
Giselle Medeiros da Costa One - Cinasama
669
1
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Giselle Medeiros da Costa One - Cinasama

1

Giselle Medeiros da Costa One Giselle Medeiros da Costa One
(Org.)
CIÊNCIAS BIOLÓGICAS: os desafios da pesquisa
na atualidade
IMEA JOÃO PESSOA-PB
2021

Instituto Medeiros de Educação Avançada - IMEA
Editor Chefe Giselle Medeiros da Costa One
Corpo Editorial
Giselle Medeiros da Costa One Bárbara Lima Rocha
Maria Luiza Souto Porto Roseanne da Cunha Uchôa
Iara Medeiros de Araújo Revisão Final
Ednice Fideles Cavalcante Anízio
FICHA CATALOGRÁFICA Dados de Acordo com AACR2, CDU e CUTTER
Laureno Marques Sales, Bibliotecário especialista. CRB -15/121
Direitos desta Edição reservados ao Instituto Medeiros de Educação Avançada – IMEAImpresso no Brasil / Printed in Brazil
One, Giselle Medeiros da Costa. O59c Ciências Biológicas:os desafios da pesquisa na atualidade /
Organizador: Giselle Medeiros da Costa One. Maria Luiza Souto Porto. IMEA. 2021.
669 fls. Prefixo editorial: 53005 ISBN: 978-65-89069-06-5 (on-line) Modelo de acesso: Word Wibe Web <http://www.cinasama.com.br> Instituto Medeiros de Educação Avançada – IMEA – João Pessoa - PB
1.Ciências Biológicas 2. Microbiologia 3. Biologia molecular I. Giselle Medeiros da Costa One II. Ciências Biológicas:os desafios da pesquisa na atualidade
CDU: 570

IMEA Instituto Medeiros de Educação
Avançada
Proibida a reprodução, total ou parcial, por qualquer meio ou processo, seja reprográfico, fotográfico,
gráfico, microfilmagem, entre outros. Estas proibições aplicam-se também às características gráficas e/ou
editoriais. A violação dos direitos autorais é punível como Crime
(Código Penal art. 184 e §§; Lei 9.895/80), com busca e apreensão e indenizações diversas (Lei
9.610/98 – Lei dos Direitos Autorais - arts. 122, 123, 124 e 126)
Todas as opiniões e textos presentes
neste livro são de inteira responsabilidade de seus autores, ficando o organizador
isento dos crimes de plágios e informações enganosas.
IMEA
Instituto Medeiros de Educação Avançada
Av Senador Ruy Carneiro, 115 ANDAR: 1; CXPST: 072; João Pessoa - PB
58032-100 Impresso no Brasil
2021

Aos participantes do CINASAMA pela
dedicação que executam suas atividades e
pelo amor que escrevem os capítulos que
compõem esse livro.

“Procure a sabedoria e aprenda a escrever os capítulos mais importantes de sua história nos momentos mais difíceis de sua vida”.
Augusto Cury

PREFÁCIO
O CINASAMA é um evento que tem como objetivo
proporcionar subsídios para que os participantes tenham
acesso às novas exigências do mercado e da educação. E ao
mesmo tempo, reiterar o intuito Educacional, Biológico, na
Saúde, Nutricional e Ambiental de direcionar todos que
formam a Comunidade acadêmica para uma Saúde Humana
e Educação socioambiental para a Vida.
Os livros “CIÊNCIAS BIOLÓGICAS: os desafios da
pesquisa na atualidade” tem conteúdo interdisciplinar,
contribuindo para o aprendizado e compreensão de varias
temáticas dentro da área em estudo. Esta obra é uma
coletânea de pesquisas de campo e bibliográfica, fruto dos
trabalhos apresentados no Congresso Internacional de Saúde
e Meio Ambiente realizado entre os dias 28 e 29 de Novembro
de 2020. No ano de 2020 devido a pandemia causada pelo
COVID – 19 o evento foi realizado em sua I Edição On-line.
Os eixos temáticos abordados no Congresso
Internacional de Saúde e Meio Ambiente e nos livros
garantem uma ampla discussão, incentivando, promovendo
e apoiando a pesquisa. Os organizadores objetivaram
incentivar, promover, e apoiar a pesquisa em geral para que
os leitores aproveitem cada capítulo como uma leitura
prazerosa e com a competência, eficiência e profissionalismo
da equipe de autores que muito se dedicaram a escrever

trabalhos de excelente qualidade direcionados a um público
vasto.
O livro CIÊNCIAS BIOLÓGICAS: os desafios da
pesquisa na atualidade” apresenta interdisciplinaridade
entre a as áreas da saúde concentrado em títulos com temas
que relatam experiência profissional e revisões bibliográficas
nas áreas afins e abrange as áreas: biologia geral,
bioquímica, ecologia, fisiologia, imunologia, microbiologia,
parasito, genética de microrganismos e biologia molecular.
Esta publicação pode ser destinada aos diversos
leitores que se interessem pelos temas debatidos.
Espera-se que este trabalho desperte novas ações,
estimule novas percepções e desenvolva novos humanos
cidadãos.
Aproveitem a oportunidade e boa leitura.

SUMÁRIO
CIÊNCIAS BIOLÓGICAS – BIOLOGIA GERAL ________________________ 14
CAPÍTULO 1 ________________________________________________ 15
AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO
DAS FOLHAS DE AVICENNIA SCHAUERIANA STAPF & LEECHMAN _________ 15
CAPÍTULO 2 ________________________________________________ 35
SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA
ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH (DANIO
RERIO) _____________________________________________________ 35
CIÊNCIAS BIOLÓGICAS - BIOQUÍMICA ___________________________ 56
CAPÍTULO 3 ________________________________________________ 57
CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO
FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE MORINGA
OLEIFERA LAM. _______________________________________________ 57
CAPÍTULO 4 ________________________________________________ 78
O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER ___________ 78
CAPÍTULO 5 ________________________________________________ 98
ESTUDO COMPARATIVO DE INIBIDORES DE UREASES _______________ 98
CAPÍTULO 6 _______________________________________________ 116
PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE MORINGA OLEIFERA
LAM. ____________________________________________________ 116

CIÊNCIAS BIOLÓGICAS - ECOLOGIA ____________________________ 137
CAPÍTULO 7 _______________________________________________ 138
O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO
DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES? ____________________ 138
CIÊNCIAS BIOLÓGICAS - FISIOLOGIA ___________________________ 162
CAPÍTULO 8 _______________________________________________ 163
PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS
PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A HIPERTENSÃO
NEUROGÊNICA ____________________________________________ 163
CAPÍTULO 9 _______________________________________________ 183
PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS
EXPERIMENTAIS DA LESÃO PULMONAR AGUDA __________________ 183
CAPÍTULO 10 ______________________________________________ 202
TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE
DOENÇAS CARDIOVASCULARES: UMA REVISÃO __________________ 202
CAPÍTULO 11 ______________________________________________ 223
TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO:
FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS ____________________________________________ 223
CIÊNCIAS BIOLÓGICAS - IMUNOLOGIA _________________________ 247

CAPÍTULO 12 ______________________________________________ 248
ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E
RINITE ALÉRGICAS COMBINADAS ______________________________ 248
CAPÍTULO 13 ______________________________________________ 266
CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
_________________________________________________________ 266
CAPÍTULO 14 ______________________________________________ 287
CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19 _______ 287
CAPÍTULO 15 ______________________________________________ 309
MACRÓFAGOS M1/M2 NA LEISHMANIOSE ______________________ 309
CAPÍTULO 16 ______________________________________________ 331
MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE
NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA ____________________ 331
CAPÍTULO 17 ______________________________________________ 353
SARS-COV-2: COMORBIDADES E COINFECÇÃO ___________________ 353
CAPÍTULO 18 ______________________________________________ 373
TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E
NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19 _ 373
CIÊNCIAS BIOLÓGICAS - MICROBIOLOGIA _______________________ 399
CAPÍTULO 19 ______________________________________________ 400
ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA
(PSEUDOBOMBAX MARGINATUM) ____________________________ 400
CAPÍTULO 20 ______________________________________________ 420

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO _________________________________________________ 420
CAPÍTULO 21 ______________________________________________ 443
ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E
RAÍZES DE MORINGA OLEIFERA LAM. (MORINGACEAE): UMA REVISÃO ____ 443
CAPÍTULO 22 ______________________________________________ 462
BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O
USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL _____ 462
CAPÍTULO 23 ______________________________________________ 484
ESTUDOS ANTIMICROBIANOS COM ALOE VERA: UMA REVISÃO _______ 484
CAPÍTULO 24 ______________________________________________ 503
FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS ____ 503
CAPÍTULO 25 ______________________________________________ 522
PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE
UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA _____________ 522
CAPÍTULO 26 ______________________________________________ 545
TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS _________________ 545
CIÊNCIAS BIOLÓGICAS -PARASITOLOGIA _______________________ 563
CAPÍTULO 27 ______________________________________________ 564
PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE
AEDES AEGYPTI: UMA REVISÃO _______________________________ 564
CAPÍTULO 28 ______________________________________________ 585

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS
PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS ________________ 585
GENÉTICA DE MICRORGANISMOS _____________________________ 604
CAPÍTULO 29 ______________________________________________ 605
MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E
DERIVADOS _______________________________________________ 605
BIOLOGIA MOLECULAR ______________________________________ 624
CAPÍTULO 30 ______________________________________________ 625
AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA
PARAÍBA: UMA REVISÃO INTEGRATIVA _________________________ 625
BIOTECNOLOGIA ___________________________________________ 645
CAPÍTULO 31 ______________________________________________ 646
APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO _________________________________________________ 646
.

14
CIÊNCIAS BIOLÓGICAS – BIOLOGIA GERAL

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
15
CAPÍTULO 1
AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO
HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF & LEECHMAN
Leandro de Lima COUTINHO1
Marllon Alex Nascimento SANTANA2 Tonny Cley Campos LEITE3
Amanda Reges de SENA4
Jéssica Andréia Pereira BARBOSA5,6
1Pós-Graduando no Centro de Investigação Translacional em Oncologia, FM/USP;2Pós-Graduando do Departamento de Ciências Biológicas, UFPE;3Técnico do Laboratório de
Bromatologia, IFPE; 4Docente do Curso de Técnico em Alimentos, IFPE; Orientadora-FACOTTUR
RESUMO: Avicennia schaueriana, espécie endêmica da vegetação de manguezal, é bastante utilizada pelas comunidades tradicionais no tratamento de reumatismo, úlceras e cicatrização de feridas. Desta forma, o objetivo deste estudo foi avaliar o potencial gastroprotetor do extrato hexânico das folhas de A. schaueriana. O extrato hexânico (As-Hex) foi preparado por maceração exaustiva, e o perfil fitoquímico analisado através de cromatografia em camada delgada (CCD). A atividade citotóxica foi realizada pelo método do MTT frente às linhagens Raw 264.7 e Vero, assim como o ensaio de toxicidade aguda. A atividade gastroprotetora foi avaliada através do modelo de úlcera induzida por etanol, seguida da

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
16
determinação dos níveis de mieloperoxidase (MPO). A CCD demonstrou a presença de terpenos no extrato As-Hex. Na determinação da toxicidade aguda do extrato orgânico não foi registrada mortes, sinais tóxicos ou alterações nos parâmetros bioquímico e hematológicos avaliados. Referente a atividade gastroprotetora, o extrato hexânico reduziu o índice de lesão ulcerativa, além de diminuir os níveis de MPO. O presente estudo revelou que o extrato hexânico das folhas de A. schaueriana possui propriedade gastroprotetora, sugerindo que esta atividade pode estar associada a presença de metabólitos como terpenos, com a capacidade de síntese de prostaglandina e outros protetores da mucosa gástrica. Portanto, o extrato hexânico deve ser considerado como uma alternativa terapêutica para o tratamento de infecções e úlcera gástrica. Palavras-chave: Avicennia schaueriana. Fitoquímica. Terpenos. Antiulcerogênica. INTRODUÇÃO
A úlcera gástrica (UG) é caracterizada pela lesão na
mucosa gástrica, que pode se estender desde danos
superficiais nos tecidos até lesões mais profundas podendo
atingir a submucosa subjacente ou muscular externa
(ELSHAZLY et al., 2018; MANJEGOWDA et al., 2017). É uma
doença associada a distúrbios gastrointestinais que afeta mais
de 10% da população mundial (SHEN et al., 2017; YOO et al.,
2018). A UG é desenvolvida a partir do desequilíbrio entre
fatores protetores , tais como, barreira de muco, secreção de
bicarbonato, aumento dos níveis de antioxidantes e produção
de prostaglandinas, fluxo sanguíneo mucoso, entre outros, e os
fatores lesivos, por exemplo, pepsina, ácido clorídrico, radicais

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
17
livres reativos e oxidantes (ATHAYDES et al., 2018; LIANG et
al., 2018).
Para o tratamento de úlcera gástrica comumente são
utilizados fármacos como os antagonistas de receptores H2,
como a cimetidina e inibidores da bomba de prótons (IBPs),
como o omeprazol. Entretanto, o uso contínuo desses podem
causar uma série de efeitos colaterais e recorrência de úlcera
ao término do tratamento (ARAÚJO, 2018). Portanto, diante das
dificuldades em relação a cura da úlcera gástrica e presença de
efeitos colaterais a partir do tratamento atual, faz-se necessário
a busca de um novo tratamento anti-úlcera que apresente alta
eficácia, baixa ou nenhuma toxicidade e efeitos adversos
mínimos.
Como alternativa seria o uso de plantas medicinais,
sendo estas bastante utilizada por comunidades tradicionais
para uma variedade de doenças (ABBAS et al., 2018). O Brasil
apresenta uma grande diversidade em relação a sua flora, com
cerca de 60 a 250 mil espécies vegetais no país, sendo destas
40% apresentando atividades biológicas. Dessa forma, muitas
são as pesquisas voltadas para identificar os possíveis
metabólitos secundários com a capacidade de tratar ou prevenir
doenças. Essas pesquisas têm confirmado o potencial de
plantas medicinais para a cura de diversas doenças, tais como
diabetes, câncer, antiulcerogênica, entre outras (CESÁRIO et
al., 2018; YOUS et al., 2018). Na literatura podemos encontrar
uma diversidade de vegetais com efeito gastroprotetor, sendo
importante alternativa para o tratamento úlcera gástrica,
estando muitas vezes seus efeitos associados a ação
antioxidante diminuindo a peroxidação lipídica, proteína e

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
18
danos no DNA, auxiliando na prevenção da inflamação que leva
a úlceras gástricas (ATHAYDES et al., 2018). Dessa forma, com
objetivo de descobrir novos tratamentos eficazes e com menos
efeitos colaterais voltados para doenças gastrointestinais,
muitas plantas veem sendo estudadas, e com isso suas
propriedades farmacológicas confirmadas e sem efeitos
adversos minimizados (ALMEIDA et al., 2019).
Avicennia schaueriana, pertence à família Acanthacea,
conhecida popularmente como mangue preto ou siriúba,
(PROFICE, et al., 2010) e na literatura já foi destacada com
algumas atividades biológicas, tais como, propriedade
cicatrizante (LOPES et al., 2019) e gastroprotetora (BARBOSA
et al., 2019).
Dessa forma, a investigação da caracterização química
e propriedades farmacológicas de Avicennia schaueriana
tornam-se importantes na obtenção de novas substâncias com
potencial terapêutico, e que apresentem efeitos colaterais
reduzidos. Além disso, ao relatar as atividades biológicas de
plantas medicinais na literatura científica, os conhecimentos
construídos pelas comunidades tradicionais são muitas vezes
reconhecidos e validados.
MATERIAL E MÉTODO
MATERIAL BOTÃNICO
As folhas de A. schaueriana foram coletadas no
mangue da ilha de Itamaracá, localizado na costa norte do
estado de Pernambuco (7º 48 '69 "S / 34º 51' 34" W), em

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
19
novembro de 2014. O material da planta foi identificado no
Herbário do Centro de Biociência da Universidade Federal de
Pernambuco, onde um espécime foi depositado sob número de
77.367.
LINHAGENS CELULARES
As linhagens Vero (fibroblastos de rim de macaco verde
africano) e RAW-264.7 (macrófagos murinos) foram
provenientes do Laboratório de Cultura de Células do
Departamento de Antibióticos/UFPE, mantidas de acordo com
o protocolo estabelecido pelo setor.
ANIMAIS EXPERIMENTAIS
Foram utilizados ratos Wistar machos (250 - 300 g) e
camundongos swiss fêmeas (25-30 g), provenientes do Biotério
do Departamento de Antibióticos da Universidade Federal de
Pernambuco (UFPE). Os animais receberam água e ração
(PRESENCE) “ad libitum” e foram mantidos em condições
normais de temperatura (21 ± 2 °C) e umidade (60 ± 1%), sob
ciclo natural claro e escuro de 12-12 horas. Todos os
tratamentos e procedimentos foram realizados de acordo com
o “Manual Prático sobre Usos e Cuidados Éticos de Animais de
Laboratório” da Sociedade Brasileira de Ciência em Animais de
Laboratório (SBCAL). Os protocolos de experimentação foram
aprovados pela Comissão de Ética em Experimentação Animal
(CEEA) da UFPE, processo nº 23076.018743/2015-42.

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
20
OBTENÇÃO DE EXTRATOS ORGÂNICOS E FRAÇÕES DAS
FOLHAS DE AVICENNIA SCHAUERIANA
As folhas de A. schaueriana foram coletadas e secas por
3 dias (72 h) em estufa com temperatura controlada (50 °C) e
renovação constante de ar. As mesmas foram moídas em
moinho de facas. Os extratos foram obtidos pelo método de
maceração por um período de 7 dias em temperatura ambiente
e protegida da incidência da luz. O solvente utilizado para a
extração foi hexano por um período de 48 h. O solvente foi
evaporado sob pressão reduzida (rotaevaporação) e em
seguida foi seco em umdessecador.
ANÁLISE DO PERFIL FITOQUÍMICO- CROMATOGRAFIA DE
CAMADA DELGADA (CCD)
Para análise do perfil fitoquímico o extrato foi testado na
concentração de 5 mg/mL em CCD, utilizando-se como fase
estacionária sílica gel F254 e fases móveis específicas de acordo
com o metabólito analisado. Para análise da prospecção
fitoquímica foram utilizadas como fases móveis diferentes
sistemas de solventes com polaridade crescente, sendo cada
fase móvel específica para o respectivo metabólito
analisado. Foram utilizados reveladores e padrões
cromatográficos adequados conforme descrito na Tabela 1.

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
21
AVALIAÇÃO CITOTÓXICA FRENTE ÀS LINHAGENS RAW
264.7 E VERO
A atividade citotóxica foi realizada através do método do
MTT brometo de 3-(4,5-dimetiltiazol-2-il)-2,5-difeniltetrazólio.
As linhagens de células normais utilizadas foram Vero (células
epiteliais do rim de macaco) e RAW 264.7 (macrófagos
murinos) mantidas em meio de cultura DMEM. Os meios foram
suplementados com 10 % de soro fetal bovino e 1 % de solução
de antibiótico (penicilina e estreptomicina). As linhagens foram
mantidas em estufa a 37 ºC em atmosfera úmida enriquecida
com 5% de CO2. As células Vero e RAW 264.7 (105 células/mL)
foram plaqueadas em placas de 96 poços e incubadas por 24
h. Em seguida o extrato foi dissolvido em DMSO (1%) e
adicionado aos poços em concentrações finais de 50, 25, 12,5
e 6,25 µg/mL. Após 72 h de reincubação foi adicionado 25 µL
de MTT (5 mg/mL) e depois de 3 h de incubação, o meio de
cultura com o MTT foram aspirados e 100 µL de DMSO foi
adicionado a cada poço. A absorbância foi medida em um leitor
de microplacas no comprimento de onda de 560 nm. O
experimento foi realizado em triplicata e a média da
percentagem de inibição e desvio padrão foram calculados. A
diferença entre os valores das medias da viabilidade celular
foram avaliadas pela Anova one way e pós teste Turkey’s (p <
0,05) no programa GraphPad Prism 6.0. demo

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
22
TOXICIDADE AGUDA
Grupos de três camundongos albinos swiss fêmeas
receberam via oral o extrato hexânico da A. schaueriana na
dose de 2000 mg/kg e o controle recebeu o veículo (salina com
5% de DMSO). Os parâmetros comportamentais observados
foram: agitação, atividade motora, locomoção, resposta ao
toque, piloereção, ptose, cianose, movimentos estereotipados,
sonolência, frêmito vocal, ataxia, contorções, agressividade,
convulsões, micção, defecação, diarreia). Durante 14 dias
seguintes, foram avaliados o consumo de água, ração, peso
corporal e mortalidade. No último dia de observação, os animais
foram anestesiados com tiopental (5 mg/kg) e posteriormente
submetidos a coleta de sangue por punção cardíaca. O material
biológico (sangue) foi submetido a análise dos parâmetros
bioquímicos e hematológicos. Este experimento foi repetido,
sendo o número final igual a 6 animais por grupo.
PARÂMETROS HEMATOLÓGICOS E BIOQUÍMICOS
Para o teste hematológico, amostras sanguíneas foram
coletadas em tubos contendo EDTA K2 e analisadas em
contador hematológico de células ABX micros 60 (HORIBE
medical). Para o teste bioquímico, as amostras foram coletas
em tubos com ativador de coágulo e centrifugados a 4000 rpm
por 5 minutos. O soro obtido foi analisado em BIOPLUS 2000
utilizando kits bioquímicos Labtest para os seguintes
parâmetros: glicose, triglicerídeos, proteínas, colesterol, ureia,
creatinina, fosfatase alcalina, TGO e TGP.

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
23
ÚLCERA GÁSTRICA INDUZIDA POR ETANOL ABSOLUTO
Os ratos Wistar foram distribuídos em grupos (n=5
animais/grupo) e submetidos a jejum de 24 horas com livre
acesso à água. Após este período, cada grupo recebeu o
tratamento com extratos hexânico das folhas de A. schaueriana
(50, 100 e 200 mg/kg v.o.). Como controle negativo os animais
receberam veículo (salina 0,9% e 5% de DMSO) e pantoprazol
(30 mg/Kg) como controle positivo. Após 30 minutos da
administração dos respectivos tratamentos, os animais
receberam por via oral 1,0 mL de etanol absoluto. Após 1 hora
da administração do etanol, os animais foram sacrificados e
seus estômagos retirados e abertos na sua curvatura maior
para contagem das lesões ulcerativas e cálculos do índice de
lesões ulcerativas (ILU). A contagem das lesões ulcerativas
foi realizada a partir de imagens digitalizadas e analisadas pelo
software ImageJ (1.51 d). As lesões foram classificadas através
dos níveis I <1 mm2; II 1 – 3 mm2; e III > 3 mm2. O índice de
lesão ulcerativa foi calculado pela fórmula: 1 x (número de
lesões nível I) + 2 x (número de lesões nível II) + 3 x (número
de lesões nível III). O % C= 100 – (IU tratado x 100/ IU controle).
DETERMINAÇÃO DOS NÍVEIS DE MIELOPEROXIDASE
(MPO)
Para avaliar os níveis da mieloperoxidase (enzima
presente nos neutrófilos, indicativas de processo inflamatório),
os tecidos dos estômagos dos animais submetidos ao modelo
de úlceras induzidas por etanol (controles e tratados) foram

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
24
colocados em tampão fosfato de potássio 50 mM (pH 6,0)
contendo 0,5% de hexadeciltrimetilamônio (1,5 mL - 100 mg de
tecido) em homogeneizador de Potter. O homogenato foi
agitado em vórtex e centrifugado durante 15 minutos a 2.500
rpm. Dez microlitros do sobrenadante obtido foram adicionados
em microplaca de 96 poços, em duplicata, sendo a seguir,
adicionado 200 µL de solução tampão contendo di-hidrocloreto
de o-dianisidina (16,7 mg), água bidestilada (90 mL), tampão
fosfato de potássio (10 mL) e H2O2 1% (50 µL). O nível da
enzima foi determinado pela medida de absorbância (460 nm)
em leitor de microplaca com comprimento de onda de 610 nm,
sendo expressa em mDO/tecido.
ANÁLISE ESTATÍSTICA
Os resultados foram expressos como valores de média ±
desvio padrão para cada grupo experimental, utilizando o
software GraphPad Prism (versão 6.0). Os dados foram
avaliados por meio da análise de variância (ANOVA) de uma
via, seguido pelo teste de Tukey, *P<0,0001, significativos após
análise de variância (ANOVA) com intervalo de confiança de
95%, quando comparados ao grupo controle. A contagem das
lesões ulcerativas foi realizada a partir de imagens digitalizadas
e analisadas pelo software ImageJ (1.51 d).

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
25
RESULTADOS E DISCUSSÃO
PROSPECÇÃO FITOQUÍMICA CROMATOGRAFIA EM
CAMADA DELGADA (CCD)
Através dos resultados obtidos verificou-se a presença
de Terpenos no extrato hexânico (Tabela1).
Barbosa et al. (2019) confirmaram a presença de
triterpenos no extrato acetato de etila das folhas de Avicennia
schaueriana corroborando os resultados obtidos neste trabalho.
Tabela 1. Prospecção fitoquímica do extrato orgânico hexânico
(As-Hex) das folhas de Avicennia schaueriana.
Metabólitos
Padrões
Revela
ções
Fase
Móvel/Proporção
Extrat
o As-
Hex
Taninos
Ácido Tâncio
Cloreto
Férrico
(1%)
Clorofórmio:
Metanol:
Água (65:30:05)
-
Alcaloides
Escopolamina
Dragen
dorff
Tolueno: Acetato de
Etila:
Dietilamina (70:20:1
0)
-
Flavanoides
Quercetina
NEU Acetato de Etila: Ác.
Fórmico: Ác. Acético
glacial: Água
(100:11:11:26)
-
Terpenóides
Lupeol
Liberm
an-
Burcha
rd
Tolueno:
Clorofórmio:
Etanol (40:40:10)
+

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
26
Cumarina
Ácido
Cumárico
KOH-
ETOH
(10%)
Clorofórmio:
Metanol:Água
(01:01)
-
(-) Não detectável; (+) Presente.
AVALIAÇÃO CITOTÓXICA FRENTE ÀS LINHAGENS DE
CÉLULAS VERO E MACRÓFAGOS (RAW 264.7)
O percentual de inibição da viabilidade celular do extrato
hexânico está demonstrado na Tabela 2. Este extrato não
apresentou citotoxicidade frente às linhagens Vero e
macrófagos (RAW-264.7).
Lopes et al. (2019) evidenciaram que o extrato aquoso
das folhas de A schaueriana não foi citotóxico frente a células
Vero, estando de acordo com os resultados encontrados neste
trabalho.
Tabela 2: Avaliação de citotoxicidade do extrato hexânico (As-
Hex) (50 µg/mL) de Avicennia schaueriana frente às células de
macrófagos (RAW-264.7) e vero.
Extrato
Concentração
(µg/ml)
Raw-264.7
% de viabilidade
Vero
% de
viabilidade
AS-HEX 50 85,0 ± 17,2 87,2 ± 3,6
*p < 0.05. Significativos após análise de variância (ANOVA) de uma via
seguida pelo teste Tukey com intervalo de confiança de 95 %, quando
comprados ao grupo controle.
TOXICIDADE AGUDA

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
27
Durante a avaliação do screening hipocrático o grupo
que recebe o extrato hexânico de A. schaueriana na dose de
2.000 mg/kg não apresentou sinais de toxicidade nas primeiras
duas horas de observação e nos 14 dias seguintes, além disso
nenhum animal veio a óbito. Também não foi observada
variação significativa no peso, consumo de ração e água em
relação aos animais do grupo controle (Tabela 3).
Não foram observadas alterações hematológicas, como
nenhuma diferença estatística destes parâmetros avaliados nos
animais tratados com o extrato hexânico quando comparado ao
grupo controle, como demonstrado na Tabela 4. Os
camundongos tratados com o extrato hexânico (2.000 mg/kg-
v.o.) apresentaram todos os parâmetros bioquímicos dentro da
faixa de referência em relação ao grupo controle (Tabela 5).
Barbosa et al. (2019) realizaram o teste de toxicidade
aguda, onde os animais foram tratados por via oral com extrato
acetato de etila na dose de 2.000 mg/kg e não apresentaram
sinais de toxicidade, como também nenhum registro de morte,
além dos parâmetros bioquímicos e hematológicos inalterados.
Tabela 3: Efeito dos extratos orgânicos hexânico (As-Hex) das
folhas de Avicennia schaueriana sobre parâmetros fisiológicos
de camundongos Swiss (n=6 animais/grupo).
Grupos
Dose/v.
o
mg/Kg
Nº
Anima
is
Nº
Morte
s
Consum
o Ração
(g/dia)
Consum
o água
(mL/dia)
Peso
corpora
l inicial
Controle - 6 0/6 14,4±1,2 21,0±2,0 33,2±2,5
As-Hex
2000
6
0/6
15,7±1,4
20,9±2,0
32,6±2,7

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
28
**** p < 0,0001 Os valores foram expressos como média ± desvio padrão e
a diferença entre os grupos foram determinados por meio da Análise de
Variância (ANOVA), seguida pelo teste de Tukey.
Tabela 4: Efeito do extrato hexânico (As-Hex) das folhas de
Avicennia schaueriana sobre parâmetros hematológicos em
camundongos submetidos à toxicidade aguda. Parâmetros Controle As-Hex
2.000 mg/kg
WBC (103/mm3) 8,7 ± 0,3 7,3 ± 0,6
RBC (106/mm3) 9,3 ± 0,5 8,0 ± 0,7
HGB (g/dL) 13,2 ± 1,0 14,5 ± 1,4
HCT (%) 48,7 ± 3,3 36,8 ± 0,9
PLT (103/mm3) 672,4 ± 47,0 661,6 ± 51,5
PCT (H %) 438,8 ± 40,6 412,6 ± 37,3
MCV (L ƒm3) 52,1 ± 1,4 51,3 ± 2,4
MCH (L g//dl) 14,6 ± 0,9 17,3 ± 1,2
RDW (%) 12,6 ± 0,8 12,5 ± 0,7
MPV (L ƒm3) 6,7 ± 0,4 7,8 ± 0,6
PDW (%) 5,9 ± 0,7 3,4 ± 0,3
****p<0,0001 Os valores foram expressos como média ± desvio padrão e a diferença
entre os grupos foram determinados por meio da Análise de Variância (ANOVA),
seguida pelo teste de Tukey.
Tabela 5: Efeito dos extratos orgânicos hexânico (As-Hex) das
folhas de Avicennia schaueriana sobre parâmetros bioquímicos
em camundongos submetidos ao teste de toxicidade aguda. Parâmetros Controle As-Hex 2.000 mg/kg
Glicose 227,6 ± 27,6 204,1 ± 79,6
Triglicerídeos 77,7 ± 14,5 62,5 ± 25,0
Proteínas 5562,4 ± 343,8 4739,0 ± 2034,0
Colesterol 79,0 ± 6,2 142,6 ± 55,9
Ureia 75,5 ± 9,1 48,1 ± 21,2
Creatinina 0,2 ± 0,08 0,19 ± 0,04
Fosfatase 240,8 ± 25,3 239,1 ± 94,8

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
29
TGO 112,0 ± 10,9 100,1 ± 37,2
TGP 44,8 ± 6,5 44,2 ± 19,8
****p<0,0001 Os valores foram expressos como média ± desvio padrão e a diferença
entre os grupos foram determinados por meio da análise de variância (ANOVA),
seguida pelo teste de Tukey.
ÚLCERAS GÁSTRICAS INDUZIDAS POR ETANOL
ABSOLUTO
Os resultados obtidos no modelo de úlcera gástrica
induzida por etanol absoluto em ratos demonstraram que o
tratamento com o extrato hexânico (50, 100 e 200 mg/kg); e
pantoprazol (30 mg/kg) administrados por via oral diminuíram
de forma significativa as lesões gástricas quando comparados
com o grupo controle (Figura 1).
Barbosa et al. (2019) avaliaram a atividade
gastroprotetora do extrato acetato de etila (50, 100 e 200 mg/kg)
das folhas de Avicennia schaueriana, e observaram que o
tratamento com o extrato orgânico reduziu o índice de lesão
ulcerativa quando comparado ao grupo controle. A atividade
gastroprotetora de terpenos também se destaca em modelos de
indução aguda de úlceras gástricas em animais (CHINEDU;
IBRAHIM, 2017; NESELLO et al., 2017)).
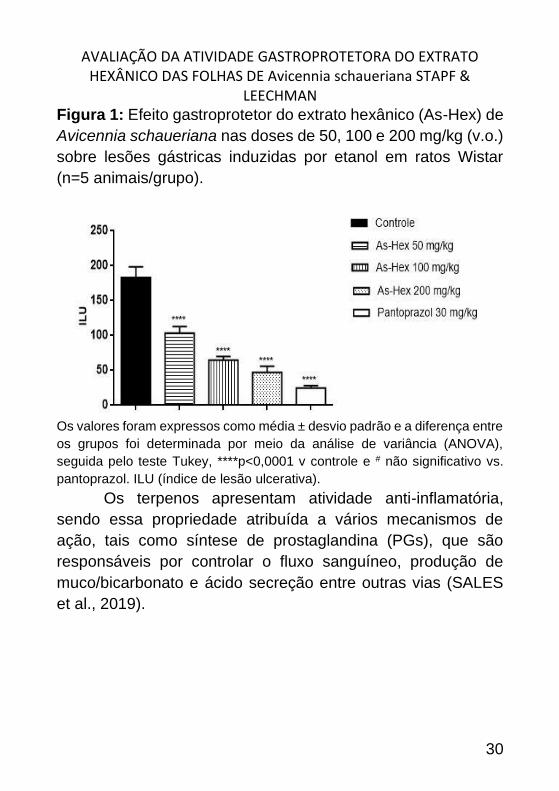
AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
30
Figura 1: Efeito gastroprotetor do extrato hexânico (As-Hex) de
Avicennia schaueriana nas doses de 50, 100 e 200 mg/kg (v.o.)
sobre lesões gástricas induzidas por etanol em ratos Wistar
(n=5 animais/grupo).
Os valores foram expressos como média ± desvio padrão e a diferença entre
os grupos foi determinada por meio da análise de variância (ANOVA),
seguida pelo teste Tukey, ****p<0,0001 v controle e # não significativo vs.
pantoprazol. ILU (índice de lesão ulcerativa).
Os terpenos apresentam atividade anti-inflamatória,
sendo essa propriedade atribuída a vários mecanismos de
ação, tais como síntese de prostaglandina (PGs), que são
responsáveis por controlar o fluxo sanguíneo, produção de
muco/bicarbonato e ácido secreção entre outras vias (SALES
et al., 2019).

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
31
DETERMINAÇÃO DOS NÍVEIS DE MIELOPEROXIDASE
(MPO)
Em relação aos níveis de mieloperoxidase no tecido da
mucosa gástrica, os grupos tratados com extrato hexânico nas
doses de 50, 100 e 200 mg/kg (maior inibição de lesão
ulcerativa) apresentaram redução nos níveis de MPO quando
comparado ao grupo controle (Figura 2).
A mieloperoxidase participa da liberação de mediadores
pró-inflamatórios, a partir de neutrófilos que ultrapassam a
barreira vascular endotelial, migrando por quimiotaxia para o
local da inflamação permitindo a liberação de elevado nível de
peptídeos, proteases e espécies reativas de oxigênio que
intensificam a resposta inflamatória (EL-BENNA et al., 2016).
Em consequência da inflamação ocorre o aumento de
citocinas pró-inflamatórias possibilitando a ativação de
neutrófilos, assim como também o acúmulo destas células nos
tecidos mucosos no processo de lesão gástrica, por sua vez
isso contribui para regulação do dano da úlcera, e essa
infiltração neutrofílica pode ser investigada a partir da atividade
da meiloperoxidase (LI et al., 2017).
Nos animais tratados com extrato hexãnico esse
aumento dos níveis de mieloperoxidase foi reduzida, o que
indica menor infiltração de neutrófilos para o tecido,
consequentemente, menor liberação de radicais livres e
substâncias lesivas no tecido, o que contribui para a melhora no
perfil gastroprotetor.
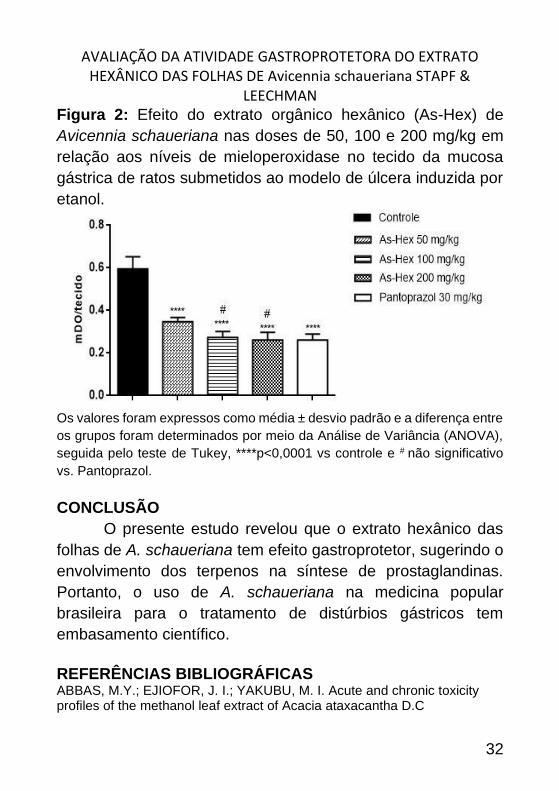
AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
32
Figura 2: Efeito do extrato orgânico hexânico (As-Hex) de
Avicennia schaueriana nas doses de 50, 100 e 200 mg/kg em
relação aos níveis de mieloperoxidase no tecido da mucosa
gástrica de ratos submetidos ao modelo de úlcera induzida por
etanol.
Os valores foram expressos como média ± desvio padrão e a diferença entre
os grupos foram determinados por meio da Análise de Variância (ANOVA),
seguida pelo teste de Tukey, ****p<0,0001 vs controle e # não significativo
vs. Pantoprazol.
CONCLUSÃO
O presente estudo revelou que o extrato hexânico das
folhas de A. schaueriana tem efeito gastroprotetor, sugerindo o
envolvimento dos terpenos na síntese de prostaglandinas.
Portanto, o uso de A. schaueriana na medicina popular
brasileira para o tratamento de distúrbios gástricos tem
embasamento científico.
REFERÊNCIAS BIBLIOGRÁFICAS ABBAS, M.Y.; EJIOFOR, J. I.; YAKUBU, M. I. Acute and chronic toxicity profiles of the methanol leaf extract of Acacia ataxacantha D.C

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
33
(Leguminosae) in Wistar rats. Bulletin of Faculty of Pharmacy, v. 56, p.185–189, 2018. ALMEIDA, G. V. B.; BALOGUN, K. A. S. O.; PAVAN, E.; ASCÊNCIO, S. D.; SOARES, I. M.; ZANATTA, A. C.; VILEGAS, W.; MACHO, A.; MARTINS, D. T. O. Chemical characterization and evaluation of gastric antiulcer properties of the hydroethanolic extract of the stem bark of Virola elongata (Benth.) Warb, Journal of Ethnopharmacology, v. 231, p. 103-124, 2019. ARAÚJO, Ruan Pablo Nunes. Investigação da atividade gastroprotetora do fitol em ratos. Dissertação (Mestrado em Farmacologia) – Universidade Federal do Piauí, Teresina, 2018. ATHAYDES, B. R.; ALVES, G. M.; ASSIS, A. L. E. M.; GOMES, J. V. D.; RODRIGUES, R. P.; CAMPAGNARO, B. P.; NOGUEIRA, B. V.; SILVEIRA, D.; KUSTER, R. M.; PEREIRA, T. M. C.; KITAGAWA, R. R.; GONÇALVES, R.C. R. Avocado seeds (Persea americana Mill.) prevents indomethacin induced gastric ulcer in mice. Food Research International.v. 119, p. 751–760, 2018. BARBOSA, J. A. P.; SANTANA, M. A. N.; OLIVEIRA, T. B.; MOTA, F. V. B.; BASTOS, I. V. G. A.; CORRÊA, A. J. C.; AMORIM, E. L. C.; VIEIRA, J. R. C.; SILVA, T. M. S.; SANTANA, A. L. B. D.; NASCKMENTO, M. S.; SILVA, T. G. Gastroprotective effect of ethyl acetate extract from Avicennia schaueriana Stapf & Leechman and underlying mechanisms, Biomedicine & Pharmacotherapy, v. 112, 2019. CESÁRIO, F. R. A. S.; ALBUQUERQUE, T.R.; LACERDA, G.M.; OLIVEIRA, M.R.C.; SILVA, B.A.F.; RODRIGUES, L.B.; MARTINS, A.O.B.P.B.; ALMEIDA, J.R.G.S.; VALE, M.L.; COUTINHO, H.D.M.; MENEZES, I.R.A. Chemical fingerprint, acute oral toxicity and anti-inflammatory activity of the hydroalcoholic extract of leaves from Tocoyena formosa (Cham. & Schlecht.) K. Schum. Saudi Journal of Biological Sciences, v. 26, p. 873-880, 2018. CHINEDU, O.; IBRAHIM, K.G. Menthol Ameliorates Secretagogue-Induced Gastric Acid Secretion in the Rat. International Journal of Current Research and Academic Review, v. 5, n.6, p. 113-118, 2017. EL-BENNA J. et al. Priming of the neutrophil respiratory burst: role in host defense and inflammation, Immunological Reviews, Copenhagen, v. 273, n. 1, p. 180-193, 2016. ELSHAZLY, S.M.; MOTTELEB, D. M. A.; IBRAHIM, I. A.A.E-H. Hesperidin protects against stress induced gastric ulcer through regulationof peroxisome proliferator activator receptor gamma in diabetic rats Chemico-Biological Interactions. v. 291, p. 153–161, 2018. LI W., WANG X., ZHI W., ZHANG H., HE Z., WANG Y. The gastroprotective effect of nobiletin against ethanol-induced acute gastric

AVALIAÇÃO DA ATIVIDADE GASTROPROTETORA DO EXTRATO HEXÂNICO DAS FOLHAS DE Avicennia schaueriana STAPF &
LEECHMAN
34
lesions in mice: impact on oxidative stress and inflammation. Immunopharmacology and Immunotoxicology, v. 39, n. 6, p. 354–363, 2017. LIANG, J.; DOU, Y.; WU, X.; LI, H.; WU, J.; HUANG, Q.; LUO, D.; YI, T.; LIU, Y.; SU, Z.; CHEN, J. Protective effect of patchoulene epoxide against ethanol-induced gastric ulcer in rats: Influence on oxidative stress, inflammation and apoptosis, Chemico-Biological Interactions, v. 283, p. 30-37, 2018. LOPES C. M. I., BARATELLA-EVÊNCIO L., SOUZA I. A., OLIVEIRA E. B., SÁ J. G. A., SANTANA M. A. N., MARCELINO NETO P. P., SANTANA E. S., SILVA L. A., VIEIRA J. R. C. Evaluation of cytotoxicity and wound healing activity of Avicennia schaueriana in cream. Anais da Academia Brasileira de Ciências, v. 91, e20180171, 2019. MANJEGOWDA S. B., RAJAGOPAL H. M., DHARMESH S.M. Polysaccharide of Black cumin (Nigella sativa) modulates molecular signaling cascade of gastric ulcer pathogenesis, International Journal of Biological Macromolecules, v. 101, p. 823-836, 2017. NESELLO, L. A. N.; CAMPOS, A.; ROSA, R. L.; ANDRADE, S. F.; CECHINEL FILHO, C. Screening of wild fruit trees with gastroprotective activity in different experimental models. Arquives Gastroenteroloy, v. 54, p. 135-138, 2017. PROFICE S. R., KAMEYAMA C., CÔRTES A. L. A., BRAZ D. M., INDRIUNAS A., VILAR T., PESSOA C., EZCURRA C., WASSHAUSEN D. Acanthaceae. In: Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro, 2010. SALES, P. F.; NÓBREGA, P. A.; NASCIMENTO, A. A.; CORRÊA, F. R. F. B.; CABRAL, N. V.; SILVA, E. G. Antiulcerogenic activity of the ethanolic extract of Licania macrophylla Benth. O Mundo da Saúde, v. 43, p. 814-833, 2019. SHEN, Y.; SUN, J.; NIU, C.; YU, D.; CHEN, Z.; CONG, W.; GENG, F. Mechanistic evaluation of gastroprotective effects of Kangfuxin on ethanol-induced gastric ulcer in mice. Chemico-Biological Interactions. v. 273, p. 115-124, 2017. YOO, J.-H.; LEE, J-S.; LEE, Y-S.; KU, S.; LEE, H-J. Protective effect of bovine milk against HCl and ethanol–induced gastric ulcer in mice. Journal of Dairy Science, v. 101, p. 1–13, 2018. YOUS, F.; ATMANI-KILANI, D.; DEBBACHE-BENAIDA, N.; CHERAFT, N.; SEBAIHI, S.; SAIDENE, N.; BENLOUKIL, M.; ATMANI, D. Anti-ulcerogenic and proton pump (H+, K + ATPase) inhibitory activity of Clematis flammula
L. extract. South African Journal of Botany, v. 119, p. 390–399, 2018..

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
35
CAPÍTULO 2
SORO DE LEITE BOVINO COMO PRODUTO
INOVADOR PARA ALIMENTAÇÃO DOS
ESTÁGIOS INICIAIS DA VIDA DO
ZEBRAFISH (Danio rerio) Maria Eduarda de Souza MAIA 1
Rafael Xavier MARTINS 2
Iris Flávia de Sousa GONÇALVES 3
Juliana Alves da Costa Ribeiro SOUZA 4
Davi Felipe FARIAS 5
1 Graduandos do curso de Biotecnologia, UFPB; 2 Mestrando em Biologia Celular e Molecular,
UFPB; 3 Mestranda em Bioquímica, UFC; 4 Mestre em Ciência Animal, UFPB; 5Orientador/Professor do DBM, UFPB.
Email: [email protected]
RESUMO: O zebrafish (Danio rerio) vem ganhando posição de destaque em vários campos da ciência. A alimentação dos estágios iniciais da vida dessa espécie tem sido um desafio, e uma alternativa que não havia sido avaliada é o soro do leite bovino, que possui uma excelente composição nutricional. Com isso, o objetivo deste estudo foi avaliar a adequação do soro de leite bovino liofilizado (SLBL) para alimentação de larvas de zebrafish. Inicialmente, a composição proximal e perfil de ácidos graxos da amostra de SLBL utilizada foi determinada. Em seguida, foram realizados dois ensaios de alimentação, com diferentes dietas formuladas a partir da substituição da ração comercial por porcentagens crescentes SLBL. O objetivo desses ensaios foi determinar o valor máximo de inclusão de SLBL na dieta capaz de garantir a melhor taxa de sobrevivência das larvas. Ao final dos experimentos, foram contabilizados o número de mortos por grupo amostral. A inclusão de SLBL na

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
36
dieta não exerceu efeitos negativos na sobrevivência ou no crescimento quando comparado à ração comercial, sendo necessário outros estudos para confirmar a adequação. Palavras-chave: Liofilização. Nutrição de peixes. Taxa de sobrevivência. Larvas de zebrafish.
INTRODUÇÃO
O zebrafish (Danio rerio), também conhecido como
“paulistinha”, é um pequeno teleósteo de 3-4 cm pertencente à
família Cyprinidae que nas últimas décadas vem ganhando
posição de destaque como organismo modelo para estudos de
biologia do desenvolvimento, bioquímica, fisiologia,
farmacologia, genética molecular, neurociência e toxicologia
(KHAN et al., 2018).
Os primeiros trabalhos utilizando o zebrafish foram
relacionados à biologia do desenvolvimento. A produção de
ovos translúcidos, a grande prole e o rápido desenvolvimento
possibilitaram um grande avanço no conhecimento da
embriogênese e do ciclo de vida dos vertebrados (KHAN et al.,
2018). Dentre as principais vantagens do uso do zebrafish
podem ser destacadas o pequeno espaço requerido para
criação e manutenção, o baixo custo e a praticidade para
triagens em larga escala. Dessa forma, pode-se dizer que o
zebrafish é um animal que combina a relevância de ser um
vertebrado na escala de um invertebrado (KHAN et al., 2018).
Esses fatores fazem com que o zebrafish esteja cada vez mais
ganhando um espaço nos laboratórios que outrora era ocupado
apenas pelos modelos tradicionais, como a mosca-dasfrutas
(Drosophila melanogaster), o rato (Rattus norvegicus) e o
camundongo (Mus musculus). Atualmente, uma ampla gama de

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
37
testes com diferentes compostos de interesse tem sido
realizada com embriões de zebrafish dessa espécie. O uso de
embriões apresenta-se como uma alternativa aos testes agudos
com juvenis e adultos, possibilitando, dessa forma, a
redução/substituição do uso desses organismos na
experimentação animal. Como principais vantagens do uso dos
embriões apontam-se o desenvolvimento embrionário rápido, a
grande quantidade de embriões produzida (média de 50
ovos/casal) e seu pequeno tamanho (BAMBINO et al, 2017;
ALESTRÕM et al., 2020). Apesar da praticidade da criação e
manejo do zebrafish, a alimentação dos estágios iniciais da vida
dessa espécie tem sido um desafio para os criadores.
O alimento vivo, especialmente paramécio, é o mais
indicado para alimentação de larvas de zebrafish do 9° ao 21°
dia. Contudo, a cultura desse protozoário é laboriosa, de fácil
contaminação e ainda inviabiliza o processo de automatização
da alimentação. Assim, várias alternativas de alimentação
artificial para larvas têm sido testadas. Dentre elas, extrato de
levedura e de fígado proporcionaram bons resultados
(ALESTRÕM et al., 2020).
Uma alternativa que ainda não havia sido avaliada é o
soro de leite bovino, subproduto da indústria queijeira. Por ter
um baixo custo e excelente composição nutricional, ele tem sido
proposto como complemento para diversos produtos da
indústria alimentícia a fim enriquecê-los do ponto de vista
nutricional (LIU et al., 2020). Além disso, a reutilização do soro
de leite evita o seu descarte errôneo, evitando a poluição do
ambiente uma vez que durante muito tempo esse subproduto
foi descartado em corpos d’água, causando um impacto

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
38
negativo, tanto a animais quanto a plantas (DE OLANDA et al.,
2017).
Tendo em vista a necessidade de busca de insumos
alternativos para a alimentação das fases inicias de vida do
zebrafish, e a excelente composição nutricional do soro de leite
bovino, uma alternativa a ser testada é a adequação desse
insumo como ingrediente para formulações de dietas para
larvas dessa espécie. Isso potencialmente pode contribuir para
o desenvolvimento de um produto inovador para ser usado na
criação desse organismo modelo, bem como para agregar valor
econômico e ambiental a esse resíduo da indústria queijeira e
importante poluente ambiental.
Sendo assim, o objetivo deste trabalho foi avaliar o
potencial do soro de leite bovino como insumo alternativo para
o desenvolvimento de uma dieta para o estágio larval do
zebrafish (D. rerio).
MATERIAIS E MÉTODOS
Obtenção dos embriões e larvas de peixe-zebra
Embriões de zebrafish (D. rerio), de uma linhagem do
tipo selvagem, com 3 h pós-fertilização (hpf) foram obtidos no
Laboratório de Biotecnologia de Organismos Aquáticos
(LaBOrA), do Centro de Biotecnologia da Universidade Federal
da Paraíba, coordenado pelo prof. Dr. Ian Amaral. Para a
obtenção dos embriões dois casais de zebrafish sexualmente
maduros foram transferidos para um dispositivo de
acasalamento na noite anterior ao dia da coleta dos embriões.

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
39
Após a coleta, os embriões foram lavados com água do
sistema de recirculação (onde estavam os parentais) e tiveram
a sua viabilidade analisada em estereomicroscópio. Os
embriões foram mantidos a 26 ±1°C e ciclo claro:escuro de
14:10h. No 9° dia de vida, as larvas viáveis foram submetidas
ao experimento de alimentação e as inviáveis foram
descartadas de acordo com protocolo n° 048/2016. aprovado
pela Comissão de Ética no Uso de Animais da Universidade
Federal da Paraíba (CEUA-UFPB).
Obtenção e liofilização do soro de leite bovino
O soro de leite bovino (SLB) utilizado foi cedido pelo
Laboratório de Inovação de Alimentos do Centro de
Biotecnologia (coordenado pela prof. Dr. Flávia Paulino) e foi
liofilizado no Laboratório de Bioprospecção de Recursos
Regionais (Bioprospec) da Universidade Federal do Ceará
(UFC), Fortaleza-CE (coordenado pela prof. Dr. Ana de Fátima
Fontenelle) e armazenado em tubos falcon, protegidos da luz
até o momento dos ensaios.
Determinação da composição proximal do SLBL
No Bioprospec, foram realizadas análises para
determinar a composição proximal (proteínas totais, lipídeos
totais, material mineral) do SLBL de acordo com as
metodologias utilizadas por Kassegn (2018). Os carboidratos
totais foram calculados por diferença dos demais constituintes.

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
40
Análise da composição de ácidos graxos do SLBL
A análise da composição de ácidos graxos foi realizada
nas instalações do Departamento de Engenharia de Alimentos,
no laboratório coordenado pela prof. Dr. Marta Suely Madruga.
Inicialmente, os ácidos graxos da amostra de SLBL foram
obtidos a partir da metodologia executada por Costa e
colaboradores (2020) e, posteriormente, metilados seguindo a
metodologia executada por Silva e colaboradores (2019). A
identificação e quantificação dos ésteres de ácidos graxos foi
realizada por cromatografia gasosa (Varian 430 - GC,
California, EUA), acoplado com detector de ionização de chama
(DIC), coluna capilar de sílica fundida (SP TM – 2560,
SUPELCO, Bellefonte, EUA) Os ácidos graxos foram
identificados por comparação dos tempos de retenção dos
ésteres metílicos das amostras com padrões Supelco ME19 -
Kit (Fatty Acid Methy Esters C4 - C24).
Determinação de sólidos solúveis totais
O SLBL e a ração comercial foram pesadas e
depositadas em microtubos de 1,5 mL. Em seguida, foi
adicionado 1 mL de água destilada em cada tubo, que
posteriormente foram submetidos a um processo de agitação
por 2 min. Posteriormente, os sobrenadantes foram transferidos
para microtubos com peso conhecido, levados para uma estufa
de secagem (50° C.) por 16 h. Os sólidos solúveis totais foram
obtidos a partir da diferença de peso dos microtubos.

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
41
Determinação de proteínas e carboidratos solúveis
O preparo da amostra foi determinado de acordo com o
item 2.5.1. A dosagem de proteínas solúveis foi feita pelo
método de Bradford (1976), utilizando uma curva padrão com
albumina séria bovina. O teor de carboidratos solúveis foi
determinado pelo método do fenol-sulfúrico executado por
Rego e colaboradores (1956). Os ensaios foram feitos em
triplicata.
Confecção das dietas com base em SLBL e experimentos de
alimentação com larvas de zebrafish
Foram realizados dois experimentos de alimentação
diferentes. Um deles um piloto para ajuste de condições
experimentais e delimitação preliminar dos níveis ideais de
inclusão de SLBL e o outro para determinação do nível máximo
de SLBL em dietas para larvas. Para o ensaio piloto foram
preparadas quatro dietas em quantidades crescentes de SLBL
na composição: 1) Dieta com 25% de SLBL e 75% de ração
comercial (SLBL 25); 2) Dieta com 50% de SLBL e 50% de
ração comercial (SLBL 50); 3) Dieta com 75% de SLBL e 50%
de ração comercial (SLBL 75); 4) Dieta com 100% de SLBL
(SLBL 100). A dieta 0% (contendo 100% de ração comercial) foi
utilizada como controle.
Foram preparados cinco grupos com 41 larvas com nove
dias de vida, mantidos em um aquário de 1 L. Durante o período
experimental, cada grupo foi alimentado duas vezes por dia
com 0,5 mg de dieta durante 12 dias. A taxa de sobrevivência

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
42
foi calculada (%sobrevivência = n° de sobreviventes/n° total x
100).
O delineamento do ensaio piloto não permitiu realizar
comparações entre a taxa de sobrevivência dos diferentes
grupos. Sendo assim, a fim de avaliar a significância estatística
das diferenças encontradas, foi realizado um segundo
experimento de alimentação, baseado nos dados encontrados
do primeiro. Para isso, foram confeccionadas novas dietas com
diferentes quantidades de SLBL: 1) dieta com 0 % de SLBL e
100% de ração comercial (SLBL 0); 2) dieta com 75% de SLBL
e 25 % de ração comercial (SLBL 75); 3) dieta com 85% de
SLBL e 15 % de ração comercial; 4) dieta com 95% de SLBL e
5% de ração comercial (SLBL 95); e 5) dieta com 100% de SLBL
e 0% de ração comercial (SLBL 100). Foram preparados cinco
grupos, onde cada um foi subdividido em três subgrupos
contendo 10 larvas cada, gerando um total de 30 indivíduos por
grupo.
Análise estatística
Foram calculadas média, desvio padrão e coeficiente de
variação dos valores obtidos. Para avaliar o grau de
similaridade entre as variáveis foram realizados testes de
comparações simples (t-Student) e de comparações múltiplas
(ANOVA seguida de Teste de Tukey). O nível de confiança das
inferências foi de 95 % (α =0,05).

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
43
RESULTADOS E DISCUSSÃO
Composição proximal do SLBL
A alimentação é um ponto fundamental e crítico para o
bom desenvolvimento do zebrafish, principalmente nas
primeiras etapas da sua vida. Tendo em vista que a alimentação
influencia na expressão de genes responsáveis pelo
crescimento, uma dieta inadequada pode acarretar um
desenvolvimento anormal de larvas e juvenis. Dessa forma, a
composição nutricional deve suprir as necessidades fisiológicas
do organismo (OPAZO,2017). Neste âmbito, procedeu-se a
determinação da composição proximal do SLBL. O teor de
proteínas quantificado no SLBL foi cerca de 5,5 vezes menor do
que na ração comercial (Tabela 1). Além disso, o teor de
proteínas encontrado no SLBL (9,4%) foi inferior ao
requerimento nutricional de 35-40% de proteína para a carpa
comum (Cyprinus carpio) (SIGNOR et al., 2017). Em
decorrência da escassez de informações sobre os
requerimentos nutricionais específicos de zebrafish,
frequentemente os dados desse outro ciprínideo de importância
comercial são utilizados para fins comparativos. Apesar do
baixo teor proteico apresentado pelo SLBL, é relatado que as
proteínas do soro do leite possuem alto valor biológico, o que
significa que possuem em sua composição quantidades
significativas de aminoácidos essenciais (KILARA e VAGHELA,
2018).
Em relação a quantidade de lipídios, normalmente o
nutriente constitui cerca de 7 a 15% da dieta dos peixes (CRAIG

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
44
et al., 2017), e o teor de lipídeos totais encontrado no SLBL foi
de 14,2%, valor 1,6 vezes superior ao encontrado na ração
comercial. Estudos demonstraram que uma dieta enriquecida
em ácidos graxos tem papel fundamental na maturação sexual
das fêmeas, influenciando na qualidade dos ovos e no melhor
desenvolvimento dos embriões (NOWOSAD et al., 2017). O
macronutriente majoritário detectado no SLBL foram os
carboidratos (69%). O teor de carboidratos totais do SLBL foi
3,4 vezes superior ao requerimento nutricional de 20%, que é o
ideal para a obtenção de uma boa taxa de crescimento,
conversão alimentar e ganho de peso (CRAIG et al., 2017).
Apesar dos peixes possuírem enzimas para a digestão de
carboidratos, esse elemento não é crucial para o bom
desenvolvimento de peixes.
Entretanto, a inserção de carboidrato na dieta pode
trazer algumas vantagens, como a redução do catabolismo
proteico para síntese de glicose, melhorar a eficiência de
retenção proteica e diminuir as perdas metabólicas de
nitrogênio no ambiente. A capacidade de digerir carboidratos
varia entre as espécies, sendo que o zebrafish é uma das que
consegue utilizar esses componentes de forma eficaz
(DAMMSKI et al., 2011). Ainda em relação a composição
proximal do SLBL, o teor de minerais encontrado foi de 7,1%
que foi muito próximo daquele da ração comercial, 7,3%.

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
45
Tabela 1. Composição proximal (%) do Soro de Leite Bovino
Liofilizado (SLBL) e de uma ração comercial para peixes.
Fonte: Autoral. 2017
Os valores são médias ± desvios-padrão. Lipídeos e matéria mineral foram
determinados em duplicata de uma mesma amostra, enquanto o teor de
nitrogênio foi feito em triplicata. Letras sobrescritas distintas na mesma linha
representam diferença significativa (t-Student; p < 0,05). **Tropical Gran
Discus Plus (Sarandi, Brasil). †N x 6,38.
Composição de ácidos graxos do SLBL
Na análise do perfil de ácidos graxos do soro de leite
bovino liofilizado foram encontrados diferentes níveis de ácidos
graxos saturados, monoinsaturados e poliinsaturados (dados
não mostrados). Dentre os poli-insaturados, foram detectados
2,89% de LA e 0,65% de ALA. Esses valores atendem à
demanda desses ácidos graxos na fase larval de zebrafish, uma
vez que, segundo Graig (2017) os requerimentos de EFA para
peixes de água doce, geralmente, podem ser atendidos pela
inclusão ALA em quantidades que variam de 0,5 a 1,5 por cento
da dieta seca para que esses possam ser alongados e
posteriormente convertidos em seus produtos bioativos. O fato
de tanto o LA quanto ALA serem metabolizados pelo mesmo
conjunto das enzimas do tipo alongase e dessaturase faz com

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
46
que o balanço entre n-6 e n-3 na composição lipídica do regime
alimentar de peixes seja um ponto crítico.
Algumas espécies exigem uma maior proporção de n-3,
alguns exigem quantidades iguais de ambos e outros exigem
maiores proporções de n-6. Uma relação desequilibrada dessas
classes de ácidos graxos poliinsaturados pode favorecer a
ocorrência de diversos tipos de doença, além do aparecimento
de diversos sintomas, como baixa eficiência alimentar,
crescimento lento, aumento do índice hepático, fígado pálido e
inchado, necrose nas nadadeiras dorsal, cauda e erosão
mandibular.
O zebrafish está inserido no grupo que possui uma
demanda maior de n-6 do que de n-3, de acordo com
experimentos que avaliaram taxa de crescimento e fertilidade,
sendo assim o perfil da relação n-6: n-3 detectado no SLBL é
adequado para os requerimentos dessa espécie (ADAM et al.,
2017).
Componentes solúveis do SLBL
A solubilidade de nutrientes é um pré-requisito crucial
para dietas processada para alimentação de larvas de
zebrafish. Fatores como tamanho, tempo de suspensão,
estabilidade na água e aceitabilidade têm relação direta com a
solubilidade da dieta, bem como são consideradas
características importantes durante a alimentação de larvas
dessa espécie (DAMMSKI et al., 2011).
De acordo com os resultados mostrados na Tabela 2, os
valores de proteínas solúveis e sólidos solúveis totais da

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
47
amostra de SLBL foram superiores (p < 0,05) àqueles da
amostra de ração comercial. O SLBL apresentou um teor de
sólidos solúveis totais de 0,38 mg/mL, valor 3,2 vezes superior
ao encontrado na ração comercial (0,12 mg/mL). Em relação às
proteínas solúveis, o valor encontrado na amostra de soro de
leite liofilizado foi de 0,11 mg/mL, enquanto o da ração não foi
possível ser quantificado pois encontrava-se abaixo da
sensibilidade mínima da metodologia empregada. Quanto ao
teor de carboidratos solúveis, o SLBL e a ração comercial não
diferiram significativamente (p> 0,05).
Tabela 2. Teores de proteínas e carboidratos solúveis e sólidos
solúveis totais de uma amostra de soro de leite bovino liofilizado
(SLBL)* e de uma ração comercial para peixes.
Fonte: Autoral. 2017
Os valores são representados como média ± desvio-padrão de uma triplicata
da mesma amostra. Letras sobrescritas distintas na mesma linha
representam diferença significativa (t-Student; p < 0,05). *Amostra
desidratada por liofilização obtida de dessora da produção de queijo. **
Tropical Discus Gran D-50 Plus (Sarandi, Brasil). †Não detectado a partir da
metodologia utilizada.
A solubilidade dos nutrientes do SLBL é uma
característica que favorece o processo digestivo das larvas de
zebrafish. Tendo em vista que em sua fase larval o zebrafish

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
48
possui um sistema digestório simples, e consequentemente a
capacidade de absorção de nutrientes limitada, quanto mais
solúvel e digerível for o alimento fornecido, mais rápida será a
absorção de nutrientes (COUTINHO, 2018).
Um ponto negativo, é que alimentos que contêm uma alta
taxa de compostos hidrossolúveis, como o SLBL, requerem um
maior cuidado durante a sua administração, uma vez que em
meio aquoso, esses nutrientes serão diluídos no ambiente
dentro de um curto período de tempo, podendo ocasionar
problemas relacionados a qualidade da água (como a
diminuição do teor de oxigênio dissolvido e o surgimento de
algas), assim como prejudicar a ingestão nutricional das larvas.
Entretanto, esses problemas podem ser contornados
através da adoção de algumas medidas preventivas, como por
exemplo a troca regular da água dos tanques e padronização
da quantidade de alimento ofertado às larvas (DAMMSKI et al.,
2011).
Experimentos de alimentação com larvas de zebrafish
As dietas preparadas apresentaram um aspecto de pó
fino, o que durante os experimentos de alimentação permitiu a
permanência visível da dieta na coluna d’água por tempo
suficiente para que as larvas conseguissem se alimentar. Isso
é um ponto positivo, pois larvas muito jovens de zebrafish não
possuem a bexiga natatória completamente formada, e isso
dificulta a mobilidade para busca de alimento. Dessa forma,
quanto mais tempo a dieta passar em suspensão na água maior
será a chance das larvas se alimentarem.

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
49
Outro aspecto importante no que diz respeito a
granulometria da dieta é que as larvas de peixe-zebra até o
primeiro mês de vida não apresentam o aparelho bucal
completamente desenvolvido. Portanto, isso dificulta o acesso
a alimentos secos ou vivos de granulometria mais elevada
(DAMMSKI et al., 2011).
Quanto ao experimento de alimentação piloto, foi
observado que o aumento da inserção de SLBL nas dietas
aumentou a taxa de sobrevivência das larvas. As dietas SLBL
75 e SLBL 50 apresentaram os resultados mais promissores,
proporcionando taxas de sobrevivência próximas a 50%.
Entretanto, foi possível observar uma diminuição da taxa de
sobrevivência tanto da SLBL 0 quanto da SLBL 100 em relação
as demais dietas. A expressiva diminuição da taxa de
mortalidade observadas na administração dessas dietas pode
ser, em parte, explicada pela excelente composição nutricional
do soro lácteo bovino (KILARA e VAGHELA, 2018). Entretanto,
alimentar as larvas somente com a ração comercial (dieta SLBL
0) ou SLBL (SLBL 100) não foi suficiente para garantir uma taxa
de sobrevivência que se equiparasse aos melhores resultados
encontrados.
Tabela 4. Efeito de dietas contendo níveis crescentes de soro
de leite bovino liofilizado (SLBL) sobre a taxa de sobrevivência

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
50
de larvas de peixe-zebra (D. rerio) durante 12 dias (do 9° ao 21°
dia de vida). Fonte: Autoral. 2017
N = número de larvas utilizadas no experimento. M = número de larvas
mortas durante o experimento. S = número de larvas sobreviventes ao final
do experimento. Taxa de sobrevivência (%) = n° de sobreviventes/ n° total x
100.
Tendo em vista que as larvas de zebrafish nas fases
iniciais da vida não são maduras o suficiente para terem um
comportamento de busca de alimento desenvolvido (OPAZO et
al., 2017), é provável que as mortes computadas durante esse
período tenham ocorrido pela não adaptação às dietas
ofertadas, uma vez que os parâmetros da água avaliados não
apresentavam condições estressantes que dificultassem o
desenvolvimento do peixe. Carvalho e colaboradores (2006)
encontraram uma taxa de sobrevivência de 86% de larvas com
21 dias alimentadas com náuplios de Artemia sp., enquanto no
presente estudo o valor mais elevado foi de 54% para as larvas
alimentadas com SLBL 75. Contudo, além de ter usado um
sistema de recirculação de água desde a fase inicial do
experimento, CARVALHO et al. (2006) usaram uma densidade
de 18 larvas/L, valor 2,3 vezes menor do que a utilizada no
presente trabalho (41 larvas/L).
O alimento ofertado para larvas de peixe possui
influência direta no seu crescimento. Fatores como o turnover
proteico (processo de síntese e degradação da proteína
corporal) são mais acentuados nas fases larval e juvenil de
peixes, fazendo com que tenham um crescimento relativamente
rápido (DHILLON et al., 2019).

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
51
Após o ensaio piloto, foi realizado o segundo
experimento para avaliar a quantidade máxima de inserção de
SLBL que garantisse a melhor taxa de sobrevivência das larvas.
Dessa forma, um segundo experimento de alimentação foi
realizado, onde cada grupo foi formado por três subgrupos de
larvas. Além disso, o segundo experimento foi realizado
também no intuito de avaliar o nível máximo de inclusão de
SLBL nas dietas que proporcionasse um aumento da taxa de
sobrevivência ou, pelo menos, não apresentasse valores
inferiores àqueles do grupo alimentado somente com ração
comercial.
Das cinco dietas administradas no segundo experimento
de alimentação, os melhores resultados foram apresentados
pela SLBL 75, onde foi possível observar uma taxa de
sobrevivência média de 43,3% (Figura 1). Esses dados,
aparentemente, estão de acordo com os resultados obtidos no
ensaio piloto, onde a inclusão de 75% de SLBL na dieta
apresentou os resultados mais promissores. Entretanto, não foi
verificada nenhuma diferença estatística (p > 0,05) entre os
dados de sobrevivência dos diferentes grupos. Um fator
importante acerca das informações encontradas nesse
experimento é que não houve diferença entre os dados de
sobrevivência dos grupos alimentados apenas com SLBL e com
ração comercial. O que reforça a possibilidade do uso desse
insumo como ingrediente alternativo na formulação de dietas
para larvas de peixe-zebra.
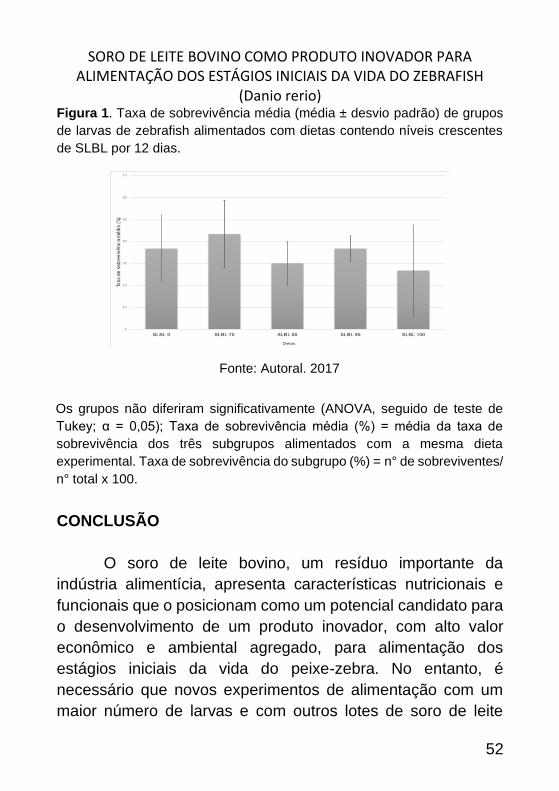
SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
52
Figura 1. Taxa de sobrevivência média (média ± desvio padrão) de grupos
de larvas de zebrafish alimentados com dietas contendo níveis crescentes
de SLBL por 12 dias.
Fonte: Autoral. 2017
Os grupos não diferiram significativamente (ANOVA, seguido de teste de
Tukey; α = 0,05); Taxa de sobrevivência média (%) = média da taxa de
sobrevivência dos três subgrupos alimentados com a mesma dieta
experimental. Taxa de sobrevivência do subgrupo (%) = n° de sobreviventes/
n° total x 100.
CONCLUSÃO
O soro de leite bovino, um resíduo importante da
indústria alimentícia, apresenta características nutricionais e
funcionais que o posicionam como um potencial candidato para
o desenvolvimento de um produto inovador, com alto valor
econômico e ambiental agregado, para alimentação dos
estágios iniciais da vida do peixe-zebra. No entanto, é
necessário que novos experimentos de alimentação com um
maior número de larvas e com outros lotes de soro de leite

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
53
sejam realizados, bem como que se amplie a caracterização
nutricional e físico-química do SLBL para que se confirme sua
adequação como um novo insumo alimentar para a aquicultura.
AGRADECIMENTOS
Ao prof. Dr. Davi Felipe Farias por não ter medido
esforços para nos orientar e apoiar o desenvolvimento desse
trabalho.
À toda a equipe do Laboratório de Avaliação de Riscos
de Novas Tecnologias (LabRisco) e da Unidade de Produção
de Organismos Modelo Não Convencionais (UniPOM) por todo
o apoio fornecido para a elaboração desse trabalho.
Ao prof. Dr. Ian Amaral, coordenador do Laboratório de
Biotecnologia de Organismos Aquáticos (LabORA) e a toda a
sua equipe por disponibilizar os embriões de peixe-zebra, bem
como a sua estrutura física para realização de parte dos
ensaios.
A prof. Dr. Marta Suelly Madruga, por se disponibilizar a
realizar a determinação do perfil de ácidos graxos da amostra
de soro de leite bovino liofilizado.
A prof. Dr. Flávia Paulino, coordenadora do Laboratório
de Inovação de Alimentos, por fornecer a amostra de soro de
leite bovino.
A toda equipe do Laboratório de Bioprospecção de
Recursos Naturais (BioProspec), coordenado pela professora
Ana de Fátima Fontenelle, por se disponibilizarem a ajudar no
processo de liofilização e determinação da composição
proximal nas amostras.

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
54
REFERÊNCIAS BIBLIOGRÁFICAS ADAM, A.-C. et al. High dietary arachidonic acid levels induce changes in complex lipids and immune-related eicosanoids and increase levels of oxidised metabolites in zebrafish (Danio rerio). The British journal of nutrition, v. 117, n. 8, p. 1075–1085, abr. 2017. ALESTROM, P., et al; Peixe-zebra: recomendações de alojamento e maneio. Laboratory Animals, v. 54, n.3, p. 213-224, 2020. BAMBINO, K.; CHU, J. Peixe-zebra em toxicologia e saúde ambiental: Tópicos atuais em biologia do desenvolvimento.
Academic Press,v. 124, p. 331-367, 2017.
CARVALHO, A. P; ARAÚJO, L; SANTOS, M. M; Rearing zebrafish (Danio rerio) larvae without live food: evaluation of a commercial, a practical and a purified starter diet on larval performance.
Aquaculture Research, v. 37, p. 1107-1111, 2006.
COSTA, C. et al. Effects of fatty acid profile of supplements on intake, performance, carcass traits, meat characteristics, and meat sensorial analysis of feedlot Bos indicus bulls offered a high-concentrate diet.
Transl Anim Sci, 2020.
COUTINHO, C. I. N.. Manutenção e reprodução em peixe-zebra (Danio rerio) –. 2018. 54 f. Tese (Doutorado) - Curso de Biomedicina, Universidade de Porto, Porto, 2018.
CRAIG, S. et al. Understanding fish nutrition, feeds, and feeding.Virginia Cooperative Extension, 420- 256, 2017.
DAMMSKI, A. P.; MULLER, B. R.; GAYA, C.; REGONATO, D. Zebrafish: manual de criação em biotério. Curitiba: Universidade Federal do Paraná, p. 101, 2011.
DE OLANDA, G. B. et al. Qualidade fisiológica de sementes de feijão tratadas com soro de leite bovino.; Revista Univap, v. 22, p.40, 2017 DHILLO, S. S. et al. Metabolic profiling of zebrafish embryo development from blastula period to early larval stages. PloS one, v.
14, p.15, 2019.
KASSEGN, H. H. Determination of proximate composition and bioactive compounds of the Abyssinian purple wheat. Cogent Food & Agriculture, v. 4, n. 1, 2018.
KHAN, F. R.; SALEH, S. A. Zebrafish (Danio rerio) as a model organism; Current Trends in Cancer Management. IntechOpen, 2018.

SORO DE LEITE BOVINO COMO PRODUTO INOVADOR PARA ALIMENTAÇÃO DOS ESTÁGIOS INICIAIS DA VIDA DO ZEBRAFISH
(Danio rerio)
55
KILARA, A; VAGHELA, M. N. Whey proteins. In: Proteins in food processing. Woodhead Publishing, p. 93-126, 2018.
LIU, Y. W., et al. Caracterizando as alterações das proteínas do soro do leite bovino após processamento industrial simulado. LWT 133
2020
NOWOSAD, J.et al. Enrichment of Zebrafish Danio rerio (Hamilton, 1822) Diet with Polyunsaturated Fatty Acids Improves Fecundity and Larvae Quality. Zebrafish, v. 00, n. 00, p. 364-370, 2017. OPAZO, R. et al. Performance of Debaryomyces hansenii as a Diet for Rotifers for Feeding Zebrafish Larvae. Zebrafish, v. 14,p.187–194, 2017.
REGO, G.C. et al. Bioconversion of pretreated sugarcane vinasse into hydrogen: new perspectives to solve one of the greatest issues of the sugarcane biorefinery. Biomass Conv. Bioref, 2020.
SILVA, T. et al. Effect of cocoa butter equivalent on cocoa butter crystallization behavior and on dark chocolate. Brazilian Journal of
Food Research. v. 10. p. 149, 2019
UDAYANGANI, R. M. C. et al.. Feeding of nano scale oats β-glucan enhances the host resistance against Edwardsiella tarda and protective immune modulation in zebrafish larvae.Fish and Selfish Immunology, v. 60, p. 60-72, 2017.
SIGNOR, Altevir et al. Lysine in the diet of common carp (Cyprinus carpio) fingerlings. Boletim do Instituto de Pesca, [S.l.], v. 43, n. 3, p. 464-473, dec. 2018

56
CIÊNCIAS BIOLÓGICAS - BIOQUÍMICA

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
57
CAPÍTULO 3
CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR
DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa oleifera Lam.
Ana Gabrielle Barros Silva 1
Matheus Cavalcanti de Barros 2
Thiago Henrique Napoleão 3 Patrícia Maria Guedes Paiva 3
1 Graduanda do curso de Bacharelado em Ciências Biológicas, UFPE; 2 Mestrando pelo Programa de Pós-graduação em Ciências Biológicas, UFPE;
3 Professor do Departamento de Bioquímica, UFPE. [email protected]
RESUMO: Moringa oleifera é uma planta nativa da Índia e bastante cultivada no nordeste brasileiro, onde é utilizada na medicina popular e alimentação humana e de outros animais. O presente trabalho avaliou o potencial fotoprotetor de extratos orgânicos de folhas de M. oleífera. Extratos foram obtidos por maceração das folhas com hexano (Mo–Hex), acetato de etila (Mo–AcoEt) e etanol (Mo-EtOH). O conteúdo de fenois totais foi mensurado através do método de Folin-Ciocalteu, a atividade antioxidante foi determinada pelo método do DPPH e o fator de proteção solar (FPS) foi calculado a partir do método espectrofotométrico na região UVB. O conteúdo (mg EAT/g) de fenóis em MO-Hex, Mo–AcoEt e Mo–EtOH foi 0,74 ± 0,01, 0,67 ± 0,38 e 1,99 ± 0,21, respectivamente. Atividade antioxidante foi detectada em todos os extratos (Mo–Hex, 51,38 ± 3,82%; Mo–AcoEt, 27,41 ± 1,00%; Mo–EtOH, 80,22 ± 0,66%). O efeito fotoprotetor foi dose-dependente sendo na concentração de 100 µg/mL, 1,38 ± 0,07 (Mo–Hex), 3,37 ± 0,22 (Mo–AcoEt) e 1,38 ± 0,07 (Mo-EtOH). O estudo revela que o conteúdo de fenóis, a

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
58
atividade antioxidante e o efeito fotoprotetor variam de acordo com a polaridade do solvente utilzado na extração dos princípios ativos. Palavras-chave: Fotoproteção. Produtos naturais. Fitoquímica
INTRODUÇÃO
Moringa oleifera Lam. (moringa ou cedro) pertence a
família Moringaceae e é natural do Noroeste da Índia, Paquistão,
Bangladesh e Afeganistão. A planta também é distribuída por
diversos países no mundo, sendo amplamente cultivada na
África (ex. Nigéria e Egito), América do Sul (ex. Nordeste do
Brasil) e Ásia (ex. Paquistão e Malásia) (LEONE et al., 2015). A
ampla faixa de cultivo desta árvore é devido à sua capacidade
de sobreviver em ambientes com baixa disponibilidade de água
e tolerar solos levemente alcalinos e pouco férteis (MANOJ;
RANJITHA, 2020).
A moringa tem relevância econômica pois folhas, frutos,
sementes e raízes são utilizados para diversos fins. Árvores de
M. oleifera são utilizadas como cercas ou para ornamentação, e
a madeira é relativamente macia, sendo bastante usada como
combustível para cozinhar e para a fabricação de papel (SEIFU;
TEKETAY, 2020). As cascas podem ser usadas como cordas ou
para o tratamento de diabetes e hipertensão e a raiz é
empregada para alimentação no leste africano, bem como anti-
helmínticos e para tratar dor de dente (POPOOLA; OBEMBE,
2013). Já as flores popularmente são utilizadas como
antiartríticas, agentes hipocolesterolêmicos e para tratar
resfriados (DABA, 2016). Além disso, O óleo das sementes pode
ser utilizado na culinária geral, na lubrificação de relógios e

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
59
outras maquinarias delicadas, na fabricação de perfumes e
como combustível (SANTOS et al., 2015).
As folhas são utilizadas como suplemento dietético para no
tratamento da desnutrição devido seu alto valor nutricional. A
relevância nutricional dessa planta tem sido bastante
documentada, uma vez que apresenta níveis significativos de
proteínas, carboidratos, vitamina C, cálcio (necessário para o
processo de coagulação), fósforo e fibras que auxiliam no
processo de digestão dos alimentos em humanos (COZ-
BOLAÑOS et al., 2018).
A planta também possui uma grande variedade de
metabólitos secundários, tais como taninos, saponinas,
esteroides, alcaloides, flavanoides e outros compostos fenólicos.
Destre esses, os flavanoides e outros fenois se destacam, pois
são os principais fitoquímicos capazes de reduzir a ação de
radicais livres com consequente diminuição do estresse
oxidativo e danos às moléculas biológicas. Os metabólitos
secundários estão relacionados às propriedades farmacológicas
das plantas medicinais e assim, estas representam uma
importante opção terapêutica para a manutenção da saúde das
pessoas, em especial para a população que não tem acesso a
medicamentos (NASCIMENTO, 2016).
Dentre os metabólitos produzidos por plantas, estão aqueles
compostos conhecidos como fotoprotetores, pois são capazes
de absorver a radiação UV, atuando no tecido cutâneo através
de moléculas fotoestáveis que possuem grupos cromóforos que
absorvem a radiação (KORKINA et al., 2018). Desta forma,
esses compostos fotoprotetores são responsáveis por minimizar

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
60
e prevenir os efeitos negativos provocados pela radiação solar
(ORLANDA, 2015)
A necessidade do uso de protetor solar é uma realidade
dos dias atuais e assim, a busca por novos compostos
fotoprotetores orgânicos pode disponibilizar alternativas para
evitar os danos causados pela radiação ultravioleta à saúde das
pessoas (SECCO, 2018). Muitos estudos tem sido
desenvolvidos com o objetivo de compreender melhor o efeito
fotoprotetor das plantas. A radiação UV é facilmente absorvida
por moléculas como o DNA e proteínas causando aumento de
espécies reativas de oxigênio, queimaduras com diferentes
graus, fotodermatoses, fotossensibilidade e efeitos mais graves
como fotocarcirnogênese (SCHNEIDER, 2020). De acordo com
Instituto Nacional do Câncer (INCA) são registrados por ano
cerca de 180 mil novos casos de câncer de pele.
A radiação ultravioleta que atinge a superfície da Terra é
apenas uma pequena parte do espectro de radiação emitido pelo
sol. A intensidade da radiação é variável, os raios UVA (315-
400nm) correspondem a cerca de 5%, os raios UVB (320-290
nm) correspondem a cerca de 0,5% e UVC (290-270 nm) que é
altamente nociva mas é absorvida pela camada de ozônio antes
de atingir a superfície terrestre (LOPES, 2015). A radiação UVA
é a radiação menos energética, mas consegue atingir a derme
profunda, e afetar negativamente a elasticidade natural da pele,
sendo responsável pelo envelhecimento cutâneo precoce. Já a
radiação UVB tem a capacidade de penetrar totalmente na
epiderme, fazendo com que haja o espessamento da camada
córnea, e por isso é a grande responsável pelos danos agudos
e crônicos a pele (CARVALHO, 2015).

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
61
Ante o exposto, o objetivo deste estudo foi avaliar o
potencial fotoprotetor de extratos orgânicos das folhas de M.
oleifera visando compreender o potencial dessa planta em
previnir danos causados pela radiação ultravioleta. Ademais, o
conteúdo de fenois e a atividade antioxidante dos extratos foram
mensurados a fim de relacioná-los com os valores de
fotoproteção obtidos.
MATERIAIS E MÉTODOS
Obtenção dos extratos
As folhas de M. oleifera foram coletadas no campus
Recife da Universidade Federal de Pernambuco. Após
separação dos folíolos, o material vegetal foi lavado, seco à
temperatura ambiente e pulverizado. Para a obtenção dos
extratos, o pó das folhas (10 g) foi mantido em contato com
diferentes solventes (100 mL) durante 48 h. Ao final do processo,
foram obtidos extratos em hexano (Mo-Hex), acetato de etila
(Mo-Acoet) e metanol (Mo-MeOH). Posteriormente, os extratos
foram concentrados com o auxílio de um rotoevaporador e
mantidos em capela até a secagem total.
Dosagem de fenois totais
O conteúdo de fenois totais foi mensurado através do
método de Folin-Ciocalteu adaptado em microplacas de 96
poços (BOBO-GARCÍA et al., 2015). Inicialmente, um total de 20
μL dos extratos diluídos (0,2 mg/mL) foram distribuídos em cada
poço. Posteriormente, 100 μL do reagente Folin-Ciocalteu
(10%) foram adicionados seguido por agitação durante 60s. Em

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
62
seguida, 75 μL de solução de carbonato de sódio (100 g/L) foram
adicionados com posterior agitação. Após 2 h de incubação no
escuro à temperatura ambiente, a absorbância foi medida a 750
nm usando o leitor de microplacas. A absorbância da mesma
reação com água ao invés dos extratos foi subtraída da
absorbância da reação com a amostra. Diluições de ácido tânico
(10–200 mg/mL) foram usadas como padrões para calibração.
Os resultados foram expressos como miligramas equivalentes
de ácido tânico por grama de extrato (mg EAT /g).
Atividade antioxidante (teste do DPPH)
O teste do DPPH (2,2-difenil-1-picril-hidrazil) baseia-se no
mecanismo de transferência de um elétron, processo que
permite detectar a capacidade de um produto em transferir um
elétron e reduzir qualquer substância (VALATINA;
NEELAMEGAM , 2015). O princípio do teste é a conversão do
radical DPPH, o qual possui coloração roxa, para o DPPH-H que
possui coloração amarela.
O teste foi realizado de acordo com os procedimentos
metodológicos descritos por Pires et al. (2017) com algumas
modificações. Para detecção do potencial antioxidante das
amostras, 20 μL dos extratos diluídos em metanol (0,2 mg/mL)
foram adicionados em placas de 96 poços. Após adição de 280
μL de DPPH em cada poço, a placa foi incubada no escuro
durante 30 min. Em seguida, a redução do DPPH foi
determinada por alteração colorimétrica a 517 nm utilizando o
leitor de microplacas. Os resultados foram representados em
porcentagem de atividade antioxidante (AAO%) de acordo com
a fórmula abaixo:

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
63
Determinação do comprimento de onda de absorção
máximo e fator de proteção solar (FPS) in vitro
Para determinação do comprimento de onda de
absorção máximo (λmax) e o fator de fotoproteção (FPS), os
extratos foram diluídos em metanol até obter as concentrações
de 12,5 µg/mL, 25 µg/mL, 50 µg/mL e 100 µg/mL.
Posteriormente, foi realizada uma varredura
espectrofotométrica em comprimentos de onda entre 260-400
nm, com intervalos de 5 nm. As leituras foram realizadas com
cubeta de quartzo e metanol foi utilizado como branco.
O cálculo do fator de fotoproteção foi realizado de acordo
com a equação abaixo:
𝑭𝑷𝑺 = 𝑭𝑪 𝒙 ∑ 𝑬𝑬
𝟑𝟐𝟎
𝟐𝟗𝟎
(𝝀) 𝒙 𝒍 (𝝀) 𝒙 𝑨𝒃𝒔 (𝝀)
Onde: EE (λ) - espectro do efeito eritemal; I (λ) -
intensidade solar espectro; Abs (λ) - absorbância do produto;
FC - fator de correção (= 10). Os valores de EE x I são
constantes e estão representados na Tabela 1.

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
64
Tabela 1. Valores de - espectro do efeito eritemal e intensidade
solar espectro (EE x I) utilizados para calcular o FPS.
Comprimento
de onda (nm) EE x I
290 0.0150
295 0.0817
300 0.2874
305 0.3278
310 0.1864
315 0.0839
320 0.0180
Total 1.0000
Análise estatística
Todos os resultados foram representados pela média
seguida pelo desvio padrão. As análises estatísticas foram
realizadas no software prism Graphpad v. 5.0. As médias
obtidas foram comparadas utilizando ANOVA de uma via.
RESULTADOS E DISCUSSÃO
O alto interesse pelo cultivo da M. oleifera deve-se aos
potenciais usos que apresenta, pois praticamente todas as suas
partes podem ser utilizadas para diversas finalidades (SANTOS
et al., 2015). Sua relevância popular é tanta que em muitos
locais a planta é comumente referida como árvore milagrosa
devido os usos múltiplos que apresenta (IDRIS et al., 2016).
Apesar de basicamente todas as partes de M. oleifera
serem empregadas para alguma finalidade, a folha geralmente

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
65
é a mais utilizada tradicionalmente. As folhas são usadas como
suplemento dietético devido ao seu conteúdo nutricional,
apresentando quantidades consideráveis de minerais
essenciais, vitaminas, aminoácidos e ácidos graxos (KOU et al.,
2018). Por isso, a planta tem sido indicada como uma
alternativa sustentável de suplemento alimentar para aqueles
que sofrem de desnutrição, especialmente nos países em
desenvolvimento (GUPTA et al., 2018). Adicionalmente,
propriedades medicinais importantes, incluindo atividade
antioxidante, anticancerígena e antimicrobiana tem sido
relatadas.
As plantas representam uma importante fonte de
moléculas que absorvem a radiação ultravioleta, em especial de
compostos fenólicos, cuja principal função é proteger as folhas
dos danos causados pela luz, atuando como antioxidantes
(NUNES et al, 2018). Diante das propriedades biológicas de M.
oleifera, este estudo avaliou o potencial fotoprotetor de extratos
orgânicos de folhas a fim de determinar a capacidade deles em
absorver a radiação solar e consequentemente evitar os danos
provocados por ela. O estresse oxidativo e as respostas
inflamatórias induzidas pela radiação ultravioleta podem causar
uma variedade de efeitos prejudiciais na pele, incluindo o
fotoenvelhecimento prematuro e a indução de
imunossupressão e carcinogênese cutânea (BERNARD;
GALLO; KRUTMANN, 2019).

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
66
Figura 1. Moringa oleifera, aspectos gerais (A), folha (B), flores
(C), fruto (D) e sementes (E)
Fonte: Rolim (2007)
Na figura 2 estão representados os perfis de absorção
espectrofotométrica dos extratos de acordo com as
concentrações testadas. Os valores de absorbância foram
dose-dependentes para todos os extratos e portanto, os
maiores valores foram detectados na concentração de 100
µg/mL. No que diz respeito ao solvente utilizado, o extrato
acetato de etila (Mo–AcoEt) foi aquele que apresentou a maior
absorção, no entanto foi na região UVA. Mo–EtOH foi o que
provocou maiores valores de absorbância na região equivalente
ao UVA e UVB. Adicionalmente, todos os extratos promoveram
absorção máxima no comprimento de onde de 400 nm, o qual

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
67
é caracterizado como radiação UVA. Resultados semelhantes
foram obtidos para os extratos em clorofórmio e acetato de etila
das folhas de Neoglaziovia variegata (Arruda), os quais
apresentaram valores de absorção máxima na região do UVA
(OLIVEIRA-JUNIOR, et al., 2015).
O fator de proteção solar (FPS) é um valor numérico
obtido a partir de cálculos e este pode ser dividido em 2 tipos a
depender dos testes utilizados para a verificação desse fator, in
vivo ou in vitro. O FPS in vitro (Figura 3) foi determinado pelo
método espectrofotométrico desenvolvido usando o região
UVB, considerada a região de maior incidência durante o dia
em que as pessoas ficam expostas por mais tempo. Esse
método é capaz de fornecer informações prévias acerca de
formulações fotoprotetoras em estágios iniciais de
desenvolvimento, além de ser de menor custo, rápido,
reprodutível e não necessitar de voluntários. De acordo com a
RDC Nº 30, de 1º de junho de 2012, da Agência Nacional
de Vigilância Sanitária (ANVISA) atribui-se uma
regulamentação técnica aplicada aos produtos e cosméticos
destinados a proteção solar da pele, que, apresenta o fator
mínimo de proteção solar igual a 6 (seis).
Mo-EtOH foi o que apresentou maior fator de
fotoproteção de maneira dose-dependente sendo obtidos
valores que variaram de 0,54 ± 0,05 a 3,37 ± 0,22. Mo-AcoEt foi
o segundo extrato mais ativo, sendo determinados valores de
FPS na faixa de 0,45 a 3,44 ± 0,12. O FPS máximo detectado
para Mo–Hex foi 1,38 ± 0,07 na concentração de 100 µg/mL.
CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
68
Figura 2. Perfil de absorção espectrofotométrica dos extratos
secos das folhas de Moringa oleifera (260 – 400 nm).
Fonte: A autora.
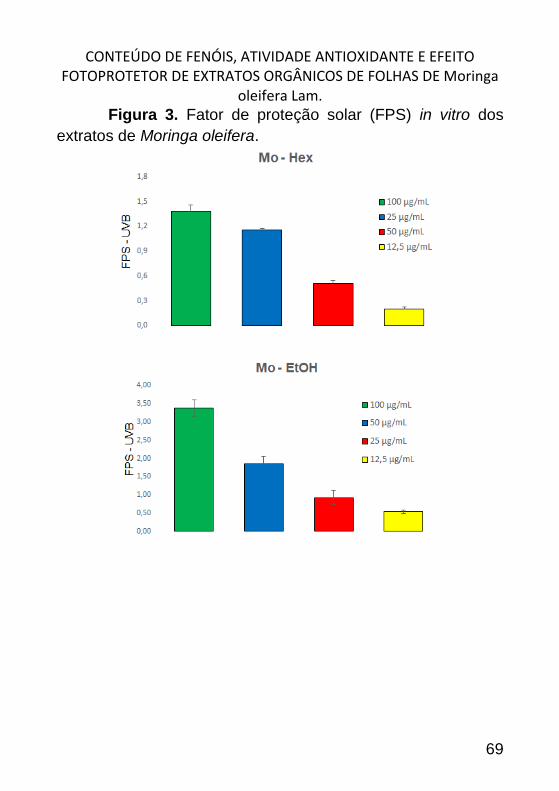
CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
69
Figura 3. Fator de proteção solar (FPS) in vitro dos
extratos de Moringa oleifera.
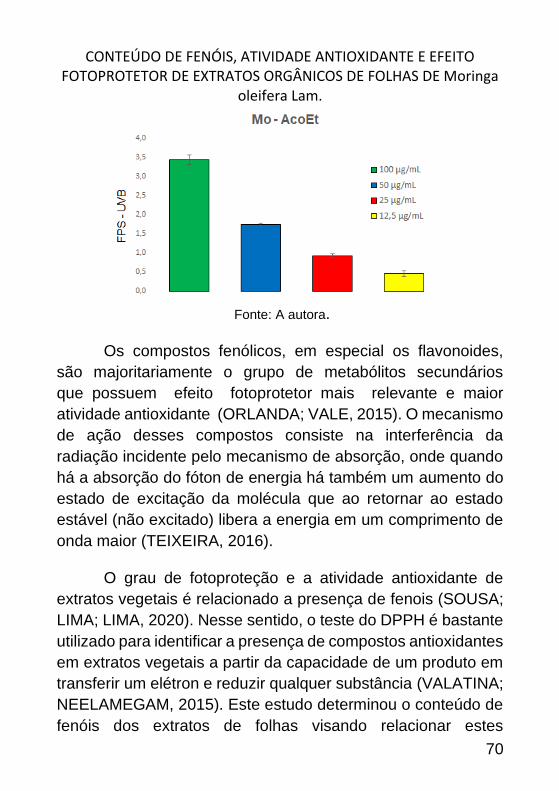
CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
70
Fonte: A autora.
Os compostos fenólicos, em especial os flavonoides,
são majoritariamente o grupo de metabólitos secundários
que possuem efeito fotoprotetor mais relevante e maior
atividade antioxidante (ORLANDA; VALE, 2015). O mecanismo
de ação desses compostos consiste na interferência da
radiação incidente pelo mecanismo de absorção, onde quando
há a absorção do fóton de energia há também um aumento do
estado de excitação da molécula que ao retornar ao estado
estável (não excitado) libera a energia em um comprimento de
onda maior (TEIXEIRA, 2016).
O grau de fotoproteção e a atividade antioxidante de
extratos vegetais é relacionado a presença de fenois (SOUSA;
LIMA; LIMA, 2020). Nesse sentido, o teste do DPPH é bastante
utilizado para identificar a presença de compostos antioxidantes
em extratos vegetais a partir da capacidade de um produto em
transferir um elétron e reduzir qualquer substância (VALATINA;
NEELAMEGAM, 2015). Este estudo determinou o conteúdo de
fenóis dos extratos de folhas visando relacionar estes

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
71
compostos com os valores de fotoproteção obtidos. Atividade
antioxidante, outra propriedade associada a fenóis, foi também
determinada. Mo-EtOH foi o extrato com maior valor de fenóis
com um conteúdo de fenois totais de 1,99 ± 0,21 mg EAT/g. Já
o conteúdo de fenois de Mo - Acoet e Mo – Hex foi de 0,67 ±
0,38 e 0,74 ± 0,01 mgEAT/g, respectivamente. Embora o
conteúdo de fenóis de Mo – Acoet tenha sido ligeiramente maior
que os obtidos para Mo – Hex, os valores obtidos não foram
diferentes dignificativamente. Todos os extratos apresentaram
atividade antioxidade, sendo Mo-EtOH o que apresentou a
maior capacidade de sequestrar o radical DPPH (80,22 ± 0,66).
A atividade antioxidante de Mo-Hex e Mo-AcoEt foi 1,56 e 2,93
vezes menor que aquela de Mo-EtOH, respectivamente (Tabela
3).
As folhas de M. oleifera tem sido referidas como boas
fontes de polifenóis, com valores que variam de 20,9 e 122 mg
GAE/1g, sendo que os principais compostos são flavonoides e
ácidos fenólicos (LEONE et al., 2015). Os dados relacionados
à quantificação desses compostos são bastante diversificados,
uma vez que podem variar de acordo com fatores como
condições ambientais, genética da planta e métodos de
secagem, extração e análise. Apesar disso, é importante
ressaltar que a maioria dos estudos empregam solventes
polares, em especial a água, para aferição dessses compostos.
Adicionalmente, kaempferol, quercetina, isoramnetina e
apigenina são os flavonoides mais comuns nas folhas de M.
oleifera, existindo em abundância como glicosídeos ligados a
um amplo espectro de açúcares (LIN; ZHANG; CHEN, 2018).
Os flavonoides apresentam efeitos protetores contra muitas

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
72
infecções (doenças bacterianas e virais) e doenças
degenerativas, como problemas cardiovasculares e câncer. Já
os ácidos fenólicos, são estudados principalmente por seus
potenciais antioxidante, anti-inflamatório, antimutagênico e
anticâncer (LEONE et al., 2015).
Tabela 3. Conteúdo de fenois totais e atividade antioxidante de
extratos de folhas de M. oleifera.
Extrato Fenois totais
(mg EAT/g)
Atividade
antioxidante
(%AAO)
Mo-EtOH 1,99 ± 0,21 80,22 ± 0,66
Mo-AcoEt 0,67 ± 0,38 27,41 ± 1,00
Mo-Hex 0,74 ± 0,01 51,38 ± 3,82
Conteúdo de fenois: Miligramas equivalentes de ácido tânico por grama
de extrato (mg EAT/g). Curva de calibração: y = 0,0767x + 0,1533 (R² =
0,9947)
O alto potencial antioxidante das folhas de M. oleifera
tem sido amplamente relatado na literatura tanto a partir de
abordagens in vitro como in vivo. Em um estudo recente
realizado in vitro, a atividade antioxidante do extrato
hidroetanólico das folhas de M. oleifera foi investigada. Os
resultados mostraram forte atividade antioxidante da
preparação a partir da utilização do ensaio de eliminação de
óxido nítrico (NO) (IC50 = 120 μg/ml) e ensaio de degradação
de desoxirribose (IC50 = 178 μg/ml). Este estudo mostrou ainda
que o potencial antioxidante das folhas da moringa pode ser
atribuído à presença de conteúdo fenólico total, conteúdo total
de flavonóides, carotenóides, licopeno, ácido ascórbico e

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
73
antocianinas nas folhas (VATS; GUPTA, 2017). In vivo, o
extrato metanólico das folhas de M. oleifera (250 mg/kg) foi
administrado por seis semanas em ratos Wistar machos
diabéticos. Os resultados mostraram alta capacidade
antioxidante do extrato e melhora nos marcadores bioquímicos
séricos do estresse oxidatico, tais como os níveis de catalase
(CAT), glutationa peroxidase (GPx), superóxido dismutase
(SOD), glutationa (GSH). Além disso, os níveis de peroxidação
lipídica (MDA) foram reduzidos em ratos não diabéticos e
diabéticos após o tratamento com o extrato das folhas quando
comparados ao controle normal (OMODANISI; ABOUA;
OGUNTIBEJU, 2017). Lin, Zhang e Chen (2018) afirmam que a
capacidade de neutralizar espécies reativas de oxigênio está
diretamente relacionada a prevenção de doenças não
transmissíveis (DNTs), como obesidade e diabetes. Saleem,
Saleem e Akhtar (2018) também avaliaram o potencial
antioxidante das folhas de moringa através do teste do DPPH.
Verificou-se que todos os extratos mostraram atividade
antioxidante em concentrações inferiores a 0,1563 mg/ml. Na
maior concentração (0,1563 mg/ml), o efeito antioxidante
observado para os extratos metanólico (88,50 ± 0,61%) e
aquoso (84,46 ± 0,34%) foi comparado ao ácido ascórbico
(82,55 ± 0,35%).
Ademais, a capacidade de neutralizar espécies reativas
de oxigênio (EROs) está diretamente relacionada a prevenção
de doenças não transmissíveis (DNTs), como obesidade e
diabetes. Nesse sentido, as atividades farmacológicas pelas
quais a moringa é reconhecida mundialmente são atribuídas,

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
74
dentre outros fatores, à presença de compostos antioxidantes,
especialmente os compostos fenólicos.
CONCLUSÕES
Extratos orgânicos obtidos a partir das folhas de M.
oleifera com diferentes soventes não apresentaram valores de
fotoproteção satisfatórios em comparação aos critérios
estabelecidos pela Agência Nacional de Vigilância Sanitária
(ANVISA). O baixo conteúdo de fenois totais nos extratos pode
ser relacionado a baixa atividade fotoprotetora. A alta
capacidade de sequestro do radical DPPH de Mo-EtOH
estimula a produção de novos extratos através da utilização de
outros solventes e métodos de extração.
REFERÊNCIAS BIBLIOGRÁFICAS
BERNARD, J. J.; GALLO, R. L.; KRUTMANN, J. Photoimmunology: how ultraviolet radiation affects the immune system. Nature Reviews Immunology, v. 19, n. 11, p. 688-701, 2019. BOBO‐GARCÍA, G.; DAVIDOV‐PARDO, G.; ARROQUI, C.; VÍRSEDA, P.;
MARÍN‐ARROYO, M. R.; NAVARRO, M. Intra‐laboratory validation of microplate methods for total phenolic content and antioxidant activity on polyphenolic extracts, and comparison with conventional spectrophotometric methods. Journal of the Science of Food and Agriculture, v. 95, n. 1, p. 204-209, 2015. CARVALHO, J. C. S.; GARCIA, P. S. P.; VIGNOL, S. R.; PEDRIALI, C. A. Estudo do impacto da utilização de ativos vegetais em fotoprotetores. InterfacEHS – Saúde, Meio Ambiente e Sustentabilidade, v. 10, n. 2, p. 62-82, 2015. COZ-BOLAÑOS, X.; CAMPOS-VEGA, R.; REYNOSO-CAMACHO, R.; RAMOS-GÓMEZ, M.; LOARCA-PIÑA, G. F.; GUZMÁN-MALDONADO. Moringa infusion (Moringa oleifera) rich in phenolic compounds and high antioxidant capacity attenuate nitric oxide pro-inflammatory mediator in vitro. Industrial Crops and Products, v. 118, p. 95-101, 2018.

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
75
DABA, M. Miracle tree: A review on multi-purposes of Moringa oleifera and its implication for climate change mitigation. J. Earth Sci. Clim. Change, v. 7, n. 4, p. 1-5, 2016. GUPTA, S.; JAIN, R.; KACHHWAHA, S.; KOTHARI, S. L. Nutritional and medicinal applications of Moringa oleifera Lam.—Review of current status and future possibilities. Journal of Herbal Medicine, v. 11, p. 1-11, 2018. KOU, X.; LI, B.; OLAYANJU, J. B.; DRAKE, J. M.; CHEN, N. Nutraceutical or pharmacological potential of Moringa oleifera Lam. Nutrients, v. 10, n. 3, p. 343, 2018. KORKINA, L.; KOSTYUK, V.; POTAPOVICH, A.; MAYER, W.; TALIB, N.; DE LUCA, C.Secondary Plant Metabolites for Sun Protective Cosmetics: From Pre-Selection to Product Formulation. Cosmetics, v. 5, n. 2, p. 32, 2018. LEONE, A.; SPADA, A.; BATTEZZATI, A.; SCHIRALDI, A.; ARISTIL, J.; BERTOLI, S. Cultivation, genetic, ethnopharmacology, phytochemistry and pharmacology of Moringa oleifera leaves: An overview. International journal of molecular sciences, v. 16, n. 6, p. 12791-12835, 2015. LIN, M.; ZHANG, J.; CHEN, X. Bioactive flavonoids in Moringa oleifera and their health-promoting properties. Journal of functional foods, 47, p. 469-479, 2018. LOPES, F. M.; CRUZ, R. O.; BATISTA, K. A. Radiação ultravioleta e ativos utilizados nas formulações de protetores solares. Ensaios e Ciência: Ciências Biológicas, Agrárias e da Saúde, v. 16, n. 4, p. 183-199, 2012. LUZ, L. A.; SANTOS, A. F. S.; PONTUAL, E. V.; NAPOLEÃO, T. H.; PAIVA, P.; COELHO, L. C. B. B. Moringa oleifera: resource management and multiuse life tree. Advances in Research, 4(6), 388-402, 2015 MANOJ, M.; RANJITHA, G. M. K.; PHYTOREMEDIATION of E-Waste using Nicotiana Tabacum & Moringa oleifera. International Journal for Scientific Research & Development. v. 7, p. 339-342, 2020. NASCIMENTO, K. S.D.; Compostos fenólicos, capacidade antioxidante e propriedades físico-químicas de méis de Apis mellifera do estado do Rio Grande do Sul. 2016. Tese de Doutorado. Universidade de São Paulo. NUNES, A. R.; RODRIGUES, A. L. M.; QUEIRÓZ, D. B.; VIEIRA, I. G. P.; NETO, J. F. C.; JUNIOR, J. T. C.; COUTINHO, H. D. M. Photoprotective potential of medicinal plants from Cerrado biome (Brazil) in relation to phenolic content and antioxidant activity. Journal of Photochemistry and Photobiology B: Biology, v. 189, p. 119-123, 2018. OLIVEIRA-JUNIOR, R. G.; SOUZA, G. R.; GUIMARÃES, A. L.; OLIVEIRA, A. P.; SOUZA, A. C.; SILVA, J. C.; CASTRO, R. N.; Photoprotective, antibacterial activity and determination of phenolic compounds of Neoglaziovia variegata (Bromeliaceae) by high performance liquid

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
76
chromatography-diode array detector (HPLC-DAD) analysis. African Journal of Pharmacy and Pharmacology, v. 9, n. 22, p. 576-584, 2015. OMODANISI, E. I.; ABOUA, Y. G.; OGUNTIBEJU, O. O. Assessment of the anti-hyperglycaemic, anti-inflammatory and antioxidant activities of the methanol extract of Moringa oleifera in diabetes-induced nephrotoxic male wistar rats. Molecules, v. 22, n. 4, p. 439, 2017. ORLANDA, J.F.F.; VALE, V.V. Análise fitoquímica e atividade fotoprotetora de extrato etanólico de Euphorbia tirucalli Linneau (Euphorbiaceae). Revista Brasileira de Plantas Medicinais, v.17, n.4, supl. I, p.730-736, 2015. PIRES, J.; TORRES, P. B.; SANTOS, D. Y. A. C.; CHOW, F. Ensaio em microplaca do potencial antioxidante através do método de sequestro do radical livre DPPH para extratos de algas. Instituto de Biociências, Universidade de São Paulo, São Paulo, p. 1- 6, 2017. POPOOLA, J. O.; OBEMBE, O. O. Local knowledge, use pattern and geographical distribution of Moringa oleifera Lam.(Moringaceae) in Nigeria. Journal of Ethnopharmacology, 150, n. 2, p. 682-691, 2013. ROLIM, Lucíola Abílio Diniz Melquiades de Medeiros. Purificação da lectina de sementes de Moringa oleifera (WSMoL) e avaliação da genotoxidade. 2007. Dissertação de Mestrado. Universidade Federal de Pernambuco.. SALEEM, A.; SALEEM, M.; AKHTAR, M. F. Antioxidant, anti-inflammatory and antiarthritic potential of Moringa oleifera Lam: An ethnomedicinal plant of Moringaceae family. South African Journal of Botany, v. 128, p. 246-256, 2020. SANGAR, V.; SAMANT, L.; PAWAR, S.; VAIDYA, S.; CHOWDHARY, A. In silico approach to combat HIV using phytoconstituents of Moringa oleifera Lam. Journal of Chemical and Pharmaceutical Research, v. 7, n. 12, p. 997-1021, 2015. SANTOS, A. F.; LUZ, L. A.; PONTUAL, E. V.; NAPOLEÃO, T. H.; PAIVA, P. M.; COELHO, L. C. Moringa oleifera: resource management and multiuse life tree. Advances in Research, v. 4, n. 6, p. 388-402, 2015. SCHNEIDER, G.; FIGUEROA, F. L.; VEGA, J.; CHAVES, P.; ÁLVAREZ G. F.; KORBEE, N.; BONOMI-BARUFI, J. Photoprotection properties of marine photosynthetic organisms grown in high ultraviolet exposure areas: Cosmeceutical applications. Algal Research, v. 49, p. 101956, 2020 SECCO, G. G.; BELTRAME, B. M.; SCHWANZ, M. Avaliação in vitro do fator de proteção solar (FPS) de cosméticos fotoprotetores manipulados. Infarma-Ciências Farmacêuticas, v. 30, n. 4, p. 242-249, 2018.

CONTEÚDO DE FENÓIS, ATIVIDADE ANTIOXIDANTE E EFEITO FOTOPROTETOR DE EXTRATOS ORGÂNICOS DE FOLHAS DE Moringa
oleifera Lam.
77
SEIFU, E.; TEKETAY, D. Introduction and expansion of Moringa oleifera Lam. in Botswana: Current status and potential for commercialization. South African Journal of Botany, v. 129, p. 471-479, 2020. SILVA L.; ALESSANDRA D. S.; GUIMARÃES, R.; LIMA, E. N. Incremento da atividade fotoprotetora e antioxidante de cosméticos contendo extratos vegetais da caatinga. Brazilian Journal of Natural Sciences, v. 3, n. 1, p. 225-225, 2020. TEIXEIRA, M. S. M. AVALIAÇÃO DA ATIVIDADE FOTOPROTETORA DE FORMULAÇÃO COSMÉTICA CONTENDO A ASSOCIAÇÃO ENTRE FRAÇÃO EM CLOROFÓRMIO DE Garcinia cambogia Desr. (Clusiaceae) E FILTRO SINTÉTICO DE AMPLO ESPECTRO. Trabalho de Conclusão de Curso(graduação) – Universidade Federal de Juiz de Fora. Faculdade de Farmácia e Bioquímica, 2016. VALANTINA, R.S., NEELAMEGAM, P.. Selective ABTS and DPPH- radical scavenging activity of peroxide from vegetable oils. International Food Research Journal, 22, 289-294, 2015. VATS, S.; GUPTA, T. Evaluation of bioactive compounds and antioxidant potential of hydroethanolic extract of Moringa oleifera Lam. from Rajasthan, India. Physiology and molecular biology of plants, v. 23, n. 1, p. 239-248, 2017. .

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
78
CAPÍTULO 4
O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
Caio César da Silva Guedes 1
Robson Raion de Vasconcelos Alves 1
Matheus Cavalcanti de Barros 2
Juliane Nancy de Oliveira Silva 3
Anderson Felipe Soares de Freitas 4 1 Doutorando no Programa de Pós-Graduação em Ciências Biológicas, UFPE; 2
Mestrando no Programa de pós-graduação em Ciências Biológicas, UFPE 3 Mestranda no Programa de pós graduação em Bioquímica e Fisiologia, UFPE;
4Graduando do curso formação pedagógica para graduados não licenciados - Biologia,
Cruzeiro do Sul virtual. [email protected]
RESUMO: O câncer é um conjunto de doenças causadas por mutações genéticas nas células, tais doenças são caracterizadas por alta taxa de divisão celular, onde estas células, podem agrupar-se resultando em tumores com elevada capacidade de invasão tecidual e metastática. As lectinas são proteínas hemaglutinantes de origem não imunológica, que possuem a capacidade de se ligar reversivelmente e especificamente a carboidratos, possuindo assim diversas atividades como, por exemplo, ação antitumoral. O objetivo desse trabalho é ressaltar o potencial das lectinas como aliadas no diagnóstico do câncer através da revisão bibliográfica de artigos científicos previamente selecionados nas plataformas: Google acadêmico, ScienceDirect e PubMed. As lectinas têm obtido resultados satisfatórios em estudos para detecção de biomarcadores do câncer, esse potencial é permitido através da capacidade de reconhecer carboidratos específicos em moléculas que aumentam seus níveis séricos durante a carinogênese ou através de ligação direta a glicanos presentes na superfície de células cancerosas tornando possível o

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
79
reconhecimento das células cancerígenas para diagnóstico, acompanhamento e estratégias terapêuticas. Palavras-chave: Diagnóstico do Câncer. Aglutininas. Biomarcadores. INTRODUÇÃO
Como definição, o câncer compreende um grupo de
mais de 100 doenças que tem como característica comum o
crescimento desordenado de células, tais células são dotadas
de grande capacidade de disseminar-se para outros tecidos e
órgãos adjacentes ao sítio que foi afetado inicialmente no
organismo. O câncer é considerado um problema de saúde
pública, enfrentado tanto pelo sistema de saúde brasileiro
como em outros paises em vista de sua amplitude
epidemiológica, social e econômica. A incidência crescente
de casos de neoplasia tem ocasionado uma transformação no
perfil epidemiológico da população, seja pelo aumento da
exposição aos fatores cancerígenos, pelo envelhecimento
populacional, pelo aprimoramento das tecnologias para o
diagnóstico, como também pela elevação do número de óbitos
por câncer (BATISTA; DE MATTOS; DA SILVA, 2015.)
O câncer é o principal problema de saúde pública no
mundo e está situado entre as quatro principais causas de
morte prematura (antes dos 70 anos de idade) na maioria dos
países. A incidência e a mortalidade por câncer vêm
aumentando no mundo, em parte pelo envelhecimento, pelo
crescimento populacional, como também pela mudança na
distribuição e na prevalência dos fatores de risco de câncer,
especialmente aos associados ao desenvolvimento
socioeconômico. Verifica-se uma transição dos principais tipos

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
80
de câncer observados nos países em desenvolvimento, com um
declínio dos tipos de câncer associados a infecções e o
aumento daqueles associados à melhoria das condições
socioeconômicas com a incorporação de hábitos e atitudes
associados à urbanização sedentarismo, alimentação
inadequada, entre outros (BRAY et al., 2018).
Com o avanço tecnológico novos estudos sempre são
realizados para desenvolver novos métodos para aprimoração
e diferenciação no diagnóstico das mais diversas patologias,
nesse aspecto o uso de biomeléculas apresenta um futuro
promissor.
As lectinas são proteínas ou glicoproteinas pertencentes
a uma diversificada superfamília, onde possuem como principal
característica a capacidade de reconhecer carboidratos de
modo específico e seletivo. Essas proteínas podem ser
encontradas em diversos organismos, desde, vírus, bactérias,
e fungos a vegetais e animais. A sua habilidade de reconhecer
uma grande quantidade de glicanos, as torna mediadoras em
diversos processos biológicos, como migração celular, defesa
imunológica, infecção por bactérias, vírus e protozoários
(OLIVEIRA, 2018).
A disponibilidade de um grande número de lectinas e
seu potencial de reconhecer diferentes carboidratos tem
introduzido essas proteínas como ferramenta de
reconhecimento em diversas pesquisas biotecnológicas.
Graças à especificidade para mono e oligossacarídeos
expressos na superfície celular, as lectinas podem ser utilizadas
para tipagem de células sanguíneas; como inseticida, ao
prejudicar o desenvolvimento e mortalidade de larvas,
bactericida e fungicida, e vêm sendo investigadas como
antivirais, por impedir a invasão e replicação viral do HIV.

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
81
Também têm sido utilizadas em matrizes de afinidade para
reconhecimento e purificação de glicoproteínas e no
desenvolvimento de sistemas de liberação de drogas em
mucosas (SILVA, 2019).
Nesse contexto, o objetivo desse trabalho é demonstrar
o potencial uso das lectinas como possíveis novas abordagens
para o diagnóstico do câncer já que devido à especificidade
com que as lectinas reconhecem os glicanos permite a
identificação de alterações tumor-específicas, levando à
descoberta de novos biomarcadores glicosilados, assim, novos
estudos demonstram a utilização dessas proteínas no
diagnóstico dos mais diversos tipos de câncer, através de
diferentes metodologias que partem desde a incorporação em
matrizes cromatográficas até imunoensaios o que possibilitando
um novo repertório, eficaz e específico para o diagnóstico de
neoplasias.
MATERIAIS E MÉTODO
O objetivo desse trabalho é apresentar uma revisão
bibliográfica sistemática a respeito do potencial anti-câncer das
lectinas. Os artigos, trabalhos de conclusão de curso,
dissertações de mestrado e teses de doutorado foram
selecionados a partir de plataformas digitais como sciencediret,
scielo, pubmed e google acadêmico. Os critérios de inclusão
foram artigos que estivessem nas línguas portuguesa e inglesa,
publicados entre 2015 e 2020. As palavras chaves utilizadas na
busca foram: Anti-tumoral Lectin, lectina anti-tumoral, câncer,
lectina anti-câncer e anti-cancer lectin. Os critérios de exclusão
adotados foram artigos que não se enquadrassem nos anos de

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
82
2015 a 2020 e artigos que não possuíam conteúdo relacionado
com o trabalho proposto.
RESULTADOS E DISCUSSÃO
O Câncer
A crescente mortalidade por neoplasias malignas vem
crescendo consideravelmente ao longo das últimas décadas o
que posiciona o câncer como a segunda principal causa de
óbito no mundo tendo levado a óbito 9,6 milhões de
pessoas no ano de 2018, em muitos países pobres e em
desenvolvimento, incluindo o Brasil as projeções sinalizam que
ocupará o primeiro lugar nas próximas décadas (INCA, 2017;
TEIXEIRA et al., 2018; CARVALHO; PAES, 2019; VERONEZ et
al., 2019). Para os anos de 2020-2022, foi estimada a
ocorrência de mais de 600 mil novos casos de câncer (figura 1),
para cada ano no Brasil (INCA, 2019).
Dentre vários tipos de câncer os mais incidentes são
os cânceres de próstata, pulmão, mama feminina, cólon e reto,
colo de útero, estômago e esôfago. Mesmo diante de avanços
científicos voltados à prevenção e detecção precoce do câncer,
essa doença ainda é um problema global de saúde pública, com
elevado ônus biopsicossocial e econômico (VERONEZ, 2019).
Diariamente as células presentes no nosso organismo
podem sofrer lesões em seu material genético ( >20.000) bem
como erros em seu processo de divisão celular (>10.000)
(MOTA, 2019). Para conter isso existe um complexo e eficiente
sistema capaz de reparar esses danos, sendo capaz de
impedir a propagação desses erros, permitindo com que as
céluas em divisão se mantenham em estado normal. Caso esse
O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
83
sistema não seja eficiente no processo de reparação, as células
descendentes transmitirão o erro e antígenos novos passam a
ser expressos na superfície dessas células.
Figura 1. Estimativa de surgimento de novos casos de câncer
para os anos de 2020-2022 pelo INCA.

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
84
Fonte: INCA, 2019
Esses antígenos podem corresponder a proto-
oncogenes mutados, genes supressores de tumores,
glicoproteínas, antígenos produzidos por vírus oncogênicos
dentre tantos outros. O termo “câncer” é utilizado para se referir
ao conjunto de doenças que tem por características
crescimento desordenado e descontrolado de células dotados
de capacidade invasiva em tecidos e órgãos (INCA, 2019).
Essas doenças podem ser multifatoriais podendo surgir em
virtude de alterações no genoma celular ou esse gênese pode
se dar a partir da combinação de fatores genéticos e fatores
externos como agentes cancerígenos de natureza física,
química e biológica.
Exitem dois tipos de genes que estão diretamente
envolvidos na gênese e no desenvolvimento de um carcinoma,
esses genes podem ser agrupados de acordo com sua natureza
funcional em dois grupos que são os proto-oncogenes e os
genes supressores tumorais, onde alterações em genes
distintos podem culminar na progressão tumoral (KLAUNIG,
2020). No decorrer da tumorigênese, células normais, podem
começar a expressar um perfil proteico em níveis anormais ou
estruturalmente diferentes podendo, a partir desse perfil
“anormal” serem distinguidas das células saudáveis e assim
podendo serem eliminadas pelo sistema imunológico
(ERRANTE, 2020).
As células cancerígenas podem escapar da destruição
pelo sistema imune, ou seja, evadir ao reconhecimento imune
por meio de diversos mecanismos. Isso é permitido devido as
células tumorais terem sido originadas a partir das células do
hospedeiro, conferindo certo grau de semelhança com as
células normais, por este motivo, existe uma dificuldade no

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
85
reconhecimento para que a essa eliminação seja efetuada
(ABBAS; LICHTMAN; PILLAI, 2015).
Devido à complexidade e à heterogeneidade dos
diversos tipos de tumores malignos, que apresentam várias
mutações genéticas na patogênese do câncer, o tratamento
dessa patologia enfrenta limitações e muitos desafios, uma vez
que esses tumores são altamente adaptáveis, onde as células
tumorais podem criar novas rotas de sinalização para contornar
a cascata de morte celular e criar resistência ao tratamento
(SCHMIDT; EFFERTH, 2016). O tratamento quimioterápico, na
maioria dos casos, é a primeira opção escolhida, porém possui
vários efeitos colaterais indesejados e danosos às células
normais e, consequentemente, ao paciente. Nesse sentido, a
biotecnologia vem auxiliar na busca por biomoléculas de origem
natural que possuam seletividade para as células cancerígenas,
como é o caso de diversas lectinas (OLIVEIRA, 2018).
Lectinas
Lectinas (do latim "lectus", que significa selecionado) são
proteínas hemaglutinantes de origem não imunológica e de
ocorrência natural que tem a capacidade de se ligar
reversivelmente e especificamente a carboidratos sejam eles
mono, oligo ou polissacarídeos. São proteínas amplamente
difundidas na natureza, sendo encontradas desde seres
unicelulares até organismos superiores (DE SCHUTTER; VAN
DAMME, 2015; MANNING et al., 2017). As lectinas são
capazes de discriminar diferentes estruturas de glicanos e são,
portanto, ferramentas muito úteis para glicoanálise, elas estão
envolvidas em eventos fisiológicos cruciais de interações
proteína-carboidrato, como adesão, migração e interação

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
86
celular (BROWN; WILLMENT; WHITEHEAD, 2018;
VILLRINGER et al., 2018).
A origem não imunológica das lectinas distinguem-as de
anticorpos anticarboidratos, que também aglutinam células.
Diferente dos anticorpos, que são estruturalmente similares, as
lectinas diferenciam-se entre si pela composição aminoacídica,
requerimento de metais, peso molecular e estrutura
tridimensional (OBAID et al., 2015; GAUTAM et al., 2018).
A habilidade de ligar carboidratos de lectinas tem sido
atribuída à existência de uma região denominada “sítio de
ligação a carboidrato” ou “domínio de reconhecimento de
carboidratos, a interação entre lectina e carboidrato pode ser
realizada principalmente através de pontes de hidrogênio (onde
os grupos hidroxila constituintes dos açúcares são capazes de
doar e receber moléculas de hidrogênio, coordenação por
grupos metil, forças de Van Der Waals e interações hidrofóbicas
(SILVA, 2017).
As lectinas podem ser divididas em vários grupos de
acordo com a sua estrutura molecular (como ilustrado na figura
2) as merolectinas, hololectinas, quimerolectinas e
superlectinas. As merolectinas possuem apenas um domínio de
renhecimento de carboidrato (DRC) o qual se liga a
carboidratos simples, são proteínas pequenas e sua natureza
monovalente as impede de precipitar glicoconjugados,
aglutinar células ou terem atividade catalítica. As hololectinas,
que compreendem o maior grupo de lectinas, possuem dois
domínios de reconhecimento de carboidratos com elevada
homologia e são capazes de aglutinar células ou precipitar
glicoconjugados; são consideradas as hemaglutininas
clássicas. As quimerolectinas, são proteínas de fusão que
dispõem de pelo menos um DRC e outro domínio com atividade

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
87
catalítica que age de forma totalmente independente do DRC,
dependendo do numero de DRC’s as quimerolectinas podem
ser comportar como uma merolectina ou hololectina (PINEDO
et al, 2015; LAVANYA et al. 2017). As superlectinas são um
grupo de quimerolectinas possuidoras de dois ou mais DRC's
distintos, conferindo a essa proteína a especificidade por
carboidratos diferentes (SAGAR e DHALL, 2018).
Figura 2. Representação dos tipos de lectinas e exemplo de
membro de cada tipo Heveína (merolectina), DWL-3SH3
(hololectina), PPL2-2GSJ (quimerolectina) e Lectina de Musa
acuminata – 3MIT (superlectina).
Fonte: Osterne (2016)
Algumas lectinas apresentam atividade biológica apenas
na presença de íons bivalentes enquanto aquelas que não

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
88
necessitam de íons metálicos já possuem a conformação
estrutural necessária para o reconhecimento aos carboidratos.
Nas lectinas de natureza glicoproteica, a porção glicídica
geralmente aumenta a estabilidade da lectina dificultando a sua
degradação proteolítica e a desnaturação por temperatura e pH
além de influenciar na interação da lectina com outras
moléculas e na solubilidade da proteína quando em solução
aquosa (GAUTAM et al., 2020.; KILPATRICK, 1986).
As lectinas são capazes de formar pontes entre
carboidratos ou glicoproteínas, que se apresentam em solução
ou ligadas a membrana (MARQUES NETO, 2015). A detecção
da presença de lectinas em uma amostra é realizada através do
ensaio de hemaglutinação (figura 3A), onde a preparação que
contém essas proteínas é incubada com uma suspensão de
eritrócitos. Os eritrócitos utilizados de humanos ou de outros
animais podem ser tratados enzimaticamente, por exemplo com
tripsina e papaína, ou quimicamente, por exemplo com
glutaraldeído ou formaldeído, visando o aumento da interação
com a lectina. A formação de uma malha de aglutinação entre
os eritrócitos indica positividade no teste e a atividade
hemaglutinante (título) é dada pelo inverso da titulação do
último poço que apresentou a malha de aglutinação (BRITO et
al., 2017).
Para a confirmação da natureza lectínica da atividade
hemaglutinante, realiza-se o teste de inibição da atividade
hemaglutinante por carboidratos (figura 3B) onde a lectina é
inicialmente incubada com soluções de diferentes carboidratos
previamente à adição da suspensão de eritrócitos. A inibição é
confirmada pela ausência de hemaglutinação, essa inibição é
causada porque os domínios de reconhecimento de
carboidratos das lectinas são ocupados pelos carboidratos

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
89
livres em solução, diminuindo a interação das lectinas com os
carboidratos presentes na membrada dos eritrócitos (LIMA et
al., 2017).
Figura 3. Representação de atividade hemaglutinante de
lectinas (A) e representação de teste de inibição de atividade
hemaglutinante por carboidratos (B)
Fonte: Paiva et al., 2011.
Dentre algumas atividades biológicas das lectinas
vegetais têm-se o armazenamento ou transporte de
carboidratos em sementes, atividade inseticida, atividade
antitumoral, atividade anti-inflamatória e atividade
antimicrobiana (PROCOPIO et al., 2017; REYES-MONTAÑO e
VEGA-CASTRO, 2018; PEREIRA; SILVA, 2019). As lectinas
além de potenciais agentes terapêuticos, são utilizadas como
ferramentas para a detecção, isolamento e caracterização
parcial de glicoproteínas, além de possibilitar a identificação de
alteração no perfil glicídico presente no glicocálice da superfície
celular durante o desenvolvimento, diferenciação e
A B

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
90
transformação tumoral através da sua capacidade de se ligar
aos antígenos associados a tumores específicos com elevada
afinidade (HASHIM; JAYAPALAN; LEE, 2017).
LECTINAS COM POTENCIAL ANTI-CÂNCER
As lectinas podem apresentar dois tipos de atividade
anti-câncer, a primeira está relacionada com a atividade anti-
tumoral e a segunda com o potencial uso dessas proteínas
como biomarcador na detecção dessas células defeituosas
(OLIVEIRA, 2018). Essas proteínas têm sido utilizadas em
glicoanálise para identificar alterações na composição de
glicanos e elucidar os mecanismos fisiopatológicos
relacionados a essas alterações, bem como as modificações
em genes reguladores das enzimas que participam da
glicosilação (MATSUDA et al., 2015; LLOP et al., 2016).
Aumento ou redução na ramificação dos glicanos e na
expressão de monossacarídeos como ácido siálico, fucose e
manose em diferentes estágios de transformação tumoral são
alterações comumente relacionadas à progressão do câncer e
metástase (PIHÍKOVÁ; KASÁK; TKAC, 2015; CHANG et al.,
2019).
A maioria dos biomarcadores glicoproteicos utilizados no
monitoramento clínico da terapia são avaliados em suas
quantidades totais de proteína, desse modo, a descoberta de
novos biomarcadores, que possibilitem diferenciar padrões de
glicosilação, é importante para o aumento da especificidade e
da seletividade no prognóstico da patologia (OLIVEIRA, 2018).
Devido a sua especificidade o emprego das lectinas na
descoberta de biomarcadores pode trazer algumas vantagens
sobre os métodos padrão, uma vez que elas são capazes de

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
91
detectar peptídeos expressos em tumores a níveis baixos em
fluidos corporais (SILVA, 2019). Algumas lectinas já tem sido
utilzadas com a finalidade de detecção de perfil de glicosilação
em células oncogênicas. Os tipos de ensaio baseados em
lectinas, comumente usados para a pesquisa de
biomarcadores, são: (a) a cromatografia de afinidade com
lectina imobilizada, (b) ensaios histoquímicos, (c) eletroforese,
(c) ensaios ELISA e, mais recentemente, (d) o formato de
microarranjos (HAAB; YUE, 2011).
Imunoensaio utilizando a lectina de Sambucus nigra,
SNA, desenvolvido para analisar o risco relacionado ao câncer
de próstata (LLOP et al., 2016) e as lectinas de Maackia
amurensis MAL-I e MAL- foi desenvolvido para comparar a
expressão de ácido siálico no PSA (um marcador para
diagnóstico do câncer de próstata), presente em soros de
câncer de próstata e em soros não cancerosos (BHANUSHALI
et al., 2016). Tais ensaios são capazes de avaliar a expressão
de ácido siálico no PSA os resultados do estudos sugerem que
os padrões de expressão de ácido siálico do PSA são mais
heterogêneos no CaP, podendo discriminar entre amostras
cancerosas e não cancerosas (SILVA 2019).
A lectina vegetal extraída das sementes da Lens culinaris
(LCA), ligante de fucose α 1,6, é um caso de sucesso como
ferramenta em kits de diagnóstico para o carcinoma
hepatocelular esta proteína tem uma afinidade específica à
isoforma glicoproteica da alfa-fetoproteína (AFP-L3), que é
específica para tumores oncogênicos ( (PERVIN et al., 2015;
COULIBALY e YOUAN, 2017).
Um estudo utilizando a aglutinina-I de Ricinus communis
demonstrou que essa lectina tem a capacidade de se ligar a
células de câncer de mama triplo negativo, onde essa ligação

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
92
está diretamente proporcional a capacidade metastática desse
tumor (ZHOU et al., 2015). Também foi desenvolvido um
imunoensaio tipo sanduíche de anticorpo – lectina para analisar
a glicosilação do antígeno CA 15-3 em pacientes com câncer
de mama, utilizando o anticorpo anti- CA 15-3 para capturar o
antígeno no soro, e a lectina concanavalina A (ConA), para
analisar a glicosilação do antígeno capturado. Os resultados
mostraram que o nível de glicosilação de CA 15-3 aumenta com
o grau de agressividade do câncer, mostrando potencial para
distiguir entre os estágios I, IIA, IIB, III, e doença benigna da
mama, além de ser uma abordagem mais sensível que a
detecção apenas dos níveis de CA 15-3 (CHOI et al., 2018).
As lectinas extraídas das sementes da Maackia
amurensis, denominadas leucoaglutinina (MAL) e
hemaglutinina (MAH), destacam-se pela especificidade de
ligação ao ácido siálico. Recentemente, estudos revelaram que
a MAL reconhece os ácidos siálicos α 2,3 ligados, presentes
principalmente em N-glicanos, e é composta por dois
monômeros unidos por pontes dissulfetos. Essa lectina pode
apresentar diversas glicoformas, pois cada monômero possui
quatro sítios de N-glicosilação (KUMAR; SUROLIA, 2017). A
aglutinina-I de Uelx Europaeus (UEA-I) em estudo demonstrou
um potencial uso na detecção de câncer de coloretal por
imagem (TIAN et al., 2018).
Um sistema de ELISA utilizando a lectina de Pholiota
squarrosa demonstrou ter capacidade de detecção de
haptoglobina para diagnóstico de câncer de pâncreas podendo
ser uma ferramenta útil no diagnóstico complementar de CA19-
9 (KUSAMA et a., 2017). Uma lectina ligadora de galactose do
soro de portadores de câncer renal foi isolada, purificada e
caracterizada, sugerindo o surgimento de um novo biomarcador

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
93
que pode auxiliar no diagnóstico dessa condição (JASIM;
HASAN; HUSAIN, 2015). Estudos ainda demonstram o possível
uso dessas proteínas na histoquímica, Lim e colaboradores em
2020 reportaram a possível utilização de lectina de Helix
pomatia como sonda para detecção de doença cervical de alto
grau.
CONCLUSÕES
Através dessa revisão, podemos concluir que as lectinas
representam um grande potencial biotecnológico pois podem
ser grandes aliadas quando se trata de detecção de diversos
tipos de câncer seja através de detecção de biomarcadores
como até mesmo pela interação com carboidratos presentes
diretamente na superfície celular das células tumorais ainda
que esses tumores apresentem grande grau de
heterogenicidade.
REFERÊNCIAS BIBLIOGRÁFICAS
ABBAS, A. K.; LICHTMAN, A. H. H.; PILLAI, S. Imunologia celular e
molecular. Elsevier Brasil, 2015.
BATISTA, D.R.R; DE MATTOS, M; DA SILVA, S. F. Convivendo com o
câncer: do diagnóstico ao tratamento. Revista de Enfermagem da UFSM,
v. 5, n. 3, p. 499-510, 2015.
BRAY, F. et al. Global cancer statistics 2018: GLOBOCAN estimates of
incidence and mortality worldwide for 36 cancers in 185 countries. CA: a
cancer journal for clinicians, Hoboken, v. 68, n. 6, p. 394-424, Nov. 2018.
BRITO, J. S. et al. Lectin from inflorescences of ornamental crop Alpinia
purpurata acts on immune cells to promote Th1 and Th17 responses, nitric
oxide release, and lymphocyte activation. Biomedicine &
Pharmacotherapy, v. 94, p. 865-872, 2017.

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
94
BROWN, G. D.; WILLMENT, J. A.; WHITEHEAD, L. C-type lectins in
immunity and homeostasis. Nature Reviews Immunology, v. 18, p. 374–
389, 2018.
CARVALHO,J.B; PAES, N. A. Taxas de mortalidade por câncer corrigidas
para os idosos dos estados do Nordeste brasileiro. Ciência & Saúde
Coletiva, v. 24, p. 3857-3866, 2019.
CAVADA, B. S. et al. Reviewing Mimosoideae lectins: A group of under
explored legume lectins. International Journal of Biological
Macromolecules, 2020.
CHANG, S. C. et al. Glycoproteomic identification of novel plasma
biomarkers for oral cancer. Journal of food and drug analysis, v. 27, n. 2,
p. 483-493, 2019.
HAAB, B. B.; YUE, T. High-throughput studies of protein glycoforms using
antibody-lectin sandwich arrays. Methods Mol Biol., v. 785, p. 223–236,
2011.
CHOI, J. W. et al. Use of CA15-3 for screening breast cancer: An antibody-
lectin sandwich assay for detecting glycosylation of CA15-3 in sera.
Oncology Reports, v.40, p. 145-154, 2018.
Coordenação de Prevenção e Vigilância, Instituto Nacional de Câncer José
Alencar Gomes da Silva. Estimativa 2018: incidência de câncer no Brasil.
COULIBALY, Fohona S.; YOUAN, Bi-Botti C. Current status of lectin-based
cancer diagnosis and therapy. 2017.
DE SCHUTTER, K; VAN DAMME, E. J. M. Protein-carbohydrate
interactions as part of plant defense and animal immunity. Molecules, v.
20, n. 5, p. 9029-9053, 2015.
ERRANTE, P.R. Imunovigilância e mecanismos de evasão
tumoral. UNILUS Ensino e Pesquisa, v. 17, n. 46, p. 239-255, 2020.
GAUTAM, A. K. et al. Current scenario of legume lectins and their practical
applications. Journal of Crop Science and Biotechnology, v. 21, n. 3, p.
217-227,2018.
HASHIM, O. H.; JAYAPALAN, J. J.; LEE, C. Lectins : an effective tool for
screening of potential cancer biomarkers. PeerJ, v. 5, p. 1–30, 2017.
INCA. Ministério da saúde. O que é câncer? [S. l.]: INCA, 2019. Disponível
em < https://www.inca.gov.br/oque-e-cancer > acesso em 20 out. 2020.
INCA: Estimativa 2020: Incidência de câncer no Brasil,<disponível em:
https://www.inca.gov.br/sites/ufu.sti.inca.local/files/media/document/estimati
va-2020-incidencia-de-cancer-no-brasil.pdf> Acesso em 30/11/200.

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
95
KILPATRICK, D. C. Carbohydrate and protein conformation. Biochemical
Journal. v. 239, p. 808-806, 1986.
KUMAR, G; SUROLIA, A. Comprehensive analysis of α 2–3-linked sialic
acid specific Maackia amurensis leukagglutinin reveals differentially
occupied N-glycans and C-terminal processing. International journal of
biological macromolecules, v. 94, p. 114-121, 2017.
KUSAMA, K. et al. Reevaluation of Pholiota squarrosa lectin-reactive
haptoglobin as a pancreatic cancer biomarker using an improved ELISA
system. Glycoconjugate journal, v. 34, n. 4, p. 537-544, 2017.
LAVANYA, V. et al. Lectins-the promising câncer
therapeutics. Oncobiology and Targets, v. 1, n. 2014, p. 1235, 2017.
LIM, A. W. W. et al. Lectins in Cervical Screening. Cancers, v. 12, n. 7, p.
1928, 2020.
LIMA, V. C. O. et al. atividade hemaglutinante em sementes exóticas
germinadas e comestíveis. Revista Brasileira de Inovação Tecnológica
em Saúde-ISSN: 2236-1103, 2017.
LLOP, E. et al. Improvement of prostate cancer diagnosis by detecting PSA
glycosylation-specific changes. Theranostics, v. 6, n. 8, p. 1190-1204,
2016.
MANNING, J. C. et al. Lectins: a primer for histochemists and cell
biologists. Histochemistry and cell biology, v. 147, n. 2, p. 199-222,
2017.
MARQUES NETO, Antonio Moreira. Uma nova lectina da esponja marinha
aplysina sp. (APLYL-1) com atividade citotóxica pra célula tumoral (HeLa) e
aglutinante de leishmania amazonensis. 2015. 68f. Dissertação (Mestrado
em Bioquímica) - Centro de Biociências, Universidade Federal do Rio
Grande do Norte, Natal, 2015.
MATSUDA, A. et al. Lectin microarray-based sero-biomarker verification
targeting aberrant O-linked glycosylation on mucin 1. Analytical
Chemistry, v. 87, n. 14, p.7274-7281, 2015.
MOTA, Augusto. Imunoterapia do Câncer: Uma Nova Era. Revista
Científica Hospital Santa Izabel, v. 3, n. 1, p. 9-19, 2019.
OBAID, Girgis et al. Cancer targeting with biomolecules: a comparative
study of photodynamic therapy efficacy using antibody or lectin conjugated
phthalocyanine-PEG gold nanoparticles. Photochemical &
Photobiological Sciences, v. 14, n. 4, p. 737-747, 2015.
OLIVEIRA, Jaqueline Franciele Caetano de. Lectinas vegetais: de
moléculas de defesa de plantas às suas diversas aplicações

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
96
biotecnológicas. 2018. 62 f. Dissertação (Mestrado - Programa de Pós-
Graduação em Ciências Farmacêuticas) - Universidade Estadual do Oeste
do Paraná, Cascavel, 2018.
Osterne, Vinícius José da Silva. Estudos de estrutura-função da lectina de
sementes de Canavalia virosa com atividade inflamatória e citotóxica: uma
potencial ferramenta anticâncer. Dissertação (mestrado) – Universidade
Federal do Ceará, Centro de Ciências Agrárias, Programa de Pós-
Graduação em Biotecnologia de Recursos Naturais, Fortaleza,2016.
PEREIRA, R; DA SILVA, A. L. C. Uma revisão do potencial biotecnológico
de lectinas do gênero Canavalia E Dioclea. Journal of Biology &
Pharmacy and Agricultural Management, v. 15, n. 3, 2019.
PIHÍKOVÁ, D.; KASÁK, P.; TKAC, J. Glycoprofiling of cancer biomarkers:
Label-free electrochemical lectin-based biosensors. Open Chem., v. 13,
p.636–655, 2015.
PINEDO, M. et al. Molecular characterization of Helja, an extracellular
jacalin-related protein from Helianthus annuus: Insights into the relationship
of this protein with unconventionally secreted lectins. Journal of plant
physiology, v. 183, p. 144-153, 2015.
PROCÓPIO, T. F. et al. CasuL: A new lectin isolated from Calliandra
surinamensis leaf pinnulae with cytotoxicity to cancer cells, antimicrobial
activity and antibiofilm effect. International journal of biological
macromolecules, v. 98, p. 419-429, 2017.
REYES-MONTAÑO, Edgar Antonio; VEGA-CASTRO, Nohora Angélica.
Plant lectins with insecticidal and insectistatic activities. Insecticides-
Agriculture and Toxicology [Prieiga per internetą 2018-11-03]. InTech,
p. 17-41, 2018.
ESTIMATIVA, I. N. C. A. et al. incidência de câncer no Brasil. Rio de
Janeiro, 2019.
SAGAR, D.; DHALL, Heena. Legumes: Potential source of entomotoxic
proteins-A review. Legume Research-An International Journal, v. 41, n.
5, p. 639-646, 2018.
GAUTAM, Ajay Kumar et al. Legume lectins: Potential use as a diagnostics
and therapeutics against the cancer. International journal of biological
macromolecules, v. 142, p. 474-483, 2020.
SILVA, Ivanice Bezerra da. Caracterização estrutural de uma lectina
nociceptiva obtida de sementes da espécie Platypodium elegans VOG.
2017. 107 f. Tese (Doutorado em Biotecnologia de Recursos Naturais) -
Universidade Federal do Ceará, Fortaleza, 2017.

O POTENCIAL DE LECTINAS NO DIAGNÓSTICO DO CÂNCER
97
SILVA, Priscila Marcelino dos Santos. Desenvolvimento de bioensaios
utilizando Cramoll 1,4 como alternativa para o diagnóstico do câncer de
próstata. 2019. Tese (Doutorado em Ciências Biológicas) – Universidade
Federal de Pernambuco, Recife, 2019.
TEIXEIRA, H. C.et al. Proteínas de checkpoint imunológico como novo alvo
da imunoterapia contra o câncer: revisão da literatura. HU Revista, v. 45,
n. 3, p. 325-333, 2019.
TIAN, R. et al. Uncovering the binding specificities of lectins with cells for
precision colorectal cancer diagnosis based on multimodal
imaging. Advanced Science, v. 5, n. 6, p. 1800214, 2018.
VERONEZ, L. C.et al. genética e imunologia do câncer para alunos do
ensino básico: relato de uma experiência. Revista Brasileira de Extensão
Universitária, v. 10, n. 2, p. 63-70, 2019.
VILLRINGER, S. et al. Lectin-mediated protocell crosslinking to mimic cell-
cell junctions and adhesion. Scientific Reports, v. 8, p. 1932-1943, 2018.
ZHOU, Shu-Min et al. Lectin RCA-I specifically binds to metastasis-
associated cell surface glycans in triple-negative breast cancer. Breast
Cancer Research, v. 17, n. 1, p. 1-14, 2015.
AGRADECIMENTOS
Agradecimentos aos amigos pelo empenho nesse trabalho,
a Universidade Federal de Pernambuco e aos órgãos de
fomento CAPES, CNPq e FACEPE que nos dão subsidos para
seguirmos em frente.
.

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
98
CAPÍTULO 5
ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
JUCILENE PEREIRA DE SOUSA1
1Doutoranda do curso de Bioquímica, UFC, [email protected]
RESUMO: As ureases fazem parte da classe de amido-hidrolases e fosfotriesterases, apresentando metal em seu sítio ativo, sendo assim denominadas de metaloenzimas níquel-dependentes. São as enzimas mais eficientes conhecidas até o momento, acelerando a taxa de reação de hidrólise da ureia em amônia e carbamato, que então se decompõe em outra molécula de amônia e dióxido de carbono.O principal papel ambiental da urease é permitir que o organismo use ureia como fonte de nitrogênio ressaltando sua importância no ciclo do nitrogênio, gerando assim nitrogênio acessível para o crescimento das plantas. Entretanto, na agricultura a alta atividade da urease causa problemas ambientais e econômicos, uma vez que a hidrólise muito rápida da ureia diminui a eficiência do uso de fertilizantes, além disso, a quantidade anormal de amônia provoca danos à germinação de sementes, mudas e plantas jovens. A abordagem mais sucedida para resolver esses problemas é a partir do uso de inibidores. Dito isto, o objetivo desse trabalho centra-se em buscar as estruturas de ureases depositadas no PDB (Protein Data Bank) tanto nativas quanto complexadas com ligantes, listar os ligantes complexados com as ureases e pesquisar trabalhos referentes à urease de Canavalia ensiformis (JBU) de 2016-2020 com esses ligantes. Para tanto, foi feita uma busca de estruturas de ureases de diferentes organismos depositadas no PDB, foram feitas análises das estruturas para mapear os ligantes complexados, os quais foram listados e utilizados como

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
99
descritores na busca de inibidores de JBU. Como resultados do presente trabalho de revisão, foram obtidas 57 estruturas de ureases com ou sem inibidores ligados. As estruturas de ureases são de cinco espécies diferentes de bactérias (Klebsiella aerogenes, Sporosarcina pasteurii, Helicobacter pylori, Helicobacter mustelae, e Yersinia enterocolítica) e duas espécies diferentes de plantas (Canavalia ensiformis e Cajanus cajan). Dentre os resultados obtidos foram encontrados trabalhos englobando a atividade inibitória de JBU com ácido bórico, AHA (ácido acetohidroxâmico), catecol, fluoreto, NBPT (N-(n-butil) tiofosfórico triamida) e sulfito de sódio, os quais se apresentaram como bons inibidores. Dentro da busca, outros inibidores também se destacaram e se apresentaram muitas vezes até mais eficiente que o AHA, inibidor padrão da urease, como o alcaloide epiberberina e derivados de quinazolinona. Logo, conclui-se que a escolha do tipo de inibidor a ser utilizado é dependente de seu possível papel, uma vez que se considerarmos como potenciais fármacos poderá ser caro, enquanto que no caso de utilização na inibição da decomposição da ureia no solo tem que ser de estrutura simples e, portanto, substancialmente mais barato. Palavras-chave: Amônia. Protein Data Bank. Canavalia ensiformis. Ácido acetohidroxâmico.
INTRODUÇÃO
As ureases (EC 3.5.1.5) são membros da superfamília de
amido-hidrolases e fosfotriesterases, apresentando metal em
seu sítio ativo, sendo assim denominadas de metaloenzimas
níquel-dependentes. São as enzimas mais eficientes
conhecidas até o momento, acelerando a taxa de reação de
hidrólise da ureia em amônia e carbamato, que então se
decompõe em outra molécula de amônia e dióxido de carbono,
por um fator de pelo ao menos 1014 (KAPPAUN et al., 2018).

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
100
A urease isolada de sementes de Canavalia ensiformis
foi a primeira enzima a ser cristalizada por James B. Sumner,
em 1926, elucidando a ideia que enzimas são proteínas, uma
descoberta a qual lhe concedeu o prêmio Nobel de Química em
1946, entretanto sua estrutura foi resolvida apenas 83 anos
depois (BALASUBRAMANIAN; PONNURAJ, 2010). Em 1999, a
estrutura proteica da urease de Klebsiella aerogenes (PDB
1EJW) foi a primeira a ser resolvida e desde então estruturas
de ureases de bactérias e plantas, com ou sem inibidores
ligados, foram determinadas como de Sporosarcina pasteurii,
Helicobacter pylori, Helicobacter mustelae, Yersinia
enterocolítica, Canavalia ensiformis e Cajanus cajan.
A urease é uma enzima chave no ciclo global do
nitrogênio pelo fato de converter a ureia gerando dióxido de
carbono e amônia. A ureia, por sua vez, é responsável de forma
significativa pelo consumo total de fertilizantes de nitrogênio em
todo o mundo. Sua aplicação é acompanhada de grandes
perdas de amônia, devido a ação de ureases (KAFARSKI;
TALMA, 2018).
O principal papel ambiental da urease é permitir que o
organismo use ureia como fonte de nitrogênio ressaltando sua
importância no ciclo do nitrogênio, gerando assim nitrogênio
acessível para o crescimento das plantas (CORDERO; SNELL;
BARDGETT, 2019). Entretanto, na agricultura a alta atividade
da urease causa problemas ambientais e econômicos. A
hidrólise muito rápida da ureia diminui a eficiência do uso de
fertilizantes, além disso, a quantidade anormal de amônia
provoca danos à germinação de sementes, mudas e plantas
jovens (GOMES et al., 2018).
A abordagem mais sucedida para resolver esses
problemas é a partir do uso de inibidores. Os inibidores da

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
101
urease pertencem a diferentes classes químicas e muitos deles
foram investigados nas últimas décadas, dentre eles os ácidos
hidroxâmicos apresentam-se como um dos melhores
candidatos (ARORA; ISSAR; KAKKAR, 2018), os derivados de
quinona são uma outra classe de compostos inibidores
(MAZZEI et al., 2016a), bem como os íons de metais pesados.
Os inibidores de urease, por sua vez, podem ser
amplamente classificados em duas categorias: (i) inibidores do
tipo substrato, como hidroxiureia e ácidos hidroxâmicos e (ii)
inibidores que afetam o mecanismo de reação, como
fosforodiamidatos e imidazóis, como inibidores da bomba de
prótons do rabeprazol, omeprazol e lansoprazol. Esses
inibidores podem ainda ser agrupados em quatro grupos: (i)
constituído por compostos tiólicos, pois os ânions tiolatos
reagem diretamente com o metalocentro da urease; (ii)
inibidores deste grupo competem com a uréia pela ligação com
o sítio ativo da urease, como o ácido hidroxâmico e seus
derivados; (iii) este grupo contém os inibidores mais eficazes,
tais como fosforodiamidatos; e (iv) o quarto grupo consiste em
ligantes e quelantes de níquel (HASSAN; ZEMLICKA, 2016).
Dito isto, o objetivo desse trabalho centra-se em buscar as
estruturas de ureases depositadas no PDB (Protein Data Bank)
tanto nativas quanto complexadas com ligantes, listar os
ligantes complexados com as ureases e pesquisar trabalhos
referentes à urease de Canavalia ensiformis (JBU) de 2016-
2020 com esses ligantes.
METODOLOGIA
Este trabalho constitui-se como uma revisão bibliográfica
a qual se desenvolveu, inicialmente, pela busca de estruturas

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
102
de ureases de diferentes organismos depositadas no PDB, quer
seja elas nativas ou complexadas com ligantes, desde a
obtenção da primeira estrutura de urease em 1995 até março
de 2020. A partir disso, foram feitas análises das estruturas para
mapear os ligantes complexados com as ureases, os quais
foram listados e utilizados como descritores na busca de
inibidores de JBU. A busca foi feita utilizando o nome do inibidor
e o termo “jbu”, com filtro para publicações dos últimos cinco
anos (2016-2020), nas principais bases de dados “Science
direct” e “Pubmed”.
RESULTADOS E DISCUSSÃO
Caracterização estrutural da Urease de Canavalia
ensiformis
A JBU possui 840 resíduos de aminoácidos com uma
única subunidade α de 90 kDa. A forma mínima dessa enzima
expressando atividade enzimática é de um trímero de 270 kDa
e a conformação mais amplamente aceita na comunidade
científica para sua forma nativa é de um hexâmero de 540 kDa
(ZERNER, 1991) .
O monômero JBU de cadeia única consiste em quatro
domínios: o domínio αβ N-terminal (1-134) que está conectado
ao domínio αβ (135–285). Este por sua vez está conectado a
um domínio β (286–401 e 702–761), o qual está conectado a
um domínio do barril TIM na porção carboxi-terminal (αβ)8 (402–
701 e 762–840). O barril αβ é comumente encontrado em
enzimas e, também, nesse domínio geralmente localiza-se o
sítio ativo. Corroborando com esse achado, o sítio ativo da JBU
encontra-se nesse domínio (αβ)8, como pode ser observado na
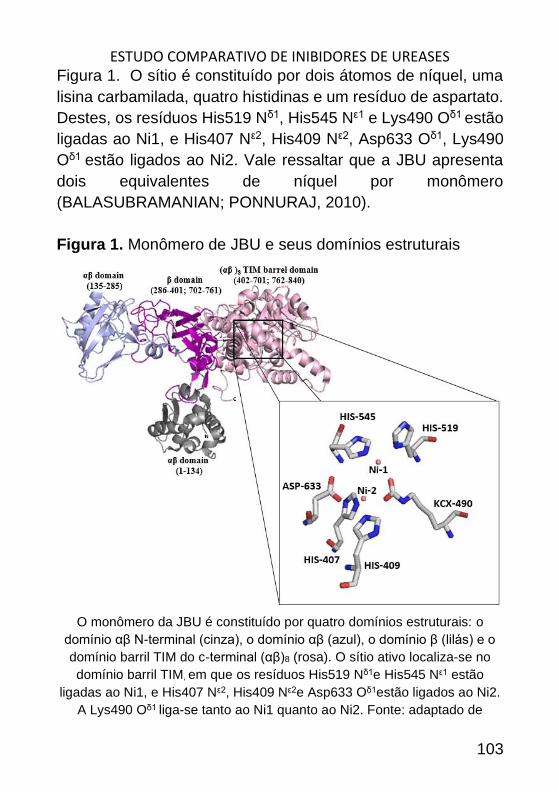
ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
103
Figura 1. O sítio é constituído por dois átomos de níquel, uma
lisina carbamilada, quatro histidinas e um resíduo de aspartato.
Destes, os resíduos His519 Nδ1, His545 Nε1 e Lys490 Oδ1 estão
ligadas ao Ni1, e His407 Nε2, His409 Nε2, Asp633 Oδ1, Lys490
Oδ1 estão ligados ao Ni2. Vale ressaltar que a JBU apresenta
dois equivalentes de níquel por monômero
(BALASUBRAMANIAN; PONNURAJ, 2010).
Figura 1. Monômero de JBU e seus domínios estruturais
O monômero da JBU é constituído por quatro domínios estruturais: o
domínio αβ N-terminal (cinza), o domínio αβ (azul), o domínio β (lilás) e o
domínio barril TIM do c-terminal (αβ)8 (rosa). O sítio ativo localiza-se no
domínio barril TIM, em que os resíduos His519 Nδ1e His545 Nε1 estão
ligadas ao Ni1, e His407 Nε2, His409 Nε2e Asp633 Oδ1estão ligados ao Ni2.
A Lys490 Oδ1 liga-se tanto ao Ni1 quanto ao Ni2. Fonte: adaptado de
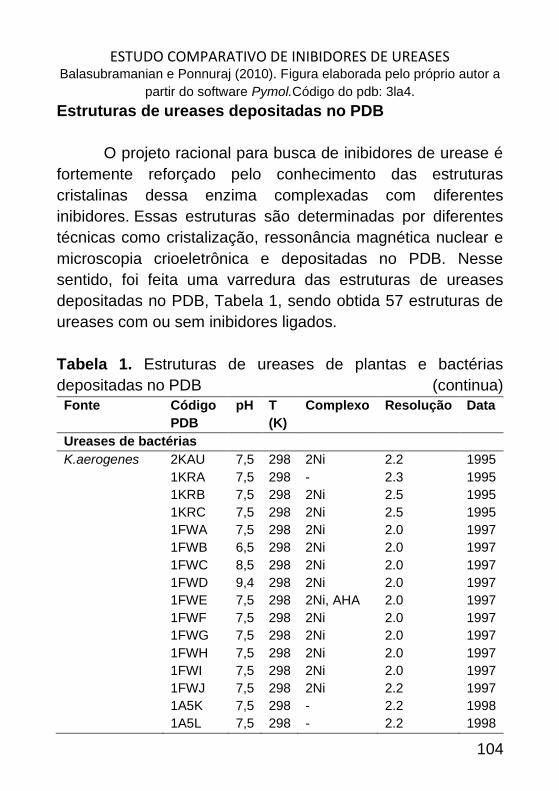
ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
104
Balasubramanian e Ponnuraj (2010). Figura elaborada pelo próprio autor a
partir do software Pymol.Código do pdb: 3la4.
Estruturas de ureases depositadas no PDB
O projeto racional para busca de inibidores de urease é
fortemente reforçado pelo conhecimento das estruturas
cristalinas dessa enzima complexadas com diferentes
inibidores. Essas estruturas são determinadas por diferentes
técnicas como cristalização, ressonância magnética nuclear e
microscopia crioeletrônica e depositadas no PDB. Nesse
sentido, foi feita uma varredura das estruturas de ureases
depositadas no PDB, Tabela 1, sendo obtida 57 estruturas de
ureases com ou sem inibidores ligados.
Tabela 1. Estruturas de ureases de plantas e bactérias
depositadas no PDB (continua)
Fonte Código
PDB
pH T
(K)
Complexo Resolução Data
Ureases de bactérias
K.aerogenes 2KAU 7,5 298 2Ni 2.2 1995
1KRA 7,5 298 - 2.3 1995
1KRB 7,5 298 2Ni 2.5 1995
1KRC 7,5 298 2Ni 2.5 1995
1FWA 7,5 298 2Ni 2.0 1997
1FWB 6,5 298 2Ni 2.0 1997
1FWC 8,5 298 2Ni 2.0 1997
1FWD 9,4 298 2Ni 2.0 1997
1FWE 7,5 298 2Ni, AHA 2.0 1997
1FWF 7,5 298 2Ni 2.0 1997
1FWG 7,5 298 2Ni 2.0 1997
1FWH 7,5 298 2Ni 2.0 1997
1FWI 7,5 298 2Ni 2.0 1997
1FWJ 7,5 298 2Ni 2.2 1997
1A5K 7,5 298 - 2.2 1998
1A5L 7,5 298 - 2.2 1998

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
105
1A5M 7,5 298 - 2.0 1998
1A5N 7,2 298 2Ni,
formiato
2.4 1998
Fonte: Tabela elaborada pelo autor com base no banco de dados PDB
As estruturas das ureases complexadas com pequenas
moléculas foram determinadas para elucidar a reatividade
química dos íons de níquel e os resíduos do sítio ativo
circundante, bem como esclarecer seu papel no mecanismo
catalítico e no mecanismo de inibição da enzima. As estruturas
de ureases correspondem a cinco espécies diferentes de
bactérias (Klebsiella aerogenes, Sporosarcina pasteurii,
Helicobacter pylori, Helicobacter mustelae, e Yersinia
enterocolítica) e duas espécies diferentes de plantas (Canavalia
ensiformis e Cajanus cajan).
A superposição dos monômeros revela que são
altamente semelhantes (Figura 2). Os resíduos de aminoácidos
que compõem o sítio catalítico dessas enzimas são altamente
conservados sendo encontrado na subunidade α, uma vez que
normalmente resíduos de aminoácidos essenciais para
atividade biológica de uma proteína são conservados ao longo
do tempo evolutivo. Como pode-se observar o sítio catalítico é
constituído por quatro resíduos de histidina, um aspartato e uma
lisina carbamilada e estão envolvidos na ligação do
metalocentro de níquel.
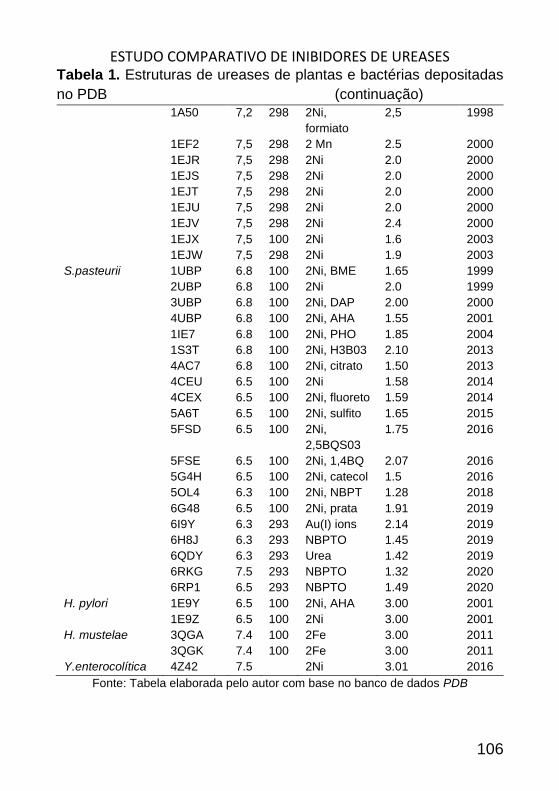
ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
106
Tabela 1. Estruturas de ureases de plantas e bactérias depositadas
no PDB (continuação)
1A50 7,2 298 2Ni,
formiato
2,5 1998
1EF2 7,5 298 2 Mn 2.5 2000
1EJR 7,5 298 2Ni 2.0 2000
1EJS 7,5 298 2Ni 2.0 2000
1EJT 7,5 298 2Ni 2.0 2000
1EJU 7,5 298 2Ni 2.0 2000
1EJV 7,5 298 2Ni 2.4 2000
1EJX 7,5 100 2Ni 1.6 2003
1EJW 7,5 298 2Ni 1.9 2003
S.pasteurii 1UBP 6.8 100 2Ni, BME 1.65 1999
2UBP 6.8 100 2Ni 2.0 1999
3UBP 6.8 100 2Ni, DAP 2.00 2000
4UBP 6.8 100 2Ni, AHA 1.55 2001
1IE7 6.8 100 2Ni, PHO 1.85 2004
1S3T 6.8 100 2Ni, H3B03 2.10 2013
4AC7 6.8 100 2Ni, citrato 1.50 2013
4CEU 6.5 100 2Ni 1.58 2014
4CEX 6.5 100 2Ni, fluoreto 1.59 2014
5A6T 6.5 100 2Ni, sulfito 1.65 2015
5FSD 6.5 100 2Ni,
2,5BQS03
1.75 2016
5FSE 6.5 100 2Ni, 1,4BQ 2.07 2016
5G4H 6.5 100 2Ni, catecol 1.5 2016
5OL4 6.3 100 2Ni, NBPT 1.28 2018
6G48 6.5 100 2Ni, prata 1.91 2019
6I9Y 6.3 293 Au(I) ions 2.14 2019
6H8J 6.3 293 NBPTO 1.45 2019
6QDY 6.3 293 Urea 1.42 2019
6RKG 7.5 293 NBPTO 1.32 2020
6RP1 6.5 293 NBPTO 1.49 2020
H. pylori 1E9Y 6.5 100 2Ni, AHA 3.00 2001
1E9Z 6.5 100 2Ni 3.00 2001
H. mustelae 3QGA 7.4 100 2Fe 3.00 2011
3QGK 7.4 100 2Fe 3.00 2011
Y.enterocolítica 4Z42 7.5 2Ni 3.01 2016
Fonte: Tabela elaborada pelo autor com base no banco de dados PDB
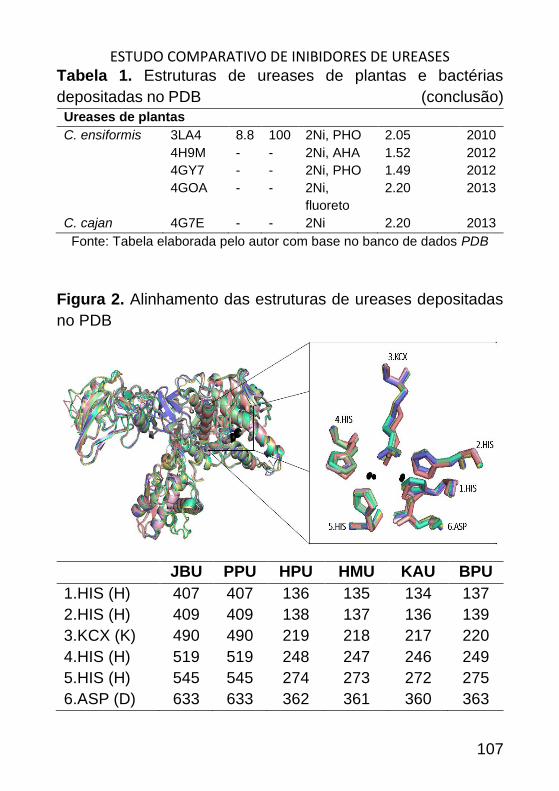
ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
107
Tabela 1. Estruturas de ureases de plantas e bactérias
depositadas no PDB (conclusão)
Ureases de plantas
C. ensiformis 3LA4 8.8 100 2Ni, PHO 2.05 2010
4H9M - - 2Ni, AHA 1.52 2012
4GY7 - - 2Ni, PHO 1.49 2012
4GOA - - 2Ni,
fluoreto
2.20 2013
C. cajan 4G7E - - 2Ni 2.20 2013
Fonte: Tabela elaborada pelo autor com base no banco de dados PDB
Figura 2. Alinhamento das estruturas de ureases depositadas
no PDB
JBU PPU HPU HMU KAU BPU
1.HIS (H) 407 407 136 135 134 137
2.HIS (H) 409 409 138 137 136 139
3.KCX (K) 490 490 219 218 217 220
4.HIS (H) 519 519 248 247 246 249
5.HIS (H) 545 545 274 273 272 275
6.ASP (D) 633 633 362 361 360 363

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
108
Tem-se o alinhamento das estruturas de JBU (amarelo claro), PPU (verde
ciano), HPU (salmão), HMU (rosa ), KAU (cinza) e BPU (verde pálido). Seis
resíduos de aminoácidos (quatro histidinas, uma lisina carbamilada e um
aspartato) compõem o sítio catalítico de cada espécie como detalhado na
tabela e dois metais estão coordenados com esses resíduos de
aminoácidos (preto). Normalmente o níquel é o metalocentro comumente
presente na maioria das ureases, dentre as seis abordadas nesse estudo a
HMU apresenta ferro como íon metálico presente no sítio ativo. Figura
elaborada pelo próprio autor a partir do software Pymol.
O número de cadeias polipeptídicas que forma o
“monômero” varia de acordo com a fonte de urease, uma vez
que para ureases vegetais e fúngicas, o monômero é
constituído por apenas uma cadeia polipeptídica (α), enquanto
que a unidade funcional das ureases de bactérias é formada por
duas (α e β) ou três (α, β e γ) cadeias polipeptídicas. A maioria
das ureases bacterianas são trímeros de trímeros ([αβγ]3), uma
exceção para a urease de Helicobacter pylori a qual foi
cristalizada como um tetrâmero de trímeros de dímeros ([αβ]3)4
(KAPPAUN et al., 2018).
Como o foco desse estudo é acerca dos inibidores
reportados para a JBU, o material buscado foi baseado nos
inibidores reportados estruturalmente para ureases de
diferentes fontes depositadas no PDB, conforme descrito na
Tabela 1. Dentre os resultados obtidos foram encontrados
trabalhos englobando o ácido bórico, AHA (ácido
acetohidroxâmico), catecol, fluoreto, NBPT (N-(N-butil)
tiofosfórico triamida) e sulfito de sódio, conforme Tabela 2.

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
109
Tabela 2. Inibidores de JBU reportados de 2016-2020 com base
em dados estruturais publicados no PDB Inibidor Referências
Ácido bórico TAN et al., 2017
AHA AKYUZ et al., 2018; HASSAN; ŠVAJDLENKA, 2017;
MENTEŞE et al., 2019; TAN et al., 2017; ZHOU et al., 2017
Catecol MAZZEI et al., 2016b
Fluoreto TAN et al., 2017
NBPT MAZZEI et al., 2019
Sulfito de sódio MAZZEI et al., 2016c
Fonte: Próprio autor
O primeiro trabalho refere-se a investigação das
atividades inibitórias da JBU e HPU (urease de Helicobacter
pylori) a partir de cinco alcalóides presentes em Rhizoma
Coptidis: a berberina, palmatina, coptisina, epiberberina e
jateorhizina. Todos eles apresentaram resultados positivos sob
a inibição das enzimas em questão. Dentre eles, a epiberberina
foi considerada o inibidor mais potente contra ambas as
ureases com valores de IC50 de 3,0±0,01 μM para HPU e 2,3 ±
0,01 μM para JBU, sendo mais eficaz do que o inibidor padrão
da urease, o AHA (83±0,01 μM para HPU e 22±0,01 μM para
JBU, respectivamente). Ainda nesse trabalho, o fluoreto de
sódio, inibidor competitivo de urease de ligação lenta, e o ácido
bórico, inibidor competitivo clássico da urease clássico, foram
empregados para determinar se a epiberberina interagiu com o
Ni2+ do sítio ativo da urease. Verificou-se que a atividade
enzimática residual da urease inativada por epiberberina na
presença desses reagentes inorgânicos foi menor do que na
ausência, indicando uma possível sinergia entre os compostos
(TAN et al., 2017).
Zhou e colaboradores (2017) avaliaram a inibição de
HPU e JBU por palmatina com valores de IC50 de 0,53±0,01
mM e 0,03±0,00 mM, respectivamente, em comparação com o

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
110
AHA (0,07±0,01 mM para HPU e 0,02±0,00 mM para JBU,
respectivamente). A inibição foi do tipo não competitiva e o
ensaio de reativação com DTT (ditiotreitol) indicou que a
inibição nas duas ureases era reversível. Já no estudo de
Hassan e Švajdlenka (2017) avaliaram as propriedades
inibitórias do ácido protocatecuico (PCA), um composto ativo
de Hibiscus sabdariffa L., contra a JBU. O composto
apresentou notável atividade anti-JBU comparado com o AHA,
com valores de IC50 de 1,7 e 3,2 µM, respectivamente. Ensaios
de docking molecular revelaram alta capacidade de ligação
espontânea do inibidor ao sítio ativo da JBU e constatou-se
ainda que a atividade inibitória está relacionada à presença de
frações hidroxila no inibidor.
Nesse estudo desenvolvido por Mentese e
colaboradores (2019), foi sintetizada uma nova série de
derivados de quinazolinona e foi analisado o potencial de
inibição in vitro desses compostos contra JBU. A maioria dos
compostos mostrou excelente atividade inibitória com valores
de IC50 variando entre 1,88±0,17 e 6,42±0,23 µg/mL, em
comparação com tioureia (IC50=15,06±0,68) e AHA (IC50 =
21,03 ± 0,94), inibidores de referência. Entre as moléculas
sintetizadas, os compostos 5c, 5e e 5a apresentaram o melhor
efeito inibitório com valores de IC50 de 1,88±0,17 µg/mL,
1,90±0,10 e 1,96±0,07 µg/mL, respectivamente.
Uma nova série de compostos de quinazolin-4 (3H)-ona
2,3-dissubstituídos incluindo anéis de oxadiazol e furano foi
sintetizada. Suas atividades inibitórias sobre a JBU foram
avaliadas in vitro. Todos os compostos sintetizados exibiram
potente atividade inibidora da urease na faixa de IC50 =
1,55±0,07-2,65±0,08 µg/mL, quando comparados com os
inibidores padrão de urease, como a tiouréia (IC50=15,08±0,71

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
111
µg/mL) e AHA (IC50=21,05±0,96 µg/mL). Os compostos
contendo anel furano foram considerados os inibidores mais
ativos quando comparados com os compostos contendo anel
oxadiazol. Dentre eles, o composto 3a foi encontrado como o
inibidor mais eficaz de urease com o valor IC50 de 1,55±0,11
µg/mL (AKIUZ et al., 2016).
A Investigação da atividade inibitória de catecol contra
JBU e SPU (Sporosarcina pasteurii) mostrou que esse polifenol
inativa irreversivelmente essas enzimas a partir de um
mecanismo complexo, o qual os autores propuseram uma
hipótese para o mecanismo de inativação da urease pelo
catecol. A estrutura cristalina do complexo SPU-catecol,
determinada na resolução de 1,50 Å, conforme Tabela 1, revela
os detalhes estruturais da inibição da enzima (MAZZEI et al.,
2016b).
O NBPTO (N-(n-butil) fosfórico triamida) é o oxo-derivado
do NBPT, que é amplamente utilizada na agricultura para
aumentar a eficiência de fertilizantes à base de ureia. A
resolução de 1,45 Å da estrutura de SPU com NBPTO mostra
a presença de DAP (ácido diamido fosfórico), gerado na
hidrólise enzimática de NBPTO com a liberação de n-butil
amina. O DAP, por sua vez, é ligado aos dois íons de níquel no
sítio ativo da urease por meio de dois átomos de oxigênio e um
grupo amida NH2. Uma caracterização cinética da inibição
baseada de NBPTO com SPU e JBU, realizada por medidas
calorimétricas, indica a ocorrência de inibição reversível. A
maior capacidade do NBPTO de inibir a urease, em
comparação com o NBPT, se deve ao fato do NBPTO reagir
com enzima sem a necessidade de converter uma das porções
de amida P − NH2 em P − OH, como no caso do NBPT,
reduzindo drasticamente a concentração da forma ativa do

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
112
inibidor NBPT. Isso indica que os inibidores de urease à base
de fosforamida devem ter pelo menos um átomo de oxigênio
ligado ao átomo de fósforo central para que a molécula se ligue
rápida e eficientemente ao centro de níquel da enzima (MAZZEI
et al., 2019).
Gomes e colaboradores (2018) ressaltam, também, que
a eficiência do NBPT depende da taxa de conversão em seu
análogo contendo oxigênio, o NBPTO. Foram analisados
quinze fertilizantes à base de uréia revestida com NBPT, ácido
húmico e zeólita, os quais foram usados em diferentes
concentrações. Os experimentos confirmaram a eficiência do
NBPT em inibir a ação enzimática mesmo em baixa
concentração (0,02% m/m). A adição de ácido húmico reduziu
a atividade da urease após 36 horas de reação, enquanto que
a zeólita não influenciou a atividade da enzima urease.
O sulfito de sódio tem sido amplamente utilizado como
conservante em soluções de urease para prevenir a oxidação
induzida por oxigênio, mas seu papel como inibidor também foi
sugerido. A estrutura do complexo urease-sulfito, determinada
na resolução de 1,65Å, mostra o inibidor ligado ao centro
dinuclear Ni (II) da urease em um modo tridentado envolvendo
ligações entre os dois íons Ni (II) no sítio ativo e todos os três
átomos de oxigênio do inibidor, apoiando a cinética de inibição
competitiva observada. Este modo de coordenação de sulfito
nunca foi observado, seja em proteínas ou em complexos de
pequenas moléculas, e poderia inspirar químicos de
coordenação sintética, bem como bioquímicos, a desenvolver
inibidores de urease com base nessa porção química (MAZZEI
et al., 2016c).
As estruturas cristalinas publicadas indicam a
necessidade de elementos indispensáveis para um inibidor

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
113
eficaz: a presença de uma fração complexante de níquel ao
lado de uma rede devidamente posicionada de doadores e
aceitadores de ligações de hidrogênio fixados a estruturas
flexíveis. O processo de concepção de inibidores de urease
também é fortemente dependente de seu possível papel. Se
considerarmos os potenciais fármacos, o arcabouço molecular
pode ser estruturalmente complexo, uma vez que o fármaco
pode ser caro, enquanto no caso de inibição da decomposição
da ureia no inibidor do solo tem que ser de estrutura simples e,
portanto, substancialmente barata (KARFASHI; TALMA, 2018).
Se faz necessária essa busca por inibidores da urease, uma vez
que são necessários para a redução da poluição ambiental,
aumento da eficiência do nitrogênio da ureia, erradicação do H.
pylori, tratamento de úlceras pépticas e outras doenças
relacionadas à urease (TAN et al., 2017).
CONCLUSÕES
Dentre os inibidores co-cristalizados com ureases de
diferentes fontes e depositadas no PDB, as moléculas de ácido
bórico, AHA, catecol, fluoreto, NBPT e sulfito de sódio
apresentaram-se como inibidores extremamente eficientes para
a JBU. Dentre eles destaca-se o AHA como um inibidor padrão
e que já vem sendo estudado a bastante tempo, sendo assim
utilizado como padrão para descoberta de novos inibidores.
Dentro da busca, outros inibidores também se destacaram e se
apresentaram muitas vezes até mais eficientes que o AHA,
como o alcaloide epiberberina e derivados de quinazolinona.
Logo, conclui-se que a escolha do tipo de inibidor a ser utilizado
é dependente de seu possível papel, uma vez que se
considerarmos como potenciais fármacos poderá ser caro,

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
114
enquanto que no caso de utilização na inibição da
decomposição da ureia no solo tem que ser de estrutura
simples e, portanto, substancialmente mais barato.
REFERÊNCIAS BIBLIOGRÁFICAS AKYÜZ, G. et al. Bioorganic Chemistry Synthesis and molecular docking study of some novel 2 , 3-disubstituted quinazolin-4 ( 3 H ) -one derivatives as potent inhibitors of urease. BioorganicChemistry, v. 80, n. June, p. 121–128, 2018. ARORA, R.; ISSAR, U.; KAKKAR, R. In Silico study of the active site of Helicobacter pylori urease and its inhibition by hydroxamic acids. Journal of Molecular Graphics and Modelling, v. 83, p. 64-73, 2018. BALASUBRAMANIAN, A.; PONNURAJ, K. Crystal Structure of the First Plant Urease from Jack Bean: 83 Years of Journey from Its First Crystal to Molecular Structure. Journal of Molecular Biology, v. 400, n. 3, p. 274-283, 2010. CORDERO, I.; SNELL, H.; BARDGETT, R. D. High throughput method for measuring urease activity in soil. Soil Biology and Biochemistry, v. 134, p. 72-77, 2019. GOMES, M. P. et al. Urease activity according to the different inhibitors. Annals of the Brazilian Academy of Sciences, v. 90, p. 1–8, 2018. HASSAN, S. T. S. Protocatechuic Acid from Hibiscus sabdariffa L . as a Potent Urease Inhibitor by an ESI-MS Based Method. Molecules, v. 22, n. 1696, p. 1-11, 2017. HASSAN, S.T.S.; ZEMLICKA, M. Plant Derived Urease Inhibitors as Alternative Chemotherapeutic. Arch. Pharm. Chem. Life Sci., 349, 1–16, 2016. KAFARSKI, P.; TALMA, M. Recent advances in design of new urease inhibitors : A review. Journal of Advanced Research, v. 13, p. 101–112, 2018. KAPPAUN, K. et al. Ureases: Historical aspects, catalytic, and non-catalytic properties – A review. Journal of Advanced Research, v. 13, p. 3–17, 2018. MAZZEI, L. et al. Inactivation of urease by 1,4-benzoquinone: chemistry at the protein surface. Dalton Trans., v. 45, n. 15, 2016a. MAZZEI, L. et al. Inactivation of urease by catechol: Kinetics and structure. Journal of Inorganic Biochemistry, n. 16, p. 1-30, 2016b. MAZZEI, L. et al. Insights into Urease Inhibition by N-(n-Butyl) Phosphoric Triamide through an Integrated Structural and Kinetic Approach. Journal of Agricultural and Food Chemistry, v. 67, p. 2127–2138, 2019.

ESTUDO COMPARATIVO DE INIBIDORES DE UREASES
115
MAZZEI, L. et al. Kinetic and structural studies reveal a unique binding mode of sul fi te to the nickel center in urease. Journal of Inorganic Biochemistry, v. 154, p. 42–49, 2016c. MENTEŞE, E. et al. Bioorganic Chemistry Synthesis , in vitro urease inhibition and molecular docking studies of some novel quinazolin-4 ( 3 H ) -one derivatives containing triazole , thiadiazole and thiosemicarbazide functionalities. Bioorganic Chemistry, v. 83, n. July 2018, p. 289–296, 2019. TAN, L. et al. European Journal of Pharmaceutical Sciences Epiberberine , a natural protoberberine alkaloid , inhibits urease of Helicobacter pylori and jack bean : Susceptibility and mechanism. European Journal of Pharmaceutical Sciences, v. 110, p. 77–86, 2017. ZERNER, B. Recent Advances in the Chemistry of an Old Enzyme, Urease. Bioorganic Chemistry, v. 19, p. 116-131, 1991. ZHOU, J. et al. Inhibition of Helicobacter pylori and Its Associated Urease by Palmatine : Investigation on the Potential Mechanism. Plos one, p. 1–15, 2017.

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
116
CAPÍTULO 6
PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE MORINGA OLEIFERA LAM.
Matheus Cavalcanti de Barros 1,2
Ana Gabrielle Barros Silva 3 Caio César da Silva Guedes 4
Thiago Henrique Napoleão 5 Patrícia Maria Guedes Paiva 5
1 Mestrando pelo Programa de Pós-graduação em Ciências Biológicas, UFPE; 2 Graduando do curso de Licenciatura em Ciências Biológicas, UNIP
3 Graduanda do curso de Bacherlado em Ciências Biológicas, UFPE; 4 Doutorando pelo Programa de Pós-graduação em Ciências Biológicas, UFPE
5 Professor do Departamento de Bioquímica, UFPE [email protected]
RESUMO: Moringa oleifera Lam. tem sido amplamente cultivada ao redor do mundo, devido aos usos biotecnológicos e farmacológicos, bem como na alimentação humana. As propriedades farmacológicas das folhas tem sido elucidadas por vários estudos científicos. Preparações obtidas a partir das folhas são tradicionalmente empregadas para o tratamento de várias doenças. O presente trabalho reuniu as principais atividades farmacológicas das folhas de M. oleifera relatadas na literatura científica buscando relacioná-las com os usos das folhas pela população. A busca por artigos relacionados às propriedades farmacológicas das folhas de M. oleifera foi realizada em diferentes bases de dados, como Science Direct, SciELO, Google Scholar e outras. Palavras-chave como Moringa, anticâncer, antimicrobiana, antiúlcera, antioxidante e atividade antiinflamatória foram usadas nas pesquisas. Foram priorizados trabalhos publicados a partir de 2015. As principais atividades farmacológicas relativas as folhas de M. oleífera foram antioxidante, hipolipidêmica, hipoglicemiante,

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
117
antiinflamatória, anticâncer, antimicrobiana, imunomoduladora e efeitos protetores. Estudos in vitro e in vivo utilizando folhas de M. oleifera certificam propriedades biológicas interessantes, demonstrando ser uma alternativa medicamentosa para o tratamento de diversas doenças. A revisão realizada estimula estudos que ampliem o leque de atividades farmacológicas das folhas de M. oleifera, bem como aqueles que identifiquem princípios ativos, elucidem mecanismos de ação das atividades já conhecidas e determinem efeitos toxicológicos de diferentes doses. Palavras-chave: Propriedades medicinais. Moringa. Fitoquímica.
INTRODUÇÃO
A família Moringaceae é formada por apenas um gênero
que apresenta treze espécies, sendo Moringa oleifera, também
conhecida como moringa, cedro ou quiabo-de-quina, a mais
cultivada e divulgada entre elas (LEONE et al., 2015). Nativa da
Índia, esta planta também é distribuída por diversos países no
mundo, sendo amplamente cultivada na Nigéria, Egito,
Nordeste do Brasil, Bangladesh, Afeganistão, Paquistão e
Malásia. Sua ampla distribuição pode ser atribuída a sua
capacidade de adaptação a ambientes com baixos índices
pluviométricos e a solos alcalinos e pouco férteis (MANOJ;
RANJITHA, 2020).
Apesar de basicamente todas as partes de M. oleifera
serem empregadas para alguma finalidade, a folha é
tradicionalmente a mais utilizada, devido à facilidade de
obtenção, alto conteúdo de metabólitos secundários bioativos e
o não comprometimento da planta durante a coleta. Ademais,
devido ao alto conteúdo nutricional das folhas, a moringa tem

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
118
sido indicada como uma alternativa sustentável de suplemento
alimentar para aqueles que sofrem de desnutrição,
especialmente nos países em desenvolvimento (VATS;
GUPTA, 2017). Infusões e outras preparações aquosas das
folhas têm sido utilizadas na medicina popular como anti-
inflamatório, hipoglicêmico e para controle de peso (COZ-
BOLAÑOS et al., 2018). Diversas outras propriedades
farmacológicas tem sido referidas para as folhas de M. oleífera.
Devido a presença de compostos bioativos e
características nutricionais, há uma crescente exploração e
compreensão da Moringa com referência à saúde humana.
Ante o exposto, o presente trabalho teve como objetivo abordar
as principais atividades farmacológicas das folhas de M. oleifera
encontradas na literatura científica, buscando evidenciar e
elucidar suas potencias aplicações populares.
MATERIAIS E MÉTODOS
A busca por artigos relacionados às propriedades
farmacológicas das folhas de M. oleifera foi realizada em
diferentes bases de dados, como Science Direct, SciELO,
Google Scholar e outras. Palavras-chave como Moringa,
anticâncer, antimicrobiana, antiúlcera, antioxidante, atividade
antiinflamatória e efeito protetor foram usadas nas pesquisas.
Além disso, foi priorizado artigos com ano de publicação entre
o período de 2015 a 2020. Para melhor organizar os dados
obtidos a partir das analises dos artigos científicos, as
informações contidas nesta revisão foram divididas por tópicos,
a fim de abordar, de maneira clara e objetiva, as diferentes
atividades biológicas..

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
119
RESULTADOS E DISCUSSÃO
Moringa oleifera Lam. (figura 1) tem sido amplamente
cultivada ao redor do mundo, devido às aplicações
biotecnológicas e farmacológicas que apresenta. As folhas de
M. oleifera são ricas em nutrientes, tais como proteínas, ácidos
graxos, fibras, carboidrato, vitaminas e aminoácidos essenciais.
Essas folhas são utilizadas popularmente para fins medicinais,
os quais tem sido elucidados por vários estudos científicos,
evidenciando o potencial nutracêutico das folhas dessa planta
(KOU et al., 2018).
Figura 2: A - Aspectos gerais de Moringa oleifera. B – Flores.
C- Sementes. D. Folhas. E – Frutos.
Fonte: Autoria Própria, 2020.
A
B
C
D
E

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
120
Estudos etnobotânicos envolvendo M. oleifera indicam que
preparações obtidas a partir das folhas dessa planta são
tradicionalmente empregadas para o tratamento de malária,
febre tifoide, pressão arterial, artrite, inchaços, cortes,
hipertensão, bem como para estimular a lactação e melhorar o
sistema imunológico (SIVASANKARI; ANANDHARAJ;
GUNASEKARAN, 2014). Infusões e outras preparações
aquosas têm sido utilizadas na medicina popular como anti-
inflamatório, hipoglicêmico e para controle de peso (COZ-
BOLAÑOS et al., 2018). Nesse sentido, devido aos potenciais
medicinais que apresenta (Figura 2), diversas pesquisas têm
sido desenvolvidas com o objetivo de comprovar as
propriedades farmacológicas das folhas dessa espécie vegetal
e elucidar os mecanismos de ação envolvidos..

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
121
Figura 2: Potenciais medicinais das folhas de Moringa
oleifera
Fonte: Autoria Própria, 2020.
Atividade antioxidante
Devido à elevada quantidade de polifenois, o alto
potencial antioxidante das folhas de M. oleifera tem sido
amplamente relatado na literatura, tanto a partir de abordagens
in vitro como in vivo. Sultana et al. (2015), avaliaram o potencial
antioxidante de folhas de moringa coletadas em três diferentes
estágios de maturação (4, 6 e 8 semanas) e observaram que o
sequestro do radical DPPH foi maior (60,1%) nas folhas com 4
semanas de maturação do que naquelas com 6 (56,0%) e 8

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
122
(53,4%) semanas. Preparações aquosas (infusão e decocção)
mostraram níveis de atividade antioxidante (em μmol
equivalente de trolox/mL) da ordem de 5,97 ± 0,08 e 5,75 ± 0,28
no ensaio do ABTS, 0,720 ± 0,01 e 0,655 ± 0,03 no teste do
DPPH e 1,469 ± 0,06 e 1,185 ± 0,08 no teste de poder de
redução do íon ferro (FRAP) (COZ-BOLAÑOS et al., 2018).
A atividade antioxidante in vitro do extrato aquoso das
folhas de M. oleifera também foi determinada por Nair et al.
(2020) através do teste do DPPH e in vivo, em ratos Wistar
machos, através do ensaio de peroxidação lipídica. A atividade
antioxidante da folha de M. oleifera apresentou um valor de IC50
de 10,47 µg/ml, enquanto que para o fármaco padrão, ácido
ascórbico, foi de 19,52 µg/ml. A análise in vivo da peroxidação
lipídica mostrou uma redução significativa da peroxidação
lipídica nos cérebros dos grupos tratados.
Atividade hipolipidêmica e hipoglicemiante
A capacidade de neutralizar espécies reativas de
oxigênio (EROs) está diretamente relacionada a prevenção de
doenças não transmissíveis (DNTs), como obesidade e
diabetes. Nesse sentido, os altos potenciais hipoglicêmico e
hipolipidêmico de preparações obtidas a partir das folhas de M.
oleifera têm sido fortemente relatados na literatura. O extrato
aquoso das folhas de M. oleifera administrado por via oral
durante 3 semanas na concentração de 100 mg/kg para ratos
com diabetes induzida por estreptozotocina e 200 mg/kg para
camundongos com dieta rica em lipídeos, restaurou a glicose
sanguínea, o perfil lipídico e o nível de enzimas hepáticas de
forma significativa em ambos os grupos (KHAN et al., 2017).
Outro estudo mostrou que o extrato das folhas de moringa

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
123
apresentou efeito hipoglicêmico em ratos diabéticos, os autores
sugeriram que esse efeito pode ser devido à presença de ácido
criptoclorogênico, quercetina 3-β-D-glicosídeo e kaempferol 3-
O- glicosídeo na preparação (IRFAN et al., 2017).
Helmy et al. (2017) revelaram que ratos albinos machos
tratados durante 60 dias com o pó de folhas de M. oleifera
(0,737% ou 1,475% de inclusão na dieta) e seu extrato (200 ou
400 mg/kg), apresentaram menor taxa de colesterol,
triglicerídeos, lipoproteína de baixa densidade e
malondialdeído; bem como menor atividade das enzimas
alanina aminotransferase e aspartato aminotransferase no soro
sanguíneo. O tratamento de ratos hipercolesterolêmicos com
extrato metanólico das folha de moringa, durante 49 dias,
resultou em redução no colesterol total, triglicerídeos, peso
corporal e dos órgãos, nos níveis de biomarcadores hepáticos
e de glicose sanguínea (PARE et al., 2016).
Atividade antiinflamatória
Fard et al. (2015), investigaram as propriedades anti-
inflamatórias do extrato hidroetanólico das folhas de M. oleifera
em macrófagos estimulados por lipopolissacarídeo (LPS). Os
autores observaram que o extrato inibiu significativamente a
secreção de óxido nítrico e outros marcadores inflamatórios,
como prostaglandina E2, fator de necrose tumoral alfa,
interleucina (IL) -6 e IL-1β. Além disso, o extrato induziu a
produção de IL-10 de forma dose-dependente e suprimiu
efetivamente a expressão de marcadores inflamatórios
proteicos, tais como óxido nítrico sintase e ciclooxigenase-2.
Resultados similares foram obtidos em outro estudo
(ARULSELVAN et al., 2016) realizado com quatro frações das

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
124
folhas de M. oleifera obtidas em diferentes solventes
(clorofórmio, hexano, butanol e acetato de etila). A avaliação do
potencial anti-inflamatório e mecanismo de ação celular foram
mensurados em macrófagos RAW264.7 induzidas por LPS. A
fração de acetato de etila suprimiu a produção de óxido nítrico
e citocinas pró-inflamatórias induzidos por LPS de maneira
dependente da concentração e foi mais eficaz do que as outras
frações. O estudo também mostrou que a fração de acetato de
etila inibiu efetivamente a expressão de mediadores
inflamatórios, incluindo ciclooxigenase-2, óxido nítrico sintase e
a via de sinalização de NF-κB. Além disso, a fração regulou
positivamente a expressão do inibidor de κB (IκBα) e bloqueou
a translocação nuclear de NF-κB.
A atividade antiinflamatória in vivo de três extratos das
folhas de moringa e suas frações mais ativas foram avaliados
em ratos com edema de pata induzido por carragenina. Dos três
extratos, o etanólico 95% (250 mg/kg) inibiu de modo
significativo o edema da pata comparativamente ao controle, no
qual o efeito teve início na primeira hora após a injeção da
carragenina, com atividade máxima em 3 horas após a
aplicação, resultando no aumento de edema de 33,377±
0,377% (MAHDI et al., 2017). Adicionalente, diferentes extratos
das folhas de moringa foram avaliados em ratos Wistar por
modelo de artrite induzida por formaldeído e edema de pata nas
doses de 150, 300 e 600 mg/kg (SALEEM; SALEEM; AKHTAR,
2018). O extrato metanólico da planta foi o que exerceu a
máxima inibição percentual do volume da pata (82,28 ± 0,07%)
na dose de 600 mg/kg. A inibição foi comparativamente maior
que aquela promovida pelo controle positivo piroxicam (77,75 ±
0,37%) e pelos extratos aquoso (80,3 ± 0,47%) e acetato de

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
125
etila (67,79 ± 0,25%). No modelo de artrite, o tratamento com
todos os extratos ou piroxicam, reduziu o nível de leucócitos
totais e plaquetas com maior atividade para o extrato
metanólico. Além disso, os tratamentos aumentaram a
contagem de hemoglobina e hemácias que havia diminuído
com a indução da artrite.
Atividade anticâncer
O potencial anticâncer das folhas de M. oleifera também
tem sido demonstrado a partir de diferentes abordagens
experimentais. Potestà et al. (2019), observaram que o efeito
citotóxico do extrato aquoso das folhas foi dependente da dose
(a partir de 0,01 mg/mL) e do tipo celular, uma vez que as
células de Jukart (linfoblastos leucêmicos de células T) e
células THP1 (monócitos leucêmicos) foram mais suscetíveis
aos tratamentos que as PBMCs (células mononucleares de
sangue periférico humano). O extrato aquoso das folhas de M.
oleifera também induziu apoptose em células HepG2
(hepatocarcinoma humano) e a administração oral do extrato,
durante quatro dias, reduziu significativamente (44‐52%) a
proliferação das células HepG2 e células A549 (câncer de
pulmão) em camundongos (JUNG; LEE; KANG, 2015).
Sadek et al. (2017), mostraram que o extrato etanólico
das folhas de moringa possui forte atividade anticâncer contra
carcinoma hepatocelular, induzido por dietil nitrosamina em
ratos Wistar, principalmente por induzir apoptose e melhorar a
atividade antioxidante.
Tragulpakseerojn et al. (2017), investigaram o efeito
antiproliferativo in vitro e o mecanismo de ação do extrato
alcoólico das folhas de M. oleifera contra células de carcinoma

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
126
de cólon humano (HCT116). Observou-se que a proliferação
celular foi significativamente inibida de forma dependente da
concentração. Além disso, a atividade antiproliferativa foi
mediada pela regulação negativa da fosforilação do ERK1/2. O
extrato aquoso das folhas também provocou a diminuição
significativa da sobrevivência de células MDA-MB-231 (câncer
de mama) e HCT-8 (adenocarcioma). Em ambas as linhagens
o percentual de inibição celular foi de 90% no tratamento na
concentração de 500 μg/mL (AL-ASMARI et al., 2015).
Atividade antimicrobiana
Elgamily et al. (2016), avaliaram três extratos de
diferentes partes de M. oleifera contra patógenos orais usando
o método de difusão em disco. Além disso, incorporaram a
preparação etanólica das folhas em formulações de creme
dental e anti-séptico bucal experimentais. Os extratos
produzidos com etanol, acetona e acetato de etila causaram,
respectivamente, zonas de inibição do crescimento de
Staphylococcus aureus (19,25 ± 0,96, 11,25 ± 0,96 e 9,00 ± 0,82
mm) e de Streptococcus mutans (13,00 ±1,15, 8,25 ± 0,50 e
7,25 ± 0,96 mm). Por outro lado, os extratos não foram
eficientes em inibir o crescimento de Candida albicans. O creme
dental experimental inibiu o crescimento de S. aureus, S.
mutans e C. albicans com zonas de inibição de 17,75 ± 0,96,
13,75 ± 0,96 e 13,25 ± 1,5 mm, respectivamente. O anti-séptico
bucal foi um agente bacteriostático menos eficiente que o creme
dental (zonas de inibição de 11,25±1,26 e 9,25±0,96 contra S.
aureus e S. mutans) e não ativo contra C. albicans.

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
127
A atividade antibacteriana do extrato etanólico das folhas
de M. oleifera (50, 100, 150 e 200 μL/poço), foi examinado
contra bactérias gram positivas (S. aureus) e gram negativas
(Escherichia coli) usando o método de difusão em disco
(MALHOTRA; MANDAL, 2018). Os resultados mostraram que o
extrato teve atividade antibacteriana contra as duas espécies,
sendo as zonas de inibição na concentração 200 μL/poço de 25
mm e 22 mm para S. aureus e E. coli, respectivamente.
Falowo et al. (2016) avaliaram a atividade antibacteriana
do extrato hidroalcoólico das folhas de M. oleifera, através da
determinação da concentração mínima inibitória (CMI) e pela
contagem do número de colônias bacterianas em carne bovina
moída, armazenada durante 6 dias. A amostra de carne fresca
moída foi tratada com o conservante químico hidroxitolueno
butilado (BHT) e extrato de M. oleifera (1g/kg). O ensaio in vitro
revelou atividade antibacteriana contra as espécies testadas,
com CMI variando de 2,5 a 10,0 mg/mL, sendo o maior efeito
detectado contra Bacillus cereus. A contaminação bacteriana e
quantidade de ácido láctico na carne moída tratada com o
extrato foram menores que aquele da carne não tratada
(controle negativo) e tratada com BHT (controle positivo).
Ademais, extratos das folhas de M. oleifera também
foram capazes de inibir a replicação do vírus da
imunodeficiência humana (HIV), do vírus herpes (HSV-1) e do
vírus da febre aftosa (FMDV) (KUROKAWA et al., 2016). Os
mecanismos de inibição da replicação viral dos extratos das
folhas envolve ligação a proteínas e enzimas, bem como
ativação da imunidade celular.

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
128
Atividade imunomoduladora
El Shanawany et al. (2019) avaliaram o efeito
imunomodulador dos extratos aquoso e hidroalcóolico das
folhas de M. oleifera, os quais aumentaram substancialmente a
resposta imune celular e humoral, o índice de neutrófilos e a
atividade fagocítica. Kurokawa et al. (2016) mostraram que o
extrato aquoso estimulou a imunidade celular, ao aumentar as
proporções de células CD11+/CD49 + entre os esplenócitos de
camundongos tratados em relação ao grupo controle.
Outro estudo avaliou os efeitos imunomoduladores do
extrato hidrometanólico das folhas de M. oleifera em ratos
Wistar machos divididos aleatoriamente em oito grupos de seis
animais. O grupo I serviu como controle positivo. Os grupos II,
III e IV receberam doses orais de 125, 250 e 500 mg/kg do
extrato, respectivamente, por 15 dias. Ciclofosfamida (CTX) na
dose de 30 mg/kg foi administrada por via oral aos Grupos V
(controle negativo), VI, VII e VIII durante os primeiros 3 dias.
Posteriormente, doses de 125, 250 e 500 mg/kg do extrato
foram administradas, respectivamente, por mais 15 dias. Nos
dias 16 e 19, amostras de sangue foram coletadas por punção
cardíaca e analisadas para contagem de glóbulos vermelhos,
concentração de hemoglobina, contagem total de leucócitos,
percentuais diferenciais de leucócitos e enzimas hepáticas ALT,
AST e fosfatase alcalina (ALP). Nos Grupos II, III e IV houve
aumento dependente da dose na contagem total de leucócitos
(TWBC) e porcentagens de neutrófilos, eosinófilos, monócitos e
linfócitos e redução dos níveis de enzimas hepáticas no soro
(ALT, AST e ALP) em comparação com o Grupo I. A
administração de CTX reduziu significativamente a contagem

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
129
de TWBC e as porcentagens de neutrófilos, eosinófilos,
monócitos, linfócitos e aumentou as enzimas hepáticas séricas
(ALT, AST, ALP) nos Grupos VI, VII e VIII quando comparados
ao Grupo V. Os efeitos do CTX no sangue e nas enzimas
hepáticas foram revertidos com a administração dos extratos de
maneira dose-dependente (OBI et al., 2018).
Polissacarídeos extraídos de M.oleifera tem ganhado
atenção científica devido ao seu excelente potencial
biotecnológico. Por exemplo, o polissacarídeo das folhas MOP-
1 (peso molecular 7,65 × 107 kDa) constituído por ramnose,
arabinose e galactose, exibiu potencial imunorregulador (HE et
al., 2018). Um novo polissacarídeo (MOP-2) também das folhas
(peso molecular 155,35 kDa) foi um agente imunomodulador
em células de macrófagos, aumentando a secreção de ROS,
NO, IL-6, TNF-α e iNOS no grupo tratado em comparação ao
grupo controle (DONG et al., 2018).
Atividade protetora
Estudos in vitro e in vivo tem relatado o potencial protetor
das folhas de M. oleifera. O extrato aquoso foi um agente
citoprotetor in vitro para eritrócitos devido sua atividade
antioxidante. Eritrócitos preparados foram incubados com o
reagente de Fenton na ausência ou presença do extrato.
Observou-se que os eritrócitos pré-tratados com o extrato exibiu
um grau reduzido de hemólise in vitro (SARKAR et al., 2017).
Abd-Elhakim et al. (2018) mostraram que o extrato (800
e 700 mg/kg) administrado por via oral uma vez por dia durante
2 semanas, neutralizou significativamente os impactos
hematoimunotóxicos promovidos pela melamina, um perigoso
contaminante alimentar e adulterante amplamente difundido.

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
130
A despeito da proteção a nível celular, preparações
obtidas a partir das folhas também apresentaram efeito protetor
para órgãos. O extrato aquoso (100 mg/kg) administrado por via
oral em dose única, restaurou o dano cardíaco promovido pelo
fosforeto de alumínio em ratos. O fosforeto de alumínio causou
alterações histopatológicas cardíacas significativas com
aumento no malondialdeído (MDA), marcador de peroxidação
lipídica, e uma depleção significativa de enzimas antioxidantes
(catalase e glutationa redutase). No entanto, o tratamento com
o extratos das folhas protegeu de forma eficiente o tecido
cardíaco de ratos intoxicados, aumentando os níveis de
antioxidantes com leve diminuição na produção de MDA em
comparação com o grupo não tratado. (GOUDA; EL-
NABARAWY; IBRAHIM, 2018).
Airaodion et al. (2019) avaliaram a eficiência
hepatoprotetora do extrato etanólico das folhas de M. oleifera
contra exposição a hidrocarbonetos (petróleo bruto) pela
determinação de biomarcadores de dano hepático. Os autores
administraram 10 g de extrato por 30 dias em ratos, detectaram
redução das enzimas alanina transaminase (ALT), fosfatase
alcalina (ALP), aspartato aminotransferase (AST) e lactato
desidrogenase. O petróleo bruto aumentou significativamente
os biomarcadores hepáticos, mas este efeito foi neutralizado
pelo extrato das folha de M. oleifera.
Outras atividades relevantes
Almuzafar (2018) avaliou o efeito antiulcerativo e
antioxidante de duas doses (100 e 500 mg/kg) do extrato
etanólico das folhas de M. oleifera em ratos Wistar machos com

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
131
lesão oxidativa gástrica induzida por indometacina
(medicamento anti-inflamatório) e etanol durante duas
semanas. O grupo tratado apenas com indometacina mostrou
aumentos significativos no índice de lesão (IL), no nível de
malondialdeído (MDA) e diminuição nas atividades de
superóxido dismutase (SOD), glutationa-S-transferase (GST) e
glutationa peroxidase (GPx), quando comparado ao grupo
controle. As duas doses do extrato testadas foram capazes de
reduzir o IL estomacal e o marcador de estresse oxidativo
(MDA), e aumentaram significativamente os biomarcadores
antioxidantes (SOD, GST e GPx) em comparação ao grupo
tratado apenas com indometacina. O autor concluiu que o
extrato tem uma atividade antiúlcera e antioxidante eficaz,
eliminando radicais livres e protegendo o estômago contra a
ulceração, efeito colateral da indometacina.
Preparações obtidas a partir das folhas também tem
apresentado atividade cicatrizante a partir do aumento
significativo da proliferação celular e viabilidade e migração de
fibroblastos dérmicos humanos. Estudo relativo ao potencial de
extrato das folhas em cicatrizar feridas de animais diabéticos,
mostraram aumento de regeneração tecidual, diminuição da
área das feridas, redução nos níveis de mediadores
inflamatórios e aumento do fator de crescimento endotelial
vascular (MUHAMMAD et al., 2016). Outro estudo mostrou que
uma formulação de creme contendo 3% do extrato das folhas
de M. oleifera aumentou o efeito de revitalização da pele em
humanos sobre parâmetros como textura, rugosidade e
suavidade da pele (ALI; A AKHTAR; CHOWDHARY, 2014).
George, Ajayi, Oyemike (2018) demonstraram a
eficiência do extrato aquoso das folhas de M. oleifera no

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
132
controle da hipertensão arterial sistêmica e ocular em trinta
humanos adultos normotensivos (idade média de 28,93 ± 1,62)
de maneira dose-dependente. O efeito anti-hipertensivo do
extrato aquoso das folhas em ratos pode ser mediado pelo
alívio da disfunção vascular e do estresse oxidativo, bem como
pela promoção do vasorrelaxamento dependente do endotélio.
Adicionalmente, Kaur et al. (2015) avaliaram os efeitos
comportamentais e antidepressivos agudos e crônicos do
extrato alcoólico das folhas de Moringa oleifera (MOE) em
modelos padronizados de depressão em camundongos albinos.
Os animais foram divididos em seis grupos: Grupos 1 e 2
serviram como controle de veículo e controle padrão de
fluoxetina (20 mg /kg), respectivamente. Os grupos 3 e 4
serviram como grupos de tratamento e foram administrados por
via oral com MOE (100 e 200 mg/kg). Os grupos 5 e 6,
respectivamente, receberam doses combinadas de MOE 100
mg/kg + 10 mg de fluoxetina e MOE 200 mg/ kg + 10 mg/kg de
fluoxetina. Após tratamentos agudos e crônicos de 14 dias,
todos os animais foram testados usando modelos
comportamentais de depressão, como teste de natação forçada
(FST), teste de suspensão da cauda (TST) e teste de atividade
locomotora (LAT). Alterações significativas em todas as
atividades testadas (FST, TST, LAT) de camundongos tratados
cronicamente foram observadas, especialmente em animais
que receberam doses combinadas simultaneamente de 200
mg/kg/dia de MOE + 10 mg/kg /dia de fluoxetina por 14 dias.
Ademais, o efeito antidepressivo do MOE pode ter sido
provocado por meio da via de neurotransmissão
noradrenérgica-serotonérgica.

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
133
CONCLUSÕES
Estudos in vitro e in vivo utilizando folhas de M. oleifera
certificam propriedades biológicas interessantes do ponto de
vista de sáude humana. O uso das folhas pode ser uma
alternativa medicamentosa para o tratamento de diversas
doenças. Adicionalmente, as folhas podem ser usadas no
combate a desnutrição por seu alto valor nutricional.
REFERÊNCIAS BIBLIOGRÁFICAS
AIRAODION, A. I.; OGBUAGU, U.; EKENJOKU, J. A.; OGBUAGU, E. O.; AIRAODION, E. O.; OKOROUKWU, V. N. Hepato-protective Efficiency of Ethanol Leaf Extract of Moringa oleifera Lam. against Hydrocarbon Exposure. International Journal of Advances in Herbal and Alternative Medicine, v. 3, p. 32-41, 2019. AL-ASMARI, A. K.; ALBALAWI, S. M.; ATHAR, M. T.; KHAN, A. Q.; AL-SHAHRANI, H.; ISLAM, M. Moringa oleifera as an anti-cancer agent against breast and colorectal cancer cell lines. PloS one, v. 10, n. 8, p. e0135814, 2015. ALI, A.; AKHTAR, N.; CHOWDHARY, F. Enhancement of human skin facial revitalization by moringa leaf extract cream. Advances in Dermatology and Allergology/Postȩpy Dermatologii i Alergologii, v. 31, n. 2, p. 71, 2014. ALMUZAFAR, H. M. Effect of Moringa oleifera leaves extract on the oxidative stress and gastric mucosal ulcer induced by indomethacin in rats. African Journal of Biotechnology, v. 17, n. 3, p. 51-56, 2018. ARULSELVAN, P.; TAN, W. S.; GOTHAI, S.; MUNIANDY, K.; FAKURAZI, S.; ESA, N. M.; KUMAR, S. S. Anti-inflammatory potential of ethyl acetate fraction of Moringa oleifera in downregulating the NF-κB signaling pathway in lipopolysaccharide-stimulated macrophages. Molecules, v. 21, n. 11, p. 1452, 2016. COZ-BOLAÑOS, X.; CAMPOS-VEGA, R.; REYNOSO-CAMACHO, R.; RAMOS-GÓMEZ; M., LOARCA-PIÑA, G. F.; GUZMÁN-MALDONADO, S. H. Moringa infusion (Moringa oleifera) rich in phenolic compounds and high antioxidant capacity attenuate nitric oxide pro-inflammatory mediator in vitro. Industrial Crops and Products, v. 118, p. 95-101, 2018. DONG, Z.; LI, C.; HUANG, Q.; ZHANG, B.; FU, X.; LIU, R. H. Characterization of a novel polysaccharide from the leaves of Moringa

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
134
oleifera and its immunostimulatory activity. Journal of Functional Foods, v. 49, p. 391-400, 2018. EL SHANAWANY, E. E.; FOUAD, E. A.; KESHTA, H. G.; HASSAN, S. E.; HEGAZI, A. G.; ABDEL-RAHMAN, E. H. Efeitos imunomoduladores do extrato aquoso das folhas de Moringa oleifera em ovinos naturalmente co-infectados com Fasciola gigantica e Clostridium novyi. Journal of Parasitic Diseases , v. 43, n. 4, pág. 583-591, 2019. ELGAMILY, H.; MOUSSA, A.; ELBORAEY, A.; EL-SAYED, H.; AL-MOGHAZY, M.; ABDALLA, A. Microbiological assessment of Moringa oleifera extracts and its incorporation in novel dental remedies against some oral pathogens. Open access Macedonian journal of medical sciences, v. 4, n. 4, p. 585, 2016. FALOWO, A. B.; MUCHENJE, V.; HUGO, C. J.; CHARIMBA, G. In vitro antimicrobial activities of Bidens pilosa and Moringa oleifera leaf extracts and their effects on ground beef quality during cold storage. Cyta-Journal of Food, v. 14, n. 4, p. 541-546, 2016. FARD, M. T.; ARULSELVAN, P.; GOVINDARAJAN, K. S. K. A.; FAKURAZI, S. Bioactive extract from Moringa oleifera inhibits the pro-inflammatory mediators in lipopolysaccharide stimulated macrophages. Pharmacognosy magazine, v. 11, n. Suppl 4, p. S556, 2015. GEORGE, G. O.; AJAYI, O. B.; OYEMIKE, A. A. Effect of Moringa oleifera leaf aqueous extract on intraocular and blood pressure of normotensive adults in Edo State, Nigeria. Journal of The Nigerian Optometric Association, v. 20, n. 2, p. 75-81, 2018. GOUDA, A. S.; EL-NABARAWY, N. A.; IBRAHIM, S. F. Moringa oleifera extract (Lam) attenuates Aluminium phosphide-induced acute cardiac toxicity in rats. Toxicology reports, v. 5, p. 209-212, 2018. HE, T. B.; HUANG, Y. P.; HUANG, Y.; WANG, X. J.; HU, J. M.; SHENG, J. Structural elucidation and antioxidant activity of an arabinogalactan from the leaves of Moringa oleifera. International journal of biological macromolecules, v. 112, p. 126-133, 2018. HELMY, S. A.; MORSY, N. F.; ELABY, S. M.; GHALY, M. A. Hypolipidemic effect of Moringa oleifera Lam leaf powder and its extract in diet-induced hypercholesterolemic rats. Journal of medicinal food, v. 20, n. 8, p. 755-762, 2017. IRFAN, H. M.; ASMAWI, M. Z.; KHAN, N. A. K.; SADIKUN, A.; MORDI, M. N. Anti-diabetic activity-guided screening of aqueous-ethanol Moringa oleifera extracts and fractions: Identification of marker compounds. Tropical Journal of Pharmaceutical Research, v. 16, n. 3, p. 543-552, 2017. JUNG, I. L.; LEE, J. H.; KANG, S. C. A potential oral anticancer drug candidate, Moringa oleifera leaf extract, induces the apoptosis of human

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
135
hepatocellular carcinoma cells. Oncology letters, v. 10, n. 3, p. 1597-1604, 2015. KAUR, G.; INVALLY, M.; SANZAGIRI, R.; BUTTAR, H. S. Evaluation of the antidepressant activity of Moringa oleifera alone and in combination with fluoxetine. Journal of Ayurveda and integrative medicine, v. 6, n. 4, p. 273, 2015. KHAN, W.; PARVEEN, R.; CHESTER, K.; PARVEEN, S.; AHMAD, S. Hypoglycemic potential of aqueous extract of Moringa oleifera leaf and in vivo GC-MS metabolomics. Frontiers in pharmacology, v. 8, p. 577, 2017. KOU, X.; LI, B.; OLAYANJU, J. B.; DRAKE, J. M.; CHEN, N. Nutraceutical or pharmacological potential of Moringa oleifera Lam. Nutrients, v. 10, n. 3, p. 343, 2018. KUROKAWA, M.; WADHWANI, A.; KAI, H.; HIDAKA, M.; YOSHIDA, H.; SUGITA, C; HAGIWARA, A. Activation of cellular immunity in herpes simplex virus type 1‐infected mice by the oral administration of aqueous extract of Moringa oleifera Lam. leaves. Phytotherapy Research, v. 30, n. 5, p. 797-804, 2016. LEONE, A.; SPADA, A.; BATTEZZATI, A.; SCHIRALDI, A.; ARISTIL, J.; BERTOLI, S. Cultivation, Genetic, Ethnopharmacology, Phytochemistry and Pharmacology of Moringa oleifera Leaves: An Overview. Int J Mol Sci. 2015. MAHDI, H. J.; KHAN, N. A. K.; MAHMUD, R.; ASMAWI, M. Z. B.; VIKNESWARAN, A.; MURUGAIYAH, L. LC/MS, GC/MS screening and in vivo anti-inflammatory activity of Malaysian Moringa oleifera Lam leaf extracts and fractions against carrageenan-induced paw oedema in rats. Journal of Innovations in Pharmaceutical and Biological Sciences, v. 4, n. 3, p. 48-54, 2017. MALHOTRA, S. P. K.; MANDAL, T. K. Phytochemical screening and in vitro antibacterial activity of Moringa oleifera (Lam.) leaf extract. Archives of Agriculture and Environmental Science, v. 3, n. 4, p. 367-372, 2018 MANOJ, M. A. D. M. R.; RANJITHA, G. M. K. PHYTOREMEDIATION of E-Waste using Nicotiana Tabacum & Moringa oleifera. International Journal for Scientific Research & Development. v. 7, 2020. MUHAMMAD, Abubakar Amali et al. Evaluation of wound healing properties of bioactive aqueous fraction from Moringa oleifera Lam on experimentally induced diabetic animal model. Drug design, development and therapy, v. 10, p. 1715, 2016. NAIR, D. A.; JAMES, T. J.; SREELATHA, S. L.; KARIYIL, B. J. Antioxidant and antiproliferative properties of Moringa oleifera Lam. leaf aqueous extract. Plant Science Today, v. 7, n. 4, p. 649–657-649–657, 2020. OBI, A.; EGWURUGWU, J. N.; OJEFA, S. O.; OHAMAEME, M. C.; EKWEOGU, C. N.; OGUNNAYA, F. U. Immunomodulatory effects of

PROPRIEDADES FARMACOLÓGICAS DAS FOLHAS DE Moringa oleifera Lam.
136
hydromethanolic extract of Moringa oleifera leaf on male wistar rats. Nigerian Journal of Experimental and Clinical Biosciences, v. 6, n. 1, p. 26, 2018. PARE, D.; HILOU, A.; OUEDRAOGO, N.; GUENNE, S. Ethnobotanical study of medicinal plants used as anti-obesity remedies in the nomad and hunter communities of Burkina Faso. Medicines, v. 3, n. 2, p. 9, 2016. POTESTÀ, M.; MINUTOLO, A.; GISMONDI, A.; CANUTI, L.; KENZO, M.; ROGLIA, V.; MONTESANO, C. Cytotoxic and apoptotic effects of different extracts of Moringa oleifera Lam on lymphoid and monocytoid cells. Experimental and Therapeutic Medicine, v. 18, n. 1, p. 5-17, 2019. SADEK, K. M.: ABOUZED, T. K.; ABOUELKHAIR, R.; NASR, S. The chemo-prophylactic efficacy of an ethanol Moringa oleifera leaf extract against hepatocellular carcinoma in rats. Pharm Biol. 2017 SALEEM, A.; SALEEM, M.; AKHTAR, M. F.; SHAHZAD, M.; JAHAN, S. Moringa rivae leaf extracts attenuate complete Freund’s adjuvant-induced arthritis in Wistar rats via modulation of inflammatory and oxidative stress biomarkers. Inflammopharmacology, v. 28, n. 1, p. 139-151, 2020. SARKAR, M.; BHOWMICK, S.; HUSSAIN, J.; HASAN, M.; HOSSAIN, S.Hot Water Extract of Moringa oleifera Leaves Protects Erythrocytes from Hemolysis and Major Organs from Oxidative Stress in vitro. J. basic appl. Res, v. 3, n. 3, p. 120-126, 2017. TRAGULPAKSEEROJN, J.; YAMAGUCHI, N.; PAMONSINLAPATHAM, P.; WETWITAYAKLUNG, P.; YONEYAMA, T.; ISHIKAWA, N.; APIRAKARAMWONG, A. Anti-proliferative effect of Moringa oleifera Lam (Moringaceae) leaf extract on human colon cancer HCT116 cell line. Tropical Journal of Pharmaceutical Research, v. 16, n. 2, p. 371-378, 2017. VATS, S.; GUPTA, T. Evaluation of bioactive compounds and antioxidant potential of hydroethanolic extract of Moringa oleifera Lam. from Rajasthan, India. Physiology and molecular biology of plants, v. 23, n. 1, p. 239-248, 2017

137
CIÊNCIAS BIOLÓGICAS - ECOLOGIA

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
138
CAPÍTULO 7
O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO
DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
Rafael Xavier MARTINS 1
Iris Flávia de Sousa GONÇALVES 2
Juliana Alves da Costa Ribeiro SOUZA 3 Marília da Guia Flor SILVA 4
Davi Felipe FARIAS 5
1 Mestrando em Biologia Celular e Molecular, UFPB; 2 Mestranda em Bioquímica, UFC; 3
Mestre em Ciência Animal, UFPB; 4 Graduanda em Farmácia, UFPB; 5 Orientador/ Professor DBM/ UFPB.
Email:[email protected]
RESUMO: O ácido 2,4 Diclorofenoxiacético (2,4-D) é um herbicida auxínico utilizado à nível mundial e é ingrediente ativo de mais de 1.500 produtos disponíveis no mercado. Presume-se que somente 0,1% dos agrotóxicos aplicados nas lavouras ao redor do mundo exerçam efeito no alvo desejado, acarretando em um grande acúmulo de resíduos de agrotóxicos e seus metabólitos. Os peixes são organismos sensíveis às alterações de parâmetros ambientais e vêm sendo impactados pela grande quantidade de resíduos da agroindústria. O objetivo desse trabalho foi realizar uma revisão sistemática sobre os efeitos adversos decorrentes da exposição ao 2,4-D em espécies de peixes. Os relatos foram identificados através de buscas na base de dados PubMed, utilizando duas combinações de palavras-chaves: (a)“2,4-D” e “fish” e (b) “2,4-D” e “zebrafish”. Posteriormente, os artigos foram filtrados de acordo com os critérios de elegibilidade. No total, foram encotrados 39 artigos, dos quais 16 se adequaram aos critérios

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
139
estabelecidos. Foram identificadas seis espéceis de peixes utilizadas como modelo, sendo Danio rerio e Cnesterodon decemmaculatus as mais utilizadas. Os efeitos adversos relatados foram: neurotoxicidade, alterações comportamentais, genotoxicidade, estresse oxidativo, cardiotoxicidade e desregulação endócrina. A exposição ao 2,4-D altera parâmetros bioquímicos, fisiológicos e comportamentais que podem colocar em risco a sobrevivência das espécies e desequilibrar os ecossistemas nos quais estão inseridas. Palavras-chave: Ecotoxicidade. Herbicida. Revisão sistemática. Vertebrados Aquáticos.
INTRODUÇÃO
No início da década de 1950, o surgimento de novas
tecnologias e a necessidade de atender à crescente demanda
de alimentos da população mundial impulsionaram a
implantação de um novo modelo de produção agrícola (LOPES
et al., 2018). Uma das principais características desse novo
formato é a utilização em massa de agrotóxicos, substâncias
que visam combater doenças e pragas causadas por insetos,
bactérias, fungos e plantas espontâneas (CARVALHO, 2017).
Dentre as diferentes classes de pesticidas, os produtos com
finalidade herbicida são os mais utilizados, representando cerca
de 47,5% dos agrotóxicos aplicados nas lavouras em todo o
mundo (SHARMA et al., 2019).
O ácido 2,4 diclorofenoxiacético (2,4-D), um químico
pertencente à classe dos ácidos fenoxiacéticos, foi um dos
primeiros herbicidas sintéticos a ser introduzido no mercado, no
ano de 1946, e atualmente é ingrediente principal de mais de
1.500 produtos disponíveis comercialmente (ISLAM et al., 2017;
TIWARI et al., 2019).

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
140
Este composto foi arquitetado para mimetizar o
mecanismo de ação das auxinas, hormônios responsáveis pelo
crescimento das plantas. Usado em baixas concentrações (20-
50 mg/L), é capaz de induzir a divisão celular, acelerando o
desenvolvimento das plantas e melhorando o aspecto visual e
durabilidade de frutos. Contudo, quando usado em altas
concentrações (maiores que 500 mg/L) exerce um efeito
herbicida, matando seletivamente plantas dicotiledôneas (LI et
al., 2017; TIWARI et al., 2019). Devido ao caráter seletivo, esse
composto é utilizado majoritariamente no combate de plantas
espontâneas em culturas monocotiledóneas, tais como trigo,
aveia, cana-de-açúcar, milho e arroz. Também é usado para
matar plantas espontâneas na fase de pré-plantio de algumas
culturas dicotiledóneas, como por exemplo a soja (GAAIED et
al., 2019).
Além do baixo custo, o 2,4-D possui uma fácil
solubilização em água e outros tipos de solventes, facilitando a
sua penetração nas estruturas das plantas o tornando um
herbicida altamente eficaz, o que explica a sua utilização
massiva em diversos tipos de lavouras ao redor do mundo
(ISLAM et al., 2017).
O principal foco mercadológico do 2,4-D são os Estados
Unidos, América do Sul, Europa e Rússia, onde o consumo
aumentou cerca de 40% em um intervalo de 9 anos (ISLAM et
al., 2017). Nos Estados Unidos, de acordo com dados referente
aos anos 1992 a 2000, foram usados mais de 20.000.000 ton.
desse composto nas lavouras do país a cada ano, ocupando
por muito tempo a posição de terceiro agrotóxico mais utilizado
pelo país (LI et al., 2017).

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
141
Já no Brasil, dados oficiais disponibilizados pelo Instituto
Brasileiro do Meio Ambiente e dos Recursos Naturais (IBAMA),
mostram que o 2,4-D foi o segundo herbicida mais
comercializado em 2017 (ficando atrás apenas do glifosato)
sendo vendidas cerca de 57.389,35 ton. desse ativo no país
(IBAMA, 2017).
Diversos estudos demonstram que os agrotóxicos não
atingem apenas as culturas em que são aplicados
(GONÇALVES et al., 2020). Presume-se que somente 0,1%
dos agrotóxicos aplicados nas lavouras ao redor do mundo
exerçam efeito no alvo desejado, acarretando em um grande
acúmulo de resíduos de agrotóxicos e seus metabólitos no
ambiente (ISLAM et al., 2017). Além da possibilidade de serem
carreados pelo ar, esses resíduos químicos podem ser
lixiviados através do solo, ter acesso a corpos aquáticos, causar
efeitos deletérios em organismos não alvo e modificar a
dinâmica dos ecossistemas (LI et al., 2017 GONÇALVES et al.,
2020).
Dados atuais mostram que em ambientes de água doce
a concentração estimada de 2,4-D pode variar entre 4 a 24 µg/
L, chegando até 4.000 µg/L em áreas próximas à culturas onde
o herbicida é aplicado. Além disso, esse herbicida possui uma
meia vida de aproximadamente 15 dias, podendo variar até 300
dias devido à parâmetros de qualidade da água e outras
condições ambientais (GAAIED et al., 2019; DEHNERT et al.,
2019).
Os peixes, componentes que integram a biota aquática,
são organismos sensíveis a alterações de parâmetros
ambientais e vêm sendo impactados diretamente pelo modelo

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
142
agroindustrial predominante no mundo atualmente (RUIZ DE
ARCAUTE et al., 2016).
Existe uma correlação positiva entre o declínio
significativo da polulação de peixes nos últimos anos em todo o
mundo e o aumento alarmante do uso de produtos pesticidas
pelo setor agrícola. Provavelmente, essa consequência está
relacionada à alta capacidade que esses organismos possuem
de acumular e metabolizar poluentes químicos (RUIZ DE
ARCAUTE et al., 2016).
Mais especificamente, existe uma preocupação a cerca
dos possíveis impactos negativos do 2,4-D sobre diferentes
espécies de peixes. Geralmente, os períodos de aplicação de
herbicidas que tem o 2,4-D como princípio ativo nas lavouras
coincidem com o período de desova desses organismos, e essa
prática os expõem em seus estágios iniciais de vida
(embrionário e larval) que são os mais vulneráveis à efeitos
adversos de xenobióticos (DEHNERT et al., 2019).
Isso pode influenciar parâmetros bioquímicos,
fisiológicos e comportamentais que sejam vitais para a
sobrevivência das espécies afetadas e alterar
significativamente a dinâmica dos ecossistemas nos quais elas
habitam. Sendo assim, essa revisão teve como objetivo analisar
a literatura recente sobre os efeitos adversos resultantes da
exposição de diferentes espécies de peixes ao 2,4-D e a
formulações comerciais contendo este princípio ativo.

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
143
MATERIAIS E MÉTODOS
Pesquisa bibliográfica
O intuito da pesquisa bibliográfica foi identificar os
trabalhos que buscaram avaliar os impactos do herbicida 2,4-D
sobre pelo menos uma espécie de peixe. As buscas foram feitas
na base de dados PubMed (https://pubmed.ncbi.nlm.nih.gov) e
foram realizadas no período de setembro de 2020 a outubro
desse mesmo ano.
Os artigos publicados nos últimos 5 anos foram reunidos
a partir de buscas usando as seguintes combinações de
palavras-chaves: “2,4-D” e “fish”; “2,4-D” e “zebrafish”. O motivo
da especificidade da segunda combinação de palavras-chave
foi a tentativa de inclusão de trabalhos de relevância que
utilizaram o Danio rerio (modelo de referência em estudos de
toxicologia) e que não foram identificados com a primeira
combinação de palavras-chaves. Inicialmente, o título e o
resumo dos trabalhos foram lidos, as duplicatas foram excluídas
e posteriormente houve a leitura dos artigos completos. Após
as buscas, as duplicatas foram excluídas.
Critérios de elegibilidade
Após a leitura dos artigos completos, os trabalhos
passaram por uma curadoria obedecendo aos seguintes
critérios de inclusão e exclusão: (i) critérios de inclusão: artigos
de pesquisa original; estudos que utilizaram o 2,4-D puro ou
alguma formulação comercial que tenha esse herbicida como
princípio ativo; estudos que utilizaram pelo menos uma espécie

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
144
de peixe como modelo; artigos que avaliaram os efeitos de
forma isolada ao 2,4-D bem como associados a outros
agrotóxicos. (ii) critérios de exclusão: artigos de revisão, artigos
que não utilizaram o 2,4-D (puro ou formulação comercial),
artigos que realizaram exposições com amostras coletadas no
ambiente; artigos que não utilizaram pelo menos uma espécie
de peixe como modelo; artigos que avaliaram os efeitos do 2,4-
D associado a outros agrotóxicos, mas não avaliaram seus
efeitos de forma isolada.
Os artigos selecionados para leitura completa foram lidos
e os dados relevantes para esta revisão foram extraídos para
uma tabela contendo as seguintes informações: nome do autor
principal, ano de publicação, espécies utilizadas, estágio de
vida, tipo de exposição (crônica e/ou aguda) e efeitos adversos
observados.
RESULTADOS E DISCUSSÃO
Pesquisa bibliográfica
As combinações de palavras-chave forneceram
diferentes perfis de resultados. Com o uso da combinação “2,4-
D” e “fish” foram encontrados 31 artigos, 16 foram selecionados
pois se adequaram aos critérios de elegibilidade. Já com o uso
da combinação “2,4-D” e “zebrafish” foram encontrados 8
artigos, dos quais 7 foram selecionados.
De forma geral, as buscas com as duas combinações de
palavras-chave resultaram em um total de 39 artigos e, após a
curadoria e exclusão das duplicatas, 16 se enquadraram no
perfil buscado para a realização desse trabalho.

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
145
Compostos químicos
Dentre os 16 artigos avaliados, além do 2,4-D em sua
forma pura, foi possível observar uma diversidade de
formulações químicas que tem esse herbicida como princípio
ativo.
Enquanto 43,35% do total de trabalhos analisados
utilizaram esse químico puro em seus experimentos,
aproximadamente 68,75% utilizaram formulação comerciais
que o tem como base. Os herbicidas comerciais utilizados
foram: DMA, DMA4, WAM40, Dedalo elite, Weedar, e Herbextra
e U46 BR.
É importante pontuar que alguns trabalhos usaram mais
de uma formulação comercial do 2,4-D, e/ou o próprio princípio
ativo, a fim de comparar os efeitos adversos provocados no
animal modelo em estudo. Além disso, também foram
identificados estudos que visaram avaliar os efeitos do 2,4-D
tanto isoladamente quanto em conjunto com outros químicos,
com o intuito de observar possíveis efeitos sinergísticos ou
antagônicos entre as substâncias.
Espécies de peixes utilizadas como modelo
Enquanto 15 dos trabalhos selecionados usaram apenas
um único tipo de animal modelo, apenas dois utilizaram uma
abordagem multi-espécie. Ambos os artigos utilizaram duas
espécies de peixes em seus ensaios de exposição ao 2,4-D.
No total, foram identificadas seis espécies de peixes
diferentes. Foram elas: Danio rerio, Cnesterodon

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
146
decemmaculatus, Pimephales promelas, Perca flavescens,
Oreochromis niloticus e Hyphessobrycon eques.
Os modelos mais utilizados foram o D. rerio (7 artigos) e
o C. decemmaculatus (5 artigos), como é possível observar na
Tabela 1.
Tabela 1. Relação entre espécie, estágio de vida, tipo de
exposição e efeitos adversos provocados pelo 2,4-D.
Referência Espécie Estágio
de vida
Tipo de
exposição Efeitos adversos
Dehnert et
al., 2019 Danio rerio
Embrião
e larva
Aguda e
crônica
Neurotoxicidade;
Alterações
comportamentais
Li et al.,
2017 Danio rerio
Embrião
e larva Aguda
Cardiotoxicidade;
Estresse oxidativo
Thiel et al.,
2020 Danio rerio Adulto Aguda
Alteração
comportamental;
Estresse oxidativo
Gaaied et
al., 2020 Danio rerio
Embrião
e larva Aguda
Alteração
comportamental;
Neurotoxicidade
Gaaied et
al., 2019 Danio rerio
Embrião
e larva Aguda Estresse oxidativo
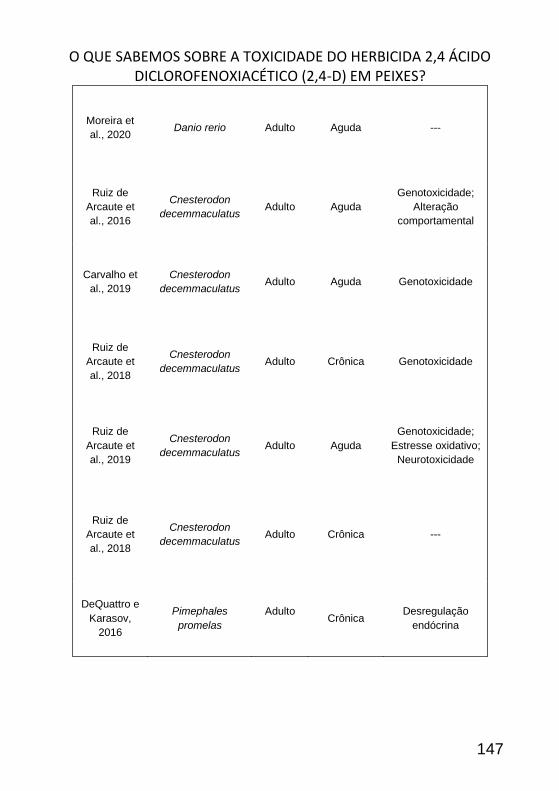
O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
147
Moreira et
al., 2020
Danio rerio
Adulto Aguda ---
Ruiz de
Arcaute et
al., 2016
Cnesterodon
decemmaculatus Adulto Aguda
Genotoxicidade;
Alteração
comportamental
Carvalho et
al., 2019
Cnesterodon
decemmaculatus Adulto Aguda Genotoxicidade
Ruiz de
Arcaute et
al., 2018
Cnesterodon
decemmaculatus Adulto Crônica Genotoxicidade
Ruiz de
Arcaute et
al., 2019
Cnesterodon
decemmaculatus Adulto Aguda
Genotoxicidade;
Estresse oxidativo;
Neurotoxicidade
Ruiz de
Arcaute et
al., 2018
Cnesterodon
decemmaculatus Adulto Crônica ---
DeQuattro e
Karasov,
2016
Pimephales
promelas
Adulto
Crônica
Desregulação
endócrina

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
148
Dehnert et
al., 2018
Pimephales
promelas
Adulto
Larva Crônica ---
Fait et al.,
2015
Oreochromis
niloticus Adulto Aguda ---
Moreira et
al., 2020
Hyphessobrycon
eques
Adulto Aguda Alteração
comportamental
Fonte: Autoral. 2020
Danio rerio
O D. rerio, popularmente conhecido como zebrafish ou
paulistinha, foi a espécie mais utilizada entre os artigos (43,75%
do total). Este é um pequeno teleósteo de 3-4 cm, pertencente
à família Cyprinidae, já estabelecido como modelo em diversas
áreas da ciência (GONÇALVES et al., 2020).
O zebrafish oferece uma gama de vantagens como
modelo animal para pesquisa. Primeiramente, a praticidade de
manejo e baixo custo de manutenção atrelados a alta taxa de
fecundidade faz desse modelo uma ferramenta promissora na
elaboração de estratégias de predição do potencial toxicante de
novos fármacos e químicos (BAMBINO et al., 2017).
Além disso, o processo de maturação do D. rerio
acontece em um período de tempo relativamente curto, no qual,
entre 48 e 72 horas evolui do estágio embrionário para o larval
e se torna sexualmente maduro por volta dos três meses de
vida, o que o torna uma ferramenta prática e viável para a

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
149
execução de trabalhos de hereditariedade, avaliação de
toxicidade crônica e triagens em larga escala (MEYERS, 2018).
Mais da metade dos artigos que utilizaram o zebrafish
como modelo fizeram uso dos seus estágios iniciais de vida
(embrionário e larval). Além de motivos como facilidade de
manejo em relação aos animais adultos, uma maior
observância em relação ao princípio dos 3Rs (replacement,
reduction and refinement – traduzidos para substituição,
redução e refinamento) pode ser a causa da preferência dos
cientistas pelo uso das fases inicias de vida. Embriões e larvas
de zebrafish possuem um desenvolvimento externo e são
translúcidos, o que permite a visualização do desenvolvimento
morfológico do animal em tempo real sem a necessidade de
equipamentos robustos e sofisticados. Com isso, em estudos
de toxicologia, é possível identificar alterações morfológicas e
comportamentais no embrião e correlacioná-las com os efeitos
deletérios da substância avaliada (GONÇALVES et al., 2020).
C. decemmaculatus
O C. decemmaculatus é um pequeno peixe ovovivíparo,
não migratório que mede entre 25 e 45 mm e está
freqüentemente presente em alta densidade em cursos d'água.
É uma espécie endêmica da Argentina e possui ampla
destribuição pela América Neotropical, especialmente no rio La
Plata e outras bacias sul-americanas (RUIZ DE ARCAUTE et
al., 2018).
Esta espécie apresenta fácil manuseio e aclimatação às
condições laboratoriais. Ela atende à condições exigidas para

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
150
testes de toxicidade, como uma grande faixa de tolerância a
diversos fatores ambientais (RUIZ DE ARCAUTE et al., 2016).
Diversos estudos utilizaram essa espécie como um
modelo in vivo para analisar a toxicidade provocada por
pesticidas em vertebrados aquáticos. Entre eles estão produtos
com finalidade pesticida, inseticida e herbicida (RUIZ DE
ARCAUTE et al., 2018) Nessa revisão, todos os estudos
encontrados utilizaram o C. decemmaculatus como modelo em
seu estágio de vida adulto.
Efeitos adversos
Na literatura existe uma extensa quantidade de trabalhos
que reportam a sensibilidade de organismos modelo a
diferentes classes de pesticidas. Além disso, é comprovada que
a exposição a esses químicos podem induzir o surgimento de
efeitos adversos, incluindo cardiotoxidiade, neurotoxicidade,
toxicidade reprodutiva, alterações comportamentais, entre
outros. Nessa revisão, foi possível observar que o 2,4-D induz
a aparição de diferentes tipos de efeitos adversos em peixes.
Também foi possível observar que fatores como idade dos
animais e tipo de exposição podem influenciar nos efeitos
induzidos por esse químico. Os dados extraídos dos artigos da
revisão são mostrados na Tabela 1.
Impactos sobre a sobrevivência e alterações morfológicas
Os dados extraídos da revisão demonstraram que o 2,4-
D (princípio ativo ou formulação comercial) foi capaz de
provocar alterações na taxa de sobrevivência de diferentes

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
151
espécies de peixes utilizadas como modelo, sendo os estágios
iniciais de vida os mais afetados.
Uma das espécies que demonstrou uma maior
sensibilidade ao herbicida foi o D. rerio. Dehnert e
colaboradores (2019), observaram que a exposição subcrônica
(21 dias) de embriões e larvas de zebrafish, tanto ao 2,4-D puro
quanto ao DAM4 (formulação comercial), foram capazes de
alterar a taxa de sobrevivência, apresentando uma CL50 entre
4 – 6 ppm e 8 – 16 ppm, respectivamente.
Já Gaaied e colaboradores (2020) e Li e colaboradores
(2017) realizaram exposições agudas (≤ 96 h) de embriões e
larvas de zebrafish ao 2,4-D puro, e encontraram diferentes
valores de CL50: 2,86 mg/L e 46,71 mg/L, respectivamente.
A grande diferença de sensibilidade entre as linhagens
utilizadas por esses dois grupos de pesquisa provavelmente
está relacionada com o alto grau de heterogeneidade dessa
espécie, inclusive em ambientes laboratoriais (PARICHY,
2015). Larvas de P. promelas submetidas a uma exposição
crônica (30 dias) também se mostraram sensíveis ao 2,4-D
puro, DMA4 e WAM40 e tiveram sua taxas de sobrevivência
diminuídas (DEHNERT et al., 2018).
Apesar de nenhum dos artigos selecionados terem
analisado o impacto do 2,4-D sobre a sobrevivência do
zebrafish adulto, dados da literatura informam que a CL50 nesse
estágio de vida é de 233,13 mg/L, valor bastante superior
àquele dos estágios embrionário e larval da espécie
(POMPERMAIER et al., 2020).
Além disso, esse valor é próximo a CL50 encontrada para
outra espécie de peixe em sua fase adulta, o O. niloticus,
quando exposto durante 96 h ao herbicida comercial Herbextra

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
152
(222,28 mg/L) (FAI et al., 2015). Já o C. decemmaculatus
adulto, apresentou uma CL50 de 1.008 mg/L em uma exposição
aguda ao DMA, se mostrando a espécie menos sensível de
acordo com os trabalhos selecionados (RUIZ DE ARCAUTE et
al., 2016).
Além do impacto sobre a sobrevivência, três dos estudos
analisados buscaram avaliar alterações morfológicas
decorrentes da exposição ao 2,4-D em embriões e larvas de
zebrafish. Dentre as deformidades identificadas, estavam:
edema de saco vitelínico, bexiga natatória sub-inflada,
curvatura na coluna vertebral e, o mais recorrente, edema
pericárdico (GAAIED et al., 2020; DEHNERT et al., 2019; LI et
al., 2017).
Diversos dados disponíveis na literatura apontam o
aparecimento de edemas de pericárdio em embriões de
zebrafish quando expostos a substâncias tóxicas, tais como
poluentes orgânicos e metais pesados. Isso sugere que essa
anomalia pode ser uma resposta geral em ambiente tóxico e
que o coração em desenvolvimento é provavelmente um alvo
para a toxicidade do 2,4-D (LI et al., 2017). O estudo realizado
por Li e colaboradores (2017) reforça essa teoria, uma vez que
avaliou a alteração de genes envolvidos com o
desenvolvimento do coração ao expor larvas de zebrafish ao
2,4-D.
Neurotoxicidade e alterações comportamentais
Alterações comportamentais foram os efeitos adversos
mais observados em decorrência da exposição dos peixes ao
2,4-D (37,5% dos artigos). Larvas de D. rerio e P. flavenscens

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
153
mostraram redução da atividade predatória quando submetidos
à uma exposição subcrônica à herbicidas comerciais a base de
2,4-D (DEHNERT et al., 2019). Esse mesmo efeito foi
observado quando adultos de D. rerio foram submetidos a uma
exposição aguda (30 min) ao 2,4-D puro (POPERMAIER et al.,
2020).
A capacidade predatória, especialmente nos estágios
iniciais de vida de peixes, é estritamente relacionada ao
desenvolvimento dos olhos, uma vez que a visão é um sentido
extremamente necessário para realizar essa atividade. Dehnert
e colaboradores (2019) observaram que larvas de zebrafish
expostas ao DMA4, apresentaram uma redução na atividade
neural no tectum óptico, uma estrutura pertencente ao
mesencéfalo de vertebrados (como peixes, aves e répteis) e
exerce uma função de extrema importância como processador
visual (DEHNERT et al., 2019).
A redução na capacidade de predação apresentada
pelos estágios larvais de peixes pode suprimir diretamente sua
sobrevivência, levar a um impacto na aptidão geral de espécies
e modificar a dinâmica dos ecossistemas em que elas habitam.
A atividade de natação anormal também foi observada
após a exposição ao 2,4-D e químicos derivados. Gaaied e
colaboradores (2020) observaram esse comportamento
anormal em larvas de D. rerio com cinco dias de vida, após
serem expostas a concentrações subletais do 2,4-D puro. Esta
alteração comportamental também foi observada em animais
adultos de outras espécies, como C. decemmaculatus e H.
eques (MOREIRA et al., 2020; RUIZ DE ARCAUTE et al., 2016).
O trabalho desenvolvido por Ruiz de Arcaute e colaboradores
(2016) evidenciou a maior diversidade de alterações

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
154
comportamentais observada. Após uma exposição aguda de
adultos de C. decemmaculatus ao DMA, eles observaram:
permanência dos animais no fundo do aquário, movimentação
lenta, diminuição da capacidade de reação, além da natação
anormal, citada anteriormente.
Geralmente, alterações comportamentais são
consequência da alteração da expressão de moléculas que
desempenham função importante sobre a atividade neural e/ou
locomotora. Uma dessas moléculas é a acetilcolinesterase,
(AChE) uma enzima encontrada principalmente no sistema
nervoso central que exerce o papel de processamento do
neurotransmissor acetilcolina (THAPA et al., 2017). Por exercer
essa função, essa enzima é um biomarcador clássico para
análise de neurotoxicidade de substâncias.
Dos artigos avaliados, dois deles mensuraram a
atividade de AChE. Larvas de D. rerio e C. decemmaculatus
adultos tiveram a atividade de AChE significativamente
diminuída após exposição ao 2,4-D (GAAIED et al., 2020; RUIZ
DE ARCAUTE et al., 2016). Somada às alterações
comportamentais observadas, a diminuição da atividade desse
biomarcador indica que um dos alvos em potencial desse
herbicida em peixes é o sistema nervoso.
Genotoxicidade
A genotoxicidade foi um dos efeitos adversos mais
observados em peixes em decorrência da exposição ao 2,4-D.
A exposição aguda de adultos de C. decemmaculatus ao DMA
foi capaz de provocar um aumento do índice de danos
cromossômicos, quebras primárias no DNA e aumento na

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
155
frequência da formação de micronúcleos (RUIZ DE ARCAUTE
et al., 2016; RUIZ DE ARCAUTE et al., 2019).
Os micronúcleos são fragmentos de cromossomos ou
cromossomos inteiros que foram perdidos durante a mitose
celular em razão de um evento aneugênico (que interfere no
fuso mitótico) ou clastogênico (que provoca quebra
cromossômica) e, por esse motivo, o teste de micronúcleo
avalia genotoxicidade (ANTÔNIO et al., 2017).
Além dos micronúcleos, foram relatados outros tipos de
anormalidades nucleares decorrentes da exposição ao 2,4-D.
Por exemplo, núcleos com bolhas e entalhes, que podem ser
consideradas evidência de instabilidade genética (RUIZ DE
ARCAUTE et al., 2016). Esses dados encontrados estão de
acordo com um estudo de avaliação de risco realizado pela
International Agency for Research on Cancer (IARC) em 2015,
que objetivou avaliar o risco da exposição a alguns agrotóxicos
organoclorados e alguns herbicidas clorofenoxi, entre eles, o
2,4-D. Como resultado, o potencial genotóxico do 2,4-D foi
fortemente abordado e confirmado por diversos ensaios in vivo
e in vitro, incluindo o potencial de induzir danos cromossômicos
e de DNA em diferentes matrizes biológicas.
Estresse oxidativo
O estresse oxidativo pode ser definido como um
desequilíbrio em radicais/espécies reativas de oxigênio (ROS)
em células ou tecidos. O estresse oxidativo pode levar à
toxicidade por meio da alteração de moléculas essenciais, tais
como proteínas, lipídios e DNA. Por exemplo, a expressão
gênica de diferentes enzimas como as enzimas do sistema de

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
156
defesa antioxidante (Glutationa s-transferase, superóxido
dismutase e catalase) funciona como um ótimo biomarcador de
efeitos tóxicos, responsáveis por estresse oxidativo e, portanto,
um parâmetro interessante em ambientes aquáticos
(GONÇALVES et al., 2020).
Nesta revisão, em 4 dos 16 estudos foi possível observar
estresse oxidativo como efeito adverso à exposição do 2,4-D
em duas espécies de peixes diferentes. Li e colaboradores
(2019) utilizaram o D. rerio como modelo para avaliar possíveis
efeitos moleculares decorrentes da exposição a diferentes
concentrações do 2,4-D. Os autores avaliaram a atividade da
superóxido dismutase (SOD), importante enzima antioxidante
para defesa contra a toxicidade de espécies reativas de
oxigênio (ROS) e o conteúdo de Malondialdeído (MDA), produto
da reação entre os radicais livres e os ácidos graxos
insaturados nas membranas celulares. O primeiro biomarcador,
não sofreu alteração, já o conteúdo de MDA foi aumentado.
Além desses biomarcadores, foi encontrada uma regulação
negativa de genes marcadores de estresse oxidativo (cat e
gpx1a).
Thiel e colaboradores (2020) também utilizaram o D.
rerio como modelo. Os autores observaram um aumento nas
atividades das enzimas citocromo c oxidase, que catalisa a
reação final da cadeia de transporte de elétrons, da catalase
(CAT) e uma redução na atividade da SOD.
Gaaied e colaboradores (2019) também utilizaram o
zebrafish como modelo, e observaram uma indução significativa
da atividade da enzima etoxiresorufina-O-desetilase (EROD) e
inibição da atividade da enzima GST, além do aumento na

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
157
atividade de CAT e Glutationa-peroxidase (GPx) e aumento no
conteúdo de MDA.
Ruiz de Arcaute e colaboradores (2019), avaliaram os
efeitos decorrentes da exposição do 2,4-D no modelo
Cnesterodon decemmaculatus. Os autores observaram o
aumento nas atividades de CAT e Glutationa S-transferase
(GST). Além disso, também existem relatos de uma regulação
negativa dos genes gpx1a e cat, responsáveis pela expressão
de enzimas importantes para o processo de desintoxicação e
combate à radicais livres (LI et al., 2019).
O aumento do conteúdo de MDA, o aumento na atividade
EROD, de CAT e de GST podem levar ao acúmulo de ROS em
embriões de zebrafish e podem induzir o estresse oxidativo ou
até mesmo comprometer as defesas antioxidantes (GAAIED et
al., 2019; LI et al., 2019).
Desta forma, alterações desses biormarcadores podem
fazer parte das vias metabólicas que levam ao estresse
oxidativo induzido pelo 2,4D.
Distúrbios endócrinos
Distúrbios endócrinos podem ser desencadeados por
substâncias que ativam ou inibem receptores de hormônios
endógenos e afetam a síntese e o transporte de hormônios,
causando desequilíbrios em seus níveis. Dessa forma,
processos como crescimento, desenvolvimento, maturação
sexual, reprodução, metabolismo energético e doenças como
diabetes podem ser diretamente impactados (GONÇALVES et
al., 2020).

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
158
Há uma limitação na literatura sobre os efeitos da
exposição de animais aquáticos ao 2,4-D e suas formulações e
o potencial que estes apresentam como desreguladores
endócrinos. No entanto, DeQuattro e Karasov (2016), avaliaram
o efeito de concentrações ambientalmente relevantes deste
composto e duas formulações herbicidas sobre reprodução,
desenvolvimento embrionário e sobrevivência de larvas de P.
promelas. Após a exposição às formulações do 2,4-D, houve
diminuição significativa na presença de tubérculos masculinos
e aumento no índice gonadossomático feminino, demonstrando
que estas formulações têm potencial para desregulação
endócrina e podem exercer algum grau de toxicidade crônica.
No entanto, mais estudos com o 2,4-D puro e suas
formulações devem ser realizados.
CONCLUSÃO
Nessa revisão, apresentamos os impactos negativos do
herbicida 2,4-D sobre espécies de peixe, abordando pontos
como o composto utilizado, tipo de exposição, espécies
estudadas e estágio de vida dos animais. Foi observado que o
2,4-D diminuiu a taxa de sobrevivência e foi capaz de provocar
a aparição de diferentes tipos de efeitos adversos, como
alterações comportamentais, neurotoxicidade e genotoxicidade
e que os estágios iniciais de vida se mostraram mais sensíveis
aos efeitos desse químico. Considerando que o 2,4-D é um
herbicida amplamente utilizado, os seus efeitos tóxicos podem
comprometer a sobrevivência de peixes e outros organismos
sensíveis, podendo gerar um desequilíbrio ecológico.
REFERÊNCIAS BIBLIOGRÁFICAS

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
159
ANTONIO, E. et al. Genotoxicidade e Enotoxicidade dos Raiox X em Crianças Submetidas à Radiografia Panorâmica. Revista Paulista de Pediatria. v.35, 2017. BAMBINO, K.; CHU, J. Zebrafish in Toxicology and Environmental Health. Current Topics in Developmental Biology. v. 124, p. 331–367, 2017. CARVALHO, F. Pesticides, environment, and food safety. Food and Energy Security. v. 6. p. 48-60, 2017. CARVALHO, W. et al. Genotoxicity of mixtures of glyphosate with 2,4-dichlorophenoxyacetic acid chemical forms towards Cnesterodon decemmaculatus (Pisces, Poeciliidae). Environmental Science and Pollution Research. v.6 , p.6515-6525, 2020. DEHNERT, G. et al. 2,4-Dichlorophenoxyacetic acid containing herbicide impairs essential visually guided behaviors of larval fish. Aquatic Toxicology. v. 209, p.1-12, 2019. DEHNERT, G. K. et al. Effects of low, subchronic exposure of 2,4-dichlorophenoxyacetic acid (2,4-D) and commercial 2,4-D formulations on early life stages of fathead minnows (Pimephales promelas). Environ. Toxicol. Chem. v.37, p.2550e2559, 2018. DEQUATTRO, Z. A.; KARASOV, W. H. Impacts of 2,4-dichlorophenoxyacetic acid aquatic herbicide formulations on reproduction and development of the fathead minnow (Pimephales promelas). Environ Toxicol Chem. v.35, p.1478-88, 2016. FAI, P. A. et al. Potential of the microbial assay for risk assessment (MARA) for assessing ecotoxicological effects of herbicides to non-target organisms. Ecotoxicology. v.24, p.1915-1922, 2015. GAAIED, S. et al. 2,4-Dichlorophenoxyacetic acid herbicide effects on zebrafish larvae: development, neurotransmission and behavior as sensitive endpoints. Environ Sci Pollut Res Int. v.27, p.3686-3696, 2020. GAAIED, S. et al. Gene expression patterns and related enzymatic activities of detoxification and oxidative stress systems in zebrafish larvae exposed to the 2,4-dichlorophenoxyacetic acid herbicide. Chemosphere. v.224, p.289-297, 2019. Gonçalves, Í.F.S. et al. Toxicity testing of pesticides in zebrafish—a systematic review on chemicals and associated toxicological endpoints. Environ Sci Pollut Res 27. p. 10185–10204, 2020. IBAMA – INSTITUTO BRASILEIRO DO MEIO AMBIENTE E DOS RECURSOS NATURAIS RENOVÁVEIS. Relatórios de comercialização de agrotóxicos. 2017. Disponível em: < http://www.ibama.gov.br/agrotoxicos/>. Acesso em: 12 Set 2018. INTERNATIONAL AGENCY FOR RESEARCH ON CÂNCER (IARC). Monographs on the Evaluation of Carcinogenic Risk of Chemicals to Man.

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
160
Some Organochlorine Insecticides and Some Chlorphenoxy Herbicides. Lyon, 2015. ISLAM, F. et al. Potential impact of the herbicide 2,4-dichlorophenoxyacetic acid on human and ecosystems. Environment international. v. 111, p. 332-351, 2017. LI, K. et al. Developmental toxicity of 2,4-dichlorophenoxyacetic acid in zebrafish embryos. Chemosphere. v. 171, 2017. MEYERS, J. Zebrafish: Development of a Vertebrate Model Organism. Current Protocols Essential Laboratory Techniques. v. 16 2018. MOREIRA, R. et al. Fipronil and 2,4-D effects on tropical fish: Could avoidance response be explained by changes in swimming behavior and neurotransmission impairments?. Chemosphere. v.263, 2020. PARICHY, D. M. Advancing biology through a deeper understanding of zebrafish ecology and evolution. Elife. v. 4, 2015. POMPERMAIER, A. et al. Waterborne agrichemicals compromise the anti-predatory behavior of zebrafish. Environmental Science and Pollution Research, v.27, p.38559-38567, 2020. RUIZ DE ARCAUTE, C. et al. Opposite effects of mixtures of commercial formulations of glyphosate with auxinic herbicides on the ten spotted live-bearer fish Cnesterodon decemmaculatus (Pisces, Poeciliidae). Environmental Pollution. v.240, p.858-866, 2018. RUIZ DE ARCAUTE, C. et al. Auxinic herbicides induce oxidative stress on Cnesterodon decemmaculatus (Pisces: Poeciliidae). Environmental Science and Pollution Research. v.26, p.20485-20498, 2019. RUIZ DE ARCAUTE, C. et al. Genotoxicity by long-term exposure to the auxinic herbicides 2,4-dichlorophenoxyacetic acid and dicamba on Cnesterodon decemmaculatus (Pisces: Poeciliidae). Environmental Pollution. v.243, p.670-678, 2018. RUIZ DE ARCAUTE, C. et al. Synergism of mixtures of dicamba and 2,4-dichlorophenoxyacetic acid herbicide formulations on the neotropical fish Cnesterodon decemmaculatus (Pisces, Poeciliidae). Environmental pollution. v. 236, p. 33-39, 2018. RUIZ DE ARCAUTE, C. et al. Toxic and genotoxic effects of the 2,4-dichlorophenoxyacetic acid (2,4-D)-based herbicide on the Neotropical fish Cnesterodon decemmaculatus. Ecotoxicol Environ Saf. v. 9, p. 128:222, 2016. SHARMA, A. et al. Worldwide pesticide usage and its impacts on ecosystem. SN Applied Sciences. v. 1, 2019. THAPA, S. et al. Acetylcholinesterase: A Primary Target for Drugs and Insecticides. Mini Rev Med Chem. v. 17, p. 1665-1676, 2017.

O QUE SABEMOS SOBRE A TOXICIDADE DO HERBICIDA 2,4 ÁCIDO DICLOROFENOXIACÉTICO (2,4-D) EM PEIXES?
161
THIEL, N. et al. Exposure to the herbicide 2,4-dichlorophenoxyacetic acid impairs mitochondrial function, oxidative status, and behavior in adult zebrafish. Environmental Science and Pollution Research. 2020. TIWARI, B. et al. Pesticides and Rice Agriculture. Cyanobacteria: Froam Basica Science to Applications. p. 303-325, 2019.

162
CIÊNCIAS BIOLÓGICAS - FISIOLOGIA

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
163
CAPÍTULO 8
PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E
MICROGLIAS E SUA RELAÇÃO COM A HIPERTENSÃO NEUROGÊNICA
Ericka Garcia LEITE 1
Leonardo Lima CARDOSO2
Joálisson da Silva CALADO3
Tatjana Souza Lima KEESEN 4
Bárbara Guimarães CSORDAS5
1Doutoranda do Programa de Pós-Graduação Multicêntrico em Ciências Fisiológicas; 2Graduando do curso de Biotecnologia, CBiotec/UFPB.3 Graduando do curso de Fármacia do
CCS-UFPB; 4Professora do Departamento de Biologia Molecular e Celular, CBiotec/UFPB; 5Orientadora/Pós Doutoranda do Programa de Pós-Graduação em Produtos Naturais e
Sintéticos Bioativos - pgPNBS - UFPB [email protected]
RESUMO: A hipertensão é considerada um dos principais problemas de saúde pública a nível mundial, apresentando alta prevalência e baixas taxas de controle. É uma condição clínica multifatorial caracterizada por elevação sustentada dos níveis pressóricos. O presente trabalho tem como objetivo fornecer uma atualização sobre a relação de células imunes com a disfunção de núcleos cerebrais e a hipertensão neurogênica. Trata-se de uma revisão de literatura de caráter exploratório e descritivo fundamentado na coleta de artigos indexados em plataformas digitais como NCBI, Elsevier e Scielo. Demonstrou-se que a Angiotensina II (Ang-II), produzida pelo sistema renina angiotensina (SRA), se liga a receptores AT1 (AT1R) que são os responsáveis pelas ações hipertensinogênicas tanto na periferia como no sistema nervoso central. A nível central, promove disfunção da barreira hematoencefálica através da

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
164
interação com AT1R, resultando em infiltração de Ang-II circulante em núcleos antes protegidos pela barreira. O sistema nervoso central possui um sistema angiotensinérgico independente e é sugerido que este sistema possa ser potencializado pelo aumento da Ang II circulante, resultando na superativação de neurônios responsáveis por determinar o fluxo simpático. Nessas regiões, células microgliais e macrófagos perivasculares residentes são ativados liberando uma variedade de substâncias pró-inflamatórias e neuroativas, incluindo citocinas, quimiocinas e espécies reativas de oxigênio (EROs), causando neuroinflamação, aumento da atividade simpática e aumento da pressão arterial. O acúmulo de ânions superóxido induzidos por Ang-II e pelas células do sistema imune são fundamentais para a patogênese da hipertensão neurogênica. Palavras-chave: Angiotensina II. Hipertensão. Macrófago perivascular. Microglia.
INTRODUÇÃO
Hipertensão arterial (HA) é uma condição clínica
multifatorial caracterizada por elevação sustentada dos níveis
pressóricos. Segundo estimativas, um terço da população
mundial será hipertensa em 2025, fato que inexoravelmente
trará sérias consequências à saúde pública global,
principalmente por danos a órgãos-alvo (ABREU et al., 2018;
RAHIMI; EMDIN; MACMAHON, 2015; WHO, 2019).
É considerada como principal fator de risco evitável para
doenças cardiovasculares (DCV) e mortes no mundo. Em 2010,
um total de 1,38 bilhão de pessoas (31,1% da população adulta
global) tinham HA, definida como pressão arterial sistólica (PA)
≥140 mmHg e/ou PA diastólica ≥90 mmHg (MILLS;

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
165
STEFANESCU; HE, 2020). As mortes relacionadas à
hipertensão ocorrem mais comumente como resultado de
doença isquêmica do coração, acidente vascular cerebral
hemorrágico e acidente vascular cerebral isquêmico, que são
estimados respectivamente em 4,9, 2,0 e 1,5 milhões de mortes
mundialmente (FOROUZANFAR et al., 2017). A prevalência da
HA está aumentando globalmente devido ao envelhecimento da
população e ao aumento da exposição a fatores de risco como:
consumo de dietas com alto teor de sódio, obesidade,
tabagismo, alcoolismo, fatores genéticos, idade, sedentarismo,
entre outos (YOUNG; DAVISSON, 2015).
Normalmente os pacientes não apresentam
sintomatologia por anos, até que um órgão vital seja afetado,
provocando diminuição da expectativa de vida entre homens e
mulheres em todo o mundo (BILAL et al., 2015).
Na maioria dos casos, cerca de 90%, a HA é de causa
indefinida, sendo classificada como hipertensão primária ou
essencial. A hipertensão essencial é atualmente entendida
como uma doença multifatorial decorrente da ação combinada
de diversos fatores genéticos (mutações), ambientais e
comportamentais. Têm seu início ainda na infância, porém,
o diagnóstico é feito apenas na idade adulta e danos
consideráveis podem já ter ocorrido no sistema arterial, coração
e em outros órgãos (GUPTA-MALHOTRA et al., 2015).
Quando a etiologia da HA é definida, classifica-se como
hipertensão secundária, tornando um importante fator para
direcionar o tratamento de acordo com a etiologia. Entre os
portadores de HA, apenas 3 a 5% são de origem secundária. A
Hipertensão renovascular é uma das causas mais comuns de

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
166
hipertensão secundária levando, muitas vezes, à hipertensão
resistente. É responsável por 1% a 5% de todos os casos de
HA na população em geral e 5,4% dos casos de hipertensão
secundária em jovens adultos. É definida como hipertensão
sistêmica que se manifesta secundariamente ao
comprometimento do suprimento sanguíneo para os rins,
geralmente devido a uma lesão oclusiva de uma ou das duas
artérias renais (HERRMANN; TEXTOR, 2019).
Muitos dos medicamentos anti-hipertensivos, incluindo
os bloqueadores do sistema renina angiotensina, nem sempre
são eficazes. Cerca de 60% dos pacientes são refratários ao
tratamento (YOUNG; DAVISSON, 2015).
Além dos processos patológicos em órgãos
cardiovasculares periféricos, o sistema nervoso central (SNC)
surgiu como o principal responsável, estando envolvido no
desenvolvimento e manutenção de diferentes formas de
hipertensão (HIROOKA, 2020; YOUNG; DAVISSON, 2015). O
sistema renina-angiotensina (SRA) desempenha um papel
crucial no controle cardiovascular, e tanto as ações da
angiotensina-II (Ang-II) sistêmica quanto o sinal gerado
centralmente através de redes neurais complexas contribuem
para o desenvolvimento da HA (YOUNG; DAVISSON, 2015). A
contribuição de células do sistema imunológico, como
microglias e macrófagos, têm sido foco de muitos estudos para
avaliar a participação dessas células na patogênese da HA
(FOULQUIER, 2020).
Desta forma, o objetivo dessa revisão é fornecer uma
breve atualização sobre os trabalhos recentes relacionados a
participação de células microgliais e macrófagos perivasculares

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
167
em núcleo hipotalâmico e bulbares envolvidos na manutenção
da pressão arterial e sua relação com a patogênese da
hipertensão neurogênica.
MATERIAIS E MÉTODOS
Este estudo trata-se de uma revisão de literatura e
apresenta caráter exploratório e descritivo, tendo como
finalidade compreender a relação do sistema imune com a
patogênese da HA. Para obtenção dos dados, utilizou-se o
levantamento bibliográfico através de artigos indexados em
plataformas digitais como NCBI, Elsevier e Scielo, através dos
descritores: “hipertensão”, “angiotensina II”, “macrófagos
perivasculares” e “microglias”. A seleção dos artigos foi
realizada por meio da leitura de títulos e resumos que se
adequavam aos critérios de inclusão, que foram: artigos que
abordassem a temática hipertensão neurogênica, disfunção de
núcleos cerebrais, macrófagos perivasculares e micróglias, nos
últimos 5 anos (2015 à 2020). Todos os estudos incluídos nas
etapas anteriores foram lidos na íntegra. Foram excluídos os
estudos que não de adequavam aos critérios de inclusão.
RESULTADOS E DISCUSSÃO
Controle da pressão arterial e hipertensão;
A pressão arterial (PA) é caracteriazada pela pressão
que o sangue exerce sobre as paredes das artérias. É
determinada pelo produto do débito cardíaco pela resistência

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
168
vascular periférica total. O débito cardíaco consiste na
quantidade de sangue que cada ventrículo ejeta na circulação
(pulmonar ou sistêmica) por minuto, sendo expresso como
produto do volume sistólico (em mL) pela frequência cardíaca
(bpm). A resistência vascular periférica é caracterizada pela
pressão que o sangue exerce na parede dos vasos (LEÃO;
SILVA, 2018).
O controle da PA é realizado por mecanismos diversos,
através de sistemas de retroalimentação que operam a curto e
longo prazo. A curto prazo, a regulação é desempenhada por
reflexos originados de pressorreceptores como
barorreceptores, quimiorreceptores e receptores
cardiopulmonares, resultando em uma resposta rápida obtida
através de um sistema de retroalimentação pelo sistema
nervoso autônomo. A longo prazo, a regulação é
desempenhada através de mecanismos renais e humorais
(JOHNSON; XUE, 2018).
Como um regulador a longo prazo, o SRA periférico e
central desempenha importantes função na manutenção da PA
através de uma diversidade de processos hemodinâmicos
renais e de transporte, contribuindo para o balanço
hidroeletrolítico do sangue (SILVA et al., 2017).
Quando ocorre uma redução da PA, as células
justaglomerulares localizadas nas artérias aferentes renais são
estimuladas a secretar renina. A secreção de renina pelos rins
é estimulada por três vias principais: 1) barorreceptores renais
que detectam diminuição da perfusão renal, 2) baixos níveis de
cloreto de sódio detectados pela mácula densa e 3) estimulação
beta-adrenérgica (SERIKI A, 2018).

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
169
A renina é uma protease que promove a clivagem do
angiotensinogênio (sintetizado no fígado) em angiotensina I
(Ang-I). A Ang-I sofre a ação da enzima conversora de
angiotensina (ECA) e é convertida em Ang-II, principal peptídeo
formado pelo SRA. A Ang-II tem um papel importante no
reestabelecimento da pressão arterial por meio de uma
variedade de mecanismos, incluindo vasoconstrição,
remodelamento cardíaco e vascular, ações diretas e indiretas
para aumentar a reabsorção renal de sódio e, principalmente, a
ativação do sistema nervoso simpático. Esses efeitos são
mediados por sua interação com receptores responsivos a Ang-
II. Embora existam duas isoformas principais de receptores
(AT1R e AT2R) o AT1R é o responsável pelas ações
hipertensinogênicas do SRA tanto na periferia como no sistema
SNC (ROSSI et al., 2019; SERIKI A, 2018).
Por ser uma molécula grande, a Ang-II circulante não
consegue atravessar a barreira hematoencefálica (BHE),
porém, consegue atuar em regiões do SNC conhecidas como
órgão circunventriculares (OCVs), que apresentam maior
permeabilidade da BHE com altas densidades de AT1R. Os
OCVs estão envolvidos em importantes mecanismos de
controle hidroeletrolítico e da função cardiovascular (BAINS;
FERGUSON, 1995; GUYENET, 2017). As ações da Ang-II
ocorrem majoritariamente no órgão subfornical (SFO) pela
sensibilidade dos neurônios dessa região a Ang-II exceder a
dos outros OCVs (BAINS; FERGUSON, 1995).
Ao se ligar aos AT1R no SFO, a Ang-II ativa projeções
angiotensinérgicas enviando sinais aos neurônios do núcleo
paraventricular (PVN), no hipotálamo. Neurônios pré-

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
170
simpáticos parvocelulares no PVN projetam-se para a medula
intermédiolateral (MIL) onde estão localizados os neurônios
motores pré-ganglionares simpáticos, influenciando
diretamente a atividade simpática (SIGMUND; GROBE, 2020).
Alguns neurônios parvocelulares do PVN também se
projetam para regiões do núcleo do trato solitário (NTS) e/ou
medula rostral ventrolateral (RVLM) responsáveis por
determinar o fluxo simpático central basal e integrar as entradas
dos barorreceptores, quimiorreceptores e receptores viscerais
através NTS. Além disso, cabe ressaltar que a ativação dos
neurônios do SFO pela Ang-II circulante, desencadeia a
produção central de Ang-II no eixo SFO-PVN-RVLM (BAINS;
FERGUSON, 1995; CRUZ et al., 2015; MISELIS, 1981).
A ligação da Ang-II ao AT1R, estimula a fosfolipase C,
que uma vez ativada, induz a formação de trifosfato de inositol,
assim como de diacilglicerol, o que provoca a liberação e
aumento da concentração citoplasmática de Ca2+, levando à
ativação da proteína quinase C (PKC). A ativação da isoforma
β da PKC fosforila domínios autoinibitórios da subunidade
regulatória da NADPH oxidase, (CRUZ et al., 2015; SERIKI A,
2018). A ativação da NADPH pela Ang-II no PVN e RVLM
produz ânion superóxido (O2●-) que reage rapidamente com o
óxido nítrico (NO), formando peroxinitrito, diminuindo a
biodisponibilidade de NO nessas regiões, levando a um
aumento no disparo neuronal (GUIMARÃES et al., 2019).
Apesar do SNC possuir um sistema angiotensinérgico
independente, é sugerido que este sistema possa ser
potencializado pelo aumento da Ang-II circulante, resultando na
superativação dos neurônios do SFO, PVN, NTS e RVLM. Um

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
171
grande conjunto de evidências sugerem um desbalanço na
sinalização de Ang-II e consequente aumento de EROs e
diminuição da biodisponibilidade de NO (BIANCARDI et al.,
2016; DE KLOET et al., 2015).
Estudos demonstraram um aumento na expressão de
AT1R e aumento na expressão de subunidades da NADPH
oxidase em áreas chaves envolvidas com a modulação
cardiovascular, como PVN e RVLM (BIANCARDI; STERN,
2016).
Posteriormente, foi demonstrado que a Ang-II promove a
disfunção da BHE através da interação com AT1R, resultando
em infiltração da Ang-II circulante no parênquima neural do PVN
e RVLM, podendo contribuir diretamente para o fluxo simpático
e a elevação da PA (BIANCARDI; STERN, 2016).
Outro estudo observou que Ang-II é capaz de ativar
células residentes do sistema imune no SNC como microgliais
e macrófagos perivasculares, responsáveis liberação de uma
variedade de substâncias ocasionando uma neuroinflamação
(BIANCARDI et al., 2016; FOULQUIER, 2020).
Relação da microglia com o desenvolvimento da
hipertensão neurogênica;
Microglias são células imunes residentes no cérebro que
funcionam em estreita associação a BHE (SHEN et al., 2015).
São um tipo celular, pertencente à neuroglia, que se diferem
das outras células gliais pela sua origem hematopoiética e pela
sua função de combater possíveis patógenos. Compreendem a
aproximadamente 10-15% de todas as células que residem no

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
172
SNC. Em um cérebro hígido as microglias geralmente se
apresentam na sua forma ramificada, ou inativa, e realizam
contínuo monitoramento do SNC. Após estímulo, essas células
sofrem uma série de alterações morfológicas e fenotípicas,
liberando mediadores que podem modular diretamente a
atividade neuronal (BIANCARDI et al., 2016; SHEN et al.,
2015).
As microglias são divididas morfológicamente e
funcionalmente em 3 fenótipos: M0, M1 e M2. Quando células
do tipo M0 (estado de repouso) são estimuladas, ocorrem
diferenciações morfológicas nas células, principalmente na
expressão de diferentes receptores. Essas modificações irão
promover reparos teciduais através da microglia anti-
inflamatória M2, com liberação de IL-10, ou efeito degenerativo,
onde ocorre aumento da citotoxicidade, causada pela transição
do estado M2 para o M1 pró-inflamatório, havendo liberação de
citocinas como: IL-6, TNF-α e IL-1β. Assim, a ativação
microglial desempenha uma atividade bifásica no SNC
(KAPOOR et al., 2016; KOIZUMI et al., 2019b).
Estudos recentes abordam sobre a relação de células
microgliais e seus processos inflamatórios com o
desenvolvimento da hipertensão neurogênica. Um dos
mecanismos atualmente estudados é a via de ativação
microglial induzida por Ang-II em AT1R (BIANCARDI et al.,
2016; FOULQUIER, 2020).
A ativação microglial, com consequente neuroinflamação
e aumento dos níveis de EROs no RVLM, pode ser um fator que
está associado ao desenvolvimento da hipertensão
neurogênica (KAPOOR et al., 2016). Além disso, sabe-se

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
173
também que a HA já desenvolvida pode induzir a ativação de
células microgliais, em regiões encefálicas que atuam como
reguladores da pressão cardiovascular, tais como PVN e NTS
(BHAT et al., 2017).
Biancardi e colaboradores (2016), ao observar em
modelo animal a ativação de células microgliais mediada por
Ang-II, foi possível afirmar que receptores do tipo TLR4,
expressos em células da microglia, são alvos moleculares que
podem associar a ativação microglial mediada por Ang-II ao
estresse oxidativo no PVN. A ativação dessas células via AT1R
é outro fator que induz a produção de EROs no PVN. Em
contrapartida, a inibição de AT1R pode levar a um processo anti-
inflamatório, indicando que antagonistas dos AT1R, além de
reduzirem a pressão arterial, podem ser benéficos no
melhoramento funcional dos vasos cerebrais (FOULQUIER,
2020).
Uma importante via pela qual a inflamação pode levar a
consequências neuronais é por meio da comunicação através
da BHE entre as moléculas inflamatórias e elementos neuronais
envolvidos na manutenção dos circuitos neurais (HARUWAKA
et al., 2019).
A BHE é um sincício fortemente regulado de células
endoteliais com baixas propriedades de transporte transcelular
e paracelular que circundam os vasos cerebrais e protegem o
delicado microambiente neuronal de substâncias neurotóxicas
(HARUWAKA et al., 2019). Uma variedade de fatores
patológicos são capazes de causar danos a BHE, incluindo
estresse oxidativo, neuroinflamação, células imunes e vários
patógenos. Esses fatores patológicos reduzem as junções de

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
174
oclusão entre as células endoteliais cerebrovasculares e
degradam as membranas basais, tudo culminando em um
aumento na permeabilidade BBB, abrindo caminho para que
moléculas grandes e substâncias prejudiciais atinjam o tecido
cerebral causando danos (SONG et al., 2020).
A inibição de células microgliais com Minociclina, é capaz
de atenuar a hipertensão em modelos de hipertensão
experimental, diminuir a atividade simpática, inflamação e
reduzir a disfunção da BHE, demonstrando a relação dessas
células com hipertensão neurogênica (HARUWAKA et al., 2019;
SANTISTEBAN et al., 2015).
Outro estudo avaliou os efeitos anti-inflamatórios do
heptapeptídeo angiotensina 1-7 [Ang-(1-7)] em células
microgliais que apresentam receptores Mas. A aplicação de
Ang-(1-7) em cultura levou a uma significativa diminuição na
transcrição de mRNA para IL-1β e TNF-α. Além disso, houve
aumento na produção da citocina imunorreguladora IL-10 (LIU;
SHI; SUMNERS, 2016). Os dados obtidos são de grande valia
para posteriores estudos da ação da ANG-(1-7) em células da
microglia e como esse peptídeo pode ser utilizado na regulação
de processos neuroinflamatórios, diminuindo efeitos
neurodegenerativos atribuídos a superativação do estado M1.
Além das investigações de células microgliais no
desenvolvimento da hipertensão, também já foram reportados
estudos abordando o potencial dessas células no surgimento
de danos cerebrovasculares (KOIZUMI et al., 2019b; ZARRUK;
GREENHALGH; DAVID, 2018).
Um estudo realizado por pesquisadores da Universidade
de Medicina de Kyoto (Japão), buscou esclarecer o processo

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
175
de danificação cerebrovascular causado por microglia pró-
inflamatória M1 em um modelo animal de hipertensão crônica.
Os ratos utilizados no processo foram submetidos a injeção de
desoxicorticosterona (DOCA) e ingestão de NaCl 1%. Os
animais foram avaliados após 2, 3 e 4 semanas de tratamento,
através de análises imuno-histoquímicas e bioquímicas.
Durante o período foi observado o aumento significativo de
células microgliais ao redor dos vasos sanguíneos e expressão
de um estado transitório caracterizado pelo marcador de
proliferação celular Ki-67, com transformação direta para o
estado M1 (pró-inflamatório), não sendo observado o estado M2
(anti-inflamatório) durante o experimento (KOIZUMI et al.,
2019b).
Porém, não foi observado no estudo se ocorreu
destruição cerebrovascular. Ademais, foram identificadas
morfologias anormais na vasculatura cerebral, ocorrendo
aumento do espaço perivascular após 3 semanas de tratamento
com DOCA-NaCl e encolhimento dos prolongamentos
citoplasmáticos de astrócitos após 4 semanas, tendo aparente
aumento da permeabilidade da BHE. A conclusões deste
trabalho apontam para uma correlação entre a ativação
microglial perivascular induzida por hipertensão crônica, com o
surgimento posterior de doenças cerebrovasculares (KOIZUMI
et al., 2019b).
Deste modo, torna-se visível a importância da microglia
na manutenção dos processos fisiológicos no SNC, sendo
válida a sua associação com o desenvolvimento da hipertensão
neurogênica através de modificações perivasculares, induzidas
por estados neuroinflamatórios e pró-oxidativos

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
176
Relação dos Macrófagos Perivasculares - PVMs com o
desenvolvimento da hipertensão neurogênica;
Os macrófagos estão presentes em todos os órgãos
desempenhando funções distintas dependendo de sua
localização e fenotipo. Durante o desenvolvimento vascular
normal, os macrófagos interagem diretamente e modulam a
vasculatura em desenvolvimento. Em alguns órgãos, como no
cérebro, esses macrófagos podem ser distinguidos de outros
macrófagos de tecido com base em sua proximidade aos vasos
sanguíneos e na expressão de certos marcadores (LAPENNA;
DE PALMA; LEWIS, 2018).
Residindo no espaço perivascular intracerebral não
parenquimatoso, são denominados de macrófagos
perivasculares (PVMs), sendo delimitadas pelas células da glia
limitantes e a membrana basal vascular (espaço de Virchow-
Robin) (FARACO et al., 2016). Diferem das microglias
associadas a vasos, que são justapostos a todos os vasos
cerebrais, mas localizados além da glia limitante. Persistem ao
longo da vida do organismo devido à sua longevidade e
capacidade de autoproliferação, ao invés da infiltração de
células mieloides periféricas (KOIZUMI et al., 2019a).
Consistente com seu envolvimento na permeabilidade
vascular em vasos sistêmicos os PVMs também desempenham
um papel no movimento de solutos, agentes infecciosos e
células imunológicas do sangue para o cérebro.Por exemplo,
na área postrema, um OCV desprovido de BHE, os PVMs
restringe a entrada de moléculas maiores que 10 kDa (FARACO
et al., 2016).

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
177
Estudos experimentais e clínicos tem demonstrado a
contribuição do sistema imunológico para a fisiopatologia da
hipertensão, evidenciando os PVMs, os quais estão envolvidas
na inflamação com relação a natriurese de pressão,
relaxamento vascular disfuncional e hiperatividade do sistema
nervoso simpático (RODRIGUEZ-ITURBE; PONS; JOHNSON,
2017).
Em um estudo com camundongos BPH/2J (modelo
neurogênico de hipertensão) foi observado que os PVMs
desempenham um papel fundamental na disfunção
cerebrovascular. Nesse modelo de estudo o aumento na
permeabilidade da BHE permitiu que a Ang-II penetrasse no
espaço perivascular tornando os PVMs alvos da Ang-II. Os
PVMs expressam AT1Rs, desta forma, a Ang-II foi capaz de se
ligar e ativar essas células, levando à produção de EROs
resultando em disfunção neurovascular e cognitiva (FARACO
et al., 2016).
Em resposta às citocinas circulantes, os PVMs cerebrais
produzem prostaglandina E2 (PGE2) por meio da enzima
biossintética ciclooxigenase-2 (COX-2). A PGE2 pode penetrar
no parênquima cerebral e ativar núcleos que expressam os
receptores de PGE2 como o PVN e RVLM, levando à uma
simpatoexcitação e elevação da pressão arterial por meio da
ativação neuronal nesses núcleos (REZQ; ABDEL-RAHMAN,
2016).
Além de secretar fatores pró-inflamatórios diretamente e
atuar como células pró-inflamatórias, os PVMs também podem
facilitar a neuroinflamação indiretamente por permeabilizar os
leucócitos circulantes que passam pela BHE

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
178
Em outro estudo, com o intuito de avaliar a contribuição
dos PVMs cerebrais para a elevação da pressão arterial, os
pesquisadores realizaram um tratamento com clodronato
encapsulado em lipossoma (que elimina macrófagos por
indução de apoptose), de forma intracerebroventricular em
modelo animal espontaneamente hipertensos propensos a
derrame (SHRSP). Foi observado que o tratamento reduziu a
atividade neuronal em núcleos reguladores simpáticos, como
PVN e RVLM e reduziu a expressão de COX-2 e PGE-E2,
demonstrando que os PVMs cerebrais contribuem para o
desenvolvimento de hipertensão por meio da ativação simpática
(IYONAGA et al., 2020).
Embora os PVMs exerçam efeitos protetores que
contribuem para a homeostasia, a ativação repetida dessas
células pela Ang-II, fazem com que sejam um componente
celular de importância para a patogênese da hipertensão
neurogênica.
5. CONCLUSÕES
A superativação do SRA e o aumento na permeabilidade
da BHE permitem que a ANG-II circulante tenha acesso a
neurônios, microglias e macrófagos perivasculares em regiões
cerebrais antes protegidas por essa barreira, como é o caso dos
neurônios pré-simpáticos do PVN e RVLM. A ativação de
células microgliais e macrófagos perivasculares pela Ang-II
induz ao acúmulo de ânions superóxido e uma processo
inflamatório aumentando a atividade simpática e
consquentemente, elevando a PA.

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
179
O estresse oxidativo é um mecanismo chave na
hipertensão neurogênica dependente de Ang-II. Desta forma,
este trabalho destaca a importância de obter informações sobre
os papéis desempenhados pelas microglias e macrófagos e sua
interação com a vasculatura do SNC, posicionando-os como
atores-chave no desenvolvimento da hipertensão e suas
consequências subseqüentes.
REFERÊNCIAS BIBLIOGRÁFICAS ABREU, D. et al. Cardiovascular disease and high blood pressure trend analyses from 2002 to 2016: After the implementation of a salt reduction strategy. BMC Public Health, v. 18, n. 1, p. 722, 11 jun. 2018. BAINS, J. S.; FERGUSON, A. V. Paraventricular nucleus neurons projecting to the spinal cord receive excitatory input from the subfornical organ. American Journal of Physiology - Regulatory Integrative and Comparative Physiology, v. 268, n. 3 37-3, 1995. BHAT, S. A. et al. Platelet CD40L induces activation of astrocytes and microglia in hypertension. Brain, Behavior, and Immunity, v. 59, p. 173–189, 1 jan. 2017. BIANCARDI, V. C. et al. Cross talk between AT1 receptors and Toll-like receptor 4 in microglia contributes to angiotensin II-derived ROS production in the hypothalamic paraventricular nucleus. American journal of physiology. Heart and circulatory physiology, v. 310, n. 3, p. H404–H415, 2016. BIANCARDI, V. C.; STERN, J. E. Compromised blood-brain barrier permeability: Novel mechanism by which circulating angiotensin II signals to sympathoexcitatory centres during hypertension. Journal of Physiology, v. 594, n. 6, p. 1591–1600, 15 mar. 2016. BILAL, M. et al. Knowledge, Awareness and Self-Care Practices of Hypertension Among Cardiac Hypertensive Patients. Global Journal of Health Science, v. 8, n. 2, p. 9–19, 1 jun. 2015. CRUZ, J. C. et al. Reactive oxygen species in the paraventricular nucleus of the hypothalamus alter sympathetic activity during metabolic syndrome. Frontiers in Physiology, v.6, p. 384, 2015. DE KLOET, A. D. et al. Role of neurons and glia in the CNS actions of the renin-angiotensin system in cardiovascular control. American Journal of

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
180
Physiology - Regulatory Integrative and Comparative, v. 309, n.5, 2015. FARACO, G. et al. Perivascular macrophages mediate the neurovascular and cognitive dysfunction associated with hypertension. Journal of Clinical Investigation, v. 126, n. 12, p. 4674–4689, 2016. FOROUZANFAR, M. H. et al. Global burden of hypertension and systolic blood pressure of at least 110 to 115mmHg, 1990-2015. JAMA - Journal of the American Medical Association, v. 317, n. 2, p. 165–182, 10 jan. 2017. FOULQUIER, S. Brain perivascular macrophages: connecting inflammation to autonomic activity in hypertension. Hypertension Research, v.43, n.2, p. 148, 2020 GUIMARÃES, D. D. et al. Dietary Nitrate Reduces Blood Pressure in Rats With Angiotensin II–Induced Hypertension via Mechanisms That Involve Reduction of Sympathetic Hyperactivity. Hypertension, v. 73, n. 4, p. 839–848, abr. 2019. GUPTA-MALHOTRA, M. et al. Essential hypertension vs. secondary hypertension among children. American Journal of Hypertension, v. 28, n. 1, p. 73–80, 2015. GUYENET, P. G. Putative Mechanism of Salt-Dependent Neurogenic HypertensionHypertensionLippincott Williams and Wilkins, , jan. 2017. HARUWAKA, K. et al. Dual microglia effects on blood brain barrier permeability induced by systemic inflammation. Nature Communications, v. 10, n. 1, p. 1–17, 1 dez. 2019. HERRMANN, S. M.; TEXTOR, S. C. Renovascular Hypertension. Endocrinology and Metabolism Clinics of North America, v.48, n.4, p. 765-778, 2019. HIROOKA, Y. Sympathetic Activation in Hypertension: Importance of the Central Nervous System. American Journal of Hypertension, 6 maio 2020. IYONAGA, T. et al. Brain perivascular macrophages contribute to the development of hypertension in stroke-prone spontaneously hypertensive rats via sympathetic activation. Hypertension Research, v. 43, n. 2, p. 99–110, 1 fev. 2020. JOHNSON, A. K.; XUE, B. Central nervous system neuroplasticity and the sensitization of hypertension. Nature Reviews Nephrology, v.14, n.12, p. 750-766, 2018. KAPOOR, K. et al. Alerted microglia and the sympathetic nervous system: A novel form of microglia in the development of hypertension. Respiratory Physiology and Neurobiology, v. 226, p. 51–62, 1 jun. 2016. KOIZUMI, T. et al. Vessel-Associated Immune Cells in Cerebrovascular Diseases: From Perivascular Macrophages to Vessel-Associated Microglia. Frontiers in Neuroscience, v. 13, n. December, p. 1–10, 2019a. KOIZUMI, T. et al. Transiently proliferating perivascular microglia harbor M1

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
181
type and precede cerebrovascular changes in a chronic hypertension model. Journal of Neuroinflammation, v. 16, n. 1, p. 79, 10 abr. 2019b. LAPENNA, A.; DE PALMA, M.; LEWIS, C. E. Perivascular macrophages in health and disease. Nature Reviews Immunology, v.18, p. 689-702, 2018. LEÃO, R. N.; SILVA, P. M. DA. Impedance Cardiography in the Evaluation of Patients with Arterial Hypertension. International Journal of Cardiovascular Sciences, v. 32, n. 1, p. 61–69, 2018. LIU, M.; SHI, P.; SUMNERS, C. Direct anti-inflammatory effects of angiotensin-(1-7) on microglia. Journal of Neurochemistry, v. 136, n. 1, p. 163–171, 1 jan. 2016. MALACHIAS, M. et al. 7a DIRETRIZ BRASILEIRA DE HIPERTENSÃO ARTERIAL. arquivos brasileiros de cardiologia, v. 107, n. 3(suppl. 3), p. 1–77, 2016. MILLS, K. T.; STEFANESCU, A.; HE, J. The global epidemiology of hypertension. Nature Reviews Nephrology, v.16, n.4, p. 223-237, 2020. MISELIS, R. R. The efferent projections of the subfornical organ of the rat: A circumventricular organ within a neural network subserving water balance. Brain Research, v. 230, n. 1–2, p. 1–23, dez. 1981. RAHIMI, K.; EMDIN, C. A.; MACMAHON, S. The Epidemiology of Blood Pressure and Its Worldwide Management. Circulation Research, v.116, n.6, p. 925-935, 2015. REZQ, S.; ABDEL-RAHMAN, A. A. Rostral ventrolateral medulla EP3 receptor mediates the sympathoexcitatory and pressor effects of prostaglandin E2 in conscious rats. Journal of Pharmacology and Experimental Therapeutics, v. 359, n. 2, p. 290–299, 1 nov. 2016. RODRIGUEZ-ITURBE, B.; PONS, H.; JOHNSON, R. J. Role of the immune system in hypertension. Physiological Reviews, v. 97, n. 3, p. 1127–1164, 1 jul. 2017. ROSSI, N. F. et al. AT 1 receptors in the subfornical organ modulate arterial pressure and the baroreflex in two-kidney, one-clip hypertensive rats. American Journal of Physiology - Regulatory Integrative and Comparative Physiology, v. 316, n. 2, p. R172–R185, 1 fev. 2019. SANTISTEBAN, M. M. et al. Involvement of bone marrow cells and neuroinflammation in hypertension. Circulation Research, v. 117, n. 2, p. 178–191, 3 jul. 2015. SERIKI A, S. Role of the Kidneys in the Regulation of Intra-and Extra-Renal Blood Pressure. Annals of Clinical Hypertension, p. 048–058, 17 jul. 2018. SHEN, X. Z. et al. Microglia Participate in Neurogenic Regulation of Hypertension. Hypertension, v. 66, n. 2, p. 309–316, ago. 2015. SIGMUND, C. D.; GROBE, J. L. A colorful view of the brain renin–angiotensin system. Hypertension Research, v.43, n.4, p,357-359, 2020.

PROCESSO INFLAMATÓRIO INDUZIDO POR MACRÓFAGOS PERIVASCULARES E MICROGLIAS E SUA RELAÇÃO COM A
HIPERTENSÃO NEUROGÊNICA
182
SILVA, A. C. S. et al. Renin angiotensin system in liver diseases: Friend or foe? World Journal of Gastroenterology, v.23, n.19, p. 3396-3406, 2017. SONG, K. et al. Oxidative Stress-Mediated Blood-Brain Barrier (BBB) Disruption in Neurological Diseases. Oxidative Medicine and Cellular Longevity, 2020. WHO. Hypertension. Disponível em: <https://www.who.int/news-room/fact-sheets/detail/hypertension>. Acesso em: 9 out. 2020. WU, K. L. H.; CHAN, S. H. H.; CHAN, J. Y. H. Neuroinflammation and oxidative stress in rostral ventrolateral medulla contribute to neurogenic hypertension induced by systemic inflammation. Journal of Neuroinflammation, v. 9, 7 set. 2012. YOUNG, C. N.; DAVISSON, R. L. Angiotensin-II, the brain, and hypertension: An update. Hypertension, v. 66, n.5, p. 920-926. ZARRUK, J. G.; GREENHALGH, A. D.; DAVID, S. Microglia and macrophages differ in their inflammatory profile after permanent brain ischemia. Experimental Neurology, v. 301, n. Pt B, p. 120–132, 1 mar. 2018.
AGRADECIMENTOS
Agradecemos à Coordenação de Aperfeiçoamento de
Pessoal de Nível Superior- CAPES pelo suporte financeiro, a Universidade Federal da Paraíba pela infraestrutura necessária para a realização deste trabalho, ao Centro de Biotecnologia (Cbiotec) e ao laboratório de imunologia de doenças infecciosas (LABIDIC).
.

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
183
CAPÍTULO 9
PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS
EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
Francisco de Assis Silveira de OLIVEIRA 1
João Batista de OLIVEIRA 2
Dayanne Tomaz Casimiro da SILVA 3 1 Mestrando do Programa de Pós-graduação em Ciências Farmacêuticas, UFPE; 2 Graduando
do curso de Farmácia, UFPB; 3 Orientadora/Professora da faculdade Rebouças de Campina Grande.
RESUMO: A Lesão Pulmonar Aguda (LPA) é uma condição presente no início da Síndrome do Desconforto Respiratório Agudo (SDRA), uma doença de condição inflamatória, que danifica a microestrutura pulmonar e permite com que esse fique cheio de fluidos, dificultando diretamente a respiração do indivíduo. Mesmo com a alta taxa de acometidos, o tratamento para LPA/SDRA ainda se mostra limitado. Logo, o objetivo desse trabalho foi realizar uma revisão exploratória dos aspectos fisiopatológicos e imunológicos da LPA, abordando também o tratamento empregado para seu manejo. Para tanto, foram utilizadas as bases de dados Pubmed, Sciente direct e Web of Science. Os resultados apresentados nesse trabalho, mostram os desafios iniciais para o desenvolvimento de diagnóstico e descrição da LPA. Em adição, aborda as manifestações fisiopatológicas da doença, e os principais alvos imunológicos envolvidos no agravamento da mesma. O tratamento farmacológico empregado também foi descrito, observando as principais problemáticas para sua utilização efetiva na prática clínica. Além disso, buscou-se elencar os

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
184
principais modelos experimentais murinos utilizados para mimetizar a manifestação da doença em humanos, a fim de entender melhor sua condição, bem como identificar moléculas alvos para o tratamento da LPA. Diante desses achados, é possível observar que a LPA/SRDA é uma condição patológica complexa, com inúmeros alvos e problemáticas terapêuticas. Palavras-chave: SDRA. Modelos murinos. Fisiopatologia. Inflamação pulmonar aguda. Revisão bibliográfica.
INTRODUÇÃO
Inúmeras são as doenças que podem acometer o trato
respiratório e, dentre elas, muitas possuem um tratamento que
desfavorecem a qualidade de vida dos acometidos, levando a
uma taxa de mortalidade de 7,5 milhões de indivíduos por ano,
em decorrência de complicações (SORIANO et al., 2017).
Dentre essas doenças, podemos citar a Lesão Pulmonar Aguda
(LPA), que é uma condição presente no início da Síndrome do
Desconforto Respiratório Agudo (SDRA), uma doença de
condição inflamatória, que danifica a microestrutura pulmonar,
permitindo aumento de fluidos nesse tecido, o que dificulta
diretamente a respiração do indivíduo (HARRINGTON et al.,
2018).
A LPA entrou em maior evidência em 2020, dada a sua
manifestação na COVID-19, que entrou em situação
pandêmica, caracterizada principalmente por danos no sistema
respiratório e cardiovascular (GALLELLI et al., 2020). Porém, a
LPA/SDRA é uma condição heterogênea no que tange a sua
variabilidade etiológica, onde pode ser desencadeada por
diversos fatores pré-definidos, como: doenças vasculares de
colágeno, sepse, drogas e álcool, choque, hemorragia

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
185
pulmonar, vasculite por irradiação, pneumonia, e, mais
recentemente, as transfusões sanguíneas e o uso de cigarros
eletrônicos (VLAAR et al., 2019; SCHUPP; PRASSE;
ERYTHROPEL, 2020). Vale salientar, entretanto, que a LPA já
foi encontrada em pacientes sem fatores pré-definidos, o que
dificulta o direcionamento de uma via terapêutica para o
tratamento das manifestações existentes (HARRINGTON et al.,
2018).
A taxa de mortalidade varia de 29% a 49,5% em
decorrência do agravamento da LPA/SDRA (CASER et al.,
2014; MOWERY; TERZIAN; NELSON, 2020). Estudos
mostraram que, em Berlin, a pneumonia de origem diversa foi
responsável por 79,4% das lesões diretas e sepse por 75,4%
das lesões indiretas na LPA/SDRA (FUJISHIMA et al., 2019),
colocando a investigação inflamatória por origem bacteriana,
um fator importante para o sucesso terapêutico de uma ampla
gama de indivíduos. No Brasil, estudos prospectivos mostram
que a incidência nacional é em torno de 6,3/100.000 hab/ano
para SDRA e cerca de 3,8/100.000 hab/ano para LPA, onde a
mortalidade para esses pacientes foi de 38,5%, em torno de 28
dias, e a mortalidade intra-hospitalar foi de 49,2%, não diferindo
entre os pacientes com LPA sem SDRA e SDRA (CASER et al.,
2014).
Mesmo com a alta taxa de acometidos, o tratamento
para LPA/SDRA ainda se mostra limitado, sendo utilizado
primariamente a ventilação mecânica, acompanhada do uso de
medicamentos como corticosteroides e broncodilatadores.
Contudo a utilização desses medicamentos pode levar a uma
série de efeitos indesejáveis que não podem ser tolerados em
pacientes de alto risco. Logo, terapias inovadoras são

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
186
investigadas a fim de garantir um melhor prognóstico aos
acometidos (MOKRA et al., 2019).
Diante disso, o objetivo desse trabalho foi realizar uma
revisão exploratória dos aspectos fisiopatológicos e
imunológicos da LPA, abordando, também, o tratamento
empregado para seu manejo. Além disso, buscou-se elencar
modelos experimentais que foram desenvolvidos, para
compreender melhor a manifestação da doença, bem como
investigar o potencial terapêuticos de novas moléculas, a fim
de garantir um melhor tratamento com o surgimento de menos
efeitos colaterais.
MATERIAIS E MÉTODO
Trata-se de uma revisão bibliográfica exploratória não
exaustiva. Para tanto, foram utilizadas as bases de dados
Pubmed, Sciente direct e Web of Science, utilizando os
seguintes descritores: ALI; acute lung injury; pathophysiology;
cytokines; therapy. Foram considerados artigos escritos em
língua portuguesa ou inglesa, publicados de 2016 à 2020.
Foram excluídos artigos que não tratavam de LPA; que não
estavam disponíveis para leitura na íntegra; que não estavam
em português ou inglês. Foram considerados artigos clássicos,
com alto impacto de citações e importantes para embasamento
do tema, publicados antes de 2016.

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
187
RESULTADOS E DISCUSSÃO
• Resgate histórico
A Acute Respiratory Distress in Adults foi um termo
cunhado em 1967 para classificar cerca de 272 pacientes com
uma forma particular de insuficiência respiratória, onde ambos
estavam sendo tratados com ventilação mecânica, e 12 desses
pacientes manifestaram sinais de dispneia grave, taquipneia,
cianose, baixa complacência pulmonar, como também
apresentaram infiltrados difusos no exame radiológico do tórax.
Entre esses 12 pacientes, sete foram à óbito e a análise
microscópica mostrou que ao menos cinco morreram em
decorrência do curso da doença, mostrando condições
histológicas de microatelectasias, congestão vascular,
hemorragia, edema pulmonar e formação de membranas
hialinas que revestem a superfície alveolar (BERNARD et al.,
1994).
A classificação dessa doença não foi nada simples e
diversas frentes de pesquisadores debateram sobre o melhor
termo para nomenclatura de tal enfermidade. Foi em 1992,
quase vinte e cinco anos após a sua primeira classificação, na
Espanha, em um evento chamado American-European
Consensus Conference on Acute Respiratory Distress
Syndrome (AECC) que o termo e os critérios para diagnóstico
clínico foram, então, estabelecidos. Nesse sentido, para
atender as exigências do acometimento epidemiológico da
doença, o termo “adulto” foi substituído para “agudo”, visto que
essa patologia acometia também crianças e jovens, ficando
então Síndrome do Desconforto Respiratório Agudo (SDRA)

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
188
(BERNARD et al., 1994; THOMPSON; CHAMBERS; LIU, 2017;
MOWERY; TERZIAN; NELSON, 2020).
Em 1994, a AECC publicou as condições clínicas para
definição e diferenciação da Lesão Pulmonar Aguda (LPA) e da
SDRA, onde ambas apresentam início agudo, hipoxemia
refratária à oxigenoterapia e são caracterizadas pelo achado de
infiltrados pulmonares bilaterais no exame radiológico de tórax.
A diferença entre elas está centrada na relação da pressão
parcial de oxigênio – PaO2/FiO2 ≤ 300 mmHg na LPA e
PaO2/FiO2 ≤ 200 mmHg para a SDRA. Ainda, as condições
clínicas dos pacientes acometidos com essa síndrome levaram
especialistas a subdividi-la de acordo com sua gravidade,
baseando-se também nas condições dos sinais vitais,
classificando-a em leve, grave e moderada. Aos poucos o termo
LPA foi descontinuado na prática médica, visto a dificuldade dos
atendentes em diferenciar a SDRA leve da LPA, considerando
a última como uma condição mais leve da síndrome. Logo “LPA”
é usada quando a intenção é avaliar as alterações
morfopatológicas presentes na enfermidade (BERNARD et al.,
1994; MOWERY; TERZIAN; NELSON, 2020).
• Aspectos fisiopatológicos
A disfunção epitélio-endotelial se mostra como um ponto
importante no desenvolvimento da LPA. Quando submetido a
exposição de agentes lesivos, as células teciduais são capazes
de liberar diversos mediadores inflamatórios importantes para o
processo inflamatório e desenvolvimento da doença, como: IL-
1β, IL-6, IL-8 e fator de necrose tumoral alfa (TNF-α)
(OLIVEIRA; PIUVEZAM, 2019).

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
189
A IL-1β e o TNF-α já possuem suas atividades bem
definidas no desenvolvimento da patologia (OLIVEIRA;
PIUVEZAM, 2019). São liberadas por células residentes
teciduais, os macrófagos alveolares, os quais são membros
importantes da imunidade inata e destacam-se por serem uma
das primeiras linhas de defesa contra microrganismos,
reconhecendo agentes indutores como padrões moleculares
associados a patógenos (PAMPs), através de receptores de
reconhecimento padrão (PRRs), a exemplo dos receptores do
tipo toll (TLR) (OLIVEIRA; PIUVEZAM, 2019). Os macrófagos
são capazes de liberar, também, exossomos, os quais podem
contribuir para o desenvolvimento da LPA, devido a sua
capacidade de interromper a expressão de proteínas de junção
das células, favorecendo a resposta inflamatória e o
rompimento da barreira estrutural (YUAN; BEDI; SADIKOT,
2018).
A liberação de citocinas pelos macrófagos é capaz,
ainda, de ativar o endotélio dos vasos sanguíneos, aumentando
a expressão de moléculas de adesão, recrutando e ativando
neutrófilos (DO NASCIMENTO XAVIER et al., 2019). Os
neutrófilos ativados são recrutados e se acumulam no espaço
aéreo e, em seguida, liberam diversos mediadores como
proteinases, espécies reativas de oxigênio, fatores de
crescimentos e citocinas. Somado a isto, a liberação de MIF
(macrophage migration inhibitory fator) dentro do ambiente local
favorece ainda mais a resposta pró-inflamatória, causando mais
lesão endotelial e epitelial alveolar. Esses eventos constituem o
centro patológico da LPA (LU et al., 2018).
A disseminação do excesso de líquido nos espaços
aéreos dos alvéolos, decorrente do aumento da permeabilidade

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
190
vascular e do dano alveolar difuso, juntamente com o declínio
na produção do surfactante, resultam no colapso alveolar,
aumento do espaço morto e dificulta as trocas gasosas
(SIEGEL; PARSONS, 2018).
O dano alveolar difuso (DAD), figura 1, é um evento
dividido em três fases: fase aguda/precoce ou exsudativa,
organizadora ou proliferativa e tardia ou fibrótica. Na fase
exsudativa, o DAD faz com que as células epiteliais e
endoteliais produzam fatores que reagem à lesão e à morte. É
possível verificar a presença de membranas hialinas distintivas
que alinham espaços alveolares, além de edema, que também
é frequentemente identificado, podendo haver ainda
hemorragia alveolar aguda (MOWERY; TERZIAN; NELSON,
2020). Devido à perda da integridade celular, os alvéolos são
preenchidos com fluido de edema proteico, o que resulta em
comprometimento das trocas gasosas (BUTT; KURDOWSKA;
ALLEN, 2016).
As membranas hialinas começam a se organizar à
medida que o DAD continua, na fase de organização, e o tecido
de granulação se desenvolve nos espaços alveolares. A
diluição e a disfunção do surfactante pulmonar levam ao
colapso dos alvéolos e à diminuição da complacência pulmonar
(MOWERY; TERZIAN; NELSON, 2020).
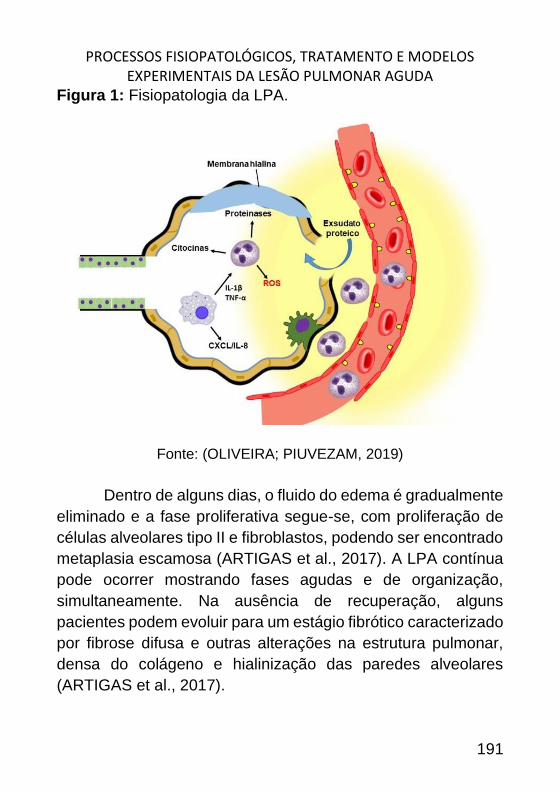
PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
191
Figura 1: Fisiopatologia da LPA.
Fonte: (OLIVEIRA; PIUVEZAM, 2019)
Dentro de alguns dias, o fluido do edema é gradualmente
eliminado e a fase proliferativa segue-se, com proliferação de
células alveolares tipo II e fibroblastos, podendo ser encontrado
metaplasia escamosa (ARTIGAS et al., 2017). A LPA contínua
pode ocorrer mostrando fases agudas e de organização,
simultaneamente. Na ausência de recuperação, alguns
pacientes podem evoluir para um estágio fibrótico caracterizado
por fibrose difusa e outras alterações na estrutura pulmonar,
densa do colágeno e hialinização das paredes alveolares
(ARTIGAS et al., 2017).

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
192
• Aspectos imunológicos
Embora a inflamação seja um processo fisiológico que
visa a restauração da homeostase, sua estimulação prolongada
pode levar a exarcebação da resposta inflamatória e causar
danos à tecidos e órgãos. Na LPA, não é diferente, havendo
complicações nos pulmões e órgãos vitais, que comprometem
a oxigenação e pode rapidamente levar os indivíduos
acometidos ao óbito (BUTT; KURDOWSKA; ALLEN, 2016).
A diminuição do processo inflamatório local, em
detrimento da migração de neutrófilos, se apresenta como um
resultado inicial importante para o manejo da LPA. Isso porque
tais células são cruciais no agravamento das manifestações
observadas em doenças pulmonares (POLVERINO et al.,
2017).
Em protocolos experimentais de LPA, a exacerbação da
resposta neutrofílica já é bem esclarecida, e tornou-se alvo
importante. O excesso de atividade da elastase de neutrófilos e
proteases semelhantes causam danos aos tecidos e alteram o
processo de remodelação em muitas outras condições clínicas
(POLVERINO et al., 2017). Em adição, os neutrófilos são fontes
importantes de espécies reativas de oxigênio (EROs), as quais
causam disfunção endotelial e lesão tecidual, e, sob as
condições inflamatórias, levam à abertura de junções
endoteliais, favorecendo a migração de células inflamatórias
(KELLNER et al., 2017). Ainda, as armadilhas extracelulares de
neutrófilos (NETs), propostas como um mecanismo de defesa
inato para depuração de patógenos, podem induzir danos
colaterais aos tecidos do hospedeiro, favorecendo a
sobrevivência e permanência de neutrófilos no tecido pulmonar
(LEFRANÇAIS et al., 2018).

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
193
Os macrófagos, por sua vez, também desempenham
papel importante na LPA (XIAOFANG et al., 2018). Os
macrófagos M1, quando ativados, liberam citocinas pró-
inflamatórias que favorecem o recrutamento de células, o
aumento da permeabilidade capilar e a expressão de moléculas
de adesão do endotélio (LU et al., 2018). Porém, os macrófagos
desempenham duplo papel dentro do processo fisiopatológico
dessa doença, pois os de perfil M2 foram relatados como
importantes para eliminar células apoptóticas e participar da
fibrose. Além disso, esse perfil é capaz de suprimir a produção
de TNF-α, IL-1β e IL-6, e aumentar a produção de IL-10,
favorecendo o reparo tecidual. Considerando isso, a
manutenção desse perfil celular é importante a depender da
fase de desenvolvimento da LPA, do grau de severidade e da
resposta ao tratamento diante de outras células principais
(SHEN et al., 2019).
O papel dos linfócitos no desenvolvimento da LPA ainda
não é bem esclarecido, mas sabe-se que alguns perfis celulares
desempenham atividades importantes (LIN et al., 2018). Os
linfócitos T auxiliares do perfil TH17, apresentam-se como
potentes auxiliares no recrutamento de macrófagos e
polimorfonucleares. Modelos experimentais já mostraram que
sua modulação negativa é importante para melhora do quadro
de LPA, onde linfócitos T regulatórios foram considerados uma
peça fundamental no balanceamento da resposta induzida
pelos linfócitos TH17 (WANG et al., 2018).
O receptor solúvel para produtos finais de glicação
avançada (sRAGE) há muito foi identificado como um
biomarcador de diagnóstico e prognóstico de SDRA. RAGE é
um receptor de membrana mais altamente expresso nas células

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
194
epiteliais alveolares do tipo I, mas também expresso em outros
tecidos. RAGE é um receptor de reconhecimento de padrões
ativado por AGEs (YEHYA, 2019).
A forma solúvel (sRAGE), resultante de splicing
alternativo ou por clivagem ativa, foi sugerida como um
marcador epitelial específico da gravidade da SDRA. A
expressão de sRAGE elevada está, consistentemente,
associada a resultados ruins na SDRA. A fonte e a função de
sRAGE não são totalmente claras e é possível que seja um
marcador de dano epitelial. Alternativamente, no cenário de
inflamação, sRAGE elevado pode atuar como um receptor
chamariz para amortecer a inflamação ao se ligar a AGEs e
outros agonistas do RAGE ligado à membrana (YEHYA, 2019).
• Tratamento
Embora a mortalidade seja elevada, os avanços na
terapêutica e intervenções ainda não se mostram tão eficazes
à curto prazo, sendo a principal intervenção o manejo do
paciente. No que tange às medidas não farmacológicas, o uso
de ventilação mecânica (VM) é comumente utilizado, porém a
sua configuração inadequada pode induzir uma lesão pulmonar
secundária. Esta, quando combinada com uma síndrome da
resposta inflamatória sistêmica (FAZZA et al., 2020), pode atuar
no agravamento da lesão pulmonar evoluindo para SDRA
(YEHYA, 2019).
A administração de terapêuticas farmacológicas, como
broncodilatadores e corticosteroides, também é realizada na
busca da melhoria das condições patológicas. São utilizados
broncodilatadores β2-agonistas, capazes de melhorar a
depuração de fluidos e favorecer a folga mucociliar (ARTIGAS

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
195
et al., 2017), e os corticosteroides, devido aos seus efeitos anti-
inflamatórios, especialmente durante a fase exsudativa inicial e
no dano hemorrágico alveolar difuso (ARTIGAS et al., 2017),
além de promover efeitos antifibróticos do cortisol. Entretanto,
o uso de corticosteroides não se mostra eficaz para pacientes
de alto risco (MOKRA et al., 2019).
A farmacoterapia da SDRA, embora ocorra por inalação,
está associada a uma série de efeitos colaterais, como tremor
muscular e desequilíbrio ventilação-perfusão. Além disso, a
SDRA enfrenta três desafios farmacológicos relativamente
únicos (MOKRA et al., 2019). Primeiramente, os pacientes com
SDRA são vulneráveis devido à disfunção de múltiplos órgãos
concomitantemente, portanto, eles podem não tolerar os efeitos
indesejados das drogas. Segundo, a administração de
medicamentos inalatórios é impedida pelo fluido proteico nos
alvéolos lesados e pela distribuição não homogênea da
ventilação, onde a área do pulmão lesada não é ventilada. Por
último, a SDRA é heterogênea em sua fisiopatologia
subjacente, portanto é improvável que o direcionamento a uma
via melhore os resultados da maioria dos pacientes (ARTIGAS
et al., 2017).
As limitações na terapia farmacológica da SDRA têm
estimulado uma busca crescente por novas alternativas mais
eficazes e seguras para tratar essa síndrome, incluindo o
reposicionamento de fármacos já utilizados na clínica. É o caso
das estatinas, uma classe de fármacos utilizada no tratamento
das dislipidemias. As mesmas foram capazes de reduzir a
inflamação e a progressão da lesão pulmonar em modelos
experimentais e melhoraram o quadro de pacientes com SDRA,
em termos de falência de órgãos e redução da necessidade de

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
196
ventilação. Porém, o tamanho da amostra utilizada nos estudos
é pequeno para avaliar, com precisão, os efeitos sobre a
mortalidade e a sepse, e investigações adicionais são
necessárias (THOMPSON; CHAMBERS; LIU, 2017; FENG,
2018).
Ainda, tem se avaliado a aplicação do fator de
crescimento de queratinócitos (KGF) no tratamento da SDRA.
O KGF está envolvido no reparo do epitélio alveolar e, portanto,
poderia ser útil para resolução dos edema e lesão pulmonar.
Trabalhos com modelos experimentais e ensaios clínicos
iniciais apontam efeito benéfico do KGF na SDRA, porém mais
estudos são necessários para conclusões mais consistentes
(THOMPSON; CHAMBERS; LIU, 2017).
• Modelos experimentais
Desde a descrição da SDRA, em 1967, os pesquisadores
têm lutado para reproduzi-la em animais de laboratório. Apenas
recentemente, houve a padronização dos critérios para que um
modelo de ALI de laboratório seja comparável ao SDRA
humano. Embora vários modelos diferentes de lesão pulmonar
aguda tenham sido desenvolvidos, nenhum captura
completamente as etiologias incitadoras, inflamação inicial,
heterogeneidade e resolução da SDRA humana. Isso tem
contribuído potencialmente para a má tradução de potenciais
terapêuticos entre modelos animais de ALI e ARDS humano
(YEHYA, 2019). No entanto, apesar da heterogeneidade dos
modelos, estes contribuíram substancialmente para a nossa
compreensão dessa síndrome respiratória (YEHYA, 2019).
Visto que a SDRA é uma doença com consideráveis
alterações patológicas, somado a uma heterogênese em sua

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
197
etiologia, é importante buscar modelos experimentais em que
seja possível avaliar tanto suas alterações morfológicas, como
a manutenção dessas. Assim, mesmo com suas limitações, os
animais de laboratório se mostram eficazes na avaliação das
alterações patológicas, bem como na experimentação de novas
moléculas para avaliação terapêutica.
Modelos de roedores, por exemplo, têm uma resposta
exagerada para produzir óxido nítrico e reconhecem diferentes
padrões moleculares associados a patógenos em diferentes
afinidades devido a diferenças em seus receptores Toll-like
(YEHYA, 2019). Ovelhas, em contrapartida, possuem
macrófagos pulmonares intravasculares que as predispõem a
pior lesão pulmonar, após o desafio de endotoxina (YEHYA,
2019). No entanto, os ratos têm a vantagem de nocautes,
relativamente, fáceis de criar, além da possibilidade de outras
manipulações genéticas para interrogar as vias de sinalização,
enquanto animais maiores permitem maior monitoramento
fisiológico e comprimentos mais longos de ventilação,
congruentes com a SDRA humana (YEHYA, 2019).
Um dos achados mais traduzíveis de modelos animais
da LPA/SDRA humana foi o dano associado à ventilação
mecânica (VM), uma entidade multifatorial conhecida como VILI
(LIU et al., 2019). A VM em VT ou pressões excessivas, leva a
alterações fisiopatológicas que lembram a SDRA do homem
(FAZZA et al., 2020). O dano depende da magnitude e da
duração, com alongamento excessivo e atelectasia (FAZZA et
al., 2020). Ainda, a LPIVM é influenciada pelo tamanho do
animal, se o tórax estava aberto ou fechado e se a pressão
expiratória final positiva extrínseca foi usada (YEHYA, 2019).

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
198
Os modelos experimentais induzidos com
lipopolissacarídeos (LPS) constituem o segundo protocolo
experimental mais utilizado na prática não clínica, perdendo
apenas para indução por ventilação mecânica (LOPES
GIACOMINI et al., 2020). Por se tratar de um componente
presente na membrana de bactérias gram-negativas, o LPS é
capaz de agir como endotoxinas e serem reconhecidos como
PAMPs pelos macrófagos teciduais, desencadeando a reposta
inflamatória (LOPES GIACOMINI et al., 2020).
Há evidências, ainda, de que a exposição à fumaça do
cigarro (CS) aumenta o risco de desenvolver a
SDRA. Streptococcus pneumoniae é a causa mais comum de
pneumonia bacteriana, a qual se relaciona com a
SDRA. Fumantes crônicos têm taxas aumentadas de
colonização pneumocócica e desenvolvem pneumonia mais
grave do que os não fumantes. As conexões entre a exposição
à SC, pneumonia e a SDRA permanecem inexploradas. Sabe-
se que a exposição aguda a CS aumenta o recrutamento de
células imunes para o pulmão durante a pneumonia bacteriana,
causando lesões pulmonares inflamatórias mais graves em
camundongos (GOTTS et al., 2018).
CONCLUSÕES
A Lesão Pulmonar Aguda (LPA) e a Síndrome do
Desconforto Respiratório Agudo (SDRA) são bastante
difundidas em todo o mundo, com altas taxas de mortalidade.
Essa revisão fornece um compilado importante sobre inúmeros
processos envolvidos na LPA. Diante desses achados, é
possível observar que a LPA/SRDA é uma condição patológica

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
199
complexa, com inúmeros alvos e problemáticas terapêuticas.
Essa revisão fornece, ainda, modelos experimentais que podem
ser aplicados no sentido de compreender melhor as
manifestações da doença, bem como pode servir como base
para investigação de moléculas com potenciais terapêuticos.
REFERÊNCIAS BIBLIOGRÁFICAS ARTIGAS, A. et al. Inhalation therapies in acute respiratory distress syndrome. Annals of Translational Medicine, v. 5, n. 14, p. 293–293, jul. 2017. BERNARD, G. R. et al. Report of the American-European consensus conference on ARDS: Definitions, mechanisms, relevant outcomes and clinical trial coordination. Consensus Committee. Intensive Care Medicine, v. 9, n. 1, p. 72–81, 1994. BUTT, Y.; KURDOWSKA, A.; ALLEN, T. C. Acute lung injury: A clinical and molecular review. Archives of Pathology and Laboratory Medicine, v. 140, n. 4, p. 345–350, 2016. CASER, E. B. et al. Impact of distinct definitions of acute lung injury on its incidence and outcomes in Brazilian ICUs: Prospective evaluation of 7,133 Patients. Critical Care Medicine, v. 42, n. 3, p. 574–582, 2014. DO NASCIMENTO XAVIER, B. M. et al. MHTP, a synthetic tetratetrahydroisoquinoline alkaloid, attenuates lipopolysaccharide-induced acute lung injury via p38MAPK/p65NF-κB signaling pathway-TLR4 dependent. Inflammation Research, v. 68, n. 12, p. 1061–1070, out. 2019. FAZZA, T. F. et al. Effect of low-level laser therapy on the inflammatory response in an experimental model of ventilator-induced lung injury. Photochemical & Photobiological Sciences, v. 19, n. 10, p. 1356–1363, 2020. FENG, Y. Efficacy of Statin therapy in patients with acute respiratory distress syndrome/acute lung injury: A systematic review and meta-analysis. European Review for Medical and Pharmacological SciencesVerduci Editore s.r.l, , 2018. FUJISHIMA, S. et al. Demographics, Treatments, and Outcomes of Acute Respiratory Distress Syndrome: The Focused Outcomes Research in Emergency Care in Acute Respiratory Distress Syndrome, Sepsis, and Trauma (Forecast) Study. Shock (Augusta, Ga.), 23 jul. 2019. GALLELLI, L. et al. Severe Acute Lung Injury Related to COVID-19 Infection: A Review and the Possible Role for Escin. Journal of Clinical PharmacologyBlackwell Publishing Inc., 2020.

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
200
GOTTS, J. E. et al. Cigarette smoke exposure worsens acute lung injury in antibiotic-treated bacterial pneumonia in mice. American Journal of Physiology - Lung Cellular and Molecular Physiology, v. 315, n. 1, p. L25–L40, 1 jul. 2018. HARRINGTON, J. S. et al. Acute respiratory distress syndrome without identifiable risk factors: A secondary analysis of the ARDS network trials. Journal of Critical Care, v. 47, p. 49–54, 2018. KELLNER, M. et al. ROS signaling in the pathogenesis of Acute Lung Injury (ALI) and Acute Respiratory Distress Syndrome (ARDS). In: Advances in Experimental Medicine and Biology. [s.l.] Springer New York LLC, 2017. v. 967p. 105–137. LEFRANÇAIS, E. et al. Maladaptive role of neutrophil extracellular traps in pathogen-induced lung injury. JCI insight, v. 3, n. 3, 8 fev. 2018. LIN, S. et al. Regulatory T cells and acute lung injury: Cytokines, uncontrolled inflammation, and therapeutic implications. Frontiers in ImmunologyFrontiers Media S.A., , 9 jul. 2018. LIU, Q. et al. JAK2/STAT1-mediated HMGB1 translocation increases inflammation and cell death in a ventilator-induced lung injury model. Laboratory Investigation, v. 99, n. 12, p. 1810–1821, 1 dez. 2019. LOPES GIACOMINI, R. et al. MODELOS EXPERIMENTAIS EM LESÃO PULMONAR AGUDA EXPERIMENTAL MODELS IN ACUTE LUNG INJURY. Revista Científica Fagoc Saúde. 2020. LU, H. L. et al. Activation of M1 macrophages plays a critical role in the initiation of acute lung injury. Bioscience Reports, v. 38, n. 2, 27 abr. 2018. MOKRA, D. et al. Corticosteroids in acute lung injury: The dilemma continuesInternational. Journal of Molecular SciencesMDPI AG, , 1 out. 2019. MOWERY, N. T.; TERZIAN, W. T. H.; NELSON, A. C. Acute lung injury. Current Problems in Surgery, v. 57, n. 5, 1 maio 2020. OLIVEIRA, J. B. DE; PIUVEZAM, M. R. Avaliação do tratamento com MHTP ([2-methoxy-4-(7-methoxy-1,2,3,4-tetrahydroisoquinolin-1-yl) phenol] na migração celular, exsudato proteico e parâmetros histopatológicos em modelo murino de Lesão Pulmonar Aguda (LPA). In: Série Iniciados 2017-2018. João Pessoa: Editora UFPB, 2019. v. 24p. 67–85. POLVERINO, E. et al. The Role of Neutrophil Elastase Inhibitors in Lung Diseases. Chest, v. 152, n. 2, p. 249–262, 1 ago. 2017. SCHUPP, J. C.; PRASSE, A.; ERYTHROPEL, H. C. E-Cigarettes - Operating Principle, Ingredients, and Associated Acute Lung Injury. Pneumologie, v. 74, n. 2, p. 77–87, 1 fev. 2020. SHEN, Y. et al. M2 macrophages promote pulmonary endothelial cells regeneration in sepsis-induced acute lung injury. Annals of Translational

PROCESSOS FISIOPATOLÓGICOS, TRATAMENTO E MODELOS EXPERIMENTAIS DA LESÃO PULMONAR AGUDA
201
Medicine, v. 7, n. 7, p. 142–142, abr. 2019. SIEGEL M, PARSONS P, F. G. Acute respiratory distress syndrome: Clinical features, diagnosis, and complications in adults. UpToDate, 2018. SORIANO, J. B. et al. Global, regional, and national deaths, prevalence, disability-adjusted life years, and years lived with disability for chronic obstructive pulmonary disease and asthma, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. The Lancet Respiratory Medicine, v. 5, n. 9, p. 691-706, 2017. THOMPSON, B. T.; CHAMBERS, R. C.; LIU, K. D. Acute respiratory distress syndrome. New England Journal of Medicine, v. 377, n. 6, p. 562-572, 2017. VLAAR, A. P. J. et al. A consensus redefinition of transfusion-related acute lung injury. Transfusion, v. 59, n. 7, p. 2465–2476, 1 jul. 2019. WANG, L. et al. Recovery from acute lung injury can be regulated via modulation of regulatory T cells and Th17 cells. Scandinavian Journal of Immunology, v. 88, n. 5, 1 nov. 2018. XIAOFANG, H. et al. The Role of Macrophages in the Pathogenesis of ALI/ARDS. Mediators of inflammation, n. 1264913, 2018. YEHYA, N. Lessons learned in acute respiratory distress syndrome from the animal laboratory. Annals of Translational Medicine, v. 7, n. 19, p. 503–503, out. 2019. YUAN, Z.; BEDI, B.; SADIKOT, R. T. Bronchoalveolar Lavage Exosomes in Lipopolysaccharide-induced Septic Lung Injury. Journal of Visualized Experiments, n. 135, p. e57737, 2018.

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
202
CAPÍTULO 10
TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE
DOENÇAS CARDIOVASCULARES: UMA REVISÃO
Júlio César Pinheiro Lúcio de OLIVEIRA 1
Rubens da Silva ARAÚJO 1 Herlanny Santana BEZERRA 1
Alline Villar de ARRUDA 2 Tays Amanda FELISBERTO 2,3
1 Graduando do curso de farmácia, UFPB; 2 Doutoranda do programa de produtos naturais e sintéticos bioativos; 3 Orientadora;
RESUMO: O tecido adiposo perivascular vem se consolidando como um órgão com atividades diversas. Várias substâncias vêm sendo relatadas como agentes que melhoram a função vascular por mediação do perivascular adipose tissue (PVAT) em modelos de doenças em animais. Assim, o objetivo desse estudo foi fazer um levantamento das substâncias-teste utilizadas em modelos animais de doenças os quadros patológicos por intermédio do PVAT. Para isso, os termos Perivascular adipose tissue + treatment foram pesquisados em bases de dados. Os artigos publicados entre 2016 e 2020, que tinham no título ou resumo os termos gordura periarterial, tecido adiposo perivascular e PVAT (termos em inglês), em inglês e que estudaram a ação de substâncias em modelos de doenças metabólicas e vasculares foram selecionados. Os 14 artigos selecionados trouxeram substâncias que atuaram na melhoria do quadro nos modelos de doenças em animais pela mediação do PVAT. Isto é, as substâncias mudaram o fenótipo ou o tipo

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
203
de substâncias que esse tecido secreta para atuar de forma parácrina nos vasos sanguíneos ou endócrina. Assim, concluímos que o PVAT é um ponto-chave no desenvolvimento de novas terapias que visem tratar as doenças que afetam o sistema cardiovascular e, principalmente, a síndrome metabólica. Palavras-chave: Síndrome metabólica. Obesidade. Tecido adiposo. Artérias.
INTRODUÇÃO
O tecido adiposo perivascular ou perivascular adipose
tissue (PVAT) é composto basicamente por tecido conjuntivo e
por células adiposas que podem ser do tipo branca, marrom ou
ambas. Antes dos anos 90 acreditava-se que a única função
para o sistema cardiovascular desse tecido era proporcionar
sustentação aos órgãos do referido sistema, porém, Soltis e
Cassis demonstraram em 1991 que o PVAT tinha capacidade
de influenciar, in vitro, a contractilidade de anéis da aorta
torácica de ratos (SOLTIS; CASSIS, 1991).
Apenas em 2002 o PVAT voltou à tona novamente,
sendo demonstrado, nesse ano, a capacidade dele de diminuir
a resposta contrátil de anéis de aorta à angiotensina II, a
fenilefrina e a serotonina. Também, foi demonstrado que esse
efeito não era causado apenas pela barreira mecânica que ele
gerava, mas sim por compostos vasoativos que causavam a
vasodilatação e exerciam ação anticontraturante nos anéis
arteriais. Ademais, ficou demonstrado que o PVAT tinha a
capacidade de liberar para o meio fatores que causavam
vasorrelaxamento de anéis da aorta torácica pré-contraídos
com fenilefrina ou noradrenalina (LOHN et al., 2001). Tais
achados fundamentaram os estudos que prosseguiram para a

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
204
elucidação da função fisiológica do PVAT sobre os vasos
sanguíneos e sobre a gênese de algumas doenças como a
hipertensão arterial sistêmica, a hipertrofia arterial, a
arteriosclerose e as doenças cardiovasculares decorrentes da
obesidade (SAXTON et al., 2019).
Os vasos sanguíneos são constituídos por três camadas:
As túnicas íntima, média e adventícia. Mas há alguns anos
diversos autores vem considerando o PVAT como uma quarta
camada que compõe os vasos – a túnica adiposa. Para tal, na
última década foi (e continua sendo) crescente o relato de
funções do PVAT sobre a regulação da fisiologia cardiovascular
e de sua importância na gênese de algumas doenças
(ZEMANČÍKOVÁ; TÖRÖK, 2019).
Hoje, o PVAT já é reconhecido devido as suas funções
parácrinas e endócrinas, secretando os fatores relaxantes
derivados do PVAT (PVAT-deriveted relaxing factors – PVRFs),
citocinas e adipocinas para a realização das suas funções sobre
o sistema cardiovascular. Os únicos vasos sanguíneos que
esse tecido não recobre são os da microcirculação e os
cerebrais (SAMUEL, 2020).
Esse tecido adiposo especial pode ser composto por
tecido adiposo branco ou marrom. O tecido adiposo branco é
composto por adipócitos com uma única gota de lipídeo, com
poucas mitocôndrias e baixa taxa de oxidação lipídica, sendo
responsável pelo armazenamento de gordura para ser utilizada
como fonte energética para o corpo. Já os adipócitos que
compões o tecido adiposo marrom são de tamanhos menores
e com gotículas lipídicas menores e têm como função principal
a produção de calor participando, assim, da regulação da
temperatura do corpo (QUEIROZ; SENA, 2020).

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
205
Hoje em dia, diversos grupos de estudos buscam meios
de tratar a obesidade (e como consequência os problemas
vasculares associados à ela) por meio de medidas não-
farmacológicas e farmacológicas.
Dentre as medidas não-farmacológicas a mais utilizada
e estudada é a realização de exercícios físicos. Tal medida é
comprovadamente eficaz para o tratamento de comorbidades
como a obesidade, a hipertensão e o diabetes (PICHÉ; MORIN;
POIRIER, 2020).
Partindo disso, o objetivo dessa revisão foi fazer um
levantamento da participação do PVAT como alvo
farmacológico de substâncias para o tratamento de doenças em
modelos de animais, buscando pontuar os achados mais
importantes por trabalho e revisar a fisiologia e fisiopatologia
desse tecido como agente ativo na gênese e manutenção de
doenças vasculares.
MATERIAIS E MÉTODO
Para a realização da presente revisão foram feitas
consultas nas seguintes bases de dados científicos eletrônicos
indexadas aos periódicos da Coordenação de Aperfeiçoamento
de Pessoal de Nível Superior (CAPES): Web of Science e
PubMed. O levantamento foi realizado durante julho a agosto
de 2020.
Para tal, utilizou-se as seguintes palavras-chave:
Perivascular adipose tissue + treatment.
Delineou-se que seriam selecionados artigos escritos
apenas em inglês e publicados entre janeiro de 2016 até julho
de 2020 e que visassem elucidar a intermediação do PVAT na
ação de substâncias em modelos de doenças em ratos.

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
206
Os artigos encontrados nas bases de dados foram
adicionados ao site Ryyan QCRI para facilitar o gerenciamento
dos artigos que seriam adicionados a revisão.
Os critérios de inclusão dos artigos para a leitura
completa foram: publicados entre 2016 e 2020, que se
caracterizam como estudos pré-clínicos em animais, que
traziam no título ou resumo os termos perivascular adipose
tissue ou PVAT.
Os artigos foram adicionados e discutidos na presente
revisão se visassem elucidar a ação de substâncias sobre o
PVAT.
RESULTADOS E DISCUSSÃO
Tecido adiposo perivascular: na saúde e na doença.
O PVAT corresponde a aproximadamente 3% de todo o
tecido adiposo corporal, revestindo quase todos os vasos
sanguíneos, com excessão dos vasos cerebrais, as vênulas e
arteríolas (SIEGEL-AXEL; HARING., 2016). Antes dos
trabalhos realizados entre os anos 1990 a 2000, acreditávasse
que o PVAT exercia função estrutural e de sustenção às veias
e artérias. Contudo, já foi confirmado que o PVAT exerce função
reguladora sobre a fisiologia vascular, dependendo do tipo de
PVAT e seu estado (LOHN et al., 2001).
Os fatores relaxantes derivados dos adipócitos (Em inglês:
adipocyte-derived relaxing factor, ADRF) compreendem um
grupo de moléculas que demonstraram atuar sobre o leito
arterial, alterando seu tônus e, ainda, de natureza não
identificada (NOSALSKI; GUZIK, 2017).

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
207
Apesar disso, algumas substâncias são candidatas a
atuarem como ADRF, tais como o sulfeto de hidrogênio
(CACANYIOVA et al., 2019), a adiponectina (ALMABROUK et
al., 2018) , a prostaciclina (AWATA et al., 2019) e a angiotensina
1-7 (AGABITI-ROSEI et al., 2018).
Há certo tempo o tecido adiposo já vem sendo
reconhecido como um órgão endócrino que secreta adipocinas
que promemovem ou reduzem a inflamação no PVAT e nos
vasos sanguíneos (NOSALSKI; GUZIK, 2017).
Dentre as adipocinas já relatadas na literatura até hoje,
duas se destacam pelos efeitos mais proeminentes: a leptina e
a adiponectina.
A leptina é uma proteína bem conservada no processo
evolutivo e que seu mecanismo de ação envolve a ativação do
receptor Ob-R, o qual atua como pivô da ativação de diversas
vias de sinalização, principalmente ada janus cinase
(JAK/STAT), a da proteína ativada por mitógeno (MAPK), a da
fosfatidilinositol-3-cinase/serina-treonina cinase (PI3K/AKT) e
da cinase ativada pelo AMP (AMPK). A ativação de tais
mecanismos irá desencadear efeitos como a proliferação
celular e a regulação imunológica. Vale salientar que como
citona a leptina exerce forte efeito inflamatório, o que pode estar
associado a disfunção vascular e a síndrome metabólica
(PHAM; PARK., 2020). Além disso, já foi demonstrado que essa
adipocina está superexpressa em pacientes que apresentam
problemas cardiovasculares associados ao diabetes
(SPYCHALSKA-ZWOLIŃSKA et al., 2020).
Também, a adiponectina constitui outra adipocinina que é
encontrada no plasma humano e de forma mais abundante que
as outras, sendo majoritariamente produzida pelos adipócitos e
tendo a produção inibida na presença de citocinas pro-

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
208
inflamatórias como o fator de necrose tumoral alfa (TNF-α) e a
interleucina 6 (IL-6) (LIM; KIM, 2020).
Essa proteína pode ser encontrada na forma oligomérica
(trímeros, pentâmeros, etc) ou pode ser clivada em
adiponectina globular, a qual exerce maior efeito biológico do
que a não-clivada e está presente em menor concentrações na
circulação sanguínea humana (PHAM; PARK., 2020).
De certo, a adiponectina é um dos principais fatores que
causam vasorrelaxamento e que está associada a melhora da
função vascular em alguns modelos de doenças (como a
obesidade e a hipertensão) em animais, sendo sugerido que
tais ações e decorrente da ativação dos receptores beta-3
adrenérgicos no PVAT que, ativados, irão desencadear a
liberação de adiponectina por esse tecido e a ativação do
receptor adipoR1 (receptor da adiponectina tipo 1) no músculo
liso vascular, o qual desencadeará a ativação da AMPK e de
canais de potássio de longa condutância ativados por cálcio
(BKCa) (BAYLIE et al., 2017).
Alguns estudos sugerem que a disfunção na expressão de
tais adipocinas podem desencadear e/ou ampliar a inflamação
e mudança de fenótipo do PVAT e, assim, o aumento da
resistência vascular, pois já foi amplamente observado que a
perda de PVAT sadio desencadeia o aumento da pressão
arterial sistêmica em modelos de obesidade em ratos (SAXTON
et al., 2019). Tal ligação torna-se um ponto de estudo animador
que liga a obesidade à gênese da hipertensão.
Na obesidade, o efeito anticontrátil do PVAT é totalmente
perdido, sendo atribuido a isso a alteração do perfil de
adipocinas e de autacoides liberados por esse tecido para o
ambiente da musculatura lisa e endotelial dos vasos
sanguíneos. A disfunção do PVAT desencadeia o aumento do

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
209
tônus vascular, o aumento da espessura da túnica média
(devido a hipertrofia e hiperplasia da musculatura lisa vascular),
o aumento do estresse oxidativo e aumento da expressão e
atividade do sistema renina-angiotensina aldosterona. Tais
alterações irão contribuir para o aparecimento da hipertensão
arterial (AGABITI-ROSEI et al., 2018).
Além disso, essa alteração nas adipocinas e do PVAT
desencadeará a diminuição da perfusão sanguínea muscular,
da sensibilidade dos tecidos à insulina, o metabolismo e,
também, a indução da inflamação no fígado. Tais fatores irão
tornar o organismo em um fenótipo favorável ao
desenvolvimento do diabetes do tipo 2 (AZUL et al., 2020).
Há uma ligação íntima entre o processo inflamatório
crônico que se estabelece no PVAT de modelos animais de
obesidade e o aparecimento de doenças cardiovasculares
(SAXTON; WITHERS; HEAGERTY, 2019). Sendo assim, a
modulação desse processo torna-se um potencial meio
terapêutico para o tratamento das doenças cardiovasculares
relacionadas à obesidade. Diversos estudos apontam os
benefícios das atividades físicas como forma de tratamento da
obesidade e de melhoria das funções cardiovasculares
comprometidas, sendo os exercícios físicos adotados como
medida complementar do tratamento farmacológico para a
hipertensão e diabetes por diversas diretrizes pelo mundo.
Todavia, a realização de tais atividades depende do grau de
empenho dos pacientes.
Interessantemente, alguns estudos apontam que a
infecção por helmintos pode ser, no futuro, uma ferramenta para
o tratamento da obesidade. Já foi demonstrado que a infecção
por Strongyloides venezuelensis em ratos obesos promoveu a
redução do peso dos animais e aumento da sensibilidade à

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
210
insulina por meio da modulação da microbiota intestinal e
diminuição, também, de algumas citocinas pró-inflamatórias
(PACE et al., 2018).
Saxton e colaboradores (2019) hipotetizam que o uso de
helmintos pode ser explorado para a restauração das funções
benéficas ao sistema vascular do PVAT. Eles se baseiam que
a infecção por helminto aumenta a concentração sérica de
eosinófilos, ao passo que a diminuição de eosinófilos no PVAT
está relacionado a sua perda de função protetora ao sistema
cardiovascular. Tais hipóteses são reforçadas pelos diversos
trabalhos que podem ser encontrados nas bases de dados e
que apontam esses efeitos.
Diante disso, o PVAT torna-se um alvo promissor para o
tratamento de doenças relacionadas à obesidade. Os achados
desde 1990 até hoje sobre o envolvimento desse tecido nos
processos vasculares e na participação da gênese de doenças
cardiovasculares abriram uma gama de possibilidades para que
a comunidade científica explore a atuação de substância e
tratamentos que revertam o estado patológico do PVAT e
restaure seu efeito anticontrátil e anti-inflamatório.
Intermédio do PVAT na ação vascular de substâncias:
achados bibliográficos dos últimos 5 anos.
Seguindo os critérios de levantamento de artigos para
serem inclusos no presente trabalho, da pesquisa com as
palavras-chave nas bases Web of Science e Pubmed entre
julho e agosto de 2020, um total de 263 artigos foram
encontrados e adicionados à plataforma Ryyan. Após a seleção
de artigos pelo critério de termos-chave no título e no resumo,
exclusão de artigos repetidos e de anos anterior a 2016, um

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
211
total de 41 artigos foram selecionados para serem lidos
integralmente. Por fim, após a realização da leitura, um total de
14 artigos foram incluidos (figura 1), os quais abordavam a
participação do PVAT na ação de substâncias bioativas e
extratos como agentes vasoprotetores em modelos de doenças
cardiovasculares em animais.
Os artigos selecionados para serem integrados a esse trabalho
foram listados na tabela 1 a qual detalha a referência, modelo
animal usado, substâcia testada e principais achados.
Em sua totalidade, os roedores, ratos e camudongos, são
os animais mais utilizados. Além disso, modelo de obesidade
foram os mais corriqueiros, tendo em vista que esse parâmetro,
em humanos, está associado a diversas alterações metabólicas
e clínica.

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
212
Figura 1. Método de busca nas bases de dados e inclusão dos
artigos pertinentes à esse capítulo de forma sistemática.
Fonte: Autor, 2020.
Os trabalhos corroboram com uma das teses para a gênese
das alterações e doenças que afetam os vasos sanguíneos,
sendo ela a de que as alteraçõs fisiológicas da região
perivascular e adventícia (como a inflamação no PVAT,
aumento do estresse oxidativo, alteração do tipo de adipócitos
e de mediadores parácrinos que esses liberam) desencadeiam
as alterações fisiológicas danosas no músculo liso e endotélio
vascular.
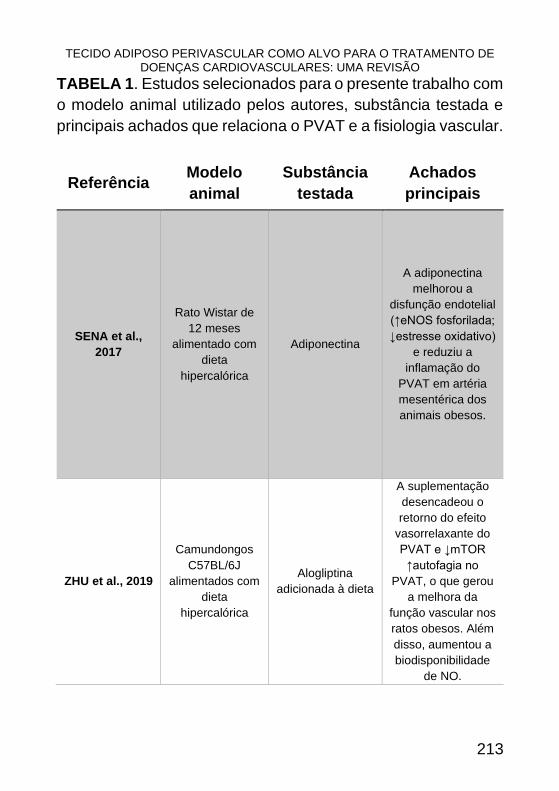
TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
213
TABELA 1. Estudos selecionados para o presente trabalho com
o modelo animal utilizado pelos autores, substância testada e
principais achados que relaciona o PVAT e a fisiologia vascular.
Referência Modelo
animal
Substância
testada
Achados
principais
SENA et al.,
2017
Rato Wistar de
12 meses
alimentado com
dieta
hipercalórica
Adiponectina
A adiponectina
melhorou a
disfunção endotelial
(↑eNOS fosforilada;
↓estresse oxidativo)
e reduziu a
inflamação do
PVAT em artéria
mesentérica dos
animais obesos.
ZHU et al., 2019
Camundongos
C57BL/6J
alimentados com
dieta
hipercalórica
Alogliptina
adicionada à dieta
A suplementação
desencadeou o
retorno do efeito
vasorrelaxante do
PVAT e ↓mTOR
↑autofagia no
PVAT, o que gerou
a melhora da
função vascular nos
ratos obesos. Além
disso, aumentou a
biodisponibilidade
de NO.
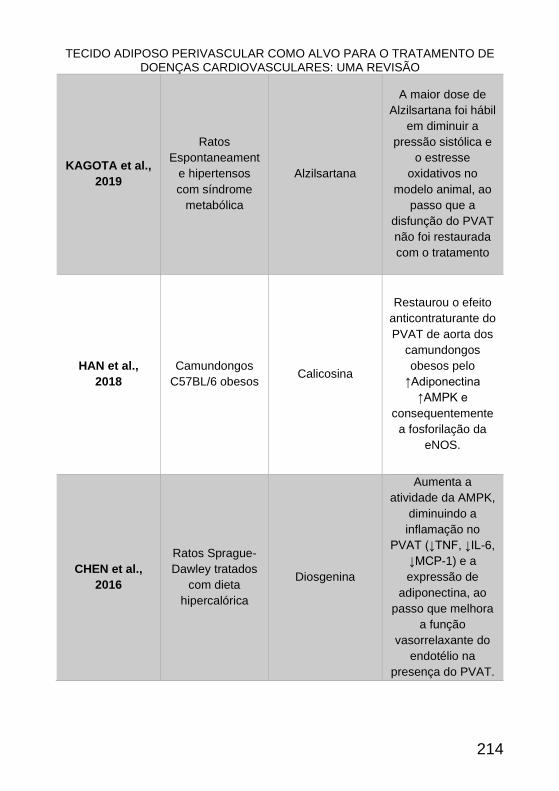
TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
214
KAGOTA et al.,
2019
Ratos
Espontaneament
e hipertensos
com síndrome
metabólica
Alzilsartana
A maior dose de
Alzilsartana foi hábil
em diminuir a
pressão sistólica e
o estresse
oxidativos no
modelo animal, ao
passo que a
disfunção do PVAT
não foi restaurada
com o tratamento
HAN et al.,
2018
Camundongos
C57BL/6 obesos Calicosina
Restaurou o efeito
anticontraturante do
PVAT de aorta dos
camundongos
obesos pelo
↑Adiponectina
↑AMPK e
consequentemente
a fosforilação da
eNOS.
CHEN et al.,
2016
Ratos Sprague-
Dawley tratados
com dieta
hipercalórica
Diosgenina
Aumenta a
atividade da AMPK,
diminuindo a
inflamação no
PVAT (↓TNF, ↓IL-6,
↓MCP-1) e a
expressão de
adiponectina, ao
passo que melhora
a função
vasorrelaxante do
endotélio na
presença do PVAT.
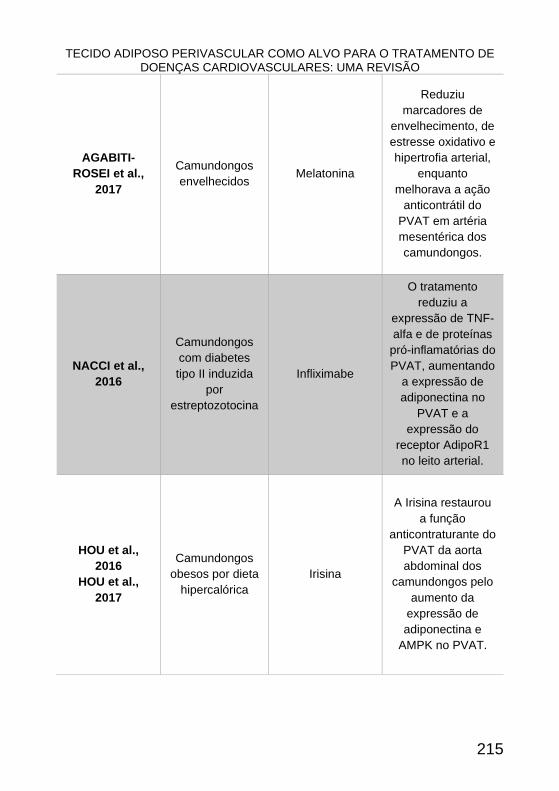
TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
215
AGABITI-
ROSEI et al.,
2017
Camundongos
envelhecidos Melatonina
Reduziu
marcadores de
envelhecimento, de
estresse oxidativo e
hipertrofia arterial,
enquanto
melhorava a ação
anticontrátil do
PVAT em artéria
mesentérica dos
camundongos.
NACCI et al.,
2016
Camundongos
com diabetes
tipo II induzida
por
estreptozotocina
Infliximabe
O tratamento
reduziu a
expressão de TNF-
alfa e de proteínas
pró-inflamatórias do
PVAT, aumentando
a expressão de
adiponectina no
PVAT e a
expressão do
receptor AdipoR1
no leito arterial.
HOU et al.,
2016
HOU et al.,
2017
Camundongos
obesos por dieta
hipercalórica
Irisina
A Irisina restaurou
a função
anticontraturante do
PVAT da aorta
abdominal dos
camundongos pelo
aumento da
expressão de
adiponectina e
AMPK no PVAT.
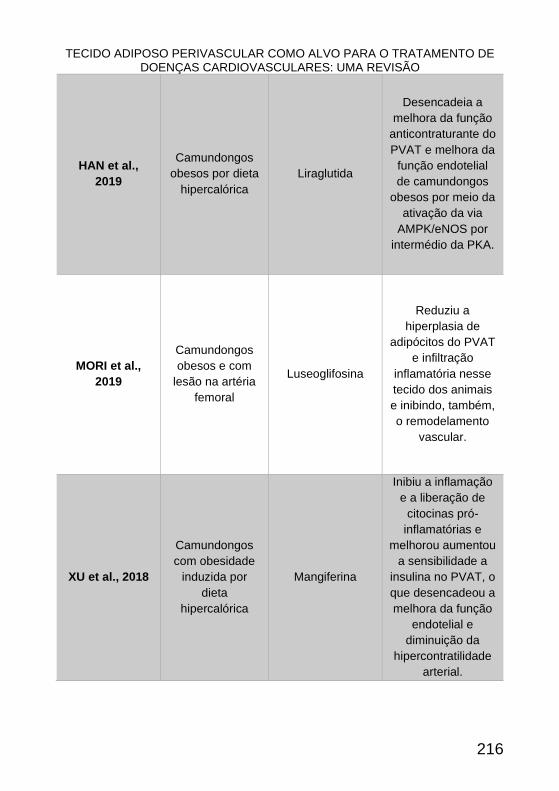
TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
216
HAN et al.,
2019
Camundongos
obesos por dieta
hipercalórica
Liraglutida
Desencadeia a
melhora da função
anticontraturante do
PVAT e melhora da
função endotelial
de camundongos
obesos por meio da
ativação da via
AMPK/eNOS por
intermédio da PKA.
MORI et al.,
2019
Camundongos
obesos e com
lesão na artéria
femoral
Luseoglifosina
Reduziu a
hiperplasia de
adipócitos do PVAT
e infiltração
inflamatória nesse
tecido dos animais
e inibindo, também,
o remodelamento
vascular.
XU et al., 2018
Camundongos
com obesidade
induzida por
dieta
hipercalórica
Mangiferina
Inibiu a inflamação
e a liberação de
citocinas pró-
inflamatórias e
melhorou aumentou
a sensibilidade a
insulina no PVAT, o
que desencadeou a
melhora da função
endotelial e
diminuição da
hipercontratilidade
arterial.
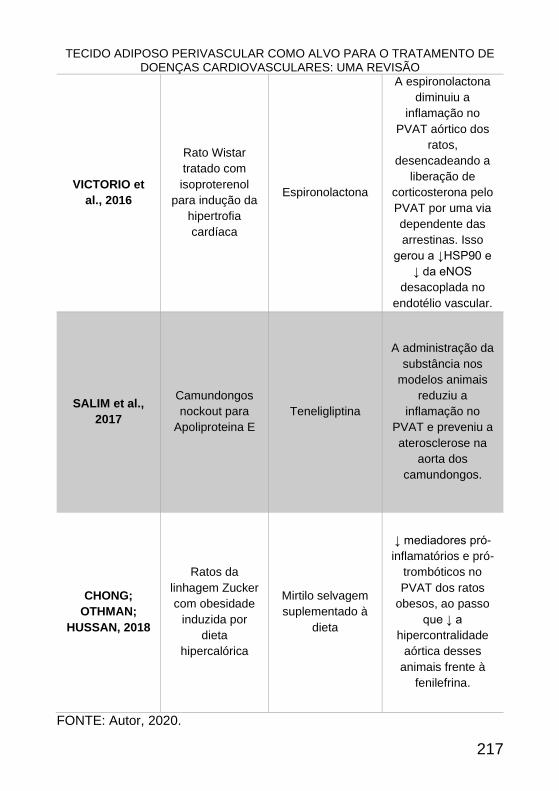
TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
217
VICTORIO et
al., 2016
Rato Wistar
tratado com
isoproterenol
para indução da
hipertrofia
cardíaca
Espironolactona
A espironolactona
diminuiu a
inflamação no
PVAT aórtico dos
ratos,
desencadeando a
liberação de
corticosterona pelo
PVAT por uma via
dependente das
arrestinas. Isso
gerou a ↓HSP90 e
↓ da eNOS
desacoplada no
endotélio vascular.
SALIM et al.,
2017
Camundongos
nockout para
Apoliproteina E
Teneligliptina
A administração da
substância nos
modelos animais
reduziu a
inflamação no
PVAT e preveniu a
aterosclerose na
aorta dos
camundongos.
CHONG;
OTHMAN;
HUSSAN, 2018
Ratos da
linhagem Zucker
com obesidade
induzida por
dieta
hipercalórica
Mirtilo selvagem
suplementado à
dieta
↓ mediadores pró-
inflamatórios e pró-
trombóticos no
PVAT dos ratos
obesos, ao passo
que ↓ a
hipercontralidade
aórtica desses
animais frente à
fenilefrina.
FONTE: Autor, 2020.

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
218
Foi observado que em todos os estudos utilizou-se
roedores para as substâncias serem testadas, sendo induzido
nesses ratos e camudongos com modelos de obesidade,
síndrome metabólica, hipertensão e diabetes. Essas doenças
estão intimamente relacionadas com a disfunção endotelial,
com alterações danosas no músculo liso vascular e também
com a perda da função vasculoprotetor do PVAT (SAXTON et
al., 2019).
Além disso, as mudanças nos hábitos da população –
principalmente a ocidental – tem desencadeado uma verdadeira
pandemia de tais doenças (PETRACCI; GABBIANELLI;
BORDONI, 2020).
Assim, nos artigos encontrados nas base de dados foi
observado que tanto substâncias candidatas a fármacos,
fármacos já usados na clínica, adipocinas e extratos vegetais
atuaram na melhoria da função vascular e melhoramento do
quadro patológico dos modelos animais por meio da mudança
de fenótipo e do tipo de adipocininas, citocinas e autacoides
secretados pelo PVAT, bem como na diminuição do quadro
inflamatório e do estresse oxidativo desse tecido.
Algumas substâncias usadas nos estudos relatados na
tabela 1 já são usados corriqueiramente na prática clínica no
dia a dia e isso aponta, de certa forma, efeitos pleiotrópicos
desses fármacos, os quais são intermediados pelo PVAT.
Outrossim, é importante salientar que alguns mecanismos
de ação desses fármacos e substâncias encontrados possuem
alguns pontos em comum, como a calicosina e adiosgenina que
possuem como ponto em comum de ação o aumento da
expressão e atividade da AMPK, o que está relacionado com o
aumento da biodisponibilidade de NO para o músculo liso

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
219
vascular e, assim, relaxamento do leito arterial. Também, foi
visto que a maioria das substâncias aumentaram a expressão
de adiponectina.
A adiponectina está diretamente relacionada, conforme
relatos da literatura, com a melhoria e manutenção da boa
função cardiovascular. Foi observado que na maioria dos
estudos, as substâncias testadas melhoravam a
biodisponibilidade ou a expressão no PVAT de adiponectina, o
que pode está relacionado, também, com a melhora da função
vascular, da obesidade, hipertensão, diabetes e síndome
metabólica nos animais dos estudos suprarrelatados.
CONCLUSÕES
Dessa maneira, pode-se concluir que o tecido adiposo
perivascular pode atuar como um mediador nas ações
benéficas de algumas substâncias testadas e que a reversão
do dano no PVAT e o resgate do seu estado fisiológico antes
da instalação das comorbidades é um importante ponto de
estudo para o desenvolvimento de novos medicamentos para o
tratamento da síndrome metabólica.
REFERÊNCIAS BIBLIOGRÁFICAS AGABITI-ROSEI, Claudia et al. Effect of long-term treatment with melatonin on vascular markers of oxidative stress/inflammation and on the anticontractile activity of perivascular fat in aging mice. Hypertension Research, v. 40, n. 1, p. 41-50, 2017. AGABITI-ROSEI, Claudia et al. Modulation of vascular reactivity by perivascular adipose tissue (PVAT). Current hypertension reports, v. 20, n. 5, p. 44, 2018. ALMABROUK, Tarek AM et al. High fat diet attenuates the anticontractile activity of aortic PVAT via a mechanism involving AMPK and reduced adiponectin secretion. Frontiers in physiology, v. 9, p. 51, 2018.

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
220
AWATA, Wanessa MC et al. Perivascular adipose tissue contributes to lethal sepsis-induced vasoplegia in rats. European Journal of Pharmacology, v. 863, p. 172706, 2019. AZUL, Lara et al. Increased inflammation, oxidative stress and a reduction in antioxidant defense enzymes in perivascular adipose tissue contribute to vascular dysfunction in type 2 diabetes. Free Radical Biology and Medicine, v. 146, p. 264-274, 2020. BAYLIE, Rachael et al. Lack of direct effect of adiponectin on vascular smooth muscle cell BKCa channels or Ca2+ signaling in the regulation of small artery pressure‐induced constriction. Physiological Reports, v. 5, n. 16, p. e13337, 2017. CACANYIOVA, S. et al. The role of perivascular adipose tissue and endogenous hydrogen sulfide in vasoactive responses of isolated mesenteric arteries in normotensive and spontaneously hypertensive rats. J. Physiol. Pharmacol, v. 70, p. 295-306, 2019. CHEN, Yan et al. Diosgenin regulates adipokine expression in perivascular adipose tissue and ameliorates endothelial dysfunction via regulation of AMPK. The Journal of Steroid Biochemistry and Molecular Biology, v. 155, p. 155-165, 2016. CHONG, C. L. G.; OTHMAN, Faizah; HUSSAN, Farida. Vascular protective effects of Morinda citrifolia leaf extract on postmenopausal rats fed with thermoxidized palm oil diet: evidence at microscopic level. International journal of vascular medicine, v. 2018, 2018. HAN, Fang et al. Calycosin directly improves perivascular adipose tissue dysfunction by upregulating the adiponectin/AMPK/eNOS pathway in obese mice. Food & function, v. 9, n. 4, p. 2409-2415, 2018. HAN, Fang et al. Liraglutide improves vascular dysfunction by regulating a cAMP-independent PKA-AMPK pathway in perivascular adipose tissue in obese mice. Biomedicine & Pharmacotherapy, v. 120, p. 109537, 2019. HOU, Ningning et al. Irisin improves perivascular adipose tissue dysfunction via regulation of the heme oxygenase-1/adiponectin axis in diet-induced obese mice. Journal of molecular and cellular cardiology, v. 99, p. 188-196, 2016. KAGOTA, Satomi et al. Angiotensin II Type 1 Receptor Antagonist Azilsartan Restores Vascular Reactivity Through a Perivascular Adipose Tissue-Independent Mechanism in Rats with Metabolic Syndrome. Cardiovascular Drugs and Therapy, v. 33, n. 5, p. 501-509, 2019. LIM, Kangil; KIM, Kijeong. Role of Exercise-induced Adiponectin Activation on Obese and Diabetic Individuals. Exercise Science, v. 29, n. 3, p. 208-213, 2020. LÖHN, MATTHIAS et al. Periadventitial fat releases a vascular relaxing factor. The FASEB Journal, v. 16, n. 9, p. 1057-1063, 2002.

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
221
MORI, Yusaku et al. Luseogliflozin attenuates neointimal hyperplasia after wire injury in high-fat diet-fed mice via inhibition of perivascular adipose tissue remodeling. Cardiovascular diabetology, v. 18, n. 1, p. 1-9, 2019. NACCI, Carmela et al. Infliximab therapy restores adiponectin expression in perivascular adipose tissue and improves endothelial nitric oxide-mediated vasodilation in mice with type 1 diabetes. Vascular pharmacology, v. 87, p. 83-91, 2016. NOSALSKI, Ryszard; GUZIK, Tomasz J. Perivascular adipose tissue inflammation in vascular disease. British journal of pharmacology, v. 174, n. 20, p. 3496-3513, 2017. PACE, Fernanda et al. Helminth infection in mice improves insulin sensitivity via modulation of gut microbiota and fatty acid metabolism. Pharmacological research, v. 132, p. 33-46, 2018. PETRACCI, Irene; GABBIANELLI, Rosita; BORDONI, Laura. The Role of Nutri (epi) genomics in Achieving the Body’s Full Potential in Physical Activity. Antioxidants, v. 9, n. 6, p. 498, 2020. PHAM, Duc-Vinh; PARK, Pil-Hoon. Recent insights on modulation of inflammasomes by adipokines: a critical event for the pathogenesis of obesity and metabolism-associated diseases. Archives of Pharmacal Research, p. 1-20, 2020. QUEIROZ, Marcelo; SENA, Cristina M. Perivascular adipose tissue in age-related vascular disease. Ageing Research Reviews, p. 101040, 2020. SALIM, Hotimah Masdan et al. Teneligliptin, a dipeptidyl peptidase-4 inhibitor, attenuated pro-inflammatory phenotype of perivascular adipose tissue and inhibited atherogenesis in normoglycemic apolipoprotein-E-deficient mice. Vascular Pharmacology, v. 96, p. 19-25, 2017. SAMUEL, Olapoju O. Review on multifaceted involvement of perivascular adipose tissue in vascular pathology. Cardiovascular Pathology: the Official Journal of the Society for Cardiovascular Pathology, v. 49, p. 107259-107259, 2020. SAXTON, Sophie N. et al. Mechanistic links between obesity, diabetes, and blood pressure: role of perivascular adipose tissue. Physiological reviews, v. 99, n. 4, p. 1701-1763, 2019. SAXTON, Sophie N.; WITHERS, Sarah B.; HEAGERTY, Anthony M. Emerging roles of sympathetic nerves and inflammation in perivascular adipose tissue. Cardiovascular Drugs and Therapy, v. 33, n. 2, p. 245-259, 2019. SENA, Cristina M. et al. Adiponectin improves endothelial function in mesenteric arteries of rats fed a high‐fat diet: role of perivascular adipose tissue. British journal of pharmacology, v. 174, n. 20, p. 3514-3526, 2017.

TECIDO ADIPOSO PERIVASCULAR COMO ALVO PARA O TRATAMENTO DE DOENÇAS CARDIOVASCULARES: UMA REVISÃO
222
SIEGEL-AXEL, D. I.; HÄRING, H. U. Perivascular adipose tissue: an unique fat compartment relevant for the cardiometabolic syndrome. Reviews in Endocrine and Metabolic Disorders, v. 17, n. 1, p. 51-60, 2016. SOLTIS, Edward E.; CASSIS, Lisa A. Influence of perivascular adipose tissue on rat aortic smooth muscle responsiveness. Clinical and Experimental Hypertension. Part A: Theory and Practice, v. 13, n. 2, p. 277-296, 1991. SPYCHALSKA-ZWOLIŃSKA, Marta et al. Blood adipocytokine concentration in patients with peripheral artery disease. International angiology: a journal of the International Union of Angiology, 2020. VICTORIO, Jamaira A. et al. Spironolactone prevents endothelial nitric oxide synthase uncoupling and vascular dysfunction induced by β-adrenergic overstimulation: role of perivascular adipose tissue. Hypertension, v. 68, n. 3, p. 726-735, 2016. XU, Xiaoshan et al. Mangiferin suppresses endoplasmic reticulum stress in perivascular adipose tissue and prevents insulin resistance in the endothelium. European journal of nutrition, v. 57, n. 4, p. 1563-1575, 2018. ZEMANČÍKOVÁ, Anna; TÖRÖK, Jozef. Influence of Age on Anticontractile Effect of Perivascular Adipose Tissue in Normotensive and Hypertensive Rats. Oxidative medicine and cellular longevity, v. 2019, 2019. ZHU, Biao et al. Alogliptin improves endothelial function by promoting autophagy in perivascular adipose tissue of obese mice through a GLP-1-dependent mechanism. Vascular pharmacology, v. 115, p. 55-63, 2019. PICHÉ, Marie-Eve; MORIN, Marie-Philippe; POIRIER, Paul. Obesity and Coronary Artery Disease: Evaluation and Treatment. In: Biochemistry of Cardiovascular Dysfunction in Obesity. Springer, Cham, 2020. p. 217-233.
AGRADECIMENTOS Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e a Universidade Federal da Paraíba (UFPB) pelo fomento e apoio.

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
223
CAPÍTULO 11
TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA
CANALOPATIAS NEUROLÓGICAS
Micheline Freire DONATO 1,2
Pedro Henrique Cardoso de CASTRO 3
Samuel José DORNELAS 4
Alessandra MATAVEL5
1 Professora Visitante da PGPNSB- DCF/UFPB; 2 Orientadora/Professora da PGPNSB-
DCF/UFPB; 3 Mestrando em Farmacologia/UFMG; 4 Graduando em Ciências
Biológicas/UFMG; 5 Pesquisadora, Fundação Ezequiel Dias/MG.
[email protected] ; [email protected]
RESUMO: Os canais iônicos operados por voltagem (VOCs) são glicoproteínas transmembrana responsáveis por inúmeros processos fisiológicos, incluindo transmissão do impulso nervoso, contração muscular e liberação de neurotransmissores e hormônios. Canais para sódio dependentes de voltagem (Nav) participam na manutenção da homeostase do sistema nervoso central e periférico, e são os responsáveis pelo impulso elétrico rápido. Doenças monogenéticas, frequentemente causadas por mutações nestes canais, causam disfunções, chamadas canalopatias, como a síndrome do QT-longo, Dravet, dor, epilepsia, enxaqueca, dentre outras. Desta forma, os Navs são alvos terapêuticos para o tratamento de várias destas doenças. A busca por substâncias com ação moduladora nestes canais é uma alternativa promissora no desenvolvimento de terapias mais eficientes e específicas. Dentre os moduladores dos Navs,

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
224
as neurotoxinas foram selecionadas ao longo da evolução para serem específicas contra regiões dos VOCs importantes para sua função, e que, portanto, não podem ser alteradas. Nesse sentido, essas toxinas têm mostrado grande potencial biotecnológico, direcionando novas abordagens farmacológicas que podem melhorar a qualidade de vida e prognóstico de pacientes portadores destas canalopatias. O objetivo desse capítulo foi revisar os dados da literatura em relação à contribuição dos Navs para fenótipos de epilepsia e dor, e apresentar os sítios de ligação das neurotoxinas sobre os Navs. Palavras-chave: Neurotoxinas de animais. Canais iônicos. Canais Nav. Epilepsia. Dor.
INTRODUÇÃO
Os canais iônicos operados por voltagem (VOCs)
pertencem à superfamília das glicoproteínas da membrana
celular e participam da homeostase iônica em diferentes tipos
celulares. Eles possuem papel fundamental na propagação do
potencial de ação neuronal, contração muscular, promovem a
secreção de neurotransmissor nas sinapses, dentre outras
funções. Os VOCs foram encontrados em todos os táxons:
vírus, procariotas e eucariotas; sugerindo um ancestral quase
tão antigo quanto à própria Terra (MORAN et al., 2015). Eles
respondem às variações rápidas na voltagem através da
membrana, permitindo a passagem seletiva de íons, o que
diferencia os canais para potássio (Kv), cálcio (Cav) e sódio
(Nav) (VAN THEEMSCHE et al., 2020).
Os Kvs estão presentes em todas as células e participam
do potencial de repouso e da repolarização. Os Cavs, além de
participarem da contração muscular, são importantes segundos

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
225
mensageiros, acionando muitas cascatas de sinalização
intracelular. Já os Navs exercem papel essencial na geração e
propagação do potencial de ação (PA) rápido em neurônios e
em outras células eletricamente excitáveis, como miócitos e
células endócrinas.
Os canais iônicos possuem pelo menos dois estados
conformacionais, aberto e fechado, sendo que o estado fechado
pode ser ainda distinto em inativado e desativado, mas
geralmente há diferentes subestados destes (AHERN et al.,
2016). As propriedades destes estados são chamadas de
propriedades de gating e são dependentes de voltagem em
maior ou menor grau, de acordo com o canal. O estado condutor
é o “aberto”, que geralmente é um estado transitório. Nos Navs,
esse estado permanece somente alguns milissegundos,
transitando muito rapidamente para o estado “inativado”.
Estruturalmente, os VOCs são constituídos por uma
subunidade principal α, que forma o canal. Esta subunidade
pode ser modulada pelas subunidades acessórias 𝛃, 𝛄 e/ou 𝛅,
que regulam as propriedades de gating, tráfico e expressão na
superfície da célula (O’LEARY & CHAHINE, 2018). A
subunidade 𝛂 do Kv é constituído por 6 segmentos
transmembrana (S1-S6) em forma de 𝛂 -hélice, separados por
alças intra e extracelulares, sendo os segmentos N- e C-
terminal voltados para o interior da célula (fig. 1).
Para formar um canal funcional os Kv necessitam de 4
subunidades, que podem ser iguais (formando um
homotetrâmero) ou diferentes (formando um heterotetrâmero)
(KIM & NIMIGEAN, 2016). Acredita-se que os Kvs foram os
primeiros VOCs que aparecerem no curso da evolução, que por

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
226
meio de dois ciclos de duplicação gênica e divergência originou
os Cav e Nav (Moran et al., 2015). Desta forma, os Cavs e Navs
possuem 4 domínios (D1-D4), homólogos às 4 subunidades dos
Kvs, no entanto, uma subunidade 𝛂 sozinha é capaz de formar
o canal funcional (VAN THEEMSCHE et al., 2020).
Figura 1. Estrutura geral da subunidade 𝛂 dos VOCs. Os Kvs
são constituídos por 6 segmentos transmembrana (ST) em 𝛂 -
hélice (S1-S6). São necessárias 4 subunidades para formar um
canal funcional. Os Navs e Cavs possuem 4 domínios (D1-D4),
possuindo 6 ST cada um.
Fonte: Autores, 2020.
O segmento S4 de cada subunidade ou domínio possui
uma carga positiva a cada 3-4 aminoácidos, sendo denominado
sensor de voltagem (KIM & NIMIGEAN, 2016; AHERN et al.,
2016). Por estar no campo elétrico da membrana, essas cargas
se movem de acordo com as mudanças na voltagem, e esse
movimento está acoplado à ativação do canal (VAN
THEEMSCHE et al., 2020). Estudos estruturais de cristalografia
por raio-X de alguns desses canais demonstraram que alças
ascendente e descendente (M1 e M2) entre os segmentos S5-
S6 de cada subunidade ou domínio, se invaginam na

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
227
membrana formando as paredes do poro, enquanto a extensão
do poro é inclinada, com arranjo de uma “tenda indígena”
invertida. Essa estrutura sugere que o poro é estreito na região
intracelular e funciona como um filtro de seletividade iônica
específica para cada íon (CLAIRFEUILLE et al., 2019). Para o
Cav a assinatura do filtro de seletividade são os aminoácidos
EEEE (4 glutamatos) presentes um em cada alça do poro de
cada domínio (ABDEREMANE-Ali et al., 2019). Da mesma
forma, os resíduos DEKA (aspartato, glutamato, lisina e alanina)
formam o filtro de seletividade dos Navs (AHERN et al., 2016).
Para o diverso grupo dos Kvs, a assinatura varia um pouco
sendo conservado o motivo GYG (glicina, tirosina, glicina) (KIM
& NIMIGEAN, 2016) que, nesse canal se encontra em
sequência na cadeia primária, se repetindo em cada
subunidade.
O estado inativado dos Navs, um estado não condutor e
não responsivo (refratário) do qual ele se recupera somente
após a repolarização, ocorre graças ao motivo IFM, constituído
por 3 aminoácidos (isoleucina, fenilalanina e metionina) na alça
intracelular entre os segmentos D3-S6 e D4-S1, e que se
movem após o movimento do sensor de voltagem D4-S4,
acoplando processo de ativação e inativação do canal (AHERN
et al., 2016).
Foram identificadas nove isoformas para a subunidade α para
os Navs em mamíferos (Nav1.1-Nav1.9) distribuídos em
diferentes tecidos, com propriedades biofísicas e sensibilidade
distinta à neurotoxina tetrodotoxina (TTX), que foi a primeira
toxina utilizada para distinguir esses canais e suas isoformas
(tabela 1) (DEUIS et al., 2017).

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
228
Devido a sua importância em muitos aspectos da
regulação celular e transdução de sinais, mutações nos VOCs
podem levar às disfunções fisiológicas envolvidas em
patologias, denominadas canalopatias (NATARAJAN &
KRISHNA, 2019). Além disso, eles são alvos moleculares de
uma extensa gama de neurotoxinas animais (NTX). As NTX que
modificam os VOCs estão divididas em duas categorias:
bloqueadoras do poro e modificadoras de gating (SHEN et al.,
2018).
Tabela 1. Subunidades α dos Navs de mamíferos.
Tipo
Gene Tecidos primários de
expressão
Nav1.1 SCN1A SNC
SNP
Nav1.2 SCN2A SNC
Nav1.3 SCN3A SNC
Nav1.4 SCN4A Músculo esquelético
Nav1.5 SCN5A Músculo cardíaco
Nav1.6 SCN8A SNC
SNP
Nav1.7 SCN9A SNP
Células de Schwann
Nav1.8 SCN10 Gânglio da raiz dorsal (DRG)
Nav1.9 SCN11 SNP

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
229
SNC: Sistema Nervoso Central; SNP: Sistema Nervoso Periférico.
Fonte: Adaptado de DEUIS et al., 2017.
Enquanto as NTX bloqueadoras ocluem fisicamente o
poro por onde os íons atravessam; as modificadoras de gating
podem alterar a cinética e/ou a dependência de voltagem da
ativação e da inativação desses canais. Por serem específicas,
NTX têm sido utilizadas para compreender a estrutura-função
das diferentes partes dos VOCs, identificar as isoformas dos
canais e auxiliar na terapia e desenvolvimento de novos
fármacos.
Neste trabalho, revisamos duas canalopatias dos Navs
no sistema nervoso central (SNC): epilepsia e dor. Além disso,
discorremos sobre os sítios de ligação das NTX nos Navs e seu
potencial biotecnológico.
MATERIAIS E MÉTODO
Trata-se de uma revisão da literatura científica
preferencialmente dos últimos cinco anos (2015 a 2020) uma
vez que revisões recentes são escassas sobre o tema em
questão.
Foram realizadas buscas em plataformas acadêmicas e
banco de dados de artigos científicos, tais como o Scielo,
Pubmed, Science Direct, Google Acadêmico através de
descritores ou palavras-chave em português e inglês, como
canais iônicos, canal para sódio dependente de voltagem, Nav,
epilepsia, dor, toxinas animais, neurotoxinas; ion channels,
voltage gated sodium channel, toxin, epilepsy, pain,
respectivamente.

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
230
RESULTADOS E DISCUSSÃO
Canalopatias associadas aos Navs
Epilepsia
A epilepsia é marcada por crises de ocorrência
transitória, focais ou generalizadas, devido atividade neuronal
anormal e excessiva no cérebro. Dependendo de quais áreas
do cérebro estão envolvidas, as crises epilépticas podem
consistir em perda de consciência com tremores, confusão
mental, movimento incontrolável dos membros e sintomas
sensoriais diversos (JOHNSON, 2019).
Compreender e investigar a etiologia da epilepsia
permite maior abrangência na conduta clínica e direcionamento
de novas terapias capazes de melhorar o prognóstico de
pacientes portadores desta doença. Embora em três quartos
dos pacientes as crises sejam idiopáticas (causas
desconhecidas), os estudos de sequenciamentos genéticos
têm evidenciado diversas mutações responsáveis pelo quadro
epiléptico, dentre elas, mutações na subunidade α do Nav1.1
(MENEZES et al., 2020). Além deste, outros Navs estão
relacionados à epilepsia (MENEZES et al., 2020), como se
segue.
Nav1.1
Mais de 1.800 mutações já foram identificadas no gene
SCN1A que codifica a proteína do canal Nav1.1 (The Human
Gene Mutation Database, 2020), sendo esse alvo o mais

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
231
comum dentre as mutações que causam epilepsia. Os Nav1.1
são expressos em neurônios inibitórios gabaérgicos, assim, a
perda de função deste canal nessas células reduz os impulsos
inibitórios, predispondo o SNC às descargas excessivas,
características da condição epileptogênica. Mutações neste
canal podem predispor o surgimento de manifestações que vão
de epilepsia generalizada com convulsão febril plus (EGCF+),
que podem persistir até a vida adulta, à síndrome de Dravet
(SD), uma encefalopatia epiléptica grave e farmacorresistente,
também caracterizada por convulsões gatilhadas por
hipertermia na infância, mas que não persistem na vida adulta,
com déficits cognitivos, comportamentais e ataxia. No caso da
EGCF+, a mutação é sítio dirigida, com consequências
funcionais imprevisíveis, dependendo do aminoácido mutado.
Em estudo feito por Mantegazza e colaboradores (2019), o
gene SCN1A foi sequenciamento em pacientes com SD e
revelou sete diferentes mutações, seis nonsense, que
acarretaram em um canal não-funcional, e uma missense
(L986F) (HUFF et al., 2020; MANTEGAZZA, et al., 2019). Em
alguns casos é possível rastrear a hereditariedade da mutação,
revelando histórico familiar em pacientes com SD e/ou EGCF+.
Mutações no SCN1A são encontradas em cerca de 80% dos
pacientes portadores de SD e em apenas 10% dos pacientes
portadores de EGCF+ (MEI et al., 2019).

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
232
Nav1.2
Codificado pelo gene SCN2A, o canal Nav1.2 é expresso
no SNC em neurônios excitatórios glutamatérgicos (SANDERS
et al., 2018). Durante o desenvolvimento inicial, Nav1.2 é
abundantemente expresso nos nódulos de Ranvier e no
segmento inicial do axônio (SIA), participando diretamente da
excitabilidade neuronal e início do potencial de ação (PA).
Nav1.2 é substituído por Nav1.6, e a expressão de Nav1.2 fica
restrita a região distal do SIA, participando do prolongamento
do PA em vez de iniciá-lo em neurônios maduros. São
conhecidas mais de 100 mutações neste gene, que causam
sintomas diversos como encefalopatias epilépticas e de
desenvolvimento, convulsões familiares neonatais benignas,
ataxia episódica, transtornos de espectro autista e deficiência
intelectual com e sem convulsões (REYNOLDS et al., 2020). As
mutações de ganho de função estão relacionadas à epilepsia,
uma vez que causam hiperexcitabilidade neuronal, e as
mutações de perda de função estão relacionadas ao autismo e
ao fenótipo de deficiência intelectual (BEN-SHALOM et al.,
2017).
Nav1.3
O Nav1.3, codificado pelo gene SCN3A, é expresso no
SNC apenas durante o desenvolvimento embrionário e
neonatal, sendo quase indetectável em indivíduos após este
período. Durante a infância estes canais são trocados pelos
Nav1.1 para manter as funções fisiológicas do corpo (ZAMAN et

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
233
al., 2018). No entanto, estudos têm demonstrado que em
tecidos que sofreram algum tipo de lesão causando dor
neuropática, há um aumento da expressão dos Nav1.3,
demonstrando sua importância nas neuropatias (MENEZES et
al., 2020). Variantes de SCN3A foram associadas à
encefalopatia epiléptica infantil precoce, que confere ganho de
função ao canal, e geralmente é responsivo ao tratamento com
fármacos bloqueadores de canal para sódio como o topiramato
de sódio (BRUNKLAUS et al., 2020).
Nav1.6
O Nav1.6 é codificado pelo gene SCN8A e expresso no
SNC e SNP. Mutações neste canal conferem fenótipos de
epilepsia que vão desde convulsões infantis benignas e
discinesias paroxísticas à encefalopatía epiléptica grave.
Geralmente essas crises epilépticas se iniciam no primeiro dia
de vida culminando entre o 4o e 5o mês com crises focais,
tônicas, clônicas, mioclônicas, crises de ausência e/ou
espasmos epilépticos. Mutações nesse gene podem gerar
problemas relacionados ao desenvolvimento cognitivo
(LARSEN et al., 2015). Em pacientes epilépticos estas
mutações conferem ganho de função para o canal, às vezes
responsivo a altas doses de bloqueadores (BRUNKLAUS et al.,
2020). Mutações no SCN8A são descritas como a causa mais
comum de morte repentina e inesperada em pacientes com
epilepsia. Uma possível correlação entre morte súbita de
pacientes epilépticos e mutações no SCN8A é a presença, em
baixos níveis, de Nav1.6 nos túbulos transversos dos miócitos

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
234
ventriculares, regulando o acoplamento excitação-contração.
Disfunções nos Nav1.6 no tecido cardíaco podem causar
alterações na função cardíaca levando à parada
cardiorrespiratória (JOHANNESEN et al., 2018).
Nav1.7
SCN9A é o gene que codifica a proteína Nav1.7,
preferencialmente expresso no SNP, mas também presente no
SNC. Yang e colaboradores (2018) observaram duas mutações
em SCN9A com manifestações epilépticas, demonstrando sua
associação às manifestações clínicas com um grande espectro
de fenótipos de convulsão, incluindo crises febris e não febris.
Pacientes com mutações de deleção (nonsense) tendem a
estar associados a tipos de crises mais graves do que mutações
missense. Sua relação com epilepsia, embora embasada por
estudos recentes ainda não foi completamente confirmada
(BRUNKLAUS et al., 2020; MENEZES et al., 2020).
Dor
A capacidade de um organismo de sentir dor se
desenvolveu ao longo da evolução das espécies como uma
forma de alerta, que tem como objetivo minimizar danos aos
tecidos e células, na tentativa de prolongar a sobrevivência
(COX et al., 2006).
Cox e colaboradores (2006) descreveram famílias
paquistanesas incapazes que sentir dor. Análise genômica
identificou mutações do gene SCN9A, que codifica o canal
Nav1.7 comumente encontrado no DRG e em neurônios

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
235
ganglionares simpáticos, como responsável pela condição.
Esta canalopatia congênita acarreta perda de função do canal.
De forma inversa, a hiperativação dos canais Nav1.7
pode levar aos distúrbios de hiperalgesia. A dor neuropática
causada por doenças como eritromelalgia hereditária,
distúrbios de dor extrema paroxística, e neuropatia fibrótica são
consequências de uma mutação de ganho de função dos canais
Nav1.7 (ZHANG et al., 2019), que diminui o limiar de ativação
destes canais tornando-os mais susceptíveis à despolarização.
Os indivíduos acometidos por essa mutação genética sentem
dores desproporcionais às lesões teciduais. Nesse caso, a dor
não desempenha uma função fisiológica de proteção e passa
ser uma condição patológica.
O Nav1.8 é codificado pelo gene SCN10A e expresso,
preferencialmente no DRG, em neurônios do nervo trigêmio e
em terminais nervosos livres na pele. Suas propriedades
biofísicas, e sua presença nas terminações nervosas sensitivas
sugerem uma importante relação com a percepção da dor
neuropática e inflamatória. Estudos recentes demonstram
associação desse canal aos efeitos na condução cardíaca.
Suas mutações de ganho de função são raras e pouco
relatadas na literatura (DUAN, 2018).
Por sua vez, o Nav1.9 é codificado pelo gene SCN11A e
expresso exclusivamente em neurônios nociceptivos, no DRG
e seus axônios. Embora o papel deste canal não esteja
completamente elucidado, Ginanneschi e colaboradores (2019)
mostraram que algumas mutações causam hipersensibilidade à
dor inflamatória periférica e neuropática.

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
236
Levando em consideração que a dor crônica é um
problema que afeta cerca de 20% da população mundial
(BAKER; NASSAR, 2020), torna-se importante realizar estudos
relacionados à prospecção de novos fármacos que atuem de
forma analgésica, inibindo ou reduzindo a função das isoformas
Nav1.7, Nav1.8. e Nav1.9.
Nesse enfoque, a toxina ProTx-II isolada da aranha
tarântula apresentou alta seletividade 100 vezes maior aos
canais Nav1.7 do que para outras isoformas, e que poderá ser
utilizada como perspectiva no tratamento de diferentes
neuropatias (LAWRENCE et al., 2019).
Neurotoxinas e seus sítios receptores nos Navs
Seis diferentes sítios receptores que se ligam NTXs
foram identificados na superfície dos Navs (ISRAEL et al., 2017)
(tabela 2). Outros sítios foram identificados, portanto eles não
se ligam NTXs e não será tratado aqui.
Farmacologicamente pode-se classificar as NTXs em
três grupos baseados em seus efeitos funcionais e pela
localização dos seus sítios receptores: (i) toxinas bloqueadoras
do poro; (ii) toxinas modificadoras do gating por se ligar à sítios
receptores intramembrana; (iii) toxinas modificadoras do gating
por se ligar à sítios receptores extracelulares (ISRAEL et al.,
2017).

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
237
Tabela 2. Identificação dos sítios receptores para as NTX nos
Nav ao domínio e o efeito sobre o canal.
Sítio Toxina Domínio Aminoácidos
Sítio 1 Tetrodotoxina
Saxitoxina
µ-conotoxina
D1-M2-S6
D2-M2-S6
D3-M2-S6
D4-M2-S6
D1-M2-S6
D2-S5-S6
D3-M2-S6
D4-M2-S6
D384, E387, Y385,
D400, Y401, E403
E758, E942, E945
K1422, D1426
A1714, D1717, D1532
D400, Y401, W402,
E403
A728, A730, E758
W1239, D1241
W1531, D1532
Sítio 2 Veratridina
Batracotoxina
Grayanotoxina
D1-S6
D4-S6
I433, N434, L437
F1579, N1584, I1760,
F1764 Y1586
Sítio 3 𝛂 -escorpiotoxina
toxina de
anêmona
toxina de aranha
D1-S5-S6
D4-S5-S6
D4-S3-S4
E1613
E1616, V1620, L1624
Sítio 4 𝛃 -escorpiotoxina D1-S5-M1
D2-S1-S2
D2-S3-S4
D3-M2-S6
G845
Sítio 5 Brevetoxinas
Ciguatoxina
D1-S6
D4-S5
Sítio 6 𝛅 -conotoxina D4-S4
Fonte: Adaptado de STEVENS et al., 2011.

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
238
Neurotoxinas Bloqueadoras do Poro do Canal
Neurotoxinas que se ligam ao sítio 1
Neurotoxinas que bloqueiam os Navs interagem com o
sítio 1, localizado na região do poro do canal. Estas toxinas se
ligam no lado extracelular do Nav bloqueando o fluxo de íons
sódio e, consequentemente, o potencial de ação em nervo e
músculo causando paralisia (ISRAEL et al., 2017).
São constituídos por dois diferentes grupos de NTXs. No
primeiro estão as toxinas guanidinas heterocíclicas
hidrossolúveis: a tetrodotoxina (TTX), inicialmente isolada do
baiacu japonês (Fugu rubripes), posteriormente encontrada em
mais de 40 espécies de peixes, moluscos, caranguejos,
cefalópodes e anfíbios da América Central; e a saxitoxina
(STX), isolada do dinoflagelado Gonyaulax catenella, podendo
ser encontrada também em mexilhões e moluscos bivalves que
deles se alimentam. No segundo grupo estão os peptídeos
tóxicos µ-conotoxina (µ-CTX), isolada do Conus geographus.
Foi mostrado que 2 toxinas de aranha também se ligam ao sítio
1 bloqueando o poro: a hainantoxin-I (HNTX-I) da Ornithoctonus
hainana; e a PnTx1 da aranha sulamericana Phoneutria
nigriventer (STEVENS et al., 2011).
A localização do sítio receptor da TTX e STX envolve
resíduos de aminoácidos de todos os domínios, aparentemente
formando anéis dentro do poro: um mais externo formado por
aminoácidos ácidos (E387, E945, D1426 e D1717) e outro mais
interno, formado pelo filtro de seletividade (D384, E942, K1422
e A1714) (ISRAEL et al, 2017). Foram identificados também
outros resíduos que reduzem a ligação da TTX, como D400,

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
239
Y401 e E403D em D1, T759 e E758 em D2 e D1532 em D4
(ISRAEL et al, 2017). A clonagem da isoforma cardíaca mostrou
que a baixa afinidade da TTX pelo canal era a substituição de
um resíduo aromático (Fen ou Tyr) na posição 385 por uma
cisteína. Desta forma, este resíduo também foi identificado
como sendo importante para a ligação da toxina guanídica
(ISRAEL et al., 2017). Porém, mutações que afetam
significativamente a ligação da TTX e STX apenas reduzem
fracamente a afinidade da µ-CTX ou PnTx1, indicando que
embora a ligação destas duas classes de toxinas seja
competitiva sobre o canal de sódio, devam haver interações
distintas (STEVENS et al., 2011), como pode ser visto na tabela
2.
Toxinas Modificadoras do Gating: Receptores
Intramembrana
As NTX lipossolúveis agem nos sítios 2 e 5 dos Navs.
Elas se ligam a sítios que são distintos ao poro ou ao sensor de
voltagem favorecendo o estado aberto do canal por meio de
interação alostérica indireta (VAN THEEMSCHE et al., 2020).
Neurotoxinas que se ligam ao sítio 2
Compostos policíclicos apolares, como a veratridina,
aconitina, batracotoxina e grayanotoxina, se ligam ao sítio 2 do
canal Nav. As toxinas antillatoxina e hoiamide isoladas de
algumas cianobactérias (Lyngbya majuscula) também se ligam
a esse sítio (STEVENS et al., 2011). Estas NTXs se ligam de

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
240
forma alostérica, preferencialmente no estado ativado,
mantendo uma ativação persistente mesmo no potencial de
repouso da membrana, que pode ser explicada pelo
deslocamento do potencial de ativação para valores negativos
pelo bloqueio da inativação e pela redução da seletividade
(VAN THEEMSCHE et al., 2020).
As regiões transmembrana da região do poro D1-S6
(I439, N440 e L443) e D4-S6 (F1586 e N1591) foram
inicialmente identificadas como sendo importantes para a
ligação da batracotoxina ao sítio 2 (VAN THEEMSCHE et al.,
2020). A ligação de toxinas ao segmento D4-S6 provavelmente
é responsável pelo bloqueio da inativação, já que este
segmento está intimamente relacionado à inativação do canal
(NIITSU et al., 2018). Posteriormente, os resíduos S1283 e
LD3-S6 do D3-S6 e N790 e L794 do D2-S6 foram incluídos
como parte do sítio receptor no poro do canal. Mutações de
outros 2 resíduos (G1282 e F1284) na dobradiça entre alça e o
segmento D3-S6, reduziu o efeito da batracotoxina no canal de
barata (VAN THEEMSCHE et al., 2020).
Neurotoxinas que se ligam ao sítio 5 do receptor
As toxinas solúveis em lipídios brevetoxina e ciguatoxina
extraídas dos dinoflagelados marinhos Karenia brevis e
Gambierdicus toxicus, respectivamente aumentam a atividade
nos Navs se ligando aos segmentos transmembranares D1-S6,
D4-S5 e D4-S6, que formam o sítio 5, resultando na inibição da
inativação e no deslocamento da dependência de potencial da
ativação para valores mais negativos. A região da ligação se

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
241
estende dentro de DI-S6 de A/T422 à K465/K477 e dentro de
DIV-S5 de E1510 à K1557 com possível extensão para K1565
(VAN THEEMSCHE et al., 2020).
Toxinas Modificadoras do Gating: Receptores
Extracelulares
Neurotoxinas que se ligam ao sítio 3
Ao sítio 3 se ligam NTXs de escorpião do tipo α, toxinas
de anêmonas e algumas toxinas de aranhas. De acordo com
suas diferentes propriedades farmacológicas, as α-toxinas
podem ser divididas em 3 subgrupos: α-clássica (tóxicas para
mamíferos), inseto α-toxina (tóxicas para insetos) e α-like
(tóxicas para ambos) (STEVENS et al., 2011).
As α-toxinas são famílias distintas de polipeptídeos que
possuem baixa homologia de sequência, mas que lentificam ou
inibem a inativação dos Navs, estabilizando o canal no estado
aberto (CLAIRFEUILLE et al., 2019).
A comparação da estrutura das NTXs de anêmonas e de
escorpião mostrou que dos cinco resíduos de aminoácidos que
estão localizados em posições similares nas duas toxinas,
quatro são básicos, constituindo dois distintos pólos carregados
positivamente sobre a superfície destas NTX. Foi identificado
um resíduo ácido na alça entre os segmentos transmembrana
D4-S3-S4 como importante na ligação eletrostática das α-
toxinas (CLAIRFEUILLE et al., 2019). Análise estrutural
identificou uma superfície hidrofóbica conservada entre as α-
toxinas que parece ser parte do sítio de ligação destas toxinas

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
242
(STEVENS et al., 2011). Modificações químicas e análise de
mutagênese em resíduos aromáticos da superfície hidrofóbica
conservada indicam que estes resíduos aromáticos
conservados estão especificamente envolvidos tanto no efeito
farmacológico quanto na estabilidade estrutural da toxina.
Embora os sítios das α-toxinas se sobreponham, estas toxinas
apresentam interação distintas com o canal (STEVENS et al.,
2011).
Neurotoxinas que se ligam ao sítio 4
As NTXs que se ligam ao sítio 4 do Nav são
representadas pelas β-toxinas de escorpião. Estas toxinas
deslocam fortemente a dependência de voltagem da ativação
para valores mais negativos (ISRAEL et al., 2017).
Por meio de quimeras de Navs de músculo esquelético e
cardíaco (que apresentam sensibilidades diferentes às toxinas
do tipo β), o domínio II foi identificado como responsável pela
ligação e efeito de uma β-toxina do escorpião Tityus serrulatus
(MARTIN-EAUCLAIRE et al., 2018). Evidências indicam que a
potente toxina do tipo do escorpião Centruroides suffusus
suffusus (Css-IV) se liga a resíduos de aminoácidos que são
exteriorizados quando o segmento D2-S4 se move em direção
ao exterior no momento da despolarização, imobilizando esse
sensor de voltagem em um estado parcialmente ativo (ISRAEL
et al., 2017). Uma vez que esse sensor de voltagem já está na
posição ativada, voltagens que normalmente não seriam
suficientes para abrir o canal, passam a abrí-lo, o que explica o
deslocamento do potencial de ativação do canal para valores

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
243
hiperpolarizados. No entanto, neste estado a inativação é
favorecida, o que pode reduzir o número de canais disponíveis
e, conseqüentemente, o pico da corrente de sódio (MARTIN-
EAUCLAIRE et al., 2018). Isto explica porque a toxina requer
um pré-pulso despolarizante para a sua ligação. Identificaram
também a mutação G845N, na alça próxima ao segmento D2-
S4, como responsável pela ligação da toxina. Outras regiões
como D2-S1-S1 e D3-S5-S6 também foram implicadas no
macro-sítio 4 (ISRAEL et al., 2017).
Neurotoxinas que se ligam ao sítio 6
A δ-toxina que se liga ao sítio 6. δ-conotoxina, é uma
toxina hidrofóbica isolada do caracol marinho do gênero Conus,
assim como as toxinas que se ligam ao sítio 3, ela lentifica a
inativação dos Navs. Estudos envolvendo estrutura e função
indicaram que a δ-conotoxina interage com um grupo de
resíduos de aminoácidos no segmento D4-S4, e compete com
a δ-hexatoxina Hv1 próximo ao sítio 3 do canal. Sua ação e
ligação são sinérgicas às α-toxinas, provavelmente porque
ambas se ligam ao sensor de voltagem D4-S4 numa
conformação similar (ISRAEL et al., 2017).
CONCLUSÕES
As manifestações clínicas entre os diferentes tipos de
mutações são, de maneira geral, muito próximas, dificultando o
diagnóstico e a escolha farmacológica ideal. Os estudos
genéticos que evidenciam as mutações presentes em pacientes

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
244
que sofrem de epilepsia se tornou ao longo dos últimos anos
uma ferramenta auxiliadora na conduta clínica, melhorando a
escolha farmacológica e a qualidade de vida dos pacientes.
As toxinas de animais com suas seletividades e alta
afinidade aos receptores e canais iônicos vêm se tornando
protótipos de fármacos indispensáveis para a melhor
compreensão farmacológica dessas canalopatias e/ou alvo
terapêutico para a melhoria do prognóstico e qualidade de vida
dos indivíduos portadores.
REFERÊNCIAS BIBLIOGRÁFICAS AHERN, C. A., et al. The hitchhiker's guide to the voltage-gated sodium channel galaxy. J Gen Physiol. 2016 Jan;147(1):1-24. doi: 10.1085/jgp.201511492. PMID: 26712848; PMCID: PMC4692491. BAKER, M. D.; NASSAR, M. A. Painful and painless mutations of SCN9A and SCN11A voltage-gated sodium channels. European Journal of Physiology, 2020, pg.865-880. BEN-SHALOM, R., et al. Opposing Effects on NaV1.2 Function Underlie Differences Between SCN2A Variants Observed in Individuals With Autism Spectrum Disorder or Infantile Seizures. Biol Psychiatry. 2017 Aug 1;82(3):224-232. doi: 10.1016/j.biopsych.2017.01.009. Epub 2017 Jan 27. PMID: 28256214; PMCID: PMC5796785. BRUNKLAUS, A., LAL, D. Sodium channel epilepsies and neurodevelopmental disorders: from disease mechanisms to clinical application. Dev Med Child Neurol. 2020 Jul;62(7):784-792. doi: 10.1111/dmcn.14519. Epub 2020 Mar 30. PMID: 32227486. CLAIRFEUILLE, T., et al,. Seizures and Epilepsy. Med Clin North Am. 2019 Mar;103(2):309-324. doi: 10.1016/j.mcna.2018.10.002. Epub 2018 Dec 3. PMID: 30704683. COX, J. J.; et al. An SCN9A channelopathy causes congenital inability to experience pain. Nature, 2006, V. 444, p.894-898. DEUIS, J. R., et al. The pharmacology of voltage-gated sodium channel activators. Neuropharmacology. 2017 Dec;127:87-108. doi: 10.1016/j.neuropharm.2017.04.014. Epub 2017 Apr 14. PMID: 28416444.

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
245
DUAN, G.; et al., A variant in the SCN10A enhancer may affect human mechanical pain sensitivity. Molecular Pain, 2018. V.14, p.1-10. GINANNESCHI, F.; et al. SCN11A variant as possible pain generator in sensory axonal neuropathy. Neurological Sdences, 2019. V. 40, p.1295-1297. HUFF, J. S., MURR, N. Seizure. [Updated 2020 Sep 25]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2020 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK430765/ JOHANNESEN, K. M., et al. Early mortality in SCN8A-related epilepsies. Epilepsy Res. 2018 Jul;143:79-81. doi: 10.1016/j.eplepsyres.2018.04.008. Epub 2018 Apr 13. PMID: 29677576. LARSEN, J., et al. The phenotypic spectrum of SCN8A encephalopathy. Neurology. 2015 Feb 3;84(5):480-9. doi: 10.1212/WNL.0000000000001211. Epub 2015 Jan 7. PMID: 25568300; PMCID: PMC4336074. LAWRENCE, N., et al. Peptide-Membrane Interactions Affect the Inhibitory Potency and Selectivity of Spider Toxins ProTx-II and GpTx-1. ACS Chem Biol. 2019 Jan 18;14(1):118-130. doi: 10.1021/acschembio.8b00989. Epub 2018 Dec 21. PMID: 30507158. MANTEGAZZA, M., BROCCOLI, V. SCN1A/NaV 1.1 channelopathies: Mechanisms in expression systems, animal models, and human iPSC models. Epilepsia. 2019 Dec;60 Suppl 3:S25-S38. doi: 10.1111/epi.14700. PMID: 31904127. MARTIN-EUCLAIRE, M. F., et al. Ts1 from the Brazilian scorpion Tityus serrulatus: A half-century of studies on a multifunctional beta like-toxin. Toxicon. 2018 Sep 15;152:106-120. doi: 10.1016/j.toxicon.2018.07.024. Epub 2018 Jul 29. PMID: 30059695. MEI, D., et al. Dravet syndrome as part of the clinical and genetic spectrum of sodium channel epilepsies and encephalopathies. Epilepsia. 2019 Dec;60 Suppl 3:S2-S7. doi: 10.1111/epi.16054. PMID: 31904125. MENEZES, L. F. S., et al. Epilepsy-Related Voltage-Gated Sodium Channelopathies: A Review. Front Pharmacol. 2020 Aug 18;11:1276. doi: 10.3389/fphar.2020.01276. PMID: 33013363; PMCID: PMC7461817. MORAN, Y., et al. Evolution of voltage-gated ion channels at the emergence of Metazoa. J Exp Biol. 2015 Feb 15;218(Pt 4):515-25. doi: 10.1242/jeb.110270. PMID: 25696815. REYNOLDS C., et al. The phenotypic spectrum of SCN2A-related epilepsy. Eur J Paediatr Neurol. 2020 Jan;24:117-122. doi: 10.1016/j.ejpn.2019.12.016. Epub 2019 Dec 12. PMID: 31924505.

TOXINAS DE ANIMAIS COM ALVO EM CANAIS PARA SÓDIO: FERRAMENTAS FARMACOLÓGICAS VALIOSAS PARA CANALOPATIAS
NEUROLÓGICAS
246
SANDERS, S. J., et al. Progress in Understanding and Treating SCN2A-Mediated Disorders. Trends Neurosci. 2018 Jul;41(7):442-456. doi: 10.1016/j.tins.2018.03.011. Epub 2018 Apr 23. PMID: 29691040; PMCID: PMC6015533. SHEN, H,et al. Structural basis for the modulation of voltage-gated sodium channels by animal toxins. Science. 2018. Oct 19;362(6412):eaau2596. doi: 10.1126/science.aau2596. Epub 2018 Jul 26. PMID: 30049784. STEVENS, M., et al. Neurotoxins and their binding areas on voltage-gated sodium channels. Front Pharmacol. 2011 Nov 9;2:71. doi: 10.3389/fphar.2011.00071. PMID: 22084632; PMCID: PMC3210964. The Human Gene Mutation Database. Disponível em: <http://www.hgmd.cf.ac.uk/ac/index.php>. Acesso em 25 out 2020. THOMAS, R. H., BERKOVIC, S. F. The hidden genetics of epilepsy: a clinically important new paradigm. Nat Rev Neurol. 2014 May;10(5):283-92. doi: 10.1038/nrneurol.2014.62. Epub 2014 Apr 15. PMID: 24733163. YANG, C., et al. Variable epilepsy phenotypes associated with heterozygous mutation in the SCN9A gene: report of two cases. Neurol Sci. 2018 Jun;39(6):1113-1115. doi: 10.1007/s10072-018-3300-y. Epub 2018 Mar 2. PMID: 29500686. ZAMAN, T., et al. Mutations in SCN3A cause early infantile epileptic encephalopathy. Ann Neurol. 2018 Apr;83(4):703-717. doi: 10.1002/ana.25188. Epub 2018 Mar 30. Erratum in: Ann Neurol. 2019 Jun;85(6):948. PMID: 29466837; PMCID: PMC5912987. ZHANG, S.; et al. SCN9A Epileptic Encephalopathy Mutations Display a Gain-offunction Phenotype and Distinct Sensitivity to Oxcarbazepine. Neurosci. Bull., 2019.
AGRADECIMENTOS
Agradecemos à Fundação Ezequiel Dias (FUNED) - MG pelo suporte à pesquisa, apoio financeiro e colaborações acadêmico-científicas institucionais; à Universidade Federal da Paraíba - UFPB, pelo ensino gratuito e de excelência; ao Instituto de Pesquisa em Fármacos e Medicamentos; ao Programa de Pós-graduação em Produtos Naturais e Sintéticos Bioativos e ao Laboratório de Ensaios Toxicológicos, pela infraestrutura e suporte à pesquisa; à CAPES, FAPEMIG e ao CNPq pelo fomento aos projetos e bolsas de estudo.

247
CIÊNCIAS BIOLÓGICAS - IMUNOLOGIA

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
248
CAPÍTULO 12
ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
Rita de Cássia de Sena dos SANTOS 1
Louise Mangueira de LIMA 2
João Bastista OLIVEIRA 2 Cosmo Isaías Duvirgens VIEIRA 2
Márcia Regina PIUVEZAM 3
1 Graduanda do curso de Biotecnologia, UFPB; 2 Graduandos do curso de Farmácia, UFPB; 3 Orientadora / Professora do Programa de Pós Graduação em Produtos Naturais e Sintéticos
Bioativos/UFPB. [email protected]
RESUMO: A rinite alérgica e asma coexistem com alta frequência em seres humanos, e suas manifestações clínicas em conjunto, levam o nome de Síndrome da Asma e Rinite Alérgicas Combinadas (CARAS - Combined allergic rhinitis and asthma syndrome) ocorrendo a polarização para o perfil de resposta imune do tipo 2 (TH2). A farmacoterapia atual promove inúmeros efeitos indesejáveis, interferindo na qualidade de vida dos acometidos. Dessa maneira, a investigação de terapias com alvos específicos, a fim de diminuir o surgimento de efeitos adversos vem sendo realizada. Diante disso, terapias alvo baseadas no uso de anticorpos monoclonais surgem como uma proposta promissora. Assim, o objetivo desse trabalho foi realizar uma revisão bibliográfica exploratória, a fim de averiguar o potencial terapêutico de anticorpos monoclonais sobre o processo fisiopatológico na CARAS. Para tanto, realizou-se um levantamento bibliográfico nas bases de dados Pubmed, Google acadêmico, Science Direct e Medline, utilizando palavras-chaces pré definidas. Os resultados

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
249
mostram que os anticorpos monoclonais específicos para citocinas do perfil 2 amenizam o agravamento da CARAS, com melhoria da função pulmona e diminuição da exarcebação do sintomas nasais e pulmonares. Portanto, os anticorpos monoclonais específicos podem atuar de maneira eficaz no tratamento da CARAS, no entanto, ánalises para avaliar a sua efetividade como monoterapia em tratamentos personalizados devem ser realizados. Palavras-chave: CARAS. Citocinas. Anticorpos Monoclonais. Terapias biológicas.
INTRODUÇÃO
As doenças respiratórias têm levado a quase 7,5 milhões
de mortes anuais no mundo, podendo afetar diretamente a
qualidade de vida dos acometidos, contribuindo diretamente
para o desgaste emocional e social desses indivíduos
(NAGHAVI e colab., 2015). Dentre as doenças respiratórias
podemos citar as doenças pulmonares obstrutivas, pneumonia,
e doenças crônicas, como rinite alérgica (RA) e asma.
A rinite alérgica (RA) é uma das afecções mais comuns
encontradas pelos otorrinolaringologistas e foi o diagnóstico
mais comumente relatado por esses profissionais nos anos
2000. A asma, por sua vez, foi também um dos distúrbios
respiratórios crônicos relatados frequentemente, com uma
prevalência estimada de 358 milhões de casos; além disso, a
Organização Mundial da Saúde prevê um aumento em mais de
100 milhões de novos casos de asma até 2025 (GBD 2015
CHRONIC RESPIRATORY DISEASE COLLABORATORS e
colab., 2017; KURUVILLA e colab., 2019). A asma e a RA
coexistem com muita frequência, podendo a RA ser um fator de
risco para o desenvolvimento de asma em adultos e crianças

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
250
(PAIVA FERREIRA e colab., 2019). Tem sido relatado que até
80% dos pacientes com asma são afetados pela RA, e até 40%
dos pacientes com RA são acometidos por asma (PAIVA
FERREIRA e colab., 2019).
A asma e a RA possuem a mesma base etiológica,
imunopatológica e terapêutica. Ambas as doencas são
caracterizadas pela polarização da resposta imune do tipo 2
(TH2), com o aumento dos níveis séricos de imunoglobulinas E
alérgeno específica (IgE), secreção de citocinas, como IL-4, IL-
5 e IL-13, o que leva ao aumento da infiltração de eosinófilos e
basófilos, além da hiperplasia das células caliciformes e
hipersecreção de muco, resultando na inflamação do tecido
nasal e pulmonar com consequente remodelamento tecidual
(BROŻEK e colab., 2017; CHAUHAN e colab., 2017)
Diversos mecanismos imunológicos têm sido
demonstrados com o objetivo de explicar a interdependência
das vias aéreas superiores e inferiores no que diz respeito ao
desenvolvimento da CARAS. E os processos inflamatórios vêm
sendo considerados o elo de correlação entre essas vias, sendo
amplamente reconhecidas como uma única via aérea. Nesse
contexto, a CARAS é considerada uma associação entre a
inflamação das vias aéreas superiores e inferiores, que causam
uma única doença das vias aéreas (PAIVA FERREIRA e colab.,
2019).
A farmacoterapia comumente usada para manejo da
CARAS baseia-se no uso de corticoides inalatórios, visando a
diminuição do processo inflamatório, além do uso de agonistas
β2-adrenérgicos, a fim de gerar alívio dos sintomas clínicos
respiratórios decorrentes da broncoconstrição (CAZZOLA et al.,
2019). Todavia, são inúmeros os efeitos adversos que essas

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
251
classes de medicamentos apresentam, entre eles observa-se
tremores, aumento de peso e taquicardia. Isso desfavorece a
qualidade de vida dos pacientes e pode interferir na adesão
terapêutica dos mesmos. Em adição, o uso prolongado de
glicocorticoides por mais de 60 dias mostrou-se propenso ao
desenvolvimento de distúrbios de natureza endócrina,
neuropsiquiátrica, gastrointestinal, muscuesquelética,
cardiovascular, dermatológica, ocular ou imunológica em até
90% dos pacientes (ORAY e colab., 2016).
Diante do exposto acima, diversos grupos de pesquisa
buscam terapias com alvos mais específicos, que venham a
diminuir o surgimento desses efeitos adversos e favorecer um
melhor prognóstico aos acometidos, permitindo um controle da
sintomatologia e proporcionando uma melhoria na qualidade de
vida dos pacientes.
Nesse contexto, a pesquisa com imunobiológicos vem
crescendo nos últimos anos, pois possuem a capacidade de
melhorar significamente diversas manifestações crônicas ou
não crônicas, podendo alterar o curso da doença (BIELORY e
colab., 2016). Imunobiológicos, como são conhecidos, são
medicamentos feitos a partir de uma variedade de moléculas,
como anticorpos monoclonais, antagonistas de receptores ou
bloqueadores de sinalização intracelular, tendo como alvos
específicos receptores de superfície, citocinas ou moléculas de
sinalização intracelular envolvidas nas respostas inflamatórias,
sendo considerados alvos terapêuticos cruciais para o
tratamento de várias doenças (BIELORY e colab., 2016).
Várias estratégias terapêuticas têm tido o foco na
intervenção das atividades das citocinas relacionadas ao perfil
de resposta imune do tipo 2 envolvidas na CARAS, tais como

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
252
IL-4, IL-5 e IL-13 que promovem a progressão da doença. O uso
de anticorpos monoclonais específicos para as citocinas,
oferece algumas vantagens em comparação às drogas
convencionais. Uma vez que, possuem alta afinidade e
especificidade ao alvo, são relativamente estáveis, com uma
meia-vida de eliminação longa, de semanas a meses, e ainda,
ao serem degradados não originam metabólitos tóxicos
(MOULD e MEIBOHM, 2016).
Nesse sentido, o objetivo dessa revisão bibliográfica foi
avaliar os mecanismos inflamatórios da CARAS e destacar as
terapias com anticorpos monoclonais específicos, direcionados
as moléculas da resposta imune do tipo 2, como potenciais
terapêuticos para a modulação e tratamento da doença.
MATERIAIS E MÉTODO
O presente trabalho trata-se de uma revisão bibliográfica
com o intuito de esclarecer o possível potêncial terapêutico de
anticorpos monoclonais sobre o processo fisiopatológico da
Síndrome da Asma e Rinite Alérgicas Combinadas (CARAS).
Para tanto, foram utilizadas as base de dados Pubmed, Google
Acadêmico, Sciente direct e Medline, utilizando os seguintes
descritores: Combined allergic rhinitis and asthma syndrome;
CARAS; allergic rhinitis; asthma; cytokines; therapy;
monoclonal antibodies. Foram considerados artigos escritos em
língua portuguesa ou inglesa, publicados de 2015 à 2019 que
tratavam do assunto investigado. Foram excluídos aqueles
trabalhos que não eram relevantes ao tema da pesquisa e que
não estavam de acordo aos objetivos propostos, tendo sido
selecionados 31 artigos relevantes para o estudo. A análise dos

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
253
dados iniciou-se em julho de 2020 e concluiu-se no mês de
outubro de 2020.
RESULTADOS E DISCUSSÃO
O perfil fisiopatológico da CARAS está diretamente
ligado a indivíduos atópicos que são predisponentes
geneticamente a estímulos (alérgenos), e desenvolvem uma
hipersensibilidade imediata. Esses alérgenos, como proteínas
presentes nos ácaros, fungos, baratas e outros, estão dispersos
no ar e, ao entrar em contato com a mucosa do trato
respiratório, podem promover um desequilíbrio entre as
respostas imunes inatas e adaptativas do indivíduo atópico, e
consequentemente desencadear a CARAS (PAIVA FERREIRA
e colab., 2019).
O processo fisiopatológico da CARAS divide-se em três
fases distintas durante processo inflamatório. O nariz e os
brônquios compartilham o mesmo tipo de epitélio,
pseudoestratificado com células cilíndricas, colunares,
apoiados sobre uma membrana basal, onde inicia-se a primeira
fase denominada de fase de sensibilização. Essa fase ocorre
quando o antígeno entra em um contato inicial com o epitélio
das vias aéreas e esse libera alarminas, como linfopoietina
estromal tímica (TSLP), que ativam células fagocitárias e
apresentadoras de antígenos (APC), como macrófagos e
células dendríticas, além de recrutar células imunes circulantes
como os basófilos (LI e colab., 2016). As APCs fagocitam,
processam e apresentam os peptídeos desse antígeno para
células TH0 através do Complexo Maior de Histocompatibilidade
de classe II (MHC-II), processo que ocorre em órgãos linfoides

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
254
secundários, os linfonodos de drenagem. Em adição, o epitélio
libera citocinas como IL-25 e IL-33, que ativam células linfoides
inatas do tipo 2 (ILC-2) que secretam IL-5 e IL-13, aumentando
assim o recrutamento local de eosinófilos e também acabam
estimulando inicialmente a produção de mucinas pelas células
caliciformes presentes no epitélio (YU e colab., 2014).
O recrutamento e ativação de basófilos e eosinófilos,
leva ao aumento da expressão local de IL-4, que favorece o
microambiente para diferenciação das células TH0 em células
TH2. As células TH2 secretam citocinas clássicas desse perfil,
como IL-4, IL-5 e IL-13, favorecendo assim a quimiotaxia de
eosinófilos, hiperplasia e aumento da produção de muco pelas
células caliciformes do epitélio, além da diferenciação de
Linfócitos B em plasmócitos secretores de IgE alérgeno
especifica (WYNN, 2015). Essas IgE agregam-se a parede de
células imunes teciduais como mastócitos, e circulantes, como
eosinófilos e basófilos (LAMBRECHT e HAMMAD, 2015)
caracterizando a fase de sensibilização (primeira fase).
A segunda fase é denominada de fase de desafio, com
exposição posterior do indivíduo ao alérgeno. Nessa fase, o
alérgeno liga-se diretamente a IgE presente na parede de
mastócitos, eosinófilos e basófilos, levando a um processo de
degranulação dessas células, com liberação de diversos
mediadores inflamatórios, tais como histamina, prostaglandinas
e leucotrienos. Nas vias aéreas superiores, esta fase está
associada aos sintomas nasais agudos (espirros, fricção nasal
e rinorreia) e à emergência de sintomas oculares (comichão,
vermelhidão e lacrimejamento), além de aumentar a
permeabilidade vascular, levando à formação de edema, sinais
clássicos da rinite alérgica (BJERMER et al., 2019). Nas vias

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
255
aéreas inferiores, os mediadores liberados geram
broncoconsctrição e hiperresponsividade brônquica, gerando
os sinais e sintomas característicos da asma, como falta de ar,
sibilos e dor no peito (FAHY, 2015).
A terceira fase, a fase tardia ou crônica, está associada
ao remodelamento tecidual das vias aéreas, aumento do
edema, além de desenvolvimento e perpetuação da congestão
nasal, considerada pelos pacientes como um dos sintomas
mais problemáticos na RA. Como resultado da inflamação da
mucosa, os tecidos tornam-se estimulados e reagem mais
vigorosamente à exposição ao alérgeno. Estas reações de fase
tardia, e as modificações na responsividade dos tecidos
contribuem para a hiperresponsividade brônquica, e
consequente remodelamento das vias aéreas inferiores,
gerando também uma densa deposição de fibras colágenas que
diminuem a capacidade de elasticidade dos pulmões,
dificultando assim o processo respiratório (BJERMER e colab.,
2019).
A co-incidência de RA e asma demonstra que as duas
doenças podem compartilhar uma mesma base genética
molecular, onde estudos já mostram uma relação importante
nos genes que transcedem a RA-asma (MAO e colab., 2018).
Outros fatores também são compartilhados em ambas as
doenças, desde influxo celular de eosinófilos, basófilos e
células TH2, como também o aumento da liberação de
mediadores inflamatórios como IL-4, IL-5, IL-13, IgE, moléculas
de adesão, histamina e leucrotrienos, processos já descritos
anteriormente (PAIVA FERREIRA e colab., 2019). Apesar do
notável progresso no diagnóstico e manejo da CARAS, a asma
alérgica grave continua a ser uma doença potencialmente fatal

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
256
com estratégias terapêuticas limitadas e com efeitos adversos
importantes (WANG, Xu e colab., 2016).
Esses fatores em potencial estão relacionados à
complexa imunopatologia que envolve a CARAS, onde células
TH2 medeiam o início da resposta alérgica e, em seguida, os
eosinófilos são atraídos com maior intensidade para o local da
inflamação pela circulação, sendo essas células identificadas
como tendo um papel crítico nos sintomas de asma e alergias,
incluindo a participação no remodelamento e regulação da
resposta inflamatória das vias aéreas (MCBRIEN e MENZIES-
GOW, 2017). Portanto, a modulação negativa desses fatores se
apresenta como importante alvo para o manejo da CARAS.
Nesse contexto, o conhecimento de que tanto a asma,
quanto a RA são frequentes distúrbios mediados por células
TH2, juntamente com o fato dessas doenças serem grandes
problemas de saúde pública, levaram a um grande investimento
de empresas farmacêuticas e de biotecnologia em programas
de desenvolvimento de drogas dirigidas a citocinas associadas
a células TH2 (LAMBRECHT e colab., 2019; PAIVA FERREIRA
e colab., 2019).
Dada as circunstâncias, o desenvolvimento de
anticorpos monoclonais específicos para moléculas e/ou
receptores envolvidos na resposta imune, sua alta afinidade e
eficiência, e a sua capacidade de aumentar ou suprimir a
resposta imune, demostram que os imunobiológicos podem ser
uma alternativa para o tratamento de pacientes com doenças
inflamatórias crônicas e de difícil regulação. Até o presente
momento foram desenvolvidos vários anticorpos monoclonais
para o tratamento da asma moderada a grave, melhorando a
eficácia dos tratamentos, chegando a reduzir os efeitos

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
257
adversos causados pela ação dos medicamentos tradicionais,
inferindo que esses imunobiológicos podem ser usados como
uma estratégia terapêutica para pacientes afetados com a
CARAS (MOULD e MEIBOHM, 2016; REGATEIRO e colab.,
2017).
O Omalizumab®, anticorpo monoclonal anti IgE alérgeno
específica, quando utilizado em pacientes demonstrou que
esses obtiveram melhora no escore dos sintomas alérgicos e
não alérgenos tanto na asma quanto nos pólipos nasais (WANG
et al., 2015). Esse anticorpo monoclonal é eficaz em minimizar
a exacerbação da asma, naqueles pacientes com o perfil de
resposta imune do tipo 2 (TH2). Acredita-se que os mecanismos
de ação além de inibir a IgE alergeno específica possa também
estar relacionado ao aumento na secreção de interferon-alfa
(IFNα) pelas células dendríticas plasmocitoides (pDCs) (FAHY,
2015). Estudos mostraram que o Omalizumab® diminuiu na
asma as doses de manutenção dos corticosteroides orais.
Estes efeitos são aparentes em crianças e adultos e foram
confirmados em vários estudos pós-registro. No mais, o
Omalizumab® mostrou-se bem tolerado e, além das reações no
local da injeção e um risco de anafilaxia em alguns indivíduos,
é geralmente considerado seguro (CAMPO e colab., 2019).
O Quilizumab® é um anticorpo monoclonal IgG1
humanizado, e tem como alvo o segmento M1 da IgE, que leva
à depleção de células B de memória convertida por IgE.
Estudos randomizados mostraram redução significativa de IgE
total e específica para alérgenos em pacientes com asma,
apresentando um perfil de segurança aceitável (HARRIS e
colab., 2016).

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
258
O Dupilumab® é um anti-receptor α para a IL-4 (IL-4Rα)
que inibe a sinalização de IL-4 e IL-13. Por estudos
randomizados, analisou-se o tratamento com Dupilumab® e
observou-se que é relativamente bem tolerado e melhorou
significativamente os sintomas clínicos da asma e a qualidade
de vida, além de reduzir o risco de exacerbação grave em
pacientes com asma não controlada (XIONG e colab., 2019).
Em um estudo de fase III, o Dupilumab® melhorou o volume
expiratório forçado pré-broncodilatador em 1 segundo
(CORREN e colab., 2018).
O Mepolizumab® é um imunobiológico humanizado anti-
IL-5, bloqueador de IgG1-k humanizado. Esse anticorpo está
sendo comercializado desde o final de 2016 como indicação
para o tratamento adjuvante da asma eosinofílica refratária
grave em adultos (REGATEIRO e colab., 2017). Ensaios
direcionados demonstraram consistentemente que o
Mepolizumab® diminuiu significativamente as taxas de
exacerbação da asma e é poupador de corticosteroides orais
(ALBERS e colab., 2019).
O Reslizumab® é um anticorpo monoclonal humanizado
anti-IL-5. Possui alta afinidade pela IL-5 humana, responsável
pela diferenciação, maturação, recrutamento e ativação de
eosinófilos humanos. Liga-se à IL-5 circulante especificamente,
impedindo-a de se ligar ao seu receptor, alterando a
sobrevivência e atividade dos eosinófilos. Obteve autorização
para comercialização em 2016, com indicação para o
tratamento adjuvante da asma eosinofílica refratária grave em
adultos. Ensaios clínicos de fase II demonstram que o
Reslizumab® é eficaz no controle da asma ou na redução das
exacerbações dos quadros asmáticos quando usados no

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
259
tratamento de pacientes com asma que apresentam eosinofilia
persistente (REGATEIRO e colab., 2017).
O Benralizumab® é um anticorpo monoclonal IgG1
recombinante humanizado que se liga ao IL-5Rα (receptor)
(CD125) (FAHY, 2015). Ele exerce a sua função por
citotoxicidade celular induzida por anticorpos (ADCC) das
células que expressam o IL-5Rα. Depleta diretamente os
eosinófilos de forma quase completa e mais duradoura (PHAM
e colab., 2016) embora, em termos teóricos, a sua
administração poderia levar a depleção de outras células além
dos eosinófilos, por exemplo, os linfócitos B e os basófilos que
expressam baixas quantidades de IL-5Rα na sua superfície.
Esses fatores poderão ter implicações no seu uso terapêutico,
tanto em termos de eficácia como de efeitos adversos. Dados
recentemente apresentados de ensaios clínicos de fase II de
Benralizumab® mostraram que ele também se mostrou eficaz
na melhora do controle da asma ou na redução das
exacerbações da asma quando usados no tratamento de
pacientes com asma que apresentam eosinofilia persistente,
resistentes ao tratamento com corticosteroides (PARK e colab.,
2018).
O Lebrikizumab® interfere diretamente com a ligação da
IL-13 ao IL-4Rα impedindo a formação de complexos
sinalizadores de IL-13 - IL-4 - IL-4Rα, e GSK679586
(GlaxoSmithKline) (FAHY, 2015). Estudos randomizados,
duplo-cegos e controlados por placebo, avaliaram doses
múltiplas de Lebrikizumab® em pacientes com asma não
controlada, resistentes á corticosteroide inalado de dose média
a alta e os resultados demonstraram a eficácia do
Lebrikizumab® na melhora das crises de asma e na função
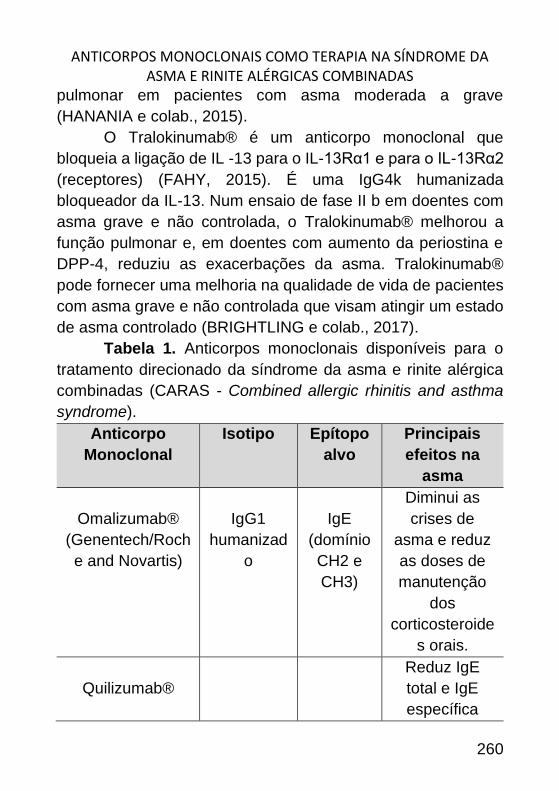
ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
260
pulmonar em pacientes com asma moderada a grave
(HANANIA e colab., 2015).
O Tralokinumab® é um anticorpo monoclonal que
bloqueia a ligação de IL -13 para o IL-13Rα1 e para o IL-13Rα2
(receptores) (FAHY, 2015). É uma IgG4k humanizada
bloqueador da IL-13. Num ensaio de fase II b em doentes com
asma grave e não controlada, o Tralokinumab® melhorou a
função pulmonar e, em doentes com aumento da periostina e
DPP-4, reduziu as exacerbações da asma. Tralokinumab®
pode fornecer uma melhoria na qualidade de vida de pacientes
com asma grave e não controlada que visam atingir um estado
de asma controlado (BRIGHTLING e colab., 2017).
Tabela 1. Anticorpos monoclonais disponíveis para o
tratamento direcionado da síndrome da asma e rinite alérgica
combinadas (CARAS - Combined allergic rhinitis and asthma
syndrome).
Anticorpo
Monoclonal
Isotipo Epítopo
alvo
Principais
efeitos na
asma
Omalizumab®
(Genentech/Roch
e and Novartis)
IgG1
humanizad
o
IgE
(domínio
CH2 e
CH3)
Diminui as
crises de
asma e reduz
as doses de
manutenção
dos
corticosteroide
s orais.
Quilizumab®
Reduz IgE
total e IgE
específica
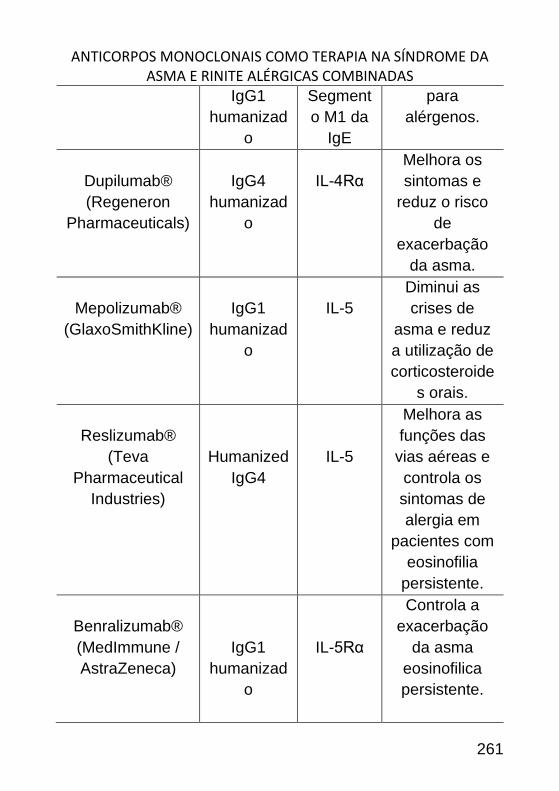
ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
261
IgG1
humanizad
o
Segment
o M1 da
IgE
para
alérgenos.
Dupilumab®
(Regeneron
Pharmaceuticals)
IgG4
humanizad
o
IL-4Rα
Melhora os
sintomas e
reduz o risco
de
exacerbação
da asma.
Mepolizumab®
(GlaxoSmithKline)
IgG1
humanizad
o
IL-5
Diminui as
crises de
asma e reduz
a utilização de
corticosteroide
s orais.
Reslizumab®
(Teva
Pharmaceutical
Industries)
Humanized
IgG4
IL-5
Melhora as
funções das
vias aéreas e
controla os
sintomas de
alergia em
pacientes com
eosinofilia
persistente.
Benralizumab®
(MedImmune /
AstraZeneca)
IgG1
humanizad
o
IL-5Rα
Controla a
exacerbação
da asma
eosinofilica
persistente.

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
262
Lebrikizumab®
(Genentech /
Roche)
IgG4
humanizad
o
IL-13
(epítopo
de
ligação
IL-4Rα)
Diminui as
crises de
asma e
melhora a
função
pulmonar.
Tralokinumab®
(MedImmune /
AstraZeneca)
IgG1
humanizad
o
IL-13Rα1
e
IL-13Rα2
Melhora a
função
pulmonar
reduz a
exacerbação
da asma.
Fonte: autoria própria.
CONCLUSÕES
Os imunobiológicos, anticorpos monoclonais dirigidos
para citocinas do perfil de resposta imune tipo 2 (IL-4, IL-5 e IL-
13) e IgE-alergeno especifica desempenham um papel
fundamental no controle dos sintomas clínicos das alergias das
vias aéreas. Diminuindo a exarcerbação da rinite / asma,
culminando na modulação da fisiopatologia da sindrome da
asma e rinite alergica combinadas (CARAS). Os estudos
realizados com os anticorpos monoclonais aqui descritos
mostram-se eficazes na redução do risco de exarcebação de
alergias das vias aéreas e melhoria da função pulmonar, sendo
capazes de diminuir o perfil de resposta TH2; portanto, são
promissores na melhora do quadro clínico da CARAS.

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
263
REFERÊNCIAS BIBLIOGRÁFICAS ALBERS, Frank C. e colab. Update: Mepolizumab treatment in patients with severe eosinophilic asthma and prior omalizumab use. Allergy, Out 2019. BIELORY, Brett P. e colab. Emerging therapeutics for ocular surface disease. Current Opinion in Allergy and Clinical Immunology, v. 16, n. 5, p. 477–486, out. 2016. BJERMER, Leif e colab. The complex pathophysiology of allergic rhinitis: scientific rationale for the development of an alternative treatment option. Allergy, Asthma & Clinical Immunology, v. 15, n. 1, p. 24, Dez 2019. BRIGHTLING, Christopher E. e colab. Effect of tralokinumab on GINA control in severe, uncontrolled asthma. European Respiratory Society, Set 2017 Disponível em: <http://erj.ersjournals.com/lookup/doi/10.1183/1393003.congress-2017.PA4683>. Acesso em: 6 jun. 2019 BROŻEK, Jan L. e colab. Allergic Rhinitis and its Impact on Asthma (ARIA) guidelines—2016 revision. Journal of Allergy and Clinical Immunology, v. 140, n. 4, p. 950–958, Out 2017. Disponível em: <http://www.ncbi.nlm.nih.gov/pubmed/28602936>. Acesso em: 16 abr 2019. CAMPO, Paloma e colab. Severe asthma phenotypes in patients controlled with omalizumab: A real-world study. Respiratory Medicine, v. 159, Nov 2019. CAZZOLA, Mario e colab. Bronchodilators in subjects with asthma-related comorbidities. Respiratory Medicine, v. 151, p. 43–48, Maio 2019. CHAUHAN, Preeti S. e DASH, D. e SINGH, Rashmi. Intranasal Curcumin Inhibits Pulmonary Fibrosis by Modulating Matrix Metalloproteinase-9 (MMP-9) in Ovalbumin-Induced Chronic Asthma. Inflammation, v. 40, n. 1, p. 248–258, 19 Fev 2017. CORREN, J. e colab. DUPILUMAB IMPROVES ASTHMA CONTROL IN PATIENTS WITH UNCONTROLLED, MODERATE-TO-SEVERE ASTHMA, REGARDLESS OF EXACERBATION HISTORY. Annals of Allergy, Asthma & Immunology, v. 121, n. 5, p. S42–S43, Nov 2018. FAHY, John V. Type 2 inflammation in asthma — present in most, absent in many. Nature reviews. Immunology, v. 15, n. 1, p. 57, 2015. GBD 2015 CHRONIC RESPIRATORY DISEASE COLLABORATORS, Joan B e colab. Global, regional, and national deaths, prevalence, disability-adjusted life years, and years lived with disability for chronic obstructive pulmonary disease and asthma, 1990-2015: a systematic analysis for the Global Burden of Disease Study 2015. The Lancet.

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
264
Respiratory medicine, v. 5, n. 9, p. 691–706, Set 2017. HANANIA, Nicola A e colab. Lebrikizumab in moderate-to-severe asthma: pooled data from two randomised placebo-controlled studies. Thorax, v. 70, n. 8, p. 748–756, Ago 2015. HARRIS, Jeffrey M. e colab. A randomized trial of the efficacy and safety of quilizumab in adults with inadequately controlled allergic asthma. Respiratory Research, v. 17, n. 1, p. 29, Dez 2016. KURUVILLA, Merin E. e colab. Epidemiology and risk factors for asthma. Respiratory Medicine. v.149, p. 16-22, 1 Mar 2019 LAMBRECHT, Bart N. e HAMMAD, Hamida. The immunology of asthma. Nature Immunology, v. 16, n. 1, p. 45–56, 18 Dez 2015. LAMBRECHT, Bart N. e HAMMAD, Hamida e FAHY, John V. The Cytokines of Asthma. Immunity, v. 50, n. 4, p. 975–991, Abr 2019. LI, Hong-tao e colab. Treatment of allergic rhinitis with CpG oligodeoxynucleotides alleviates the lower airway outcomes of combined allergic rhinitis and asthma syndrome via a mechanism that possibly involves in TSLP. Experimental Lung Research, v. 42, n. 6, p. 322–333, Jul 2016. MAO, Zhengdao e colab. Transcriptional regulation on the gene expression signature in combined allergic rhinitis and asthma syndrome. Epigenomics, v. 10, n. 2, p. 119–131, Fev 2018. MCBRIEN, Claire N e MENZIES-GOW, Andrew. The Biology of Eosinophils and Their Role in Asthma. Frontiers in medicine, v. 4, p. 93, Jun 2017. MOULD, Diane R. e MEIBOHM, Bernd. Drug Development of Therapeutic Monoclonal Antibodies. BioDrugs: Springer International Publishing, v. 30, n. 4, p. 275–293, 24 ago. 2016. NAGHAVI, Mohsen e colab. Global, regional, and national age-sex specific all-cause and cause-specific mortality for 240 causes of death, 1990-2013: A systematic analysis for the Global Burden of Disease Study 2013. The Lancet, v. 385, n. 9963, p. 117–171, 2015. ORAY, Merih e colab. Long-term side effects of glucocorticoids. Expert Opinion on Drug Safety, v.15, n.4, p. 457-465, 2016 PAIVA FERREIRA, Laércia K.D. e colab. Combined allergic rhinitis and asthma syndrome (CARAS). International Immunopharmacology, v.74, p. 105718, Sep 2019. PARK, Hae-Sim e colab. Benralizumab Reduces Exacerbations and Improves Lung Function in Patients From Republic of Korea With Severe, Uncontrolled Asthma: Subgroup Analysis of the SIROCCO Trial. Journal of Allergy and Clinical Immunology, v. 141, n. 2, p. AB14, Fev 2018. PHAM, Tuyet-Hang e colab. Reductions in eosinophil biomarkers by

ANTICORPOS MONOCLONAIS COMO TERAPIA NA SÍNDROME DA ASMA E RINITE ALÉRGICAS COMBINADAS
265
benralizumab in patients with asthma. Respiratory Medicine, v. 111, p. 21–29, Fev 2016. REGATEIRO, Frederico e LUÍSA MOURA, Ana e FARIA, Emília. Novel biologicals for the treatment of asthma. Revista Portuguesa de Imunoalergologia, v. 25, n. 2, p. 99–113, 2017. WANG, Min e colab. Association of periostin expression with eosinophilic inflammation in nasal polyps. Journal of Allergy and Clinical Immunology, v. 136, n. 6, p. 1700- 1703.e9, Dez 2015. WANG, Xu e colab. Potent ameliorating effect of Hypoxia-inducible factor 1α (HIF-1α) antagonist YC-1 on combined allergic rhinitis and asthma syndrome (CARAS) in Rats. European journal of pharmacology, v. 788, p. 343–350, Out 2016. WYNN, Thomas A. Type 2 cytokines: mechanisms and therapeutic strategies. Nature Reviews Immunology, v. 15, n. 5, p. 271–282, Maio 2015. XIONG, Xiao-feng e colab. Efficacy and safety of dupilumab for the treatment of uncontrolled asthma: a meta-analysis of randomized clinical trials. Respiratory Research, v. 20, n. 1, p. 108, Dez 2019. YU, Sanhong e colab. Innate lymphoid cells and asthma. Journal of Allergy and Clinical Immunology. [S.l: s.n.]. , Abr 2014
AGRADECIMENTOS Agradeço à primordial orientação da minha orientadora
Márcia Piuvezam, bem como aos colaboradores na construção desse trabalho.

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
266
CAPÍTULO 13
CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
Pedro Henrique de Sousa PALMEIRA 1
Fernanda Silva ALMEIDA 2
Fernando Cézar Comberlang Queiroz Davis dos SANTOS 3
Bárbara Guimarães CSORDAS 4 Tatjana Souza Lima KEESEN 5
1 Mestrando do Programa Multicêntrico de Pós-Graduação em Ciências Fisiológicas, UFPB; 2 Doutoranda do Programa Renorbio de Biotecnologia, UFPB; 3 Graduando do curso de Biotecnologia, UFPB; 4 Pós Doutoranda do Programa de Pós-Graduação em Produtos
Naturais e Sintéticos Bioativos - PgPNBS - UFPB; 5 Orientadora/Professora do CBIOTEC/UFPB.
RESUMO: A atual pandemia desencadeada pelo SARS-CoV-2 vem gerando diversos transtornos em todo o mundo. Batizada de COVID-19, a infecção provocada pelo novo coronavírus, mobilizou diversos pesquisadores na busca pela sua compreensão e por um tratamento adequado. Com elevada capacidade de disseminação, o vírus afetou um grande número de pessoas conduzindo, até o momento, a mais de um milhão óbitos. Com um amplo espectro sintomatológico, uma densa maioria de infectados apresentam sintomas leves enquanto que uma pequena parcela é acometida por sinais clínicos mais graves necessitando, muitas vezes, de hospitalização. O sistema imune inato caracteriza-se como primeira linha de defesa sendo de extrema importância no combate a diversos patógenos. No entanto, embora desempenhe papel protetivo, células inatas parecem contribuir de modo significativo com sintomas mais graves associadas a doença. Nesse contexto, o presente trabalho compilou informações associadas as

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
267
principais características da patologia incluindo epidemiologia e estrutura viral apresentando como foco, no entanto, a participação de leucócitos componentes da imunidade inata em aspectos leve, moderado e grave. São observadas a mobilização de diferentes subpopulações celulares no sangue periférico e em alguns órgãos como o pulmão. Além disso, alterações na frequência de determinadas populações celulares bem como a síntese de mediadores inflamatórios parecem se correlacionar com a forma mais grave da doença. Palavras-chave: COVID-19. Leucócitos. Infecção. Coronavírus. Imunopatologia.
INTRODUÇÃO
No final de dezembro de 2019, uma série de casos de
pneumonia de etiologia até então desconhecida, em Whuan, na
China, passaram a ser notificados. Análises posteriores
identificaram a presença de um novo coronavírus como o
responsável pelas ocorrências. Devido a sua homologia de 80%
com o SARS-CoV, responsável por surtos respiratórios entre
2002-2003, o “novo vírus” foi nomeado em fevereiro de 2020 de
Coronavírus da Síndrome Respiratória Aguda Grave-2 (SARS-
CoV-2) (YUKI; FUJIOGI; KOUTSOGIANNAKI, 2020).
A patologia foi nomeada posteriormente de Doença do
Coronavírus 2019 (COVID-19). Tendo início mediante
transmissão zoonótica, a rápida disseminação viral ocorreu
através do contato indivíduo-indivíduo. Como consequência, a
Organização Mundial da Saúde (OMS) declarou situação de
pandemia em 11 de março de 2020, por conta da presença do
vírus em mais de 200 países e territórios (ZHENG, et. al. 2020).
Com a rápida progressão da doença, medidas
preventivas passaram a ser adotadas conferindo proteção

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
268
social, principalmente aos indivíduos enquadrados nos grupos
de risco. Assim o isolamento e distanciamento social somado à
abertura somente de serviços essenciais como supermercados,
farmácias e hospitais e o uso da máscara facial para a
contenção de partículas infectantes passaram a compor a rotina
populacional (OMS, 2020; ZHOU et al., 2020).
Desenvolvendo uma resposta primária e “inespecífica"
frente a diversos patógenos, a imunidade inata compreende um
sistema de moléculas e células presentes no sangue e em
tecidos que podem agir e se adaptar a diferentes condições
fisiológicas. Embora desempenhe papel protetivo e fucionando
como ponte para a ação da resposta adaptativa, componentes
da imunidade inata podem contribuir com o desenvolvimento de
quadros mais graves no organismo (MELLA et al., 2013;
ABBAS; LITCHMAN; PILLAI, 2019).
OBJETIVOS
Diante do exposto, o presente trabalho objetivou avaliar
e compilar informações acerca da infecção desencadeada pelo
SARS-CoV-2 no qual aspectos clínicos e epidemiológicos foram
inseridos no capítulo. Ademais, compreendendo seu papel
como primeira linha de defesa no organismo, a revisão trouxe
como foco a ação da imunidade inata na COVID-19 em
questões associadas não apenas ao seu papel protetivo mas
também sua relação com o agravamento do quadro clínico
observado em alguns pacientes.

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
269
MATERIAIS E MÉTODOS
O estudo está caracterizado como uma revisão de
literatura cuja metodologia empregada baseou-se em dados
provenientes de fontes relacionadas aos aspectos clínicos,
epidemiológicos e imunológicos associados, principalmente, à
COVID-19. O critério de inclusão para a escolha dos 38 artigos
científicos aqui inseridos foram a escrita em língua inglesa com
período de publicação de até 5 anos. Todos os artigos utilizados
no capítulo foram coletados dos bancos de dados online
Elsevier, Pubmed e Google Scholar, mediante aplicação de
palavras-chave associadas aos assuntos de interesse como,
“epidemiologia, imunologia, SARS-CoV-2, imunidade inata e
imunopatologia”. Objetivando a aquisição de dados atualizados
acerca de número de infectados e mortos durante a pandemia,
foram consultados apenas páginas confiáveis associadas aos
governos Federal e Estadual como o Ministério da Saúde e
Secretaria da Saúde, respectivamente bem como de entidades
internacionais como Organização Mundial da Saúde (WHO).
Por fim, para a obtenção de informações básicas associadas ao
campo imunológico, fez-se uso de livro referência na área da
imunologia, Imunologia Celular e Molecular, 9° edição.
RESULTADOS E DISCUSSÃO
EPIDEMIOLOGIA
Com o início da pandemia, uma grande reportação de
casos de COVID-19 ocorreu nos EUA, França, Alemanha,
Espanha e Itália, sendo esta a mais afetada após do surto na
China (YUKI et. al. 2020). Até a presente data, já foram

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
270
notificados mais de 54 milhões de casos com um número de
óbitos ultrapassando a marca de um milhão em todo o mundo
(WHO, 2020).
No Brasil, o primeiro registro de COVID-19 ocorreu em
26 de fevereiro havendo, posteriormente, uma rápida
disseminação do vírus no país no qual, em meados de
novembro, quase 6 milhões de indivíduos já foram infectados.
Igualmente preocupante, o número de óbitos, por sua vez,
ultrapassou a marca de 160.000 (WHO, 2020). Analisando as
regiões brasileiras, Sudeste e Nordeste são as mais afetadas
pela COVID-19. De acordo com o Ministério da Saúde, no dia
14 de agosto de 2020, a região Nordeste alcançou a marca de
mais de um milhão de infectados com um saldo de 32 mil
mortes. Até 17 de novembro, foram notificados 1.547.926 casos
enquanto que 43.450 pessoas já perderam a vida frente a
infecção pelo SARS-CoV-2.
ESTRUTURA E INTERAÇÃO VÍRUS-CÉLULA
O SARS-CoV-2 pertence a família Coronaviridae e ao
gênero Betacoronavirus no qual também estão inseridos o
SARS-CoV e MERS-CoV (ICTV, 2020). Seu genoma é
caracterizado por ser um RNA de fita simples com sentido
positivo codificador de proteínas não estruturais e estrurais no
qual, dentres estas, destaca-se a proteína do envelope (E),
proteína de membrana (M) e a do nucleocapsídeo (N) e a
proteína Spike (S) (LI et. al. 2020). Esta, por sua vez, é de
fundamental importância no processo de infecção celular
mediante sua interação com o receptor da enzima conversora
de angiotensina 2 (ECA2) (LAN et. al. 2020). O fusionamento
do vírus na célula é reforçado pela serina protease

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
271
transmembranar 2 (TMPRSS2), uma estrutura altamente
expressa nos pulmões e ramos bronquiais, na qual atua como
um cofator de entrada do SARS-CoV-2 ao clivar e ativar a
proteína S facilitando, assim, a interação desta com a ECA2 (LI
et. al. 2020).
Uma vez na célula, o RNA viral dará origem aos
principais componentes do SARS-CoV-2 no qual a estrutura S
determina a infectividade do vírus e sua transmissibilidade para
o hospedeiro. Estruturalmente, essa proteína é composta pelas
subunidades S1 e S2 na qual a primeira apresenta é formada por
domínios N-terminal (NTD) e C-terminal (CTD) sendo este, no
caso dos coronavírus (CoV), utilizado como região de interação
com o ECA2, A associação do S1 com a ECA2 faz com que haja
uma dissociação da S2 e esse passe de uma forma pré-fusão
para uma forma pró-fusão, uma mudança conformacional
essencial para a fusão de membrana. Além disso, sendo alvo
dos anticorpos neutralizantes além de ser um dos principais
promotores das respostas imunes protetoras, a proteína S
tornou-se alvo da maioria das vacinas em desenvolvimento
(LAN et. al. 2020; LI et. al. 2020; OU et al., 2020; YUKI et. al.
2020).
FORMAS DE TRANSMISSÃO E SINAIS E SINTOMAS
Uma das principais formas de propagação da COVID-19
ocorre mediante perdigotos expelidos durante a fala, espirro ou
tosse de indivíduos infectados. Em aspectos comparativos, a
exposição prolongada a grupos sintomáticos parece apresentar
maior risco de contaminação comparado aos indivíduos
assintomáticos. Além disso, devido a ausência temporária de
sinais clínicos, indivíduos pré-sintomáticos também

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
272
compreendem um importante fator de transmissibilidade viral.
Com relação a transmissão vertical (que ocorre da mãe para o
filho durante a gestação) relatos de mulheres infectadas cujos
filhos nasceram saudáveis sugerem um menor risco para essa
forma de propagação. Não menos importante, a contaminação
pode ocorrer também pela presença de aerossóis suspensos
bem como o contato com superfícies contaminadas no qual
nestas, a duração das partículas virais parece variar
dependendo da composição da superfície. (WIERSINGA et. al.
2020).
Uma vez no organismo, o vírus passa por um período de
incubação de aproximadamente 5 dias (2-7) no qual, logo após,
os indivíduos podem apresentar sinais clínicos bastante
variados com duração de cerca de 14 dias (WIERSINGA et. al.
2020). Os sintomas mais comuns incluem febre, tosse seca,
falta de ar, fadiga, nauséa, diarreia e mialgia. Ademais, alguns
sintomas isolados como manifestações gastrointestinais e falta
de paladar e olfato podem ser observados sendo, os dois
últimos, bem característicos remetendo a infecção pelo SARS-
CoV-2. Felizmente, uma grande parcela dos acometidos
desenvolvem sintomas leves, enquanto uma minoria apresenta
um quadro mais crítico. Tais complicações podem acometer
órgãos como o coração, pulmão, cérebro, pâncreas, rim além
do sistema de coagulação (CHINA-CDC, 2020; WHO, 2020;
WIERSINGA et. al. 2020).
DIAGNÓSTICO
Atualmente há algumas formas de detecção da infecção
pelo SARS-CoV-2. No entanto, o RT-qPCR (Reação em Cadeia
de Polimerase - Transcrição Reversa quantitativa) se enquadra

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
273
como o diagnóstico padrão pela sua alta sensibilidade.
Importante destacar que este teste é indicado a ser realizado
até 7 dias após o inicio dos sintomas em virtude da diminuição
da carga viral. Assim, alguns fatores influenciam nos falsos
resultados como a técnica empregada na coleta, tempo de
exposição e a fonte da amostra sendo, o fluido broncoalveolar,
de extrema relevância.(WIERSINGA et. al. 2020).
Além do RT-qPCR, testes sorológicos também são
utilizados para detectar infecções pelo novo coronavírus. O
período para a coleta da amostra biológica é de extrema
importância para um resultado confiável. Nesse contexto,
anticorpos IgM são detectáveis dentro de 5 dias apresentando,
no entanto, altos níveis entre 2 a 3 semanas da doença. O
anticorpo IgG, por sua vez, é detectável após 14 dias após o
início dos sintomas. (WIERSINGA et. al. 2020).
Por fim, a elevação na proteína C-reativa, lactato
desidrogenase, alanino aminotransferase e aspartato
aminotransferase são marcadores prognósticos clínicos que
também podem indicar a infecção pelo COVID-19. Indivíduos
acometidos pela SARS-CoV-2 geralmente apresentam quadro
de linfopenia, trombocitopenia e valores elevados do D-dímero.
No entanto, todos esses achados laboratoriais se assemelham
a um quadro de pneumonia, sendo necessária a conjugação
com outras metodologias de diagnóstico (WIERSINGA et. al.
2020).
SISTEMA IMUNE INATO E COVID-19
Células Mielóides
Compondo cerca de 10% de células nucleadas
presentes no sangue humano, monócitos são uma população

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
274
heterogênia de fagócitos circulantes que, ao migrarem para os
tecidos se diferenciam em macrófagos (GINHOUX; JUNG,
2014; ABBAS; LITCHMAN; PILLAI, 2019). Ambas
desempenham importante ação moduladora de respostas
inflamatórias podendo, no entanto, apresentar participação no
desenvolvimento de imunopatologias (GINHOUX; JUNG, 2014;
MA et al., 2019).
Acerca da COVID-19, embora uma parcela significativa
de indivíduos apresentarem manifestações clínicas leves,
alguns pacientes desenvolvem complicações no qual sistema
imune desregulado pode contribuir com o quadro. Nesse
contexto, monócitos e macrófagos parecem desempenhar um
importante papel na gravidade da doença mediante alterações
na frequência celular além da síntese de mediadores
inflamatórios (MERAD; MARTIN, 2020).
A análise do líquido de lavagem broncoalveolar (LLB) de
indivíduos acometidos pela doença mostrou um aumento
significativo de quimiocinas envolvidas com o recrutamento de
monócitos, como CCL2 e CCL7, no qual macrófagos parecem
ser uma importante fonte em indivíduos com agravamento do
quadro clínico (LIAO et al., 2020; ZHOU et al., 2020). Tais
observações corroboram com o expressivo aumento de
monócitos derivados de macrófagos pro-inflamatórios em
indivíduos com a forma grave da infecção (LIAO et al., 2020).
No entanto, embora o número de monócitos totais pareça
não ser alterado, a modulação em subpopulações destas
células são observadas em sangue periférico de indivíduos
infectados pelo SARS-CoV-2. Nesse contexto, Zhang e
colaboradores (2020) observaram um aumento expressivo de
monócitos com fenótipo intermediário (CD14++CD16+) e não

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
275
clássico (CD14++CD16++) em detrimento a redução do perfil
clássico (CD14++CD16-). De modo semelhante, um estudo
desenvolvido por Zhou e colegas (2020) revelou aumento no
número de monócitos CD14+CD16+, principalmente naqueles
que apresentaram a forma grave.
Não menos importante, pesquisas mostram que,
principalmente, indivíduos com um quadro clínico mais grave da
COVID-19 podem desenvolver a síndrome da tempestade de
citocinas. Esta é caracterizada por uma produção exacerbada
de citocinas mediante um processo de desbalanço da resposta
imune frente a diferentes estímulos e, no âmbito da infecção
pelo SARS-CoV-2, monócitos parecem contribuir com esse
quadro (TANG et al., 2020; ZHANG et al., 2020). Zhou e
colaboradores (2020) revelaram um crescimento de monócitos
produtores de IL-6 nas formas moderada e grave da doença, de
modo mais expressivo para este último. Ademais, monócitos
produtores de GM-CSF também aumentaram em ambas as
formas sugerindo uma importante contribuição dessas células
no processo mencionado.
Células Linfóides Inatas (ILC)
Células linfóides inatas são componentes de
fundamental papel na homeostase tecidual bem como nas
respostas imunes. Esse grupo celular é formado por
subpopulações leucocitárias distinguidas mediante expressão
de receptores, síntese de citocinas e fatores de transcrição.
Curiosamente, algumas carcaterísticas fenotípicas inerentes as
ILCs assemelham-se a grupos celulares componentes da
imunidade adaptativa, os linfócitos T, demonstrando algum grau

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
276
de equivalência entre os grupos supracitados (BAL,
GOLEBSKI, SPITS, 2020).
Dentre as ILCs, as células Natural Killers (NKs), ou
assassinas naturais correspondem a cerca de 5%-15% dos
leucócitos nucleados presentes no sangue e no baço. Seu
nome remete ao papel citotóxico desempenhado por elas
sendo, portanto, importantes na eliminação de fontes de
infecções virais e bacterianas. Além da atividade citotóxica, as
NKs são fonte de citocinas pró-inflamatórias, como IFN-γ, que
aumentam a capacidade microbicida desempenhada por
macrófagos (ABBAS; LITCHMAN; PILLAI, 2019).
De modo semelhante aos monócitos, infecções pelo
SARS-CoV-2 podem alterar alguns aspectos inerentes as NKs.
Com relação à frequência populacional, estudos mostram
resultados variados acerca de alterações no número dessas
células na infecção pelo novo coronavírus (SOKOLOWSKA et
al., 2020). No entando, a redução na frequência de NKs no
sangue periférico de indivíduos acometidos pela COVID-19
parece estar relacionada com as formas mais graves (VABRET
et al., 2020).
Além da frequência celular, a análise fenotípica na
doença revela alterações significativas na proporção de
subpopulações de NKs bem como na funcionalidade das
mesmas. Alguns estudos observaram não apenas uma
acentuada redução no número destas células no sangue
periférico de pacientes graves como houve uma considerável
atenuação nas subpopulações CD56dimCD16+ e
CD56brightCD16- (KURI-CERVANTES et al., 2020; WILK et al.,
2020). Com fundamental ação citotóxica as primeiras estão
presentes em elevadas proporções no sangue periférico em

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
277
detrimento das CD56brightCD16-. Estas, por sua vez, modulam
as respostas imunes principalmente mediante a síntese de
citocinas como IFN-γ e IL-10 (ORTEGA-RODRÍGUEZ et al.,
2020).
Em conjunto com os dados anteriormente mencionados,
marcadores relacionados com a atividade citotóxica além da
expressão de citocinas IFN-γ e TNFα encontraram-se reduzidas
em sangue periférico de pacientes infectados (ZHENG et al.,
2020). No entando, tanto as células NKs totais quanto as
subpopulações CD56dim e CD56bright apresentaram
característica fenotípica de células ativadas não havendo,
porém, diferenças significativas entre as formas moderada e
grave da doença (MAUCOURANT et al., 2020).
Como observado, alguns dados demonstram que as
células NKs podem ter suas funções efetoras atenuadas
durante a fase aguda da COVID-19. Esse processo pode estar
relacionado ao crescimento na expressão de proteínas
envolvidas com a modulação de suas atividades. Nesse
contexto, o aumento na síntese de receptores inibitórios a
exemplo do LAG-3, TIM-3 e NKG2A possivelmente contribuem
com os resultados já explicitados (VABRET et al., 2020).
Portanto, analisando as células NKs frente a infecção pelo
SARS-CoV-2, observa-se que esta pode desencadear
modulação celular favorável para a evasão do vírus no
organismo.
Em consonância com o supracitado, é observado que
durante a infecção pelo novo coronavírus, pode ocorrer um
processo de infiltração celular pulmonar, observado também em
monócitos/macrófagos, o qual pode estar relacionado com o
agravamento do quadro clínico do paciente. Ao contrário do

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
278
observado em alguns dados do sangue periférico, um aumento
na frequêcia de células NKs no LLB foi demonstrado.
Curiosamente, pacientes mais graves apresentaram menor
frequência comparado a indivíduos com a forma moderada
(LIAO et al., 2020).
Em conjunto com as NKs, o grupo das ILCs é integrado
por três subpopulações celulares diferenciadas mediante
presença de fatores de transcrição e síntese de citocinas. As
ILC1 são caracterizadas pela síntese de INF-γ e do fator de
transcrição Tbet. As ILC2 apresentam o GATA-3 como fator de
transcrição e produzem as citocinas IL-4, IL-5 e IL-13. Por fim,
as ILC17 são produtoras de IL-22 e IL-17 sendo o RORγt,
produzido pelas mesmas (Th1, Th2 e Th17) (ABBAS;
LITCHMAN; PILLAI, 2019; BAL, GOLEBSKI, SPITS, 2020).
No contexto de infecções desencadeadas pelo SARS-
CoV-2, embora ainda não se tenha dados robustos envolvendo
as ILCs logo acima mencionadas, estudos mostram que
indivíduos com forma grave da COVID-19 apresentaram
redução na frequência destas células em sangue periférico. No
entanto, de modo semelhante as NKs, o processo de
recuperação parece conduzir a um retorno dos níveis basais
das ILCs como observado em comparações realizadas entre os
grupos infectado, recuperado e controle (KURI-CERVANTES et
al., 2020). Contudo, mais estudos são necessários para melhor
compreender o papel dessas células em infecções pelo SARS-
CoV-2.
Granulócitos
Granulócitos são células do sistema imune inato, de
origem mielóide, que apresentam como característica

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
279
morfológica a presença de grânulos em seu citoplasma e
desenvolvem funções vitais para a resposta imune. Se
enquadram dentro deste grupo os neutrófilos, eosinófilos e
basófilos presentes nos tecidos e corrente sanguínea. Os
mastócitos, por sua vez, estão concentrados no pulmão, vias
aéreas/trato respiratório, trato gastrointestinal, pele, passagem
nasal e meninges (ABBAS; LICHTMAN; PILLAI, 2019;
KEMPURAJ et al., 2020).
Os mastócitos são células imunes inatas que também
participam de respostas adaptativas. Esses leucócitos estão
envolvidos em infecções virais, doenças inflamatórias
sistêmicas, asma, doenças neuroinflamatórias, traumatismo
cranioencefálico (TCE), acidente vascular cerebral e vários
transtornos de estresse. Essas células participam da resposta
protetora, bem como de uma resposta deletéria e
desempenham um papel significativo na primeira linha de
defesa contra vírus e bactérias (KEMPURAJ et al., 2020).
Durante a infecção, o SARS-CoV-2 pode ativar os
mastócitos presentes no trato respiratório na fase inicial da
doença. Como consequência, essas células liberam várias
moléculas pró-inflamatórias, incluindo histamina, triptase, IL-1β,
CCL2, IL-6, GM-CSF e TNF-α, que estão implicadas na doença
COVID-19, levando ao estado mais grave. (KEMPURAJ et al.,
2020).
Em um estudo realizado em pacientes com COVID-19
leve/moderado e grave, a imunofenotipagem de leucócitos de
sangue total foi realizada em indivíduos na admissão em
enfermaria ou unidade de terapia intensiva e em controles
saudáveis. Marcadores granulocíticos (neutrófilos, eosinófilos e
basófilos) foram analisados. Houve contagens aumentadas de

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
280
neutrófilos CD15 + CD16 +, expressão granulocítica diminuída
de integrina CD11b e regulação negativa de CRTH2
relacionada a Th2 em eosinófilos e basófilos. A gravidade foi
associada ao surgimento da expressão do ponto de verificação
PDL1 em basófilos e eosinófilos (VITTE et al., 2020).
Os eosinófilos, por sua vez, são leucócitos circulantes e
residentes em tecidos que têm efeitos pró-inflamatórios
potentes em várias doenças. Essas células mostraram ter
várias outras funções, incluindo imunorregulação e atividade
antiviral sendo os níveis celulares variáveis em diversos
cenários clínicos (LINDSLEY et al., 2020). No contexto da
COVID-19, dados da literatura demonstram a presença de
leucocitose, com contagens totais de neutrófilos aumentadas,
associada a apresentações mais graves de COVID-19 (LI et al.,
2020; LIU et al., 2020; MO et al ., 2020; QIN et al., 2020). Porém,
Liu Fang et al (2020) relataram que mais da metade dos
pacientes admitidos com COVID-19 tinham eosinopenia, onde
os níveis de eosinófilos melhoraram em todos os pacientes
antes da alta, sugerindo que a resolução da eosinopenia pode
ser um indicador de melhora do estado clínico (LINDSLEY et
al., 2020).
Por fim, os neutrófilos, também chamados de leucócitos
polimorfonucleares, constituem a população mais abundante de
células brancas sanguíneas circulantes e medeiam as fases
iniciais das reações inflamatórias (ABBAS; LITCHMAN; PILLAI,
2019). Evidências indicam que a resposta desregulada ao
SARS-CoV-2 se estende aos neutrófilos na manifestação grave
da COVID-19. Uma contagem absoluta elevada de neutrófilos
e a razão neutrófilos/linfócitos são prognósticos para SRAG
(Síndrome Respiratória Aguda Grave) e morte em COVID-19.

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
281
(RUAN et al., 2020; DEL VALLE et al., 2020). Além disso,
pacientes com COVID-19 grave também têm níveis
aumentados de armadilhas extracelulares (NETs) de
neutrófilos, que são compostas de material de DNA com
antimicrobianos e enzimas oxidantes expelidas por essas
células para controlar infecções. É importante ressaltar que as
NETs podem contribuir para o desenvolvimento de lesão
pulmonar e microtrombos na COVID-19 (GUSTINE; JONES et
al., 2020)
Em conjunto com os dados acima mencionados, a
proporção de neutrófilos-linfócitos (NLR) mostrou servir como
um indicador confiável de COVID-19 grave. A alta proporção da
relação NLR leva a complicações graves nos pacientes
infectados, como fadiga, tosse seca, febre, pneumonia,
infecção respiratória aguda, insuficiência de órgãos e baixa
qualidade de vida. Por outro lado, a manutenção das
concentrações sanguíneas de NLR pode melhorar a resposta
ao comprometimento do sistema imunológico. Portanto, o NLR
é um marcador útil na rotina clínica para rastrear o prognóstico
em pacientes infectados com COVID-19 (PIMENTEL; VEGA;
LAVIANO, 2020).
Finalmente, um estudo feito por Barnes e colaboradores
(2020) em uma amostra de autópsia dos pulmões de um
paciente que foi a óbito por COVID-19, foi observado infiltração
neutrofílica extensa nos capilares pulmonares, com capilarite
aguda, deposição de fibrina e extravasamento para o espaço
alveolar. Uma imagem foi escolhida para enfatizar as lesões
dos capilares e mucosite neutrofílica da traqueia. Todas as vias
aéreas foram afetadas. A neutrofilia prediz, portanto, resultados

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
282
ruins em pacientes com COVID-19 e a proporção de neutrófilos
para linfócitos é um fator de risco para doença grave.
Sistema Complemento
Componentes da imunidade inata, o sistema
complemento consiste um conjunto de proteínas plasmáticas
cujas funções assemelham-se, em alguns aspectos, aos
desenvolvidos pelos anticorpos. O processo de ativação ocorre
através de três vias (clássica, alternativa e da lectina) no qual,
em todas, uma cascata proteolíca é requerida. Uma vez
ativada, moléculas do complemento passam a desempenhar
seu papel na resposta imune no qual inclui opsonização e
eliminação de algumas estruturas além do recrutamento celular
aos sítios de infecção (ABBAS; LITCHMAN; PILLAI, 2019).
O papel desse sistema na COVID-19 vem sendo
estudada e algumas pesquisas sugerem a participação de
proteínas do complemento na doença. Biópsia de tecido
pulmonar ou da pele de 5 pacientes com a forma grave da
patologia apresentando falha respiratória e erupções cutâneas
purpúricas (3 pacientes) revelou um acúmulo de (MASP)2, C5b-
9 e C4d sendo, os dois últimos, também acumulados em análise
epitelial (SOKOLOWSKA et al., 2020). Além disso, um estudo
comparativo conduzido por Magro et al (2020) revelaram um
aumento considerável dos níveis séricos da proteína C5a em
pacientes graves da doença. A melhora no quadro clínico de
dois pacientes graves tratados com anticorpos monoclonais
anti-C5a sugerem um importante papel de proteínas do
complemento na COVID-19 (MAGRO et al., 2020).

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
283
CONCLUSÕES
Por fim observa-se que, embora de fundamental
importância no combate a microorganismos patogênicos, a
resposta desencadeada pela imunidade inata, quando
desregulada, pode se associar ao agravamento do quadro
clínico de uma doença. Como observado nos estudos aqui
explicitados, tais alterações, sejam em células circulantes ou
presentes em órgãos, somados a análises fenotípicas, podem
auxiliar no prognóstico de gravidade da COVID-19. Assim, tais
pesquisas podem contribuir na escolha de medidas mais
eficazes para o tratamento da patologia além de auxiliar no
desenvolvimento de terapêuticas ideiais.
REFERÊNCIAS BIBLIOGRÁFICAS ABBAS, Abul K.; LICHTMAN, Andrew H.; PILLAI, Shiv. Imunologia celular e molecular. Elsevier Brasil, 2019. BAL, Suzanne. M.; GOLEBSKI, Korneliusz; SPITS, Hergen. Plasticity of innate lymphoid cell subsets. Nature Reviews Immunology, v. 20, p. 1-14, 2020. BARNES, Betsy J. et al. Targeting potential drivers of COVID-19: Neutrophil extracellular traps. Journal of Experimental Medicine, v. 217, n. 6, p. 1-8, 2020. CHINA-CDC. China Centers for Disease Control and Prevention. Global Health-China. Disponível em: https://www.cdc.gov/globalhealth/countries/china/default.htm. Acesso em: 15.10.2020. DEL VALLE, Diane Marie et al. An inflammatory cytokine signature helps predict COVID-19 severity and death. medRxiv, 2020. DOI: https://doi.org/10.1101/2020.05.28.20115758 GINHOUX, Florent; JUNG, Steffen. Monocytes and macrophages: developmental pathways and tissue homeostasis. Nature Reviews Immunology, v. 14, n. 6, p. 392-404, 2014.

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
284
GUSTINE, Joshua N.; JONES, Dennis. Immunopathology of Hyperinflammation in COVID-19. The American Journal of Pathology, 2020. DOI:https://doi.org/10.1016/j.ajpath.2020.08.009 ICTV. International Committee on Taxonomy of Viruses. Disponível em: https://talk.ictvonline.org/taxonomy/. Acesso em: 15.10.2020 IBRAHIM, Ibrahim M. et al. COVID-19 spike-host cell receptor GRP78 binding site prediction. Journal of Infection, v. 80, n. 5, p. 554-562, 2020. KEMPURAJ, Duraisamy et al. COVID-19, mast cells, cytokine storm, psychological stress, and neuroinflammation. The Neuroscientist, v. 26, p. 402-414, 2020. KURI-CERVANTES, Leticia. et al. Immunologic perturbations in severe COVID-19/SARS-CoV-2 infection. bioRxiv, 2020. LAN, Jun et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature, v. 581, n. 7807, p. 215-220, 2020. LI, Xiaochen et al. Risk factors for severity and mortality in adult COVID-19 inpatients in Wuhan. Journal of Allergy and Clinical Immunology, v. 146, n. 1, p. 110-118, 2020. LI, Qianqian et al. The impact of mutations in SARS-CoV-2 spike on viral infectivity and antigenicity. Cell, v. 182, n. 5, p. 1284-1294. e9, 2020. LIAO, Mingfeng. et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nature medicine, v. 26, n. 6, p. 1-3, 2020. LINDSLEY, Andrew W.; SCHWARTZ, Justin T.; ROTHENBERG, Marc E. Eosinophil responses during COVID-19 infections and coronavirus vaccination. Journal of Allergy and Clinical Immunology, v. 146, n. 1, p. 1-7, 2020. LIU, Fang et al. Patients of COVID-19 may benefit from sustained lopinavir-combined regimen and the increase of eosinophil may predict the outcome of COVID-19 progression. International Journal of Infectious Diseases, v. 95; p: 183–191, 2020. LIU, Yuwei et al. Neutrophil-to-lymphocyte ratio as an independent risk factor for mortality in hospitalized patients with COVID-19. Journal of Infection, v. 81, n. 1, p. e6-e12, 2020. MA, Wen-Tao. et al. The role of monocytes and macrophages in autoimmune diseases: a comprehensive review. Frontiers in immunology, v. 10, n.1140, p. 1-24, 2019. MAGRO, Cynthia. et al. Complement associated microvascular injury and thrombosis in the pathogenesis of severe COVID-19 infection: a report of five cases. Translational Research, v. 220, p. 1-13, 2020. MAUCOURANT, Christopher et al. Natural killer cell immunotypes related to COVID-19 disease severity. Science Immunology, v. 5, n. 50, p. 1-22, 2020.

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
285
MERAD, Miriam; MARTIN, Jerome C. Pathological inflammation in patients with COVID-19: a key role for monocytes and macrophages. Nature Reviews Immunology, v. 20, n. 6, p. 355-362, 2020. MELLA, Cesar et al. Innate immune dysfunction is associated with enhanced disease severity in infants with severe respiratory syncytial virus bronchiolitis. The Journal of infectious diseases, v. 207, n. 4, p. 564-573, 2013. MO, Pingzheng et al. Clinical characteristics of refractory COVID-19 pneumonia in Wuhan, China. Clinical Infectious Diseases, 2020. OU, Xiuyuan et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nature communications, v. 11, n. 1, p. 1-12, 2020. PIMENTEL, Gustavo D.; VEGA, Maria CM Dela; LAVIANO, Alessandro. High neutrophil to lymphocyte ratio as a prognostic marker in COVID-19 patients. Clinical Nutrition ESPEN, 2020. QIN, Chuan et al. Dysregulation of immune response in patients with COVID-19 in Wuhan, China. Clinical Infectious Diseases, v. 71, n. 15, p. 762-768, 2020. RODRÍGUEZ, Alma Celeste et al. Altered NK cell receptor repertoire and function of natural killer cells in patients with acute myocardial infraction: A three-month follow-up study. Immunobiology, v. 225, n. 3, p.1-10, 2020. RUAN, Qiurong et al. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive care medicine, v. 46, n. 5, p. 846-848, 2020. SOKOLOWSKA, Milena et al. Immunology of COVID‐19: mechanisms, clinical outcome, diagnostics and perspectives–a report of the European Academy of Allergy and Clinical Immunology (EAACI). Allergy, 2020. TANG, Yujun et al. Cytokine storm in COVID-19: the current evidence and treatment strategies. Frontiers in immunology, v. 11, n. 1708, p. 1-13, 2020. VABRET, Nicolas et al. Immunology of COVID-19: current state of the science. Immunity, v. 52, n.6, p. 910-941, 2020. VITTE, Joana et al. A granulocytic signature identifies COVID-19 and its severity. The Journal of Infectious Diseases, 2020. WIERSINGA, W. Joost et al. Pathophysiology, transmission, diagnosis, and treatment of coronavirus disease 2019 (COVID-19): a review. Jama, v. 324, n. 8, p. 782-793, 2020. WHO. World Health Organization. Coronavirus disease (COVID-19) pandemic. Disponível em: who.int/emergencies/diseases/novel-coronavirus-019?gclid=Cj0KCQjw28T8BRDbARIsAEOMBcz4gXwqpHZjIuaVSn3xSbwevkxXZ_uKdlkwMijN_X4aaAYY5QoJ2ZAaAkl7EALw_wcB. Acesso em: 15.10.2020.

CARACTERÍSTICAS DA COVID-19 E A MODULAÇÃO DA IMUNIDADE INATA
286
WILK, Aaron J. et al. A single-cell atlas of the peripheral immune response in patients with severe COVID-19. Nature Medicine, v. 26, n. 7, p. 1070-1076, 2020. WU, Di et al. The SARS-CoV-2 outbreak: what we know. International Journal of Infectious Diseases, v. 94, p. 44-48, 2020. YUKI, Koichi; FUJIOGI, Miho; KOUTSOGIANNAKI, Sophia. COVID-19 pathophysiology: A review. Clinical immunology, v. 215, p. 1-7, 2020. ZHANG, Dan et al. COVID-19 infection induces readily detectable morphological and inflammation-related phenotypic changes in peripheral blood monocytes, the severity of which correlate with patient outcome. medRxiv, 2020. ZHENG, Meijuan et al. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cellular & molecular immunology, v. 17, n. 5, p. 533-535, 2020. ZHOU, Yonggang et al. Pathogenic T-cells and inflammatory monocytes incite inflammatory storms in severe COVID-19 patients. National Science Review, v. 7, n. 6, p. 998-1002, 2020. ZHOU, Zhou et al. Overly exuberant innate immune response to SARS-CoV-2 infection. Cell Host & Microbe, p. 1-32, 2020.
AGRADECIMENTOS
Agradecemos aos órgãos de fomento CAPES e Fapesq-
PB, pelo apoio. Agradecemos também a todos os que ajudaram com a pesquisa de artigos científicos bem como na revisão do conteúdo aqui explicitado.

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
287
CAPÍTULO 14
CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
Fernando Cézar Comberlang Queiroz Davis dos SANTOS1
Pedro Henrique de Sousa PALMEIRA2
Leonardo Lima CARDOSO3
Bárbara Guimarães CSORDAS4 Tatjana Souza Lima KEESEN5
1 Graduando do curso de Biotecnologia, UFPB; 2Mestrando do Programa Multicêntrico de Pós-Graduação em Ciências Fisiológicas, UFPB; 3 Graduando do curso de Biotecnologia,
UFPB;4Doutoranda do Programa de Pós-Graduação em Produtos Naturais e Sintéticos Bioativos-PgPNBS-UFPB 5Orientadora/Professora do CBIOTEC/UFPB.
RESUMO: O surgimento do novo coronavírus denominado de SARS-CoV-2, agente causador da COVID-19, instaurou uma pandemia mundial. Descoberto no final de dezembro de 2019 a partir de supostos casos de pneumonia em Wuhan, China, até o presente momento já infectou mais de 35 milhões de pessoas no mundo. Pela sua recém-descoberta, os mecanismos fisiopatológicos envolvidos da doença e sua correlação com a imunidade adaptativa ainda são obscuros. No entanto, com o avançar das pesquisas se observou que essa desempenha importante papel frente à infecção, diante disso o trabalho objetivou compilar informações e estudos associados ao papel da imunidade adaptativa na COVID-19, por meio da revisão da literatura encontrada até o presente momento acerca do SARS-CoV-2. A imunidade adquirida é caracterizada por ser a linha de defesa específica (do corpo), abarcando ambas respostas humorais e celulares. A imunidade celular é o principal mecanismo de combate aos microrganismos intracelulares, como os vírus a exemplo do SARS-CoV-2. No entanto, diante à infecção causadora da COVID-19, diversos aspectos associados aos habituais mecanismos de ação dos linfócitos T

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
288
CD4, T CD8 e B sofrem alterações fenotípicas e populacionais, podendo contribuir com o amplo espectro de casos clínicos. A elucidação acerca da participação do sistema imune adaptativo na doença bem como na compreensão de uma memória protetiva após a resolução da patologia ainda não está totalmente elucidada. Palavras-chave: SARS-CoV-2. Imunidade adaptativa. Linfócitos T. Infecção. COVID-19.
INTRODUÇÃO
O sistema imune adaptativo (ou adquirido) é constituído
essencialmente por células de origem linfoide denominadas
linfócitos. Ativados mediante estímulos antigênicos e outros
mecanismos de feedback positivo, a imunidade adquirida pode
ser dividida em humoral, em que se tem a ação de células B
secretando anticorpos que neutralizarão o patógeno, e
imunidade mediada por células, sendo esta realizada por
linfócitos T auxiliares e citotóxicos, que promovem a modulação
de outros leucócitos e morte de células infectadas,
respectivamente (ABBAS; LICHTMAN; PILLAI, 2015).
Assim, a imunidade adquirida se adapta a infecção que
está sendo combatida, para que os agentes invasores sejam
neutralizados da melhor forma possível. Ademais, reinfecções
pelo mesmo patógeno gerarão respostas imunológicas mais
robustas e eficazes, devido à memória celular e especificidade
obtidas durante a primeira infecção (ABBAS; LICHTMAN;
PILLAI, 2015). Com isso, a manutenção da imunidade
adaptativa, tal como o bom funcionamento dos linfócitos T, é um
fator de extrema importância no combate a doenças
infecciosas.

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
289
O Coronavírus da Síndrome Respiratória Aguda Grave 2
(SARS-CoV-2), pertencente ao gênero betacoronavírus, é o
agente causador da Doença do Coronavírus 2019 (COVID-19),
que prejudica principalmente o trato respiratório, trazendo
sintomas como dispneia, tosse seca e em casos mais graves,
dificuldade respiratória (BOURGONJE et al., 2020).
Estudos relatam que infecções virais graves e
sustentadas podem ocorrer devido ao esgotamento funcional
de linfócitos T citotóxicos (ZHANG et al., 2019). Consoante a
isso, é verificado que a evolução da COVID-19 para estágios
mais graves da doença, pode estar associada à exaustão de
células T CD8+ e CD4+ (CHEN; WHERRY, 2020; ZHENG et
al., 2020).
Além disso, linfopenia também demonstrou ser uma
característica recorrente em pessoas com agravamento da
doença, sendo observada uma redução nas populações de
células T CD4+, CD8+ e subconjuntos de células B (KURI-
CERVANTES et al., 2020; TAN et al., 2020; YU et al., 2020).
Segundo Carsetti e colaboradores (2020) outra
observação importante no espectro de casos de COVID-19 é o
aumento passageiro, em indivíduos assintomáticos, das
Imunoglobulinas IgA, IgM e IgG, este, porém, em menor grau.
Em pacientes acometidos por COVID-19 grave, no entanto,
níveis altos e sustentados de IgA e IgG foram observados.
Ademais, marcadores inflamatórios se apresentam como
bons auxiliadores na previsão de quadros clínicos mais graves.
Dessa forma, estudos a respeito dos mesmos são essenciais
para o desenvolvimento de tratamentos precoces que garantam
uma maior chance de remissão da doença. (LUCAS et al.,
2020). Do mesmo modo, casos de pessoas em estágios de
convalescença e recuperação total também são investigados a

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
290
fim de se obter novas descobertas sobre os efeitos de longo
prazo que a doença pode trazer para o corpo.
Em suma, estudos que visem compreender o papel da
imunidade adaptativa na COVID-19, são essenciais para o
desenvolvimento de prognósticos e novas formas de manejo
clínico que possuam maior eficácia no tratamento da doença.
OBJETIVOS
Diante do exposto, o trabalho visa reunir algumas
informações já presentes na literatura acerca da correlação da
imunidade adaptativa com a infecção causada pelo SARS-CoV-
2. O trabalho tem como foco principal o papel da imunidade
adaptativa no combate da COVID-19, trazendo à tona os
mecanismos imunológicos envolvidos nos diferentes espectros
clínicos da doença.
MATERIAIS E MÉTODOS
Este trabalho se refere a uma revisão da literatura acerca
da COVID-19. A partir disso, foi realizada uma busca
documental em quatro bancos de dados eletrônicos: PubMed,
Nature, Cell, MedRxiv. Foram selecionados 42 artigos de língua
inglesa referente ao tema SARS-CoV-2, do período de
novembro de 2015 até outubro de 2020, no qual palavras-chave
associadas a aspectos imunológicos inerentes a resposta
adaptativa foram aplicadas. Dados epidemiológicos atualizados
foram adquiridos mediante buscas em sites do governo federal
e estadual bem como de instituições não governamentais como
a Organização Mundial de Saúde.

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
291
RESULTADOS E DISCUSSÃO
SISTEMA IMUNE ADAPTATIVO NA COVID-19
A infecção pelo SARS-CoV-2 provoca alterações
responsivas em células componentes do sistema imunológico.
No âmbito da imunidade adaptativa, compreende-se que
diversos aspectos associados aos mecanismos de ação dos
linfócitos T e B como frequência populacional, produção de
citocinas e expressão de receptores são modulados. Com isso,
as alterações no sistema imune ocasionadas pelo novo
Coronavírus podem levar a um amplo espectro clínico durante
seu processo infeccioso.
Células T e COVID-19
Os linfócitos T, são componentes do sistema imune
adaptativo que ao serem ativados mediante estímulo antigênico
proporcionado pelas células apresentadoras de antígeno
(APC), atuam através de vários mecanismos de sinalização.
Estes, por sua vez, são fundamentais no combate a patógenos,
na modulação da resposta imune além de conferir ao organismo
especificidade e memória imunológica após a resolução de uma
patologia (ABBAS; LICHTMAN; PILLAI, 2015).
Dentre as subpopulações de linfócitos, estão as células
T CD4+. Também denominadas auxiliares ou “T helper”, estas
células auxiliam na proliferação e diferenciação de outros tipos
de células T, além de ativarem células B, macrófagos e outros
leucócitos, promovendo estímulos. Por outro lado, células T
CD8+ são conhecidas pela sua função citotóxica, na qual
induzem a morte celular mediante o reconhecimento de células
alvo e posterior liberação de grânulos citolíticos. Além disso,
estes tipos específicos de linfócitos ativam a proteína FAS, que

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
292
induz apoptose nas células infectadas com vírus. (ABBAS;
LICHTMAN; PILLAI, 2015).
No contexto da COVID-19, pode-se observar a partir de
diversos estudos que os números de linfócitos T CD4+ e CD8+
de forma geral diminuem drasticamente (VABRET et. al., 2020;
SUN et. al., 2020; GANJI et. al., 2020; ODAK et. al., 2020). A
ocorrência de linfopenia, embora presente em todos os
espectros da doença, são mais proeminentes em pacientes
moderados e graves como observado por Fan Wang et. al.
(2020). Em sua pesquisa, a comparação do grupo controle com
pacientes acometidos pela COVID-19 demonstrou uma
diminuição de células T CD4+ e T CD8+, sendo tal perfil mais
acentuado no grupo de pacientes leves e graves.
Uma pesquisa realizada em pacientes acometidos pela
COVID-19 para observação da porcentagem de linfócitos no
sangue durante os dias de progressão da doença, observou
que indivíduos acometidos pelas formas leve e moderada
tiveram queda na porcentagem de linfócitos sanguíneos
seguida de posterior aumento. Já em pacientes que vieram a
óbito devido ao agravamento das complicações da doença,
houve diminuição progressiva da frequência de linfócitos, em
um tempo médio de 20 dias. A partir disso, sugere-se que a
linfopenia esteja estritamente relacionada com casos mais
graves de COVID-19 (TAN et al., 2020).
O quadro de linfopenia parece ter uma estreita relação
com os níveis das citocinas IL-6, IL-10, TNF-α e INF-I, sendo
estas capazes de inibir a recirculação de linfócitos T e promover
a retenção desses em tecidos linfoides e ligação ao endotélio
(VABRET et. al., 2020). Já em pacientes convalescentes o
número de linfócitos T retorna aos níveis basais enquanto que

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
293
o nível de citocinas pró-inflamantórias diminui, como observado
em um estudo desenvolvido por Fan Wang et. al. (2020).
O recrutamento de células T CD8+ nos sítios de infecção
propicia também a linfopenia, diminuindo a presença desses no
compartimento do sangue periférico. Em um estudo feito com o
lavado broncoalveolar (LBA) de um paciente que foi a óbito pela
COVID-19, foi revelado a presença aumentada de infiltrados
celulares como linfócitos T CD8+ em expansão clonal (VABRET
et. al., 2020).
É notório em pacientes acometidos pelo SARS-CoV-2
que as células T CD8+ são mais ativadas que as T CD4+
(VABRET et. al., 2020). Muitos estudos demonstram a alta
expressão de moléculas inibitórias nas CTLs (linfócitos T
citotóxicos), como o PD-1, TIM-3 e NKG2A, no qual esta é
diminuída após administração de terapia antiviral (PACES et.
Al., 2020). Zheng e colaboradores (2020) observaram a
redução na expressão de NKG2A em pacientes que se
encontravam em estágio de convalescença, levando a
recuperação funcional dos CLTs. Além disso, um úmero
reduzido de linfócitos T CD8+ CD28+, além de altas frequências
de células T CD8 PD-1/TIM-3 em pacientes em unidades de
tratamento intensivo também foi observado em um estudo feito
por Vabret e colaboradores (2020).
Apesar do esgotamento funcional das células CD8+ na
COVID-19, outros estudos apontam de forma alternativa para
uma hiperativação desses linfócitos, tal como os linfócitos T
CD8+ invariantes de mucosa (MAIT CD8+), sendo identificados
altos níveis de expressão de CD38, CD69 e HLA-DR nessas
células. Esses fatores podem implicar em respostas mais
agressivas de CLTs na COVID-19 (KURI-CERVANTES et al.,
2020; CHEN; WHERRY, 2020).

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
294
As células T CD8+ em pacientes acometidos pelo SARS-
CoV-2 apresentaram uma menor degranulação (diminuição da
externalização do CD107, marcador de citotoxidade) e também
diminuição na produção de IL-2, INF-γ e Granzima B quando
comparados com os pacientes controle. Em outro trabalho, foi
observado a alta produção de granzima B e perforina pelas
células T CD8+ em pacientes graves. Uma pesquisa feita por
Liao et. al. (2020) demonstrou que células T CD8+ retiradas do
LBA de pacientes graves expressaram genes citotóxicos como
GZMA, GZMB e GZMK em altos níveis, enquanto KLRC1 e
XCL1, genes também citotóxicos, têm destaque em pacientes
com quadros leves (VABRET et. al., 2020). Com isso, os dados
aqui explicitados sugerem que essas células são um excelente
fator de prognóstico da severidade da doença, como também
de uma eficácia clínica na COVID-19.
Em relação à funcionalidade de células T CD4+ na
COVID-19, há evidências de exaustão da função dos linfócitos
T auxiliares através de receptores inibitórios, tal como nas
CLTs, com concomitante hiper expressão de receptores de
ativação (CHEN; WHERRY, 2020).
Sobre os aspectos de reatividade cruzada entre
diferentes tipos de Coronavírus, foram encontradas células T
CD4+ reativas a glicoproteína Spike do SARS-CoV-2, usando a
expressão de marcadores de ativação 4-1BB e CD40-L, tanto
em pacientes acometidos pela COVID-19 quanto em pessoas
saudáveis. A presença de células CD4+ reativas em pessoas
não infectadas pelo SARS-CoV-2 pode indicar essa reatividade
cruzada, adquirida através do contato com outros tipos de
coronavírus que também infectam humanos (BRAUN et al.,
2020). Ademais, um estudo realizado para investigar as ações
de células T CD4+ na imunidade contra o vírus da SARS,

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
295
estabeleceu o direcionamento de células T em relação a 3
peptídeos derivados da proteína Spike do SARS-CoV-1, sendo:
S729-745,S358-374 e S 427-444 (LIBRATY et al., 2007). Os dados
dessas pesquisas podem ser de grande valia para futuros
estudos de reatividade cruzada entre linfócitos de memória
gerados pela infecção com SARS-CoV-1 e outros tipos de
Coronavírus, contra epítopos da proteína Spike de SARS-CoV-
2, a partir das suas semelhanças estruturais.
Por fim, mais estudos precisam ser realizados para
elucidar cada vez mais o papel das células T na imunidade
contra o SARS-CoV-2. Análises de exaustão funcional de
linfócitos T, superativação, diferenciação fenotípica, quadros de
linfopenia e reatividade cruzada com outros tipos de
coronavírus humanos, devem continuar sendo investigados
para o melhor entendimento das alterações fisiológicas que o
SARS-CoV-2 pode trazer ao organismo. Com isso, tais
pesquisas poderiam contribuir com o desenvolvimento de
alternativas mais eficazes no tratamento da doença.
Células B e anticorpos neutralizantes em pacientes
convalescentes na COVID-19
As células B estão no centro do sistema imune humoral
adaptativo e são responsáveis por mediar a produção de
imunoglobulina (Ig) específica para estruturas da superfície
(antígeno), dirigida contra patógenos invasivos (normalmente
conhecidos como anticorpos). As respostas de células B aos
antígenos (Ag), ocorrem quando as células FO B maduras
voltam a circular entre órgãos linfoides secundários em busca
de antígeno. Após esse encontro, as células B que recebem
ajuda de células T podem entrar em algumas possibilidades de

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
296
desenvolvimento diferenciado (COOPER, 2015; CROTTY,
2015).
As células B podem sofrer diferenciação plasmocítica,
formar plasmablastos extrafoliculares e formar células
plasmáticas secretoras de IgM. Essas células não têm genes Ig
mutados somaticamente e têm vida curta, mas fornecem uma
resposta inicial rápida ao antígeno. A segunda possibilidade de
desenvolvimento é o estabelecimento de um centro
germinativo, uma estrutura especializada dentro da qual as
células B sofrem rodadas de proliferação acompanhadas por
maturação de afinidade: um processo iterativo de mutação e
seleção do gene Ig resultando em um pool de células B que
podem se ligar a Ag com maior afinidade. Os anticorpos
também sofrem recombinação de troca de classe (COOPER,
2015; CROTTY, 2015).
Os anticorpos neutralizantes fazem parte da resposta
humoral do sistema imune adaptativo contra COVID-19, além
de outros vírus, bactérias intracelulares e toxinas microbianas.
Ao se ligarem especificamente aos Ag. Anticorpos
neutralizantes defendem a célula de um patógeno ou partícula
infecciosa, neutralizando qualquer efeito que tenha
biologicamente. A neutralização torna uma partícula não-
infecciosa ou o microrganismo perde a capacidade patogênica
(BROUWER et al., 2020; PAYNE, 2017).
Na população convalescente, a análise do repertório de
seis pacientes representativos do estudo de coorte (n=149),
revelou a presença de células B IgG+ e em expansão clonal.
Anticorpos neutralizantes potentes foram encontrados em
indivíduos, independentemente de sua capacidade geral de
neutralização do soro. Além disso, os anticorpos têm como alvo
epítopos neutralizantes para regiões antigênicas de SARS-

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
297
CoV-2. Esses achados indicam que, apesar da baixa
capacidade de neutralização do plasma, a maioria dos
indivíduos podem gerar anticorpos neutralizantes de IgG
robustos, independentemente da gravidade de seus sintomas
(UHL; CHAUVEAU, 2020).
Estudos em 17 pacientes, em que 14 amostras de
sangue (6-7 meses após a infecção), apresentaram atividades
neutralizantes significativas em um ensaio utilizando
pseudovírus (TAN et al., 2020). Foi possível detectar imunidade
humoral e celular específica para SARS-CoV-2 em oito
pacientes recém-dispensados. A análise de acompanhamento
em outra coorte de seis pacientes, 2 semanas após a alta
também revelou altos títulos de anticorpos IgG. Em todos os 14
pacientes testados, 13 apresentaram atividades de
neutralização do soro em um ensaio utilizando pseudovírus (NI
et al., 2020). Todos estes trabalhos puderam observar uma forte
correlação entre os títulos de anticorpos de neutralização e o
número de células T específicas para o vírus.
TRATAMENTO E VACINAS NA COVID-19: potenciais
estratégias contra ADE
O potencial terapêutico dos anticorpos monoclonais tem
sido bem reconhecido no tratamento de muitas doenças. Os
anticorpos monoclonais representam a principal classe de
bioterapêuticos para imunoterapia passiva no combate à
infecção viral (SHANMUGARAJ et al., 2020).
IL-6 é uma citocina que desempenha um papel
importante na reação inflamatória e resposta imunológica. As
experiências clínicas na China sugeriram que a IL-6 é uma das
citocinas mais importantes envolvidas nas tempestades de
citocinas induzidas pela COVID-19. Por esta razão,

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
298
tocilizumabe (TCZ), um anticorpo monoclonal humanizado
contra o receptor de interleucina-6 (IL-6R), foi recomendado em
pacientes graves e com IL-6 elevada. No entanto, existem
dados limitados sobre o efeito do TCZ sobre a atividade
inflamatória em pacientes com COVID-19.
Outras implicações tanto no tratamento quanto na
produção de vacinas contra COVID-19, devem ser ressaltadas.
Drogas baseadas em anticorpos e vacinas contra SARS-CoV-2
estão sendo aceleradas através do desenvolvimento pré-clínico
e clínico. Estudos anteriores sobre o vírus sincicial respiratório
e a vacina contra o vírus da dengue revelaram riscos de
segurança clínica em humanos relacionados a infecção
dependente de anticorpos (Antibody-dependent enhancement–
ADE), resultando em testes de vacinas fracassados. Os
mecanismos não manifestados à proteção dos anticorpos
contra qualquer vírus têm um potencial teórico para amplificar a
infecção ou desencadear imunopatologia prejudicial, tornando-
se a ADE uma preocupação geral para o desenvolvimento de
vacinas e terapias de anticorpos (ARVIN et al., 2020). Essa
possibilidade requer consideração cuidadosa, principalmente
no ponto crítico na pandemia da COVID-19. Estudos que
avaliem a probabilidade de revelar a ADE na patologia da
doença COVID-19 e que examinem como os dados podem
influenciar as intervenções clínicas são de extrema relevância.
As vacinas com um alto risco teórico de induzir ADE ou
ERD patológico incluem vacinas virais inativadas, que podem
conter alvos de antígeno não neutralizantes e/ou a proteína da
espícula viral de SARS-CoV-2 em conformações não
neutralizantes, fornecendo uma infinidade de alvos não
protetores para anticorpos que podem conduzir ao fracasso
vacinal (LEE et al., 2020).

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
299
As estratégias de imunização em humanos para SARS-
CoV-2 que induzem altos títulos de anticorpos neutralizantes
podem ter grande chance de sucesso e com risco mínimo de
ADE. Vacinas de subunidade que podem induzir anticorpos
neutralizantes específicos da proteína da espícula
(especialmente na conformação de pré-fusão, para reduzir a
apresentação de epítopos não neutralizantes) devem
apresentar riscos de ADE mais baixos (WRAPP et al., 2020).
Essas abordagens modernas de projeto de imunógenos devem
reduzir a imunopatologia potencial associada a anticorpos não
neutralizantes, aumentando a segurança das vacinas.
Biomarcadores na COVID-19
Os biomarcadores imunológicos têm sido fundamentais
no prognóstico do curso da COVID-19. Lucas et. al. (2020) fez
uma investigação acerca do aparecimento inicial de respostas
de citocinas e sua correlação com a severidade da doença.
Inicialmente foram obtidos três grupos distintos ao quadro da
patologia, sendo caracterizados por quatro assinaturas
imunológicas.
Pôde ser observado um exame abrangente da dinâmica
inflamatória diversificada durante COVID-19 e possíveis
contribuições de conjuntos distintos de mediadores
inflamatórios para a progressão da doença, com possibilidade
de que as intervenções imunológicas precoces os quais visam
marcadores inflamatórios são preditivos de pior evolução da
doença. Essas análises fornecem bases para um tratamento
mais direcionado de pacientes com COVID-19 com base em
marcadores precoces de citocinas, bem como terapias
destinadas a melhorar a reparação do tecido e promover a
tolerância à doença (LUCAS et. al., 2020).

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
300
Ademais, houve a percepção de altos níveis de IL-6 em
pacientes críticos, geralmente hospitalizados, sendo
associados à admissão na UTI e falha respiratória. Foi visto
também o aumento de IL-2R, IL-8, IL-10, e GM-CSF em
pacientes com quadros severos, porém estudos posteriores
serão necessários para elucidar os potenciais prognósticos
desses indicadores (VABRET et. al., 2020).
Geração de resposta da imunidade adaptativa em pacientes
convalescentes e recuperados
Como previamente mencionado, o processo de infecção
pelo SARS-CoV-2 fornece alterações fenotípicas e
populacionais podendo, em alguns aspectos, apresentar
durabilidade durante e após a fase de recuperação. Nesse
contexto, a compreensão acerca destas mudanças
imunológicas torna-se de fundamental importância na busca por
terapêuticas eficazes como o desenvolvimento de vacinas.
Em um estudo desenvolvido por Juno e colaboradores
(2020), foi demonstrado similaridade entre células T foliculares
circulantes (cTfh) entre indivíduos convalescentes e o grupo
controle. Embora um leve aumento de linfócitos T totais em
sangue periférico de indivíduos recuperados tenham sido
encontrados, alterações estatísticas significativas em células T
CD4 e T CD8 não foram observadas em amostras não
estimuladas com estruturas do SARS-CoV-2 (GRIFONI et al.,
2020). Esses resultados sugerem um possível retorno da
frequência celular de linfócitos aos níveis basais.
Uma pesquisa comparativa entre as fases inicial (FI) e
tardia (FT) de recuperação da COVID-19 sugeriram que os
últimos apresentavam maior frequência de células B e células
T comparado às primeiras. Ademais, subpopulações celulares

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
301
de células T auxiliares mostraram um aumento expressivo de
linfócitos T CD4 de memória central e atenuação das
subpopulações de memória efetora, células T reguladoras e
células T CD4 naive sendo, tais modificações, mais expressivas
em indivíduos FI (WEN et al., 2020).
A observância da presença de células específicas para
porções de um patógeno após a resolução da doença são
relevantes para a compreensão acerca do curso da mesma.
Amostras de células mononucleares do sangue periférico
(PBMC) de indivíduos recuperados da COVID-19 mostraram
uma frequência de 100% na presença de células T CD4
específicas para a proteína Spike (GRIFONI et al., 2020).
A proteína S demonstra ser umas das principais
estruturas proteícas do SARS-CoV-2 indutoras de respostas
adaptativas. Corroborando com os dados acima mencionados,
subpopulações de linfócitos T CD4 parecem ser moduladas
frente ao estímulo. Nesse contexto, as células cTfh bem como
o subtipo cTfh17 (CCR6+CXCR3-) apresentaram um aumento
considerável após exposição à estrutura Spike. Este resultado,
porém, não foi observado para a porção RBD (JUNO et al.,
2020). Em conjunto com os resultados supracitados, a proteína
Spike parece direcionar à presença as células T CD4 a um
padrão Th1 (GRIFONI et al., 2020).
O estímulo com proteínas não-Spike de PBMC de
pacientes convalescentes para COVID-19 induziram ao
aumento responsivo de células T CD4 com tendência ao perfil
Th1 (GRIFONI et al., 2020). De modo semelhante, Sekine e
colaboradores (2020) mostraram maior expressão de citocinas
características dessa subpopulação (IFN-γ, IL-2 e TNF) quando
estimuladas com peptídeos Spike, de membrana e do
nucleocapsídeo.

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
302
De fato, diferentes estruturas virais parecem conduzir a
distintas subpopulações celulares inseridas no grupo das
células T auxiliadoras. Enquanto as proteínas de membrana e
do nuclecapsídeo pareçam direcionar a um perfil Th1/Th17, a
sobreposição de peptídeos Spike inclinam ao subtipo cTfh em
pacientes convalescentes. A indução de células cTfh por
estrutura Spike sugere envolvimento chave na síntese de
anticorpos de alta afinidade (JUNO et al., 2020; SEKINE et al.,
2020). Corroborando com os estudos anteriormente expostos,
Sekine e colaboradores (2020) indicaram um aumento na
expressão de marcadores de ativação em células T CD4.
Não apenas os linfócitos T auxiliares apresentaram
resposta protetora mediante uma possível reexposição ao
SARS-CoV-2. O estímulo peptídico inserido em PBMC de
pacientes recuperados pela COVID-19 induziu os linfócitos T
CD8 (CLT) a alterações fenotípicas e populacionais. Um estudo
comparativo desenvolvido por Peng et al. (2020) demonstraram
que tanto a proteína Spike quanto as estruturas de membrana
(M) e do nucleocapsídeo (N) conduziram a uma maior
reatividade de linfócitos citotóxicos em indivíduos que
apresentaram a forma leve comparado com a forma grave.
A pesquisa acima mencionada também sugere uma
maior capacidade das estruturas M e N de conduzirem os
linfócitos T citotóxicos a apresentarem maior funcionalidade
(mediante expressão de múltiplas citocinas) quando comparado
a proteína Spike, em indivíduos recuperados que apresentaram
a forma leve. No entanto, não foram observadas alterações
estatísticas significativas na citotoxicidade nas formas leve e
grave dos voluntários (PENG et al., 2020).
A reatividade dos linfócitos CLT em indivíduos
recuperados também foi explicitada pela presença elevada de

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
303
células T CD8+ expressando CD69+ e CD137+, estes, usados
como marcadores de ativação celular, além da maior expressão
de IFN-γ. Uma expressiva quantidade de células T CD8
produtoras de IFN-γ também expressaram granzima e TNF
mostrando, assim, que as células de uma parcela considerável
de indivíduos geraram resposta contra o vírus após o desafio
com pools de peptídeos relacionados ao SARS-CoV-2
(GRIFONI et al., 2020). Em outro estudo foi possível observar
que células T CD4 desencadearam um aumento na produção
de INF-γ, ao serem estimuladas com proteínas S, M e N de
SARS-CoV-2. Além disso, foi revelado nos linfócitos T CD8 um
aumento na mobilização do marcador de degranulação CD107a
(SEKINE et al., 2020), sugerindo aumento na atividade
citotóxica dessas células.
Uma análise detalhada acerca das estruturas virais
responsáveis pela resposta de células T CD8 revelaram uma
distribuição variável. Embora a proteína S ainda se mostrasse
dominante com 26% da reatividade celular, as estruturas M
(com 22%), nsp6 (com 15%) e N (com 12%) também auxiliaram
na resposta. Por outro lado, 59% da reatividade de células T
CD4 se concentrou nas estruturas S, M e N sendo, a Spike,
mais prevalente com 27% (GRIFONI et al., 2020).
Por fim, compreender a dinâmica de ação desenvolvida
pelas células B durante e após o processo de recuperação de
doenças torna-se de fundamental importância. Assim como
observado para as células T os linfócitos B também parecem
ser modulados durante a fase de recuperação da COVID-19
como observado por Wen e colegas (2020). Nesse estudo,
células B plasmáticas estavam aumentadas enquanto que
linfócitos B naive sofreram atenuação.

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
304
Por outro lado, a análise sorológica de pacientes em fase
aguda bem como indivíduos recuperados mostram que nestes
os níveis de anticorpos IgM, IgG e IgA reduz significativamente
quando comparado com os primeiros. Além disso, a presença
de anticorpos anti-RBD parece decair em pacientes
recuperados como observado em 6 indivíduos, 2-3 meses após
a primeira coleta sanguínea (VAISMAN-MENTESH et al.,
2020).
Em contrapartida, a presença de células B de memória
específicas para a porção RBD parece se manter estável por
pelo menos 6 meses como observado em amostras de 5
indivíduos recuperados da infecção desencadeada pela SARS-
CoV-2 (VAISMAN-MENTESH et al., 2020)8. Esse estudo
sugere uma persistência de resposta humoral após a COVID-
19. No entanto, e não menos importante, o autor destaca a
necessidade de estudos futuros para compreender o papel
protetivo desses achados.
CONCLUSÕES
Semelhante a diversas patogenias, a infecção pelo
SARS-CoV-2 desencadeia alterações imunológicas
importantes que podem contribuir com o quadro clínico da
doença. Nesse contexto, diversos estudos sugerem uma
correlação entre alterações fenotípicas e populacionais de
componentes da imunidade adaptativa com as formas mais
graves da infecção. Porém, como demonstrado por estudos
aqui explicitados, observa-se que após a resolução da doença
uma possível reinfecção pode mobilizar linfócitos a uma
resposta mais rápida, na qual determinadas estruturas do
SARS-CoV-2 apresentam notável imunogeinicidade.

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
305
Por fim, mesmo com o direcionamento de uma parcela
expressiva das pesquisas científicas voltadas para o estudo da
COVID-19, diversas questões ainda precisam ser elucidadas.
Dentres estas, o entendimento acerca do desenvolvimento de
uma memória imunológica protetiva de longa duração será
esclarecida com o tempo. Assim, a compreensão dos aspectos
imunopatológicos inerentes à COVID-19 podem, de fato,
auxiliar no desenvolvimento de terapêuticas mais eficazes à
serem aplicadas no tratamento de pessoas acometidas pela
infecção.
REFERÊNCIAS BIBLIOGRÁFICAS ABBAS, A. K.; LICHTMAN, A. H.; PILLAI, S. Imunologia Celular e Molecular. 8. ed. Rio de Janeiro: Elsevier Editora Ltda, 2015. Cap. 1, p. 21-47. ABBAS, Abul K.; LICHTMAN, Andrew H.; PILLAI, Shiv. Imunologia celular e molecular. Elsevier Brasil, 2019. ALI, Amanat; VIJAYAN, Ranjit. Dynamics of the ACE2–SARS-CoV-2/SARS-CoV spike protein interface reveal unique mechanisms. Scientific reports, v. 10, n. 1, p. 1-12, 2020. ARVIN, A. M. et al. A perspective on potential antibody-dependent enhancement of SARS-CoV-2. Nature, v. 584, n. 7821, p. 353-363, 2020. BOURGONJE, Arno R. et al. Angiotensin‐converting enzyme‐2 (ACE2),
SARS‐CoV‐2 and pathophysiology of coronavirus disease 2019 (COVID‐19). The Journal of Pathology, 2020. BRAUN, Julian et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature, v. 2020, 2020. BROUWER, P. et al. Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. Science, v. 369, n. 6504, p. 643-650, 2020. CARSETTI, Rita et al. Spectrum of innate and adaptive immune response to SARS CoV 2 infection across asymptomatic, mild and severe cases; a longitudinal cohort study. medRxiv, 2020. CHEN, Zeyu; WHERRY, E. John. T cell responses in patients with COVID-19. Nature Reviews Immunology, p. 1-8, 2020. COOPER, M. D. The early history of B cells. Nature Reviews Immunology, v. 15, n. 3, p. 191-197, 2015.

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
306
CROTTY, S. A brief history of T cell help to B cells. Nature Reviews Immunology, v. 15, n. 3, p. 185-189, 2015. GANJI, Ali et al. Increased expression of CD8 marker on T-cells in COVID-19 patients. Blood Cells, Molecules, and Diseases, p. 102437, 2020. GRIFONI, Alba et al. Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals. Cell, v. 181, n. 7, p. 1489-1501, 2020. HUANG, Chaolin et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. The lancet, v. 395, n. 10223, p. 497-506, 2020. JUNO, Jennifer A. et al. Immunogenic profile of SARS-CoV-2 spike in individuals recovered from COVID-19. medRxiv, 2020. KURI-CERVANTES, Leticia et al. Comprehensive mapping of immune perturbations associated with severe COVID-19. Science Immunology, v. 5, n. 49, 2020. LAKE, Mary A. What we know so far: COVID-19 current clinical knowledge and research. Clinical Medicine, v. 20, n. 2, p. 124, 2020. LEE, W. S. et al. Antibody-dependent enhancement and SARS-CoV-2 vaccines and therapies. Nature microbiology, v. 5, n. 10, p. 1185-1191, 2020. LIBRATY, Daniel H. et al. Human CD4+ memory T-lymphocyte responses to SARS coronavirus infection. Virology, v. 368, n. 2, p. 317-321, 2007. LUCAS, Carolina et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature, v. 584, n. 7821, p. 463-469, 2020. MATHEW, Divij et al. Deep immune profiling of COVID-19 patients reveals distinct immunotypes with therapeutic implications. Science, v. 369, n. 6508, p. 1-19, 2020. NI, L.; et al. Detection of SARS-CoV-2-specific humoral and cellular immunity in COVID-19 convalescent individuals. Immunity, 2020. ODAK, Ivan et al. Reappearance of effector T cells is associated with recovery from COVID-19. EBioMedicine, v. 57, p. 102885, 2020. ORGANIZAÇÃO PAN-AMERICANA DA SAÚDE / ORGANIZAÇÃO MUNDIAL DA SAÚDE. Brasília: OPAS/OMS, 2020. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Folha Informativa COVID-19 – Escritório da OPAS e da OMS no Brasil. 7 de outubro, 2020. PACES, Jan et al. COVID-19 and the immune system. Physiological Research, 2020. PAYNE, S. Chapter 6-Immunity and Resistance to Viruses. Viruses; Payne, S., Ed.; Academic Press: Cambridge, MA, USA, p. 61-71, 2017. PENG, Yanchun et al. Broad and strong memory CD4+ and CD8+ T cells induced by SARS-CoV-2 in UK convalescent individuals following COVID-19. Nature immunology, v. 21, n. 11, p. 1336-1345, 2020.

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
307
PHAN, Lan T. et al. Importation and human-to-human transmission of a novel coronavirus in Vietnam. New England Journal of Medicine, v. 382, n. 9, p. 872-874, 2020. SEKINE, Takuya et al. Robust T cell immunity in convalescent individuals with asymptomatic or mild COVID-19. Cell, v. 183, n. 1, p. 158-168, 2020. SHANMUGARAJ, B. et al. Perspectives on monoclonal antibody therapy as potential therapeutic intervention for Coronavirus disease-19 (COVID-19). Asian Pac J Allergy Immunol, v. 38, n. 1, p. 10-18, 2020. SUN, Da-wei et al. The underlying changes and predicting role of peripheral blood inflammatory cells in severe COVID-19 patients: a sentinel?. Clinica Chimica Acta, 2020. TAN, Li et al. Lymphopenia predicts disease severity of COVID-19: a descriptive and predictive study. Signal transduction and targeted therapy, v. 5, n. 1, p. 1-3, 2020. TAN, Y.; et al. Durability of neutralizing antibodies and T-cell response post SARS-CoV-2 infection. Frontiers of Medicine, p. 1-6, 2020. UHL, L. F. K.; CHAUVEAU, A. Neutralizing antibodies in convalescent patients. 2020. VABRET, Nicolas et al. Immunology of COVID-19: current state of the science. Immunity, 2020. VAISMAN-MENTESH, Anna et al. SARS-CoV-2 specific memory B cells frequency in recovered patient remains stable while antibodies decay over time. medRxiv, 2020. WANG, Fan et al. Characteristics of peripheral lymphocyte subset alteration in COVID-19 pneumonia. The Journal of infectious diseases, v. 221, n. 11, p. 1762-1769, 2020. WANG, Dawei et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus–infected pneumonia in Wuhan, China. Jama, v. 323, n. 11, p. 1061-1069, 2020. WEN, Wen et al. Immune cell profiling of COVID-19 patients in the recovery stage by single-cell sequencing. Cell discovery, v. 6, n. 1, p. 1-18, 2020. WRAPP, D. et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science, v. 367, n. 6483, p. 1260-1263, 2020. YU, Kuai et al. Dysregulated adaptive immune response contributes to severe COVID-19. Cell Research, v. 30, n. 9, p. 814-816, 2020. ZHANG, Ji-Yuan et al. Single-cell landscape of immunological responses in patients with COVID-19.Nature immunology, v. 21, n. 9, p. 1107-1118, 2020. ZHANG, Chao et al. NKG2A is a NK cell exhaustion checkpoint for HCV persistence. Nature communications, v. 10, n. 1, p. 1-11, 2019. ZHENG, Meijuan et al. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cellular & molecular immunology, v. 17, n. 5, p. 533-535, 2020.

CORRELAÇÃO DA IMUNIDADE ADAPTATIVA COM A COVID-19
308
ZHOU, Fei et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. The lancet, 2020. ZHOU, Y. et al. Pathogenic T-cells and inflammatory monocytes incite inflammatory storms in severe COVID-19 patients. National Science Review, v. 7, n. 6, p. 998-1002, 2020.
AGRADECIMENTOS
Gostaríamos de agradecer às agências de
fomento CAPES e Fapesq-PB, as quais estão financiando nossos estudos acerca do SARS-CoV-2 e todos que contribuíram com este trabalho.

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
309
CAPÍTULO 15
MACRÓFAGOS M1/M2 NA LEISHMANIOSE
Edson dos Santos SILVA 1
Júlia Ondrusch de Moraes COSTA 2
Raquel Rocha VENANCIO 3
Fernanda Silva ALMEIDA 4 1 Mestrando do Programa de Pós Graduação Multicêntrico em Ciências Fisiológicas/UFPB; 2
Graduanda do Curso de Medicina, Faculdade de Ciências Médicas da Paraíba (FCM-PB) ; 3
Graduanda do Curso de Biotecnologia/UFPB; 4 Orientadora/ Doutoranda do Programa de
Pós-Graduação em Biotecnologia- RENORBIO/ UFPB;
RESUMO: A leishmaniose se caracteriza como um complexo de doenças infecto-parasitárias causadas por protozoários pertencentes ao gênero Leishmania, e estão inseridas no grupo das doenças tropicais negligenciadas. Esta patologia é considerada um agravante de saúde pública, presente em diversos países, em especial o Brasil, como um dos países com maior prevalência da doença. Os macrófagos são células fagocíticas que são encontradas em vários tecidos. Sendo assim, o presente trabalho objetivou revisar a bibliografia existente acerca da ação do parasita na plasticidade dos macrófagos e os papéis da polarização M1 e M2 na parasitose em questão, utilizando um total de 28 referências entre os anos de 2016 a 2020. Na Leishmaniose, em especial, os macrofágos tornam-se células hospedeiras finais para a proliferação de parasitas de Leishmania, sendo uma chave para a resposta controlada ou exacerbada que culmina em manifestações clínicas. M1 e M2 são os dois principais fenótipos de macrófagos nos quais M1 é um subtipo clássico com perfil pró-inflamatório e M2, ou alternativamente ativado, é um subtipo anti-inflamatório / regulatório. A compreensão dos principais

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
310
mecanismos imunomodulatórios presentes na leishmaniose, possibilitam o desenvolvimento de novas abordagens terapêuticas que garantam melhor qualidade no tratamento dos pacientes. Palavras-chave: Leishmaniose. Imunidade Inata. Macrófagos M1 e M2. INTRODUÇÃO
Leishmaniose é uma doença infecciosa causada por
espécies de protozoários do gênero Leishmania. A transmissão
ocorre pelo repasto sanguíneo realizado pela fêmea de
flebotomíneos e as principais formas de manifestações clínicas
das leishmanioses são: tegumentar (LT) e visceral (LV). A LV,
possui como agentes etiológicos a Leishmania donovani e a
Leishmania infantum e é considerada a forma mais grave da
doença. A LT por sua vez, possui, dentre seus agentes
etiológicos no Brasil, a Leishmania amazonensis e Leishmania
braziliensis (STEVERDING, 2017; WHO, 2016).
Os parasitas da Leishmaniose apresentam um ciclo de
vida do tipo heteroxênico e com morfologia variável, que se
alterna entre dois estágios de desenvolvimento distintos: a
forma promastigota, a qual se apresenta de maneira flagelada
de vida livre, e a forma amastigota, caracterizada por serem
desprovidas de flagelo e serem intracelulares obrigatórias,
sendo encontradas nas vesículas fagolisossômicas dos
vertebrados, principalmente em macrófagos (SERAFIM, 2018).
Os macrófagos desempenham um duplo papel na
infecção por Leishmania. Os macrófagos são responsáveis pela
destruição de parasitas internalizados, mas também fornecem
um local seguro para replicação da Leishmania. Dessa forma,
os macrófagos são importantes para a progressão da doença,

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
311
e o sucesso ou falha da infecção depende da interação do
imunometabolismo do patógeno com a resposta imunológica do
hospedeiro. Esses fatores estão relacionados com às formas
clínicas da doença (SCOTT, NOVAIS, 2016; SRIVASTAVA et
al., 2016). Os macrófagos normalmente estão em repouso com
o fenótipo M0, e através de sinais diferentes e necessários
fornecidos pelo microambiente, há ativação de vias de
sinalização que levam ao desenvolvimento de fenótipos de
macrófagos funcionalmente distintos: os macrófagos ativados
de modo clássico, chamados macrófagos M1 ou macrófagos
ativados de forma alternativa, os M2 (TOMIOTTO-PELLISSIER
et al., 2018)
Macrófagos M0 são diferenciados em M1 pela ação de
citocinas secretadas por linfócitos Th1, como interferon (IFN-y)
e fator de necrose tumoral (TNF-α). Os macrófagos ativados
classicamente são cruciais para eliminação do patógeno
intracelular pela produção de espécies reativas de oxigênio
(ERO) e óxido nítrico (NO), que exibem um potencial
microbicida (TOMIOTTO-PELLISSIER et al., 2018).
Por outro lado, a ativação de linfócitos Th2 levam a
produção de citocinas como IL-4 IL-13 que induzem o perfil M2
de macrófagos, que são caracterizados pela biossíntese de
poliamina pela ativação da enzima arginase, produção de uréia
e L-ornitina, que são benéficos para o crescimento da
Leishmania no macrófago, favorecendo a sobrevivência do
parasita em macrófagos infectados e a progressão da doença
(MAXUEL et al., 2017; TOMIOTTO-PELLISSIER et al., 2018).

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
312
OBJETIVOS
Revisar as informações existentes na literatura sobre a
polarização de macrófagos no amplo aspecto da Leishmaniose.
MATERIAIS E MÉTODO
Este estudo trata-se de uma revisão sistemática da
literatura de caráter descritivo fundamentado na coleta de
dados por meio de uma revisão bibliográfica. Sendo assim,
refere-se a uma pesquisa documental com base em dados
secundários obtidos a partir de cinco bancos de dados online:
Elsevier, Google acadêmico, PubMed, Nature, com um
somatório de 19 artigos no total, 4 dissertações e 2 teses.
Com base nos critérios de exclusão e inclusão, os artigos
incluídos nesta pesquisa foram todos originais de língua
portuguesa ou inglesa, que abordaram a temática, Imunologia
da Leishmaniose, Diferenciação de Macrófagos, Terapêutica
com foco na imunologia, além disso, foram utilizados
referências no período de tempo entre 2016 e 2020.
RESULTADOS E DISCUSSÃO
IMUNIDADE INATA
Classicamente a resposta imune é dividida em dois tipos,
imunidade inata e imunidade adaptativa. A imunidade inata é
composta por algumas moléculas e células que promovem
mecanismos que compõem a resposta inata, sendo essas
moléculas mediadoras de reações inflamatórias (sistema
complemento) além de estimular algumas células com

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
313
propriedades fagocíticas (neutrófilos e macrófagos), células
Natural Killer (NK) (NETEA et. al, 2019).
O sistema complemento também faz parte da barreira
inata a infecções primárias. Na leishmaniose o componente C3
apresenta uma função crucial no controle e morte do parasita.
Imediatamente após a inoculação do parasita pelo vetor, são
formados os primeiros produtos de clivagem, os componentes
C3a e C5a, agentes quimiotáticos que desempenham um papel
preponderante na promoção da atração de neutrófilos e
macrófagos (AMBROSIO et. al. 2020).
A relação de morte da Leishmania pela ativação do
sistema complemento é obtida através de alguns componentes
pertencentes ao mesmo. A destruição do parasita ocorre via
ativação do componente C3b do sistema complemento, por sua
vez, o C3b recruta a formação do complexo de ataque à
membrana (MAQ), constituído pelos componentes C5b-C9
(VERMA et. al. 2018).
Em contrapartida, o parasita apresenta em sua estrutura
superficial uma metaloproteinase, a gp63 (glicoprotease de 63
kDa), na qual apresenta função primordial na sobrevivência da
Leishmania. Essa glicoproteina transforma o componente C3b
em C3bi (inativo), esse agente inativo promove a fagocitose
mediada por macrófagos sem ativá-los, essa relação
macrófago-Leishmania está diretamente ligada ao
estabelecimento da infecção parasitária (VERMA et. al. 2018).
Os neutrófilos são uma das primeiras células a serem
recrutados pós evento de infecção, são elas as responsáveis
por combaterem inicialmente o processo infeccioso, sendo
estes eventos moleculares a promoção de morte por NETosis
(armadilha extracelular de neutrófilos), os
monócitos/macrófagos a posteriori são recrutados, estas

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
314
células possuem fundamentalmente dotadas a capacidade de
fagocitose (GARDINASSI et. al. 2017). Adicionalmente, os
mecanismos pelo qual nosso sistema imune inato responde à
infecção pela Leishmania sp. consiste na produção de ROS
(espécies reativas de oxigênio), NO (óxido nítrico) e ativação do
sistema complemento (SCOTT & NOVAIS, 2016).
Na infecção por Leishmania, os neutrófilos são
estimulados por padrões derivados do patógeno, e isso
promove a descondensação da proteína, fazendo com que haja
a junção de eucromatina e heterocromatina. Esse DNA é
exteriorizado formando redes que aprisionam o patógeno e
promovem a sua destruição mediada por proteases liberadas
por gránulos citoplasmáticos. Os neutrófilos também eliminam
a Leishamnia através do metabolismo mitocondrial, a citar a
produção de espécies reativas de oxigênio (ERO/ROS) e
explosão respiratória atuando em sinergia com oxido nítrico na
depuração parasitária (Figura 1) (SCOTT & NOVAIS, 2016).
Figura 1. Mecanismo de morte de parasita via NETosis
Fonte: adaptado de Scott & Novais, 2016.

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
315
DIFERENCIAÇÃO DE MACRÓFAGOS M1/M2
Os macrófagos são células altamente plásticas
responsivas a uma variedade de sinais ambientais que têm o
poder de alterar seu fenótipo e função (Shapouri-Moghaddam
et. al. 2018). Os monócitos, que segundo Chistiakov et al.
(2018) “...neutralizam patógenos e moléculas tóxicas, envolvem
células mortas, danificadas e exógenas, secretam citocinas...”
são os precursores dos macrófagos. Estes amadurecem em
tecidos periféricos passando primeiramente pelo endotélio
vascular. O amadurecimento destes macrófagos se dará
dependendo de estímulos específicos e fatores endógenos e
exógenos (SHAPOURI-MOGHADDAM et. al. 2018).
Doenças inflamatórias possuem a habilidade de ativar ou
desativar funções importantes para a regulação celular, e com
isso modificam processos intracelulares e extracelulares. Neste
contexto, estão inseridos os macrófagos como alvos destas
doenças inflamatórias, e estes estão intimamente associados à
capacidade do hospedeiro de reagir imunologicamente, e
reagem a Leishmania bem como à moléculas danificadas do
próprio hospedeiro, de modo que haja a remoção do tecido
danificado (ATRI, et. al. 2018).
Atualmente está em vigência o conceito de polarização
de macrófagos, que consiste na sua diferenciação fenotípica
entre macrófagos classicamente ativados ou M1, e macrófagos
alternativamente ativados ou M2. A diferenciação fenotípica
dessas células é dependente de estímulos microambientais, de
citocinas produzidas e duração da exposição a estas, e cada
subtipo possuirá então uma resposta específica baseada no
ambiente no qual está inserido, podendo resultar na resolução
da inflamação ou em sua progressão (ATRI, et. al. 2018;
TOMIOTTO-PELLISSIER et. al. 2018).

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
316
Segundo Ferrari (2016), os macrófagos são células de
característica plástica, sendo assim passíveis de serem
estimulados a polarizar-se principalmente pelo fator estimulador
de colônias de macrófagos (FCM), sendo a diferenciação em
M1 induzida por citocinas liberadas por meio de linfócitos TH1,
como por exemplo IFN-y e TNF-α e M2 induzida por citocinas
liberadas por linfócitos TH2 como a IL-4, IL-10 e IL-13 (ATRI, et.
al. 2018).
De acordo com Vogel et. al., (2014) a ativação de
macrófagos varia em diferentes tipos de lesões, por exemplo,
em lesões ativas eles contém mielina e expressam marcadores
para o fenótipo M1 e M2, enquanto na ativação crônica é
evidenciado apenas marcadores para o fenótipo M1.
Marcadores normalmente utilizados para as populações de
macrófagos do tipo M1 e M2 em humanos são CD40, CD64,
CXCL11, CCR7 e MR, estabilina-1, CD180, CD163 e TREM2
respectivamente, entretanto não são específicos para cada um
dos fenótipos, M1 ou M2. (VOGEL et al., 2014).
Os macrófagos classicamente ativados, M1, possuem
uma alta capacidade de apresentação antigênica e alta
produção de fatores promotores da proliferação celular e
ativação de células T, além da produção de substâncias tóxicas
como espécies reativas de oxigênio e intermediários do óxido
nítrico, e aumento da expressão do complexo maior de
histocompatibilidade do tipo 2 (MHC II). Desse modo, a
produção de citocinas pró-inflamatórias e a apresentação de
antígenos configura macrófagos M1 como bons mediadores no
combate a microrganismos e células tumorais. Macrófagos do
tipo M1 orientam nas respostas inflamatórias agudas, e podem
produzir IL-1β, IL-6, IL-12, IL-18 e IL-23, TNF-α e IFN tipo I; e
várias quimiocinas como CXCL1, CXCL3, CXCL5, CXCL8,

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
317
CXCL9, CXCL10, CXCL11, CXCL13 e CXCL16; CCL2, CCL3,
CCL4, CCL5, CCL8, CCL15, CCL11, CCL19 e CCL20;
induzindo assim a resposta de linfócitos Th1 (ATRI, et. al. 2018).
Macrófagos do tipo M2 possuem como característica a
expressão de CD64 e CD209, e sua diferenciação pode se dar
através da presença de IL-4, IL-10, IL-13, e fator estimulador de
colônias de macrófagos (M-CSF). Eles possuem atividade
moduladora, regulam negativamente citocinas pró-inflamatórias
e induzem a produção de mediadores anti-inflamatórios (ATRI,
et. al. 2018). De acordo com Paião (2016) macrófagos M2 são
fenotipicamente caracterizados por expressarem em sua
superfície CD206 , CD301, CD163 e baixa expressão de MHC
de classe II, CD80 e CD86.
Macrófagos alternativamente ativados, M2, possuem
ainda subdivisões denominadas M2a, M2b e M2c. A ativação
do M2a se dá por meio da presença de IL-4 e IL-13 (como
mostra figura 2) e são expressos altos níveis de CD86 e
CD200R enquanto é observada uma baixa expressão de CD14
e TLR4. Este subtipo é responsável por induzir a produção de
IL-10, CCL13, CCL17 e CCL22. O subtipo M2b por sua vez tem
sua resposta estimulada por lipopolissacarídeo (LPS), IL-1β
(como mostra figura 2), agonistas de TLR e complexos imunes,
sendo observada a maior expressão de CD80, CD14, CCL1, IL-
10 e a produção de citocinas pró-inflamatórias, e baixa na
secreção de IL-12 e expressão de antígeno leucocitário humano
(HLA-DR) (ATRI, et. al. 2018). Por outro lado, os macrófagos do
tipo M2c são estimulados por glucocorticóides e IL-10, sendo
considerado por alguns autores também a estimulação via fator
de transformação do crescimento beta (TGF-β) (como mostra
figura 2). A caracterização deste subtipo envolve alta expressão

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
318
de CD163 e diminuição na expressão de CD86 e HLA-DR, além
da produção de CCL16 e CCL18 (ATRI, et. al. 2018).
Existem ainda os macrófagos associados ao tumor
(TAMs) que configuram um novo subtipo de M2, tido como M2d.
Estes macrófagos são estimulados por IL-6, ligantes do
receptor toll-like (TLR) e agonistas do receptor da adenosina
A2A e são caracterizados por uma alta produção de IL-10,
VEGF (como mostra figura 2) e baixa produção de IL-12, TGF-
β, iNOS TNF-α e secreta quimiocinas como CXCL10, CXCL16
e CCL5. Os M2d contribuem para a inflamação do tecido
tumoral, consequente metástase do tumor e colaboram com a
inibição da resposta imune (ATRI, et. al. 2018; TOMIOTTO-
PELLISSIER et. al. 2018).
Figura 2. Polarização de macrófagos não ativados (M0) para
classicamente ativados (M1) e macrófagos alternativamente
ativados (M2).
Fonte: adaptado de Chistiakov et al., 2018.

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
319
IMUNOMETABOLISMO DOS MACRÓFAGOS NA
LEISHMANIOSE
Todas as células do sistema imune são caracterizadas
por sua capacidade de responder a sinais através de receptores
de reconhecimento de padrões, receptores de antígenos,
receptores de citocinas, fator de crescimento, e esses sinais
podem induzir uma mudança dos estados de repouso para
ativado (VAN TEIJLINGEN BAKKER; PEARCE, 2020).
Sabe-se que os macrófagos alteram seu metabolismo
em resposta ao microambiente. Por sua vez, essas alterações
metabólicas direcionam funções efetoras específicas em
macrófagos. Alguns dos eventos mais comuns que
desencadeiam a reprogramação metabólica de macrófagos
incluem a ativação de receptores de reconhecimento de
patógenos, como receptores toll-like (TLRs) ou sinais baseados
em nutrientes que envolvem receptores nucleares lipídicos
[nutrient based signals that engage lipid nuclear receptors
(PPAR, ERR)] e / ou cinases (mTOR ou AMP quinase)
(ARTYOMOV; SERGUSHICHEV; SCHILLING, 2016).
A estimulação de lipopolissacarídeo (LPS) leva à
regulação negativa da expressão da enzima isocitrato
desidrogenase (IDH), que leva à produção de excesso de citrato
para apoiar a síntese de ácidos graxos e a geração de
intermediários lipídicos pró-inflamatórios. A regulação negativa
do IDH também promove o acúmulo de precursores à base de
citrato para alimentar a produção de itaconato, que inibe o
complexo succinato desidrogenase (complexo SDH) nas
mitocôndrias. Para compensar a expressão reduzida de IDH, as

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
320
células hospedeiras ativadas por LPS aumentam o
metabolismo da glutamina para a produção dos principais
intermediários metabólicos α-cetoglutarato e succinato
(RUSSELL; HUANG; VANDERVEN, 2019).
Além disso, essas células também apresentam atividade
aumentada da via de pentose fosfato necessária para converter
glicose-6-fosfato em nucleotídeos e proteínas enquanto gera
NADPH, que suporta a biossíntese lipídica, bem como a
produção de espécies reativas de oxigênio (ROS) e
intermediários reativos de nitrogênio (RUSSELL; HUANG;
VANDERVEN, 2019).
As infecções causadas por patógenos causam
mudanças na fisiologia e metabolismo da célula hospedeira, as
quais são mediadas pelo sistema imune inato e por fatores
derivados de patógenos. Durante a invasão pela leishmania, os
macrófagos ativam vias citotóxicas na tentativa de erradicar o
patógeno. Essas vias incluem a indução da biossintese de óxido
nítrico (NO) e a liberação de espécies reativas de oxigênio
(ERO). Para combater os mecanismos citotóxicos do
macrófago, a leishmania desenvolve diversas estratégias para
suprimir a resposta do hospedeiro. A infecção por leishmania
regula positivamente a atividade da arginase I em macrófagos
hospedeiros, limitando a arginina disponível para NO sintase,
que são enzimas que catalisam a produção de óxido nítrico a
partir da arginina. Ademais, a forma amastigota da leishmania
aumenta a produção de antioxidante como a tripanotiona, que
neutraliza as espécies reativas de oxigênio permitindo o
desenvolvimento de amastigotas nos fagolisossomos de
macrófagos (GOLDMAN-PINKOVICH, 2016).
Os macrófagos pró-inflamatórios são caracterizados por
uma diminuição da fosforilação oxidativa mitocondrial

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
321
(OXPHOS), contudo produz uma maior quantidade de ATP
através da cadeia de transporte de elétrons. Mesmo com o
aumento da taxa glicolítica, o OXPHOS nos macrófagos M1 é
bastante prejudicado. O óxido nítrico (NO) não é apenas um
agente antimicrobiano, mas também desempenha um papel na
modulação metabólica dos macrófagos M1. O NO modifica a
cadeia de transporte de elétrons e inibe a respiração
mitocondrial. Uma mudança metabólica em direção à glicólise
e OXPHOS instrui a diferenciação de macrófagos em diferentes
subconjuntos funcionais. Embora a oxidação mitocondrial de
ácidos graxos (FAO) e OXPHOS sejam essenciais para a
manutenção de células quiescentes "em repouso", como
células T naive, a biossíntese lipídica é um pré-requisito para
respostas pró-inflamatórias de macrófagos (ARTYOMOV;
SERGUSHICHEV; SCHILLING, 2016; THAPA; LEE, 2019).
Nos macrófagos M2, a principal assinatura metabólica é
o consumo de ácido graxo e o aumento da capacidade
respiratória mitocondrial. Os macrófagos M2 também são
freqüentemente associados a um metabolismo distinto da
arginina. A indução da óxido nítrico sintase 2 (NOS2) nos
macrófagos M1 metaboliza a arginina para produzir NO, porém
nos macrófagos alternativamente ativados, a regulação positiva
da atividade da arginase nos macrófagos M2 processa
alternativamente a arginina em ornitina, que é usada para
produzir poliaminas (THAPA; LEE, 2019).
Macrófagos alternativamente ativados (M2) utilizam
ácidos graxos, glicose e glutamina para alimentar o ciclo TCA e
produzir ATP por fosforilação oxidativa. Sabe-se que essas
células possuem mitocôndrias alongadas e,
consequentemente, produção de energia mais eficiente. (VAN

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
322
TEIJLINGEN BAKKER; PEARCE, 2020; DE SOUZA BREDA et
al, 2019). Os ácidos graxos exógenos e triacilgliceróis são
absorvidos pelo CD36 e a lipólise é usada para gerar ácidos
graxos para a oxidação de ácidos graxos (FAO). Os ácidos
graxos ativados entram na mitocôndria, que gera acetil-CoA,
para contribuir para o ciclo do TCA. O carbono da glicose
também é crítico para a síntese de citrato. Diferentemente dos
macrófagos inflamatórios, o isocitrato desidrogenase é
expresso em macrófagos alternativamente ativados e, além de
ser usado na produção de acetil-CoA, usado na acetilação de
histonas, o citrato é convertido em α-KG, que também é gerado
a partir de glutamina pela glutaminólise. (VAN TEIJLINGEN
BAKKER; PEARCE, 2020).
Em um estudo realizado por Ty, Maureen e
colaboradores (2019), foi investigado o status metabólico e
imunológico dos macrófagos humanos após a infecção por
Leishmania donovani e L. amazonensis e sua capacidade de
responder a um estímulo polarizador clássico (LPS e IFN-γ). A
infecção dos macrófagos Leishmania resultou na ativação da
fosforilação oxidativa, um padrão também encontrado em
macrófagos polarizados em M2. Ademais, a adição de
estímulos de ativação (LPS + IFN-γ) aos macrófagos infectados
por L. donovani resultou em um aumento significativo na
glicólise, indicando que as células infectadas ainda respondem
à ativação, uma característica muito importante durante a
infecção. Houve uma secreção ainda mais alta de citocinas
inflamatórias em comparação aos macrófagos não infectados,
indicando que os macrófagos infectados aumentaram a
capacidade metabólica, correspondendo ao aumento da
produção de citocinas em resposta aos estímulos.

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
323
MACRÓFAGOS M1/M2 NA LEISHMANIOSE TEGUMENTAR
O papel dos subconjuntos de macrófagos na
Leishmaniose ainda não foi investigado completamente. No
entanto, é necessário entender o papel dessas células no
desenvolvimento da lesão pela infecção por Leishmania a fim
de compreender os perfis M1 e M2 como uma ferramenta
importante na patogênese da leishmaniose.
A Leishmaniose Tegumentar (LT) é a forma mais comum
da leishmaniose. A LT causa lesões cutâneas que deixam
cicatrizes ao longo da vida e sérias deficiências, tornando-se
um grave problema de saúde pública. Os agentes etiológicos
primários incluem as espécies Leishmania tropica e L. major, L.
amazonensis e L. (v.) braziliensis (WHO, 2020).
Pacientes com LT possuem níveis plasmáticos mais
altos de arg-1 (arginase 1), TGF-β (fator de transformação do
crescimento beta) e PGE2 (prostaglandina E2), além de um
número aumentado de arginina nas lesões, sugerindo que os
macrófagos M2 podem desempenhar um papel na patogênese
da doença. Nesse sentido, um estudo in vitro demonstrou que
apenas os macrófagos M2 permitem o crescimento de L. major
e L. amazonensis, uma vez que o lipofosfoglicano (GLP) e a
glicoproteína GP63 dos parasitas agem em macrófagos M2 e
suprimem os RNAs não codificadores, o que deixa essas
células permissivas à infecção (TOMIOTTO-PELLISSIER ET
AL., 2018). A infecção preferencial das células M2 tem papel
crucial na gravidade da doença cutânea.
De acordo com Tomiotto e colaboradores (2018) que
realizaram um estudo de revisão, estudos investigaram o papel
dos receptores ativados por proliferadores de peroxissoma
(PPARs) em macrófagos infectados por Leishmania em

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
324
camundongos resistentes e suscetíveis à essa infecção. Os
PPARs são fatores de transcrição dependentes de ligantes que
são expressos em macrófagos e regulam a expressão de genes
relacionados com a resposta inflamatória. Assim,
pesquisadores demonstraram que camundongos deficientes de
PPARγ, tiveram um progresso retardado da doença, com
menos edema na pata e carga parasitária reduzida. É
importante ressaltar que os genes expressos preferencialmente
em macrófagos M2, como arg-1, Mrc1 e Clec7a, também foram
diminuídos no tecido de camundongos deficientes para PPARγ
Esses dados sugerem que os receptores PPARγ são
necessários para aquisição e manutenção dos macrófagos M2
no modelo L. major. Em contraste, a expressão de PPARs
induziu a ativação da linhagem celular de macrófagos murinos
J774A.1 para polarização em direção ao perfil M1, com alta
produção de mediadores pró-inflamatórios (TNF-α, IL-1β, IL-6,
TLR4 e ROS) e aumento da atividade microbicida contra L.
mexicana. Sendo assim, os receptores ativados por
proliferadores de peroxissoma favorecem o fenótipo M1 dos
macrófagos.
MACRÓFAGOS M1/M2 NA LEISHMANIOSE VISCERAL
A leishmaniose visceral é uma doença sistêmica crônica
que é causada por protozoários intracelulares do gênero
Leishmania, e seus agentes infecciosos são as espécies
Leishmania donovani, Leishmania infantum e Leishmania
infantum / chagasi (OPAS, 2019).
De acordo com o Ministério da Saúde (2019) a
Leishmaniose Visceral é uma zoonose de evolução crônica e
caso não tratada possui 90% de chance de evolução dos casos

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
325
para óbito. O Brasil, é uma região endêmica para a doença e
possui como vetor responsável pela transmissão dos
protozoários a espécie Lutzomyia longipalpis, também
conhecido como mosquito-palha, e a disseminação ocorre
através da picada da fêmea deste flebotomíneo ao realizar o
repasto sanguíneo.
Devido a Leishmania ser um parasito intracelular a forma
de defesa contra este patógeno é observada majoritariamente
como uma resposta imune mediada por células (GONÇALVES,
2015). É sabido que o reconhecimento inicial destes parasitas
possui a participação de receptores encontrados em células
NK, células dendríticas (CDs), monócitos, macrófagos e células
polimorfonucleares, entretanto ainda há muitos aspectos da
resposta que necessitam ser estudados mais profundamente
(GATTO, 2018). Os macrófagos que se encontram presentes
no sítio de infecção são estimulados a ativação por meio do
reconhecimento de PAMPs (Padrões moleculares ligados a
patógenos) via receptores TLRs (Toll Like Receptors)
(GONÇALVES, 2015).
Macrófagos são as células hospedeiras da Leishmania
no hospedeiro vertebrado, e aqueles infectados pelas espécies
viscerotrópicas se localizam essencialmente no fígado, baço e
medula óssea, além disso, atuam também como importante
forma de controle da infecção. (MAGALHÃES, 2016). Espécies
de Leishmania são capazes de induzir a polarização de ambos
os fenótipos de macrófagos, sendo eles do tipo M1 relacionados
a resposta Th1/IFN-γ, ou do tipo M2, relacionados a resposta
Th2/IL-4 e reparação tecidual (MAGALHÃES, 2016).
Estudos têm demonstrado que macrófagos M2 são
permissivos a proliferação de Leishmania, sendo relacionado
ao agravamento da infecção, enquanto o fenótipo M1 é

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
326
relacionado a resistência e a expressão do radical
leishmanicida, óxido nítrico (SILVA, 2017; MAGALHÃES, 2016).
Entretanto, as formas promastigotas induzem a apoptose por
meio do aumento da síntese de TGF-β e supressão da síntese
de óxido nítrico (NO), o que torna a atividade inflamatória das
células M1 menor e possibilita a sobrevivência dos parasitas
(SILVA, 2017).
O controle da infecção da maioria das espécies de
Leishmania é mediada por células Th1, estas produzem IL-12 e
IFN-γ conjuntamente com a produção de citocinas pró
inflamatórias e a ativação de macrófagos infectados,
promovendo produção de NO e oxigênio intermediário reativo
(ROI). A resposta por meio de células Th2 não é ainda bem
esclarecida, mas sabe-se que há produção de IL-4 e
estimulação de plasmócitos para que haja a produção de
anticorpos anti-Leishmania. Durante a infecção por LV observa-
se o aumento de IL-10, e juntamente com a IL-4 aparenta ser
responsável pelo agravamento da doença, devido a capacidade
de inibição da ação de macrófagos por estas citocinas,
participando também deste processo o TGF-β e prostaglandina
E2 e devido a isto possa se dar a incapacidade dos macrófagos
de eliminar formas amastigotas do protozoário (ARAÚJO,
2019).
A IL-10 produzida por células Treg, caracterizadas por
serem CD4+, CD25+, Foxp3+, também está atrelada a
suscetibilidade de desenvolvimento da doença, a leishmaniose
visceral. Participam ainda desta resposta imune células do
perfil TH17, que são linfócitos do tipo CD4+ e produzem
citocinas pró-inflamatórias como a IL-17. As células com este
perfil se diferenciam em microambiente contendo TGF-β, IL-6,
IL-1β e IL-23, que em conjunto colaboram para a manutenção

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
327
e expressão dos receptores de quimiocinas nestas células
(GATTO, 2018).
Segundo Dos Santos (2016) pacientes clássicos com LV
apresentam comprometimento da produção de IL-2, IFN-γ e IL-
12 a partir de células T específicas para antígenos da
leishmania in vitro, mas é passível de restauração após o
tratamento caso este seja bem-sucedido. Contudo, foi
observado que pacientes com LV ativa apresentam altos níveis
de IFN-γ e IL-12 no soro, isto poderia ajudar no controle da
infecção, todavia, foi relatado também uma elevação de
mediadores anti-inflamatórios, por exemplo, IL-10, que podem
interferir negativamente no combate ao parasita e possibilitar a
sua multiplicação induzindo assim o agravo da infecção.
Citocinas como a IL-6, IL-21 e IL-27 são referenciadas
como moduladoras da resposta imune no hospedeiro, além do
parasita induzir a resposta inflamatória essa resposta também
pode se dar através de co-infecções, geralmente por bactérias
que provocam a ativação do sistema imune (DOS SANTOS,
2016).
CONCLUSÕES
Vimos que os macrófagos desempenham um papel
duplo na infecção por Leishmania. Essas células são
responsáveis pela destruição de parasitas internalizados,
porém também podem fornecer um local seguro para a
replicação da Leishmania. Portanto, os macrófagos são
fundamentais para a progressão da doença e o sucesso ou
retrocesso dela, e esses processos dependem da interação
entre o parasito e o microambiente na resposta imune do

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
328
hospedeiro. Esses dois fatores estão intimamente relacionados
às formas clínicas da leishmaniose.
Diferentes espécies de Leishmania exibem fatores de
virulência que podem subverter os mecanismos microbicidas do
hospedeiro, favorecendo sua proliferação devido à polarização
M2. No entanto, os macrófagos M1 agem durante períodos
posteriores da infecção e desencadeiam uma resposta imune
exacerbada que leva a um agravamento das lesões, apesar do
papel dessas células como poderosos agentes microbióticos.
Mais estudos são necessários para elucidar completamente o equilíbrio entre essas duas principais populações de macrófagos no controle da infecção por Leishmania. A compreensão aprimorada das vias de interação essenciais na infecção por macrófagos por Leishmania auxiliam do desenho de novos fármacos que possam interromper as interações que favorecem o parasita, aumentar as funções microbicidas e imunológicas dos macrófagos que apoiam a eliminação do parasita. REFERÊNCIAS BIBLIOGRÁFICAS Ambrosio AR, Bavia L, Borges BS, Hiraiwa PM, Pietsch JLM, Ribeiro MCVDC, Figueiredo FB, Messias-Reason IJ. Novel findings on the role of ficolins and colectins in the innate response against Leishmania braziliensis. Acta Trop. v. 19, n. 212:105673, p. 1-5. 2020. ARTYOMOV, Maxim N.; SERGUSHICHEV, Alexey; SCHILLING, Joel D. Integrating immunometabolism and macrophage diversity. In: Seminars in immunology. Academic Press, 2016. p. 417-424. ATRI, Chiraz; GUERFALI, Fatma Z.; LAOUINI, Dhafer. Role of human macrophage polarization in inflammation during infectious diseases. International journal of molecular sciences, v. 19, n. 6, p. 1801, 2018. CHISTIAKOV, Dimitry A. et al. The impact of interferon-regulatory factors to macrophage differentiation and polarization into M1 and M2. Immunobiology, v. 223, n. 1, p. 101-111, 2018. DOS SANTOS, Priscila L. et al. The severity of visceral leishmaniasis correlates with elevated levels of serum IL-6, IL-27 and sCD14. PLoS neglected tropical diseases, v. 10, n. 1, p. e0004375, 2016.

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
329
FERRARI, Pâmela. Polarização M1 e M2 da linhagem U-937 de macrófagos em meio de soro de pacientes com transtorno bipolar. 2016. GARDINASSI, LG et al. Assinaturas moleculares de armadilhas extracelulares de neutrófilos na leishmaniose visceral humana. Parasites Vectors. v. 10, n. 285, p. 1-7. 2017. GATTO, Mariana. Análise do perfil transcricional de células THP-1 infectadas com Leishmania infantum/chagasi: ênfase no inflamassoma e receptores NODs. 2018. GOLDMAN-PINKOVICH A et al. An Arginine Deprivation Response Pathway Is Induced in Leishmania during Macrophage Invasion. PLoS Pathog. 2016. GONÇALVES, A. C. Avaliação do efeito do laser terapêutico de 660nm na leishmaniose tegumentar experimental: expressão de citocinas e perfil funcional de macrófagos. 2015. 90 f. Dissertação (Mestrado em Biologia da Relação Parasito-Hospedeiro) - Universidade Federal de Goiás, Goiânia, 2015. MAGALHÃES, Lucas Sousa. Caracterização fenotípica de Leishmania infantum obtidas de pacientes refratários ao tratamento com antimonial. 2016. 49 f. Dissertação (Mestrado em Biologia Parasitária) – Universidade Federal de Sergipe, São Cristóvão, SE, 2016. MINISTÉRIO DA SAÚDE. Leishmaniose Visceral: O que é, causas, sintomas, tratamento, diagnóstico e prevenção. Disponível em: <http://saude.gov.br/saude-de-a-z/leishmaniose-visceral>. Acesso em: 20 out. 2020. Netea, M. G.; Schlitzer, A.; Placek, K.; Joosten, L. A. B.; Schultze, J. L. Innate and Adaptive Immune Memory: an Evolutionary Continuum in the Host's Response to Pathogens. Cell Host Microbe. v. 9, n. 25(1), p. 13-26. 2019. PAIÃO, Munir Regini. Avaliação da participação de macrófagos M1 e M2 na resposta contra o fungo dimórfico Paracoccidioides brasiliensis. 2016. recurso online ( 106 p.). Tese (doutorado) - Universidade Estadual de Campinas, Faculdade de Ciências Médicas, Campinas, SP. RUSSELL, David G.; HUANG, Lu; VANDERVEN, Brian C. Immunometabolism at the interface between macrophages and pathogens. Nature Reviews Immunology, v. 19, n. 5, p. 291-304, 2019. SCOTT, Phillip; NOVAIS, Fernanda O. Cutaneous leishmaniasis: immune responses in protection and pathogenesis. Nature Reviews Immunology, v. 16, n. 9, p. 581-592, 2016. SERAFIM TD, Coutinho-Abreu IV, Oliveira F, Meneses C, Kamhawi S, Valenzuela JG. Sequential blood meals promote Leishmania replication and reverse metacyclogenesis augmenting vector infectivity. Nat Microbiol. 2018. Shapouri-Moghaddam A, Mohammadian S, Vazini H, Taghadosi M, Esmaeili SA, Mardani F, Seifi B, Mohammadi A, Afshari JT, Sahebkar A. Macrophage

MACRÓFAGOS M1/M2 NA LEISHMANIOSE
330
plasticity, polarization, and function in health and disease. J Cell Physiol. v. 233, n. 9, p. 6425-6440. 2018. SILVA, Mayara Cristina Pinto da. Micro/Nanopartículas poliméricas e biodegradáveis de mesocarpo de babaçu: ação imunomoduladora na polarização de macrófagos e efeito anti-leishmania. 2017. [91 f.]. Tese( PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DA SAÚDE/CCBS) - Universidade Federal do Maranhão, [São Luís]. SRIVASTAVA, Saumya et al. Possibilities and challenges for developing a successful vaccine for leishmaniasis. Parasites & vectors, v. 9, n. 1, p. 277, 2016. THAPA, Bikash; LEE, Keunwook. Metabolic influence on macrophage polarization and pathogenesis. BMB reports, v. 52, n. 6, p. 360, 2019. STEVERDING, Dietmar. The history of leishmaniasis. Parasites & vectors, v. 10, n. 1, p. 82, 2017. TOMIOTTO-PELLISSIER, Fernanda et al. Macrophage polarization in leishmaniasis: broadening horizons. Frontiers in immunology, v. 9, p. 2529, 2018. VAN TEIJLINGEN BAKKER, Nikki; PEARCE, Edward J. Cell‐intrinsic metabolic regulation of mononuclear phagocyte activation: Findings from the tip of the iceberg. Immunological Reviews, v. 295, n. 1, p. 54-67, 2020. VERMA S, et al. Leishmania donovani Inhibitor of Serine Peptidases 2 Mediated Inhibition of Lectin Pathway and Upregulation of C5aR Signaling Promote Parasite Survival inside Host. Front Immunol. v. 29, n. 9:63, p. 1-20. 2018. VOGEL, Daphne YS et al. Human macrophage polarization in vitro: maturation and activation methods compared. Immunobiology, v. 219, n. 9, p. 695-703, 2014. World Health Organization. Leishmaniasis. Disponível em: http://www.who.int/news-room/fact-sheets/detail/leishmaniasis. Acesso em: 22.out. 2020. World Health Organization Leishmaniasis in high-burden countries: an epidemiological update based on data reported in 2014. Relev Epidemiol Hebd. 2016. AGRADECIMENTOS
Os autores agradecem aos órgãos de fomento: Fundação de apoio a Pesquisa na Paraíba - UFPB, à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior- CAPES e ao Conselho Nacional de Pesquisa - CNPq pelo suporte financeiro.

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
331
CAPÍTULO 16
MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA
PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
Letícia de Oliveira SOUZA1
Claudio Roberto Bezerra dos SANTOS2 1 Graduanda do curso de Ciências Biológicas, UFPB; 2 Orientador/Professor do DPF/UFPB.
RESUMO: A inflamação é um mecanismo de defesa orquestrado pelo sistema imunológico para combater infecções através do recrutamento de leucócitos. Este processo é mediado pela liberação de citocinas por células residentes de tecidos como macrófagos, mastócitos e células dendríticas (CDs), ativação do endotélio vascular por proteínas de adesão e quimiocinas, transmigração endotelial, fagocitose e eliminação do patógeno. A resposta inflamatória é um componente marcante das doenças crônicas, a exemplo das doenças cutâneas com manifestações predominante ou exclusivamente na pele. Objetivou-se fazer uma revisão acerca dos mecanismos celulares envolvidos na inflamação de pele, com foco na psoríase (PsO). Ao todo foram selecionadas 54 referências, utilizando-se palavras-chave e artigos entre 2015 e 2020, encontradas nas bases de dados PubMed e ScienceDirect. A PsO é definida como uma doença inflamatória sistêmica, crônica e com manifestações cutâneas, caracterizada por um infiltrado inflamatório, hiperproliferação epidérmica e alta diferenciação dos queratinócitos. Esta patologia é multifatoral, sendo afetada por fatores genéticos, imunológicos e ambientais. As lesões são identificadas pela

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
332
presença de macrófagos, CDs e populações de linfócitos Th1 e Th17, e suas citocinas secretadas como interferon-γ, fator de necrose tumoral-α e interleucinas 17 e 23. Logo, entender a imunopatogênese da psoríase é essencial para melhor compreensão da função e atividade dos alvos terapêuticos, para elaboração de tratamentos eficazes para todas as variantes dentro do espectro da PsO. Palavras-chave: Inflamação na pele. Imunopatogênese. Psoríase.
INTRODUÇÃO
O epitélio é a primeira linha de defesa do organismo
contra infecções (GRAVITZ, 2018), com uma área de superfície
que está exposta a patógenos ambientais de diferentes
naturezas. É composto por queratinócitos, importantes para
regular as respostas imunes na pele e formar uma barreira
mecânica, células de Merkel, melanócitos, células de
Langerhans, linfócitos T gama/delta, e outros leucócitos
(KUMARI & PASPARAKIS, 2015), que são tipos celulares
especializados em ativar a barreira imunológica para impedir a
entrada de patógenos, e juntamente com o sistema imune
adaptativo, criar uma memória imunológica (O’MALLEY;
CLARK; WIDLUND, 2019).
As células imunes são ativadas por um contato
permanente com um estímulo antigênico, como microrganismos
quando há ruptura da barreira, ou antígenos não microbianos,
como agentes físico-químicos (também capazes de
desenvolver uma resposta de hipersensibilidade), ou estímulos
endógenos. Quando há persistência dos estímulos e
predisposição genética, ou os mecanismos regulatórios não
atuam corretamente, o local é infiltrado por células do sistema

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
333
imunológico que promovem uma ativação crônica envolvendo o
tecido infiltrado (SABAT et al., 2019).
A resposta inflamatória é o mecanismo utilizado pelo
sistema imunológico na defesa contra infecções (ISAILOVIC et
al., 2015), que envolve o recrutamento, marginalização e
migração dos leucócitos para o local da infecção, mediados por
quimiocinas, proteínas de adesão, como selectinas e integrinas
(ABBAS; LICHTMAN; PILLAI, 2019; KORNILUK et al., 2016),
além de mediadores pró-inflamatórios lipídicos, como
prostaglandinas e leucotrienos (BASIL & LEVY, 2016). Esse
processo inato é geralmente autolimitado, com liberação de
substâncias anti-inflamatórias que promovem uma resolução
ativa da inflamação, com apoptose e liberação de resíduos
celulares (CRUSZ & BALKWILL, 2015). Entretanto, essa
resolução comumente requer uma resposta adicional do
sistema imune adaptativo, que é mediado por linfócitos T e B
(TODD; SPICKETT; FAIRCLOUGH, 2015).
Sendo assim, a inflamação pode ser desencadeada por
uma ruptura no tecido e entrada de microrganismos
patogênicos, como é o caso das bactérias do gênero
Streptococcus, as quais residem no epitélio, e no rompimento
da barreira epitelial, entram nos tecidos ou caem na corrente
sanguínea, dando início a uma série de respostas imunológicas
ativadas para combater a infecção. Os estímulos exógenos
primeiramente são reconhecidos por células residentes de
tecido ou do sangue, como mastócitos, macrófagos (M1) e
células dendríticas (GERHARDT & LEY, 2015), através de
padrões moleculares associados ao patógeno (PAMPs),
secretando citocinas, como TNF, IL-1 e outros mediadores
inflamatórios, estimulando a ativação do endotélio por proteínas

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
334
de adesão, e permitindo o rolamento de leucócitos através do
endotélio. Proteínas do sistema complemento tanto se ligam
aos organismos antigênicos no processo de opsonização
facilitando a fagocitose ou destruição por apoptose dos
patógenos, quanto seus fragmentos induzem o recrutamento de
fagócitos no sangue (PASTWINSKA et al., 2017; TODD;
SPICKETT; FAIRCLOUGH, 2015).
Assim, o presente trabalho objetiva fazer um
levantamento bibliográfico acerca dos princípios e mecanismos
celulares envolvidos na inflamação de pele, elucidar a atividade
desses processos nas doenças inflamatórias com
manifestações cutâneas, com foco na Psoríase (Pso), bem
como discutir suas características celulares e patológicas.
MATERIAIS E MÉTODO
Trata-se de uma revisão bibliográfica com o objetivo de
integrar as informações disponíveis sobre os mecanismos
imunológicos da inflamação de pele e seu envolvimento na
imunopatogênes de doenças inflamatórias com manifestações
cutâneas, como a PsO.
As bases de dados utilizadas para pesquisa foram
PubMed e ScienceDirect. Ao todo foram utilizadas 54
referências. A seleção de artigos e demais fontes bibliográficas
foi feita a partir de dois critérios: (1) Os trabalhos que foram
publicados entre 2015 e 2020 foram selecionados, com
exceção de um trabalho de 2014 considerado essencial para
esta revisão; (2) Os trabalhos foram selecionados perante a
leitura dos resumos e busca por palavras-chave (pele,
inflamação, citocinas, mediadores inflamatórios, genes,

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
335
psoríase), e escolhidos quando a abordagem e metodologia
coincidiam com a temática do presente trabalho, para que
houvesse precisão no tema tratado nesta revisão.
RESULTADOS E DISCUSSÃO
Rolamento e transmigração dos leucócitos
O rolamento e transmigração dos leucócitos são eventos
cruciais na inflamação aguda, ocorrem em diferentes etapas
desde o recrutamento das células no sangue até a passagem
das moléculas através do endotélio, processo nomeado de
diapedese (GERHARDT & LEY, 2015). Os eventos são básicos
e semelhantes para todos os tipos de leucócitos, porém,
moléculas e vias de sinalização variam dependendo do tipo
celular, do leito vascular e do estímulo inflamatório (MULLER,
2016).
A liberação de TNF e IL-1 pelos macrófagos e células
dendríticas estimula a expressão da E-selectina, enquanto que
a histamina secretada pelos mastócitos induz a expressão de
P-selectina pelas células endoteliais (BORSING, 2018). A baixa
afinidade das selectinas com seus ligantes, aliada ao conjunto
de alterações nos vasos sanguíneos, como a dilatação e o
cisalhamento do fluxo sanguíneo, permitem o rolamento dos
leucócitos ao longo da superfície endotelial. Após a interação
de alta afinidade das integrinas leucocitárias com seus ligantes
nas células endoteliais, e a forte ligação dos leucócitos ao
endotélio, o complexo VE-caderina que une as células
endoteliais é fosforilado por cinases, e assim a ligação entre as
células é rompida, permitindo a passagem dos leucócitos
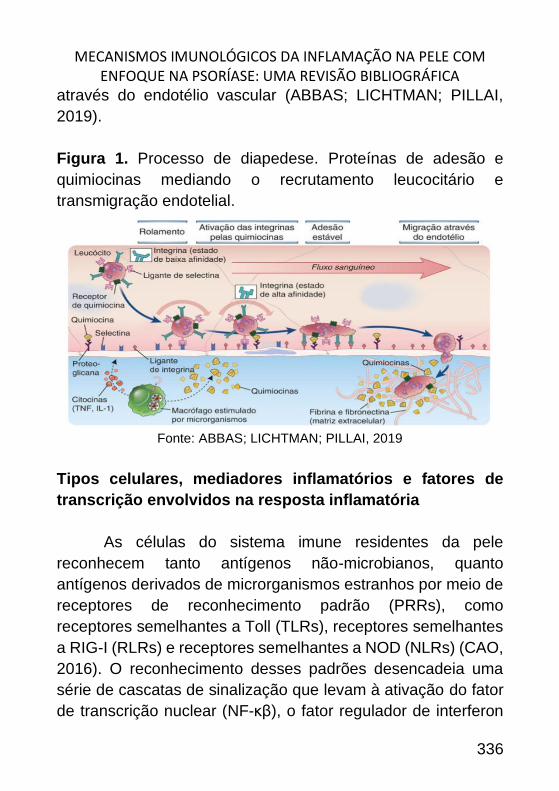
MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
336
através do endotélio vascular (ABBAS; LICHTMAN; PILLAI,
2019).
Figura 1. Processo de diapedese. Proteínas de adesão e
quimiocinas mediando o recrutamento leucocitário e
transmigração endotelial.
Fonte: ABBAS; LICHTMAN; PILLAI, 2019
Tipos celulares, mediadores inflamatórios e fatores de
transcrição envolvidos na resposta inflamatória
As células do sistema imune residentes da pele
reconhecem tanto antígenos não-microbianos, quanto
antígenos derivados de microrganismos estranhos por meio de
receptores de reconhecimento padrão (PRRs), como
receptores semelhantes a Toll (TLRs), receptores semelhantes
a RIG-I (RLRs) e receptores semelhantes a NOD (NLRs) (CAO,
2016). O reconhecimento desses padrões desencadeia uma
série de cascatas de sinalização que levam à ativação do fator
de transcrição nuclear (NF-κβ), o fator regulador de interferon

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
337
(IRF) e a proteína ativadora-1 (AP-1) (LIU & CAO, 2016),
resultando na expressão de diversas moléculas inflamatórias, e
dando início a resposta inflamatória inata e adaptativa em
paralelo.
Os macrófagos são as principais células produtoras de
mediadores inflamatórios nas doenças autoinflamatórias e
autoimunes. Possuem uma complexa via de sinalização
dependente de cinases que está ligada aos PRRs da célula, e
ativam fatores de transcrição NF-κβ, CREB, AP-1, e IRFs, os
quais iniciam a transcrição de genes codificadores de citocinas
como IL-12/13 e interferon tipo I e interleucina-6 (IL-6)
(HAMIDZADEH et al., 2017). O aumento da produção destas
citocinas está associado com os mecanismos intracelulares
para eliminação do patógeno, como a produção de espécies
reativas de oxigênio (ROS) e óxido nítrico (NO), e formação da
imunidade celular de longa duração (HAMIDZADEH et al.,
2017; CHEN; LIU; CAO, 2017).
As células dendríticas (cCDs) são as principais células
apresentadoras de antígeno (APCs) (ABBAS; LICHTMAN;
PILLAI, 2019). Quando imaturas, as CDs são sentinelas para
patógenos e expressam IL-12, IL-1β, IL-6, e ao reconhecerem
organismos patogênicos por meio dos receptores diversificados
TRLs, iniciam vias de processamento e apresentação do
antígeno às células T naive nos linfonodos, no processo de
ativação da resposta adaptativa (QIAN & CAO, 2018). Várias
moléculas de sinalização intracelular estão envolvidas na
regulação de DCs, como ubiquitinas, cinases (CaMKII), e outras
vias que sinalizam para ativação dos fatores de transcrição
STAT3, BLIMP1 e NFκβ que é ubiquitinado via K27 para

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
338
produção de IL-6, TNF e IL-1, principais citocinas liberadas na
resposta inflamatória, além de IL-23 (DMITRIEVA et al., 2016).
Os mastócitos estão entre as principais células
envolvidas na defesa contra infecções parasitárias, nas reações
de hipersensibilidades e tanto na inflamação aguda quanto na
inflamação crônica. São ativados pelo reconhecimento de
PAMPs e DAMPs através dos receptores TRLs, pela ligação de
anticorpos através dos receptores de alta afinidade FcεRI, para
IgE; FcγRI, para IgG; FcγRIIA e FcγRIII, além de receptores de
hormônios e múltiplos receptores associados à proteína G
(GPCRs). Os mastócitos possuem grânulos citoplasmáticos
contendo aminas vasoativas, como a histamina e enzimas
proteolíticas liberadas em diferentes respostas inflamatórias,
que afetam o início, a manutenção e a resolução da inflamação
(PASTWINSKA et al, 2017; LICHTMAN; PILLAI, 2019).
Os fatores de transcrição GATA1, GATA2, STAT5, NFκβ
e MITF foram descritos como essenciais no desenvolvimento e
função dos mastócitos, bem como o fator de transcrição
C/EBPy da família C/EBP, que é altamente expresso e
importante nas funções dos mastócitos (KARDOSOVA et al.,
2017; SASAKI et al., 2016). Martín-Ávila et al. (2016) também
comprovaram o papel da cinase Fyn como importante fator na
via de sinalização de TRL4 para produção de TNF por
mastócitos. Até o momento era conhecido o papel da cinase Src
nessa via. Nesse sentido, a ativação dos mastócitos via
receptores e ativação dos fatores de transcrição por diferentes
vias levam à produção de mediadores lipídicos, como
prostaglandinas (PGD2, PGE2) e LTC4, além de outras
moléculas envolvidas nas diferentes funções da célula, como
TNF, TGF-β, VEGF, CXCL8 e NO (PASTWINSKA et al, 2017).

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
339
Além das células mieloides envolvidas na inflamação
aguda, diferentes subpopulações de células linfoides inatas
(ILCs) que são residentes nos tecidos, participam na linha de
frente para eliminar patógenos nas barreiras epiteliais. ILC1,
ILC2 e ILC3 são caracterizados pela ausência de receptores
antigênicos específicos (TRABANELLI et al., 2018), por esse
motivo, são ativados por citocinas e produzem diferentes
moléculas semelhantes as produzidas por subpopulações de
linfócitos T (Th1, Th2 e Th17) (ABBAS; LICHTMAN; PILLAI,
2019). As células Natural Killer (NK) e as células indutoras de
tecido linfoide (LTi) também fazem parte das subpopulações de
ILCs, sendo elas dos grupos ILC1 e ILC3, respectivamente
(TRABANELLI et al., 2018).
As ILCs são ativadas pela ligação de diferentes citocinas
liberadas nas respostas inatas, e sua classificação e função
dependem dos fatores de transcrição ativados e das citocinas
produzidas. O grupo ILC1 é caracterizado pela ativação de Tbet
e do gene EOMES para produzir IFN-γ. ILC 2 tem ativação do
fator de transcrição GATA-3, levando à produção de IL-4, IL-5
e IL-13, enquanto que ILC-3 tem ativação de RORγt e NF-Κβ e
produção de IL-17, IL-22 e IL-12. Todos os grupos sofrem
ativação do fator de transcrição NF-κβ, ativando os membros da
família de IL-1, como IL-1β, IL-18 ou IL-33, e em conjunto com
as vias de sinalização de STAT através das vias de IL-23, IL-
12, IFN-α/β e TSLP, desencadeiam a produção de citocinas
auxiliares de ILCs (GASTEIGER et al., 2016).
Os neutrófilos são os primeiros leucócitos a serem
recrutados para o local da inflamação. Quando as células
residentes de tecido são ativadas por DAMPs, são liberados
mediadores inflamatórios (TNF e IL-1) que estimulam a

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
340
produção das quimiocinas CXCL1, CXCL2 e CXL8, as quais se
ligam aos GPCRs nos neutrófilos e em monócitos. Os PRRs,
TLRs e NLRs e receptores Fc, os quais são capazes de
reconhecer mediadores solúveis do sistema complemento e
imunoglobulinas que opsonizam patógenos, também estão
presentes na superfície dos neutrófilos (CASTANHEIRA &
KUBES, 2019; ABBAS; LICHTMAN; PILLAI, 2019).
A liberação das citocinas estimula a expressão das
proteínas de adesão para transmigração dos leucócitos
recrutados. Os neutrófilos utilizam diferentes mecanismos para
eliminar os patógenos, entre eles, a fagocitose e produção de
ROS, proteases (CASTANHEIRA & KUBES, 2019; ABBAS;
LICHTMAN; PILLAI, 2019), e produção intracelular de AMPs no
processo de NETosis. Este processo consiste em armadilhas
extracelulares de neutrófilos (NETs), que são redes
extracelulares de estruturas compostas de proteínas citosólicas
e grânulos, em forma de cromatina descondensada, que
eliminam os patógenos e também estão presentes na patologia
de doenças inflamatórias (PAPAYANNOPOULOS, 2017), tendo
sido identificados na PsO (LOWES; SUÁREZ-FARIÑAS;
KRUEGER, 2014). Em lesões psoríaticas, os neutrófilos são
encontrados predominantemente na epiderme junto com as
quimiocinas CXCL1, CXCL2 e CXCL8/IL-8 (KIM & KRUEGER,
2015).
Psoríase como doença inflamatória com manifestações
cutâneas
O conceito de autoinflamação tem ganhado atenção na
área de dermatologia já que a maioria das doenças com essas

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
341
características apresentam manifestações cutâneas.
Autoinflamação se caracteriza por respostas imunes inatas
mediadas por vias de interleucina-1 (IL-1). Nesse sentido,
evidências comprovam a autoinflamação na patogênese dos
tipos mais clássicos de PsO (MURTHY & LESLIE, 2016).
Lowes, Suárez-Fariña e Krueger (2014), caracterizaram
a patologia da PsO pela presença de queratinócitos que se
movem através da epiderme dez vezes mais rápido que o
normal no processo de diferenciação, com um aumento na
síntese de queratina 16, e presença de neutrófilos na epiderme
e no estrato córneo, formando os microabscessos de Munro. Na
derme, estão presentes células mononucleares e populações
de células T. Logo, o aparecimento de eritemas nas lesões
psoríaticas é resultado da sucessão de mecanismos
imunológicos que levam à dilatação dos vasos sanguíneos
dérmicos. Apesar de alguns mecanismos já serem conhecidos,
ainda não se sabe se a desregulação na biologia de células
imunes e alta produção de citocinas, ou a acelerada
diferenciação de queratinócitos e produção exagerada de
quimiocinas são a raiz desencadeadora da patologia
(MOUNSEY & KULAKOV, 2018). No entanto, por não se
conhecer a origem da imunopatogênese da doença, ainda é
discutido se PsO é uma doença autoimune ou é desencadeada
por estímulos externos (FRY et al., 2015).
A PsO entra no leque de doenças inflamatórias que
afetam a população mundial, e caracteriza-se como uma
doença inflamatória, sistêmica e crônica com manifestações na
pele, ou manifestações não dermatológicas, na forma de
inflamação articular, no caso da Artrite Psoríatica (PsA).
Mesmo afetando cerca de 5% da população mundial

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
342
(MOUNSEY & KULAKOV, 2018), e no Brasil 4,4% (MIOT et al.,
2018), é considerada uma condição comum, onde cerca de 70-
80% dos pacientes apresentam lesões leves e são tratados
apenas com terapias tópicas (BOEHNCKE & SCHON, 2015).
A PsO é uma doença multifatorial, capaz de ser
desencadeada por fatores intrínsecos e extrínsecos, como
injúrias teciduais, traumas ocasionando o fenômeno de
Koebner, infecções por vírus (HIV), ou bactérias do gênero
Streptococcus, fármacos, fatores ambientais (exposição solar e
clima), além de fatores genéticos (BOEHNCKE, 2015), e
epigenéticos. Assim, sua patogênese é resultado dos efeitos
combinados de múltiplos loci de susceptibilidade gênica
associado com fatores ambientais (ZENG et al., 2017).
As lesões psoríaticas se apresentam em cinco tipos de
manifestações clínicas: PsO de placas (vulgaris), forma mais
comum da doença que afeta cerca de 90% dos casos; gutata,
caracterizada por pequenas pápulas eritematosas espalhadas
pelo corpo; invertida, que envolve áreas de dobras, como axilas
e pregas inguinais; postular, associada com comprometimento
das funções cutâneas e geralmente generalizada;
eritrodérmica, caracterizada por eritemas cutâneos distribuídos
pelo corpo. As duas últimas, contudo, são formas raras e mais
severas da doença (BOEHNCKE & SCHON, 2015; LEBWOHL,
2018; SCHLEICHER, 2016). Entretanto, outra manifestação
clínica é citada por Mounsey & Kulakov (2018), a PsO
palmoplantar, que prevalece em 1,5% da população, é uma
variante clínica da PsO de placas e compromete a qualidade de
vida dos pacientes por afetar as mãos e pés (RAPOSO &
TORRES, 2016).

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
343
A manifestação da PsO está associada com o aumento
de risco de diversas comorbidades que impactam física e
emocionalmente a qualidade de vida dos acometidos da
doença. Embora Takeshita et al. (2017) tenham apontado a
disfunção sexual, apneia obstrutiva do sono e doença pulmonar
obstrutiva crônica como comorbidades emergentes na
epidemiologia da doença, condições como diabetes,
hipertensão, síndrome metabólica, infecções, PsA, desordens
cardiovasculares, doença de Crohn, depressão e ansiedade já
são conhecidas e mais comuns nos estudos de relação com a
PsO (SCHLEICHER, 2016; BOEHNCKE & SCHON, 2015).
Imunopatogênese: Células T na Psoríase
As células T têm mostrado desempenhar um papel
principal na imunopatogênese da doença. Além da participação
de células T CD4+ (auxiliares - Th), são encontrados linfócitos
T CD8+ (citotóxicos) nas lesões psoríaticas, capazes de
produzir IL-2, IFN-γ, TNF-α, IL-22 e IL-17 (VOLARIC et al.,
2019). Os linfócitos T reguladores (Tregs) atuam suprimindo e
regulando respostas inflamatórias por meio da liberação de
citocinas anti-inflamatórias como IL-10 e TGF-B (KIM &
KRUEGER, 2015). Karczewski et al. (2016) discutem que
mesmo havendo a presença de células Tregs nas lesões de
PsO, estas não são capazes de controlar a inflamação,
sugerindo um defeito qualitativo na regulação de outras células
imunes. Dessa forma, acredita-se que a ativação das
subpopulações Th1 e Th17 medeia o processo inflamatório na
psoríase (ZEICHNER & ARMSTRONG, 2016).

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
344
As citocinas IL-12 e INF-γ estimulam a diferenciação de
células T CD4+ em Th1, as quais produzem IFN-γ e TNF-α. A
diferenciação de TCD4+ em Th17 é estimulada por IL-6, IL-1 e
IL-23. Assim, a subpopulação produz IL-22, IL-6, IL-20 e TNF-
α. (KARCZEWSKI et al., 2016; ABBAS; LICHTMAN; PILLAI,
2019). A liberação destas citocinas promove a proliferação de
queratinócitos, que produzem citocinas pró-inflamatórias.
Queratinócitos na Psoríase
Os queratinócitos são a principal fonte de citocinas
inibitórias, as quais mantém o potencial inflamatório da pele em
equilíbrio. Produzem peptídeos antimicrobianos, como
catelicidina LL-37, defensinas, proteínas S100 (KIM &
KRUEGER, 2015), e controlam respostas imunes pela liberação
de CXCL8, a qual é relacionada com o recrutamento de
neutrófilos; CCL2, CCL5, CXCL10, responsáveis por atrair
monócitos e células Th1; CCL20 e IL-8, que recrutam CDs e
células de Langerhans (BENHADOU; MINTOFF; DEL
MARMOL, 2019).
A IL-17R é expressa principalmente por queratinócitos na
PsO. Os genes IL-17, IFN-α, IL-22, IL-1, TNF, e IFN-γ foram
observados sendo expressos em queratinócitos em resposta a
citocinas presentes nas lesões psoríaticas, enquanto que
destes, IL-17, IL-22, TNF e IFN-γ mostraram causar a
proliferação dos queratinócitos, e induzir expressão de
citocinas, quimiocinas e AMPs (LOWES; SUÁREZ-FARIÑAS;
KRUEGER, 2014).

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
345
Células Dendríticas na Psoríase
As principais populações de células dendríticas na pele
são as células de Langerhans na epiderme (LC), células
dendríticas mieloides (mCD) e células dendríticas
plasmocitoides (pCD) na derme. Estudos recentes têm
apontado que as CDs exercem um papel no início da psoríase,
pois são encontrados múltiplos subtipos celulares nas lesões
(WANG & BAI, 2019). Tendo em vista que a citocina IL-23 é
necessária para sobrevivência de células Th17, pesquisadores
mostraram que injeções de IL-23 em camundongos aumentam
a população de CD103+ cCDs, monócitos e moCDs (CDs
dérmicas derivadas de monócitos) na derme. Dessa forma,
estas atuam diretamente produzindo citocinas como IL-23, IL-
22 e TNF que ativam células T αβ e γδ para produzirem IL-22 e
IL-17A/F, as quais estimulam a proliferação de queratinócitos e
liberação de fatores de crescimento, proteínas S100 e β-
defensinas, amplificando a cascata da doença (SINGH et al.,
2016).
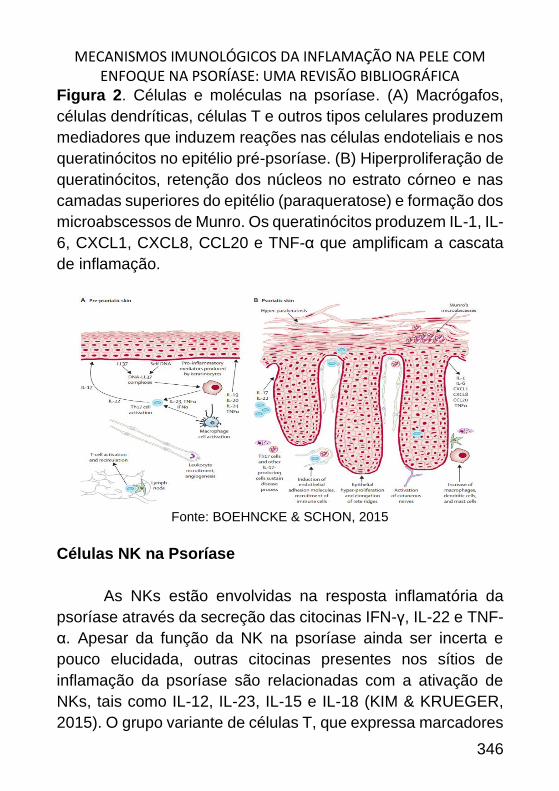
MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
346
Figura 2. Células e moléculas na psoríase. (A) Macrógafos,
células dendríticas, células T e outros tipos celulares produzem
mediadores que induzem reações nas células endoteliais e nos
queratinócitos no epitélio pré-psoríase. (B) Hiperproliferação de
queratinócitos, retenção dos núcleos no estrato córneo e nas
camadas superiores do epitélio (paraqueratose) e formação dos
microabscessos de Munro. Os queratinócitos produzem IL-1, IL-
6, CXCL1, CXCL8, CCL20 e TNF-α que amplificam a cascata
de inflamação.
Fonte: BOEHNCKE & SCHON, 2015
Células NK na Psoríase
As NKs estão envolvidas na resposta inflamatória da
psoríase através da secreção das citocinas IFN-γ, IL-22 e TNF-
α. Apesar da função da NK na psoríase ainda ser incerta e
pouco elucidada, outras citocinas presentes nos sítios de
inflamação da psoríase são relacionadas com a ativação de
NKs, tais como IL-12, IL-23, IL-15 e IL-18 (KIM & KRUEGER,
2015). O grupo variante de células T, que expressa marcadores

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
347
(CD56) presentes em células NK (ABBAS; LICHTMAN; PILLAI,
2019), chamado NKT, possivelmente desempenha algum papel
no processo inflamatório liberando IFN-γ (KIM & KRUEGER,
2015).
Loci de susceptibilidade gênica
O alelo HLA-cw6 está associado com as características
fenotípicas da psoríase e está presente em cerca de 40 a 80%
dos casos. O alelo apresenta resposta positiva ao anticorpo
monoclonal Ustecinumab, utilizado para neutralizar IL-17A,
tratando dos casos mais moderados aos mais severos da PsO
(COSTANZO et al., 2018). De Keyser et al. (2019), ao
verificarem o papel do Ustecinumab, genotiparam HLA-cw6 em
seus experimentos e obtiveram o status de atividade do HLA-
cw6 em 61 pacientes dentro de uma população de 137. Dos
41% da população, os HLA-cw6-positivo desenvolveram
psoríase cedo, enquanto que a prevalência de Artrite Psoríatica
em pacientes HLA-cw6-negativo foi maior, mas não
significativo.
Chen & Tsai (2018) também apontaram a relação do
HLA-cw6 com dois autoantígenos recém identificados na
psoríase, catelicidina LL-37, peptídeo derivado de
queratinócitos, com um papel antimicrobiano, e capacidade de
formar um complexo com o DNA/RNA próprio ou não próprio e
ativar células dentríticas; e ADAMTSL5, uma proteína
importante para função das microfibrilas, que é apresentada por
HLA-cw6 resultando na expansão clonal de células T CD8+ nas
lesões psoríaticas. (FUENTES-DUCULAN et al., 2017).

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
348
Outros loci de susceptibilidade gênica da PsO (PSORS)
têm sido estudados, sendo 12 já identificados. Um locus
localizado no cromossomo 6p, PSORS1, é fortemente
associado com HLA-cw6, se tornando o alelo mais evidenciado
na PsO até o momento (CHEN & TSAI, 2018; LOWES;
SUÁREZ-FARIÑAS; KRUEGER, 2014; BOEHNCKE & SCHON,
2015). Genes que são encontrados nestes loci, contudo, estão
envolvidos com a síntese de citocinas presentes na psoríase
(BOEHNCKE & SCHON, 2015). O gene CARD14 é altamente
expresso em quaratinócitos e foi identificado como gene no
locus de susceptibilidade gênica da psoríase 2 (PSORS2).
Variantes e mutações desse gene estão associados com as
variações no espectro da psoríase (ISRAEL & MELLETT,
2018).
CONCLUSÕES
Os recentes estudos do funcionamento do sistema
imunológico, do background genético, bem como dos princípios
e mecanismos dos mediadores celulares no processo
inflamatório, têm sido fundamentais para elucidação da
imunopatologia de diversas doenças inflamatórias, cutâneas e
autoimunes. A psoríase entra nesse leque de doenças
inflamatórias com manifestações cutâneas, com a presença de
células e moléculas imunes envolvidas na imunidade inata e
adaptativa. Embora algumas vias de mediadores inflamatórios
já tenham sido identificadas na doença, não se sabe ao certo
qual o mecanismo desencadeador, mas sim que pode ser
afetada por fatores intrínsecos e extrínsecos. Nesse sentido,
entender a imunopatogênese da psoríase, assim como

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
349
entender como se dá a relação da condição com outras
comorbidades no paciente, é essencial para melhor
compreensão da função e atividade dos alvos terapêuticos,
para elaboração de tratamentos eficazes para todas as
variantes dentro do espectro da psoríase.
REFERÊNCIAS BIBLIOGRÁFICAS ABBAS, A. K.; LICHTMAN, A. H.; PILLAI, S. H. I. V. Imunologia celular e molecular. 9. ed. Rio de Janeiro: Elsevier, 2019. and Jandus C. Human Innate Lymphoid Cells (ILCs): Toward a Uniform Immune-
Phenotyping. Cytometry Part B
and Jandus C. Human Innate Lymphoid Cells (ILCs): Toward a Uniform Immune-
Phenotyping. Cytometry Part B
and Jandus C. Human Innate Lymphoid Cells (ILCs): Toward a Uniform Immune-
Phenotyping. Cytometry Part B
and Jandus C. Human Innate Lymphoid Cells (ILCs): Toward a Uniform Immune-
Phenotyping. Cytometry Part B
and Jandus C. Human Innate Lymphoid Cells (ILCs): Toward a Uniform Immune-
Phenotyping. Cytometry Part B
and Jandus C. Human Innate Lymphoid Cells (ILCs): Toward a Uniform Immune-
Phenotyping. Cytometry Part B
BASIL, M. C.; LEVY, B. D. Specialized pro-resolving mediators: endogenous regulators of infection and inflammation. Nature Reviews Immunolology, v.16, n. 1, p.51–67, 2016. BENHADOU, F.; MINTOFF, D.; DEL MARMOL, V. Psoriasis: Keratinocytes or Immune Cells – Which Is the Trigger? Dermatology, v. 235 ,n. 2, p.91–100, 2019. BOEHNCKE, W.H. Etiology and Pathogenesis of Psoriasis. Rheumatic Disease Clinics of North America, v. 41, n.4, p. 665–675, 2015. BOEHNCKE, W.H.; SCHÖN, M. P. Psoriasis. The Lancet, v. 386, n. 9997, p. 983–994, 2015. BORSIG, L. Selectins in cancer immunity. Glycobiology, v. 28, n.9, p. 648–655, 2018. CAO, X. Self-regulation and cross-regulation of pattern-recognition receptor signalling in health and disease. Nature Reviews Immunology, v. 16, n.1, p. 35–50, 2016. CASTANHEIRA, F. V. S.; KUBES, P. Neutrophils and NETs in modulating acute and chronic inflammation. Blood, v. 133, n. 20, p. 2178-2185, 2019.

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
350
CHEN, K.; LIU, J.; CAO, X. Regulation of type I interferon signaling in immunity and inflammation: A comprehensive review. Journal of Autoimmunity, v. 83, p. 1–11, 2017. CHEN, L.; TSAI, T. F. HLA-Cw6 and psoriasis. British Journal of Dermatology, v. 178, n.4, p. 854–862, 2018. COSTANZO, A. et al. Secukinumab shows high efficacy irrespective of HLA-Cw6 status in patients with moderate-to-severe plaque-type psoriasis: SUPREME study. British Journal of Dermatology, v. 179, n. 5, v. 1072-1080, 2018. CRUSZ, S. M.; BALKWILL, F. R. Inflammation and cancer: advances and new agents. Nat Rev Clin Oncol, v .12, n. 10, p. 584–596, 2015. DE KEYSER, E. et al. Clinical Consequences of Antibody Formation, Serum Concentrations, and HLA-Cw6 Status in Psoriasis Patients on Ustekinumab. Therapeutic drug monitoring, v. 41, n. 5, p. 634-639, 2019. DMITRIEVA, O. S. et al. Interleukins 1 and 6 as main mediators of inflammation and cancer. Biochemistry, v.81, n.2, p. 80–90, 2016. e B, Michaud K, Mavilio D, Landis BN, Jandus P.
e B, Michaud K, Mavilio D, Landis BN, Jandus P.
e B, Michaud K, Mavilio D, Landis BN, Jandus P.
e B, Michaud K, Mavilio D, Landis BN, Jandus P.
e B, Michaud K, Mavilio D, Landis BN, Jandus P.
e B, Michaud K, Mavilio D, Landis BN, Jandus P.
FRY, L. et al. Psoriasis is not an autoimmune disease?. Experimental dermatology, v. 24, n. 4, p. 241-244, 2015. FUENTES-DUCULAN, J. et al. Autoantigens ADAMTSL5 and LL37 are significantly upregulated in active Psoriasis and localized with keratinocytes, dendritic cells and other leukocytes. Experimental Dermatology, v. 26, n.11, p. 1075–1082, 2017. GASTEIGER, G. et al. Cellular Innate Immunity: An Old Game with New Players. Journal of Innate Immunity,v. 9, n. 2, p. 111–125, 2016. GERHARDT, T.; LEY, K. Monocyte trafficking across the vessel wall. Cardiovascular Research, v. 107, n. 3, p. 321–330, 2015. GRAVITZ, L. Skin. Nature outlook, v. 563, n. 7732, p. S83, 2018. HAMIDZADEH, K. et al. Macrophages and the Recovery from Acute and Chronic Inflammation. Annual Review of Physiology, v.79, n.1, p. 567–592, 2017. ISAILOVIC, N. et al. Interleukin-17 and innate immunity in infections and chronic inflammation. J. Autoimmun, v. 60, p. 1-11, 2015. ISRAEL, L.; MELLETT, M. Clinical and Genetic Heterogeneity of CARD14 Mutations in Psoriatic Skin Disease. Front. Immunol, v. 9, p. 2239, 2018.

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
351
KARCZEWSKI, J. et al. New insights into the role of T cells in pathogenesis of psoriasis and psoriatic arthritis. Autoimmunity, v. 49, n.7, p. 435–450, 2016. KARDOSOVA, M. et al. The transcription factor C/EBPγ regulates mast cell development and function. Experimental Hematology, v. 53, p. S92, 2017. KIM, J.; KRUEGER, J. G. The Immunopathogenesis of Psoriasis. Dermatologic Clinics, v. 33, n.1, p. 13–23, 2015. KORNILUK, A. et al. From inflammation to cancer. Ir J Med Sci, v. 186, n.1, p. 57–62, 2017. KUMARI, S.; PASPARAKIS, M. Epithelial Cell Death and Inflammation in Skin. Apoptotic and Non-Apoptotic Cell Death, v. 403, p. 77–93, 2015. LEBWOHL, M. Psoriasis. Annals of Internal Medicine, v. 168, n. 7, p.49-64, 2018. LIU, J.; CAO, X. Cellular and molecular regulation of innate inflammatory responses. Cellular & Molecular Immunology, v. 13, n. 6, p. 711–721, 2016. LOWES, M. A.; SUÁREZ-FARIÑAS, M.; KRUEGER, J. G. Immunology of psoriasis. annual review of immunology, v. 32, n. 1, p. 227–255, 2014. MARTÍN-ÁVILA, A. et al. Protein Tyrosine Kinase Fyn Regulates TLR4-Elicited Responses on Mast Cells Controlling the Function of a PP2A-PKCα/β Signaling Node Leading to TNF Secretion. The Journal of Immunology, v. 196, n.12, p. 5075–5088, 2016. MIOT, H. A. et al. Profile of dermatological consultations in Brazil. Anais Brasileiros de Dermatologia, v. 93, n. 6, p. 916–928, 2018. MOUNSEY, S. J.; KULAKOV, E. Psoriasis. British Journal of Hospital Medicine, v. 79, n. 8, p. C114–C117, 2018. MULLER, W. A. Transendothelial migration: unifying principles from the endothelial perspective. Immunological Reviews, v. 273, n.1, p. 61–75, 2016. MURTHY, A. S.; LESLIE, K. Autoinflammatory Skin Disease: A Review of Concepts and Applications to General Dermatology. Dermatology, v. 232, n. 5, p. 534–540, 2016. O’MALLEY, J. T.; CLARK, R. A.; WIDLUND, H. R. Meeting report - Skin Inflammation in Human Health and Disease: 2018 International Conference. Journal of Investigative Dermatology, v. 139, n.5, p. 991-994, 2019. PAPAYANNOPOULOS, V. Neutrophil extracellular traps in immunity and disease. Nature Reviews Immunology, v. 18, n.2, p.134–147, 2017. PASTWIŃSKA, J. et al. Mast cells as the strength of the inflammatory process. Polish Journal of Pathology, v. 68, n.3, p.187–196, 2017. QIAN, C.; CAO, X. Dendritic cells in the regulation of immunity and inflammation. Seminars in Immunology, v. 35, p. 3–11, 2018.

MECANISMOS IMUNOLÓGICOS DA INFLAMAÇÃO NA PELE COM ENFOQUE NA PSORÍASE: UMA REVISÃO BIBLIOGRÁFICA
352
rabanelli S, Gomez-Cadena A, Salom
rabanelli S, Gomez-Cadena A, Salom
rabanelli S, Gomez-Cadena A, Salom
RAPOSO, I.; TORRES, T. Palmoplantar Psoriasis and Palmoplantar Pustulosis: Current Treatment and Future Prospects. American Journal of Clinical Dermatology, v. 17, n. 4, p. 349–358, 2016. SABAT, R. et al. T cell pathology in skin inflammation. Semin Immunopathol, v. 41, n. 3, p. 359–377, 2019. SASAKI, H.; KUROTAKI, D.; TAMURA, T. Regulation of basophil and mast cell development by transcription factors. Allergology International, v. 65, n. 2, p. 127–134, 2016. SCHLEICHER, S. M. Psoriasis. Clinics in Podiatric Medicine and Surgery, v. 33, n. 3, p. 355–366, 2016. SINGH, T. P. et al. Monocyte-derived inflammatory Langerhans cells and dermal dendritic cells mediate psoriasis-like inflammation. Nature Communications, v. 7, p. 13581, 2016. TAKESHITA, J. et al. Psoriasis and comorbid diseases. Journal of the American Academy of Dermatology, v. 76, n. 3, p. 377–390, 2017. TODD, I.; SPICKETT, G.; FAIRCLOUGH, L. Lecture Notes: Immunology. 7. ed. Nova York: John Wiley & Sons, 2015. Trabanelli S, Gomez-Cadena A, Salom
Trabanelli S, Gomez-Cadena A, Salom
Trabanelli S, Gomez-Cadena A, Salom
TRABANELLI, S. et al. Human Innate Lymphoid Cells (ILCs): Toward a Uniform Immune-Phenotyping. Cytometry B Clin Cytom, v. 94, n. 3, p. 392-399, 2018. VOLARIĆ, I,; VIČIĆ, M.; PRPIĆ-MASSARI, L. The Role of CD8+ T-Cells and their Cytokines in the Pathogenesis of Psoriasis. Acta Dermatovenerol Croat, v. 27, n. 3, p.159-162, 2019. WANG, A; BAI, Y. Dendritic cells: The driver of psoriasis. J Dermatol, v. 47, n. 2, p. 104-113, 2020. ZEICHNER, J. A.; ARMSTRONG, A. The Role of IL-17 in the Pathogenesis and Treatment of Psoriasis. The Journal of clinical and aesthetic dermatology, v. 9, n. 6, p. S3–S6, 2016. ZENG, J. et al. Critical role of environmental factors in the pathogenesis of psoriasis. The Journal of Dermatology, v. 44, n. 8, p. 863–872, 2017.

SARS-COV-2: COMORBIDADES E COINFECÇÃO
353
CAPÍTULO 17
SARS-COV-2: COMORBIDADES E COINFECÇÃO
Fernanda Silva ALMEIDA1
Bárbara Guimarães CSORDAS2 Ericka Garcia LEITE3
Daniele Idalino JANEBRO4 Tatjana Souza Lima KEESEN5
1 Doutoranda do Programa de Pós-Graduação em Biotecnologia- RENORBIO/ UFPB; 2Pós Doutoranda do Programa de Pós-Graduação em Produtos Naturais e Sintéticos Bioativos - pgPNBS - UFPB; 2 Doutoranda do Programa de Pós-Graduação Multicêntrico em Ciências
Fisiológicas 3Doutora em produtos naturais e sintéticos bioativos Professora de hematologia e citologia clínica DCF/ CCS/ UFPB 4Orientadora/Professora do
Departamento de Biologia Molecular e Celular, CBiotec/UFPB [email protected]
RESUMO: Em março de 2020 foi declarado pela Organização mundial de Saúde a COVID-19, causada pelo novo coronavírus da síndrome respiratória aguda grave 2 (SARS-CoV-2) como pandemia global. O impacto causado pelo COVID-19 tem sido devastador, sendo contabilizado mais de 38 milhões de casos confirmados e mais de 1 milhão de mortos, onde o Brasil está entre os países com maior número de casos. Uma base de evidências emergentes identificou vários fatores associados a resultados adversos para pessoas com COVID-19. O presente trabalho tem como objetivo uma revisão acerca da infeção por SARS-Cov-2 e sua relação com comorbidades e coinfecção com a tuberculose que são responsáveis pelo agravamento da doença. Trata-se de uma revisão sistemática de caráter descritivo fundamentado na coleta de dados por meio de uma revisão bibliográfica. Observou-se que a idade avançada é o fator de risco mais consistente para a gravidade da doença. Indivíduos idosos estão mais propensos a desenvolverem a forma grave da doença por apresentarem comorbidades crônicas associadas, sendo a hipertensão a doença de base

SARS-COV-2: COMORBIDADES E COINFECÇÃO
354
mais comum, seguida do diabetes mellitus. A obesidade está entre uma das comorbidades que influenciam na gravidade da doença por afetar a maioria dos processos fisiológicos e modificar as funções do sistema imunológico. Ademais, manifestações graves podem também estar associadas a coinfecções com outros patógenos pulmonares como Mycobacterium tuberculosis. Palavras-chave: SARS-CoV-2. Hipertensão. Diabetes. Obesidade. Tuberculose.
INTRODUÇÃO
Coronavírus são vírus que possuem partículas
envelopadas com genomas de RNA de fita simples que
pertencem a uma grande família de vírus respiratórios, podem
causar doenças que variam desde sintomas de resfriado
comum à forma mais grave, a Síndrome Respiratória do Oriente
Médio (MERS) e à Síndrome Respiratória Aguda Grave
(SARS). O coronavírus SARS (SARS-CoV-1) surgiu em
novembro de 2002 no sudeste da China, e causou síndrome
respiratória aguda grave (SARS), desaparecendo em 2004. Já
a síndrome respiratória do Oriente Médio (MERS) é causada
pelo coronavírus MERS (MERS-CoV), ambos pertencentes a
família beta-coronavirus. O MERS foi identificado em setembro
de 2012, possui uma taxa de mortalidade de 35% e continua a
causar surtos esporádicos e localizados. O SARS-CoV-2 é um
novo membro do gênero betacoronavírus distinto do SARS-CoV
e do MERS-CoV (CHEN et al., 2020).
A COVID-19, nome dado a doença causada pelo SARS-
CoV-2 colocou as autoridades de saúde em alerta máximo na
China (CHEN et al., 2020). Já foram confirmados até o dia 25
de outubro do ano corrente, mais de 42 milhões de casos para
COVID-19 sendo mais de 1 milhão de mortes, onde o Brasil está

SARS-COV-2: COMORBIDADES E COINFECÇÃO
355
entre os países com maior número de casos, aproximadamente
5 milhões de casos e 156 mil mortes até a presente data (WHO,
2020). Como uma emergência de saúde pública de interesse
internacional, a COVID-19 foi declarada uma pandemia pela
Organização Mundial da Saúde.
Quanto a sintomatologia, a COVID-19 apresenta
manifestações clínicas presentes: febre, tosse seca, mialgia e
fadiga mais comumente observadas no início da doença;
menos comumente observados são a produção de escarro,
hemoptise, cefaleia, tontura, dor abdominal, diarreia, náuseas e
vômitos. Semelhante à SARS e MERS, alguns pacientes
desenvolvem a síndrome respiratória aguda grave (SRAG).
Cerca de 81% dos pacientes infectados desenvolvem apenas
sintomas leves, com pneumonia leve ou ausência de
pneumonia; os que apresentam estado grave e crítico são 14 e
5%, respectivamente (CHEN et al., 2020; HARAPAN et al,
2020).
Embora os fatores de risco de COVID-19 permaneçam
obscuros, estudos relataram que uma proporção significativa de
pacientes possuía doenças subjacentes. Estudos mostram que
pacientes com pneumonia causada por SARS-CoV
apresentavam comorbidades crônicas, sendo a hipertensão a
doença de base mais comum, seguida do diabetes mellitus. A
maioria dos pacientes com COVID-19 são adultos entre 34 e 59
anos. Os pacientes que requerem cuidados em UTI tendem a
ser idosos. Ademais, há uma prevalência maior de homens com
COVID-19 do que de mulheres. A presença de comorbidades e
mortalidade da doença, portanto, têm sido associadas as
doenças crônicas não transmissíveis e são importantes fatores
de risco para a gravidade e prognóstico da doença (CHEN et
al., 2020; LAI et al., 2020).

SARS-COV-2: COMORBIDADES E COINFECÇÃO
356
Além disso, manifestações graves podem também estar
associadas a coinfecções de bactérias e fungos (LAI et al.,
2020). A coinfecção com outros patógenos pulmonares já foi
observada em pacientes com SARS. Além disso, casos de
coinfecção entre tuberculose e MERS também foram descritos,
demonstrando que ocorrem coinfeções em vírus semelhantes a
SARS-CoV-2, causadores de problemas respiratórios e/ou
pulmonares (ALFARAJ et., 2017).
OBJETIVOS
Este trabalho tem como objetivo, trazer uma revisão
sistemática da literatura da relação entre pacientes com
COVID-19, comorbidades e coinfecção.
MATERIAIS E MÉTODO
Este estudo trata-se de uma revisão sistemática da
literatura de caráter descritivo fundamentado na coleta de
dados por meio de uma revisão bibliográfica. Sendo assim,
refere-se a uma pesquisa documental com base em dados
secundários obtidos a partir de cinco bancos de dados online:
Elsevier, Google Scholar, PubMed, Nature.
Os artigos incluídos nesta pesquisa foram todos originais
de língua inglesa, que abordaram a temática COVID-19,
comorbidades, diabetes, hipertensão, obesidade e coinfecção
entre SARS-Cov-2 e Mycobacterium tuberculosis.

SARS-COV-2: COMORBIDADES E COINFECÇÃO
357
RESULTADOS E DISCUSSÃO
HIPERTENSÃO
A hipertensão arterial (HA) é o principal fator de risco
modificável para doenças cardiovasculares e morte prematura
em todo o mundo. É uma condição clínica multifatorial
caracterizada por elevação sustentada dos níveis pressóricos
(PA) ≥ 140 e/ou 90 mm Hg (MALACHIAS et al., 2016). De
acordo com a Organização Mundial de Saúde cerca de 1,13
bilhão de pessoas têm HA no mundo, onde menos de 1 a cada
5 pessoas têm o problema sob controle (WHO, 2019).
A HA pode ser dividida em dois tipos: primária e
secundária. A hipertensão secundária é provocada
secundariamente por uma causa identificável. Porém na grande
maioria dos pacientes a etiologia não é reconhecida. Esses
pacientes são agrupados coletivamente no que é conhecido
como hipertensão primária ou essencial que, apesar das
melhores opções terapêuticas, não é controlada em 8–12% dos
pacientes (HERRMANN; TEXTOR, 2019).
A incidência de HA aumenta significativamente com o
aumento da idade (afetando aproximadamente 70% dos adultos
mais velhos). Portanto, é mais prevalente entre adultos mais
velhos do que na população jovem e de meia-idade. Numerosos
inquéritos de base populacional em várias regiões do mundo
estimam que 7 em cada 10 adultos, com 65 anos ou mais, foram
diagnosticados com pressão arterial elevada (KARIO et al.,
2020; MULI et al., 2020).
A HA é uma comorbidade mais frequentemente
observadas em pacientes com COVID-19 levantando muitas

SARS-COV-2: COMORBIDADES E COINFECÇÃO
358
questões sobre a maior suscetibilidade desses pacientes ao
novo coronavírus (CHAN et al., 2020).
Os dados provenientes predominantemente da China
mostram um percentual significativo de hipertensão arterial e
doenças cardiovasculares (DCV) entre os pacientes com
COVID-19. Em Wuhan na China onde o primeiro surto de
COVID-19 ocorreu, em dezembro de 2019, um estudo
envolvendo 548 pacientes internados, a prevalência de
hipertensão foi significativamente maior em pacientes com
COVID-19 grave do que em casos não graves (38,7% vs.
22,2%) (SHIBATA et al., 2020). Em outro estudo realizado por
Wang e colaboradores, analisaram 344 pacientes na UTI com
COVID-19. Dos 344 indivíduos, 133 pacientes morreram no 28º
dia com uma sobrevida média de 25 dias. Desses 344
pacientes, haviam 141 pacientes com HA, significativamente
distribuída entre não sobreviventes (69 [52,3%]) e
sobreviventes (72 [34,1%]) (WANG et al., 2020).
A Itália foi o primeiro, e por muito tempo, o país europeu
mais gravemente afetado pela COVID-19. Em 20 de março de
2020, o Instituto Italiano de Saúde anunciou que haviam 3.200
mortos por COVID‐19. Os pacientes que morreram tinham
idade média de 78,5 anos e 98,7% tinham pelo menos uma
comorbidade, sendo a HAuma comorbidade comum nos casos
italianos, afetando 73,8% dos pacientes (KARIO et al., 2020).
Nos Estados Unidos, em um estudo de uma série de
casos de COVID-19, onde foram avaliados 5700 pacientes
hospitalizados sequencialmente com idade média de 63 anos,
3.026 (56,6%) tinham hipertensão (RICHARDSON et al., 2020).
A prevalência de hipertensão é clinicamente relevante
em pacientes com COVID-19 e, particularmente, em idosos. O
impacto da hipertensão e no resultado ainda é incerto. Não está

SARS-COV-2: COMORBIDADES E COINFECÇÃO
359
claro se a HA está relacionada com a gravidade da doença ou
se a pressão arterial controlada é ou não um fator de risco
menor. No entanto, várias organizações já enfatizaram o fato de
que o controle da pressão arterial continua sendo uma
consideração importante para reduzir a carga da doença,
mesmo que não tenha efeito sobre a suscetibilidade à infecção
viral SARS-CoV-2 (SCHIFFRIN et al., 2020).
DIABETES
Risco, gravidade, presença de comorbidades e
mortalidade da doença têm sido associados a doenças crônicas
não transmissíveis, como diabetes mellitus (DM), a qual tem
uma elevada morbidade no Brasil, o que tem causado grande
preocupação. Muitos estudos permitirão esclarecer os perfis
clínicos da infecção por SARS-CoV-2, sendo verificado que a
presença de morbidades crônicas como o DM e suas doenças
associadas (obesidade, hipertensão, doenças cardiovasculares
e doença renal crônica) são importantes fatores de risco para a
gravidade e prognóstico da doença (PITITT; FERREIRA, 2020).
O Brasil é o quarto país com o maior número de pessoas
diabéticas e no ano de 2019 foi estimado que 16,8 milhões de
indivíduos, com idade entre 20 e 79 anos, tinham DM no Brasil,
com projeção de aumento de 55% até 2045 (ATLAS, 2019).
Outro aspecto do DM no Brasil é que metade dos
brasileiros diabéticos não tem diagnóstico, as frequências
existentes são subestimadas e as mesmas variam de acordo
com a faixa etária e escolaridade, sendo 23,1% para maiores
de 65 anos e 15,2% para aqueles com menos de 8 anos de
escolaridade total (BRASIL, 2019).

SARS-COV-2: COMORBIDADES E COINFECÇÃO
360
Também a falta do controle glicêmico dos pacientes com
DM no Brasil é preocupante. A Sociedade Brasileira de
Diabetes recomenda como meta a dosagem da hemoglobina
glicada (HbA1c) <7%, no entanto, este valor é alcançado por
uma minoria de pacientes prejudicando vários aspectos da
resposta imune inata e adaptativa às infecções virais e ao
potencial de infecção bacteriana secundária nos pulmões
(CAREY et al., 2018; FERLITA et al., 2019). Ou seja, a ação
inadequada das células T, o comprometimento da atividade das
células natural killer e defeitos na ação do complemento, podem
reduzir a depuração viral (NYAMBUYA et al., 2020). Além disso,
o DM representa um importante fator de risco para as doenças
cardiovasculares (DCV), responsáveis por 31,3% das mortes no
Brasil (SCHMIDT et al., 2011).
Este cenário de alta prevalência de DM associado ao
diagnóstico tardio e controle metabólico alterado tem um efeito
importante nas complicações a longo prazo da doença
(macroangiopatia, retinopatia, nefropatia e neuropatia) e desta
forma, considerando a pandemia da COVID-19, estes fatores,
pioram o prognóstico dos diabéticos afetados pela infecção por
SARS-Cov-2 (PITITT; FERREIRA, 2020).
Desde os primeiros relatos da COVID-19 em Wuhan,
China, foram verificadas altas frequências de indivíduos
diabéticos entre os pacientes internados e com desfecho para
óbito, mostrando o DM como fator de risco importante, seguido
também por outros fatores, como idade avançada, obesidade,
hipertensão e DCV. Vários estudos relatam estes achados:
estudo chinês mostra que de 1.099 indivíduos com COVID-19,
dos 173 pacientes com doença severa, 23,7% tinha
hipertensão, 16,2% DM, 5,8% doença cardíaca coronária e
2,3% doença cerebrovascular (GUAN et al., 2020). Outra

SARS-COV-2: COMORBIDADES E COINFECÇÃO
361
pesquisa com 140 pacientes internados relata taxas
semelhantes, 30% de hipertensão e 12% de DM (ZHANG et al.,
2020). Wu; Mcgoogan (2020) avaliaram um total de 44.672
casos de COVID-19, no qual a letalidade ocorreu em 14,8% dos
pacientes com mais de 80 anos, 8,0% entre 70 e 79 anos e em
49,0% dos casos graves. Entre aqueles que morreram e tinha
doenças crônicas, 10,5% tinha DCV, 7,3% DM, 6,3% doença
respiratória crônica, 6,0% hipertensão e 5,6% câncer.
Com o avanço da pandemia nos outros continentes, a
mesma situação foi confirmada em relação ao fator de risco DM
e mau prognóstico para COVID-19. Na Itália, entre 1.591
pacientes em UTI, 68% apresentavam pelo menos uma
comorbidade, 49% hipertensão sistêmica, 21% DCV, 17% DM
tipo 2, 8% câncer e 4% DPOC (GRASSELLI et al., 2020).
Nos EUA, foi observado que 89,3% de 178 adultos
apresentaram uma ou mais comorbidades; a mais comum foi
hipertensão (49,7%), seguida de obesidade (48,3%), doença
pulmonar crônica (34,6%), DM (28,3%) e DCV (27,8%) (GARG
et al., 2020). O mesmo cenário foi visto no Brasil, dados do
Ministério da Saúde identificaram DM e DCV prévia como
comorbidades associadas à morte em indivíduos infectados
pelo SARS-Cov-2 (BRASIL, 2020).
A relação entre COVID-19 e doenças cardiometabólicas
é baseado em estudos anteriores que mostraram a associação
de DM e CVD em epidemias de influenza, H1N1 ou MERS
(MADIID et al., 2020), e este conhecimento esclarece os
mecanismos subjacentes da doença associados aos fatores de
risco, permitindo assim o desenvolvimento de estratégias de
prevenção mais eficazes para enfrentar epidemias virais, dentre
elas, o controle da pandemia do coronavírus.

SARS-COV-2: COMORBIDADES E COINFECÇÃO
362
É visto relação direta da alteração do metabolismo da
glicose com a gravidade da COVID-19, sendo assim, o controle
glicêmico adequado reduz a predisposição e melhora o
prognóstico das infecções em pessoas com DM, diminuindo os
resultados ruins e morte nestes pacientes pela COVID-19, uma
vez que DM causa um estado pró-inflamatório podendo
acentuar a tempestade de citocinas, bem como disfunção de
múltiplos órgãos (MADDALONI; BUZZETTI, 2020),
Assim, como um alto grau de inflamação sistêmica pode
ocorrer na COVID-19, visto por elevações em importantes
marcadores inflamatórios, como proteína C-reativa, dímero-D e
ferritina (YANG et al., 2020), além de níveis aumentados de
plasminogênio (JI et al., 2020). Desta forma, o DM e suas
doenças associadas intensificam o processo inflamatório,
contribuindo para a progressão da COVID-19 em indivíduos
diabéticos.
No mesmo contexto, a infecção por SARS-CoV-2 e o
mecanismo de ação de alguns medicamentos foram baseados
nas vias fisiopatológicas do DM e das comorbidades
associadas, como, por exemplo, a hipertensão. Os coronavírus
são capazes de se ligar às células-alvo por meio da enzima
conversora de angiotensina 2 (ECA 2) que é expresso em
vários tecidos. Estudos mostram que a ECA 2 está aumentada
em indivíduos com DM tipo 1 e tipo 2 e que alguns
medicamentos podem aumentar a expressão de ECA 2, como
inibidores de ECA, bloqueadores do receptor de angiotensina
2, tiazolidinedionas e ibuprofeno (FANG; KARAKIULAKIS;
ROTH, 2020). No entanto, nenhuma evidência indica potenciais
efeitos adversos desses agentes no prognóstico de indivíduos
diabéticos e hipertensos com COVID-19, e desta forma, a

SARS-COV-2: COMORBIDADES E COINFECÇÃO
363
comunidade científica não recomendam a retirada dessas
drogas devido à infecção por SARS-CoV-2 (SBD, 2020).
O processo fisiopatológico do DM e COVID-19 precisa ser
bem entendido para que medidas preventivas e estratégias
terapêuticas eficazes possam ser estabelecidas, incluindo as as
variações individuais, regionais e étnicas na prevalência e no
curso natural da doença. A gravidade da infecção por DM e
SARS-CoV-2 foi demonstrado por diversos estudos e a
susceptibilidade e o prognóstico variam de acordo a presença
do DM e outras comorbidades associadas, além de fatores
associados a idade, duração da doença, controle glicêmico,
presença de complicações e medicamentos em uso. O estudo
destas associações é um desafio que precisa ainda de melhor
investigação para que tenhamos uma prevenção eficaz por
meio da vacinação ou tratamento com agentes farmacológicos.
OBESIDADE
Um crescente corpo de dados sugere que os resultados
graves pela Doença do Coronavírus 2019 (COVID-19) são
piores naqueles que sofrem de obesidade. Isso ocorre devido a
uma proporção significativa daqueles que precisam de cuidados
intensivos sofrem de sobrepeso ou obesidade. A obesidade
está afetando a maioria dos processos fisiológicos e
modificando as funções do sistema imunológico (MUSCOGIURI
et al., 2020)
Uma explicação sugerida é que pacientes com
obesidade são conhecidos por terem um sistema imunológico
defeituoso que os torna vulneráveis a um tipo de infecção que
requer especificamente uma resposta imediata da imunidade
celular. A obesidade prejudica a imunidade ao alterar a resposta
das citocinas, resultando em uma diminuição na resposta das

SARS-COV-2: COMORBIDADES E COINFECÇÃO
364
células citotóxicas que têm um papel antiviral fundamental,
além de perturbar o equilíbrio dos hormônios endócrinos, como
a leptina, que afetam a interação entre as células metabólicas
e sistemas imunológicos (ROJAS-OSORNIO et al., 2019).
Esta comorbidade leva ao envolvimento de moléculas
específicas do tecido adiposo (adipocinas) na geração de um
ambiente favorável para doenças com causa imune. As células
dendríticas (CDs) tem o papel de ligação essencial entre a
imunidade inata e adaptativa, produziram duas vezes mais
citocina interleucina imunossupressora do que em controles e,
estimularam quatro vezes mais a produção de IL-4 a partir de
células T alogênicas. Existem também impactos negativos da
capacidade das CDs de amadurecer e induzir respostas de
células T adequadas a um estímulo geral como a infecção viral
(O'SHEA et al., 2013).
Um estudo de coorte com 3.615 indivíduos que
apresentaram resultado positivo para Covid-19, 775 (21%)
apresentaram índice de massa corporal (IMC) 30-34 e 595
(16% da coorte total) tinham IMC> 35. Pacientes com idade
inferior a 60 anos, com IMC de 30 a 34 apresentaram 2,0 vezes
mais chances de serem admitidos na emergência e 1,8 vezes
mais chances de admissão em terapia intensiva, em
comparação com indivíduos com IMC abaixo de 30. Para
pacientes na mesma faixa etária com IMC acima de 35, o risco
foi 2,2 e 3,6 vezes maior, respectivamente (LIGHTER et al.,
2020).
Embora pacientes com idade menor que 60 anos sejam
geralmente considerados um grupo de risco mais baixo para
COVID-19, com base nos dados do estudo, a obesidade parece
ser uma doença que aumenta o fator de risco para internação e
necessidade de cuidados intensivos (LIGHTER et al., 2020).

SARS-COV-2: COMORBIDADES E COINFECÇÃO
365
Existe a hipótese de que a mortalidade de indivíduos com
COVID-19 é alta entre pacientes com esta comorbidade em
específico, devido à grande massa de tecido adiposo com alta
expressão de receptores ACE2. A enzima conversora de
angiotensina humana 2 (ACE2) é o receptor putativo de SARS-
CoV-2 que possui alta afinidade com esse receptor e o utiliza
para a entrada nas células-alvo (KASSIR, 2020).
O nível de expressão de ACE2 no tecido adiposo é
relatado como maior do que no tecido pulmonar. Apesar da
expressão dos receptores ACE2 serem a mesma para o tecido
adiposo em pacientes obesos e não obesos, a diferença está
na massa do tecido adiposo que faz com que os pacientes com
obesidade expressem alto número de receptores ACE2. Isso
poderia explicar porque os pacientes com obesidade estão
apresentando a forma grave da COVID-19 (LI et al., 2020).
Outros fatores podem contribuem de forma significativa
para o aumento do risco de COVID‐19 em pacientes com
obesidade. O tecido adiposo pode servir como reservatório para
outros patógenos como o adenovírus humano (Ad-36), vírus da
influenza A, HIV, citomegalovírus, Trypanosoma gondii e
Mycobacterium tuberculosis anteriormente mencionado
BOURGEOIS et al., 2019).
Em resumo, a obesidade pode levar a condições graves
da COVID-19, como: i) A obesidade e a síndrome metabólica
podem causar danos aos órgãos causando falha de função; ii)
A obesidade foi acompanhada por expressão aumentada de
ACE2, onde a porção da proteína S do vírus se liga firmemente
e tornaria o tecido adiposo uma porta de entrada para a invasão
do vírus, tornado vulneráveis ao ataque do vírus tanto órgãos
como pulmão e coração; iii) A obesidade foi acompanhada por
um estado de inflamação superativada e resposta imune

SARS-COV-2: COMORBIDADES E COINFECÇÃO
366
desregulada, que pode induzir a resposta inflamatória
excessiva e exaustão de células imune em COVID-19; iv)
Pacientes obesos apresentam aumento da pressão abdominal,
expansão e movimento torácico limitados e função
compensatória respiratória insuficiente. No caso de infecção
pulmonar, é mais provável que evoluam para insuficiência
respiratória (CAUSSY et al., 2020; GHEBLAWI et al., 2020;
KASSIR, 2020).
Um ponto a ser observado é a cautela ao analisar
estudos nos casos de obesidade ao observar se foram incluídos
as comorbidades detalhadas de pacientes obesos, como
diabetes e hipertensão. Isso pode confundir o papel da
obesidade como um fator de risco independente em COVID-19.
Entretanto, mesmo estes trabalhos apresentando algum tipo de
viés, puderam concluir que a obesidade pode agravar os casos
de infecções por COVID-19 (HUSSAIN et al., 2020).
TUBERCULOSE
A tuberculose (TB) é uma doença infecciosa causada
pelo bacilo de Koch, Mycobacterium tuberculosis, de fácil
transmissão, afetando principalmente os pulmões. Quanto às
formas clínicas da doença, os pacientes com TB são
classificados como portadores de infecção latente por TB, que
é um estado assintomático e não transmissível; e a forma clínica
ativa da tuberculose, transmissível (na TB pulmonar ativa) onde
o diagnóstico molecular ou baseado em cultura celular pode ser
usado. Pacientes com doença ativa da TB apresentam
sintomas gerais, como febre, fadiga, falta de apetite e perda de
peso, e aqueles com doença pulmonar podem ter tosse
persistente e hemoptise (tosse com sangue) na sua forma

SARS-COV-2: COMORBIDADES E COINFECÇÃO
367
avançada. No entanto, alguns pacientes na sua forma ativa da
doença e de diagnóstico positivo podem ser assintomáticos e
são mais bem descritos como portadores de tuberculose
subclínica (PAI et al., 2016).
Pacientes acometidos por TB, devido ao
comprometimento pulmonar, fazem parte do grupo de risco
para o COVID -19 (LIU et al., 2020), e podem apresentar
sintomas comuns entre as duas doenças, como febre e tosse,
por esse motivo, precisam ser vistas com uma atenção especial
voltada ao diagnóstico diferencial e direcionamentos
adequados sem causar prejuízos ao paciente. Manter as ações
de vigilância e controle da TB durante a Pandemia faz parte da
estratégia de saúde orientada pelo Ministério da Saúde
(BRASIL, 2020).
Foi relatado ainda uma queda de mais de 50 % na
detecção de casos novos de tuberculose no mês de abril de
2020, seguindo decrescente até o último mês de avaliação. O
isolamento social pode, em determinadas situações, contribuir
com a transmissão da tuberculose em domicílios que têm a
presença de pessoas bacilíferas não diagnosticadas, ou ainda,
por aqueles que não usam os medicamentos de forma
adequada ou abandonaram o tratamento e continuam
transmitindo o bacilo de Koch, sobre tudo, em locais com difícil
acesso à água limpa e aglomerados urbanos, e se não forem
identificados oportunamente os desfechos podem ser
desfavoráveis devido a complicações severas, principalmente
se houver coinfecção por HIV/aids e/ou COVID-19 (BELL;
NOURSADEGHI, 2018).
Já foi demonstrado anteriormente coinfecção de SARS-
CoV-1 durante a epidemia em 2002 com tuberculose (TB), os
pacientes com TB hospitalizados e infectados com SARS-COV-

SARS-COV-2: COMORBIDADES E COINFECÇÃO
368
1 foram considerados com maior risco de morte. As respostas
imunes do hospedeiro para eliminar patógenos são estimuladas
na infecção por TB, e a infecção pelo SARS-CoV pode
aumentar a carga imunológica, levando a um sistema
imunológico em desequilíbrio ocasionando as tempestades de
citocinas, induzindo, em última instância, disfunção tanto
pulmonar como em outros órgãos. A infecção por M.
tuberculosis, em pacientes com a COVID-19, já foi relatado. Ao
se comparar os pacientes com TB e COVID-19 com pneumonia,
22% dos pacientes com TB avaliados apresentaram formas
clínicas leve/moderada da doença, sendo que os demais 78%
desenvolveram formas mais graves da COVID-19 (LIU et al.,
2020). Isso nos mostra a grande necessidade de avaliar a TB
como fator de risco para a COVID-19.
O fornecimento de tratamento adequado para pessoas
com diagnóstico de tuberculose também pode ser afetado,
especialmente para pessoas com tuberculose resistente a
medicamentos, devido a interrupções na produção e transporte
de medicamentos e suprimentos, suporte nutricional e de saúde
mental reduzido, acesso limitado a instalações de saúde e
redução do atendimento clínico para o controle de reações
adversas a medicamentos e comorbidades, como HIV, diabetes
e câncer. Além disso, o manejo prejudicado dessas
comorbidades também pode aumentar significativamente o
risco de progressão da fase latente para a forma ativada TB na
população em geral (SAUNDERS; EVANS, 2020).
CONCLUSÕES
A presença de comorbidades e mortalidade da doença
têm sido associados a doenças crônicas não transmissíveis,
como hipertensão, diabetes mellitus, os quais são causa de

SARS-COV-2: COMORBIDADES E COINFECÇÃO
369
uma elevada morbidade no Brasil, o que tem causado grande
preocupação. Muitos estudos verificaram que a presença
dessas morbidades crônicas e suas doenças associadas
(obesidade, hipertensão, doenças cardiovasculares e doença
renal crônica) são importantes fatores de risco para a gravidade
e prognóstico da doença.
Indivíduos com tuberculose são considerados grupo com
risco de morte devido a própria evolução clínica da doença e
por coinfecção com o SARS-CoV-2, devido ao histórico anterior
de coinfecção de SARS-CoV-1. É necessário a continuação de
políticas públicas de saúde assistencial com a realização da
testagem dos pacientes devido à semelhança já mencionada da
sintomatologia clínica por COVID, e a necessidade de
acompanhamento clínico durante o tratamento nos casos
positivos de TB.
REFERÊNCIAS BIBLIOGRÁFICAS ALFARAJ, Sarah H. et al. Middle East respiratory syndrome coronavirus and pulmonary tuberculosis coinfection: implications for infection control. Intervirology, v. 60, n. 1-2, p. 53-55, 2017. ATLAS, Diabetes. International diabetes federation. IDF Diabetes Atlas, 9th edn. Brussels, Belgium: International Diabetes Federation, 2019. BOURGEOIS, C. et al. Specific biological features of adipose tissue, and their impact on HIV persistence. Frontiers in microbiology. v. 10, 2019. BRASIL. Ministério da Saúde-MS. Vigilância de fatores de risco e proteção para doenças crônicas por inquérito telefônico. Brasília, DF; 2019. BRASIL. Secretaria de Vigilância em Saúde, Ministério da Saúde. Boletim Epidemiológico Especial - COE-COVID-19. Brasília: Ministério da Saúde, 2020. CAUSSY, Cyrielle et al. Obesity is associated with severe forms of COVID‐19. Obesity, 2020. CAREY, I.M. et al. Glycemic control and risk of infections among people with type 1 or type 2 diabetes in a large primary care cohort study. Diabetes Care, v. 41, n.10, p. 513–521, 2018. CHAN, C.-K. et al. Renin-angiotensin-aldosterone System Inhibitors and

SARS-COV-2: COMORBIDADES E COINFECÇÃO
370
Risks of SARS-CoV-2 Infection: A Systematic Review and Meta-analysis. Hypertension, set. 2020. FANG, L.; KARAKIULAKIS, G.; ROTH, M. Are patients with hypertension and diabetes mellitus at increased risk for COVID-19 infection? Lancet, v.8, n. 4, p. e21, 2020. FERLITA, S. et al. Type 2 diabetes mellitus and altered immune system leading to susceptibility to pathogens, especially mycobacterium tubercu- losis. Journal of Clinical Medicine. V. 8, n. 12, p.2219, 2019. GHEBLAWI, Mahmoud et al. Angiotensin-converting enzyme 2: SARS-CoV-2 receptor and regulator of the renin-angiotensin system: celebrating the 20th anniversary of the discovery of ACE2. Circulation research, v. 126, n. 10, p. 1456-1474, 2020. GARG, S. et al. Hospitalized rates and characteristics of patients with laboratory-confirmed Coronavirus Disease 2019 - COVID-NET, 14 States, March 1-30, 2020. MMWR. Morbidity and Mortality Weekly Report, v.69, n.15, p. 458-464, 2020. GRASSELLI, G. et al. Baseline characteristics and outcomes of 1591 patients infected with SARS-CoV-2 admitted to ICUs of the Lombardy Region, Italy. JAMA - Journal of the American Medical Association, v. 323, n.6, p. 1574-1581, 2020. GUAN, W. et al. Clinical features of coronavirus disease 2019 in China. New England Journal of Medicine, v. 382, n. 18, p. 1708-1720, 2020. HARAPAN, H. et al. Coronavirus disease 2019 (COVID-19): A literature review. Journal of Infection and Public Health, v. 13, n. 5, p. 667-673, 2020. HERRMANN, S. M.; TEXTOR, S. C. Renovascular Hypertension. Endocrinology and Metabolism Clinics of North America, v. 48, n. 4, p. 765-778, 2019. JI, H.L. et al. Elevated Plasmin(ogen) as a common risk factor for COVID-19 susceptibility. Physiological Reviews, v.100, n.3, p. 1065-1075, 2020. KARIO, K. et al. COVID‐19 and hypertension—evidence and practical management: Guidance from the HOPE Asia Network. The Journal of Clinical Hypertension, v. 22, n. 7, p. 1109–1119, 2020. KASSIR, Radwan. Risk of COVID‐19 for patients with obesity. Obesity Reviews, v. 21, n. 6, 2020. LI, M.-Y. et al. Expression of the SARS-CoV-2 cell receptor gene ACE2 in a wide variety of human tissues. Infectious diseases of poverty, v. 9, p. 1-7, 2020. LIGHTER, J. et al. Obesity in patients younger than 60 years is a risk factor for Covid-19 hospital admission. Clinical Infectious Diseases, v. 71, n. 15, p. 896-897, 2020. LIU, Yongyu et al. Active or latent tuberculosis increases susceptibility to COVID-19 and disease severity. Medrxiv, 2020.

SARS-COV-2: COMORBIDADES E COINFECÇÃO
371
MADDALONI, E.; BUZZETTI, R. Covid-19 and diabetes mellitus: unveiling the interaction of two pandemics. Diabetes/Metabolism Research and Reviews, 2020. MALACHIAS, M. V. B. et al. Concept, epidemiology and primary prevention. Arquivos Brasileiros de Cardiologia, v. 107, n. 3, p. 1-6, 2016. MULI, S. et al. Prevalence, awareness, treatment, and control of hypertension in older people: results from the population-based KORA-age 1 study. BMC public health, v. 20, n. 1, p. 1049, 2020. MUSCOGIURI, Giovanna et al. Comentary: Obesity: The “Achilles heel” for COVID-19?. Metabolism-Clinical and Experimental, v. 108, 2020. NYAMBUYA, T.M. et al. T-cell activation and cardiovascular risk in adults with type 2 diabetes mellitus: A systematic review and meta-analysis Clinical Immunology, v. 210, 2020. O'SHEA, Donal et al. Changes in human dendritic cell number and function in severe obesity may contribute to increased susceptibility to viral infection. International journal of obesity, v. 37, n. 11, p. 1510-1513, 2013. PAI, M., BEHR, M. A., DOWDY, D., DHEDA, K., Divangahi, M., Boehme, C. C., & Raviglione, M.. Tuberculosis. Nature Reviews Disease Primers, v. 2, n. 16076, 2016. RICHARDSON, S. et al. Presenting Characteristics, Comorbidities, and Outcomes among 5700 Patients Hospitalized with COVID-19 in the New York City Area. JAMA - Journal of the American Medical Association, v. 323, n. 20, p. 2052–2059, 2020. ROJAS-OSORNIO, S. A. et al. Immunity to influenza: Impact of obesity. Obesity Research & Clinical Practice, v. 13, n. 5, p. 419-429, 2019. SAUNDERS, M. J.; EVANS, C. A. COVID-19, tuberculosis, and poverty: preventing a perfect storm. European Respiratory Journal, v.56, n.1, 2020. SCHIFFRIN, E. L. et al. Hypertension and COVID-19. American Journal of Hypertension, v.33, n. 5, p. 373–374, 2020. SHIBATA, S. et al. Hypertension and related diseases in the era of COVID-19: a report from the Japanese Society of Hypertension Task Force on COVID-19. Hypertension Research, v. 43, p. 1028-1046, 2020. SCHMIDT, M.I.; et al. Chronic non-communicable diseases in Brazil: current burden and challenges. The Lancet, v. 377, n. 9781, p. 1949-1961, 2011. Sociedade Brasileira de Diabetes. Notas de esclarecimentos da Sociedade Brasileira de Diabetes sobre o coronavírus (COVID-19). São Paulo: SBD; 2020. Disponível em: <https://www.diabetes.org.br/publico/notas-de-esclarecimentos-da-sociedade-brasileira-de-diabetes-sobre-o-coronavirus-covid-19> WANG, Y. et al. Clinical course and outcomes of 344 intensive care patients with COVID-19. American Journal of Respiratory and Critical

SARS-COV-2: COMORBIDADES E COINFECÇÃO
372
Care Medicine, v. 201, n. 11, 2020. WHO, Word Health Organization. Hypertension. Disponível em: <https://www.who.int/news-room/fact-sheets/detail/hypertension>. Acesso em: 16 out. 2020. WHO. Word Health Organization. WHO Coronavirus Disease (COVID-19) Dashboard Disponível em: < https://covid19.who.int/>. Acesso em: 25. Out.2020. WU, Z,.; MCGOOGAN, J.M. Characteristics of and Important Lessons from the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72314 Cases from the Chinese Center for Disease Control and Prevention. JAMA - Journal of the American Medical Association, v. 323, n. 13, p. 1239-1242, 2020. HUSSAIN, Abdulzahra et al. Obesity and mortality of COVID-19. Meta-analysis. Obesity research & clinical practice, v. 14, n. 4, p. 295-300, 2020. YANG, J.K. et al. Blood glucose is a representative of the clusteredindicators of multi-organ injury for predicting mortality of COVID-19 in Wuhan, Chin. SSRN Electronic Journal, 2020. ZHANG, J.J. et al. Clinical characteristics of 140 patients infected with SARS-CoV-2 em Wuhan, China. Allergy: European Journal of Allergy and Clinical Immunology, v. 75, n. 7, p. 1730-1741, 2020.
AGRADECIMENTOS
Os autores agradecem aos órgãos de fomento: Fundação de apoio a Pesquisa na Paraíba - UFPB, à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior- CAPES e ao Conselho Nacional de Pesquisa - CNPq pelo suporte financeiro.

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
373
CAPÍTULO 18
TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO:
PROCESSOS FISIOPATOLÓGICOS NA COVID-19
Bárbara Guimarães CSORDAS1
Fernanda Silva ALMEIDA2
Edson dos Santos SILVA3
Tatjana de Souza Lima KEESEN4
Daniele Idalino JANEBRO5
1Pós Doutoranda do Programa de Pós-Graduação em Produtos Naturais e Sintéticos Bioativos - PgPNBS - UFPB; 2 Doutoranda da Rede Nordeste de Biotecnologia (RENORBIO)-
UFPB; 3 Mestrando do Programa de Pós Graduação em Ciências Fisiológicas da UFPB - UFPB ; 4Professora do Centro de Biotecnologia no Laboratório de Imunologia das doenças
Infecciosas - UFPB; 5Orientadora/Professora do Centro de Ciências da Saúde no Departamento de Ciências Farmacêuticas- UFPB.
RESUMO: A Doença por coronavírus 2019 (COVID-19) é uma doença infecciosa que desencadeou a pandemia no ano de 2020 e que causa a síndrome respiratória aguda grave (SRAG). Este trabalho teve como objetivo revisar os principais achados sobre tempestades de citocinas, cascata de coagulação e neuroinflamação, ressaltando os aspectos fisiopatológicos na doença COVID-19. Para isso, foi realizado um estudo de revisão bibliográfica de caráter descritivo com o total de 41 artigos examinados nas

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
374
plataformas digitais PubMed, Google Scholar, MedRxiv, BioRxiv. Um dos fatores de mortalidade tem sido associado à tempestade de citocinas induzida pelo vírus. A produção excessiva de citocinas pró-inflamatórias leva ao agravamento da SRAG e dano generalizado aos tecidos, resultando em falência múltipla de órgãos e morte. Processos de distúrbios de coagulação que geram problemas vasculares e cardíacos podem ser resultantes da infecção cardiotrópica que estão presentes na maioria dos pacientes que foram a óbito pela COVID-19. A capacidade que alguns vírus possuem de apresentar tropismo pelo córtex cerebral e ativar células imunes reativas nesse ambiente, promovem uma situação conhecida como neuroinflamação, acarretando muitas das vezes em uma disfunção do sistema nervoso. A necessidade de obter maior compreensão sobre tempestade de citocinas, doenças cardíacas e vasculares ou de neuroinflamação em pacientes com COVID-19 para um manejo clínico adequado, é um fator crítico no tratamento dos casos graves da doença. Palavras-chave: Síndrome da Liberação de Citocina. Cardiomiopatia dilatada. Neurofisiologia. Coronavírus da Síndrome Respiratória Aguda Grave 2.

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
375
1. INTRODUÇÃO
A doença infecciosa coronavírus 2019
(COVID-19), que está se espalhando em escala global
é causada por um vírus denominado de síndrome
respiratória aguda grave coronavírus 2 (SARS-CoV-2),
este vírus de RNA de fita simples positiva de tamanho
30Kb e envelopado, pertence à família Coronaviridae,
gênero beta-coronavírus. As proteínas não estruturais
deste vírus desempenham um papel crucial na
replicação do vírus, enquanto as proteínas estruturais
e auxiliares estão envolvidas na morfogênese e
interferem na resposta imunológica do hospedeiro
(CHEN et al., 2020; GIOVANETTI et al., 2020).
As reações imunológicas em COVID-19
grave, podem caracterizar a tempestade de citocinas
que está associada a consequências clínico-
patológicas desfavoráveis. A tempestade de citocinas
é uma liberação fora de controle de citocinas que tem
sido observada em algumas doenças infecciosas e
não infecciosas, levando a uma condição de
hiperinflamação no hospedeiro. Esta resposta
descontrolada de citocinas pode ser acompanhada por
mais ativação de células imunes, incluindo a
diferenciação de células T helper 17 (Th17) de
linfócitos CD4+, as respostas Th17 com nível

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
376
aumentado foram relatadas em MERS-CoV, SARS-
CoV e SARS-CoV-2 (XU et al., 2020; WU; YANG,
2020).
Outro fator relacionado as complicações no
agravamento da doença, podem levar à ativação da
coagulação sistêmica e complicações trombóticas.
Desde a propagação da pandemia de SARS‐CoV‐2,
aconteceram diversos relatos sobre uma alta
incidência de complicações trombóticas, incluindo
trombose de circuitos extracorpóreos para
hemofiltração venosa contínua, trombose associada à
cateter venoso central, trombose venosa profunda e
embolia pulmonar. A maioria dessas complicações,
mas não todas, ocorreram em pacientes internados na
unidade de terapia intensiva (UTI), com a maioria dos
pacientes recebendo profilaxia de trombose de rotina
(GUAN et al., 2019; MIDDELDORP et al., 2020;
WANG et al., 2020; ZHOU et al., 2020)
O papel fisiopatológico fundamental dessas
complicações no progresso clínico da COVID-19 é
ainda apoiado pela presença de trombos fibrinosos,
tumefação endotelial, megacariócitos em pequenas
artérias pulmonares e capilares pulmonares em
autópsias de pacientes que foram a óbito causados
pela COVID-19 grave (DOLHNIKOFF et al., 2020). A
miocardite é uma causa frequente de cardiomiopatia

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
377
dilatada com apresentações clínicas heterogêneas e
uma ampla gama de desfechos clínicos. Após a
infecção por um vírus cardiotrópico, ocorre uma
resposta pós-viral desadaptada, que pode causar
disfunção das células miocárdicas e comprometimento
da contratilidade (POLLACK et al., 2015).
O córtex cerebral é um ambiente estéril protegido por
diversas camadas celulares, dentre elas a barreira
hematoencefálica. Algumas populações celulares são
residentes característicos desse tecido, como a
micróglia e os astrócitos, ambos desempenhando
atividades protetivas e reparadoras no Sistema
Nervoso Central (SNC). Entretanto alguns agentes
infecciosos virais são capazes de invadir esse
ambiente utilizando diversas estratégias como porta
de entrada ao SNC. Outros coronavírus apresentaram
esse perfil neuro invasivo, produzindo um ambiente
infeccioso crônico e uma inflamação acima do nível
basal (MCGAVERN; KANG, 2011; KOYUNCU;
HOGUE; ENQUIST, 2013).
Nesse contexto, existe a necessidade de ampliar
o conhecimento dos fatores responsáveis pelo
desenvolvimento de complicações clínicas
significativas em um subgrupo de pacientes com
COVID-19, desvendando a alta gravidade da doença,
podendo levar à identificação de melhores terapias.

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
378
Isso melhoraria os resultados clínicos dos pacientes e
reduziria a carga atual sobre os sistemas de saúde,
enquanto se aguarda o desenvolvimento de vacinas
eficazes.
2. OBJETIVOS
Revisar os principais achados sobre
tempestades de citocinas, cascata de coagulação e
neuroinflamação, ressaltando os aspectos
fisiopatológicos na doença COVID-19.
3. MÉTODOS
Foi realizado um estudo de revisão bibliográfica
de caráter descritivo. O critério de exclusão utilizado
para esta revisão foram trabalhos que não
apresentassem o diagnóstico adequado para SARS-
CoV-2; idioma que não fosse inglês; ausência de
dados a serem extraídos, comentários ou cartas;
estudos caso controle; resultados redundantes e
casuísticas repetidas. Para isso, o total de 41 artigos
foram examinados nas plataformas digitais PubMed,
Google Scholar, MedRxiv, BioRxiv e publicações que
incluíram dados sobre a temática aqui abordada, tanto
para COVID‐19 ou doença similar SARS.

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
379
4. RESULTADOS E DISCUSSÃO
4.1. Tempestade de Citocinas
As citocinas desempenham um papel importante
na imunopatologia contra infecções. No entanto,
respostas imunológicas desreguladas e/ou excessivas
podem causar danos tanto em células teciduais como
as próprias células do sistema imune. Por serem parte
essencial do processo inflamatório são produzidas por
diversas células imunológicas, incluindo os
macrófagos, células dendríticas, células assassinas
naturais (Natural Killer-NK) e os linfócitos T e B
(RAGAB et al., 2020; YE; WANG; MAO, 2020).
Três das citocinas pró-inflamatórias mais
importantes da resposta imune inata são IL-1, TNF-α
e IL-6. Macrófagos teciduais, mastócitos, células
endoteliais e epiteliais são a principal fonte dessas
citocinas durante a resposta imune inata. A
“tempestade de citocinas” resulta então de um
aumento agudo súbito nos níveis circulantes de
diferentes citocinas pró-inflamatórias, incluindo IL-1,
IL-6, IL-18, IFN-γ e TNF-α. Este aumento nas citocinas
resulta no influxo de várias células imunes, como
macrófagos, neutrófilos e células T da circulação para
o local da infecção com efeitos destrutivos no tecido
humano resultante da desestabilização da célula

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
380
endotelial para as interações celulares, dano da
barreira vascular, capilar dano, dano alveolar difuso,
falha multiorgânica e, finalmente, morte (HUANG et al.,
2020; MAHMUDPOUR et al., 2020; YE; WANG; MAO,
2020; RAGAB et al.,2020)
A síndrome respiratória aguda grave (SRAG)
que leva a baixos níveis de saturação de oxigênio é
uma das principais causas de mortalidade na COVID-
19. Embora o mecanismo exato da SRAG em
pacientes com COVID-19 não seja totalmente
compreendido, a produção excessiva de citocinas pró-
inflamatórias é considerada um dos principais fatores
contribuintes (RAGAB et al., 2020).
Os estudos de que pacientes que agravaram da
COVID-19, sugerem que as respostas pró-
inflamatórias desempenham um papel na patogênese
desses vírus. Devido a regulação negativa na resposta
de células Th2, resposta imune Th1 (parcialmente)
inadequada e níveis elevados de expressão de
citocinas inflamatórias IL-1α, IL-1β, IL-8 e quimiocina
quimioatraente de neutrófilos (CXCL8) no trato
respiratório inferior de pacientes infectados por MERS-
CoV (ALOSAIMI et al., 2020).
Assim com SARS, MERS-CoV infecta células
epiteliais das vias aéreas, macrófagos derivados de
monócitos de sangue periférico humano e induz níveis

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
381
elevados de citocinas pró-inflamatórias e quimiocinas
e são induzidos a produzir altas concentrações de
IFNs (SCHEUPLEIN et al., 2015; YE; WANG; MAO,
2020).
A produção de IFN é a resposta imune natural
chave contra infecções virais, e IFN-I é a molécula
chave que desempenha um papel antiviral nas fases
iniciais da infecção viral. A liberação tardia de IFNs nos
estágios iniciais da infecção por coronavírus dificulta a
resposta antiviral do corpo. Posteriormente, o rápido
aumento de citocinas e quimiocinas atraem muitas
células inflamatórias, como neutrófilos e monócitos,
resultando em infiltração excessiva das células
inflamatórias no tecido pulmonar e, portanto, lesão
pulmonar. Respostas desreguladas e/ou exageradas
de citocinas e quimiocinas por células infectadas com
SARS-CoV ou MERS-CoV podem desempenhar um
papel importante na patogênese dessas viroses (YE;
WANG; MAO, 2020).
O agravamento da doença por fatores
associados a tempestade de citocinas pode levar ao
risco de vida e requer internação em Unidade de
Terapia Intensiva (UTI). Caracterizada por uma
apresentação clínica de inflamação sistêmica
predominante, hiperferritinemia, instabilidade
hemodinâmica e falência de múltiplos órgãos, deve ser

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
382
tratada imediatamente. O tratamento pode ser
realizado por terapias antivirais que podem ter como
alvo direto o vírus, as terapias anti-inflamatórias que
diminuem as respostas das citocinas são sugeridas
para diminuir a morbidade e a mortalidade em
pacientes com COVID-19 (RAGAB et al., 2020).
4.2 TRATAMENTO
Tocilizumabe (TCZ) é um anticorpo monoclonal
antagonista do receptor de IL-6 humano recombinante,
que impede a ligação da IL-6 ao seu receptor para
exercer a imunossupressão promovida pela IL-6.
Atualmente, o tocilizumabe é aplicado principalmente
em doenças autoimunes, como a artrite reumatoide e
tem um efeito terapêutico na tempestade de citocinas
induzida por infecção (SAHA et al., 2020; TANG et al.,
2020a; YE; WANG; MAO, 2020).
Relatos de caso mostraram que o TCZ é uma
abordagem de eficácia e segurança na COVID-19,
mesmo em pacientes com outras doenças, como
mieloma múltiplo, doença renal em estágio terminal ou
anemia falciforme (TANG et al., 2020a). Estudos
clínicos demonstraram que o tocilizumabe é eficaz no
tratamento de pacientes graves com lesões
pulmonares bilaterais extensas, que apresentam
níveis elevados de IL-6 (YE; WANG; MAO, 2020).

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
383
A terapia com glicocorticoides é amplamente
utilizada entre pacientes em estado grave com outras
infecções por coronavírus (por exemplo, SARS,
MERS). Os corticosteroides são anti-inflamatórios
esteroidais comumente usados para suprimir a
inflamação. Durante a epidemia de SARS de 2003, os
corticosteroides foram o principal meio de
imunomodulação e levaram a melhorias precoces,
como redução da febre, alívio da infiltração de
radiação no pulmão e melhora da oxigenação (TANG
et al., 2020a; YE; WANG; MAO, 2020).
No entanto, ainda é controverso se os
corticosteroides são benéficos no tratamento de
pacientes graves com COVID-19 (SOY et al., 2020;
TANG et al., 2020a). Deve-se notar que altas doses
podem atrasar a eliminação do coronavírus devido à
imunossupressão (YE; WANG; MAO, 2020).
No entanto, as intervenções relatadas são ainda
experimentais para o uso em média ou grande escala
da população em geral, sendo necessários mais
estudos em relação ao manejo da tempestade de
citocinas e suas complicações em pacientes graves,
este tipo de tratamento na COVID-19, com o uso de
anticorpos monoclonais é de alto custo, o que
inviabiliza muitas vezes esta forma de tratamento.

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
384
4.2 Cascata de coagulação: é possível que COVID-
19 seja principalmente uma doença cardíaca e
vascular.
A miocardite, uma doença inflamatória do
músculo cardíaco, é uma causa importante de
cardiomiopatia dilatada em todo o mundo. A infecção
viral também é uma causa importante de miocardite, e
o espectro de vírus conhecidos por causar miocardite
mudou nas últimas 2 décadas. Vários novos métodos
de diagnóstico, como a ressonância magnética
cardíaca, são úteis para o diagnóstico de miocardite
(ROSE, 2016).
A biópsia endomiocárdica pode ser usada para
pacientes com cardiomiopatia dilatada aguda
associada a comprometimento hemodinâmico,
aqueles com arritmia com risco de vida e aqueles cuja
condição não responde à terapia de suporte
convencional. Variáveis prognósticas importantes
incluem o grau de disfunção ventricular esquerdo e
direito, bloqueio cardíaco e formas histopatológicas
específicas de miocardite (ROSE, 2016).
Uma hipótese crescente de que SARS-CoV-2 seja
principalmente um vírus cardiovascular, e não
pulmonar está aumentando com diversos estudos. O
contexto em que as pessoas afetadas geralmente são
assintomáticas e têm doença muito leve ou nenhuma

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
385
doença podem ser considerado na verdade uma
doença resultante que leva a miocardite, mas que
pode levar muitos anos para se manifestar (POLLACK
et al., 2015; REYNOLDS et al., 2020; PÉREZ-
BERMEJO et al., 2020).
A relação de complicações cardiovasculares com o
vírus SARS-CoV-2 estão presentes desde o início da
pandemia. Na China, 20% dos pacientes
hospitalizados pela COVID-19 desenvolveram
doenças cardíacas e tromboembolismo ocorreram em
31% dos pacientes na UTI (SHI et al., 2020). Na
Alemanha, em um estudo ainda em andamento, foi
relatado inflamação do miocárdio em 60% de 100
pacientes recentemente recuperados da COVID-19.
No entanto, a doença cardiovascular foi considerada
por muitos como uma manifestação de COVID-19
grave e não por infecção direta do vírus, mas sim por
doença de pequenos vasos causada pela
hipercoagulação do sangue (PUNTMANN et al., 2020).
Cardiomiócitos expressam a enzima conversora de
angiotensina 2 (ECA2), necessária para a entrada viral
nas células. Cardiomiócitos cultivados em células,
desenvolveram alterações específicas que
correspondiam às alterações observadas em
autópsias de pacientes com COVID-19. Sugerindo que
o vírus também pode atacar e destruir o coração

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
386
diretamente, não apenas indiretamente com estresse
hemodinâmico, hipoxemia e trombose de pequenos
vasos (PÉREZ-BERMEJO et al., 2020).
Outro estudo realizado por Reynolds et al. (2020),
ofereceram uma explicação para o achado clínico de
que pacientes assintomáticos pela COVID-19 que não
apresentam sintomas pulmonares, mas ainda estão
com hipóxia, indicam que algo importante pode estar
ocorrendo. Foi observado que há uma desconexão
entre a troca gasosa e a mecânica pulmonar na
COVID-19. Ou seja, a oxigenação diminuída do
paciente não corresponde à capacidade do paciente
em respirar.
Em pacientes assintomáticos, foi traçado
semelhanças com o que é observado na síndrome
hepatopulmonar e, ao usar doppler com contraste, foi
observado a dilatação vascular pulmonar com
aumento do fluxo sanguíneo resultante, sugerindo que
a dilatação vascular pulmonar pode ser uma causa
significativa de hipoxemia em pacientes com
insuficiência respiratória causada pela COVID-19
(PÉREZ-BERMEJO et al., 2020).
Ou seja, pacientes considerados assintomáticos,
podem de fato estar realmente doentes. Uma porção
significativa deles pode desenvolver miocardite junto
com vasodilatação vascular com hipoxemia resultante.

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
387
Este processo patológico pode ser mais significativo
do que os efeitos pulmonares do vírus, uma vez que
está presente tanto em pacientes sintomáticos quanto
assintomáticos. Podendo explicar os sintomas
observados de fadiga e confusão mental que
determinados grupos de pacientes tiveram na doença
que inicialmente foi leve (PÉREZ-BERMEJO et al.,
2020).
4.2.1 Mecanismos patogênicos em SARS: O que
aprendemos?
Evidências de surtos epidêmicos anteriores
documentaram que em pacientes com SARS-CoV-1,
diversos genes com efeito pró-coagulante altamente
expresso nas células mononucleares infectadas como
receptor Toll-like 9 (TLR9). Expresso em plaquetas, o
TLR9 mediante ligação ao ligante, promove a ativação,
desgranulação e agregação plaquetária (GIANNIS;
ZIOGAS; GIANNI, 2020).
Estudos em modelo animal para SARS-CoV,
demonstraram que a expressão de genes pró-
coagulantes (trombina, VII, XI, XII), expressão de
ativadores do plasminogênio e desregulação da via da
uroquinase foram associadas à infecção fatal por
SARS-CoV. O SARS-CoV e o SARS-CoV-2
compartilham até 85% da identidade genômica e

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
388
ambos utilizam o mesmo receptor, a enzima ECA2,
para entrar nas células-alvo (GRALINSKI, BARIC,
2015; PERROTTA et al., 2020). Devido porcentagem
altamente homóloga, é esperado que ocorram
aspectos clínicos e imunopatológicos semelhantes.
Contudo, pode existir diferenças e é necessário
cautela ao extrapolar dados de SARS para COVID-19
Tanto nos casos de SARS como COVID-19, um
desarranjo do sistema renina angiotensina-
aldosterona é observado, que está intrinsecamente
ligado à cascata de coagulação e pode exacerbar o
processo de trombose imunológica, conduzindo ao
aumento da formação de microtrombos na COVID-19.
A angiotensina II (AngII) induz a expressão do inibidor
de ativação do plasminogênio tipo 1 (PAI-1) pelas
células endoteliais via receptor AT1, contribuindo para
um desequilíbrio PAI-1/tPA e um estado
hipercoagulativo (GOYAL et al., 2020).
E da mesma forma, é observado que a AngII estimula
a liberação de PAI-1 em adipócitos por meio dos
receptores AT1 e pode em parte, ser responsável pelo
aumento da gravidade observada nos casos com o
índice IMC alto. Além disso, a ECA metaboliza a
bradicinina, que finalmente estimula a vasodilatação e
liberação de PAI-1 das células endoteliais (GOYAL et
al., 2020). No entanto, mais estudos são necessários

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
389
para elucidar o papel da fisiopatologia da coagulação
podendo auxiliar no tratamento e melhora no
prognóstico da COVID-19.
4.2.2 Implicações para o manejo clínico e terapêutico
As melhores práticas para o tratamento de miocardites
atualmente precisam ser extrapoladas de experiências
anteriores não-COVID-19 e dos dados atuais de
COVID-19. Terapias experimentais que tentam limitar
a replicação do SARS-CoV-2 ou a resposta imune
estão em ensaios clínicos em andamento. Entretanto,
ainda não há terapias com eficácia clinicamente
comprovada para COVID-19 ou para tratamento de
miocardites causada por vírus especificamente
(HENDREN et al., 2020).
Estratégias terapêuticas como terapia
imunomoduladora, no caso de imunoadsorção de
anticorpos, tem sido utilizado. Alguns estudos já
relataram que a imunoadsorção em pacientes com
cardiomiopatia dilatada demonstrou melhorar a função
sistólica do ventrículo esquerdo e os níveis de
biomarcadores de insuficiência cardíaca, reduzir a
inflamação miocárdica e melhorar parâmetros
hemodinâmicos importantes, incluindo resistência
vascular sistêmica e índices de débito cardíaco
(POLLACK et al., 2015).

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
390
Já nos distúrbios de coagulação encontrados em
pacientes com COVID-19 um estudo retrospectivo de
Tang et al.(2020b), analisaram 449 pacientes
classificados com COVID-19 grave, comparando a
mortalidade em 28 dias entre usuários de heparina, 99
dos quais 94 receberam heparina de baixo peso
molecular, HBPM (40-60 mg enoxaparina/dia) e 5
receberam heparina não fracionada, HNF (10.000-
15.000 U/dia), por 7 dias ou mais e controles não
usuários. A mortalidade em 28 dias também foi
avaliada de acordo com o risco de coagulopatia.
Quando os pacientes foram estratificados de acordo
com os níveis de dímero-D, a mortalidade em usuários
de heparina basicamente permaneceu no mesmo
nível, mas em não usuários a mortalidade aumentou
proporcionalmente com os níveis de dímero-D.
Quando o dímero-D excedeu 3μg/ml (6 vezes do limite
superior normal), foi encontrada redução de
aproximadamente 20% na mortalidade com o
tratamento com heparina (32,8%; 52,4%, p = 0,017).
A heparina inibe a ativação e função dos neutrófilos
além de neutralizar a expressão de mediadores
inflamatórios que iniciam e conduzem a ativação do
sistema imune inato por meio da redução da
translocação do fator de transcrição nuclear kappa-
beta (NF-κß) do citoplasma para o núcleo; da mesma

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
391
forma, níveis reduzidos de TNF-α, IL-6, IL-8 e IL-1ß
são observados (POTERUCHA et al., 2017).
Uma vez que a ativação da coagulação também
contribui para a compartimentação de patógenos e
reduz sua invasão, o tratamento com anticoagulantes
em pacientes sem coagulopatia significativa tem um
risco potencial. Portanto, os anticoagulantes não
devem ser estendidos a todos os pacientes com
COVID-19, mas apenas àqueles que atendem aos
critérios dos perfis de risco trombótico e hemorrágico
e de dímero-D elevado.
4.3 Neuroinflamação nas doenças infecciosas
As pontes que formam o Sistema Nervoso e o Sistema Imune
são amplamente estudadas, tanto em condições de homeostase,
quanto em desordens homeostáticas, tais como em infecções virais
e doenças autoimunes. Neste contexto, o acionamento de uma
resposta protetiva com característica inflamatória tem a finalidade de
promover o papel defensivo contra diversos estímulos como
patógenos e toxinas (LI et al. 2019).
As respostas imunológicas periféricas frente a um desafio são
previamente não específicas, desenvolvida pelo reconhecimento por
células da imunidade inata, as quais podemos citar, macrófagos,
células NK, neutrófilos, mastócitos entre outras, estimulando uma
resposta de caráter inflamatório que em paralelo ativa células da
imunidade adaptativa possuidoras de especificidade contra o
estímulo, como os linfócitos T e B, a fim de promover a depuração do
evento patológico. A recíproca interação do sistema imune com o

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
392
microambiente neuronal estabelece uma relação harmônica
favorecendo a defesa imunológica (TOHIDPOUR et. al 2017).
No Sistema Nervoso Central (SNC) as células imunes
residentes predominantemente são as micróglias e os astrócitos,
ambas as populações contribuem sinergicamente para a homeostase
tecidual do SNC e funcionamento ideal do cérebro. A micróglia exerce
funções biológicas como imunovigilância neuronal, rápidas respostas
ao dano tecidual além do desenvolvimento de progenitores
neuronais. Os astrócitos são células imunocompetentes que
apresentam duas vias distintas de resposta a depender do ambiente
e contexto, a via caracterizada por um perfil regulador, e uma via
caracterizada por um perfil pró-inflamatório. É evidente o papel crucial
destas populações celulares no protagonismo da atividade protetiva
do CNS, entretanto a depender do microambiente neuronal sua
atividade pode ser agravada, tendenciado a uma resposta aberrante
(COLOMBO; FARINA, 2016; NORRIS; KIPNIS, 2019).
O Sistema Nervoso Central pode ser susceptível a entrada de
alguns agentes infecciosos de origem viral, denominados de vírus
neurotróficos, em adição, a ruptura da homeostase acarretada por um
processo infeccioso pode promover um ambiente capaz de induzir
uma resposta imune descontrolada. Os vírus podem utilizam de
algumas estratégias evasivas para adentrar ao tecido do SNC, uma
dessas estratégias é o rompimento da camada que protege o tecido
nervoso do SNC, a barreira hematoencefálica, uma estrutura formada
por multicamadas de células endoteliais justapostas e células
imunológicas como os astrócitos. A capacidade de alteração das
características das células endoteliais é efetuada através da
modulação infecciosa que promove o aumento de citocinas e
quimocinas, e de proteínas acessórias, como proteínas de junção e
de adesão, que de modo direto aumenta a permeabilidade da célula
(MCGAVERN; KANG, 2011; KOYUNCU; HOGUE; ENQUIST, 2013.)

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
393
Uma vez que o potencial vírus consegue invadir o córtex
cerebral, promove a liberação de padrões moleculares, como DNA ou
RNA viral, que são reconhecidos por receptores toll-like expresso nas
células micróglias. Nessa situação patológica aguda, a micróglia
reativa libera citocinas características pró-inflamatórias levando a
depuração do agente infeccioso viral, desempenhando um papel
substancial no processo de neuroinflamação. Em uma condição
infecciosa crônica, essa ativação prolongada e persistente decorrente
da infecção, ativa de modo contínuo a liberação de fatores pró-
inflamatórios, acarretando um processo inflamatório acima do basal
no SNC e disfunção neuronal (LI, et. al. 2019).
A utilização de rotas virais intracelulares também pode
acometer os neurônios dos sistemas sensitivos, de modo que o vírus
possa ser propagado no tecido nervoso. Os vírus utilizam as células
epiteliais do sistema olfatório periférico para chegar aos terminais
nervosos axonais dos neurônios do bulbo olfatório e assim invadir o
córtex cerebral. Como diversas células periféricas do sistema imune
podem migrar para o SNC, os vírus também usufruem dessas
habilidades celulares com a finalidade de ultrapassar a barreira
hematoencefálica e produzir efeitos no córtex cerebral (MCGAVERN;
KANG, 2011; MANGLANI; MCGAVERN, 2018).
O atual conhecimento sobre o potencial neurotrófico do
SARS-CoV-2 ainda permanece insuficiente, entretanto, em um
estudo MAO et al. (2020), identificaram sintomas como dor de
cabeça, anosmia, ageusia, consciência prejudicada, convulsão e
encefalopatia em 36,4% dos pacientes infectados pelos novo
coronavírus emergente. Os significativos danos cerebrais associados
a alterações no paladar e olfato na COVID-19 pode sugerir um
potencial neuro mecanismo de invasão, baseados em estudos
anteriores com MERS-CoV e SARS-CoV e outros coronavírus, foi
relatado a presença de partículas virais em neurônios cerebrais (LI et
al., 2016).

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
394
A via neuroimune tem funcionalidade bidirecional,
respondendo tanto aos estímulos decorrentes da região periférica,
quanto estimulando esta área por meio de moléculas liberadas. Uma
vez que esses mediadores inflamatórios são produzidos e
reconhecidos, as células podem adotar uma característica hiper-
responsiva com efeitos inflamatórios sistêmicos (DOSSANTOS et. al.
2020).
5. CONCLUSÕES
Em resumo, a infecção pelo novo tipo de coronavírus
causa uma tempestade inflamatória de citocinas nos
pacientes. A tempestade de citocinas leva à SRAG ou
insuficiência extrapulmonar de múltiplos órgãos e é um
fator importante que causa a exacerbação da COVID-
19 ou mesmo a morte. O gatilho para a tempestade de
citocinas é uma resposta imune descontrolada que
resulta em ativação e expansão contínuas de células
imunes, linfócitos e macrófagos, que produzem
quantidades imensas de citocinas. Os achados
clínicos da tempestade de citocinas são atribuídos à
ação de citocinas pró-inflamatórias como IL-1, IL-6, IL-
18, IFN-γ e TNF-α.
Os estudos que ressaltam a extrema importância
de testagens preventivas da infecção e identificação
de portadores assintomáticos da COVID-19 devem
continuar incluindo a população jovem. As

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
395
preocupações em relação às consequências desse
vírus a longo prazo são crescentes, como no caso de
complicações vasculares e miocardites. O mecanismo
fisiopatológico exato do tromboembolismo induzido
pela SARS-CoV-2 precisa ser mais estudado para
fornecer bases biológicas para melhorar a terapia
clínica com anticoagulantes.
Atualmente o conhecimento acerca do perfil
neuroinvasivo do SARS-CoV-2 permanece incerto,
haja vista as lacunas existentes por se tratar de uma
infecção viral emergente. Em comparação com outros
coronavírus, o SARS-CoV-2 pode ter capacidade
neurotrófica e neuro inflamatória por desencadear
imunidade reativa através da micróglia e astrócitos no
córtex cerebral e estimular a produção de fatores com
características pró-inflamatória. Contudo, são
necessários mais estudos para a real compreensão
desse processo.

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
396
REFERÊNCIAS BIBLIOGRÁFICAS ALOSAIMI, B. et al. MERS-CoV infection is associated with downregulation of genes encoding Th1 and Th2 cytokines/chemokines and elevated inflammatory innate immune response in the lower respiratory tract. Cytokine, v. 126, p. 154895, 2020. CHEN, Y.; LIU, Q.; GUO, D. Emerging coronaviruses: genome structure, replication, and pathogenesis. Journal of medical virology, v. 92, n. 4, p. 418-423, 2020. COLOMBO, E.; FARINA, C. Astrocytes: Key Regulators of Neuroinflammation. Trends Immunol. v. 37, n. 9, p. 1-13. 2016. DOLHNIKOFF, M. et al. Pathological evidence of pulmonary thrombotic phenomena in severe COVID‐19. Journal of Thrombosis and Haemostasis, 2020. DOSSANTOS, M. F. et al. Neuromechanisms of SARS-CoV-2: a review. Frontiers in neuroanatomy, v. 14, p. 37, 2020. GIANNIS, D.; ZIOGAS, I. A.; GIANNI, P. Coagulation disorders in coronavirus infected patients: COVID-19, SARS-CoV-1, MERS-CoV and lessons from the past. Journal of Clinical Virology, p. 104362, 2020. GIOVANETTI, M. et al. A doubt of multiple introductions of SARS‐CoV‐2 in Italy: a preliminary overview. Journal of Medical Virology, 2020. GOYAL, P. et al. Clinical characteristics of Covid-19 in New York city. New England Journal of Medicine, 2020. GRALINSKI, L. E.; BARIC, R. S. Molecular pathology of emerging coronavirus infections. The Journal of pathology, v. 235, n. 2, p. 185-195, 2015. GUAN, W. et al. Clinical characteristics of coronavirus disease 2019 in China. New England journal of medicine, v. 382, n. 18, p. 1708-1720, 2020. HENDREN, N. S. et al. Description and proposed management of the acute COVID-19 cardiovascular syndrome. Circulation, 2020. HUANG, C. et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. The lancet, v. 395, n. 10223, p. 497-506, 2020. KOYUNCU, O. O; HOGUE, I. B; ENQUIST, L. W. Virus infections in the nervous system. Cell Host Microbe. v. 13, n. 4, p. 379-393. 2013. Li, K. et al. Jr. Middle East Respiratory Syndrome Coronavirus Causes Multiple Organ Damage and Lethal Disease in Mice Transgenic for Human Dipeptidyl Peptidase 4. The Journal of infectious diseases, v. 1;213, n. 5, p. 12-22. 2016.

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
397
Li, L. et al. Viral infection and neurological disorders-potential role of extracellular nucleotides in neuroinflammation. ExRNA. v.1, n.26, p. 1-5. 2019. MAHMUDPOUR, M. et al. COVID-19 cytokine storm: The anger of inflammation. Cytokine, p. 155151, 2020. MANGLANI, M; MCGAVERN, D. B. New advances in CNS immunity against viral infection. Current opinion in virology. n. 28, p. 116–126. 2018. MAO, L. et al. Neurologic Manifestations of Hospitalized Patients with Coronavirus Disease 2019 in Wuhan, China. JAMA Neurology, v. 1;77, n. 6 p. 683-690. 2020. MCGAVERN, D.; KANG, S. Illuminating viral infections in the nervous system. Nat Rev Immunol. v. 11, p. 318–329. 2011. MIDDELDORP, S. et al. Incidence of venous thromboembolism in hospitalized patients with COVID‐19. Journal of Thrombosis and Haemostasis, 2020. NORRIS, G. T.; KIPNIS; J. Immune cells and CNS physiology: Microglia and beyond. J Exp Med. v. 216, n. 1, p. 60-70. 2019. PÉREZ-BERMEJO, J. A. et al. SARS-CoV-2 infection of human iPSC-derived cardiac cells predicts novel cytopathic features in hearts of COVID-19 patients. BioRxiv, 2020. PERROTTA, F. et al. Severe respiratory SARS-CoV2 infection: Does ACE2 receptor matter? Respiratory Medicine, p. 105996, 2020. POLLACK, A. et al. Viral myocarditis-diagnosis, treatment options, and current controversies. Nature Reviews Cardiology, v. 12, n. 11, p. 670, 2015. POTERUCHA, Timothy J.; LIBBY, Peter; GOLDHABER, Samuel Z. More than an anticoagulant: Do heparins have direct anti-inflammatory effects? Thrombosis and haemostasis, v. 117, n. 03, p. 437-444, 2017. PUNTMANN, V. O. et al. Outcomes of cardiovascular magnetic resonance imaging in patients recently recovered from coronavirus disease 2019 (COVID-19). JAMA cardiology, 2020. RAGAB, Dina et al. The COVID-19 cytokine storm; what we know so far. Frontiers in immunology, v. 11, p. 1446, 2020 REYNOLDS, A. S. et al. Pulmonary vascular dilatation detected by automated transcranial doppler in COVID-19 pneumonia. American journal of respiratory and critical care medicine, n. ja, 2020. ROSE, N. R. Viral myocarditis. Current opinion in rheumatology, v. 28, n. 4, p. 383, 2016. SAHA, Abinit et al. Tocilizumab: A therapeutic option for the treatment of cytokine storm syndrome in COVID-19. Archives of Medical Research, 2020.

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
398
SCHEUPLEIN, V. A. et al. High secretion of interferons by human plasmacytoid dendritic cells upon recognition of Middle East respiratory syndrome coronavirus. Journal of virology, v. 89, n. 7, p. 3859-3869, 2015. SHI, S. et al. Association of cardiac injury with mortality in hospitalized patients with COVID-19 in Wuhan, China. JAMA cardiology, 2020. SOY, M. et al. Cytokine storm in COVID-19: pathogenesis and overview of anti-inflammatory agents used in treatment. Clinical Rheumatology, p. 1, 2020. TANG, N. et al. Anticoagulant treatment is associated with decreased mortality in severe coronavirus disease 2019 patients with coagulopathy. Journal of thrombosis and haemostasis, v. 18, n. 5, p. 1094-1099, 2020a. TANG, Yujun et al. Cytokine storm in COVID-19: the current evidence and treatment strategies. Frontiers in immunology, v. 11, p. 1708, 2020b. TOHIDPOUR, A. et al. Neuroinflammation and infection: molecular mechanisms associated with dysfunction of neurovascular unit. Frontiers in cellular and infection microbiology, v. 7, p. 276, 2017. WANG, D. et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus–infected pneumonia in Wuhan, China. Jama, v. 323, n. 11, p. 1061-1069, 2020. WU, D.; YANG, X. O. TH17 responses in cytokine storm of COVID-19: An emerging target of JAK2 inhibitor Fedratinib. Journal of Microbiology, Immunology and Infection, 2020. XU, Z. et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. The Lancet respiratory medicine, v. 8, n. 4, p. 420-422, 2020. YE, Q.; WANG, B.; MAO, J. The pathogenesis and treatment of theCytokine Storm'in COVID-19. Journal of infection, v. 80, n. 6, p. 607-613, 2020. ZHOU, F. et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. The lancet, 2020.
AGRADECIMENTOS
Os autores agradecem aos órgãos de fomento: Fundação de apoio
a Pesquisa da Paraíba-Fapesq/PB, à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior- CAPES pelo suporte financeiro e a Universidade Federal da Paraíba pela infraestrutura necessária para a realização deste trabalho.

TEMPESTADE DE CITOCINAS, CASCATA DE COAGULAÇÃO E NEUROINFLAMAÇÃO: PROCESSOS FISIOPATOLÓGICOS NA COVID-19
399
CIÊNCIAS BIOLÓGICAS - MICROBIOLOGIA

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
400
CAPÍTULO 19
ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA
(PSEUDOBOMBAX MARGINATUM)
Maria Isabela Ferreira de Araújo 1
Izaqueu Rodrigues da Silva 2
Raquel Bezerra da Silva 3
Amanda Regis de Sena 4
Tonny Cley Campos leite 5 1 Doutoranda em Ciências Biológicas, UFPE; 2 Graduando em Licencitura em Química/ IFPE;
3 Graduanda em Licenciatura em Ciências Biológicas, UFPB; 4 Professora do Instituto Federal de Educação, Ciências e Tecnologia de Pernambuco (IFPE); Professor/Orientador do Instituto
Federal de Educação, Ciências e Tecnologia de Pernambuco (IFPE) 4. [email protected]
RESUMO: A multirresistência desencadeada pelos microrganismos possibilitou o surgimento de novas perspectivas no campo de pesquisa de produtos naturais, entre essas está a Pseudobombax marginatum, cuja prospecção fitoquímica e atividade microbiana foram avaliadas nesse estudo. Foi determinado o perfil fitoquímico de extrato hexânico (Hex), acetato de etila (AcOEt), metanólico (MeOH) e aquoso das folhas de Embiratanha por cromatografia em camada delgada (CCD), e análises espectrofotometrica. A concentração inibitória mínima (MIC) foi avaliada por microdiluição. A concentração microbicina mínima (CMM) dos extratos foi determinada pela técnica de esgotamento a partir de alíquotas dos extratos que apresentaram MIC. A análise do perfil fitoquímico identificou esteroides, flavonoides, taninos e triterpenos nos extratos hexânico (Hex), acetato de etila (AcOEt), metanólico (MeOH) e aquoso, com exceção verificada com o triterpeno na triagem fitoquimica, na qual não foi detectado nos extratos mencionados. O MIC e MBC para

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
401
Micrococcus luteus (ATCC 06), Enterococcus faecalis (ATCC 138) submetido ao extrato de Hex foram de 1 mg/mL, 0.5 mg/mL, respectivamente, as cepas de Staphylococcus aureus padrão (ATCC 01) e clínica (NCCLS 700) a CIM foi 1 mg/mL e a CBM 2 mg/mL, M. luteus (ATCC 06) e E. faecalis (ATCC138) submetidos ao extrato adquirido com o solvente AcoEt foram sensíveis na concentração 1 mg/mL. Portanto, a Embiratanha é composta por classes diversas de metabólitos secundários, além apresentar-se bactericida para as cepas utilizadas neste estudo. Palavras-chave: Pseudobombax marginatum. Métabolitos secundários. Fitoquímica. Antibacterianos.
INTRODUÇÃO
A introdução de diversas classes de antibióticos durante
processos infecciosos contribui para o surgimento de
microrganismos com elevados parâmetros de seletividade
(WESTHOFF et al., 2017). A resistência a múltiplas drogas
(MDR) está relacionada a fatores como alterações na absorção,
eliminação e inativação enzimática dos fármacos (VELÁSQUEZ
et al, 2020).
Tais parâmetros reforçam a necessidade de estudos
voltados a esses patógenos e introduzem a ascensão de novas
perspectivas farmacológicas, dentre as quais o uso de plantas
medicinais, capazes de promover reorganização molecular e
ação contra microrganismos (KUPPUSAMY et al., 2016).
Nesse âmbito, se encontra os vegetais inseridos na
família Malvaceae, subdividida em 30 gêneros e 290 espécies
distribuídas pelo mundo. Dentre as quais destaca-se a
Pseudobombax marginatum conhecida popularmente como
Embiratanha (DIÓRGENES, et al. 2019), Paineira-imbiriçu,

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
402
Sumaúma, Imbiruçu (PAIVA, 2013). Explorada para fins
farmacológicos no tratamento de problemas intestinais,
infecciosos, inflamatórios e alergias (ALMEIDA; OLIVEIRA;
FALCÃO, 2015).
Os metabólitos secundários, como os flavonoídes,
alcaloides, terpenos dentre outros, produzidos pela
Embiratanha impulsionam benefícios fisiológicos, efeito
antioxidante, manutenção celular, atividade anti-inflamatória,
além de auxiliar na prevenção do câncer (TIWARI; RANA,
2015), quando introduzidos no organismo (MENEZES et al.,
2015). Tais compostos bioativos interagem sinergicamente,
apresentando potencial anticâncer e antioxidante, além de,
permitir a inibição de cepas bacterianas de interesse clínico
(VASUDEVAN; ARULMOORTHY; SURESH, 2020).
Neste contexto, sabendo-se da importância da
descoberta de produtos naturais com efeitos terapêuticos, o
objetivo do trabalho é determinar o perfil fotoquímico das
folhas de P.marginatum, bem como verificar a suscetibilidade
de diferentes cepas bacterianas frente a extratos obtidos da
referida matéria prima.
MATERIAIS E MÉTODO
MATERIAL VEGETAL
As folhas de Pseudobombax marginatum foram
coletadas na Zona Rural do município de São José do Egito –
Pernambuco, Brasil, entre os meses de abril e maio de 2018.
Coletou-se 2 Kg do material fresco, em seguida, essa amostra
foi seca em estufa de 40 °C durante quadro dias e moída em
moinho elétrico do tipo Willey, obtendo-se uma farinha fina que

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
403
foi estocada em recipientes hermeticamente fechados
vedados e utilizada para análises posteriores.
PREPARAÇÃO DOS EXTRATOS
Para extração dos metabólitos secundários presentes
nas folhas de Embiratanha, foram utilizadas diferentes
soluções extratoras: hexano (Hex), acetato de etila (AcOEt) e
metanol (MeOH). Produziram os extratos: hexânico (Hex, 2g),
acetato de etila (AcOEt, 3g) e metanólico (MeOH, 10g).
PERFIL FITOQUÍMICO
O perfil fotoquímico foi realizado segundo Wagner e
Bladt (2001). Os extratos foram aplicados em da placa de sílica
e, posteriormente colocados em cuba cromatográfica com os
eluentes. Posteriormente as placas foram aspergidas com os
reveladores específicos para cada classe de composto (Tabela
1). Os aspectos visíveis foram observados, usando os
comprimentos de onda de 254 nm e 365 nm. A mudança de cor
visível e da radiação ultravioleta (UV) é indicativo da
positividade da reação.
Tabela 1. Classe de compostos químicos e reveladores
CLASSES REVELADOR COLORAÇÃO UV/VIS
Alcaloides Dragendorff Laranja VIS
Mayer Branco VIS
Wagner Marrom VIS
Antraquinonas KOH 5% Laranja-avermelhadas UV/VIS
Cumarinas KOH 5% Verde-azuladas UV/VIS
Flavonoides AlCl3 Amarelo-esverdeado UV
Anisaldeído-
sulfúrico
Amarelo VIS
Sulfato cérico Amarelo VIS

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
404
AlCl3: cloreto de alumínio, FeCl3: cloreto férrico, KOH: hidróxido de
potássio.
TRIAGEM FITOQUÍMICA
Seguindo a metodologia de Silva et al. (2019), o extrato foi
submetido à análise fotoquímica qualitativa para detecção das
principais classes de metabólitos secundários.
ATIVIDADE ANTICROBIANA
MICRORGANISMOS
As linhagens de bactérias e fungos leveduriformes
utilizados foi obtida a partir da coleção de Microrganismos do
Departamento de Antibióticos da Universidade Federal de
Pernambuco (UFPEDA). Para este ensaio foram utilizadas
quatro bactérias gram-positivas: Staphylococcus aureus
(UFPEDA 01), Micrococcus luteus (UFPEDA 06), Bacillus
subtilis (UFPEDA 16), e Enterococcus faecalis (UFPEDA 138);
três bactérias gram-negativas: Escherichia coli (UFPEDA 224),
uma álcool-ácido-resistente: Mycobcterium smegmatis
(UFPEDA 71) e uma cepa clínica Staphylococcus aureus
(NCCLS 700).
Óleos
essenciais
Anisaldeído
sulfúrico
Vermelho-
amarronzadas
VIS
Terpenoides Liebermann-
Burchad
Rosa/roxo: triterpenos,
Azul/verde: esteroides
VIS
Vanilina-sulfúrica Roxo: terpenos VIS
Taninos FeCl3 Preto VIS

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
405
INÓCULO
As suspensões foram preparadas a partir de culturas
bacterianas recentes semeadas em Ágar Nutriente (NA) e
incubadas a 35±2°C durante 24-48h. Após a incubação, foi
transferida cerca de 4-5 colônias de bactérias (com uma alça
microbiológica estéril) para tubos de ensaios contendo 5.0 mL
de solução salina estéril (NaCl 0.85%). As suspensões
resultantes foram agitadas durante 15 segundos com o auxílio
de um aparelho Vortex (Fanem Ltd., Guarulhos, SP, Brasil). A
turbidez do inoculo final foi normalizada utilizando uma
suspensão de sulfato de bário (tubo 0.5 na escala de
McFarland). A concentração final obtida foi de 1-5 × 108
unidades formadoras de colônias por mililitros (UFC/mL)
(ARAUJO et al., 2017).
CONCENTRAÇÃO INIBITÓRIA MÍNIMA (CIM) E
CONCENTRAÇÃO BACTERICIDA MÍNIMA (CBM)
A determinação da Concentração Inibitória Mínima
(CIM) e Concentração Bactericida Mínima (CBM) seguiram a
metodologia de (INGROFF et al., 2002). Estas medidas foram
determinadas utilizando placas com 96 poços estéreis,
próprias para microdiluição.
Inicialmente adicionou-se em cada poço 90 µL do meio
Caldo Muller Hinton (CMH) e posteriormente a partir da terceira
coluna (A3) 90 µL do extrato na concentração de 16 mg/mL
esta alíquota foi homogeneizada e transferida para a quarta
coluna (A4) e assim por diante até a décima segunda coluna
(A12) que recebe o extrato na concentração de 0,03 mg/mL,
neste último a alíquota (90 µL) depois de homogeneizada é

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
406
descartada. Por último foi adicionada uma alíquota de 10 µL
da suspensão do microrganismo.
As placas com o meio, extrato e microrganismo foram
incubadas por 24 horas (37º C) para bactérias e 48 horas (28º
C) para leveduras. Posteriormente foi adicionada 30 µL de
Rezasurina para análise quantitativa do crescimento
microbiano nos poços de ensaio e determinação da atividade
antimicrobiana relativa de cada diluição das amostras. Para a
determinação da Concentração Mínima Bactericida (CBM) foi
repicada uma alíquota de 5 µL, das concentrações que
apresentaram atividade na placa do CIM, em placas de Petri
contendo Ágar Mueller Hinton (AMH). Ao mesmo tempo, os
controles foram feitos para a viabilidade bacteriana e para a
suscetibilidade ao antibiótico padrão cloranfenicol (100UI /
mL). As placas foram incubadas a 35 ± 2° C durante 24 horas.
Após o tempo de incubação apropriado, a presença (ou
ausência) de crescimento foi observada visualmente, por meio
da formação de aglomerados celulares ou “botões” nos poços,
sugestivo de positividade. A CBM foi considerada a menor
concentração do extrato onde não houve crescimento celular
sobre a superfície do AMH.
RESULTADOS
ANÁLISE FITOQUÍMICA
PERFIL FOTOQUÍMICO
O perfil e triagem fitoquímica das folhas do vegetal foram
realizados, permitindo a detecção dos principais metabolitos
secundários (Tabela 2 e 3).
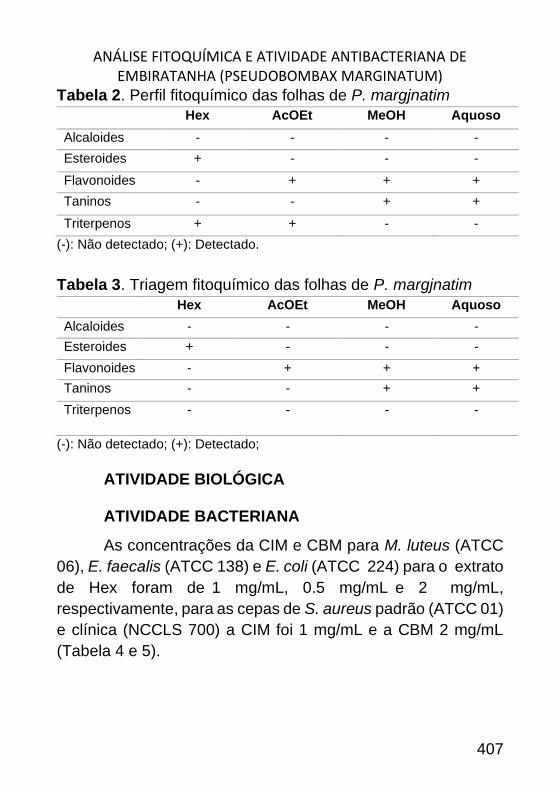
ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
407
Tabela 2. Perfil fitoquímico das folhas de P. margjnatim
Hex AcOEt MeOH Aquoso
Alcaloides - - - -
Esteroides + - - -
Flavonoides - + + +
Taninos - - + +
Triterpenos + + - -
(-): Não detectado; (+): Detectado.
Tabela 3. Triagem fitoquímico das folhas de P. margjnatim
Hex AcOEt MeOH Aquoso
Alcaloides - - - -
Esteroides + - - -
Flavonoides - + + +
Taninos - - + +
Triterpenos - - - -
(-): Não detectado; (+): Detectado;
ATIVIDADE BIOLÓGICA
ATIVIDADE BACTERIANA
As concentrações da CIM e CBM para M. luteus (ATCC
06), E. faecalis (ATCC 138) e E. coli (ATCC 224) para o extrato
de Hex foram de 1 mg/mL, 0.5 mg/mL e 2 mg/mL,
respectivamente, para as cepas de S. aureus padrão (ATCC 01)
e clínica (NCCLS 700) a CIM foi 1 mg/mL e a CBM 2 mg/mL
(Tabela 4 e 5).
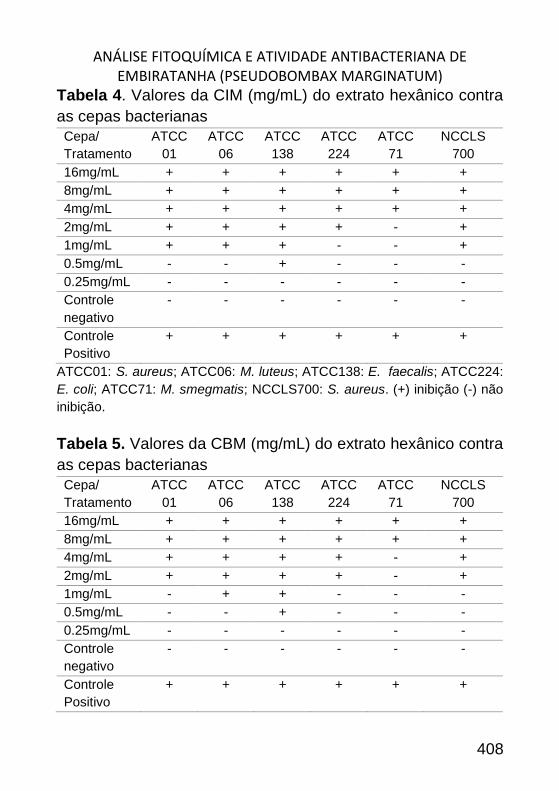
ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
408
Tabela 4. Valores da CIM (mg/mL) do extrato hexânico contra
as cepas bacterianas
Cepa/
Tratamento
ATCC
01
ATCC
06
ATCC
138
ATCC
224
ATCC
71
NCCLS
700
16mg/mL + + + + + +
8mg/mL + + + + + +
4mg/mL + + + + + +
2mg/mL + + + + - +
1mg/mL + + + - - +
0.5mg/mL - - + - - -
0.25mg/mL - - - - - -
Controle
negativo
- - - - - -
Controle
Positivo
+ + + + + +
ATCC01: S. aureus; ATCC06: M. luteus; ATCC138: E. faecalis; ATCC224: E. coli; ATCC71: M. smegmatis; NCCLS700: S. aureus. (+) inibição (-) não
inibição.
Tabela 5. Valores da CBM (mg/mL) do extrato hexânico contra
as cepas bacterianas
Cepa/
Tratamento
ATCC
01
ATCC
06
ATCC
138
ATCC
224
ATCC
71
NCCLS
700
16mg/mL + + + + + +
8mg/mL + + + + + +
4mg/mL + + + + - +
2mg/mL + + + + - +
1mg/mL - + + - - -
0.5mg/mL - - + - - -
0.25mg/mL - - - - - -
Controle
negativo
- - - - - -
Controle
Positivo
+ + + + + +
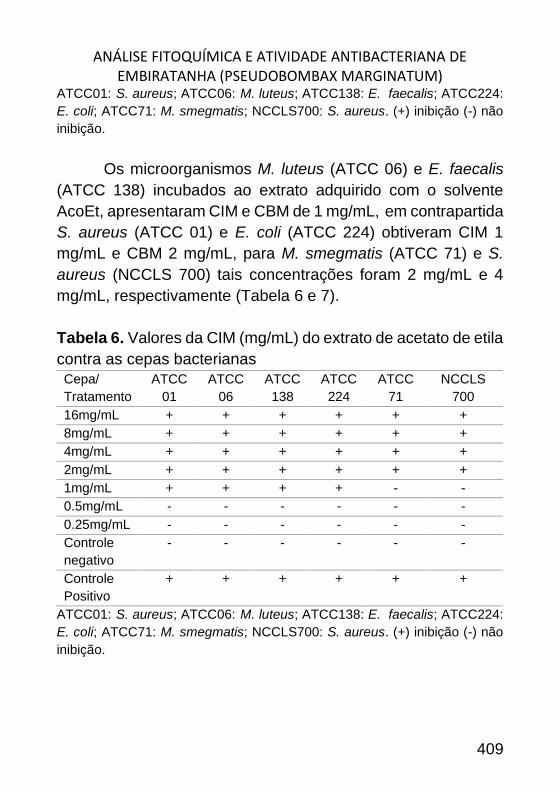
ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
409
ATCC01: S. aureus; ATCC06: M. luteus; ATCC138: E. faecalis; ATCC224: E. coli; ATCC71: M. smegmatis; NCCLS700: S. aureus. (+) inibição (-) não
inibição.
Os microorganismos M. luteus (ATCC 06) e E. faecalis
(ATCC 138) incubados ao extrato adquirido com o solvente
AcoEt, apresentaram CIM e CBM de 1 mg/mL, em contrapartida
S. aureus (ATCC 01) e E. coli (ATCC 224) obtiveram CIM 1
mg/mL e CBM 2 mg/mL, para M. smegmatis (ATCC 71) e S.
aureus (NCCLS 700) tais concentrações foram 2 mg/mL e 4
mg/mL, respectivamente (Tabela 6 e 7).
Tabela 6. Valores da CIM (mg/mL) do extrato de acetato de etila
contra as cepas bacterianas
Cepa/
Tratamento
ATCC
01
ATCC
06
ATCC
138
ATCC
224
ATCC
71
NCCLS
700
16mg/mL + + + + + +
8mg/mL + + + + + +
4mg/mL + + + + + +
2mg/mL + + + + + +
1mg/mL + + + + - -
0.5mg/mL - - - - - -
0.25mg/mL - - - - - -
Controle
negativo
- - - - - -
Controle
Positivo
+ + + + + +
ATCC01: S. aureus; ATCC06: M. luteus; ATCC138: E. faecalis; ATCC224: E. coli; ATCC71: M. smegmatis; NCCLS700: S. aureus. (+) inibição (-) não
inibição.
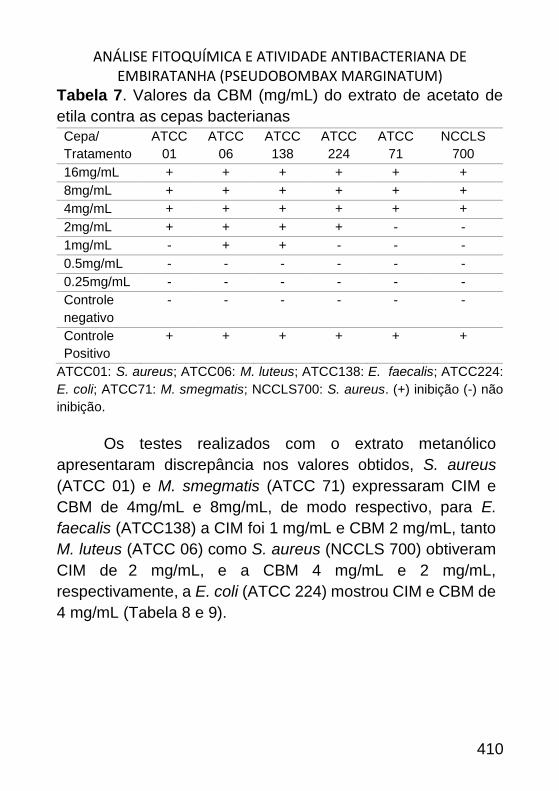
ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
410
Tabela 7. Valores da CBM (mg/mL) do extrato de acetato de
etila contra as cepas bacterianas
Cepa/
Tratamento
ATCC
01
ATCC
06
ATCC
138
ATCC
224
ATCC
71
NCCLS
700
16mg/mL + + + + + +
8mg/mL + + + + + +
4mg/mL + + + + + +
2mg/mL + + + + - -
1mg/mL - + + - - -
0.5mg/mL - - - - - -
0.25mg/mL - - - - - -
Controle
negativo
- - - - - -
Controle
Positivo
+ + + + + +
ATCC01: S. aureus; ATCC06: M. luteus; ATCC138: E. faecalis; ATCC224: E. coli; ATCC71: M. smegmatis; NCCLS700: S. aureus. (+) inibição (-) não
inibição.
Os testes realizados com o extrato metanólico
apresentaram discrepância nos valores obtidos, S. aureus
(ATCC 01) e M. smegmatis (ATCC 71) expressaram CIM e
CBM de 4mg/mL e 8mg/mL, de modo respectivo, para E.
faecalis (ATCC138) a CIM foi 1 mg/mL e CBM 2 mg/mL, tanto
M. luteus (ATCC 06) como S. aureus (NCCLS 700) obtiveram
CIM de 2 mg/mL, e a CBM 4 mg/mL e 2 mg/mL,
respectivamente, a E. coli (ATCC 224) mostrou CIM e CBM de
4 mg/mL (Tabela 8 e 9).
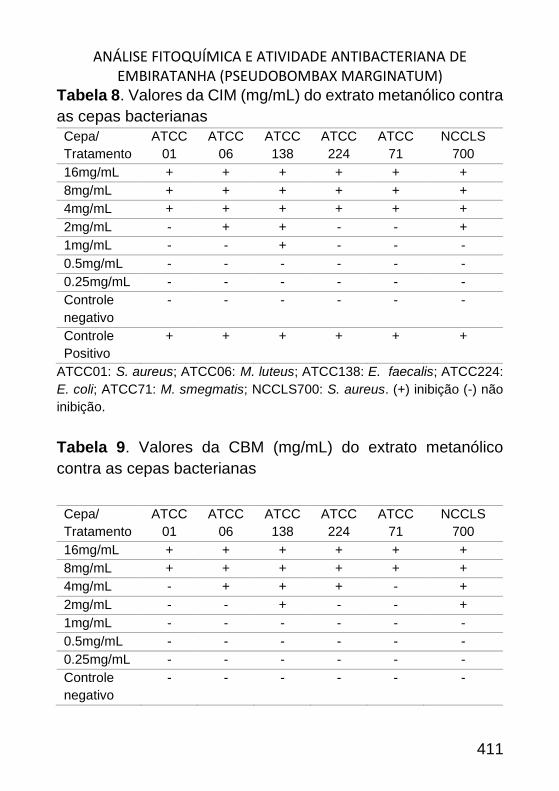
ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
411
Tabela 8. Valores da CIM (mg/mL) do extrato metanólico contra
as cepas bacterianas
Cepa/
Tratamento
ATCC
01
ATCC
06
ATCC
138
ATCC
224
ATCC
71
NCCLS
700
16mg/mL + + + + + +
8mg/mL + + + + + +
4mg/mL + + + + + +
2mg/mL - + + - - +
1mg/mL - - + - - -
0.5mg/mL - - - - - -
0.25mg/mL - - - - - -
Controle
negativo
- - - - - -
Controle
Positivo
+ + + + + +
ATCC01: S. aureus; ATCC06: M. luteus; ATCC138: E. faecalis; ATCC224: E. coli; ATCC71: M. smegmatis; NCCLS700: S. aureus. (+) inibição (-) não
inibição.
Tabela 9. Valores da CBM (mg/mL) do extrato metanólico
contra as cepas bacterianas
Cepa/
Tratamento
ATCC
01
ATCC
06
ATCC
138
ATCC
224
ATCC
71
NCCLS
700
16mg/mL + + + + + +
8mg/mL + + + + + +
4mg/mL - + + + - +
2mg/mL - - + - - +
1mg/mL - - - - - -
0.5mg/mL - - - - - -
0.25mg/mL - - - - - -
Controle
negativo
- - - - - -

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
412
Controle
Positivo
+ + + + + +
ATCC01: S. aureus; ATCC06: M. luteus; ATCC138: E. faecalis; ATCC224: E. coli; ATCC71: M. smegmatis; NCCLS700: S. aureus. (+) inibição (-) não
inibição.
DISCUSSÃO
A resistência desenvolvida pelos patógenos a múltiplas
drogas ocorre de forma progressiva como resultado da meia-
vida dos medicamentos ou devido à distribuição espacial das
drogas, permitindo que, apenas em determinadas regiões do
organismo um subconjunto de fármacos alcance o nível
terapêutico (MORENO-GAMEZ et al., 2015; BAYM; STONE;
KISHONY, 2016).
Esses fatores reforçam a necessidade de estudos
voltados a agentes multirresistentes, além de contribuir para a
ascensão de novas perspectivas farmacológicas (AHMED et al.,
2017), dentre as quais o uso de plantas medicinais, que possuem
compostos bioativos, capazes de promover alterações
fisiológicas no organismo (NAIK; AL–KHAYRI, 2016; YUE et al.,
2016).
A presunção dos biocompostos é realizada por
cromatografia em camada delgada (CCD) com o emprego de
reveladores específicos e sistemas de eluição sob diferentes
proporções, tornando o método mais eficiente (WAGNER;
BLADT, 2001). O revelador de Liebermann-Burchard
(ADAMOVICS, 2017) induz a formação de bandas em tons
característicos, remetendo a existência de triterpenos ou
esteroides no extrato (ELBERRY et al., 2015).

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
413
Notoriedade identificada na eluição com hexano, essa
exibiu coloração violeta sugestiva da ação de esteroides e
triterpenos (Tabela 2). A falta de polaridade dos esteroides ou
compostos derivados de terpenos, permite que ambos fiquem
evidenciados em solventes polares como o hexano, ou, em
menor quantidade, em solventes com polaridade intermediaria
como o acetato de etila (AcOEt) (SIMÕES et al., 2016).
Segundo Das et al. (2016), a análise de flavonoides em
extratos eluidos com AcOEt levam a formação da cor violeta ao
reagirem com o revelador de Liebermann, em contrapartida em
meio a sulfato cérico e anisaldeído formam uma tonalidade
amarelada. Desse modo, é evidente que a revelação sob
diferentes comprimentos de ondas promove a diversificação da
coloração no gel, tais características também estão relacionadas
ao tipo de estrutura presente na solução (ISHIHARA et al., 2015).
Similaridade observada neste estudo, na qual houve alterações
na coloração, sugestivas da presença de triterpenos e
flavonoides nos extratos de AcOEt (Tabela 2).
Os dados inferidos na triagem fitoquímica das folhas de
Embiratanha, mostrou resultados positivos para esteroides,
flavonoides e taninos extratos de hexano, AcOEt, MeOH e água
(Tabela 3). Similaridade analisada por Paiva (2013), em estudos
com a entrecasca do vegetal, na qual foi detectado a presença
de compostos através de reações químicas que induziam o
aparecimento de colocaração e/ou precipitado característicos
para cada substância.
Almeida, Oliveira e Falcão (2015), traçaram o perfil
fitoquímico das folhas e entrecasca da Pseudobombax
marginatum, utilizaram extratos hidroalcoólicos, na qual se

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
414
destacou além dos achados propostos por Paiva (2013), a
presença de alcaloides nas amostras.
No entanto, os testes realizados nesta pesquisa
remeteram a negatividade de alcaloides nas folhas de
Embiratanha, tanto no teste do perfil (Tabela 2), como na triagem
fitoquímica (Tabela3). Segundo Zhang et al. (2018), esse
resultado apresentado pode estar relacionado a metodologia
usada na identificação, capazes de gerar interferências na
quantificação do componente, visto que o percentual encontrado
nos vegetais é limitado.
Os compostos biologicamente ativos encontrados nas
plantas, conseguem promover modificações em
microrganismos, induzem lesões na membrana citoplasmática
do mesmo, desde modo, permitem a degradação da parede
celular, danos as proteínas de membrana, liberação de
conteúdo celulares e consequentemente perda de força protón-
motriz (DHOLVITAYAKHUN, et al. 2017).
Chaves et al. (2013), relacionou a variação sazonal de
metabolitos secundários produzidos por P. marginatum quanto
aos efeitos antibacterianos, inferindo que as concentrações
testadas dos extratos conduziram a completa inibição no
crescimento das cepas testadas, exceto para Pseudomonas
aeroginosa, no entanto, os valores da CIM contra K.
pneumoniae mostraram-se significativos com valores de 12,5
mg.mL-1.
Similaridade observada nesta pesquisa, na qual diluição
de 0,5 µg/mL apresentada no teste com hexano mostrou
bactericida contra cepas de Enterococcus faecalis (ATCC 138)
(Tabela 5), sugerindo que, concentrações relativamente baixas
do extrato possuem atividade antimicrobiana. Os valores para

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
415
CIM e CBM para extratos de AcOEt foram de 1,0 mg/mL (Tabela
6 e 7), respectivamente, entretanto para as soluções que
continham MeOH as concentrações de CIM foi de 1,0 µg/mL e
CBM de 2,0 µg/mL, para essa mesma espécie (Tabela 8 e 9).
A técnica utilizada no ensaio, meio de crescimento,
microrganismos e os meios extrativos são fatores relacionados
a efetividade da atividade antimicrobiana (AMPARO, et al.
2018), permitindo, portanto, possíveis discrepâncias nos valores
de CIM e CBM para diferentes espécies analisadas.
Segundo Bölgen et al. (2020), a CIM sofre variações de
acordo com a sensibilidade e as espécies de microrganismos.
Notoriedade analisada entre as cepas padrões Micrococcus
luteus (ATCC 06), Escherichia coli (ATCC 224) e Mycobacterium
smegmatis (ATCC 71), na qual valores em torno de 2,0 µg/mL
foram suficientes para inibir o crescimento do patógeno teste. A
CBM do extrato seguiu a metodologia proposta por Hafidh et al.
(2011), considerando-se que fitocompostos apresentam
propriedades bactericida quando o coeficiente entre CBM/CIM
está entre 1 e 2, efeito capaz de provocar eventual morte do
microrganismo. Desta forma os extratos provenientes das folhas
de P. marginatum apresentam efeito bactericida, mostrando ser
efetivo contra todas as cepas nesse estudo.
CONCLUSÕES
Através desse estudo é possível inferir que o extrato de
P. marginatum comportam como uma nova alternativa
farmacológica, visto que é constituído flavonoídes, alcaloides,
taninos e triterpenos. Os métodos extrativos utilizados para as
análises fitoquímicas auxiliaram na prospecção desses

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
416
constituintes. Além disso, os extratos obtidos induziram
resposta antimicrobiana contra as cepas bacterianas testadas.
Dessa forma, o extrato proveniente da P. marginatum é um
promissor agente antibacteriano, sendo necessárias
investigações mais minuciosas acerca dessa espécie.
REFERÊNCIAS BIBLIOGRÁFICAS ADAMOVICS, J. A. (Ed.). Chromatographic analysis of pharmaceuticals, ed. 2, Routledge, 2017. AHMED, E.; ELKHATEEB, W.; TAIE, H.; RATEB, M.; FAYAD, W. Biological capacity and chemical composition of secondary metabolites from representatives Japanese lichens. Journal of Applied Pharmaceutical Science, v. 7, n. 1, p. 98-103, 2017. ALMEIDA, H. M.; OLIVEIRA, F. F. M.; FALCÃO, D. A. Análise fitoquímica dos extratos hidroalcoólico da entrecasca e da folha de pseudobombax marginatum (St. Hill) Rob. Blucher Chemistry Proceedings, v. 3, n. 1, p. 301-308, 2015. AMPARO, T. R.; BRAGA, V. C.C.; SEIBERT, J. B.; SOUZA, G. H. B.; TEXEIRA, L. F. M. Métodos para avaliação in vitro da atividade antimicrobiana de plantas medicinais: a necessidade da padronização. Infarma Ciências Farmacêuticas, v. 30, n. 1, p. 50-59, 2018. ARAUJO, M. I. F.; FREITAS, F. O. R.; MORAIS, A. M. B.; BRUSTEN, V. P.; NOGUEIRA, R. B. SÁ. S.; NOGUEIRA, T. B. SÁ. S.; SOUSA, M. N. A.; ALMEIDA-FILHO, G. G.; LIMA, E. O.; MEDEIROS, C. I. S. Antibacterial Activity and Toxicological Analysis in silico of 7-Hydroxycitronellal. International Journal of Toxicological and Pharmacological Research, v. 9, n. 1, p. 1-5, 2017. BAYM, MICHAEL.; STONE, L. K.; KISHONY, R. Multidrug evolutionary strategies to reverse antibiotic resistance. Science, v. 351, n. 6268, p. aad3292, 2016. BÖLGEN, N.; DEMIR, D.; YALÇIN, M. S.; ÖZDEMIR, S. Development of Hypericum perforatum oil incorporated antimicrobial and antioxidant chitosan cryogel as a wound dressing material. International Journal of Biological Macromolecules, v. 161, n. 15, p. 1581-1590, 2020. CHAVES, T. P.; SANTANA, C. P.; VÉRAS, G.; BRANDÃO, D. O.; FELISMINO, D. C.; MEDEIROS, A. C. D.; TROVÃO, D. M. D. B. Seasonal variation in the production of secondary metabolites and antimicrobial

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
417
activity of two plant species used in Brazilian traditional medicine. African Journal of Biotechnology, v. 12, n. 8, 2013. DAS, M.; MALIPEDDI, H.; NAMBIRAJ, N. A.; RAJAN, R. Phytochemical analysis, antioxidant activity and in vitro growth inhibition of struvite crystals by Ipomoea eriocarpa leaf extracts. Journal of Food Biochemistry, v. 40, n. 2, p. 148-160, 2016. DIÓRGENES, F. E. G.; MAIA, J. H.; SOUZA, L. S.; SOUSA, T. M.; CASTRO, V. G. Caracterizaçãi física e determinação do teor de extrativos da madeira de Embiratanha. Advances in Forestry Science. v. 6, n. 3, p. 757-761, 2019. DHOLVITAYAKHUN, A.; TRACHOO, N.; NARKKONG, N.; CUSHNIE, T. P. T. Using scanning and transmission electron microscopy to investigate the antibacterial mechanism of action of the medicinal plant Annona squamosa Linn. Jornal of Herbal Medicine. v. 7, p. 31-36, 2017. ELBERRY, A. A.; HARRAZ, F. M.; GHAREIB, S. A.; GABR, S. A.; NAGY, A. A.; ABDEL-SATTAR, E. Methanolic extract of Marrubium vulgare ameliorates hyperglycemia and dyslipidemia in streptozotocin-induced diabetic rats. International Journal of Diabetes Mellitus, v. 3, n. 1, p. 37-44, 2015. HAFIDH, R. R.; ABDULAMIR, A. S.; VERN, L. S.; BAKAR, F. A.; ABAS, F.; JAHANSHIRI, F.; SEKAWI, Z. Inhibition of growth of highly resistant bacterial and fungal pathogens by a natural product. The open microbiology jornal, v. 5, n. 1, p. 96-106, 2011. INGROFF, A. E.; CHATURVEDI, V.; FOTHERGILL, A.; RINALDI, M. G. Optimal testing conditions for determining MICs and minimum fungicidal concentrations of new and established antifungal agents for uncommon molds: NCCLS collaborative study. Journal of Clinical Microbiology, v. 40, n. 10, p. 3776-3781, 2002. ISHIHARA, H.; TOHGE, T.; VIEHÖVER, P.; FERNIE, A. R.; WEISSHAAR, B.; STRACKE, R. Natural variation in flavonol accumulation in Arabidopsis is determined by the flavonol glucosyltransferase BGLU6. Journal of experimental botany, v. 67, n. 5, p. 1505-1517, 2015. KUPPUSAMY, P.; YUSOFF, M. M.; MANIAM, G. P.; GOVINDAN, N. Biosynthesis of metallic nanoparticles using plant derivatives and their new avenues in pharmacological applications–An updated report. Saudi Pharmaceutical Journal, v. 24, n. 4, p. 473-484, 2016. MENEZES, M. A. G.; OLIVEIRA NETO, F. B.; BERTINO, L. M.; SILVA, F. F. M.; ALVES,L. A. Quantificação de antocianinas dos extratos de embiratanha (Pseudobombax marginatum). HOLOS, v. 1, p. 30-35, 2015.. MORENO-GAMEZ, S.; HILL, A. L.; ROSENBLOOM, D. I.; PETROV, D. A.; NOWAK, M. A.; PENNINGS, P. S. Imperfect drug penetration leads to

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
418
spatial monotherapy and rapid evolution of multidrug resistance. Proceedings of the National Academy of Sciences, v. 112, n. 22, 2015. NAIK, P. M.; AL–KHAYRI, J. M. Abiotic and biotic elicitors–role in secondary metabolites production through in vitro culture of medicinal plants. In: Abiotic and Biotic Stress in Plants- Recent Advances and Future Perspectives. InTech, 2016. PAIVA, D. C. C.; SANTOS, C. A.; DINIZ, J. C.; VIANA, F. A.; THOMAZZI, S. M.; FALCÃO, D. A. Anti-inflammatory and antinociceptive effects of hydroalcoholic extract from Pseudobombax marginatum inner bark from caatinga potiguar. Journal of ethnopharmacology, v. 149, n. 2, p. 416-421, 2013. ZHANG, C.; CHEN, J.;ZHANG, J.; WEI, S.; JI, H.; WU, X.; MA, W.; CHEN, J. Different processing methods change the oral toxicity induced by Sophora alopecuroides seeds and the contents of five main toxic alkaloids from ethanol extracts determined by a validated UHPLC-MS/MS assay. Brasilian Journal of Pharmacognosy. v. 28, ed. 4, p. 481-488, 2018. SILVA, F. R. G.; MATIAS, T. M. S.; SOUZA, L. I. O.; MATOS-ROCHA, T. J.; FONSECA, S. A.; MOUSINHO, K. C.; SANTOS, A. F. Phytochemical screening and in vitro antibacterial, antifungal, antioxidant and antitumor activities of the red propolis Alagoas. Brazilian Journal of Biology, v. 79, n. 3, p. 452-459, 2019. SIMÕES, C. M. O.; SCHENKEL, E. P.; MELLO, J. C. P.; MENTZ, L. A.; PETROVICK, P. R. Farmacognosia: do produto natural ao medicamento. Artmed Editora, 2016. TIWARI, R.; RANA, C. S. Plant secondary metabolites: a review. International Journal of Engineering Research and General Science, v. 3, n. 5, p. 661-670, 2015. VASUDEVAN, S.; ARULMOORTHY, M. P.; SURESH, R. Isolation, purification and structural elucidation of seconday metabolites from Microcystis aeryginosa bloom from Muttukadu estuary and its in vitro antibacterial, antioxidant and anticancer potency. South African Journal of Botany, v. 132, p. 59-67, 2020. VELÁSQUEZ, S. C. J; HIGUERRA, L. D. T.; ARANGO, J. L. P.; BAUTISTA, J. L. R.; CASTRO, F. E. G.; BURBANO, R. E. P. Profile of antimicrobial resistance in isolates os Staphylococcus spp. obtained from bovine milk in Colombia. Revista Argentina de Microbiología, v.52, n. 2, p. 121-130, 2020. WAGNER, H; BLADT, S. Plant drug analysis: a thin layer chromatography atlas. Nova York: Springer, ed. 2, 2001. WESTHOFF, S.; LEEUWE, T. M. V.; QACHACH, O.; ZHANG, Z.; WEZEL, G. P. V.; ROZEN, D. E. The evolution of no-cost resistance at sub-MIC concentrations of streptomycin in Streptomyces coelicolor.

ANÁLISE FITOQUÍMICA E ATIVIDADE ANTIBACTERIANA DE EMBIRATANHA (PSEUDOBOMBAX MARGINATUM)
419
Multidisciplinary Journal of Microbial Ecology, v. 11, p. 1168-1178, 2017. YUE, W.; MING, Q. L.; LIN, B.; RAHMAN, K.; ZHENG, C. J.; HAN, T.; QIN, L. P. Medicinal plant cell suspension cultures: pharmaceutical applications and high-yielding strategies for the desired secondary metabolites. Critical reviews in biotechnology, v. 36, n. 2, p. 215-232, 2016.

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
420
CAPÍTULO 20
APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
Héberly Fernandes BRAGA 1
Heloiza Ferreira Alves do PRADO 2
1 Doutorando em Microbiologia, UNESP; 2 Orientadora/Professora Doutora, UNESP.
RESUMO: A biodiversidade e potenciais aplicações dos fungos do Cerrado ainda são pouco conhecidas e positivamente exploradas. Assim, a presente revisão objetivou elucidar e apontar a importância de se conservar, conhecer e se utilizar do potencial biotecnológico oferecido pela micobiota desse bioma, de modo a dar um caráter ainda mais valorativo à região, e mostrar os importantes recursos que podem e estão sendo perdidos por meio seu do manejo inadequado. Por meio de pesquisa exploratória nas bases de dados Google Acadêmico, Scielo, Portal Capes e DOAJ, foram selecionados diferentes trabalhos em inglês e português publicados entre 2015 e 2019, empregando-se os seguintes descritores de busca: fungos do cerrado, (bio)prospecção de fungos do cerrado, propriedades de fungos do cerrado, manejo de fungos do cerrado. As diferentes pesquisas encontradas demonstram uma variedade de possibilidades de uso dos fungos do bioma seja no setor agropecuário, industrial, medicinal, alimentício, químico entre outros e apotam para a necessidade de ações e investimentos que visem não somente à conservação da região, e por consequência da sua micobiota, que vêm ao longo de décadas sendo alterada, devido principalmente ao manejo inadequado

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
421
do solo; como também do estímulo à novas pesquisas sistematizadas que possam gerar mais conhecimento e descobertas. Palavras-chave: Biodiversidade fúngica. Bioprospecção. Biotecnologia. Savana brasileira.
INTRODUÇÃO
O Cerrado é um bioma que apresenta uma enorme
riqueza e abundância de vegetais, animais e micro-organismos.
A alta biodiversidade associada às ameças de destruição dessa
região tem colocado a mesma no grupo dos habitats naturais
considerados como hotspots ecológicos (SAWYER et al.,
2016).
Mesmo apresentando e ofertando distintos serviços
ecossitêmicos, como por exemplo: abastecimento hídrico;
alimentos e matérias-primas de valor econômico; recursos
genéticos; sequestro e armazenamento de gás carbônico (CO2)
atmosférico; turismo e lazer; substâncias farmacológicas e
bioprodutos derivados de plantas e outros organismos,
observa-se que sua área vem sofrendo constantes alterações,
consequentes principalmente da urbanização, agricultura e
pecuária desenfreadas.
Os desequilíbrios e desordens vêm historicamente
propiciando a redução e/ou mesmo a perda das riquezas
genéticas e de biodiversidade, especialmente aquelas com
potencial farmacológico e tecnológico destinados ao tratamento
de doenças e/ou aplicações biotecnológicas, como por exemplo
micro-organismos e seus biocompostos derivados (RESENDE
et al. 2019).

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
422
Nesse contexto a presente revisão têm por objetivo
elucidar e apontar a importância de se conservar, conhecer e
se utilizar das distintas pontencialidades biotecnológicas dos
fungos existentes no Cerrado, de modo a dar a essa região um
caráter ainda mais valorativo, e mostrar os importantes recursos
que podem e estão sendo perdidos com o manejo inadequado
desse bioma.
MATERIAIS E MÉTODO
O material bibliográfico empregado na pesquisa foi
selecionado nas bases de dados Google Acadêmico, Scielo,
Portal Capes e DOAJ, e em sítios eletrônicos de órgãos
governamentais, por meio de pesquisa qualitativa exploratória
realizada em trabalhos publicados entre 2015 a 2019.
Como critérios de inclusão foram considerados artigos
em inglês e português que indicassem possíveis aplicações
biotecnológicas de fungos do Cerrado e/ou pesquisas que
elucidassem as perdas da micobiota nesse determinado bioma,
por manejos inadequados.
Os seguintes descritores foram empregados na busca:
fungos do cerrado, (bio)prospecção de fungos do cerrado,
propriedades de fungos do cerrado, manejo de fungos do
cerrado.
Trabalhos não enquadrados nos critérios de inclusão,
assim como aqueles que somente relatassem quantificação da
micobiota local; ou descobertas de novas espécies sem elucidar
as possíveis aplicações biotecnológicas foram excluídos da
busca.

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
423
POTENCIAIS APLICAÇÕES BIOTECNOLÓGICAS E
AMEAÇAS À MICODIVERSIDADE DO CERRADO
As alterações fitofisionômicas derivadas do
desmatamento, que historicamente vêm assolando o bioma
Cerrado, podem promover mudanças na composição do solo e
sua maior exposição aos raios solares, procipando por
exemplo, alterações físico-químicas na acidez e umidade,
fatores esses essenciais a permanência e desenvolvimento da
microbiota residente.
Dessa forma, estudar e entender a dinamicidade,
funcionamento e interação dos micro-organismos existentes no
Cerrado, e em especial no solos dessa região, não só propicia
um maior entendimento dos impactos ambientais e formas
inadequadas de manejo que o bioma vem sofrendo
(VALADARES-PEREIRA, et al.; 2017), como também viabiliza
a possibilidade se prospectar espécies com ponteciais
aplicações de interesse biotecnológico, por exemplo (NORILER
et al., 2018).
Estudo realizado por Jacinto, Moura e Uto (2018), em
diferentes áreas de cerrado (nativo; de cultivo agrícola; e de
reserva ambiental urbana/reflorestamento), visando determinar
o grau da antropicidade do solo frente à biomassa de bactérias
e fungos, evidencia os impactos que o bioma têm sofrido. Eles
verificaram contagens significativamente distintas (p < 0,05) de
bactérias em unidades formadoras de colônia por grama
(UFC.g-1) de solo, em valores de 4,2 x 105; 1,1 x 105 e 1,8 x 105,
respectivamente, nos solos das áreas de cerrado nativo, cultivo
agrícola e reflorestamento. As contagens fúgincas (UFC.g-1)
também foram significativamente distintas, 9 x 104 (áera nativa),

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
424
3 x 104 (cultivo agrícola) e 6,6 x 103 (reflorestamento), mas com
valores inferiores à da população bacteriana, indicando
segundo os autores, a maior sensibilidade dos fungos à
atividade antrópica.
Valadares-Pereira et al. (2017), também evidenciaram
mudanças na comunidade fúngica em solo do cerrado
tocantinense, ao comparar uma área nativa com outra
destinada ao plantio de soja. Analisando o ácido
desoxirribonucleico (DNA) do solo, por meio de Análise
Espacial Intergênica Ribossomal Automatizada (ARISA), e
comparando os resultados aos teores de cobre, cálcio, ferro,
capacidade de troca de cátions, saturação por bases e relação
cálcio/magnésio, de ambas as regiões, os pesquisadores
observaram mudanças na estrutura da comunidade fúgica
como efeito do manejo agrícola.
Araujo et al. (2017), avaliaram a diversidade fúngica de
uma área de cerrado preservado, na região nordeste do estado
do Piauí, dentro do Parque Nacional das Sete Cidades, ao
longo de distintos gradientes fitofisionômicos: formação
campestre, cerrado stricto sensu, cerradão e floresta decidual.
Os pesquisadores identificaram a maior riqueza de espécies na
floresta decidual, sendo a maior porcentagem dos isolados
pertencentes ao filo Ascomycota (45%), seguido do
Basidiomycota (32%). Conforme os autores, as classes mais
abundantes encontradas incluindo espécies com potencial
biotecnológico foram: Agaricomycetes, Eurotiomycetes,
Lecanoromycetes, Basidiobolus, Dothideomycetes e
Taphrinomycetes.
Conforme Bononi et al. (2017), a diversidade de micro-
organismos, e especialmente dos fungos presentes em regiões

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
425
do Cerrado, ainda não é totalmente conhecida. Em coletas
realizadas pelos pesquisadores, na região central do estado de
Mato Grosso do Sul (MS), centro-oeste do Brasil, foram
identificadas 18 espécies de fungos (Conocybe crispa,
Gymnopus dryophilus, Marasmius androsaceus, Marasmius
oreades, Mycena pura, Rhodocollybia butyraceae, Pleurotus
eryngii, Coprinellus micaceus, Panaeolus rickenii, Hypholoma
fasciculare, Clitocybe infundibuliformis, Tricholoma
argyraceum, Fomitiporia robusta, Cymatoderma elegans,
Athelopsis galzinii, Trametes ellipsospora, Guepinia
helvelloides, Tremella mesentérica) ainda não citadas e listadas
nacionalmente, e 36 espécies com primeira citação para o
estado. Do total de 62 espécies de basidiomicetos coletados, os
autores identificaram 15 comestíveis, quatro tóxicas, 10 com
propriedades medicinais, duas com potencialidade de aplicação
em processos de biorremediação, e uma com propriedade
bioluminescente, conforme comparações à literatura disponível.
Plantas medicinais têm sido muito estudadas como
reservatório de cepas de micro-organismos endofíticos com
potencial biotecnológico. Nesse sentido, Noriler et al. (2018),
estudaram comunidades fúngicas endofíticas associadas a
folhas e pecíolos da planta medicinal Stryphnodendron
adstringens, de ocorrência no Cerrado. Dentre os 117
morfotipos fúngicos (95,6% deles pertencentes ao filo
Ascomycota) isolados pelos pesquisadores, cinco (Diaporthe
sp. LGMF1530, Diaporthe cf. heveae LGMF1631,
Colletotrichum siamense LGMF1604, Fusarium sp. LGMF1630
e Epicoccum sp. LGMF1641) apresentaram atividade inibitória
(maior que 60%) sobre o crescimento micelial de Phyllosticta

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
426
citricarpa (LGMF06) e Colletotrichum abscissum (CA142),
patógenos estes de grande importância na citricultura.
Isolados endofíticos também apresentaram efeito
inibitório frente a patógenos de interesse clínico (NORILER et
al., 2018), demonstrando as potencialidades biotecnológicas
dos fungos encontrados, não só como alternativa ao controle
biológico de fitopatógenos, como também de micro-organismos
associados a saúde humana. Phaeophleospora sp.2
LGMF1513, Epicoccum sp.1 LGMF1598 e Epicoccum sp.2
LGMF1600 tiveram alta atividade de ação (+++) contra
Staphylococcus aures (American Type Culture Collection -
ATCC 33591) meticilino-resistentes (MRSA). Alguns isolados
fúngicos (Diaporthe sp. LGMF1548 e LGMF1614, Diaporthe cf.
heveae LGMF1631, Fusarium sp. LGMF1630 e Hypoxylon sp.1
LGMF1613) se mostraram também ativos contra S. aures
(MRSA), Pseudomonas aeruginosa (ATCC 27853),
Acinetobacter baumannii (BACHAC-ABA) e Klebsiella
pneumoniae (BACHC-KPC) produtora de carbapenemase
(KPC).
Em um estudo realizado com Miconia albicans (Sw.)
Triana (Melastomataceae), planta fanerógama edêmica do
Cerrado, conhecida popularmente como “quaresmeira-branca”,
Piza, Hokka e Sousa (2015) também isolaram e verificaram alta
atividade antagonista do fungo actinomiceto Amycolotopsis
orientalis frente a S. aureus, Enterococcus faecalis, Candida
albicans, Shigella sonnei.
Alternativas de controle biológico de artrópodes-praga
frente a culturas agrícolas, também vêm sendo muito aplicadas.
Por exemplo, em sistemas agroflorestais da região neotropical,
alguns organismos destacam-se como as principais pragas que

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
427
assolam as culturas agrosilvícolas, sendo as formigas
cortadeiras dos gêneros Atta e Acromyrmex, muito citadas. Tais
formigas, conhecidas popularmente como saúvas e quenquéns
atacam as partes frescas das plantas e as carregam para o
interior dos seus ninhos, onde servirão de substrato ao fungo
simbionte Leucoagaricus gongylophorus (Möller) Singer
(Basidiomycota: Agaricales: Lepiotaceae) do qual elas se
alimentam (NYGAARD et al., 2016).
A forma mais usual de controle desses insetos é pelo
método químico, entretanto a busca por alternativas
ambientalmente corretas tem sido estimulada. Nesse sentido,
Dornelas et al. (2016) realizaram estudo prospectivo de fungos
filamentosos, associados a operárias de Atta sexdens e A.
laevigata da região do cerrado tocantinense, com propriedades
patogênicas. Os autores isolaram de A. sexdens os fungos
Metarhizium anisopliae (Metchnikoff) Sorokin, Aspergillus flavus
Link, Acremonium sp.1, Aspergillus sp.1, Colletotrichum sp. e
Acremonium sp. 2, e de A. laevigata, Mucor sp., A. flavus,
Fusarium solani (Martius) Saccardo, A. niger van Tieghem;
entretanto o teste de patogenicidade contra a formiga A.
sexdens foi realizada somente com os fungos A. flavus, A. niger
e M. anisopliae, pois esses apresentaram alta produção de
conídios e elevada prevalência no estudo. Foi verificada uma
taxa de letalidade de 50% das operárias em apenas cinco dias,
nos testes com A. niger e M. anisopliae, sendo essa duração
significativamente inferior ao tratamento controle.
Estudo realizado por Zanardo (2015) em solos dos cinco
biomas do Brasil corrobora a importância que o Cerrado tem em
abrigar uma das maiores quantidades de fungos
entomopatogênicos. A autora determinou a abundância de

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
428
fungos ao longo de dois anos em amostras de solos de cultivos
anuais, perenes e vegetação nativa, durante estação climática
seca e úmida, de cinco áreas representativas dos biomas
brasileiros, e verificou no geral, maiores densidades de
entomopatogênicos no Cerrado (2,27 ± 1,58 x 103 a 3,46 ± 1,51
x 104 UFC.g-1 de solo), sendo o gênero Metarhizium spp. o de
maior predominância.
Conforme mencionado anteriormente, além do emprego
dos fungos no controle biológico e como bioindicadores da
qualidade do solo em manejos diversos, esses micro-
organismos tem importância preponderante no setor agrícola
atuando na melhoria da produtividade das culturas, não só
porque realizam a ciclagem de nutrientes, ao decompor e
transformar materiais orgânicos em minerais, que serão assim
aproveitados pelas plantas, mas também pela associação
simbionte (micorrizas) com raízes de muitas espécies vegetais,
proporcionando a essas aumento da superfície de contato com
o solo, para absorção de água e sais minerais.
Dentre os grupos de micorrizas destacam-se os fungos
micorrízicos arbusculares (FMA), também denominados
endomicorrizas, que tem como característica diferencial
desenvolver um micélio que penetra internamente no córtex da
raiz, invadindo tanto o espaço intra quanto intercelular. Berude
et al. (2015), em pesquisa bibliográfica, compilaram outros
distintos benefícios às plantas quando de sua associação aos
FMA: redução ao estresse hídrico e salino; diminuição de sítios
de infecção por patógenos; aumento na absorção de fósforo e
nitrogênio do solo e maior tolerância a metais pesados.
Experimento realizado por Salgado et al. (2016), ilustra a
potencialidade de fungos do Cerrado em propiciar o

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
429
crescimento inicial de plantas de feijoeiro e soja. Os
pesquisadores coletaram, isolaram e idetificaram FMA
(Acaulospora scrobiculata, Claroideoglomus etunicatum,
Dentiscutata heterogama, Gigaspora margarita e Rhizophagus
clarus) provenientes de solo do Cerrado (região sudoeste de
Goiás), os quais foram associados ao estimulante de
colonização (7-hidroxi,4’-metoxi-isoflavona) e inoculados sobre
ambas culturas. Eles observaram aumento da biomassa nas
plantas, mais de 200% para o feijoeiro e mais de 80% para a
soja, partindo-se dos seguintes parâmetros de avaliação:
massa seca da parte aérea, massa seca da raiz, colonização
micorrízica, massa de nódulos e acúmulo de cálcio, zinco e
fósforo na massa seca da parte aérea.
Em outro experimento, partindo das mesmas
condicionantes, Salgado et al. (2017), determinaram os efeitos
de tal combinação frente ao crescimento inicial de plantas de
algodão e milho, e verificaram que baixa densidade de
propágulos de FMA inoculados e associados a estimulante de
colonização foram também benéficos ao desenvolvimento de
ambas culturas.
Santos et al. (2018), avaliaram os efeitos da inoculação
de FMA, provenientes de solo de Cerrado conservado, sobre
uma região degradada pela construção de uma usina
hidrelétrica, no município de Selvíria-MS. Eles verificaram que
a combinação de resíduos orgânicos/minerais com a aplicação
dos fungos influenciou positivamente o solo degradado, pois o
teor de carbono da massa microbiana (CBM), carbono de gás
cabônico liberado (C-CO2) e quociente microbiano (qMic)
elevaram-se. Todos esses parâmetros são indicadores de
qualidade do solo.

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
430
Outro aspecto a se considerar são os estudos sobre
emprego da atividade fúngica para a melhoramento de
processos de produção, extração, solubilização e
transformação no setor industrial. Os mesmos vêm crescendo
e propiciando vantagens em termos ambientais e de obtenção
de produtos com maior valor econômico agregado. A produção
de biossurfactantes por micro-organismos, por exemplo, é uma
opção promissora em termos de obtenção de produtos
biotecnológicos, haja visto a baixa toxicidade, alta
biodegrabilidade e melhor compatibilidade ambiental quando
comparados aos surfactantes sintetizados quimicamente.
Partindo disso, Passone et al. (2015) avaliaram o
potencial biossurfactante de três fungos isolados de ambientes
do Cerrado na região do Triângulo Mineiro, codificados como
C18, C35 e C40, frente a diferentes concentrações de glicerol,
empregando fermentação submersa. Os autores observaram
índices de emulsificação variando de 28,6% a 71,4%. Foi
notado também aumento nesse parâmetro (em média 10%),
com o aumento do tempo de fermentação (48 h para 72 h), nos
cultivos contendo concentrações de glicerol 2,5; 5,0 e 10% (v/v)
inoculados com o fungo C35, além um dos menores valores de
tensão supercial (40 mN/m-1) na condição de 1,5% (v/v) de
glicerol a 48 h. Altos íncides de emulsificação e baixos valores
de tensão superficial são bons indicativos de atividade
surfactante.
A produção e oferta interna de fertilizantes fosfatados no
Brasil ainda tem sido um gargalo no setor agrícola,
especialmente porque a rocha fosfática, principal insumo da
indústria de fertilizantes, é um mineral pouco disponível no país
e ainda apresenta baixo teor de pentóxido de fósforo (P2O5).

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
431
Solos aráveis, como os do Cerrado, demandam fósforo que é
pouco disponibilizado e muitas vezes insolúvel para ser
utilizado pelas culturas agrícolas, necessitando assim serem
suplementados com fertilizantes. Associado a isso, a extração
dos fosfatos é realizada empregando-se ácido fosfórico (H3PO4)
ou sulfúrico (H2SO4), o que gera resíduos. Tais condições
tornam o custo da produção nacional elevado e ambientalmente
pouco sustentável, suscitando pesquisas que se utilizem de
métodos naturais renováveis e que agridam menos o ambiente,
tal como a biossolubilização realizada por micro-organismos
(EMBRAPA, 2015).
Nesse sentido, Cabral et al. (2017) isolaram Trichoderma
harizianum, um fungo filamentoso do filo Ascomycota, de um
complexo minerador do município de Araxá-MG, e testaram sua
capacidade de biossolubilizar fósforo a partir de concentrado de
rocha fosfática (teor de P2O5 igual a 37,5%) originária do
complexo de mineração do município de Tapira-MG.
Empregando-se fermentação submersa em biorreator air-lift
com reciclo do meio reacional, e variando o fluxo volumétrico de
ar ascendente e a concentração de rocha fosfática, os
pesquisadores obtiveram uma concentração de fósforo solúvel
em meio líquido igual a 244,9 partes por milhão (ppm) ou
33,78% de solubilização, num período de três dias a 28 ± 0,5°C.
Gomes et al. (2017), obtiveram 11,95% (94,8 ppm) de
solubilização de fósforo em 14 dias, e 6,81% (52 ppm) em 9
dias, empregando o mesmo concentrado de rocha fosfática, na
fermentação sólida e líquida, respectivamente, com o fungo T.
ressei, coletado no Cerrado.
Estudo realizado por Rocha (2017), com a levedura
Torulaspora globosa, isolada da rizosfera de milho, no

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
432
município de Araras-SP, evidenciou também a rápida
solubilização de fosfato tricálcio (Ca3(PO4)2) em fermentação
submersa. A autora obteve quantidade máxima de fósforo
solúvel (42%) em apenas 48 h, demonstrando a potencialidade
do fungo em aplicações industriais e agrícolas.
Propiciar o aumento e variedade da quimiodiversidade,
isto é, da obtenção de novas substâncias derivadas da ação
metabólica de micro-organismos e/ou enzimas isoladas, sobre
compostos orgânicos ou sintéticos, têm sido uma nova
estratégia biotecnológica alternativa de síntese química, dentro
da área de química verde. Para Hegazy et al. (2015), o uso
desse processo apresenta vantagens quando comparado a
síntese tradicional: baixo custo; quimio, estereo e
regioseletividade; manutenção das cepas; redução no número
de animais experimentais; maior quantidade e variedade de
compostos produzidos; consumo mínimo de energia e menor
geração de resíduos, especialmente compostos tóxicos e
danosos ao ambiente.
Silva (2015), testaram cepas de Beauveria sp. isoladas
de solo do Cerrado da região de Goiás, frente a ácido gálico
(AG) e elágico (AE), utilizando cultivo submerso em meio
glicose e Corn Steep, durante 96 h a 27ºC e 200 rotações por
minuto (rpm). Empregando-se Cromatografia em Camada
Delgada (CCD) e Cromatografia Líquida de Alta Eficiência
(CLAE), foi notado que todas as cepas foram eficientes na
biotransformação dos compostos, sendo que o meio contendo
Corn Steep proporcionou maior variedade e quantidade de
compostos formados para ambos substratos. Os derivados do
AG obtidos e caracterizados estruturalmente por métodos
espectroscópicos foram: ácido benzoico, 3-O-metil-β-

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
433
glucopiranosil-galatoester (Labiocon 465); ácido benzoico,4-[[6-
O-(3,4-diidroxi-5-metoxibenzoico)-α-D-glucopiranosideo]oxi]-3-
hidroxi-5metoxi (Labiocon 484); ácido gálico 3-O-β-
glucopiranoside (Labiocon 510) e ácido 3,4-Diidroxi-5-metoxi-
benzoico-2-hidroxi-5-metoxicarbonil-fenilester (Labiocon 530).
E os do AE: 3,8-Dimetoxi-dibenzo[b,d]piran-6-one (Labiocon
511) e o ácido elágico 4-O,3’-O-metil-β-D-glicose (Labiocon
534).
Na indústria de alimentos, algo que tem se tornado
corriqueiro é o emprego de insumos e/ou ingredientes com
caráter mais natural para a elaboração de produtos
alimentícios, dada a mudança de atitudes do consumidor e sua
maior preocupação com a saúde. Para isso, tem-se por
exemplo substituído o emprego de aditivos químicos artificiais,
por compostos naturais derivados de animais, plantas ou
mesmo micro-organismos.
Conforme Rao, Xiao e Li (2017), a demanda por corantes
alimentícios naturais derivados de micro-organismos, por
exemplo, tem aumentado dia a dia, não somente pelos efeitos
nocivos cientificamente relatados da ingestão de alguns
corantes sintéticos, mas também porque pigmentos de origem
microbiana apresentam vantagens quando comparados a
outros produtos naturais: rapidez na produção; fácil
processamento; independência das condições climáticas; e
propriedades antioxidantes, antimicrobianas e
anticancerígenas.
Entre os distintos pigmentos microbianos, aqueles
derivados de fungos ambientais e especialmente do solo, tem
sido pesquisados e vem apresentando-se como alternativa
favorável não só na medicina, como também na indústria

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
434
cosmética, têxtil e principalmente alimentícia. Nesse sentido,
Machado (2018) isolou leveduras com potencial carotenogênico
do ambiente (folhas, solo, flores, frutos silvestres e insetos) do
Cerrado e verificou que as espécies Rhodotorula glutinis, R.
graminis, R. aurantiaca, R. lactosa e R. mucilagionosa se
destacaram na produção de carotenoides. Realizando testes de
otimização em condições de cultivo experimental submerso
com variação de agitação (130 a 230 rpm), temperatura (25 a
35ºC), pH (4,5 a 6,5) e componentes do meio de cultivo, o autor
observou que a levedura R. mucilaginosa foi a mais efetiva na
bioprodução do pigmento (4164,45 µg.L-1).
Em outro estudo bioprospectivo de fungos endofítifos de
folhas de Hymenaeae coubaril (jatobá), árvore típica do
cerrado, Romano (2015) identificou pontencial produção de
pigmento vermelho pelo fungo Talaromyces minioluteus,
empregando-se diferentes meios de cultivo a 28ºC, por 72 h e
a 200 rpm, ao se comparar a mensuração da biomassa após os
cultivos, com índices de absorbâncias obtidos a 510 nm.
Conforme o autor, a intensa coloração vermelha obtida em
todos os cultivos realizados evidencia o possível emprego
biotecnológico da espécie, entretanto sugere-se a continuidade
das pesquisas visando-se não somente identificar
quimicamente o pigmento, como verificar a micotoxidade,
empregar substratos alternativos de menor custo, e também
otimizar o processo de produção.
No setor industrial, especialmente alimentício, a
produção biotecnológica de frutooligossacarídeos (FOS) e
exopolissacarídeos (EPS) empregando-se micro-organismos
têm sido muito demandada, não somente por serem
considerados naturais; como também pela praticididade e

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
435
facilidade de obtenção quando comparado a extração realizada
de fontes vegetais; além desses polissacarídeos apresentarem
benefícios à saúde, bem como propriedades tecnológicas que
melhoram as características físico-químicas, reológicas e
sensoriais dos produtos destinados ao consumidor
(BARCELOS et al., 2019).
Diante dos benefícios e aplicações dos FOS, Zaninette
et al. (2019) observaram atividade hidrolítica e capacidade de
transfrutosilação de solução de sacarose (150 g.L-1), com
produção de 8 g.L-1 de 1-cestose (FOS do tipo inulina) e 7,3 g.L-
1 de neocestose (FOS do tipo neolevana, comumente não
produzido por fungos filamentosos), pelo fungo Penicillium
janczewskii Zaleski CCIBt 3352, isolado da rizosfera de uma
planta do Cerrado (Chrysolaena obovata), evidenciando a
promissora capacidade sintetizadora de FOS por uma cepa
selvagem. Outra pesquisa realizada com diferentes frutos do
Cerrado (SILVEIRA, 2018), corrobora a necessidade de
estudos prospectivos com viés biotecnológico mais
sistematizados sobre o bioma. A autora isolou 17% de micro-
organismos (de um total de 59), com potencialialidades de
produção de EPS, sendo que uma das linhagens alcançou uma
produção máxima desses bioprodutos de 1,2 g.L-1 após 2 h de
processo a 30ºC e 150 rpm.
No setor de energias renováveis, em especial na
produção e melhoria de processos para obtenção
biocombustíveis e/ou biodiesel, os fungos também vêm
apresentando suas potenciais aplicações.
Nos últimos anos o crescimento incessante da população
tem demandado o consumo de recursos energéticos para suprir
o bem-estar individual, desempenho industrial ou mesmo

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
436
serviços de manutenção; promovendo a escassez de fontes de
energia não renováveis como o petróleo, por exemplo, e a
intensa emissão de gases poluentes para a atmosfera. Assim,
o uso da bionconversão de diferentes resíduos empregando-se
fermentação microbiana veêm sendo adotado como alternativa
para se otimizar a aquisição de combustíveis mais sustentáveis,
e ao mesmo tempo minimizar alterações ambientais
decorrentes da disposição de subprodutos e rejeitos com
características poluentes (ZANONI et al., 2015).
Um dos gargalos da produção de biocombustíveis,
especialmente os de segunda geração, deve-se às dificuldades
em se despolimerizar as longas cadeias carbônicas que
constituem a parede celular vegetal, para se disponibilizar os
açúcares fermentescíveis que serão convertidos em álcool.
Assim, diferentes estudos e esforços públicos e privados têm
sido realizados, visando selecionar micro-organismos que
apresentem tal capacidade bioconversora, de forma a se
reduzir o custo atual do refinamento da biomassa vegetal.
Diante disso, Dabaja et al. (2019), testaram 43 culturas
de fungos endofíticos isolados de Annona crassiflora (araticum),
uma planta nativa e exclusiva do Cerrado, e verificaram que 8%
apresentaram atividade esterásica; 34,43% atividade
celulolítica; 73,91% atividade fosfatásica; 78,26% atividade
lipolítica; e 91,3% atividade β-frutocfuranosidásica. Os autores
ainda realizaram a fermentação submersa para determinação
da atividade lipolítica, visto serem as lipases enzimas muito
aplicadas em biocombustíveis (principalmente na produção de
biodiesel), e constataram uma atividade expressiva de 27,34
U.g-1 em 60 h de fermentação a 28ºC.

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
437
Bessa (2018) isolou fungos do solo do Cerrado e
verificou atividades lipolíticas promissoras de 9,268 U.mg-1 e
7,032 U.mg-1 para duas cepas Paecilomyces formosus (isolado
I e II); e 6,447 U.mg-1 para Aspergillus tubingensis. A autora
ainda observou que o extrato enzimático bruto proveniente do
isolado II de P. formosus conseguiu atingir 41,60% de
conversão de óleo:metanol em ésteres metílicos. Tal
capacidade de transesterificação demonstra o potencial das
enzimas para emprego na produção de biodiesel,
principalmente porque esse processo é considerado mais limpo
quando comparado ao uso de catalizadores químicos.
Dentre os distintos micro-organismos presentes no solo
do Cerrado, bactérias e fungos que apresentam capacidade de
excretar enzimas para o meio, tem sido alvo de interesse nos
estudos relacionados à bioprospecção e biotecnologia. Isso
porque, a secreção extracelular enzimática, realizada
naturalmente pelo micro-organismo com objetivo de promover
a degradação de compostos complexos para sua posterior
absorção e nutrição, reduz custos nos sistemas de isolamento
de tais bioprodutos quando comparado aos processos onde é
necessário o rompimento celular (ANDRADE-LINHARES;
VERESOGLOU; RILLIG, 2015). Além disso, biocompostos
excretados tendem, de forma geral, a ser mais estáveis aos
distintos fatores ambientais, quando comparados àqueles que
se mantêm no interior da célula, em condições fisiológicas
favoráreis.
Silva et al. (2015), por exemplo, estudando a capacidade
de 66 estirpes de Trichoderma spp. isoladas do solo e madeira
em decomposição da região do cerrado tocantinense,
verificaram que dois dos isolados (UFTJCO85 e UFTMG15)

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
438
apresentaram atividades enzimáticas celulásicas satisfatórias
para FPase (1,34 UI.mL-1) e CMCase (0,81 UI.mL-1), haja visto
que não foram empregados tratamentos prévios do sabugo de
milho testado.
Oliveira et al. (2015) testaram a atividade amilolítica de
enzimas produzidas por leveduras isoladas de ambientais
naturais (frutas e serrapilheira) e industriais (mosto de usina de
cana-de-açúcar) da região de Dourados (MS), e identificaram
três cepas de um total de 54, com boas capacidades de produzir
amilases, a partir de farelo de trigo a 70% de umidade. Entre as
cepas destacaram-se Candida parapsilosis com atividade
máxima 14,68 U.mL-1; Rhodotorula mucilaginosa com 25,0
U.mL-1 e Candida glabrata com 25,39 U.mL-1, empregando
cultivo em estado sólido por 120 h a 28ºC. A amilases
produzidas por tais cepas exibiram atividade máxima em pH 7,0
e temperatura de 60ºC; apresentaram atividade mesmo após 1
h de incubação a 50°C; mativeram atividade catalítica residual
maior que 70% após exposição a solução de 15% de etanol; e
ainda apresentaram maiores atividades enzimáticas frente ao
amido de milho, quando comparado aos amidos de batata, trigo
e mandioca.
Santos et al. (2016) destacaram o potencial
biotecnológico de outro fungo também isolado do solo da região
de Dourados (MS), com capacidade de produção de β-
glicosidades, enzimas empregadas na produção de
biocombustíveis; vinificação e desenvolvimento de alimentos
funcionais. A cepa de G. butleri isolada apresentou a máxima
produção de β-glicosidades (21,5 U.mL-1), em cultivo em estado
sólido a 30°C por 96 h, sobre farelo de trigo a 55% de umidade.
Os autores verificaram ainda uma atividade catalítica máxima

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
439
em pH 4,5 e temperatura de 65°C, com retenção de 80% da
atividade mesmo após manutenção a 50ºC por 1 h.
Em pesquisa realizada por Melo et al. (2018), em
amostras de solo e serrapilheira coletados no Parque Estadual
do Cerrado em Jagariúna (PR), 89,3% dos isolados fúngicos
pertencentes ao filo Ascomycota e Basiodiomycota,
apresentaram atividade celulásica em meio de cultura contendo
carboximetilcelulose (CMC), tendo se destacado Colletotrichum
boninense e Trichoderma sp. Os autores observaram ainda que
todas as cepas apresentaram alguma atividade lipolítica,
demonstrando potencialidades para serem empregadas em
processos biotecnológicos que envolvam degradação de
biomassa vegetal e produção de biocombustíveis.
CONCLUSÕES
As diferentes pesquisas relatadas apontam a
importância e potencialidades do emprego dos fungos do
Cerrado em diferentes produtos, processos, e atividades, sejam
elas agropecuárias, industriais, alimentícias, medicinais,
químicas, entre outras; e elucidam a necessidade de pesquisas
e ações de maior conservação desse bioma, que ainda possui
uma enorme e variada biodiversidade fúngica ainda pouca
conhecida e inexplorada.
REFERÊNCIAS ARAUJO, A.S.F. et al. Fungal diversity in soils across a gradient of preserved Brazilian Cerrado. J Microbiol, v.55, n.4, p. 273-79, 2017. BARCELOS, M.C.S. et al. Current status of biotechnological production and applications of microbial exopolysaccharides. Crit Rev Food Sci Nutr, v.60, n.9, p.1475-95, 2019.

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
440
BECHARA, R. et al. Methodologya for the optimal design of integrated first and second generation ethanol production plant combined with power cogeneration. Bioresour Technol, v.214, p.441-49, 2016. BESSA, D.H.R.F. Prospecção de enzimas lipolíticas em fungos para a produção de biodiesel. Dissertação (Mestrado). Instituto Federal de Educação, Ciência e Tecnologia Goiano. 51fl. Rio Verde - GO, 2019. BERUDE, M.C. et al. Microrrizas e sua importância agroecológica. Encicl Biosf, v.11, n.22, p.132-46, 2015. BONONI, V.L.R. et al. Agaricomycetes (Basidiomycota, Fungi) diversity in a protected area in the Maracaju Mountains, in the brazilian central region. Hoehnea, v.44, n.3, p. 361-77, 2017. CABRAL, B.V. et al. Recycling influence in phosphorus biossolubilization from rock phosphate concentrate in air-lift bioreactor. Chem Eng Trans, v.57, p. 673-78, 2017. DABAJA, M.Z. et al. Avaliação da atividade enzimática de fungo endofítico isolado de Annona crassiflora (Marolo) com interesse biotecnológico. Rev Univ Vale do Rio Verde, v.17, n.1, p.1-10, 2019. DAVIS, R. et al. Process design and economics for the conversion of lignocellulosic biomass to hydrocarbons: dilute-acid and enzymatic desconstruction of biomass to sugars and catalytic conversion of sugars to hydrocarbons. Golden: National Renewable Energy Laboratory, 2015. DORNELAS, A.S.P. et al. Fungos filamentosos associados às espécies Atta sexdens (Linnaeus) e Atta laevigata (F. Smith) (Hymenoptera: Formicidae). EntomoBrasilis, v.9, n.1, p. 26-30, 2016. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA – EMBRAPA. Biossolubilização de potássio in vitro a partir da rocha fonolito por microrganismos do solo. Org.: SILVA, U. C. et al. Sete Lagoas: Embrapa Milho e Sorgo, 2015. (Documentos/Embrapa Milho e Sorgo 177). GOMES, B.C.C. et al. Avaliação da biossolubilização de fósforo proveniente de concentrado de rocha fosfática por fermentação sólida e líquida utilizando Trichoderma ressei. In: CONGRESSO BRASILEIRO DE ENGENHARIA QUÍMICA EM INICIAÇÃO CIENTÍFICA, 12., 2017, São Carlos. Anais... São Paulo: Blucher Chemical Engineering Proceedings, v.1, n.4, 2017. HEGAZY, M.F. et al. Microbial biotransformation as a tool for drug development based on natural products from mevalonic acid pathway: A review. J Adv Res, v.6, n.1, p. 17-33, 2015. JACINTO, H.R.; MOURA, J.B.; UTO, F.N. Avaliação da influência antrópica sobre a microbiota edáfica de cerrado no município de Anápolis, Goiás. Ipê Agron J, v.2, n.2, p. 4-17, 2018. MACHADO, W.R.C. Bioprospecção de leveduras para a produção de carotenpides microbianos. Tese (Doutorado). Universidade Estadual

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
441
Paulista. Instituto de Biociências, Letras e Ciências Exatas. 123fl. São José do Rio Preto - SP, 2018. MELO, M. et al. Cellulolytic and lipolytic fungi isolated from soil and leaf litter samples from the Cerrado (Brazilian Savanna). Rev Biol Trop, v.66, n.1, p.237-45, 2018. NORILER, S.A. et al. Bioprospecting and structure of fungal endophyte communities found in the brazilian biomes, Pantanal, and Cerrado. Front Microbiol, v.9, e1526, 2018. NYGAARD, S. et al. Reciprocal genomic evolution in the ant-fungus agricultural symbiosis. Nat Commun, v.7, n.12233, p.1-9, 2016. OLIVEIRA, A.P.A. et al. Bioprospecting of yeasts for amylase production in solid state fermentation and evaluation of the catalytic properties of enzymatic extracts. Afr J Biotechnol, v.14, n.14, p. 1215-23, 2015. PASSONE, J.M. et al. Avaliação da produção de biossurfactante empregando fungos e glicerol como substrato. In: CONGRESSO BRASILEIRO DE ENGENHARIA QUÍMICA EM INICIAÇÃO CIENTÍFICA, 11., 2015, Campinas. Anais... São Paulo: Blucher Chemical Engineering Proceedings, v.1, n.3, 2015. PIZA, A.C.M.T.; HOKKA, C.O.; SOUSA, C.P. Endophytic actinomycetes from Miconia albicans (Sw.) Triana (Melastomataceae) and evaluation of its antimicrobial activity. J Sci Res Rep, v.4, n.4, p.281-91, 2015. RAO, M.P.N.; XIAO, M.; LI, J-W. Fungal and bacterial pigments: secondary metabolites with wide applications. Front Microbiol, v.8, 1113, 2017. RESENDE, F.M. et al. Consequences of delaying actions for safeguarding ecosystem services in the Brazilian Cerrado. Biol Conserv, v.234, p.90-9, 2019. ROCHA, R.K. Solubilização in vitro de fosfato por Torulaspora globosa. Dissertação (Mestrado). Universidade Federal de São Carlos. Centro de Ciências Agrárias. 59fl. Araras - SP, 2017. ROMANO, L.H. Bioprospecção de microrganismos endofíticos isolados de Tabebuia spp. e Hymenaeae courbaril e identificação da produção de metabólitos de interesse biotecnológico. Tese (Doutorado). Universidade Federal de São Carlos. 134fl. São Carlos - SP, 2015. SALGADO, F.H.M. et al. Arbuscular mycorrhizal fungi and mycorrhizal stimulant affect dry matter and nutrient accumulation in bean and soybean plants. Pesq Agropec Trop, v.46, n.4, p.367-73, 2016. SALGADO, F.H.M. et al. Arbuscular mycorrhizal fungi and colonization stimulant in cotton and maize. Ciênc Rural, v.47, n.6, e20151535, 2017. SANTOS, A.A. et al. Addition of waste and introduction of microorganisms after 45 years of soil degradation. Rev Ciênc Agron, v.49, n.3, p.363-70, 2018.

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO CERRADO
442
SANTOS, F.R.S. et al. Production and characterization of β-glucosidase from Gongronella butleri by solid-state fermentation. Afr J Biotechnol, v.15, n.16, p.633-41, 2016. SAWYER, D. et al. Perfil do ecossistema hotspot de biodiversidade do Cerrado. Critical Ecosystem Partnership Fund, 2016. SILVA, D.B. et al. Bioprospecção de linhagens de Trichoderma sp. na degradação de resíduos lignocelulósicos de sabugo de milho. Rev Verde Agroecologia Desenvolv Sustent, v.10, n.1, p.252-58, 2015. SILVA, L.S. Biotransformação dos ácidos gálico e elágico por microorganismo do Cerrado brasileiro. Dissertação (Mestrado). Universidade Federal de Goiás. Faculdade de Farmácia. 157fl. Goiânia - GO, 2015. SILVEIRA, B.M.P. Seleção de biocatalizador com potencial para a produção de exopolissacarídeos. Dissertação (Mestrado). Universidade Federal dos Vales do Jequitinhonha e Mucuri. 136fl. Diamantina - MG, 2018. VALADARES-PEREIRA, A.A. et al. Fungal community structure as na indicator of soil agricultural management effects in the Cerrado. Rev Bras Ciênc Solo, v.41, e0160489, p.1-12, 2017. ZANARDO, A.B.R. Abundância de fungos entomopatogênicos da ordem Hypocreales e diversidade genética de Metarhizium spp. isolados de amostras de solo de áreas representativas de cinco biomas brasileiros. Tese (Doutorado). Universidade de São Paulo. Escola Superior de Agricultura “Luiz de Queiroz”. 153fl. Piracicaba - SP, 2015. ZANINETTE, F. et al. Production of inulin and neolevan-type fructooligosaccharides by Penicillum janczewskii Zaleski CCIBt 3352. Biotechnol Appl Biochem, v.66, n.3, p. 419-25, 2019. ZANONI, P.R.S. et al. Produção de etanol: uma opção para aproveitamento de resíduos gerados nas indústrias de reciclagem de papel. Colombo: Embrapa Florestas, 2015.

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
443
CAPÍTULO 21
ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E
RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
Maria Helena do NASCIMENTO 1
Fernanda Pereira SANTOS 2
Maria Vanessa Pontes da Costa ESPÍNOLA 3
Rádamis Barbosa CASTOR 1 Krystyna Gorlach LIRA 4
1 Mestrandos do Programa de Pós Graduação em Biologia Celular e Molecular, UFPB; ² Graduanda em Biotecnologia, UFPB; 3 Graduanda em Ciências biológicas, UFPB;
4Orientadora/Professora do DBM/ UFPB. [email protected]
RESUMO: Diante do surgimento de microrganismos resistentes aos compostos antimicrobianos, as plantas tornaram-se uma alternativa viável para o desenvolvimento de novos compostos biologicamente ativos, pois apresentam baixos ou nulos efeitos colaterais, além de possuírem um custo benefício melhor quando comparado aos medicamentos sintéticos. Nos últimos anos, todas as partes da Moringa oleifera (Moringaceae) têm sido amplamente exploradas para fins nutricionais, medicinais e industriais. O foco desta revisão integrativa foi a atividade antimicrobiana das folhas, sementes e raízes da M. oleifera realizada na base de dados de PubMed, Scielo, ScienceDirect, Google Scholar e Lilacs. Foi feito o levantamento de artigos científicos publicados entre 2016-2020. As pesquisas científicas apontam que extratos de folhas de M. oleifera possuem atividade antibacteriana e antifúngica frente a diversos patógenos como, Escherichia coli, Klebsiella pneumoniae, Candida albicans e Aspergillus fumigatus. Quanto aos extratos de sementes foi verificada a atividade

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
444
antimicrobiana frente as bactérias E. coli, K. pneumoniae, Pseudomonas aeruginosa, Staphylococcus aureus e Salmonella typhii. Existem poucos dados sobre a atividade antimicrobiana de raízes de moringa, as quais relataram ação de extratos de raízes contra P. aeruginosa e Erwinia carotovora e de óleo essencial contra Enterococcus faecalis e K. pneumoniae. Deste modo, os dados mencionados nesta revisão indicam grande potencial antimicrobiano de extratos e óleos essenciais da M. oleifera, constituindo novas perspectivas nas pesquisas de síntese de compostos antimicrobianos. Palavras-chave: Resistência. Produtos naturais. Compostos ativos. Fitoquímicos. Plantas medicinais.
INTRODUÇÃO
A resistência microbiana tem sido um dos maiores e mais
consideráveis obstáculos para a saúde pública a nível mundial
(SILVA; AQUINO, 2018). Fatores como o uso indiscriminado e
excessivo desses compostos, desde a utilização por indivíduos
à, principalmente, sua utilização em maior escala nos âmbitos
de saúde, agricultura e agropecuária, intensificaram ao longo
dos anos a resistência aos antimicrobianos existentes no
mercado, favorecendo o surgimento de linhagens microbianas
multirresistentes (MDR - “Multidrug-resistant”) (MARSTON et
al., 2016; KALEO et al., 2019).
Ainda que a resistência microbiana seja natural aos
microrganismos, a utilização em massa de antimicrobianos têm
direcionado o desenvolvimento de linhagens dispostas de
mecanismos de resistência contra as principais classes desses
fármacos. Nesse cenário, a rapidez e diversidade com que
microrganismos se multiplicam torna-se preocupante
(MARSTON et al., 2016; REMPEL et al., 2019).

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
445
Diante disso, a busca por novos fármacos com atividade
antimicrobiana tornou-se indispensável, e as plantas medicinais
são consideradas fontes promissoras para o desenvolvimento
dessas novas terapias (BRÍGIDO et al., 2020). Uma vez que
estudos científicos demonstram que fármacos provenientes de
plantas são farmacologicamente eficazes, possuem efeitos
colaterais baixos ou nulos, além de ampla margem de
segurança para consumo humano e animal e um custo-
benefício melhor em comparação aos medicamentos ditos
alopáticos. De forma geral, os fitoterápicos encontram-se
facilmente disponíveis, o que os tornam candidatos valiosos
para o desenvolvimento de diversos tipos de medicamentos
efetivos e seguros no tratamento de doenças (SALEEM;
SALEEM; AKHTAR, 2020).
Atualmente, estima-se que existem cerca de meio milhão
de espécies de plantas ainda não estudadas quanto aos seus
efeitos medicinais. Dentre uma das mais importantes
propriedades medicinais das plantas está a atividade
antimicrobiana (MORENO et al., 2018; KALEO et al., 2019), a
qual pode ser uma importante alternativa terapêutica quando
comparada aos medicamentos tradicionais, uma vez que a
maioria dos microrganismos patogênicos aos humanos já
desenvolveram inúmeros mecanismos de resistência (REMPEL
et al., 2019).
A Moringa é um dos gêneros de plantas mais importantes
devidos ao seu valor econômico e sua multiplicidade de usos
de folhas, flores e sementes para fins terapêuticos e outros. O
gênero compreende 13 espécies, a M. concanensis, M.
arborea, M. ovalifolia, M. borziana, M. pygmaea, M. ruspoliana,
M. rivae, M. hildebrandtii, M. drouhardii, M. stenopetala, M.

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
446
longituba, M. peregrine e a Moringa oleifera (SINGH et al.,
2019).
Também conhecida como “árvore milagrosa”,
“Nebedave”, “árvore de baqueta”, “raiz-forte” ou “árvore da
vida”, a Moringa oleifera pertence à família Moringaceae,
oriunda do sub-Himalaia, no noroeste da Índia, Afeganistão,
Bangladesh e Paquistão, porém atualmente está distribuída em
diversas regiões tropicais e subtropicais (MATIC et al., 2018;
SINGH et al., 2019). No Brasil, a moringa foi introduzida em
meados de 1950 sendo cultivada para fins medicinais e
ornamentais. Encontrada principalmente no Nordeste do Brasil,
em estados como Maranhão, Ceará e Piauí, a moringa é
conhecida como “quiabo de quina” ou “lírio-branco” (SARAIVA
et al., 2018).
A M. oleifera possui diversas características como
capacidade de adaptar-se a diferentes climas, crescendo em
ambientes quentes, úmidos ou até mesmo secos, com
precipitação anual entre 1.000 - 2.000 mm. A temperatura para
o crescimento da árvore varia entre 25 - 35 °C e pH 5,0 - 9,0,
sob luz solar e uma altitude de aproximadamente 550 metros
(RAJENDRAN; SUDESHRAJ; SURESHKUMAR, 2019; SU;
CHEN, 2020). Portanto, a moringa consegue adaptar-se a
diferentes tipos de solos, no entanto, apresenta melhor
tendência de crescimento em solos franco-arenosos levemente
alcalinos (SU; CHEN, 2020).
Caracterizada por ser uma árvore delgada, perene
com altura de aproximadamente 5 a 12 metros, com as folhas
compostas tripinadas nas pontas dos galhos, possui flores com
pétalas branco-amareladas e fragrância suave, o seu fruto é
uma cápsula trilobada possuindo morfologia delgada e longa de
vagem. As sementes secas da moringa possuem o formato

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
447
redondo ou triangulares, e cada grão é rodeado por uma casca
ligeiramente arborizada (SU; CHEN, 2020).
Cada parte da M. oleifera possui fitoquímicos específicos
que lhes oferecem variadas ações medicinais (TSHABALALA
et al., 2020). Além do uso medicinal, a moringa é muito
valorizada por possuir diversas aplicações industriais, por
exemplo na produção de cosméticos e na nutrição alimentar,
pois as folhas apresentam altas taxas proteicas, as sementes
são ricas em lipídios (especialmente ácido palmítico, ácido
esteárico e ácido oleico), e assim como as sementes, as vagens
possuem altos níveis de ferro, cálcio, potássio e sódio (MITIC
et al., 2018; YAMAGUCHI et al., 2020).
Diversos estudos foram feitos sobre a moringa, no qual
foram verificadas suas propriedades antidiabética (UMAR et
al., 2018); antiartrítica e antioxidante (SALEEM; SALEEM;
AKHTAR, 2020); diurética (TAHKUR et al., 2016); anti-
inflamatória e antitumoral (BARHOI et al., 2020).
Adicionalmente, a moringa é uma das plantas mais bem
estudadas devido a sua propriedade antimicrobiana, sendo que
extratos de todas as partes da M. oleifera, principalmente
extratos metanólicos, apresentaram forte atividade
antimicrobiana contra bactérias patogênicas como, Bacillus
cereus, Escherichia coli, Staphylococcus aureus (MILLER et
al., 2017), Streptococcus mutans, Serratia marcescens,
Staphylococcus resistente à Meticilina (MRSA), e fungos
patogênicos, Fusarium oxysporum, Aspergillus flavus,
Polysphondylium pallidum, Alternaria alternata (REHMAN et al.,
2020).
A M. oleifera tornou-se objeto de diversas pesquisas
devido à presença de substâncias biologicamente ativas com
efeitos antibacterianos e antifúngicos, os quais possuem

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
448
potencial para serem utilizados na formulação de produtos
aplicados na área de saúde. Portanto, esse trabalho busca
enfatizar a atividade antimicrobiana de M. oleifera, a partir de
um levantamento bibliográfico sobre seus compostos bioativos,
ressaltando avaliações de diferentes extratos de folhas,
sementes e raízes da planta.
MATERIAIS E MÉTODO
O presente estudo trata-se de uma revisão de literatura
realizada por meio de levantamento bibliográfico no período
entre 2016 e 2020, buscando publicações em plataformas como
PubMed, Scielo, ScienceDirect, Google Scholar e Lilacs.
Utilizou-se palavras chaves como: Moringa oleifera, compostos
bioativos, tratamento de água, seeds, leaves, compostos
químicos, antimicrobianos, antimicrobial activity, entre outros.
Para selecionar os artigos utilizados nesta revisão, inicialmente,
foram avaliados os respectivos resumos, em seguida,
considerado o fator de impacto, relevância na área e o ano de
publicação.
RESULTADOS E DISCUSSÃO
Várias partes da moringa como folhas (Figura 1A),
sementes (Figura 1B) e raízes (Figura 1C) possuem atividade
antimicrobiana frente a bactérias e fungos, pois cada parte da
planta possui fitoquímicos que lhes oferecem um amplo
espectro para atividades farmacológicas (PAIKRA et al., 2017;
TSHABALALA et al., 2020).

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
449
Figura 1: Folhas, flores, vagens (A), sementes (B) e raízes (C)
da M. oleifera.
Fonte: ANDRADE; GORLACH-LIRA, 2018 (A); PAIKRA;
DHONGADE; GIDWAHI, 2017 (B); <https://amarnatureza.org.br/raiz-da-
moringa/> (C).
As folhas de moringa possuem compostos fenólicos,
flavonoides e taninos, e acredita-se que devido a sua
composição fitoquímica apresenta uma relevante atividade
antimicrobiana frente a bactérias patogênicas, quando
comparada a outras partes da planta (IDOWU; OSENI, 2015).
As folhas apresentam também compostos como as saponinas,
que agem interferindo na permeabilidade da parede celular
bacteriana (SOPANDANI et al., 2020), carotenoides,
terpenoides, alcaloides e glucosinolatos (MITIC et al., 2018).
Entre os flavonoides encontrados na M. oleifera, os flavonoides
glicosídeos (glicosídeos, malonil glicosídeos e rutinídeos) de
quercetina > caempferol > isorhamnetina são os mais
encontrados nas folhas. Enquanto que o maior teor de

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
450
glucosinolato é encontrado em sementes e folhas (SAINI;
SIVANESAN; KEUM, 2016).
Quanto às sementes da moringa, sua atividade
antimicrobiana, geralmente está associada a presença dos
compostos moringina, piterigosperma e isotiocianato de benzila
(FAROOQ; KOUL, 2019).
Por fim, estudos sobre raízes ainda são poucos, no entanto,
extratos de raízes de moringa possuem em sua composição os
fitoquímicos: carboidratos, glicosídeos, alcaloides, saponinas,
terpenoides, esteroides, taninos e flavonoides, principal
constituinte responsável pela atividade antimicrobiana
(RAMAKRISHNA et al., 2018; TSHABALALA et al., 2019).
Os flavonoides podem formar complexos com proteínas
por meio de ligações de hidrogênio, de maneira que, a estrutura
terciária da proteína seja alterada. Isto pode ocasionar danos
ou desnaturação de ácidos nucleicos e proteínas.
Consequentemente, essa desnaturação interfere nas funções
biológicas e no metabolismo das bactérias. Ademais, os
flavonoides podem impedir a síntese da membrana celular e os
efeitos agregados em todas as células bacterianas
(SOPANDANI et al., 2020).
Os taninos são compostos que podem inibir a síntese
proteica para a formação da parede celular e encolher as
paredes celulares, cessando a permeabilidade celular e,
consequentemente, causando a morte do microrganismo
(SOPANDANI et al., 2020).
Quanto aos alcaloides, estes possuem um mecanismo
inibitório antibacteriano que age nos constituintes do
peptidoglicano nas células bacterianas, de maneira que as
camadas intactas da parede celular não são construídas,
causando a morte celular (SOPANDANI et al., 2020).

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
451
Já os compostos terpenoides podem agir em contato
com as porinas presentes na membrana externa da parede
celular bacteriana e, portanto, formar fortes ligações poliméricas
que causam danos a pilha. Isto resulta na entrada de
compostos que podem reduzir a permeabilidade da parede
celular bacteriana, de maneira que a célula bacteriana
necessite de nutrientes e o crescimento bacteriano é inibido ou
interrompido, causando a morte (SOPANDANI et al., 2020).
Extratos de folhas de M. oleifera: atividade antimicrobiana.
Dzotam e colaboradores (2016) realizaram triagem
fitoquímica em extrato metanólico de folhas de M. oleifera,
evidenciando a presença metabólitos secundários advindos da
planta, como alcaloides, esteróis, cumarinas, entre outros.
Esses metabólitos foram indicados como responsáveis pela
eficácia desse extrato contra linhagens de bactérias Gram-
negativas multirresistentes de E. coli, Enterobacter aerogenes,
K. pneumoniae, Pseudomonas aeruginosa e Providencia
stuartli. A ação sinérgica entre antibióticos existentes e o extrato
produzido contra as linhagens patogênicas também esteve em
evidência nesse estudo.
Reforçando esses resultados, Matic e colaboradores (2018),
relataram que extratos de folhas de M. oleifera contém diversos
compostos bioativos, principalmente taninos, flavonoides e
saponinas, indicando uma atividade antimicrobiana frente a
microrganismos patogênicos.
Botero et al. (2019) indicaram as folhas de M. oleifera
como valiosa fonte de peptídeos antimicrobianos. Extratos
etanólicos de folhas de M. oleifera apresentaram, em diferentes
concentrações, eficácia antimicrobiana frente a bactérias

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
452
patógenas transmitidas através de alimentos como P.
aeruginosa, S. aureus e Candida albicans.
Dalukdeniya e colaboradores (2016) avaliaram a
atividade de folhas de M. oleifera frente a patógenos
microbianos de origem alimentar, S. enteritica, V.
parahaemolyticus, E. coli e L. monocytogenes, no qual os
resíduos obtidos após a extração aquosa foram utilizados para
a extração com o clorofórmio. Como resultado, foi demonstrada
a maior atividade antibacteriana contra a S.enteritica (superior
a 20 mm), no entanto a L. monocytogenes demonstrou a maior
resistência ao extrato (halo entre 10-15 mm).
Elgamily e colaboradores (2016), reforçaram a
propriedade antimicrobiana da folha da moringa empregando
como método a fissão em disco, utilizando extratos obtidos a
partir da extração com etanol, acetona e acetato de etila. Como
resultado, os autores observaram inibição do crescimento de S.
aureus, S. mutans e C. albicans. Ressalta-se que os extratos
etanólicos apresentaram maior halo de inibição contra os três
microrganismos patogênicos.
Corroborando com os resultados, o estudo de Yee (2019)
demonstrou que diferentes extratos de folhas de M. oleifera
extraídas com acetato de etila (EtOAc), etanol (EtOH), e
metanol (MeOH) foram eficazes contra linhagens bacterianas
de Bacillus subtilis, S. aureus, B. pumilus; e fúngicas de
Candida albicans. No entanto, não foi observado nenhuma
atividade dos extratos acima citados, frente a P. aeruginosa.
Adicionalmente, extratos aquosos não demonstraram nenhuma
atividade antimicrobiana frente aos microrganismos
patogênicos.
Os experimentos realizados por Chakraborty e
colaboradores (2019) indicaram a eficácia de extratos de folhas

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
453
de M. oleifera contra E. coli (19,6 mm), S. typhimurium (18,55
mm), S. aureus (13,80 mm) e Aeromonas hydrophila (8,35 mm),
apontando o potencial dessa planta como agente
antimicrobiano a ser utilizado pela indústria alimentícia, uma
vez que a partir deste estudo a moringa provou ser um
conservante natural promissor devido a inibição contra
bactérias Gram-positivas e Gram-negativas.
Em linha de pesquisa semelhante, Tirado-Torres et al.
(2019) demonstraram a eficácia de extratos aquosos e
etanólicos de folhas de M. oleifera frente a vinte isolados de S.
aureus multirresistentes a 12 antibióticos. Todas as cepas
foram sensíveis aos extratos, sendo que os extratos etanólicos
exibiram uma maior atividade antimicrobiana em relação aos
extratos aquosos e ao antibiótico controle, penicilina.
Extratos de folhas de M. oleifera preparados com auxílio
dos solventes metanol, hexano, acetato de etila e água,
demonstraram eficácia contra linhagens patogênicas de
bactérias, E. coli, S. aureus, P. aeruginosa, S. typhi e B. subtillis
e fúngicas, Saccharomyces cerevisiae e A. fusarium, as quais
frequentemente são associadas a infecções transmitidas por
alimentos, ou identificadas como contaminantes de ambientes
industriais e hospitalares (JANVIER et al., 2018).
Em um estudo recente, Adadallah e Muhammad (2019)
avaliaram a atividade antibacteriana do extrato aquoso das
folhas de M. oleifera, em diferentes concentrações, frente a
cinco microrganismos, E. coli, S. typhi, Shigella spp., S. aureus
e E. faecalis. Os resultados demonstraram que todos os
isolados foram inibidos pelo extrato aquoso, ressalta-se que a
zona média de inibição foi de 8,20 mm, com maior halo de
inibição obtido frente a Shigella spp. (11,09 ± 0,26 mm).

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
454
Oladeji, Odelade e Oloke (2019) investigaram a atividade
antimicrobiana de extratos etanólicos, clorofórmico e acetato de
etila de folhas de M. oleifera frente as bactérias P. aeruginosa,
Klebsiella sp., E. coli, S. pneumoniae e B. cereus e os fungos
A. ninger, A. flavus, Trichoderma sp. e Candida sp.
Todos os extratos demonstraram atividade contra as cepas
testadas e foi observado que o extrato etanólico apresentou
maior inibição do crescimento dos microrganismos quando
comparado aos extratos clorofórmico e acetato de etila. Os
autores sugerem que atividade antimicrobiana dos extratos
analisados ocorrem devido aos metabólitos secundários
presentes nas folhas da moringa, como as saponinas, taninos,
alcaloides, terpenoides, flavonoides, entre outros.
Extratos de sementes de M. oleifera: atividade
antimicrobiana.
Sharma, Wichaphon e Klangpetch (2020) investigaram os
efeitos antimicrobianos do extrato etanólico de farinha de
sementes de M. oleifera, obtido por extração assistida por
ultrassom, contra as bactérias E. coli, B. cereus, S. typhimurium
e S. aureus a partir dos testes de difusão em placa de ágar e de
microdiluição. Nos testes de difusão, os maiores valores das
zonas de inibição foram observadas no caso de B. cereus
(19,75 ± 0,3 mm) e S. aureus (19,25 ± 0,9 mm), enquanto que
nos testes de microdiluição as melhores concentrações
inibitórias mínimas (MIC) foram de 3,13 mg/ml contra E. coli e
B. cereus, respectivamente.
Os estudos de Andrade e Gorlach-Lira (2018) mostraram
que o extrato aquoso das sementes apresentou capacidade
inibitória do crescimento de S. aureus, enquanto o extrato

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
455
hidroalcoólico inibiu S. aureus, S. typhimurium e V. cholerae,
utilizando o teste de difusão no meio sólido. Entretanto, as
cepas de P. aeruginosa, E. coli e E. faecalis foram resistentes
aos dois extratos.
Fernandes e colaboradores (2020) investigaram os perfis
fitoquímicos, nutricionais e as atividades antimicrobianas dos
extratos aquosos e hidroetanólicos das sementes de M. oleifera
contra as bactérias Gram-negativas E. coli, S. typhimurium e E.
cloacae, Gram-positivas S. aureus, B. cereus e L.
monocytogenes e dos fungos A. fumigatus, A. ochraceus, A.
niger, Penicillium funiculosum, P. ochrochloron e P.
aurantiogriseum. Os valores de MIC e de concentração
bactericida mínima (MBC) ou de concentração fungicida mínima
(MFC) foram estabelecidos a partir do método de microdiluição
e, no geral, o melhor valor de MIC e de MBC/MFC encontrados
contra todas as espécies foi de 0,037 mg/mL e de 0,075 mg/mL,
respectivamente, mostrando o potencial antimicrobiano deste
extrato (FERNANDES et al., 2020).
Brilhante e colaboradores (2015) estudaram a eficácia dos
extratos de sementes de M. oleifera contra espécies de Vibrio
patogênicas provenientes do camarão Macrobrachium
amazonicum. Os extratos etanólicos apresentaram baixa
eficácia, aferida a partir do método de microdiluição, na inibição
do crescimento de V. cholerae, V. vulnificus e V. mimicus.
O estudo de Patel e Mohan (2018) demonstrou a
atividade antimicrobiana de extratos de acetato de etila de
sementes da M. oleifera, frente as bactérias Gram-positivas e
Gram-negativas Bacillus cereus, B. subtilis, E. coli, E. faecalis,
K. pneumoniae, Micrococcus luteus, P. aeruginosa, S. aureus,
S. epidermidis, S. marcescens, Salmonella paratyphi e S. typhi,
e o fungo Trichoderma harzanium. Os autores sugerem que os

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
456
flavonoides presentes nas sementes foram responsáveis pela
ação antimicrobiana.
Além dos diferentes extratos de sementes da moringa
possuírem atividade antimicrobiana, o óleo obtido de sementes
também apresenta atividade antagonista frente a
microrganismos patogênicos. Bichi e Shehu (2018) avaliaram
que o óleo de sementes em quatro concentrações (100%, 75%,
50% e 25%) apresentou potencial de inibição contra E. coli.
Extratos de raízes de M. oleifera: atividade antimicrobiana.
Abdulkadir e colaboradores (2015), avaliaram a atividade
antimicrobiana de extrato etanólico de raízes de moringa pelo
método de difusão em ágar e de microdiluição, e observaram
ação antimicrobiana contra os patógenos E. coli, S. aureus e C.
albicans.
Extratos das raízes de M. oleifera obtidos com diversos
solventes (acetona, metanol, etanol, acetato de etila, água)
apresentaram significativa ação antimicrobiana contra P.
aeruginosa e Erwinia carotovora. Os extratos de metanol e
acetato, na concentração de 100 mg/mL, apresentaram maior
inibição contra ambas as cepas (PRABAKARAN et al., 2018).
Os experimentos de Tshabalala e colaboradores (2020)
indicaram a atividade antimicrobiana dos extratos aquosos,
etanólicos e acetônicos de raízes da M. oleifera contra B.
subtilis, E. coli e S. aureus e C. albicans.
Dalukdeniya e colaboradores (2016) indicaram atividade
de extratos aquosos, alcoólicos e clorofórmicos das raízes de
moringa contra S. enteritica, V. parahaemolyticus, E. coli e L.
monocytogenes, as quais são frequentemente relacionados a
doenças adquiridas a partir de alimentos.

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
457
Estudo realizado por Ramakrishna e colaboradores
(2018) corrobora com os resultados já descritos, em que
extratos etanólicos de raízes de moringa demonstraram
atividade antimicrobiana frente aos patógenos E. coli (halo de
35 mm) e S. aureus (halo de 15 mm), demonstrando ser um
potencial fitoterápico contra bactérias.
Adeleye e colaboradores (2018), estudaram a atividade
antifúngica de extratos metanólicos, etanólicos e extratos com
água quente e fria de raízes de M. oleifera, frente ao fungo
patogênico C. albicans. Os autores observaram que os extratos
metanólicos apresentaram maior atividade antimicrobiana,
enquanto os extratos obtidos com água fria foram menos
eficazes.
A atividade antimicrobiana dos extratos metanólicos e
aquosos de cascas de raízes da moringa também foi testada
frente E. coli e S. aureus. No entanto, os melhores resultados
foram obtidos a partir da atividade dos extratos aquosos
empregando diferentes concentrações (AMANZE et al., 2020).
Óleos essenciais extraídos a partir de raízes de M. oleifera
demonstraram atividade antimicrobiana contra linhagens Gram-
positivas (Enterococcus faecalis) e Gram-negativas (K.
pneumoniae, Proteus vulgaris e Shigella flexineri),
demonstrando que a presença de compostos bioativos são
evidências de seus potenciais efeitos farmacológicos
(OTUNOLA; AFOLAYAN, 2018).
CONCLUSÃO
A partir desta revisão integrativa é possível concluir que
os extratos de folhas, sementes e raízes da M. oleifera
revelaram-se promissores para aplicação na área de saúde

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
458
como fontes de compostos biologicamente ativos contra
microrganismos resistentes aos antimicrobianos. M. oleifera
apresenta uma significativa atividade antagonista frente a
diversos patógenos, tanto bactérias como fungos, destacando
S. aureus resistente à meticilina (MRSA).
REFERÊNCIAS BIBLIOGRÁFICAS ABADALLAH, M. S.; ALI, M. Antibacterial activity of Moringa oleifera leaf extracts against bacteria isolated from patients attending General Sani Abacha Specialist Hospital Damaturu. Journal of Allied Pharmaceutical Sciences, p. 61-66, 2019. ABDULKADIR, I. S. et al. Phytochemical screening and antimicrobial activities of ethanolic extracts of Moringa oleifera Lam on isolates of some pathogens. Journal of Applied Pharmacy, v. 7, n. 4, p. 2-7, 2015. ADELEYE, S. A. et al. Phytochemistry and antifungal activity of root and seed extracts of Moringa oleifera. International Journal of Advanced Research in Biological Sciences, v. 5, n. 3, p. 169-176, 2018. AMANZE, E. K. et al. Antibacterial activity of Moringa oleifera root bark extract against some pathogenic organisms. Asian Journal of Immunology, v. 4, n. 3 p. 21-27, 2020. AMAR A NATUREZA. Raiz da moringa. Disponível em: https://amarnatureza.org.br/raiz-da-moringa/. Acesso em: 30 out. 2020. ANDRADE, T.M.; GORLACH-LIRA, K. Antibacterial activity of the white lily Moringa oleifera seed extract and its use in water treatment. Brazilian Journal of Biological Sciences, v. 5, p. 699-707, 2018. BARHOI, D. et al. Aqueous extract of Moringa oleifera exhibit potential anticancer activity and can be used as a possible cancer therapeutic agent: a study involving in vitro and in vivo approach. Journal of the American College of Nutrition, p. 1-16, 2020. BIHI, M.H.; SHEHU, U. H. Antimicrobial properties of Moringa oleifera seed oil. International Journal of Research in Earth & Environmental Sciences, v. 13, n. 1, 2018. BOTERO, K., et al. Morinpeptides: in silico identification of antimicrobial peptides and biological activity from Moringa oleifera leaves. Pharmacology Online, v.2, p.199-211, 2019. BRILHANTE, R. et al. Vibrio spp. from Macrobrachium amazonicum prawn farming are inhibited by Moringa oleifera extracts. Asian Pacific Journal of Tropical Medicine, v. 8, n. 11, p. 919-922, 2015.

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
459
BRÍGIDO, H. P. C. et al. Atividade antimicrobiana de Aspidosperma nitidum benth (Apocynaceae). Revista Eletrônica Acervo Saúde, v. 12, n. 10, p. e4123-e4123, 2020. CHAKRABORTY, K. et al. Antimicrobial activity of Moringa oleifera leaf extract in Pangasianodon hypophthalmus minced meat. Environment and Ecology, v. 37, n. 3, p. 812-816, 2019. DALUKDENIYA, D.A.C.K.; DE SILVA, K.L.S.R.; RATHNAYAKA, R.M.U.S.K. Antimicrobial activity of different extracts of leaves bark and roots of Moringa oleifera (Lam). International Journal of Current Microbiology and Applied Sciences, v. 5, n. 7, p. 687-691, 2016. DZOTAM, J. K.; TOUANI, F. K.; KUETE, V. Antibacterial and antibiotic-modifying activities of three food plants (Xanthosoma mafaffa Lam., Moringa oleifera (L.) Schott and Passiflora edulis Sims) against multidrug-resistant (MDR) Gram-negative bacteria. BMC Complementary and Alternative Medicine, v. 16, n. 1, p. 9, 2016. ELGAMILY, H. et al. Microbiological assessment of Moringa oleifera extracts and its incorporation in novel dental remedies against some oral pathogens. Open access Macedonian Journal of Medical Sciences, v. 4, n. 4, p. 585, 2016. FERNANDES, A. et al. Nutritional and phytochemical profiles and biological activities of Moringa oleifera Lam. edible parts from Guinea-Bissau (West Africa). Food Chemistry, v. 341, p. 128229-128267, 2020. FAROOQ, B.; KOUL, B. Comparative analysis of the antioxidant, antibacterial and plant growth promoting potential of five Indian varieties of Moringa oleifera L. South African Journal of Botany, v. 129, p. 47-55, 2020. IDOWU, K.; OSENI, O. A. Compositional investigation of phytochemical and antioxidant properties of various parts of Moringa oleifera plant. European Journal of Basic and Applied Sciences, v. 2, n. 2, p. 1-10, 2015. JANVIER, H. et al. Antimicrobial and antifungal activity of three selected homegrown vegetables consumed in Rwanda. European Journal of Medicinal Plants, v. 23, n. 1, p. 1-8, 2018. KALEO, I. V. et al. Effects of Moringa oleifera leaf extract on growth performance, physiological and immune response, and related immune gene expression of Macrobrachium rosenbergii with Vibrio anguillarum and ammonia stress. Fish & Shellfish Immunology, v. 89, p. 603-613, 2019. MARSTON, H. D. et al. Antimicrobial resistance. Jama, v. 316, n. 11, p. 1193-1204, 2016. MILLER, G. J. et al. A laboratory activity demonstrating the antibacterial effects of extracts from two plant species, Moringa oleifera and Allium sativum (garlic). Journal of Microbiology & Biology Education, v. 18, n. 3, 2017.

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
460
MATIC, I. et al. Investigation of medicinal plants traditionally used as dietary supplements: A review on Moringa oleifera. Journal of Public Health in Africa, v. 9, n. 3, 2018. MORENO, A. H., et al. Avaliação da atividade antimicrobiana e citotoxicidade hemolítica em diferentes extratos vegetais. Arquivos de Ciências da Saúde, v. 25, n. 1, p. 11-12, 2018. OLADEJI, O. S.; ODELADE, K. A.; OLOKE, J. K. Phytochemical screening and antimicrobial investigation of Moringa oleifera leaf extracts. African Journal of Science, Technology, Innovation and Development, v. 12, n. 1, p. 79-84, 2020. OTUNOLA, G. A.; AFOLAYAN, A. J. Chemical composition, antibacterial and in vitro anti-inflammatory potentials of essential oils from different plant parts of Moringa oleifera Lam. American Journal of Biochemistry and Biotechnology, v. 14, n. 3, p. 210-220, 2018. PATEL, N.; MOHAN, J. S. S. Antimicrobial activity and phytochemical analysis of Moringa oleifera Lam. crude extracts against selected bacterial and fungal strains. International Journal of Pharmacognosy and Phytochemical Research, v. 10, n. 2, p. 68-72, 2018. PAIKRA, B. K., DHONGADE, H. K. J., GIDWANI, B. Phytochemistry and pharmacology of Moringa oleifera Lam. Journal of Pharmacopuncture, v. 20, n. 3, p. 194, 2017. PRABAKARAN, M. et al. Polyphenol composition and antimicrobial activity of various solvent extracts from different plant parts of Moringa oleifera. Food Bioscience, v. 26, p. 23-29, 2018. RAJENDRAN, A.; SUDESHRAJ, R.; SURESHKUMAR, S. Phytonutrients: Moringa oleifera leaf extracts an incredible health super food supplement. The Pharma Innovation Journal, v. 8, n. 2, p. 29-33, 2019. RAMAKRISHNA, C. et al., Antimicrobial, phytochemical and quantitative HPLC analysis of Moringa oleifera root. Innoriginal International Journal of Scienses, v. 5, n. 6, 2018. REHMAN, K. et al. Competence of benzoil tree (Moringa oleifera L.) as antibacterial and antifungal agent. Advancements in Life Sciences – International Quarterly Journal of Biological Sciences, v. 7, n. 3, p. 135-139, 2020. REMPEL, C. et al. Efeito antimicrobiano de plantas medicinais: uma revisão de estudos científicos. Revista Ibero-Americana de Ciências Ambientais, v. 10, n. 4, p. 57-82, 2019. SAINI, R. K.; SIVANESAN, I.; KEUM, Y.S. Phytochemicals of Moringa oleifera: a review of their nutritional, therapeutic and industrial significance. 3 Biotech, v. 6, n. 2, p. 203, 2016. SALEEM, A.; SALEEM, M.; AKHTAR, M. F. Antioxidant, anti-inflammatory and antiarthritic potential of Moringa oleifera Lam: an ethnomedicinal plant

ATIVIDADE ANTIMICROBIANA DE EXTRATOS DE FOLHAS, SEMENTES E RAÍZES DE Moringa oleifera LAM. (Moringaceae): UMA REVISÃO
461
of Moringaceae family. South African Journal of Botany, v. 128, p. 246-256, 2020. SARAIVA, L. C. F. et al. Triagem fitoquímica das folhas de Moringa oleifera. Boletim Informativo Geum, v. 9, n. 2, p. 12, 2018. SHARMA, P.; WICHAPHON, J.; KLANGPETCH, W. Antimicrobial and antioxidant activities of defatted Moringa oleifera seed meal extract obtained by ultrasound-assisted extraction and application as a natural antimicrobial coating for raw chicken sausages. International Journal of Food Microbiology, v. 332, p. 108770-108811, 2020. SILVA, M. O.; AQUINO, S. Resistência aos antimicrobianos: uma revisão dos desafios na busca por novas alternativas de tratamento. Revista de Epidemiologia e Controle de Infecção, v. 8, n. 4, p. 472-482, 2018. SOPANDANI, P. et al. Antibacterial effects of Moringa oleifera leaf extract against Enterococcus faecalis in vitro. Scientific Dental Journal, v. 4, n. 1, p. 16, 2020. SU, B.; CHEN, X. Current status and potential of Moringa oleifera leaf as an alternative protein source for animal feeds. Frontiers in Veterinary Science, v. 7, 2020. TAHKUR, R. S. et al. Diuretic activity of Moringa oleifera leaves extract in swiss albino rats. The Pharma Innovation, v. 5, n. 3, Part A, p. 8, 2016. TIRADO-TORRES, D. et al. Antimicrobial Activity of Moringa oleifera Against multidrug-resistant Staphylococcus aureus isolated from raw milk. Applied Ecology and Enviromental Research, v. 17, n. 1, p. 587-599, 2019. TSHABALALA, T. et al. Potential substitution of the root with the leaf in the use of Moringa oleifera for antimicrobial, antidiabetic and antioxidant properties. South African Journal of Botany, v. 129, p. 106-112, 2020. UMAR, S. A. et al. Evaluation of hypoglycaemic and antioxidant activity of Moringa oleifera root in normal and alloxan-induced diabetic rats. Tropical Journal of Natural Product Research, v. 2, n. 8, p. 401-408, 2018. YAMAGUCHI, N. U. et al. A review of Moringa oleifera seeds in water treatment: Trends and future challenges. Process Safety and Environmental Protection, v. 147, p. 405-420, 2020. YEE, M. M. A comparative studies on antimicrobial activity and antioxidant activity on different extracts of leaf, bark and root of Moringa oleifera Lamk (Drumstick tree). International Journal of Recent Innovations in Academic Research, v. 3, n. 7, p. 24-34. 2019.

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
462
CAPÍTULO 22
BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA
EXPERIMENTAL
Raquel Bezerra da SILVA 1
Maria Isabela Ferreira ARAÚJO 2
Paula Perazzo de Souza BARBOSA 3
1 Graduanda do curso de Ciências Biológicas, UFPB; 2 Doutoranda em Ciências Biológicas, UFPE; 3 Doutora em Ciência e Tecnologia de Alimentos, UFPB.
RESUMO: O presente trabalho trata-se de uma breve revisão comparativa sobre o SARS-CoV, o MERS-CoV e o SARS-CoV-2 e algumas pesquisas com plasma convalescente, que tem sido usado no tratamento das doenças provocadas por esses vírus. Para tanto, foi feita uma busca de trabalhos científicos nas bases de dados eletrônicas utilizando palavras-chave específicas para o tema, e selecionadas publicações sobre o estudo com esses coronavírus entre 2015 a 2020. Os SARS-CoV, o MERS-CoV e, possivelmente, o SARS-CoV-2 originaram-se de morcegos, mas possuem hospedeiros intermediários diferentes, estando o deste último ainda sob investigação. Os coronavírus possuem genoma de RNA de fita simples, com comprimento entre 25 e 32 kb; e contêm pelo menos quatro proteínas estruturais: Spike (S), a de membrana (M), a do envelope (E); e a do nucleocapsídeo (N), que participam dos processos de infecção, montagem e liberação das partículas virais da célula hospedeira, mas cujas sequências primárias variam entre eles. O plasma convalescente é uma das primeiras opções de tratamento para

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
463
doenças infecciosas emergentes na ausência de uma terapia otimizada. Porém, não há evidências clínicas bem fundamentadas para que seu uso possa ser amplamente recomendado. As experiências com esses três coronavírus devem assistir as projeções de experimentos futuros e orientar os esforços de prevenção para reduzir a transmissão e para conter surtos futuros de doenças zoonóticas. Palavras-chaves: Betacoronavírus. Pandemia. COVID-19.
INTRODUÇÃO
Em dezembro de 2019 foi notificada em Wuhan, na
China, uma nova infecção respiratória viral, semelhante às
provocadas pelos agentes causadores da síndrome respiratória
aguda grave (Severe Acute Respiratory Syndrome – SARS) e
da síndrome respiratória do Oriente Médio (Middle East
Respiratory Syndrome – MERS) (SANDERS et al, 2020).
O novo patógeno, nomeado de Severe Acute Respiratory
Syndrome Coronavirus 2 (SARS-CoV-2), rapidamente se
disseminou, tornando-se o terceiro tipo de coronavírus que
causa manifestações clínicas mais graves em humanos a surgir
na população; e a doença por ele provocada,
Coronavirus Disease 2019 (COVID-19), teve seu caráter de
pandemia declarado em 11 de março de 2020 (WHO, 2020).
Outros coronavírus como os Human Coronavirus 229E,
NL63, OC43 e HKU1 (HCOV-229E, HCOV-NL63, HCOV-OC43
e HCOV-HKU1, respectivamente) provocam infecções
respiratórias autolimitadas, mas podem tornar-se graves em
bebês, pacientes imunossuprimidos e idosos (LIU et al., 2020).
O surgimento dos surtos e/ou reinfecções provocados
por coronavírus está relacionado a sua capacidade de mutação

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
464
e/ou recombinação, característica que os auxilia a se tornarem
patogênicos para humanos. Assim, uma vez que essas
mudanças no perfil genético são comuns por fazerem parte do
processo evolutivo viral, torna-se essencial a busca por
terapêuticas eficazes para o combate dos próprios (RABAAN et
al., 2020).
Embora as pessoas tenham como medidas preventivas
a higienização, a quarentena, o isolamento e o distanciamento
social para evitar a propagação da COVID-19, ainda em curso,
um antiviral específico para uma intervenção otimizada ainda
não foi encontrado, o que coloca a virose em estado de
urgência, exigindo investigações mais aprofundadas (BADEN;
RUBIN, 2020).
Para o SARS-CoV-2, têm sido consideradas terapias que
tiveram atividade confirmada contra o Severe Acute Respiratory
Syndrome Coronavirus (SARS-CoV) e o Middle East
Respiratory Syndrome Coronavirus (MERS-CoV), por serem
seus parentes próximos (KO et al., 2020), como é o caso do
plasma convalescente, normalmente usado como opção
terapêutica na ausência de um tratamento específico quando
do surgimento de uma doença infecciosa.
Nesse contexto, o presente trabalho objetivou fazer uma
breve revisão comparativa sobre o SARS-CoV, o MERS-CoV e
o SARS-CoV-2, e alguns dos avanços das pesquisas com
plasma convalescente, que vem sendo usado na terapia
experimental da COVID-19. Dessa forma, pretende-se auxiliar
na troca de informações sobre o tema, conduzindo o
desenvolvimento de novas perspectivas de aprendizado e
impulsionar futuras investigações clínicas.

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
465
MATERIAIS E MÉTODO
Para realizar a presente revisão bibliográfica, foi feita
uma busca de trabalhos científicos nos bancos de dados
Google Scholar, Medline (PubMed), ScienceDirect, Scopus e
Scielo através dos descritores, isolados ou em combinação:
“terapia medicamentosa”, “plasma convalescente”,
“coronavírus”, “pandemia”, “síndrome respitatória aguda grave”,
“síndrome respitatória do Oriente Médio”, “doença do novo
coronavírus”, “relações evolutivas entre os vírus”, e seus
correspondentes na língua inglesa, além das siglas quando se
aplicavam.
Para o refinamento do material foram considerados os
trabalhos na forma de artigo científico e capítulos de livro que
atendessem aos critérios: (i) ano de publicação – de 2015 a
2020; (ii) abordagem experimental do plasma convalescente
para o tratamento da MERS ou SARS ou COVID-19; e (iii)
características gerais e filogenia dos três coronavírus em
questão. Os resultados compreenderam uma narrativa
compendiada das informações fornecidas pelos trabalhos
selecionados segundo os parâmetros mencionados.
RESULTADOS E DISCUSSÃO
MERS-CoV, SARS-CoV e SARS-CoV-2: CONSIDERAÇÕES
GERAIS
Apesar de alguns coronavírus já serem conhecidos como
as principais causas de morbidade e mortalidade em animais,
sua importância obteve destaque apenas após o surgimento da

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
466
SARS, no ano de 2002, em Hong Kong, provocada pelo SARS-
CoV (FANI; TEIMOOR; GHAFARI, 2020). Em 2012, na Arábia
Saudita, Oriente Médio, foi identificado o MERS-CoV (de WIT
et al., 2016). Ambos infectam o trato respiratório inferior,
podendo causar síndrome respiratória aguda frequentemente
fatal, assim como o SARS-CoV-2 (KHAILANY; SAFDAR;
OZASLAN, 2020). Os três vírus levantaram grande
preocupação global e tornaram-se problemas de saúde
emergentes em todo o mundo em seus respectivos períodos
(MEO et al., 2020), como mostrado na tabela 1.
Tabela 1. Comparação entre SARS-CoV, MERS-CoV e SARS-
CoV-2 com relação ao ano de identificação, local da primeira
notificação, número de países atingidos e situação atual.
Vírus Ano de
notificação
Local da
primeira
notificação
Número
de
países
atingidos
Situação atual
SARS-CoV 2002 Hong Kong
(China) 26
Sem casos
relatados
desde 2004
MERS-CoV 2012 Arábia Saudita
(Oriente médio) 27
Redução de
casos desde
2016, mas
com riscos
significantes
no Oriente
Médio
SARS-CoV-2 2019 Wuhan
(China) 214 Em curso
Fontes: FANI; TEIMOOR; GHAFARI, 2020; de WIT et al., 2020; NHI, 2020.

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
467
O SARS-CoV, o MERS-CoV e o SARS-CoV-2 pertencem
ao gênero Betacoronavirus, da família Coronaviridae, ordem
Nidovirales (PAYNE 2017). Os primeiros dados sequenciados
de genoma mostraram que o SARS-CoV-2 é membro do gênero
em questão e se enquadra no subgênero Sarbecovírus,
juntamente com o SARS-CoV. O MERS-CoV pertence ao
subgênero Merbecovírus (ZHANG; HOLMES, 2020).
A família Coronaviridae é constituída por grandes vírus
envelopados e com genoma de RNA de fita simples, cujo
comprimento varia de 25 a 32 kb e possui estrutura de cap 5′ e
cauda poli-A 3′. Os vírions desse grupo são aproximadamente
esféricos e possuem diâmetro de 118 a 136 nm (PAYNE 2017).
O motivo de a família ser assim denominada é a presença das
moléculas de proteína Spike (S) na superfície viral, o que dão
aos vírions um formato de coroa (ABDELRAHMAN; LI; WANG,
2020).
Coronavírus que usam animais como reservatórios
naturais são conhecidos desde o final de 1930. Por milhares de
anos, esses agentes vêm cruzando as barreiras das espécies e
alguns emergiram como importantes patógenos humanos.
Evidências filogenéticas mostraram que morcegos e roedores
servem como fonte de genes da maioria dos betacoronavírus
(YE et al., 2020), no entanto, outros animais também podem
servir de reservatório.
Estudos revelaram que tanto o SARS-CoV quanto o
MERS-CoV têm origem zoonótica. Outros relataram a
possibilidade de o SARS-CoV-2 também ter procedência
animal, proposta baseada na semelhança de sua sequência
genética com a de outros coronavírus conhecidos. Assim como
o SARS-CoV e o MERS-CoV, o SARS-CoV-2 pode ter sido

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
468
transmitido a humanos por um hospedeiro animal intermediário
(LI et al., 2020).
Comparações genéticas entre os vírus humanos e os
que infectam outros animais fundamenta os estudos sobre a
origem desses patógenos. As sequências genômicas de
coronavírus de pangolins (pangolin-CoV) de Guangdong, na
China, foram mais estreitamente relacionadas ao SARS-CoV-2,
mas as evidências também apontam ratos e algália de bambu,
da família Rhizomyidae, como seus prováveis hospedeiros
intermediários, mas nenhuma prova absoluta foi encontrada
(VINODHKUMAR et al., 2020).
Outro estudo analisou a dinâmica de transmissão e
história evolutiva do SARS-CoV-2 comparando os genomas de
cepas de diferentes países com o pangolin-CoV de duas
amostras de metagenoma de pangolin e com vírus do tipo
SARS de morcego, focando as sequências de aminoácidos das
respectivas proteínas S. Os autores verificaram que o vírus das
duas amostras de pangolins eram filogeneticamente
relacionados o SARS-CoV-2, mas este foi mais semelhante ao
vírus do tipo SARS de morcego (LI et al., 2020).
Outra análise mostrou que o SARS-CoV-2 pertence,
juntamente com o SARS-CoV e o vírus do tipo SARS de
morcego, a um clado diferente do MERS-CoV, sendo
filogeneticamente mais relacionado àqueles isolados de
morcegos ferradura, na China, entre 2015 e 2018, do que ao
SARS-CoV, o que apóia a hipótese de que a cadeia de
transmissão começou a partir do morcego e atingiu o ser
humano (BENVENUTO et al., 2020).
Uma comparação da origem zoonótica, dos hospedeiros
intermediários, do tamanho dos genomas juntamente com o

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
469
receptor celular em humanos de cada coronavírus aqui
discutido é apresentada na tabela 2.
Tabela 2. Origem zoonótica, hospedeiro intermediário, tamanho
do genoma e receptor celular humano de SARS-CoV, MERS-
CoV e SARS-CoV-2. ECA-2: Enzima Conversora de
Angiotensina 2; CD26: Enzima Dipeptidil peptidase 4.
*Provavelmente.
Vírus Hospedeiro
primário
Hospedeiro
intermediário
Tamanho do
Genoma (kb)
Receptor
principal em
humanos
SARS-CoV Morcego
Civeta de
palma
mascarada
29,75 kb ECA-2
MERS-CoV Morcego Dromedário 30,11 kb CD26
SARS-CoV-2 Morcego* Desconhecido 29,8 – 29,9 kb ECA-2
Fontes: Zhu et al., 2020; Yuan et al., 2015; Khailany; Safdar; Ozaslan, 2020;
Wang et al., 2020.
O SARS-CoV, o MERS-CoV e, possivelmente, o SARS-
CoV-2 originaram-se de morcegos, sendo que para o primeiro,
a civeta de palma mascarada constitui o hospedeiro
intermediário que desencadeou o transbordamento para os
humanos, e os camelos dromedários, a ponte para o MERS-
CoV (LAUXMANN; SANTUCCI; AUTRAN-GÓMEZ, 2020). O
interventor do SARS-CoV-2 ainda permanece desconhecido.
O SARS-CoV-2 compartilha aproximadamente 79,5% de
homologia genômica com o SARS-CoV, enquanto apenas
cerca de 50% de similaridade com o MERS-CoV, indicando
proximidade maior com o primeiro (ZHU et al., 2020).
Sanders et al. (2020) encontraram 98 áreas de
sobreposição de regiões altamente estruturadas ao comparar

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
470
os genomas de SARS-CoV e SARS-CoV-2. Dentro dessas
regiões conservadas na região de codificação do genoma de
SARS-CoV-2, foram identificadas 65 com estruturas
secundárias semelhantes, estando a maioria, 26, dentro da
Open Reading Frames 1b (Fase de leitura aberta – ORF1b).
Todos os coronavírus contêm pelo menos quatro
proteínas estruturais, que são essenciais para desencadear a
infecção viral e promover a montagem do vírion (CHEN; LIU;
GUO, 2020), sendo três delas associadas à membrana: Spike
(S), necessária à ligação na células-alvo e à fusão da
membrana celular do vírus durante a infecção; as proteínas de
membrana (M) e do envelope (E); e uma do nucleocapsídeo (N)
(WU et al., 2020).
A proteína M tem três domínios transmembrana, dá
forma aos vírions, promove a curvatura da membrana e se liga
ao nucleocapsídeo. A proteína E desempenha um papel na
montagem, na liberação do vírus da célula e na patogênese
viral. A proteína N contém dois domínios, os quais podem ligar-
se ao RNA genômico do vírus (CHEN; LIU; GUO, 2020).
A ligação do RNA a N estabiliza o nucleocapsídeo e o
núcleo interno dos vírions, promovendo a conclusão da
montagem e o brotamento viral (PAYNE et al., 2017). Uma
comparação entre as massas moleculares das proteínas
estruturais de MERS-CoV, SARS-CoV e SARS-CoV-2 é
apresentada na tabela 3.

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
471
Tabela 3. Comparação do tamanho em Dálton (Da) das
proteínas Spike, de Membrana, de Nucleocapsídeo e de
Envelope de SARS-CoV, MERS-CoV e SARS-CoV-2 e seus
respectivos códigos de entrada na base de dados UniProtKB.
Vírus
Tamanho da
proteína S
(Da)
Tamanho da
proteína M
(Da)
Tamanho da
proteína N
(Da)
Tamanho da
proteína E
(Da)
SARS-CoV 139,125 Da
(P59594)
25,061 Da
(P59596)
45,626 Da
(P59595)
8,361 Da
(P59637)
SARS-CoV-2
141,178 Da
(P0DTC2)
25,147 Da
(P0DTC5)
46,025 Da
(P0DTC9)
8,365 Da
(P0DTC4)
MERS-CoV 149,384 Da
(K9N5Q8)
24,551 Da
(T2BB40 )
45,048 Da
(R9UM87)
9,354Da
(K9N5R3)
Fonte: UniProt Knowledgebase (UniProtKB).
A proteína S dos coronavírus compreende duas
subunidades: S1, que contém o domínio de ligação ao receptor
(RBD), e S2, que auxilia na fusão da membrana viral com a da
célula-alvo (WU et al., 2020), sendo ambas unidas de forma não
covalente. A ligação com o receptor induz mudanças
conformacionais na proteína S, expondo os locais de clivagem
das duas subunidades (TRIGUEIRO-LOURO et al., 2020),
processo este que ativa a proteína de fusão. O resultado é uma
entrada complexa nas células suscetíveis, que requer ação
combinada de ligação ao receptor e proteólise da proteína S
para promover a fusão vírus-célula (WALL et al., 2020).
Em MERS-CoV, SARS-CoV e SARS-CoV-2, a clivagem
proteolítica de iniciação é realizado pela Protease Serina
Transmembrana humana 2 (TMPRSS2), embora que para o
MERS-CoVs uma pré-clivagem é necessária para a ativação

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
472
subsequente da proteína S pela TMPRSS2, o que é realizada
por furina nas células infectadas (PARK et al., 2016).
A sequência de aminoácidos da proteína S em SARS-
CoV-2 é 76,47% idêntica à do SARS-CoV, com a mesma
confirmação estrutural e propriedades eletrostáticas na
interface de interação (XU et al., 2020). Esse valor é promovido
por cerca de 380 subsitituições de aminoácidos entre as duas,
que se traduz em diferenças em cinco dos seis aminoácidos
vitais do RBD com o ECA-2 humano expresso na superfície
(GHEBLAWI et al., 2020).
A afinidade de ligação da proteína S à ECA-2 é
comparável ou ainda mais forte do que no SARS-CoV. Isso
pode explicar o rápido desenvolvimento e forte capacidade de
transmissão humano-humano em COVID-19 (XIE; CHEN,
2020).
As alterações em vários resíduos de aminoácidos
permitem aumento das interações hidrofóbicas e formações de
pontes salinas, o que pode explicar a influência global
consideravelmente maior do COVID-19 do que o SARS
(GHEBLAWI et al., 2020).
A proteína S é o principal alvo de anticorpos
neutralizantes durante a infecção viral, bem como o foco da
projeção de vacinas (WANG et al., 2020). Porém, nenhum
medicamento ou vacina específica ainda foram aprovados para
o tratamento desses coronavírus humanos (WU et al., 2020),
mas a sobreposição genética entre eles (LI et al., 2020) fez com
que as pesquisas estejam reaproveitando para COVID-19
terapias antivirais usadas no cenário das infecções por SARS-
CoV e MERS-CoV, como é o caso do plasma convalescente.

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
473
A TERAPIA DE PLASMA CONVALESCENTE E O SEU USO
PARA O TRATAMENTO DA SARS
Plasma convalescente é um tipo de imunoterapia passiva
baseada em plasma ou derivados obtidos de doadores
recuperados de infecções graves (GARRAUD, 2017), que são
transfundidos em pacientes recém-infectados como a primeira
opção terapêutica na ausência de um tratamento específico
disponível para a doença em questão (KO et al., 2018).
O princípio da infusão de plasma convalescente foi
estabelecido em 1880, ao ser demonstrada a presença de
anticorpos contra difteria no sangue de animais imunizados com
doses não letais de toxinas, e que poderiam ser transferidos
para animais que sofriam de infecções ativas (ROJAS et al.,
2020).
Os produtos do plasma fornecem imunidade passiva,
neutralizando o patógeno (MARANO et al., 2016) ou ativando o
sistema complemento, a citotoxicidade celular dependente de
anticorpo e/ou fagocitose, erradicando eventualmente o
patógeno da circulação sanguínea do paciente (BLOCH et al.,
2020).
Dessa forma, a administração de plasma convalescente
no período de janela imunológica, antes que um paciente
desenvolva seu próprio anticorpo, pode fornecer esses
produtos prontos para medeiar a atividade antiviral e aumentar
o desenvolvimento da resposta imune (CASADEVALL;
JOYNER; PIROFSKI, 2020).
A imunoterapia passiva foi aprimorada e a tecnologia
molecular desenvolveu e estabeleceu o uso de anticorpos
monoclonais direcionados contra fatores envolvidos na

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
474
patogênese de doenças (WALKER; BURTON, 2018),
anticorpos policlonais, imunoglobulinas intravenosas, entre
outros, para tratar condições infecciosas. Porém, em situações
emergenciais, a obtenção desses produtos é mais difícil,
principalmente por conta dos custos, o que faz com que o
plasma convalescente seja a opção imediata (ROJAS et al.,
2020).
O plasma convalescente foi usado pela primeira para
tratar a gripe espanhola, em 1918, tendo sido aproveitado em
surtos de SARS (BROWN, McCULLOUGH, 2020; CAO, SHI,
2020). O procedimento era aplicado como último recurso para
melhorar a taxa de sobrevivência de pacientes com SARS cuja
condição continuava a piorar apesar de outros tratamentos
administrados. Além disso, os estudos admitem abreviação do
tempo de internação e redução da mortalidade em pacientes
tratados com plasma convalescente em comparação com os
que não foram (CHEN et al., 2020).
No entanto, a eficácia em relação à SARS é motivo de
controvérsias. A maioria das pesquisa que testaram a terapia
admite limitações como: falta de grupos-controle (AL-TAWFIQ;
ARABI, 2020), número pequeno de pacientes na amostra e falta
de padronização dos títulos de anticorpos no plasma
convalescente dos doadores (SULLIVAN; ROBACK, 2020).
PLASMA CONVALESCENTE NA TERAPIA DA MERS
A literatura também relata o uso de plasma
convalescente no tratamento de MERS, mas assim como para
SARS, os trabalhos relacionados são escassos e
inconcludentes com relação a sua eficácia e segurança.

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
475
Arabi et al (2016) fizeram um estudo sobre a viabilidade
de se conduzir um ensaio clínico randomizado de terapia de
plasma convalescente obtido de pacientes previamente
recuperados de MERS. Os autores estabeleceram um protocolo
para o seu uso nessa situação e que consistia de duas fases: a
primeira para coleta do plasma convalescente e análise dos
títulos de anticorpos anti-MERS-CoV; e a segunda, o
tratamento dos pacientes.
No entanto, ao executarem a primeira fase planejada,
Alrashid et al (2017) relataram baixa soroprevalência de
anticorpos MERS-CoV entre coortes de pacientes com infecção
suspeita ou confirmada. Foram testados para anticorpos anti-
MERS 170 doadores na Arábia Saudita com idades entre 18-32
anos e nenhum foi positivo.
Os autores acham que os níveis de anticorpos têm uma
rápida redução durante a recuperação dos pacientes, chegando
a desaparecer após cerca de 8 meses. Dessa forma, neste
estudo, através dos resultados foi verificado que, para infecção
por MERS-CoV, o plasma obtido dos doadores saudáveis não
tem anticorpos suficientes que permitam o seu uso como
tratamento.
Em contrapartida, Ko et al (2018) testaram o efeito do
plasma convalescente de quatro doadores recuperados de
MERS em três pacientes coreanos e foi constatada atividade
neutralizante no soro, apesar de ter sido em apenas dois. Um
deles teve piora na oxigenação 2 h depois de receber a terapia
e desenvolveu lesão pulmonar aguda relacionada à transfusão
(transfusion-related acute lung injury – TRALI), mas cinco dias
depois seu soro apresentou atividade neutralizante e teve alta
com 102 dias após o início da doença. O outro paciente recebeu

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
476
infusão de plasma convalescente no 14º dia da doença, quando
teve piora na oxigeniação, e no dia seguinte seu soro
apresentou atividade neutralizante, recebendo alta no 39º dia
da MERS.
Os dois indivíduos receberam infusão de plasma
convalescente com atividade de neutralização de um título
PRNT de 1:80 (verificada através do plaque reduction
neutralization test – PRNT, teste de neutralização por redução
de placas de lise). O que não teve atividade recebeu um plasma
convalescente com título PRNT de 1:20. Os autores concluíram
que, para uma infusão eficaz na infecção por MERS-CoV, o
plasma do doador deve ser testado quanto à atividade de
anticorpos e a atividade de neutralização de título PRNT ≥ 1:80
pode ser necessária.
PLASMA CONVALESCENTE NA TERAPIA PARA COVID-19
A terapia de plasma convalescente direcionada a
pacientes da COVID-19, pandemia ainda em curso, é de amplo
interesse, pois, teoricamente, imunoglobulinas anti-SARS-CoV-
2 neutralizantes existem no plasma dos pacientes recuperados
(SEGHATCHIANA; LANZA, 2020).
YE et al., 2020 usaram plasma convalescente em seis
pacientes com COVID-19 em curso tardio. Os resultados
apontaram que eles tiveram os sintomas aliviados, alterações
nas anormalidades radiológicas e nos exames laboratoriais, e
não houve efeito adverso óbvio durante o tratamento. O vírus
foi eliminado em cinco pacientes que apresentaram SARS-CoV-
2 no swab da garganta, e houve aumento imediato nos títulos
de anticorpos anti-SARS-CoV-2 em dois dos pacientes. Os

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
477
autores atribuem ao plasma os benefícios clínicos em todos os
seis casos, provavelmente devido aos anticorpos anti-SARS-
CoV-2 IgM e IgG que neutralizam diretamente o vírus, além do
conteúdo anti-inflamatório que pode ter prevenido a liberação
de citocinas.
SHEN et al., 2020 descreveram a melhora do estado
clínico de cinco pacientes (duas mulheres e três homens, de 36
a 76 anos) com COVID-19 grave após a transfusão de plasma
convalescente de cinco doadores entre 18 e 60 anos que
haviam se recuperado da infecção por SARS-CoV-2. Os
pacientes transfundidos tiveram a temperatura corporal
normalizada e redução da carga viral, além de desmame da
ventilação mecânica.
Em outro estudo, com 189 pacientes de COVID-19,
Alboghasemi et al. (2020) administraram em 115 deles,
tratamento com plasma convalescente, e em 74, terapia
antiviral de rotina semelhante, incluindo lopinavir/ritonavir,
hidroxicloroquina e agente anti-inflamatório. Os autores
concluíram que o plasma convalescente deve ser considerada
como uma terapia segura e eficaz para pacientes com COVID-
19, pois melhorou a sobrevida dos pacientes e reduziu
significativamente o período de hospitalização e as
necessidades de intubação em comparação com o grupo de
controle.
Os dois primeiros estudos aqui descritos com plasma
convalescente direcionado ao tratamento de COVID-19 têm
limitações: (i) o número da amostra, que é pequeno; (ii) a falta
de controles; (iii) a administração de antivirais nos pacientes
durante a transfusão, não sendo possível determinar se a
melhora observada poderia estar relacionada a outras terapias

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
478
do que plasma convalescente ou se ele potencializou a ação
dos antivirais; (iv) o período em que a transfusão de plasma foi
realizada: 10 a 22 dias após a admissão dos pacientes nos dois
casos, não podendo ser conclusivo se a melhora pode ser
resultante também do curso natural do organismo.
No terceiro estudo, embora tenha sido executado com
uma amostra maior e com grupo-controle, o plasma
convalescente foi administrado em menos de três dias da
admissão hospitalar, o que não deixa claro se seria o momento
ideal. Os pacientes que receberam a terapia experimental
também receberam terapia antiviral de rotina, incluindo
lopinavir/ritonavir, hidroxicloroquina e agente anti-inflamatório,
também sendo inconclusivo se o plasma potencializou a ação
dos antivirais.
Questões éticas também impõem limitações às
pesquisas relacionadas à terapêutica experimental em qualquer
situação. Segundo Alboghasemi et al. (2020), em seu trabalho,
houve relutância em privar os pacientes da terapia, o que fez o
grupo-controle ser formado de pacientes com apresentação
clínica leve e não compatíveis de plasma convalescente do
grupo sanguíneo, estando em menor número que no grupo de
tratamento; além de ser invitável neste a administração dos
medicamentos antivirais
Para eficácia da terapia, a administração do plasma
convalescente em pacientes, seja de COVID-19 ou de
quaisquer outras enfermidades, os estudos devem ser bem
projetados e considerar possíveis reações adversas como
TRALI, sobrecarga circulatória associada à transfusão
(Transfusion Associated Circulatory Overload – TACO),
reações alérgicas à transfusão, transmissão de doenças

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
479
infecciosas e riscos associados a leucócitos e aloimunização de
glóbulos vermelhos (PANDEY; VYAS, 2020).
O suporte laboratorial para a realização de testes
sorológicos e moleculares, que incluem medição da
neutralização viral e resposta imune devem ser considerados,
assim como o alinhamento governamental e institucional que
permitam as instituições de saúde e a indústria de plasma
seguirem estratégias perfeitas (LANGHI-JÚNIOR; DE SANTIS;
BORDIN, 2020).
Como se pode perceber, embora seu conceito esteja
bem estabelecido enquanto terapia, os ensaios clínicos com
plasma convalescente em larga escala, randomizados e bem
projetados são escassos, o que faz com que o próprio ainda
seja considerado empírico (CAO, SHI, 2020), estando as
questões éticas envolvidas com essas restirções.
CONCLUSÕES
O SARS-CoV-2 resultou em uma pandemia de maior
proporção do que as provocadas pelo SARS-CoV e pelo MERS-
CoV, atingido mais regiões do mundo. Apesar da origem dos
dois primeiros estarem estabelecidas, a do SARS-CoV-2 ainda
permanece obscura. Conhecer sua procedência é um passo
importante para prevenir possíveis surtos futuros semelhantes,
bem como a transmissão de doenças zoonóticas aos humanos,
tema este que também deve ser prioridade em pesquisa.
O combate com plasma convalescente aos coronavírus
que ainda configuram ameaça para humanidade ainda é algo
desafiador, pois existem consideráveis obstáculos, entre eles
as questões éticas em relação aos cuidados fornecidos para os

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
480
pacientes, a serem superados para que o procedimento possa
ser recomendado como opção terapêutica.
A capacidade de mutação subsidia o surgimento de
novas linhagens de coronavírus, e a falta de evidências clínicas
para fundamentar as intervenções medicativas, tendo em vista
que nenhum antiviral está disponível para tratar as viroses
ainda em andamento, caso da MERS e da COVID-19, torna
clara a necessidade de desenvolvimento de protocolos a serem
usados em ensaios clínicos randomizados.
As experiências com a SARS, a MERS e, agora, com a
COVID-19 devem assistir as projeções de experimentos futuros
e orientar os esforços de prevenção para reduzir a transmissão
e conter surtos posteriores de doenças zoonóticas.
REFERÊNCIAS BIBLIOGRÁFICAS ABDELRAHMAN, Z.; LI, M.; WANG, X. Comparative Review of SARS-CoV-2, SARS-CoV, MERS-CoV, and Influenza A Respiratory Viruses. Frontiers in Immunology, v. 11, p. 2309, 2020. ABOLGHASEMI, H. et al. Clinical efficacy of convalescent plasma for treatment of COVID-19 infections: Results of a multicenter clinical study. Transfusion and Apheresis Science, p. 102875, 2020. ALRASHID, M. et al. Prevalence of antibodies against the Middle East Respiratory Syndrome coronavirus, influenza A and B viruses among blood donors, Saudi Arabia. Annals of Thoracic Medicine, v. 12, n. 3, 2017. AL-TAWFIQ, J. A.; ARABI, Y. Convalescent plasma therapy for coronavirus infection: experience from MERS and application in COVID-19. Human vaccines & immunotherapeutics, p. 1-7, 2020. ARABI, Y. et al. Feasibility, safety, clinical, and laboratory effects of convalescent plasma therapy for patients with Middle East respiratory syndrome coronavirus infection: a study protocol. Springerplus, v. 4, n. 1, p. 1-8, 2015. BADEN, L. R.; RUBIN, E. J. Covid-19 - the search for effective therapy. 2020. BENVENUTO, D. et al. The 2019‐new coronavirus epidemic: evidence for virus evolution. Journal of medical virology, v. 92, n. 4, p. 455-459, 2020.

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
481
BESTLE, D. et al. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life science alliance, v. 3, n. 9, 2020. BLOCH, E. M. et al. Deployment of convalescent plasma for the prevention and treatment of COVID-19. The Journal of clinical investigation, v. 130, n. 6, p. 2757-2765, 2020. CANRONG, W. et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharmaceutica Sinica B, 2020. CAO, H.; SHI, Y. Convalescent plasma: possible therapy for novel coronavirus disease 2019. Transfusion, v. 60, n. 5, p. 1078, 2020. CASADEVALL, A. et al. SARS-CoV-2 viral load and antibody responses: the case for convalescent plasma therapy. The Journal of clinical investigation, v. 130, n. 10, 2020. CHUNYAN, W. et al. A human monoclonal antibody blocking SARS-CoV-2 infection. Nature communications, v. 11, n. 1, p. 1-6, 2020. DE WIT, E. et al. SARS and MERS: recent insights into emerging coronaviruses. Nature Reviews Microbiology, v. 14, n. 8, p. 523, 2016. FANI, M.; TEIMOORI, Ali; GHAFARI, Shokouh. Comparison of the COVID-2019 (SARS-CoV-2) pathogenesis with SARS-CoV and MERS-CoV infections. Future Virology, n. 0, 2020. GARRAUD, O. Use of convalescent plasma in Ebola virus infection. Transfusion and apheresis science, v. 56, n. 1, p. 31-34, 2017. GHEBLAWI, M. et al. Angiotensin-converting enzyme 2: SARS-CoV-2 receptor and regulator of the renin-angiotensin system: celebrating the 20th anniversary of the discovery of ACE2. Circulation research, v. 126, n. 10, p. 1456-1474, 2020. KHAILANY, R. A.; SAFDAR, M.; OZASLAN, M. Genomic characterization of a novel SARS-CoV-2. Gene reports, p. 100682, 2020. KO, J. et al. Challenges of convalescent plasma infusion therapy in Middle East respiratory coronavirus infection: a single centre experience. Antiviral Therapy, v. 23, n. 7, p. 617-22, 2018. LAUXMANN, M. A.; SANTUCCI, N. E.; AUTRÁN-GÓMEZ, A. M. The SARS-CoV-2 Coronavirus and the COVID-19 Outbreak. International Brazilian Journal of Urology, v. 46, p. 6-18, 2020. LI, X. et al. Evolutionary history, potential intermediate animal host, and cross‐species analyses of SARS‐CoV‐2. Journal of medical virology, v. 92, n. 6, p. 602-611, 2020. LIU, J. et al. A comparative overview of COVID-19, MERS and SARS. International Journal of Surgery, v. 81, p. 1-8, 2020.RABAAN et al., 2020

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
482
MARANO, G. et al. Convalescent plasma: new evidence for an old therapeutic tool?. Blood Transfusion, v. 14, n. 2, p. 152, 2016. MEO, S. A. et al. Novel coronavirus 2019-nCoV: prevalence, biological and clinical characteristics comparison with SARS-CoV and MERS-CoV. European Review for Medical and Pharmacological Sciences, v. 24, n. 4, p. 2012-2019, 2020. NATIONAL INSTITUTE OF ALLERGY AND INFECTIOUS DISEASES. COVID-19, MERS & SARS [2020]. Disponível em: https://www.niaid.nih.gov/diseases-conditions/covid-19. Acesso em: 11 out. 2020. PANDEY, A. et al. Potential therapeutic targets for combating SARS-CoV-2: Drug repurposing, clinical trials and recent advancements. Life Sciences, p. 117883, 2020. PARK, J. et al. Proteolytic processing of Middle East respiratory syndrome coronavirus spikes expands virus tropism. Proceedings of the National Academy of Sciences, v. 113, n. 43, p. 12262-12267, 2016. PAYNE, S. Family Coronaviridae. Viruses, p. 149, 2017. ROJAS, M. et al. Convalescent plasma in Covid-19: Possible mechanisms of action. Autoimmunity Reviews, p. 102554, 2020. SANDERS, W. et al. Comparative analysis of coronavirus genomic RNA structure reveals conservation in SARS-like coronaviruses. BioRxiv, 2020. SEGHATCHIAN, J.; LANZA, F. Convalescent plasma, an apheresis research project targeting and motivating the fully recovered COVID 19 patients: A rousing message of clinical benefit to both donors and recipients alike. Transfusion and Apheresis Science, v. 59, n. 3, 2020. SHEN, C. et al. Treatment of 5 critically ill patients with COVID-19 with convalescent plasma. Jama, v. 323, n. 16, p. 1582-1589, 2020. TRIGUEIRO-LOURO, J. et al.. Unlocking COVID therapeutic targets: A structure-based rationale against SARS-CoV-2, SARS-CoV and MERS-CoV Spike. Computational and structural biotechnology journal, v. 18, p. 2117-2131, 2020. UniProt Knowledgebase. Disponível em: https://www.uniprot.org/. Acesso em 11 out. 2020. VINODHKUMAR, O. R.. et al. SARS-CoV-2 (COVID-19): Zoonotic Origin and Susceptibility of Domestic and Wild Animals. Journal of Pure and Applied Microbiology, v. 14, 2020. WALKER, L. M.; BURTON, D. R. Passive immunotherapy of viral infections:'super-antibodies' enter the fray. Nature Reviews Immunology, v. 18, n. 5, p. 297, 2018. WALLS, A. et al. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell, 2020. WHO. WHO Director-General's opening remarks at the media briefing on COVID-19 - 11 March 2020. 2020. Disponível em:

BREVE REVISÃO SOBRE O SARS-COV, O MERS-COV E O SARS-COV-2 E O USO DO PLASMA CONVALESCENTE NA TERAPIA EXPERIMENTAL
483
https://www.who.int/dg/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020. Acesso em: 11 out. 2020. WONG, G. et al. Zoonotic origins of human coronavirus 2019 (HCoV-19/SARS-CoV-2): why is this work important?. Zoological research, v. 41, n. 3, p. 213, 2020. XIE, M.; CHEN, Q. Insight into 2019 novel coronavirus—an updated intrim review and lessons from SARS-CoV and MERS-CoV. International Journal of Infectious Diseases, 2020. XU, P. et al. Arbidol/IFN-α2b therapy for patients with corona virus disease 2019: a retrospective multicenter cohort study. Microbes and infection, v. 22, n. 4-5, p. 200-205, 2020. XUEJIAO, C.; JUNZHANG, T.; GUANMING, L.; GUOWEI, L. Convalescent plasma as a potential therapy for COVID-19. The Lancet Infectious Diseases, v. 20, n. 4, p. 398-400, 2020. YE, M. et al. Treatment with convalescent plasma for COVID‐19 patients in Wuhan, China. Journal of Medical Virology, 2020. YUAN, Y. et al. Molecular basis of binding between Middle East Respiratory Syndrome Coronavirus and CD26 from seven bat species. Journal of Virology, v. 94, n. 5, 2020. ZHANG, Y.; HOLMES, E. C. A genomic perspective on the origin and emergence of SARS-CoV-2. Cell, 2020. ZHIXING, Z. et al. From SARS and MERS to COVID-19: a brief summary and comparison of severe acute respiratory infections caused by three highly pathogenic human coronaviruses. Respiratory research, v. 21, n. 1, p. 1-14, 2020.

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
484
CAPÍTULO 23
ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
Juliane Nancy de Oliveira SILVA 1
Caio César da Silva GUEDES 2
Suellen Pedrosa da SILVA 3 Anderson Felipe Soares de FREITAS 4
Robson Raion de Vasconcelos ALVES 5
1 Mestranda em Bioquímica e Fisiologia, UFPE; 2 Doutorando em Ciências Biológicas, UFPE; 3 Doutoranda em Ciências Biológicas, UFPE; 4 Mestrando em Ciências Biológicas, UFPE; 5
Doutorando em Ciências Biológicas, UFPE [email protected]
RESUMO: A Aloe vera é uma planta conhecida pelo seu grande uso na medicina popular e apresenta diversas atividades biológicas que podem estar relacionadas com os compostos biologicamente ativos encontrados nas folhas, raiz e gel. Com o aumento na busca por compostos capazes de diminuir o crescimento de microrganismos e inibir a formação de cepas resistentes é de suma importância estudos que demostrem diferentes mecanismos e aplicações na indústria farmacêutica e alimentícia. Com isso, o objetivo deste trabalho foi analisar estudos dos últimos cinco anos sobre a atividade antimicrobiana com a utilização da A. vera em suas preparações/formulações. A partir das bases de dados do Science Direct e Google Acadêmico, foram identificados trabalhos que abordaram o tema proposto visando a elaboração desse capítulo de livro. Preparações dos diferentes tecidos vegetais de A. vera apresentaram atividade contra várias espécies bacterianas, a exemplo de Escherichia coli, Pseudomonas aeruginosa e Staphylococcus aureus. Também foi visto que essas preparações também apresentaram ação contra fungos, como Penicillium digitatum, Candida glabrata e Aspergillus niger. Além disso, o gel da A. vera foi bastante

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
485
utilizado na formação de um filme comestível, que pode ser empregado para o melhoramento dos alimentos e eliminação dos microrganismos, aumentando a vida de prateleira dos alimentos. Sendo assim, a A. vera pode ser utilizada como um agente com grande potencial antimicrobiano. Palavras-chave: Antibacteriano. Antifúngico. Sinergismo.
INTRODUÇÃO
O gênero Aloe possui mais de 400 espécies. De acordo
com as regras internacionais de nomenclatura botânica, o nome
científico da “Aloe vera” é Aloe barbadensis Mill. e Aloe vera (L.)
Burm. F., sendo esses dois nomes sinônimos. Ela é uma planta
pertencente à família das Aloeaceae, sendo uma planta perene,
possuindo resistência à seca. As suas folhas são suculentas e
formam uma roseta em sua haste. Esta planta sobrevive a uma
variedade de climas, principalmente em regiões temperadas e
subtropicais, porém em temperaturas baixas ela não consegue
sobreviver. A denominação “aloe” deriva do grego alóe, do
árabe alloeh e do hebraico halal, apresentando o mesmo
significado nos três casos, que corresponde à “substância
amarga e brilhante”, enquanto “vera” significa verdadeira. São
nativas do norte da África, habitam os desertos e estepes
africanas e adotam a forma de cacto (SÁNCHEZ-MACHADO et
al., 2017).
A A. vera foi bastante utilizada pelos povos do Antigo
Egito, principalmente pela rainha Cleópatra que a utilizava para
fins estéticos, tais como massagear a pele e na forma de
máscara de beleza. Apesar de ter um nome de origem arábica,
a A. vera, por intermédio dos indígenas, foi disseminada para
outras localidades como os Estados Unidos e o Brasil
(SANTOS, 2019).

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
486
As folhas da A. vera são constituídas principalmente por
água, e o restante dos compostos incluem carboidratos,
antraquinonas, compostos fenólicos, proteínas, lipídios, ácidos
orgânicos e outros. Muitos dos compostos isolados já foram
reportados como possuindo atividades anticâncer, antioxidante,
antimicrobiana e anti-inflamatória (BARUAH et al., 2016).
Devido ao aumento da resistência microbiana aos
antibióticos existentes, há uma busca por novos fármacos que
possam aumentar a gama de drogas destinadas aos
microrganismos ou que possam diminuir essa resistência
(MORRISON; ZEMBOWER, 2020).
As limitações desses medicamentos estabelecem a
necessidade de desenvolver novos agentes antimicrobianos,
particularmente, com mecanismos de ação inovadores. Um dos
artifícios que está se destacando com o propósito de contornar
essas limitações é o uso de inibidores de fatores de virulência
do patógeno, uma vez que estes fatores causam vários danos
ao hospedeiro, aumentando o poder de infecção destes
microrganismos (SANTOS, 2019).
Devido a sua grande quantidade de compostos com
atividades biológicas, a A. vera é utilizada em diversas áreas da
pesquisa, como por exemplo, na indústria alimentícia, onde a
planta fornece um revestimento comestível nos alimentos,
livrando-os de microrganismos e aumentando a vida útil do
alimento. Além disso, ela também é utilizada na área da
medicina, onde é aplicada de diferentes formas em cada órgão
ou sistema, como na pele, na cavidade oral, no sistema
gastrointestinal, entre outras aplicações (AKABERI et al., 2016;
GIANNAKOUDAKIS et al., 2018).

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
487
Com isso, o objetivo deste trabalho foi analisar estudos
dos últimos cinco anos sobre a atividade antimicrobiana com a
utilização da A. vera em suas preparações/formulações.
MATERIAIS E MÉTODO
Para a realização deste trabalho, foi utilizada abordagem
qualitativa com enfoque bibliográfico e cunho descritivo, que
tem uma função integradora e facilitadora possibilitando
agregar conhecimentos aos leitores, dando uma maior
compreensão sobre o assunto identificado por autores
presentes no arcabouço da literatura científica.
A busca eletrônica da literatura foi realizada em bases de
dados de referência, nas seguintes fontes de pesquisa: Science
Direct e Google Acadêmico onde foram incluídos manuais e
cadernos de Órgãos de Referência. Os descritores utilizados
para levantamento da pesquisa foram: Atividade
antimicrobiana, atividade antibacteriana, atividade antifúngica,
indústria de alimentos, sinergismo, estudo in vivo e Aloe vera
com o indicador booleano AND.
Os critérios de seleção dos artigos e periódicos foram:
estudos realizados entre o ano de 2015 a 2020, nos idiomas
português, inglês ou espanhol, pesquisas on-line disponíveis na
íntegra, por atender a margem de período de publicação. Foram
excluídos da pesquisa: artigos que não abordassem a temática
relevante para o alcance do objetivo, estudos incompletos, que
não estivessem disponíveis eletronicamente e escritos fora do
período delimitado. A busca dos artigos se deu entre os meses
de junho a outubro de 2020.
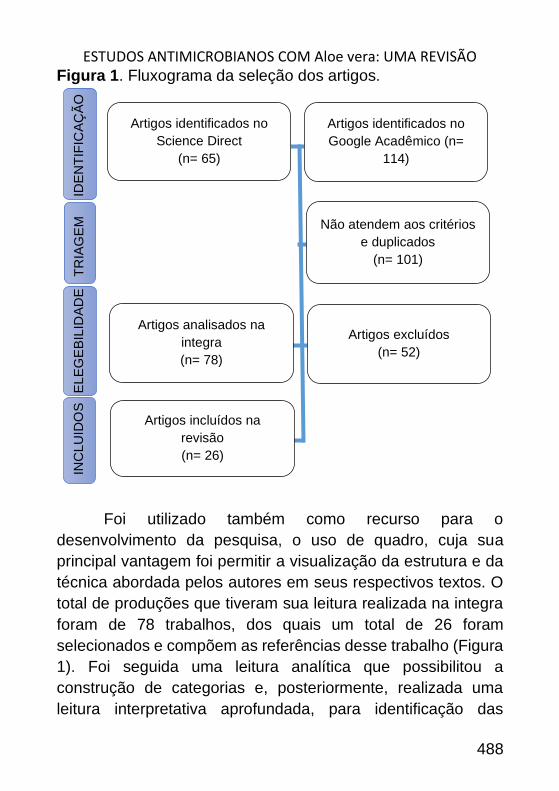
ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
488
Figura 1. Fluxograma da seleção dos artigos.
Foi utilizado também como recurso para o
desenvolvimento da pesquisa, o uso de quadro, cuja sua
principal vantagem foi permitir a visualização da estrutura e da
técnica abordada pelos autores em seus respectivos textos. O
total de produções que tiveram sua leitura realizada na integra
foram de 78 trabalhos, dos quais um total de 26 foram
selecionados e compõem as referências desse trabalho (Figura
1). Foi seguida uma leitura analítica que possibilitou a
construção de categorias e, posteriormente, realizada uma
leitura interpretativa aprofundada, para identificação das
IDE
NT
IFIC
AÇ
ÃO
T
RIA
GE
M
EL
EG
EB
ILID
AD
E
INC
LU
IDO
S
Artigos identificados no
Science Direct
(n= 65)
Artigos analisados na
integra
(n= 78)
Artigos identificados no
Google Acadêmico (n=
114)
Não atendem aos critérios
e duplicados
(n= 101)
Artigos incluídos na
revisão
(n= 26)
Artigos excluídos
(n= 52)
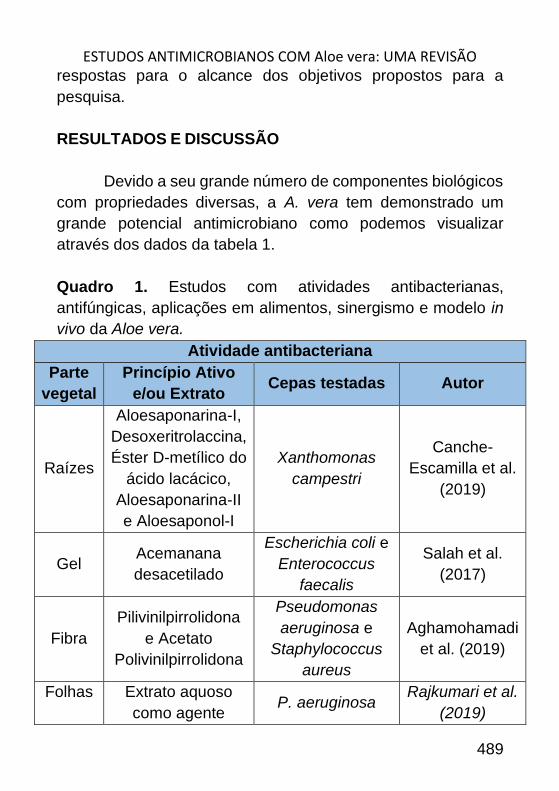
ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
489
respostas para o alcance dos objetivos propostos para a
pesquisa.
RESULTADOS E DISCUSSÃO
Devido a seu grande número de componentes biológicos
com propriedades diversas, a A. vera tem demonstrado um
grande potencial antimicrobiano como podemos visualizar
através dos dados da tabela 1.
Quadro 1. Estudos com atividades antibacterianas,
antifúngicas, aplicações em alimentos, sinergismo e modelo in
vivo da Aloe vera.
Atividade antibacteriana
Parte
vegetal
Princípio Ativo
e/ou Extrato Cepas testadas Autor
Raízes
Aloesaponarina-I,
Desoxeritrolaccina,
Éster D-metílico do
ácido lacácico,
Aloesaponarina-II
e Aloesaponol-I
Xanthomonas
campestri
Canche-
Escamilla et al.
(2019)
Gel Acemanana
desacetilado
Escherichia coli e
Enterococcus
faecalis
Salah et al.
(2017)
Fibra
Pilivinilpirrolidona
e Acetato
Polivinilpirrolidona
Pseudomonas
aeruginosa e
Staphylococcus
aureus
Aghamohamadi
et al. (2019)
Folhas
Extrato aquoso
como agente P. aeruginosa
Rajkumari et al.
(2019)
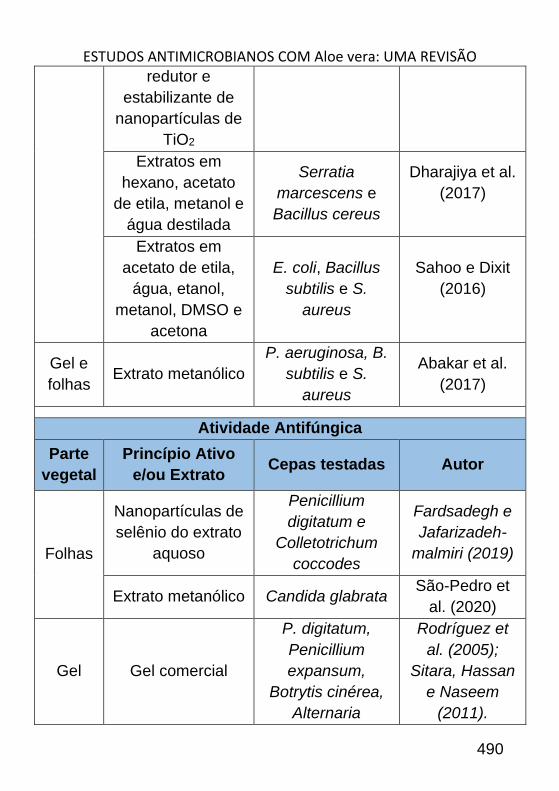
ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
490
redutor e
estabilizante de
nanopartículas de
TiO2
Extratos em
hexano, acetato
de etila, metanol e
água destilada
Serratia
marcescens e
Bacillus cereus
Dharajiya et al.
(2017)
Extratos em
acetato de etila,
água, etanol,
metanol, DMSO e
acetona
E. coli, Bacillus
subtilis e S.
aureus
Sahoo e Dixit
(2016)
Gel e
folhas Extrato metanólico
P. aeruginosa, B.
subtilis e S.
aureus
Abakar et al.
(2017)
Atividade Antifúngica
Parte
vegetal
Princípio Ativo
e/ou Extrato Cepas testadas Autor
Folhas
Nanopartículas de
selênio do extrato
aquoso
Penicillium
digitatum e
Colletotrichum
coccodes
Fardsadegh e
Jafarizadeh-
malmiri (2019)
Extrato metanólico Candida glabrata São-Pedro et
al. (2020)
Gel Gel comercial
P. digitatum,
Penicillium
expansum,
Botrytis cinérea,
Alternaria
Rodríguez et
al. (2005);
Sitara, Hassan
e Naseem
(2011).
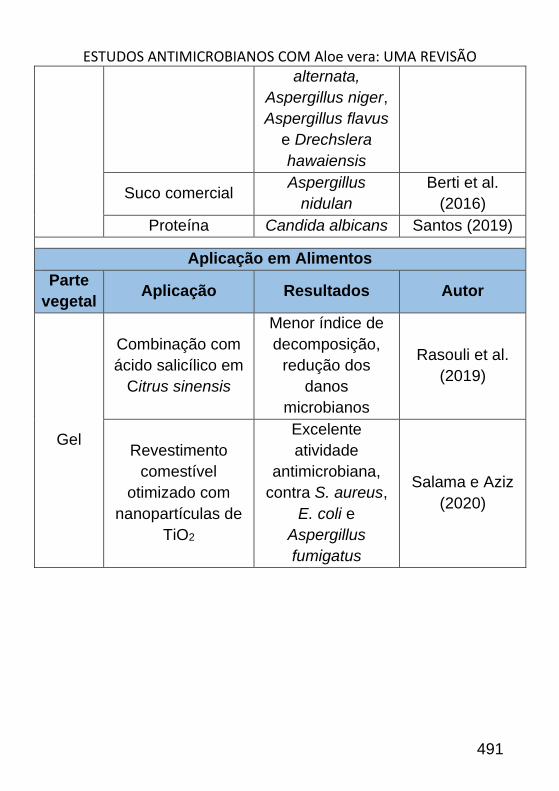
ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
491
alternata,
Aspergillus niger,
Aspergillus flavus
e Drechslera
hawaiensis
Suco comercial Aspergillus
nidulan
Berti et al.
(2016)
Proteína Candida albicans Santos (2019)
Aplicação em Alimentos
Parte
vegetal Aplicação Resultados Autor
Gel
Combinação com
ácido salicílico em
Citrus sinensis
Menor índice de
decomposição,
redução dos
danos
microbianos
Rasouli et al.
(2019)
Revestimento
comestível
otimizado com
nanopartículas de
TiO2
Excelente
atividade
antimicrobiana,
contra S. aureus,
E. coli e
Aspergillus
fumigatus
Salama e Aziz
(2020)
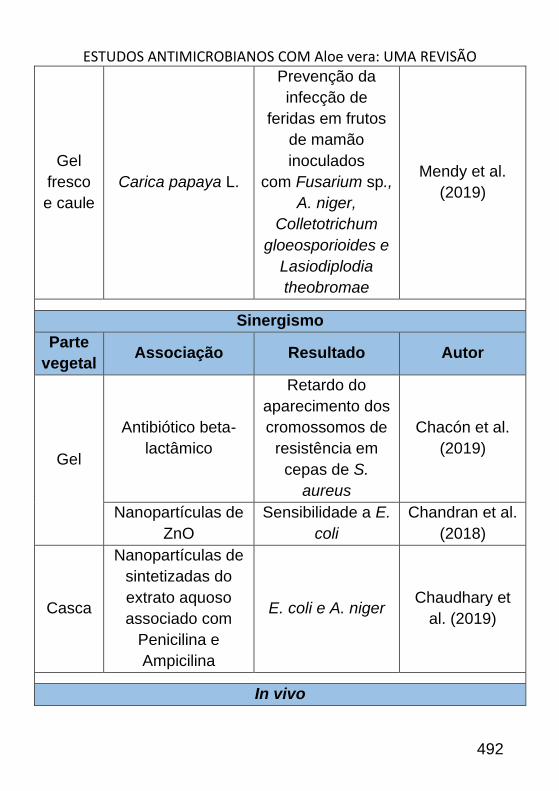
ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
492
Gel
fresco
e caule
Carica papaya L.
Prevenção da
infecção de
feridas em frutos
de mamão
inoculados
com Fusarium sp.,
A. niger,
Colletotrichum
gloeosporioides e
Lasiodiplodia
theobromae
Mendy et al.
(2019)
Sinergismo
Parte
vegetal Associação Resultado Autor
Gel
Antibiótico beta-
lactâmico
Retardo do
aparecimento dos
cromossomos de
resistência em
cepas de S.
aureus
Chacón et al.
(2019)
Nanopartículas de
ZnO
Sensibilidade a E.
coli
Chandran et al.
(2018)
Casca
Nanopartículas de
sintetizadas do
extrato aquoso
associado com
Penicilina e
Ampicilina
E. coli e A. niger Chaudhary et
al. (2019)
In vivo

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
493
Parte
vegetal
Via de
administração Modelo animal Autor
Gel Via Oral
Camundongos
infectados com
Shingella
dysenteride
Hussain et al.
(2017)
Fonte: Dados da pesquisa
Atividade antibacteriana da Aloe vera
As raízes de A. vera oriundas de uma plantação pós-
colheita foram utilizadas para a obtenção de cinco metabólitos
(aloesaponarina-I, desoxeritrolaccina, éster D-metílico do ácido
lacácico, aloesaponarina-II e aloesaponol-I), sendo testados
contra cepas bacterianas fitopatogênicas. Desses metabólicos
dois deles apresentaram ação contra X. campestri, o
desoxeritrolaccina e o éster D-metílico do ácido lacácico, com
CMI (concentração Mínima inibitória) de 46,86 e 93,75 µg/mL,
respectivamente. Demonstrando que, esses componentes
podem atuar como agentes antimicrobianos (CANCHE-
ESCAMILLA et al., 2019).
Um polissacarídeo extraído do gel da A. vera, chamado
de acemanana, possui algumas funcionalidades, com isso foi
feito um estudo em que esse polissacarídeo foi levemente
desacetilado, gerando alguns polímeros. Através do ensaio de
contagem, o polímero que recebeu uma menor desacetilação
apresentou inibição no crescimento bacteriano e inibiu a
formação de biofilmes de duas cepas, E. coli e E. faecalis,
apresentando uma CIB50 (Concentração inibição de biofilme) ≤
7,6 ± 0,7 e CIB90 ≤ 68,2 ± 4,5. Com isso, conclui-se que a

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
494
desacetilação do acemanana pode diminuir o seu potencial
antibacteriano e antibiofilme (SALAH et al., 2017).
Existem fibras derivadas da A. vera que foram isoladas
através do método de eletrofiação, sendo elas o
Pilivinilpirrolidona e Acetato-Polivinilpirrolidona. Com essas
fibras foi feito o teste para saber sua capacidade antibacteriana
em cepas que podem gerar inflamações em cicatrizações de
feridas. Como resultado, observou-se que não houve
crescimento bacteriano nas placas inoculadas com P.
aeruginosa e S. aureus, em diferentes concentrações das
fibras, sendo um candidato promissor para ajudar na
cicatrização de feridas (AGHAMOHAMADI et al., 2019).
Outra molécula extraída do extrato aquoso de Aloe vera
foram as nanopartículas de dióxido de titânio (TiO2-NPs). Elas
foram testadas em cepas de P. aeruginosa, para saber a
possível capacidade antibiofilme. Sendo assim, foi obtido como
resultado uma redução na formação de biofilme da cepa, que
foi incubada junto com o TiO2-NPs na concentração de CMI de
31,25 µg/mL, demonstrando uma grande capacidade de
inibição na formação do biofilme (RAJKUMARI et al., 2019).
Utilizando diferentes métodos de extração, extratos da
seiva e folhas de A. vera foram avaliados quanto a sua ação
antimicrobiana. De acordo com esse estudo, ambos os extratos
apresentaram atividade antimicrobiana, sendo o extrato da
seiva mais eficaz do que o das folhas. A CMI se mostrou ≤ 6,25
µg/mL para a maioria dos microrganismos (P. aeruginosa, B.
subtilis e S. aureus) e os diferentes extratos, concluindo assim
que os extratos de seiva e folhas de A. vera podem ser
utilizados como agentes antimicrobianos (ABAKAR et al.,
2017).

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
495
Diferentes tipos de extrato da folha de A. vera (hexano,
acetato de etila, metanol e água destilada) na concentração de
100 mg/mL foram testados contra algumas espécies de
bactérias, utilizando o método de difusão em poços de ágar.
Todos os extratos, com exceção do acetato de etila,
apresentaram atividade. O maior efeito foi contra S.
marcescens com uma zona de inibição de 13,67 ± 0,57 mm pelo
extrato de hexano e contra B. cereus com 12,33 ± 0,57 mm pelo
extrato de metanol (DHARAJIYA et al., 2017).
Outro estudo utilizando diferentes tipos de extratos da
folha de A. vera, também observou sua ação contra três
espécies de bactérias (E. coli, B. subtilis e S. aureus). Diferente
do trabalho anterior o extrato que possuiu uma maior área de
inibição do crescimento foi o de acetato de etila sendo 6 mm
para E. coli, 4,8 mm para B. subtilis e 2 mm para S. aureus
(SAHOO; DIXIT, 2016).
Atividade antifúngica da A. vera
Através do extrato aquoso de folhas de A. vera são
obtidas nanopartículas de selênio (Se-NPs) utilizando o método
de síntese hidrotermal. As Se-NPs inibiram o crescimento de
micélios de P. digitatum, porém apresentaram baixo efeito
fungicida em C. coccodes. Os autores associaram tal achado
ao mecanismo dos efeitos fungicidas dos íons selênio que inclui
a absorção e o acúmulo dos mesmos pela célula do fungo o que
provocava o encolhimento da membrana citoplasmática e
inibição das atividades vitais da célula (FARDSADEGH;
JAFARIZADEH-MALMIRI, 2019).
A atividade antifúngica do gel de A. vera na
concentração de 10-1 µg/L contra quatro patógenos comumente

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
496
encontrados em frutas pós-colheita: P. digitatum, P. expansum,
B. cinerea e A. alternata foi determinada. As espécies P.
digitatum e A. alternata foram as mais sensíveis. O efeito
inibitório foi baseado na supressão da germinação e
crescimento micelial. A sobrevivência dos esporos de P.
digitatum, A. alternata e B. cinerea foi reduzida em 15-20%. No
entanto, o gel também foi similarmente eficaz contra P.
expansum apenas em concentração mais elevada. O efeito
supressor do gel para P. digitatum e P. expansum, foi mais
elevado conforme o aumento da concentração, com uma
redução de 95% na sobrevivência dos esporos (RODRÍGUEZ
et al., 2005).
Sitara, Hassan e Naseem (2011), também testaram o gel
de A. vera nas concentrações de 0,15%, 0,25% e 0,35% frente
aos fitopatógenos A. niger, A. flavus, A. alternata, D. hawaiensis
e P. digitatum. Houve redução significativa no crescimento dos
fungos proporcionalmente à concentração testada. O gel na
concentração de 0,15% apresentou notável atividade
antifúngica para todos os fungos em comparação ao controle.
São-Pedro et al. (2020) averiguaram os extratos
metabólicos das folhas verde (FV), folhas secas ao sol (FSS) e
secas na estufa (FSE) frente a espécie de C. glabrata. FSS e
FSE não apresentaram inibição do crescimento frente ao
microrganismo testado. O extrato FV foi fungistata para C.
glabrata apresentando o valor de CMI de 250μg/mL. Os autores
do trabalho atribuíram essa atividade às propriedades químicas
das substâncias presentes na planta, acreditando assim que
elas possam ter atuado de forma sinérgica. Algumas destas
substâncias foram identificadas como o pirocatecol, ácido
cinâmico, ácido ascórbico e ácido p-cumárico.

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
497
Berti et al. (2016) avaliaram o efeito do suco comercial
da A. vera, nas concentrações de 4% e 8% (v/v), frente a
germinação e antimutagênese em A. nidulan. A maior
concentração do suco se mostrou a mais eficiente, estimulando
a germinação em todas as linhagens de A. nidulans. Entretanto,
em relação à diminuição de esporos mortos e malformados,
ambas as concentrações se mostraram benéficas, porém, de
forma aleatória. A antimutagenicidade foi demonstrada
somente pelo extrato a 8%.
Santos (2019), relatou a presença de uma proteína
obtida do gel das folhas de A. vera com massa molecular de
14,9 kDa, presença de açúcares detectáveis por RMN
(ressonância magnética nuclear) e potente ação antifúngica
frente à C. albicans. A partir do método de diluição em
microplacas, realizou-se a avaliação da formação de biofilme.
Estas frações nas concentrações de 250; 125; 31,2 e 7,8 g/mL,
apresentaram atividade antifúngica no que concerne à inibição
da formação de biofilme de C. albicans, inibindo cerca de 70%
do mesmo.
Atividade antimicrobiana da A. vera nos alimentos
O gel de A. vera tem sido bastante utilizado nos estudos
recentes, o qual é constituído por polissacarídeos, minerais,
açúcares, vitaminas e compostos fenólicos, e têm demonstrado
diferentes propriedades biológicas. Um exemplo disso é sua
utilização no tratamento de frutos que podem ser
comercializados com a qualidade maior. Um estudo feito com o
gel de A. vera a 30 % junto com ácido salicílico a 2,0 mM em
Citrus sinensis L. Osbeck, demonstrou resultados significativos,
onde a fruta tratada apresentou menor índice de decomposição,

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
498
redução da carga microbiana e dos danos causados pelo frio,
em comparação com o controle. Concluindo assim que a
combinação desses dois compostos pode aumentar a
qualidade e o tempo de vida de prateleira das laranjas
(RASOULI et al., 2019).
Outro estudo feito com a A. vera no tratamento de frutos,
foi a utilização de dois extratos da planta, que foram submetidos
para investigar o efeito sobre fungos encontrados em mamão
(Carica papaya L.) pós-colheita. Os procedimentos foram
utilizados em concentrações diferentes dos extratos (0, 15, 25
e 50% v/v), tendo como resultado a diminuição da infecção
fúngica (Fusarium sp., L. theobromae, A. niger e C.
gloeosporioides) inoculados no mamão (MENDY et al., 2019).
Um revestimento comestível foi preparado com a A. vera
(AV), utilizando 1,94 g, junto com Alginato (1,014 g), além desse
gel, foi feito também a incorporação de nanopartículas de óxido
de titânio (TiO2-NPs) nas concentrações de 1, 3 e 5 %. Esses
filmes foram testados contra três espécies microbiológicas, S.
aureus, E. coli e A. fumigatus, utilizando o método de zona de
inibição. Percebeu-se que os filmes demonstraram uma
excelente atividade antimicrobiana, comparada com o controle
(apenas o alginato), sendo maior nos filmes contendo o TiO2-
NPs (SALAMA; AZIZ, 2020).
Sinergismo
A A. vera também possui estudos em que foram
avaliados sua ação antimicrobiana junto com outra substância.
Um exemplo disso foi um estudo em que seu objetivo foi avaliar
o aparecimento de resistência cromossômica em bactérias
gram-positivas causadoras de mastite em gado, quando são

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
499
expostas a A. vera (AV), junto com o antibiótico beta-lactâmico.
Como resultado a combinação de A. vera e ceftiofur ou
cloxacilina (20 mg/mL AV + 0,06 µg/mL antibiótico) retardou o
aparecimento dos cromossomos de resistência em cepas de S.
aureus, reduzindo a dose de antibióticos que são utilizados no
tratamento de gado leiteiro que possuem Mastite (CHACÓN et
al., 2019).
Para avaliar a eficiência do gel de A. vera foi realizado a
combinação do gel junto com nanopartículas de óxido de zinco
(ZnO-NPs) e avaliação in vitro dessa combinação contra
patógenos humanos utilizando em concentrações de 20 e 40
µg/mL. Resultados mostraram que a mistura dos dois
componentes aumentou a ação antibacteriana e antifúngica,
comparado com o padrão de Cloranfenicol e Estreptociclina,
sendo a E. coli o patógeno mais sensível a mistura
(CHANDRAN et al., 2018).
Nanopartículas de óxido de zinco (ZnO-NPs) também
foram sintetizadas a partir do extrato aquoso da casca de A.
vera e foi feito sua avaliação antimicrobiana contra alguns
patógenos e junto com dois antibióticos (Penicilina e Ampicilina)
obtendo uma concentração de 1 mg/mL. De acordo com os
resultados as nanopartículas apresentaram atividade contra
cepas de E. coli (MTCC-41), tendo uma zona de inibição de 14
± 0,58 mm e para o fungo A. niger com 15 ± 1,53. Porém a
junção do ZnO-NPs com os antibióticos apresentaram uma
zona de inibição menor do que apenas o antibiótico sozinho
(CHAUDHARY et al., 2019).

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
500
Estudos in vivo
Existem também estudos in vivo utilizando a A. vera
como a utilização do prebiótico Lassi suplementado com Aloe
vera (APL) que foi testado na prevenção de infecção por S.
dysenteride em camundongos administrados por via oral. Como
resultado, este trabalho apresentou que houve uma redução na
contagem de S. dysenteride em todos os órgãos comparados
com outros grupos de tratamento e aumentou também os níveis
de anticorpos IgA, demonstrando assim que o APL apresentou
efeito imunoprotetor contra a bactéria nos camundongos
(HUSSAIN et al., 2017).
CONCLUSÕES
Dessa forma concluímos que a A. vera é uma planta com
grande potencial antimicrobiano, podendo ser utilizada em
diversos procedimentos voltados as indústrias alimentícias e
farmacêuticas.
REFERÊNCIAS BIBLIOGRÁFICAS ABAKAR, H. O. M.; BAKHIET, S. E. A.; ABADI, R. S. M. Antimicrobial activity and minimum inhibitory concentration of Aloe vera sap and leaves using different extracts. Journal of Pharmacognosy and Phytochemistry, v. 6, n. 3, p. 298-303, 2017. AGHAMOHAMADI, N. et al. Preparation and characterization of Aloe vera acetate and electrospinning fibers as promising antibacterial properties materials. Materials Science and Engineering: C, v. 94, p. 445-452, 2019. AKABERI, M. et al. Therapeutic effects of Aloe spp. in traditional and modern medicine: A review. Biomedicine & Pharmacotherapy, v. 84, p. 759-772, 2016. BARUAH, A.; BORDOLOI, M.; BARUAH, H. P. D. Aloe vera: A multipurpose industrial crop. Industrial Crops and Products, v. 94, p. 951-963, 2016.

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
501
BERTI, A. P. et al. Efeitos do suco comercial de Aloe vera L. na germinação e antimutagênese em Aspergillus nidulans e pelo ensaio cometa em ratos Wistar. Revista Brasileira de Biociências, v. 14, n. 2, 2016. CANCHE-ESCAMILLA, G. et al. Extraction of phenolic components from an Aloe vera (Aloe barbadensis Miller) crop and their potential as antimicrobials and textile dyes. Sustainable Chemistry and Pharmacy, v. 14, p. 100168, 2019. CHACÓN, O. et al. Effect of Aloe barbadensis Miller (Aloe vera) associated with beta-lactam antibiotics on the occurrence of resistance in strains of Staphylococcus aureus and Streptococcus uberis. European Journal of Integrative Medicine, v. 32, p. 100996, 2019. CHANDRAN, S. et al. Enhanced Antimicrobial activity of Aloe vera blended Zinc Oxide Nanoparticles in PVA matrix. Materials Today: Proceedings, v. 5, n. 8, p. 16190-16198, 2018. CHAUDHARY, A. et al. Antimicrobial activity of zinc oxide nanoparticles synthesized from Aloe vera peel extract. SN Applied Sciences, v. 1, n. 1, p. 136, 2019. DHARAJIYA, D. et al. Antimicrobial activity and phytochemical screening of Aloe vera (Aloe barbadensis Miller). Int. J. Curr. Microbiol. App. Sci, v. 6, n. 3, p. 2152-2162, 2017. FARDSADEGH, B.; JAFARIZADEH-MALMIRI, H. Aloe vera leaf extract mediated green synthesis of selenium nanoparticles and assessment of their in vitro antimicrobial activity against spoilage fungi and pathogenic bacteria strains. Green Processing and Synthesis, v. 8, n. 1, p. 399-407, 2019. GIANNAKOUDAKIS, D. A. et al. Aloe vera waste biomass-based adsorbents for the removal of aquatic pollutants: a review. Journal of environmental management, v. 227, p. 354-364, 2018. HUSSAIN, S. A. et al. Aloe vera (Aloe barbadensis Miller) supplemented probiotic lassi prevents Shigella infiltration from epithelial barrier into systemic blood flow in mice model. Microbial pathogenesis, v. 102, p. 143-147, 2017. MENDY, T. K. et al. Antifungal properties of Aloe vera through in vitro and in vivo screening against postharvest pathogens of papaya fruit. Scientia Horticulturae, v. 257, p. 108767, 2019. MORRISON, L.; ZEMBOWER, T. R. Antimicrobial Resistance. Gastrointestinal endoscopy clinics of North America, v. 30, p. 619-635, 2020. RAJKUMARI, J. et al. Synthesis of titanium oxide nanoparticles using Aloe barbadensis mill and evaluation of its antibiofilm potential against Pseudomonas aeruginosa PAO1. Journal of Photochemistry and Photobiology B: Biology, v. 201, p. 111667, 2019.

ESTUDOS ANTIMICROBIANOS COM Aloe vera: UMA REVISÃO
502
RASOULI, M.; SABA, M. K.; RAMEZANIAN, A. Inhibitory effect of salicylic acid and Aloe vera gel edible coating on microbial load and chilling injury of orange fruit. Scientia horticulturae, v. 247, p. 27-34, 2019. RODRIGUEZ, D. J. et al. Antifungal activity in vitro of Aloe vera pulp and liquid fraction against plant pathogenic fungi. Industrial Crops and Products, v. 21, n. 1, p. 81-87, 2005. SAHOO, S.; DIXIT, S. Analysis of Qualitative and Quantitive Constituents and Antimicrobial Activities of Aloe Vera L. against Some Selected Pathogens. 2016. SAKS, Y.; BARKAI-GOLAN, R. Aloe vera gel activity against plant pathogenic fungi. Postharvest Biology and Technology, v. 6, n. 1-2, p. 159-165, 1995. SALAH, F. et al. Effect of the deacetylation degree on the antibacterial and antibiofilm activity of acemannan from Aloe vera. Industrial Crops and Products, v. 103, p. 13-18, 2017. SALAMA, H. E.; AZIZ, M. S. A. Optimized alginate and Aloe vera gel edible coating reinforced with nTiO2 for the shelf-life extension of tomatoes. International Journal of Biological Macromolecules, 2020. SÁNCHEZ-MACHADO, D. I. et al. Aloe vera: Ancient knowledge with new frontiers. Trends in Food Science & Technology, v. 61, p. 94-102, 2017. SANTOS, K. F. S. Potencial antifúngico de frações do gel de Aloe vera (l.) burm. f. frente à Candida albicans. 2019. SÃO-PEDRO, G. R. B. et al. Avaliação Da Atividade Antimicrobiana Do Extrato Metanólico Da Aloe vera. Revista Cientefico, v. 20, n. 41, p. 195-210, 2020. SITARA, U.; HASSAN, N.; NASEEM, J. Antifungal activity of Aloe vera gel against plant pathogenic fungi. Pakistan Journal of Botany, v. 43, n. 4, p. 2231-2233, 2011.

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
503
CAPÍTULO 24
FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
Suelen Cristina Gomes Dos SANTOS1
Lara Nardi BARONI1
Carolina Maria Da SILVA2
1Graduação em Biomedicina pelo Centro Universitário UniFBV Wyden, Brasil; 2Orientadora/Professora da UniFBV e Doutorado em Biologia de Fungos pela Universidade
Federal de Pernambuco, UFPE, Brasil. [email protected]
RESUMO: Arboviroses são doenças transmitidas por arbovírus, a exemplo do vírus denominado Zika, que foi isolado pela primeira vez no macaco rhesus em 1947 na Uganda. A doença causada por esse vírus é transmitida a partir da picada de fêmeas infectadas do gênero Aedes, sendo o Aedes aegypti o principal vetor no Brasil. Recentemente a infecção pelo Zika tem sido associada ao nascimento de bebês com microcefalia, que se caracteriza pela observação do crânio e cérebro pequenos, com calcificações e atrofia cerebral e má formação ventricular. De acordo com a Organização Mundial de Saúde é uma malformação congênita e multifatorial, pode envolver fatores genéticos e ambientais. Estudos têm demonstrado que o vírus pode estar relacionado com a doença, pois é encontrado no líquido amniótico de gestantes infectadas por ele. Diante dos exposto, o objetivo deste trabalho foi verificar o mecanismo fisiopatológico do vírus Zika frente ao sistema nervoso central correlacionando ao desenvolvimento da microcefalia. Tratou-se de trabalho de revisão de literatura, onde foram utilizadas as plataformas Google Acadêmico, Pubmed, Scielo e Periódicos Capes para se pesquisar artigos publicados entre os anos de 2015 e 2020. A intensa urbanização intensifica o aumento da incidência de doenças como arboviroses. Podendo afetar a

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
504
saúde, bem estar, segurança da população afetando também o meio ambiente ao seu redor. Ao longo dos últimos anos, tem se observado a ocorrência de diversos surtos de doenças virais transmitidas por vetores como o A. aegypty. Dentre as arboviroses transmitidas por fêmeas do A. aegypty destaca-se o Zika vírus, que foi isolado pela primeira vez no macaco Rhesus em 1947 em Uganda. O vírus é novo nas Américas, porém não há imunidade frente a ele dentro da população. Uma correlação do Zika vírus com sequelas neurológicas como a microcefalia começou a ser destacada recentemente. Os casos de microcefalia aumentou em outubro de 2015 com início no estado de Pernambuco e posteriormente se espalhando pelos outros estados região Nordeste. A microcefalia se caracteriza pela observação do crânio e cérebro pequenos, com calcificações, atrofia cerebral e má formação ventricular. De acordo com a Organização Mundial de Saúde é uma malformação congênita e multifatorial, pode envolver fatores genéticos e ambientais. Palavras-chave: Arbovirose. Zika. Microcefalia. Fisiopatologia. Infecção.
INTRODUÇÃO
O Vírus Zika é um flavivírus da família Flaviviridae e foi o
responsável pelo surto de infecção que ocorreu em 2015 no
Nordeste do Brasil. Junto com esse surto se observou o
aumento dos casos de microcefalia, principalmente No Estado
de Pernambuco. A Comprovação da correlação existente entre
os recém-nascidos com microcefalia e suas mães infectadas
pelo vírus, ocorreu através da detecção do vírus no líquido
amniótico em gestantes pela Fiocruz na Paraíba (NUNES, et al,
2016).
A microcefalia segundo a Organização Mundial de
Saúde (OMS) é caracterizada por uma redução na

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
505
circunferência da cabeça, com a medida occipitofrontal
(circunferência do crânio) de um recém-nascido com 37
semanas de gestação igual ou menor que 31,9 cm para
meninos e 31,5 para meninas. Pode ser diagnosticada através
de exames de imagens, como a ultrassonografia, durante a
gestação e teste inicial para neonatos (RUSSO, JUNGMANN e
BELTRÃO, 2017; GOODFELLOW et al, 2018).
O vírus Zika tem a capacidade de ser transmitido pela
forma vertical (da mãe para o feto) através da passagem pela
placenta. Além disso, dependendo do período gestacional em
que a mulher adquiriu o vírus, aumenta a chance de o feto
apresentar microcefalia e outros problemas principalmente
neurológicos (NUNES, et al., 2016; PRATA, et al., 2019).
Adicionalmente, essa infecção pode causar outros
problemas que atrapalham no desenvolvimento da criança,
como por exemplo, o baixo peso e a baixa estatura para a idade,
e o retardo no desenvolvimento neurológico. Essas
consequências pós-infecção atrapalham no desenvolvimento
da criança (PRATA, et al., 2019).
Desta forma, o objetivo do trabalho é verificar o
mecanismo fisiopatológico do Zika vírus frente ao sistema
nervoso central correlacionado ao desenvolvimento da
microcefalia em neonatos.
MATERIAIS E MÉTODO
Esse trabalho tratou-se de uma revisão de literatura
sobre a fisiopatologia da infecção pelo Zika vírus no sistema
nervoso central de neonatos.
Para a coleta de dados foram utilizadas as plataformas
Google Acadêmico, Pubmed, Scielo e Periódicos Capes. Os

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
506
descritores para busca foram: “Arbovirose”, “Zika”,
“Microcefalia”, “Fisiopatologia” e “Infecção”. Foram incluídos no
estudo artigos publicados entre os anos de 2015 e 2020, na
língua inglesa ou portuguesa.
Foram excluídos os trabalhos que não estavam
disponíveis na íntegra e cujo tema central divergia do objetivo
deste trabalho. seno utilizados 30 atigos nas línguas portuguesa
e inglesa.
RESULTADOS E DISCUSSÃO
A intensa urbanização em conjunto com o uso do solo de
forma inadequada intensificam ameaças como enchentes,
tempestades e deslizamentos, ou até mesmo o aumento da
incidência de doenças como arboviroses. No Brasil, a ocupação
de áreas de risco e condições socioeconômicas deficientes
auxiliam no aparecimento de grupos sociais sob essas
ameaças devido a vulnerabilidade presente no local.Os
impactos ambientais podem ser observados nas cidades, assim
como, a degradação ambiental que é uma consequência da
formação de uma urbanização descomprometida com os
aspectos físico-ambientais (ALMEIDA, 2020).
A legislação brasileira caracteriza o impacto ambiental
como qualquer tipo de alteração, de caráter físico, químico ou
biológico do meio ambiente. Essas alterações podem afetar a
saúde, bem estar, segurança da população afetando também o
meio ambiente ao seu redor. Os diversos impactos ambientais
oferecem riscos à saúde humana, um exemplo disso é o
saneamento ineficaz que pode vir acarretar doenças
transmitidas pela água como esquistossomose, hepatite e
diarreias. Além de, propiciar a reprodução de vetores de outras

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
507
doenças como no caso do Aedes aegypti, principal responsável
pela transmissão das arboviroses (ALMEIDA, 2020).
As doenças infecciosas são consideradas de grande
importância para a saúde pública, além de, estarem associadas
à pobreza e a condições de vida inadequadas. No Brasil, é
possível observar que essas doenças persistem, devido a
transição epidemiológica e demográfica. Tendo como
predominantes as doenças transmissíveis, podendos ser
observadas em diferentes regiões do país (DE SOUZA, 2020).
Dentro de um contexto social diverso, se torna complexo
o trabalho da vigilância em saúde em avaliar e disseminar
informações para obter a tomada de decisão. Portanto, a
identificação de áreas susceptíveis para doenças infecciosas e
o entendimento da relação com indicadores socioeconômicos é
muito importante para determinar as ações da vigilância.
Ajudando também, no planejamento de medidas de controle e
intervenção da doença, bem como articular ações intersetoriais
de mitigação das causas que determinam o aparecimento
dessas patologias (DE SOUZA, 2020).
Ao longo dos últimos anos tem se observado a
ocorrência de diversos surtos de doenças virais transmitidas por
mosquitos, como a Chikungunya, a dengue e o Zika. Esta
ocorrência tem sido principalmente associada ao
desmatamento ambiental e ao acumulo de água que favorecem
a proliferação de vetores como o A. aegypty no ambiente
domiciliar. A trajetória desse vetor é caracterizada como um
comportamento sinantrópico e antropófilico, acompanhando o
homem de acordo com seu deslocamento (LIMA, 2016).
A distribuição geográfica do A. aegypty, pode ser
associada ao clima, sendo os climas tropical e subtropical os
mais favoráveis para o seu desenvolvimento. Porém existem

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
508
outros aspectos que influenciam no aparecimento do mosquito
como o aumento do fluxo populacional, acumulo de lixo,
moradias inapropriadas e abastecimento de água inadequado
(LIMA, 2016).
Dentre as arboviroses transmitidas por fêmeas do A.
aegypty destaca-se o Zika vírus, que foi isolado pela primeira
vez no macaco rhesus em 1947 em Uganda. Já os primeiros
casos do isolamento deste vírus em humanos foram relatados
na Nigéria, e os primeiros relatos no Brasil surgiram por volta
do ano de 2015 (DONALISIO et al., 2017).
Da mesma forma que as outras arboviroses, o Zika vírus
se espalha de acordo com as condições favoráveis à
proliferação do mosquito, que deposita ovos em recipientes de
água e as fêmeas são hematófagas. Por isso a eliminação dos
criadouros é tarefa complexa nas cidades brasileiras,
especialmente em lugares com condições precárias e falta de
saneamento além da coleta de lixo irregular (HENRIQUES et
al., 2016).
Além da transmissão vetorial a partir da picada de
fêmeas infectadas pelo gênero Aedes, principalmente o A.
aegypti, a patologia pode ser transmitida por outras formas
como por via transfusional, perinatal, e há relatos que podem
ocorrer transmissão ocupacional em laboratórios e por relação
sexual (DONALISIO et al., 2017; TRINDADE et al., 2019).
Em relação a morfologia e classificação taxonômica
Wang e colaboradores (2017) destacam que o vírus Zika é um
arbovírus que pertence ao Gênero Flavivirus e família
Flaviviridae. O genoma da cepa africana ZIKV (MR766) é
composto por uma molécula simples de RNA de sentido
positivo, possuinte de um tamanho de ~10.9-kb. A região de
leitura codifica uma poliproteína que é posteriormente

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
509
transformada em proteínas da membrana, do capsídeo,
envelope viral, e sete proteínas não estruturais: NS1, NS2A,
NS2B, NS3, NS4A, NS4B e NS5.
O vírus Zika é caracterizado por ser um vírus envelopado
de RNA de fita simples e apresenta uma poliproteína que é
clivada em dez proteínas, sendo três delas responsáveis por
formar a cápsula, o envelope e a membrana precursora, e as
outras sete são proteínas não estruturais. E ele consegue entrar
na célula graças a interação das glicoproteínas da sua cápsula
com os receptores celulares. Além de fatores que facilitam sua
entrada como os TAM que são receptores da família da
tirosinoquinase como o TYRO3, AXL e MER (LI, et al., 2016).
O capsídeo viral possui um formato icosaedro envolto de
uma bicamada lipídica derivada da célula hospedeira. As
proteínas M e E são localizadas na parte superior do vírus e
possuem hélices transmembranares, responsáveis pela
ancoragem na parte externa da membrana. À medida que as
partículas virais são liberadas, a proteína prM é clivada por uma
protease presente na região trans do complexo de Golgi,
formando peptídeos pr e a proteína M (WANG et al., 2017).
Já as proteínas não estruturais (NS1-NS5) são
responsáveis por formar um complexo replicativo, além de
desempenharem um importante papel de antagonista do
sistema imune inato do hospedeiro. Sendo que a estrutura da
proteína NS1 é altamente semelhante ao de outros flavovírus e
apresenta um potencial eletrostático notavelmente divergente
na superfície de seu loop. E esta região divergente está
intimamente ligada aos fatores de ligação à célula hospedeira e
a proteção contra os anticorpos. Ela ainda participa da evasão
imune e juntamente com a NS4A parece que desempenha um
papel na replicação viral. Além disso a NS4A junto com as

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
510
proteínas NS2A, NS2B, NS4B formam um grupo de proteínas
hidrofóbicas que parecem estar associadas a membrana
plasmática embora não parecem ter função de atividade
enzimática, mas suas funções precisam ser melhor elucidadas
(ZHU et al., 2016).
Enquanto que a proteína NS3 é responsável e
necessária para a replicação e o processamento da poliproteina
por causa do sua porção N-terminal da cadeia polipeptídica que
apresenta a protease e na sua região C-terminal que apresenta
a helicase. Já a proteína NS5 é uma RNA polimerase
dependente de RNA que é realizada pela sua região C-terminal,
além de apresentar um domínio de atividade metiltrasferase
que se localiza na região N-terminal que tem a função de fazer
o capeamento do RNA viral (MANDACARU,2019).
A proteína E é considerada a principal proteína presente
na superfície do vírus e possui a função de ligação molecular
entre receptores da célula hospedeira e fusão de suas
membranas. Esta proteína no Zika possui uma diferença
quando comprado Ao vírus da dengue (DENV), pois no vírus
DENV a proteína E é glicosilada na asparagina 67
apresentando uma interação importante com o receptor DC-
SIGN da célula hospedeira. Já no Zika a glicosilação no
aminoácido asparagina 67 parece ser ausente (MANDACARU,
2019).
Adicionalmente, Mandacaru (2019) destaca que quando
se observa a evolução do vírus Zika e dos outros membros do
seu gênero, se percebe que o mesmo apresenta uma alta
similaridade genômica com o vírus da dengue (DENVs) em um
alto nível hierárquico. Mesmo sendo afirmado por Wang et al.
(2017) que o Zika é mais próximo filogeneticamente do
Spondweni vírus.

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
511
Além disso, o Zika vírus apresenta um cluster (grupo
genético) único e essa separação de seu cluster dos cluster
similares em seu nível hierárquico sugeriu que o Zika
desenvolveu mecanismos distintos de outras viroses. Outra
característica deste vírus é a sua elevada atividade de
recombinação homóloga que é incomum nos demais flavovirus.
Por isso essa sua característica é atribuída a sua necessidade
de se adaptar aos hospedeiros (MANDACARU, 2019).
O vírus é considerado novo nas Américas, porém não
há imunidade frente a ele dentro da população, além disso o
número total de casos não é totalmente comprovado devido à
falta de procura por assistência em saúde, por isso os casos de
ocorrência da patologia são baseados em casos suspeitos
(TRINDADE et al., 2019).
Nos últimos ano tem se dado uma importante atenção às
medidas de controle contra o Zika vírus, sobretudo em
gestantes. O Ministério da Saúde recomenda especialmente à
esta população que se mantenha portas e janelas fechadas,
uso de roupas compridas ou de mangas e utilizar repelentes.
Pois, a melhor forma de proteção contra o vírus é se proteger
do vetor que o transmite (HENRIQUES et al., 2016).
A sintomatologia da doença aparece em poucos dias em
relação a picada do mosquito, durando de 3 a 12 dias e incluem
febre baixa, artralgia, mialgia, dor de cabeça, conjuntivite e
exantema maculopapular (DONALISIO et al., 2017).
Além disso, alguns autores relatam que a infecção pelo
Zika vírus pode resultar em graves falhas neurológicas que
causam paralisia cerebral, retardo mental e convulsões
(ASHWAL et al., 2009; RUSSO et al., 2017). Porém, o
percentual dessa doença em relação a hospitalização é baixo,

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
512
e mais de 80% dos infectados são assintomáticos (TRINDADE
et al., 2019).
Uma importante correlação do Zika vírus com sequelas
neurológicas começou a ser destacada recentemente. O
aumento do número de casos de microcefalia aumentou em
outubro de 2015 com início no estado de Pernambuco e
posteriormente se espalhando pelos outros estados região
Nordeste, coincidentemente meses depois da confirmação da
ocorrência de surtos de transmissão pelo vírus Zika nessas
áreas (HENRIQUES et al., 2016).
No mesmo ano, foi criado o comitê Internacional de
Regulação de Emergências em Saúde que se reuniu dia 1° de
fevereiro e declarou que os casos de microcefalia estavam
inteiramente relacionados a febre pelo vírus Zika, essa
declaração foi feita a partir de casos similares na polinésia
Francesa no ano de 2014, considerados emergência de saúde
pública (HENRIQUES et al., 2016).
A microcefalia se caracteriza pela observação do crânio
e cérebro pequenos, com calcificações e atrofia cerebral e má
formação ventricular. De acordo com a Organização Mundial de
Saúde é uma malformação congênita e multifatorial, pode
envolver fatores genéticos e ambientais. A microcefalia pode
observada através da medição do perímetro cefálico do
neonato, sendo estabelecida quando a medição é igual ou
inferior a 31,9 cm para o sexo masculino e igual ou inferior a
31,5 cm para o sexo feminino (BRASIL et al., 2016; de FÁTIMA
et al., 2016; RASMUSSEN et al., 2016; RUSSO et al., 2017;
TRINDADE et al., 2019).
A partir de 2015 diversos estudos passaram a ser
conduzidos para tentar explicar a correlação da infecção pelo
vírus Zika em gestante e o nascimento de bebês com

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
513
microcefalia. Onorati e colaboradores (2016) e Russo et al.
(2017) destacam que cérebro é menor porque o vírus transloca
a proteína quinase de ligante TANK fosforilada (pTBK1) do
centrossoma para a mitocôndria causando a morte celular pela
mudança do ciclo celular. Além disso, Herling et al. (2016)
afirmam que o vírus infecta as células progenitoras corticais do
feto causando a redução da zona de proliferação,
interrompendo a zona cortical.
Outras infecções virais como por citomegalovírus, vírus
do oeste do Nilo, rubéola, Chikungunya e HIV também estão
associadas a má formação neural durante o desenvolvimento
do feto. Dentro deste contexto tem sido observado que más
formações neurológicas na infecção por Zika, apresentam
semelhanças com as causadas pelo citomegalovírus
(MANDACARU, 2019).
O primeiro trimestre da gravidez é considerado bastante
crucial no desenvolvimento neurológico do feto. A infecção pelo
Zika na mãe nesse período é mais susceptível para que ocorra
danos ao sistema nervoso central do feto (BLAZQUEZ; SAIZ,
2016).
O vírus já foi isolado do sangue e líquido amniótico
(LUSTIG et al., 2016; CALVET et al., 2016), assim como do leite
materno (BESNARD et al., 2014), cérebro de feto (MLAKAR et
al., 2016), e tecido placentário (TABATA et al., 2016).
Os fibroblastos dérmicos e os queratinócitos epidérmicos
são os alvos da infecção pelo vírus Zika. As células dendríticas
dérmicas estão em segundo lugar na preferência pelo vírus,
fazendo com que facilite a disseminação do vírus por diferentes
órgãos através do sistema circulatório. A transmissão do ZIKV
de mãe para feto ocorre devido à infecção de macrófagos

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
514
placentários células Hofbauer e citotrofoblastos (HAMEL et al.,
2015; MANDACARU, 2019).
Na placenta, os macrófagos são as principais células
atingidas pelo zika. A infecção nesses macrófagos ativa a
produção de interferon I e citosinas pró-inflamatórias, levando a
uma expressão de genes e proteínas antivirais (OLAGNIER et
al., 2016).
Estudos conduzidos por Tang e colaboradores (2016)
demonstraram que o Zika pode ser replicar principalmente nas
células progenitoras neurais (NPCs) de ratos. Assim como,
podem também infectar NPCs humana (hNPCs). A infecção
causa uma desregulação do ciclo celular, provocando uma
ativação de vias apoptóticas em hNPCs, tendo como resultado
o crescimento reduzido dessas células.
O vírus entra mediante receptores celulares como
proteínas de choque térmico, DC-SIGN, TYRO3, AXL e TIM-1.
Dentre estes, AXL é uma fosfatidilserina e é super expressa
durante o desenvolvimento de células cerebrais humanas,
incluindo células glia radial, micróglias e astrócitos
(NOWAKOWSKI et al., 2016).
Células que possuem AXL altamente expressa são
vulneráveis ao ZIKV. A entrada do vírus na célula através da
formação de endossomos é facilitada quando ocorre a ligação
das partículas virais com o receptor AXL. O envelope viral é
degradado devido ao baixo pH dos endossomos e o material
genético do vírus é liberado no citoplasma da célula infectada.
Alguns RNAs virais podem ser replicados logo após sua
liberação no citoplasma, formando assim, novos vírus a partir
da utilização da própria maquinaria da célula infectada. Os
novos RNAs e proteínas produzidos, formarão novas partículas
virais que serão liberadas da célula (FAIZAN et al., 2016).

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
515
O vírus foi encontrado a partir de exames de ultrassom e
a presença do ácido ribonucléico (RNA) do vírus no líquido
amniótico. Um potencial indicador da relação entre o vírus e a
microcefalia é o relato do aparecimento de erupções cutâneas
na mãe durante a gestação (TRINDADE et al., 2019).
Estudos sugerem que a placenta seria o principal
mediador da transmissão vertical do zika, por causa das
propriedades estruturais, celulares e imunológicas únicas que
possui. A placenta atua como barreira de proteção contra
infecções, mas há sugestões de que este vírus ultrapasse a
barreira placentária infectando e destruindo células, alcançando
assim, o feto (BAYER et al., 2016; QUICKE et al., 2016).
Um passo importante para entender os mecanismos de
infecção do Zika no desenvolvimento do cérebro e a ligação do
vírus com a microcefalia é encontrar as células que são
afetadas ou são vulneráveis à infecção depois que o ZIKV
rompe a barreira placentária. (ZHEXING WEN; HONGJUN
SONG; GUO-LI MING, 2017.). Há algumas evidências de que
o vírus atinge as células do complexo do poro nuclear (NPC)
cortical, mas não atingem tanto os neurônios (TANG et al.,
2016).
Além das NPCs, alguns estudos de Zika vírus em
culturas que reunem tipos diferentes de células do tecido
cerebral de necropsia de fetos, descobriram que astrócitos,
microglia e células precursoras de oligodendrócitos ao longo do
córtex, podem ser infectados pelo vírus. Algumas dessas são
consideradas essenciais no desenvolvimento do cérebro e suas
funções (NORONHA et al., 2016; RETALLACK et al., 2016).
Além do Zika causar microcefalia em fetos, ele pode
causar outros problemas associados ao sistema nervoso, a
exemplo da Síndrome da Zika congênita que é uma

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
516
encefalopatia crônica não progressiva. Além disso outros danos
têm sido relatados como retardo do crescimento intrauterino,
malformações cerebrais como a calcificação subcorticais e
defeito de migração cortical, desproporção craniofacial, entre
outros problemas neurológicos (PRATA et al., 2019).
As crianças que apresentam a microcefalia ou a
síndrome da Zika congênita, em geral apresentam estatura e
peso menor que as crianças saudáveis na mesma idade. Isso
pode ocorrer por fatores nutricionais e pela ingestão inferior de
carboidratos que o necessário ou fatores não nutricionais. Mas,
não se pode definir qual deles causou o menor crescimento da
criança por sua causa ser multifatorial. Além disso esse baixo
crescimento pode ser uma causa secundária da má formação
cerebral. (PRATA et al., 2019).
Além do que exposto anteriormente as crianças que
apresentam a síndrome da Zika congênita, apresentam outras
anomalias neurológicas além da microcefalia, dentre elas a
hipertonia grave com hiper-reflexia, irritabilidade,
hiperexcitabilidade, choro excessivo, distúrbio de deglutição e
respostas auditivas e visuais comprometidas. Adicionalmente,
algumas bebês apresentam crises de convulsão que vão
aumentando sua frequência com o tempo, além de após o
terceiro mês começarem a apresentar crises epilépticas
principalmente do tipo espasmos epiléticos (EICKMANN et al,
2016).
Santana (2018) ainda destacam que as crianças
acometidas apresentam espasticidade em diferentes grupos
musculares que dificulta o desenvolvimento motor, e
deformidades musculoesqueléticas que levam a postura e
padrões de movimentos anormais e o atraso na aquisição das
habilidades motoras (sentar, engolir, ficar em pé e caminhas).

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
517
Mesmo a microcefalia sendo a principal descoberta nos
recém-nascidos acometidos com a infecção congênita por zika,
também outras má-formações tem sido associadas como
perdas auditivas e achados de anomalias oculares (MIURA; DA
SILVA; DE ALMEIDA, 2020).
Observando todos esses agravantes, o Ministério da
Saúde produziu protocolos de vigilância e de atenção à saúde
devido às ocorrências de microcefalia relacionada à infecção
pelo vírus Zika, lançou também um protocolo de estimulação de
crianças com microcefalia e tem atuado como ajudante no
empenho de esforços e recursos para esta finalidade
(HENRIQUES et al., 2016).
O Governo Federal Brasileiro proporcionou algumas
estratégias para o enfrentamento à microcefalia: (1) combate ao
mosquito; (2) atendimento às pessoas; e (3) desenvolvimento
tecnológico, educação e pesquisa. Para que sejam realizadas
as referidas funções, foi implantada a Sala Nacional de
Coordenação e Controle para o Enfrentamento à Microcefalia.
O Governo anunciou que as mães de crianças com microcefalia
com renda familiar inferior a R$ 220,00 podem realizar a
inscrição para o Benefício de Prestação Continuada (BPC), com
o valor de um salário mínimo (HENRIQUES et al., 2016).
Em relação ao diagnóstico uma importante questão é
utilizar um método que permita diferenciar o Zika das demais
arboviroses. Na literatura são demonstrados alguns estudos
relacionados ao diagnóstico do vírus, a exemplo de um kit
desenvolvido pela Fundação Oswaldo Cruz (Fiocruz) que
permite realizar a identificação e diferenciação do vírus da
dengue, Zika e Chikungunya (HENRIQUES et al., 2016).
Além do desenvolvimento de estudos a cerca da
fisiopatologia, consequências neurológicas e meios

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
518
diagnósticos, a população precisa ser informada de como se
prevenir contra o vírus, por isso foi criada a revista
Epidemiologia e Serviços de Saúde: revista do Sistema Único
de Saúde do Brasil (RESS) que possui o objetivo de publicar
artigos e informações relacionados ao tema. Ajudando assim
nas ações de vigilância na saúde, como controlar o vetor Aedes
além de informações sobre microcefalia (HENRIQUES et al.,
2016).
CONCLUSÕES
O Zika vírus se expandiu de forma rápida pelo mundo,
porém como apresenta sintomas da infeção semelhantes a
outros arbovírus sua prevalência foi subestimada. A partir de
dados presentes na literatura é possível observar que o vírus
Zika possui proteínas em seu arcabouço viral que promovem
reações com proteínas do hospedeiros, essas envolvidas com
a proliferação celular e resposta Imune.
Em relação a associação do vírus com o
desenvolvimento de microcefalia, já é observado que o vírus
pode estar envolvido como a diminuição da produção e
maturação de células progenitoras neurais, além de possuir a
habilidade de acessar barreiras imunes como a barreira
hematoencefálica e placentária.
A microcefalia causada pelo vírus é caracterizada por um
malformação cerebral, no qual, o neonato afetado não possui
um cérebro desenvolvido de maneira adequada. Nesta
condição, os neonatos nascem com perímetro cefálico (PC)
menor que o normal, porém não existe um tratamento
específico para microcefalia. Mas ações de suporte podem
ajudar no desenvolvimento do bebê e da criança.

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
519
Esse auxilio é disponibilizado pelo Sistema Único da
Saúde (SUS). E devido a aparição de complicações diferentes
entre as crianças (motoras, neurológicas e respiratórias), o
acompanhamento multidisciplinar vai depender das funções
que ficarem comprometidas na criança.
REFERÊNCIAS BIBLIOGRÁFICAS ALMEIDA, Lorena Sampaio; COTA, Ana Lídia Soares; RODRIGUES, Diego Freitas. Saneamento, Arboviroses e Determinantes Ambientais: impactos na saúde urbana. Ciência & Saúde Coletiva, v. 25, p. 3857-3868, 2020. BAYER, Avraham et al. Type III interferons produced by human placental trophoblasts confer protection against Zika virus infection. Cell host & microbe, v. 19, n. 5, p. 705-712, 2016. BLÁZQUEZ, Ana-Belén; SAIZ, Juan-Carlos. Neurological manifestations of Zika virus infection. World journal of virology, v. 5, n. 4, p. 135, 2016. CALVET, Guilherme et al. Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: a case study. The Lancet infectious diseases, v. 16, n. 6, p. 653-660, 2016. CHIRAMEL, Abhilash I.; BEST, Sonja M. Role of autophagy in Zika virus infection and pathogenesis. Virus research, v. 254, p. 34-40, 2018. DE SOUZA, Helen Paredes et al. Doenças infecciosas e parasitárias no Brasil de 2010 a 2017: aspectos para vigilância em saúde. Revista Panamericana de Salud Pública, v. 44, p. e10, 2020. DONALISIO, Maria Rita; FREITAS, André Ricardo Ribas; VON ZUBEN, Andrea Paula Bruno. Arboviroses emergentes no Brasil: desafios para a clínica e implicações para a saúde pública. Revista de Saúde Pública, v. 51, p. 1-6, 2017. EICKMANN, Sophie Helena et al. Síndrome da infecção congênita pelo vírus Zika. Cadernos de Saúde Pública, v. 32, p. e00047716, 2016. FAIZAN, Md Imam et al. Zika virus-induced microcephaly and its possible molecular mechanism. Intervirology, v. 59, n. 3, p. 152-158, 2016. GOODFELLOW, Forrest et al. Strain-Dependent Consequences of Zika Virus Infection and Differential Impact on Neural Development. Viruses, v. 10, n. 10, p. 550, 2018. GOUVEIA, Nelson. Resíduos sólidos urbanos: impactos socioambientais e perspectiva de manejo sustentável com inclusão social. Ciência & saúde coletiva, v. 17, p. 1503-1510, 2012.

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
520
HENRIQUES, Cláudio Maierovitch Pessanha; DUARTE, Elisete; GARCIA, Leila Posenato. Desafios para o enfrentamento da epidemia de microcefalia. 2016. HAMEL, Rodolphe et al. Biology of Zika virus infection in human skin cells. Journal of virology, v. 89, n. 17, p. 8880-8896, 2015. HERLING, Juliana Debei et al. Infecção por Zika Vírus e nascimento de crianças com microcefalia: revisão de literatura. Revista Ciência e Estudos Acadêmicos de Medicina, v. 1, n. 05, 2016. LI, Hongda et al. The neurobiology of Zika virus. Neuron, v. 92, n. 5, p. 949-958, 2016. LIMA-CAMARA, Tamara Nunes. Emerging arboviruses and public health challenges in Brazil. Revista de saude publica, v. 50, p. 36, 2016. LUSTIG, Yaniv et al. Detection of Zika virus RNA in whole blood of imported Zika virus disease cases up to 2 months after symptom onset, Israel, December 2015 to April 2016. Eurosurveillance, v. 21, n. 26, p. 30269, 2016. MANDACARU, Samuel C. Eventos moleculares e fisiopatologia da infecção por Zika vírus. Revista Ciências da Saúde e Educação IESGO, v. 1, n. 2, 2019. MIURA, Heloisa; DA SILVA, Odenilson José; DE ALMEIDA, Vilson Rosa. Diagnóstico de alterações refracionais em crianças com microcefalia, presumidamente por síndrome congênita do Zika vírus. Research, Society and Development, v. 9, n. 10, p. e4429108674-e4429108674, 2020. MLAKAR, Jernej et al. Zika virus associated with microcephaly. New England Journal of Medicine, v. 374, n. 10, p. 951-958, 2016. NORONHA L, Zanluca C, Azevedo ML, Luz KG, Santos CN. Zika virus damages the human placental barrier and presents marked fetal neurotropism. Mem Inst Oswaldo Cruz 111: 287–293. 2016. NOWAKOWSKI, Tomasz J. et al. Expression analysis highlights AXL as a candidate Zika virus entry receptor in neural stem cells. Cell stem cell, v. 18, n. 5, p. 591-596, 2016. NUNES, Magda Lahorgue et al. Microcephaly and Zika virus: a clinical and epidemiological analysis of the current outbreak in Brazil. Jornal de pediatria, v. 92, n. 3, p. 230-240, 2016. OLAGNIER, David et al. Mechanisms of Zika virus infection and neuropathogenesis. DNA and cell biology, v. 35, n. 8, p. 367-372, 2016. PRATA‐BARBOSA, Arnaldo et al. Effects of Zika infection on growth. Jornal de Pediatria (Versão em Português), v. 95, p. 30-41, 2019. RUSSO, Fabiele Baldino; JUNGMANN, Patricia; BELTRÃO‐BRAGA, Patricia Cristina Baleeiro. Zika infection and the development of neurological defects. Cellular microbiology, v. 19, n. 6, p. e12744, 2017.

FISIOPATOLOGIA DA INFECÇÃO PELO ZIKA VÍRUS EM NEONATOS
521
SANTANA, Anne Galindo. Desenvolvimento motor de crianças portadoras da síndrome congênita do Zika vírus. Caderno de Graduação-Ciências Biológicas e da Saúde-UNIT-ALAGOAS, v. 5, n. 1, p. 131, 2018 TABATA, Takako et al. Zika virus targets different primary human placental cells, suggesting two routes for vertical transmission. Cell host & microbe, v. 20, n. 2, p. 155-166, 2016. TANG, Hengli et al. Zika virus infects human cortical neural progenitors and attenuates their growth. Cell stem cell, v. 18, n. 5, p. 587-590, 2016. TRINDADE, Adriana Gomes; LOPES, Victor de Oliveira. Microcefalia e Zika vírus no Brasil: um estudo de revisão. 2019. WANG, Anyou et al. Zika virus genome biology and molecular pathogenesis. Emerging microbes & infections, v. 6, n. 1, p. 1-6, 2017. WEN, Zhexing; SONG, Hongjun; MING, Guo-li. How does Zika virus cause microcephaly? Genes & development, v. 31, n. 9, p. 849-861, 2017.
AGRADECIMENTOS
Agradecemos a nossa orientadora Carolina Silva, pela ajuda e pela paciência com as correções que nos permitiram alcançar um melhor desempenho na nossa formação profissional. Agradecemos também, a nossa instituição Centro Universitário Unifbv por auxiliar no nosso processo de formação e por ter cedido as instalações que contribuíram para o desenvolvimento da pesquisa.

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
522
CAPÍTULO 25
PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS
ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
Rádamis Barbosa CASTOR 1
Maria Helena do NASCIMENTO 1
Krystyna Gorlach LIRA 2 1 Mestrandos do Programa de Pós-graduação em Biologia Celular e Molecular, UFPB;
2 Orientadora/Professora do DBM/UFPB. [email protected]
RESUMO: As bactérias do solo são conhecidas por produzir substâncias biologicamente ativas de apelo industrial, a exemplo das enzimas hidrolíticas extracelulares. Elas são as principais catalisadoras de processos industriais e aquelas de origem microbiana oferecem uma série de vantagens frente às produzidas por plantas e animais. Em virtude da demanda crescente, a procura por novos microrganismos produtores de enzimas hidrolíticas apresenta um elevado interesse biotecnológico. Desse modo, o objetivo do trabalho foi avaliar a produção de enzimas extracelulares (proteases, celulases, amilases e lipases) por 20 isolados de bactérias provenientes de um solo tratado com óleo vegetal residual. Para tal, utilizou-se meios sólidos contendo gelatina, carboximetilcelulose, amido e óleo vegetal residual para verificação da produção de proteases, celulases, amilases e lipases, respectivamente, pelos isolados. Pôde-se constatar que 85% dos isolados produziram, pelo menos, duas classes de enzimas extracelulares. A maioria das bactérias testadas foi capaz de

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
523
produzir lipases (17 isolados), proteases (14 isolados) e celulases (13 isolados). Amilases foram produzidas por apenas cinco isolados. As bactérias Gram-positivas formadores de endósporos foram as mais ativas na produção de todas as enzimas analisadas, com destaque para o isolado O27. Os resultados obtidos indicam que alguns isolados de bactérias analisados apresentam um potencial a ser explorado na produção de enzimas extracelulares. Palavras-chave: Biotecnologia. Bactérias. Enzimas extracelulares. INTRODUÇÃO
O solo enquanto habitat é o ambiente mais complexo da
Terra e estima-se conter um terço de toda a vida do planeta. É
definido como a totalidade de seres vivos que o habitam
(plantas, animais e microrganismos) e seu ambiente abiótico
(rochas e minerais, água e ar). Um grama de solo pode conter
quilômetros de hifas fúngicas e mais de 109 células de bactérias
e arqueias, além de protozoários e vírus em menor proporção
(VORONEY; HECK, 2015; FIERER, 2017). Baseado na
literatura científica, Fierer (2017) aproximou o conteúdo de
biomassa microbiana do solo em torno de uma tonelada por
hectare, com predominância de fungos e bactérias,
corroborando o fato de que a pedosfera mantém uma rica
biodiversidade de microrganismos.
Estes são considerados os principais agentes
decompositores da matéria orgânica do solo, participando de
processos essenciais como a ciclagem de nutrientes, dentre os
quais, carbono, nitrogênio e fósforo (PLANTE et al., 2015;
LLADÓ et al., 2017). Isso se deve ao fato de microrganismos
secretarem enzimas extracelulares capazes de converter

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
524
biopolímeros complexos do meio em nutrientes assimiláveis
para manutenção do seu metabolismo e crescimento celular
(LUO et al., 2017). Allison et al. (2010) propôs que a seleção
natural tende a promover mecanismos que minimizem os
custos empregados e maximizem os benefícios associados à
produção enzimática, de modo que, a síntese de enzimas
extracelulares requeira apenas uma parcela muito pequena do
carbono e nitrogênio assimilados do meio (1-5%). Desta forma,
a produção de enzimas extracelulares se mostra um
mecanismo valioso à sobrevivência dos microrganismos que
habitam o solo por aumentar a disponibilidade de nutrientes
minerais e orgânicos de baixo peso molecular a um custo
pequeno para a célula (BURNS et al., 2013).
Enzimas extracelulares, como relatado, são produzidas
por uma ampla variedade de microrganismos do solo.
Pertencentes às classes das hidrolases e oxirredutases
(PLANTE et al., 2015), elas garantem a sobrevivência dos
microrganismos e desempenham papéis ecológicos vitais.
Hidrolases catalisam reações de clivagem pela adição de água
a ligações covalentes específicas, como C-O, C-N, O-P e C-C
e são consideradas as enzimas de maior importância industrial
(BANERJEE; RAY, 2017; LIU; KOKARE, 2017; SINGH et al.,
2019). Segundo Li et al. (2012), 75% das enzimas utilizadas na
indústria são hidrolíticas e aproximadamente 90% delas são de
origem microbiana (GUERRAND, 2018).
Desse total, aproximadamente 64% são produzidas por
fungos, 26% por bactérias, 6% por animais e 4% por plantas
(KAUR; GIL, 2019). A preferência por enzimas hidrolíticas
microbianas decorre de uma série de vantagens: elas são mais
ativas e estáveis que suas análogas animais e vegetais,

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
525
podendo ser produzidas em larga escala, dependendo do
microrganismo escolhido; são secretadas extracelularmente, o
que facilita o processo de purificação; apresentam elevada
especificidade pelos substratos, sejam eles orgânicos ou
sintéticos; possuem baixo custo de produção; não apresentam
toxicidade; podem ser reutilizadas e são ecologicamente
sustentáveis. Por último, microrganismos dispõem de facilidade
no manejo, multiplicam-se rapidamente sob condições
controladas e são suscetíveis à manipulação genética, além de
serem naturalmente uma rica fonte de enzimas hidrolíticas (LIU;
KOKARE, 2017; KAUR; GIL, 2019; SINGH et al., 2019; SINGH;
KUMAR, 2019).
Segundo previsão do relatório “Industrial Enzymes – A
Global Market Overview” (2018) publicado pela empresa
Research and Markets, o mercado mundial de enzimas deve
movimentar cerca de 7,7 bilhões de dólares até 2024. Ademais,
foi apontado que a indústria de alimentos e bebidas foi o setor
que mais utilizou enzimas no ano de 2017, movimentando 1,4
bilhões de dólares, seguido pela indústria de biocombustíveis
(969,3 milhões) e de sabões e detergentes (754,4 milhões).
Proteases, celulases, amilases e lipases estão entre as
enzimas hidrolíticas mais utilizadas na indústria (LIU; KOKARE,
2017; GUERRAND, 2018).
Proteases são enzimas que catalisam a hidrólise de
proteínas pela quebra de suas ligações peptídicas. Classificam-
se quanto ao mecanismo de ação em endopeptidases (EC
3.4.21 – 3.4.24, 3.4.99) e exopeptidases (EC 3.4.11 – 3.4.19).
Endopeptidases clivam ligações peptídicas de aminoácidos
localizados no interior das cadeias polipeptídicas, enquanto
exopeptidases possuem afinidade pelas regiões aminoterminal

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
526
e carboxiterminal de aminoácidos localizados nas extremidades
de tais cadeias (CONTESINI et al., 2017; SHARMA et al., 2017;
GURUMALLESH et al., 2019).
Proteases contribuem com aproximadamente 60% do
mercado mundial de enzimas hidrolíticas, sendo usadas em sua
maioria na indústria de detergentes (GLOBAL MARKET
INSIGHTS, 2017). Algumas bactérias do gênero Bacillus
secretam uma grande quantidade de proteases alcalinas (mais
de 20 g/L), sendo as maiores produtoras destas enzimas em
nível comercial (CONTESINI et al., 2017). Algumas de suas
aplicações envolvem a formulação de detergentes para
remoção de manchas em roupas; remoção de pelos em
carcaças de couro de animais, melhorando sua resistência e
qualidade, além de permitir o reaproveitamento dos pelos para
confecção de fibras sintéticas, aminoácidos para a indústria
farmacêutica e de cosméticos, biogás e biofertilizantes.
Também são muito utilizadas pela indústria de alimentos para
o amaciamento de carnes, hidrólise do glúten, produção e
maturação de queijos. Na indústria farmacêutica, proteases são
empregadas na produção de peptídeos bioativos com ação
antioxidante, antimicrobiana, anti-hipertensiva e antitumoral,
por exemplo (BANERJEE; RAY, 2017; CONTESINI et al., 2017;
GURUMALLESH et al., 2019).
Celulases são enzimas que catalisam a hidrólise da
celulose, o biopolímero mais abundante na superfície da Terra.
Ela é formada pela união de moléculas de glicose através de
ligações glicosídicas do tipo β-1,4, criando cadeias com 15.000
- 25.000 unidades de glicose, cada. Estas, se alinham
paralelamente umas às outras, por meio de ligações de
hidrogênio e interações de van der Waals, formando uma

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
527
estrutura paracristalina, com regiões amorfas e cristalinas
alternadas (MOHITE; PATIL, 2016; BEHERA et al., 2017).
Em virtude dessa complexidade, a hidrólise completa da
celulose é feita por meio da ação conjunta de três classes de
enzimas, nomeadas de acordo com seu mecanismo de ação e
região do substrato que catalisam: endoglucanases (EC
3.2.1.4), exoglucanases (EC 3.2.1.74) e celobiohidrolases (EC
3.2.1.91) e β-glicosidases (EC 3.2.1.21) (MOHITE; PATIL,
2016; SHARMA; YAZDANI, 2016). Endoglucanases catalisam
a hidrólise de ligações glicosídicas das regiões amorfas da
molécula de celulose, enquanto exoglucanases possuem
afinidade pelas ligações glicosídicas das regiões cristalinas,
clivando-as. A ação conjunta dessas duas enzimas leva à
formação de moléculas de celobiose, que quebradas pelas β-
glicosidases, liberam glicose (MOHITE; PATIL, 2016).
Celulases são produzidas por uma ampla variedade de
microrganismos e apresentam elevado potencial biotecnológico
para uso em uma série de processos industriais. Na indústria
de alimentos, podem ser utilizadas para extração e clarificação
de sucos de frutas e vegetais, reduzindo a viscosidade e
aumentando a estabilidade dos mesmos. Na indústria têxtil, são
comumente empregadas no tratamento da lã e amaciamento do
algodão, além da fabricação de jeans com aparência desbotada
ou envelhecida. Nas biorrefinarias, enzimas celulolíticas são
usadas na conversão de biomassa celulósica em açúcares
fermentescíveis para produção de bioetanol. Além disso,
celulases são amplamente utilizadas na fabricação de papel,
detergentes e ração animal (BEHERA et al., 2016; MOHITE;
PATIL, 2016; LIU; KOKARE, 2017).

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
528
Amido é o segundo polissacarídeo mais abundante na natureza
depois da celulose. Produzido pelas plantas, é armazenado em
frutas, sementes, rizomas e tubérculos na forma de grânulos
semicristalinos compostos por dois tipos de molécula: amilose
e amilopectina. Amilose é um polímero linear, insolúvel em
água, formado por 100-10.000 unidades de glicose unidas por
ligações glicosídicas do tipo α-1,4 e compõe cerca de 20-27%
do amido. Amilopectina é um polímero ramificado, solúvel em
água, formado por cadeias lineares de 10-60 unidades de
glicose unidas por ligações do tipo α-1,4, ligadas à cadeias de
15-45 unidades de glicose unidas por ligações do tipo α-1,6 e
compõe aproximadamente 75-85% do amido (SINDHU et al.,
2016; MOHANAN; SATYANARAYANA, 2018).
As enzimas que catalisam a hidrólise do amido são
chamadas de amilases e são classificadas de acordo com seu
mecanismo de ação em α-amilases (EC 3.2.1.1), β-amilases
(EC 3.2.1.2) e glicoamilases (EC 3.2.1.3). As α-amilases clivam
ligações glicosídicas do tipo α-1,4 de forma aleatória no interior
das moléculas de amido, sendo, portanto, endoamilases. As β-
amilases também hidrolisam as ligações glicosídicas do tipo α-
1,4, mas o fazem nas extremidades das moléculas de amido,
sendo, deste modo, exoamilases. Por fim, as glicoamilases são
as únicas enzimas capazes de quebrar ligações glicosídicas do
tipo α-1,4 e α-1,6 e também são do tipo exoamilases (ABD-
ELHALEM et al., 2015; MOHANAN; SATYANARAYANA, 2018).
Amilases são produzidas por uma grande variedade de
organismos, vista a importância nutritiva do amido na natureza
(SINDHU et al., 2016). Dentre as bactérias produtoras, aquelas
pertencentes ao gênero Bacillus são as mais importantes (LIU;
KOKARE, 2017; SINGH et al., 2019). Amilases representam 15-

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
529
25% do mercado mundial de enzimas hidrolíticas (MOHANAN;
SATYANARAYANA, 2018; SINGH et al., 2019), apresentando
inúmeras aplicações industriais, tais qual seu uso na
formulação de detergentes junto com outras hidrolases. Na
indústria têxtil, o amido é usado para proteção dos fios durante
o processamento dos tecidos, sendo posteriormente removido
com auxílio de enzimas amilolíticas. O amido também é
utilizado para revestimento do papel nas indústrias de papel e
celulose, melhorando sua qualidade e capacidade de
apagamento. Para isso, α-amilases são empregadas na
diminuição de sua viscosidade antes de vir a revesti-lo. Por fim,
a indústria de alimentos e bebidas emprega α-amilases e
glicoamilases para produção de glicose, frutose e
oligossacarídeos, utilizados na grande maioria dos alimentos e
bebidas industrializados (SINDHU et al., 2016; MOHANAN;
SATYANARAYANA, 2018).
Lipases (EC 3.1.1.3) são enzimas que catalisam a
hidrólise das ligações éster de triacilgliceróis em meio aquoso
(interface óleo-água) liberando moléculas de diacilglicerol,
monoacilglicerol, ácidos graxos livres e glicerol (FERRAZ et al.,
2018; JAVED et al., 2018). A atividade enzimática das lipases
depende de três fatores principais: posição do ácido graxo no
esqueleto de glicerol, comprimento da cadeia do ácido graxo e
presença de insaturações. Desta forma, lipases apresentam
regiosseletividade, quimiosseletividade e enantiosseletividade
(BORRELLI; TRONO, 2015; FERRAZ et al., 2018).
Lipases são as enzimas hidrolíticas mais utilizadas
industrialmente depois de proteases e amilases, ocupando a
terceira colocação em vendas no mercado mundial de enzimas
(HASSAN et al., 2006; ÜLKER et al., 2011). Fungos dos

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
530
gêneros Rhizopus, Aspergillus, Penicillium, Geotrichum, Mucor,
Rhizomucor e bactérias dos gêneros Bacillus, Pseudomonas e
Burkholderia são os principais produtores de lipases
comerciais. A indústria de detergentes é o setor que mais
emprega enzimas lipolíticas, consumindo aproximadamente
1000 toneladas de lipases todos os anos (HASSAN et al.,
2006). Isso decorre de sua capacidade de hidrolisar moléculas
de gordura, favorecendo a solubilização das mesmas em água,
e desta forma, facilitar o processo de limpeza. Na indústria de
alimentos, são utilizadas para obtenção de álcoois e ácidos
graxos de cadeia curta, a partir da hidrólise de óleos e gorduras
presentes em diversos alimentos, conferindo aroma e sabor
característicos. Outras aplicações envolvem a produção de
cosméticos, síntese de medicamentos, tratamento do couro e
produção de biodiesel (HASAN et al., 2006; LIU; KOKARE,
2017; JAVED et al., 2018).
Em virtude dos fatos mencionados, a busca por novos
microrganismos produtores de enzimas hidrolíticas
extracelulares apresenta elevado potencial biotecnológico.
Assim sendo, o objetivo deste trabalho foi avaliar a produção de
enzimas extracelulares (proteases, celulases, amilases e
lipases) por bactérias isoladas de um solo utilizado para o
descarte experimental de óleo vegetal residual com o intuito de
selecionar os isolados mais promissores para produção de
enzimas em larga escala.

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
531
MATERIAL E MÉTODOS
Microrganismos
Analisou-se vinte isolados bacterianos da coleção de
microrganismos do Laboratório de Biologia de Microrganismos
(BIOMICRO) no Departamento de Biologia Molecular (DBM) da
Universidade Federal da Paraíba (UFPB). Os isolados foram
provenientes do solo de uma área experimental tratada com
óleo vegetal de fritura e caracterizados na base de coloração de
Gram e formação de endósporos (GONÇALVES; GORLACH
LIRA, 2018).
Atividade Enzimática de Bactérias
Os isolados foram testados quanto às atividades
proteolítica, celulolítica, amilolítica e lipolítica utilizando meios
de cultura sólidos contendo os respectivos substratos para cada
enzima: gelatina, carboximetilcelulose (CMC), amido e óleo de
cozinha residual. Os experimentos descritos nos itens 3.2.1 -
3.2.4 foram realizados em triplicata e os isolados foram
previamente incubados, para crescimento, em meio sólido ágar
nutriente por 48h a 30ºC. Ao final dos testes, os halos de
degradação dos substratos foram medidos em mm e os
resultados expressos em média aritmética.
Atividade Proteolítica
Para detecção de proteases, foi usado o meio descrito
por Smibert e Krieg (1994) contendo 20,0 g/L de gelatina; 0,1

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
532
g/L de NaCl; 0,5 g/L de K2HPO4; 0,2 g/L de MgSO4.7H2O; 0,19
g/L de CaCl2.6H2O; um traço de FeSO4 e 15 g/L de ágar
bacteriológico (Kasvi); pH 7,0. Os isolados foram incubados por
48h a 30ºC e em seguida, foi adicionada uma alíquota da
solução de Frazier (12,0 g de HgCl2; 16 ml de HCl concentrado
e 80 ml de água destilada). Esta, desnatura as proteínas da
gelatina, que precipitam, dando ao meio uma coloração
esbranquiçada e revelando os halos resultantes de sua
hidrólise.
Atividade Celulolítica
A produção de celulases foi verificada através do meio
desenvolvido por Teather e Wood (1982) contendo 1,0 g/L de
CMC; 0,5 g/L de NaNO3; 1,0 g/L de K2HPO4; 0,5 g/L de
MgSO4.7H2O; 0,01g/L de FeSO4.7H2O; 1,0 g/L de extrato de
levedura (Kasvi) e 15,0 g/L de ágar bacteriológico; pH 7,0. Após
96h de incubação a 30ºC, os halos resultantes da hidrólise do
substrato foram revelados pela adição da solução de Lugol (1,0
g de I2; 2,0 g de KI e 300 ml de água destilada).
Atividade Amilolítica
Para detecção de amilases, utilizou-se o meio proposto
por Smibert e Krieg (1994) contendo 2,0 g/L de amido; 0,5 g/L
de NaNO3; 1,0 g/L de K2HPO4; 0,01 g/L de MgSO4.7H2O e 15
g/L de ágar bacteriológico; pH 7,0. Os isolados foram incubados
por 96h a 30ºC e a formação de halos, decorrente da hidrólise
do amido, foi verificada pela adição da solução de Lugol (ver
item 3.2.2) diluída cinco vezes.

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
533
Atividade Lipolítica
Utilizou-se, para produção de lipases, uma adaptação do
meio descrito por Kouker e Jaeger (1987), composto pelo
corante Rodamina B (0,050 g de Rodamina B; 50 ml de água
destilada), emulsão de óleo (30 ml de óleo de cozinha residual;
50 ml de água destilada e 250 µL de Tween 80) e meio básico
[6,0 g de caldo nutriente (HiMedia); 1,25 g de extrato de
levedura (HiMedia); 10,0 g de ágar bacteriológico e 450 ml de
água destilada)]; pH 7,0. Após 96 h de incubação a 30ºC, a
atividade lipolítica foi verificada pela formação de halos
alaranjados e fluorescentes através da irradiação das placas
por luz ultravioleta (UV) com comprimento de onda de 350
nanômetros (nm).
RESULTADOS E DISCUSSÃO
Análises de atividade enzimática dos 20 isolados
bacterianos indicaram que 85% dos isolados produziram, pelo
menos, duas classes de enzimas hidrolíticas (Tabelas 1 e 2).
Os isolados O16, O27 e O65 (bastonetes Gram positivos
formadores de endósporos) se destacaram por degradar todos
os substratos utilizados, ou seja, gelatina, carboximetilcelulose,
amido e óleo de cozinha residual (Tabela 1). Apenas dois
isolados, O10 e O20 (bastonetes Gram negativos), não foram
capazes de produzir as enzimas analisadas.
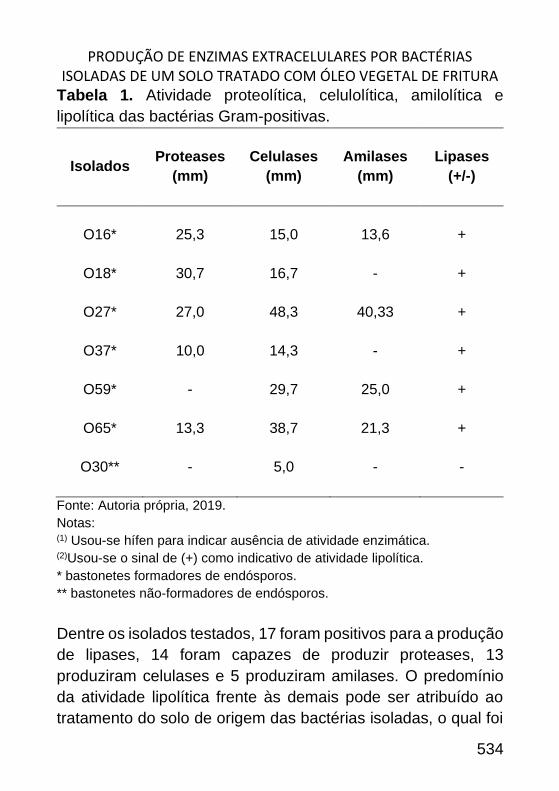
PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
534
Tabela 1. Atividade proteolítica, celulolítica, amilolítica e
lipolítica das bactérias Gram-positivas.
Isolados
Proteases
(mm)
Celulases
(mm)
Amilases
(mm)
Lipases
(+/-)
O16*
25,3
15,0
13,6
+
O18*
30,7
16,7
-
+
O27*
27,0
48,3
40,33
+
O37*
10,0
14,3
-
+
O59*
-
29,7
25,0
+
O65*
13,3 38,7 21,3 +
O30**
-
5,0
-
-
Fonte: Autoria própria, 2019.
Notas: (1) Usou-se hífen para indicar ausência de atividade enzimática. (2)Usou-se o sinal de (+) como indicativo de atividade lipolítica.
* bastonetes formadores de endósporos.
** bastonetes não-formadores de endósporos.
Dentre os isolados testados, 17 foram positivos para a produção
de lipases, 14 foram capazes de produzir proteases, 13
produziram celulases e 5 produziram amilases. O predomínio
da atividade lipolítica frente às demais pode ser atribuído ao
tratamento do solo de origem das bactérias isoladas, o qual foi

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
535
utilizado para o descarte experimental de óleo vegetal residual
durante três anos, com o objetivo de enriquecimento do mesmo
para seleção de microrganismos produtores de lipases
(GONÇALVES; GORLACH LIRA, 2018).
O óleo vegetal residual se mostrou um bom indutor da
atividade lipolítica das bactérias testadas e um dos isolados de
bactérias Gram-positivas (O27) apresentou alta atividade
lipolítica evidenciada pela presença de halos alaranjados de
forte intensidade (Figura 1). Bharathi et al. (2018) afirma que
óleos vegetais e derivados do petróleo são bons estimulantes
da atividade lipolítica de microrganismos, corroborando os
resultados apresentados. Microrganismos produtores de
lipases têm sido encontrados em lixos de resíduos
agroindustriais, esgotos de indústrias alimentícias, cursos
d’água e solos contaminados por óleos vegetais e derivados do
petróleo.
No que diz respeito à atividade proteolítica, segunda
mais frequente entre os isolados testados, destacaram-se
aqueles cuja média dos halos de degradação da gelatina
ultrapassou 20 mm de diâmetro: O2, O5, O6 (bastonetes Gram-
negativos), O16, O18 e O27 (bastonetes Gram-positivos
formadores de endósporos), como elencado nas Tabelas 1 e 2
e apresentado na Figura 2. Além da gelatina, outros substratos
proteicos têm sido utilizados para triagem de microrganismos
produtores de proteases, tais como: caseína, albumina,
elastina, queratina, extrato de levedura, peptona, leite
desnatado, extratos de soja, carne e peixe (SHARMA et al.,
2017). A exemplo disso, os ensaios realizados por Pant et al.
(2015) apontaram a gelatina como substrato de escolha para a
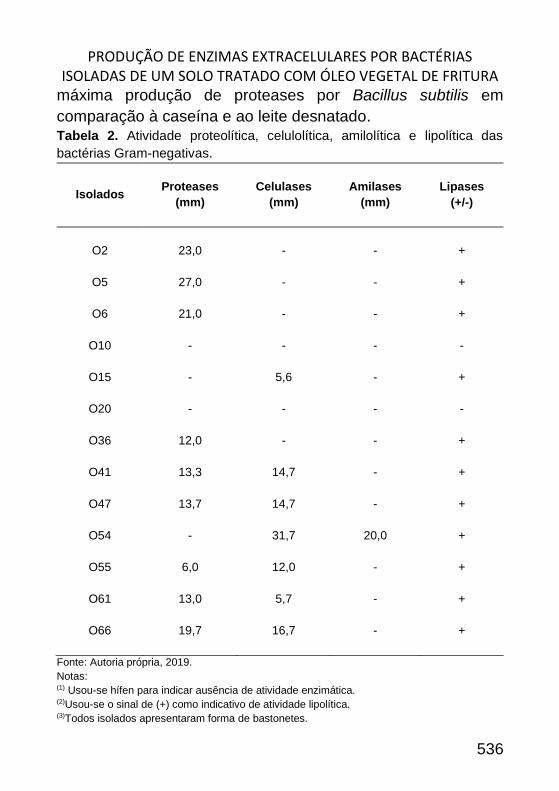
PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
536
máxima produção de proteases por Bacillus subtilis em
comparação à caseína e ao leite desnatado. Tabela 2. Atividade proteolítica, celulolítica, amilolítica e lipolítica das
bactérias Gram-negativas.
Isolados
Proteases
(mm)
Celulases
(mm)
Amilases
(mm)
Lipases
(+/-)
O2
23,0
-
-
+
O5
27,0
-
-
+
O6
21,0
-
-
+
O10
-
-
-
-
O15
-
5,6
-
+
O20
-
-
-
-
O36
12,0
-
-
+
O41
13,3
14,7
-
+
O47
13,7
14,7
-
+
O54
-
31,7
20,0
+
O55
6,0
12,0
-
+
O61
13,0
5,7
-
+
O66
19,7
16,7
-
+
Fonte: Autoria própria, 2019.
Notas: (1) Usou-se hífen para indicar ausência de atividade enzimática. (2)Usou-se o sinal de (+) como indicativo de atividade lipolítica. (3)Todos isolados apresentaram forma de bastonetes.

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
537
Observou-se que 65% dos isolados foram capazes de
degradar a carboximetilcelulose e que as linhagens O27 e O65
(bastonetes Gram-positivos) apresentaram elevado potencial
de hidrolise desse substrato, indicado pelas zonas de
degradação de 48,3 e 38,6 mm de diâmetro, respectivamente
(Tabela 1). Em virtude do alto grau de polimerização e da baixa
cristalinidade, a CMC é o substrato preferencial para triagem de
microrganismos secretores de endoglucanases, enzimas que
iniciam o processo de hidrólise da celulose (FLORENCIO,
2011). Por sua vez, exoglucanases e ß-glicosidases não
possuem especificidade pela CMC, e desta forma, faz-se
necessária a realização de ensaios complementares utilizando
substratos específicos para as mesmas, como a celulose
cristalina (Avicel) e a celobiose (BEHERA et al., 2016).
Fungos filamentosos são os principais produtores das
enzimas celulolíticas de importância industrial, mas apesar
disso, bactérias também são uma importante fonte de celulases
e embora menos estudadas, exibem uma série de vantagens
como rápido crescimento, estabilidade celular e secreção de
enzimas celulolíticas estáveis, altamente específicas e que
atuam de modo sinérgico para despolimerização da celulose
(SHARMA; YAZDANI, 2016; PATEL et al., 2019).
O amido foi a fonte de carbono menos utilizada pelas
bactérias, como observado nas Tabelas 1 e 2. Entretanto,
dentre os cinco isolados com atividade amilolítica, quatro
exibiram halos de degradação superiores a 20 mm de diâmetro,
todos pertencentes ao biotipo bastonetes Gram-positivos
formadores de endósporos, com destaque para O27, que
apresentou o maior halo de hidrólise do amido - 40,3 mm de
diâmetro (Figura 4). Amido, amilopectina, maltose, galactose,
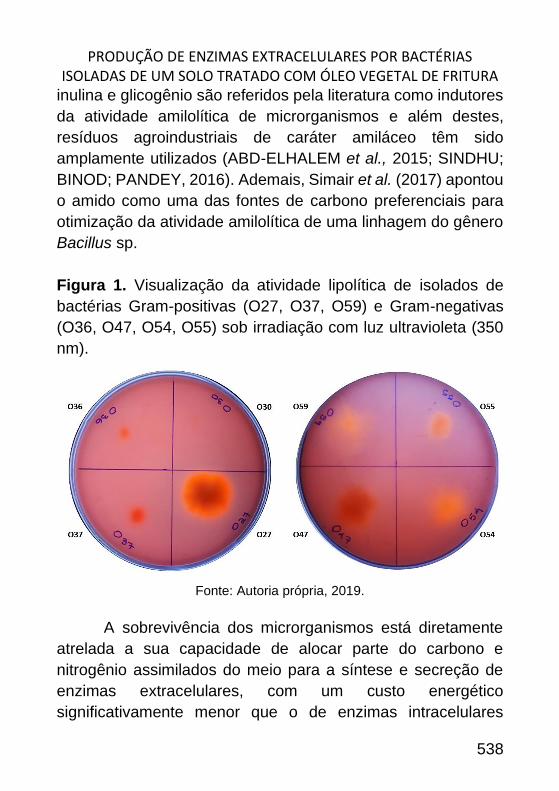
PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
538
inulina e glicogênio são referidos pela literatura como indutores
da atividade amilolítica de microrganismos e além destes,
resíduos agroindustriais de caráter amiláceo têm sido
amplamente utilizados (ABD-ELHALEM et al., 2015; SINDHU;
BINOD; PANDEY, 2016). Ademais, Simair et al. (2017) apontou
o amido como uma das fontes de carbono preferenciais para
otimização da atividade amilolítica de uma linhagem do gênero
Bacillus sp.
Figura 1. Visualização da atividade lipolítica de isolados de
bactérias Gram-positivas (O27, O37, O59) e Gram-negativas
(O36, O47, O54, O55) sob irradiação com luz ultravioleta (350
nm).
Fonte: Autoria própria, 2019.
A sobrevivência dos microrganismos está diretamente
atrelada a sua capacidade de alocar parte do carbono e
nitrogênio assimilados do meio para a síntese e secreção de
enzimas extracelulares, com um custo energético
significativamente menor que o de enzimas intracelulares

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
539
(BURNS et al., 2013). Desta forma, a habilidade de utilizar mais
de um substrato, como observado, pode ser vista como uma
forma de prevenir a desnutrição e consequente morte celular e
se deve ao fato de bactérias poderem detectar a presença de
moléculas-sinal do ambiente, que podem indicar, entre outras
coisas, o tipo, localização, qualidade e quantidade dos
substratos disponíveis, num fenômeno conhecido como quorum
sensing (GOO et al., 2015).
Figura 2. Atividade proteolítica dos isolados de bactérias Gram-
positivas (O16, O18, O27) e Gram-negativas (O36).
Fonte: Autoria própria, 2019.
Nota: As setas indicam as zonas de hidrólise da gelatina.
Figura 3. Atividade celulolítica dos isolados de bactérias Gram-
positivas (O27, O30, O37, O59) e Gram-negativas (O41, O47,
O54, O55.

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
540
Fonte: Autoria própria, 2019.
Nota: As setas indicam as zonas de hidrólise da carboximetilcelulose.
Figura 4. Atividade amilolítica dos isolados de bactérias Gram-
positivas O27 e O65.
Fonte: Autoria própria, 2019.
Nota: As setas indicam as zonas de hidrólise do amido.

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
541
CONCLUSÃO
A maioria dos isolados bacterianos testados apresentou
produção de enzimas extracelulares. As atividades lipolítica,
proteolítica e celulolítica foram as mais comuns. Os bastonetes
Gram-positivos formadores de endósporos foram mais
expressivos na produção das hidrolases testadas com destaque
para o isolado O27, capaz de produzir proteases, amilases,
celulases e lipases.
REFERÊNCIAS BIBLIOGRÁFICAS ABD-ELHALEM, B.T. et al. Production of amylases from Bacillus amyloliquefaciens under submerged fermentation using some agro-industrial by-products. Annals of Agricultural Sciences, v. 60, n. 2, p. 193-202, 2015. ALLISON, S. D.; WEINTRAUB, M. N.; GARTNER, T. B.; WALDROP, M. P. Evolutionary-Economic Principles as Regulators of Soil Enzyme Production and Ecosystem Function. In: SHUKLA, G. e VARMA, A. (Ed.). Soil Enzymology. Berlin, Heidelberg: Springer Berlin Heidelberg, 2011. p. 229-243. BANERJEE, G.; RAY, A.K. Impact of microbial proteases on biotechnological industries. Biotechnology and Genetic Engineering Reviews, v. 33, n. 2, p. 119-143, 2017. BEHERA, B.C. et al. Microbial cellulases – Diversity & biotechnology with reference to mangrove environment: A review. Journal of Genetic Engineering and Biotechnology, v. 15, n. 1, p. 197-210, 2016. BHARATHI, D.; RAJALAKSHMI, G.; KOMATHI, S. Optimization and production of lipase enzyme from bacterial strains isolated from petrol spilled soil. Journal of King Saud University - Science, v. 31, n. 4, p. 898-901, 2019.
BORRELLI, G.M.; TRONO, D. Recombinant lipases and phospholipases and their use as biocatalysts for industrial applications. International Journal of Molecular Sciences, v. 16, n. 9, p. 20774-20840, 2015. BURNS, R. G.; DEFOREST, J. L.; MARXSEN, J.; SINSABAUGH, R. L. et al. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biology and Biochemistry, 58, p. 216-234.

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
542
GONÇALVES, L.; LIRA, K. Lipases and biosurfactants production by the newly isolated Burkholderia sp. Brazilian Journal of Biological Sciences, v. 5, p. 57-68, 2018.
CONTESINI, F. J.; MELO, R.G.; SATO, H.H. An overview of Bacillus proteases: from production to application. Critical Reviews in Biotechnology, p. 321-334, 2017. FERRAZ, J.L.A.A. et al. Obtenção de lipases microbianas: Uma breve revisão. Revista Ciências Exatas e Naturais, v. 20, n. 1, p. 30-53, 2018. FIERER, N. Embracing the unknown: disentangling the complexities of the soil microbiome. Nature Reviews Microbiology, v.15, n. 10, p. 579-590, 2017.
FLORENCIO, C. Microrganismos Produtores de Celulases : Seleção de Isolados de Trichoderma spp. 2011. 86 f. Dissertação de Mestrado (Mestrado em Biotecnologia). Universidade Federal de São Carlos, 2012. GOO, E. et al. Control of bacterial metabolism by quorum sensing. Trends in Microbiology, v. 23, n. 9, p. 567-576, 2015. GUERRAND, D. Economics of food and feed enzymes: status and prospectives. In: KUMAR, V.; NUNES, C. Enzymes in Human and Animal Nutrition. Academic Press, 2018. p. 487-514. GURUMALLESH, P. et al. A systematic reconsideration on proteases. International Journal of Biological Macromolecules, v. 128, p. 254-267, 2019. HASAN, F.; SHAH, A.A.; HAMEED, A. Methods for detection and characterization of lipases: A comprehensive review. Biotechnology Advances, v. 27, n. 6, p. 782-798, 2009. Industry Experts. Industrial enzymes – a global market overview. IE, 2018. Disponível em: < http://industry-experts.com/verticals/biotechnology/industrial-enzymes-a-global-market overview. Acesso em: 25 ago. 2020.
JAVED, S. et al. Bacterial lipases: A review on purification and characterization. Progress in Biophysics and Molecular Biology, v. 132, p. 23-34, 2017. KAUR, H.; GILL, P.K. Microbial enzymes in food and beverages processing. In: GRUMEZESCU, A.; HOLBAN, A-M. Engineering Tools in the Beverage Industry. 1. ed. Woodhead Publishing, 2019. p. 255-282. KOUKER, G.; JAEGER, K.E. Specific and sensitive assay for bacterial lipases. Applied and Environmental Microbiology, v. 53, n. 1, p.211-213, 1987.

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
543
LI, S. et al. Technology prospecting on enzymes: application, marketing and engineering. Computational and Structural Biotechnology Journal, v. 2, n. 3, p. 1-11, 2012. LIU, X.; KOKARE, C. Microbial enzymes of use in industry. In: BRAHMACHARI, G. Biotechnology of Microbial Enzymes. Academic Press, 2016. p. 267-298. LUO, L.; MENG, H.; GU, J.-D. Microbial extracellular enzymes in biogeochemical cycling of ecosystems. Journal of Environmental Management, v. 197, p. 539-549, 2017.
MOHANAN, N.; SATYANARAYANA, T. Amylases. Reference Module in Life Sciences, p. 1-20, 2018. MOHITE, B.B.; PATIL, S.V. Impact of microbial cellulases on microbial cellulose biotechnology. In: GUPTA, V. New and future developments in microbial biotechnology and bioengineering: microbial cellulase system properties and applications. 1. ed. Elsevier, 2016. p. 31-40. PANT, G.; PRAKASH, A.; PAVANI, J. V. P.; BERA, S. et al. Production, optimization and partial purification of protease from Bacillus subtilis. Journal of Taibah University for Science, v.9, n. 1, p. 50-55, 2015. PATEL, A.K. et al. Thermostable cellulases: Current status and perspectives. Bioresource Technology, v. 279, p. 385-392, 2019.
PLANTE, A.F.; STONE, M.S.; MCGILL, W.B. The metabolic physiology of soil microorganisms. In: PAUL, E. Soil Microbiology, Ecology and Biochemistry. 4. ed. Academic Press, 2015. p. 245-272. SHARMA, P. et al. Purification and characterization of lipase by Bacillus methylotrophicus PS3 under submerged fermentation and its application in detergent industry. Journal of Genetic Engineering and Biotechnology, v. 15, n. 2, p. 369-377, 2017. SHARMA, S.; YAZDANI, S.S. Diversity of microbial cellulase system. In: GUPTA, V. New and future developments in microbial biotechnology and bioengineering: microbial cellulase system properties and applications. 1. ed. Elsevier, 2016. p. 49-64. SIMAIR, A.H. et al. Production and partial characterization of α-amylase enzyme from Bacillus sp. BCC 01-50 and potential applications. Biomedical Research International, p. 1-10. SINDHU, R.; BINOD, P.; PANDEY, A. α-amylases. In: PANDEY, A.; NEGI, S.; SOCCOL, C. Current developments in biotechnology and bioengineering: production, isolation and purification of industrial products. 1. ed. Elsevier, 2016. p. 3-24, 2016. SINDHU, R.; BINOD, P.; PANDEY, A. α-amylases. In: PANDEY, A.; NEGI, S.; SOCCOL, C. Current developments in biotechnology and

PRODUÇÃO DE ENZIMAS EXTRACELULARES POR BACTÉRIAS ISOLADAS DE UM SOLO TRATADO COM ÓLEO VEGETAL DE FRITURA
544
bioengineering: production, isolation and purification of industrial products. 1. ed. Elsevier, 2016. p. 3-24, 2016. SINGH, P.; KUMAR, S. Microbial enzyme in food biotechnology. In: SINGH, R. S. et al. Advances in enzyme technology. 1. ed. Science Direct, 2019. p. 19-28. SMIBERT, R.M.; KRIEG, N.R. Phenotypic characterization. In: Gerhardt, P. et al. Methods for General and Molecular Bacteriology. Washington DC, 1994. p. 607-654. TEATHER, R.M.; WOOD, P.J. Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Applied and Environmental Microbiology, v. 43, n. 4, p. 777-780, 1982. ÜLKER, S. et al. Isolation, production, and characterization of an extracellular lipase from Trichoderma harzianum isolated from soil. Turkish Journal of Biology, v. 35, p. 543-550, 2010. VORONEY, R. P.; HECK, R. J. The Soil Habitat. In: PAUL, E. A. (Ed.). Soil Microbiology, Ecology and Biochemistry. 4. ed. Boston: Academic Press, 2015. p. 15-39.

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
545
CAPÍTULO 26
TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
Shayenne Eduarda Ramos VANDERLEY 1,2
Wilias Silva Santos GREISON 1,2
Anne Beatriz Cunha BARROS 1,2
Matheus Soares da Silva MELO 1,2
Sildivane Valcácia SILVA 1,3
1 Laboratório de Biotecnologia em Reprodução Animal, CBIOTEC/ UFPB; 2 Graduandos do curso de Biotecnologia, UFPB; 3 Orientadora/Professora do Departamento de
Biotecnologia/UFPB. [email protected]
RESUMO: Vírus respiratórios são os agentes causadores de doenças mais frequentes em humanos e animais, com impacto significativo na morbidade e mortalidade em todo o mundo. Os agentes respiratórios comuns de várias famílias de vírus estão bem adaptados à transmissão eficiente de indivíduo para indivíduo e circulam em escala global. Dentre esses vírus, o Coronavírus, membro da família Coronaviridae, se destaca por possuir o maior número de espécies catalogadas e causar a mortalidade do seu hospedeiro. O enfoque dessa família ainda se deu no final de 2019, quando diversos casos de infecção por esse vírus foram relatados na China. Com isso, o objetivo deste trabalho foi fazer uma revisão de literatura, sobre as principais espécie de Coronavírus que afetam tanto a saúde humana quanto a animal. A revisão foi baseada em artigos nacionais e internacionais, utilizando-se das bases de dados como MEDLINE/PUBMED, Scientific Electronic Library Online (SCIELO), Science Direct e Google Scholar, dentre outros. Apesar da evolução das pesquisas acerca desses vírus, pouco se sabe da atuação dos mesmo em animais não domésticos,

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
546
sendo necessário pesquisas mais aprofundadas para melhor entendê-lo. Palavras-chave: Coronaviridae. HCoVs. Doenças infecciosas. INTRODUÇÃO
Coronavírus são um grupo de vírus membro da família
Coronaviridae e da ordem Nidovirales, que apresentam RNA de
fita simples positiva no seu genoma, utilizado para síntese
proteica. Essa família de vírus possui grandes RNAs
envelopados sendo todos de fita simples, e são considerados
os maiores vírus de RNA conhecidos. Seus genomas podem
variar de 25 a 32 kb e viriões de 118 a 140 nm de diâmetro Eles
podem ser classificados em duas subfamílias: (1) Coronaviridae
compreendendo os gêneros Alphacoronavirus,
Betacoronavirus, Gammacoronavirus e Deltacoronavirus e (2)
Torovirinae compreendendo os gêneros Torovirus, Bafinivirus e
um gênero não atribuído (SAIF et al., 2019).
Os membros da subfamília Coronaviridae são comuns
entre os mamíferos, geralmente causando apenas infecções
respiratórias ou infecções entéricas leves. Dentre os quatro
gêneros, apenas seis espécies causam infecção em humanos
(BRASIL, 2020). No gênero Alphacoronavirus as espécies mais
conhecidas são coronavírus canino (Ccov) e o coronavírus
felino (FcoV), ambos causadores da gastrenterite, isto é,
inflamação do revestimento do estômago e dos intestinos
grosso e delgado - transmissible gastroenteritis virus (TGEV).
Os coronavírus humanos (HcoV) mais comuns são os das
espécies HCoV-229E e HCoV-NL63, que causam infecções
leves a moderadas comumente (JAIMES et al., 2020).
O gênero Betacoronavírus faz parte da linhagem mais
antiga presente na literatura, por apresentar a maior quantidade

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
547
de espécies que infectam homens e animais de interesse
zootécnico. O primeiro caso registrado em animais foi em 1930
em galinhas e posteriormente identificada em bovinos, equinos
e lagomorfos (coelhos e lebres). Os principais sintomas são
febres e sintomas respiratórios, podendo incluir pneumonia
aguda. As espécies que infectam humanos são HCoV-HKU1,
HCoV-OC43, SARS-CoV e MERS-CoV (LIM et al., 2016).
O HCoV-HKU1 e o HCoV-OC43 são capazes de causar
infecções leves a moderadas. O HCoV-HKU1 é originário de
ratos infectados e foi identificado pela primeira vez em um
paciente idoso na cidade de Hong Kong, China, no ano de 2005.
O HCov-OC43 é capaz de infectar tanto humanos quanto
bovinos, e foi isolado pela primeira vez de um voluntário doente
em 1967 na cidade de Salisbúria, Inglaterra (KIN et al., 2016).
Outra subespécie conhecida é o MERS-CoV.
Encontrado em dromedários, esse vírus é o causador da
Síndrome Respiratória do Oriente Médio, do inglês Middle East
respiratory syndrome coronavirus (MERS), e foi identificado no
ano de 2012, após a obtenção de escarros de pacientes
doentes, após um surto no Oriente Médio. No ano de 2014,
casos de MERS-CoV foram relatados em vários países, como
Arábia Saudita, Malásia, Jordânia, Qatar, Emirados Árabes
Unidos, Tunísia e Filipinas (KHUDHAIR et al., 2019).
O SARS-CoV é considerado o vírus mais prejudicial a
saúde humana e é o agente responsável pela Síndrome
Respiratória Aguda Grave (SARS, do inglês Severe Acute
Respiratory Syndrome, ou SRAG, em português. Foi
identificado pela primeira vez no fim de 2002 na China. Entre
2002 e 2003, um surto da doença resultou em mais de 8000
casos e cerca de 800 mortes em todo o mundo (RU et al., 2020).
No ano de 2019, um novo vírus foi identificado na cidade de

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
548
Wuhan, China. Os pacientes por ele afetado apresentavam
febre, tosse, falta de ar e dificuldade para respirar, indicativo do
quadro clínico de pneumonia (CDC, 2020).
Este coronavírus foi inicialmente nomeado como o novo
coronavírus de 2019 (2019-nCoV), pela Organização Mundial
da Saúde (OMS)(GUO et al., 2020). Rapidamente, instalou-se
uma epidemia de infecção aguda do trato respiratório, tendo a
OMS nomeado oficialmente a doença como doença pelo
coronavírus 2019 (Coronavírus disease 2019 - Covid-19), e o
novo coronavírus como o vírus da síndrome respiratória aguda
grave do coronavírus 2 (severe acute respiratory syndrome
coronavirus 2 – Sars-CoV-2). Até março de 2020, um total de
79.968 casos de Covid-19 foram confirmados na China, com
2.873 mortes. Ao redor do globo SARS-CoV-2 foi responsável
por mais de 110.000 infecções e 4000 mortes até aquele mês,
afetando diretamente a economia de diversos países (LU et al.,
2020).
Outro coronavírus responsável por grandes perdas
econômicas é o do genêro Gamacoronavírus. Esse grupo
possui apenas um subgênero conhecido, o Igacovirus,
causador da bronquite infecciosa aviária (do inglês Avian
Infectious bronchitis - BIG). Esse vírus não afeta humanos
(SANTOS et al., 2018), porém, a infecção é responsável por
provocar grandes perdas econômicas no mercado aviário.
Apesar da existência da sua vacina, a mutação viral, formação
de novos sorotipos e variações antigênicas dificultam seu
controle da BIG. O mesmo acontece com o gênero
Deltacoronavírus, que apesar de causar grande perdas
econômicas devido a sua infecção em aves (avian
coronaviruses - CoVs) e suínos (porcine deltacoronavirus -
PDCoV) , não possui relato de infecção em humanos. Esse

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
549
gênero viral também foi identificado em leopardos, furões e
texugos (LIANG et al., 2019) .
Dado o exposto, o objetivo deste trabalho é reunir
informações acerca das diferentes espécies de coronavírus,
descrevendo as particularidades de cada gênero, com o intuito
de facilitar o entendimento acerca das diferentes espécies e
fisiopatologias acometidas por esta família de vírus.
MATERIAL E MÉTODOS
Este estudo constitui-se de uma revisão bibliográfica
descritiva, limitada às obras e estudos publicados entre 2016 e
2020, em inglês e português. As bases de dados do
MEDLINE/PUBMED, Scientific Electronic Library Online
(SCIELO), Science Direct e Google Scholar foram consultadas
para o levantamento de artigos científicos publicados em
periódicos indexados. Na estratégia de busca, foram utilizados
os descritores: Coronaviridae; Alphacoronavirus;
Betacoronavirus; Gammacoronavirus; Deltacoronavirus.
A partir dos descritores, diversas fontes foram
selecionadas com base na leitura dos títulos e resumos, e,
posteriormente, com base no texto e qualidade dos trabalhos
na íntegra. Para a construção de tal revisão bibliográfica foi
utilizado como critério de inclusão trabalhos em português e
inglês intrínsecos ao tema, com ênfase na problemática
proposta.

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
550
RESULTADOS E DISCUSSÃO
Alfacoronavírus
Os alfacoronavírus (antigo grupo CoV 1) são vírus
encontrados exclusivamente em mamíferos, causando
geralmente gastroenterite em animais e doenças respiratórias
em humanos, estes, em sua maioria, interagem e entram nas
células hospedeiras usando CD13 (aminopeptidase N). Dentre
os componentes desse gênero estão o FIPV (vírus da peritonite
infecciosa felina), o NL63, que acomete humanos e o CCoV, o
coronavírus canino (CUI; LI; SHI et al., 2019).
A peritonite infecciosa felina (FIP), é uma doença
sistêmica letal causada pelo vírus FIP (FIPV), um mutante
virulento do coronavírus entérico apatogênico felino (FECV).
Os achados patológicos característicos da FIP em gatos
evidenciam fluído seroso nas cavidades peritoneal e pleural e
lesões piogranulomatosas nos órgãos internos e no cérebro
(TAKANO et al., 2020).
O vírus se espalha de forma eficiente pela via fecal-oral
e, como as infecções podem persistir subclinicamente por um
período de um ano ou mais, a prevalência de FECV é alta,
podendo chegar até 90% de soropositividade em ambientes
multi-gatos. O progresso na pesquisa da FIP tem sido limitado,
não apenas por causa das dificuldades em isolar e propagar o
sorotipo I FCoV, mas ainda devido às mutações ocorridas in
vitro para estudos mecanísticos de patogênese em nível
molecular, que são urgentemente necessários para o
desenvolvimento de vacinas e terapêuticas (YANG et al., 2020).
O HCoV-NL63 (Coronavírus humano NL63) é um dos
coronavírus humanos mais comuns e um dos principais HCoVs
circulantes no outono e inverno em todo o mundo, sendo

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
551
isolado pela primeira vez em Amsterdã, no ano de 2004 (WENG
et al., 2019). O vírus ancora-se nas células ciliadas por meio de
proteoglicanos de sulfato de heparana, na sequência, interage
com o receptor de entrada ACE2 (ação compartilhada com o
SARS-CoV), esta interação desencadeia o recrutamento de
clatrina, seguido por endocitose mediada por clatrina,
dependente de dinamina, que requer remodelação do córtex de
actina. Sua infecção acomete principalmente em crianças e
imunocomprometidos, podendo causar sintomas respiratórios
superiores leves (tosse, febre e rinorreia) ou envolvimento mais
grave do trato respiratório inferior, como bronquiolite e crupe
(CHOUDHERY; KHATIB; 2020).
O último vírus desse gênero é o CCov (Coronavírus
canino). Responsável pela coronavirose canina entérica, uma
doença infecciosa endêmica com distribuição mundial e alta
morbidade em filhotes (DESAI et al., 2020). A doença intestinal
pode estar relacionada à apoptose induzida por vírus de células
da mucosa epitelial do intestino delgado, pois, sabe-se que o
vírus invade e se replica nas vilosidades do intestino delgado,
resultando em ruptura da área de superfície absortiva, o que
ocasiona má absorção dos nutrientes e consequente
debilidade. Ainda de acordo com o autor, os sinais clínicos
geralmente incluem anorexia, vômito e diarreia, e, a maioria dos
cães adultos infectados se recuperam em uma semana por
meio de tratamento regular intenso junto com a fluidoterapia e
raramente resultam em mortalidade.
Betacoronavírus
O gênero Betacoronavirus é responsável por abrigar pelo
menos três espécies com reconhecida patogenicidade em
humanos. Neste gênero estão inclusos o vírus responsável pela
síndrome respiratória aguda do Oriente Médio (MERS-CoV), e
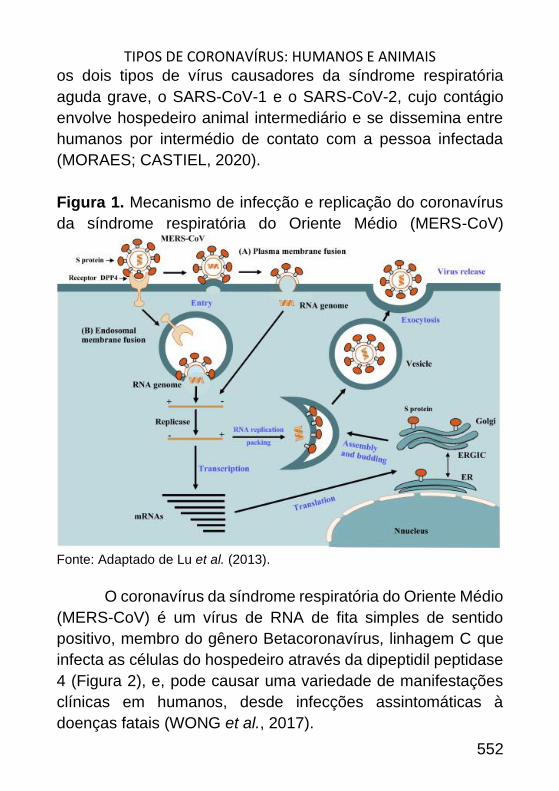
TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
552
os dois tipos de vírus causadores da síndrome respiratória
aguda grave, o SARS-CoV-1 e o SARS-CoV-2, cujo contágio
envolve hospedeiro animal intermediário e se dissemina entre
humanos por intermédio de contato com a pessoa infectada
(MORAES; CASTIEL, 2020).
Figura 1. Mecanismo de infecção e replicação do coronavírus
da síndrome respiratória do Oriente Médio (MERS-CoV)
Fonte: Adaptado de Lu et al. (2013).
O coronavírus da síndrome respiratória do Oriente Médio
(MERS-CoV) é um vírus de RNA de fita simples de sentido
positivo, membro do gênero Betacoronavírus, linhagem C que
infecta as células do hospedeiro através da dipeptidil peptidase
4 (Figura 2), e, pode causar uma variedade de manifestações
clínicas em humanos, desde infecções assintomáticas à
doenças fatais (WONG et al., 2017).
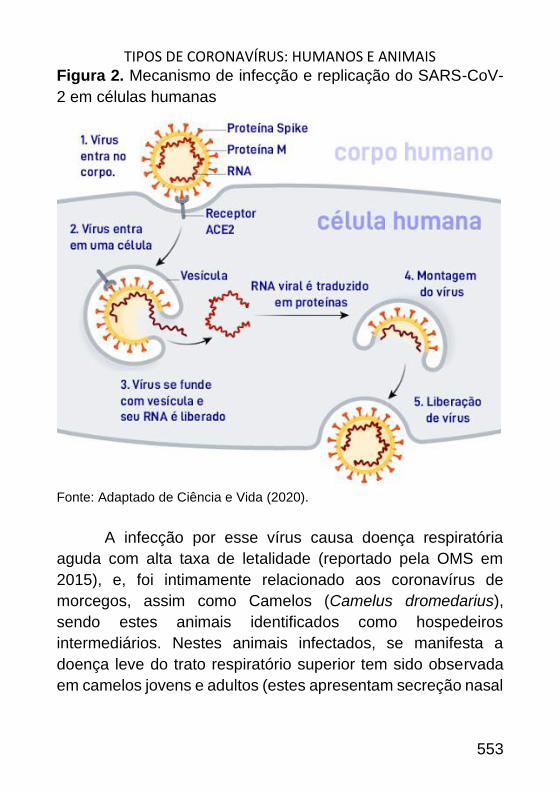
TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
553
Figura 2. Mecanismo de infecção e replicação do SARS-CoV-
2 em células humanas
Fonte: Adaptado de Ciência e Vida (2020).
A infecção por esse vírus causa doença respiratória
aguda com alta taxa de letalidade (reportado pela OMS em
2015), e, foi intimamente relacionado aos coronavírus de
morcegos, assim como Camelos (Camelus dromedarius),
sendo estes animais identificados como hospedeiros
intermediários. Nestes animais infectados, se manifesta a
doença leve do trato respiratório superior tem sido observada
em camelos jovens e adultos (estes apresentam secreção nasal

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
554
purulenta), naturalmente infectados com MERS-CoV ou sem
doença aparente (MOSTAFA et al., 2020).
Em 2003, a população chinesa foi infectada com um
vírus que causa a Síndrome Respiratória Aguda Grave (SARS),
na província de Guangdong. Os pacientes infectados exibiam
sintomas de pneumonia com lesão alveolar difusa que levou à
síndrome do desconforto respiratório agudo (SDRA).
Inicialmente o SARS-CoV infectou cães-guaxinins (Nyctereutes
procyonoides) e civetas-palmeira (Paradoxurus
hermaphroditus), e estes animais tornaram reservatório-chave
de infecção e hospedeira secundária, respectivamente. O
SARS-CoV-2 é o mais novo membro da família, identificado em
dezembro de 2019. Trata-se de um vírus de RNA envelopado
esférico, com genoma de fita simples positiva com cerca de
30.000 nucleotídeos bem adaptado à infecção de seres
humanos e de propagação rápida, o que culminou em expansão
global e início da doença do coronavírus de 2019 (Coronavírus
Disease 2019: COVID-19) (SHEREEN et al., 2020).
Além das proteínas não estruturais envolvidas no
processo de sua replicação, o coronavírus possui outras de
natureza estrutural, como a glicoproteína de membrana na
forma de espícula (“Spike”), que se mostra necessária para
infecção da célula hospedeira. Neste sentido, o acesso
intracelular do SARS-CoV-2 ocorre através da interação desta
proteína “Spike” com o receptor da enzima conversora de
angiotensina 2 (ECA2) (Figura 2) (BOOPATHI; POMA;
KOLANDAIVEL, 2020). O SARS-CoV-2 tem período de
incubação variável, que pode alcançar cinco dias antes da
manifestação dos sintomas, e o fato de que nem todos os
infectados apresentam sintomatologia incapacitante ou bem

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
555
definidos, o que eleva a sua taxa de transmissibilidade em
vários grupos populacionais (MORAES; CASTIEL, 2020).
Gamacoronavírus
Os gamacoronavírus são geralmente detectados em
espécies galináceas e não galináceas saudáveis, e, são
relacionados à bronquite aviária infecciosa (BAGHERI;
PEIGHAMBARI; LANGEROUDI, 2019). A espécie mais
conhecida é o vírus da bronquite aviária infecciosa (IBV). Esse
é um vírus envelopado com cerca de 120 nm de diâmetro e o
seu genoma consiste em um RNA de sentido positivo de fita
simples de aproximadamente 27,6 kb. O vírus apresenta em
seu material genético o gene 1, que é indispensável para
replicação e transcrição de RNA, cujos produtos fornecem
funções de RNA polimerase dependentes de RNA (MOHARAM
et al., 2020; FERNANDES, 2016).
Além do gene 1, o IBV compreende quatro genes que
codificam proteínas não estruturais (3, 4, 5 e 6b) e quatro genes
que codificam proteínas estruturais, ou seja, a proteína spike
(S), que é proteoliticamente clivada em duas subunidades: S1
e S2, que são responsáveis pela formação da porção bulbar do
ectodomínio amino-terminal da proteína S, formação de
sincícios e pela fusão de membranas celulares; a glicoproteína
de membrana integral (M); a proteína do envelope associada à
membrana (E) e a nucleoproteína (N) (FERNANDES, 2016;
MOHARAM et al., 2020).
O IBV invade o hospedeiro aviário pelo trato respiratório,
por apresentar tropismo pelas superfícies epiteliais; em seguida
causa a destruição do epitélio, levando ao desconforto
respiratório e ao início de infecções bacterianas secundárias. A
infecção é caracterizada pelo aumento da secreção oculonasal

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
556
e excesso de muco na traqueia das aves, juntamente com a
redução do ganho de peso e da eficiência alimentar. Porém as
mutações e recombinações no genoma do IBV resultaram em
vírus com diferentes tropismos de tecidos, aumento da
virulência, aumento da persistência da infecção, além da
possibilidade de transmissão assintomática. O IBV, também
pode se disseminar para os ovidutos, rins e trato
gastrointestinal, características que afetam negativamente a
indústria avícola (BAGHERI; PEIGHAMBARI; LANGEROUDI,
2019).
Deltacoronavírus
Deltacoronavírus é um gênero que abrange cerca de três
espécies oficialmente reconhecidas que são capazes de
infectar diferentes mamíferos e aves. A principal espécie desse
gênero é deltacoronavírus porcino, CoV HKU 15, agora
conhecido como PDCoV, e, foi encontrado em amostras fecais
de porcos em Hong Kong (WOO et al., 2017). O vírus
apresenta, em seu material genético, os genes característicos
da família Coronaviridae (ORF1A; ORF1B; S, spike; E,
envelope; M, membrana; N, nucleocapsídeo), todavia, contém
alguns genes acessórios como o NS6 e o NS7, que originam
proteínas não estruturais do vírus. De acordo com análises
moleculares realizadas, foi constatado que o PDCoV se
originou relativamente de um evento de troca de hospedeiro
entre espécies de aves e mamíferos (WANG et al., 2019).
A via de infecção do patógeno dá-se pelas vias nasais, e
no tecido epitelial nasal há a replicação do PDCoV e, então, são
transportados por células dendríticas e transmitidos para
células T CD3+, que transportam o vírus para o intestino,
infectando assim, os enterócitos. A lesão dessas células

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
557
causam vacuolização do citoplasma, necrose, condensação
estromal, além de morfologia escamosa cuboidal a achatada
dos enterócitos das vilosidades afetadas e presença de detritos
nucleares(WANG et al., 2019).
Os sintomas apresentados pelos suínos após infecção
são diarreia e vômito, que podem levar à desidratação; perda
de peso corporal, letargia e morte. A doença é geralmente mais
grave e letal em animais neonatos. Em alguns suínos, já foram
relatadas a presença do PDCoV no sangue, fígado, pulmão,
rins, saliva ou fluido oral, e nos linfonodos mesentéricos, o que
implica que o vírus pode causar infecções sistêmicas
ocasionais (WOO et al., 2017). Desde 2014 este vírus é
considerado um dos principais patógenos entéricos que
acomete diarreia em porcos, com taxa de mortalidade que varia
de 30% a 50%, sendo associado entre os anos de 2014 e 2017
a surtos fatais em porcos no Leste/Sudeste Asiático e na
América do Norte (WOO et al., 2017).
Outros vírus
Ainda há outros vírus que compõem a subfamília
Coronaviridae, como por exemplo os Alfacoronavírus FCoV e
229E. De acordo com Santana-Clavijo et al. (2020), destes, o
FCov causa infecção em felinos, principalmente gatos,
ocasionado enterite leve nos animais. O vírus também é
conhecido como “coronavírus entérico leve felino” ou “vírus da
peritonite felina altamente virulenta - FIPV“ e é altamente
virulento, dois genótipos de FCoV são conhecidos: FCoV-I e
FCoV-II. Foi proposto que o FCoV tipo I evolua de um vírus
ancestral comum, enquanto as cepas de FCoV tipo II surgiram
de vários eventos de recombinação (SANTANA-CLAVIJO et
al., 2020). O HCoV-229E, por outro lado, circula na população

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
558
humana e seus sintomas são relacionados ao resfriado comum,
porém, devido a mudanças na sequência genética e sorologia,
sugere-se que o vírus esteja em mutação ao longo de sua
interação com o sistema imunológico humano (WONG et al.,
2017).
No gênero Betacoronavírus, os vírus CRCoV, HKU1 e
OC43 também são responsáveis por infecções relacionadas ao
trato respiratório de diferentes espécies animais. O CRCoV, ou
coronavírus respiratório canino, está associado a sinais
respiratórios leves a graves em cães e é o agente etiológico da
doença respiratória infecciosa canina (do inglês canine
infectious respiratory disease - CIRD). As linhagens HKU1 e
OC43 são comumente associadas ao resfriado comum em
humanos, porém já foi relatado que estes vírus podem causar
infecções pulmonares graves e/ou fatais em pessoas que
apresentam alguma comorbidade, e que, em casos raros, o
vírus OC43 que usa ácido 9-O-acetil siálico como seu receptor,
pode ocasionar encefalite letal(WONG et al., 2017).
O vírus SW1 (do gênero gamacoronavírus) e o HKU11
(do gênero deltacoronavírus) também acarreta infecções do
trato respiratório. O SW1 ou Beluga Whale CoV, é responsável
por doenças pulmonares e insuficiência hepática aguda
terminal em baleias brancas e é o primeiro coronavírus a ser
relatado em animais marinhos (WANG et al., 2019). A linhagem
HK11 é responsável por causar infecções relacionadas à família
Pycnonotidae, que compreende pássaros canoros
passeriformes, levando a doenças pulmonares nestas aves
(CHEN; LIU; GUO, 2020).

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
559
CONSIDERAÇÕES FINAIS
Os vírus respiratórios geralmente têm dois modos
principais de transmissão, grandes partículas de aerossóis de
gotículas respiratórias transmitidas diretamente por contato,
através de tosse ou espirro, ou através de fômites, que são
materiais contaminados por secreções que contenham o vírus.
Esta característica de transmissão eleva potencialmente a
propagação da doença, permite a interação do vírus com
diferentes espécies e pode apresentar comportamento
zoonótico.
Assim, este trabalho objetivou elencar as principais
espécies de vírus da família Coronaviridae devido a sua
capacidade de infectar diferentes espécies, causar grandes
impactos econômicos e também devido ao seu ressurgimento
no final do ano de 2019, sendo responsável por uma das
maiores pandemias mundiais. Como percebido na revisão, o
coronavírus permeia o meio e existem profilaxias para o seu
combate em espécies animais, como as de animais de
companhia, o que gerou no início da pandemia uma busca pela
vacina já utilizada em animais para que fossem aplicados em
humanos. Ao constatar que temos diferentes classes deste
vírus, esta prática não surte efeito.
Apesar de muitos materiais encontrados voltados para
vírus que infectam humanos, quando se depara com vírus que
infectam apenas animais que não são de interesse
zootecnológico e/ou domésticos, as pesquisas são bem
escassas. Com isso, ainda é preciso mais pesquisas em
diferentes espécies para entender o mecanismo completo de
infecção desse vírus para futuramente criar meios para
combatê-los.

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
560
REFERÊNCIAS BAGHERI, S. S.; PEIGHAMBARI, S. M.; LANGEROUDI, A. G. Detection and Molecular Characterization of Gammacoronavirus in Quail Population in Iran. Irian Journal of Veterinary Medicine, v.13, n.3, p. 251-259, 2019. Boopathi S.; Poma A. B.; Kolandaivel P. Novel 2019 coronavirus structure, mechanism of action, antiviral drug promises and rule out against its treatment. Journal of Biomolecular Structure and Dynamics, p. 1-10, 2020. BRASIL. Ministério da Saúde. Coronavírus Covid-19. 2020. Disponível em:<https://coronavirus.saude.gov.br/>Acesso em 06 de julho de 2020. CDC - Chinese Center for Disease Control and Prevention. [The Epidemiological Characteristics of an Outbreak of 2019 Novel Coronavirus Diseases (COVID-19) in China]. Zhonghua Liu Xing Bing Xue Za Zhi. v. 41, n. 2, p. 145-151. 2020. CHOUDHERY, A. A.; KHATIB, S. Covid-19 - A New Pandemic for 137 million Indian-impact, interventions and solutions. Uttar Pradesh Journal of Zoology, p. 1-17, 2020. CUI, J; LI, F.; SHI, Z.-L. Origin and evolution of pathogenic coronaviruses. Nature Reviews Microbiology, v. 17, p. 181–192, 2019. DESAI, D.; KALYANI, I.; PATEL, D.; MAKWANA, P; SOLANKI, J.; VALA, J. Rapid Detection based Prevalence of Canine Corona Virus (CCoV) and Canine Parvo Virus (CPV) Infection in Diarrheic Dogs in South Gujarat. Indian Journal of Veterinary Science and Biotechnology, v. 16, n. 1, p. 41-43, 2020. DONG, S.;SUN, J.; MAO, Z.; WANG, L.; LU, Y.I.; LI, J. A guideline for homology modeling of the proteins from newly discovered betacoronavirus, 2019 novel coronavirus (2019‐nCoV). J. Medical Virology. n. 1, p. 1-7. March, 2020. FERNANDES, A. M. Genealogia de Amostras Brasileiras de Coronavírus Bovino. Dissertação (Mestrado em Sanidade, Segurança Alimentar e Ambiental no Agronegócio). Instituto Biológico, Agência Paulista de Tecnologia dos Agronegócios, Secretaria de Agricultura e Abastecimento do Estado de São Paulo. São Paulo. 2016. FTERJ - Federação de Triathlon do Estado do Rio de Janeiro, 2020. Disponível em <http://www.triathlon.org.br/docs/artigo_corona.pdf>. Acesso em 15 de setembro de 2020. GUO, Y.R.; CAO, Q.D.; HONG, Z.S.; TAN, Y.Y.; CHEN, S.D.; JIN, H. J.; TAN, K. S.; WANG, D. Y.; YAN, Y. The origin, transmission and clinical therapies on coronavirus disease 2019 (COVID-19) outbreak - an update on the status. Military Medical Research. v. 7, n. 1, p. 1-11.2020. JAIMES , J.A.; MILLET, J.K.; STOUT, A.E.; ANDRE, N.M.; WHITTAKER , G.R. A Tale of Two Viruses: The Distinct Spike Glycoproteins of Feline Coronaviruses. Viruses. v. 12, n. 83, p. 1-14, 2020.

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
561
KHUDHAIR, A.; KILLERBY, M.E.; MULLA, M.A.; ELKHEIR, K.A.; TERNANNI, W.; BANDAR, Z.; WEBER, S.; KHOURY, M.; DONNELLY, G.; AL MUHAIRI, S.; KHALAFALLA, A. I.; TRIVEDI, S.; TAMIN, A.; THORNBURG, N. J.; WATSON, J. T.; GERBER, S. I.; AL HOSANI, F.; HALL, A. J. Risk Factors for MERS-CoV Seropositivity among Animal Market and Slaughterhouse Workers, Abu Dhabi, United Arab Emirates, 2014–2017. Emerging Infectious Diseases, n. 25, v. 5, p. 927–35, may 2019. KIN, N.; MISZCZAK. F.; DIANCOURT, L.; CARO. V.; MOUTOU F.; VABRET, A.A.R.; GOUILH. M. Comparative molecular epidemiology of two closely related coronaviruses, bovine coronavirus (BCoV) and human coronavirus OC43 (HCoV-OC43), reveals a different evolutionary pattern.Infect Genet Evol. n. 40, p. 186-191. Jun, 2016. LIANG, Q.; ZHANG, H.; LI, B.; DING,Q.; WANG, Y.; GAO, W.; GUO, D.; WEI, Z.; HU, H. Susceptibility of Chickens to Porcine Deltacoronavirus Infection. Viruses. n. 11, v. 6, p. 1-13. 2019. LIM, Y.X.; NG, Y.L.; TAM, J.P.; LIU, D.X. Human Coronaviruses: A Review of Virus–Host Interactions. Diseases. n. 4, v. 26, p. 1-28. July, 2016. LU, X.; ZHANG, L.; DU, H.; ZHANG, J.; LI, Y. Y.; QU, J.; ZHANG, W.; WANG, Y.; BAO, S.; LI, Y.; WU, C.; LIU, H.; LIU, D.; SHAO, J.; PENG, X.; YANG, Y.; LIU, Z.; XIANG, Y.; ZHANG, F.; SILVA, R. M.; PINKERTON, K. E.; SHEN, K.; XIAO, H.; XU, S.; WONG, G. W. K. SARS-CoV-2 Infection in Children. The New England Journal of Medicine. v. 382, n. 17. p. 1-3. Abril de 2020. MOHARAM, I.; SULTAN, H.; HASSAN, K.; MAHMOUD, I.; SHANY, S.; SHEHATA, A. A.; ABO-ELKHAIR, M.; PFAFF, F.; HOPER, D.; EL KADY, M.; BEER, M.; HARDER, TIMM.; HAFEZ, H.; GRUND, C. Emerging infectious bronchitis virus (IBV) in Egypt: Evidence for an evolutionary advantage of a new S1 variant with a unique gene 3ab constellation. Infection, Genetics and Evolution, v.85, p.1567-1348, 2020.
MORAES, R.; CASTIEL, L. D. Infecção pelo coronavírus e disfunção vascular: impactos na saúde e economia e na prática de atividades físicas.
MORAWSKA, L.; CAO, J. Airborne transmission of SARS-CoV-2: The world should face the reality. Environment International, v. 139, p. 105730, 2020. MOSTAFA, A.; KANDEIL, A.; SHEHATA, M.; SHESHENY, R.E.; SAMY, A.M.; KAYALI, G. ALI, M.A. Middle East Respiratory Syndrome Coronavirus (MERS-CoV): State of the Science. Microorganisms, v. 8, n. 7, p. 99, 2020. RU, H.; YANG, E.; ZOU, K. Combating the COVID-19 Pandemic: The Role of the SARS Imprint. 2020. SSRN online. Disponível em:<https://ssrn.com/abstract=3569330>. Acesso em 07 de julho de 2020. SAIF, L.J.; WANG, Q.; VLASOVA, A.N.; JUNG, K.; XIAO, S. Coronaviruses. in: ZIMMERMAN, J.J.; KARRIKER, L.A.; RAMIREZ, A.; KENT J. SCHWARTZ, K.J,; STEVENSON, G.W.; ZHANG, J. Diseases of Swine, ed. 11, p. 488-523, mar de 2019.

TIPOS DE CORONAVÍRUS: HUMANOS E ANIMAIS
562
SANTANA-CLAVIJO, N. F.; ROMERO, D. P. R.; FAJARDO, D. F. A.; MUÑOZ, A. V.; TANIWAKIA, S. A.; SILVA, S. O. S.; BRANDÃO, P. E. Molecular diversity of Alphacoronavirus 1 in dogs and cats in Colombia. Heliyon, v.9, ed.7, jul 2020. SANTOS, R. M., FERNANDO, F. S.; MONTASSIER, M. F. S.; SILVA, K. R. LOPES, P. D.; BORZI, M. M.; OLIVEIRA, E. S.; MONTASSIER, H. J. Vírus da bronquite infecciosa aviária: perfil de imunogenicidade e novos patótipos no Brasil – revisão de literatura. Nucleus Animalium. v. 10, n. 1, p. 45-64. 2018. SHEREEN, M. A.; KHAN, S.; KAZMI, A.; BASHIR, N.; SIDDIQUE, R. COVID-19 infection: origin, transmission, and characteristics of human coronaviruses. Journal of Advanced Research, v. 24, p. 91-98, 2020. TAKANO, T.; SATOH, K; DOKI, T.; TANABE, T; HOHDATSU, T. Antiviral Effects of Hydroxychloroquine and Type I Interferon on In Vitro Fatal Feline Coronavirus Infection. Viruses, v. 12, n. 5, p. 576, 2020. WANG, L.; MADDOX, C; TERIO, K; LANKA, S.; FREDRICKSON, R. NOVICK, B.; PARRY, C.; MCCLAIN, A.; ROSS, K. Detection and Characterization of New Coronavirus in Bottlenose Dolphin, United States, 2019. Emerging Infectious Disease, v. 26, ed. 7, p.1610-1612, jul 2020. WANG, Q.; VLASOVA, A. N.; KENNEY, S. P.; SAIF, L. J. Emerging and re-emerging coronnaviruses in pigs. Current Opinion in Virology, v.34, p.39-49, 2019. WENG, J.-R.; LIN, C.-S.; LAI, H.-C.; LIN, Y.-P.; WANG, C.-Y.; TSAI, Y.-C.; WU, K.-C.; HUANG, S.-H.; LIN, C.-W. Antiviral activity of Sambucus FormosanaNakai ethanol extract and related phenolic acid constituents against human coronavirus NL63. Virus Research, v. 273, p. 197767, 2019. WONG, A.H.M.; TOMLINSON, A.C.A.; ZHOU, D.; SATKUNARAJAH, M.; CHEN, K.; SHARON, C.; DESFORGES, M.; TALBOT, P. J.; RINI, J. M. Receptor-binding loops in alphacoronavirus adaptation and evolution. Nature Communication, v.8, n. 1735, nov. 2017. WOO, P. C.; LAU, S. K.; TSANG, C. C.; LAU, C. C.; WONG, P. C.; CHOW, F. W; FONG, J. Y.; YUEN, K. Y. Coronavirus HKU15 in the respiratory tract of pigs and first discovery of coronavirus quasispecies in 5′-untranslated regions. Emerging Microbes & Infections, v.6, n.6, ed. 53, 2017. YANG, T.J.; CHANG, Y,C; KO, T.P.; DRACZKOWSKI, P.; CHIEN, Y.C.; CHANG, Y.C.; WU, K.P.; KHOO, K.H.; CHANG, H.W.; HSU, S.T. D. Cryo-EM analysis of a feline coronavirus spike protein reveals a unique structure and camouflaging glycans. Proceedings of the National Academy of Sciences, v. 117, n. 3, p. 1438-1446, 2020.

563
CIÊNCIAS BIOLÓGICAS -PARASITOLOGIA

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
564
CAPÍTULO 27
PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE
AEDES AEGYPTI: UMA REVISÃO
Karine da Silva CARVALHO1
Romulo Carlos Dantas da CRUZ2
Lais da Silva CARVALHO3 Laiane da Silva CARVALHO3
Ivone Antonia de SOUZA4
1Departamento de Antibióticos, UFPE; 2Pós-doutorando no Departamento de Antibióticos, UFPE; 3Graduanda em medicina, UFS, 4Orientadora/professora do Departamento de Antibióticos, UFPE. [email protected]
RESUMO: O Brasil possui uma grande biodiversidade difundida em diferentes biomas, dentre eles a Caatinga. A singularidade climática desse bioma permite a sobrevivência de plantas que produzem diversos metabólitos secundários, algunss dos quais podem ser tóxicos para os insetos. A busca por plantas ativas sobre mosquitos, como Aedes aegypti, principal transmissor do dengue, chikungunya e Zika, torna-se essencial. Assim, o objetivo desse trabalho foi realizar uma revisão integrativa sobre o potencial inseticida de plantas da Caatinga sobre o mosquito Ae. aegypti. Para tal, realizou-se um levantamento de artigos indexados, nos últimos cinco anos, às plataformas do Pubmed, Science Direct e Scielo utilizando os descritores “Caatinga”, “Insecticides” “Plants” e “Aedes aegypti”. Diversos estudos demonstram a eficácia inseticida de plantas da Caatinga sobre as fases de ovo, larva, pupa e adulta do mosquito Ae. aegypti, com predominância da atividade de plantas na fase larval. A maioria dos estudos utilizam como órgão vegetal as folhas. Além da atividade inseticida, alguns

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
565
estudos também demonstram a toxicidade destas plantas sobre mamíferos visando estabelecer o uso seguro dessas. Assim, a Caatinga é um bioma promissor quanto à presença de plantas detentoras de compostos bioativos, úteis ao controle de insetos vetores, o que poderá contribuir para a caracterização e valorização desse bioma, bem como a produção de inseticidas de origem natural. Palavras-chave: Biodiversidade. Dengue. Inseticida.
INTRODUÇÃO
O Brasil é um país de grande extensão territorial com
diversas zonas climáticas que produzem variações ecológicas,
formando distintos biomas. A Caatinga possui uma vegetação
típica denominada Floresta e Arbustais Tropicais Sazonalmente
Secos, FATSS, sendo a maior em extensão e mais contínua
área do bioma do Novo Mundo (FERNANDES; QUEIROZ,
2018). Esse bioma dispõe de elevado índice de endemismo, o
que favorece a sua grande biodiversidade, e localiza-se na
região semiárida nordestina, caracterizado pelo clima tropical
semiárido (QUEIROZ et al., 2017). Apesar da importância
biológica desse bioma, diante da diversidade de fisionomias
que possui, esta é uma das regiões fitogeográficas menos
protegidas por unidades de conservação no país, e
consequentemente um dos mais devastados.
Diante da marcante sazonalidade da Caatinga, muitas
espécies vegetais desenvolvem mecanismos de adaptação,
como a deciduidade, alterações morfológicas nas folhas como
a presença de espinhos, espessamento foliar e alteração na
síntese dos metabólitos secundários. Tais metabólitos podem
ser úteis na produção de inseticidas, visto que constituem a

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
566
defesa química das plantas contra as intempéries, como a
herbivoria (SOUZA et al., 2018; CRUZ et al., 2020).
A descoberta de inseticidas originados das plantas tem
como vantagens um menor impacto sobre a biodiversidade e
uma maior seletividade, ao apresentarem um baixo espectro de
ação (CRUZ et al., 2020; PAVELA 2016). Pesquisas sobre
inseticidas botânicos, torna-se essencial tendo em vista a
relevância epidemiológica, entre esses, destaca-se Aedes
aegypti, o principal transmissor do vírus da dengue, doença que
acomete milhares de pessoas no mundo. A transmissão do
vírus Zika (ZIKV) no Brasil, também por esse vetor, representou
um grave problema de saúde pública internacional, o qual foi o
epicentro da Síndrome Congênita do ZIKV (WHO, 2018;
MESSINA et al., 2019). O Ae. aegypti poderá transmitir o vírus
chikungunya e febre amarela urbana (MESSINA et al., 2019).
Diante da ausência de vacinas eficazes e disponíveis
para a maioria dessas arboviroses, é imperativo a busca por
formas de controle alternativas a serem implementadas no
controle integrado de insetos vetores. Dentre as práticas, se
destaca a aplicação de larvicidas, importantes ferramentas no
controle de mosquitos (ACHEE et al., 2019). Assim, é
indispensável a busca por formas alternativas de controle de
mosquitos, a exemplo daqueles de origem botânica por serem
ambientalmente seguros e podem retardar mecanismos de
resistências. Nesse contexto, esse estudo tem por objetivo
realizar uma revisão sobre o potencial de plantas do bioma
Caatinga quanto à propriedade inseticida sobre o mosquito Ae.
aegypti.

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
567
MATERIAIS E MÉTODO
A presente revisão integrativa ocorreu mediante a busca
eletrônica de artigos científicos indexados nas bases de dados
Science Direct, National Library of Medicine (PubMed) e
ScientificElectronic Library Online (Scielo). Utilizou-se o
operador booleano “AND” e os descritores em inglês e
português: “Caatinga”, “Inseticidas”-“Insecticides, “plantas”-
“plants” “Aedes aegypti”. Os artigos publicados nos últimos
cinco anos foram selecionados relacionados a pesquisa
científica de plantas do bioma Caatinga e seu potencial
inseticida sobre o mosquito Aedes aegypti.
Os critérios de inclusão dos estudos primários
delimitados para a presente revisão foram artigos científicos
completos e comunicações (short communication) publicados
no período de 2015 a 2020. Foram excluídas as publicações
que não foram realizadas com plantas da Caatinga e
necessariamente coletadas desse bioma, ou o tema do estudo
não fosse relacionado à atividade inseticida.
RESULTADOS E DISCUSSÃO
A Caatinga e a síntese de metabólitos secundários
A Caatinga ocupa uma área de aproximadamente
912.529 km2, distribuído nos estados do Nordeste brasileiro
(Paraíba (PB), Piauí (PI), Maranhão (MA), Sergipe (SE),
Pernambuco (PE), Bahia (BA) e Rio Grande do Norte (RN) e
também no norte de Minas Gerais (MG), na região sudeste.
Esse bioma dispõe de uma grande variedade de ambientes,

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
568
apresentando um clima com longa estação seca, irregularidade
pluviométrica, luminosidade e temperaturas intensas (GARCEZ
et al., 2016). O termo “caatinga” de origem Tupi significa
“floresta branca”, que corresponde ao aspecto da vegetação
durante a estação seca, onde a maioria das árvores perdem
suas folhas e expõem seus troncos esbranquiçados (SILVA et
al., 2017). Todavia, tal aspecto contrasta com o período das
chuvas, onde o verde predomina e as flores exibem um colorido
diversificado. Esse contraste do aspecto vegetacional em
diferentes períodos pode ser visualizado na Figura 1.
Figura 1. Diferença da vegetação na Floresta Nacional do Açu
(RN) na época da seca (a) e no período chuvoso (b).
Fonte: Coe; Souza, 2014.
As mudanças abruptas ao longo do ano na Caatinga
condicionam nos seus grupos biológicos padrões de
distribuição, singularidade e de diversidade. Apesar de nas
últimas décadas, ter sido descrito como pobre em espécies e
baixo grau de endemismo, o que reflete no pouco conhecimento
da região, e estudos recentes demonstram a sua vasta
biodiversidade, e elevado número de endemismo
(FERNANDES; QUEIROZ, 2018; SOUZA et al., 2018).
A vegetação da Caatinga inclui árvores de pequeno
porte, com cobertura descontínua de copas e as herbáceas são
anuais com destaque para euforbiáceas, bromeliáceas e
a b

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
569
cactáceas (CAVALCANTE; DUARTE; OMETTO, 2020). Entre
os mecanismos fisiológicos das plantas da Caatinga, mediante
a anatomia, morfologia e os mecanismos fisiológicos
adequados às condições locais, destaca-se a produção de
metabólitos secundários, os quais representam uma interface
química entre os vegetais e o meio que as abrangem, sendo a
sua síntese, portanto, consideravelmente afetada por
condições ambientais (SHITAN et al., 2016).
A propriedade inseticida dos compostos secundários das
plantas deve-se principalmente a estratégia de defesa, contra o
ataque de insetos e/ou patógenos (ISMAN, 2020). Tais
compostos são representados pelas classes dos compostos
fenólicos, tais como os taninos e flavonoides, classe dos
terpenos, já bem estabelecidos, quanto a atividade inseticida, e
aos alcaloides, sobretudo os alcaloides esteroidais
(CARVALHO et al., 2016; SOUZA et al., 2015). Nesse contexto,
os metabólitos secundários sintetizados pelas plantas da
Caatinga, constituem fontes de substâncias bioativas, que
podem ser utilizadas como inseticida, podendo dessa forma,
atuar no controle populacional de muitos insetos a exemplo de
Ae. aegypti que transmitem arbovírus causadores de doenças
de grande impacto na saúde pública mundial.
Aedes aegypti: importância médica e métodos de controle
Aedes aegypti é um mosquito holometábolo, ou seja,
apresenta ciclo de vida completo incluindo as fases de ovo,
larva e pupa, as quais desenvolvem-se no meio aquático, e a
fase adulta, que se desenvolve no meio aéreo. A diversificação
de nicho ecológico possibilita a esse inseto adaptação e

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
570
ocupação de diferentes ambientes, o que auxilia
consideravelmente na sua dispersão e resistência em
diferentes habitats (CARVALHO; MOREIRA, 2017). Os ovos ao
entrarem em contato com a água dará origem às larvas, as
quais possuem quatro ínstares (L1-L4). A fase larval constitui o
período de alimentação. Em seguida, a larva se transformará
em pupa, a qual não se alimenta e há modificações internas e
externas que possibilitará o desenvolvimento do adulto. O
adulto é o estágio reprodutivo da espécie. Machos e fêmeas se
alimentam de solução açucarada e apenas as fêmeas
necessitam realizar repasto sanguíneo para a maturação de
seus ovos, e nessa condição, caso estejam infectadas com
arbovírus, poderão ser capazes de transmiti-los através da sua
picada (CONSOLI; OLIVEIRA 1994).
Nas Américas, os arbovírus são transmitidos
principalmente pelo Ae. aegypti e estes são o vírus da dengue,
doença que acomete milhares de pessoas no mundo, além do
vírus Zika (ZIKV), vírus chikungunya e vírus amarílico, que
causam arboviroses de grande impacto na saúde pública. A
dengue caracteriza-se como doença febril e pode até ser letal,
através da febre hemorrágica da dengue. No Brasil, a dengue é
uma doença endêmica, responsável por um elevado número de
óbitos, anualmente. No Brasil, a Síndrome Congênita do ZIKV,
enfermidade que acomete neonatais incluindo a microcefalia,
no ano de 2016, representou um sério problema de saúde
pública nacional e internacional, segundo a Organização
Mundial da Saúde (WHO, 2018; MESSINA et al., 2019). O vírus
chikungunya causa infecção caracterizada por uma intensa
artralgia, ou seja, dor articular (MESSINA et al., 2019). Dentre
os arbovírus transmitidos pelo Ae. aegypti somente para o vírus

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
571
amarílico (causador da febre amarela) há vacina disponível no
país. Diante da ausência de vacinas disponíveis para a maioria
das doenças causadas por vírus transmitidos pelo mosquito Ae.
aegypti e de tratamentos específicos para tais arboviroses,
torna-se fundamental a implementação do controle integrado de
insetos vetores, onde diferentes práticas são adotadas visando
reduzir a população de mosquitos. Diferentes métodos de
controle incluem o controle mecânico, controle químico,
controle genético e controle biológico, os quais são os principais
métodos de controle para esse vetor. É importante destacar que
em cada um destes é muito importante haver inclusão e
mobilização social, sendo a comunidade componente essencial
no controle para este vetor. Entre essas práticas, se destaca a
aplicação de larvicidas, importante ferramenta no controle de
mosquitos (ACHEE et al., 2019).
No Brasil, o Programa Nacional de Controle da Dengue
(PNCD) recomenda, nos dias atuais, a utilização de inseticidas
químicos para o controle de mosquitos vetores. Todavia, o uso
excessivo e constante destes compostos seleciona resistência
nas populações de insetos. Além de promover a seleção da
resistência aos insetos, os inseticidas químicos causam danos
ecológicos.
Plantas da Caatinga: investigação da atividade inseticida
No que diz respeito aos inseticidas botânicos, várias
pesquisas voltadas para o potencial das espécies nativas da
Caatinga têm sido realizadas, apresentando resultados
promissores em relação a um método de controle alternativo
para o Ae. aegypti e, ao mesmo tempo, menos impactante para

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
572
o ambiente. A Tabela 1 demonstra o potencial de plantas da
Caatinga quanto sua atividade inseticida sobre o mosquito Ae.
aegypti, entre os anos 2015 e 2020.
Os vegetais do bioma Caatinga que possuem atividade
inseticida sobre o mosquito Ae. aegypti incluem espécies das
famílias Euphorbiaceae, Apocynaceae, Anacardiaceae,
Myrtaceae, Agavaceae, Verbenacea e Fabaceae (Tabela 1).
Das partes vegetais usadas para a avaliação da atividade
inseticida há uma predominância da utilização das folhas,
sendo possível também usar sementes, caules e raiz, sendo
essa última com uma menor frequência. É importante notar que
na utilização das folhas não há destruição completa das
plantas, sendo uma prática, portanto altamente sustentável.
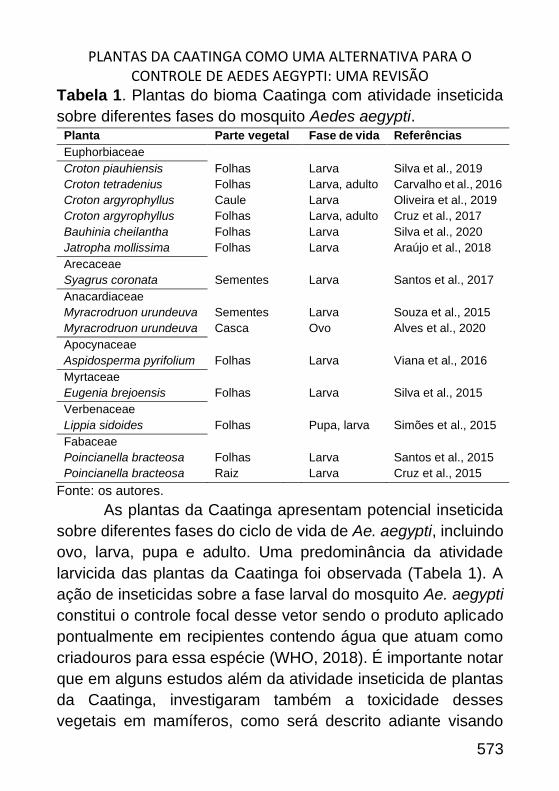
PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
573
Tabela 1. Plantas do bioma Caatinga com atividade inseticida
sobre diferentes fases do mosquito Aedes aegypti. Planta Parte vegetal Fase de vida Referências
Euphorbiaceae
Croton piauhiensis Folhas Larva Silva et al., 2019
Croton tetradenius Folhas Larva, adulto Carvalho et al., 2016
Croton argyrophyllus Caule Larva Oliveira et al., 2019
Croton argyrophyllus Folhas Larva, adulto Cruz et al., 2017
Bauhinia cheilantha Folhas Larva Silva et al., 2020
Jatropha mollissima Folhas Larva Araújo et al., 2018
Arecaceae
Syagrus coronata Sementes Larva Santos et al., 2017
Anacardiaceae
Myracrodruon urundeuva Sementes Larva Souza et al., 2015
Myracrodruon urundeuva Casca Ovo Alves et al., 2020
Apocynaceae
Aspidosperma pyrifolium Folhas Larva Viana et al., 2016
Myrtaceae
Eugenia brejoensis Folhas Larva Silva et al., 2015
Verbenaceae
Lippia sidoides Folhas Pupa, larva Simões et al., 2015
Fabaceae
Poincianella bracteosa Folhas Larva Santos et al., 2015
Poincianella bracteosa Raiz Larva Cruz et al., 2015
Fonte: os autores.
As plantas da Caatinga apresentam potencial inseticida
sobre diferentes fases do ciclo de vida de Ae. aegypti, incluindo
ovo, larva, pupa e adulto. Uma predominância da atividade
larvicida das plantas da Caatinga foi observada (Tabela 1). A
ação de inseticidas sobre a fase larval do mosquito Ae. aegypti
constitui o controle focal desse vetor sendo o produto aplicado
pontualmente em recipientes contendo água que atuam como
criadouros para essa espécie (WHO, 2018). É importante notar
que em alguns estudos além da atividade inseticida de plantas
da Caatinga, investigaram também a toxicidade desses
vegetais em mamíferos, como será descrito adiante visando

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
574
estabelecer o seu uso seguro (CARVALHO et al., 2016; CRUZ
et al., 2020).
Dessa maneira, estudos realizados nos últimos cinco
anos com plantas coletadas no bioma Caatinga, inserido em
diferentes estados do nordeste brasileiro, demonstram
propriedade inseticida (Tabela 1), o que é de grande relevância
na caracterização e valorização desse bioma, muitas vezes
desconhecido e desprezado. O desconhecimento prejudica a
conservação da biodiversidade desse bioma (LESSA et al.,
2019). A flora da Caatinga constitui uma rica fonte de pesquisa,
contendo exemplares botânicos promissores quanto ao seu
efeito tóxico sobre Ae. aegypti.
Diversas plantas da Caatinga têm apresentado
propriedade inseticida sobre Ae. aegypti, e essa atividade se
deve à presença de metabólitos secundários presentes em
diferentes partes dos vegetais. Esses metabólitos podem ser
obtidos de diferentes partes anatômicas dos vegetais da
Caatinga, e a seguir será feita uma descrição detalhada por
agrupamento de cada parte anatômica, iniciando pelas folhas,
seguida de semente, caules e raízes. Em relação às folhas Silva
et al. (2019), após analisarem a composição química do óleo
essencial de folhas de Croton piauhiensis identificaram β-
cariofileno, D-limoneno, γ-terpineno e germacreno D como os
componentes químicos majoritários. Os autores sugerem que o
ativo β-cariofileno, o principal dentre estes, provavelmente seja
o responsável pelo potencial inseticida da planta C. piauhiensis
sobre larvas de Ae. aegypti (CL50=0,34 mg/mL) (SILVA et al.,
2019).
Silva et al (2015) também demonstraram que o óleo
essencial obtido das folhas de E. brejoensis foi ativo sobre

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
575
larvas de Ae. aegypti (CL50= 0,21 mg/mL) e a avaliação
fitoquímica desse óleo essencial revelou a presença de 41
compostos químicos diferentes. Entre estes, cadineno, beta-
cariofileno, alfa-murolol e biciclogermacreno foram os
principais. Essa planta foi coletada no estado de Pernambuco,
em uma região de brejo, e seus frutos e folhas são
popularmente conhecidos como “cutia”. Essa planta tem sido
usada na medicina tradicional pela população como
antidiarreica e antifebril (SILVA et al., 2015).
O óleo essencial da planta da Caatinga Bauhinia
cheilantha conhecida popularmente como “mororó” também foi
avaliada quanto ao seu potencial larvicida sobre Ae. aegypti
(SILVA et al., 2020). Esse vegetal é popularmente utilizado
como hipoglicemiante. O óleo essencial obtido das folhas de
Bauhinia cheilantha, coletadas no estado do Ceará, apresentou
atividade larvicida sobre Ae. aegypti CL50= 0,04 mg/mL. A
avaliação da química desse óleo essencial identificou 44
compostos, com dominância de sesquiterpenos (78,6%) os
quais têm relato sobre atividade inseticida. E-cariofileno, alfa-
pineno, espatulenol e óxido de cariofileno, foram os principais
compostos químicos identificados (SILVA et al., 2020).
As folhas da planta Jatropha mollissima popularmente
conhecida como “pinhão bravo” também foram avaliadas
quanto ao seu potencial larvicida sobre Ae. aegypti (ARAÚJO
et al., 2018). As folhas da planta foram coletadas no estado do
Piauí, e a partir dessa foi obtido o extrato aquoso, o qual
ocasionou nas concentrações de 0,08 e 0,1 mg/mL taxas
superiores a 75% de mortalidade larval. Os autores, na
discussão, sugerem que esta atividade inseticida possa estar
relacionada à presença dos compostos taninos, saponinas,

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
576
alcaloides e cumarinas. Além disso ensaio de genotoxicidade
utilizando o tecido meristemático de Allium cepa foi realizado e
o extrato aquoso das folhas de Jatropha mollissima foi
considerado atóxico (ARAÚJO et al., 2018).
Da parte aérea da planta A. pyrifolium, popularmente
conhecida como “pereiro”, avaliou-se o extrato etanólico e suas
respectivas frações (diclorometânica e hidroalcoólica), assim
como as misturas das compostas pelas frações diclorometânica
mais acetato de etila (D/AE) e diclorometânica mais hexânica
(D/H) quanto ao seu potencial larvicida sobre Ae. aegypti
(VIANA et al., 2016). Como resultado a fração diclorometânica
(CL50= 3,09 mg/mL e CL90= e 6,74 mg/mL), seguida da mistura
D/H, (CL50 = 5,35 mg/mL e CL90= 7,31 mg/mL) foram mais
tóxicas comparadas ao extrato etanólico e as demais frações.
Paralelamente, os autores mostraram que o extrato etanólico foi
submetido à prospecção fitoquímica e as frações à análise de
Ressonância Magnética Nuclear 1H, e a presença majoritária
de heterosídeos cianogênicos e taninos condensados na fração
diclorometânica, os quais podem estar envolvidos na
propriedade inseticida desse vegetal (VIANA et al., 2016).
Além dos metabólitos secundários estarem presentes
em folhas de vegetais, também poderão ser encontrados nas
sementes. Souza et al. (2015), demonstraram que extrato
etanólico de sementes de M. urundeuva possui potente
atividade sobre Ae. aegypti. Essa planta foi coletada na região
da Caatinga do estado do Ceará, e é popularmente conhecida
como “aroeira do sertão”. M. urundeuva é tradicionalmente
utilizada pela população no tratamento de dores em geral e no
combate a infecções do trato urinário e digestivo. Diante do
potencial da atividade inseticida foi realizado o processo de

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
577
separação bioguiada a fim de isolar o(s) composto(s) ativo(s).
O processo de isolamento resultou em apenas uma substância
ativa, identificada como m‐pentadecadienil‐fenol. Esse
composto fenólico apresentou uma potente atividade larvicida e
pupicida (CL50=0,01 e 0,10 mg/mL, respectivamente) e
atividade inibitória na eclosão de ovos (CL50=0,49 mg/mL). Em
um trabalho anterior os autores investigaram o modo de ação
de m‐pentadecadienil‐fenol em larvas tratadas e algumas
alterações morfológicas em órgãos relacionados ao trato
respiratório foram observadas, tais como inundação do sistema
traqueal, alterações nas válvulas sifonais e nas brânquias
anais, além de letargia. Nesse estudo foi possível observar uma
diferença na expressão de 12 proteínas, entre elas a enzima
detoxificadora glutationa S-transferase, além de proteínas
envolvidas na proteção da membrana peritrófica e no
metabolismo de lipídeo, o que indicou um possível efeito tóxico
sistêmico após as larvas serem expostas ao m‐pentadecadienil‐
fenol (SOUZA et al., 2015).
O óleo essencial obtidos das sementes da planta da
Caatinga Syagrus coronata conhecida pela população como
“ouricuri” ou “licuri” foi investigada quanto ao seu potencial
inseticida sobre o mosquito Ae. aegypti (SANTOS et al., 2017).
Essa planta possui valor comercial e social, de importância na
alimentação, construção, arte popular e combustível. Sementes
da Syagrus coronata, coletadas no estado de Pernambuco,
apresentaram atividade larvicida (CL50= 0,02 mg/mL, após 48h)
e deterrente-oviposição de fêmeas grávidas. A análise
fitoquímica desse óleo essencial demonstrou a presença
predominante de ácidos graxos, e os compostos ácidos
decanóico e dodecanóico são os responsáveis pela atividade

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
578
larvicida. Enquanto, o ácido octanóico é o responsável pela
ação deterrente-oviposição (SANTOS et al., 2017).
Em relação a estudos realizados com plantas do bioma
Caatinga, utilizando os caules como a parte anatômica,
diferentes extratos vegetais foram obtidos e investigado sua
propriedade inseticida. Paralelamente nesses estudos foram
identificados uma grande diversidade de princípios ativos. Por
exemplo, Oliveira et al. (2019), ao realizarem a prospecção
fitoquímica dos extratos etanólicos de caules de C.
argyrophyllus coletados em períodos chuvoso e seco,
identificaram substâncias químicas majoritárias diferentes entre
os extratos de diferentes períodos. No extrato obtido a partir de
caules do período chuvoso, as flavonas, flavonois, flavanonas
e xantonas, foram as principais substâncias encontradas,
enquanto no extrato oriundo de caules do período seco os
ácidos fixos fortes, esteróides livres e triterpenóides foram
identificados como majoritários. O extrato do período chuvoso,
mostrou-se mais tóxico sobre larvas de Ae. aegypti (CL50=6,16
mg/mL) quando comparado ao extrato seco (CL50=7,34
mg/mL), o que poderá está relacionado aos metabólitos
secundários da estação chuvosa, e consequentemente no seu
efeito tóxico sobre as larvas de Ae. aegypti. Ainda em relação
ao extrato obtido no período chuvoso os autores chamam a
atenção, que o período de chuva levaria a uma maior
disponibilidade de alimentos para os insetos herbívoros e o
ataque destes poderia estimular a produção de componentes
químicos de defesa do vegetal, com destaque para os
flavonoides que apenas foram detectados no extrato do período
chuvoso (OLIVEIRA et al., 2019).

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
579
O composto lectina obtido da casca de M. urundeuva, o
qual já tinha demonstrado atividade larvicida sobre Ae. aegypti,
apresentou também atividade ovicida sobre esse inseto. As
lectinas obtidas da casca e cerne de M. urundeuva exibiram
CL50 = 0,28 mg/mL e de CL50 = 0,80 mg/mL, repectivamente.
Através da análise ultraestrutural por microscopia eletrônica,
após os ovos de Ae. aegypti manterem contato com as lectinas
da casca e cerne de M. urundeuva por um período de 24 horas
a 72 horas, foi possível observar alterações morfológicas na
superfície de ovos de Ae. aegypti. Essas alterações incluíram o
surgimento de protuberâncias nos ovos, desaparecimento do
exocórion e redução dos tubérculos, o que indica, portanto, que
a lectina obtida dessa planta da Caatinga, conseguiu atingir o
trato digestivo do embrião (ALVES et al., 2020). A
caracterização desse efeito na fase de ovo do mosquito é de
grande importância, uma vez que esse representa o estágio de
maior resistência. Nesse estudo, os autores chamam a atenção
para o potencial dessa lectina em Programa Nacional de
Controle da Dengue, o qual não possui, no momento, nenhuma
substância com ação ovicida (ALVES et al., 2020).
Embora, não seja muito comum, há estudos que
utilizaram a raiz de planta da Caatinga para avaliar o potencial
larvicida sobre Ae. aegypti (CRUZ et al., 2015). Da raiz de
Poincianella bracteosa se obteve o extrato etanólico e a partir
desse foram obtidas as frações acetato de etila, hexânica e
diclorometânica. Dentre estas, a fração diclorometânica foi a
mais tóxica sobre as larvas desse mosquito. Através da análise
fitoquímica da raiz dessa planta os constituintes químicos
encontrados foram alcaloides, bases quaternárias, flavonoides,
xantonas, triterpenoides e taninos. Os autores sugerem que a

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
580
atividade larvicida seja devido a presença de triterpenoides,
flavonoides e alcaloides (CRUZ et al., 2015).
Algumas plantas além de possuir determinada atividade
biológica poderá apresentar também efeitos nocivos ao
Homem, e por isso torna-se imperativo conhecer também sobre
sua toxicidade em mamíferos, para assim promover o seu uso
como inseticida de forma segura à população. O óleo essencial
obtido de folhas de C. tetradenius foram muito eficazes contra
larvas e adultos de Ae. aegypti sendo necessária uma
concentração letal média de 0,15 mg/mL e de 1,85 mg/mL,
respectivamente. O principal componente identificado nesse
óleo essencial foi a cânfora, de um total de 26 compostos
encontrados. Os autores também investigaram a toxicidade
desse óleo essencial sobre mamíferos, o qual foi encontrada
uma Dose Letal média (DL50) de 500 mg/kg em camundongos
(CARVALHO et al., 2016). O óleo essencial obtido das folhas
de C. argyrophyllus também foi eficaz sobre o mosquito Ae.
aegypti, ao apresentar atividade sobre larvas e adultos com
uma concentração letal média de 0,3 mg/mL e 5,92 mg/mL,
respectivamente. Também nesse estudo foi avaliado a
toxicidade em camundongos, o que demonstrou sua segurança
frente a mamíferos (DL50= 2500 mg/kg). A análise fitoquímica
do óleo de C. argyrophyllus identificou um total de 45
compostos sendo o espatulenol, E-cariofileno alfa-pineno e
biciclogermacreno os componentes majoritários (CRUZ et al.,
2017). Em seguida, Cruz et al. (2020) ao avaliarem a atividade
conjunta destes dois óleos essenciais em forma de blend, com
vistas na obtenção de um produto inseticida mais ativo e mais
seguro, verificaram que quanto à atividade inseticida houve um
efeito sinérgico e aditivo sobre adultos e larvas de Ae. aegypti,

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
581
respectivamente. Em relação à toxicidade sobre mamíferos a
união desses óleos essenciais foi cinco vezes mais segura
quando comparada ao óleo individual de C. tetradenius (DL50=
2500 mg/kg), demonstrando assim que o blend se apresentou
mais seguro comparado ao óleo individual. Os autores
relacionaram essa menor toxicidade a uma redução quantitativa
de alguns componentes que podem ser neurotóxicos presentes
em maior proporção no óleo individual de C. tetradenius. A
análise fitoquímica do blend demonstrou que a cânfora,
isofenocampeol, E-cariofileno e espatulenol foram os principais
componentes, de um total de 36 compostos detectados.
Interessantemente este valor (36) é justamente o valor médio
do número de compostos somados das plantas individuais
(CRUZ et al., 2020). C. tetradenius e C. argyrophyllus foram
coletadas na região de Caatinga do estado da Bahia e são
popularmente conhecidas como “velame do campo” ou
“velandinho” para C. tetradenius e “velame falso” para C.
argyrophyllus (CRUZ et al., 2017, CARVALHO et al., 2016).
Nesse contexto, o bioma Caatinga constitui uma flora
diversificada e revela-se como fonte de espécies botânicas
detentoras de compostos bioativos, os quais podem ser úteis
para a formulação de inseticidas naturais, que podem vir atuar
no controle de insetos vetores, especialmente de Ae. aegypti
(OLIVEIRA et al., 2019). Além disso, a descoberta de novos
inseticidas de origem botânica poderá simultaneamente,
agregar valor e expandir conhecimentos sobre a biodiversidade
da Caatinga, e através da utilização sustentável dessa flora,
contribuir com o uso e a conservação da biodiversidade desse
bioma, bem como com o desenvolvimento socioeconômico da
região. A região nordeste do país, local que está inserida a

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
582
Caatinga, é uma das regiões mais pobres do Brasil e apresenta
os menores valores de Índice de Desenvolvimento Humano
(FERNANDES; QUEIROZ, 2018).
CONCLUSÕES
A diversidade vegetal encontrada na Caatinga estimula
a busca por compostos naturais detentores de atividade
biológicas. Nos últimos cinco anos diversos estudos têm
demonstrado o potencial inseticida de plantas da Caatinga
sobre Ae. aegypti, sendo a maioria deles verificado o potencial
sobre larvas desse mosquito. Além disso, alguns estudos
secundários recentes demonstram o mecanismo de ação
desses subprodutos em Ae. aegypti. Dentre as plantas
avaliadas observa-se que a maioria dos estudos utiliza as folhas
desses vegetais. A partir especialmente das folhas de plantas
da Caatinga, é detectada atividade inseticida principalmente
sobre larvas de Ae. aegypti, o que poderá simultaneamente,
agregar valor e expandir conhecimentos sobre a Caatinga.
REFERÊNCIAS ACHEE, N. L. et al. Alternative strategies for mosquito-borne arbovirus control. PLoS Negl.Trop. Dis. San Francisco, v. 13, n. 1, p. e0006822, 2019. ALVES, R. R. V et al. Ovicidal lectins from Moringa oleifera and Myracrodruon urundeuva cause alterations in chorionic surface and penetrate the embryos of Aedes aegypti eggs. Pest Management Sci. v. 76, n. 2, p. 730-736, 2020. AMOABENG, B. W. et al. Natural enemy enhancement and botanical insecticide source: a review of dual use companion plants. Applied Entomol. Zool. v. 54, n. 1, p. 1-19, 2019. ARAUJO, A. P. et al. Screening Aedes aegypti (Diptera: Culicidae) populations from Pernambuco, Brazil for resistance to temephos,

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
583
diflubenzuron, and cypermethrin and characterization of potential resistance mechanisms. J. Insect Sci. v. 19, n. 3, p. 16, 2019. ARAÚJO, J. R. S. et al. Larvicidal, cytotoxic and genotoxic effects of aqueous leaf extract of Jatropha mollissima (Pohl) Baill. Acta Scientiarum. Biological Sciences, v. 40, p. e34501-e34501, 2018. CARVALHO, K. S. et al. Toxicological evaluation of essential oil from the leaves of Croton tetradenius (Euphorbiaceae) on Aedes aegypti and Mus musculus. Parasitol. Res. v. 115, n. 9, p. 3441-3448, 2016. CAVALCANTE, A.; DUARTE, A. S.; OMETTO, J. P. H. Modeling the potential distribution of Epiphyllum phyllanthus (L.) Haw. under future climate scenarios in the Caatinga biome. Anais Acad. Bras. Ciências, v. 92, n. 2, p. 1-12, 2020. COE, H. H. G.; SOUSA, L. O. F. The Brazilian "Caatinga": Ecology and Vegetal Biodiversity of a Semiarid Region. In: GREER, F. E. (Ed.). Dry Forests: Ecology, Species Diversity and Sustainable Management. New York: Nova Science, 2014. p. 81-103. CONSOLI, R.A.G.B.; OLIVEIRA, R. L. Principais mosquitos de importância sanitária no Brasil. Rio de Janeiro: Editora Fiocruz, 1994. 228p. CRUZ, R. C. D.; et al. Phytochemical and toxicological evaluation of a blend of essential oils of Croton species on Aedes aegypti and Mus musculus. S. African J. Botany, v. 132, p. 188-195, 2020. CRUZ, R. C. D. et al. Bioatividade da raiz de Poincianella bracteosa (Tul.) LP Queiroz (Fabaceae) sobre larvas do Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae). Rev.Bras. Bioc. v. 13, n. 4, 2015. CRUZ, R. C. D. et al. Toxicological evaluation of essential oil from the leaves of Croton argyrophyllus (Euphorbiaceae) on Aedes aegypti (Diptera: Culicidae) and Mus musculus (Rodentia: Muridae). J. Med. Entomol. v. 54, n. 4, p. 985-993, 2017. FERNANDES, M. F.; QUEIROZ, L. P. D. Vegetação e flora da Caatinga. Ciência e Cultura, v. 70, n. 4, p. 51-56, 2018. GARCEZ, F. R. et al. A diversidade dos constituintes químicos da flora de Mato Grosso do Sul e sua relevância como fonte de substâncias bioativas. Rev. Virtual Quím. v. 8, n. 1, p. 97-129, 2016. ISMAN, M. B. Botanical insecticides in the twenty-first century-fulfilling their promise?. Annual Rev. Entomol. v. 65, p. 233-249, 2020. LESSA, T. et al. Known unknowns: Filling the gaps in scientific knowledge production in the Caatinga. PloS one, v. 14, n. 7, p. 219- 359, 2019. MESSINA, J. P. et al. The current and future global distribution and population at risk of dengue. Nat. Microbiol. v. 4, n. 9, p. 1508-1515, 2019. OLIVEIRA, G. P. et al. Avaliação do potencial larvicida de extratos etanólicos do caule de Croton argyrophyllus (Euphorbiaceae:

PLANTAS DA CAATINGA COMO UMA ALTERNATIVA PARA O CONTROLE DE AEDES AEGYPTI: UMA REVISÃO
584
Crotonoideae) sobre Aedes aegypti (Diptera: Culicidae). Rev. Bras. Bioc. v. 17, n. 2, p. 87-91, 2019. PAVELA, R. History, presence and perspective of using plant extracts as commercial botanical insecticides and farm products for protection against insects–a review. Plant Protection Sci. v. 52, n. 4, p. 229-241, 2016. QUEIROZ, L. P. et al. Diversidade e Evolução de Plantas com Flores do Domínio da Caatinga. In: SILVA, J. M. C.; LEAL, I. R.; TABARELLI, M. (eds) Caatinga. Springer, Cham. 2017. p. 23-63. SANTOS, L. M. M. et al. Fatty acid-rich volatile oil from Syagrus coronata seeds has larvicidal and oviposition-deterrent activities against Aedes aegypti. Physiol. Mol. Plant pathol. v. 100, p. 35-40, 2017. SILVA, A. M. A. et al. Chemical composition, larvicidal and cytotoxic activities of the leaf essential oil of Bauhinia cheilantha (Bong.) Steud. South African J. Botany, v. 131, p. 369-373, 2020. SILVA, J. M. C. et al. Caatinga: Compreendendo os desafios. In: SILVA, J. M. C, LEAL, I. R, TABARELLI, M. (eds) Caatinga. Springer, Cham. 2017. p. 3-19. SILVA, L. N. et al. Anti-infective effects of Brazilian Caatinga plants against pathogenic bacterial biofilm formation. Pharm. Biol. v. 53, n. 3, p. 464-468, 2015. SILVA, P. T. et al. Circadian Rhythm and larvicidal activity against Aedes aegypti of essential oils from Croton piauhiensis. Rev. Virtual Quím. v. 11, n. 6, p. 1682-1692, 2019. SHITAN, N. Secondary metabolites in plants: transport and self-tolerance mechanisms. Biosci. Biotechno. Biochem. v. 80, n. 7, p. 1283-1293, 2016. SIMÕES, E. R. et al. Biomedical properties and potentiality of Lippia microphylla Cham. and its essential oils. J. Intercultural Ethnopharm. v. 4, n. 3, p. 256-263. 2015. SOUZA, A. V. V. et al. Composição química do óleo essencial de folhas de Lippia schaueriana Mart. coletados na área da Caatinga. Molecules, v. 23, n. 10, p. 2480, 2018. SOUZA, T. M. et al. Further evidences for the mode of action of the larvicidal m-pentadecadienyl-phenol isolated from Myracrodruon urundeuva seeds against Aedes aegypti. Acta tropica, v. 152, p. 49-55, 2015. VIANA, F. F. O. et al. CARVALHO, M. G. Bioatividade dos extratos obtidos da parte aérea de Aspidosperma pyrifolium (Martius) sobre larvas de Aedes aegypti. Rev. Bras. Plantas Med. v. 18, n. 4, p. 804-810, 2016. WORLD HEALTH ORGANIZATION (WHO). Global vector control response 2017–2030. 2018. Disponível em: file:///C:/Users/R%C3%B4/Downloads/WHO_Resolution70_16%20(2).pdf. Acesso em: 19 ago. 2020..

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
585
CAPÍTULO 28
REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS
PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
Dayane Maximiano da SILVA1
Ronaldo Rodrigues SARMENTO2
Ana Carolina Bernardes DULGHEROFF3
1Aluna do curso técnico de Análises Clínicas, UFPB; 2Professor da ETS/UFPB; 3Orientadora/Professora da ETS/UFPB.
RESUMO: As enteroparasitoses são infecções causadas por helmintos e protozoários, que podem ser transmitidas por via fecal-oral ou por meio de penetração ativa da larva na pele. A areia das praias é uma potencial fonte de infecção em humanos, pois são comumente contaminadas por fezes de animais e humanos. No entanto, na maioria dos estados brasileiros, não é exigido o controle e divulgação da qualidade da areia das praias. Este estudo teve como objetivo, fazer uma revisão bibliográfica narrativa acerca da frequência de contaminação das areias das praias brasileiras por parasitos, descrevendo os principais enteroparasitos encontrados e metodologias utilizadas para detecção. Foi realizado um levantamento bibliográfico nas bases de dados SciELO, Google Acadêmico, Biblioteca Virtual da Saúde e MedLine de estudos publicados nos últimos 5 anos sobre a temática. Os estudos descrevem altos índices de contaminação das areias por parasitos, variando de cerca de 30% no Rio de Janeiro a 100% em Luís Correia. Os métodos mais comumente utilizados para detecção dos parasitos foram Lutz, Rugai e Baermann-Moraes. Os ancilostomídeos e ascarídeos, foram os parasitos mais

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
586
frequentemente encontrados. Diante dos resultados, se faz necessário a implementação de políticas eficazes e contínuas para controle da qualidade da areia das praias, como, assegurar um destino adequado as fezes, impedir a circulação de animais na areia das praias e tratar animais e humanos doentes. Palavras-chave: Contaminação Ambiental. Doenças Parasitárias. Educação Ambiental. Helmintos.
INTRODUÇÃO
As parasitoses intestinais são doenças cujos agentes
etiológicos são helmintos ou protozoários, os quais, em pelo
menos uma das fases do ciclo evolutivo, localizam-se no
aparelho digestivo do homem, e pode afetar todas as faixas
etárias, sendo mais predominante em crianças. Os danos que
os enteroparasitos podem causar em pessoas acometidas
incluem: diarreia, dores abdominais, inapetência, perda de
peso, e até processos obstrutivos, o que pode levar, em casos
extremos, ao óbito (CORRÊA et al., 2018).
As infecções parasitárias constituem um sério problema
de saúde pública no mundo, afetando cerca de 3,5 bilhões de
pessoas, e sendo responsáveis por 2 a 3 milhões de óbitos
anualmente (NETO; FARIA; MATOS-ROCHA, 2017).
No Brasil, este problema encontra-se presente em todas
as regiões, com destaque para as zonas rurais e periféricas
devido à ausência de saneamento básico. A prevalência
absoluta real dos casos de parasitoses no Brasil é
desconhecida, uma vez que essas infecções não são de
notificação compulsória, assim os dados de frequência são
decorrentes de estudos pontuais e de um inquérito nacional

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
587
sobre a prevalência dos geo-helmintos e de Schistosoma
mansoni. No último inquérito nacional publicado em 2018,
realizado de 2011 a 2015 em escolares de 7 a 17 anos, de
ambos os sexos, avaliou-se a presença de ovos de
ancilostomídeos nas fezes de 197.564 escolares (KATZ, 2018).
O parasito foi detectado nas fezes de 5.192 estudantes (2,73%),
sendo que as prevalências foram maiores nas regiões Norte e
Nordeste. As maiores frequências foram observadas no
Maranhão (15,79%), Pará (7,21%), Tocantins (6,06%), Sergipe
(6,62%), Paraíba (5,09%) e Bahia (4,23%).
A transmissão das enteroparasitoses, como amebíase,
giardíase e ascaridíase, ocorre através da ingestão de água ou
alimentos contaminados com suas respectivas formas
parasitárias e de estrongiloidíase e ancilostomose,
principalmente, pela penetração ativa das larvas infectantes
(presentes no solo) na pele. Assim condições higiênico-
sanitárias precárias, associadas à falta de tratamento adequado
de água e esgoto favorecem sua disseminação (CORRÊA et
al., 2018).
As praias são importantes focos de infecção humana por
parasitas intestinais, tanto através da água, quanto da areia,
pelo fato de poderem estar contaminadas com formas
infectantes dos parasitos. A contaminação desses locais pode
ocorrer pela má educação ambiental dos banhistas, por fezes
de animais, como pombos, cães, gatos e ratos, como também
pelo lixo e despejos de esgoto (NETO; FARIA; MATOS-
ROCHA, 2017).
A areia e a água são fortes indicadores da balneabilidade
das praias, no Brasil o Conselho Nacional do Meio Ambiente
(CONAMA) através da Resolução nº 274, de 29 de novembro

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
588
de 2000, no artigo 8°, recomenda que órgãos ambientais
avaliem as condições parasitológicas e microbiológicas da areia
e da água. Porém, como a legislação não exige o
monitoramento da areia, na grande maioria dos municípios
brasileiros, somente a qualidade da água é monitorada,
aumentando o risco de contaminação da população.
Altos índices de contaminação das areias das praias
brasileiras com parasitos de importância clínica têm sido
descritos (NETO; FARIA; MATOS-ROCHA, 2017; CORRÊA et
al., 2018). Na praia de Maceió foi observada uma frequência de
parasitos de 83,8% entre 2012 e 2015. O enteroparasito mais
frequentemente encontrado foi o Ancylostoma sp. (NETO;
FARIA; MATOS-ROCHA, 2017).
Dado o exposto, o objetivo deste trabalho foi realizar um
levantamento bibliográfico acerca do índice de contaminação
das areias das praias brasileiras por parasitos, relatando
também os principais enteroparasitos encontrados, podendo
dessa forma, servir de base para a implementação de uma
política nacional de controle da qualidade das areias das praias
brasileiras.
MATERIAIS E MÉTODO
Tratou-se de uma revisão bibliográfica narrativa com o
intuito de se fazer um levantamento de dados acerca do índice
de contaminação das areias do litoral brasileiro por parasitos.
As seguintes etapas foram seguidas para a elaboração
desta revisão: (1) definiu-se o tema a ser pesquisado; (2)
estabeleceu-se os descritores que seriam utilizados para
pesquisa e critérios de inclusão e exclusão de estudos; (3)

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
589
realizou-se a busca na literatura; (4) definiu-se quais as
informações que seriam extraídas dos trabalhos; (5) avaliou-se
quais estudos seriam incluídos na revisão; (6) realizou-se a
análise, síntese, comparação e exibição dos resultados; (7)
apresentou-se os resultados da pesquisa.
O levantamento bibliográfico foi realizado nas bibliotecas
virtuais: Scientific Eletronic Library Online (SciELO), Google
Acadêmico; Biblioteca Virtual da Saúde (BVS) e na National
Library of Medicine (MedLine).
Foram utilizados os seguintes descritores e suas
combinações na língua portuguesa e inglesa: “Parasitos”,
“Helmintos”, “Protozoários”, “Enteroparasitoses’’, “Praias” e
“Areias”. Como critérios de inclusão determinaram-se: trabalhos
de acesso livre na íntegra, em formato de artigo científico ou
trabalho de conclusão de curso, dissertações e teses,
publicados no idioma português e inglês. Foi escolhido como
recorte temporal o período de 2015 a 2020.
Tanto à análise quanto à síntese dos dados extraídos
dos artigos foram realizadas de forma descritiva, permitindo
notar, contar, descrever e classificar os dados, com o objetivo
de agrupar o conhecimento sobre o tema abordado nesta
revisão.
Foram extraídos dos trabalhos a cidade de realização do
estudo, o número de praias analisadas, o ano e método de
análise, a frequência absoluta de detecção de parasitos e os
parasitos intestinais mais frequentes em cada estudo.

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
590
RESULTADOS E DISCUSSÃO
Após a busca nas bases de dados e leitura dos trabalhos,
5 estudos foram incluídos nesta revisão, 1 descrevendo dados
da região Sudeste, 1 da região Norte e 3 da região Nordeste
(Tabela 1). Esses estudos descreveram a análise da areia de 2
praias do sudeste, uma praia do Norte e 9 praias do litoral
nordestino.
Todos os estudos descreveram um alto índice de
contaminação das areias das praias variando de cerca de 30%
(dependendo da estação do ano) nas praias do Rio de Janeiro,
RJ (AMARAL et al., 2015), a 100% nas praias de Luís Correia,
PI (CORRÊA et al., 2018).Apesar de não haver muitos estudos
disponíveis analisando a frequência de parasitos nas areias das
praias brasileiras e a grande maioria deles descrever uma
análise pontual da areia, altos índices de contaminação foram
descritos, dessa forma, servindo de alerta para o risco de
transmissão não só de doenças parasitárias, mas também de
outros agentes de transmissão fecal-oral, como os vírus de
gastroenterite (rotavírus, norovírus e enterovírus), o vírus da
hepatite A e bactérias relacionadas à diarreia (AN et al., 2020).
Em alguns estados e municípios do Brasil, foram criadas
leis que obrigam a divulgação da qualidade da areia das praias,
como por exemplo, a Lei nº 3.210 de 05 de abril de 2001, do
município do Rio de Janeiro e a Lei nº 14.366, de 15 de março
de 2011, do Estado de São Paulo. No entanto, na grande
maioria dos municípios brasileiros, a qualidade da areia não é
monitorada, somente da água.
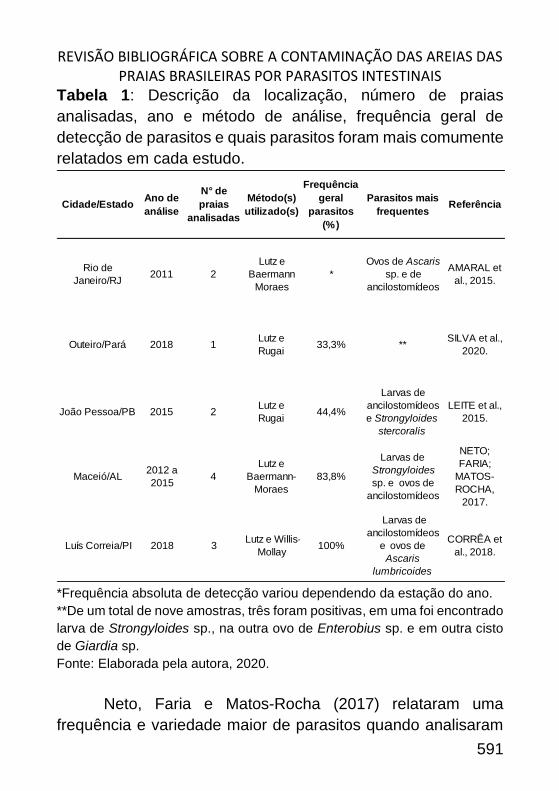
REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
591
Tabela 1: Descrição da localização, número de praias
analisadas, ano e método de análise, frequência geral de
detecção de parasitos e quais parasitos foram mais comumente
relatados em cada estudo.
Cidade/EstadoAno de
análise
N° de
praias
analisadas
Método(s)
utilizado(s)
Frequência
geral
parasitos
(%)
Parasitos mais
frequentes Referência
Rio de
Janeiro/RJ2011 2
Lutz e
Baermann
Moraes
*
Ovos de Ascaris
sp. e de
ancilostomídeos
AMARAL et
al., 2015.
Outeiro/Pará 2018 1Lutz e
Rugai 33,3% **
SILVA et al.,
2020.
João Pessoa/PB 2015 2Lutz e
Rugai 44,4%
Larvas de
ancilostomídeos
e Strongyloides
stercoralis
LEITE et al.,
2015.
Maceió/AL2012 a
20154
Lutz e
Baermann-
Moraes
83,8%
Larvas de
Strongyloides
sp. e ovos de
ancilostomídeos
NETO;
FARIA;
MATOS-
ROCHA,
2017.
Luís Correia/PI 2018 3Lutz e Willis-
Mollay100%
Larvas de
ancilostomídeos
e ovos de
Ascaris
lumbricoides
CORRÊA et
al., 2018.
*Frequência absoluta de detecção variou dependendo da estação do ano.
**De um total de nove amostras, três foram positivas, em uma foi encontrado
larva de Strongyloides sp., na outra ovo de Enterobius sp. e em outra cisto
de Giardia sp.
Fonte: Elaborada pela autora, 2020.
Neto, Faria e Matos-Rocha (2017) relataram uma
frequência e variedade maior de parasitos quando analisaram

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
592
a areia de dentro das barracas de praia, isto provavelmente se
deve ao maior fluxo de pessoas e animais neste local, o que
acaba favorecendo a contaminação do ambiente.
Leite et al. (2015) relataram uma frequência maior de
detecção de parasitos na profundidade de 10 cm (39,3%) e 20
cm (51,8%), quando comparada a análise da areia da superfície
(8,9%). O que provavelmente pode ser explicado, pelo fato de
as larvas migrarem para partes mais profundas da areia
buscando condições de umidade e temperatura mais favoráveis
para seu desenvolvimento. Em contraste com esse resultado,
Neto, Faria e Matos-Rocha (2017) também analisaram
amostras de areias de diferentes profundidades e não
observaram diferenças significativas de detecção dos parasitos.
No entanto, eles não fizeram uma análise separada da
frequência de protozoários e helmintos na areia de diferentes
profundidades, o que pode explicar essa discrepância entre os
resultados, já que são as larvas que migram para a parte mais
profunda da areia em busca de abrigo. Portanto, mais estudos
precisam ser realizados para avaliar se essa condição,
realmente se correlaciona com a variação de frequência de
larvas de parasitos.
Os métodos mais comumente utilizados para pesquisa
das formas parasitárias na areia foram: Lutz, Rugai e
Baermann-Moraes. Segundo Neto, Faria e Matos-Rocha (2017)
é de suma importância a utilização de vários métodos
parasitológicos ao analisar uma amostra, uma vez que as
formas parasitárias variam quanto ao seu peso e sobrevida no
meio exterior.
O método de Lutz ou Hoffman, Pons & Janer (HPJ) tem
como fundamento a sedimentação espontânea das formas
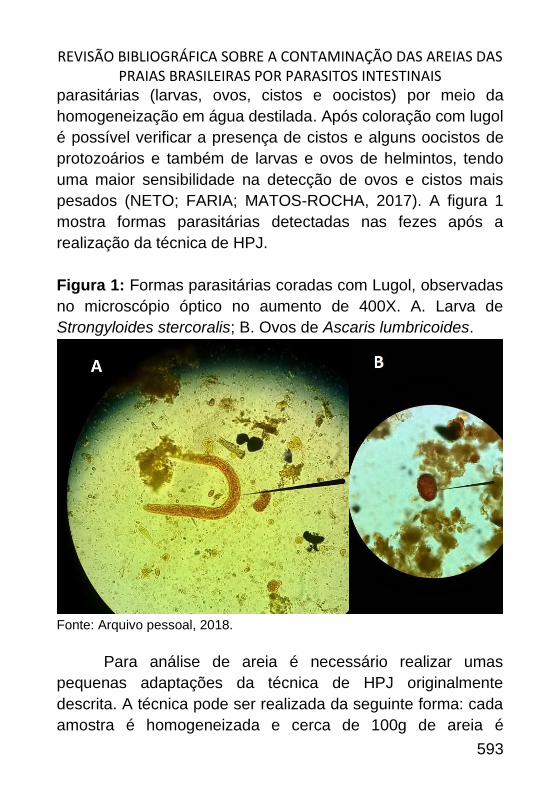
REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
593
parasitárias (larvas, ovos, cistos e oocistos) por meio da
homogeneização em água destilada. Após coloração com lugol
é possível verificar a presença de cistos e alguns oocistos de
protozoários e também de larvas e ovos de helmintos, tendo
uma maior sensibilidade na detecção de ovos e cistos mais
pesados (NETO; FARIA; MATOS-ROCHA, 2017). A figura 1
mostra formas parasitárias detectadas nas fezes após a
realização da técnica de HPJ.
Figura 1: Formas parasitárias coradas com Lugol, observadas
no microscópio óptico no aumento de 400X. A. Larva de
Strongyloides stercoralis; B. Ovos de Ascaris lumbricoides.
Fonte: Arquivo pessoal, 2018.
Para análise de areia é necessário realizar umas
pequenas adaptações da técnica de HPJ originalmente
descrita. A técnica pode ser realizada da seguinte forma: cada
amostra é homogeneizada e cerca de 100g de areia é

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
594
transferida para um recipiente contendo 100mL de detergente
neutro comercial a 0,5%. A solução é homogeneizada e deixada
em repouso por 5 minutos. O sobrenadante é filtrado com o uso
de gazes dobradas em quatro para um cálice de sedimentação
tendo seu volume completado até 250mL com água destilada.
Após 2 horas de sedimentação espontânea é feita a análise do
sedimento com lugol utilizando os aumentos de 100X e 400X.
De cada amostra devem ser realizadas no mínimo 2 lâminas
(AMARAL et al., 2015).
O método de Baermann-Moraes baseia-se no
hidrotropismo e termotropismo positivo das larvas vivas, sendo
usado para encontrar larvas vivas (NETO; FARIA; MATOS-
ROCHA, 2017). Amaral et al. (2015) observaram em seu
trabalho que a técnica de Baermann-Moraes foi mais sensível
para detecção de larvas, quando comparada com o método de
Lutz, identificando a presença de 31 larvas, enquanto no
método de Lutz o número de larvas encontradas foram 18.
A técnica de Rugai também é uma técnica utilizada para
detecção de larvas, que tem como princípio o hidrotropismo e
termotropismo das mesmas, porém de execução mais fácil que
a técnica de Baermann-Moraes (LEITE et al., 2015).
Os helmintos, especialmente os ancilostomídeos e
ascarídeos, foram os parasitos mais frequentemente
detectados. No entanto, cistos de protozoários também foram
detectados nas praias do Rio de Janeiro, Outeiro, Maceió e Luís
Correia (AMARAL et al., 2015; NETO; FARIA; MATOS-ROCHA,
2017; CORRÊA et al., 2018; SILVA et al., 2020). No caso do
estudo realizado por Leite et al. (2015) nas praias de João
Pessoa, somente foram detectadas larvas de helmintos. Na
grande maioria dos estudos, a frequência de detecção de

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
595
helmintos, é superior à de protozoários. Esses achados
reforçam a contaminação por diferentes formas parasitárias da
areia das praias brasileiras por fezes de humanos e animais.
Amaral et al. (2015) utilizando as técnicas de Lutz e
Baermann-Moraes, não conseguiram detectar protozoários na
areia das praias do Rio de Janeiro. No entanto, descrevam
elevadas frequências de oocistos de Cryptosporidium sp.
(82,9%) e de cistos de Giardia sp. (3,1%), quando analisaram
as mesmas amostras utilizando o ensaio imunoenzimático
(ELISA). A técnica de ELISA tem como princípio a detecção de
antígenos dos parasitos, portanto, não é necessário que os
parasitos estejam íntegros. No entanto, no caso dos métodos
de microscopia tradicionais é necessário que as formas
parasitárias estejam íntegras para que sejam identificadas.
Dessa forma, a técnica de ELISA é mais sensível para detecção
dos parasitos e, portanto, recomenda-se sua utilização para
monitoramento das areias das praias (AMARAL et al., 2015).
Os ancilostomídeos e ascarídeos são geo-helmintos, ou
seja, necessitam, obrigatoriamente, do solo para completar o
seu ciclo evolutivo. A Organização Panamericana da Saúde,
estima que no mundo 820 milhões de pessoas estão infectadas
por Ascaris lumbricoides, 460 milhões por Trichuris trichiura e
440 milhões por ancilostomídeos. Ancilostomose, conhecida
popularmente como Amarelão, a Ascaridíase e a Tricuríase são
algumas doenças causadas por esses parasitos, além da Larva
migrans cutânea (LMC) que é uma manifestação na pele de
infestação por ancilóstomo que causa intenso prurido, e os
sinais são pápulas e eritema no local da penetração, seguida
de um trajeto sinuoso por um túnel filiforme subcutâneo de cor
vermelho-amarronzada. Entre as complicações que podem

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
596
causar em humanos estão: o retardo do crescimento e a
redução do desenvolvimento mental em crianças e anemia,
especialmente entre mulheres em idade fértil (OPAS, 2018).
A infecção humana ocorre por meio da ingestão de água
ou alimentos contaminados com ovos embrionados ou pela
penetração, através da pele ou mucosas, de larvas infectantes
que se encontram no solo (OPAS, 2018). A alta frequência de
detecção desses parasitos nas praias reflete a contaminação
da areia com fezes humanas e/ou de animais. Ferraz et al.,
(2018) analisaram as fezes de cães presentes na orla das
praias do município de Pelotas/RS e dentre as 420 amostras
analisadas, detectaram parasitas em 266 (63,3%), sendo o
Ancylostoma o gênero mais frequentemente encontrado. Os
ovos de Ancylostoma sp. presentes no solo eclodem após
alguns dias, liberando a larva rabditóide em seu primeiro
estágio (L1), que diante de condições favoráveis de solo,
temperatura e umidade evolui para estágio L2 e posteriormente
L3, estágio em que a larva se encontra em sua forma infectante
e portanto são capazes de penetrar pela pele e causar doença.
Nesse estágio a larva recebe o nome de larva filarióide. Essa
forma pode sobreviver por longos períodos no solo em
condições ambientais favoráveis (PETENUSSE, 2018).
Entre os ascarídeos e ancilostomídeos relatados nos
artigos revisados, é possível destacar o isolamento de Toxocara
e Ancylostoma, respectivamente. Algumas espécies, como, T.
cati, T. canis, A. braziliense e A. caninum são parasitos comuns
de cães e gatos e podem acabar infectando humanos que
entram em contato. Apesar desses parasitos não serem
capazes de completar seu ciclo de desenvolvimento em seres
humanos, eles podem causar uma reação inflamatório no local

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
597
de penetração (A. braziliense e A. caninum), desencadeando a
síndrome da larva migrans cutânea; ou na superfície dos órgãos
ou olhos (T. cati e T. canis), desenvolvendo as síndromes da
larva migrans visceral ou larva migrans ocular, respectivamente
(ROCHA; WEBER; COSTA, 2019).
Os ancilostomídeos são pequenos vermes da família
Ancylostomatidae (classe Nematoda) e parasitos obrigatórios
de mamíferos, possuem aproximadamente 1 cm de tamanho,
tem uma ampla cavidade bucal, provida de lâminas (N.
americanus) e dentes (A. duodenale) que lhes permitem aderir
à parede do intestino do hospedeiro em busca de alimento,
tecido e/ou sangue (BRASIL, 2018; OPAS, 2018).
Enquanto os ascarídeos pertencem à família Ascarididae
do gênero Ascaris. É o geo-helminto que apresenta o maior
tamanho corporal, podendo atingir 30 cm. Conhecido
popularmente como “lombriga” ou “bicha”, é responsável pela
contaminação de 1,3 bilhões de pessoas no mundo. Tendo a
maior prevalência em países tropicais, onde o clima contribui
para o seu desenvolvimento. O ser humano é o único
hospedeiro do A. lumbricoides. A transmissão ocorre
geralmente ao se ingerir os ovos embrionados presentes em
água e/ou alimentos crus contaminados, como frutas, verduras
e legumes. As larvas passam por vários órgãos, como fígado e
pulmões e quando presentes em grandes quantidades no
intestino podem formar um bolo de parasitas que obstruindo a
passagem dos alimentos. Da mesma forma, pode haver
obstrução da faringe, com consequente crise de asfixia
(BRASIL, 2018; OPAS, 2018).
As larvas de Strongyloides sp. também foram
frequentemente relatadas nas areias das praias brasileiras. A

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
598
estrongiloidíase afeta cerca de 30 a 100 milhões de pessoas em
todo o mundo, principalmente de regiões tropicais e
subtropicais e de locais onde há exposição da pele ao solo
contaminado e condições higiênicas precárias. A infecção se
adquire quando as larvas filariformes, presentes na areia ou em
material contaminado com fezes humanas, penetram na pele e
migram até os pulmões, invadindo os alvéolos e seguem até o
intestino delgado, onde se transformam em parasitas adultos.
Do ponto de vista clínico o indivíduo pode apresentar sintomas
leves, diarreia moderada até hiperinfecção com choque séptico
em indivíduos com imunocomprometimento celular, podendo
ocasionar óbito (SANTANA; LOUREIRO, 2017).
Nesse contexto, faz-se necessário a adoção de medidas
de controle das parasitoses, começando pela conscientização
da população acerca da educação sanitária e ambiental através
de ações educativas, a fim de reduzir a contaminação do meio
(BRASIL, 2018; TEIXEIRA et al., 2020). Essas medidas incluem
por exemplo, não permitir que os animais domésticos defequem
nas areias das praias, que são áreas públicas destinadas à
recreação humana. Ao passear com os animais, é
recomendado utilizar as calçadas e levar sacos plásticos para
coletar as fezes do animal que devem ser descartadas no vaso
sanitário ao retornar para casa. Uma iniciativa interessante que
tem sido adotada em algumas cidades, é a criação de espaços
próprios para recreação dos animais. Ações de controle de
animais abandonados também precisam ser desenvolvidas. É
importante também, adotar as medidas de higiene pessoal,
como lavar as mãos com frequência e os alimentos antes
consumi-los. Além de, não descartar lixo em locais
inadequados. A contaminação do ambiente também pode ser

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
599
evitada, através do tratamento de humanos e animais doentes
com antiparasitário (BRASIL, 2018; OPAS, 2018).
O saneamento básico adequado, também é essencial
para impedir a contaminação da superfície da água e do solo
com ovos e larvas dos parasitos provenientes de fezes
humanas. O acesso a saneamento básico é um direito de todos
previsto na constituição federal, no entanto, precisa ser
efetivado. É necessário também, que seja realizada não só a
coleta, mas o tratamento do esgoto, antes do despejo nas
águas dos rios e do mar (TEIXEIRA et al., 2020).
As conhecidas línguas negras, que são o
extravasamento na praia de efluentes de galerias pluviais,
misturados ou não com esgotos, ou de esgoto bruto, de
qualquer origem, representam também uma importante fonte de
contaminação das praias (NETO; FARIA; MATOS-ROCHA,
2017). A Lei nº 11.445/2007, conhecida como Lei do
Saneamento, estabelece que a drenagem e o transporte da
água da chuva são realizados por meio das galerias pluviais,
que transportam essa água até os mananciais, evitando, assim,
alagamentos nas áreas urbanas. Porém, a falta de manutenção
e o acúmulo de lixo dos bueiros pode atrapalhar o manejo da
água da chuva, entupindo as galerias pluviais, impedindo que a
água da chuva seja transportada de forma adequada e poluindo
os rios e até as areia e água das praias. Nas principais praias
de João Pessoa por exemplo, é possível observar 8 galerias
pluviais, 3 na praia de Cabo Branco, uma em Tambaú, 3 em
Manaíra e duas na praia do Bessa (Figura 2).

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
600
Figura 2: Locais onde se encontram as galerias pluviais das
praias do Cabo Branco, Tambaú, Manaíra e Bessa de João
Pessoa, PB.
Fonte: Google Earth, 2020. Imagem gentilmente cedida por Glória Cristina
Cornélio do Nascimento.
Dessa forma, é indispensável uma atenção maior por
partes dos gestores no que diz respeito ao saneamento básico,
principalmente pela dificuldade de limpar a areia das praias
depois de contaminada, visto que, nos parques e lugares de
recreação é possível trocar a areia, mas em relação as praias a
única forma é a coleta de lixo, o saneamento adequado e a
educação ambiental da população.
CONCLUSÕES
Apesar de não haver muitos estudos disponíveis
analisando a frequência de parasitos nas praias brasileiras nos
últimos anos, foi possível observar altos índices de
contaminação das praias brasileiras por parasitos, tanto
protozoários, quanto helmintos. Foi descrita também, uma

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
601
frequência maior de parasitos nos locais com maior fluxo de
pessoas.
A maioria dos estudos utilizou a técnica de Lutz para
detecção das formas parasitárias em geral, mas também uma
técnica mais sensível para detecção de larvas, como Rugai ou
Baermann-Moraes. Portanto, recomenda-se a utilização de
mais de uma técnica para monitoramento das areias das praias,
além da utilização da técnica de ELISA, que é mais sensível
que as técnicas tradicionais de microscopia.
As formas parasitárias mais frequentemente
encontradas foram os ovos e larvas de ascarídeos e
ancilostomídeos, servindo dessa forma, como um importante
marcador de contaminação das praias por fezes de humanos e
animais.
Este trabalho aponta para a necessidade de: melhorias
das condições de saneamento básico, proibição de animais nas
praias, se trabalhar a educação ambiental e sanitária com às
pessoas que frequentam as praias do Brasil e implantação de
políticas de controle e divulgação da qualidade da areia das
praias do litoral brasileiro, que até o momento é facultativa, visto
que uma infecção por parasitas além de ocasionar prejuízo à
saúde da população em geral, especialmente crianças, trazem
problemas ao estado, em virtude dos gastos com o tratamento
da população afetada.
REFERÊNCIAS BIBLIOGRÁFICAS AMARAL, L. S et al. Parasitological Indicators of Contamination at Sand of Beaches and Monitoring by Traditional Methods and Immunoenzymatic Assay. Brazilian Journal of Biosystems Engineering, v.9, n.4, p. 304-311, 2015.

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
602
AN, X. L. et al. High-throughput diagnosis of human pathogens and fecal contamination in marine recreational water. Environ Res, v. 190, n.109982, 2020. BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Departamento de Vigilância das Doenças Transmissíveis. Guia Prático para o Controle das Geo-helmintíases [recurso eletrônico] / Ministério da Saúde, Secretaria de Vigilância em Saúde, Departamento de Vigilância das Doenças Transmissíveis. – Brasília: Ministério da Saúde, 2018. 33 p. : il. Modo de acesso: World Wide Web: http://bvsms.saude.gov.br/bvs/publicacoes/guia_pratico_controle_geohelmintiases.pdf. Acesso em: 02 out. 2020. CORRÊA, L. S. et al. Análise parasitológica das areias das praias do litoral de Luís Correia – Piauí. 2018. 46f. TCC (Graduação) - Curso de Biomedicina, Uninovafapi, Piauí, 2018. FERRAZ, A. et al. Ocorrência de parasitos gastrointestinais, em fezes de cães, encontradas na orla das praias de Pelotas, RS, Brasil. Atas de Saúde Ambiental (São Paulo, online), v. 6, p. 226-234, 2018. LEITE, L. C. A. Avaliação do nível de contaminação da areia de praias do Município de João Pessoa - pb por larvas de helmintos. 2015. 55f TCC (Graduação) – Curso de Farmácia, Universidade Federal da Paraíba, João Pessoa, 2015. KATZ, N. Inquérito nacional de prevalência da esquistossomose mansoni e geo-helmintoses. Belo Horizonte: CPqRR; 2018. Disponível em: <https://www.arca.fiocruz.br/bitstream/icict/25662/2/Inqu%c3%a9rito%20Nacional%20de%20Preval%c3%aancia%20da%20Esquistossomose%20mansoni%20e%20Geo-helmintoses.pdf>. Acesso em: 30 abr. 2020. NETO, J. J. G.; DE FARIAS, J. A. C.; MATOS-ROCHA, T. J. Contaminação de areia por parasitos de importância humana detectados nas praias da orla marítima de Maceió-AL. Arq Med Hosp Fac Cienc Med Santa Casa São Paulo, v.62, n. 2, p.81-84, 2017. ORGANIZAÇÃO PAN-AMERICANA DE SAÚDE (OPAS). Quimioterapia preventiva para o controle de infecções por helmintos transmitidos pelo contato com o solo em grupos de risco. Genebra, 2018. Disponível em:<https://iris.paho.org/bitstream/handle/10665.2/49072/9789275719947_por.pdf?sequence=5&isAllowed=y.> Acesso em: 30 abr. 2020. PETENUSSE, C. S. Contaminação por helmintos em parques e bosques municipais de Goiânia, Goiás. 65p. Dissertação (Mestrado) - Universidade Federal de Goiás, Escola de Veterinária e Zootecnia (EVZ), Programa de Pós-Graduação em Ciência Animal, Goiânia, 2018.

REVISÃO BIBLIOGRÁFICA SOBRE A CONTAMINAÇÃO DAS AREIAS DAS PRAIAS BRASILEIRAS POR PARASITOS INTESTINAIS
603
ROCHA, M. J.; WEBER, D. M.; COSTA, J. P. Prevalência de larvas migrans em solos de parques públicos da cidade de Redenção, estado do Pará, Brasil. Rev Pan-Amaz Saude, Ananindeua, v. 10, e201901607, 2019. SANTANA, A. T. T.; LOUREIRO, M. B. Síndrome de hiperinfecção e/ou disseminação por Strongyloides stercoralis em pacientes imunodeprimidos. RBAC, v. 49, n. 4, p.351-8, 2017. TEIXEIRA, P. A. et al. Parasitoses intestinais e saneamento básico no Brasil: estudo de revisão integrativa. Braz. J. of Develop., Curitiba, v. 6, n. 5, p.22867-22890, 2020.
.

604
GENÉTICA DE MICRORGANISMOS

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
605
CAPÍTULO 29
MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E
DERIVADOS
Aline Cardoso CARLOS1
1 Doutora em Bioquímica, UFRN;
RESUMO: Os resíduos gerados de atividades industriais relacionadas a exploração do petróleo representam um passivo ambiental de grandes proporções, os quais carecem de tratamento adequado. Desta forma, tem sido crescente nos últimos anos o desenvolvimento de estratégias voltadas para a biorremediação ambiental. Nesse contexto, o presente trabalho teve com objetivo isolar microrganismos associados à biodegradação de hidrocarbonetos e síntese de biossurfactantes, a partir de consórcios microbianos provenientes do resíduo de fluido de perfuração de poço de petróleo. Os microrganismos isolados foram analisados quanto as atividades de degradação de hidrocarbonetos e síntese de biossurfactantes, por meio do crescimento em petróleo, índice de emulsificação e hidrofobicidade. Todos os testes utilizados mostraram atividades positivas para degradação do petróleo. Portanto, a abordagem metagenômica utilizada no presente trabalho, mostrou-se uma boa estratégia para a prospecção de microrganismos atuantes na degradação de hidrocarbonetos do petróleo.
Palavras-chave: Biorremediação. Degradação de
hidrocarbonetos. Síntese de biossurfactantes.

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
606
INTRODUÇÃO
O petróleo é uma das principais fontes de energia e
matéria-prima nos dias atuais, com grande importância em
diversos setores, presente tanto em combustíveis, como em
fertilizantes, plásticos, tintas, borracha, entre outros materiais.
Entretanto, sua produção em larga escala, uso, transporte e
disposição no mundo inteiro tornaram-no um importante
contaminante no meio ambiente (WANG et al., 2010).
Vazamentos acidentais com navios petroleiros ou
plataformas de petróleo, descarte, estocagem inadequados e
uso incorreto de seus derivados, provocam uma série de
problemas ambientais como a contaminação do solo e da água
por hidrocarbonetos. Dessa forma, o
tratamento/descontaminação de ambientes contaminados com
esses compostos é um grande desafio na atualidade (RAHMAN
et al., 2002).
Dentre os resíduos que são gerados pela exploração de
petróleo está a lama do fluído de perfuração, que tem como
função o fornecimento da pressão hidrostática para impedir que
os fluídos do reservatório entrem na abertura do poço durante
a perfuração, manter a broca de perfuração resfriada e limpa
durante esse processo, e suspender o cascalho gerado quando
a perfuração for interrompida (HERZHAFTet al., 2006).
Anualmente, milhões de toneladas desses resíduos são
gerados em todo mundo. Em 2010, a Shell Brasil Petróleo Ltda.
gerou cerca de 2.960,870 toneladas de resíduo de fluido de
perfuração. Embora não exista uma lei para gerenciamento e
descarte desse resíduo, este é classificado como perigoso
(classe I), de acordo com a norma NBR: 10004:2004 da
Associação Brasileira de Normas Técnicas (ABNT). O descarte

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
607
inadequado desses resíduos no meio ambiente, pode causar
sérios problemas ambientais, como contaminação do solo e da
água (GUIMARÃES, 2007), uma vez que podem conter
hidrocarbonetos alifáticos e policíclicos, bifenispoliclorinados e
metais pesados, tais como bário, chumbo, zinco, mercúrio,
cromo, arsênico e níquel (WOJTANOWICZ, 1997; NEFF, 2005),
dependendo dos compostos químicos do fluido utilizado e da
composição das rochas que serão perfuradas.
O manejo do cascalho gerado durante a perfuração
representa um problema para a indústria de óleo e gás devido
à quantidade gerada e seu conteúdo de contaminantes
orgânicos e inorgânicos. A cada ano, milhões de toneladas
desses resíduos são geradas em todo mundo, e a atenuação
do dano ambiental vem sendo realizada com a utilização de
diferentes metodologias como: a solidificação e estabilização
(LEONARD E STEGEMANN, 2010), tratamentos térmicos
(ZUPAN e KAPILA, 2000) e com microondas (ROBINSON et al.,
2010), fitodegradação (JI et al., 2004) e bioremediação
(OKPARANMA et al., 2009). As opções de manejo utilizadas
são dispendiosas, como a reinjeção do fluído (MINTON, 2004),
tratamento térmico (STEPHENSON et al., 2004) e incineração
(VEIL, 2002), requerendo uso intensivo de energia, ou
ambientalmente insustentável, como a disposição em aterros
(VEIL et al.,1996) e a reutilização na construção sem um
tratamento prévio (CORDAH, 1999).
Nesse contexto, os processos biológicos são
interessantes alternativas, principalmente devido à simplicidade
e eficiência de custo quando comparados a outros métodos
tradicionais (MARIANO, 2005). A biorremediação microbiana
vem sendo pesquisada como processo biológico promissor
para degradação de componentes do petróleo e derivados bem

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
608
como para a recuperação ambiental (IWAMOTO; NASU, 2001).
Ela consiste na utilização de microrganismos com potencial
metabólico para transformar ou mineralizar diversos
contaminantes ambientais, como os hidrocarbonetos do
petróleo. A biotransformação consiste na degradação parcial do
contaminante com formação de outros compostos que podem
ser ou não menos tóxicos que o composto original. A
mineralização, representa a degradação completa do composto
em substâncias inertes, como CO2 e H2O (SILVA, 2002). Esta
estratégia tem sido utilizada em escala comercial no tratamento
de diversos ambientes contaminados (BAMFORTH;
SINGLETON, 2005) e pode ser realizada in situ ou ex situ. Na
biorremediação in situ não há remoção do material a ser
remediado, sendo o processo realizado no próprio local
contaminado. Esta técnica tem sido comumente utilizada por
apresentar maior custo benefício, podendo ser aplicada nos
mais diversos ambientes, mesmo naqueles onde a
biorremediação ex situ seria inacessível (PANDEY; CHAUHAN;
JAIN, 2009). Na técnica de biorremediação ex situ, ocorre com
a remoção do solo, sendo realizada em outro local, como em
um biorreator, por exemplo (NANO et al., 2003). Nesta técnica
há a independência de fatores ambientais que poderiam afetar
a eficácia da degradação. Sendo assim, esse é o processo mais
utilizado para remediação em ambientes que não apresentam
condições favoráveis á degradação por processos biológicos
(PANDEY; CHAUHAN; JAIN, 2009).
Espécies microbianas que atuam na biorremediação dos
hidrocarbonetos são largamente distribuídas na natureza e não
são restritas a apenas alguns gêneros (MORAIS e TAUK-
TORNISIELO, 2009). Uma ampla variedade de
microrganismos, genes e enzimas têm sido descritos como

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
609
envolvidos em degradação de hidrocarbonetos, incluindo
bactérias aeróbias, anaeróbias facultativos, redutoras de sulfato
e metanogênicas (SETTE et al., 2007; TAKAI et al., 2002;
FUCHS et al., 2011).
O processo de biorremediação pode ocorrer por: a)
atenuação natural: processo no qual a própria microbiota nativa
dos ambientes contaminados, possui capacidade de degradar
e transformar os contaminantes em substâncias inertes
(JACQUES et al., 2007); b) bioestimulação: usada com o intuito
de acelerar e favorecer o processo de degradação. Na
bioestimulação podem ser adicionados nutrientes e corrigidos
os parâmetros físico-químicos como, umidade, temperatura, pH
(SILVA, 2002; JOHNSEN et al., 2005); c) bioaumentação:
processo que acelera o processo de degradação por meio da
introdução de microrganismos autóctones (próprios do
ambiente) ou alóctones (advindos de outros locais) com
reconhecido potencial metabólico de degradação ou de
produção de biossurfactantes (IWAMOTO; NASU, 2001;
ŁAWNICZAK et al., 2013). Alguns autores tem sugerido a
utilização da bioaumentação autóctone, através da adição de
microrganismos exclusivamente nativos ao local a ser
remediado (HOSOKAWA et al., 2009).
Dessa forma, conhecer a biodiversidade microbiana
autóctone e sua capacidade de degradação de hidrocarboneto
e outros contaminantes ambientais têm sido considerados
essenciais para o desenvolvimento de estratégias de
biorremediação mais eficientes e seguras.
Apesar de a maioria dos componentes do petróleo serem
biodegradáveis, cerca de 10% a 40% em estado bruto ou
refinado é recalcitrante, o que representa toneladas de
poluentes impactando ecossistemas e sendo bioacumulados e

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
610
biomagnificados na cadeia trófica (CRAPEZ et al., 2002).
Ademais a biodegradação do petróleo pelos microrganismos no
solo é muitas vezes prejudicada devido à baixa solubilidade de
alguns dos hidrocarbonetos do petróleo, ocasionando menor
taxa de biodegradabilidade. Assim, a utilização de
biossurfactantes auxilia na biodisponibilidade dos
hidrocarbonetos, aumentando, dessa forma, a eficiência de
biodegradação. Os biossurfactantes são moléculas anfifílicas
que consistem em subprodutos metabólicos de bactérias,
fungos e leveduras (NITSCHKE; PASTORE, 2002). Essas
moléculas possuem como principais vantagens em relação aos
surfactantes sintéticos, baixa toxicidade, alta degradabilidade,
alta capacidade emulsificante e excelente atividade interfacial e
superficial (DA SILVA et al., 2014; NITSCHKE; PASTORE,
2002). A utilização de biossurfactantes têm se mostrado
bastante promissora na indústria do petróleo, seja como
coadjuvante em processos de biorremediação ou mesmo em
processos de recuperação avançada em reservatórios de
petróleo (DA SILVA et al., 2014). A principal função dos
biossurfactantes nos processos de biorremediação é a redução
da tensão superficial e interfacial, aumentando a disponibilidade
do substrato, permitindo que os microrganismos cresçam
através da absorção e metabolismo desses substratos. Além
disso, possuem capacidade de emulsionar a fonte de carbono
e estão envolvidos na adesão de células microbianas a
hidrocarbonetos (SANTOS et al., 2016).
Muitos são os estudos que têm relatado a eficiência dos
biossurfactantes em processos de biorremediação (SOBRINHO
et al., 2008; BATISTA et al., 2010; YU et al., 2014; LUNA et al.,
2015). Zhang e colaboradores (2012) mostraram que a
suplementação com ramnolipídeos em solo contaminado com

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
611
petróleo, reduziu em 86,97% a presença de hidrocarbonetos no
solo. Outro estudo utilizando suplementação com
ramnolipídeos foi realizado por Sponza e Gök (2010), que
observaram redução de 90% de hidrocarbonetos
poliaromáticos. Remoção de óleo motor do solo e da água do
mar por biossurfactante produzido por Candida sphaericapara,
mostrou redução de 75% e 92% respectivamente. Na água do
mar o biossurfactante mostrou eficiência de 75% de
espalhamento do óleo, apresentando potencial de aplicação em
processos de descontaminação ambiental (LUNA et al., 2015).
A biodegradação é uma técnica viável e econômica para
a remediação de ambientes contaminados com
hidrocarbonetos do petróleo. Entretanto, fatores como
temperatura, nutrientes, salinidade e tipo de solo, influenciam
as taxas de biodegradação (VARJANI et al., 2017). A
temperatura exerce uma função primordial, ao provocar
alterações no estado físico dos hidrocarbonetos e na taxa de
crescimento dos microrganismos. Além de aumentar a
solubilidade dos poluentes e diminuir a viscosidade. Em baixas
temperaturas a viscosidade do poluente aumenta, a
volatilização de alcanos tóxicos de cadeia curta e a solubilidade
na água diminuem (VARJANI; UPASANI, 2017). Concentrações
dos nutrientes fósforo, nitrogênio e carbono devem ser
significativas para o crescimento microbiano adequado e
consequente sucesso do processo de degradação. A proporção
C:N:P de 100:10:1 tem sido comumente utilizada afim de
garantir um meio equilibrado para o crescimento microbiano
(DIAS et al., 2012). Elevadas concentrações salinas diminuem
a disponibilidade dos hidrocarbonetos, em razão dos sais
solúveis reduzirem a solubilidade dos compostos orgânicos
hidrofóbicos na água. Apesar disso, ambientes hipersalinos não

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
612
são um impedimento ao processo de degradação, uma vez que
a atividade microbiana só é afetada por flutuações de sal em
concentrações abaixo ou acima do crescimento ótimo dos
microrganismos em estudo (MARTINS; PEIXOTO, 2012). Os
tipos de solos são de grande importância para o sucesso da
biodegradação. Solos argilosos possuem baixa
permeabilidade, o que dificulta a distribuição e transporte de
nutrientes, oxigênio e água, ocasionando baixa taxa de
degradação. Já os solos arenosos, possibilitam melhor
transferência de oxigênio e consequentemente maior taxa de
degradação.
Dentro deste contexto, o objetivo do nosso trabalho foi
isolar microrganismos que tenham atividade de degradação dos
principais constituintes do petróleo (hidrocarbonetos) e
produção de biossurfactantes. Uma vez que microrganismos
com essas propriedades são de fundamental importância no
estabelecimento das condições ótimas para a degradação
destes compostos, podendo contribuir com o desenvolvimento
de estratégias em processos de biorremediação e
consequentemente redução dos impactos ambientais.
MATERIAIS E MÉTODO
A amostra utilizada consiste no resíduo de fluido de
perfuração de poço de petróleo, que foi gentilmente cedido pela
empresa Brasil Química e Mineração Industrial Ltda (BQMIL),
localizada no município de Mossoró-RN (05°11′16″S de latitude
e 37°20′38″W de longitude). A amostra foi coletada pela
empresa durante a perfuração da fase de um poço terrestre. A
coleta foi realizada do início até o fim da perfuração do poço.

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
613
Após a coleta, esse resíduo foi transferido para embalagens
plásticas e transportado até o laboratório obtenção dos isolados
microbianos.
Obtenção de bactérias isoladas
As bactérias isoladas foram obtidas a partir dos consórcios
L e Y provenientes do resíduo de fluido de perfuração de poço
de petróleo (GUERRA et al., 2018). Para tanto, foram
adicionados em frascos Erlenmeyers 50 mL de meio Bushnell-
Haas (BH), 1% da cultura dos consórcios e 1% de petróleo bruto
estéril. Em seguida, foram levados ao shaker a 180 rpm e 37 °C
durante 24 horas. Após esse período, 100 µL da cultura foi
utilizada no plaqueamento pelo método spread plate em placas
com BH ágar, 1% de petróleo e cloreto de trifeniltetrazólio (TTC)
(10 mL de solução a 1% / L), reagente indicador da atividade de
degradação do petróleo (OLGA et al., 2008). As placas foram
transferidas para a estufa a 37°C durante 7 dias. Após esse
período, as colônias com coloração rósea (indicativo de
degradação) foram selecionadas e transferidas para tubos
cônicos contendo 2 mL de meio LB ou YPD e levadas ao shaker
a 37 °C e 180 rpm. Após 48 horas (período com maior
crescimento), os isolados foram novamente plaqueados em
meio rico ao qual foram inicialmente selecionados (LB ou YPD)
contendo ágar, visando à confirmação do isolamento mediante
a observação das colônias. Após a confirmação do isolamento
das culturas, estoques em glicerol (50%) foram preparados e,
em seguida, foram armazenados a -20 °C até a utilização.
Crescimento das bactérias isoladas em petróleo

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
614
O crescimento das bactérias isoladas na presença de
petróleo foi determinado em meio Bushnell Haas (BH). Para
tanto, as bactérias foram inicialmente ativadas (pré-inóculo) em
frascos Erlenmeyers de 250 mL, contendo 50 mL de meio
mineral Bushnell Haas (BH), 1% da bactéria isolada (estoque)
e 1% de petróleo bruto estéril (v/v) como única fonte de carbono.
Os frascos foram levados ao shaker à 180 rpm e 37 ºC. A cada
24 horas a densidade óptica a 600 nm era medida em
espectrofotômetro. Como controles, foram preparados
Erlenmeyers nas mesmas condições acima, entretanto, sem a
presença de petróleo ou nenhuma outra fonte de carbono
adicional.
Índice de emulsificação (IE24)
A bactérias isoladas foram avaliadas quanto à capacidade
de produção de biossurfactantes por meio do teste de
emulsificação seguindo o protocolo de Iqbal et al. (1995) com
modificações. Inicialmente, 2 mL de querosene foram
adicionados em 2 mL de cultura crescida em diferentes fontes
de hidrocarbonetos (hexano, hexadecano, diesel, querosene,
petróleo, tolueno, xileno) e, em seguida agitados em vortex a
alta velocidade durante 2 min deixando-se repousar durante 24
horas. O índice de emulsificação (IE24%) foi calculado pela
razão entre a altura da camada de emulsão e a altura total do
sistema (medidas com paquímetro) multiplicada por 100.
E24% = altura da camada emulsificada x 100
altura total do sistema

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
615
Hidrofobicidade
A hidrofobicidade celular dos microrganismos isolados foi
determinada através da medida de adesão a hidrocarbonetos
(MATH), seguindo o protocolo descrito por Rosenberg (1984)
com modificações, conforme descrito a seguir. Para tanto,
inicialmente os isolados foram crescidos e as células lavadas
em tampão fosfato/ureia/magnésio (composição: 22,2 g/L de
K2HPO4.H2O; 7,26g/L de KH2PO4; 0,2 g/L de MgSO4.7H2O e
1,8g/L de ureia). Esse mesmo tampão foi utilizado para
ressuspensão das células dos isolados após lavagem para
correção da densidade óptica inicial (DO600) entre 0,6 a 0,8.
Foram adicionados 2,4 mL de suspensão celular em tubos de
vidro contendo 0,4 mL de cada hidrocarboneto testado
(hexadecano, diesel, querosene, xileno, tolueno). Em seguida,
os tubos foram agitados em vortex por 2 minutos e repousaram
durante 20 minutos. Após esse tempo, a densidade óptica foi
novamente medida e os valores de hidrofobicidade calculados
através da seguinte equação:
100 x {1- (DO da fase aquosa/DO da suspensão celular
inicial)}
Análise estatística
Todos os ensaios realizados foram feitos em triplicatas.
A significância estatística dos resultados, foram determinadas
utilizando a análise de variância (ANOVA) e aplicação de teste
para comparação de médias (de triplicatas) pelo teste de Tukey

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
616
(p<0,05). As análises estatísticas e gráficos, foram gerados
utilizando o programa GraphPad Prism5 e 7.
RESULTADOS E DISCUSSÃO
Ambientes contaminados com petróleo têm se revelado
como grandes bancos genéticos, contendo uma vasta
diversidade de microrganismos, que possibilita a descoberta de
novos microrganismos de interesse biotecnológico. Dessa
forma, a proposta do presente estudo foi identificar
microrganismos que possam contribuir com estratégias de
biorremediação de ambientes contaminados com petróleo.
Para isso, foram isolados microrganismos a partir de consórcios
provenientes do resíduo de perfuração de petróleo,
caracterizado por possuir espécies e vias metabólicas
relacionadas com a degradação de petróleo e síntese de
biossurfactantes (GUERRA et al., 2018).
Dois dos isolados obtidos (isolados R4 e R7) foram
submetidos a testes preliminares de degradação de
hidrocarbonetos e produção de biossurfactantes, os resultados
são apresentados a seguir. O sequenciamento e análise
taxonômica é apresentado na sequência.
Avaliação do crescimento na presença de petróleo
A presença de petróleo parece ser essencial para o
crescimento dos isolados, uma vez que maiores crescimentos
foram alcançados ou mantiveram-se na presença deste.
Ambos os isolados (R4 e R7) revelaram aumento do
crescimento na presença de petróleo e diminuição na sua
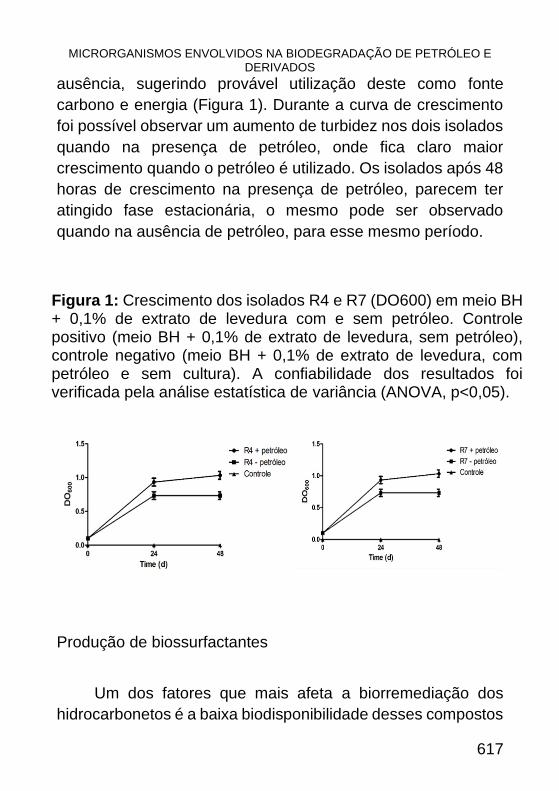
MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
617
ausência, sugerindo provável utilização deste como fonte
carbono e energia (Figura 1). Durante a curva de crescimento
foi possível observar um aumento de turbidez nos dois isolados
quando na presença de petróleo, onde fica claro maior
crescimento quando o petróleo é utilizado. Os isolados após 48
horas de crescimento na presença de petróleo, parecem ter
atingido fase estacionária, o mesmo pode ser observado
quando na ausência de petróleo, para esse mesmo período.
Produção de biossurfactantes
Um dos fatores que mais afeta a biorremediação dos
hidrocarbonetos é a baixa biodisponibilidade desses compostos
Figura 1: Crescimento dos isolados R4 e R7 (DO600) em meio BH + 0,1% de extrato de levedura com e sem petróleo. Controle positivo (meio BH + 0,1% de extrato de levedura, sem petróleo), controle negativo (meio BH + 0,1% de extrato de levedura, com petróleo e sem cultura). A confiabilidade dos resultados foi verificada pela análise estatística de variância (ANOVA, p<0,05).

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
618
aos microrganismos. Os hidrocarbonetos, principalmente os
HPAs possuem baixa solubilidade aquosa e forte tendência a
sorção no solo, que reduzem sua biodisponibilidade (GHOSAL
et al., 2016). A utilização de biossurfactantes produzidos por
alguns microrganismos, promove redução das tensões super e
interfacial, aumentando a solubilidade e biodisponibilidade dos
hidrocarbonetos (PACWA-PŁOCINICZAK et al., 2011) e
consequentemente, contribuindo para o processo de
biorremediação. Bosch et al (1988) consideram cepas como
boas produtoras de biossurfactantes com propriedade
emulsificante aquelas que formam emulsões superiores a 40%.
Neste estudo, foram observados índices de emulsificação
superiores à 40%, sugerindo produção de
biossurfactantes/bioemulsificantes. Os isolados R4 e R7
apresentaram emulsificação de 45% e 55%, respectivamente.
O SDS (10%), utilizado como controle, apresentou valor médio
de 70% (Figura 2).
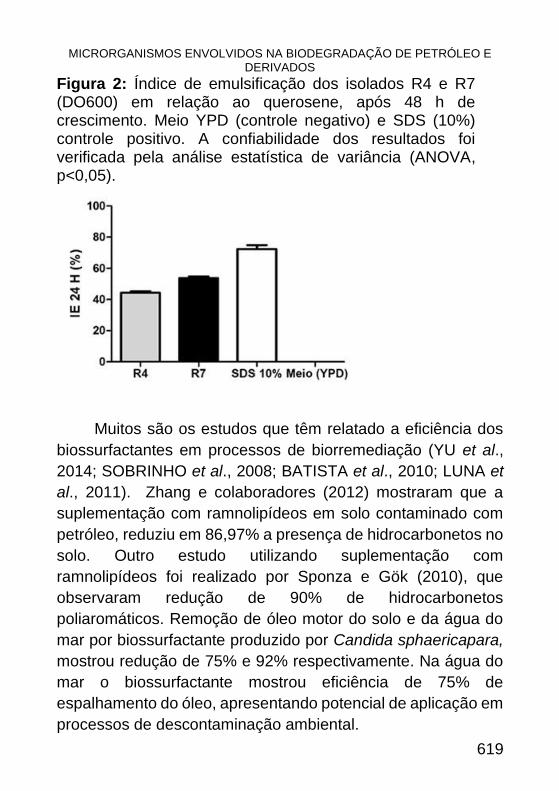
MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
619
Figura 2: Índice de emulsificação dos isolados R4 e R7 (DO600) em relação ao querosene, após 48 h de crescimento. Meio YPD (controle negativo) e SDS (10%) controle positivo. A confiabilidade dos resultados foi verificada pela análise estatística de variância (ANOVA, p<0,05).
Muitos são os estudos que têm relatado a eficiência dos
biossurfactantes em processos de biorremediação (YU et al.,
2014; SOBRINHO et al., 2008; BATISTA et al., 2010; LUNA et
al., 2011). Zhang e colaboradores (2012) mostraram que a
suplementação com ramnolipídeos em solo contaminado com
petróleo, reduziu em 86,97% a presença de hidrocarbonetos no
solo. Outro estudo utilizando suplementação com
ramnolipídeos foi realizado por Sponza e Gök (2010), que
observaram redução de 90% de hidrocarbonetos
poliaromáticos. Remoção de óleo motor do solo e da água do
mar por biossurfactante produzido por Candida sphaericapara,
mostrou redução de 75% e 92% respectivamente. Na água do
mar o biossurfactante mostrou eficiência de 75% de
espalhamento do óleo, apresentando potencial de aplicação em
processos de descontaminação ambiental.
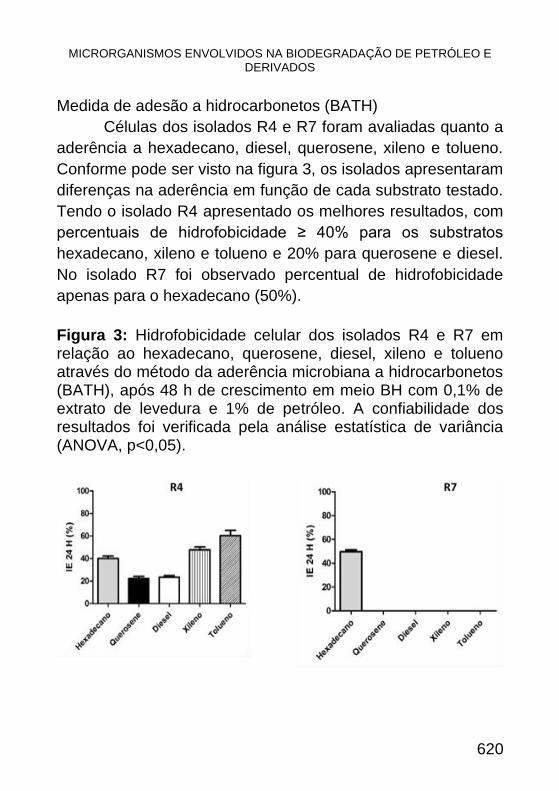
MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
620
Medida de adesão a hidrocarbonetos (BATH)
Células dos isolados R4 e R7 foram avaliadas quanto a
aderência a hexadecano, diesel, querosene, xileno e tolueno.
Conforme pode ser visto na figura 3, os isolados apresentaram
diferenças na aderência em função de cada substrato testado.
Tendo o isolado R4 apresentado os melhores resultados, com
percentuais de hidrofobicidade ≥ 40% para os substratos
hexadecano, xileno e tolueno e 20% para querosene e diesel.
No isolado R7 foi observado percentual de hidrofobicidade
apenas para o hexadecano (50%).
Figura 3: Hidrofobicidade celular dos isolados R4 e R7 em relação ao hexadecano, querosene, diesel, xileno e tolueno através do método da aderência microbiana a hidrocarbonetos (BATH), após 48 h de crescimento em meio BH com 0,1% de extrato de levedura e 1% de petróleo. A confiabilidade dos resultados foi verificada pela análise estatística de variância (ANOVA, p<0,05).

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
621
CONCLUSÕES
Os isolados obtidos visando a degradação de
hidrocarbonetos e síntese de biossurfactantes, mostraram-se
bastante promissores nos testes, sendo, portanto, de grande
interesse em processos de biorremediação.
REFERÊNCIAS BIBLIOGRÁFICAS
BOSCH, M.P., et al. Surface-active compounds on microbial cultures. Tenside Surfactants Detergents. 25 (4): 208-211, 1988.
CORDAH, Research on the re-use of drill cuttings onshore. Anne Manning and Marianne Lang, Report No: Cordah/COR012/1999, CORDAH Research Limited.1999
CRAPEZ, M.A.C., BORGES, A.L.N., BISPO, M.G.S., PEREIRA, D.C.. Biorremediação: tratamento para derrames de petróleo. Ciência hoje, 30: 179.2002
FUCHS, G.; BOLL, M.; HEIDER, J. Microbial degradation of aromatic compounds — from one strategy to four. Nature Reviews Microbiology, v. 9, n. 11, p. 803–816, nov. 2011. GHOSAL, D. et al. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Frontiers in Microbiology, v. 7, 31 ago. 2016. GUERRA, A. B. et al. Metagenome enrichment approach used for selection of oil-degrading bacteria consortia for drill cutting residue bioremediation. Environmental Pollution, v. 235, p. 869–880, abr. 2018. HERZHAFT, B., RAGOUILLAUX, A., COUSSOT, P. How to Unify Low-shear-rate Rheology and Gel Properties of Drilling Muds: A Transient Rheological and Structural Model for Complex Well Applications. IADC/SPE Paper 99080.2006 IQBAL, S., KHALID, Z.M., MALIK, K.A.. Enhanced biodegradation and emulsification of crude oil and hyperproduction of biosurfactants by a gamma ray-induced mutant of Pseudomonas aeruginosa. Lett Appl Microbiol. v.21, n.3, Sep, p.176-9.1995 IWAMOTO, T.; NASU, M. Current bioremediation practice and perspective. Journal of Bioscience and Bioengineering, v. 92, n. 1, p. 1–8, 1 jan. 2001. JI, G.D., YANG, Y.S., ZHOU, Q., SUN, T., NI, J.R.. Phytodegradation of extra heavy oil-based drill cuttings using mature reed wetland: an in situ pilot study. Environ. Int. 30, 509–517.2004

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
622
LEONARD, S.A., STEGEMANN, J.A.. Stabilization/solidification of petroleum drill cuttings: leaching studies. J. Hazard. Mater. 174, 484–491. 2010 LUNA, J. et al. Properties of the biosurfactant produced by candida sphaerica cultived in lowcost substrates. Chemical Engineering Transactions, p. 67–72, jun. 2012 MARIANO, A. P.; ANGELIS, D. DE F. DE; BONOTTO, D. M. Monitoring of geochemical indicators and biodegradation evaluation at a site contaminated with diesel oil. Engenharia Sanitaria e Ambiental, v. 12, n. 3, p. 296–304, set. 2007. MINTON, R. Strategic management of waste streams generated by exploration and production drilling operations, in: Business Briefing: Exploration & Production: The Oil & Gas Review. 2004. MORAIS, E. B., TAUK-TORNISIELO, S. M. Biodegradation of oil refinery residues using mixedculture of microorganisms isolated from a landfarming. Braz. arch. biol. technol., Curitiba, v. 52, n. 6, p. 1571-1578.2009 NITSCHKE, M.; PASTORE, G. M. Biossurfactantes: propriedades e aplicações. Química Nova, v. 25, n. 5, p. 772–776, set. 2002. OKPARANMA, R.N., AYOTAMUNO, J.M., ARAKA, P.P.Bioremediation of hydrocarbon contaminated-oil field drill-cuttings with bacterial isolates. Afr. J. Environ. Sci. Technol. 3, 131–140.2009 PACWA-PŁOCINICZAK, M. et al. Environmental Applications of Biosurfactants: Recent Advances. International Journal of Molecular Sciences, v. 12, n. 1, p. 633–654, 18 jan. 2011. RAHMAN, K. S. . et al. Towards efficient crude oil degradation by a mixed bacterial consortium. Bioresource Technology, v. 85, n. 3, p. 257–261, dez. 2002. ROBINSON, J.P., KINGMAN, S.W., SNAPE, C.E., BRADSHAW, S.M., BRADLEY, M.S.A., SHANG, H., BARRANCO, R.. Scale-up and design of a continuous microwave treatment system for the processing of oil-contaminated drill cuttings. Chem. Eng. Res. Des. 88, 146–154.2010 SANTOS, D. et al. Biosurfactants: Multifunctional Biomolecules of the 21st Century. International Journal of Molecular Sciences, v. 17, n. 3, p. 401, 18 mar. 2016a. SETTE, L.D., SIMIONI, K.C., VASCONCELLOS, S.P., DUSSAN, L.J., NETO, E.V., OLIVEIRA, V.M.Analysis of composition of bacterial communities in oil reservoirs from a southern offshore brazilian basin. Antoine van Leewenhoek. v. 91, 253-266.2007 STEPHENSON, R. L. et al. Thermal Desorption of Oil from Oil-Based Drilling Fluids Cuttings: Processes and Technologies. Society of Petroleum Engineers, 2004a. SILVA, EJ; SILVA, NMPR; RUFINO, RD; LUNA, JM; SILVA RO, SARRUBO LA. Characterization of a biosurfact ant produced by

MICRORGANISMOS ENVOLVIDOS NA BIODEGRADAÇÃO DE PETRÓLEO E DERIVADOS
623
Pseudomonas cepacia CCT6659 in the presence of industrial wastes and its application in the biodegradation of hydrophobic compounds in soil. Colloid Surface B 117:36–41.2014 SOBRINHO, H. B. S. et al. Utilization of two agroindustrial by-products for the production of a surfactant by Candida sphaerica UCP0995. Process Biochemistry, v. 43, n. 9, p. 912–917, set. 2008. SPONZA, D. T.; GÖK, O. Effect of rhamnolipid on the aerobic removal of polyaromatic hydrocarbons (PAHs) and COD components from petrochemical wastewater. Bioresource Technology, v. 101, n. 3, p. 914–924, fev. 2010. TAKAI, K., INOUE, A., HORIKOSHI, K. Methanothermococcus okinawensis sp. nov., a thermophilic, methane-producing archae on isolated from a Western Pacific deep-sea hydrothermal vent system. International Journal of Systematic and Evolutionary Microbiology. v. 52, 1089–1095. 2002 VEIL, J.A. Drilling waste management: past, present, and future, in: Proceedings of the SPE Annual Technical Conference and Exhibition, San Antonio, TX, United States. 2002 VEIL, J., ELCOCK, D., RAIVEL, M., CAUDLE, D., AYERS, R.C., GRUNEWALD, B. Disposal of nonhazardous oil field wastes into salt caverns, in: Proceedings of the International Conference on HSE in Oil and Gas Exploration and Production, vol. 2, pp.767–768. 1996 WANG, H., XU, R., LI, F., QIAO, J., ZHANG, B. Efficient degradation of lube oil by a mixed bacterial consortium. Journal of Environmental Sciences. v. 22(3), p.381–388, 2010. WOJTANOWICZ, A.K. Environmental control technology in petroleum drilling and production, in: S.T. Orszulik (Ed.), Environmental Technology in the Oil Industry, Blackie Academic & Professional, London, UK, pp. 73–170. 1997.

624
BIOLOGIA MOLECULAR

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
625
CAPÍTULO 30
AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA:
UMA REVISÃO INTEGRATIVA
Felipe Gonçalves BEZERRA1
Karolyne de Melo SOARES2
Maryanne Pereira Franco ADRIANO3
Maria Soraya Pereira Franco ADRIANO4
1 Mestrando em Biologia Celular e Molecular, UFPB; 2 Graduanda do Curso Técnico em Prótese Dentária, UFPB e Odontologia, UNIESP; 3 Graduanda do Curso de Medicina, UNIPÊ;
4 Orientadora/Professora do PGBCM/UFPB. [email protected]
RESUMO: O flúor é um halogênio que forma compostos inorgânicos e orgânicos chamados fluoretos. A deficiência de flúor pode resulta em retardo do crescimento e desenvolvimento, enquanto a exposição excessiva pode levar a distúrbios no osso que chamamos de fluorose esquelética. Nesse contexto, é importante frisar que são diversos os fatores que podem influenciar o tipo de alteração provocada pela intoxicação com o flúor nos ossos, dentre eles a dose e duração da exposição, herança genética, influência da natureza, idade, sexo e hábitos nutricionais. Essa pesquisa teve como objetivo realizar uma avaliação molecular da fluorose esquelética no estado da Paraíba, através de uma revisão integrativa de literatura. As informações foram obtidas numa seleção de 37 artigos. O instrumento utilizado para coleta de dados incluiu-se a utilização de consulta na base de dados do Google acadêmico, Scielo, Bireme e Biblioteca Virtual em Saúde de forma gratuita. Através dos dados obtidos, observou-se que os efeitos do flúor sobre as células depende da concentração, duração da exposição, e do tipo de célula envolvida. Além

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
626
disso, os estudos mostram que a fluorose esquelética envolve efeitos combinados de vários fatores, como fatores genéticos, ambientais e comportamentais. Palavras-chave: Análise molecular. Fluoreto. Fluorose.
INTRODUÇÃO
Durante décadas, a fluretação tem sido uma medida de
saúde pública adotada em países de todo o mundo, sendo
aceita como um importante fator para a diminuição da
prevalência da cárie dentária (LEAL; CARVALHO; CARVALHO,
2015; MOURA, et al., 2016; CURY, et al., 2019; QIN, et al.,
2017).
Nos Estados Unidos, país precursor na utilização do
fluoreto para prevenir cárie dentária, sua tecnologia se
desenvolveu e se firmou como sendo apropriada e segura no
uso em Saúde Pública, a medida foi considerada pelas
autoridades sanitárias e investigadores como uma das dez
maiores conquistas do país, nesse campo, no século
XX. Alguns centros de pesquisa que acompanhavam na época,
por meio de criteriosas revisões sistemáticas, aprovam a sua
evidência científica e segurança para a saúde humana
(FERREIRA; NARVAI, 2015).
No Brasil, a fluoretação só foi utilizada pela primeira vez
em 1953. Duas décadas depois, no auge do período histórico,
onde não existia democracia no País, foi aprovada a lei (Lei nº
6.050/1974) tornando o uso do fluoreto como uma medida
obrigatória nos locais onde exista Estação de Tratamento de
Água (GARBIN, et al., 2014; FERREIRA; NARVAI, 2015).
As primeiras investigações relacionando à utilização do
flúor estavam direcionadas na água, por ser um recurso de fácil

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
627
acesso à população, porém, com o avanço da tecnologia e a
melhoria das condições de higiene bucal, essa prática tem sido
diminuída em vários países (DEATH, et al., 2015; LI, et al.,
2017). Em contra partida, os EUA e o Brasil mantiveram a
prática de adicionar flúor na água destinada ao abastecimento
público, que é aconselhada devido a sua segurança, facilidade
de administração e baixo custo populacional, sendo esse um
dos recursos mais seguros para manter a regularização
continua da presença de flúor na cavidade bucal (GARBIN, et
al., 2014; SCALIZE, et al., 2018).
Segundo Scalize (2018) o uso da fluoretação na água,
considera-se justificado pelo índice de redução de cárie
dentária citado em mais de 160 trabalhos distribuídos em mais
de 10 países. A utilização de água fluoretada na concentração
0,70 ppm de F é um método seguro, pois é capaz de produzir o
máximo de benefícios com o mínimo efeito colateral (PEI, et al.,
2017; LEAL; CARVALHO; CARVALHO, 2015; LIMA, et al.,
2019). Entretanto, a Organização Mundial da Saúde (OMS)
alerta para o risco de concentrações superiores a 1,5 mg F/L
que podem acarretar a saúde humana, como o aparecimento
de fluorose (BACHANEK, et al., 2019; PRAMANIK; SAHA,
2017).
A água que é destinada ao consumo humano deve
atender a certos requisitos de qualidade, para não colocar em
risco a saúde da população que a utiliza. Mesmo sendo límpida,
ela contém misturado a sua composição impurezas que podem
ser de natureza química, física ou biológica. No Brasil, o padrão
de qualidade da água para consumo humano é determinado
pela Portaria 518 do Ministério da Saúde, que com base nos
valores encontrados de impurezas aconselha ou não a sua

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
628
utilização para o consumo humano (CURY, et al., 2015;
PRIETTO, et al., 2015; SCHARDONG, et al., 2018; SILVA, et
al., 2018).
Estudos mais recentes mostraram que, em países
desenvolvidos, a contribuição do flúor na água potável poderia
ser 2 a 3,5 vezes mais elevado do que o recomendado
(ZABALA, et al., 2018; QIN, et al., 2017). A Agência de Proteção
Ambiental (EPA) recomendou reduzir o valor alvo de flúor para
0,7 mg/l em água, como objetivo de minimizar os efeitos
indesejável causado pelo produto. Por esse motivo, entre
outros, fluoretação da água potável representa uma das
intervenções de saúde pública com mais controvérsias. Além
disso, alguns municípios não dispõem de meios para adição do
flúor ou de métodos de vigilância para o controle de sua
concentração nos sistemas de abastecimento público
(SCALIZE, et al., 2018).
No Brasil, existem relatos de fluorose endêmica devido à
presença de altos níveis de fluoreto nas águas, nos Estados de
São Paulo (Marília e Bauru), Paraná (Ponta Grossa), Minas
Gerais (Belo Horizonte), Santa Catarina, Paraíba (Catolé do
Rocha e São João do Rio do Peixe) e Rio Grande do Sul (Santa
Tereza) (RIGO; LOD; GARBIN, 2015; ADRIANO, et al., 2017;
SCHARDONG, et al., 2018).
Outras fontes de exposição ao flúor são pelos
suplementos alimentares, enxaguatórios bucais e cremes
dentais, chamados de dentifrícios fluoretados (PRIETTO, et al.,
2015; MOURA, et al., 2016; ZABALA, et al., 2018). Estudos
mostram que desde 1989 no Brasil, o flúor também tem sido
adicionado em mais de 90% dos dentifrícios (LEAL, S. D;
CARVALHO, F. C; CARVALHO, 2015). Com isso, a ingestão

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
629
acidental de dentifrícios, também podem contribuir para o
aumento do risco de fluorose (SCALIZE, et al., 2018).
Entretanto, em populações expostas ao flúor, através do
consumo de dentifrícios, a ocorrência de fluorose está
restringida aos graus leves e muito leves (OLIVEIRA, et al.,
2016; SILVA, et al., 2018; LIMA, et al., 2019).
Como já mencionado, o flúor possui características
benéficas como tóxicas, isso vai variar de acordo com dose. A
terapia com flúor é complexa, pois pode apresentar uma
resposta bifásica dependente da dose, isto é, em doses altas
estimulando a formação óssea, além de preventiva no combate
a cárie e tóxico em dose alta (maior que 1,5 mg /L) levando a
deformidades esqueléticas e dentárias chamadas fluorose
(MOURA, et al., 2017; AMARAL, et al., 2018; LIU, et al., 2018).
De fato, quando se ingere fluoreto é esperado o
desenvolvimento de algum grau de fluorose. O grau de
intolerância da fluorose está diretamente relacionado à dose de
ingestão e ao tempo de uso, podendo variar de graus leves até
graus mais severos (JORDÃO, et al., 2015). Isto implica dizer
que todo flúor ingerido poderá causar algum grau de fluorose,
entretanto, o conceito clínico dependerá da dose e do seu
tempo de duração (LIMA, et al., 2019).
Os efeitos adversos mais estudados pelo flúor são
fluorose dentária e esquelética. A diferença entre a fluorose
dentária e a esquelética e mostrada através do processo pelo
qual o esmalte e o esqueleto sofrem mineralização totalmente
distintos (DEATH, et al., 2015; LV; KANG; WU, 2016; AMARAL,
et al., 2018). Tanto o esmalte dentário como o esqueleto
humano, em sua formação são mediados por células,
ameloblasto (esmalte) e osteoblasto (esqueleto),

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
630
respectivamente. Ambos iniciam o processo de esmalte e osso
através da formação de uma sintese e secreção de uma matriz
de proteína que é depositado no espaço extracelular (WEI, et
al., 2018). A adição do composto protéico na matriz de ambos
os composto também apresentam-se totalmente diferente.
Enquanto o esmalte é mineralizado e é precendida pela
reabsorção da matriz de proteínas, a matriz de colágeno do
osso é mineralizada sem reabsorção. No esmalte, o flúor
circulante no corpo e presente nos fluidos da matriz extracelular
inibe a reabsorção de proteínas, produzindo um mineral
hipomineralizado de tecido, e no osso, o tecido fica
hipermineralizado devido a diferentes mecanismos. Portanto,
enquanto o esmalte em desenvolvimento é sensível em baixas
concentrações de fluoreto em exposição à água (menor que 1,0
ppm F) em curto periodo de tempo, as alterações ósseas são
causadas apenas por 5 a 10 vezes maior concentração de
exposição prolongada de flúor. Contudo, sabe-se que a
concentração 1,0 ppm F flúor na água não acomete a
resistência e densidade óssea (RIGO; LOD; GARBIN, 2015;
JORDÃO, et al., 2015).
Os estudos sobre fluorose dentária são normalmente
mais frequentes do que a fluorose esquelética (PEI, et al., 2017;
LI, et al., 2017). O motivo para a existência de poucos estudos
com a fluorose esquelética, se explica pelo fato dessa tipo
somente ocorrer em situações onde a concentração de flúor na
água de consumo atinge níveis acima de 3,5 ppm (MOURA, et
al., 2017; SCARPETTA, et al., 2014).
Embora a fluorose esquelética seja típica em alguns
países, o Brasil tem sido considerado um país poucos relatos
da doença. Em contra partida, no semiárido nordestino, as altas

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
631
temperaturas e escassez dos recursos hídricos, a população
rural recorre às águas subterrâneas para satisfazer suas
necessidades básicas, onde muitas das vezes, as águas
subterrâneas podem apresentar concentrações de flúor acima
dos níveis recomendados para consumo humano,
representando um risco para o desenvolvimento da doença
(AMARAL, et al., 2018; ADRIANO, et al., 2017; LIU, et al.,
2018).
Nesse sentido, o estado da Paraíba destaca-se na
epidemiologia da fluorose, por apresentar áreas endêmica de
fluorose devido aos elevados níveis de flúor nas águas de
consumo para população rural. No momento atual, estima-se
que cerca de 30 mil pessoas estão sendo sujeitas a níveis
elevados de flúor no interior da Paraíba, sendo 20% destes,
com risco de fluorose esquelética (SOUZA, 2012).
Mediante a proposta escolhida, essa pesquisa teve como
objetivo de realizar uma avaliação molecular da fluorose
esquelética no estado da Paraíba, através de uma revisão
integrativa de literatura.
MATERIAIS E MÉTODO
A metodologia de uma pesquisa é um dos primeiros passos a
ser decidido, através da escolha de ferramentas e estratégias
adequadas para que seja estabelecida a sistemática de
trabalho, respondendo os quesitos iniciais, como mostrar
(ANTUNES, et al., 2018). Desta forma, o trabalho proposto
trata-se de uma pesquisa de revisão integrativa de literatura. As
informações foram obtidas numa seleção de 46 artigos,
referindo-se ao respectivo tema, mas apenas 37 dos 46 artigos

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
632
que foram selecionados, estavam dentro dos critérios de
inclusão para serem utilizados na construção do estudo. O
instrumento utilizado para coleta de dados incluiu-se a
utilização de consulta na base de dados do Google acadêmico,
Scielo, Bireme e Biblioteca Virtual em Saúde (BVS) de forma
gratuita por meio das palavras-chave: análise molecular,
fluoreto e fluorose. O Critério de elegibilidade incluiu artigos
registrados entre 2012 a 2020 no idioma Português, Inglês e
Espanhol. Dentre os critérios observados foram considerados
os seguintes aspectos: disponibilidade do texto integral do
estudo e clareza no detalhamento metodológico utilizado.
RESULTADOS E DISCUSSÃO
Após leitura criteriosa dos artigos nas bases de dados
nos referidos bancos de dados, realizou-se o fichamento
catalográfico do material bibliográfico com o intuito de dinamizar
a leitura e compreensão dessa revisão, optouse por dividí-la em
três tópicos principais: Flúor; Fluorose óssea e Considerações
moleculares acerca da fluorose óssea.
FLUOR
O flúor é um gás amarelo claro, considerado o 13°
elemento mais abundante na natureza e está presente no ar, no
solo, nas rochas e nas águas. Apresenta-se sob a forma de
moléculas covalentes apolares F2, nas quais cada átomo de
flúor atinge uma configuração eletrônica de oito elétrons na
camada mais externa este elemento químico descoberto no ano
de 1886 por Henri Mossan, está incluído como componente

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
633
natural da biosfera (BACHANEK, et al., 2019; AMARAL, et al.,
2018). É o elemento mais eletronegativo dos halogênios, reativo
à temperatura ambiente e possui a capacidade de reagir
consideravelmente com a maioria dos compostos que contêm
hidrogênio, como água e todas as substâncias orgânicas, sejam
líquidos, sólidos ou gases, formado compostos orgânicos e
inorgânicos, sendo frequentemente encontrado na forma de
fluoreto (QIN, et al., 2017; PRAMANIK; SAHA, 2017; DEATH,
et al., 2015).
Nos seres humanos, o flúor é instantaneamente
absorvido pelo estômago e intestino delgado, seguido de sua
deposição em tecidos calcificados (aproximadamente 50% é
incorporado nos ossos na forma de hidroxifluoroapatita) e por
fim, excretado pela urinária. É o principal agente anticárie e tem
sido indicado como o maior responsável pela redução dos
índices de cárie ao redor do mundo (OLIVEIRA, et al., 2016;
LIMA, et al., 2019; LV; KANG; WU, 2016; SILVA, et al., 2018).
É um elemento importante para a manutenção da integralidade
de ossos e dentes, sendo que, aproximadamente 99% do flúor
presente no corpo está nos tecidos mineralizados. O flúor não
é um componente essencial para o crescimento e
desenvolvimento humano, mas devido aos seus benefícios
recebe uma atenção semelhante, enquanto a sua ingestão
recomendada for estabelecida (GARBIN, et al., 2014; WANG;
XU, 2017; VÁSQUEZ; AZAÑEDO, 2019).
No entanto, quando as concentrações de F ultrapassam
os valores recomendáveis, o flúor pode ocasionar reações
indesejáveis. Tais efeitos se referem aos níveis de exposição a
essa substância, e não a substância propriamente dita. Seus
efeitos preventivos ou mesmo terapêutico, tornam-se ineficazes

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
634
causando reações indesejáveis. Nesse sentido, os efeitos
colaterais do flúor, ocorrem pela administração de doses
pequenas e constantes, atingindo principalmente os tecidos
mineralizados (PEI, et al., 2017; PRIYANKA; VANDANA, 2019;
QIN, et al., 2017).
Portanto, o flúor é uma substância química que é capaz
de prevenir uma doença e provocar outra, isso a depender da
concentração na qual é utilizado. Com isso, o flúor é o único
elemento químico capaz de promover a prevenção da cárie
dentária e, simultaneamente, provocar o aparecimento da
fluorose (MOURA, et al., 2017; WEI, et al., 2018; LV; KANG;
WU, 2016).
Uma vez que o flúor intervém nas principais vias
metabólicas do sistema biológico, ele pode ocasionar
alterações bioquímicas e/ou moleculares e algumas fatóres
influenciando nos efeitos tóxicos na via metabólica, como idade,
deficiência de minerais (cálcio, magnésio) e/ou vitamina C,
distúrbios ácido-básico, presença de problemas
cardiovasculares ou renais, alterações genéticas, temperatura
ambiente, entre outros, fazendo com que algumas pessoas
sejam mais vulneráveis à utilização desse elemento e seus
compostos, uma vez que são condições que podem agravar ou
reduzir os efeitos no organismo humano (AMARAL, et al.,
2018).
FLUOROSE ESQUELÉTICA
A fluorose esquelética é definida como sendo uma
doença óssea metabólica crônica causada pela inalação ou
ingestão duradoura de quantidades elevadas de fluoreto

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
635
(ingestão de mais de 10mg por dia, por pelo menos 10 anos),
em que as células ósseas (osteoblastos e osteoclastos) são
sensibilizadas pelos níveis elevados de flúor, desencadeando o
desenvolvimento de lesões ósseas incluindo osteosclerose,
osteoporose, osteomalácia e alterações articulares
degenerativas (WU, J. et al., 2015; YANG et al., 2017). Outros
efeitos como osteossarcoma, hipotireoidismo e
neurotoxicidade, também foram descritos (ZABALA, et al.,
2018; PEI, et al., 2017).
A sua gravidade está relacionada com qualidade de vida
e condições de trabalho do indivíduo, sendo responsável por
diversos tipos de problemas, incluindo a posturais e a limitações
de movimentos (LIU, et al., 2018). A doença provoca nos seres
humanos desconforto, dor, rigidez do pescoço, costas, joelho,
ombro, ossos e articulações, levando a várias deformidades
ósseas, aumenta o risco de fraturas e outros problemas
associados, afetando jovens e idosos (TEIXEIRA, et al., 2015;
PURANIK, et al., 2015). É considerada um problema de saúde
mais agravante do que a fluorose dentária, porque tem
capacidade de causar incapacidade permanente nos indivíduos
(WU, J. et al., 2015).
A severidade da fluorose esquelética e definida em três
graus: grau I (leve), os exames de imagens indicam sinais de
fluorose esquelética sem apresentar sintomatologia clínica,
como limitação de movimentos e deformação de juntas. O grau
II (moderada), os exames de imagens indicam alterações como
enfraquecimento e desmineralização de ossos maduros
(osteomalacia), apresentando sintomatologia (dor nas
articulações e malformações ósseas), sendo que as limitações
dos movimentos não impede o trabalho e Grau III (severo), os

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
636
exames de imagans indicam diminuições severas na matriz
óssea (osteomalacia e osteopenia) e sinais de curva de
lateralidade na postura, dor renal crônica e perda de habilidade
para o trabalho (QIN, et al., 2017; LI, et al., 2017; PRAMANIK;
SAHA, 2017).
A sintomatologia das pessoas com fluorose esquelética
é incerto, dependendo assim, do grau de alteração. Em alguns
casos pode ser assintomático, onde as mudanças são visíveis
apenas em exames de imagens. No início do doença, os
pacientes queixam-se repetidamente de um desconforto nos
membros e no tronco. A maioria dos casos mais graves,
desenvolve dores ósseas constante e inflexbilidade de todo o
corpo. Cifose constante e limitação de movimentos tornam-se
aparentes, piorando gradualmente para o estado mais grave
(MOURA, et al., 2017; LV; KANG; WU, 2016; DEATH, et al.,
2015).
O comprometimento neurológico é observado no estágio
mais grave da doença, devido à compressão das raízes da
medula espinhal e nervos, separadamente ou por combinação,
e frequentemente se desenvolve depois da exposição elevada
ao fluoreto durante mais de 10 anos (WEI, et al., 2018).
CONSIDERAÇÕES MOLECULARES ACERCA DA
FLUOROSE ÓSSEA
Até o presente momento, poucos estudos tem explorado
os mecanismos moleculares responsáveis pela patogenicidade
da fluorose esquelética, embora seja aceitável que o flúor tenha
alta afinidade com tecidos mineralizados (AMARAL, et al., 2018;
PRAMANIK; SAHA, 2017).

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
637
Estudos moleculares utilizando variações de espécies de
camundongos têm apresentado resposta significativamente
aceitável do efeito do flúor no desenvolvimento e na
homeostase do tecido ósseo (PURANIK, et al., 2015; QIN, et
al., 2017).
Em um estudo realizado por Scarpetta et al (2014) e Wei
et al (2018) com duas espécies de camundongos (A/J e
129P3/J) apresentaram diferenças intrínsecos em osso cortical
e trabecular que não são afetados pelo tratamento com F. As
diferenças estão relacionadas à densidade mineral óssea
(DMO), relação a superfície óssea e o volume do tecido, relação
volume de osso para volume de tecido, número trabecular,
espessura e separação. O estudo revelou que os camundongos
A/J apresentaram menos, mais finos, menos conectados e mais
trabéculas amplamente espaçadas em relação aos
camundongos 129P3/J. Além disso, a ALP (enzima envolvida
no processo de mineralização), foi aumentada no tipo 129P3/J.
Os mecanismos são baseados em proteínas desencadeados
por F em osteoblastos e osteoclastos. Em 129P3/J, as
proteínas são identificadas com maior abundância em relação
a A/J.
A sensibilidade do osso é influenciado pela genética. Isso
explicar por que os tratamentos com F para reduzir a massa
óssea não é universalmente eficaz (AMARAL, et al., 2018;
PRIYANKA; VANDANA, 2019; WEI, et al., 2018; MOURA, et
al., 2017).
O flúor interage com uma imensa quantidade de
processos celulares, como os genes que estão relacionados
com a formação e desenvolvimento do osso (BGLAP, ESR,
COL1A2), genes que codificam receptores envolvidos na

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
638
formação óssea e metabolismo (VDR, CTR), genes que
codificam hormônios (PTH, PRL), genes que codificam enzimas
desintoxicantes (GSTP1), genes que codificam proteínas da
matriz extracelular (MMP), genes com funções cognitivas
funciona (COMT) e genes que têm papéis em funções imunes
(MPO). Alguns outros estudos revelaram associação
significante entre polimorfismos em (GSTP1, MMP-2,
PRL,VDR), genes (MPO) (PRAMANIK; SAHA, 2017; LV;
KANG; WU, 2016).
Os polimorfismos podem estar possivelmente ligado a
fluorose esquelética. Estudos mostram que a fluorose
esquelética envolve efeitos combinados de vários fatores, como
fatores genéticos, ambientais e comportamentais (PEI, et al.,
2017; LI, et al., 2017).
Os estudos realizados por Yang et al (2016) e Wu et al
(2015) mostraram que o polimorfismo do gene da
mieloperoxidase está relacionado com a fluorose do tipo carvão
queima, o que propõe que o gene o polimorfismo pode
desempenhar um papel importante na patogênese da fluorose
esquelética. No mesmo estudo, foi observado que entre os
genes potenciais relevantes para o metabolismo ósseo, o gene
do receptor de vitamina D (VDR) é considerado ser um
candidato importante. Além disso, o gene VDR, localizado no
cromossomo 12 (12q12-14), desempenha um papel importante
no metabolismo ósseo, incluindo a absorção intestinal de cálcio.
O gene superóxido dismutase 2 (SOD2) codifica uma
enzima eliminadora de radicais livres, que retira o superoxidase
e catalisa a produção de peróxido de hidrogênio (LI, et al.,
2017). Em um estudo realizado por Liu et al (2018), os
indicativos mostraram que fluorose por queima de carvão foi

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
639
relacionada com a redução da atividade em enzimas
superóxido dismutase e gene expressão.
O flúor apresenta diversas propriedades relevantes no
seu mecanismo molecular de ação: in vitro, apresenta ação
mitogênica que é específica das células ósseas e resulta em
mudanças na expressão gênica, estresse e morte celular, in
vivo, apresenta ação anabólico que é específico de tecidos
mineralizados (PRAMANIK; SAHA, 2017).
Segundo Liu et al (2018) o consumo excessivo de
fluoreto causa estresse oxidativo, que é um estado de
desequilíbrio entre o pró-oxidantes e antioxidantes que levam à
lesão química dos lipídios, proteínas e DNA. A formação
demasiada de espécies reativas de oxigênio resulta em
oxidação de macromoléculas, quebra de fosfolipídios da
membrana, peroxidação lipídica, despolarização da membrana
mitocondrial e apoptose.
Alguns pesquisadores, ao estudar os mecanismos
moleculares da toxicidade causada pelo fluoreto, demostraram
que os genes associado à transdução de sinal (proteína G, JNK,
ERK, MEK1, MEK2), imunidade (IL6, IL8), apoptose (c-fos, c-
jun, bax, p53, bcl2), câncer e fatores de transcrição (CREB, NF-
kappaβ) poderiam estar fortemente ligados a ocorrência e
desenvolvimento de fluorose. Além disso, a intoxicação pode
afetar eventos intracelulares como a expressão gênica, síntese
proteica, secreção, tráfico, reabsorção e degradação de
produtos e também eventos extracelulares como a interação
entre matriz proteica, cristais e componentes fluidos (PEI, et al.,
2017; PRAMANIK; SAHA, 2017).
Os estudos apontam que o flúor interferir na síntese de
DNA, RNA e proteínas, causando danos ao DNA. Ao realizar

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
640
análise in vitro com células HaLa, os achados mostraram que o
NaF afeta a síntese de proteínas, impedindo o início de cadeia
peptídica. A exposição das células HeLa com NaF resultou na
inibição da síntese protéica, desagregação dos
polirribossomos, acúmulo dos rinossomos 80 S e diminuição
das subunidades ribossômicas livres. Logo após ao retirar NaF,
os níveis normais das subunidades ribossômicas livres foram
restaurados (QIN, et al., 2017; AMARAL, et al., 2018; LV;
KANG; WU, 2016).
Glutationa S-transferases (GST) são enzimas
metabolizadoras da fase II que desempenham funções
importantes na proteção celular, pela liberação de substâncias
nocivas, incluindo radicais redox. a atividade da classe pi GST
(GSTP1), codificada pelo gene GSTP1, está diretamente
associado com as características clínica da fluorose. Foi
observado que a expressão e atividade de GSTP1 esta
relacionado com polimorfismos de nucleotídeo único, incluindo
GSTP1 rs1695 (p.105 Ile> Val), resultando em atividade
catalítica reduzida e capacidade de desintoxicação da enzima
(WU, J. et al., 2015; YANG; WANG; XU, 2017).
CONCLUSÕES
Considerando que a fluorose esquelética seja definida como uma doença óssea metabólica crônica causada pela inalação ou ingestão prolongada de quantidades elevadas de fluoreto, a sua sintomatologia pode simular outras patologias. Nesse sentido, muitos indivíduos encontram-se em risco de desenvolver a doença, e em outros casos, a doença ainda não fora diagnosticada corretamente.

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
641
O efeito deletério do flúor sobre as células depende da concentração, duração da exposição, e do tipo de célula envolvida. Como visto, o fluoreto inibe a secreção e/ou síntese de proteínas e influencia distintas vias de sinalização envolvidas no processo celular, a utilização da análise proteômica poderá mostrar como os processos metabólicos, regulatórios e de sinalização ocorrrem nessa morbidade, permitindo uma maior compreensão dessa patologia.
Embora a concentração de flúor seja um critério importante que pode influenciar a sua toxicidade, pressupõe que os níveis das doses não é o único fator. Fatores genéticos, sexo, idade, e origem da população podem contribuir com a suscetibilidade ou resistência a exposição ao flúor Assim, esses achados sugerem que os determinantes podem influenciar a suscetibilidade ou resistência aos efeitos do fluoreto.
Dessa forma, a compreensão da doença somado a utilização dessa metodologia, viabilizará a detecção precoce, constituindo um ponto chave no tratamento em pacientes com fluorose esquelética. Este trabalho caracteriza-se como um passo inicial nos estudo molecular com a possibilidade de introduzir uma análise sistemática em indivíduos portadores de fluorose esquelética. Espera-se que com a identificação de proteínas envolvidas nos processos metabólicos do flúor, possamos esclarecer melhor patogenicidade desse doença, bem como identificar indivíduos mais vulneráveis à fluorose esquelética.
REFERÊNCIAS BIBLIOGRÁFICAS ADRIANO, M. S. P. F. et al. Fluorose óssea: conhecimento de uma população acerca da doença. Revista Família, v. 5, n. 1, p. 125-130, 2017. AMARAL, S. L. et al. Effect of chronic exercise on fluoride metabolism in fluorosis-susceptible mice exposed to high fluoride. SCientifiC Reports, v. 8, n. 32, p. 1-7, 2018.

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
642
ANTUNES J. et al. Diagnóstico rápido participativo como método de pesquisa em educação. Avaliação (Campinas), v. 23, n. 3, p. 590-610, 2018. BACHANEK, T. et al. Condition of mineralized tooth tissue in a population of 15-year-old adolescents living in a region of Ukraine with slightly exceeded fluorine concentration in the water. Annals of Agricultural and Environmental Medicine, v. 26, n. 4, p. 623-629, 2019. CURY, J. A. et al. Concentração de fluoreto nos dentifrícios a base de MFP/CaCO3 mais vendidos no Brasil, ao final dos seus prazos de validade. Rev. Assoc. Paul. Cir. Dent, v. 69, n. 3, p. 248-251, 2015. CURY, J. A. et al. Systemic Effects (Risks) of Water Fluoridation. Braz. Dent J, v. 30, n. 5, p. 421-428, 2019. DEATH, C. et al. Dental fluorosis and skeletal fluoride content as biomarkers of excess fluoride exposure in marsupials. Science of The Total Environment, v. 533, n. 2, p. 528-541, 2015. FERREIRA, R. G. L. A; NARVAI, P. C. Fluoretação da água: significados e lei da obrigatoriedade na visão de lideranças em saúde. Rev. Assoc. Paul. Cir. Dent, v. 69, n. 3, p. 266-271, 2015. GARBIN, C. A. S. et al. Concentração de flúor em águas envasadas: análise laboratorial e da legislação relacionada. RFO UPF, v. 19, n. 3, p. 323-328, 2014. JORDÃO, L. M. R. et al. Fluorose dentária: prevalência e fatores associados em escolares de 12 anos de Goiânia, Goiás. Revista Brasileira de Epidemiologia, v. 18, n. 3, p. 568 – 577, 2015. LEAL, S. D; CARVALHO, F. C; CARVALHO, C. A. P. Conhecimento de alunos do Curso de Odontologia sobre o uso racional do flúor. Rev. odontol. UNESP, v. 44, n. 1, p. 51-58, 2015. LIMA, I. F. P. et al. Prevalência de fluorose dental em regiões abastecidas com água sem suplementação de flúor no território brasileiro: uma revisão sistemática e metanálise. Ciênc. Saúde Coletiva, v. 24, n. 8, p. 2909-2922, 2019. LI, B. Y. et al. ClC-7/Ostm1 contribute to the ability of tea polyphenols to maintain bone homeostasis in C57BL/6 mice, protecting against fluorosis. International Journal of Molecular Medicine, v. 39, n. 1, p. 1155-1163, 2017. LIU, Jun. et al. Association of Dietary Carotenoids Intake with Skeletal Fluorosis in the Coal-burning Fluorosis Area of Guizhou Province. Biomed Environ Sci, v. 31, n. 6, p. 438-447, 2018. LV, Y. G; KANG, L; WU, G. Fluorosis increases the risk of postmenopausal osteoporosis by stimulating interferon γ. Biochemical and Biophysical Research Communications, v. 479, n. 2, p. 372-379, 2016.

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
643
MOHAMMADI, A. A. et al. Skeletal fluorosis in relation to drinking water in rural areas of West Azerbaijan, Iran. Scientific Reports, v. 7, n. 17, p. 1-7, 2017. MOURA, M. S. et al. Vigilância epidemiológica da fluorose dentária em município de clima tropical com água de abastecimento público fluoretada. Ciência & Saúde Coletiva, v. 21, n. 4, p. 1247-1254, 2016. OLIVEIRA, L. F. B. et al. A presença de fluorose dentária estaria associada ao traumatismo dentário entre escolares? Ciênc. saúde coletiva, v. 21, n. 3, p. 967-976, 2016. PRIETTO, N. R. et al. Atitudes e conhecimento dos pais quanto ao uso de dentifrícios fluoretados em crianças de um a 65 meses de idade. RFO UPF, v. 20, n. 2, p. 216-221, 2015. PEI, J. et al. Matrix Metallopeptidase-2 Gene rs2287074 Polymorphism is Associated with Brick Tea Skeletal Fluorosis in Tibetans and Kazaks, China. Scientific Reports, v. 7, n. 4, p. 1-8, 2017. PRAMANIK, S; SAHA, D. The genetic influence in fluorosis. Environmental Toxicology and Pharmacology, v. 56, n. 3, p. 157-162, 2017. PRIYANKA, D; VANDANA, K. L. Oestrogen receptor Rsa I gene polymorphism in osteoporosis periodontitis patients with or without dental fluorosis. Indian J Med Res, v. 3, n. 2, p. 364-368, 2019. PURANIK, C. P. et al. Fluoride Modulates Parathyroid Hormone Secretion in vivo and in vitro. Cells Tissues Organs, v. 200, n. 6, p. 413-423, 2015. QIN, J. H. et al. Excessive fluoride increases the expression of osteocalcin in the mouse testis. Zhonghua Nan Ke Xue, v. 23, n. 9, p. 782-785, 2017. RIGO, L; LODI, L; GARBIN, R. R. Diagnóstico diferencial de fluorose dentária por discentes de odontologia. Einstein, v. 13, n. 4, p. 547-554, 2015. SCALIZE, P. S. et al. Heterocontrole da fluoretação da água de abastecimento público em cidades do estado de Goiás, Brasil. Ciência e Saúde Coletiva, v. 23, n. 11, p. 3849-3860, 2018. SCARPETTA, R. A. G. et al. Prevalencia de fluorosis dental en infantes de 8 a 12 años de colegios públicos, villavicencio 2013. Hacia promoc. Salud, v. 19, n. 1, p. 25-38, 2014. SCHARDONG, B. A. et al. Eficácia de um dentifrício sem flúor no controle de Streptococcus mutans in vitro. RFO UPF, Passo Fundo, v. 23, n. 3, p. 268-273, 2018. SILVA, A. E. R. et al. Concentração de íons flúor em águas minerais envasadas no Rio Grande do Sul. RFO UPF, Passo Fundo, v. 23, n. 2, p. 156-160, 2018.

AVALIAÇÃO MOLECULAR DA FLUOROSE ESQUELÉTICA NO ESTADO DA PARAÍBA: UMA REVISÃO INTEGRATIVA
644
SOUZA, C. F. M. et al. Assement of groundwater quality in a region of endemic fluorosis in the northeast of Brazil. Environmental Monitoring and Assessment, v. 184, n. 11, p. 1-11, 2012. TEIXEIRA, A. O. et al. Blood and bone parameters of cattle fed diets containing different phosphates and phosphorus: fluorine relations. Rev. bras. saúde prod. Anim, v. 16, n. 1, p. 93-105, 2015. VÁSQUEZ, H. A; AZAÑEDO D. Cepillado dental y niveles de flúor en pastas dentales usadas por niños peruanos menores de 12 años. Rev Peru Med Exp Salud Publica, v. 36, n. 4, p. 646-52, 2019. WEI,Y. et al. Comparative proteomic analysis of fluoride treated rat bone provides new insights into the molecular mechanisms of fluoride toxicity. Toxicology Letters, v. 291, n.1 , p. 39-50, 2018. WU, J. et al. Modifying Role of GSTP1 Polymorphism on the Association between Tea Fluoride Exposure and the Brick-Tea Type Fluorosis. PLoS ONE, v. 10, n. 6, p. 1-14, 2015. YANG, C; WANG, Y; XU, H. Treatment and Prevention of Skeletal Fluorosis. School of Pharmaceutical Sciences, v. 4, n. 2, p. 147-149, 2017. YANG, D. et al. Association between vitamin D receptor gene FokI polymorphism and skeletal fluorosis of the brick-tea type fluorosis: a cross sectional, case control study. BMJ Open, v. 6, n. 11, p. 1-6, 2016. ZABALA, A. J. et al. Ingesta de flúor a través del consumo de agua de abastecimiento público en la cohorte INMA-Gipuzkoa. Gaceta Sanitaria, v. 32, n. 5, p. 418-424, 2018.

645
BIOTECNOLOGIA

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
646
CAPÍTULO 31
APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
Héberly Fernandes BRAGA 1 1 Doutorando em Microbiologia, UNESP;
RESUMO: A biodiversidade e potenciais aplicações dos fungos do Cerrado ainda são pouco conhecidas e positivamente exploradas. Assim, a presente revisão objetivou elucidar e apontar a importância de se conservar, conhecer e se utilizar do potencial biotecnológico oferecido pela micobiota desse bioma, de modo a dar um caráter ainda mais valorativo à região, e mostrar os importantes recursos que podem e estão sendo perdidos por meio seu do manejo inadequado. Por meio de pesquisa exploratória nas bases de dados Google Acadêmico, Scielo, Portal Capes e DOAJ, foram selecionados diferentes trabalhos em inglês e português publicados entre 2015 e 2019, empregando-se os seguintes descritores de busca: fungos do cerrado, (bio)prospecção de fungos do cerrado, propriedades de fungos do cerrado, manejo de fungos do cerrado. As diferentes pesquisas encontradas demonstram uma variedade de possibilidades de uso dos fungos do bioma seja no setor agropecuário, industrial, medicinal, alimentício, químico entre outros e apotam para a necessidade de ações e investimentos que visem não somente à conservação da região, e por consequência da sua micobiota, que vêm ao longo de décadas sendo alterada, devido principalmente ao manejo inadequado do solo; como também do estímulo à novas pesquisas sistematizadas que possam gerar mais conhecimento e descobertas.

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
647
Palavras-chave: Biodiversidade fúngica. Bioprospecção. Biotecnologia. Savana brasileira.
INTRODUÇÃO
O Cerrado é um bioma que apresenta uma enorme
riqueza e abundância de vegetais, animais e micro-organismos.
A alta biodiversidade associada às ameças de destruição dessa
região tem colocado a mesma no grupo dos habitats naturais
considerados como hotspots ecológicos (SAWYER et al.,
2016).
Mesmo apresentando e ofertando distintos serviços
ecossitêmicos, como por exemplo: abastecimento hídrico;
alimentos e matérias-primas de valor econômico; recursos
genéticos; sequestro e armazenamento de gás carbônico (CO2)
atmosférico; turismo e lazer; substâncias farmacológicas e
bioprodutos derivados de plantas e outros organismos,
observa-se que sua área vem sofrendo constantes alterações,
consequentes principalmente da urbanização, agricultura e
pecuária desenfreadas.
Os desequilíbrios e desordens vêm historicamente
propiciando a redução e/ou mesmo a perda das riquezas
genéticas e de biodiversidade, especialmente aquelas com
potencial farmacológico e tecnológico destinados ao tratamento
de doenças e/ou aplicações biotecnológicas, como por exemplo
micro-organismos e seus biocompostos derivados (RESENDE
et al. 2019).
Nesse contexto a presente revisão têm por objetivo
elucidar e apontar a importância de se conservar, conhecer e
se utilizar das distintas pontencialidades biotecnológicas dos
fungos existentes no Cerrado, de modo a dar a essa região um

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
648
caráter ainda mais valorativo, e mostrar os importantes recursos
que podem e estão sendo perdidos com o manejo inadequado
desse bioma.
MATERIAIS E MÉTODO
O material bibliográfico empregado na pesquisa foi
selecionado nas bases de dados Google Acadêmico, Scielo,
Portal Capes e DOAJ, e em sítios eletrônicos de órgãos
governamentais, por meio de pesquisa qualitativa exploratória
realizada em trabalhos publicados entre 2015 a 2019.
Como critérios de inclusão foram considerados artigos
em inglês e português que indicassem possíveis aplicações
biotecnológicas de fungos do Cerrado e/ou pesquisas que
elucidassem as perdas da micobiota nesse determinado bioma,
por manejos inadequados.
Os seguintes descritores foram empregados na busca:
fungos do cerrado, (bio)prospecção de fungos do cerrado,
propriedades de fungos do cerrado, manejo de fungos do
cerrado.
Trabalhos não enquadrados nos critérios de inclusão,
assim como aqueles que somente relatassem quantificação da
micobiota local; ou descobertas de novas espécies sem elucidar
as possíveis aplicações biotecnológicas foram excluídos da
busca.
POTENCIAIS APLICAÇÕES BIOTECNOLÓGICAS E
AMEAÇAS À MICODIVERSIDADE DO CERRADO
As alterações fitofisionômicas derivadas do
desmatamento, que historicamente vêm assolando o bioma

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
649
Cerrado, podem promover mudanças na composição do solo e
sua maior exposição aos raios solares, procipando por
exemplo, alterações físico-químicas na acidez e umidade,
fatores esses essenciais a permanência e desenvolvimento da
microbiota residente.
Dessa forma, estudar e entender a dinamicidade,
funcionamento e interação dos micro-organismos existentes no
Cerrado, e em especial no solos dessa região, não só propicia
um maior entendimento dos impactos ambientais e formas
inadequadas de manejo que o bioma vem sofrendo
(VALADARES-PEREIRA, et al.; 2017), como também viabiliza
a possibilidade se prospectar espécies com ponteciais
aplicações de interesse biotecnológico, por exemplo (NORILER
et al., 2018).
Estudo realizado por Jacinto, Moura e Uto (2018), em
diferentes áreas de cerrado (nativo; de cultivo agrícola; e de
reserva ambiental urbana/reflorestamento), visando determinar
o grau da antropicidade do solo frente à biomassa de bactérias
e fungos, evidencia os impactos que o bioma têm sofrido. Eles
verificaram contagens significativamente distintas (p < 0,05) de
bactérias em unidades formadoras de colônia por grama
(UFC.g-1) de solo, em valores de 4,2 x 105; 1,1 x 105 e 1,8 x 105,
respectivamente, nos solos das áreas de cerrado nativo, cultivo
agrícola e reflorestamento. As contagens fúgincas (UFC.g-1)
também foram significativamente distintas, 9 x 104 (áera nativa),
3 x 104 (cultivo agrícola) e 6,6 x 103 (reflorestamento), mas com
valores inferiores à da população bacteriana, indicando
segundo os autores, a maior sensibilidade dos fungos à
atividade antrópica.
Valadares-Pereira et al. (2017), também evidenciaram
mudanças na comunidade fúngica em solo do cerrado

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
650
tocantinense, ao comparar uma área nativa com outra
destinada ao plantio de soja. Analisando o ácido
desoxirribonucleico (DNA) do solo, por meio de Análise
Espacial Intergênica Ribossomal Automatizada (ARISA), e
comparando os resultados aos teores de cobre, cálcio, ferro,
capacidade de troca de cátions, saturação por bases e relação
cálcio/magnésio, de ambas as regiões, os pesquisadores
observaram mudanças na estrutura da comunidade fúgica
como efeito do manejo agrícola.
Araujo et al. (2017), avaliaram a diversidade fúngica de
uma área de cerrado preservado, na região nordeste do estado
do Piauí, dentro do Parque Nacional das Sete Cidades, ao
longo de distintos gradientes fitofisionômicos: formação
campestre, cerrado stricto sensu, cerradão e floresta decidual.
Os pesquisadores identificaram a maior riqueza de espécies na
floresta decidual, sendo a maior porcentagem dos isolados
pertencentes ao filo Ascomycota (45%), seguido do
Basidiomycota (32%). Conforme os autores, as classes mais
abundantes encontradas incluindo espécies com potencial
biotecnológico foram: Agaricomycetes, Eurotiomycetes,
Lecanoromycetes, Basidiobolus, Dothideomycetes e
Taphrinomycetes.
Conforme Bononi et al. (2017), a diversidade de micro-
organismos, e especialmente dos fungos presentes em regiões
do Cerrado, ainda não é totalmente conhecida. Em coletas
realizadas pelos pesquisadores, na região central do estado de
Mato Grosso do Sul (MS), centro-oeste do Brasil, foram
identificadas 18 espécies de fungos (Conocybe crispa,
Gymnopus dryophilus, Marasmius androsaceus, Marasmius
oreades, Mycena pura, Rhodocollybia butyraceae, Pleurotus
eryngii, Coprinellus micaceus, Panaeolus rickenii, Hypholoma

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
651
fasciculare, Clitocybe infundibuliformis, Tricholoma
argyraceum, Fomitiporia robusta, Cymatoderma elegans,
Athelopsis galzinii, Trametes ellipsospora, Guepinia
helvelloides, Tremella mesentérica) ainda não citadas e listadas
nacionalmente, e 36 espécies com primeira citação para o
estado. Do total de 62 espécies de basidiomicetos coletados, os
autores identificaram 15 comestíveis, quatro tóxicas, 10 com
propriedades medicinais, duas com potencialidade de aplicação
em processos de biorremediação, e uma com propriedade
bioluminescente, conforme comparações à literatura disponível.
Plantas medicinais têm sido muito estudadas como
reservatório de cepas de micro-organismos endofíticos com
potencial biotecnológico. Nesse sentido, Noriler et al. (2018),
estudaram comunidades fúngicas endofíticas associadas a
folhas e pecíolos da planta medicinal Stryphnodendron
adstringens, de ocorrência no Cerrado. Dentre os 117
morfotipos fúngicos (95,6% deles pertencentes ao filo
Ascomycota) isolados pelos pesquisadores, cinco (Diaporthe
sp. LGMF1530, Diaporthe cf. heveae LGMF1631,
Colletotrichum siamense LGMF1604, Fusarium sp. LGMF1630
e Epicoccum sp. LGMF1641) apresentaram atividade inibitória
(maior que 60%) sobre o crescimento micelial de Phyllosticta
citricarpa (LGMF06) e Colletotrichum abscissum (CA142),
patógenos estes de grande importância na citricultura.
Isolados endofíticos também apresentaram efeito
inibitório frente a patógenos de interesse clínico (NORILER et
al., 2018), demonstrando as potencialidades biotecnológicas
dos fungos encontrados, não só como alternativa ao controle
biológico de fitopatógenos, como também de micro-organismos
associados a saúde humana. Phaeophleospora sp.2
LGMF1513, Epicoccum sp.1 LGMF1598 e Epicoccum sp.2

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
652
LGMF1600 tiveram alta atividade de ação (+++) contra
Staphylococcus aures (American Type Culture Collection -
ATCC 33591) meticilino-resistentes (MRSA). Alguns isolados
fúngicos (Diaporthe sp. LGMF1548 e LGMF1614, Diaporthe cf.
heveae LGMF1631, Fusarium sp. LGMF1630 e Hypoxylon sp.1
LGMF1613) se mostraram também ativos contra S. aures
(MRSA), Pseudomonas aeruginosa (ATCC 27853),
Acinetobacter baumannii (BACHAC-ABA) e Klebsiella
pneumoniae (BACHC-KPC) produtora de carbapenemase
(KPC).
Em um estudo realizado com Miconia albicans (Sw.)
Triana (Melastomataceae), planta fanerógama edêmica do
Cerrado, conhecida popularmente como “quaresmeira-branca”,
Piza, Hokka e Sousa (2015) também isolaram e verificaram alta
atividade antagonista do fungo actinomiceto Amycolotopsis
orientalis frente a S. aureus, Enterococcus faecalis, Candida
albicans, Shigella sonnei.
Alternativas de controle biológico de artrópodes-praga
frente a culturas agrícolas, também vêm sendo muito aplicadas.
Por exemplo, em sistemas agroflorestais da região neotropical,
alguns organismos destacam-se como as principais pragas que
assolam as culturas agrosilvícolas, sendo as formigas
cortadeiras dos gêneros Atta e Acromyrmex, muito citadas. Tais
formigas, conhecidas popularmente como saúvas e quenquéns
atacam as partes frescas das plantas e as carregam para o
interior dos seus ninhos, onde servirão de substrato ao fungo
simbionte Leucoagaricus gongylophorus (Möller) Singer
(Basidiomycota: Agaricales: Lepiotaceae) do qual elas se
alimentam (NYGAARD et al., 2016).
A forma mais usual de controle desses insetos é pelo
método químico, entretanto a busca por alternativas

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
653
ambientalmente corretas tem sido estimulada. Nesse sentido,
Dornelas et al. (2016) realizaram estudo prospectivo de fungos
filamentosos, associados a operárias de Atta sexdens e A.
laevigata da região do cerrado tocantinense, com propriedades
patogênicas. Os autores isolaram de A. sexdens os fungos
Metarhizium anisopliae (Metchnikoff) Sorokin, Aspergillus flavus
Link, Acremonium sp.1, Aspergillus sp.1, Colletotrichum sp. e
Acremonium sp. 2, e de A. laevigata, Mucor sp., A. flavus,
Fusarium solani (Martius) Saccardo, A. niger van Tieghem;
entretanto o teste de patogenicidade contra a formiga A.
sexdens foi realizada somente com os fungos A. flavus, A. niger
e M. anisopliae, pois esses apresentaram alta produção de
conídios e elevada prevalência no estudo. Foi verificada uma
taxa de letalidade de 50% das operárias em apenas cinco dias,
nos testes com A. niger e M. anisopliae, sendo essa duração
significativamente inferior ao tratamento controle.
Estudo realizado por Zanardo (2015) em solos dos cinco
biomas do Brasil corrobora a importância que o Cerrado tem em
abrigar uma das maiores quantidades de fungos
entomopatogênicos. A autora determinou a abundância de
fungos ao longo de dois anos em amostras de solos de cultivos
anuais, perenes e vegetação nativa, durante estação climática
seca e úmida, de cinco áreas representativas dos biomas
brasileiros, e verificou no geral, maiores densidades de
entomopatogênicos no Cerrado (2,27 ± 1,58 x 103 a 3,46 ± 1,51
x 104 UFC.g-1 de solo), sendo o gênero Metarhizium spp. o de
maior predominância.
Conforme mencionado anteriormente, além do emprego
dos fungos no controle biológico e como bioindicadores da
qualidade do solo em manejos diversos, esses micro-
organismos tem importância preponderante no setor agrícola

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
654
atuando na melhoria da produtividade das culturas, não só
porque realizam a ciclagem de nutrientes, ao decompor e
transformar materiais orgânicos em minerais, que serão assim
aproveitados pelas plantas, mas também pela associação
simbionte (micorrizas) com raízes de muitas espécies vegetais,
proporcionando a essas aumento da superfície de contato com
o solo, para absorção de água e sais minerais.
Dentre os grupos de micorrizas destacam-se os fungos
micorrízicos arbusculares (FMA), também denominados
endomicorrizas, que tem como característica diferencial
desenvolver um micélio que penetra internamente no córtex da
raiz, invadindo tanto o espaço intra quanto intercelular. Berude
et al. (2015), em pesquisa bibliográfica, compilaram outros
distintos benefícios às plantas quando de sua associação aos
FMA: redução ao estresse hídrico e salino; diminuição de sítios
de infecção por patógenos; aumento na absorção de fósforo e
nitrogênio do solo e maior tolerância a metais pesados.
Experimento realizado por Salgado et al. (2016), ilustra a
potencialidade de fungos do Cerrado em propiciar o
crescimento inicial de plantas de feijoeiro e soja. Os
pesquisadores coletaram, isolaram e idetificaram FMA
(Acaulospora scrobiculata, Claroideoglomus etunicatum,
Dentiscutata heterogama, Gigaspora margarita e Rhizophagus
clarus) provenientes de solo do Cerrado (região sudoeste de
Goiás), os quais foram associados ao estimulante de
colonização (7-hidroxi,4’-metoxi-isoflavona) e inoculados sobre
ambas culturas. Eles observaram aumento da biomassa nas
plantas, mais de 200% para o feijoeiro e mais de 80% para a
soja, partindo-se dos seguintes parâmetros de avaliação:
massa seca da parte aérea, massa seca da raiz, colonização

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
655
micorrízica, massa de nódulos e acúmulo de cálcio, zinco e
fósforo na massa seca da parte aérea.
Em outro experimento, partindo das mesmas
condicionantes, Salgado et al. (2017), determinaram os efeitos
de tal combinação frente ao crescimento inicial de plantas de
algodão e milho, e verificaram que baixa densidade de
propágulos de FMA inoculados e associados a estimulante de
colonização foram também benéficos ao desenvolvimento de
ambas culturas.
Santos et al. (2018), avaliaram os efeitos da inoculação
de FMA, provenientes de solo de Cerrado conservado, sobre
uma região degradada pela construção de uma usina
hidrelétrica, no município de Selvíria-MS. Eles verificaram que
a combinação de resíduos orgânicos/minerais com a aplicação
dos fungos influenciou positivamente o solo degradado, pois o
teor de carbono da massa microbiana (CBM), carbono de gás
cabônico liberado (C-CO2) e quociente microbiano (qMic)
elevaram-se. Todos esses parâmetros são indicadores de
qualidade do solo.
Outro aspecto a se considerar são os estudos sobre
emprego da atividade fúngica para a melhoramento de
processos de produção, extração, solubilização e
transformação no setor industrial. Os mesmos vêm crescendo
e propiciando vantagens em termos ambientais e de obtenção
de produtos com maior valor econômico agregado. A produção
de biossurfactantes por micro-organismos, por exemplo, é uma
opção promissora em termos de obtenção de produtos
biotecnológicos, haja visto a baixa toxicidade, alta
biodegrabilidade e melhor compatibilidade ambiental quando
comparados aos surfactantes sintetizados quimicamente.

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
656
Partindo disso, Passone et al. (2015) avaliaram o
potencial biossurfactante de três fungos isolados de ambientes
do Cerrado na região do Triângulo Mineiro, codificados como
C18, C35 e C40, frente a diferentes concentrações de glicerol,
empregando fermentação submersa. Os autores observaram
índices de emulsificação variando de 28,6% a 71,4%. Foi
notado também aumento nesse parâmetro (em média 10%),
com o aumento do tempo de fermentação (48 h para 72 h), nos
cultivos contendo concentrações de glicerol 2,5; 5,0 e 10% (v/v)
inoculados com o fungo C35, além um dos menores valores de
tensão supercial (40 mN/m-1) na condição de 1,5% (v/v) de
glicerol a 48 h. Altos íncides de emulsificação e baixos valores
de tensão superficial são bons indicativos de atividade
surfactante.
A produção e oferta interna de fertilizantes fosfatados no
Brasil ainda tem sido um gargalo no setor agrícola,
especialmente porque a rocha fosfática, principal insumo da
indústria de fertilizantes, é um mineral pouco disponível no país
e ainda apresenta baixo teor de pentóxido de fósforo (P2O5).
Solos aráveis, como os do Cerrado, demandam fósforo que é
pouco disponibilizado e muitas vezes insolúvel para ser
utilizado pelas culturas agrícolas, necessitando assim serem
suplementados com fertilizantes. Associado a isso, a extração
dos fosfatos é realizada empregando-se ácido fosfórico (H3PO4)
ou sulfúrico (H2SO4), o que gera resíduos. Tais condições
tornam o custo da produção nacional elevado e ambientalmente
pouco sustentável, suscitando pesquisas que se utilizem de
métodos naturais renováveis e que agridam menos o ambiente,
tal como a biossolubilização realizada por micro-organismos
(EMBRAPA, 2015).

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
657
Nesse sentido, Cabral et al. (2017) isolaram Trichoderma
harizianum, um fungo filamentoso do filo Ascomycota, de um
complexo minerador do município de Araxá-MG, e testaram sua
capacidade de biossolubilizar fósforo a partir de concentrado de
rocha fosfática (teor de P2O5 igual a 37,5%) originária do
complexo de mineração do município de Tapira-MG.
Empregando-se fermentação submersa em biorreator air-lift
com reciclo do meio reacional, e variando o fluxo volumétrico de
ar ascendente e a concentração de rocha fosfática, os
pesquisadores obtiveram uma concentração de fósforo solúvel
em meio líquido igual a 244,9 partes por milhão (ppm) ou
33,78% de solubilização, num período de três dias a 28 ± 0,5°C.
Gomes et al. (2017), obtiveram 11,95% (94,8 ppm) de
solubilização de fósforo em 14 dias, e 6,81% (52 ppm) em 9
dias, empregando o mesmo concentrado de rocha fosfática, na
fermentação sólida e líquida, respectivamente, com o fungo T.
ressei, coletado no Cerrado.
Estudo realizado por Rocha (2017), com a levedura
Torulaspora globosa, isolada da rizosfera de milho, no
município de Araras-SP, evidenciou também a rápida
solubilização de fosfato tricálcio (Ca3(PO4)2) em fermentação
submersa. A autora obteve quantidade máxima de fósforo
solúvel (42%) em apenas 48 h, demonstrando a potencialidade
do fungo em aplicações industriais e agrícolas.
Propiciar o aumento e variedade da quimiodiversidade,
isto é, da obtenção de novas substâncias derivadas da ação
metabólica de micro-organismos e/ou enzimas isoladas, sobre
compostos orgânicos ou sintéticos, têm sido uma nova
estratégia biotecnológica alternativa de síntese química, dentro
da área de química verde. Para Hegazy et al. (2015), o uso
desse processo apresenta vantagens quando comparado a

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
658
síntese tradicional: baixo custo; quimio, estereo e
regioseletividade; manutenção das cepas; redução no número
de animais experimentais; maior quantidade e variedade de
compostos produzidos; consumo mínimo de energia e menor
geração de resíduos, especialmente compostos tóxicos e
danosos ao ambiente.
Silva (2015), testaram cepas de Beauveria sp. isoladas
de solo do Cerrado da região de Goiás, frente a ácido gálico
(AG) e elágico (AE), utilizando cultivo submerso em meio
glicose e Corn Steep, durante 96 h a 27ºC e 200 rotações por
minuto (rpm). Empregando-se Cromatografia em Camada
Delgada (CCD) e Cromatografia Líquida de Alta Eficiência
(CLAE), foi notado que todas as cepas foram eficientes na
biotransformação dos compostos, sendo que o meio contendo
Corn Steep proporcionou maior variedade e quantidade de
compostos formados para ambos substratos. Os derivados do
AG obtidos e caracterizados estruturalmente por métodos
espectroscópicos foram: ácido benzoico, 3-O-metil-β-
glucopiranosil-galatoester (Labiocon 465); ácido benzoico,4-[[6-
O-(3,4-diidroxi-5-metoxibenzoico)-α-D-glucopiranosideo]oxi]-3-
hidroxi-5metoxi (Labiocon 484); ácido gálico 3-O-β-
glucopiranoside (Labiocon 510) e ácido 3,4-Diidroxi-5-metoxi-
benzoico-2-hidroxi-5-metoxicarbonil-fenilester (Labiocon 530).
E os do AE: 3,8-Dimetoxi-dibenzo[b,d]piran-6-one (Labiocon
511) e o ácido elágico 4-O,3’-O-metil-β-D-glicose (Labiocon
534).
Na indústria de alimentos, algo que tem se tornado
corriqueiro é o emprego de insumos e/ou ingredientes com
caráter mais natural para a elaboração de produtos
alimentícios, dada a mudança de atitudes do consumidor e sua
maior preocupação com a saúde. Para isso, tem-se por

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
659
exemplo substituído o emprego de aditivos químicos artificiais,
por compostos naturais derivados de animais, plantas ou
mesmo micro-organismos.
Conforme Rao, Xiao e Li (2017), a demanda por corantes
alimentícios naturais derivados de micro-organismos, por
exemplo, tem aumentado dia a dia, não somente pelos efeitos
nocivos cientificamente relatados da ingestão de alguns
corantes sintéticos, mas também porque pigmentos de origem
microbiana apresentam vantagens quando comparados a
outros produtos naturais: rapidez na produção; fácil
processamento; independência das condições climáticas; e
propriedades antioxidantes, antimicrobianas e
anticancerígenas.
Entre os distintos pigmentos microbianos, aqueles
derivados de fungos ambientais e especialmente do solo, tem
sido pesquisados e vem apresentando-se como alternativa
favorável não só na medicina, como também na indústria
cosmética, têxtil e principalmente alimentícia. Nesse sentido,
Machado (2018) isolou leveduras com potencial carotenogênico
do ambiente (folhas, solo, flores, frutos silvestres e insetos) do
Cerrado e verificou que as espécies Rhodotorula glutinis, R.
graminis, R. aurantiaca, R. lactosa e R. mucilagionosa se
destacaram na produção de carotenoides. Realizando testes de
otimização em condições de cultivo experimental submerso
com variação de agitação (130 a 230 rpm), temperatura (25 a
35ºC), pH (4,5 a 6,5) e componentes do meio de cultivo, o autor
observou que a levedura R. mucilaginosa foi a mais efetiva na
bioprodução do pigmento (4164,45 µg.L-1).
Em outro estudo bioprospectivo de fungos endofítifos de
folhas de Hymenaeae coubaril (jatobá), árvore típica do
cerrado, Romano (2015) identificou pontencial produção de

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
660
pigmento vermelho pelo fungo Talaromyces minioluteus,
empregando-se diferentes meios de cultivo a 28ºC, por 72 h e
a 200 rpm, ao se comparar a mensuração da biomassa após os
cultivos, com índices de absorbâncias obtidos a 510 nm.
Conforme o autor, a intensa coloração vermelha obtida em
todos os cultivos realizados evidencia o possível emprego
biotecnológico da espécie, entretanto sugere-se a continuidade
das pesquisas visando-se não somente identificar
quimicamente o pigmento, como verificar a micotoxidade,
empregar substratos alternativos de menor custo, e também
otimizar o processo de produção.
No setor industrial, especialmente alimentício, a
produção biotecnológica de frutooligossacarídeos (FOS) e
exopolissacarídeos (EPS) empregando-se micro-organismos
têm sido muito demandada, não somente por serem
considerados naturais; como também pela praticididade e
facilidade de obtenção quando comparado a extração realizada
de fontes vegetais; além desses polissacarídeos apresentarem
benefícios à saúde, bem como propriedades tecnológicas que
melhoram as características físico-químicas, reológicas e
sensoriais dos produtos destinados ao consumidor
(BARCELOS et al., 2019).
Diante dos benefícios e aplicações dos FOS, Zaninette
et al. (2019) observaram atividade hidrolítica e capacidade de
transfrutosilação de solução de sacarose (150 g.L-1), com
produção de 8 g.L-1 de 1-cestose (FOS do tipo inulina) e 7,3 g.L-
1 de neocestose (FOS do tipo neolevana, comumente não
produzido por fungos filamentosos), pelo fungo Penicillium
janczewskii Zaleski CCIBt 3352, isolado da rizosfera de uma
planta do Cerrado (Chrysolaena obovata), evidenciando a
promissora capacidade sintetizadora de FOS por uma cepa

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
661
selvagem. Outra pesquisa realizada com diferentes frutos do
Cerrado (SILVEIRA, 2018), corrobora a necessidade de
estudos prospectivos com viés biotecnológico mais
sistematizados sobre o bioma. A autora isolou 17% de micro-
organismos (de um total de 59), com potencialialidades de
produção de EPS, sendo que uma das linhagens alcançou uma
produção máxima desses bioprodutos de 1,2 g.L-1 após 2 h de
processo a 30ºC e 150 rpm.
No setor de energias renováveis, em especial na
produção e melhoria de processos para obtenção
biocombustíveis e/ou biodiesel, os fungos também vêm
apresentando suas potenciais aplicações.
Nos últimos anos o crescimento incessante da população
tem demandado o consumo de recursos energéticos para suprir
o bem-estar individual, desempenho industrial ou mesmo
serviços de manutenção; promovendo a escassez de fontes de
energia não renováveis como o petróleo, por exemplo, e a
intensa emissão de gases poluentes para a atmosfera. Assim,
o uso da bionconversão de diferentes resíduos empregando-se
fermentação microbiana veêm sendo adotado como alternativa
para se otimizar a aquisição de combustíveis mais sustentáveis,
e ao mesmo tempo minimizar alterações ambientais
decorrentes da disposição de subprodutos e rejeitos com
características poluentes (ZANONI et al., 2015).
Um dos gargalos da produção de biocombustíveis,
especialmente os de segunda geração, deve-se às dificuldades
em se despolimerizar as longas cadeias carbônicas que
constituem a parede celular vegetal, para se disponibilizar os
açúcares fermentescíveis que serão convertidos em álcool.
Assim, diferentes estudos e esforços públicos e privados têm
sido realizados, visando selecionar micro-organismos que

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
662
apresentem tal capacidade bioconversora, de forma a se
reduzir o custo atual do refinamento da biomassa vegetal.
Diante disso, Dabaja et al. (2019), testaram 43 culturas
de fungos endofíticos isolados de Annona crassiflora (araticum),
uma planta nativa e exclusiva do Cerrado, e verificaram que 8%
apresentaram atividade esterásica; 34,43% atividade
celulolítica; 73,91% atividade fosfatásica; 78,26% atividade
lipolítica; e 91,3% atividade β-frutocfuranosidásica. Os autores
ainda realizaram a fermentação submersa para determinação
da atividade lipolítica, visto serem as lipases enzimas muito
aplicadas em biocombustíveis (principalmente na produção de
biodiesel), e constataram uma atividade expressiva de 27,34
U.g-1 em 60 h de fermentação a 28ºC.
Bessa (2018) isolou fungos do solo do Cerrado e
verificou atividades lipolíticas promissoras de 9,268 U.mg-1 e
7,032 U.mg-1 para duas cepas Paecilomyces formosus (isolado
I e II); e 6,447 U.mg-1 para Aspergillus tubingensis. A autora
ainda observou que o extrato enzimático bruto proveniente do
isolado II de P. formosus conseguiu atingir 41,60% de
conversão de óleo:metanol em ésteres metílicos. Tal
capacidade de transesterificação demonstra o potencial das
enzimas para emprego na produção de biodiesel,
principalmente porque esse processo é considerado mais limpo
quando comparado ao uso de catalizadores químicos.
Dentre os distintos micro-organismos presentes no solo
do Cerrado, bactérias e fungos que apresentam capacidade de
excretar enzimas para o meio, tem sido alvo de interesse nos
estudos relacionados à bioprospecção e biotecnologia. Isso
porque, a secreção extracelular enzimática, realizada
naturalmente pelo micro-organismo com objetivo de promover
a degradação de compostos complexos para sua posterior

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
663
absorção e nutrição, reduz custos nos sistemas de isolamento
de tais bioprodutos quando comparado aos processos onde é
necessário o rompimento celular (ANDRADE-LINHARES;
VERESOGLOU; RILLIG, 2015). Além disso, biocompostos
excretados tendem, de forma geral, a ser mais estáveis aos
distintos fatores ambientais, quando comparados àqueles que
se mantêm no interior da célula, em condições fisiológicas
favoráreis.
Silva et al. (2015), por exemplo, estudando a capacidade
de 66 estirpes de Trichoderma spp. isoladas do solo e madeira
em decomposição da região do cerrado tocantinense,
verificaram que dois dos isolados (UFTJCO85 e UFTMG15)
apresentaram atividades enzimáticas celulásicas satisfatórias
para FPase (1,34 UI.mL-1) e CMCase (0,81 UI.mL-1), haja visto
que não foram empregados tratamentos prévios do sabugo de
milho testado.
Oliveira et al. (2015) testaram a atividade amilolítica de
enzimas produzidas por leveduras isoladas de ambientais
naturais (frutas e serrapilheira) e industriais (mosto de usina de
cana-de-açúcar) da região de Dourados (MS), e identificaram
três cepas de um total de 54, com boas capacidades de produzir
amilases, a partir de farelo de trigo a 70% de umidade. Entre as
cepas destacaram-se Candida parapsilosis com atividade
máxima 14,68 U.mL-1; Rhodotorula mucilaginosa com 25,0
U.mL-1 e Candida glabrata com 25,39 U.mL-1, empregando
cultivo em estado sólido por 120 h a 28ºC. A amilases
produzidas por tais cepas exibiram atividade máxima em pH 7,0
e temperatura de 60ºC; apresentaram atividade mesmo após 1
h de incubação a 50°C; mativeram atividade catalítica residual
maior que 70% após exposição a solução de 15% de etanol; e
ainda apresentaram maiores atividades enzimáticas frente ao

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
664
amido de milho, quando comparado aos amidos de batata, trigo
e mandioca.
Santos et al. (2016) destacaram o potencial
biotecnológico de outro fungo também isolado do solo da região
de Dourados (MS), com capacidade de produção de β-
glicosidades, enzimas empregadas na produção de
biocombustíveis; vinificação e desenvolvimento de alimentos
funcionais. A cepa de G. butleri isolada apresentou a máxima
produção de β-glicosidades (21,5 U.mL-1), em cultivo em estado
sólido a 30°C por 96 h, sobre farelo de trigo a 55% de umidade.
Os autores verificaram ainda uma atividade catalítica máxima
em pH 4,5 e temperatura de 65°C, com retenção de 80% da
atividade mesmo após manutenção a 50ºC por 1 h.
Em pesquisa realizada por Melo et al. (2018), em
amostras de solo e serrapilheira coletados no Parque Estadual
do Cerrado em Jagariúna (PR), 89,3% dos isolados fúngicos
pertencentes ao filo Ascomycota e Basiodiomycota,
apresentaram atividade celulásica em meio de cultura contendo
carboximetilcelulose (CMC), tendo se destacado Colletotrichum
boninense e Trichoderma sp. Os autores observaram ainda que
todas as cepas apresentaram alguma atividade lipolítica,
demonstrando potencialidades para serem empregadas em
processos biotecnológicos que envolvam degradação de
biomassa vegetal e produção de biocombustíveis.
CONCLUSÕES
As diferentes pesquisas relatadas apontam a
importância e potencialidades do emprego dos fungos do
Cerrado em diferentes produtos, processos, e atividades, sejam
elas agropecuárias, industriais, alimentícias, medicinais,

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
665
químicas, entre outras; e elucidam a necessidade de pesquisas
e ações de maior conservação desse bioma, que ainda possui
uma enorme e variada biodiversidade fúngica ainda pouca
conhecida e inexplorada.
REFERÊNCIAS ARAUJO, A.S.F. et al. Fungal diversity in soils across a gradient of preserved Brazilian Cerrado. J Microbiol, v.55, n.4, p. 273-79, 2017. BARCELOS, M.C.S. et al. Current status of biotechnological production and applications of microbial exopolysaccharides. Crit Rev Food Sci Nutr, v.60, n.9, p.1475-95, 2019. BECHARA, R. et al. Methodologya for the optimal design of integrated first and second generation ethanol production plant combined with power cogeneration. Bioresour Technol, v.214, p.441-49, 2016. BESSA, D.H.R.F. Prospecção de enzimas lipolíticas em fungos para a produção de biodiesel. Dissertação (Mestrado). Instituto Federal de Educação, Ciência e Tecnologia Goiano. 51fl. Rio Verde - GO, 2019. BERUDE, M.C. et al. Microrrizas e sua importância agroecológica. Encicl Biosf, v.11, n.22, p.132-46, 2015. BONONI, V.L.R. et al. Agaricomycetes (Basidiomycota, Fungi) diversity in a protected area in the Maracaju Mountains, in the brazilian central region. Hoehnea, v.44, n.3, p. 361-77, 2017. CABRAL, B.V. et al. Recycling influence in phosphorus biossolubilization from rock phosphate concentrate in air-lift bioreactor. Chem Eng Trans, v.57, p. 673-78, 2017. DABAJA, M.Z. et al. Avaliação da atividade enzimática de fungo endofítico isolado de Annona crassiflora (Marolo) com interesse biotecnológico. Rev Univ Vale do Rio Verde, v.17, n.1, p.1-10, 2019. DAVIS, R. et al. Process design and economics for the conversion of lignocellulosic biomass to hydrocarbons: dilute-acid and enzymatic desconstruction of biomass to sugars and catalytic conversion of sugars to hydrocarbons. Golden: National Renewable Energy Laboratory, 2015. DORNELAS, A.S.P. et al. Fungos filamentosos associados às espécies Atta sexdens (Linnaeus) e Atta laevigata (F. Smith) (Hymenoptera: Formicidae). EntomoBrasilis, v.9, n.1, p. 26-30, 2016. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA – EMBRAPA. Biossolubilização de potássio in vitro a partir da rocha fonolito por

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
666
microrganismos do solo. Org.: SILVA, U. C. et al. Sete Lagoas: Embrapa Milho e Sorgo, 2015. (Documentos/Embrapa Milho e Sorgo 177). GOMES, B.C.C. et al. Avaliação da biossolubilização de fósforo proveniente de concentrado de rocha fosfática por fermentação sólida e líquida utilizando Trichoderma ressei. In: CONGRESSO BRASILEIRO DE ENGENHARIA QUÍMICA EM INICIAÇÃO CIENTÍFICA, 12., 2017, São Carlos. Anais... São Paulo: Blucher Chemical Engineering Proceedings, v.1, n.4, 2017. HEGAZY, M.F. et al. Microbial biotransformation as a tool for drug development based on natural products from mevalonic acid pathway: A review. J Adv Res, v.6, n.1, p. 17-33, 2015. JACINTO, H.R.; MOURA, J.B.; UTO, F.N. Avaliação da influência antrópica sobre a microbiota edáfica de cerrado no município de Anápolis, Goiás. Ipê Agron J, v.2, n.2, p. 4-17, 2018. MACHADO, W.R.C. Bioprospecção de leveduras para a produção de carotenpides microbianos. Tese (Doutorado). Universidade Estadual Paulista. Instituto de Biociências, Letras e Ciências Exatas. 123fl. São José do Rio Preto - SP, 2018. MELO, M. et al. Cellulolytic and lipolytic fungi isolated from soil and leaf litter samples from the Cerrado (Brazilian Savanna). Rev Biol Trop, v.66, n.1, p.237-45, 2018. NORILER, S.A. et al. Bioprospecting and structure of fungal endophyte communities found in the brazilian biomes, Pantanal, and Cerrado. Front Microbiol, v.9, e1526, 2018. NYGAARD, S. et al. Reciprocal genomic evolution in the ant-fungus agricultural symbiosis. Nat Commun, v.7, n.12233, p.1-9, 2016. OLIVEIRA, A.P.A. et al. Bioprospecting of yeasts for amylase production in solid state fermentation and evaluation of the catalytic properties of enzymatic extracts. Afr J Biotechnol, v.14, n.14, p. 1215-23, 2015. PASSONE, J.M. et al. Avaliação da produção de biossurfactante empregando fungos e glicerol como substrato. In: CONGRESSO BRASILEIRO DE ENGENHARIA QUÍMICA EM INICIAÇÃO CIENTÍFICA, 11., 2015, Campinas. Anais... São Paulo: Blucher Chemical Engineering Proceedings, v.1, n.3, 2015. PIZA, A.C.M.T.; HOKKA, C.O.; SOUSA, C.P. Endophytic actinomycetes from Miconia albicans (Sw.) Triana (Melastomataceae) and evaluation of its antimicrobial activity. J Sci Res Rep, v.4, n.4, p.281-91, 2015. RAO, M.P.N.; XIAO, M.; LI, J-W. Fungal and bacterial pigments: secondary metabolites with wide applications. Front Microbiol, v.8, 1113, 2017. RESENDE, F.M. et al. Consequences of delaying actions for safeguarding ecosystem services in the Brazilian Cerrado. Biol Conserv, v.234, p.90-9, 2019.

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
667
ROCHA, R.K. Solubilização in vitro de fosfato por Torulaspora globosa. Dissertação (Mestrado). Universidade Federal de São Carlos. Centro de Ciências Agrárias. 59fl. Araras - SP, 2017. ROMANO, L.H. Bioprospecção de microrganismos endofíticos isolados de Tabebuia spp. e Hymenaeae courbaril e identificação da produção de metabólitos de interesse biotecnológico. Tese (Doutorado). Universidade Federal de São Carlos. 134fl. São Carlos - SP, 2015. SALGADO, F.H.M. et al. Arbuscular mycorrhizal fungi and mycorrhizal stimulant affect dry matter and nutrient accumulation in bean and soybean plants. Pesq Agropec Trop, v.46, n.4, p.367-73, 2016. SALGADO, F.H.M. et al. Arbuscular mycorrhizal fungi and colonization stimulant in cotton and maize. Ciênc Rural, v.47, n.6, e20151535, 2017. SANTOS, A.A. et al. Addition of waste and introduction of microorganisms after 45 years of soil degradation. Rev Ciênc Agron, v.49, n.3, p.363-70, 2018. SANTOS, F.R.S. et al. Production and characterization of β-glucosidase from Gongronella butleri by solid-state fermentation. Afr J Biotechnol, v.15, n.16, p.633-41, 2016. SAWYER, D. et al. Perfil do ecossistema hotspot de biodiversidade do Cerrado. Critical Ecosystem Partnership Fund, 2016. SILVA, D.B. et al. Bioprospecção de linhagens de Trichoderma sp. na degradação de resíduos lignocelulósicos de sabugo de milho. Rev Verde Agroecologia Desenvolv Sustent, v.10, n.1, p.252-58, 2015. SILVA, L.S. Biotransformação dos ácidos gálico e elágico por microorganismo do Cerrado brasileiro. Dissertação (Mestrado). Universidade Federal de Goiás. Faculdade de Farmácia. 157fl. Goiânia - GO, 2015. SILVEIRA, B.M.P. Seleção de biocatalizador com potencial para a produção de exopolissacarídeos. Dissertação (Mestrado). Universidade Federal dos Vales do Jequitinhonha e Mucuri. 136fl. Diamantina - MG, 2018. VALADARES-PEREIRA, A.A. et al. Fungal community structure as na indicator of soil agricultural management effects in the Cerrado. Rev Bras Ciênc Solo, v.41, e0160489, p.1-12, 2017. ZANARDO, A.B.R. Abundância de fungos entomopatogênicos da ordem Hypocreales e diversidade genética de Metarhizium spp. isolados de amostras de solo de áreas representativas de cinco biomas brasileiros. Tese (Doutorado). Universidade de São Paulo. Escola Superior de Agricultura “Luiz de Queiroz”. 153fl. Piracicaba - SP, 2015. ZANINETTE, F. et al. Production of inulin and neolevan-type fructooligosaccharides by Penicillum janczewskii Zaleski CCIBt 3352. Biotechnol Appl Biochem, v.66, n.3, p. 419-25, 2019.

APLICAÇÕES BIOTECNOLÓGICAS E AMEAÇAS À DIVERSIDADE FÚNGICA DO
CERRADO
668
ZANONI, P.R.S. et al. Produção de etanol: uma opção para aproveitamento de resíduos gerados nas indústrias de reciclagem de papel. Colombo: Embrapa Florestas, 2015.

669
Este livro foi publicado em 2021 IMEA
Intituto Medeiros de Educação Avançada Av Senador Ruy Carneiro, 115 ANDAR: 1; CXPST: 072;
Joao Pessoa - PB 58032-100




















