
Genetic differentiation of Elysia timida (Risso, 1818) populations in the Southwest Mediterranean...
14
Genetic differentiation of Elysia timida (Risso, 1818) populations in the Southwest Mediterranean and Mar Menor coastal lagoon Mercedes Gonza ´lez-Wangu ¨emert a, * , Francisca Gime ´nez-Casalduero b , Angel Pe ´rez-Ruzafa a a Departamento de Ecologı ´a e Hidrologı ´a, Facultad de Biologı ´a, Universidad de Murcia, Campus de Espinardo, 30100 Murcia, Spain b Departamento de Ciencias del Mar y Biologı ´a Aplicada, Universidad de Alicante, Campus de San Vicente de Raspeig, Ap. 99, E-03080 Alicante, Spain Received 23 July 2005; accepted 22 December 2005 Abstract Genetic variation at 10 enzyme loci was analysed in Elysia timida sacoglossan mollusc samples, originating from both coastal lagoon and marine sites. The observed heterozygosity ranged from 0.390 (Los Urrutias) to 0.277 (Tabarca). Marine and coastal lagoon populations were characterised by exclusive alleles. Examination of the spatial structure was performed using Nei’s distances and F-statistics, and indicated significant genetic dif- ferences between marine groups and coastal lagoon populations. Multivariate analysis of allele frequencies also suggested that dif- ferent arrays of genotypes prevail in coastal lagoon samples compared to marine ones. Ó 2006 Elsevier Ltd. All rights reserved. Keywords: Elysia timida; Population genetics; Gene flow; Coastal lagoon; Allozymes 1. Introduction Estuaries and coastal lagoons constitute transitional environments, whose main characteristic is the instability of their physicalechemical parameters, mainly the concentration of salt (Cognetti and Maltagliati, 2000; Gamito et al., 2005). Physiological adaptations are very important for survival in these extreme environments. These adaptations imply a high energetic cost that is reflected in growth and reproduction rates, thus, genetic selection aimed at mini- mizing such costs can be assumed (Wright, 1977; Remmert, 1988). * Corresponding author. Present address: Centro de Investigac ¸~ ao em Biodiversidade e Recursos Gene ´ticos (CIBIO), Universidade do Porto, Campus Agra ´rio de Vair~ ao, R. Monte-Crasto, 4485-661 Porto, Portugal. Tel.: þ351 252 660 416; fax: þ351 252 661 780. E-mail addresses: [email protected], [email protected] (M. Gonza ´lez-Wangu ¨emert), [email protected] (F. Gime ´nez-Casalduero), [email protected] ( A. Pe ´rez-Ruzafa). 0305-1978/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.bse.2005.12.009 Biochemical Systematics and Ecology 34 (2006) 514e527 www.elsevier.com/locate/biochemsyseco
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Genetic differentiation of Elysia timida (Risso, 1818) populations in the Southwest Mediterranean...

Biochemical Systematics and Ecology 34 (2006) 514e527www.elsevier.com/locate/biochemsyseco
Genetic differentiation of Elysia timida (Risso, 1818)populations in the Southwest Mediterranean and
Mar Menor coastal lagoon
Mercedes Gonzalez-Wanguemert a,*, Francisca Gimenez-Casalduero b,�Angel Perez-Ruzafa a
a Departamento de Ecologıa e Hidrologıa, Facultad de Biologıa, Universidad de Murcia, Campus de Espinardo,
30100 Murcia, Spainb Departamento de Ciencias del Mar y Biologıa Aplicada, Universidad de Alicante, Campus de San Vicente de Raspeig,
Ap. 99, E-03080 Alicante, Spain
Received 23 July 2005; accepted 22 December 2005
Abstract
Genetic variation at 10 enzyme loci was analysed in Elysia timida sacoglossan mollusc samples, originating from both coastallagoon and marine sites. The observed heterozygosity ranged from 0.390 (Los Urrutias) to 0.277 (Tabarca). Marine and coastallagoon populations were characterised by exclusive alleles.
Examination of the spatial structure was performed using Nei’s distances and F-statistics, and indicated significant genetic dif-ferences between marine groups and coastal lagoon populations. Multivariate analysis of allele frequencies also suggested that dif-ferent arrays of genotypes prevail in coastal lagoon samples compared to marine ones.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Elysia timida; Population genetics; Gene flow; Coastal lagoon; Allozymes
1. Introduction
Estuaries and coastal lagoons constitute transitional environments, whose main characteristic is the instability oftheir physicalechemical parameters, mainly the concentration of salt (Cognetti and Maltagliati, 2000; Gamito et al.,2005). Physiological adaptations are very important for survival in these extreme environments. These adaptationsimply a high energetic cost that is reflected in growth and reproduction rates, thus, genetic selection aimed at mini-mizing such costs can be assumed (Wright, 1977; Remmert, 1988).
* Corresponding author. Present address: Centro de Investigac~ao em Biodiversidade e Recursos Geneticos (CIBIO), Universidade do Porto,
Campus Agrario de Vair~ao, R. Monte-Crasto, 4485-661 Porto, Portugal. Tel.: þ351 252 660 416; fax: þ351 252 661 780.
E-mail addresses: [email protected], [email protected] (M. Gonzalez-Wanguemert), [email protected] (F. Gimenez-Casalduero),
[email protected] (�A. Perez-Ruzafa).
0305-1978/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.bse.2005.12.009

515M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
Numerous studies based on allozymes and DNA sequence variations have revealed patterns of allelic distributionassociated with clinal variations in environmental factors, such as temperature and salinity (reviewed in Mitton, 1997;Eanes, 1999; Watt, 2000; Veliz et al., 2004). Furthermore, evidence of correlation between changes in allele frequen-cies and different habitat types or heterogeneous environmental parameters has been used to infer the effect of selec-tion in natural populations of molluscs and fishes (Allegrucci et al., 1997; Camilli et al., 2001).
Elysia timida (Risso, 1818) is a gregarious Mediterranean opisthobranch mollusc that lives in shallow waters char-acterised by low-energy hydrodynamic conditions. They inhabit areas covered by either stones or a thick layer of sand(Gimenez-Casalduero et al., 2002) along the Mediterranean, Caribbean and subtropical eastern Atlantic coasts, in-cluding coastal lagoons (Ortea et al., 1997). E. timida is a simultaneous hermaphrodite species with possible self-fertilization under stressful situations (Gimenez-Casalduero, pers. obs.). In the eastern Mediterranean, E. timidahas been described as having direct development (Rahat, 1976). However, other authors have described it as a possiblepoecilogonous species in the western Mediterranean (Gimenez-Casalduero, 1999; Marın and Ros, 2004).
Two morphs of E. timida have been described in the southwest Mediterranean (Gimenez-Casalduero, 1999). Thesemorphs show differences in body size, chlorophyll concentration, colour, number of spawnings per individual, numberof eggs per spawning and larval development. In the aforementioned study, the dun morph was only found in the MarMenor coastal lagoon, close to the natural communication channels with the Mediterranean Sea (Lo Pagan); mean-while the green morph was found both in the Mediterranean Sea and the Mar Menor lagoon.
This study investigates the relationship between genetic structure and habitat type in E. timida, estimating the de-gree of allozyme genetic divergence between coastal lagoon and adjacent marine populations.
2. Material and methods
2.1. Study areas and sampling
A total of 192 specimens of E. timida (Table 1) were collected in the period 2001e2002 at three localities: MarMenor lagoon, Tabarca island and Mazarron. Two sites, hundreds of metres apart, were sampled for each locality(Fig. 1).
Specimens were transferred live to the laboratory. They were classified according to the site where they werecollected.
2.2. Electrophoresis
Sea slugs feed by slicing or puncturing algal cells and sucking out the cell contents. Sea slugs can extract a largenumber of chloroplasts with minimal perturbation simply by puncturing the thin cell wall with their specially adaptedradular tooth and sucking out the contents (Jensen, 1993). All of the contents are discarded except for the chloroplasts,which are engulfed phagocytically into the cells (Rumpho et al., 2000). For this reason, the rinophores were chosen asa homogenate tissue in order to avoid the possible contamination of the symbiotic chloroplasts in the mantle. Also, theenzymes that were chosen to run the electrophoresis do not exist in chloroplasts.
The rinophores, which were extracted from each specimen and stored at �70 �C, were homogenised in an equalvolume of TriseCitrate buffer (pH 9). The homogenates were centrifuged at 7000 g for 12 min at 4 �C. The super-natant was stored at �80 �C. Electrophoresis was performed on starch-gel using four buffers [TriseBorateeEDTA8.6 (TBE); TriseCitrate 8.0 (TC); TriseCitrateeEDTA 7.0 (TCE); and TriseMaleate 7.4 (TM)] according to Harrisand Hopkinson (1976) and Aebersold et al. (1987). The following 10 loci were screened (nomenclature according toShakle et al., 1990): glucose phosphate isomerase, EC 5.3.1.9 (GPI*, TM 7.4); phosphoglucomutase, EC 5.4.2.1(PGM-1* and PGM-2*; TM 7.4); phosphogluconate dehydrogenase, EC 1.1.1.44 (PGDH*; TBE 8.6); isocitratedehydrogenase, EC 1.1.1.42 (IDH*; TCE 7.0); aspartate amino transferase, EC 2.6.1.1 (AAT-1*; TBE 8.6); malatedehydrogenase, EC 1.1.1.37 (MDH-1* and MDH-2*; TM 7.4); and leucine aminopeptidase, EC 3.4.11 (LAP-1*and LAP-2*; TC 8.0).
For the electrophoretic runs and staining, slight modifications in the procedures according to Harris and Hopkinson(1976) and Aebersold et al. (1987) were applied. Isozymes coded by separate loci were numbered in the order ofdecreasing anodal mobility. The most common allele was given the value 100, whereas slower or faster bands, rep-resenting other alleles, were given lower or higher numbers, respectively (Mamuris et al., 1998).
516 M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
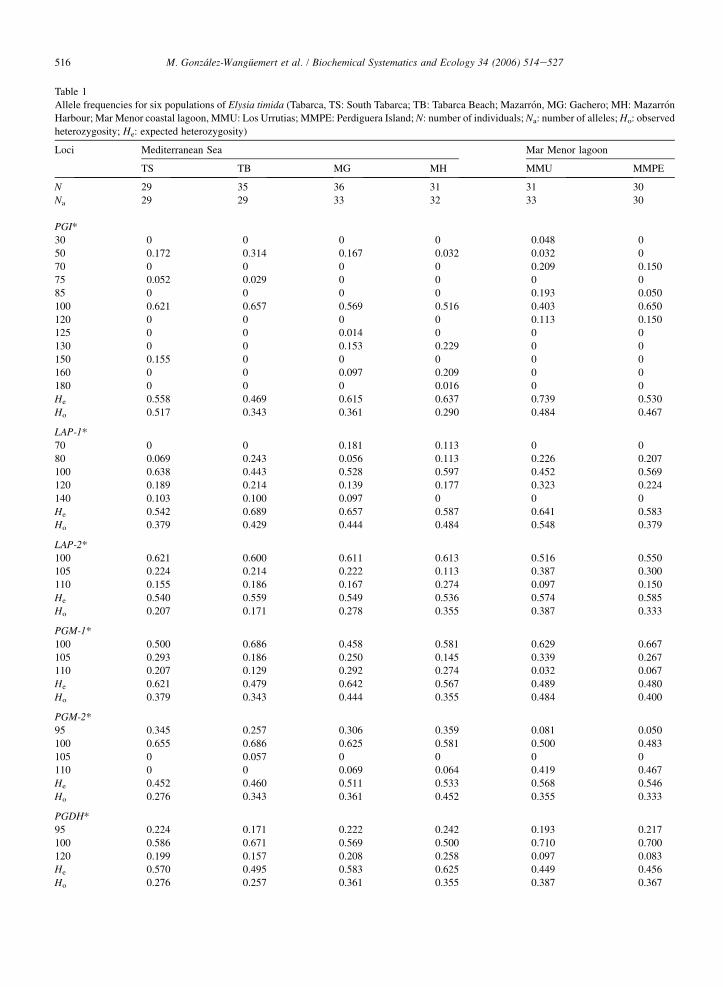
Table 1
Allele frequencies for six populations of Elysia timida (Tabarca, TS: South Tabarca; TB: Tabarca Beach; Mazarron, MG: Gachero; MH: Mazarron
Harbour; Mar Menor coastal lagoon, MMU: Los Urrutias; MMPE: Perdiguera Island; N: number of individuals; Na: number of alleles; Ho: observed
heterozygosity; He: expected heterozygosity)
Loci Mediterranean Sea Mar Menor lagoon
TS TB MG MH MMU MMPE
N 29 35 36 31 31 30
Na 29 29 33 32 33 30
PGI*
30 0 0 0 0 0.048 0
50 0.172 0.314 0.167 0.032 0.032 0
70 0 0 0 0 0.209 0.150
75 0.052 0.029 0 0 0 0
85 0 0 0 0 0.193 0.050
100 0.621 0.657 0.569 0.516 0.403 0.650
120 0 0 0 0 0.113 0.150
125 0 0 0.014 0 0 0
130 0 0 0.153 0.229 0 0
150 0.155 0 0 0 0 0
160 0 0 0.097 0.209 0 0
180 0 0 0 0.016 0 0
He 0.558 0.469 0.615 0.637 0.739 0.530
Ho 0.517 0.343 0.361 0.290 0.484 0.467
LAP-1*
70 0 0 0.181 0.113 0 0
80 0.069 0.243 0.056 0.113 0.226 0.207
100 0.638 0.443 0.528 0.597 0.452 0.569
120 0.189 0.214 0.139 0.177 0.323 0.224
140 0.103 0.100 0.097 0 0 0
He 0.542 0.689 0.657 0.587 0.641 0.583
Ho 0.379 0.429 0.444 0.484 0.548 0.379
LAP-2*
100 0.621 0.600 0.611 0.613 0.516 0.550
105 0.224 0.214 0.222 0.113 0.387 0.300
110 0.155 0.186 0.167 0.274 0.097 0.150
He 0.540 0.559 0.549 0.536 0.574 0.585
Ho 0.207 0.171 0.278 0.355 0.387 0.333
PGM-1*
100 0.500 0.686 0.458 0.581 0.629 0.667
105 0.293 0.186 0.250 0.145 0.339 0.267
110 0.207 0.129 0.292 0.274 0.032 0.067
He 0.621 0.479 0.642 0.567 0.489 0.480
Ho 0.379 0.343 0.444 0.355 0.484 0.400
PGM-2*
95 0.345 0.257 0.306 0.359 0.081 0.050
100 0.655 0.686 0.625 0.581 0.500 0.483
105 0 0.057 0 0 0 0
110 0 0 0.069 0.064 0.419 0.467
He 0.452 0.460 0.511 0.533 0.568 0.546
Ho 0.276 0.343 0.361 0.452 0.355 0.333
PGDH*
95 0.224 0.171 0.222 0.242 0.193 0.217
100 0.586 0.671 0.569 0.500 0.710 0.700
120 0.199 0.157 0.208 0.258 0.097 0.083
He 0.570 0.495 0.583 0.625 0.449 0.456
Ho 0.276 0.257 0.361 0.355 0.387 0.367

517M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
2.3. Population genetic analysis
Genetic variability between marine and coastal lagoon populations was recorded as observed (Ho) and expected(He) mean heterozygosity, the proportion of polymorphic loci at the 5% and 1% criterion (P), and the mean numberof alleles per locus (A).
Genotypic frequencies were tested for conformity with HardyeWeinberg expectations. Probabilities of random de-parture from the HardyeWeinberg equilibrium and significance of divergence among groups were calculated usingthe ‘‘exact test’’ method (Raymond and Rousset, 1995a).
F-statistics according to Wright (1951) were applied in order to quantify both the within-sample and between-samplegenetic heterogeneities (FIS and FST). Both statistics were calculated by the Weir and Cockerham (1984) method. Un-biased estimators were obtained by jack-knifing across loci and tested for difference from zero via permutation tests.
Genetic distance (Nei, 1972) was computed between pairwise samples and the resulting matrix was clustered using theEuclidean distance and the unweighted pair-group method of arithmetic averages (UPGMA) algorithm (Statistica, version6.0). Probabilities of random departure from zero for Nei’s values, according to the null hypothesis, were read directlyfrom the distribution of 1000 randomised matrices computed by permutation. Nei’s D were associated with geographicdistances measured as the shortest marine route between the locations sampled. Also, correlation between geographicand genetic distances was tested using Mantel’s test for association between two parameters in data matrices with internalcorrelation (Mantel, 1967). Probabilities were read directly from the distribution of 1000 randomised matrices computedby permutation. Mantel’s test was performed using the MANTEL procedure from the ‘Genetix’ package.
Data were analysed using GENEPOP software (Raymond and Rousset, 1995b) and the GENETIX package(Bonhomme et al., 1993).
Genetic differences were also compared in a multivariate ordination, using principal component analysis (PCA)on the allele frequencies. These analyses have been described by She et al. (1987) and were calculated using theCANOCO package.
Table 1 (continued )
Loci Mediterranean Sea Mar Menor lagoon
TS TB MG MH MMU MMPE
IDH*
80 0.172 0.200 0.208 0.193 0.193 0.067
92 0 0 0 0 0.210 0.283
96 0.190 0.229 0.194 0.129 0.016 0
100 0.638 0.571 0.597 0.677 0.581 0.650
He 0.527 0.581 0.562 0.487 0.581 0.493
Ho 0.207 0.257 0.472 0.290 0.419 0.500
MDH-1*
90 0 0 0 0 0.161 0.250
100 0.552 0.529 0.667 0.661 0.693 0.650
120 0.448 0.471 0.333 0.339 0.145 0.100
He 0.495 0.498 0.444 0.448 0.472 0.505
Ho 0.276 0.257 0.389 0.419 0.226 0.400
MDH-2*
100 0.517 0.729 0.736 0.774 0.581 0.583
110 0.483 0.271 0.167 0.048 0.419 0.417
115 0 0 0.097 0.177 0 0
He 0.499 0.395 0.421 0.367 0.487 0.486
Ho 0.276 0.143 0.361 0.323 0.323 0.300
AAT*92 0 0 0 0 0.113 0.183
98 0.069 0.200 0.153 0.097 0 0
100 0.741 0.657 0.625 0.548 0.645 0.667
108 0.190 0.143 0.222 0.355 0.242 0.150
He 0.410 0.508 0.537 0.564 0.512 0.499
Ho 0.172 0.229 0.250 0.258 0.290 0.233

518 M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
Gene flow among samples was estimated as the number of migrants exchanged between populations per generationat equilibrium (Nem). Values for Nem were derived from one approach with FST values, following Wright’s (1951)island model with a small level of migration, where:
Nem¼ ð1�FSTÞ=4FST
To study the genetic differences among morphs, a factorial correspondence analysis (FCA) was carried out. This typeof canonical analysis is the correct one for describing the associations between qualitative variables (She et al., 1987).
In order to determine the most likely geographic origin of each E. timida morph, and assign the stock origin ofa sacoglossan using genotype data only, individual probabilities described by Cornuet et al. (1999) were used. Usingthe Bayesian method (GENECLASS package), the probability for all individuals was calculated with a threshold of0.01. Individuals were then assigned to the populations for which the highest probability was found.
2.4. Morphological analysis
To compare lengths for dun and green morphs of E. timida, a variance analysis (ANOVA) was carried out for in-dividuals from Tabarca Beach locality. A p-value of 0.05 or less was considered statistically significant. This localitywas chosen because all individuals (dun and green) were sampled at the same time and there were a similar number ofspecimens for each morph.
3. Results
3.1. Allozyme variation
Electrophoresis performed on seven enzymes gave a total of 10 polymorphic loci. Allele richness in E. timida pop-ulations ranged from 29 alleles in Los Urrutias to 33 alleles in Gachero (Table 1). The sampled populations showedsome exclusive alleles (PGI*30, Los Urrutias; PGI*125, Gachero; PGI*150, South Tabarca; PGI*180, Mazarron and
TB∗∗
∗ ∗
∗∗
TS
MMU
MMPE
MH
MG
Mediterranean Sea
37º26’N1º29’W 1º07’W 0º28’E
38º14’N
Mazarrón
Mar Menor
Tabarca
Fig. 1. Map of the six sites studied in the SW Mediterranean and the Mar Menor.

519M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
PGM-2*105, Tabarca Beach). Some alleles characterised the coastal lagoon populations such as PGI*70, PGI*85,PGI*120, IDH*92, MDH-1*90 and AAT*92; while all Mediterranean populations showed only one exclusive allele(AAT*98).
All loci were polymorphic (p� 0.99) (Table 2). The average number of alleles per population (A) varied from 3.30for Los Urrutias and Gachero to 2.90 for Tabarca. Heterozygosity values revealed a high degree of within-sample ge-netic variability. Observed heterozygosity (Ho) ranged from 0.277 for the Tabarca Beach population to 0.390 for LosUrrutias.
Genic and genotypic differentiation was detected among all populations. The PGI* locus showed significant genicdifferentiation for all population pairs (p< 0.05). Allelic frequencies between Los Urrutias and Perdiguera Island(both coastal lagoon sites) are very similar and show no significant differences at nine loci. The loci LAP-2* andPGDH* show no significant differences among populations, except for Mazarron harbour in relation to the two coastallagoon populations (Los Urrutias and Perdiguera Island). Seven loci (LAP-1*, PGM-1*, PGM-2*, IDH*, MDH-1*,MDH-2* and AAT*) also had differences in allele frequencies between coastal lagoon and marine samples. Genotypefrequencies among the six populations are not randomly distributed, only the PGDH* locus shows significant differ-ences among populations.
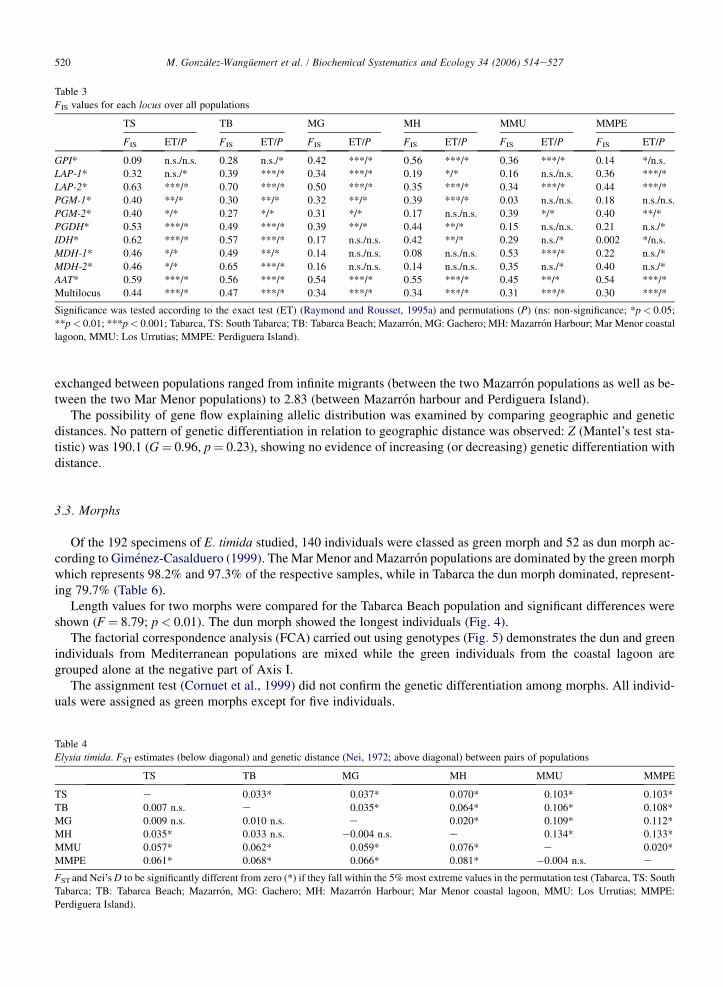
The positive and significant FIS values indicate a general departure from the HardyeWeinberg equilibrium due toa heterozygote deficit (Table 3). Overall FIS (Weir and Cockerham, 1984) was 0.368 with a 95% confidence intervalusing bootstrapping over loci from 0.322 to 0.422. These values indicate a higher heterozygote deficit for marine sam-ples than for lagoon ones. FIS values per locus ranged from 0.002 to 0.701 and no loci showed heterozygote excess.
3.2. Genetic differentiation
FST (Weir and Cockerham, 1984) was 0.041 with a 95% confidence interval using bootstrapping over loci from0.021 to 0.063. Table 4 reports FST estimates between pairs of populations for which values ranged from �0.004, be-tween the two Mazarron samples, to 0.081 between Perdiguera Island and Mazarron harbour. Some allozyme loci suchas PGM-2*, PGI*, MDH-2* and MDH-1* showed greater genetic differentiation from other loci and would be respon-sible for the FST differences.
Nei’s genetic distances (Nei, 1972) were estimated using 10 polymorphic loci (Table 4). Values ranged from 0.020to 0.134, with all distances being significant at the 0.05 level. Lowest D-values were obtained between populationsfrom each locality and the highest value (D¼ 0.134) was shown between coastal lagoon and marine samples.
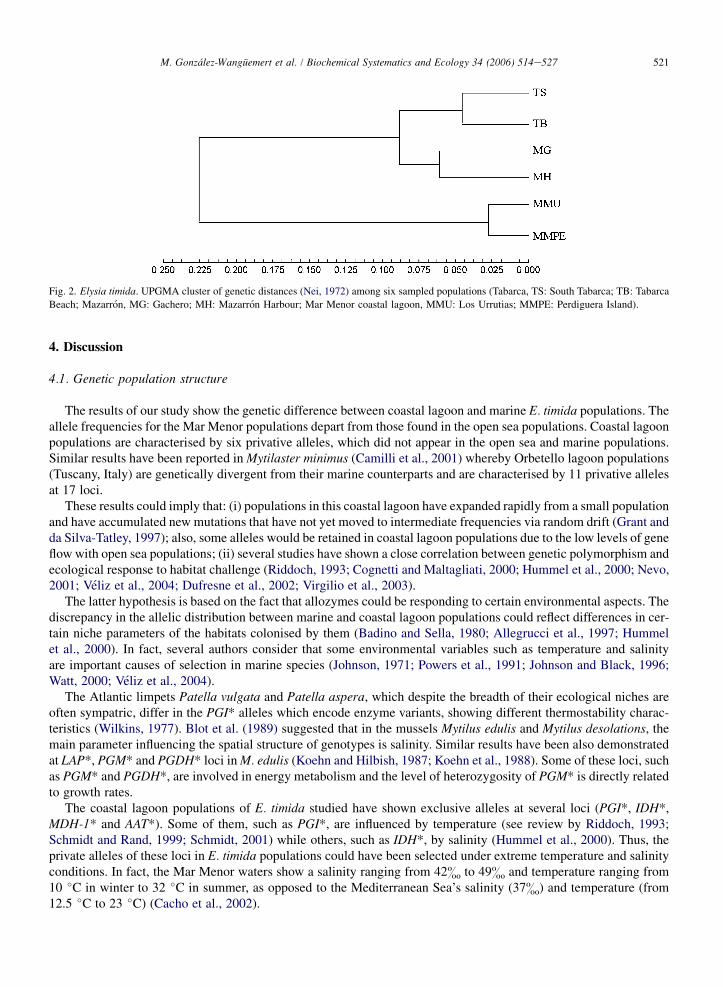
The levels of genetic distance between coastal lagoon and marine populations of E. timida are highlighted by theUPGMA dendrogram (Fig. 2). The mean genetic distance between coastal lagoon and marine populations was 0.113.Coastal lagoon populations (Los Urrutias and Perdiguera Island) are grouped in the same cluster and the marine sam-ples in two separate sub-groups.
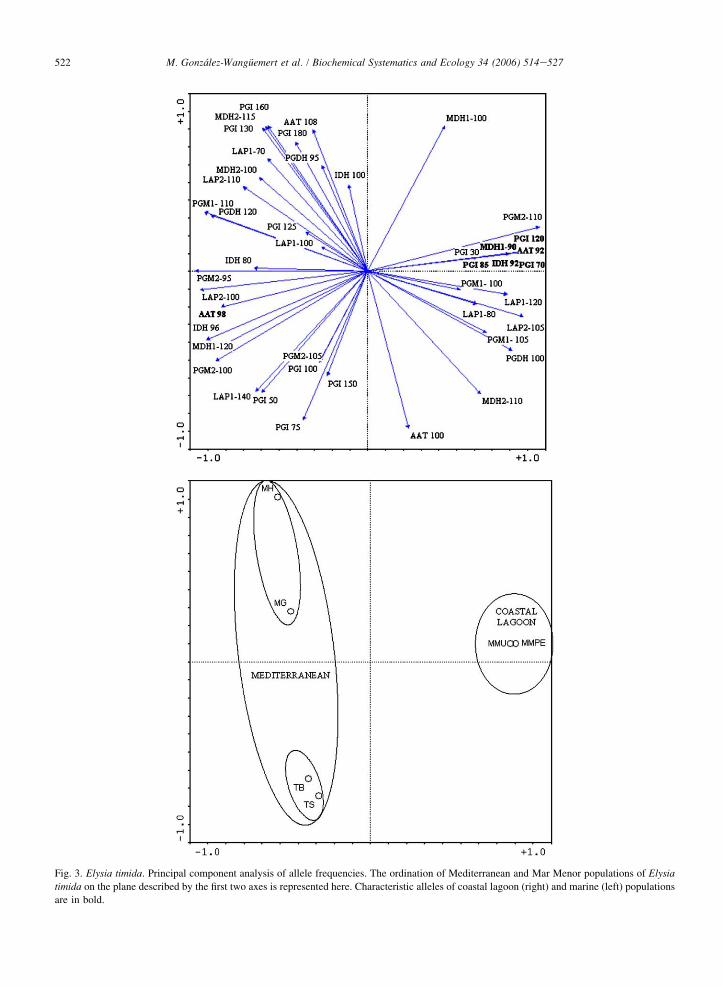
Further analysis on the spatial structuring of genetic diversity was attempted by means of a principal componentanalysis carried out on allele frequencies (Fig. 3). When all alleles are considered, the first axis, which explains 82.6%of the total variance, clearly separates the coastal lagoon samples in the positive part from the marine samples in thenegative part.
Assuming equilibrium between genetic drift and migration, the number of migrants (Nem) per generation was cal-culated using the FST values and according to the island model (Table 5). Estimates of the number of migrants
Table 2
Description of genetic diversity in sampled populations (TS: South Tabarca; TB: Tabarca Beach; MG: Gachero; MH: Mazarron Harbour; MMU: Los
Urrutias; MMPE: Perdiguera Island)
Population N AT P95% P99% Ho� SE He� SE FIS
TS 29 2.90 1.00 1.00 0.297� 0.10 0.521� 0.06 0.445
TB 35 2.90 1.00 1.00 0.277� 0.09 0.513� 0.08 0.472
MG 36 3.30 1.00 1.00 0.372� 0.07 0.552� 0.08 0.338
MH 31 3.20 1.00 1.00 0.358� 0.07 0.535� 0.08 0.345
MMU 31 3.30 1.00 1.00 0.390� 0.09 0.551� 0.09 0.307
MMPE 30 3.00 1.00 1.00 0.372� 0.08 0.516� 0.04 0.301
N: number of individuals analysed; AT: average number of alleles; P95% and P99%: polymorphism; Ho: observed heterozygosity; He: expected het-
erozygosity; SE: standard error.
520 M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
exchanged between populations ranged from infinite migrants (between the two Mazarron populations as well as be-tween the two Mar Menor populations) to 2.83 (between Mazarron harbour and Perdiguera Island).
The possibility of gene flow explaining allelic distribution was examined by comparing geographic and geneticdistances. No pattern of genetic differentiation in relation to geographic distance was observed: Z (Mantel’s test sta-tistic) was 190.1 (G¼ 0.96, p¼ 0.23), showing no evidence of increasing (or decreasing) genetic differentiation withdistance.
3.3. Morphs
Of the 192 specimens of E. timida studied, 140 individuals were classed as green morph and 52 as dun morph ac-cording to Gimenez-Casalduero (1999). The Mar Menor and Mazarron populations are dominated by the green morphwhich represents 98.2% and 97.3% of the respective samples, while in Tabarca the dun morph dominated, represent-ing 79.7% (Table 6).
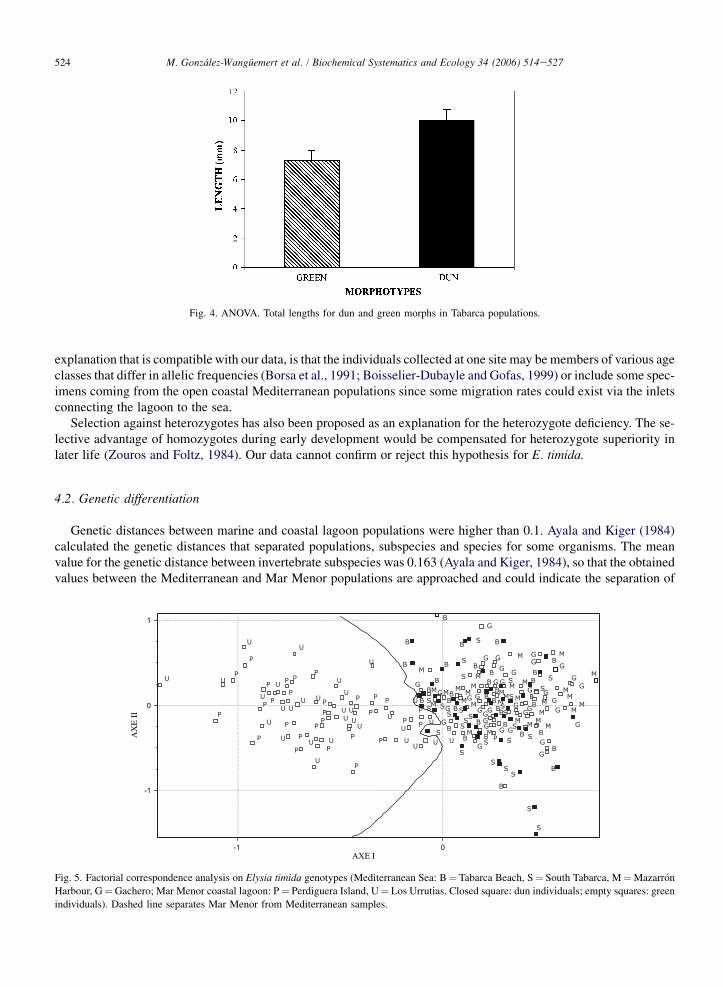
Length values for two morphs were compared for the Tabarca Beach population and significant differences wereshown (F¼ 8.79; p< 0.01). The dun morph showed the longest individuals (Fig. 4).
The factorial correspondence analysis (FCA) carried out using genotypes (Fig. 5) demonstrates the dun and greenindividuals from Mediterranean populations are mixed while the green individuals from the coastal lagoon aregrouped alone at the negative part of Axis I.
The assignment test (Cornuet et al., 1999) did not confirm the genetic differentiation among morphs. All individ-uals were assigned as green morphs except for five individuals.
Table 3
FIS values for each locus over all populations
TS TB MG MH MMU MMPE
FIS ET/P FIS ET/P FIS ET/P FIS ET/P FIS ET/P FIS ET/P
GPI* 0.09 n.s./n.s. 0.28 n.s./* 0.42 ***/* 0.56 ***/* 0.36 ***/* 0.14 */n.s.
LAP-1* 0.32 n.s./* 0.39 ***/* 0.34 ***/* 0.19 */* 0.16 n.s./n.s. 0.36 ***/*
LAP-2* 0.63 ***/* 0.70 ***/* 0.50 ***/* 0.35 ***/* 0.34 ***/* 0.44 ***/*
PGM-1* 0.40 **/* 0.30 **/* 0.32 **/* 0.39 ***/* 0.03 n.s./n.s. 0.18 n.s./n.s.
PGM-2* 0.40 */* 0.27 */* 0.31 */* 0.17 n.s./n.s. 0.39 */* 0.40 **/*
PGDH* 0.53 ***/* 0.49 ***/* 0.39 **/* 0.44 **/* 0.15 n.s./n.s. 0.21 n.s./*
IDH* 0.62 ***/* 0.57 ***/* 0.17 n.s./n.s. 0.42 **/* 0.29 n.s./* 0.002 */n.s.
MDH-1* 0.46 */* 0.49 **/* 0.14 n.s./n.s. 0.08 n.s./n.s. 0.53 ***/* 0.22 n.s./*
MDH-2* 0.46 */* 0.65 ***/* 0.16 n.s./n.s. 0.14 n.s./n.s. 0.35 n.s./* 0.40 n.s./*
AAT* 0.59 ***/* 0.56 ***/* 0.54 ***/* 0.55 ***/* 0.45 **/* 0.54 ***/*
Multilocus 0.44 ***/* 0.47 ***/* 0.34 ***/* 0.34 ***/* 0.31 ***/* 0.30 ***/*
Significance was tested according to the exact test (ET) (Raymond and Rousset, 1995a) and permutations (P) (ns: non-significance; *p< 0.05;
**p< 0.01; ***p< 0.001; Tabarca, TS: South Tabarca; TB: Tabarca Beach; Mazarron, MG: Gachero; MH: Mazarron Harbour; Mar Menor coastal
lagoon, MMU: Los Urrutias; MMPE: Perdiguera Island).
Table 4
Elysia timida. FST estimates (below diagonal) and genetic distance (Nei, 1972; above diagonal) between pairs of populations
TS TB MG MH MMU MMPE
TS e 0.033* 0.037* 0.070* 0.103* 0.103*
TB 0.007 n.s. e 0.035* 0.064* 0.106* 0.108*
MG 0.009 n.s. 0.010 n.s. e 0.020* 0.109* 0.112*
MH 0.035* 0.033 n.s. �0.004 n.s. e 0.134* 0.133*
MMU 0.057* 0.062* 0.059* 0.076* e 0.020*
MMPE 0.061* 0.068* 0.066* 0.081* �0.004 n.s. e
FST and Nei’s D to be significantly different from zero (*) if they fall within the 5% most extreme values in the permutation test (Tabarca, TS: South
Tabarca; TB: Tabarca Beach; Mazarron, MG: Gachero; MH: Mazarron Harbour; Mar Menor coastal lagoon, MMU: Los Urrutias; MMPE:
Perdiguera Island).
521M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
4. Discussion
4.1. Genetic population structure
The results of our study show the genetic difference between coastal lagoon and marine E. timida populations. Theallele frequencies for the Mar Menor populations depart from those found in the open sea populations. Coastal lagoonpopulations are characterised by six privative alleles, which did not appear in the open sea and marine populations.Similar results have been reported in Mytilaster minimus (Camilli et al., 2001) whereby Orbetello lagoon populations(Tuscany, Italy) are genetically divergent from their marine counterparts and are characterised by 11 privative allelesat 17 loci.
These results could imply that: (i) populations in this coastal lagoon have expanded rapidly from a small populationand have accumulated new mutations that have not yet moved to intermediate frequencies via random drift (Grant andda Silva-Tatley, 1997); also, some alleles would be retained in coastal lagoon populations due to the low levels of geneflow with open sea populations; (ii) several studies have shown a close correlation between genetic polymorphism andecological response to habitat challenge (Riddoch, 1993; Cognetti and Maltagliati, 2000; Hummel et al., 2000; Nevo,2001; Veliz et al., 2004; Dufresne et al., 2002; Virgilio et al., 2003).
The latter hypothesis is based on the fact that allozymes could be responding to certain environmental aspects. Thediscrepancy in the allelic distribution between marine and coastal lagoon populations could reflect differences in cer-tain niche parameters of the habitats colonised by them (Badino and Sella, 1980; Allegrucci et al., 1997; Hummelet al., 2000). In fact, several authors consider that some environmental variables such as temperature and salinityare important causes of selection in marine species (Johnson, 1971; Powers et al., 1991; Johnson and Black, 1996;Watt, 2000; Veliz et al., 2004).
The Atlantic limpets Patella vulgata and Patella aspera, which despite the breadth of their ecological niches areoften sympatric, differ in the PGI* alleles which encode enzyme variants, showing different thermostability charac-teristics (Wilkins, 1977). Blot et al. (1989) suggested that in the mussels Mytilus edulis and Mytilus desolations, themain parameter influencing the spatial structure of genotypes is salinity. Similar results have been also demonstratedat LAP*, PGM* and PGDH* loci in M. edulis (Koehn and Hilbish, 1987; Koehn et al., 1988). Some of these loci, suchas PGM* and PGDH*, are involved in energy metabolism and the level of heterozygosity of PGM* is directly relatedto growth rates.
The coastal lagoon populations of E. timida studied have shown exclusive alleles at several loci (PGI*, IDH*,MDH-1* and AAT*). Some of them, such as PGI*, are influenced by temperature (see review by Riddoch, 1993;Schmidt and Rand, 1999; Schmidt, 2001) while others, such as IDH*, by salinity (Hummel et al., 2000). Thus, theprivate alleles of these loci in E. timida populations could have been selected under extreme temperature and salinityconditions. In fact, the Mar Menor waters show a salinity ranging from 42& to 49& and temperature ranging from10 �C in winter to 32 �C in summer, as opposed to the Mediterranean Sea’s salinity (37&) and temperature (from12.5 �C to 23 �C) (Cacho et al., 2002).
Fig. 2. Elysia timida. UPGMA cluster of genetic distances (Nei, 1972) among six sampled populations (Tabarca, TS: South Tabarca; TB: Tabarca
Beach; Mazarron, MG: Gachero; MH: Mazarron Harbour; Mar Menor coastal lagoon, MMU: Los Urrutias; MMPE: Perdiguera Island).
522 M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
Fig. 3. Elysia timida. Principal component analysis of allele frequencies. The ordination of Mediterranean and Mar Menor populations of Elysia
timida on the plane described by the first two axes is represented here. Characteristic alleles of coastal lagoon (right) and marine (left) populations
are in bold.

523M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
Furthermore, these differences have been higher over the lagoon’s recent history, reaching 53& salinity and a ther-mal range from 7.5 �C to 29 �C (Perez-Ruzafa, 1989; Perez-Ruzafa et al., 1991).
E. timida populations showed mean observed heterozygosity values higher than those reported in other molluscstudies (Wilkins, 1977; Tatarenkov, 1995; Gajardo et al., 2002). Some authors (Badino and Sella, 1980; Ignacioet al., 2000; Johnson et al., 2001; Mariani et al., 2002) found values of similar heterozygosity in populations ofP. aspera (Ho¼ 0.437), Crassostrea rhizophorae (Ho¼ 0.280), Siphonaria kurracheensis (Ho¼ 0.239) and Cerasto-derma glaucum (Ho¼ 0.270).
Furthermore, coastal lagoon populations of E. timida showed higher heterozygosity than marine populations. Val-entine (1976) suggested that species inhabiting temporally variable or spatially heterogeneous environments exhibitedhigher levels of genetic variability than those from less variable or more monotonous environments. Veliz et al. (2004)affirm that the ability of organisms to maximize their fitness in a new environment depends on the existence of var-iation in ecologically relevant genes. Species living in coastal lagoons often have to cope with heterogeneous envi-ronments that differ both in time and space. In such cases, it is expected that balancing selection will maintaingenetic polymorphism in genes that differentially affect fitness in contrasting environments (Maynard, 1998; Velizet al., 2004). E. timida seems to confirm this hypothesis in agreement with the results obtained for other mollusc spe-cies such as Crassostrea angulata, M. edulis and M. minimus (Michinina and Rebordinos, 1997; Raymond et al., 1997;Camilli et al., 2001).
Despite their high heterozygosity, E. timida populations presented a heterozygote deficit. Departure from HardyeWeinberg expectations, expressed as a deficiency of heterozygous phenotypes, is common in marine invertebrates(Arculeo et al., 1998; Boisselier-Dubayle and Gofas, 1999). Explanations for this may take into account inbreedingresulting from fertilization between relatives, as E. timida is a gregarious species. Also, self-fertilization could clearlycontribute to heterozygote deficiency in a natural population. In fact, there is some evidence of sporadic self-fertiliza-tion in Physa heterostropha and other pulmonate species (Rollison et al., 1989; Jarne et al., 1993; Dillon and Wething-ton, 1994) which showed an important heterozygote deficit. This process could occur in E. timida populations asa survival strategy under extreme conditions.
Electrophoretic artefacts such as the misclassification of heterozygotes as homozygotes (null alleles) could be an-other cause of heterozygote deficit, although our data do not provide any evidence for the presence of null alleles.Another possible explanation for heterozygote deficiencies is the spatial and temporal Wahlund effect, particularlyfor species in which allele frequencies change dramatically over small geographic distances. A hypothetical
Table 5
Estimates of Nem using FST values (Tabarca, TS: South Tabarca; TB: Tabarca Beach; Mazarron, MG: Gachero; MH: Mazarron Harbour; Mar Menor
coastal lagoon, MMU: Los Urrutias; MMPE: Perdiguera Island)
TB MG MH MMU MMPE
TS 36.93 27.53 6.92 4.14 3.82
TB 27.53 6.92 4.14 3.82
MG N 4.01 3.55
MH 3.05 2.83
MMU N
Table 6
Number of observed individuals of different morphs (TS: South Tabarca; TB: Tabarca Beach; MG: Gachero; MH: Mazarron Harbour; MMU: Los
Urrutias; MMPE: Perdiguera Island)
Locality Site Total Morphs (%)
Green Dun
Tabarca South Tabarca (TS) 29 7 93
Tabarca Beach (TB) 35 37 63
Mazarron Gachero (MG) 36 97 3
Mazarron Harbour (MH) 31 97 3
Mar Menor Los Urrutias (MMU) 31 100 0
Perdiguera Island (MMPE) 30 97 3
524 M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
explanation that is compatible with our data, is that the individuals collected at one site may be members of various ageclasses that differ in allelic frequencies (Borsa et al., 1991; Boisselier-Dubayle and Gofas, 1999) or include some spec-imens coming from the open coastal Mediterranean populations since some migration rates could exist via the inletsconnecting the lagoon to the sea.
Selection against heterozygotes has also been proposed as an explanation for the heterozygote deficiency. The se-lective advantage of homozygotes during early development would be compensated for heterozygote superiority inlater life (Zouros and Foltz, 1984). Our data cannot confirm or reject this hypothesis for E. timida.
4.2. Genetic differentiation
Genetic distances between marine and coastal lagoon populations were higher than 0.1. Ayala and Kiger (1984)calculated the genetic distances that separated populations, subspecies and species for some organisms. The meanvalue for the genetic distance between invertebrate subspecies was 0.163 (Ayala and Kiger, 1984), so that the obtainedvalues between the Mediterranean and Mar Menor populations are approached and could indicate the separation of
Fig. 4. ANOVA. Total lengths for dun and green morphs in Tabarca populations.
AXE I0-1
AX
E I
I
1
0
-1
Fig. 5. Factorial correspondence analysis on Elysia timida genotypes (Mediterranean Sea: B¼ Tabarca Beach, S¼ South Tabarca, M¼Mazarron
Harbour, G¼Gachero; Mar Menor coastal lagoon: P¼ Perdiguera Island, U¼ Los Urrutias. Closed square: dun individuals; empty squares: green
individuals). Dashed line separates Mar Menor from Mediterranean samples.

525M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
E. timida. Similar results have been shown for other species such as M. minimus, in which a population, differentiatedin a coastal lagoon, presented a 0.25 genetic distance from the adjacent marine population (Cognetti, 1994; Cognettiand Maltagliati, 2000).
On the other hand, we found that the values for genetic identity between the Mediterranean and coastal lagoon pop-ulations ranged from 0.90 to 0.87. These values indicate a subspecies-rank separation according to several authors(Avise, 1974; Thorpe, 1983; Mariani et al., 2002). In fact, the minimum empirical threshold beyond which twotaxa are likely to represent subspecies is 0.90.
However, it is important to stress that there are different methods of delimiting species which show limitations(Knowlton, 2000; Sites and Marshall, 2003). Future molecular genetic analyses using mitochondrial DNA and micro-satellite markers should help to resolve many of these issues, particularly if coupled with other biological approach.
FST values suggest a moderate genetic differentiation, within marine and coastal lagoon populations, and a highdifferentiation between marine and coastal lagoon populations. These FST values could be due to coastal lagoon pop-ulations being relatively isolated by a gene flow barrier, resulting from the confinement of the lagoon environment.
On the other hand, some authors have described a direct development of E. timida (Gimenez-Casalduero, 1999;Marin and Ros, 2004). Current theories postulate that molluscs with non-planktotrophic development possess limiteddispersal abilities and are therefore subject to greater genetic differentiation among populations (Hoskin, 1997;Boisselier-Dubayle and Gofas, 1999).
Low gene flow has also been detected between coastal lagoon and marine populations of E. timida. Local currentsand small-scale eddies could retain larvae close to or within parental habitats, reducing or eliminating the possibilityof dispersal from the natal site, just as some authors have described for other mollusc species (Boxshall, 2000; Wareset al., 2001). The low levels of Nem detected may indicate insufficient cohesion among sampled populations, whichcould lead to speciation (Makholela et al., 2003).
4.3. Morphs
Our results shows that no genetic differences exist between the two previously described E. timida morphs. In fact,according to data from Gimenez-Casalduero (1999) and the present work, both morphs can occur either in the Med-iterranean or in Mar Menor populations. Differences in coloration could be related to the feeding preferences of E.timida, or food availability depending on the algae assemblage for each season or locality. In fact, Gimenez-Casal-duero (1999) expressed the possibility of E. timida having greater feeding versatility than assumed by other authorssuch as Marın and Ros (1993). Further studies should be done to confirm these aspects.
5. Conclusion
The present genetic analysis resembles the picture observed in other marine molluscs separated by geographic bor-ders (Quesada et al., 1995; Laudien et al., 2003) and supports the hypothesis that environmental forces may be themajor causal elements determining genetic differentiation (Makholela et al., 2003).
Acknowledgements
We are grateful to C. Muniain and M. Varela for the field assistance. This study received support in the form ofa research grant (Ecologıa y Ordenacion de Ecosistemas Marinos Costeros, University of Murcia) awarded to M.G.W.
References
Aebersold, P.B., Winans, G.A., Teel, D.J., Miller, G.B., Utter, F.M., 1987. Manual for starch gel electrophoresis: a method for detection of genetic
variation. NOAA Tech. Rep. NMFS 61, 1e19.
Allegrucci, G., Fortunato, C., Sbordoni, V., 1997. Genetic structure and allozyme variation of sea bass (Dicentrarchus labrax and D. puctatus) in
the Mediterranean Sea. Mar. Biol. 128, 347e358.
Arculeo, M., Lo Brutto, S., Pancucci, M.P., Cammarata, M., Parrinello, M., 1998. Allozyme similarity in two morphologically distinguishable
populations of Paracentrotus lividus (Echinodermata) from distinct areas of the Mediterranean. J. Mar. Biol. Assoc. 78 (1), 231e238.
Avise, J.C., 1974. Systematic value of electrophoretic data. Syst. Zool. 23, 465e481.
Ayala, F.J., Kiger, J.A., 1984. Genetica moderna. Ediciones Omega, S.A., Barcelona, Espana.

526 M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
Badino, G., Sella, G., 1980. Phosphoglucose isomerase variability in sympatric populations of Mediterranean species of Patella (Gastropoda:
Prosobranchia). Mar. Ecol. 2, 315e320.
Blot, M., Thiriot-Quievreux, S., Soyer, J., 1989. Genetic differences and environments of mussel populations in Kerguelen Islands. Polar Biol. 10,
167e174.
Boisselier-Dubayle, M.C., Gofas, S., 1999. Genetic relationships between marine and marginal-marine populations of Cerithium species from the
Mediterranean Sea. Mar. Biol. 135, 671e682.
Bonhomme, F., Belkhir, K., Borsa, P., Mathieu, E., Roux, M., 1993. GENETIX e Logiciel D’analyse Des Donnees Du Groupe de Genetique Des
Populations de Montpellier. Version 0.1. Universite Montpellier II, France.
Borsa, P., Zainuri, M., Delay, B., 1991. Heterozygote deficiency and population structure in the bivalve Ruditapes decussatus. Heredity 66, 1e8.
Boxshall, A.J., 2000. The importance of flow and settlement cues to larvae of the abalone, Haliotis rufescens Swainson. J. Exp. Mar. Biol. Ecol.
254, 143e167.
Cacho, I., Grimalt, J.O., Canals, M., 2002. Response of the Western Mediterranean Sea to rapid climatic variability during the last 50,000 years:
a molecular biomarker approach. J. Mar. Syst. 33e34, 253e272.
Camilli, L., Castelli, A., Lardicci, C., Maltagliati, F., 2001. Evidence for high levels of genetic divergence between populations of the bivalve
Mytilaster minimus from a brackish environment and two adjacent marine sites. J. Mollus. Stud. 67, 506e510.
Cognetti, G., 1994. Colonization of brackish waters. Mar. Pollut. Bull. 28, 583e586.
Cognetti, G., Maltagliati, F., 2000. Biodiversity and adaptive mechanisms in brackish water fauna. Mar. Pollut. Bull. 40, 7e14.
Cornuet, J.-M., Piry, S., Luikartm, G., Estoup, A., Solignac, M., 1999. New method multilocus genotypes to select or exclude populations as or-
igins of individuals. Genetics 153, 1989e2000.
Dillon, R.T., Wethington, A.R., 1994. Inheritance at five loci in the freshwater snail Physa heterostropha. Biochem. Genet. 32, 75e82.
Dufresne, F., Bourget, E., Bernatchez, L., 2002. Differential patterns of spatial divergence in microsatellite and allozyme alleles: further evidence
for locus-specific selection in the acorn barnacle, Semibalanus balanoides? Mol. Ecol. 11, 113e123.
Eanes, W.F., 1999. Analysis of selection on enzyme polymorphism. Annu. Rev. Ecol. Syst. 30, 301e326.
Gajardo, G., Cancino, J.M., Navarro, J.M., 2002. Genetic variation and population structure in the marine snail Chorus giganteus (Gastropod:
Muricidae), an overexploited endemic resource from Chile. Fish. Res. 55, 329e333.
Gamito, S., Gilabert, J., Marcos, C., Perez-Ruzafa, A., 2005. Effects of changing environmental conditions on lagoon ecology. In: Gonenc, I.E.,
Wolflin, J. (Eds.), Coastal Lagoons: Ecosystem Processes and Modelling for Sustainable Use and Development. CRC Press, Boca Raton, Flor-
ida, pp. 193e229.
Gimenez-Casalduero, F., 1999. Comparative study in different populations of Elysia timida Risso, 1818 (Gastropoda, Opistobranchia, Sacoglossa)
in open sea and lagoon environment. Iberus 17, 137e146.
Gimenez-Casalduero, F., Muniain, C., Garcıa-Charton, J.A., 2002. Elysia timida (Risso, 1818) (Gastropoda, Opistobranchia): relationship and
feeding deterrence to a potential predator on the South-western Mediterranean coast. Mar. Biol. 141, 1051e1057.
Grant, W.S., da Silva-Tatley, F.M., 1997. Lack of genetically-subdivided population structure in Bullia digitalis, a southern African marine
gastropod with lecithotrophic development. Mar. Biol. 129, 123e137.
Harris, H., Hopkinson, D.A., 1976. Handbook of Enzyme Electrophoresis in Human Genetics. American Elsevier, New York, 284 pp.
Hoskin, M.G., 1997. Effects of contrasting modes of larval development on the genetic structures of populations of three species of prosobranch
gastropods. Mar. Biol. 127, 647e656.
Hummel, H., Sokolowski, A., Bogaards, R., Wolowicz, M., 2000. Ecophysiological and genetic traits of the Baltic clam Macoma balthica
in the Baltic: differences between populations in the Gdansk Bay due to acclimatization or genetic adaptation? Int. Rev. Hydrobiol. 85,
621e637.
Ignacio, B.L., Absher, T.M., Lazoski, C., Sole-Cava, A.M., 2000. Genetic evidence of the presence of two species of Crassostrea (Bivalvia:
Ostreidae) on the coast of Brazil. Mar. Biol. 136, 987e991.
Jarne, P., Vianey-Liaud, M., Delay, B., 1993. Selfing and outcrossing in hermaphrodite freshwater gastropods (Basommatophora): where, when
and why. Biol. J. Linn. Soc. 49, 99.
Jensen, K.R., 1993. Morphological adaptations and plasticity of radular teeth of the Sacoglossa (¼Ascoglossa) (Mollusca: Opistobranchia) in
relation to their food plants. Biol. J. Linn. Soc. 48, 135e155.
Johnson, G.B., 1971. The relationship of enzyme polymorphism to metabolic function. Nature 232, 347e348.
Johnson, M.S., Black, R., 1996. Geographic cohesiveness versus associations with habitat genetic subdivision of Bembicium vittatum Philippi
(Gastropoda: Littorinidae) in the Houtman Abrolhos Islands. Biol. J. Linn. Soc. 58, 57e74.
Johnson, M.S., Bentley, S.L., Ford, S.S., Ladyman, M.T., Lambert, G.J., 2001. Effects of a complex archipelago on genetic subdivision of the
intertidal limpet Siphonaria kurracheensis. Mar. Biol. 139, 1087e1094.
Knowlton, N., 2000. Molecular genetic analyses of species boundaries in the sea. Hydrobiologia 420, 73e90.
Koehn, R.K., Hilbish, T.J., 1987. The adaptive importance of genetic variation. Am. Sci. 75, 134e141.
Koehn, R.K., Diehl, W.J., Scott, M.T., 1988. The differential contribution by individual enzymes of glycolisis and protein catabolism to the
relationship between heterozygosity and growth rate in the coot clam, Mulinia lateralis. Genetics 118, 121e130.
Laudien, J., Flint, N.S., van der Bank, F.H., Brey, T., 2003. Genetic and morphological variation in four populations of the surf clam Donax serra
(Roding) from southern African sandy beaches. Biochem. Syst. Ecol. 31, 751e772.
Makholela, T., van der Bank, H., Balkwill, K., 2003. A preliminary study of allozyme variation in three rare and restricted endemic Barleriagreenii (Acanthaceae) populations. Biochem. Syst. Ecol. 31, 141e154.
Mamuris, Z., Apostolidis, A.P., Theodorou, A.J., Triantaphyllidis, C., 1998. Genetic protein variation in red mullet (Mullus barbatus) and striped
red mullet (M. surmuletus) populations from the Mediterranean Sea. Mar. Biol. 130, 353e360.
Mantel, N., 1967. The detection of disease clustering and generalized regression approach. Cancer Res. 27, 209e220.

527M. Gonzalez-Wanguemert et al. / Biochemical Systematics and Ecology 34 (2006) 514e527
Mariani, S., Ketmaier, V., de Matthaeis, E., 2002. Genetic structuring and gene flow in Cerastoderma glaucum (Bivalvia: Cardiidae): evidence
from allozyme variation at different geographic scales. Mar. Biol. 140, 687e697.
Marın, A., Ros, J.D., 1993. Ultrastructural and ecological aspects of the development of chloroplast retention in the sacoglossan gastropod Elysiatimida. J. Molluscan Stud. 59, 95e104.
Marın, A., Ros, J.D., 2004. Chemical defences in Sacoglossan Opistobranchs: taxonomic trends and evolutionary implications. Sci. Mar. 68, 227e
241.
Maynard, S.J., 1998. Evolution in Natural Populations. Oxford Press, Oxford.
Mitton, J.B., 1997. Selection in Natural Population. Oxford University Press, New York.
Michinina, S.R., Rebordinos, L., 1997. Genetic differentiation in marine and estuarine natural populations of Crassostrea angulata. Mar. Ecol.
Prog. Ser. 154, 167e174.
Nei, M., 1972. Genetic distance between populations. Am. Nat. 106, 283e292.
Nevo, E., 2001. Evolution of genomeephenome diversity under environmental stress. Proc. Natl Acad. Sci. U.S.A. 98, 6233e6240.
Ortea, J., Moro, L., Espinosa, J., 1997. Nuevos datos sobre el genero Elysia Risso 1818 (Opistobranchia, Sacoglossa) en el Atlantico. Rev. Acad.
Canaria Cienc. 9, 141e155.
Perez-Ruzafa, A., 1989. Estudio ecologico y bionomico de los poblamientos bentonicos del Mar Menor (Murcia, SE de Espana). Tesis de Doc-
torado, Universidad de Murcia, 751 pp.
Perez-Ruzafa, A., Marcos-Diego, C., Ros, J.D., 1991. Environmental and biological changes related to recent human activities in the Mar Menor
(SE of Spain). Mar. Pollut. Bull. 23, 747e751.
Powers, D.A., Lauerman, T., Crawford, D., Dimichele, L., 1991. Genetic mechanisms for adapting to a changing environment. Annu. Rev. Genet.
25, 629e659.
Quesada, H., Zapata, C., �Alvarez, G., 1995. A multilocus allozyme discontinuity in the mussel Mytilus galloprovincialis: the interaction of eco-
logical and life-history factors. Mar. Ecol. Prog. Ser. 116, 99e115.
Rahat, M., 1976. Direct development and symbiotic chloroplasts in Elysia timida (Mollusca: Opisthobranchia). Isr. J. Zool. 25, 186e193.
Raymond, M., Rousset, F., 1995a. An exact test for population differentiation. Evolution 49, 1280e1283.
Raymond, M., Rousset, F., 1995b. GENEPOP version 1.2: a population genetics software for exact test and ecumenicism. J. Hered. 86, 248e249.
Raymond, M., Vaanto, R.L., Thomas, F., Rousset, F., de Meeus, T., Renaud, F., 1997. Heterozygote deficiency in the mussel Mytilus edulis species
complex revisited. Mar. Ecol. Prog. Ser. 156, 225e237.
Remmert, H., 1988. Ecologıa, autoecologıa, ecologıa de poblaciones y estudio de ecosistemas. Blume, Barcelona.
Riddoch, B.J., 1993. The adaptive significance of electrophoretic mobility in phosphoglucose isomerase (PGI). Biol. J. Linn. Soc. 50, 1e17.
Rollison, D., Kane, R.A., Lines, J.R.L., 1989. An analysis of fertilization in Bulinus cernicus (Gastropoda: Planorbidae). J. Zool. (Lond.) 226, 311.
Rumpho, M.E., Summer, E.J., Manhart, J.R., 2000. Solar-powered sea slugs. Mollusc/algal chloroplast symbiosis. Plant Physiol. 123, 29e38.
Schmidt, P.S., Rand, D.M., 1999. Intertidal microhabitat and selection at MPI: interlocus contrasts in the northern acorn barnacle. Evolution 53,
135e146.
Schmidt, P.S., 2001. The effects of diet and physiological stress on the evolutionary dynamics of an enzyme polymorphism. Proc. R. Soc. Lond. B
268, 9e14.
Shakle, J.B., Allendorf, F.W., Morizot, D.C., Whitt, G.S., 1990. Gene nomenclature for protein-coding loci in fish. Trans. Am. Fish. Soc. 119,
2e15.
She, J.X., Autem, M., Kotulas, G., Pasteur, N., Bonhomme, F., 1987. Multivariate analysis of genetic exchanges between Solea segalensis (Tel-
eosts, Soleidae). Biol. J. Linn. Soc. 32, 357e371.
Sites, J.W., Marshall, J.C., 2003. Delimiting species: a Renaissance issue in systematic biology. Trends Ecol. Evol. 18 (9), 462e470.
Tatarenkov, A., 1995. Habitat related allozyme variation on a microgeographic scale in the marine snail Littorina mariae (Prosobranchia, Littor-
inacea). Biol. J. Linn. Soc. 63, 105e125.
Thorpe, J.P., 1983. Enzyme variation, genetic distance and evolutionary divergence in relation to levels of taxonomic separation. In: Oxford, G.S.,
Rollison, D. (Eds.), Protein Polymorphism: Adaptive and Taxonomic Significance. Academic, London, pp. 131e152.
Valentine, J.W., 1976. Genetic strategies of adaptation. In: Ayala, F.J. (Ed.), Molecular Evolution. Sinauer Associates, Sunderland, MA,
pp. 78e94.
Veliz, D., Bourget, E., Bernatchez, L., 2004. Regional variation in the spatial scale of selection at MPI* and GPI* in the acorn barnacle Semi-balanus balanoides (Crustacea). J. Evol. Biol. 17, 953e966.
Virgilio, M., Baroncini, N., Trombini, C., Abbiati, M., 2003. Relationships between sediments and tissue contamination and allozymic patterns in
Hediste diversicolor (Polychaeta, Nereididae) in the Pialassa lagoons (north Adriatic Sea). Oceanol. Acta 26, 85e92.
Wares, J.P., Gaines, S.D., Cunningham, C.W., 2001. A comparative study of asymmetric migration events across a marine biogeographic bound-
ary. Evolution 55, 295e306.
Watt, W.B., 2000. Avoiding paradigm-cased limits to knowledge of evolution. Evol. Biol. 32, 73e96.
Weir, B.S., Cockerham, C.C., 1984. Estimating F-statistics for the analysis of population structure. Evolution 38, 1358e1370.
Wilkins, N.O., 1977. Genetic variability in littoral gastropods: phosphoglucose isomerase and phosphoglucomutase in Patella vulgata and Patellaaspera. Mar. Biol. 40, 151e155.
Wright, S., 1951. The genetic structure of populations. Ann. Eugen. 15, 323e354.
Wright, S., 1977. Experimental Results and Evolutionary Deductions. In: Evolution and Genetics of Populations, vol. 3. University of Chicago
Press, Chicago.
Zouros, F., Foltz, D.W., 1984. Possible explanations of heterozygote deficiency in bivalve molluscs. Malacologia 25, 583e591.




















