
Post Transaction Changes in 2014-15 – Playbook for CPs and ...

2000, 38(9):3420. J. Clin. Microbiol.
NorgrenMats Sellin, Carin Olofsson, Stellan Håkansson and Mari Serotypes III and VII
Allelic Variants ofcpsReveals Two Major) in Nontypeable Group B Streptococcicps
Genotyping of the Capsule Gene Cluster (
http://jcm.asm.org/content/38/9/3420Updated information and services can be found at:
These include:
REFERENCEShttp://jcm.asm.org/content/38/9/3420#ref-list-1at:
This article cites 36 articles, 25 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

JOURNAL OF CLINICAL MICROBIOLOGY,0095-1137/00/$04.0010
Sept. 2000, p. 3420–3428 Vol. 38, No. 9
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Genotyping of the Capsule Gene Cluster (cps) in NontypeableGroup B Streptococci Reveals Two Major cps Allelic
Variants of Serotypes III and VIIMATS SELLIN,1* CARIN OLOFSSON,1 STELLAN HÅKANSSON,2 AND MARI NORGREN1
Department of Clinical Bacteriology1 and Department of Pediatrics,2 Umeå University, S-901 85 Umeå, Sweden
Received 21 December 1999/Returned for modification 29 May 2000/Accepted 12 July 2000
Forty group B Streptococcus (GBS) isolates obtained from Europe and the United States previously reportedto be nontypeable (NT) by capsule serotype determination were subjected to buoyant density gradient centrif-ugation. From nearly half of the isolates capsule-expressing variants could be selected. For characterization ofthe remaining NT-GBS isolates, the capsule operon (cps) was amplified by the long-fragment PCR techniqueand compared by restriction fragment length polymorphism (RFLP) analysis. The patterns from serotypereference isolates (n 5 32) were first determined and used as a comparison matrix for the NT-GBS isolates.Using two restriction enzymes, SduI and AvaII, cluster analysis revealed a high degree of similarity withinserotypes but less than 88% similarity between serotypes. However, serotypes III and VII were each split in twodistant RFLP clusters, which were designated III1 and III2 and VII1 and VII2, respectively. Among the isolatesthat remained NT after repeated Percoll gradient selections, two insertional mutants were revealed. Both werefound in blood isolates and harbored insertion sequence (IS) elements within cpsD: one harbored IS1548, andthe other harbored IS861. All other NT-GBS isolates could, by cluster analysis, be referred to differentserotypes by comparison to the RFLP reference matrix. In pulsed-field gel electrophoresis of SmaI-restrictedchromosomal DNA, patterns from allelic type 1 and 2 isolates were essentially distributed in separate clustersin serotypes III and VII. A covariation with insertion sequence IS1548 in the hylB gene was suggested forserotype III, since allelic type III1 harboring IS1548 in hylB, clustered separately. The variation in serotype VIIwas not dependent on the presence of IS1548, which was not detected at any position in the type VIIchromosome.
Group B Streptococcus (GBS) is the most common cause ofneonatal bacteremia and meningitis but can also cause seriousinfectious disease among adults (3, 6, 28). However, commen-sal intestinal or genital colonization is probably the most com-mon manifestation of contact between GBS and humans (3, 5).Apart from pregnancy, several predisposing host factors im-portant for the pathogenesis of invasive GBS infection inadults have been proposed, such as advanced age and chronicdebilitating disease (6, 15). Individual bacterial virulence fac-tors have been identified, and the clonal dispersal of GBS withincreased virulence has been suggested as a factor in neonataldisease (12, 25, 29, 36).
Clinical isolates of GBS generally have a polysaccharidecapsule, which is the basis for the serotyping traditionally usedfor epidemiological purposes. Nine immunologically distinctserotypes have so far been described: serotypes Ia, Ib, and IIthrough VIII (16, 18, 19, 35, 37, 38). Research, e.g., for devel-oping vaccines, has focused on serotype III, since it has beenthe serotype most commonly isolated in invasive neonatal dis-ease (3). While type III together with serotypes Ia, Ib, and IIremains important, the most dramatic change in prevalenceduring the last decade has been the increase in type V amonginvasive isolates (4, 11, 14). Evidently, GBS serotype preva-lence fluctuates over time and also with geographical location.In Japan, the most common serotypes are VI and VIII, whilereports of those serotypes from the rest of the world are rare(22). Furthermore, the relative distribution of serotypes III
and V differs among isolates from laboratories representingdifferent areas of the United States (24).
The polysaccharide capsule is a major virulence factor inGBS, and consequently most invasive GBS isolates are type-able and hence encapsulated. However, nontypeable GBSs(NT-GBSs) are infrequently reported as a cause of invasiveinfection and may be an increasing problem in adult invasiveGBS infection (11). We have previously described an inverserelationship between the capsule thickness and the buoyantdensity (9). Clinical GBS isolates have often proved to beheterogeneous, and subpopulations with thick or thin capsulesmay arise in a phase shift-like manner (32). Reversible NT-GBS phase variants have previously been encountered in bloodisolates at our laboratory, where enrichment of low-densitysubpopulations with hypotonic Percoll gradient centrifugationbefore typing has been successful (33).
Apart from technical reasons, we hypothesize three causesfor a GBS isolate to be nontypeable: (i) the isolate is a revers-ible nonencapsulated phase variant, (ii) the isolate produces anuncharacterized polysaccharide for which antibodies not yetare available (i.e., a new serotype), and (iii) the isolate has aninsertion or a mutation in genes essential for capsule expres-sion. The aim of this study was to develop methods to classifyNT-GBSs and hopefully find the etiology explaining the loss ofcapsule expression. To that end, GBS isolates previously re-ported to be NT, gathered from well-known typing laboratoriesin Scandinavia and the United States, were analyzed to revealphase variants, supported by a newly devised method of re-striction fragment length polymorphism (RFLP) analysis. ThisRFLP method also revealed major subdivision of serotypes IIIand VII, so the study was extended to compare the impact of
* Corresponding author. Mailing address: Department of ClinicalBacteriology, Umeå University, S-901 85 Umeå, Sweden. Phone: 46 907851123. Fax: 46 90 7852225. E-mail: [email protected].
3420
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from
allelic variation on pulsed-field gel electrophoresis (PFGE)patterns for the two serotypes.
MATERIALS AND METHODS
Bacterial strains and culture conditions. Clinical GBS isolates from Sweden,Norway, Denmark, and the United States that were previously reported as NTwere collected (6, 10, 13, 21). Together with additional isolates from our ownlaboratory, a total of 40 isolates from adults and neonates (20 invasive and 20noninvasive isolates) were gathered (Table 1). Reference type strains for each ofserotypes I through VIII were kindly supplied by J. Motlova at the CzechNational Collection of Type Cultures (CNCTC) in Prague, Czechoslovakia. GBSblood isolates from our laboratory were added to make up three isolates of eachof serotypes Ia, Ib, II, IV, V, VI, VII, and VIII. For serotype III, a total of sixisolates were investigated; for serotype VII, five isolates were investigated (Table2). Bacteria were plated on blood agar plates (Columbia II agar base [BBL,Cockeysville, Md.] supplemented with 5% horse blood) or in Todd-Hewitt broth(Difco, Detroit, Mich.) and incubated overnight at 37°C.
Serotyping. Serotyping was performed by coagglutination as previously de-scribed, with a kit for serotypes I through VIII (10).
Hypotonic Percoll gradient centrifugation. Gradient centrifugation was usedto estimate the buoyant density of GBS isolates and to select populations oflower buoyant density, as previously described (32). If an NT-GBS isolate turnedreactive in serotyping assays, it was considered a “reverting” NT-GBS isolate. Ifan isolate remained NT during three cultivation and centrifugation cycles, it wasconsidered a “nonreverting” NT-GBS isolate.
Sialic acid analysis. All GBS capsule polysaccharides known so far containsialic acid; therefore, sialic acid analysis can be used for interstrain comparisonsof capsule polysaccharide levels. The pellet from a 15-ml culture of GBS (opticaldensity at 500 nm 5 1.0) was used to extract sialic acid by HCl hydrolysis (0.1 NHCl at 84°C for 20 min). Analysis was performed with N-acetylneuraminic acid(Sigma Chemical Co., St. Louis, Mo.) as the standard, with the thiobarbituricacid method modified from that of Skoza and Mohos (34). Protein analysis wasperformed with the hydrolysate by the bicinchoninic acid method (Pierce, BouleAB, Huddinge, Sweden). All analyses were made in duplicate. To comparemeans, Student’s t test was used.
Preparation of chromosomal DNA. GBS isolates were grown overnight at 37°Cin 15 ml of Todd-Hewitt yeast broth supplemented with 20 mM threonine.Bacteria were washed once and resuspended in 5 ml of 0.02 M Tris-HCl (pH 6.8)with 1 M NaCl and 0.05% Triton X-100. One hundred microliters of mutanolysin
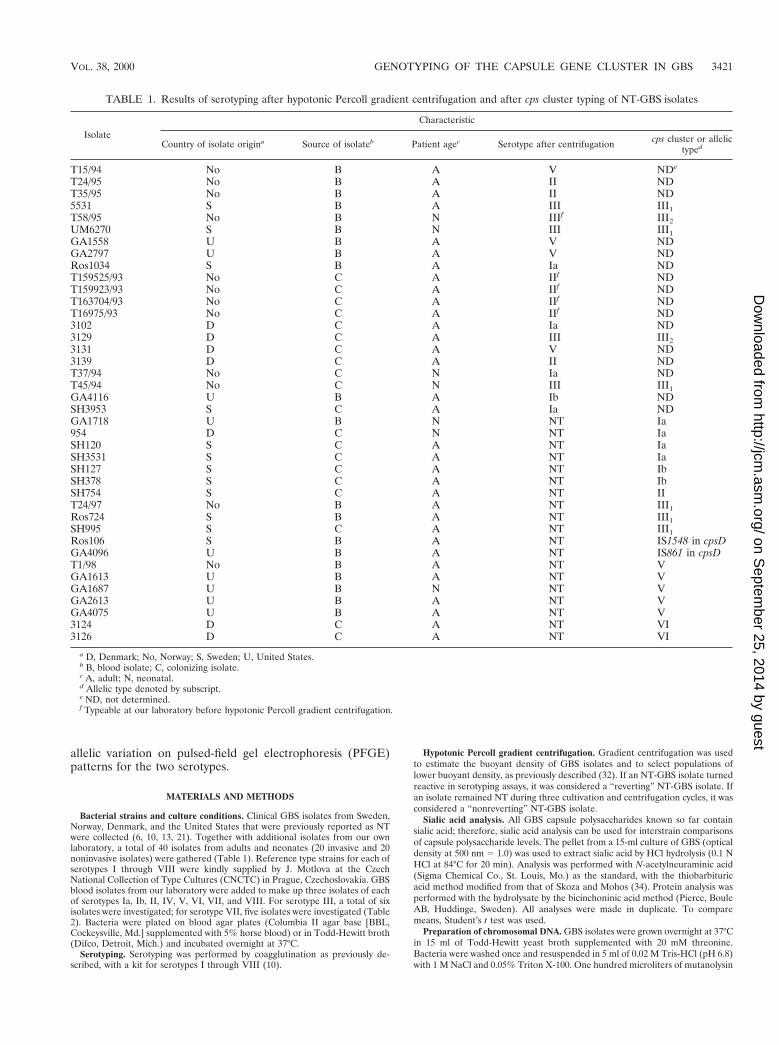
TABLE 1. Results of serotyping after hypotonic Percoll gradient centrifugation and after cps cluster typing of NT-GBS isolates
Isolate
Characteristic
Country of isolate origina Source of isolateb Patient agec Serotype after centrifugation cps cluster or allelictyped
T15/94 No B A V NDe
T24/95 No B A II NDT35/95 No B A II ND5531 S B A III III1T58/95 No B N IIIf III2UM6270 S B N III III1GA1558 U B A V NDGA2797 U B A V NDRos1034 S B A Ia NDT159525/93 No C A IIf NDT159923/93 No C A IIf NDT163704/93 No C A IIf NDT16975/93 No C A IIf ND3102 D C A Ia ND3129 D C A III III23131 D C A V ND3139 D C A II NDT37/94 No C N Ia NDT45/94 No C N III III1GA4116 U B A Ib NDSH3953 S C A Ia NDGA1718 U B N NT Ia954 D C N NT IaSH120 S C A NT IaSH3531 S C A NT IaSH127 S C A NT IbSH378 S C A NT IbSH754 S C A NT IIT24/97 No B A NT III1Ros724 S B A NT III1SH995 S C A NT III1Ros106 S B A NT IS1548 in cpsDGA4096 U B A NT IS861 in cpsDT1/98 No B A NT VGA1613 U B A NT VGA1687 U B N NT VGA2613 U B A NT VGA4075 U B A NT V3124 D C A NT VI3126 D C A NT VI
a D, Denmark; No, Norway; S, Sweden; U, United States.b B, blood isolate; C, colonizing isolate.c A, adult; N, neonatal.d Allelic type denoted by subscript.e ND, not determined.f Typeable at our laboratory before hypotonic Percoll gradient centrifugation.
VOL. 38, 2000 GENOTYPING OF THE CAPSULE GENE CLUSTER IN GBS 3421
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

(5 IU/ml) (Sigma) was added, and the sample was incubated overnight at 4°C.Two sequential incubations at 37°C for 15 min each were done, with 100 ml ofRNase (10 mg/ml) and 100 ml of proteinase K (10 mg/ml), respectively. Threesubsequent extraction steps were performed with volumes equal to the buffer-DNA solution: (i) with redistilled phenol and chloroform in a 50/50 mixture, (ii)with chloroform, and (iii) with isobutanol. DNA was precipitated by the additionof a 1/10 volume of 3 M sodium acetate (pH 4.8) and 2 volumes of cold 95%ethanol and incubation for 30 min at 270°C, followed by centrifugation at10,000 3 g for 15 min. The precipitated DNA was washed twice in cold 70%ethanol, dried, and then resuspended in 0.5 ml of 10 mM Tris-HCl (pH 8.0) with1 mM EDTA.
Capsule gene cluster RFLP analysis. A total of 19 nonreverting and 32 type-able GBS isolates were investigated (3 for each serotype or allelic type except forallelic type VII2, where only 2 isolates were retrieved). Long-fragment PCRswere set up with the Boehringer Mannheim Expand Long Fragment PCR kit(Roche Diagnostics Scandinavia AB, Bromma, Sweden) with 3 to 400 mM ofeach primer, lorfXfo (59-CAAGGAGGCGATAACGATAG AGGTAGAAATCAAG-39) and loEFrev (59-TTTGACCCTG ATCGCGCAGGAATAATAC-39)and 350 mM dinucleoside triphosphates in buffer 1 with a final concentration of1.75 mM MgCl2 with 2.6 U of enzyme mixture in a reaction mixture volume of50 ml. As a template, 2 to 5 ml of chromosomal DNA prepared as above was used.Thermocycling was performed on a PTC-200 thermocycler (MJ Research, Falk-enberg, Sweden) at 92°C for 2 min for 1 cycle, followed by 92°C for 20 s, 65°C for30 s, and 68°C for 6 min for 10 cycles, followed by 92°C for 10 s, 65°C for 30 s,and 68°C for 6 min extended by 20 s for each cycle for a total of 20 cycles,followed by 68°C for 7 min. Separate digestions with AvaII and SduI (MBI;Tamrolab AB, Molndal, Sweden) of the long-fragment PCR amplicons wereperformed, and fragments were separated on 0.8% agarose gels. Molecularweight standard IV (Roche) was included at multiple positions on the gel toallow optimal normalization between runs in the subsequent conversion to com-puter format prior to cluster analysis (see Fig. 2).
PFGE. The serotypes displaying allelic variation in the cps cluster RFLPanalysis, namely serotypes III and VII, were analyzed by PFGE. In all, 14 type IIIsensu lato (s.l.), reference, reverting NT-GBS, and cluster type III strains wereincluded in a PFGE comparison. The seven serotype VII strains in the study wereanalyzed on a separate PFGE gel.
Chromosomal DNA from GBS isolates embedded in agarose plugs was pre-pared as previously described (7, 8). An agarose slice was incubated overnight at
30°C with 20 U of SmaI (MBI) in the enzyme buffer provided. Plugs were washedin Tris-EDTA buffer for 1 h at 5°C before being mounted into wells of a 1%Pulsed Field Certified Agarose gel (Bio-Rad Laboratories AB, Sundbyberg,Sweden) in 0.53 Tris-borate-EDTA buffer (pH 8.3). Electrophoresis was per-formed on an automated PFGE apparatus, the Gene Path strain typing system(Bio-Rad). The standard program for fragment sizes of 50 to 600 kb was used. Astandard l ladder (New England Biolabs, Inc., Beverly, Mass.) was includedalongside the samples. The agarose gel was stained with 0.2% ethidium bromide,washed in tap water, visualized, and photographed under UV light.
Cluster analysis. Restriction profile photographs were analyzed with GelCom-par software, version 4.0 (Applied Maths, Kortrijk, Belgium). The scans of theAvaII and SduI restriction gels were combined into one RFLP pattern for eachisolate. AvaII bands of less than 0.7 kb were often faint and were not included inthe clustering. The unweighted pair-group method with arithmetic averages(UPGMA) clustering method was used, using the Dice coefficient with a bandposition tolerance of 1 to 1.2%. The electrophoresis patterns of both the capsulegene cluster RFLP and the PFGE of chromosomal DNA were analyzed with thesame program settings.
Sequencing technique. Direct PCR product sequencing was performed withABI PRISM products (Perkin-Elmer Applied Biosystems, Stockholm, Sweden)as previously described (20).
PCR mapping. The area immediately upstream of cpsA and cpsA-D wasmapped with PCR primers, using previously published amplification protocols(20). All serotype III and VII isolates were explored for the insertion of IS1548in the hylB gene by PCR analysis as previously described (8).
RESULTS
Serotyping before and after hypotonic Percoll gradient cen-trifugation. Without pretreatment, five of the isolates previ-ously classified as NT-GBS isolates could be typed by coagglu-tination at our laboratory. After the isolation of phase variantsby gradient centrifugation, the serotypes of 16 additional NT-GBS isolates could be determined (Table 1). A total of 19 of 35NT-GBS isolates were still nontypeable after hypotonic Percollgradient centrifugation.
Sialic acid and buoyant density estimations. No NT-GBSisolate with low buoyant density and high levels of sialic acid,suggestive of a new serotype, was found. The nonrevertingisolates had significantly (P , 0.05) lower sialic acid levels thanreverting isolates, with a mean of 0.7 (standard deviation [SD,0.3]) to 1.8 (SD, 1.9) mg/mg of protein, respectively. The fivetypeable NT-GBS isolates did not differ significantly in sialicacid content compared to the reference isolates, with a meanof 3.6 (SD, 1.3) to 2.9 (SD, 1.1) mg/mg of protein. The refer-ence isolates displayed significantly higher amounts of sialicacid than both reverting and nonreverting NT-GBS isolates(P , 0.001).
Capsule locus-targeted RFLP. The cps locus-specific RFLPmethod was devised not only as an alternative typing methodbut also to evaluate the gross structure of the capsule genecluster. Long-fragment PCR generated an approximately 13.3-kb-long amplicon spanning cpsX to cpsE (Fig. 1). By visualinspection of the AvaII restrictions gels, a distinction betweenserotypes started to appear, but the addition of a parallel SduIrestriction gel is needed to allow classification in relation toserotype (Fig. 2). In order to use cluster analysis of the gels fortyping purposes, a suitable cutoff value had to be established,i.e., a similarity index above which similar organization of thecapsule gene locus is suggested. When only reference isolateswere included in the dendrogram, comparison of the combinedAvaII and SduI restriction patterns showed the highest per-centage of similarity between two separate serotypes, 88%,between serotypes V and VI (data not shown). All other se-rotypes differed more with similarities of 60 to 80%; therefore,88% was set as the cutoff value in the comparison when NT-GBS isolates were added to the dendrogram (Fig. 3). All threereference serotype isolates clustered with 100% similarity withthe exception of serotype Ib, where a minor diversity (within90% similarity) was displayed. However, serotypes III and VII
TABLE 2. Type reference strains
Serotype and allelic typea Strain designation
Ia.................................................................CNCTC index 58/59Ia.................................................................B88-4568Ia.................................................................B91-5963Ib.................................................................CNCTC 59/59Ib.................................................................B89-1803Ib.................................................................B95-4854II .................................................................CNCTC 60/59II .................................................................B88-543II .................................................................B93-6268III1 ..............................................................CNCTC 4/70III1 ..............................................................B4877III1 ..............................................................B90-261III2 ..............................................................B88-1975III2 ..............................................................M732III2 ..............................................................B90-1259IV................................................................CNCTC 1/82IV................................................................B95-2516IV................................................................CNCTC B114877V .................................................................CNCTC 10/84V .................................................................B96-2781V .................................................................B97-360VI................................................................CNCTC 2/86VI................................................................J-B114862VI................................................................J-B114852VII1.............................................................CNCTC 4/89VII1.............................................................S2-03104VII1.............................................................B132157/rVII2.............................................................B123348VII2.............................................................B124055VIII.............................................................CNCTC 1/92VIII.............................................................130609VIII.............................................................CNCTC 130669
a Index denotes allelic type, determined as a result of this study.
3422 SELLIN ET AL. J. CLIN. MICROBIOL.
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

were each split in two distant clusters with a similarity ofapproximately only 60%. When the RFLP of nonrevertingNT-GBS isolates was added to the a dendrogram, four maindivisions became apparent. The first division contains sero-types Ia, III2, and VII2; the second division contains serotypesV, VI, and IV; the third division contains serotypes Ib, VIII,and III1; and the fourth division contains serotypes II and VII1(Fig. 3). Of the individual isolates, 17 of 19 clustered with.88% similarity to a reference serotype. In fact, 9 of 19 clus-tered with 100% similarity, 6 more isolates displayed $90%similarity, and 2 isolates clustered close to the cutoff at 89%(Fig. 3). The ascribed cluster types for NT-GBS isolates ac-
cording to the closest clustering reference serotype in the den-drogram are compiled in Table 1.
PFGE analysis. Visual inspection of the type III s.l. PFGEgels secludes one group of III1 isolates that form more scat-tered patterns with several weak partially digested bands com-plicating comparison and transformation to computer formatpreceding cluster analysis (Fig. 4A). In this process, visualguidance and careful scrutiny of negatives or gels are of par-amount importance. The PFGE patterns were reproducible,resisting variation in the digestion protocol. The dendrogramanalysis matched the visual impression in that three main clus-ters could be seen: one cluster contained allelic type III2 iso-
FIG. 1. Open reading frames in the GBS capsule gene cluster and positions for long-fragment PCR primers. Positions are based on two entries in the GenBank:for serotype Ia, accession no. AB028896 (39), and for serotype III, accession no. AF163833. Restriction sites for the enzymes used in the RFLP, investigation, AvaIIand SduI, are denoted above the genes.
FIG. 2. RFLP analyses of long-fragment PCR amplicons from the cps capsule gene cluster from two reference type strains each for every GBS serotype. Allelicvariants for serotypes III and VII are designated 1 and 2. M, molecular weight standard in kilobases.
VOL. 38, 2000 GENOTYPING OF THE CAPSULE GENE CLUSTER IN GBS 3423
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

lates only, one cluster contained the scattered allelic type III1pattern that was PCR positive for IS1548 in hylB, and a middlecluster contained IS1548-negative allelic type III1 isolates andone allelic type III2 isolate (Fig. 4B).
For the serotype VII isolates, the dendrogram analysis of the
gel picture resulted in a split of the two allelic variants intodifferent PFGE clusters, which both lacked IS1548 (Fig. 5).
PCR mapping. In search of insertions or gross rearrange-ments, nonreverting NT-GBS isolates were mapped using PCRprimers for sequences from cpsY to cpsD (20). Two of 19
FIG. 3. Gel-dendrogram diagram of cps long-fragment PCR amplicon RFLP from 19 NT-GBS and 32 typeable GBS isolates. Each isolate is represented by onelane where the schematic band profile is combined from separate digestions with AvaII (left) and SduI (right). The horizontal bar (upper left) represents the percentsimilarity coefficient. Clustering was performed using the UPGMA method.
3424 SELLIN ET AL. J. CLIN. MICROBIOL.
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

nonreverting NT-GBS isolates, Ros106 and GA4096, gener-ated approximately 1.5-kb-larger amplicons with primers forcpsA and cpsD then the other isolates. Nucleotide sequencingof the amplicon flanks revealed the insertion of IS1548 (8) inRos106 and the insertion of IS861 (30) in GA4096 into cpsD(Fig. 6).
DISCUSSION
Following the classical description in the 1930s by R. Lance-field of GBS serotyping based on immunoprecipitation in gel(23), many additional serotyping techniques have been de-vised, such as immunodiffusion, coagglutination, or enzymeimmunoassays (1, 10, 17, 23). The theoretical detection levelsof these techniques differ by 10- to 100-fold (26). Furthermore,different extraction procedures for the antigenic preparations
and differences in antisera may also influence the sensitivity ofthe test and thus the rate of NT-GBS isolates found (27). Inthis study, more than 10% of GBS isolates previously reportedas NT could be ascribed to a serotype merely by applying analternative method of analysis. The technical reasons discussedabove may have contributed to the initial nonreactivity in thosecases. In roughly 50% of the NT-GBS isolates gathered for thisstudy, a shift to lower buoyant density after hypotonic Percollgradient centrifugation accompanied a positive capsule-sero-typing reaction (Table 2). It seems that the primary NT reac-tion may be due to the fact that the NT-GBSs were isolated ina reversible non- or thin-encapsulated phase. All the majorserotypes could be detected among the reverting isolates,which implies that the phase variation is a common phenom-enon in GBSs regardless of serotype. A variable level of sero-
FIG. 4. Analysis of SmaI-digested chromosomal DNA from GBS serotype III s.l. NT/PV III denotes NT-GBS where phase variant serotype III was found aftergradient centrifugation; NT/CT III, nonreverting isolates with cluster type III. III1 and III2, allelic types III1 and III2, respectively, according to cps typing. (A) PFGEgel. M, molecular weight standards in kilobases. (B) Cluster analysis and presence of IS1548 in the hylB gene. The horizontal bar (upper left) represents the percentsimilarity coefficient. Clustering was performed using UPGMA method.
VOL. 38, 2000 GENOTYPING OF THE CAPSULE GENE CLUSTER IN GBS 3425
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

type antigen expression has previously also been reported byPalacios et al., where low levels of serotype polysaccharidehave been found in subsets of NT-GBS isolates (27).
The RFLP method described here allowed 17 of 19 nonre-verting isolates to be assigned a cluster type in relation toserotype reference strains. Thus, a genotyping method com-patible with the prevailing serotyping system is proposed. Thedescribed genotyping system still needs preparation of chro-mosomal DNA as a template, and DNA-primer relations forsome of the isolates need to be individually optimized to obtaina long-fragment PCR amplicon. The major advantages are thatgenotyping seems to work where other methods so far havefailed with NT-GBS and that allelic variants of type III and VIImay be diagnosed. Furthermore, the suggested relations be-tween serotypes within the divisions may be interesting from anevolutionary point of view. Reports on an evolutionary split forserotype III have previously been made based on populationgenetic approaches (12, 25). The split of the low-prevalenceserotype VII into two distinct lineages has to our knowledgenot been published before. If allelic variation in the cps locushas any functional implications at all, it remains to be shown;but nevertheless, it has the potential to be a genotypic markerfor epidemiological investigations of GBS serotype III and VIIinfections.
A close similarity of RFLP patterns in NT-GBS isolates andtypeable reference strains suggests a similar gross organizationof the capsule gene clusters. Although the decreased similarityindex seen for NT-GBS isolates in the cps dendrogram ascompared to reference isolates (Fig. 3) may conceal causes of
defective capsule expression, no definite explanation is offered.However, in two of the nonreverting isolates a rationale wasproposed, since insertion sequences (ISs) were identifiedwithin the capsule gene cluster (Fig. 6). Both insertions af-fected cpsD, and loss of the corresponding product, galactosyltransferase, has previously been demonstrated to be detrimen-tal to capsule expression (31). That the search for IS elementsis relevant when loss of gene function occurs in wild-type GBSisolates was also exemplified in a previous report by Granlundet al., where inactivation of the hyaluronidase gene by theinsertion of IS1548 was described (8). The target sequences forthe IS elements in our report differ from those previouslypublished, indicating that variation in the target site is allowed(Fig. 6). Furthermore, short target sequences (as in IS861)increase the probability for multiple insertions in the GBSgenome with possible interference with other genes. Up toeight copies of IS861 and IS1548 have previously been re-ported in serotype III GBS isolates (8, 32), and the overallimpact of insertion elements on the phenotype, genome size,and PFGE patterns could consequently be substantial. Al-though the two isolates which harbored insertion elements,Ros106 and GA4096, had a similarity index below the chosencutoff value for assigning a cluster type, the closest clusteringreference (serotype II) still seems possible. These insertionelements add one or two restriction sites for the enzymes used,which may explain the deviation. No candidate with a putativenew serotype was discovered among the NT-GBS isolates. Thiswould have been suggested if a low-density isolate was found
FIG. 5. Analysis of SmaI-digested chromosomal DNA from GBS serotypeVII reference strains. VII1 and VII2, allelic variants. (A) PFGE gel. M, molecularweight standards in kilobases. (B) Cluster analysis and presence of IS1548 in thehylB gene.
3426 SELLIN ET AL. J. CLIN. MICROBIOL.
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

with a high level of sialic acid and low similarity to referencestrains in the cluster typing.
DNA-based typing methods are important tools in the epi-demiology of infectious diseases. Due to high discriminatorypower and reproducibility, PFGE has been established as themethod of choice for tracking common-source outbreaks ofinfection (2). Some correspondence to serotype can be seen incluster analysis following PFGE of GBS genomic DNA (4).Since the cps cluster RFLP described here was designed to becongruent to the existing serotyping scheme, it allows an in-creased resolution, compared to PFGE, for ascribing a type toNT-GBS isolates. The combination of locus-based andgenomic DNA typing methods, together with accessory PCRand phenotyping methods, may be needed to cover all aspectsof variability that GBS can display.
ACKNOWLEDGMENTS
A. Kvam, R. Helmig, M. Farley, and I. Julander are gratefullyacknowledged for supplying nontypeable GBS isolates. Reference typestrains were kindly supplied by J. Motlova at the CNTCC in Prague,Czechoslovakia. M. Wagner kindly supplied type reference strains aswell as anti-type VIII antiserum and P. Cleary kindly supplied typereference strains.
This work was supported by grants from the Swedish Medical Re-search Council (08675), The Wiberg Foundation, The Swedish Medi-cal Society, The Sven Jerring Foundation, and The Umeå UniversityInsamlings Fonden to M.N.
ADDENDUM IN PROOF
After completion of this report, the gene designations in theserotype III cps cluster (GenBank accession no. AF163833)were revised by Chaffin et al. (D. O. Chaffin, S. B. Beres, H. H.Yim, and C. E. Rubens, J. Bacteriol. 182:4466–4477, 2000) andare now congruent to that of serotype Ia (39).
REFERENCES
1. Arakere, G., A. E. Flores, P. Ferrieri, and C. E. Frasch. 1999. Inhibitionenzyme-linked immunosorbent assay for serotyping of group B streptococcal
isolates. J. Clin. Microbiol. 37:2564–2567.2. Arbeit, R. D. 1999. Laboratory procedures for the epidemiologic analysis of
microorganisms, p. 116–137. In P. R. Murray, E. J. Baron, M. A. Pfaller,F. C. Tenover, and R. H. Yolken (ed.), Manual of clinical microbiology, 7thed. American Society for Microbiology, Washington, D.C.
3. Baker, C. J., and M. S. Edwards. 1995. Group B streptococcal infections, p.980–1028. In J. S. Remington and J. S. Klein (ed.), Infectious diseases of thefetus and newborn infant. 4th ed. W. B. Saunders Co., Philadelphia, Pa.
4. Blumberg, H. M., D. S. Stephens, M. Modansky, M. Erwin, J. Elliot, R. R.Facklam, A. Schuchat, W. Baughman, and M. M. Farley. 1996. Invasivegroup B streptococcal disease: the emergence of serotype V. J. Infect. Dis.173:365–373.
5. Dillon, H. C., Jr., E. Gray, M. A. Pass, and B. M. Gray. 1982. Anorectal andvaginal carriage of group B streptococci during pregnancy. J. Infect. Dis.145:794–799.
6. Farley, M. M., R. C. Harvey, T. Stull, J. D. Smith, A. Schuchat, J. D. Wenger,and D. S. Stephens. 1993. A population-based assessment of invasive diseasedue to group B Streptococcus in nonpregnant adults. N. Engl. J. Med. 328:1807–1811.
7. Fasola, E., C. Livdahl, and P. Ferrieri. 1993. Molecular analysis of multipleisolates of the major serotypes of group B streptococci. J. Clin. Microbiol.31:2616–2620.
8. Granlund, M., L. Oberg, M. Sellin, and M. Norgren. 1998. Identification ofa novel insertion element, IS1548, in group B streptococci, predominantly instrains causing endocarditis. J. Infect. Dis. 177:967–976.
9. Håkansson, S., M. Granlund-Edstedt, M. Sellin, and S. E. Holm. 1990.Demonstration and characterization of buoyant density subpopulations ofgroup B Streptococcus type III. J. Infect. Dis. 161:741–746.
10. Håkansson, S., L. G. Burman, J. Henrichsen, and S. E. Holm. 1992. Novelcoagglutination method for serotyping group B streptococci. J. Clin. Micro-biol. 30:3268–3269.
11. Harrison, L. H., J. A. Elliott, D. M. Dwyer, J. P. Libonati, P. Ferrieri, L.Billmann, and A. Schuchat. 1998. Serotype distribution of invasive group Bstreptococcal isolates in Maryland: implications for vaccine formulation.Maryland Emerging Infections Program. J. Infect. Dis. 177:998–1002.
12. Hauge, M., C. Jespersgaard, K. Poulsen, and M. Kilian. 1996. Populationstructure of Streptococcus agalactiae reveals an association between specificevolutionary lineages and putative virulence factors but not disease. Infect.Immun. 64:919–925.
13. Helmig, R., N. Uldbjerg, J. Boris, and M. Kilian. 1993. Clonal analysis ofStreptococcus agalactiae isolated from infants with neonatal sepsis or men-ingitis and their mothers and from healthy pregnant women. J. Infect. Dis.168:904–909.
14. Hickman, M. E., M. A. Rench, P. Ferrieri, and C. J. Baker. 1999. Changingepidemiology of group B streptococcal colonization. Pediatrics 104:203–209.
15. Jackson, L. A., R. Hilsdon, M. M. Farley, L. H. Harrison, A. L. Reingold,B. D. Plikaytis, J. D. Wenger, and A. Schuchat. 1995. Risk factors for group
FIG. 6. Schematic diagram of IS elements in the capsule gene cluster in twoNT-GBS blood isolates. (A) Isolate GA4096 harboring IS861 in cpsD. (B) IsolateRos106 harboring IS1548 in cpsD. The sequences for the direct repeats (DR) attarget sites are shown in boxes. The inverted repeat (IR) sequences for thepresent isolates are noted in open boxes and were identical with sequencespreviously reported for IS861 (accession no. M22449) (30) and IS1548 (accessionno. Y14270) (8). Arrows show directions of transcription for IS elements.
VOL. 38, 2000 GENOTYPING OF THE CAPSULE GENE CLUSTER IN GBS 3427
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

B streptococcal disease in adults. Ann. Intern. Med. 123:415–420.16. Jennings, H. J. 1990. Capsular polysaccharides as vaccine candidates. Curr.
Top. Microbiol. Immunol. 150:97–127.17. Jensen, N. E. 1979. Production and evaluation of antisera for serological type
determination of group-B streptococci by double diffusion in agarose gel.Acta Pathol. Microbiol. Scand. B 87:77–83.
18. Kogan, G., D. Uhrin, J. R. Brisson, L. C. Paoletti, A. E. Blodgett, D. L.Kasper, and H. J. Jennings. 1996. Structural and immunochemical charac-terization of the type VIII group B Streptococcus capsular polysaccharide.J. Biol. Chem. 271:8786–8790.
19. Kogan, G., J. R. Brisson, D. L. Kasper, C. von Hunolstein, G. Orefici, andH. J. Jennings. 1995. Structural elucidation of the novel type VII group BStreptococcus capsular polysaccharide by high resolution NMR spectroscopy.Carbohydr. Res. 277:1–9.
20. Koskiniemi, S., M. Sellin, and M. Norgren. 1998. Identification of two genes,cpsX and cpsY, with putative regulatory function on capsule expression ingroup B streptococci. FEMS Immunol. Med. Microbiol. 21:159–168.
21. Kvam, A. I., A. Efstratiou, L. Bevanger, B. D. Cookson, I. F. Marticorena,R. C. George, and J. A. Maeland. 1995. Distribution of serovariants of groupB streptococci in isolates from England and Norway. Med. Microbiol. 42:246–250.
22. Lachenauer, C. S., D. L. Kasper, J. Shimada, Y. Ichiman, H. Ohtsuka, M.Kaku, L. C. Paoletti, P. Ferrieri, and L. C. Madoff. 1999. Serotypes VI andVIII predominate among group B streptococci isolated from pregnant Jap-anese women. J. Infect. Dis. 179:1030–1033.
23. Lancefield, R. 1933. A serological differentiation of human and other groupsof hemolytic streptococci. J. Exp. Med. 57:571–595.
24. Lin, F. Y., J. D. Clemens, P. H. Azimi, J. A. Regan, L. E. Weisman, J. B.Philips III, G. G. Rhoads, P. Clark, R. A. Brenner, and P. Ferrieri. 1998.Capsular polysaccharide types of group B streptococcal isolates from neo-nates with early-onset systemic infection. J. Infect. Dis. 177:790–792.
25. Musser, J. M., S. J. Mattingly, R. Quentin, A. Goudeau, and R. K. Selander.1989. Identification of a high-virulence clone of type III Streptococcus aga-lactiae (group B Streptococcus) causing invasive neonatal disease. Proc. Natl.Acad. Sci. USA 86:4731–4735.
26. Nichols, W. S., and R. M. Nakamura. 1986. Agglutination and agglutinationinhibition assays, p. 49–56. In N. R. Rose, H. Friedman, and J. L. Fahey(ed.), Manual of clinical laboratory immunology, 3rd ed. American Societyof Microbiology, Washington, D.C.
27. Palacios, G. C., E. K. Eskew, F. Solorzano, and S. J. Mattingly. 1997.Decreased capacity for type-specific-antigen synthesis accounts for highprevalence of nontypeable strains of group B streptococci in Mexico. J. Clin.
Microbiol. 35:2923–2926.28. Pfaller, M. A., R. N. Jones, S. A. Marshall, M. B. Edmond, and R. P. Wenzel.
1997. Nosocomial streptococcal blood stream infections in the SCOPE pro-gram: species occurrence and antimicrobial resistance. The SCOPE HospitalStudy Group. Diagn. Microbiol. Infect. Dis. 29:259–263.
29. Rolland, K., C. Marois, V. Siquier, B. Cattier, and R. Quentin. 1999. Geneticfeatures of Streptococcus agalactiae strains causing severe neonatal infec-tions, as revealed by pulsed-field gel electrophoresis and hylB gene analysis.J. Clin. Microbiol. 37:1892–1898.
30. Rubens, C. E., L. M. Heggen, and J. M. Kuypers. 1989. IS861, a group Bstreptococcal insertion sequence related to IS150 and IS3 of Escherichia coli.J. Bacteriol. 171:5531–5535.
31. Rubens, C. E., L. M. Heggen, R. F. Haft, and M. R. Wessels. 1993. Identi-fication of cpsD, a gene essential for type III capsule expression in group Bstreptococci. Mol. Microbiol. 8:843–855.
32. Sellin, M., S. Håkansson, and M. Norgren. 1995. Phase-shift of polysaccha-ride capsule expression in group B streptococci, type III. Microb. Pathog.18:401–415.
33. Sellin, M., M. Linderholm, M. Norgren, and S. Håkansson. 1992. Endocar-ditis caused by a group B Streptococcus strain, type III, in a nonencapsulatedphase. J. Clin. Microbiol. 30:2471–2473.
34. Skoza, L., and S. Mohos. 1976. Stable thiobarbituric acid chromophore withdimethyl sulphoxide. Application to sialic acid assay in analytical de-O-acetylation. Biochem. J. 159:457–462.
35. von Hunolstein, C., S. D’Ascenzi, B. Wagner, J. Jelinkova, G. Alfarone, S.Recchia, M. Wagner, and G. Orefici. 1993. Immunochemistry of capsulartype polysaccharide and virulence properties of type VI Streptococcus aga-lactiae (group B streptococci). Infect. Immun. 61:1272–1280.
36. Wessels, M. R., C. E. Rubens, V. J. Benedi, and D. L. Kasper. 1989. Defi-nition of a bacterial virulence factor: sialylation of the group B streptococcalcapsule. Proc. Natl. Acad. Sci. USA 86:8983–8987.
37. Wessels, M. R., J. L. DiFabio, V. J. Benedi, D. L. Kasper, F. Michon, J. R.Brisson, J. Jelinkova, and H. J. Jennings. 1991. Structural determination andimmunochemical characterization of the type V group B Streptococcus cap-sular polysaccharide. J. Biol. Chem. 266:6714–6719.
38. Wessels, M. R., W. J. Benedi, H. J. Jennings, F. Michon, J. L. DiFabio, andD. L. Kasper. 1989. Isolation and characterization of type IV group BStreptococcus capsular polysaccharide. Infect. Immun. 57:1089–1094.
39. Yamamoto, S., K. Miyake, Y. Koike, M. Watanabe, Y. Machida, M. Ohta,and S. Iijima. 1999. Molecular characterization of type-specific capsularpolysaccharide biosynthesis genes of Streptococcus agalactiae type Ia. J. Bac-teriol. 181:5176–5184.
3428 SELLIN ET AL. J. CLIN. MICROBIOL.
on Septem
ber 25, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















