
Genetic characterization of the honeybee (Apis mellifera) population of Rodrigues Island, based on...
9
Apidologie 37 (2006) 547–555 547 c INRA/DIB-AGIB/ EDP Sciences, 2006 DOI: 10.1051/apido:2006029 Original article Genetic characterization of honey bee (Apis mellifera cypria) populations in northern Cyprus* Irfan K a,b , Marina D. M b , Ayca O a , Walter S. S b a Department of Biology, Zonguldak Karaelmas University, Incivez 67100 Zonguldak, Turkey b Department of Entomology, Washington State University, Pullman 99164-6382 WA, USA Received 15 April 2005 – revised 16 December 2005 – accepted 22 December 2005 Abstract – The variability of the honey bees of northern Cyprus was investigated using morphometric, mitochondrial DNA (mtDNA) and microsatellite analyses. Morphometric analysis resulted in a clear classi- fication of the Cyprus bees as Apis mellifera cypria, but showed the influence of imported A. m. anatoliaca in some areas. In eastern Cyprus, several samples showed a similarity to A. m. meda, possibly corroborating a published report of similarity between A. m. cypria and Mediterranean A. m. meda. However, the importa- tion of A. m. meda into Cyprus could not be ruled out. MtDNA analysis showed that most Cyprian samples belonged to the mitochondrial C lineage, but a small proportion of samples displayed restriction patterns typical for the mitochondrial O lineage. Population differentiation between Cyprus and honey bees from ad- jacent mainland populations was low, but the northwestern Cyprus population appeared to be introgressed to a larger extent by alleles from the Turkish mainland. Apis mellifera cypria / mtDNA / microsatellites / morphometry / Cyprus 1. INTRODUCTION Traditionally, subspecific classification and phylogeographic inferences in Apis mellifera L. have been based on the variation of behav- ior and morphology within the endemic range of the species. Using morphometric analy- ses, Ruttner (1988, 1992) hypothesized the ex- istence of four evolutionary lineages within the species: M in northern and western Eu- rope, A in Africa, C in southeastern Europe, and O in western Asia. Subsequent studies, based on variation of mitochondrial DNA, confirmed Ruttner’s hypotheses about the phy- logeographic structure of Apis mellifera to a large extent (Garnery et al., 1992, 1993; Arias and Sheppard, 1996; Franck et al., 2000a). The most widely used marker in these stud- ies was variation in the intergenic region be- tween the COI and the COII gene in Apis mel- Corresponding author: I. Kandemir, [email protected] * Manuscript editor: Stefan Fuchs lifera mtDNA, as determined by sequencing or restriction analysis (Garnery et al., 1992, 1993, 1998; Franck et al., 1998). However, us- ing these methods, the morphological C and O branches were undistinguishable and were subsumed into a single mitochondrial lineage (C). Recently, Franck et al. (2000a) reported the existence of a previously unknown mtDNA re- striction enzyme pattern in honey bees sam- pled from Lebanon and inferred the existence of a fourth mitochondrial lineage of honey bees (‘mitochondrial O’). This lineage may be analogous to the mtDNA lineage hypothe- sized based on restriction enzyme data (Palmer et al., 2000) and mitochondrial ND2 gene se- quences (Arias and Sheppard, 1996). The dis- tribution of the mitochondrial O lineage re- mains unknown, but may extend from Syria to Egypt (Arias and Sheppard, 1996). The island of Cyprus is situated at the east- ern end of the Mediterranean Sea, south of Turkey (75 km), west of Syria and Lebanon Article published by EDP Sciences and available at http://www.edpsciences.org/apido or http://dx.doi.org/10.1051/apido:2006029
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Genetic characterization of the honeybee (Apis mellifera) population of Rodrigues Island, based on...

Apidologie 37 (2006) 547–555 547c© INRA/DIB-AGIB/ EDP Sciences, 2006DOI: 10.1051/apido:2006029
Original article
Genetic characterization of honey bee (Apis mellifera cypria)populations in northern Cyprus*
Irfan Ka,b, Marina D. Mb, Ayca Oa, Walter S. Sb
a Department of Biology, Zonguldak Karaelmas University, Incivez 67100 Zonguldak, Turkeyb Department of Entomology, Washington State University, Pullman 99164-6382 WA, USA
Received 15 April 2005 – revised 16 December 2005 – accepted 22 December 2005
Abstract – The variability of the honey bees of northern Cyprus was investigated using morphometric,mitochondrial DNA (mtDNA) and microsatellite analyses. Morphometric analysis resulted in a clear classi-fication of the Cyprus bees as Apis mellifera cypria, but showed the influence of imported A. m. anatoliacain some areas. In eastern Cyprus, several samples showed a similarity to A. m. meda, possibly corroboratinga published report of similarity between A. m. cypria and Mediterranean A. m. meda. However, the importa-tion of A. m. meda into Cyprus could not be ruled out. MtDNA analysis showed that most Cyprian samplesbelonged to the mitochondrial C lineage, but a small proportion of samples displayed restriction patternstypical for the mitochondrial O lineage. Population differentiation between Cyprus and honey bees from ad-jacent mainland populations was low, but the northwestern Cyprus population appeared to be introgressedto a larger extent by alleles from the Turkish mainland.
Apis mellifera cypria / mtDNA /microsatellites / morphometry / Cyprus
1. INTRODUCTION
Traditionally, subspecific classification andphylogeographic inferences in Apis melliferaL. have been based on the variation of behav-ior and morphology within the endemic rangeof the species. Using morphometric analy-ses, Ruttner (1988, 1992) hypothesized the ex-istence of four evolutionary lineages withinthe species: M in northern and western Eu-rope, A in Africa, C in southeastern Europe,and O in western Asia. Subsequent studies,based on variation of mitochondrial DNA,confirmed Ruttner’s hypotheses about the phy-logeographic structure of Apis mellifera to alarge extent (Garnery et al., 1992, 1993; Ariasand Sheppard, 1996; Franck et al., 2000a).The most widely used marker in these stud-ies was variation in the intergenic region be-tween the COI and the COII gene in Apis mel-
Corresponding author: I. Kandemir,[email protected]* Manuscript editor: Stefan Fuchs
lifera mtDNA, as determined by sequencingor restriction analysis (Garnery et al., 1992,1993, 1998; Franck et al., 1998). However, us-ing these methods, the morphological C andO branches were undistinguishable and weresubsumed into a single mitochondrial lineage(C).
Recently, Franck et al. (2000a) reported theexistence of a previously unknown mtDNA re-striction enzyme pattern in honey bees sam-pled from Lebanon and inferred the existenceof a fourth mitochondrial lineage of honeybees (‘mitochondrial O’). This lineage maybe analogous to the mtDNA lineage hypothe-sized based on restriction enzyme data (Palmeret al., 2000) and mitochondrial ND2 gene se-quences (Arias and Sheppard, 1996). The dis-tribution of the mitochondrial O lineage re-mains unknown, but may extend from Syria toEgypt (Arias and Sheppard, 1996).
The island of Cyprus is situated at the east-ern end of the Mediterranean Sea, south ofTurkey (75 km), west of Syria and Lebanon
Article published by EDP Sciences and available at http://www.edpsciences.org/apido or http://dx.doi.org/10.1051/apido:2006029

548 I. Kandemir et al.
Figure 1. Map indicating sampling lo-cations in Cyprus.
Table I. Sampling locations, geographical positions, and number of colonies sampled for this study.
Location Geographical position # Colonies sampled year1-Omorfo 35o12’N 32o59’E 5 20002-Lefke 35o06’N 32o51’E 4 20003-Gaziveren 35o11’N 33o01’E 6 20004-Kalecik 35o20’N 34o00’E 3 20005-Iskele 35o16’N 33o54’E 3 20006-Ardahan 35o21’N 33o52’E 47 2000, 20027-Yedikonuk 35o24’N 34o01’E 10 20028-Taslica 35o23’N 34o04’E 5 20029-Kantara 35o23’N 33o53’E 3 200210-Mersinlik 35o24’N 33o55’E 4 200211-Kaplica 35o23’N 33o54’E 5 200212-Girne 35o19’N 33o19’E 6 2004
(105 km) and north of Egypt (380 km). Thehoney bees of Cyprus were described as aseparate subspecies, A. m. cypria, by Pollman(1879) and shown by Ruttner (1988) to be-long to the morphological O lineage of Apismellifera. While other island populations andsubspecies of honey bees in the Mediterraneanhave received more scientific interest (Crete:Ruttner, 1980; Sicily: Badino et al., 1985;Sinacori et al., 1998; Franck et al., 2000b;Malta: Sheppard et al., 1997; Balearics: De laRúa et al., 2001, 2003), very little is knownabout the honey bee of Cyprus. The geo-graphic location of Cyprus positions A. m.cypria in close proximity to subspecies to boththe mitochondrial C and O lineages and thegeographic region of transition between them.In this paper we report the results of an ex-tensive morphometric and genetic analysis ofthe honey bees of Cyprus and compare their
morphometric and genetic variability to that ofneighboring subspecies.
2. MATERIALS AND METHODS
2.1. Collection of bee samples
A total of 101 colonies were sampled from 12locations in northern Cyprus in the years 2000 (40),2002 (55), and 2004 (6) (Fig. 1, Tab. I). Sampleswere stored in 90% ethanol (2000, 2004) or in dryice (2002).
2.2. Morphometric analysis
A total of 18 colonies (3 from each location ofthe 2000 collection) were subjected to morphome-tric analysis. Between 11–15 worker bees per sam-ple were dissected and measured for 39 morphome-tric characters according to methods of Ruttner et al.

Genetic characterization of Apis mellifera cypria 549
(1978) and Ruttner (1988, 1992). Characters of pi-losity and pigmentation were assessed with a stere-omicroscope and an ocular micrometer. All othercharacters were measured with a CCD camera com-bined with a morphometric measurement program(Bee2, c©Meixner, 2004). Reference data of honeybee subspecies of the eastern Mediterranean regionwere obtained from the database of the Institut fürBienenkunde, Oberursel. These included A. m. car-nica (20 samples), A. m. macedonica (10), A. m. ce-cropia (10), A. m. anatoliaca (13), A. m. syriaca (9),A. m. adami (5), and A. m. meda (25). Referencedata for A. m. meda came from samples of A. m.meda collected in Turkey and Syria (Ruttner, 1988;Ftayeh et al., 1994). Data were subjected to prin-cipal component analysis and discriminant analysisusing the SPSS 12.0.1 statistical software.
2.3. Restriction and sequence analysisof mitochondrial DNA
Total nucleic acids of one bee per sample wereisolated with a modified phenol-chloroform ex-traction (Arias and Sheppard, 1996) or a modi-fied CTAB extraction protocol (Doyle and Doyle,1987). A mitochondrial fragment containing theintergenic region between the tRNAleu gene andthe second subunit of the cytochrome oxidasegene was amplified using the primer pair E2-H2(Garnery et al., 1993): E2: 5’-GGC AGA ATAAGT GCA TTG-3’, H2: 5’-CAA TAT CAT TGATGA CC-3’. The 25 µL reaction mix consistedof 0.8 µM of each primer, 0.2 mM of PCR Nu-cleotide mix (Boehringer Mannheim), 1.5 mMMgCl2 (Promega), 1X Reaction Buffer (Promega),1 U Taq Polymerase (Promega) and 1 µL of tem-plate. The amplification cycle consisted of an initialdenaturation step of 2 min at 92 ◦C, followed by 35cycles of 30 s at 92 ◦C, 30 s at 47 ◦C and 2 min at63 ◦C, followed by a final extension step of 10 minat 63 ◦C. Five µL of the PCR products were runon a 1.5% agarose gel, stained with ethidium bro-mide and photographed under UV illumination. A20 µL aliquot of each positive reaction was digestedwith the restriction enzyme DraI at 37 ◦C overnight.Restriction fragments were separated on 10% poly-acrylamide gels, stained with ethidium bromide andphotographed under UV illumination.
Among the samples expressing restriction pro-files of the C and O mitochondrial lineages, we se-quenced the COI-COII region of one sample andthe NADH dehydrogenase subunit 2 gene of two
samples each, using a cycle sequencing protocol(Craxton, 1991) and an ABI 377 automated se-quencer. The ND2 sequences were aligned withcorresponding published sequence data from otherApis mellifera subspecies (Arias and Sheppard,1996) using Clustal X (Thompson et al., 1997).Phylogenetic analyses using both neighbor-joiningand parsimony methods were performed withMEGA 3.1 (Kumar et al., 2004). Sequences weredeposited in GenBank under the accession numbersAY618919–AY618921.
2.4. Microsatellite analysis
The samples were analyzed for nine microsatel-lite loci: A7, A24, A28, A88, A113, B124 (Estoupet al., 1995), Ap55, Ap66, and Ap81 (Garnery et al.,unpubl. data). Amplifications were performed in10 µL reactions containing 1 µL extracted DNA, 1Xreaction buffer, 3 mM dNTPs, 0.001 mg BSA, 1–4 mM of respective primers and 1.5 units Taq poly-merase. Microsatellite primers were combined intotwo multiplex reactions with optimized concentra-tions of MgCl2: 1.2 mM for A7, A113, Ap55 andAp81; and 1.5 mM for loci A24, A28, A88, Ap66and B124. The PCR reaction conditions were iden-tical for all loci and consisted of 7 min at 95 ◦C,followed by 30 cycles of 95 ◦C (30 s), 54 ◦C (30 s),72 ◦C (30 s), and a final 60 min cycle at 72 ◦C. For-ward primers were fluorescent labeled and amplifi-cation products were separated on an ABI 3730 au-tomatic sequencer. The resulting electropherogramswere analyzed using GeneMapper Software (Ap-plied Biosystems).
For analysis, the microsatellite data were com-bined with unpublished reference data from popula-tions in Turkey (n= 47), Syria (n = 22) and Iran (n =43). Exact tests for genetic structure and genetic dif-ferentiation between populations using unbiased es-timates of Fst were calculated using the Genepoppackage version 3.4 (Raymond and Rousset, 1995).A neighbor-joining tree based on the microsatellitedata and the chord distance of Cavalli-Sforza andEdwards was constructed using the Phylip programpackage (Felsenstein, 2005) with bootstrap valuescomputed over 2000 replications.
3. RESULTS
3.1. Morphometry
In a principal component analysis basedon three factors describing 38.8%, 11.9% and
550 I. Kandemir et al.
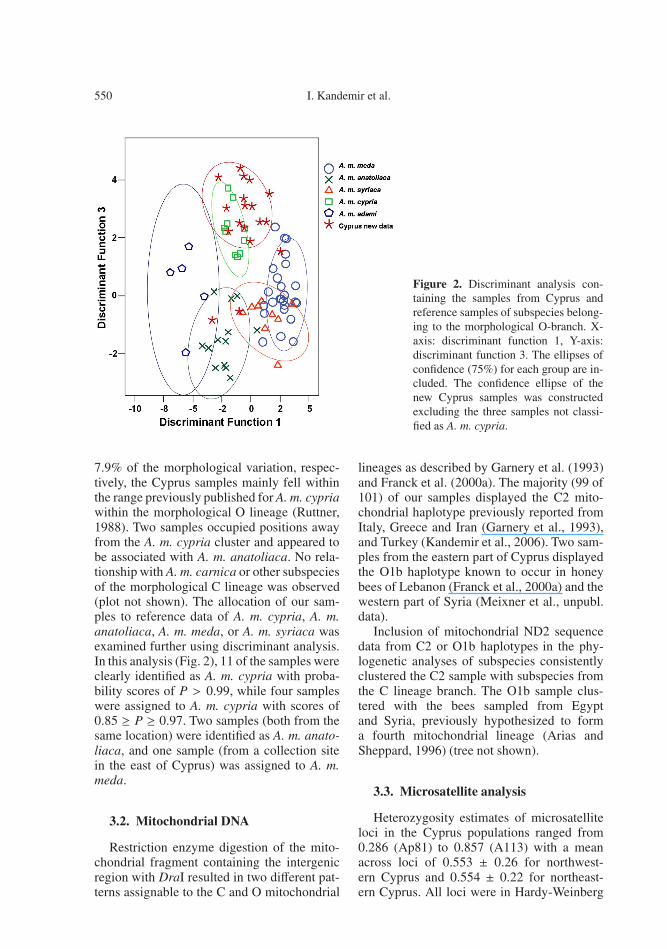
Figure 2. Discriminant analysis con-taining the samples from Cyprus andreference samples of subspecies belong-ing to the morphological O-branch. X-axis: discriminant function 1, Y-axis:discriminant function 3. The ellipses ofconfidence (75%) for each group are in-cluded. The confidence ellipse of thenew Cyprus samples was constructedexcluding the three samples not classi-fied as A. m. cypria.
7.9% of the morphological variation, respec-tively, the Cyprus samples mainly fell withinthe range previously published for A. m. cypriawithin the morphological O lineage (Ruttner,1988). Two samples occupied positions awayfrom the A. m. cypria cluster and appeared tobe associated with A. m. anatoliaca. No rela-tionship with A. m. carnica or other subspeciesof the morphological C lineage was observed(plot not shown). The allocation of our sam-ples to reference data of A. m. cypria, A. m.anatoliaca, A. m. meda, or A. m. syriaca wasexamined further using discriminant analysis.In this analysis (Fig. 2), 11 of the samples wereclearly identified as A. m. cypria with proba-bility scores of P > 0.99, while four sampleswere assigned to A. m. cypria with scores of0.85 ≥ P ≥ 0.97. Two samples (both from thesame location) were identified as A. m. anato-liaca, and one sample (from a collection sitein the east of Cyprus) was assigned to A. m.meda.
3.2. Mitochondrial DNA
Restriction enzyme digestion of the mito-chondrial fragment containing the intergenicregion with DraI resulted in two different pat-terns assignable to the C and O mitochondrial
lineages as described by Garnery et al. (1993)and Franck et al. (2000a). The majority (99 of101) of our samples displayed the C2 mito-chondrial haplotype previously reported fromItaly, Greece and Iran (Garnery et al., 1993),and Turkey (Kandemir et al., 2006). Two sam-ples from the eastern part of Cyprus displayedthe O1b haplotype known to occur in honeybees of Lebanon (Franck et al., 2000a) and thewestern part of Syria (Meixner et al., unpubl.data).
Inclusion of mitochondrial ND2 sequencedata from C2 or O1b haplotypes in the phy-logenetic analyses of subspecies consistentlyclustered the C2 sample with subspecies fromthe C lineage branch. The O1b sample clus-tered with the bees sampled from Egyptand Syria, previously hypothesized to forma fourth mitochondrial lineage (Arias andSheppard, 1996) (tree not shown).
3.3. Microsatellite analysis
Heterozygosity estimates of microsatelliteloci in the Cyprus populations ranged from0.286 (Ap81) to 0.857 (A113) with a meanacross loci of 0.553 ± 0.26 for northwest-ern Cyprus and 0.554 ± 0.22 for northeast-ern Cyprus. All loci were in Hardy-Weinberg

Genetic characterization of Apis mellifera cypria 551
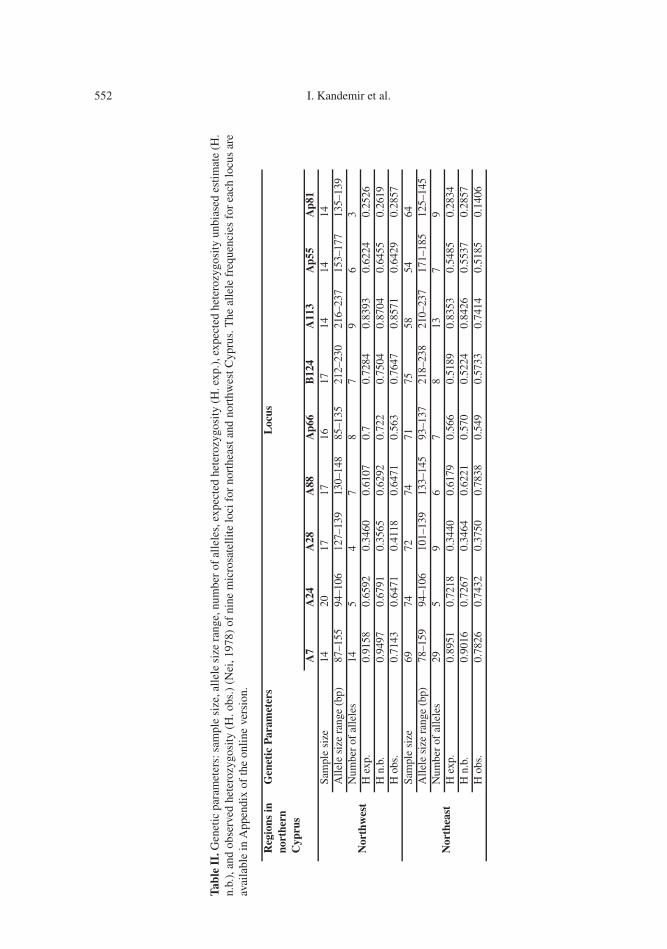
equilibrium with respect to the populationsstudied. The number of alleles, the allele sizerange in bp and the expected and observed het-erozygosities (H. exp. and H. obs.) and the al-lele frequencies for each individual locus arepresented in Table II. The results of pairwisepopulation comparisons using multilocus F-statistics between the northwestern and north-eastern Cyprus populations and the referencepopulations were low and ranged between0.003 (northwestern Cyprus, Turkey) and0.081 (northeastern Cyprus, Syria) (Tab. III).
The populations of northwestern and north-eastern Cyprus showed significant differencesin their microsatellite variability (P < 0.001,Fisher exact test). When compared to sur-rounding mainland populations, the allelic dis-tribution of the northwestern Cyprus popula-tion was not significantly different from thepopulation of Turkey, but different from Syriaand Iran (P < 0.001). The bees of easternCyprus differed significantly from all adja-cent mainland honey bee populations (Turkey,Syria, Iran) (P < 0.001).
A neighbor-joining tree based on theCavalli-Sforza and Edwards chord distance re-sulted in low resolution between the popu-lations from Iran and the branch combiningthe other groups from the Near East. Withinthis branch, the honey bee populations fromCyprus were incorporated into a subclusterwith Syria (Fig. 3).
4. DISCUSSION
Several different subspecies of honey beesbelonging to two different evolutionary lin-eages (C and O) come together in the easternMediterranean and the Near East. Althoughthese two evolutionary lineages are distin-guishable by morphological methods, thedelineation based on restriction analysis of mi-tochondrial DNA is incongruent and seem-ingly confusing. The honey bee subspeciesof the entire Near East, including Turkey,morphologically belong to the O evolution-ary lineage sensu Ruttner (1988). However,in the southern portion of this range a divi-sion between mitochondrial lineages C (sensuGarnery et al., 1993) and O (sensu Franck
et al., 2000a) occurs further south and east.Thus, C mitochondrial haplotypes occur inmany subspecies belonging to the O morpho-logical lineage sensu Ruttner, including mostof the honey bees of Turkey (Kandemir et al.,2006) and those that occur east into Iran andCentral Asia at the eastern edge of the Apismellifera range (unpublished data; Sheppardand Meixner, 2003). Further south, extendingfrom southern Turkey (Kandemir et al., 2006)through Lebanon (Franck et al., 2000a), Syriaand Egypt (Arias and Sheppard, 1996; unpubl.data), honey bee populations are characterizedby haplotypes belonging to the (perhaps unfor-tunately named) mitochondrial lineage O (asdescribed and named by Franck et al., 2000a).
Our results show that the contemporaryhoney bee population of (northern) Cyprus re-tain A. m. cypria characteristics as describedby Ruttner (1988), although in some areasthe influence of other subspecies, especiallyA. m. anatoliaca, can be detected. Beekeepersin Cyprus predominantly use primitive hives,but the use of modern equipment, migratorybeekeeping and commercial pollination prac-tices are increasing (Kandemir, 2003). The twoCyprian samples that were morphometricallyclassified as A. m. anatoliaca and the one withan intermediate score between A. m. anatoli-aca and A. m. cypria all came from modernbeekeeping operations involved in citrus pol-lination (located in northwestern Cyprus) andmay reflect past or recent importation of A.m. anatoliaca queens. In contrast, three othersamples with intermediate scores showed anaffinity to A. m. meda and, together with theone sample classified as A. m. meda, originatedfrom the eastern part of the island where tra-ditional beekeeping in trunk hives is still pre-dominant.
Mitochondrial analysis predominantlyplaced our Cyprus collection into the mito-chondrial C lineage, but also showed a smallproportion of restriction profiles characteristicfor the mitochondrial O lineage. Whetherthis observation reflects a mixed ancestryof the Cyprus population or a more recentintroduction of honey bees from the easternshore of the Mediterranean is unknown. WhileO mitochondrial lineage haplotypes might bea remnant of the Pleistocene fauna of Cyprus
552 I. Kandemir et al.
Tabl
eII
.Gen
etic
para
met
ers:
sam
ple
size
,all
ele
size
rang
e,nu
mbe
rof
alle
les,
expe
cted
hete
rozy
gosi
ty(H
.exp
.),e
xpec
ted
hete
rozy
gosi
tyun
bias
edes
tim
ate
(H.
n.b.
),an
dob
serv
edhe
tero
zygo
sity
(H.o
bs.)
(Nei
,197
8)of
nine
mic
rosa
telli
telo
cifo
rno
rthe
asta
ndno
rthw
estC
ypru
s.T
heal
lele
freq
uenc
ies
for
each
locu
sar
eav
aila
ble
inA
ppen
dix
ofth
eon
line
vers
ion.
Reg
ions
inno
rthe
rnC
ypru
s
Gen
etic
Par
amet
ers
Loc
us
A7
A24
A28
A88
Ap6
6B
124
A11
3A
p55
Ap8
1
Nor
thw
est
Sam
ple
size
1420
1717
1617
1414
14A
llel
esi
zera
nge
(bp)
87–1
5594
–106
127–
139
130–
148
85–1
3521
2–23
021
6–23
715
3–17
713
5–13
9N
umbe
rof
alle
les
145
47
87
96
3H
exp.
0.91
580.
6592
0.34
600.
6107
0.7
0.72
840.
8393
0.62
240.
2526
Hn.
b.0.
9497
0.67
910.
3565
0.62
920.
722
0.75
040.
8704
0.64
550.
2619
Hob
s.0.
7143
0.64
710.
4118
0.64
710.
563
0.76
470.
8571
0.64
290.
2857
Nor
thea
st
Sam
ple
size
6974
7274
7175
5854
64A
llel
esi
zera
nge
(bp)
78–1
5994
–106
101–
139
133–
145
93–1
3721
8–23
821
0–23
717
1–18
512
5–14
5N
umbe
rof
alle
les
295
96
78
137
9H
exp.
0.89
510.
7218
0.34
400.
6179
0.56
60.
5189
0.83
530.
5485
0.28
34H
n.b.
0.90
160.
7267
0.34
640.
6221
0.57
00.
5224
0.84
260.
5537
0.28
57H
obs.
0.78
260.
7432
0.37
500.
7838
0.54
90.
5733
0.74
140.
5185
0.14
06

Genetic characterization of Apis mellifera cypria 553
Table III. Fst results from Genepop.
Northwest NortheastCyprus Cyprus
Northeast Cyprus 0.018Iran 0.021 0.048Turkey 0.003 0.050Syria 0.069 0.081
Figure 3. Neighbor-joining tree based on theCavalli-Sforza and Edwards chord distance be-tween populations (based on nine microsatelliteloci). Bootstrap values are based on 2000 replica-tions.
that contained African elements such as dwarfhippos and elephants (Schüle, 1993), it is morelikely that the presence of these O lineagecolonies in Cyprus resulted from more recenthuman-mediated introductions. For example,it is known that extensive importations ofhoney bee queens were made from presentday Syria and Lebanon to Cyprus in the 19thcentury (Strange, 2001).
In contrast to the high differentiationobserved using mitochondrial markers, mi-crosatellite analysis indicated a relatively lowlevel of differentiation among the Near East-ern populations studied, irrespective of theirassignment to mitochondrial lineages C or O.While overall Fst values between Cyprus andall reference populations were low, the honeybee population of northwestern Cyprus wasintrogressed to a larger extent by microsatel-lite alleles from Turkey, suggesting the roleof queen importation from the Turkish main-land. Thus, while our results confirm the dis-tinctness of A. m. cypria as island subspeciesof Cyprus, they also show that importation ofbees from adjacent mainland areas may be-come a threat to its conservation in the future.
ACKNOWLEDGEMENTS
This project was partially supported by USDA-IFAFS grant 2001-52103-11417 to WSS, and ZKU-2003-13-06-04, TUBITAK-TBAG 2403 grantsand TUBITAK-BAYG 2219 scholarship to IK.We gratefully acknowledge the Institut für Bi-enenkunde in Oberursel, Germany, for use of mor-phometric reference data. We thank Stefan Fuchs,Lionel Garnery and three anonymous reviewers forcomments on earlier versions of the manuscript.
Résumé – Caractérisation génétique des popu-lations d’abeilles domestiques (Apis mellifera cy-pria) à Chypre. L’île méditerranéenne de Chyprepossède sa propre sous-espèce d’abeille domes-tique, Apis mellifera cypria Pollmann 1879, maison connaît peu de choses concernant sa variabilitégénétique et ses relations avec les sous-espèces voi-sines. De par la position géographique de Chypre,A. m. cypria avoisine directement les sous-espècesdes lignées mitochondriales C et O. Nous avonsétudié la variabilité des abeilles de Chypre par lesméthodes morphométriques et par des analyses del’ADN mitochondrial et des microsatellites. Au to-tal 101 échantillons ont été prélevés dans 12 loca-lités du nord de Chypre ; 18 d’entre eux ont faitl’objet d’une analyse morphométrique. Les mesuresde 39 caractères ont été analysées par les méthodesde statistique multivariées. Des échantillons de réfé-rence provenant des régions continentales voisinesont également été analysés. Un fragment mitochon-drial contenant la région intergénique entre le gèneARNtleu et la seconde sous-unité du gène de lacytochrome oxydase a été amplifié et digéré parl’enzyme de restriction DraI. Le fragment conte-nant la région intergénique et un fragment conte-nant l’ARNt pour l’isoleucine et une partie du Gènemitochondrial ND2 ont été séquencés pour deuxéchantillons représentant chacun des haplotypes ob-servés dans l’analyse de restriction. Pour l’analysede la variabilité des microsatellites neuf locus diffé-rents ont été amplifiés avec des amorces marquéespar une substance fluorescente et analysés dans unséquenceur automatique.L’analyse morphométrique a nettement classé lesabeilles de Chypre comme étant A. m. cypria, maisa montré également l’influence dans certaines ré-gions des importations d’A. m. macedonica. Dansla partie orientale de Chypre plusieurs échantillonsprésentaient des similitudes avec A. m. meda et lànon plus l’importation d’A. m. meda à Chypre n’apu être écartée. Les analyses de restriction commecelles de la séquence de l’ADNmt ont montré quela plupart des échantillons chypriotes appartenaientà la lignée mitochondriale C, mais une petite pro-portion d’échantillons présentait des profils de res-triction typiques de la lignée mitochondriale O.D’après la fréquence allélique des microsatellites

554 I. Kandemir et al.
la différenciation entre les échantillons chyprioteset les populations voisines du continent était faible.La population du nord-ouest de Chypre sembleavoir subi une large introgression par des allèlesvenant du continent turque. Ainsi, alors que nosrésultats confirment la particularité d’A. m. cypriacomme sous-espèce de l’île de Chypre, ils montrentaussi que l’importation d’abeilles du continent voi-sin peut devenir une menace pour sa conservation àl’avenir.
Apis mellifera cypria / ADNmt / microsatellite /morphométrie / Chypre
Zusammenfassung – Genetische Charakterisie-rung von Populationen der Honigbiene in Nord-zypern (Apis mellifera cypria). Die Mittelmeerin-sel Zypern besitzt ihre eigene Unterart der Honig-biene, Apis mellifera cypria Pollmann 1879, aberbisher ist noch wenig über ihre genetische Varia-bilität und ihre Beziehungen zu benachbarten Un-terarten bekannt. Bedingt durch die geographischeLage Zyperns ist A. m. cypria sowohl zu Unter-arten der mitochodrialen C als auch der O Liniedirekt benachbart. Wir untersuchten die Variabili-tät von A. m. cypria sowohl mit morphometrischenMethoden als auch mit Analysen der mitochondria-len DNA und von neun Mikrosatellitenloci. Insge-samt wurden 101 Proben an 12 Orten in Nordzy-pern gesammelt, wovon 18 einer morphometrischenAnalyse unterzogen wurden. Es wurden 39 mor-phometrische Merkmale gemessen und mit multiva-riaten statistischen Methoden analysiert, wobei Re-ferenzproben von umliegenden Festlandspopulatio-nen einbezogen wurden. Ein mitochondriales Frag-ment, das die nichtkodierende Region zwischendem tRNAleu Gen und dem CytochromoxidaseIIGen enthält, wurde amplifiziert und mit dem Re-striktionsenzym DraI verdaut. Für jeweils zwei Pro-ben mit den im Restriktionsversuch beobachtetenverschiedenen Haplotypen wurde das Fragment mitder nichtkodierenden Region sowie ein Fragmentdas die tRNAileu sowie einen Teil des mitochon-drialen ND2 Gens enthält, sequenziert. Für die Un-tersuchung der Mikrosatellitenvariabilität wurdenneun verschiedenen Loci mit Fluoreszenzfarbstoffmarkierten Primern amplifiziert und in einem auto-matischen Sequenzierer analysiert.Die morphomterische Analyse ergab eine eindeuti-ge Zuordnung der Proben aus Zypern zu A. m. cy-pria, aber in einigen Gegenden war auch ein Ein-fluss von importierter A. m. anatoliaca deutlich. Imöstlichen Zypern zeigten einige Proben Ähnlichkeitmit A. m. meda, aber auch hier kann ein Import vonA. m. meda nach Zypern nicht ausgeschlossen wer-den. Sowohl Restriktions- als auch Sequenzanalyseder mtDNA ergaben, dass die meisten zyprischenProben zur mitochondrialen C Linie gehören, je-doch wies ein kleiner Prozentsatz der Proben typi-sche Haplotypen der O Linie auf. Auf der Basis der
Allelfrequenzen der Mikrosatelliten war die Diffe-renzierung zwischen den Proben aus Zypern undden benachbarten Festlandspopulationen gering, je-doch erschien die Population aus Nordwestzypernstärker von Allelen des türkischen Festlands beein-flusst. Damit bestätigen unsere Ergebnisse zwar dieBesonderheit von A. m. cypria als Unterart der In-sel Zypern, sie zeigen aber auch, dass Importe vonBienen vom benachbarten Festland eine potentielleBedrohung für die Erhaltung dieser Biene darstel-len.
Apis mellifera cypria / mtDNA / Morphometrie /Zypern /Mikrosatelliten
REFERENCES
Arias M.C., Sheppard W.S. (1996) Molecular phyloge-netics of honey bee subspecies (Apis mellifera L.)inferred from mitochondrial DNA sequence, Mol.Phyl. Evol. 5, 557–566.
Badino G., Celebrano G., Manino A., Longo S. (1985)Enzyme polymorphism in the Sicilian honeybee,Experientia 41, 752–754.
Bee2 Morphometric Software c© 1997–2005, A.Meixner and M.D., Frankfurt, Germany.
Craxton M. (1991) Linear amplification sequencing: Apowerful method for sequencing DNA, Methods3, 20–24.
De la Rúa P., Galian J., Serrano J., Moritz R.F.A.(2001) Molecular characterization and populationstructure of the honeybees from the Balearic is-lands (Spain), Apidologie 32, 417–427.
De la Rúa P., Galian J., Serrano J., Moritz R.F.A.(2003) Genetic structure of Balearıc honeybeepopulations based on microsatellite polymor-phism, Genet. Sel. Evol. 35, 339–350.
Doyle J.J., Doyle L.L. (1987) A rapid DNA isolationprocedure for small quantities of fresh leaf tissue,Phytochem. Bull. 19, 11–15.
Estoup A., Garnery L., Solignac M., Cornuet J.-M.(1995) Microsatellite variation in honey bee (Apismellifera L.) populations: hierarchical geneticstructure and test of the infinite allele and stepwisemutation models, Genetics 140, 679–695.
Felsenstein J. (2005) Phylip (Phylogeny InterferencePackage) version 3.6, Distributed by theauthor, Department of Genome Sciences,University of Washington, Seattle, USA.http://evolution.genetics.washington.edu/phylip.html(accessed on 13 April 2006).
Franck P., Garnery L., Solignac M., Cornuet J.-M.(1998) The origin of west European subspecies ofhoney bees (Apis mellifera): New insights frommicrosatellite and mitochondrial data, Evolution52, 1119–1134.

Genetic characterization of Apis mellifera cypria 555
Franck P., Garnery L., Solignac M., Cornuet J.M.(2000a) Molecular confirmation of a fourth lin-eage in honeybees from the Near East, Apidologie31, 167–180.
Franck P., Garnery L., Celebrano G., Solinac M.,Cornuet J.M. (2000b) Hybrid origins of the Italianhoneybees, Apis mellifera ligustica and A. m. sic-ula, Mol. Ecol. 9, 907–923.
Ftayeh A., Meixner M., Fuchs S. (1994)Morphometrical investigation in Syrian hon-eybees, Apidologie 25, 396–401.
Garnery L., Cornuet J.-M., Solignac M. (1992)Evolutionary history of the honey bee Apis mel-lifera inferred from mitochondrial DNA analysis,Mol. Ecol. 1, 145–154.
Garnery L., Solignac M., Celebrano G., Cornuet J.-M. (1993) A simple test using restricted PCR-amplified mitochondrial DNA to study the ge-netic structure of Apis mellifera L., Experientia 49,1016–1021.
Garnery L., Franck P., Baudry E., Vautrin D., CornuetJ.-M., Solignac M. (1998) Genetic biodiversity ofthe west European honey bee (Apis mellifera mel-lifera and A. m. iberica). I. Mitochondrial DNA,Genet. Sel. Evol. 30, 31–47.
Kandemir I. (2003) Beekeeping experience and devel-opments in Turkey and in Northern Cyprus, Am.Bee J. 143, 464–467.
Kandemir I., Kence M., Sheppard W.S., Kence A.(2006) Mitochondrial DNA variation in honey bee(Apis mellifera L.) populations from Turkey, J.Apic. Res. and Bee World 45, 33–38.
Kumar S., Tamura K., Nei M. (2004) MEGA3:Integrated software for Molecular EvolutionaryGenetics Analysis and sequence alignment,Briefings in Bioinformatics 5, 150–163.
Nei M. (1978) Estimation of average heterozygosityand genetic distance from a small number of indi-viduals, Genetics 89, 583–590.
Palmer M.R., Smith D.R., Kaftanoglu O. (2000)Turkish Honey bees: Genetic variation and
evidence for a fourth lineage of Apis melliferamtDNA, J. Hered. 91, 42–46.
Pollmann A. (1879) Wert der verschiedenenBienenrassen und deren Varietäten, 2nd ed.,Voigt, Berlin Leipzig (1st ed. with description ofA. m. carnica).
Raymond M., Rousset F. (1995) GENEPOP (Version1.2): Population genetics software for exact testsand ecumenicism, J. Hered. 86, 248–249.
Ruttner F. (1980) Apis mellifera adami (n. ssp.), diekretische Biene, Apidologie 11, 385–400.
Ruttner F. (1988) Biogeography and Taxonomy ofHoney bees, Springer-Verlag, Berlin, Heidelberg.
Ruttner F. (1992) Naturgeschichte der Honigbienen,Ehrenwirth Verlag, München.
Ruttner F., Tassencourt L., Louveaux J. (1978)Biometrical-statistical analysis of the geographicvariability of Apis mellifera L. 1. Materials andMethods, Apidologie 9, 363–381.
Schüle W. (1993) Mammals, vegetation and the initialhuman settlement of the Mediterranean islands: apaleoecological approach, J. Biogeogr. 20, 399–411.
Sheppard W.S., Arias M.C., Grech A., Meixner M.D.(1997) Apis mellifera ruttneri, a new honeybeesubspecies from Malta, Apidologie 28, 287–293.
Sheppard W.S., Meixner M.D. (2003) Apis mellif-era pomonella, a new honey bee subspecies fromCentral Asia, Apidologie 34, 367–375.
Sinacori A., Rinderer T.E., Lancaster V., SheppardW.S. (1998) A morphological and mitochondrialassessment of Apis mellifera from Palermo, Italy,Apidologie 29, 481–592.
Strange J.P. (2001) “A severe stinging and much fa-tigue” – Frank Benton and his 1881 search for Apisdorsata, Am. Entomol., Summer 2001, 112–116.
Thompson J.D., Gibson T.J., Plewniak F., JeanmouginF., Higgins D.G. (1997) The ClustalX windowsinterface: flexible strategies for multiple sequencealignment aided by quality analysis tools, NucleicAcids Res. 24, 4876–4882.




















