
Geographical variation and taxonomy of the snail-eating snake Dipsas albifrons (Sauvage, 1884), with...
16
1013 Accepted by S. Carranza: 15 Jun. 2005; published: 30 Jun. 2005 19 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2005 Magnolia Press Zootaxa 1013: 19–34 (2005) www.mapress.com/zootaxa/ Geographical variation and taxonomy of the snail-eating snake Dipsas albifrons (Sauvage, 1884), with comments on the systematic status of Dipsas albifrons cavalheiroi Hoge, 1950 (Serpentes: Colubridae: Dipsadinae) PAULO PASSOS 1, 3 , RONALDO FERNANDES 1 & MARCOVAN PORTO 2 1 Departamento de Vertebrados, Museu Nacional, Universidade Federal do Rio de Janeiro, Quinta da Boa Vista, Rio de Janeiro, RJ, 20940-040, Brazil. 2 Faculdade de Veterinária, Fundação Educacional Serra dos Órgãos, Teresópolis, RJ, 25976-340 and Pro- grama de Pós-graduação em Biologia Animal, Universidade Federal Rural do Rio de Janeiro, Seropédica, RJ, 23851-970, Brazil. 3 E-mail: [email protected] Abstract The taxonomic status of Dipsas albifrons is revised and geographic variation of external morphol- ogy and hemipenis are described. Multivariate analyses suggest that the pattern of geographical variation is correlated with gap sampling, probably indicating a northernward cline in segmental counts (mainly ventral and subcaudal scales). Quantitative and qualitative analyses reveal that the island population (Dipsas albifrons cavalheiroi) is not divergent from mainland populations and, herein, is considered a junior synonymy of Dipsas albifrons. A lectotype of Dipsas albifrons is des- ignated. Key words: Serpentes, Dipsadinae, Dipsas albifrons, intraespecific variation, population status, taxonomy Introduction The gastropod-eating dipsadine snakes constitute a monophyletic group characterized by a series of morphological features (Brongesma 1958; Peters 1960; Fernandes 1995). Peters (1960) provided a comprehensive taxonomic revision of these snakes and recognized seven species groups for Dipsas Laurenti, 1768, placing D. albifrons (Sauvage, 1884) in the D. variegata group with D. variegata (Duméril, Bibron & Duméril, 1854) and D. incerta (Jan, 1863). This group was defined by Peters (1960) to include species with 15
Transcript of Geographical variation and taxonomy of the snail-eating snake Dipsas albifrons (Sauvage, 1884), with...

1013
Accepted by S. Carranza: 15 Jun. 2005; published: 30 Jun. 2005 19
ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2005 Magnolia Press
Zootaxa 1013: 19–34 (2005) www.mapress.com/zootaxa/
Geographical variation and taxonomy of the snail-eating snake Dipsas albifrons (Sauvage, 1884), with comments on the systematic status of Dipsas albifrons cavalheiroi Hoge, 1950 (Serpentes: Colubridae: Dipsadinae)
PAULO PASSOS1, 3, RONALDO FERNANDES1 & MARCOVAN PORTO2
1Departamento de Vertebrados, Museu Nacional, Universidade Federal do Rio de Janeiro, Quinta da Boa Vista, Rio de Janeiro, RJ, 20940-040, Brazil.2Faculdade de Veterinária, Fundação Educacional Serra dos Órgãos, Teresópolis, RJ, 25976-340 and Pro-grama de Pós-graduação em Biologia Animal, Universidade Federal Rural do Rio de Janeiro, Seropédica, RJ, 23851-970, Brazil.3E-mail: [email protected]
Abstract
The taxonomic status of Dipsas albifrons is revised and geographic variation of external morphol-ogy and hemipenis are described. Multivariate analyses suggest that the pattern of geographicalvariation is correlated with gap sampling, probably indicating a northernward cline in segmentalcounts (mainly ventral and subcaudal scales). Quantitative and qualitative analyses reveal that theisland population (Dipsas albifrons cavalheiroi) is not divergent from mainland populations and,herein, is considered a junior synonymy of Dipsas albifrons. A lectotype of Dipsas albifrons is des-ignated.
Key words: Serpentes, Dipsadinae, Dipsas albifrons, intraespecific variation, population status,taxonomy
Introduction
The gastropod-eating dipsadine snakes constitute a monophyletic group characterized by aseries of morphological features (Brongesma 1958; Peters 1960; Fernandes 1995). Peters(1960) provided a comprehensive taxonomic revision of these snakes and recognizedseven species groups for Dipsas Laurenti, 1768, placing D. albifrons (Sauvage, 1884) inthe D. variegata group with D. variegata (Duméril, Bibron & Duméril, 1854) and D.incerta (Jan, 1863). This group was defined by Peters (1960) to include species with 15

PASSOS ET AL.20 © 2005 Magnolia Press
1013ZOOTAXA dorsal scales rows and pattern of dorsal blotches higher than wide and, at least posteriorly,
much narrower than interspaces.Recently, some studies have focused on the taxonomic status of species from Dipsas
variegata species group and other related taxa. Porto and Fernandes (1996) reported mor-phological variation in D. neivai Amaral, 1926, and suggested a close relationship with D.variegata. Cadle and Myers (2003) revised specimens referred to D. variegata in Panamaand western South America, recognizing D. andiana (Boulenger, 1896) and D. nicholsi(Dunn, 1933) as valid species. Finally, Passos et al. (2004) revised D. incerta, restrictingthis species to the Guianas, recognizing the Atlantic Forest populations as D. alternans
(Fischer, 1885), and synonymizing D. copei (Günther, 1872) with D. incerta.The taxonomic status of the Atlantic Rain Forest species Dipsas albifrons remains
poorly knows since the original description by Sauvage (1884). All earlier identificationswere based only a brief original description, without further examination of the type spec-imens. This may explain why this species has been associated with several genera, such asDipsadomorus, Leptognathus, Dipsas, Conchliophagus, and Sibynomorphus, by differentauthors (Sauvage 1884; Boulenger 1896; Mocquard 1908; Ihering 1911; Amaral 1929a,b). In the last revision, Peters (1960) questioned the association of the material he exam-ined with the Sauvage’s types, as he could not exam the latter.
Based on four female specimens, Hoge (1950) described Dipsas albifrons cavalheiroifrom Queimada Grande Island. He distinguished his subspecies from the nominate race bylower numbers of ventral and subcaudal scales and by the rounded shape of the dorsalscales. Without examining a single specimen from Queimada Grande Island, Peters (1960)synonymized D. a. cavalheroi with the nominal species based on the overlapping scalecounts and the unreliability of the dorsal scale shape as a character. Nevertheless, in thecurrent edition of the Brazilian Red List of Endangered Species (BRASIL 2003), D. a.cavalheiroi was cited as critically threatened.
In this paper we intend to evaluate the taxonomic status of continental and island pop-ulations referred to Dipsas albifrons in order to infer boundaries among putative naturalentities.
Materials and Methods
Material and charactersSpecimens examined are housed in the Instituto Butantan, São Paulo, Brazil (IBSP),
Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil (MNRJ),and Museu de Zoologia, Universidade de São Paulo, São Paulo, Brazil (MZUSP).Referred specimens and localities are listed in the Appendix.
Terminology for Dipsas cephalic shields follows Peters (1960), whereas the method ofventral counting follows Dowling (1951); hemipenial terminology is based on Dowlingand Savage (1960), as augmented by Myers and Campbell (1981). Measurements were

© 2005 Magnolia Press 21DIPSAS ALBIFRONS
1013ZOOTAXAtaken with a dial caliper to the nearest 0.1 mm, except for snout-vent (SVL) and tail
lengths, which were taken with a flexible ruler to nearest 1.0 mm. Sex was determined bysubcaudal incision. Description of the hemipenis is based on five organs examined in situand seven organs everted using the procedures of Pesantes (1994). Specimens with evertedhemipenis are also listed in the appendix.
FIGURE 1. Geographical distribution of Dipsas albifrons, showing the three morphometricgroups.
Statistical analysesWe partitioned the sample into three groups, reflecting the disjointed geographical dis-
tribution of the species (Fig. 1). Group 1 included specimens from Santa Catarina andParaná, as well as a single specimen from the continental portion of São Paulo. Group 2included specimens from Queimada Grande Island, São Paulo. Group 3 included speci-mens from the Espírito Santo. We excluded three specimens from the Rio de Janeiro, asthey were too isolated to be lumped in one of the three groups and too few to form theirown group. For the statistical analyses we used number of blotches on the body and tailand numbers of ventral, subcaudal, supralabial, and infralabial scales. We employed ananalysis of variance (ANOVA) using segmental counts to test for sexual dimorphismwithin each group and within the whole sample. We evaluated the assumptions of univari-ate normality and homocedasticity with the Kolmogorov-Smirnov’s and Levene’s tests,

PASSOS ET AL.22 © 2005 Magnolia Press
1013ZOOTAXA respectively (Zar 1999). In cases where characters showed insufficient variation to justify
these assumptions we performed non-parametric tests such as Mann-Whitney andKruskal-Wallis (Zar 1999). We performed a principal component analysis (PCA) at grouplevel for males and females separately to evaluate differentiation between the groups with-out priori definition (Manly 2000). We projected the first two principal components ontoorthogonal axes and computed 95% confidence regions from the simulation of 1000 pseu-doreplicate data matrices obtained by parametric bootstrap. All principal-component load-ings are portrayed as vector correlations (directional cosines), which are estimated for eachcharacter by correlations with projection scores across individuals (Wright 1954; Strauss1985).
We also selected localities for which many specimens were available in order to inferthe relationship between geographical distribution and segmental counts. We chose ninelocalities for the analysis of females (Blumenau, Colatina, Corupá, Jaguará do Sul, Joen-ville, Massaranduba, Queimada Grande Island, São Francisco do Sul, and São Gabriel daPalha), although only six of these could be used for males (all the above except Joenville,Massaranduba, and São Gabriel da Palha). We computed Mahalanobis D distancesbetween each pair of localities and plotted them against the geographic distance (in km) inorder to indirectly assess whether discrimination patterns revealed by the PCA analysiswould fit an isolation-by-distance model (De Queiroz & Good 1997). If such a modelcould not be refuted, a clinal pattern of variation could be assumed. The test of Mantel forcorrelation between matrices (Manly 2000) was employed to estimate the correlationbetween geographical and morphological distances among localities within populations(De Queiroz & Good 1997; Reis et al. 2002).
We performed all computations using the software MATLAB 4.2c1 (MathWorks1994), except normality and homocedasticity tests, for which we used STATISTICA 5.0(Statsoft 1995).
Results
Dipsas albifrons shows significant sexual dimorphism in the number of ventral scales(F1,137 = 5.80; p < 0.02; N = 139), subcaudal scales (F1,121 = 32.40; p < 0.0001; N = 123),
and number of blotches over the tail (F1,108 = 21.38; p < 0.0001; N = 110), and therefore
these characters were treated separately for each sex in all subsequent analyses.Males from group 2 differed from those in group 3 in number of ventral scales (H =
4.41; p < 0.04; N = 11); and males from group 1 differed from those in 3 in number of sub-caudal scales (H = 4.73; p < 0.04; N = 42). Females in groups 1 and 2 differ in number ofventral scales (H = 8.80; p < 0.004; N = 75), and females from groups 1 and 3 differ inboth number of ventrals (H = 16.61; p < 0.0001; N = 67) and subcaudals (H = 9.81; p <0.002; N = 63). Similarly, females from groups 2 and 3 differ in both ventral (H = 21.04; p< 0.0001; N = 34) and subcaudal (H = 12.46; p < 0.001; N = 29) counts.

© 2005 Magnolia Press 23DIPSAS ALBIFRONS
1013ZOOTAXAThe projections of the bivariate plots based on scores of principal component analyses
(Figs. 2a–b) discriminate group 3 (State of Espírito Santo) from the remaining groups, allof which overlap. Discrimination occurred mainly along the first principal componentaxis, which was strongly correlated with the number of ventrals and subcaudal scales(Figs. 3a–b).
FIGURE 2. Bivariate plot with 95% confidence regions for the first 2 axes derived from scores ofPCA analysis of female (A) and male (B) populations of Dipsas albifrons. (C, D). Black dots =States of Paraná and Santa Catarina; black squares = Queimada Grande Island; and black triangles3 = State of Espírito Santo.

PASSOS ET AL.24 © 2005 Magnolia Press
1013ZOOTAXA
FIGURE 3. Correlations of meristic characters with corresponding principal components (direc-tional cosines) for females (A) and males (B) of Dipsas albifrons; 1 = number of ventral scales, 2 =number of subcaudal scales, 3 = number of supralabial scales, 4 = number of infralabial scales, 5 =number of blotches on the body, and 6 = number of blotches on the tail.
Bivariate plots of geography versus Mahalanobis D distances show a strong correla-tion for females (r = 0.83; p = 0.034), but not for males (r = 0.78; p = 0.153) (Figs. 4a–b).
Despite the suggestive evidence from the ANOVA and PCA analyses for a distinctpopulation in the State of Espírito Santo (group 3), the strong correlation between the geo-
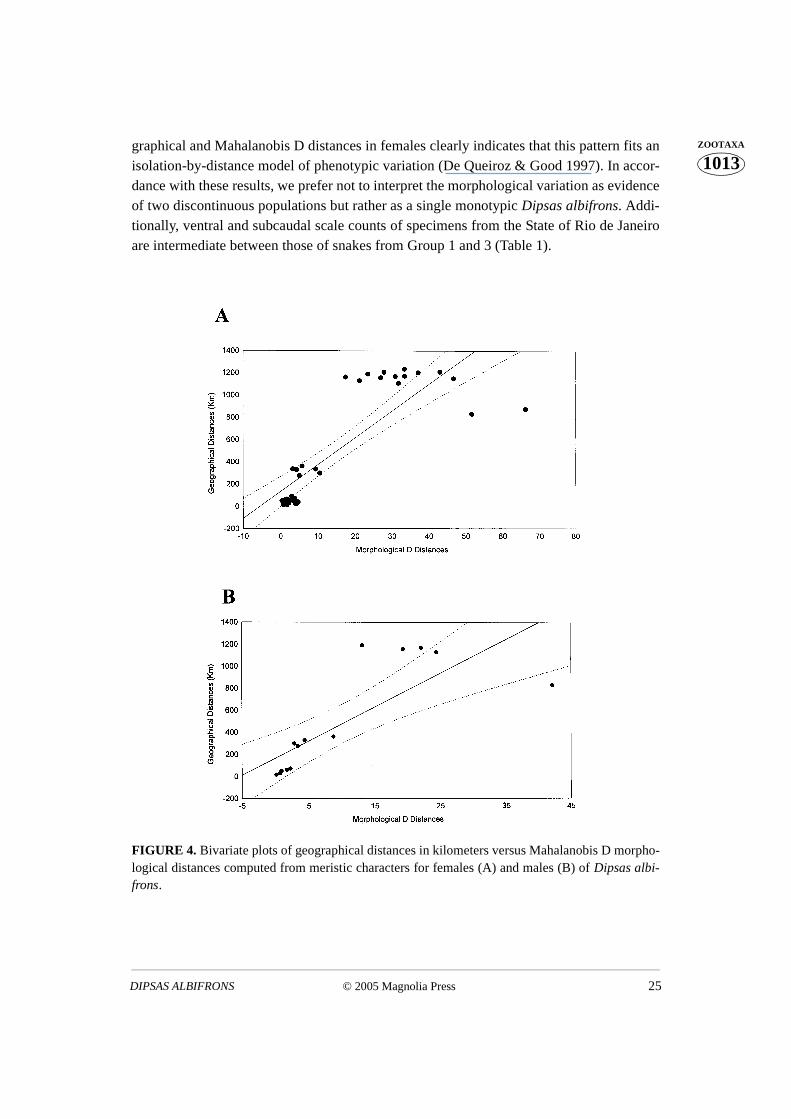
© 2005 Magnolia Press 25DIPSAS ALBIFRONS
1013ZOOTAXAgraphical and Mahalanobis D distances in females clearly indicates that this pattern fits an
isolation-by-distance model of phenotypic variation (De Queiroz & Good 1997). In accor-dance with these results, we prefer not to interpret the morphological variation as evidenceof two discontinuous populations but rather as a single monotypic Dipsas albifrons. Addi-tionally, ventral and subcaudal scale counts of specimens from the State of Rio de Janeiroare intermediate between those of snakes from Group 1 and 3 (Table 1).
FIGURE 4. Bivariate plots of geographical distances in kilometers versus Mahalanobis D morpho-logical distances computed from meristic characters for females (A) and males (B) of Dipsas albi-frons.

PASSOS ET AL.26 © 2005 Magnolia Press
1013ZOOTAXA TABLE 1. Descriptive statistics for groups of Dipsas albifrons. Values corresponding to average ±
standard deviations. Group 1 = Specimens from States of Paraná and Santa Catarina; Group 2 =Specimens from Queimada Grande Island; Group 3 = Specimens from State of Espírito Santo;Group 4 = Specimens from State of Rio de Janeiro.
Taxonomic Account
Dipsas albifrons (Sauvage, 1884) (Fig. 5)
Dipsadomorus albifrons Sauvage, 1884Leptognathus albifrons — Boulenger, 1896Dipsas albifrons — Mocquard, 1908Cochliophagus albifrons — Ihering, 1911Dipsas albifrons — Amaral, 1923Dipsas albifrons albifrons — Hoge, 1950Dipsas albifrons cavalheiroi Hoge, 1950Dipsas albifrons — Peters, 1960
Lectotype: Instituto Butantan IBSP 17746 (previously Muséum National d'HistoireNaturelle de Paris MNHN 6106), adult female, SVL 390 mm, TL 139 mm, collected byFerry from Brazil [designated herein].
Paralectotype: Muséum National d'Histoire Naturelle de Paris MNHN 6293, adultfemale, collected by Ferry from Brazil [designated herein].
Diagnosis: Distinguished from all congeners by the combination of 15 dorsal scalesrows; vertebral scale row not or slightly enlarged; maxillary teeth 11–15; pterigoyd short,toothless, not curved and lacking the quadrate process; hemipenis single; and dorsal pat-tern of irregular blotches extending less than half of length of interspaces.
Comparisons: Species groups of Dipsas were diagnosed manly by color pattern byPeters (1960). The Dipsas variegata group is distinguished from the other groups by a dor-sal ground color generally light with dark, narrow (usually higher than long), irregular, lat-eral blotches not as wide as interspaces over the greater part of the body. The interspaces in
Groups Sex Ventral Scales N Subcaudal Scales N
Group 1 & 168 ± 3.42 54 79.12 ± 3.10 50
% 171.9 ± 3.75 37 84.66 ± 4.50 33
Group 2 & 163.40 ± 5.10 21 77 ± 4.67 16
% 168 ± 7.30 6 84.20 ± 6.10 4
Group 3 & 184 ± 3.65 13 87.10 ± 5.92 13
% 182.80 ± 6.0 5 92 ± 3.90 4
Group 4 & 167.50 ± 2.12 2 81 ± 5.65 2
% 177 ± 6.36 2 93 1

© 2005 Magnolia Press 27DIPSAS ALBIFRONS
1013ZOOTAXAthis group are rarely unicolored and frequently marked with streaks, spots, and stippling
throughout the body.
FIGURE 5. Dorsal (A) and lateral (B) views of the head of Dipsas albifrons (Sauvage, 1884)
(IBSP 17746, lectotype). (Ruler equals to 5 mm).

PASSOS ET AL.28 © 2005 Magnolia Press
1013ZOOTAXA Dipsas albifrons can be distinguished from others species of the D. variegata group by
a suite of diagnostic features. It may be distinguished from D. andiana, D. incerta, D.neivai, D. nicholsi, and D. variegata by lacking pterigoydal teeth; from D. alternans, D.variegata, D. andina, and D. nicholsi by having a pair of parallel dark brown spots on thehead that are not connected anteriorly (instead of conspicuous white bordered dorsal ocelliin D. alternans, irregular blotches in D. variegata and a n-shaped black marking on D.andina and D. nicholsi); from D. neivai, D. andina, D. nicholsi, and D. variegata by hav-ing a single hemipenis (instead of a deeply bilobed in the first or slightly bilobed in theothers); from D. andiana, D. nicholsi, and D. incerta by a lower number of ventral andsubcaudal scales (see Cadle & Myers 2003 and Passos et al. 2004, respectively); from D.alternans and D. nicholsi by possessing a conspicuous series of small diamond shapeddots in the mid portion of interspaces.
Description of lectotype (Fig. 5): Head distinct from body, head length 15.3 mm(3.9% of SVL) from rostral scale to posterior tip of mandibles, head width 8.4 mm (55.3%of head length) at broadest point; interocular distance 4.9 mm; snout-orbit distance 2.7 mm(0.60 times interocular distance); rostral twice broader than high, visible from above;internasals and prefrontals slightly broader than long, the former about 0.75 times longerthan the latter; prefrontals not entering orbit; frontal as broad as long; parietals about 0.75times longer than wide; nasal divided on right side and semi-divided on left; one preocu-lar; loreal higher than long entering orbit; distance across eyes 3.25 mm; one postocular;temporal formula 1+2; seven supralabials, third to fifth entering orbit; symphysial broaderthan long; eleven infralabials on left and twelve infralabials on right side, two pairs in con-tact behind symphysial, second to fifth on left and third to fifth on right side contactingfirst pair of genials, fifth to seventh on left and fifth to eighth on right side contacting sec-ond pair of genials; three pairs of genials; 15 dorsal scales rows along entire body, dorsalscales smooth; vertebral row scarcely enlarged; seven dorsal scales rows on tail, at level ofsecond subcaudal; 177 ventrals; anal plate entire; 83/84 subcaudals. Thirteen maxillaryand six palatine teeth; pterygoid toothless. Coloration is mostly faded, blotches two scaleslong in anterior and posterior regions, and three scales long at midbody; interspaces aboutsix scales long along body.
Color in preservative: The color pattern of preserved specimens is compatible withthat of the lectotype. Dorsum of head uniformly light brown except for faded dot overfrontal and two parallel dark-brown spots over parietals, often diverging posteriorly;supralabials light-brown, almost creamish white along ventral edge; infralabials and gularregion uniformly creamish white; dorsal ground color of body beige; dorsum of body with17–32 ( = 23.04; SD = 2.37; N = 122) well-defined, nearly elliptical, dark-brown, whitebordered dorsal blotches; blotches smaller than interspaces along body, generally with 3/2/2 scales long; blotches eventually join in vertebral region and extend to paraventral area;interspaces generally six scales long; venter generally with anterior portion light-brown,posteriorly becoming brown under tail, irregular streaks arising on mid and/or posterior

© 2005 Magnolia Press 29DIPSAS ALBIFRONS
1013ZOOTAXAportion of venter to tail; streaks of different tones and size, sometimes forming irregular
dots between interspaces in paravertebral region; tail with 11–19 ( = 15.27; SD = 1.64; N= 39) blotches in males and 11–18 ( = 13.76; SD = 1.63; N = 71) in females, with samepattern as body.
Hemipenis (Fig. 6) [everted organ N = 7]: Organ single, cylindrical, with bulbousshape, slightly larger at apices; strongly capitate; capitulum completely encircles organand occupies more than distal half of hemipenial body on sulcate side; capitulum occupiesless than distal half of hemipenial body on asulcate side; capitulum ornamented with highnumber of papillate calyces, more concentrated on the sulcate side; sulcus spermaticusdivides approximately on most basal region of capitulum; branches in centrolineal orienta-tion terminating almost at distal region of capitulum; basal portion of hemipenial bodywith large spines (ca. 30) arranged in several rows and concentrated on portion adjacent tocapitulum; some spines on lateral region; both sides with spinules among large spines,despite being more concentrated on sulcate side.
FIGURE 6. Sulcate (A) and asulcate (B) sides of hemipenes of Dipsas albifrons. (A, from munici-
pality of Corupá (IBSP 64929), State of Santa Catarina, Brazil (Ruler equals to 5 mm).

PASSOS ET AL.30 © 2005 Magnolia Press
1013ZOOTAXA Hemipenis variation: The organ extends (in situ) to the level of the 5–10th ( = 8.07;
SD = 1.23; N = 34) subcaudal; the musculus retractor penis magnus originates betweenthe 20–26th subcaudal. In northern populations, the papillate calyces are less conspicuousin both sides.
Meristic variation: Largest male 559 mm SVL, 215 mm TL; largest female 690 mmSVL, 227 mm TL. Ventrals 154–191 ( = 169.88; SD = 7.46; N = 90) in females, 159–187 ( = 172.87; SD = 6.08; N = 49) in males; subcaudals 70–96 ( = 80.08; SD = 5.03;N = 81) in females, 68–97 ( = 85.62; SD = 5.26; N = 42) in males; preoculars 1 (N = 158sides), 2 (N = 4 sides) or none (N = 116 sides); supralabials 6–10 ( = 7.81; SD = 0.63; N= 139), supralabials touching the orbit 3–4 (N = 7), 3–5 (N = 23), 3–6 (N = 4), 4–5 (N =71), 4–6 (N = 27), 5–6 (N = 7); infralabials 8–13 ( = 10.85; SD = 0.71; N = 138), 1–4 (= 2.34; SD = 0.62; N = 136) pairs in contact behind symphysial and generally 3-6 touchingfirst pair of genials and 5-8 touching second pair of genials; nasal divided (N = 53) orsemidivided (N = 31); 1 postoculars (N = 13 sides), 2 postoculars (N = 164 sides); tempo-rals 1+1 (N = 3), 1+2 (N =116), 2+1 (N = 4), 2+2 (N = 13), 2+3 (N = 3); maxillary teeth11–15 ( = 13,71; SD = 0.82; N = 75); palatine teeth 6–9 ( = 7.38; SD = 0.76; N = 13);dentary teeth 20–25 ( = 22.63; SD = 1.56; N = 11).
Distribution (Fig. 1): Dipsas albifrons occurs in southeastern and southern Brazil,from the states of Espírito Santo to Santa Catarina with a large gap along the continentalportion of the State of São Paulo.
Discussion
Peters (1960) provided a comprehensive historical taxonomy of Dipsas albifrons andestablished that generally this species has no pterygoidal teeth, although he observed twospecimens with three pterygoidal teeth (specimens’ numbers not cited). However, we havefailed to observe any specimen of D. albifrons with pterygoidal teeth (N = 20), whereas thepresence of three teeth in the pterygoid is characteristic of the closely related species D.neivai and D. variegata (Fernandes 1995; Porto & Fernandes 1996). As already pointedout by Porto and Fernandes (1996), Peters (1960) misidentified two specimens of D.neivai from Museu Nacional, Brazil (MNRJ 708 and 714) as D. albifrons, which mayexplain his two specimens with pterygoid teeth.
A question that emerges from these results is whether two groups of populations iden-tified on the basis of variance and PCA analyses represent distinct geographic units withinDipsas albifrons. This question must be evaluated within the framework of geographicsampling of the populations (De Queiroz & Good 1997). The known distribution of D.albifrons includes a large gap along most of the continental portion of the State of SãoPaulo (Fig. 1). Considering that these states are the two most populated States in Braziland that the Instituto Butantan has received constant samples of São Paulo for more than ahundred years, it is likely that the disjunct distribution is not an artifact of poor sampling.

© 2005 Magnolia Press 31DIPSAS ALBIFRONS
1013ZOOTAXAInterestingly, the same pattern was found by Porto and Fernandes (1996) for D. neivai, and
suggests a natural event that caused regional extinctions, perhaps caused by retraction ofsea level in the Quaternary (Ab’Saber 1965; Bigarella 1965; Vanzolini and Ab’Saber1968; Vanzolini, 1973).
The groups north (group 3) and south (group 1) of the gap have a remarkable differ-ence in the numbers of ventral and subcaudal scales (Fig. 2a-b). However, we found a pos-itive significant relationship between morphological and geographic distances amongfemales (Fig. 4a). A similar, but not significant, relationship was found for males (Fig. 4b).We believe that the smaller sample of localities in the analysis of males provided less sta-tistical support, which, nevertheless, displayed a good correlation (r=0.78). Following DeQueiroz and Good (1997), we interpret the phenetic clusters in the plots (Figs. 2 and 4) asnot necessarily indicative of a pattern of two different species.
The first two principal component variables (strongly correlated with the number ofventral and subcaudal scales) may be correlated with climatic factors. It is well known(Blanchard & Blanchard 1940; Fox 1948; Fox & Fox 1961; Hoge et al. 1977) that in manysnakes, humidity and temperature is correlated with segmental counts. As such, the factthat snakes from the northern group has more segmental counts may be just a reflection ofthe higher temperatures at the localities from which those snakes were sampled.
Dipsas albifrons cavalheroi was originally defined (Hoge 1950) by low segmentalcounts and the shape of the dorsal scales. Our data corroborate Peters (1960) synonymiza-tion of this subspecies with the nominate race. Peters (1960) dismissed the shape of thedorsal scales as a character. We found that, although some D. albifrons do possess roundeddorsal scales, these specimens are from both continental and insular populations.
Acknowledgements
We are grateful to G. Puorto, I. Laporta-Ferreira, and F. L. Franco (IBSP), and P. E. Vanzo-lini and H. Zaher (MZUSP) for allowing us to exam specimens in their institutions; D. S.Fernandes, P. R. Gonçalves, J. P. Pombal Jr., and M. B. Harvey for critically reviewing themanuscript; G. Scrocchi and a anonymous referee for useful suggestions; P. R. Nascimentofor rendering the line arts; V. J. Germano for valuable help with IBSP collection and O. A.V. Marques for insightful discussions on the biology of insular snake populations. Theauthors thank the Conselho Nacional de Desenvolvimento Científico e Tecnológico(CNPq), and the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Riode Janeiro (FAPERJ) for financial support.
References
Ab’Saber, A.N. (1965) A evolução geomorfológica. In: Azevedo, H. (Ed) A baixada Santista.

PASSOS ET AL.32 © 2005 Magnolia Press
1013ZOOTAXA Aspectos geográficos, São Paulo, Brasil, pp 50–66.
Amaral, A. (1923) New genera and species of snakes. Proceedings of New England ZoologicalClub, 8, 85–105.
Amaral, A. (1925) Comunicações das linhas telegráficas estratégicas de Mato Grosso ao Amazo-nas. História Natural, Zoologia, São Paulo, 84 (Annexo 5), 1–29.
Amaral, A. (1930a) Contribuição ao conhecimento dos ophidios do Brasil IV. Lista remissiva dosophidios do Brasil. Memórias do Instituto Butantan, 4, 69–126 [dated 1929].
Amaral, A. (1930b) Estudo sobre ophidios neotropicos XVIII. Lista remissiva dos ophidios daregião Neotropica. Memórias do Instituto Butantan, 4, 127–271 [dated 1929].
Bigarella, J.L. (1965) Subsídios para o estudo das variações de nível oceânico no quaternáriobrasileiro. Anais da Academia Brasileira de Ciências, 37, 263–278.
Blanchard, F.N. & Blanchard, F.C. (1940) Factor determining time of birth in the garter snakeThamnophis sirtalis sirtalis (Linnaeus). Papers of Michigan Academic Science Arts and Let-ters, 26, 161–176.
Boulenger, G.A. (1896) Catalogue of the snakes in the British Museum (Natural History). Vol. 3.Trustees of British Museum (Natural History), London, 727 pp.
BRASIL (2003) Lista da fauna brasileira ameaçada de extinção. Ministério do Meio-ambiente,Brasília. Available from: http://www.mma.gov.br/port/sbf/fauna/index.cfm
Brongesma, L.D. (1958) Some features of Dipsadinae and Pareinae (Serpentes: Colubridae). Pro-ceedings of the Koninklijke Nederlandse Akadenie van Wetenschappen, 61, 7–12.
Cadle, J.E. & Myers, C.W. (2003) Systematic of snakes referred to Dipsas variegata in Panama andwestern South America, with revalidation of two species and notes on defensive behaviors inthe Dipsadini (Colubridae). American Museum Novitates, 3409, 1–47.
De Queiroz, K. & Good, D.A. (1997) Phenetic clustering in biology: A critique. The QuarterlyReview of Biology, 72, 3–30.
Dowling, H.G. (1951) A proposed standard system of counting ventrals in snakes. British Journalof Herpetology, 1, 97–99.
Dowling, H.G. & Savage, J.M. (1960) A guide to the snake hemipenis: A survey of basic structureand systematic characters. Zoologica: New York Zoological Society, 45, 17–28.
Fernandes, R. (1995) Phylogeny of Dipsadine Snakes. Arlington. Unpublished Ph. D. Dissertation,University of Texas, Arlington, Texas, U.S.A., 115 pp.
Fox, W. (1948) Effects of temperature on development of scutellation in the garter snake Thamno-phis elegans atratus. Copeia, 1948, 252–262.
Fox, W. & Fox, M.H. (1961) Morphological effects of low temperatures during the embryonicdevelopment of the garter snake, Thamnophis elegans. Zoologica: New York Zoological Soci-ety, 46, 57–71.
Hoge, A.R. (1950) Notas erpetológicas: Fauna erpetológica da Ilha da Queimada Grande.Memórias do Instituto Butantan, 22, 151–172.
Hoge, A.R., Belluomini, H.E. & Fernandes, W. (1977) Variação do número de placas ventrais deBothrops jararaca em função dos climas (Viperidae, Crotalinae). Memórias do Instituto Butan-tan, 41/42, 11–17.
Ihering, R.von (1911) As cobras do Brazil. Primeira Parte. Revista do Museu Paulista, 8, 273–379.Manly, B.F.J. (2000) Multivariate statistical methods: A primer, 2nd ed., Chapman & Hall Press,
New York, U.S.A., 215 pp.MathWorks, Inc. (1994) Matlab for windows 4.2c1. Natick: The MathWorks, Texas Tech Univer-
sity, U.S.A.Mocquard, F. (1908) Reptiles In: Duméril, A.M.C. & Bocourt, M.F. (Eds.), Mission scientific au
Mexique et dans l’Amérique Central, vol. 2, France, pp 863–898.Myers, C.W. & Campbell, J.A. (1981) A new genus and species of colubrid snake from Sierra
Madre del Sul of Guerrero, Mexico. Americam Museum Novitates, 2708, 1–20.

© 2005 Magnolia Press 33DIPSAS ALBIFRONS
1013ZOOTAXAPassos, P., Fernandes, D.S. & Caramaschi, U. (2004) The taxonomic status of Leptognathus incer-
tus Jan, 1863, with revalidation of Dipsas alternans (Fischer, 1885) (Serpentes: Colubridae:Dipsadinae). Amphibia-Reptilia, 25, 381–393.
Peters, J.A. (1960) The snakes of subfamily Dipsadinae. Miscellaneous Publications Museum ofZoology, University of Michigan, 114, 1–224.
Pesantes, O.S. (1994) A method for preparing the hemipenis of preserved snakes. Journal of Herpe-tology, 28, 93–95.
Porto, M. & Fernandes, R. (1996) Variation and natural history of the snail-eating snake Dipsasneivai (Colubridae: Xenodontinae). Journal of Herpetology, 30, 269–271.
Reis, S.F. dos, Duarte, L.C., Monteiro, L.R. & Von Zuben, F.J. (2002) Geographical variation incranial morphology in Thrichomys apereoides (Rodentia: Echimydae). II. Geographical units,morphological discontinuities, and sampling gaps. Journal of Mammalogy, 83, 345–353.
Sauvage, M.H.-E. (1884) Sur quelques Reptiles de la collection du Muséum d’Histoire Naturelle.Bulletain Societé Philomathic, 7, 142–147.
Statsoft, Inc (1995) Statistica for windows version 5.1. Tulsa, U.S.A.Strauss, R.E. (1985) Evolutionary allometry and variation in body form in the South American cat-
fish genus Corydoras (Callichthyidae). Systematic Zoology, 34, 381–396.Vanzolini, P.E. (1973) Distribution and differentiation of animals along the coast and in continental
island of the state of S. Paulo, Brasil. I. Introdution to the area and problem. Papéis Avulsos deZoologia, São Paulo, 26, 281–294.
Vanzolini, P.E. & Ab’Saber, A.N. (1968) Divergence rate in South American lizards of the genusLiolaemus (Sauria, Iguanidae). Papéis Avulsos de Zoologia, São Paulo, 21, 205–208.
Wright, S. (1954) The interpretation of multivariate systems. In: Kempthorne, O., Bancroft, T.A.,Gowen, J.W. & Lush, J.L. (Eds) Statistics and mathematics in biology, Iowa State CollegePress, U.S.A., pp 11–13.
Zar, J.H. (1999) Bioestatistical analysis, 4th ed., Prentice-Hall Press, New Jersey, U.S.A., 633 pp.

PASSOS ET AL.34 © 2005 Magnolia Press
1013ZOOTAXA Appendix Material examined
Specimens with an asterisk (*) are those for which hemipenes were examined.
Dipsas albifrons: BRAZIL: Unknown locality: IBSP 17746 (lectotype of Dipsadomorus albifrons,formerly MNHN 6106); ESPÍRITO SANTO: Afonso Claudio: IBSP 50462*, IBSP 50463; Castelo:IBSP 53543; Colatina, IBSP 22872, IBSP 34178, IBSP 37490*, IBSP 37496, 37521, IBSP 45915,IBSP 45938; Guandú: IBSP 50015; Santa Teresa: MNRJ 1265; São Gabriel da Palha: IBSP49194, IBSP 49656, IBSP 50070, IBSP 50658-59, IBSP 50661, IBSP 51197-98; Vila Velha: Morrodo Moreno: MNRJ 11027; RIO DE JANEIRO: Angra dos Reis: IBSP 27608; Cabo Frio: MNRJ7353; Maricá: MNRJ 7372; SÃO PAULO: Apiaí: IBSP 56275; Peruíbe: Ilha da Queimada Grande:IBSP 11486 (holotype of Dipsas albifrons cavalheiroi), IBSP 1638 (paratype of D. a. cavalheiroi),IBSP 3468 (paratype of D. a. cavalheiroi), IBSP 11487 (paratype of D. a. cavalheiroi), IBSP11488, IBSP 11489 (paratype of D. a. cavalheiroi), IBSP 15808-09, IBSP 17151, IBSP 17213,IBSP 18426-7, IBSP 29727, IBSP 30088-92, IBSP 30093*, IBSP 30094-95, IBSP 52670, IBSP55723; Santos: Ilha de Alcatrazes: IBSP 13029, IBSP 16972, IBSP 62175; São Sebastião: IlhaBela: IBSP 60345; PARANÁ: Antonina: IBSP 24952, IBSP 30236; Itaperuçu: IBSP 37553; Jus-sara: IBSP 49657*; Mallet: IBSP 18971; Palmeira: IBSP 11363; Paranaguá: IBSP 23025; PontaGrossa: IBSP 44124; SANTA CATARINA: Unknown locality: IBSP 27574; Bananal: IBSP 6510-12, IBSP 6870; Blumenau, IBSP 1639, IBSP 20954, IBSP 26777-78, IBSP 40360 IBSP 41411,IBSP 55089, IBSP 55718*, IBSP 55831, IBSP 55901, IBSP 55904, IBSP 55014, IBSP 55948*,IBSP 56212*, IBSP 57557, IBSP 67918-19; Campos: IBSP 14584-85; Corupá: IBSP 5077-78,IBSP 5546, IBSP 5557, IBSP 5572, IBSP 6227, IBSP 6721, IBSP 6994-7000, IBSP 10331, IBSP11840, IBSP 12069, IBSP 13225, IBSP 13575, IBSP 13587, IBSP 15610, IBSP 15755-7, IBSP15845-6, IBSP 17367, IBSP 17375, IBSP 20953, IBSP 22417-31, IBSP 22428-31, IBSP 22767,IBSP 33202, IBSP 55902, IBSP 56347, IBSP 64928; IBSP 64929*, IBSP 65015, MNRJ 709-10;Garúva: IBSP 31181; Indaial: IBSP 47325; Jaraguá do Sul: IBSP 1194-7, IBSP 1548, IBSP 1549-50, IBSP 1594-5, IBSP 1601, IBSP 1639, IBSP 4315, IBSP 4520, IBSP 4525, IBSP 4531, IBSP4575-6, IBSP 4600, IBSP 4609, IBSP 4650, IBSP 5308, IBSP 5816, IBSP 5859-60, IBSP 5924,IBSP 5989, IBSP 6056, IBSP 6066, IBSP 6069, IBSP 6098, IBSP 6109, IBSP 6109-A, IBSP 6338,IBSP 6370, IBSP 6376-78, IBSP 6495-97, IBSP 6510, IBSP 6721, IBSP 6869, IBSP 6870-73,IBSP 6899, IBSP 6911-14, IBSP 6963-64, IBSP 6980-83, IBSP 6992-93, IBSP 7040, IBSP 7064,IBSP 7079-81, IBSP 7112-16, IBSP 7283, IBSP 10072, IBSP 58679, IBSP 58739, IBSP 62058-59,IBSP 62401; Joinville: IBSP 22888, IBSP 22900, IBSP 25990, IBSP 29312, MNRJ 711-13, MNRJ715-16; Massaranduba: IBSP 5188-89, IBSP 5211-13, IBSP 5269, IBSP 5331-33, IBSP 5446,IBSP 6362, IBSP 6366-67, IBSP 6959, IBSP 6989; Rio Natal: IBSP 576, IBSP 13284, IBSP 15516;Pomerode: IBSP 64279; Porto Belo: IBSP 52448; Rio Natal: Estação: IBSP 5576, IBSP 13284,IBSP 15516; São Bento do Sul: IBSP 4832, MZUSP 9440; São Francisco do Sul: IBSP 602, IBSP602-A, IBSP 6238, IBSP 6253-54, IBSP 6258, IBSP 15471, IBSP 16400, IBSP 20953, IBSP21099, IBSP 27327, IBSP 29096, IBSP 40469, IBSP 55306.
Dipsas alternas, D. incerta, and D. variegata: See section Appendix in Passos et al. (2004).
Dipsas neivai: See section Material Examined in Porto & Fernandes (1996).




















