
Genetic analysis of avian influenza A viruses isolated from domestic waterfowl in live-bird markets...
13
ORIGINAL ARTICLE Genetic analysis of avian influenza A viruses isolated from domestic waterfowl in live-bird markets of Hanoi, Vietnam, preceding fatal H5N1 human infections in 2004 Samadhan J. Jadhao Doan C. Nguyen Timothy M. Uyeki Michael Shaw Taronna Maines Thomas Rowe Catherine Smith Lien P. T. Huynh Ha K. Nghiem Diep H. T. Nguyen Hang K. L. Nguyen Hanh H. T. Nguyen Long T. Hoang Tung Nguyen Lien S. Phuong Alexander Klimov Terrence M. Tumpey Nancy J. Cox Ruben O. Donis Yumiko Matsuoka Jacqueline M. Katz Received: 19 January 2009 / Accepted: 9 June 2009 / Published online: 4 July 2009 Ó US Government 2009 Abstract The first known cases of human infection with highly pathogenic avian influenza (HPAI) H5N1 viruses in Vietnam occurred in late 2003. However, HPAI H5N1 and low-pathogenic avian influenza (LPAI) H5N2 and H9N3 viruses were isolated from domestic waterfowl during live- bird market (LBM) surveillance in Vietnam in 2001 and 2003. To understand the possible role of these early viruses in the genesis of H5N1 strains infecting people, we per- formed sequencing and molecular characterization. Phylo- genetic analysis revealed that the hemagglutinin (HA) genes of two geese HPAI H5N1 strains belonged to clade 3, and their surface glycoprotein and replication complex genes were most closely related (98.5–99.7% homologous) to A/duck/Guangxi/22/01 (H5N1) virus, detected contem- porarily in southern China, whilst the M and NS genes were derived from an A/duck/Hong Kong/2986.1/00 (H5N1)-like virus. The H5 HA gene of the duck HPAI H5N1 strain belonged to clade 5 and acquired a gene constellation from A/quail/Shantou/3846/02 (H5N1), A/teal/China/2978.1/02 (H5N1) and A/partridge/Shantou/2286/03 (H5N1)-like viruses. The phylogenetic analysis further indicated that all eight gene segments of goose and duck HPAI H5N1 and LPAI H5N2 viruses were distinct from those of H5N1 clade-1 viruses known to have caused fatal human infec- tions in Vietnam since late 2003. The duck H9N3 isolates derived genes from aquatic-bird influenza viruses, and their H9 HA belonged to the Korean lineage. The PB2 gene of A/ duck/Vietnam/340/01 (H9N3) virus had lysine at position 627. Based on the molecular characterization of specific amino acid residues in the surface and relevant internal S. J. Jadhao Á D. C. Nguyen Á T. M. Uyeki Á M. Shaw Á T. Maines Á T. Rowe Á C. Smith Á A. Klimov Á T. M. Tumpey Á N. J. Cox Á R. O. Donis (&) Á Y. Matsuoka Á J. M. Katz Influenza Division, MS-G16, National Center for Immunization and Respiratory Diseases, Centers for Disease Control and Prevention, 1600 Clifton Road, NE, Atlanta, GA 30333, USA e-mail: [email protected] D. C. Nguyen Á L. P. T. Huynh Á H. K. Nghiem Á H. K. L. Nguyen Á H. H. T. Nguyen Á L. T. Hoang National Institute of Hygiene and Epidemiology, Hanoi, Vietnam D. H. T. Nguyen Á T. Nguyen Á L. S. Phuong National Center for Veterinary Diagnosis, Hanoi, Vietnam Present Address: S. J. Jadhao Southeast Poultry Research Laboratory, USDA-ARS, 934 College Station Road, Athens, GA 30605, USA e-mail: [email protected] Present Address: D. C. Nguyen Department of Biology, Georgia State University, Atlanta, GA 30302, USA Present Address: T. Rowe Department of Immunology, University of Toronto, Toronto, ON, Canada Present Address: Y. Matsuoka Laboratory of Infectious Diseases, National Institute of Allergy and Infectious Diseases, National Institute of Health, Bethesda, MD 20892, USA 123 Arch Virol (2009) 154:1249–1261 DOI 10.1007/s00705-009-0429-2
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Genetic analysis of avian influenza A viruses isolated from domestic waterfowl in live-bird markets...

ORIGINAL ARTICLE
Genetic analysis of avian influenza A viruses isolatedfrom domestic waterfowl in live-bird markets of Hanoi, Vietnam,preceding fatal H5N1 human infections in 2004
Samadhan J. Jadhao Æ Doan C. Nguyen Æ Timothy M. Uyeki Æ Michael Shaw Æ Taronna Maines ÆThomas Rowe Æ Catherine Smith Æ Lien P. T. Huynh Æ Ha K. Nghiem Æ Diep H. T. Nguyen Æ Hang K. L. Nguyen ÆHanh H. T. Nguyen Æ Long T. Hoang Æ Tung Nguyen Æ Lien S. Phuong Æ Alexander Klimov Æ Terrence M. Tumpey ÆNancy J. Cox Æ Ruben O. Donis Æ Yumiko Matsuoka Æ Jacqueline M. Katz
Received: 19 January 2009 / Accepted: 9 June 2009 / Published online: 4 July 2009
� US Government 2009
Abstract The first known cases of human infection with
highly pathogenic avian influenza (HPAI) H5N1 viruses in
Vietnam occurred in late 2003. However, HPAI H5N1 and
low-pathogenic avian influenza (LPAI) H5N2 and H9N3
viruses were isolated from domestic waterfowl during live-
bird market (LBM) surveillance in Vietnam in 2001 and
2003. To understand the possible role of these early viruses
in the genesis of H5N1 strains infecting people, we per-
formed sequencing and molecular characterization. Phylo-
genetic analysis revealed that the hemagglutinin (HA)
genes of two geese HPAI H5N1 strains belonged to clade 3,
and their surface glycoprotein and replication complex
genes were most closely related (98.5–99.7% homologous)
to A/duck/Guangxi/22/01 (H5N1) virus, detected contem-
porarily in southern China, whilst the M and NS genes were
derived from an A/duck/Hong Kong/2986.1/00 (H5N1)-like
virus. The H5 HA gene of the duck HPAI H5N1 strain
belonged to clade 5 and acquired a gene constellation from
A/quail/Shantou/3846/02 (H5N1), A/teal/China/2978.1/02
(H5N1) and A/partridge/Shantou/2286/03 (H5N1)-like
viruses. The phylogenetic analysis further indicated that all
eight gene segments of goose and duck HPAI H5N1 and
LPAI H5N2 viruses were distinct from those of H5N1
clade-1 viruses known to have caused fatal human infec-
tions in Vietnam since late 2003. The duck H9N3 isolates
derived genes from aquatic-bird influenza viruses, and their
H9 HA belonged to the Korean lineage. The PB2 gene of A/
duck/Vietnam/340/01 (H9N3) virus had lysine at position
627. Based on the molecular characterization of specific
amino acid residues in the surface and relevant internal
S. J. Jadhao � D. C. Nguyen � T. M. Uyeki � M. Shaw �T. Maines � T. Rowe � C. Smith � A. Klimov �T. M. Tumpey � N. J. Cox � R. O. Donis (&) �Y. Matsuoka � J. M. Katz
Influenza Division, MS-G16, National Center for Immunization
and Respiratory Diseases, Centers for Disease Control and
Prevention, 1600 Clifton Road, NE, Atlanta, GA 30333, USA
e-mail: [email protected]
D. C. Nguyen � L. P. T. Huynh � H. K. Nghiem �H. K. L. Nguyen � H. H. T. Nguyen � L. T. Hoang
National Institute of Hygiene and Epidemiology, Hanoi,
Vietnam
D. H. T. Nguyen � T. Nguyen � L. S. Phuong
National Center for Veterinary Diagnosis, Hanoi, Vietnam
Present Address:S. J. Jadhao
Southeast Poultry Research Laboratory, USDA-ARS,
934 College Station Road, Athens, GA 30605, USA
e-mail: [email protected]
Present Address:D. C. Nguyen
Department of Biology, Georgia State University,
Atlanta, GA 30302, USA
Present Address:T. Rowe
Department of Immunology, University of Toronto, Toronto,
ON, Canada
Present Address:Y. Matsuoka
Laboratory of Infectious Diseases, National Institute of Allergy
and Infectious Diseases, National Institute of Health, Bethesda,
MD 20892, USA
123
Arch Virol (2009) 154:1249–1261
DOI 10.1007/s00705-009-0429-2

protein-coding genes, the Vietnamese H5N1 and H9N3
virus isolates indicated specificity to avian cell surface
receptor and susceptibility for currently licensed anti-
influenza A virus chemotherapeutics. Our findings suggest
that the H5N1 and H5N2 viruses that circulated among
geese and ducks in LBMs in Hanoi, Vietnam, during 2001
and 2003 were not the immediate ancestors of the clade-1
viruses associated with fatal human infections in Vietnam.
The clade-1 HPAI H5N1 viruses were independently
introduced into Vietnam.
Introduction
Widespread outbreaks of highly pathogenic avian influenza
(HPAI) H5N1 viruses among poultry in Vietnam began in
late 2003 [1, 20]. HPAI H5N1 outbreaks were also reported
among domestic poultry and migratory birds in Asian,
African and some European countries during subsequent
years [1, 2, 9, 18–20, 26, 28, 36, 38]. From November 2003
through June 2008, HPAI H5N1 viruses caused fatal
human infections in 257 of 418 confirmed cases in 15
countries in Asia, Africa, the Middle East, and Europe [39].
The origins of the HA genes of all viruses isolated from
humans has been traced to domestic poultry in southern
China [1, 8, 12, 20, 32]. A limited number of cloacal and
tracheal swabs were collected from domestic waterfowl in
the course of epidemiological study of avian influenza A
viruses in live bird markets (LBMs) in Hanoi, Vietnam, in
2001 [24]. On the basis of this pilot surveillance in LBMs,
we reported isolation of HPAI H5N1, low-pathogenic
avian influenza (LPAI) H5N2, H4N6 and H9N3 viruses
from apparently healthy geese and ducks [24]. The H5N1
avian influenza viruses from geese and ducks possessed
polybasic amino acid sequence at the HA cleavage site.
One of the goose H5N1 isolates was tested and proven
highly pathogenic in a chicken intravenous pathotyping
experiment based on the criteria of Office International des
Epizooties, World Organization for Animal Health [24].
Concurrent circulation of several genotypes of HPAI H5N1
viruses was reported among domestic ducks between 1999
and 2002 in southern China [6]. Since late 2003, HPAI
H5N1 outbreaks among domestic poultry in Vietnam have
resulted in zoonotic transmission and fatal human infec-
tions in 56 of 111 cases to date [39]. A partial nucleotide-
sequence-based phylogeny of the HA and NA genes of the
goose H5N1, duck H5N2 and duck H9N3 avian influenza
viruses isolated in 2001 in the LBMs of Hanoi, Vietnam,
was reported previously [24]. However, information on the
origin of the internal genes of these avian influenza virus
isolates and complete HA and NA nucleotide sequence
data were lacking. Phylogenetic and molecular analysis of
these H5N1 and H9N3 avian influenza viruses isolated in
2001 in the LBMs of Hanoi, Vietnam, could enhance our
understanding about the origin of the H5N1 viruses from
late 2003 and subsequent outbreaks in poultry and fatal
H5N1 human infections in Vietnam. In this study, we
investigated whether the HPAI H5N1 viruses isolated in
2001 and 2003 from domestic waterfowl in LBMs of Ha-
noi, Vietnam, were the precursors of the HPAI H5N1
viruses that caused fatal human infections in 2003. Phylo-
genetic analysis of the LPAI H5N2 and H9N3 viruses
was performed to understand if these viruses contributed
genes to the HPAI H5N1 viruses that caused fatal human
infections after late 2003. We found that the HPAI H5N1
viruses isolated from domestic waterfowl in 2001 and 2003
were not the closest common ancestors of the clade-1 viruses
that caused human infections in Vietnam in late 2003–2004.
The clade-1 H5N1 viruses were independently introduced
into Vietnam.
Materials and methods
Avian influenza viruses
The avian influenza A virus strains isolated from geese and
ducks at LBMs in Hanoi, Northern Vietnam, or during
routine surveillance in domestic ducks are shown in
Table 1. Viruses were propagated in 10-day-old embryo-
nated chicken eggs. RNA was extracted from allantoic
fluids using a commercial kit per manufacturer instructions
(Qiagen Inc., Valencia, CA). All research with these
Vietnamese avian influenza viruses was conducted under
biosafety level 3 containment, including enhancements
required by the US Department of Agriculture and the
Select Agent Program (see interim guidance at http://www.
cdc.gov/od/ohs/biosfty/bmbl5/bmbl5toc.htm).
Nucleotide sequencing and phylogenetic analysis
Influenza virus genes were amplified with segment-specific
oligonucleotides using the One-Step RT-PCR kit (Qiagen
Inc., Valencia, CA) [16]. The nucleotide sequences were
determined on an automated Applied Biosystems ABI 3100
sequencer using a BigDye terminator v3.1 cycle sequenc-
ing kit (Applied Biosystems Inc. Foster City, CA).
Sequence outputs were analyzed with the SeqMan module
of the Lasergene software (DNAStar Inc., Madison, WI).
Nucleotide sequences were aligned by the Clustal W
algorithm using the MegAlign 5.07 module in Lasergene
molecular biology software (DNAStar Inc., Madison, WI).
The influenza virus gene nucleotide sequences have been
submitted to the Global Initiative on Sharing Avian Influ-
enza Data (GISAID) database [3], and the sequence
accession numbers EPI168444 through EPI168481 can be
1250 S. J. Jadhao et al.
123
accessed at http://platform.gisaid.org. Phylogenetic analy-
sis was performed by the neighbor-joining method with the
maximum composite likelihood model and 10,000 boot-
strap replicate analyses using MEGA 4.0 software [34].
The sequence regions used for phylogenetic analysis of
various gene segments were based on the availability of the
reference virus sequences in GenBank: PB2 (1029-2135),
PB1 (92-1408), PA (52-2151), H5 HA (42-1640), NP (133-
1034), N1 NA (63-1073), M (32-782), NS (78-719), H9
HA (148-1551), N2 NA (89-1271), N3 NA (19-1449). The
phylogenetic trees for PB2, N1 NA, NS, H9 HA, N2 NA
and N3 NA genes are not included in this report but are
available upon request. The analysis and assignment of
genotypes to the influenza virus isolates was performed
using a web tool as described previously [21]. Viruses
representing relevant HA clades were included as a point of
reference in the phylogenetic tree in Fig. 1 [11].
Molecular analysis
H5 HA amino acid sequences comparison of all H5N1
viruses known to have infected people between the years
1997 and 2005 were aligned using the Clustal W algorithm
(DNAStar Inc., Madison, WI). The highest nucleotide/
amino acid identities to the published influenza A virus
sequences were determined using FluGenome BLAST
search [21].
Results and discussion
Phylogenetic and genotype analyses
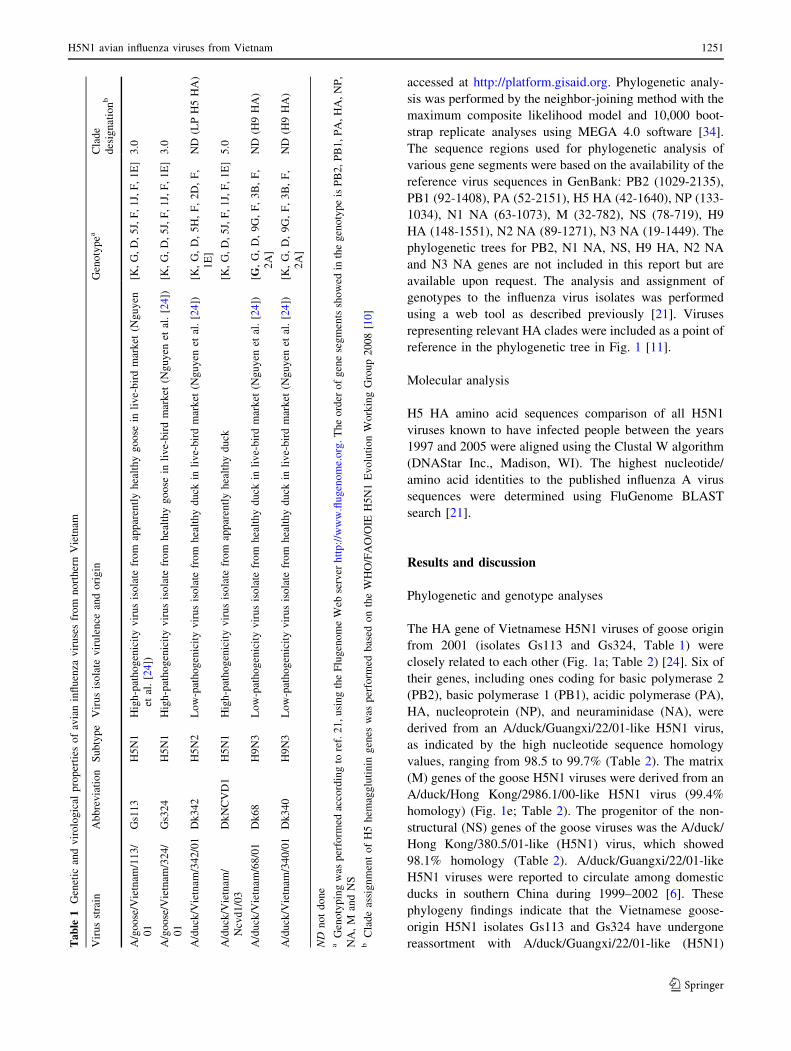
The HA gene of Vietnamese H5N1 viruses of goose origin
from 2001 (isolates Gs113 and Gs324, Table 1) were
closely related to each other (Fig. 1a; Table 2) [24]. Six of
their genes, including ones coding for basic polymerase 2
(PB2), basic polymerase 1 (PB1), acidic polymerase (PA),
HA, nucleoprotein (NP), and neuraminidase (NA), were
derived from an A/duck/Guangxi/22/01-like H5N1 virus,
as indicated by the high nucleotide sequence homology
values, ranging from 98.5 to 99.7% (Table 2). The matrix
(M) genes of the goose H5N1 viruses were derived from an
A/duck/Hong Kong/2986.1/00-like H5N1 virus (99.4%
homology) (Fig. 1e; Table 2). The progenitor of the non-
structural (NS) genes of the goose viruses was the A/duck/
Hong Kong/380.5/01-like (H5N1) virus, which showed
98.1% homology (Table 2). A/duck/Guangxi/22/01-like
H5N1 viruses were reported to circulate among domestic
ducks in southern China during 1999–2002 [6]. These
phylogeny findings indicate that the Vietnamese goose-
origin H5N1 isolates Gs113 and Gs324 have undergone
reassortment with A/duck/Guangxi/22/01-like (H5N1)Ta
ble
1G
enet
ican
dv
iro
log
ical
pro
per
ties
of
avia
nin
flu
enza
vir
use
sfr
om
no
rth
ern
Vie
tnam
Vir
us
stra
inA
bb
rev
iati
on
Su
bty
pe
Vir
us
iso
late
vir
ule
nce
and
ori
gin
Gen
oty
pea
Cla
de
des
ign
atio
nb
A/g
oo
se/V
ietn
am/1
13
/
01
Gs1
13
H5
N1
Hig
h-p
ath
og
enic
ity
vir
us
iso
late
fro
map
par
entl
yh
ealt
hy
go
ose
inli
ve-
bir
dm
ark
et(N
gu
yen
etal
.[2
4])
[K,
G,
D,
5J,
F,
1J,
F,
1E
]3
.0
A/g
oo
se/V
ietn
am/3
24
/
01
Gs3
24
H5
N1
Hig
h-p
ath
og
enic
ity
vir
us
iso
late
fro
mh
ealt
hy
go
ose
inli
ve-
bir
dm
ark
et(N
gu
yen
etal
.[2
4])
[K,
G,
D,
5J,
F,
1J,
F,
1E
]3
.0
A/d
uck
/Vie
tnam
/34
2/0
1D
k3
42
H5
N2
Lo
w-p
ath
og
enic
ity
vir
us
iso
late
fro
mh
ealt
hy
du
ckin
liv
e-b
ird
mar
ket
(Ng
uy
enet
al.
[24
])[K
,G
,D
,5
H,
F,
2D
,F
,
1E
]
ND
(LP
H5
HA
)
A/d
uck
/Vie
tnam
/
Ncv
d1
/03
Dk
NC
VD
1H
5N
1H
igh
-pat
ho
gen
icit
yv
iru
sis
ola
tefr
om
app
aren
tly
hea
lth
yd
uck
[K,
G,
D,
5J,
F,
1J,
F,
1E
]5
.0
A/d
uck
/Vie
tnam
/68
/01
Dk
68
H9
N3
Lo
w-p
ath
og
enic
ity
vir
us
iso
late
fro
mh
ealt
hy
du
ckin
liv
e-b
ird
mar
ket
(Ng
uy
enet
al.
[24
])[G
,G
,D
,9
G,
F,
3B
,F
,
2A
]
ND
(H9
HA
)
A/d
uck
/Vie
tnam
/34
0/0
1D
k3
40
H9
N3
Lo
w-p
ath
og
enic
ity
vir
us
iso
late
fro
mh
ealt
hy
du
ckin
liv
e-b
ird
mar
ket
(Ng
uy
enet
al.
[24
])[K
,G
,D
,9
G,
F,
3B
,F
,
2A
]
ND
(H9
HA
)
ND
no
td
on
ea
Gen
oty
pin
gw
asp
erfo
rmed
acco
rdin
gto
ref.
21
,u
sin
gth
eF
lug
eno
me
Web
serv
erh
ttp
://w
ww
.flu
gen
om
e.o
rg.
Th
eo
rder
of
gen
ese
gm
ents
sho
wed
inth
eg
eno
typ
eis
PB
2,
PB
1,
PA
,H
A,
NP
,
NA
,M
and
NS
bC
lad
eas
sig
nm
ent
of
H5
hem
agg
luti
nin
gen
esw
asp
erfo
rmed
bas
edo
nth
eW
HO
/FA
O/O
IEH
5N
1E
vo
luti
on
Wo
rkin
gG
rou
p2
00
8[1
0]
H5N1 avian influenza viruses from Vietnam 1251
123
A/duck/Hong Kong/205/77A/duck/Hong Kong/698/79
A/duck/Potsdam/22164/84A/duck/Mongolia/54/01
A/duck/Mongolia/500/0186
84 Early aquatic bird isolates
A/duck/Vietnam/342/01 (H5N2)A/duck/Primoriae/2633/01
A/duck/Hokkaidio/101/04A/mallard/Sweden/58/02
A/goose/Guangdong/1/96A/Hong Kong/156/97
A/duck/Anyang/AVL-1/01A/silky chicken/Hong Kong/YU100/02
A/ hi k /H K /FY150/01
8299
100
100100
81
Recent aquatic bird isolates
Clade 0
A/chicken/Hong Kong/FY150/01A/duck/Hong Kong/2986.1/00A/duck/Fujian/17/01A/goose/Hong Kong/3014.5/00
A/duck/Guangxi/22/01A/goose/Vietnam/113/01 (H5N1)A/goose/Vietnam/324/01 (H5N1)
A/chicken/HongKong/YU822.2/01A/ hi k /H K /8791/01
100
99
9993
Clade 3
A/chicken/Hong Kong/8791/01A/goose/Guiyang/337/06A/chicken/Guiyang/846/06
A/chicken/Shanxi/2/06A/teal/China/2978.1/02A/quail/Shantou/3846/02
A/duck/Vietnam/NCVD1/03 (H5N1)A/chicken/Vietnam/Ncvd8/03
A/goose/Guangxi/914/04
100
100
98
98
9486
0.01
Clade 4
Clade 7
Clade 5
A/goose/Guangxi/914/04A/chicken/Guangxi/2439/04
A/duck/Zhejiang/52/00A/swine/Fujian/F1/01A/swine/Fujian/1/03A/duck/Hubeiwg/02/6
A/chicken/Jilin/9/04A/duck/Guangxi/2775/05
A/chicken/Hong Kong/YU22/02
100
98
94
84
86
Cl d 8
Clade 6
Clade 9A/chicken/Hong Kong/YU22/02
A/chicken/Henan12/04A/mallard/Guangxi/wt/04
A/duck/China/E319-2/03A/goose/Guangxi/3316/05A/duck/Hunan/139/05
A/duck/Guiyang/3009/05A/Anuhai/1/05A/ bh goose/Qinghai/12/05
86
91
9760
82
Clade 2
Clade 8
A/ bh goose/Qinghai/12/05A/chicken/Indonesia/7/03
A/duck/Indonesia/MS/04A/Indonesia/283H/06
A/goose/Hong Kong/739.2.02A/Hong Kong/213/03
A/duck/Yunnan/215/03A/chicken/Vietnam/27/03A/chicken/Vietnam/C58/04
9899
68
98
85
A/quail/Vietnam/036/04A/Vietnam/3062/04 (H5N1)
A/Vietnam/1194/04 (H5N1)A/Vietnam/1203/04 (H5N1)
A/duck/Vietnam/NCVD07/0590
90 Clade 1
A: H5 HA
1252 S. J. Jadhao et al.
123

virus to acquire surface glycoprotein genes and four repli-
cation complex genes. The matrix and NS segments were
acquired from an A/duck/Hong Kong/380.5/01-like (H5N1)
virus. The reassortment events were likely occurred in
southern China before introduction of these viruses in geese
and their detection in the LBMs of Hanoi, Vietnam, because
no additional avian influenza A viruses could be isolated
from these geese. Three different viruses were identified as
the ancestors of the different genes found in DkNCVD1
(Fig. 1a–e; Table 2). The PB2, PB1, NP and NA genes of
DkNCVD1 were derived from A/teal/China/2978.1/02-like
(H5N1) virus (Table 2). The ancestor for the HA, M and NS
genes of DkNCVD1 was an A/quail/Shantou/3846/02-like
H5N1 virus (Fig. 1a, e; Table 2). The PA gene of
DkNCVD1 virus had a more distant ancestor, namely A/
partridge/Shantou/2886/03-like (H5N1) virus, showing
97% sequence homology (Fig. 1c; Table 2).
The identification of contemporary isolates from the
Guangxi and Shantou provinces of southern China as the
progenitors of the Vietnamese goose and duck viruses
characterized in this study suggests a possible virus intro-
duction from southern China to northern Vietnam [7, 24, 25,
40]. Phylogenetic analysis suggested that the genome of
LPAI Dk342 (H5N2) virus had multiple ancestors among
viruses from aquatic birds (Fig. 1a–e; Table 2). The H5 HA
and N2 NA descended from an A/duck/Primorie/2633/01-
like (H5N2) virus (98.0 and 98.6% homology, respectively)
[22]. The internal genes of Dk342 (H5N2) virus were derived
from aquatic-bird influenza viruses from southern China and
the Hokkaido region of Japan (Fig. 1b–e; Table 2).
The phylogenetic analysis of the genomes of LPAI duck
H9N3 viruses isolated from LBMs in Hanoi, Vietnam, in
2001 suggested that these viruses were reassortants and
derived their genes from multiple subtypes of avian influ-
enza viruses from aquatic birds and domestic poultry in
southern China, Hong Kong, Japan or Europe (Fig. 1b–e;
Table 2). None of the genes from Vietnamese LPAI H9N3
viruses appeared to be immediate ancestors of the HPAI
H5N1 viruses responsible for causing fatal H5N1 human
infections in Vietnam in late 2003. The HA of Dk68 and
Dk340 H9N3 virus isolates showed highest nucleotide
sequence homology with H9 HA of A/duck/Hokkaido/9/99
(H9N2) virus (96.98 and 98.73%, respectively). The N3
NA of Dk68 and Dk340 viruses were most closely related
to A/mallard/Italy/208/00 (H5N3) virus (96.88 and 95.83%
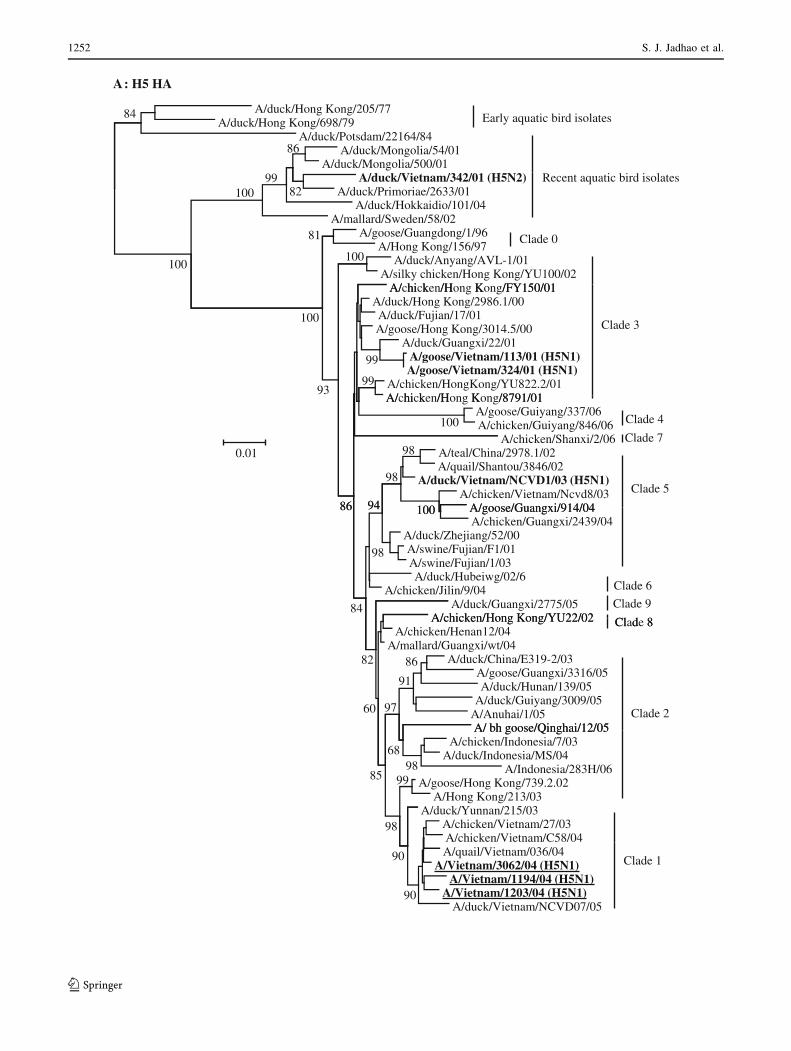
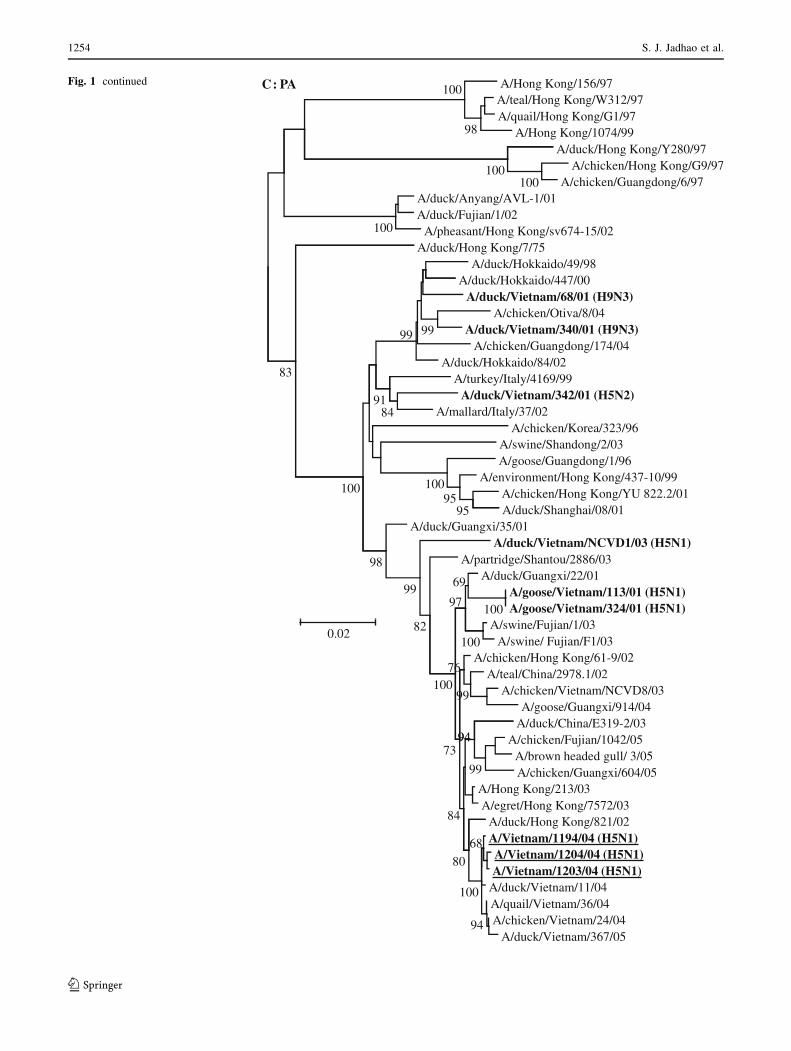
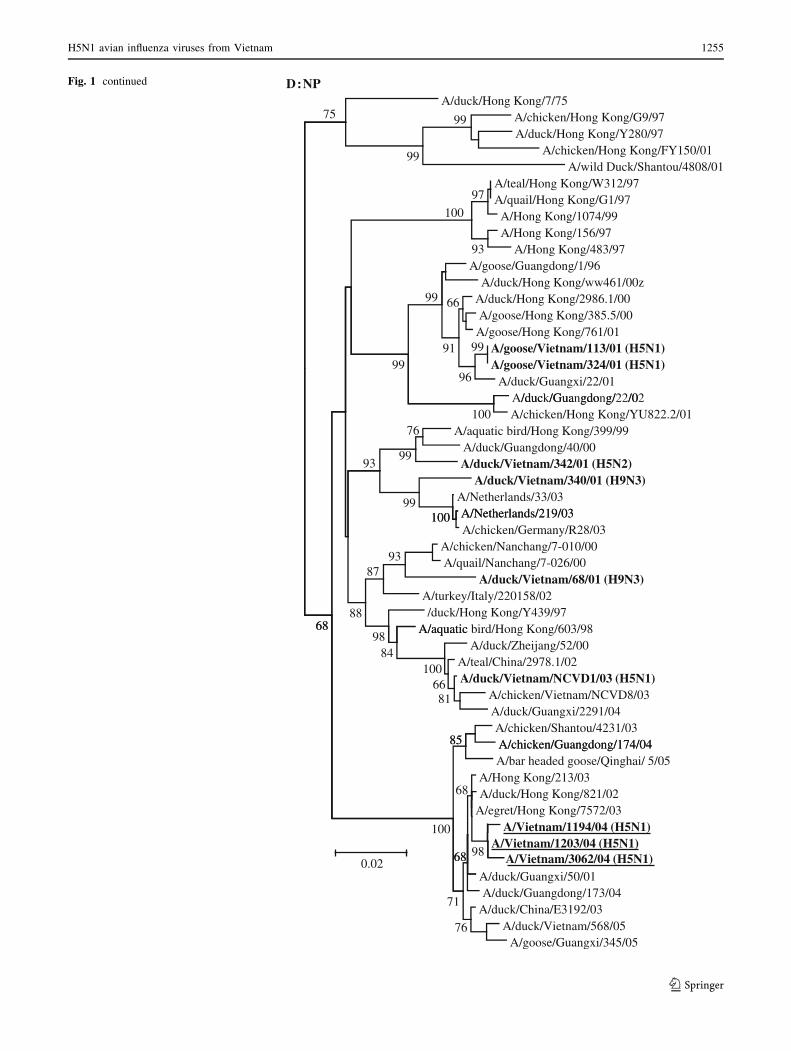
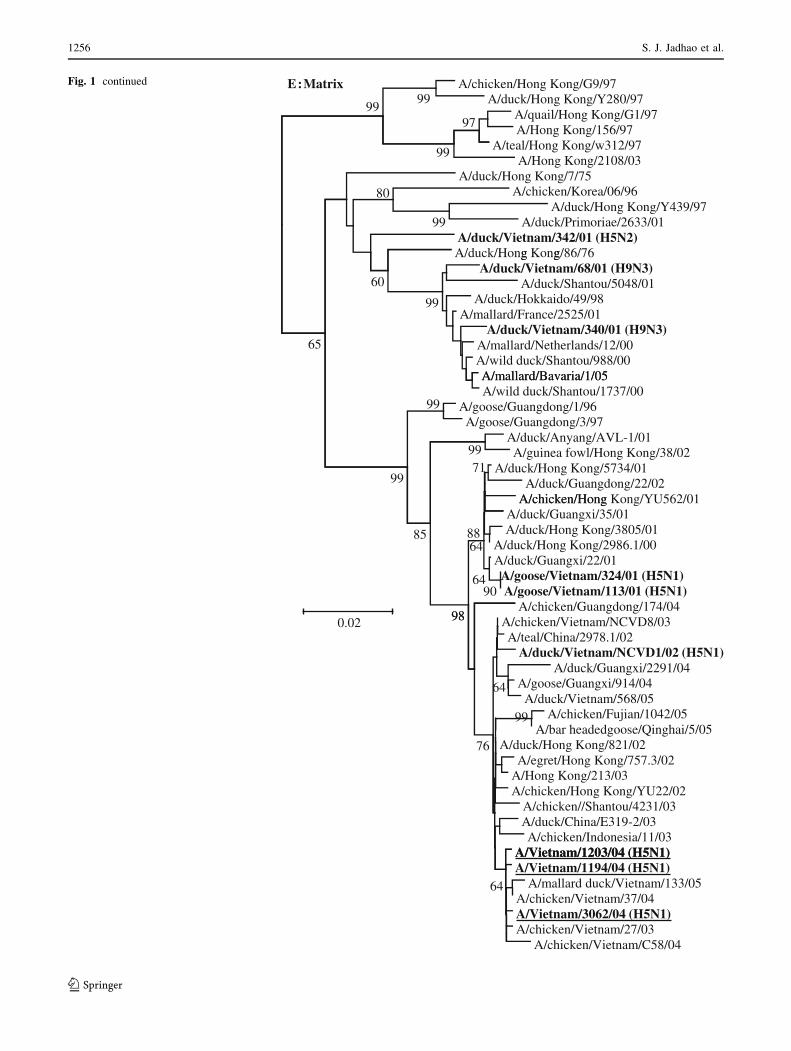
Fig. 1 Phylogenetic analysis of the genes of influenza A viruses
isolated from geese and ducks in Hanoi, Northern Vietnam, in 2001
and 2003. Phylogenetic trees were generated using neighbor-joining
analysis with the maximum composite model in the MEGA 4.0
program [33]. The reliability of the trees was assessed by bootstrap
analysis with 10,000 replicates, and the bootstrap values C60% are
indicated at tree nodes. The scale bar on each tree indicates
nucleotide substitution/site. Vietnam viruses from this study are
shown in bold font. The H5N1 viruses responsible for causing fatal
human infections in Vietnam in 2004 are shown in bold type and are
underlined. The outgroup virus isolates used for the different gene
segment trees are indicated in parentheses, H5 HA tree (A/tern/South
Africa/61); PB1 and PA tree (A/equine/London/1416/73); NP and M
tree (A/equine/Prague/1/56)
b
A/duck/Hong Kong/Y280/97A/chicken/Guangdong/10/00
A/duck/Hong Kong/7/75A/duck/Hong Kong/Y439/97
A/duck/Hokkaido/49/98
100
99 A/duck/Hokkaido/49/98Achicken/Germany/R28/03
A/duck/Fujian/19/00A/duck/Vietnam/68/01 (H9N3)
A/duck/Nanchang/4-165/00A/guinea fowl/Hong Kong/38/02
A/duck/Vietnam/342/01 (H5N2)A/d k/Ji i/1742/03
99
84A/duck/Jiangxi/1742/03
A/duck/Vietnam/340/01 (H9N3)A/chicken/Korea/06/96
A/quail/Hong Kong/G1/97A/chicken/Guangxi/9/99
A/Hong Kong/2108/03A/teal/Hong Kong/W312/97
8268
100
99
A/Hong Kong/156/97A/chicken/Hong Kong/G9/97
A/goose/Guangdong/3/97A/goose/Guangdong/1/96
A/duck/Hong Kong/2986.1/00A/chicken/Hong Kong/715.5/01
A/duck/Guangxi/35/01
85
100
100
92
100
A/duck/Guangxi/35/01A/duck/Guangxi/22/01A/goose/Vietnam/113/01 (H5N1)A/goose/Vietnam/324/01 (H5N1)
A/duck/Zheijang/52/00A/chicken/Hong Kong/YU777/02A/duck/Guangxi/50/01
A/ hi k /H K /96 1/02
9099
77
10066
A/chicken/Hong Kong/96.1/02A/duck/Vietnam/NCVD1/03 (H5N1)
A/teal/China/2978.1/02A/chicken/Vietnam/NCVD8/03A/duck/Guangxi/2291/04
A/duck/China/E319-2/03A/duck/Vietnam/568/05
7586
0.02
A/chicken/Shantou/4231/03A/chicken/Korea/ES/03A/egret/Hong Kong/7573/02
A/Hong Kong/213/03A/duck/Hong Kong/821/02
A/Vietnam/3062/04 (H5N1)A/Vietnam/1203/04 (H5N1)
99
93A/Vietnam/1203/04 (H5N1)
A/chicken/Vietnam/27/03A/Vietnam/1194/04 (H5N1)
A/chicken/Vietnam/37/04A/mallard duck/Vietnam/133/0564
92
B: PB1
Fig. 1 continued
H5N1 avian influenza viruses from Vietnam 1253
123
A/Hong Kong/156/97A/teal/Hong Kong/W312/97A/quail/Hong Kong/G1/97
A/Hong Kong/1074/99A/duck/Hong Kong/Y280/97
98
100
A/chicken/Hong Kong/G9/97A/chicken/Guangdong/6/97
A/duck/Anyang/AVL-1/01A/duck/Fujian/1/02
A/pheasant/Hong Kong/sv674-15/02A/duck/Hong Kong/7/75
A/duck/Hokkaido/49/98
100100
100
A/duck/Hokkaido/447/00A/duck/Vietnam/68/01 (H9N3)
A/chicken/Otiva/8/04A/duck/Vietnam/340/01 (H9N3)
A/chicken/Guangdong/174/04A/duck/Hokkaido/84/02
A/turkey/Italy/4169/9983
9999
A/duck/Vietnam/342/01 (H5N2)A/mallard/Italy/37/02
A/chicken/Korea/323/96A/swine/Shandong/2/03A/goose/Guangdong/1/96
A/environment/Hong Kong/437-10/99A/chicken/Hong Kong/YU 822.2/0195
100
8491
100
A/duck/Shanghai/08/01A/duck/Guangxi/35/01
A/duck/Vietnam/NCVD1/03 (H5N1)A/partridge/Shantou/2886/03
A/duck/Guangxi/22/01A/goose/Vietnam/113/01 (H5N1)A/goose/Vietnam/324/01 (H5N1)
95
98
100
99 69
97
A/swine/Fujian/1/03A/swine/ Fujian/F1/03
A/chicken/Hong Kong/61-9/02 A/teal/China/2978.1/02
A/chicken/Vietnam/NCVD8/03A/goose/Guangxi/914/04
A/duck/China/E319-2/03
82100
99
94
10076
0.02
A/chicken/Fujian/1042/05A/brown headed gull/ 3/05A/chicken/Guangxi/604/05
A/Hong Kong/213/03A/egret/Hong Kong/7572/03
A/duck/Hong Kong/821/02A/Vietnam/1194/04 (H5N1)
99
9473
84
68A/Vietnam/1204/04 (H5N1)A/Vietnam/1203/04 (H5N1)
A/duck/Vietnam/11/04A/quail/Vietnam/36/04A/chicken/Vietnam/24/04
A/duck/Vietnam/367/05
80
94
100
C: PAFig. 1 continued
1254 S. J. Jadhao et al.
123
A/duck/Hong Kong/7/75A/chicken/Hong Kong/G9/97A/duck/Hong Kong/Y280/97
A/chicken/Hong Kong/FY150/01A/wild Duck/Shantou/4808/01
99
99
75
A/teal/Hong Kong/W312/97A/quail/Hong Kong/G1/97A/Hong Kong/1074/99A/Hong Kong/156/97
A/Hong Kong/483/97A/goose/Guangdong/1/96
A/duck/Hong Kong/ww461/00z
97
93
100
A/duck/Hong Kong/2986.1/00A/goose/Hong Kong/385.5/00
A/goose/Hong Kong/761/01A/goose/Vietnam/113/01 (H5N1)A/goose/Vietnam/324/01 (H5N1)
A/duck/Guangxi/22/01A/duck/Guangdong/22/02
99
96
66
91
99
99
/duc /Gua gdo g/ /0A/chicken/Hong Kong/YU822.2/01
A/aquatic bird/Hong Kong/399/99A/duck/Guangdong/40/00A/duck/Vietnam/342/01 (H5N2)
A/duck/Vietnam/340/01 (H9N3)A/Netherlands/33/03A/Netherlands/219/03
100
10099
76
9993
A/Netherlands/219/03A/chicken/Germany/R28/03
A/chicken/Nanchang/7-010/00A/quail/Nanchang/7-026/00
A/duck/Vietnam/68/01 (H9N3)A/turkey/Italy/220158/02/duck/Hong Kong/Y439/97
A/aquatic bird/Hong Kong/603/98
100
9387
8868 A/aquatic
A/duck/Zheijang/52/00A/teal/China/2978.1/02A/duck/Vietnam/NCVD1/03 (H5N1)
A/chicken/Vietnam/NCVD8/03A/duck/Guangxi/2291/04A/chicken/Shantou/4231/03A/chicken/Guangdong/174/04
8166
10084
9868
85 A/chicken/Guangdong/174/04A/bar headed goose/Qinghai/ 5/05
A/Hong Kong/213/03A/duck/Hong Kong/821/02
A/egret/Hong Kong/7572/03A/Vietnam/1194/04 (H5N1)
A/Vietnam/1203/04 (H5N1)
85
100
9868
68
A/duck/Guangxi/50/01A/duck/Guangdong/173/04
A/duck/China/E3192/03A/duck/Vietnam/568/05
A/goose/Guangxi/345/0576
71
680.02 A/Vietnam/3062/04 (H5N1)
D:NPFig. 1 continued
H5N1 avian influenza viruses from Vietnam 1255
123
A/chicken/Hong Kong/G9/97A/duck/Hong Kong/Y280/97
A/quail/Hong Kong/G1/97A/Hong Kong/156/9797
9999
A/teal/Hong Kong/w312/97A/Hong Kong/2108/03
A/duck/Hong Kong/7/75A/chicken/Korea/06/96
A/duck/Hong Kong/Y439/97A/duck/Primoriae/2633/01
A/duck/Vietnam/342/01 (H5N2)A/duck/Hong Kong/86/76
99
99
80
g gA/duck/Vietnam/68/01 (H9N3)
A/duck/Shantou/5048/01A/duck/Hokkaido/49/98
A/mallard/France/2525/01A/duck/Vietnam/340/01 (H9N3)
A/mallard/Netherlands/12/00A/wild duck/Shantou/988/00A/mallard/Bavaria/1/05
99
60
65
A/mallard/Bavaria/1/05A/wild duck/Shantou/1737/00
A/goose/Guangdong/1/96A/goose/Guangdong/3/97
A/duck/Anyang/AVL-1/01A/guinea fowl/Hong Kong/38/02
A/duck/Hong Kong/5734/01A/duck/Guangdong/22/02
A/chicken/Hong Kong/YU562/01
99
99
9971
A/chicken/HongA/duck/Guangxi/35/01A/duck/Hong Kong/3805/01
A/duck/Hong Kong/2986.1/00A/duck/Guangxi/22/01
A/goose/Vietnam/324/01 (H5N1)A/goose/Vietnam/113/01 (H5N1)
A/chicken/Guangdong/174/04
85
90
64
64
88
98 A/chicken/Vietnam/NCVD8/03A/teal/China/2978.1/02
A/duck/Vietnam/NCVD1/02 (H5N1)A/duck/Guangxi/2291/04
A/goose/Guangxi/914/04A/duck/Vietnam/568/05
A/chicken/Fujian/1042/05A/bar headedgoose/Qinghai/5/05
99
98
64
0.02
A/duck/Hong Kong/821/02A/egret/Hong Kong/757.3/02
A/Hong Kong/213/03A/chicken/Hong Kong/YU22/02
A/chicken//Shantou/4231/03A/duck/China/E319-2/03A/chicken/Indonesia/11/03
A/Vietnam/1203/04 (H5N1)
76
A/Vietnam/1203/04 (H5N1)A/Vietnam/1194/04 (H5N1)
A/mallard duck/Vietnam/133/05A/chicken/Vietnam/37/04A/Vietnam/3062/04 (H5N1)A/chicken/Vietnam/27/03
A/chicken/Vietnam/C58/04
64
E:MatrixFig. 1 continued
1256 S. J. Jadhao et al.
123
nucleotide homology, respectively) (phylogenetic tree not
shown). The M gene of Dk340 was most closely related to
A/mallard/Netherlands/12/00 (H7N3), and the NP gene
showed highest nucleotide sequence homology (Table 2)
with the A/Netherlands/33/03 (H7N7) and A/Netherlands/
219/03 (H7N7) viruses, which are known to have caused
human infection in the Netherlands in 2003 [10, 17].
Therefore, the NP and M gene phylogenies indicate that
Vietnamese Dk340 (H9N3) virus, A/mallard/Netherlands/
12/00 (H7N3) and A/Netherlands/33/03 (H7N7) virus had
similar ancestors for these two genes. Since Vietnam and
the the Netherlands are located on different continents, a
role of migratory birds and virus reassortments for inter-
continental spread of the progenitors of the NP and M gene
is suggested. The NS gene segment of the H9N3 viruses
belonged to allele B similar to that of HPAI A/goose/
Guangdong/1/96 (H5N1) virus. The genotyping of the H5
subtype viruses using the FluGenome tool [21] showed that
Dk68, a Vietnamese duck-origin H9N3 virus isolate
acquired its PB2 gene from a different lineage than the
H5N1 viruses (Table 1). Phylogenetically, the H5 HA gene
of the HPAI Gs113 and Gs324 viruses belonged to phylo-
genetic clade 3, whereas DkNCVD1 virus belonged to
phylogenetic clade 5 (Fig. 1a). Unlike H5N1 viruses bear-
ing clade 0, 1, 2 or 7 H5 HA, viruses with clade-3 and -5 HA
have not been reported to infect people to date [1, 11].
Molecular characterization
Hemagglutinin cleavage site and receptor binding domains
The H5 HA from geese and a duck H5N1 virus isolates
analyzed in this study have the polybasic amino acids motif
at the HA0 cleavage site, a characteristic of the HPAI
viruses in poultry (Fig. 2). The H5 HA of Gs113 and
Gs324 viruses possessed novel polybasic site, PRI-
ERRRKKR/GLF, with arginine (R) at the -9 position
(counting from the C terminus of HA1) instead of Q, which
is commonly observed in H5N1 HA cleavage sites of HPAI
viruses. The H5 HA cleavage site (PQREIRRKKR) of
DkNCVD1 H5N1 virus was identical to that of A/teal/
China/2978.1/02 [20]. The HA cleavage site sequence
PQRETR/GLF of DkVn342 virus was characteristic of
LPAI H5 virus and identical to that of A/duck/Primorie/
2633/01 (H5N3) virus. The H9 HA protein of Dk68 and
Dk340 H9N3 viruses exhibited connecting peptide
sequences that differed by a single amino acid at position
-2, [Dk68 (PAASSR) versus Dk340 (PAASGR)]. The
analysis of the available 948 H9 HA sequences of H9 virus
isolates from avian and mammalian species in the NCBI
GenBank indicated that although the significance is not
obvious, Serine at position -2 in the PAASSR/GLF
cleavage site is unique to the Dk68 H9 virus.Ta
ble
2H
igh
est
nu
cleo
tid
ese
qu
ence
ho
mo
log
yo
fH
5N
1v
iru
sg
enes
fro
mV
ietn
amto
the
infl
uen
zaA
vir
use
sse
qu
ence
sav
aila
ble
inG
enB
ank
Vir
us
PB
2P
B1
PA
HA
NP
NA
MN
S
gs1
13
(H5
N1
)
dk
/Gx
/22
/01
(H5
N1
)(9
9.0
0)a
dk
/Gx
/22
/01
(H5
N1
)(9
9.6
9)
dk
/Gx
/22
/01
(H5
N1
)(9
9.5
3)
dk
/Gx
/22
/01
(H5
N1
)(9
8.8
1)
dk
/Gx
/22
/01
(H5
N1
)
(98
.54
)
dk
/Gx
/22
/01
(H5
N1
)
(99
.08
)
dk
/HK
/29
86
.1/0
0
(H5
N1
)(9
9.3
9)
dk
/HK
/38
0.5
/01
(H5
N1
)(9
8.1
3)
gs3
24
(H5
N1
)
dk
/Gx
/22
/01
(H5
N1
)(9
9.3
5)
dk
/Gx
/22
/01
(H5
N1
)(9
9.7
7)
dk
/Gx
/22
/01
(H5
N1
)(9
9.4
8)
dk
/Gx
/22
/01
(H5
N1
)(9
8.9
3)
dk
/Gx
/22
/01
(H5
N1
)
(99
.13
)
dk
/Gx
/22
/01
(H5
N1
)
(99
.15
)
dk
/HK
/29
86
.1/0
0
(H5
N1
)(9
9.3
0)
dk
/HK
/38
0.5
/01
(H5
N1
)(9
8.1
3)
dk
NC
VD
1
(H5
N1
)
teal
/Ch
ina/
29
78
.1/
02
(H5
N1
)
(99
.30
)
teal
/Ch
ina/
29
78
.1/
02
(H5
N1
)
(99
.06
)
par
trid
ge/
ST
/
28
86
/03
(H5
N1
)(9
7.0
)
qu
ail/
Sh
anto
u/
38
46
/02
(H5
N1
)
(99
.14
)
teal
/Ch
ina/
29
78
.1/0
2
(H5
N1
)(9
9.6
0)
teal
/Ch
ina/
29
78
.1/0
2
(H5
N1
)(9
9.6
2)
qu
ail/
Sh
anto
u/3
84
6/0
2
(H5
N1
)(9
9.5
6)
qu
ail/
Sh
anto
u/
38
46
/02
(H5
N1
)
(99
.51
)
dk
34
2
(H5
N2
)
dk
/Tai
wan
/WB
29
/
99
(H6
N1
)
(98
.25
)
dk
/Gx
/22
/01
(H5
N1
)(9
6.7
0)
mal
lard
/Ita
ly/3
7/
02
(H5
N3
)
(97
.54
)
dk
/Pri
mo
rie/
26
21
/
01
(H5
N2
)
(98
.62
)
aqu
atic
bir
d/H
on
g
Ko
ng
/39
9/9
9(H
3N
8)
(97
.86
)
Du
ck/E
aste
rnC
hin
a/
72
9/0
3(H
6N
2)
(98
.37
)
dk
/Ho
kk
aid
o/8
6/0
1
(H2
N3
)(9
8.0
5)
mal
lard
/Ita
ly/4
3/0
1
(H7
N3
)(9
7.9
7)
dk
68
(H9
N3
)
ck/H
K/3
1.2
/02
(H5
N1
)(9
8.3
9)
dk
/Nan
chan
g/4
-
16
5/0
0(H
4N
6)
(96
.99
)
dk
/Ho
kk
aid
o/8
4/
02
(H5
N3
)
(98
.20
)
dk
/Ho
kk
aid
o/9
/99
(H9
N2
)(9
6.9
8)
ck/N
anch
ang
/7-0
10
/00
(H3
N6
)(9
7.7
2)
mal
lard
/Ita
ly/2
08
/00
(H5
N3
)(9
6.8
8)
dk
/Mo
ng
oli
a/5
4/0
1
(H5
N2
)(9
9.6
9)
dk
/Ho
kk
aid
o/1
20
/
01
(H6
N2
)
(99
.02
)
dk
34
0
(H9
N3
)
dk
/Zh
ejia
ng
/11
/00
(H5
N1
)(9
6.8
6)
dk
/Jia
ng
xi/
17
42
/03
(H7
N7
)(9
8.2
6)
dk
/Ho
kk
aid
o/4
47
/
00
(H5
N3
)
(98
.20
)
dk
/Ho
kk
aid
o/9
/99
(H9
N2
)(9
8.7
3)
Th
eN
eth
erla
nd
s/3
3/0
3
(H7
N7
)(9
7.7
2)
mal
lard
/Ita
ly/2
08
/00
(H5
N3
)(9
5.8
3)
mal
lard
/th
e
Net
her
lan
ds/
12
/00
(H7
N3
)(9
9.1
9)
dk
/Ho
kk
aid
o/4
47
/
00
(H5
N3
)
(99
.39
)
aO
nly
the
infl
uen
zav
iru
ses
wit
hh
igh
est
deg
ree
of
nu
cleo
tid
ese
qu
ence
ho
mo
log
yb
ased
on
the
NC
BI
Gen
Ban
kn
ucl
eoti
de
BL
AS
Tse
arch
anal
ysi
sar
ein
clu
ded
inth
eta
ble
H5N1 avian influenza viruses from Vietnam 1257
123

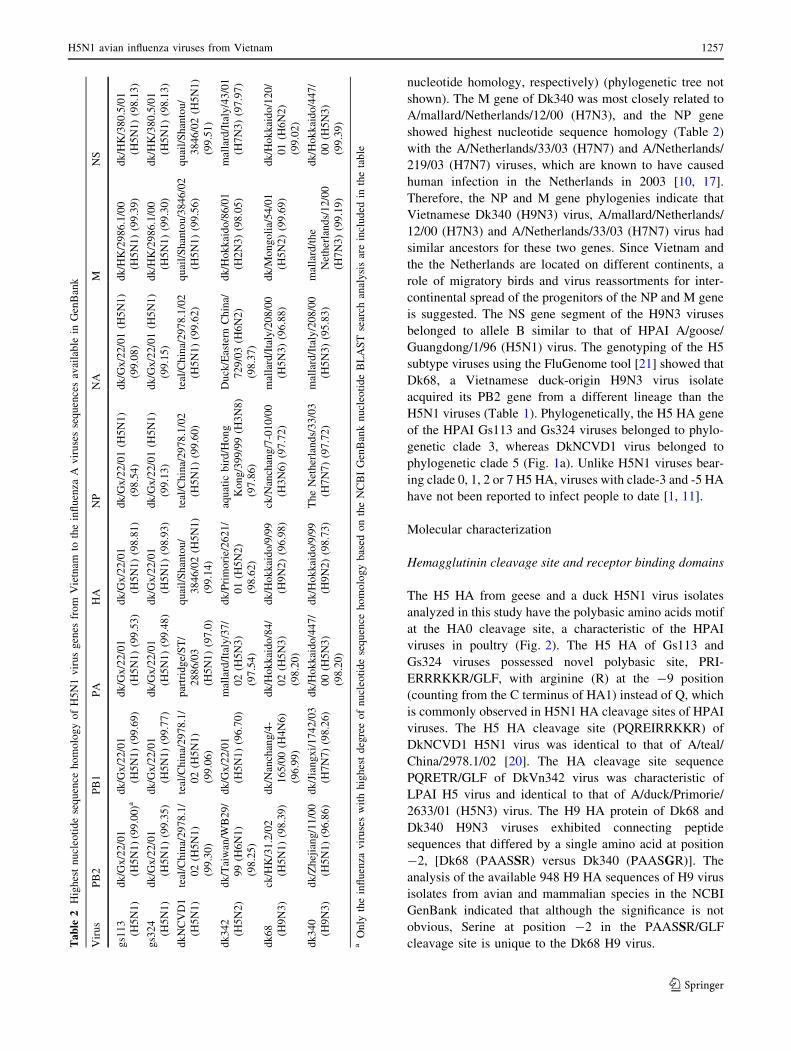
Fig. 2 Alignment and
comparison of HA amino acid
sequence of representative
H5N1 poultry viruses from
Vietnam in 2001 and 2003 and
in 2004–2005. The H5N1 virus
isolates from fatal human cases
are shown in bold type. Amino
acid sequences with a greenbackground represent potential
glycosylation sites, yellowindicates loss of glycosylation,
light blue indicates residues
involved in the receptor-binding
site, red/brick represents HA
sequence at the cleavage site,
and violet shows substitutions
among genotypes/lineages. A
consensus H5 HA amino acid
sequence was generated for
viruses known to have infected
humans in Vietnam during
2003–2005 (color figure online)
1258 S. J. Jadhao et al.
123
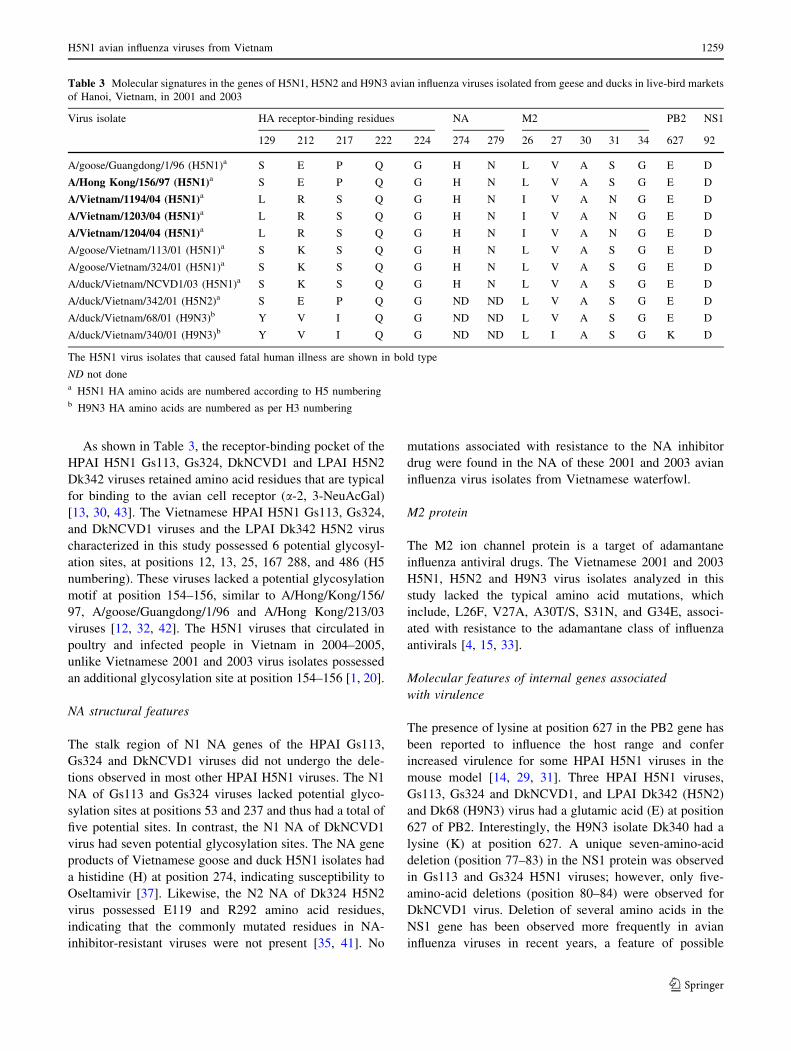
As shown in Table 3, the receptor-binding pocket of the
HPAI H5N1 Gs113, Gs324, DkNCVD1 and LPAI H5N2
Dk342 viruses retained amino acid residues that are typical
for binding to the avian cell receptor (a-2, 3-NeuAcGal)
[13, 30, 43]. The Vietnamese HPAI H5N1 Gs113, Gs324,
and DkNCVD1 viruses and the LPAI Dk342 H5N2 virus
characterized in this study possessed 6 potential glycosyl-
ation sites, at positions 12, 13, 25, 167 288, and 486 (H5
numbering). These viruses lacked a potential glycosylation
motif at position 154–156, similar to A/Hong/Kong/156/
97, A/goose/Guangdong/1/96 and A/Hong Kong/213/03
viruses [12, 32, 42]. The H5N1 viruses that circulated in
poultry and infected people in Vietnam in 2004–2005,
unlike Vietnamese 2001 and 2003 virus isolates possessed
an additional glycosylation site at position 154–156 [1, 20].
NA structural features
The stalk region of N1 NA genes of the HPAI Gs113,
Gs324 and DkNCVD1 viruses did not undergo the dele-
tions observed in most other HPAI H5N1 viruses. The N1
NA of Gs113 and Gs324 viruses lacked potential glyco-
sylation sites at positions 53 and 237 and thus had a total of
five potential sites. In contrast, the N1 NA of DkNCVD1
virus had seven potential glycosylation sites. The NA gene
products of Vietnamese goose and duck H5N1 isolates had
a histidine (H) at position 274, indicating susceptibility to
Oseltamivir [37]. Likewise, the N2 NA of Dk324 H5N2
virus possessed E119 and R292 amino acid residues,
indicating that the commonly mutated residues in NA-
inhibitor-resistant viruses were not present [35, 41]. No
mutations associated with resistance to the NA inhibitor
drug were found in the NA of these 2001 and 2003 avian
influenza virus isolates from Vietnamese waterfowl.
M2 protein
The M2 ion channel protein is a target of adamantane
influenza antiviral drugs. The Vietnamese 2001 and 2003
H5N1, H5N2 and H9N3 virus isolates analyzed in this
study lacked the typical amino acid mutations, which
include, L26F, V27A, A30T/S, S31N, and G34E, associ-
ated with resistance to the adamantane class of influenza
antivirals [4, 15, 33].
Molecular features of internal genes associated
with virulence
The presence of lysine at position 627 in the PB2 gene has
been reported to influence the host range and confer
increased virulence for some HPAI H5N1 viruses in the
mouse model [14, 29, 31]. Three HPAI H5N1 viruses,
Gs113, Gs324 and DkNCVD1, and LPAI Dk342 (H5N2)
and Dk68 (H9N3) virus had a glutamic acid (E) at position
627 of PB2. Interestingly, the H9N3 isolate Dk340 had a
lysine (K) at position 627. A unique seven-amino-acid
deletion (position 77–83) in the NS1 protein was observed
in Gs113 and Gs324 H5N1 viruses; however, only five-
amino-acid deletions (position 80–84) were observed for
DkNCVD1 virus. Deletion of several amino acids in the
NS1 gene has been observed more frequently in avian
influenza viruses in recent years, a feature of possible
Table 3 Molecular signatures in the genes of H5N1, H5N2 and H9N3 avian influenza viruses isolated from geese and ducks in live-bird markets
of Hanoi, Vietnam, in 2001 and 2003
Virus isolate HA receptor-binding residues NA M2 PB2 NS1
129 212 217 222 224 274 279 26 27 30 31 34 627 92
A/goose/Guangdong/1/96 (H5N1)a S E P Q G H N L V A S G E D
A/Hong Kong/156/97 (H5N1)a S E P Q G H N L V A S G E D
A/Vietnam/1194/04 (H5N1)a L R S Q G H N I V A N G E D
A/Vietnam/1203/04 (H5N1)a L R S Q G H N I V A N G E D
A/Vietnam/1204/04 (H5N1)a L R S Q G H N I V A N G E D
A/goose/Vietnam/113/01 (H5N1)a S K S Q G H N L V A S G E D
A/goose/Vietnam/324/01 (H5N1)a S K S Q G H N L V A S G E D
A/duck/Vietnam/NCVD1/03 (H5N1)a S K S Q G H N L V A S G E D
A/duck/Vietnam/342/01 (H5N2)a S E P Q G ND ND L V A S G E D
A/duck/Vietnam/68/01 (H9N3)b Y V I Q G ND ND L V A S G E D
A/duck/Vietnam/340/01 (H9N3)b Y V I Q G ND ND L I A S G K D
The H5N1 virus isolates that caused fatal human illness are shown in bold type
ND not donea H5N1 HA amino acids are numbered according to H5 numberingb H9N3 HA amino acids are numbered as per H3 numbering
H5N1 avian influenza viruses from Vietnam 1259
123

adaptation of these viruses to poultry [5, 44, 45]. The LPAI
Gs342 H5N2 and Dk68 H9N3 viruses did not show a
deletion in the NS1 gene. The NS1 protein of domestic-
waterfowl HPAI H5N1 and LPAI H5N2 and H9N3 viruses
analyzed in this study possessed aspartic acid (D) at posi-
tion 92. Pathogenesis studies using A/Hong Kong/156/97
(H5N1) 9 A/Puerto Rico/8/34 (H1N1) reassortant virus
carrying the A/Hong Kong/156/97 (H5N1) NS1 gene with
glutamic acid (E) at position 92 have shown enhanced
virulence in pigs [27].
In summary, our genetic analysis suggests that the
HPAI H5N1 viruses isolated from domestic waterfowl in
Hanoi, northern Vietnam, in 2001 and 2003 are not the
nearest progenitors of the HPAI H5N1 viruses that
caused widespread outbreaks in poultry and human
infections in Vietnam beginning in late 2003. The
phylogenetic analyses indicates that the progenitors of
these Vietnamese 2001 goose and 2003 duck HAPI
H5N1 virus isolates were most closely related to viruses
contemporarily prevalent among domestic waterfowl in
southern China. HPAI H5N1 viruses with HA genes
belonging to clade 3 and clade 5 have not been found to
infect people in Vietnam or elsewhere to date. Our data
suggest that clade-1 HPAI H5N1 viruses that caused
widespread H5N1 outbreaks in poultry and human
infections beginning in late 2003 did not evolve from
these 2001 goose and 2003 duck H5N1 viruses that were
detected in Hanoi, northern Vietnam. The phylogenetic
analysis and epidemiological data are consistent with the
notion that HPAI H5N1 clade 1 viruses were introduced
independently into Vietnam after 2002 [23, 40]. More
recently, clade 2.3.4 H5N1 viruses that were prevalent in
southern China have been introduced into northern
Vietnam, further underscoring the role of domestic
waterfowl and poultry movement in the spread of variant
H5N1 viruses [25].
Acknowledgments We thank the National Institute of Hygiene and
Epidemiology and National Center for Veterinary Diagnosis, Hanoi,
Vietnam, for invaluable leadership in conducting of outbreak inves-
tigations. SJJ and DCN were supported by International Emerging
Infectious Disease Fellowship at the US Centers for Disease Control
and Prevention and administered by the Association of Public Health
Laboratories, USA. We thank P. Rivailler for sequence data annota-
tion and management. The findings and conclusions in this report are
those of the authors and do not necessarily represent the views of the
Centers for Disease Control and Prevention or the Agency for Toxic
Substances and Disease Registry.
References
1. W.H.O Working Group (2005) Evolution of H5N1 avian influ-
enza viruses in Asia. Emerg Infect Dis 11:1515–1521
2. Abolnik C, Cornelius E, Bisschop SP, Romito M, Verwoerd D
(2006) Phylogenetic analyses of genes from South African LPAI
viruses isolated in 2004 from wild aquatic birds suggests intro-
duction by Eurasian migrants. Dev Biol 124:189–199
3. Bogner PCI, Lipman DJ, Cox NJ et al (2006) A global initiative
on sharing avian flu data. Nature 442:981
4. Bright RA, Medina MJ, Xu X, Perez-Oronoz G, Wallis TR, Davis
XM, Povinelli L, Cox NJ, Klimov AI (2005) Incidence of ada-
mantane resistance among influenza A (H3N2) viruses isolated
worldwide from 1994 to 2005: a cause for concern. Lancet
366:1175–1181
5. Cauthen AN, Swayne DE, Sekellick MJ, Marcus PI, Suarez DL
(2007) Amelioration of influenza virus pathogenesis in chickens
attributed to the enhanced interferon-inducing capacity of a virus
with a truncated NS1 gene. J Virol 81:1838–1847
6. Chen H, Deng G, Li Z, Tian G, Li Y, Jiao P, Zhang L, Liu Z,
Webster RG, Yu K (2004) The evolution of H5N1 influenza
viruses in ducks in southern China. Proc Natl Acad Sci USA
101:10452–10457
7. Chen H, Smith GJ, Li KS, Wang J, Fan XH, Rayner JM,
Vijaykrishna D, Zhang JX, Zhang LJ, Guo CT, Cheung CL, Xu
KM, Duan L, Huang K, Qin K, Leung YH, Wu WL, Lu HR,
Chen Y, Xia NS, Naipospos TS, Yuen KY, Hassan SS, Bahri S,
Nguyen TD, Webster RG, Peiris JS, Guan Y (2006) Establish-
ment of multiple sublineages of H5N1 influenza virus in Asia:
implications for pandemic control. Proc Natl Acad Sci USA
103:2845–2850
8. Claas EC, Osterhaus AD, van Beek R, De Jong JC, Rimmelzwaan
GF, Senne DA, Krauss S, Shortridge KF, Webster RG (1998)
Human influenza A H5N1 virus related to a highly pathogenic
avian influenza virus. Lancet 351:472–477
9. Coven F (2006) The situation of highly pathogenic avian influ-
enza (HPAI) outbreaks in Turkey. J Vet Med 53:34
10. Fouchier RA, Schneeberger PM, Rozendaal FW, Broekman JM,
Kemink SA, Munster V, Kuiken T, Rimmelzwaan GF, Schutten
M, Van Doornum GJ, Koch G, Bosman A, Koopmans M,
Osterhaus AD (2004) Avian influenza A virus (H7N7) associated
with human conjunctivitis and a fatal case of acute respiratory
distress syndrome. Proc Natl Acad Sci USA 101:1356–1361
11. Group WOFHNEW (2008) Toward a unified nomenclature sys-
tem for highly pathogenic avian influenza virus (H5N1). Emerg
Infect Dis 14:e1
12. Guan Y, Poon LL, Cheung CY, Ellis TM, Lim W, Lipatov AS,
Chan KH, Sturm-Ramirez KM, Cheung CL, Leung YH, Yuen
KY, Webster RG, Peiris JS (2004) H5N1 influenza: a protean
pandemic threat. Proc Natl Acad Sci USA 101:8156–8161
13. Ha Y, Stevens DJ, Skehel JJ, Wiley DC (2001) X-ray structures
of H5 avian and H9 swine influenza virus hemagglutinins bound
to avian and human receptor analogs. Proc Natl Acad Sci USA
98:11181–11186
14. Hatta M, Gao P, Halfmann P, Kawaoka Y (2001) Molecular basis
for high virulence of Hong Kong H5N1 influenza A viruses.
Science (New York, NY 293:1840–1842
15. Hay AJ, Zambon MC, Wolstenholme AJ, Skehel JJ, Smith MH
(1986) Molecular basis of resistance of influenza A viruses to
amantadine. J Antimicrob Chemother 18(Suppl B):19–29
16. Hoffmann E, Stech J, Guan Y, Webster RG, Perez DR (2001)
Universal primer set for the full-length amplification of all
influenza A viruses. Arch Virol 146:2275–2289
17. Koopmans M, Wilbrink B, Conyn M, Natrop G, van der Nat H,
Vennema H, Meijer A, van Steenbergen J, Fouchier R, Osterhaus
A, Bosman A (2004) Transmission of H7N7 avian influenza A
virus to human beings during a large outbreak in commercial
poultry farms in the Netherlands. Lancet 363:587–593
18. Kwon YK, Joh SJ, Kim MC, Sung HW, Lee YJ, Choi JG, Lee
EK, Kim JH (2005) Highly pathogenic avian influenza (H5N1) in
the commercial domestic ducks of South Korea. Avian Pathol
34:367–370
1260 S. J. Jadhao et al.
123

19. Lee CW, Suarez DL, Tumpey TM, Sung HW, Kwon YK, Lee YJ,
Choi JG, Joh SJ, Kim MC, Lee EK, Park JM, Lu X, Katz JM,
Spackman E, Swayne DE, Kim JH (2005) Characterization of
highly pathogenic H5N1 avian influenza A viruses isolated from
South Korea. J Virol 79:3692–3702
20. Li KS, Guan Y, Wang J, Smith GJ, Xu KM, Duan L, Rahardjo
AP, Puthavathana P, Buranathai C, Nguyen TD, Estoepangestie
AT, Chaisingh A, Auewarakul P, Long HT, Hanh NT, Webby RJ,
Poon LL, Chen H, Shortridge KF, Yuen KY, Webster RG, Peiris
JS (2004) Genesis of a highly pathogenic and potentially pan-
demic H5N1 influenza virus in eastern Asia. Nature 430:209–213
21. Lu G, Rowley T, Garten R, Donis RO (2007) FluGenome: a web
tool for genotyping influenza A virus. Nucleic Acids Res
35:W275–W279
22. Lvov DK, Yamnikova SS, Fedyakina IT, Lomakina NF (2000–
2002) Evolution of H4, H5 influenza A viruses in natural eco-
systems in Northern Eurasia. In: KawaokaY (ed) Proceedings
of the international conference on options for the Control of
influenza V Okinawa, Japan, 7–11 October 2003, International
Congress Series (2004), pp 169–173
23. Muramoto Y, Le TQ, Phuong LS, Nguyen T, Nguyen TH, Sakai-
Tagawa Y, Iwatsuki-Horimoto K, Horimoto T, Kida H, Kawaoka
Y (2006) Molecular characterization of the hemagglutinin and
neuraminidase genes of H5N1 influenza A viruses isolated from
poultry in Vietnam from 2004 to 2005. J Vet Med Sci 68:527–531
24. Nguyen DC, Uyeki TM, Jadhao S, Maines T, Shaw M, Matsuoka
Y, Smith C, Rowe T, Lu X, Hall H, Xu X, Balish A, Klimov A,
Tumpey TM, Swayne DE, Huynh LP, Nghiem HK, Nguyen HH,
Hoang LT, Cox NJ, Katz JM (2005) Isolation and characteriza-
tion of avian influenza viruses, including highly pathogenic
H5N1, from poultry in live bird markets in Hanoi, Vietnam, in
2001. J Virol 79:4201–4212
25. Nguyen TD, Nguyen TV, Vijaykrishna D, Webster RG, Guan Y,
Peiris JSM, Smith GJD (2008) Multiple sublineages of influenza
A virus (H5N1), Vietnam, 2005–2007. Emerg Infect Dis.
http://www.cdc.gov/EID/content/14/4/632.htm
26. Oprisan G, Coste H, Lupulescu E, Oprisoreanu AM, Szmal C,
Onita I, Popovici N, Ionescu LE, Bicheru S, Enache N, Ceianu C,
Czobor F, Olaru E, Alexandrescu V, Radu DL, Onu A, Popa MI
(2006) Molecular analysis of the first avian influenza H5N1
isolates from fowl in Romania. Roum Arch Microbiol Immunol
65:79–82
27. Seo SH, Hoffmann E, Webster RG (2004) The NS1 gene of
H5N1 influenza viruses circumvents the host anti-viral cytokine
responses. Virus Res 103:107–113
28. Shestopalov AM, Durimanov AG, Evseenko VA, Ternovoi VA,
Rassadkin YN, Razumova YV, Zaykovskaya AV, Zolotykh SI,
Netesov SV (2006) H5N1 influenza virus, domestic birds,
Western Siberia, Russia. Emerg Infect Dis 12:1167–1169
29. Shinya K, Hamm S, Hatta M, Ito H, Ito T, Kawaoka Y (2004)
PB2 amino acid at position 627 affects replicative efficiency, but
not cell tropism, of Hong Kong H5N1 influenza A viruses in
mice. Virology 320:258–266
30. Stevens J, Blixt O, Tumpey TM, Taubenberger JK, Paulson JC,
Wilson IA (2006) Structure and receptor specificity of the hem-
agglutinin from an H5N1 influenza virus. Science (New York)
312:404–410
31. Subbarao EK, London W, Murphy BR (1993) A single amino
acid in the PB2 gene of influenza A virus is a determinant of host
range. J Virol 67:1761–1764
32. Subbarao K, Klimov A, Katz J, Regnery H, Lim W, Hall H,
Perdue M, Swayne D, Bender C, Huang J, Hemphill M, Rowe T,
Shaw M, Xu X, Fukuda K, Cox N (1998) Characterization of an
avian influenza A (H5N1) virus isolated from a child with a fatal
respiratory illness. Science (New York) 279:393–396
33. Suzuki H, Saito R, Masuda H, Oshitani H, Sato M, Sato I (2003)
Emergence of amantadine-resistant influenza A viruses: epide-
miological study. J Infect Chemother 9:195–200
34. Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular
evolutionary genetics analysis (MEGA) software version 4.0.
Mol Biol Evol 24:1596–1599
35. Tisdale M (2000) Monitoring of viral susceptibility: new chal-
lenges with the development of influenza NA inhibitors. Rev Med
Virol 10:45–55
36. Tosh C, Murugkar HV, Nagarajan S, Bhatia S, Pateriya AK,
Behera P, Jain R, Kumar S, Khandia R, Vanamayya PR, Dubey
SC, Ahlawat SP (2007) Outbreak of avian influenza virus H5N1
in India. Vet Rec 161:279
37. Treanor JJ, Hayden FG, Vrooman PS, Barbarash R, Bettis R, Riff
D, Singh S, Kinnersley N, Ward P, Mills RG (2000) Efficacy and
safety of the oral neuraminidase inhibitor oseltamivir in treating
acute influenza: a randomized controlled trial. US Oral Neur-
aminidase Study Group. J Am Med Assoc 283:1016–1024
38. Van Borm S, Thomas I, Hanquet G, Lambrecht B, Boschmans M,
Dupont G, Decaestecker M, Snacken R, van den Berg T (2005)
Highly pathogenic H5N1 influenza virus in smuggled Thai
eagles, Belgium. Emerg Infect Dis 11:702–705
39. W.H.O. (2009) http://www.who.int/csr/disease/avian_influenza/
country/cases_table_2009_04_17/en/index.html
40. Wang J, Vijaykrishna D, Duan L, Bahl J, Zhang JX, Webster RG,
Peiris JS, Chen H, Smith GJ, Guan Y (2008) Identification of the
progenitors of Indonesian and Vietnamese avian influenza A
(H5N1) viruses from southern China. J Virol 82:3405–3414
41. Ward P, Small I, Smith J, Suter P, Dutkowski R (2005) Osel-
tamivir (Tamiflu) and its potential for use in the event of an
influenza pandemic. J Antimicrob Chemother 55(Suppl 1):i5–i21
42. Xu X, Subbarao CoxNJ, Guo Y (1999) Genetic characteriza-
tion of the pathogenic influenza A/Goose/Guangdong/1/96
(H5N1) virus: similarity of its hemagglutinin gene to those of
H5N1 viruses from the 1997 outbreaks in Hong Kong. Virol-
ogy 261:15–19
43. Yamada S, Suzuki Y, Suzuki T, Le MQ, Nidom CA, Sakai-
Tagawa Y, Muramoto Y, Ito M, Kiso M, Horimoto T, Shinya K,
Sawada T, Kiso M, Usui T, Murata T, Lin Y, Hay A, Haire LF,
Stevens DJ, Russell RJ, Gamblin SJ, Skehel JJ, Kawaoka Y
(2006) Haemagglutinin mutations responsible for the binding of
H5N1 influenza A viruses to human-type receptors. Nature
444:378–382
44. Zhu Q, Yang H, Chen W, Cao W, Zhong G, Jiao P, Deng G, Yu
K, Yang C, Bu Z, Kawaoka Y, Chen H (2008) A naturally
occurring deletion in its NS gene contributes to the attenuation of
an H5N1 swine influenza virus in chickens. J Virol 82:220–228
45. Zohari S, Gyarmati P, Thoren P, Czifra G, Brojer C, Belak S,
Berg M (2008) Genetic characterization of the NS gene indicates
co-circulation of two sub-lineages of highly pathogenic avian
influenza virus of H5N1 subtype in Northern Europe in 2006.
Virus Genes 36:117–125
H5N1 avian influenza viruses from Vietnam 1261
123




















