
Genotoxicity of environmental air pollution in three European cities: Prague, Košice and Sofia
11
Mutation Research 563 (2004) 49–59 Genotoxicity of environmental air pollution in three European cities: Prague, Košice and Sofia Alena Gábelová a,∗ , Zuzana Valoviˇ cová a , Eva Horváthová a , Darina Slame ˇ nová a , Blanka Binková b , Radim J. Šrám b , Peter B. Farmer c a Laboratory of Mutagenesis and Carcinogenesis, Cancer Research Institute of the Slovak Academy of Sciences, Bratislava, Slovak Republic b Laboratory of Genetic Ecotoxicology, Institute of Experimental Medicine of the Academy of Sciences of the Czech Republic, Prague, Czech Republic c Cancer Biomarkers and Prevention Group, Biocentre, University of Leicester, UK Received 1 March 2004; received in revised form 20 April 2004; accepted 30 May 2004 Available online 27 July 2004 Abstract The genotoxic potential of extractable organic matter (EOM) associated with the respirable particulate matter (PM <10 m) of atmospheric pollution has been determined in three European cities—Prague (Czech Republic, two monitoring sites, Libuš and Sm´ ıchov), Košice (Slovak Republic) and Sofia (Bulgaria) using the alkaline single-cell gel electrophoresis (the comet assay). The EOM samples were extracted by dichloromethane from ambient airborne particles collected daily (24 h intervals) during 3-month sampling periods in winter and summer seasons. The human metabolically competent cell line Hep G2 was used as a test system and benzo[a]pyrene (BaP), a known carcinogen, was applied as a positive control (internal standard) in each electrophoretic run. Two-hour exposure of Hep G2 cells to equivalent EOM concentrations ranging from 5 to 150 g EOM/ml resulted in a linear dose-dependent increase of DNA migration (r > 0.9, P < 0.01). A less significant dose-response (r = 0.61) was only induced by the EOM sample from the locality Prague-Libuš (PRG-LB) in the winter. Generally, a 1.5 to four-fold increase of DNA strand breaks over the background control level was determined in EOM-exposed cells. In order to compare the genotoxic potential of individual EOMs, a mathematical model was used to correct the ‘real’ data. No substantial location- or season-related differences were found in EOM genotoxicity (EOM g/ml), except for the EOM sample from Sofia, collected in the summer. This EOM sample induced a nearly two-fold lower level of DNA damage in comparison with other EOMs. On the other hand, clear statistically significant location- and season-related differences (P < 0.001) in ambient air geno- toxicity were determined when the EOM quantity per cubic meter of air (g/m 3 ) was taken into account. In that case, the genotoxicity of winter air pollution was six- to 10-fold higher in comparison with summer air. The air pollution genotoxicity in individual localities rose during the winter season in the order: PRG-LB < Košice < Prague-Sm´ ıchov (PRG-SM) < Sofia, while during the summer season the highest ambient air genotoxicity was revealed in the locality Prague-Sm´ ıchov and approx- imately equal air pollution genotoxicity was determined among localities Prague-Libuš, Košice and Sofia (PRG-LB ∼ Košice ∼ Sofia < PRG-SM). The greatest overall air pollution genotoxicity was determined in the locality Sofia during the winter season. ∗ Corresponding author. Tel.: +421-2-59327-512; fax: +421-2-59327-506. E-mail address: [email protected] (A. G´ abelov´ a). 1383-5718/$ – see front matter © 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.mrgentox.2004.05.014
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Genotoxicity of environmental air pollution in three European cities: Prague, Košice and Sofia

Mutation Research 563 (2004) 49–59
Genotoxicity of environmental air pollution in three Europeancities: Prague, Košice and Sofia
Alena Gábelováa,∗, Zuzana Valovicováa, Eva Horváthováa, Darina Slamenováa,Blanka Binkováb, Radim J. Šrámb, Peter B. Farmerc
a Laboratory of Mutagenesis and Carcinogenesis, Cancer Research Institute of the Slovak Academy of Sciences,Bratislava, Slovak Republic
b Laboratory of Genetic Ecotoxicology, Institute of Experimental Medicine of the Academy of Sciences of the Czech Republic,Prague, Czech Republic
c Cancer Biomarkers and Prevention Group, Biocentre, University of Leicester, UK
Received 1 March 2004; received in revised form 20 April 2004; accepted 30 May 2004
Available online 27 July 2004
Abstract
The genotoxic potential of extractable organic matter (EOM) associated with the respirable particulate matter (PM<10�m)of atmospheric pollution has been determined in three European cities—Prague (Czech Republic, two monitoring sites, Libušand Smıchov), Košice (Slovak Republic) and Sofia (Bulgaria) using the alkaline single-cell gel electrophoresis (the cometassay). The EOM samples were extracted by dichloromethane from ambient airborne particles collected daily (24 h intervals)during 3-month sampling periods in winter and summer seasons. The human metabolically competent cell line Hep G2 wasused as a test system and benzo[a]pyrene (BaP), a known carcinogen, was applied as a positive control (internal standard) ineach electrophoretic run. Two-hour exposure of Hep G2 cells to equivalent EOM concentrations ranging from 5 to 150�gEOM/ml resulted in a linear dose-dependent increase of DNA migration (r > 0.9,P < 0.01). A less significant dose-response(r = 0.61) was only induced by the EOM sample from the locality Prague-Libuš (PRG-LB) in the winter. Generally, a 1.5 tofour-fold increase of DNA strand breaks over the background control level was determined in EOM-exposed cells. In order tocompare the genotoxic potential of individual EOMs, a mathematical model was used to correct the ‘real’ data. No substantiallocation- or season-related differences were found in EOM genotoxicity (EOM�g/ml), except for the EOM sample fromSofia, collected in the summer. This EOM sample induced a nearly two-fold lower level of DNA damage in comparison withother EOMs.
On the other hand, clear statistically significant location- and season-related differences (P < 0.001) in ambient air geno-toxicity were determined when the EOM quantity per cubic meter of air (�g/m3) was taken into account. In that case, thegenotoxicity of winter air pollution was six- to 10-fold higher in comparison with summer air. The air pollution genotoxicityin individual localities rose during the winter season in the order: PRG-LB< Košice< Prague-Smıchov (PRG-SM)< Sofia,while during the summer season the highest ambient air genotoxicity was revealed in the locality Prague-Smıchov and approx-imately equal air pollution genotoxicity was determined among localities Prague-Libuš, Košice and Sofia (PRG-LB∼ Košice∼ Sofia< PRG-SM). The greatest overall air pollution genotoxicity was determined in the locality Sofia during the winterseason.
∗ Corresponding author. Tel.:+421-2-59327-512; fax:+421-2-59327-506.E-mail address:[email protected] (A. Gabelova).
1383-5718/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.mrgentox.2004.05.014

50 A. Gabelova et al. / Mutation Research 563 (2004) 49–59
In a time course study to evaluate the kinetics of DNA strand break rejoining it was shown that the level of DNA strandbreaks in EOM-exposed cells has returned to near the background level within 24 h after the treatment.© 2004 Elsevier B.V. All rights reserved.
Keywords:Airborne pollution; PM10; Extractable organic matter; Genotoxicity; Single-cell gel electrophoresis; Hep G2 cells
1. Introduction
Epidemiological studies have consistently demon-strated that inhalable and respirable particulate atmo-spheric pollution (particulate matter<10�m, PM10)is associated with an increased incidence of respira-tory and cardiovascular mortality and morbidity[1,2],intrauterine growth retardation[3], embryotoxicity[4], and probably increased rates of lung cancer[5–7].Till now, however, the biological mechanisms thatmight be responsible for these adverse health effectsare unknown. PM10 is a complex mixture of aggre-gates of organic and inorganic compounds such as sul-phates, halogenated and nitrated organic compounds,polycyclic aromatic hydrocarbons (PAHs), and theirnitro- and oxy-derivatives, strong acids, trace elementsincluding metals, and water. More than 2800 chemi-cals have been identified in ambient air[8], many ofwhich are known or suspected human mutagens andcarcinogens[9]. The main sources of these substancesare anthropogenic (man-made) emissions—fossil fuelcombustion, factories, household, and automobileexhausts. PM10 vary in abundance and compositionboth spatially and temporally, with concentrationstypically highest in urban atmosphere[10]. Around100 PAHs have been identified in airborne particles;most of them are frequently found in highly pollutedambient air of large cities[11,12]. Nowadays, envi-ronmental air pollution is a matter of great interestbecause millions of people are chronically exposedto low doses of noxious chemicals, e.g. PAHs, fre-quently present in the complex mixtures. Thereforethere is a need to improve our knowledge aboutthe mechanisms underlying air pollution-inducedadverse effects on biological systems in order tobetter assess the impact of air pollution on humanhealth.
Mutagenicity and genotoxicity of organic complexmixture of urban airborne particles have been demon-strated with different short-term tests on prokaryoticand eukaryotic organisms[4,12–15], including mam-
malian cells cultured in vitro[16,17] and laboratoryanimals [6]. However, rather a limited number ofstudies were aimed to assess EOM genotoxicity usinghuman cells cultured in vitro. A most promising toolfor detection and risk assessment of compounds thatmight pose a health hazard to man are human-derivedmetabolically competent cell lines. Such cell linescan reflect the metabolism of genotoxins in mam-mals better than other in vitro models that requireaddition of exogenous activation mixtures. Severalstudies have indicated that the human hepatoma cellline Hep G2 has retained the activities of certainphase I and phase II enzymes, which play a key rolein the activation/detoxification of various promuta-gens/procarcinogens[18,19].
DNA strand breakage represents only one type oflesion caused by genotoxic compounds. However, be-cause the strand breaks are so readily detected, despitethe fact that most of them are also readily repaired,their measurement has become a standard method ofDNA damage expression. Up to now, a number of bio-chemical techniques have been developed for DNAstrand break measurement, based on different mecha-nisms and performed under different conditions[20].Nowadays, the single-cell gel electrophoresis (SCGE)assay, a rapid and sensitive fluorescence microscopicmethod, is most frequently used as a genotoxicityscreening test[21]. Depending on the experimentalconditions, this technique allows detection of varioustypes of DNA damage including single and doublestrand breaks, oxidative DNA damage, alkali-labilesites and DNA cross-links.
The focus of this work, which is a part of anEC-funded study[22], was to characterise the geno-toxic properties of air pollution mixtures in vitro. Forthis purpose, airborne particles (PM10) were collecteddaily (24 h intervals) by high-volume samplers at fourmonitoring sites during a 3-month period in summerand winter seasons, and extracted by dichloromethane.Three monitoring sites, Prague-Smıchov (PRG-SM),Košice and Sofia, are classified according to the

A. Gabelova et al. / Mutation Research 563 (2004) 49–59 51
European Environmental Register of monitoring sta-tions as commercial, residential, traffic, and urban,while the locality Prague-Libuš (PRG-LB) is char-acterized as background, residential and suburban.To determine the correlation between the genotoxicproperties of EOM and PAHs, the concentrations of13 PAHs in each organic extract were determinedby an HPLC method with fluorimetric detection. Forthe in vitro experiments, the dichloromethane organicextracts of PM10 were evaporated to dryness undera stream of nitrogen and re-dissolved in dimethylsulfoxide (DMSO).
The aims of this study were: (i) to determine thegenotoxic potential of organic complex mixtures ex-tracted from PM10 using SCGE; (ii) to elucidateseason- or location-related variability in the biologi-cal effects of individual EOMs due to variation in thecrude organic mixture compositions and air pollutantconcentration per cubic meter air. Part of this studywas focused on the kinetics of DNA strand breakrejoining induced by EOMs.
2. Materials and methods
2.1. Cell line
The human hepatoma cell line Hep G2 was gener-ously provided by Prof. Andrew R. Collins (Universityof Oslo, Norway). Hep G2 cells were maintained inWilliam‘s modified medium supplemented with 10%fetal calf serum and antibiotics (penicillin 200 U/ml;streptomycin and kanamycin 100�g/ml) in humidi-fied 5% CO2 atmosphere.
2.2. Chemicals
B[a]P (CAS No. 50-32-8) was purchased fromSigma. The stock solutions of B[a]P (2 mM in DMSO)and organic complex mixture (EOM, 50 mg/ml inDMSO) from individual samplings were kept at−20◦C and diluted immediately before use. Allchemicals used for cell cultivation (MEM, FCS, an-tibiotics) and agarose LMP (low melting point) werepurchased from GIBCO BRL; agarose electrophore-sis grade (NMP, normal melting point) and ethidiumbromide were purchased from Sigma. All other chem-icals and solvents were of analytical grade.
2.3. Collection of air samples, extraction of EOMand chemical analysis
Air particles PM10 (particulate matter<10�m)were collected uniformly by the HiVol air samplers(Anderson) daily (24 h period) during 3-month sam-pling periods at four monitoring sites—PRG-SM,PRG-LB, Košice and Sofia—during the winter andsummer seasons. The summer sampling period inindividual localities was as follows: in PRG-SMand PRG-LB from 15/06/2000 to 15/09/2000; inKošice from 20/05/2002 to 17/08/2002; in Sofia from10/06/2002 to 07/09/2002. The winter sampling pe-riod in these localities was: in PRG-SM and PRG-LBfrom 04/12/2000 to 07/03/2001; in Košice from01/12/2001 to 28/02/2002; in Sofia from 08/01/2002to 08/04/2002. Both the dichloromethane extractionsof organic complex mixture (EOM) from the filterswith PM10 samples and the chemical analyses of PAHswere performed uniformly in the laboratories of thecertified company Ecochem a.s., Prague (EN ISO CSNIEC 17025). Details of these procedures have beenalready described elsewhere[4]. The concentrationsof thirteen polycyclic aromatic hydrocarbons (PAHs)including phenanthrene (PHE), anthracene (ANT),fluoranthene (FLU), pyrene (PYR), coronene (COR),benz[a]anthracene (B[a]A), chrysene (CHRY), benzo-[b]fluoranthene (B[b]F), benzo[k]fluoranthene (B[k]-F), benzo[a]pyrene (B[a]P), dibenzo[a,h]anthracene(DB[a,h]A), benzo[g,h,l]perylene (B[g,h,l]P), andindeno[1,2,3-cd]pyrene (I[c,d]P) were analysed ineach EOM sample. For the in vitro experiments,the dichloromethane organic extracts of PM10 wereevaporated to dryness under a stream of nitrogenand re-dissolved in dimethyl sulfoxide (DMSO). Thestock solution of each individual EOM sample con-tained 50 mg of EOM per ml of DMSO (50 mg/ml).
2.4. Treatment of human cells
Hep G2 cells (1.5× 105) were plated on severalculture vials (Ø= 20 mm). The cells were grownfor 48 h until they were semi-confluent, prior to ex-position to mixtures and positive control. The stocksolutions of individual EOM (50 mg/ml) were freshlydiluted in DMSO before use and added to serum-freemedium to reach final concentrations ranging from5 to 150�g/ml. The concentration of DMSO neverexceeded 0.5%. Control cells (negative control) were

52 A. Gabelova et al. / Mutation Research 563 (2004) 49–59
exposed to 0.5% DMSO. After 2 h exposure themedium was removed and cells were washed twicewith PBS buffer. Then, the cells were either harvestedimmediately and used for SCGE (the comet assay) orcultivated in fresh medium. To measure the kineticsof DNA break rejoining, Hep G2 cells were harvestedafter 2, 4, 16 and 24 h of post-cultivation and used forSCGE. Benzo[a]pyrene (BaP, 7.5�M) was used as apositive control (internal standard) and 0.5% DMSOas negative control in each electrophoretic run.
2.5. Single-cell gel electrophoresis
The procedure of Singh et al.[23], modified byCollins et al. [24] and Slamenová et al.[25] was
followed. A base layer of 100�l of 1.0% NMP agarosein PBS buffer (Ca2+- and Mg2+-free) was placed onmicroscope slides. Instead of fully frosted microscopeslides plain slides were used covered by a thin layerof 1% NMP agarose in water and dried at 50◦C.Cells suspended in 0.75% LMP agarose were spreadon the base layer on each of three slides. When theagarose had solidified the slides were placed in a ly-sis mixture (2.5 M NaCl, 100 mM Na2EDTA, 10 mMTris–HCl, pH 10 and 1% Triton X-100) at 4◦C for1 h to remove cellular proteins. Slides were then trans-ferred to an electrophoretic box containing an alka-line solution (300 mM NaOH, 1 mM Na2EDTA, pH> 13). After 40 min unwinding time at 4◦C, a currentof 25 V (300 mA) was applied for 30 min. The slideswere removed, neutralised with 3× 5 min washes withTris–HCl (0.4 M, pH 7.5), and stained with 20�l ofethidium bromide (EtBr, 10�g/ml). EtBr-stained nu-cleoides were examined with a Zeiss Jenalumar flu-orescence microscope by image analysis using thesoftware Komet 3.0 (Kinetic Imaging Ltd.). The per-centage of DNA in the tail (% tail DNA) was usedas a parameter to analyse induced DNA damage. Atotal of 100 nucleoids were scored per each sam-ple in one electrophoretic run. The data from all in-dependent experiments (at least four) were pooledtogether and evaluated statistically by the Student’st-test.
2.6. Mathematical analys´ıs
A mathematical model proposed by De Boecket al. [26] was used in order to eliminate the experi-mental variability of individual measurements (elec-trophoretic runs) and variations in the response of thecells to treatment. This model takes into account boththe negative (neg) and positive (pos) standard datafrom the appropriate electrophoresis sessions for eachmeasurement (M). The mean data of individual EOMgenotoxicity measurement were corrected accordingto the formula:
F1 = M − neg
pos− neg;
S.D.F1 = [(|pos− neg| × S.D.(M)] + [(|M − pos| × S.D.(neg)] + [(|M − neg| × S.D.(pos)]
(pos− neg)2;
whereM is the measurement; neg, the negative stan-dard; pos, the positive standard and; S.D., the standarddeviation. Data were expressed in relative units.
3. Results
3.1. Particulate matter (PM10) and EOM
The average seasonal concentrations of PM10,EOM, benzo[a]pyrene, carcinogenic PAHs (c-PAHs)and total PAHs as analyzed by HPLC with fluores-cence detection in crude extracts collected in indi-vidual localities during 3-month winter and summersampling periods have been already published[22].Extractable organic matter per PM10 was at least2-fold higher in winter than in summer, and carcino-genic PAHs over 10-fold higher in winter air thansummer air. The PM10 concentration in individuallocalities during winter sampling rose in the order:PRG-LB< Košice< PRG-SM< Sofia, while duringsummer sampling the order was: Košice< PRG-LB< Sofia < PRG-SM. The highest concentrations ofc-PAHs and total PAHs were determined in Sofia dur-ing the winter, the lowest concentrations were seenin the localities Košice and PRG-LB during summersampling.Table 1shows the variations in PAH con-centrations in crude extracts according to the localityand season.

A. Gabelova et al. / Mutation Research 563 (2004) 49–59 53
Table 1PAHs content in the EOM samples from the four monitoring sites—PRG-SM, PRG-LB, Košice and Sofia
PAHsa EOM (ng/mg) Summer Winter
PRG-LB PRG-SM Košice Sofia PRG-LB PRG-SM Košice Sofia
PHE 19.9 25.8 47.2 41.7 121.5 131.2 187.8 191.1ANT 3.1 4.8 10.9 8.7 7.4 7.4 30.8 23.7FLU 36.7 51.6 116.2 85.6 561.7 515.7 356.0 467.3PYR 39.8 51.3 105.0 59.5 681.4 529.1 306.3 393.3COR 24.5 37.1 54.0 39.1 30.4 24.1 10.8 8.9B[a]A 21.4 33.9 71.3 51.1 377.5 348.3 181.8 249.4CHRY 27.5 38.7 17.9 65.3 340.7 314.8 176.3 375.6B[b]F 62.7 74.2 159.4 121.7 340.7 308.1 165.2 212.6B[k]F 29.1 35.5 65.5 55.1 156.5 134.0 58.4 120.9B[a]P 44.3 50.0 90.4 92.4 267.0 234.4 89.7 196.8DB[a,h]A 8.3 9.5 15.4 9.8 23.9 19.4 6.4 34.8B[g,h,l]P 81.0 132.3 148.9 134.3 165.7 127.3 48.1 128.8I[c,d]P 79.5 87.1 149.3 85.4 202.6 167.4 49.8 162.3Total c-PAHs 353.8 461.1 718.1 615.6 1874.8 1653.7 775.7 1481.2Total PAHs 477.7 631.8 1051.3 849.6 3277.2 2861.3 1667.5 4046.6
c-PAH: benz[a]anthracene B[a]A, chrysene CHRY, benzo[b]fluoranthene B[b]F, benzo[k]fluoranthene B[k]F, benzo[a]pyrene B[a]P,dibenzo[a,h]anthracene DB[a,h]A, benzo[g,h,l]perylene B[g,h,l]P, indeno[1,2,3-cd]pyrene I[c,d]P.
a Reliability of PAHs analysis expressed as percent relative standard deviation (% R.S.D.) was less than 20% with the exception ofPHE (23%), ANT (25%) and DB[a,h]A (±26%).
3.2. Genotoxicity of EOM
On the basis of preliminary experiments (data notshown), a 2 h exposure period was chosen for the as-sessment of the genotoxicity of individual EOM sam-ples. Using these treatment conditions the viability ofexposed Hep G2 cells never decreased below 80%.Human metabolically competent cells were treatedwith equivalent EOM concentrations ranging from 5 to150�g EOM/ml. Benzo[a]pyrene (BaP) at concentra-tion of 7.5�M was used as a positive control in eachelectrophoretic run (internal standard). In general, a1.5- to four-fold increase in DNA strand breaks overthe background level was determined in EOM-exposedcells. Nearly all complex mixtures induced a statisti-cally significant dose-dependent increase of DNA mi-gration (P < 0.05–0.001,r > 0.9) in Hep G2 cells(Table 2). The EOM sample from Prague (PRG-LB)collected in the winter showed a clear increase ofstrand breaks, which reached a plateau at a concen-tration of about 20�g/ml. A slightly higher level ofDNA damage was estimated for EOM samples fromwinter air in comparison with summer air, however,these differences were not statistically significant.
In order to compare the EOM genotoxicity be-tween individual localities and seasons the real data
were corrected according to the formula mentionedabove (Section 2) and expressed in relative units(F1). Benzo[a]pyrene, the known PAH carcinogen,was used as an internal standard in our experi-ments. Genotoxicity of B[a]P in individual elec-trophoretic runs ranged between 23.07 and 31.53%tail DNA. No location- or season-related substan-tial differences in EOM genotoxicity except for thelocality in Sofia were determined (Table 2). TheEOM sample from Sofia, summer period, inducedan approximately two-fold lower extent of DNA mi-gration in Hep G2 cells compared with the wintersample.
3.3. Air genotoxicity
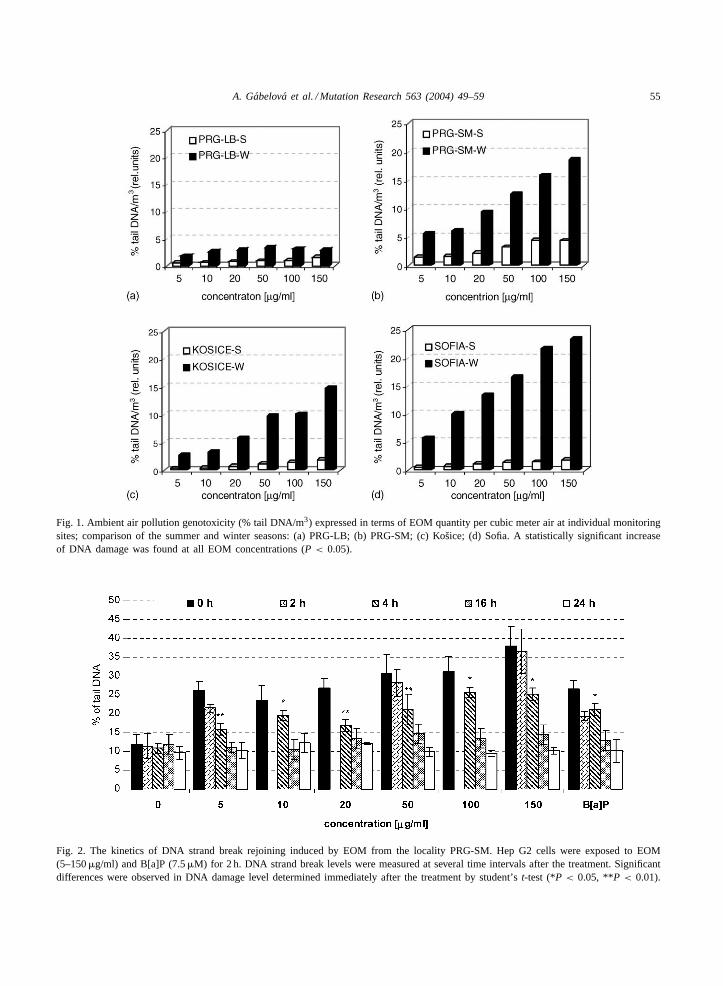
Genotoxicity of the ambient air pollution based onthe amount of EOM associated with particulate mat-ter per unit volume of air (EOM�g/m3) is shown inFig. 1. In general, significantly higher genotoxicityof ambient air was demonstrated during the winterseason in comparison with the summer period in allfour monitoring sites. The ambient air genotoxic-ity during the winter season was three- to four-foldhigher in the localities Prague-Libuš (Fig. 1a) andPrague-Smıchov (Fig. 1b); eight- to nine-fold higher

54 A. Gabelova et al. / Mutation Research 563 (2004) 49–59
Table 2DNA damage level detected in Hep G2 cells exposed to EOM using the alkaline single-cell gel electrophoresis
EOM sample (�g/ml) Mean % tail DNA (real data) Percentage of tail DNA corrected [F1]
Winter Summer Winter Summer
PRG-LB0 12.13± 2.53 10.79± 1.655 19.07± 2.56∗ 18.40± 2.67∗∗ 0.48 ± 0.43 0.62± 0.4410 25.65± 2.33∗∗ 19.44± 2.55∗∗ 0.93 ± 0.49 0.70± 0.4520 27.95± 2.39∗∗∗ 19.98± 2.60∗∗ 1.09 ± 0.55 0.75± 0.4450 29.27± 3.19∗∗∗ 24.98± 4.57∗∗∗ 1.18 ± 0.66 1.16± 0.69100 29.79± 2.95∗∗∗ 25.18± 3.02∗∗∗ 1.21 ± 0.66 1.17± 0.57150 28.73± 3.54∗∗∗ 28.33± 4.02∗∗∗ 1.14 ± 0.66 1.43± 0.70
B[a]P (7.5�M) 26.70 ± 5.03∗∗∗ 23.07± 3.29∗∗∗
PRG-SM0 11.02± 3.11 13.96± 2.325 20.19± 5.24∗∗ 17.54± 4.16∗ 0.62 ± 0.63 0.38± 0.7310 21.08± 3.57∗∗∗ 19.59± 3.33∗ 0.69 ± 0.51 0.60± 0.7120 26.13± 2.54∗∗∗ 23.64± 3.53∗∗ 1.06 ± 0.47 1.02± 0.8350 30.91± 3.51∗∗∗ 25.09± 5.14∗∗ 1.40 ± 0.71 1.18± 1.00100 36.02± 6.60∗∗∗ 29.25± 5.90∗∗∗ 1.78 ± 1.12 1.62± 1.37150 40.24± 5.14∗∗∗ 31.45± 5.63∗∗∗ 2.08 ± 1.16 1.85± 1.41B[a]P (7.5�M) 25.38 ± 3.58∗∗∗ 23.41± 3.43∗∗
KOŠICE0 11.02± 0.73 9.05± 0.535 16.63± 3.05∗ 14.17± 2.64∗ 0.29 ± 0.24 0.30± 0.2110 18.28± 2.70∗ 15.94± 2.65∗ 0.39 ± 0.24 0.39± 0.2320 22.79± 2.66∗∗ 20.98± 3.02∗∗ 0.68 ± 0.27 0.63± 0.2850 28.89± 4.18∗∗∗ 28.67± 2.72∗∗∗ 1.12 ± 0.39 0.95± 0.32100 30.96± 3.56∗∗∗ 32.94± 2.33∗∗∗ 1.36 ± 0.38 1.06± 0.34150 39.63± 5.67∗∗∗ 40.28± 3.02∗∗∗ 1.78 ± 0.59 1.53± 0.45B[a]P (7.5�M) 29.75 ± 3.23∗∗∗ 26.62± 2.52∗∗∗
SOFIA0 11.51± 1.04 12.63± 1.095 16.30± 2.01∗ 16.40± 2.05∗ 0.39 ± 0.29 0.22± 0.1910 19.97± 1.82∗∗ 18.94± 2.66∗ 0.68 ± 0.30 0.24± 0.2220 22.77± 1.88∗∗ 21.81± 3.73∗∗ 0.91 ± 0.33 0.46± 0.3250 25.43± 1.13∗∗∗ 24.49± 3.13∗∗ 1.12 ± 0.32 0.53± 0.32100 29.78± 3.06∗∗∗ 26.35± 3.07∗∗∗ 1.47 ± 0.57 0.57± 0.32150 31.15± 5.20∗∗∗ 28.78± 4.05∗∗∗ 1.58 ± 0.77 0.71± 0.40B[a]P (7.5�M) 23.92 ± 2.39∗∗ 31.53± 7.36∗∗∗
Columns 2 (winter) and 3 (summer) represent ‘real’ data for % tail DNA, including internal standard, B[a]P-treated Hep G2 cells, andcontrol cells. Columns 4 and 5 show values after correction with mathematical model (F1) proposed by De Boeck et al.[26].
∗ P < 0.05 as compared with DMSO-treated cells.∗∗ P < 0.01 as compared with DMSO-treated cells.∗∗∗ P < 0.001 as compared with DMSO-treated cells.
in the locality Košice (Fig. 1c) and even 12- to15-fold higher in the monitoring site Sofia (Fig. 1d)in comparison with the appropriate summer season.Environmental air genotoxicity in the monitoring sitesrose during the winter season in the order: PRG-LB< Košice < PRG-SM < Sofia, while during the
summer sampling period the order was: PRG-LB∼ Košice ∼ Sofia < PRG-SM. Overall, the great-est genotoxicity of the environmental air pollutionwas determined in the locality Sofia, winter seasonand Prague, PRG-SM, during the summer samplingperiod.
A. Gabelova et al. / Mutation Research 563 (2004) 49–59 55
Fig. 1. Ambient air pollution genotoxicity (% tail DNA/m3) expressed in terms of EOM quantity per cubic meter air at individual monitoringsites; comparison of the summer and winter seasons: (a) PRG-LB; (b) PRG-SM; (c) Košice; (d) Sofia. A statistically significant increaseof DNA damage was found at all EOM concentrations (P < 0.05).
Fig. 2. The kinetics of DNA strand break rejoining induced by EOM from the locality PRG-SM. Hep G2 cells were exposed to EOM(5–150�g/ml) and B[a]P (7.5�M) for 2 h. DNA strand break levels were measured at several time intervals after the treatment. Significantdifferences were observed in DNA damage level determined immediately after the treatment by student’st-test (*P < 0.05, **P < 0.01).

56 A. Gabelova et al. / Mutation Research 563 (2004) 49–59
3.4. Kinetics of DNA break rejoining
A time course study was performed with the EOMsample from Prague (PRG-SM), winter season, in or-der to optimize the experimental condition for thestudy of DNA repair kinetics. In this pilot study sev-eral time intervals after the treatment (2, 4, 16 and24 h) were chosen to measure the residual DNA dam-age level in exposed cells (Fig. 2). No, or a very slight,reduction of DNA damage was detected in Hep G2cells 2 h after the treatment in fresh medium. Althougha significant decrease of DNA migration was deter-mined 4 h after the treatment, still approx. 60–80% ofDNA strand breaks persisted in the treated cells. Six-teen hours post-cultivation in fresh medium resulted ina 80–90% reduction of DNA damage in treated cellsand the level of DNA strand breaks had returned tonear the background level within 24 h after the treat-ment. On the basis of these preliminary experiments,two sampling intervals, 4 and 24 h after the treatment,were chosen to evaluate the kinetics of DNA break re-joining in cells exposed to EOM samples. In general,a reduction of DNA damage of about 20% was deter-mined in EOM-exposed cells 4 h after the treatment re-gardless of the season or locality. No increase of DNAmigration over control cells was detected in treatedcells after 24 h post-cultivation in fresh medium (datanot shown).
4. Discussion
Although several studies have pointed to a causal re-lationship between ambient air pollution and adversehealth effects, the biological mechanisms that could beresponsible for these biological effects are unknown.In addition, it is not clear whether the effects are pro-duced by the PM, by the organic complex mixturesadsorbed onto inhalable and respirable particles, or byparticular chemical classes/compounds in the mixture[2,27].
Bioassay-directed chemical analysis of the crude or-ganic extracts has suggested that polycyclic aromatichydrocarbons (PAHs) might be responsible for most ofthe mutagenic and genotoxic activity[28]. Several invitro studies with fractionated crude organic complexmixtures clearly demonstrated that the highest muta-genic and DNA adduct-forming potential is associated
with the neutral and slightly polar fractions, whichcontain most of the PAHs and nitro-PAHs derivatives[14,17,29,30]. In order to verify the key role of PAHsin ambient air genotoxicity and adverse health effectsan EC-funded study has been initiated[22]. Part ofthis study has been focused on the genotoxic prop-erties of the ambient air pollution mixture in vitro.Complex organic mixtures were prepared by extrac-tion from the airborne particles (PM10) collected atfour monitoring sites during the summer and winterseason. In each organic extract the concentrations of13 PAHs were determined by an HPLC method withfluorimetric detection. As has been shown recently, thetotal mass of extractable organic matter per PM10 wasat least two-fold higher in winter than in summer, andcarcinogenic PAHs were over 10-fold higher in winterair than in summer air[22].
Approximately a 1.5- to four-fold increase of DNAstrand breaks over the background level was foundin Hep G2 cells exposed to equivalent (5–150�g/ml)EOM concentrations (Table 2). A 2 h exposure toEOMs resulted in weak cytotoxicity, however, cellviability did not decrease below 80% (data notshown). A linear dose-response effect (P < 0.01, r> 0.9) was determined using each EOM, though thedose-dependent increase of DNA migration was lesssignificant (r = 0.61) for the EOM sample from thelocality Prague-Libuš, winter season (Table 2). Aslightly higher level of DNA damage was estimatedfor EOM samples from winter air in comparisonwith summer air, however, these differences werenot statistically significant. In order to compare theEOM genotoxicity from individual monitoring sitesthe real data (% tail DNA) were corrected accord-ing to De Boeck et al.[26]. Despite this correction,which should have eliminated any variations in elec-trophoretic runs and cell-response to exposure, nosubstantial location- or season-related differences inEOM genotoxicity were measured except for the lo-cality Sofia. Genotoxicity of EOM from Sofia, winterseason, was approximately two-fold higher comparedwith EOM from the summer season (Table 2). Themutagenic and genotoxic potential of EOM from air-borne particles has been already demonstrated withseveral short-term tests on prokaryotic and eukaryoticorganisms[11–14,31,32]. Our data are in agreementwith those of others who did not find any significantdifferences in EOM genotoxicity in dependence on the

A. Gabelova et al. / Mutation Research 563 (2004) 49–59 57
locality and season[4,17,29,30]. The failure of in vitrosystems to detect any substantial variation in EOMgenotoxicity due to differences in the crude mixturecomposition may be explained by the saturation ofmetabolic pathways needed to activate pro-mutagenicagents. More than 2800 chemicals have been iden-tified in ambient air [8], many of which requiremetabolic activation to electrophilic species beforethey can interact with DNA and exert their mutagenicand carcinogenic effects. Although Hep G2 cellshave retained activities of various phase I and phaseII enzymes involved in the activation/detoxificationof various promutagens/procarcinogens, the enzymelevels are lower than in vivo[18,33]. A time coursestudy that focused on standardisation of treatmentconditions using B[a]P showed that comet formationincreased gradually as a function of dose and reachedmaximum values between 7.5–10�M. Concentra-tions over 7.5�M resulted in substantial reduction ofcomet formation (data not shown).
Clear, statistically highly significant differences(P < 0.001) in the ambient air genotoxicity wererevealed between summer and winter seasons andamong monitoring sites if the EOM concentrationper cubic meter air (�g/m3) at individual monitor-ing sites was taken into account. The genotoxicityof environmental air pollution was 6–10-fold higherduring the winter period as compared to the summerseason (Fig. 1a-d). Comparing the monitoring sites,the genotoxicity rose during the winter season in theorder: Prague-Libuš< Košice< Prague-Smıchov <
Sofia, while during the summer season approximatelyequal ambient air genotoxicity was determined amonglocalities Prague-Libuš, Košice and Sofia, whereasthe locality Prague-Smıchov had higher genotoxixity(PRG-LB∼Košice∼Sofia < PRG-SM). The overallgreatest air pollution genotoxicity was determined inthe locality Sofia during winter season, the lowestone in the localities Prague-Libuš and Košice. Thesedata were more or less in good correlation with theconcentration of B[a]P, cPAHs and total PAHs percubic meter of air.
A time course study to evaluate the kinetics of DNAstrand break (sb) rejoining in cells exposed to EOMshas shown that the repair is relatively slow (Fig. 2);between 60–80% of DNA damage still persisted inthe cells 4 h after the treatment. DNA repair kinet-ics differs depending on the type of lesions and their
location. Single strand break rejoining is very rapidwith the initial t1/2 values of 5.0 min for X-ray and6.8 min for hydrogen peroxide exposure[34], whilethe oxidised bases could persist in the cells for over 4 hof post-incubation[35]. Although the level of DNAstrand breaks in EOM-treated cells has returned tonear the background level within 24 h, it is hard to saywhether DNA damage was removed by the error-proneor error-free DNA repair systems.
In conclusion, this study confirmed the geno-toxic properties of organic complex mixtures asso-ciated with respirable airborne particles collectedin three European cities. No substantial season- orlocation-related variation in EOM genotoxicity (EOM�g/ml) were determined, however, significant dif-ferences in ambient air genotoxicity in dependenceon locality and season were assessed, if EOM con-centration per cubic meter of air (EOM�g/m3) wastaken into account. A DNA repair kinetics studyshowed that the removal of strand breaks induced byEOMs is relatively slow, taking around 24 h. Furtherexperiments are required to determine whether theerror-prone or error-free DNA repair systems are in-volved in DNA damage removal. The SCGE assaywith Hep G2 cells appears to be a promising approachfor the detection of genotoxic properties of ambientair pollution mixture in vitro.
Acknowledgements
The authors express their appreciation to Prof. An-drew R. Collins, University of Oslo, Norway, whokindly offered the human hepatoma Hep G2 cells,Prof. Ivan Kalina, University of P.J. Šafárik, SlovakRepublic, and Prof. Todor A. Popov, National Centerof Hygiene, Bulgaria, who performed airborne parti-cle sampling for EOM extraction. The authors wish tothank Mrs. Alžbeta Vokáliková for an excellent tech-nical assistance. The authors acknowledge the Euro-pean Commission ‘Quality of life and management ofliving resources’ programme (QLK4-CT-2000-00091)for financial support.
References
[1] N. Kunzli, I.B. Tager, Long-term health effects of particulateand other ambient air pollution: research can progress faster

58 A. Gabelova et al. / Mutation Research 563 (2004) 49–59
if we want it to, Environ. Health Perspect. 108 (2000) 915–918.
[2] C.A. Pope III, R.T. Burnett, M.J. Thun, E.E. Calle, D.Krewski, K. Ito, G.D. Thurston, Lung cancer, cardiopul-monary mortality, and long-term exposure to fine particulateair pollution, JAMA 287 (2002) 1132–1141.
[3] J. Dejmek, I. Solansky, I. Benes, J. Lenicek, R.J. Sram, Theimpact of polycyclic aromatic hydrocarbons and fine particleson pregnancy outcome, Environ. Health Perspect. 108 (2000)1159–1164.
[4] B. Binkova, M. Cerna, A. Pastorkova, R. Jelinek, I. Benes, J.Novak, R.J. Sram, Biological activities of organic compoundsadsorbed onto ambient air particles: comparison between thecities of Teplice and Prague during the summer and winterseasons 2000–2001, Mutat. Res. 525 (2003) 43–59.
[5] F. Nyberg, P. Gustavsson, L. Jarup, T. Bellander, N. Berglind,R. Jakobsson, G. Pershagen, Urban air pollution and lungcancer in Stockholm, Epidemiology 11 (2000) 487–495.
[6] P.M. Cury, A.J. Lichtenfels, M.S. Reymao, G.M. Conceicao,V.L. Capelozzi, P.H. Saldiva, Urban levels of air pollutionmodifies the progression of urethane-induced lung tumoursin mice, Pathol. Res. Pract. 196 (2000) 627–633.
[7] X.S. Zhao, Z. Wan, H.G. Zhu, R.P. Chen, The carcinogenicpotential of extractable organic matter from urban airborneparticles in Shanghai, China, Mutat. Res. 540 (2003) 107–117.
[8] J. Lewtas, J. Gallagher, Complex mixtures of urban airpollutants: identification and comparative assessment ofmutagenic and tumorigenic chemicals and emission sources,IARC Sci. Publ. (1990) 252–260.
[9] IARC. The International Agency for Research on Cancer(http://www.iarc.fr/).
[10] M.A. Mazurek, Molecular identification of organic com-pounds in atmospheric complex mixtures and relationship toatmospheric chemistry and sources, Environ. Health Perspect.110 (Suppl 6) (2002) 995–1003.
[11] A. Ducatti, V.M.F. Vargas, Mutagenic activity of airborneparticulate matter as an indicative measure of atmosphericpollution, Mutat. Res. 540 (2003) 67–77.
[12] X. Zhao, Z. Wan, G. Chen, H. Zhu, S. Jiang, J. Yao, Genotoxicactivity of extractable organic matter from urban airborneparticles in Shanghai, China, Mutat. Res. 514 (2002) 177–192.
[13] M. Cerna, A. Pastorkova, V. Vrbikova, J. Smid, P. Rossner,Mutagenicity monitoring of airborne particulate matter(PM10) in the Czech Republic, Mutat. Res. 444 (1999) 373–386.
[14] A. Buschini, F. Cassoni, E. Anceschi, L. Pasini, P.Poli, C. Rossi, Urban airborne particulate: genotoxicityevaluation of different size fractions by mutagenesis tests onmicroorganisms and comet assay, Chemosphere 44 (2001)1723–1736.
[15] A. Delgado-Rodriguez, R. Ortiz-Marttelo, R. Villalobos-Pietrini, S. Gomez-Arroyo, U. Graf, Genotoxicity of organicextracts of airborne particles in somatic cells of Drosophilamelanogaster, Chemosphere 39 (1999) 33–43.
[16] W.L. Hsiao, Z.Y. Mo, M. Fang, X.M. Shi, F. Wang,Cytotoxicity of PM(2.5) and PM(2.5--10) ambient air
pollutants assessed by the MTT and the Comet assays, Mutat.Res. 471 (2000) 45–55.
[17] J. Topinka, L.R. Schwarz, F.J. Wiebel, M. Cerna, T. Wolff,Genotoxicity of urban air pollutants in the Czech Republic.Part II. DNA adduct formation in mammalian cells byextractable organic matter, Mutat. Res. 469 (2000) 83–93.
[18] S. Knasmuller, W. Parzefall, R. Sanyal, S. Ecker, C. Schwab,M. Uhl, V. Mersch-Sundermann, G. Williamson, G. Hietsch,T. Langer, F. Darroudi, A.T. Natarajan, Use of metabolicallycompetent human hepatoma cells for the detection ofmutagens and antimutagens, Mutat. Res. 402 (1998) 185–202.
[19] M. Uhl, C. Helma, S. Knasmuller, Evaluation of the singlecell gel electrophoresis assay with human hepatoma (HepG2) cells, Mutat. Res. 468 (2000) 213–225.
[20] A. Gabelova, D. Slamenova, L. Ruzekova, T. Farkasova, E.Horvathova, Measurement of DNA strand breakage and DNArepair induced with hydrogen peroxide using single cell gelelectrophoresis, alkaline DNA unwinding and alkaline elutionof DNA, Neoplasma 44 (1997) 380–388.
[21] R.R. Tice, E. Agurell, D. Anderson, B. Burlinson, A.Hartmann, H. Kobayashi, Y. Miyamae, E. Rojas, J.C. Ryu,Y.F. Sasaki, Single cell gel/comet assay: guidelines for invitro and in vivo genetic toxicology testing, Environ. Mol.Mutagen. 35 (2000) 206–221.
[22] P.B. Farmer, R. Singh, B. Kaur, R.J. Sram, B. Binkova,I. Kalina, T.A. Popov, S. Garte, E. Taioli, A. Gabelova,A. Cebulska-Wasilewska, Molecular epidemiology studies ofcarcinogenic environmental pollutants. Effects of polycyclicaromatic hydrocarbons (PAHs) in environmental pollution onexogenous and oxidative DNA damage, Mutat. Res. Rev.Mutat. Res. 544 (2003) 397–402.
[23] N.P. Singh, M.T. McCoy, R.R. Tice, E.L. Schneider, A simpletechnique for quantitation of low levels of DNA damage inindividual cells, Exp. Cell Res. 175 (1988) 184–191.
[24] A.R. Collins, S.J. Duthie, V.L. Dobson, Direct enzymicdetection of endogenous oxidative base damage in humanlymphocyte DNA, Carcinogenesis 14 (1993) 1733–1735.
[25] D. Slamenova, A. Gabelova, L. Ruzekova, I. Chalupa,E. Horvathova, T. Farkasova, E. Bozsakyova, R. Stetina,Detection of MNNG-induced DNA lesions in mammaliancells; validation of comet assay against DNA unwindingtechnique, alkaline elution of DNA and chromosomalaberrations, Mutat. Res. 383 (1997) 243–252.
[26] M. De Boeck, N. Touil, G. De Visscher, P.A. Vande, M.Kirsch-Volders, Validation and implementation of an internalstandard in comet assay analysis, Mutat. Res. 469 (2000)181–197.
[27] F.J. Kelly, Oxidative stress: its role in air pollution and adversehealth effects, Occup. Environ. Med. 60 (2003) 612–616.
[28] D.M. DeMarini, Mutation spectra of complex mixtures,Mutat. Res. 411 (1998) 11–18.
[29] M. Cerna, D. Pochmanova, A. Pastorkova, I. Benes, J.Lenicek, J. Topinka, B. Binkova, Genotoxicity of urbanair pollutants in the Czech Republic. Part I. Bacterialmutagenic potencies of organic compounds adsorbed onPM10 particulates, Mutat. Res. 469 (2000) 71–82.
[30] B. Binkova, D. Vesely, D. Vesela, R. Jelinek, R.J. Sram,Genotoxicity and embryotoxicity of urban air particulate

A. Gabelova et al. / Mutation Research 563 (2004) 49–59 59
matter collected during winter and summer period in twodifferent districts of the Czech Republic, Mutat. Res. 440(1999) 45–58.
[31] A. Baulig, M. Sourdeval, M. Meyer, F. Marano, A. Baeza-Squiban, Biological effects of atmospheric particles on humanbronchial epithelial cells. Comparison with diesel exhaustparticles, Toxicol. In Vitro 17 (2003) 567–573.
[32] M. Lazarova, D. Slamenova, Genotoxic effects of a complexmixture adsorbed onto ambient air particles on human cellsin vitro; the effects of Vitamins E and C, Mutat. Res. 557(2004) 167–175.
[33] J. Rueff, C. Chiapella, J.K. Chipman, F. Darroudi, I.D.Silva, M. Duvergervanbogaert, E. Fonti, H.R. Glatt, P. Isern,
A. Laires, A. Leonard, M. Llagostera, P. Mossesso, A.T.Natarajan, F. Palitti, A.S. Rodrigues, A. Schinoppi, G. Turchi,G. Werleschneider, Development and validation of alternativemetabolic systems for mutagenicity testing in short-termassays, Mutat. Res. 353 (1996) 151–176.
[34] M.O. Bradley, L.C. Erickson, Comparison of the effects ofhydrogen peroxide and x-ray irradiation on toxicity, mutation,and DNA damage/repair in mammalian cells (V-79), Biochim.Biophys. Acta 654 (1981) 135–141.
[35] A.R. Collins, A.G. Ma, S.J. Duthie, The kinetics of repairof oxidative DNA damage (strand breaks and oxidisedpyrimidines) in human cells, Mutat. Res. 336 (1995) 69–77.




















