
General relations of stomatal responses to xylem sap abscisic acid under stress in the rooting zone...
13
Flora 202 (2007) 624–636 General relations of stomatal responses to xylem sap abscisic acid under stress in the rooting zone – A global perspective Hermann Heilmeier a, , Ernst-Detlef Schulze b , Jiang Fan c , Wolfram Hartung c a Interdisziplina¨resO ¨ kologisches Zentrum, TU Bergakademie Freiberg, Leipziger Str. 29, D-09599 Freiberg, Germany b Max-Planck-Institut fu¨r Biogeochemie, POB 100164, D-07701 Jena, Germany c Julius-von-Sachs-Institut der Universita¨t Wu¨rzburg, Lehrstuhl Botanik I, Julius-von-Sachs-Platz 2, D-97082 Wu¨rzburg, Germany Received 7 June 2007; accepted 7 June 2007 Dedicated to Prof. Dr. Dr. h.c. mult. Otto Ludwig Lange on the occasion of his 80th birthday Abstract A sensitivity factor that quantifies the responsiveness of stomata to xylem sap abscisic acid concentration ([ABA] xyl ) is described, using the relation between [ABA] xyl and maximum leaf conductance (g max ). Plotting g max against this factor results in a common linear relationship for woody and herbaceous species from boreal to (semi-) arid climates. The global distribution of the sensitivity factor reveals an unexpected pattern which is inverse to rainfall, i.e., plants in humid climates respond more sensitively to ABA than plants in arid areas. The implications for the response of natural vegetation and consequences for agriculture are discussed. r 2007 Elsevier GmbH. All rights reserved. Keywords: ABA; Irrigation; Leaf conductance; Plant functional types; Plant life forms; Stomatal sensitivity Introduction The role of abscisic acid (ABA) as a plant stress hormone is well established. Under a range of stress conditions ABA biosynthesis is initiated in different plant organs. After transport to target cells, such as stomata, plant water relations can be maintained or even improved by reduction of stomatal transpiration avoiding damage by desiccation (Davies and Zhang, 1991; Hartung and Davies, 1991; Schulze, 1993). Under mild stress conditions whenever the soil starts to dry, ABA is being transported from the root to the shoot and it thus acts as a long distance stress signal. Zhang and Davies (1987) were the first to describe the relation between ABA concentration in the xylem sap ([ABA] xyl ) and leaf conductance (g). For field-grown plants, the first relation between [ABA] xyl and maximum leaf conductance (g max ) was shown by Wartinger et al. (1990) in desert grown almond trees. They found a biphasic relation between [ABA] xyl and g max with concentration ranges of [ABA] xyl for both highly and less sensitively responding stomata. The relation between [ABA] xyl and g max has been investigated since then for a large range of plants, both herbaceous and woody, originating from moderate and extreme habitats (for references see Table 1). It turned out that in many cases the relationship between [ABA] xyl and g max was very similar – indicating a general principle for the stress physiological role of ABA within terrestrial plants. ARTICLE IN PRESS www.elsevier.de/flora 0367-2530/$ - see front matter r 2007 Elsevier GmbH. All rights reserved. doi:10.1016/j.flora.2007.06.002 Corresponding author. Tel.: +49 3731 39 3208; fax: +49 3731 39 3012. E-mail address: [email protected] (H. Heilmeier).
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of General relations of stomatal responses to xylem sap abscisic acid under stress in the rooting zone...

ARTICLE IN PRESS
0367-2530/$ - se
doi:10.1016/j.flo
�Correspondfax: +49 3731 3
E-mail addr
Flora 202 (2007) 624–636
www.elsevier.de/flora
General relations of stomatal responses to xylem sap abscisic acid under
stress in the rooting zone – A global perspective
Hermann Heilmeiera,�, Ernst-Detlef Schulzeb, Jiang Fanc, Wolfram Hartungc
aInterdisziplinares Okologisches Zentrum, TU Bergakademie Freiberg, Leipziger Str. 29, D-09599 Freiberg, GermanybMax-Planck-Institut fur Biogeochemie, POB 100164, D-07701 Jena, GermanycJulius-von-Sachs-Institut der Universitat Wurzburg, Lehrstuhl Botanik I, Julius-von-Sachs-Platz 2, D-97082 Wurzburg, Germany
Received 7 June 2007; accepted 7 June 2007
Dedicated to Prof. Dr. Dr. h.c. mult. Otto Ludwig Lange on the occasion of his 80th birthday
Abstract
A sensitivity factor that quantifies the responsiveness of stomata to xylem sap abscisic acid concentration ([ABA]xyl)is described, using the relation between [ABA]xyl and maximum leaf conductance (gmax). Plotting gmax against thisfactor results in a common linear relationship for woody and herbaceous species from boreal to (semi-) arid climates.The global distribution of the sensitivity factor reveals an unexpected pattern which is inverse to rainfall, i.e., plants inhumid climates respond more sensitively to ABA than plants in arid areas. The implications for the response of naturalvegetation and consequences for agriculture are discussed.r 2007 Elsevier GmbH. All rights reserved.
Keywords: ABA; Irrigation; Leaf conductance; Plant functional types; Plant life forms; Stomatal sensitivity
Introduction
The role of abscisic acid (ABA) as a plant stresshormone is well established. Under a range of stressconditions ABA biosynthesis is initiated in differentplant organs. After transport to target cells, such asstomata, plant water relations can be maintained oreven improved by reduction of stomatal transpirationavoiding damage by desiccation (Davies and Zhang,1991; Hartung and Davies, 1991; Schulze, 1993).
Under mild stress conditions whenever the soil startsto dry, ABA is being transported from the root to theshoot and it thus acts as a long distance stress signal.
e front matter r 2007 Elsevier GmbH. All rights reserved.
ra.2007.06.002
ing author. Tel.: +49 3731 39 3208;
9 3012.
ess: [email protected] (H. Heilmeier).
Zhang and Davies (1987) were the first to describe therelation between ABA concentration in the xylem sap([ABA]xyl) and leaf conductance (g). For field-grownplants, the first relation between [ABA]xyl and maximumleaf conductance (gmax) was shown by Wartinger et al.(1990) in desert grown almond trees. They found abiphasic relation between [ABA]xyl and gmax withconcentration ranges of [ABA]xyl for both highly andless sensitively responding stomata.
The relation between [ABA]xyl and gmax has beeninvestigated since then for a large range of plants, bothherbaceous and woody, originating from moderate andextreme habitats (for references see Table 1). It turnedout that in many cases the relationship between [ABA]xyland gmax was very similar – indicating a general principlefor the stress physiological role of ABA within terrestrialplants.

ARTICLE IN PRESS
Table
1.
Listofplants
includingtheirreferenceswhichwereusedto
establish
therelationofFig.3
Species
Life
form
PFT
Location
ofgrowth
Rooting
condition
Drivingfactor
Sampling
ofxylem
sapat
gmax
Methodof
xylem
sap
sampling
gmax
Sensitivity
factor
Reference
Aca
cia
con
fusa
WT
GP
Soildrought,split
root
No
PP
0.14
�0.0835
LiangandZhang(1999)
Aca
cia
con
fusa
WT
GP
Soilcompaction,soil
drought
No
PP
0.08
�0.8618
Lianget
al.(1996)
Aca
cia
con
fusa
WT
GP
Splitroot
No
PP
0.09
�0.0470
Lianget
al.(1997)
Ace
rp
seu
do
pla
tan
us
WN
GP
Splitroot
Yes
PS
0.10
�0.1993
KhalilandGrace
(1993)
Ace
rru
bru
mW
NF
SSoildrought
No
PS
0.16
�0.1115
Augeet
al.(2000)
Ace
rsa
cch
aru
mW
NG
PSoildrought
No
PS
0.14
�0.0420
Loew
ensteinandPallardy
(1998)
Bet
ula
pen
du
laW
NG
PSplitroot
No
PS
0.38
�0.2592
Fort
etal.(1998)
Bu
xu
sm
icro
ph
yll
aW
SF
PSoildrought
Yes
PS
0.09
�0.0374
AbrilandHanano(1998)
Ca
psi
cum
an
nu
um
HL
GP
Rootrestriction,soil
drought
Yes
PS
0.60
�0.4378
IsmailandDavies(1998)
Ca
psi
cum
an
nu
um
HL
GP
Soildrought
Yes
PS
0.50
�0.3968
Ismailet
al.(2002)
Ca
psi
cum
an
nu
um
HL
GP
Soildrought
Yes
PS
0.40
�0.3205
Ismailet
al.(2002)
Ca
sta
nea
sati
vaW
WG
PSplitroot
Yes
PS
0.50
�0.4522
Maurelet
al.(2004)
Cea
no
thu
sth
yrs
iflo
rus
WS
FP
Soildrought
No
PS
0.10
�0.0545
Tenhunen
etal.(1994)
Cea
no
thu
sth
yrs
iflo
rus
WS
FP
Soildrought
Yes
PS
0.15
�0.0508
AbrilandHanano(1998)
Co
rnu
sfl
ori
da
WN
FS
Soildrought
No
PS
0.16
�0.1150
Augeet
al.(2000)
Gly
cin
em
ax
HL
GP
Soildrought
No
PP
0.54
�0.2468
Liu
etal.(2003)
Gly
cin
em
ax
HL
GP
Soildrought
No
PP
0.56
�0.2100
Liu
etal.(2005a)
Hel
ian
thu
sa
nn
uu
sH
LG
PPressurizedpot
No
PC
0.65
�0.3673
Schurr
etal.(1992)
Hel
ian
thu
sa
nn
uu
sH
LG
PSoildrought
No
PS
2.25
�1.4382
TardieuandSim
onneau
(1998)
Hel
ian
thu
sa
nn
uu
sH
LF
PSoildrought
No
PS
2.25
�1.6579
TardieuandSim
onneau
(1998)
Ho
rdeu
mvu
lga
reH
3G
PSoildrought
No
PS
0.55
�0.3673
Borelet
al.(1997)
Ho
rdeu
mvu
lga
reH
3G
PSoildrought
No
PS
0.55
�0.3976
Borelet
al.(1997)
Ju
gla
ns
nig
raW
NG
PSoildrought
No
PS
0.04
�0.0255
Loew
ensteinandPallardy
(1998)
Leu
caen
ale
uco
cep
ha
laW
TG
PSplitroot,soil
drought
No
PP
0.21
�0.1420
LiangandZhang(1999)
Leu
caen
ale
uco
cep
ha
laW
TG
PSplitroot,soil
drought
No
PP
0.21
�0.2284
LiangandZhang(1999)
H. Heilmeier et al. / Flora 202 (2007) 624–636 625

ARTICLE IN PRESS
Table
1.(c
on
tin
ued
)
Species
Life
form
PFT
Location
ofgrowth
Rooting
condition
Drivingfactor
Sampling
ofxylem
sapat
gmax
Methodof
xylem
sap
sampling
gmax
Sensitivity
factor
Reference
Leu
caen
ale
uco
cep
ha
laW
TG
PSplitroot
No
PP
0.18
�0.0953
Lianget
al.(1997)
Liq
uid
am
ba
rst
yra
cifl
ua
WN
FS
Soildrought
No
PS
0.50
�0.1676
Augeet
al.(2000)
Lit
sea
glu
tin
osa
WT
GP
Soilcompaction,soil
drought
No
PP
0.10
�0.0787
Lianget
al.(1996)
Lu
pin
us
alb
us
HL
GP
Soildrought
Yes
PS
0.80
�0.4210
CorreiaandPereira
(1995)
Nic
oti
an
a
plu
mb
ag
inif
oli
a
HL
GP
Soildrought
No
PS
1.50
�0.7314
Borelet
al.(2001)
Pic
easi
tch
ensi
sW
EF
PSoildrought
No
PS
0.15
�0.1959
Jacksonet
al.(1995)
Pin
us
sylv
estr
isW
EF
PSoildrought
No
PS
0.80
�0.4521
Jacksonet
al.(1995)
Pin
us
sylv
estr
isW
EF
SSoildrought
No
PS
0.26
�0.2100
Sturm
etal.(1998)
Po
pu
lus
trem
ula
WN
FS
Soildrought
Yes
PS
0.28
�0.0663
Aasamaaet
al.(2004)
Po
pu
lusspec.
Wn.d.
Fn.d.
Soildrought
Yes
n.d.
0.40
�0.2351
TardieuandSim
onneau
(1998)
Pru
nu
sa
viu
mW
NF
SSoildrought
No
PS
0.50
�0.1701
HartungandHeilm
eier
(1993)
Pru
nu
sd
ulc
isW
WF
SSoildrought
Yes
PS
0.40
�0.2948
Wartinger
etal.(1990)
Pru
nu
sp
ersi
caW
WF
SSoildrought
Yes
PS
0.20
�0.2513
Correiaet
al.(1997)
Qu
ercu
sa
lba
WN
GS
Soildrought
No
PS
0.60
�0.2976
Augeet
al.(2000)
Qu
ercu
sp
rin
us
WN
FS
Soildrought
No
PS
0.50
�0.3667
Augeet
al.(2000)
Sa
lix
nig
raW
NG
PSoildrought
No
PS
1.12
�0.5352
Loew
ensteinandPallardy
(1998)
So
lan
um
tub
ero
sum
HL
GP
Soildrought
No
PP
0.75
�0.6102
Liu
etal.(2005b)
Til
iaco
rda
taW
NF
SSoildrought
No
PS
0.05
�0.0607
Niinem
etset
al.(1999)
Tri
foli
um
sub
terr
an
eum
HL
FS
Soildrought
No
PS
1.00
�0.6096
Sociaset
al.(1997)
Vig
na
un
gu
icu
lata
HL
GP
Soildrought
Yes
PS
0.34
�0.1631
Duanet
al.(1996)
Vig
na
un
gu
icu
lata
HL
GP
Soildrought
No
PS
0.22
�0.0922
Ebel
etal.(1997)
Vit
isvi
nif
era
WW
FS
Soildrought
Yes
PS
0.46
�0.5804
Correiaet
al.(1995)
Zea
ma
ys
H4
GP
Osm
oticstress
Yes
PP
0.08
�0.7474
Stikic
andDavies(2000)
Zea
ma
ys
H4
GP
Osm
oticstress
Yes
PP
0.07
�0.0544
Stikic
andDavies(2000)
Zea
ma
ys
H4
GS
Soilcompaction,soil
drought
Yes
PS
0.24
�0.1163
TardieuandDavies(1992)
Zea
ma
ys
H4
FS
Soilcompaction,soil
drought
No
PS
0.27
�0.1740
Tardieuet
al.(1992)
Zea
ma
ys
H4
FS
Soildrought,ABA
feeding
Yes
PS
0.28
�0.1396
Tardieuet
al.(1993)
Zea
ma
ys
H4
FP
ABA
feeding
No
Exudate
0.13
�0.0429
ZhangandDavies(1990)
Additionally,life
form
(herbaceous,H,andwoody,W)andplantfunctionaltype(PFT;3:cultivatedC3grasses,4:cultivatedC4grasses,L:broad-leaved
crops,N:temperate
deciduousbroad-leaved
trees,W:warm
deciduousbroad-leaved
trees,S:sclerophyllousscrub,E:evergreen
coniferoustrees,T:tropicaltrees),locationofgrowth
(greenhouse,G,vs.field,F),rootingconditions(pots,P,or
unrestricted
naturalsoil,S),drivingfactorforthevariationin
xylem
sapABA
concentrationandleafconductance,coincidence
ofxylem
sapsamplingandmaxim
um
leafconductance
(gmax)and
methodofxylem
sapsampling(PC:balancingpressure
inPassioura-typepressure
chamber,PP:pressurizingdetopped
potted
plantin
pressure
chamber,PS:pressurizingexcisedshootorleafwith
pressure
chamber)are
given
(n.d.:nodata).
H. Heilmeier et al. / Flora 202 (2007) 624–636626

ARTICLE IN PRESSH. Heilmeier et al. / Flora 202 (2007) 624–636 627
In the following paper, (1) it will be tested whethersuch a general relation exists, (2) such a relationship willbe quantified for individual species on the basis of anewly defined stomatal sensitivity factor, and (3) thehypothesis will be tested that the stomatal sensitivityfactor is related to physiological performance of theplants as indicated by maximum leaf conductance. Inaddition the relationship between the stomatal sensitiv-ity factor and global vegetation patterns will beinvestigated, and practical agricultural implications ofsuch a sensitivity factor will be discussed.
The importance of stomatal sensitivity to
[ABA]xyl
Already in 1975, Raschke pointed out that stomataresponded to ABA when intercellular CO2 was increasedalthough bulk ABA content of the leaves remainedunchanged (Raschke, 1975). He explained this phenom-enon by a significantly increased stomatal sensitivity toABA. Later, Burschka et al. (1983) confirmed theimportance of changing sensitivity of stomata forArbutus unedo plants growing in a natural macchia.Despite the high amounts of leaf ABA which againshowed small diurnal fluctuations only, the injection oftiny amounts of ABA only to the petiole resulted indrastic responses to the applied ABA. This responsediffered during the course of the day pointing to achange of sensitivity. Diurnal variation in the sensitivityof stomata to xylem sap ABA under field conditions hasalso been found by Tardieu and Davies (1992) andCorreia et al. (1995, 1997). The more pronouncedstomatal response to [ABA]xyl in the afternoon in maizeplants in the experiments of Tardieu and Davies (1992)was attributed to sensitising of stomata by lowepidermal water potentials. In contrast, in field-grownalmond trees leaf conductance was suppressed by high[ABA]xyl during daily courses at leaf water potentials(cl) below �3.2MPa only, whereas for a less critical leafwater status stomatal response to diurnal changes in[ABA]xyl was independent of cl (Hartung and Heilmeier,1993). This agrees with a unique relationship betweenleaf conductance and [ABA]xyl regardless of cl in therange from 0 to �3MPa for sunflower plants subjectedto a variety of treatments (Tardieu et al., 1996). On theother hand, for the same species Schurr et al. (1992)found contrasting sensitivities of stomatal opening to[ABA]xyl as dependent on xylem sap pH, nitrate orcalcium concentration. Alkalinisation of xylem sap is acommon effect of various environmental stressors(Hartung and Radin, 1989; Wilkinson et al., 1998);however, stomatal response to elevated xylem pH isvariable (Dodd, 2005). Bahrun et al. (2002) propose asequence of events for maize plants responding to
progressive soil moisture deficit, starting with a decreasein xylem nitrate concentration and a corresponding shiftin xylem pH, which initiate de novo ABA synthesis inroots and ABA release into the xylem, causing stomatalconductance to decrease before a fall in leaf waterpotential. Also environmental conditions such astemperature and vapour pressure deficit can influencethe apparent stomatal sensitivity to ABA. These andother examples are cited and discussed by Trejo et al.(1993, 1995), Dodd et al. (1996) and Dodd (2003, 2005).
As for diurnal courses, a number of studies shows aclose relationship between xylem sap ABA concentra-tion and leaf conductance for an extended period, e.g.,during soil drying for several days to weeks. Periods ofhigh maximum leaf conductances (gmax) and low[ABA]xyl during early stages of soil drying followed bylow gmax and high [ABA]xyl during periods with waterdeficits have been documented both for woody (Augeet al., 2000; Jackson et al., 1995; Khalil and Grace, 1993;Liang and Zhang, 1999; Liang et al., 1996, 1997; Sturmet al., 1998; Wartinger et al., 1990) and herbaceousspecies (Correia and Pereira, 1995; Ismail and Davies,1998; Ismail et al., 2002; Liu et al., 2003, 2005a; Sociaset al., 1997). In these cases, it is essential to excludeshort-term sources of environmental variability like thetremendous effect of humidity on stomata first docu-mented by the group of Otto L. Lange (e.g., Lange et al.,1971). However, using a standardized protocol in long-term experiments, e.g., measurement of g at saturatinglight intensity, but non-limiting VPD, should allow toreveal a possible general relationship between gmax and[ABA]xyl. Dodd (2005) concludes that ‘‘in a givenspecies, this relationship is commonly unified acrossdifferent growing conditions, from leaf to leaf onindividual plants, and from day to day as the plantdevelops’’. In the following these individual responsepatterns for a number of plant species from theliterature will be synthesised into a general stresssensitivity framework.
Processing the data
A literature study was performed to select experi-ments in which plants subjected to different stresstreatments (e.g., drought, soil compaction, split-rootsystems, osmotic stress, feeding of ABA) were assayedfor both leaf conductance and xylem sap ABAconcentration during the whole treatment period. Table1 gives the references of those data analysed in thisstudy. Three case studies only did not show a relation-ship between g and [ABA]xyl (Triboulot et al., 1996:Quercus spec.; Niinemets et al., 1999: Populus tremula;Sobeih et al., 2004: Lycopersicon esculentum). Because asensitivity factor could not be calculated, these studies
ARTICLE IN PRESSH. Heilmeier et al. / Flora 202 (2007) 624–636628
were not included. At present we have no explanationfor these missing relation, special experimental condi-tions may be the reason. When there were several datasets available from any one publication (Correia et al.,1995, 1997; Tardieu and Simonneau, 1998; Tardieuet al., 1992, 1993), e.g., for diurnal courses, data fromthe morning were selected when minimum influence ofVPD or cl was assumed.
Experiments were classified into the driving factor forvariation in g and [ABA]xyl, location of growth (green-house or field), rooting conditions (grown in pots withsoil or in unrestricted soil at the natural site). In additionplants were separated into herbaceous or woody growthforms, and into eight plant functional types (cultivatedC3 and C4 grasses, broad-leaved crops, temperatedeciduous broad-leaved trees, warm deciduous broad-leaved trees, tropical trees, sclerophyllous scrub, ever-green coniferous trees). Thus plant species in this studycover a wide range of life forms from annual crop plantsto long-living trees, and they grow from humid to semi-arid climates (Table 1). Nevertheless, among herbaceousplants the database shows a bias towards agriculturalspecies, especially broadleaved crops. For trees, broad-leaved deciduous species predominate.
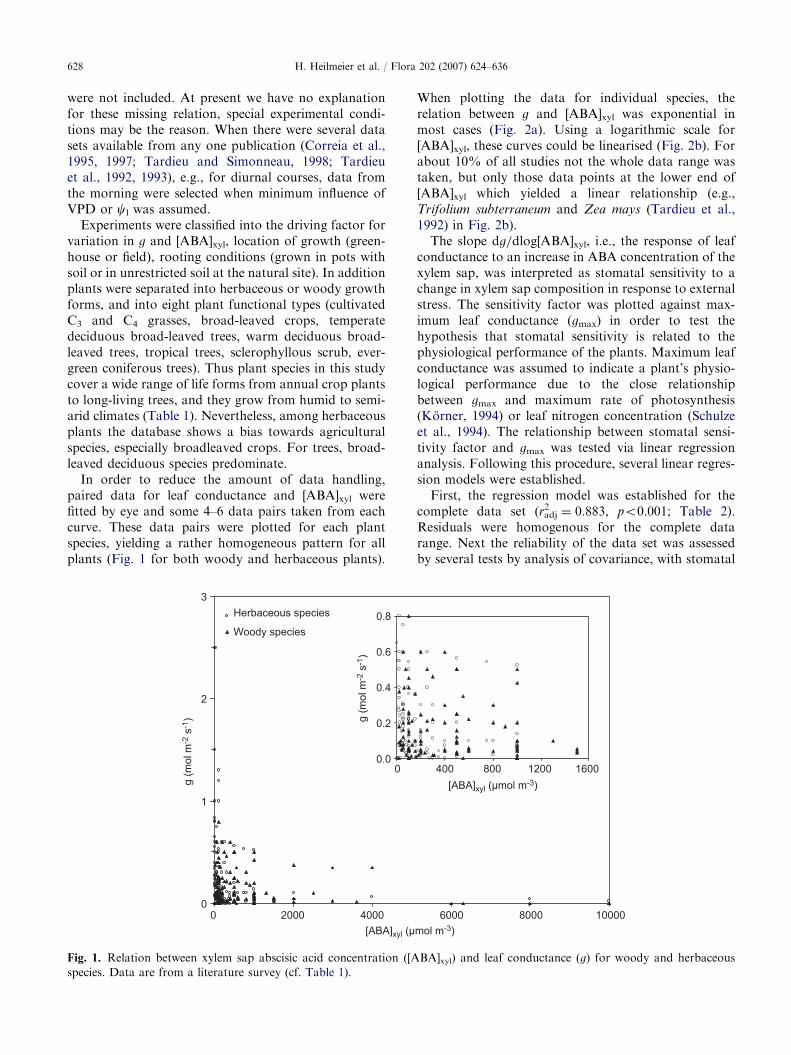
In order to reduce the amount of data handling,paired data for leaf conductance and [ABA]xyl werefitted by eye and some 4–6 data pairs taken from eachcurve. These data pairs were plotted for each plantspecies, yielding a rather homogeneous pattern for allplants (Fig. 1 for both woody and herbaceous plants).
0
1
2
3
0
0
g (
mol m
-2 s
-1) g (
mol m
-2 s
-1)
[ABA]xyl (µ
40002000
Herbaceous species
Woody species
0.8
0.6
0.4
0.2
0.0
Fig. 1. Relation between xylem sap abscisic acid concentration ([A
species. Data are from a literature survey (cf. Table 1).
When plotting the data for individual species, therelation between g and [ABA]xyl was exponential inmost cases (Fig. 2a). Using a logarithmic scale for[ABA]xyl, these curves could be linearised (Fig. 2b). Forabout 10% of all studies not the whole data range wastaken, but only those data points at the lower end of[ABA]xyl which yielded a linear relationship (e.g.,Trifolium subterraneum and Zea mays (Tardieu et al.,1992) in Fig. 2b).
The slope dg/dlog[ABA]xyl, i.e., the response of leafconductance to an increase in ABA concentration of thexylem sap, was interpreted as stomatal sensitivity to achange in xylem sap composition in response to externalstress. The sensitivity factor was plotted against max-imum leaf conductance (gmax) in order to test thehypothesis that stomatal sensitivity is related to thephysiological performance of the plants. Maximum leafconductance was assumed to indicate a plant’s physio-logical performance due to the close relationshipbetween gmax and maximum rate of photosynthesis(Korner, 1994) or leaf nitrogen concentration (Schulzeet al., 1994). The relationship between stomatal sensi-tivity factor and gmax was tested via linear regressionanalysis. Following this procedure, several linear regres-sion models were established.
First, the regression model was established for thecomplete data set (radj
2¼ 0.883, po0.001; Table 2).
Residuals were homogenous for the complete datarange. Next the reliability of the data set was assessedby several tests by analysis of covariance, with stomatal
mol m-3)
[ABA]xyl (µmol m-3)
1000080006000
16001200800400
BA]xyl) and leaf conductance (g) for woody and herbaceous

ARTICLE IN PRESS
3
2
1
00 100 200 300 400 500 600 700 800 900
g (
mol m
-2 s
-1)
3
2
1
00 1 2 3 4
g (
mol m
-2 s
-1)
Helianthus annuus (Tardieu and Simonneau, 1998)
Trifolium subterraneum (Socias et al., 1997)
Zea mays (Tardieu and Davies, 1992)
Zea mays (Tardieu et al., 1992)
Zea mays (Tardieu et al., 1993)
Helianthus annuus (Tardieu and Simonneau, 1998)
Trifolium subterraneum (Socias et al., 1997)
Zea mays (Tardieu and Davies, 1992)
Zea mays (Tardieu et al.,1992)
Zea mays (Tardieu et al., 1993)
ABAxyl (µmol m-3)
log (ABAxyl (µmol m-3))
Fig. 2. Relation between xylem sap abscisic acid concentration ([ABA]xyl) and leaf conductance (g) for single-herbaceous species
grown in the field (a) on a linear scale, (b) on a logarithmic scale for [ABA]xyl.
Table 2. Linear regression model for stomatal sensitivity as
dependent on maximum leaf conductance (gmax)
Effect Coefficient Std error t P
Constant 0.000 0.020 �0.015 0.988
gmax �0.624 0.031 �19.846 o0.001
Dependent variable: stomatal sensitivity, N: 54, adjusted R2: 0.883.
H. Heilmeier et al. / Flora 202 (2007) 624–636 629
sensitivity as the dependent factor, different experimen-tal procedures or plant types as grouping factor andgmax as covariate.
Goodger et al. (2005) had pointed out that differenttechniques of xylem sap sampling may yield contrastingconcentrations e.g., of ABA for both well-watered andwater-stressed plants. Therefore, first the effect of xylemsap sampling on stomatal sensitivity was tested.Although the main effect of sampling technique wasnot significant, there was a significant interactionbetween sampling method and gmax (Table 3a), due toa relatively low sensitivity for Helianthus annuus xylemsap recovered under balancing pressure in a Passioura-type pressure chamber (Schurr et al., 1992).
Second, in view of contrasting patterns of diurnalcourses for both g and [ABA]xyl documented in theliterature it was necessary to test whether the time ofsampling xylem sap and measuring g was of relevance.For example, Tardieu et al. (1991), Tenhunen et al.(1994) and Jackson et al. (1995) showed an increasingtrend for [ABA]xyl during a daily course with g
decreasing in the afternoon, whereas Correia et al.(1997) found a steady decrease in g with constant[ABA]xyl during the day. Therefore, the complete dataset was divided into experiments in which, according tothe long-term pattern between [ABA]xyl and gmax firstestablished by Wartinger et al. (1990), both variableswere measured at the time of gmax on the one handand the rest on the other hand (see Table 1). Therewas no significant effect of the experimental protocol(Table 3b). Therefore, we conclude for our data set thatboth long-term experiments in which there was a clearrelationship between gmax and [ABA]xyl and experimentsin which there was an impact of environmental factorsvarying on a short time scale like VPD or plant-internalfactors like leaf water potential do not differ withrespect to their effect on stomatal sensitivity.

ARTICLE IN PRESS
Table 3. Analysis of covariance for the effect of method of xylem sap sampling, time of xylem sap sampling, driving factor for
variation in gmax and xylem sap ABA concentration, growth conditions, plant growth form and plant functional type (grouping
factors) on stomatal sensitivity as dependent on maximum leaf conductance (covariate)
Source Sum-of-squares df Mean-square F-ratio P
(a) Effect of method of xylem sap sampling: pressurizing leaf (n ¼ 16) or shoot (n ¼ 23), pressurizing detopped plants (n ¼ 12),
Passioura-type pressure chamber (n ¼ 1), root exudate (n ¼ 1)
Sampling method 0.004 2 0.002 0.199 0.820
gmax (covariate) 0.035 1 0.035 3.731 0.060
Sampling method� gmax 0.171 4 0.043 4.612 0.003
Error 0.416 45 0.009
Dependent variable: stomatal sensitivity, N: 53, squared multiple R: 0.918
(b) Effect of time of xylem sap sampling: coincidence with maximum leaf conductance (n ¼ 18) or not (n ¼ 32)
Sampling time 0.004 1 0.004 0.448 0.507
gmax (covariate) 1.097 1 1.097 118.5 o0.001
Sampling time� gmax 0.001 1 0.001 0.073 0.789
Error 0.426 46 0.009
Dependent variable: stomatal sensitivity, N: 50, squared multiple R: 0.913
(c) Effect of driving factor: soil drought (n ¼ 37), split-root system+soil drought (n ¼ 8), soil compaction (n ¼ 4), pressurized pot
(n ¼ 1), osmotic stress (n ¼ 2), ABA feeding (n ¼ 2)
Driving factor 0.003 3 0.001 0.074 0.990
gmax (covariate) 0.001 1 0.001 0.132 0.718
Driving factor� gmax 0.036 5 0.007 0.743 0.596
Error 0.423 44 0.010
Dependent variable: stomatal sensitivity, N: 54, squared multiple R: 0.916
(d) Effect of location of growth: greenhouse (n ¼ 31) vs. field (n ¼ 23)
Location of growth 0.009 1 0.009 0.872 0.355
gmax (covariate) 4.488 1 4.488 422.4 o0.001
Growth location� gmax 0.051 1 0.051 4.799 0.033
Error 0.531 50 0.011
Dependent variable: stomatal sensitivity, N: 54, squared multiple R: 0.895
(e) Effect of rooting conditions: pots (n ¼ 37) vs. unrestricted soil (n ¼ 16)
Rooting conditions 0.011 1 0.011 1.067 0.307
gmax (covariate) 4.349 1 4.349 407.8 o0.001
Rooting� gmax 0.059 1 0.059 5.546 0.023
Error 0.523 49 0.011
Dependent variable: stomatal sensitivity, N: 53, squared multiple R: 0.896
(f) Effect of plant growth form: woody (n ¼ 32) vs. herbaceous (n ¼ 22)
Growth form 0.041 1 0.041 3.736 0.059
gmax (covariate) 2.064 1 2.064 189.5 o0.001
Growth form� gmax 0.034 1 0.034 3.132 0.083
Error 0.544 50 0.011
Dependent variable: stomatal sensitivity, N: 54, squared multiple R: 0.892
(g) Effect of plant functional type PFT: cultivated C3 (n ¼ 2) and C4 (n ¼ 6) grasses, broad-leaved crops (n ¼ 14), temperate
deciduous broad-leaved trees (n ¼ 14), warm deciduous broad-leaved trees (n ¼ 4), tropical trees (n ¼ 7), sclerophyllous scrub
(n ¼ 3), evergreen coniferous trees (n ¼ 3)
Plant functional type 0.056 6 0.009 0.922 0.490
gmax (covariate) 0.035 1 0.035 3.467 0.070
PFT� gmax 0.084 7 0.012 1.188 0.333
Error 0.384 38 0.010
Dependent variable: stomatal sensitivity, N: 53, squared multiple R: 0.924
H. Heilmeier et al. / Flora 202 (2007) 624–636630

ARTICLE IN PRESS
0
y (
dg d
(logA
BA
)-1)
H. Heilmeier et al. / Flora 202 (2007) 624–636 631
Third, the driving factor that caused variation in g
and [ABA]xyl was categorised into the six groups soildrying, split-root experiments with soil drought, soilcompaction, Passioura-type pressurized pot, osmoticstress and ABA feeding (Table 1). There was nosignificant effect of the driving factor on stomatalsensitivity nor a significant interaction between drivingfactor and gmax (Table 3c).
Fourth, it was tested whether stomatal sensitivity ofplants grown in the greenhouse differed from thosegrown under field conditions, using maximum leafconductance as a covariate. There was no significantmain effect; however a significant interaction betweengrowth location and gmax (Table 3d). Plants grown inthe field (n ¼ 23) had a higher stomatal sensitivity factor(�0.705, s.e. ¼ 0.047) compared to greenhouse-grownplants (�0.569, s.e. ¼ 0.040; n ¼ 31).
Fifth, when plants grown in pots were compared tothose grown in unrestricted soil at the natural site, therewas no significant main effect (Table 3e). However, againthe interaction between rooting medium and gmax wassignificant, with plants grown in unrestricted soil (n ¼ 16)responding more sensitive (�0.719, s.e. ¼ 0.058) thanplants grown in pots (�0.569, s.e. ¼ 0.036; n ¼ 37).
Finally, the effect of plant growth form and func-tional type (PFT) on stomatal sensitivity was tested.There was no difference in stomatal sensitivity betweenherbaceous and woody plants (Table 3f) nor betweendifferent PFT (Table 3g).
Thus, we could not find a main effect of experimentalprocedure nor plant type on stomatal sensitivity inour data set. Since interaction effects, when present,were of minor significance in most cases (p40.01),we conclude that for the complete data set sensitivityto ABA did not change dramatically with environmentalor experimental conditions nor depend on speciesidentity.
-2
-1
0 1 2 3
Sto
mata
l se
nsitiv
it
Maximum leaf conductance (mol m-2 s-1)
cultivated C3 grasses
warm deciduous
cultivated C4 grasses
broadleaved trees
evergreen coniferous
trees tropical trees
sclerophyllous scrub
broadleaved treestemperate deciduous
broadleaved crops
Fig. 3. Relation between maximum leaf conductance (gmax)
and stomatal sensitivity (dg/d log[ABA]xyl) for different plant
functional types (PFT). The effect of PFT was statistically not
significant (cf. Table 3g).
Responsiveness of stomata to stress in the
rooting zone: stomatal sensitivity
Examples of the relation between g and [ABA]xylshow the similarity between plants (Fig. 1). For bothherbaceous and woody plants, leaf conductance de-creases dramatically at xylem sap ABA concentrationsabove 1mmolm�3. However, herbaceous plants reachhigher maximum leaf conductances, and they respondearlier to an increase in xylem sap ABA as shown by thesteeper decline. Woody species, on the other hand,maintain higher leaf conductance at [ABA]xyl between0.5 and 4mmolm�3.
Linearisation of these curves (cf. Fig. 2 for herbaceousplants grown in the field) reveals a distinct pattern: InH. annuus, the species with the highest maximum leaf
conductance (2.25molm�2 s�1) leaf conductance de-creases much steeper with increasing [ABA]xyl than inother species with lower gmax. The three experimentswith Z. mays yielded similar gmax and slopes for g versuslog[ABA]xyl (Fig. 2b).
As may be inferred from Fig. 1, the relationshipbetween g and log[ABA]xyl showed similar patternsfor all species studied: for species with higher gmax, theslope (dg/d log[ABA]xyl) was steeper and vice versa ascan be concluded from Fig. 2. Plotting dg/d log[ABA]xylas a function of gmax for all data available from theliterature survey resulted in a common linear relation-ship for all plant functional types (Fig. 3). The slopedg/d log[ABA]xyl describes the responsiveness of thestomata to various external stresses that affect the waterrelations of the plant (drought, salt, heat, coldness, etc.).It indicates whether plants would respond with high(more negative slope) or low sensitivity (less negativeslope) to stress. It is called ‘‘sensitivity factor’’.
The significance of stomatal sensitivity to ABA hasalready been discussed by Trejo et al. (1993, 1995)without showing a general relationship. In the presentstudy, the relationship between plant physiologicalperformance and stomatal sensitivity seems to beindependent of genetic or environmental influences(see Table 3). Rather it may be general for a wide rangeof plant and vegetation types.

ARTICLE IN PRESSH. Heilmeier et al. / Flora 202 (2007) 624–636632
Ecological relevance of stomatal sensitivity
The general relationship between gmax as an indicatorof plant physiological performance and stomatal sensi-tivity as shown in Fig. 3 evokes a number of ecologicalimplications. Schulze et al. (1994) have presented a closelinear relationship between gmax and leaf nitrogen (N)concentration for a broad range of vegetation types withdifferent leaf longevities. Based on the global distribu-tion of leaf N concentration, they produced a globalmap for gmax. Because of the linear relationship betweengmax and the sensitivity factor the latter can betransferred to the global map of gmax by Schulze et al.(1994) as shown in Fig. 4. Regions with the mostsensitive stomata are those dominated by intensiveagricultural land use in the mid latitudes of NorthAmerica, Europe, Western and Eastern Asia, andsubtropical Southern Asia (India). Furthermore, thetropical deciduous forests in Central and South Americaand Africa show high stomatal sensitivity. Cold-temperate grasslands in Central Asia are major ecosys-tems with medium stomatal sensitivity. For major partsof tropical rainforests, boreal deciduous and evergreenforests, semiarid and arid savannahs and deserts andalpine ecosystems stomatal sensitivity is low. Theseareas are characterized by various stress factors in therooting zone, e.g., low nutrient availability, soil droughtand low temperatures.
In the case of plants growing in dry or extremely dryhabitats, no direct relationships between gmax and[ABA]xyl have been published, mainly because theharvest of xylem sap from these plants is not possibleor extremely difficult. This is (1) because of the anatomyof the plants and (2) because of the extremely low xylemwater potential that makes the harvest of xylem sap bypressurising such plants a serious problem. We areaware of only one experiment in which leaf conductancewas recorded after feeding three different concentrationsof ABA to the xylem sap of a desert plant (Anastatica
hierochuntica, Hartung et al., 1990). However, since thein planta [ABA]xyl is not known in this case, data werenot included in the present study.
It is known that in plants from arid habitatsABA concentrations in the xylem sap often areextremely high with small fluctuations only (ownunpublished data for Hammada scoparia: 670 mmolm�3,Pulicaria crispa: 700 mmolm�3, Reaumuria negevensis:1400 mmolm�3, Zygophyllum dumosum: 1400 mmolm�3,Atriplex halinus: 1000–2000 mmolm�3, Artemisia herba-
alba: 1800–2000 mmolm�3). For these desert plantsgrowing in the Negev (Israel) extremely high leafdiffusion resistances have been measured by Schulzeet al. (1972, 1973). Similar relations also have beendetected with poikilohydric angiosperms from SouthernAfrica and Western Australia (Schiller et al., 1997). Toour surprise, it emerges that the ABA root signal may
not be very effective in desert plants as can be inferredfrom their low sensitivity factor. Under severe stressconditions, as they are typical for deserts, ABA mayplay a more important role for acquiring or maintaininga high desiccation tolerance by formation of dehydrinsor other protective compounds rather than in regulatingleaf conductance (Hartung et al., 1998; Heilmeier andHartung, 2001). A significant part of this ABA may besynthesised in the leaves.
If in agricultural areas gmax of crop plants would bemeasured with a porometer on a sunny day duringthe late morning hours, the linear relation of Fig. 3could be taken as a calibration line to estimate thesensitivity coefficient. In the range of gmax from 0.8 to1.8molm�2 s�1, the sensitivity would be sufficiently highto indicate further stomatal closure. Thus, a moderateirrigation would be sufficient to prevent negativeconsequences for productivity. Consequently, gmax
values below 1molm�2 s�1 could be avoided where thesmall sensitivity of the stomatal system would requiremuch higher irrigation to reverse the response. Thiswould be relatively ineffective. The sunflower plantswithin the gmax range above 1.8molm�2 s�1 originatefrom a greenhouse and a field study on sufficientlyirrigated plots where drought stress does not haveserious negative effects on crop production. Since therelation of Fig. 3 seems to be valid for nearly all climatesand plant functional types, the calibration line could beused for terrestrial plants nearly worldwide and could behelpful for farmers especially in semiarid and aridclimates. Thus besides modern irrigation techniquessuch as partial root zone drying (PRD, Davies et al.,2005) the use of the sensitivity factor could play a partfor performing a sustainable agriculture.
From an ecosystem point of view, it would beinteresting whether a general relationship exists betweensoil water content and stomatal response to xylem sapABA. However, only few data have been publishedshowing the relation between the soil water content (y)and [ABA]xyl. The five data sets evaluated yielded ahyperbolic shape which could be linearised by plottinglog[ABA]xyl against log(y). Although these data trans-formation resulted in straight lines with different slopes,no pattern related to plant life form or experimentalconditions emerged. In greenhouse experiments withplants grown in pots Vigna unguiculata (Duan et al.,1996; Ebel et al., 1997) reacted more sensitively thanHordeum vulgare (Borel et al., 1997) and sycamoreseedlings (Acer pseudoplatanus, Khalil and Grace, 1993).An intermediate response was found for Scots pine(Pinus sylvestris) growing at its natural site (Sturm et al.,1998). Because of the very small number of data, it is notpossible to perform a similar analysis as in the case of g
vs. [ABA]xyl, where a large body of data is available.However, taking in mind that soil water status and g areclosely related (Schulze, 1986) one also may expect

ARTICLE IN PRESS
Fig.4.Theglobaldistributionofthesensitivityfactord
g/d
log[A
BA] xyl.Productionofthemapwasbasedontheglobaldistributionmapfor
gmaxbySchulzeet
al.(1994)and
applyingalinearregressionto
thedata
from
Fig.3yieldingthemodel
dg
/dlog[A
BA] xyl¼
0.000–0.624
gmax(cf.Table
2).
H. Heilmeier et al. / Flora 202 (2007) 624–636 633

ARTICLE IN PRESSH. Heilmeier et al. / Flora 202 (2007) 624–636634
a similarly close relation between y and [ABA]xyl.Consequently, the global distribution of the sensitivity[ABA]xyl (d log[ABA]xyl/d log y) versus log(ymax) shouldresemble that of dg/d log[ABA]xyl vs. gmax as shown inFig. 3.
Following this analysis, it emerges that the ABA rootsignal is mainly important for fast-growing herbaceousspecies from mesic environments with high rates ofwater flow and a fast root-to-shoot communication. Incontrast, if the relationship depicted in Fig. 3 is furthersubstantiated with experimental data and proves to bewidely applicable one may conclude that for plants frommore stressful habitats with low water flow and aconservative water use, just as much as for very tallspecies (e.g., Sequoiadendron) the ABA root signal maynot be very effective. In these cases, other factors, suchas ABA production or release within the leaf, may bemore important.
Acknowledgement
We thank Kerstin Sickel, MPI for Biogeochemistry,for help to prepare the global ABA-sensitivity map.
References
Aasamaa, K., Sober, A., Hartung, W., Niinemets, U., 2004.
Drought acclimation of two deciduous tree species of
different layers in a temperate forest canopy. Trees 18,
93–101.
Abril, M., Hanano, R., 1998. Ecophysiological responses of
three evergreen woody Mediterranean species to water
stress. Acta Oecol. 19, 377–387.
Auge, R.M., Green, C.D., Stodola, A.J.W., Saxton, A.M.,
Olinick, J.B., Evans, R.M., 2000. Correlations of stomatal
conductance with hydraulic and chemical factors in several
deciduous tree species in a natural habitat. New Phytol.
145, 483–500.
Bahrun, A., Jensen, C.R., Asch, F., Mogensen, V.O., 2002.
Drought-induced changes in xylem pH, ionic composition,
and ABA concentration act as early signals in field-grown
maize (Zea mays L.). J. Exp. Bot. 53, 251–263.
Borel, C., Simonneau, T., This, D., Tardieu, F., 1997.
Stomatal conductance and ABA concentration in the xylem
sap of barley lines of contrasting genetic origin. Austr.
J. Plant Physiol. 24, 607–615.
Borel, C., Frey, A., Marion-Poll, A., Tardieu, F., Simonneau, T.,
2001. Does engineering abscisic acid biosynthesis in Nicotiana
plumbaginifolia modify stomatal response to drought? Plant
Cell Environ. 24, 477–489.
Burschka, C., Tenhunen, J.D., Hartung, W., 1983. Diurnal
variations in abscisic acid content and stomatal response to
applied abscisic acid in leaves of irrigated and non-irrigated
Arbutus unedo plants under naturally fluctuating environ-
mental conditions. Oecologia 58, 128–131.
Correia, M.J., Pereira, J.S., 1995. The control of leaf
conductance of white lupin by xylem ABA concentration
decreases with the severity of water deficits. J. Exp. Bot. 46,
101–110.
Correia, M.J., Pereira, J.S., Chaves, M.M., Rodrigues, M.L.,
Pacheco, C.A., 1995. ABA xylem concentrations determine
maximum daily leaf conductance of field grown Vitis
vinifera L. plants. Plant Cell Environ. 18, 511–521.
Correia, M.S., Rodrigues, M.L., Ferreira, M.I., Pereira, J.S.,
1997. Diurnal change in the relationship between stomatal
conductance and abscisic acid in the xylem sap of field
grown peach trees. J. Exp. Bot. 48, 1727–1736.
Davies, W.J., Zhang, J., 1991. Root signals and the
regulation of growth and development of plants in
drying soil. Annu. Rev. Plant Physiol. Plant Mol. Biol.
42, 55–76.
Davies, W.J., Kudoyarova, G., Hartung, W., 2005. Long
distance ABA signalling and its relation to other signalling
pathways in the detection of soil drying and the mediation
of the plant’s response to drought. J. Plant Growth Regul.
24, 285–295.
Dodd, I.C., 2003. Hormonal interactions and stomatal
responses. J. Plant Growth Regul. 22, 32–46.
Dodd, I.C., 2005. Root-to-shoot signalling: assessing the roles
of ‘up’ in the up and down world of long-distance signalling
in planta. Plant Soil 274, 251–270.
Dodd, I.C., Stikic, R., Davies, W.J., 1996. Chemical regulation
of gas exchange and growth of plants in drying soil in the
field. J. Exp. Bot. 47, 1475–1490.
Duan, X., Nemann, D.S., Reiber, J.M., Green, C.D.,
Saxton, A.M., Auge, R.M., 1996. Mycorrhizal influence
on hydraulic and hormonal factors implicated in the
control of stomatal conductance during drought. J. Exp.
Bot. 47, 1541–1550.
Ebel, R.C., Duan, X., Still, D.W., Auge, R.M., 1997. Xylem
sap abscisic acid concentration and stomatal conductance
of mycorrhizal Vigna unguiculata in drying soil. New
Phytol. 135, 755–761.
Fort, C., Muller, F., Label, P., Granier, A., Dreyer, E., 1998.
Stomatal conductance, growth and root signalling in Betula
pendula seedlings subjected to partial soil drying. Tree
Physiol. 18, 769–777.
Goodger, J.Q.D., Sharp, R.E., Marsh, E.L., Schachtman, D.P.,
2005. Relationships between xylem sap constituents and
leaf conductance of well-watered and water-stressed maize
across three xylem sap sampling techniques. J. Exp. Bot. 56,
2389–2400.
Hartung, W., Davies, W.J., 1991. Drought-induced changes in
physiology and ABA. In: Davies, W.J., Jones, H.G. (Eds.),
Abscisic Acid – Physiology and Biochemistry. Bios
Scientific Publishers, Oxford, pp. 63–79.
Hartung, W., Heilmeier, H., 1993. Stomatal responses to
abscisic acid in natural environments. In: Jackson, M.B.,
Black, C.R. (Eds.), Interacting Stresses on Plants in a
Changing Climate. NATO-ASI Series, I 16. Springer,
Heidelberg, pp. 524–542.
Hartung, W., Radin, J.W., 1989. Abscisic acid in the
mesophyll apoplast and in the root xylem sap of water-
stressed plants: the significance of pH gradients. Curr.
Topics Plant Biochem. Physiol. 8, 110–124.

ARTICLE IN PRESSH. Heilmeier et al. / Flora 202 (2007) 624–636 635
Hartung, W., Heilmeier, H., Wartinger, A., Kettemann, I.,
Schulze, E.-D., 1990. Ionic and abscisic acid relationships
of Anastatica hierochuntica L. under arid conditions. Israel
J. Bot. 39, 373–382.
Hartung, W., Schiller, P., Dietz, K.J., 1998. Physiology of
poikilohydric plants. Progr. Bot. 59, 299–327.
Heilmeier, H., Hartung, W., 2001. Survival strategies under
extreme and complex environmental conditions: the aquatic
resurrection plant Chamaegigas intrepidus. Flora 196,
245–260.
Ismail, M.R., Davies, W.J., 1998. Root restriction affects leaf
growth and stomatal response: the role of xylem sap ABA.
Sci. Horticult. 74, 257–268.
Ismail, M.R., Davies, W.J., Awad, M.H., 2002. Leaf growth
and stomatal sensitivity to ABA in droughted pepper
plants. Sci. Horticult. 96, 313–327.
Jackson, G.E., Irvine, J., Grace, J., Khalil, A.A.M., 1995.
Abscisic acid concentrations and fluxes in droughted
conifer saplings. Plant Cell Environ. 18, 13–22.
Khalil, A.A.M., Grace, J., 1993. Does xylem sap ABA control
the stomatal behaviour of water stressed sycamore (Acer
pseudoplatanus L.) seedlings. J. Exp. Bot. 44, 1127–1134.
Korner, C., 1994. Leaf diffusive conductances in the
major vegetation types of the globe. In: Schulze, E.-D.,
Caldwell, M.M. (Eds.), Ecophysiology of Photosynthesis.
Ecol. Stud. 100. Springer, Berlin-Heidelberg-New York,
pp. 463–490.
Lange, O.L., Losch, R., Schulze, E.-D., Kappen, L., 1971.
Responses of stomata to changes in humidity. Planta 100,
76–86.
Liang, J., Zhang, J., 1999. The relation of stomatal closure and
reopening to xylem ABA concentration and leaf water
potential during soil drying and rewatering. Plant Growth
Regul. 29, 77–86.
Liang, J., Zhang, J., Wong, M.H., 1996. Stomatal conductance
in relation to xylem sap ABA concentrations in two tropical
trees, Acacia confusa and Litsea glutinosa. Plant Cell
Environ. 19, 93–100.
Liang, J., Zhang, J., Wong, M.H., 1997. Can stomatal closure
caused by xylem ABA explain the inhibition of leaf
photosynthesis under soil drying? Photosynth. Res. 51,
149–159.
Liu, F., Jensen, C.R., Andersen, M.N., 2003. Hydraulic and
chemical signals in the control of leaf growth and stomatal
conductance in soybean exposed to drought stress. Funct.
Plant Biol. 30, 65–73.
Liu, F., Andersen, M.N., Jacobsen, S.E., Jensen, C.R., 2005a.
Stomatal control and water use efficiency of soybean
(Glycine max L. Merr.) during progressive soil drying.
Environ. Exp. Bot. 54, 33–40.
Liu, F., Jensen, C.R., Shahanzari, A., Andersen, M.N.,
Jacobsen, S.E., 2005b. ABA regulated stomatal control
and photosynthetic water use efficiency of potato (Solanum
tuberosum L.) during progressive soil drying. Plant Sci. 168,
831–836.
Loewenstein, N.J., Pallardy, S.G., 1998. Drought tolerance,
xylem sap abscisic acid and stomatal conductance
during soil drying. A comparison of young plants of
four temperate deciduous angiosperms. Tree Physiol. 18,
421–430.
Maurel, M., Robin, C., Simonneau, T., Loustau, D.,
Dreyer, E., Desprez-Loustau, M.L., 2004. Stomatal con-
ductance and root-to-shoot signalling in chestnut saplings
exposed to Phytophtora cinnamomi or partial soil drying.
Funct. Plant Biol. 31, 41–51.
Niinemets, U., Sober, A., Kull, O., Hartung, W., Tenhunen, J.D.,
1999. Apparent controls on leaf conductance by soil water
availability and via light-acclimation of foliage structural and
physiological properties in a mixed deciduous, temperate
forest. Int. J. Plant Sci. 160, 707–721.
Raschke, K., 1975. Simultaneous requirement of carbon
dioxide and abscisic acid for stomatal closure in Xanthium
strumarium. Planta 125, 243–259.
Schiller, P., Heilmeier, H., Hartung, W., 1997. Abscisic acid
(ABA) relations of the aquatic resurrection plant Chamae-
gigas intrepidus under naturally fluctuating environmental
conditions. New Phytol. 136, 603–611.
Schulze, E.-D., 1986. Carbon dioxide and water vapor
exchange in response to drought in the atmosphere and in
the soil. Annu. Rev. Plant Physiol. 37, 247–274.
Schulze, E.-D., 1993. Soil water deficits and atmospheric
humidity as environmental signals. In: Smith, J.A.C.,
Griffith, H. (Eds.), Water Deficits: Plant Responses from
Cell to Community. Bios Scientific Publishers, Oxford,
pp. 129–145.
Schulze, E.-D., Lange, O.L., Koch, W., 1972. Okophysiolo-
gische Untersuchungen an Wild- und Kulturpflanzen der
Negev-Wuste. III. Tageslaufe von Nettophotosynthese und
Transpiration am Ende der Trockenzeit. Oecologia 9,
317–340.
Schulze, E.-D., Lange, O.L., Kappen, L., Buschbom, U.,
Evenari, M., 1973. Stomatal responses to changes in
temperature at increasing water stress. Planta 110, 29–42.
Schulze, E.-D., Kelliher, F.M., Korner, C., Lloyd, J.,
Leuning, R., 1994. Relationships among maximum stoma-
tal conductance, ecosystem surface conductance, carbon
assimilation rate, and plant nitrogen nutrition: a global
ecology scaling exercise. Annu. Rev. Ecol. Syst. 25,
629–660.
Schurr, U., Gollan, T., Schulze, E.-D., 1992. Stomatal
response to drying soil in relation to changes in the xylem
sap composition of Helianthus annuus. II. Stomatal
sensitivity to abscisic acid imported from the xylem sap.
Plant Cell Environ. 15, 561–567.
Sobeih, W.Y., Dodd, I.C., Bacon, M.A., Grierson, D.,
Davies, W.J., 2004. Long-distance signals regulating
stomatal conductance and leaf growth in tomato (Lyco-
persicon esculentum) plants subjected to partial root-zone
drying. J. Exp. Bot. 55, 2353–2363.
Socias, X., Correia, M.J., Chaves, M., Medrano, H., 1997.
The role of abscisic acid and water relations in
drought responses of subterranean clover. J. Exp. Bot. 48,
1281–1288.
Stikic, R., Davies, W.J., 2000. Stomatal reactions of two
different maize lines to osmotically induced drought stress.
Biol. Plant. 43, 399–405.
Sturm, N., Kostner, B., Hartung, W., Tenhunen, J.D., 1998.
Environmental and endogenous controls on leaf- and
stand-level water conductance in a Scots pine plantation.
Ann. Sci. For. 55, 237–253.

ARTICLE IN PRESSH. Heilmeier et al. / Flora 202 (2007) 624–636636
Tardieu, F., Davies, W.J., 1992. Stomatal responses to abscisic
acid is a function of current plant water status. Plant
Physiol. 98, 540–545.
Tardieu, F., Simonneau, T., 1998. Variability among species of
stomatal control under fluctuating soil water status and
evaporative demand: modelling isohydric and anisohydric
behaviours. J. Exp. Bot. 49, 419–432.
Tardieu, F., Katerji, N., Bethenod, O., Zhang, J., Davies, W.J.,
1991. Maize stomatal conductance in the field – its relation-
ship with soil and plant water potentials, mechanical
constraints and ABA concentration in the xylem sap. Plant
Cell Environ. 14, 121–126.
Tardieu, F., Zhang, J., Katerji, N., Bethenod, O., Palmer, S.,
Davies, W.J., 1992. Xylem ABA controls the stomatal
conductance of field grown maize subjected to soil compac-
tion and soil drying. Plant Cell Environ. 15, 193–197.
Tardieu, F., Zhang, J., Gowing, D.J.G., 1993. Stomatal
control by both [ABA] in the xylem sap and leaf water
status: a test of a model for droughted or ABA fed field
grown maize. Plant Cell Environ. 16, 413–420.
Tardieu, F., Lafarge, T., Simonneau, T., 1996. Stomatal
control by fed or endogenous xylem ABA in sunflower:
interpretation of correlations between leaf water potential
and stomatal conductance in anisohydric species. Plant Cell
Environ. 19, 75–84.
Tenhunen, J.D., Hanano, R., Abril, M., Weiler, E.W.,
Hartung, W., 1994. Above- and belowground environmen-
tal influences on leaf conductance of Ceanothus thyrsiflorus
growing in a chaparral environment. Drought response and
the role of abscisic acid. Oecologia 99, 306–314.
Trejo, C.L., Davies, W.J., Ruiz, L.P., 1993. Sensitivity of
stomata to abscisic acid. An effect of the mesophyll. Plant
Physiol. 102, 497–502.
Trejo, C.L., Clephan, A.L., Davies, W.J., 1995. How do stomata
read abscisic acid signals? Plant Physiol. 109, 803–811.
Triboulot, M.B., Fauveau, M.L., Breda, N., Label, P., Dreyer, E.,
1996. Stomatal conductance and xylem-sap abscisic acid
(ABA) in adult oak trees during a gradually imposed drought.
Ann. Sci. For. 53, 207–220.
Wartinger, A., Heilmeier, H., Hartung, W., Schulze, E.-D.,
1990. Daily and seasonal courses of leaf conductance and
abscisic acid in the xylem sap of almond trees (Prunus dulcis
(Miller) D.A.Webb) under desert conditions. New Phytol.
116, 581–587.
Wilkinson, S., Corlett, J.E., Oger, L., Davies, W.J., 1998.
Effects of xylem pH on transpiration from wild-type and
flacca-tomato leaves: a vital role for abscisic acid in
preventing excessive water loss even from well-watered
plants. Plant Physiol. 117, 703–709.
Zhang, J., Davies, W.J., 1987. Increased synthesis of ABA in
partially dehydrated root tips and ABA transport from
roots to leaves. J. Exp. Bot. 38, 2015–2023.
Zhang, J., Davies, W.J., 1990. Changes of concentration of
ABA in the xylem sap as a function of changing soil water
status can account for changes in leaf conductance and
growth. Plant Cell Environ. 13, 277–285.






![Drought responses of two gymnosperm species with contrasting stomatal regulation strategies under elevated [CO2] and temperature](https://static.fdokumen.com/doc/165x107/63365de8cd4bf2402c0b78c2/drought-responses-of-two-gymnosperm-species-with-contrasting-stomatal-regulation.jpg)













