351e354.
Stevens CE, Hume ID, 1995. Comparative Physiology of the VertebrateDigestive System. Cambridge University Press, Cambridge.
Tory MK, May TW, Keane PJ, Bennett AF, 1997. Mycophagy in smallmammals: a comparison of the occurrence and diversity ofhypogeal fungi in the diet of the long-nosed potoroo Potoroustridactylus and the bush rat Rattus fuscipes from southwesternVictoria, Australia. Australian Journal of Ecology 22: 460e470.
Gut-retention time in mycophagous mammals 209

Author's personal copy
Troy S, Coulson G, 1993. Home range of the swamp wallaby,Wallabia bicolor. Wildlife Research 20: 571e577.
Ud�en P, Colucci PE, Van Soest PJ, 1980. Investigation of chromium,cerium and cobalt as markers in digesta. Rate of passagestudies. Journal of the Science of Food and Agriculture 31: 625e632.
Vernes K, 2010. Mycophagy in a community of macropod species.In: Coulson G, Eldridge M (eds), Macropods: The Biology ofKangaroos, Wallabies and Rat-Kangaroos. CSIRO Publishing,Collingwood, Victoria, Australia, pp. 155e169.
Vernes K, Blois S, Baerlocher F, 2004. Seasonal and yearly changesin consumption of hypogeous fungi by northern flyingsquirrels and red squirrels in old-growth forest, NewBrunswick. Canadian Journal of Zoology 82: 110e117.
Vernes K, McGrath K, 2009. Are introduced black rats (Rattusrattus) a functional replacement for mycophagous nativerodents in fragmented forests? Fungal Ecology 2: 145e149.
Wallis IR, 1994. The rate of passage of digesta through thegastrointestinal tracts of potoroine marsupials: more evidenceabout the role of the potoroine foregut. Physiological Zoology 67:771e795.
Warner A, 1981a. The mean retention times of digesta markers inthe gut of the tammar, Macropus eugenii. Australian Journal ofZoology 29: 759e771.
Warner ACI, 1981b. The rate of passage of digesta through the gutof mammals and birds. Nutrition Abstracts and Reviews Series B51: 789e820.
Wellard G, Hume I, 1981. Digestion and digesta passage in thebrushtail possum, Trichosurus vulpecula (Kerr). AustralianJournal of Zoology 29: 157e166.
Young Owl M, Batzli GO, 1998. The integrated processingresponse of voles to fibre content of natural diets. FunctionalEcology 12: 4e13.
210 M.A. Danks




















