
Growth of bell pepper plants (Capsicum annuum) affected by coloured covers
9
1 23 Gesunde Pflanzen Pflanzenschutz - Verbraucherschutz - Umweltschutz ISSN 0367-4223 Volume 66 Number 4 Gesunde Pflanzen (2014) 66:149-155 DOI 10.1007/s10343-014-0328-7 Growth of bell pepper plants (Capsicum annuum) affected by coloured covers Fánor Casierra-Posada, Y. A. Matallana- Díaz & E. Zapata-Casierra
Transcript of Growth of bell pepper plants (Capsicum annuum) affected by coloured covers

1 23
Gesunde PflanzenPflanzenschutz - Verbraucherschutz -Umweltschutz ISSN 0367-4223Volume 66Number 4 Gesunde Pflanzen (2014) 66:149-155DOI 10.1007/s10343-014-0328-7
Growth of bell pepper plants (Capsicumannuum) affected by coloured covers
Fánor Casierra-Posada, Y. A. Matallana-Díaz & E. Zapata-Casierra

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

1 3
Original article
Received: 10 March 2014 / Accepted: 15 September 2014 / Published online: 9 October 2014© Springer-Verlag Berlin Heidelberg
Growth of bell pepper plants (Capsicum annuum) affected by coloured covers
Fánor Casierra-Posada · Y. A. Matallana-Díaz · E. Zapata-Casierra
Gesunde Pflanzen (2014) 66:149–155DOI 10.1007/s10343-014-0328-7
Keywords Light quality · Spectral quality · Monochromatic filters · Coloured films · Photomorphogenesis
Das Wachstum von Gemüsepaprikapflanzen (Capsicum annuum), beeinflusst durch farbige Folienbedeckungen
Zusammenfassung Während die Pflanze das gesam-te Spektrum des sichtbaren Lichtes nutzt, weisen einige Lichtfarben grössere Bedeutungen als andere auf. Das Ziel dieser Untersuchung war es, die Reaktionen der Gemüse-paprikapflanzen, die unter farbigen Abdeckungen wuch-sen, zu evaluieren. Paprikapflanzen (Capsicum annuum) in Hydrokultur wurden in einem Gewächshaus in Tunja (Kolumbien) verschiedenen Lichtintensitäten ausgesetzt, welche durch Abdeckung mit Polypropylen-Folien (gelb, grün, blau, transparent, rot, und ohne Abdeckung [Kont-rolle]), erzeugt wurden. Herkömmliche Wachstumsindizes der Pflanzen wurden gemessen oder berechnet. Alle ausge-werteten Parameter wurden in unterschiedlicher Weise von der Farbe der Abdeckungen beeinflusst, sodass z. B. die Blattfläche unter den grünen und blauen Abdeckungen am höchsten war und auch die Pflanzen unter der blauen Fo-lie die grösste Wassermenge aufnahmen. Paprikapflanzen unter den gelben Abdeckungen zeigten eine höhere Was-sernutzungseffizienz als die anderen Behandlungen, wäh-rend der Chlorophyllgehaltsindex unter den blauen, grünen und transparenten Abdeckungen höher war. Die Ergebnisse zeigen, dass nicht eine bestimmte farbige Abdeckung die meisten der gemessenen Wachstumsparameter begünstigt, jedoch kann je nach den Bedürfnissen der Erzeuger eine bestimmte Abdeckung verwendet werden, um einen ge-wünschten Effekt zu erzielen.
Abstract While a plant utilizes the full spectrum of visible light, some colours of light have more important applica-tions than others. The aim of this study was to evaluate the responses of bell pepper plants grown under coloured covers. Bell pepper plants (Capsicum annuum) were grown hydroponically in a greenhouse in Tunja, Colombia, un-der different light quality regimes that were obtained with polypropylene films (yellow, green, blue, transparent, red, and a control without a plastic film cover). Conventional growth indices were calculated or measured as response variables. All of the evaluated parameters were influenced in a different ways by the colour of the covers, e.g. the leaf area was higher under the green and blue covers. The highest water uptake was found in the plants grown un-der the blue film. Plants under the yellow cover presented higher water use efficiency than the other treatments. The chlorophyll content index was higher under the blue, green and transparent covers. Based on these results, a coloured cover that favors most of the growth indices of plants can-not be selected; however, according to the needs of grow-ers, the supplemental light quality can be used to achieve a specific effect.
F. Casierra-Posada () · Y. A. Matallana-DíazFaculty of Agricultural Sciences, Research Group in Plant Ecophysiology, Pedagogical and Technological University of Colombia – UPTC,Tunja, Colombiae-mail: [email protected]
E. Zapata-CasierraFaculty of Industrial Engineering, Technological University of Pereira,Pereira, Colombia
Author's personal copy

150
1 3
F. Casierra-Posada et al.
Schlüsselwörter Lichtqualität · Spektrale Qualität · Monochromatische Filter · Farbfolien · Photomorphogenese
Introduction
Plants have sophisticated photosensitive mechanisms to capture light energy for photosynthesis; nevertheless, the light environment also strongly influences plant growth and development. Such responses, collectively known as photo-morphogenesis, modulate the entire life cycle of plants. To increase their fitness, plants sense ambient light conditions and modulate their developmental processes accordingly by utilizing an array of photoreceptors such as phytochromes, cryptochromes and phototropins (Jiao et al. 2007). Spectral light changes induce different morphogenetic and photosyn-thetic responses, which can vary among plant species. Such photoresponses are of practical importance in recent plant cultivation technologies because the feasibility of purpose-fully tailoring illumination spectra enables one to control plant growth, development, and nutritional quality (Lin et al. 2013). Some of these physiological responses are spe-cifically triggered by a single photoreceptor but often mul-tiple light sensors ensure a coordinated response (Kami et al. 2010).
There is increasing interest in the physiological responses of plants to variations in light quality. To obtain variations in light quality, diverse sources have been used, such as the exposure of plants to coloured films (Casierra-Posada et al. 2012a, b; Hückstädt et al. 2013), light-emitting diodes (LEDs) (Jao et al. 2005) and fluorescent lamps (Piszczek & Głowacka 2008). These illumination systems allow wave-lengths to be matched to plant photoreceptors to produce optimal yield and influence plant morphology and physi-ological responses (Li et al. 2013). Similarly, the results of these studies are both varied and contradictory; therefore, researchers have not yet agreed on a numerical parameter for correlating plant responses and light quality.
The combination of lighting sources, providing different wavelengths, was effective for plant growth and develop-ment. Spectrum, intensity, and duration of light can easily be controlled by growers in artificial growing environments and supplemental light quality can be used strategically to enhance the nutritional value and growth of plants (Lin et al. 2013). Altering the quality of light in a specified band spectrum can improve the quality of cut flowers (Casierra-Posada et al. 2012a) and promote the plant growth of leaf vegetables (Johkan et al. 2012; Kim et al. 2004a); however, in some cases, the use of alterations in light quality reduces plant growth and appears to have no practical benefits (Hückstädt et al. 2013).
The objective of this study was to investigate the effects of light quality for growing bell pepper plants under coloured polypropylene films. An optimal strategy of light quality regulation will contribute to the knowledge of the physiology of plants grown under controlled conditions and in designing growth chamber or greenhouse light environ-ments to obtain higher economic profits for growers.
Materials and Methods
The experiment was carried out in greenhouse conditions at the Pedagogical and Technological University of Colom-bia (UPTC) in Tunja, Colombia, located at the coordi-nates 5°33′16.25″N and 73°21′9.14″W and an altitude of 2790 m above sea level. Inside the greenhouse, the aver-age temperature was 16.9 °C, with 72.4 % relative humid-ity, and the photosynthetic photon flux density (PPFD) was 1380 μmol m−2 s−1, as determined with a Light Scout Quan-tum Meter 3415 (Spectrum Technologies, Inc. Plainfield, IL, USA). The Light reduction, PPFDs, and red/far red ratios (R/FR was measured at 660 ± 20/730 ± 15 nm) registered beneath the different covers are shown in Table 1. The R/FR ratio was determined with a Field Scout Red/Far red Meter 3412 (Spectrum Technologies, Inc. Plainfield, IL, USA). The planting material that was used consisted of seedlings of the bell pepper (Capsicum annuum) hybrid ‘Nathalie’ (Syn-genta®). The seedlings were placed in 4 L glass containers filled with a nutrient solution with the following composi-tion in mg L−1: nitrate nitrogen 40.3; ammonium nitrogen 4.0; phosphorus 20.4; potassium 50.6; calcium 28.8; mag-nesium 11.4; sulfur 1.0; iron 1.12; manganese 0.112; copper 0.012; zinc 0.0264; boron 0.106; molybdenum 1.2E−3, and cobalt 3.6E−4.
Ten plants per treatment were subjected to solar radia-tion filtered through 15 μm-thick polypropylene films of different colours: red, yellow, blue, green, and transparent. The control plants were grown in the greenhouse without any polypropylene covering. Frames were covered with polyethylene film of the different experimental colours and
Table 1 Characteristics of the light recorded under the coloured films under which the bell pepper plants (Capsicum annuum) grew.Cover colour Photosynthetic
photon flux densityLight reductiona R: FR ratiob
µmol m−b s−a %Control 1380 0.00 1.10Yellow 376 27.25 0.92Blue 486 35.22 0.92Green 452 32.78 0.86Red 578 41.88 1.06Transparent 622 45.07 1.05aCalculated in respect to the control treatmentbMeasured at 660 ± 20:730 ± 15 nm
Author's personal copy

151
1 3
Growth of bell pepper plants (Capsicum annuum) affected by coloured covers
exposed to green and red light, indicating a morphogenic response of the plants to light quality. In this response, phy-tochromes, blue-light receptors, and UV-B photoreceptors are involved (Jiao et al. 2007). For green light mediated responses in plants, Folta & Maruhnich (2007) proposed a zeaxanthin based compound as a green-light receptor; similarly, phytochromes and cryptochromes are green-light receptors. Therefore, as discussed by Macedo et al. (2011), the plants grown under green light were in some respects comparable to those grown in red light and, in other respects, similar to those grown in blue light, as was found in the current study for leaf area in plants exposed to blue and green light.
Regarding the LAR, similar to findings for leaf area, no significant difference between the control plants and those grown under the transparent cover was found, but, between these two treatments and other colour covers, there were dif-ferences, among which the treatment with green cover stood out, whose plants had the highest LAR value (Table 2). The average LAR of the plants exposed to this treatment was 94.3 % above the average of the control plants. Although green light has been thought to be of no use to plant growth, particularly for photomorphogenesis and photosynthesis, it may affect plant morphology and physiology, as well as leaf growth, stomatal conductance and early stem elongation (Folta, 2004; Kim et al. 2004a, b); on the other hand, high-intensity green LED light was effective in promoting plant growth and, in particular, short-wavelength green light was available for active plant growth of Lactuca sativa (Johkan et al. 2012).
Typically, the LAR increases with low light intensities (Paulilo et al. 2007), yet in the current study, the highest LAR average was seen in the plants under the green cover, higher than in the red and transparent covers, which induced more light reduction than the green film (Table 1); there-fore, presumably, the plants located under the green film were affected more by the light enriched in the green colour range of the incident spectrum than by light reduction, due to the fact that the exposure of plants to green light induces
fitted over the glass jars on tables in the greenhouse with each frame covering 10 plants at a height of 1 m above the plants. The greenhouse was equipped with pipes and hoses connected to an aeration system to oxygenate the nutrient solution of the plants growing in the glass containers. At the beginning of the experiment, the dry weight was measured in 20 seedlings for an initial value in the calculation of the growth indices, according to the methodology reported by Hunt (1990).
The experiment lasted 65 days; at the end of which, the dry weight of the different plant organs (dried in an 80 °C oven) was measured. The chlorophyll content index of the leaves was measured with a CCM-200 chlorophyll meter device (Opti-sciences, Inc. USA). The leaf area was deter-mined with a Li-cor 3000 A analyzer (LI-COR Biosciences, USA). With this information, it was possible to calculate the leaf area ratio (LAR), leaf weight ratio (LWR), specific leaf area, root to shoot ratio, absolute growth rate, and rela-tive growth rate, based on the procedures outlined by Hunt (1990). The dry matter allocation was expressed as the por-tion of dry matter determined in a particular organ in rela-tion to the dry mass of the whole plant. Every week, the amount of water missing from the containers was measured and taken as the water consumed by the plant. The water use efficiency (WUE) was calculated as the amount of dry mat-ter produced per liter of evapotranspired water.
The experiment had a totally-randomized one-factor design in which the plants were exposed to light filtered through coloured films. Each treatment consisted of 10 plants and the analyses were performed individually for each plant. In this way, each plant was taken as a repeti-tion (n = 10). The data were subjected to a classical variance analysis (P < 0.05) and a Tukey’s range test using the IBM-SPSS Statistics program, version 19.0.0 (2010; IBM Corpo-ration, N.Y.—USA).
Results and Discussion
No statistically significant differences were found for the variables: dry mass production per plant, specific leaf area (SLA), absolute growth rate (AGR), relative growth rate (RGR) and leaf weight ratio (LWR). However, statistical differences for other variables related to plant growth were found. In regards to leaf area, all of the average values of this variable measured in the plants exposed to coloured covers exceeded the means of the control plants (Table 2). The highest average value of this variable was presented by the plants placed under green and blue covers, which exceeded the average of the control plants by 118.9 and 112.0 % respectively.
Macedo et al. (2011) reported the smallest leaf area in Alternanthera brasiliana cultured in vitro for plantlets
Table 2 Effect of coloured covers on leafiness and plant-water rela-tionships in the bell pepper plants (Capsicum annuum) (n = 10)Cover colour
Leaf area* Leaf area ratio*
Water uptake*
Water use efficiency*
cm2 cm2 g−1 L g L−1
Control 324.4 ± 80.4 b 57.7 ± 19.9 c 4.3 ± 0.4 ab 1.2 ± 0.3 bYellow 617.9 ± 129.9 a 87.1 ± 18.2 b 3.5 ± 0.5 bc 2.0 ± 0.4 aBlue 689.8 ± 148.0 a 91.8 ± 11.2 ab 4.4 ± 0.5 a 1.6 ± 0.2 abGreen 710.2 ± 193.6 a 112.3 ± 21.0 a 3.9 ± 0.4 abc 1.6 ± 0.4 abRed 561.5 ± 176.0 a 76.6 ± 15.8 bc 4.3 ± 0.7 ab 1.6 ± 0.2 abTransparent 345.0 ± 126.7 b 60.3 ± 12.6 c 3.4 ± 0.8 c 1.6 ± 0.5 ab*(P < 0.01)Means in columns with different letters are significantly different from one another (Tukey s̓ HSD test, P < 0.01)
Author's personal copy
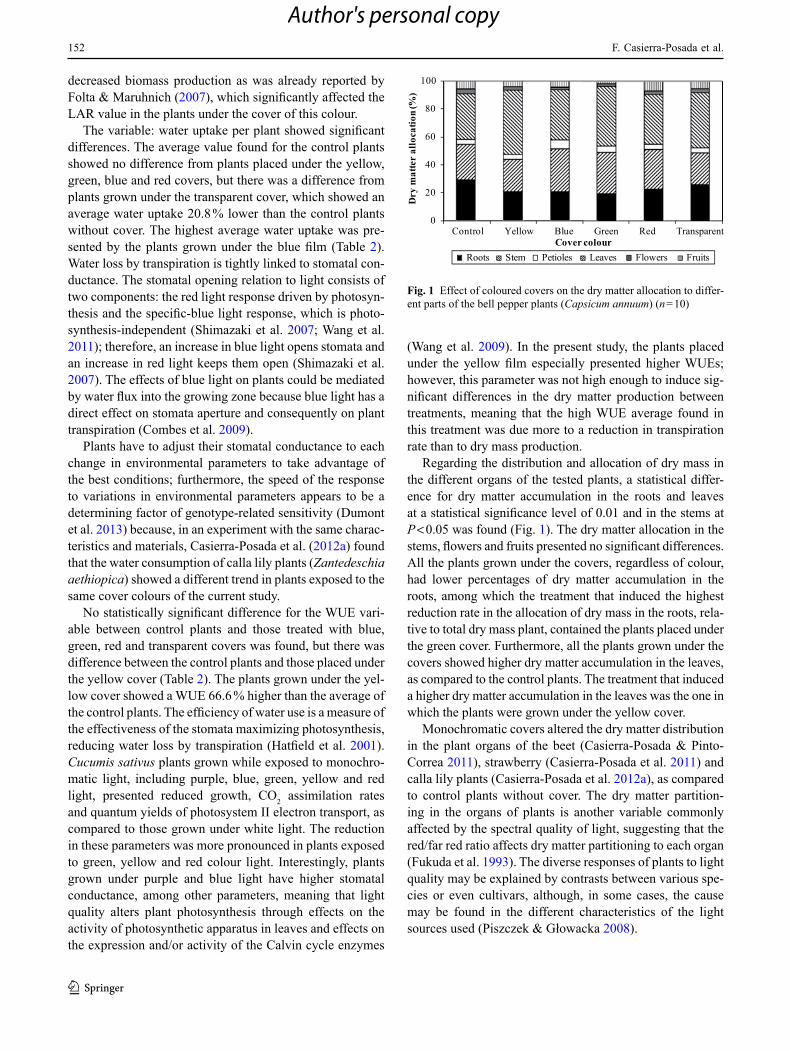
152
1 3
F. Casierra-Posada et al.
(Wang et al. 2009). In the present study, the plants placed under the yellow film especially presented higher WUEs; however, this parameter was not high enough to induce sig-nificant differences in the dry matter production between treatments, meaning that the high WUE average found in this treatment was due more to a reduction in transpiration rate than to dry mass production.
Regarding the distribution and allocation of dry mass in the different organs of the tested plants, a statistical differ-ence for dry matter accumulation in the roots and leaves at a statistical significance level of 0.01 and in the stems at P < 0.05 was found (Fig. 1). The dry matter allocation in the stems, flowers and fruits presented no significant differences. All the plants grown under the covers, regardless of colour, had lower percentages of dry matter accumulation in the roots, among which the treatment that induced the highest reduction rate in the allocation of dry mass in the roots, rela-tive to total dry mass plant, contained the plants placed under the green cover. Furthermore, all the plants grown under the covers showed higher dry matter accumulation in the leaves, as compared to the control plants. The treatment that induced a higher dry matter accumulation in the leaves was the one in which the plants were grown under the yellow cover.
Monochromatic covers altered the dry matter distribution in the plant organs of the beet (Casierra-Posada & Pinto-Correa 2011), strawberry (Casierra-Posada et al. 2011) and calla lily plants (Casierra-Posada et al. 2012a), as compared to control plants without cover. The dry matter partition-ing in the organs of plants is another variable commonly affected by the spectral quality of light, suggesting that the red/far red ratio affects dry matter partitioning to each organ (Fukuda et al. 1993). The diverse responses of plants to light quality may be explained by contrasts between various spe-cies or even cultivars, although, in some cases, the cause may be found in the different characteristics of the light sources used (Piszczek & Głowacka 2008).
decreased biomass production as was already reported by Folta & Maruhnich (2007), which significantly affected the LAR value in the plants under the cover of this colour.
The variable: water uptake per plant showed significant differences. The average value found for the control plants showed no difference from plants placed under the yellow, green, blue and red covers, but there was a difference from plants grown under the transparent cover, which showed an average water uptake 20.8 % lower than the control plants without cover. The highest average water uptake was pre-sented by the plants grown under the blue film (Table 2). Water loss by transpiration is tightly linked to stomatal con-ductance. The stomatal opening relation to light consists of two components: the red light response driven by photosyn-thesis and the specific-blue light response, which is photo-synthesis-independent (Shimazaki et al. 2007; Wang et al. 2011); therefore, an increase in blue light opens stomata and an increase in red light keeps them open (Shimazaki et al. 2007). The effects of blue light on plants could be mediated by water flux into the growing zone because blue light has a direct effect on stomata aperture and consequently on plant transpiration (Combes et al. 2009).
Plants have to adjust their stomatal conductance to each change in environmental parameters to take advantage of the best conditions; furthermore, the speed of the response to variations in environmental parameters appears to be a determining factor of genotype-related sensitivity (Dumont et al. 2013) because, in an experiment with the same charac-teristics and materials, Casierra-Posada et al. (2012a) found that the water consumption of calla lily plants (Zantedeschia aethiopica) showed a different trend in plants exposed to the same cover colours of the current study.
No statistically significant difference for the WUE vari-able between control plants and those treated with blue, green, red and transparent covers was found, but there was difference between the control plants and those placed under the yellow cover (Table 2). The plants grown under the yel-low cover showed a WUE 66.6 % higher than the average of the control plants. The efficiency of water use is a measure of the effectiveness of the stomata maximizing photosynthesis, reducing water loss by transpiration (Hatfield et al. 2001). Cucumis sativus plants grown while exposed to monochro-matic light, including purple, blue, green, yellow and red light, presented reduced growth, CO2 assimilation rates and quantum yields of photosystem II electron transport, as compared to those grown under white light. The reduction in these parameters was more pronounced in plants exposed to green, yellow and red colour light. Interestingly, plants grown under purple and blue light have higher stomatal conductance, among other parameters, meaning that light quality alters plant photosynthesis through effects on the activity of photosynthetic apparatus in leaves and effects on the expression and/or activity of the Calvin cycle enzymes
Fig. 1 Effect of coloured covers on the dry matter allocation to differ-ent parts of the bell pepper plants (Capsicum annuum) (n = 10)
Author's personal copy

153
1 3
Growth of bell pepper plants (Capsicum annuum) affected by coloured covers
highest root to shoot ratio when they were placed under a green cover in comparison to yellow, red, blue and transpar-ent films (Casierra-Posada et al. 2012b); while in the cur-rent study, the bell pepper plants presented the lowest root to shoot ratio in response to exposure to yellow, blue and green films, indicating a notable effect of light quality on the growth of the shoots and roots of treated plant species.
In regards to the chlorophyll content index (CCI), a statistically significant difference between treatments was found. In fact, all the plants grown under cover presented higher CCI values, as compared to the average in the control plants (Fig. 3). This increase ranged from 31.5 to 50.5 % higher than that presented by the uncovered plants (control). However, the most marked difference was found between the control plants and those that were under the blue, green and transparent covers, with increases of 49.1, 50.5 and 39.9 % beyond the control. Some studies have investigated the effects of monochromatic light on the pig-ment content in plants. In broccoli plants grown in vitro, Wilson et al. (1998) discovered an increased dry weight and chlorophyll content of plantlets grown under red light, while blue light reduced chlorophyll and led to elongated stems, but Jao et al. (2005) found a higher chlorophyll content in Zantedeschia plantlets exposed to red + blue light, showing that blue light may be involved in chloro-phyll development control. In Triticum aestivum, it was found that the formation of green pigments in the plants was inhibited by red light, but this was reversed when the plants were exposed to blue light or a mixture of blue and red light (Gupta & Tripathy 2010); on the other hand, in onion plants, exposure to red light increased chlorophyll content in comparison with plants exposed to blue light and controls without coloured light filters (Alam et al. 2007).
The manipulation of the light spectrum produces changes in the chlorophyll content of green plant tissues. However, studies with a more physiological focus demonstrate that the effects of light quality vary among different plant spe-cies, e.g., in Cucumis sativus plants, the chlorophyll content
Generally, the light quality had no significant effect on the photosynthetic rate per leaf or per unit dry weight, but rates of transpiration, stomatal conductance and water use efficiency increased in Eucalyptus grandis plants exposed to diverse red/far red ratios, indicating that compensatory changes in plant morphology and gas exchange caused equality in total dry weight per plant between treatments. These effects of light quality on dry matter partitioning and gas exchange had important effects on the morphology and physiology of the exposed plants (Hoad & Leakey 1994).
As for the root to shoot ratio, all the plants grown under covers, regardless of colour, presented averages below those found for the control plants which grew without cover. This reduction ranged from 18.7 to 44.7 %, as compared to the controls; however, a significant difference was only found between the control plants and the treatments with yel-low, blue and green films (Fig. 2). Hückstädt et al. (2013) reached similar results using coloured film covers on some herbs and attributed their finding to a reduction in PAR and to an increase in red/far red ratio due to the covers. In fact, in their experiment, the shoot of the plants was reduced as the red/far red ratio increased. Nevertheless, in our study, the low root to shoot rate observed in the plants grown under the yellow, blue and green covers can most likely be attrib-uted to the colour of the covers rather than to the reduced light or red/far red ratio because these cover colours induced both less light reduction and a low red/ far-red ratio between all the evaluated cover colours.
The relationship between root growth and whole plant growth is called allometry or relative growth (Fagaria et al. 2006) and it indicates the balance between the root and shoot components of a whole plant (Hunt 1990). A shift in root to shoot biomass allocation in response to external resource availability therefore allows plants to minimize an imbalance in any critical resource that is limiting growth (Grechi et al. 2007). The responses of plants to light quality differs among different plant materials (Piszczek & Głowacka 2008), e. g. strawberry plants grown under coloured films presented the
Fig. 2 Effect of coloured covers on the root to shoot ratio in the bell pepper plants (Capsicum annuum) (n = 10)
Fig. 3 Effect of coloured covers on the chlorophyll content index (CCI) in the bell pepper plants (Capsicum annuum) (n = 10)
Author's personal copy

154
1 3
F. Casierra-Posada et al.
Casierra-Posada F, Peña-Olmos JE, Ulrichs C (2012b) Basic growth analysis in strawberry plants (Fragaria sp.) exposed to different radiation environments. Agron Colomb 30(1):25–33
Casierra-Posada F, Peña-Olmos JE, Ulrichs C (2011) Crecimiento y eficiencia fotoquímica del fotosistema II en plantas de fresa (Fragaria sp.) afectadas por la calidad de la luz: implicaciones agronómicas. Rev UDCA Act & Div Cient 14(2):43–53
Casierra-Posada F, Pinto-Correa JR (2011) Crecimiento de plantas de remolacha (Beta vulgaris L. var. Crosby Egipcia) bajo coberturas de color. Rev Fac Nal Agr Medellín 64(2):6081–6091
Combes, D, Barillot R, Durand JL, Escobar-Gutiérrez AJ, Frak E (2009) Impending effects of transpiration in blue light regulation of leaf growth. Comp Biochem Phys A 153(2):S204
Dumont J, Spicher F, Montpied P, Dizengremel P, Jolivet Y, Le Thiec D (2013) Effects of ozone on stomatal responses to environmental parameters (blue light, red light, CO2 and vapour pressure deficit) in three Populus deltoides × Populus nigra genotypes. Environ Pollut 173:85–96
Facella P, Daddiego L, Giuliano G, Perrotta G (2012) Gibberellin and auxin influence the diurnal tran-scription pattern of photoreceptor genes via CRY1a in tomato. PLoS ONE 7:e30121
Fagaria NK, Baligar VC, Clark RB (2006) Root architecture. In: Physiology of crop production. The Haworth Press, Binghamton, pp. 23–59
Folta KM (2004) Green light stimulates early stem elongation, an-tagonizing light-mediated growth inhibition. Plant Physiol 135:1407–1416
Folta KM, Maruhnich SA (2007) Green light: a signal to slow down or stop. J Exp Bot 58:3099–3111
Fukuda N, Ikeda H, Nara M (1993) Effects of light quality on the growth of tomato and kidney bean cultured by hydroponics under controlled environment. J Soc Agri Struct—Japan 23(3):127–134
Grechi I, Vivin P, Hilbert G, Milin S, Robert T, Gaudillère JP (2007) Effect of light and nitrogen supply on internal C:N balance and control of root-to-shoot biomass allocation in grapevine. Environ Exp Bot 59:139–149
Gupta V, Tripathy BC (2010) Effect of light quality on chlorophyll ac-cumulation and protein expression in wheat (Triticum aestivum L.) seedlings. Int J Biotechn Biochem 6(4):521–536
Hatfield Jl, Sauer Tj, Prueger Jh (2001) Managing soils to achieve greater water use efficiency: a review. Agr J 93:271–280
Hoad SP, Leakey RRB (1994) Effects of light quality on gas exchange and dry matter partitioning in Eucalyptus grandis W. Hill ex Maid-en. Forest Ecol Manag 70(1–3):265–273
Hückstädt AB, Mortensen LM, Gislerød HR (2013) The effect of high maximum day temperatures and coloured film cover on growth and morphogenesis of some herbs in a CO2 enriched greenhouse atmosphere. Eur J Hortic Sci 78(5):203–208
Hunt, R. 1990. Basic growth analysis: Plant growth analysis for begin-ners. Unwin Hyman, Boston
Jao RC, Lai CC, Fang W, Chang SF (2005) Effects of red light on the growth of Zantedeschia plantlets in vitro and tuber formation using light-emitting diodes. Hortscience 40(2):436–438
Jiao Y, Lau OS, Deng XW (2007) Light-regulated transcriptional net-works in higher plants. Nat Rev Genet 8:217–230
Johkan M, Shoji K, Goto F, Hahida S, Yoshihara T (2012) Effect of green light wavelength and intensity on photomorphogenesis and photosynthesis in Lactuca sativa. Environ Exp Bot 75:128–133
Kami C, Lorrain S, Hornitschek P, Fankhauser C (2010) Light-regu-lated plant growth and development. Curr Top Dev Biol 91:29–66
Kim, HH, Goins, GD, Wheeler, RM, Sager, JC (2004a) Green-light supplementation for enhanced lettuce growth under red- and blue-light-emitting diodes. Hortscience 39:1617–1622
reached its greatest value with white light and the lowest value with yellow light. Chl a/b ratios were the highest in plants grown under purple and blue light (Wang et al. 2009); these results differ from those previously reported and even from the findings of the current study.
Some recent findings suggest that plant growth and development might be co-regulated by photoreceptors and other endogenous factors such as hormones and a tempera-ture sensing system (Facella et al. 2012), indicating a very complex relationship between different factors that regulate plant morphology and physiology and that plant species present differences in their interactions with the surround-ing environment. The results of the present study indicate that it may be essential to choose a suitable light quality for maximizing plant photosynthetic performance, growth and, eventually, production, reducing plant stress, when artifi-cial supplemental light is used in protected farmlands.
Conclusions
Our results demonstrate that bell pepper plants grown under coloured polyethylene films may respond to relatively small changes in the light environment that are induced by mono-chromatic films. Nevertheless, the responsiveness of these plants to light stimulation was less evident than in other plant species grown under the same light conditions. Based on the results of most of the analyzed variables, no clear trend was found that would indicate that a colour has a marked posi-tive or negative effect on plant growth because each evalu-ated variable was affected differently by the tested coloured covers. Hence, these results can be taken as a basis to induce a specific effect in bell pepper plants, according to the needs of farmers, after analyzing the benefit-cost ratio.
Acknowledgments The team gratefully acknowledges the generous support of the Research Directorate (Dirección de Investigaciones—DIN) of the Pedagogical and Technological University of Colombia (UPTC) for providing us with the funding and opportunity to conduct this research project. We also thank Dr. Gerhard Fischer for his critical reading of this manuscript and for his helpful suggestions, as well as Carlos Fernandez for helping during data collection.
References
Alam MN, Islam MS, Ali MK, Barkotulla MAB, Khandaker SMAT (2007) Effect of light qualities on dry matter production, crop growth performance and chlorophyll content in onion plant. Res J Agr Biol Sci 3(6):871–875
Casierra-Posada F, Nieto PJ, Ulrichs C (2012a) Crecimiento, produc-ción y calidad de flores en calas (Zantedeschia aethiopica (L.) K. Spreng) expuestas a diferente calidad de luz. Rev UDCA Act & Div Cient 15(1):97–105
Author's personal copy

155
1 3
Growth of bell pepper plants (Capsicum annuum) affected by coloured covers
Wang Y, Nogushi K, Terashima I (2011) Photosynthesis-dependent and –independent responses of stomata to blue, red and green monochromatic light: differences between the normally oriented and inverted leaves of sunflower. Plant Cell Physiol 52:479–489
Wilson SB, Iwabuchi K, Rajapakse N, Young RE (1998) Responses of broccoli seedlings to light quality during low-temperature storage in vitro: I. Morphology and survival. Hortsci 33(7):1253–1257
Fánor Casierra-Posada, ist am 22.12.1959 in Cartago/Kolumbien geboren. Er ist Agrar-ingenieur (1989) und arbeitet an der Univer-sidad Pedagógica y Tecnológica de Colombia als Professor für Pflanzenphysiologie und Pflanzenvermehrung. An der Humboldt Uni-versität zu Berlin promovierte er 1999. Von 1989 bis 1995 war er als Geschäftsleiter in verschiedenen Obstbaubetrieben in Kolumb-ien tätig. Er hat viele Artikel in nationalen und internationalen wissenschaftlichen
Zeitschriften veröffentlicht.Von 2007 bis 2011 war er als Chefredak-teur der kolumbianischen wissenschaftlichen Zeitschrift „Revista Colombiana de Ciencias Hortícolas“ tätig.
Kim, HH, Goins, GD, Wheeler, RM, Sager, JC (2004b) Stomatal con-ductance of lettuce grown under or exposed to different light qual-ity. Ann Bot 94:691–697
Li H, Tang C, Xu Z (2013) The effects of different light qualities on rapeseed (Brassica napus L.) plantlet growth and morphogenesis in vitro. Sci Hortic-Amsterdam 150:117–124
Lin KH, Huang MY, Huang WD, Hsu MH, Yang ZW, Yang CM (2013) The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. Capitata). Sci Hortic-Amsterdam 150:86–91
Macedo AF, Leal-Costa MV, Tavares ES, Lage CLS, Esquibel MA (2011) The effect of light quality on leaf production and develop-ment of in vitro-cultured plants of Alternanthera brasiliana Kun-tze. Environ Exp Bot 70(1):43–50
Paulilo MTS, Duz SR, Siminski A, Santos M. (2007) Responses to light changes in tropical seedlings of the Brazilian Atlantic rain-forest tree species Cecropia glazioui (Cecropiaceae) and Cedrela fissilis (Meliaceae). Aust J Bot 55:795–802
Piszczek P, Głowacka B (2008) Effect of the colour of light on cucum-ber (Cucumis sativus L.) seedlings. Veg Crops Res Bull 68:71–80
Shimazaki KI, Doi M, Assmann SM, Kinoshita T (2007) Light regula-tion of stomatal movement. Annu Rev Plant Biol 58:219–247
Wang H, Gu M, Cui J, Shi K, Zhou Y, Yu J (2009) Effects of light quality on CO2 assimilation, chlorophyll-fluorescence quenching, expression of Calvin cycle genes and carbohydrate accumulation in Cucumis sativus. J Photoch Photobio B 96:30–37
Author's personal copy




















