
G F J I H K M L N Q O P D B E A C
80
Bradleya 25/2007 45 Summary: The genus Haageocereus (Cactaceae) is almost endemic to Peru, occupying arid areas of river valleys draining westwards off the Andes. Extensive fieldwork, morphological studies, liter- ature revision and a study of the types led to the delimitation of 9 species and 6 heterotypic sub- species. Nomenclatural innovations include H. acranthus subsp. backebergii N. Calderón subsp. nov., H. versicolor subsp. pseudoversicolor (Rauh & Backeb.) N. Calderón comb. nov. and the recog- nition of H. repens Rauh & Backeb. Descriptions and keys are accompanied by line drawings and plates including SEM photos of the seeds for almost every taxon. Exsiccata prepared during this study constitute approximately 80% of exist- ing Haageocereus herbarium samples from wild origin. Conservation assessments based on the IUCN Red List categories (2001) determined 3 taxa as Critically Endangered (CR), 8 taxa as Endangered (EN) and 5 taxa as Vulnerable (VU) in Peru. Zusammenfassung: Die Gattung Haageocereus (Cactaceae) ist mehrheitlich in Peru endemisch und kommt in den trockenen Teilen der die Anden nach Westen entwässernden Flusstäler vor. Ausgedehnte Feldstudien, morphologische Untersuchungen und Literaturauswertungen sowie die Unterschung der Typmaterialien ver- anlassen uns, 9 Arten und 6 heterotypische Unterarten zu unterscheiden. H. acranthus subsp. backebergii N. Calderón subsp. nov., H. versicolor subsp. pseudoversicolor (Rauh & Backeb.) N. Calderón comb. nov. sowie die Anerkennung von H. repens Rauh & Backeb. sind nomenklatorische Innovationen. Beschreibungen und Bestimmungsschüssel werden für fast alle Taxa von Strichzeichnungen sowie SEM- Samenfotos begleitet. Die während des Projektes präparierten Herbarbelege machen ungefähr 80% aller Haageocereus-Herbarbelege vom natürlichen Fundort aus. Auf der Basis der Rote- Listen-Kategorien der IUCN (2001) werden 3 peruanische Taxa als Critically Endangered (CR), 8 als Endangered (EN) und 5 als Vulnerable (VU) eingestuft. Resumen: El género Haageocereus (Cactaceae) es, en su mayor parte, endémico de Perú, ocupando zonas áridas de la vertiente occidental de los Andes. Extenso trabajo de campo, estudios mor- fológicos, revisión de literatura y estudio de tipos concluyeron en la delimitación de 9 especies y 6 subespecies heterotípicas. Innovaciones de nomenclatura incluyen H. acranthus subsp. bac- kebergii N. Calderón subsp. nov., H. versicolor subsp. pseudoversicolor (Rauh & Backeb.) N. Calderón comb. nov. y el reconocimiento de la especie H. repens Rauh & Backeb. Las descrip- ciones y claves son acompañadas por dibujos científicos y fotografías incluyendo fotos SEM de las semillas para casi todos los taxa. Las exsicca- ta preparadas durante este estudio constituyen aproximadamente el 80% de las muestras de her- barios existentes de Haageocereus de origen sil- vestre. El estado de conservación se estimó de acuerdo a las categorías de la Lista Roja IUCN (2001) determinando 3 taxa como Críticamente En Peligro (CR), 8 taxa como En Peligro (EN) y 5 taxa como Vulnerables (VU) en Perú. Introduction The family Cactaceae, represented mostly by suc- culent plants with areoles and spines, is placed by Cronquist (1981) as the single family of the Order Cactales in the Sub-class Caryophyllidae. Bradleya 25/2007 pages 45 – 124 Taxonomy and conservation of Haageocereus Backeb. (Cactaceae) in Peru Natalia Calderón 1 , Daniela Zappi 2 , Nigel Taylor 2 and Aldo Ceroni 1 1 Universidad Nacional Agraria La Molina, Lima, Peru (email: [email protected]). 2 Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3AB, UK.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of G F J I H K M L N Q O P D B E A C

Bradleya 25/2007 45
Summary: The genus Haageocereus (Cactaceae)is almost endemic to Peru, occupying arid areasof river valleys draining westwards off the Andes.Extensive fieldwork, morphological studies, liter-ature revision and a study of the types led to thedelimitation of 9 species and 6 heterotypic sub-species. Nomenclatural innovations include H.acranthus subsp. backebergii N. Calderón subsp.nov., H. versicolor subsp. pseudoversicolor (Rauh& Backeb.) N. Calderón comb. nov. and the recog-nition of H. repens Rauh & Backeb. Descriptionsand keys are accompanied by line drawings andplates including SEM photos of the seeds foralmost every taxon. Exsiccata prepared duringthis study constitute approximately 80% of exist-ing Haageocereus herbarium samples from wildorigin. Conservation assessments based on theIUCN Red List categories (2001) determined 3taxa as Critically Endangered (CR), 8 taxa asEndangered (EN) and 5 taxa as Vulnerable (VU)in Peru.
Zusammenfassung: Die Gattung Haageocereus(Cactaceae) ist mehrheitlich in Peru endemischund kommt in den trockenen Teilen der dieAnden nach Westen entwässernden Flusstälervor. Ausgedehnte Feldstudien, morphologischeUntersuchungen und Literaturauswertungensowie die Unterschung der Typmaterialien ver-anlassen uns, 9 Arten und 6 heterotypischeUnterarten zu unterscheiden. H. acranthussubsp. backebergii N. Calderón subsp. nov., H.versicolor subsp. pseudoversicolor (Rauh &Backeb.) N. Calderón comb. nov. sowie dieAnerkennung von H. repens Rauh & Backeb. sindnomenklatorische Innovationen. Beschreibungenund Bestimmungsschüssel werden für fast alleTaxa von Strichzeichnungen sowie SEM-
Samenfotos begleitet. Die während des Projektespräparierten Herbarbelege machen ungefähr80% aller Haageocereus-Herbarbelege vomnatürlichen Fundort aus. Auf der Basis der Rote-Listen-Kategorien der IUCN (2001) werden 3peruanische Taxa als Critically Endangered(CR), 8 als Endangered (EN) und 5 als Vulnerable(VU) eingestuft.
Resumen: El género Haageocereus (Cactaceae) es,en su mayor parte, endémico de Perú, ocupandozonas áridas de la vertiente occidental de losAndes. Extenso trabajo de campo, estudios mor-fológicos, revisión de literatura y estudio de tiposconcluyeron en la delimitación de 9 especies y 6subespecies heterotípicas. Innovaciones denomenclatura incluyen H. acranthus subsp. bac-kebergii N. Calderón subsp. nov., H. versicolorsubsp. pseudoversicolor (Rauh & Backeb.) N.Calderón comb. nov. y el reconocimiento de laespecie H. repens Rauh & Backeb. Las descrip-ciones y claves son acompañadas por dibujoscientíficos y fotografías incluyendo fotos SEM delas semillas para casi todos los taxa. Las exsicca-ta preparadas durante este estudio constituyenaproximadamente el 80% de las muestras de her-barios existentes de Haageocereus de origen sil-vestre. El estado de conservación se estimó deacuerdo a las categorías de la Lista Roja IUCN(2001) determinando 3 taxa como CríticamenteEn Peligro (CR), 8 taxa como En Peligro (EN) y 5taxa como Vulnerables (VU) en Perú.
IntroductionThe family Cactaceae, represented mostly by suc-culent plants with areoles and spines, is placedby Cronquist (1981) as the single family of theOrder Cactales in the Sub-class Caryophyllidae.
Bradleya 25/2007pages 45 – 124
Taxonomy and conservation of Haageocereus Backeb.(Cactaceae) in Peru
Natalia Calderón1, Daniela Zappi 2, Nigel Taylor 2 and Aldo Ceroni 1
1Universidad Nacional Agraria La Molina, Lima, Peru (email: [email protected]).2Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3AB, UK.

Bradleya 25/200746
According to the Angiosperm Phylogeny Group(APG, 1998, 2003), the Cactaceae is placed in thecore Caryophyllales, being more related to theunresolved “Portulacaceae” (Judd et al., 2002).
Genetic studies (Wallace, 1995; Wallace et al.,1996, 2002a) as well as seed anatomical, pollenand gross morphological data have led to the def-inition of four subfamilies: Cactoideae,Opuntioideae, Pereskioideae and Maihuen -ioideae. Although recent molecular studies(Edwards et al., 2005) have cast doubts upon therelative importance of these, presenting a newhypothesis of phylogenetic relationships at thebase of the Cactaceae, supporting a basal split inCactaceae – thus, possible paraphyly – betweena clade of eight Pereskia species, centred aroundthe Caribbean basin, and all other cacti.However, additional genes are still going to needto be tested before reclassifying the Cactaceae.
The Cactaceae are a neotropical family withapproximately 1500 species (Hunt, 1999), distrib-uted in a broad variety of environments, fromtropical rainforest to extremely arid deserts.After Mexico and the south-western USA, thesecond most important geographical centre ofdiversity for Cactaceae is within the Andeanchain, with Peru and Bolivia being especiallyrich, where the taxonomic complexities of thefamily are considerable and inadequately under-stood (Taylor & Zappi, 2004).
Monographic treatments of PeruvianCactaceae were published by Britton & Rose(1919–23), Rauh (1958), Backeberg (1958–62)and Ritter (1981). The most recent species check-lists including Peruvian cacti are provided byHunt (1999) and Hunt et al. (2006).
In Peru, the cactus family is found along thePacific coast, mostly on western slopes of theAndean mountains, and also inter-Andean val-leys, with an altitude range between sea leveland 5100 m. With over 250 species and 34 genera(Hunt, 1999), the Cactaceae form an importantpart of the arid and semi-arid zone landscape.Important communities of cacti have alreadybeen identified in the north (Depts. Amazonasand Cajamarca) and in the south (Depts.Arequipa and Cuzco) (Pennington, 2004).
Taxonomically, the majority of Peruvian cac-tus species belong to the subfamily Cactoideae,tribe Trichocereeae that together with the tribesBrowningieae and Cereeae form the “BCT” clade,characterised by a shared deletion in the rpl16intron (Wallace & Gibson, 2002). However, thecircumscription among these three tribes lacksresolution. Considering traditional circumscrip-
tion, Trichocereeae are more closely related toCereeae than Browningieae, where unlike theCereeae with naked flowers, Trichocereeae pre-sent flowers covered by hairs, spines and/or woolborne on the hypanthial areoles (Taylor & Zappi,2004).
According to the phylogenetic system ofBuxbaum (1962), Haageocereus Backeb. (1934)belongs to tribe Trichocereae. Allied genera ofHaageocereus supported by genetic studies arepostulated as Espostoa, Weberbauerocereus andCleistocactus (Arakaki, 2002); they all havecolumnar stems and similar spination charactersbut the flowers are distinctively different, espe-cially in shape and size, corresponding with thedifferent pollination syndromes. In nature,Haageocereus hybridizes with Espostoa, formingthe intergeneric hybrid ×Haagespostoa that dis-plays a range of characters inherited from bothparent genera.
Haageocereus is mostly endemic in Peru, dis-tributed over much of the country, with its north-ern limit in Piura (Peru) and extending south-wards to northernmost Chile. In Peru, the genuscan be found in most of the arid areas of river val-leys draining westwards and in coastal dunes,between 50 and 2800 m altitude, including thePacific coastal desert, the northern dry forest andwestern Andean valleys. These habitats have avery low annual rainfall, especially along thecoast and in the southern Andes (18–100 mm). Inthe central Andes and in the northern dry forestthe precipitation is higher (100–500 mm).
The species of Haageocereus are generallycharacterized by their shrubby habit, branchingtypically from the base, crepuscular nocturnalflowers that may remain open until the nextmorning, a pericarpel covered by areolar scaleswith axillary trichomes, and ovoid to globosefruits, which are generally indehiscent. The phe-nology is highly variable amongst species, flower-ing being ephemeral, unpredictable and not easyto observe.
For conservation purposes, it is a prerequisiteto establish a stable taxonomy and nomenclatu-re, something that Haageocereus species havelacked during recent decades.
In Peru, there are serious threats to membersof the family Cactaceae and especially toHaageocereus, due to the location of many popu-lations close to major cities. Disturbance of thehabitats of cactus populations is caused byhuman expansion, environmental pollution andagricultural development (Ostolaza, 1995a). Inrelation to human disturbance, population stu-

Bradleya 25/2007 47
dies of Haageocereus have been undertaken(Calderón, 2002) showing that the closer thesecacti are to human settlements, the poorer istheir health, particularly in terms of longevityand reproductive effectiveness.
Establishing the status of these species underthe Red List categories of the InternationalUnion for the Conservation of the Nature andNatural Resources (IUCN, 2001) will enable newconservation proposals and action plans for thenatural habitats to be developed for these taxa ashas been achieved for other kinds of habitatsinvolving many non-cacti plant species in Peru.
Materials and MethodsStudy areaThe area of study is mainly located in diversedeserts, valleys and dry forests along the lengthof Peru (Map 1), between 50 m and 2800 m alti-tude, from Haageocereus versicolor in the north(79˚54’W, 5˚7’S) to Haageocereus decumbens(70˚52’W, 18˚0’S) in the south.
Data collectionBibliographic records were provided by thePeruvian Cactus & Succulent Society (SPECS),and especially by the main library of the RoyalBotanic Gardens, Kew. Field studies were carriedout between 2001 and 2005, recording not onlymorphological characters of the species but alsodata regarding the distribution range and envi-ronmental quality of the habitats involved.
Study of herbarium material was initiatedwith the collections from the Herbarium of theBotanic Garden of La Molina University (Herb.B. G. La Molina). Herbarium specimens werealso prepared following the methodology propo-sed by Zappi (1994). These specimens are held atthe Herb. B. G. La Molina and La MolinaUniversity Herbarium (MOL). Visits to theEuropean herbaria of Utrecht (U) and theSucculent Collection of Zürich (ZSS) were under-taken. These European herbaria include 21 holo-types, 6 isotypes and several non-type exsiccataof Haageocereus. Loans requested from ZSS andBerlin (B) collections were also provided for studyat the Kew Herbarium. Additional seed samplesof Haageocereus were provided by GrahamCharles from his large private collection ofknown wild provenance.
It is estimated that this study embraces 80% ofthe current Haageocereus exsiccata from wildprovenance, but the genus still remains poorlyrepresented in most herbaria.
Study methodologyFor morphological studies, fresh and dried cactusspecimens were carefully measured to enable thecomparison of characters and elaboration ofdescriptions. A stereoscopic Leica MZ6 microsco-pe was used to observe small structures like sta-mens and trichomes. Records of rib number, stemdiameter and distance between areoles weretaken from fresh samples, since these structuresshrink when dried, while flowers and fruit cha-racters were recorded, where possible, from sam-ples preserved in spirit. It is important to recordthe colour, texture and odour of reproductivestructures before these are preserved. Areoles,spines and hairs can be measured reliably fromeither fresh or dry material.
For making line drawings, the stereoscopeLeica MZ6 and a professional vernier were espe-cially useful to determine diameters of hairs, spi-nes, and floral indumentation. The figures illust-rate the typical growth habit, detail of spines andareoles, as well as complete flowers and fruit lon-gitudinal sections. Diagnostic taxonomic charac-ters are given especial attention in the figures.
The study of Haageocereus seeds was carriedout with the aid of the Scanning ElectronMicroscope (HITACHI S-2400 SEM) at thePalynology Unit of the Jodrell Laboratory, Kew.The treatment of seeds prior to scanning startedwith ultrasonic cleaning using distilled waterwith a few drops of industrial detergent (dil. to1%). Seeds were dried and mounted on stubswith double-sided sticky tape. After a period of1–2 days in a desiccation capsule, mounted seedswere coated with platinum using the EMITECHK550X coater for 2×4 minutes prior to scanning.In the SEM, seeds were scanned and photogra-phed at ×60 and ×600 to determine the variabili-ty of seed morphology. Side views were selectedin order to show as many characters as possibleand details of testa surface were observed byclose-ups of the peripheral region of the seed, asproposed by Barthlott & Hunt (2000).
The Geographic Information System (GIS)Unit of the Kew Herbarium provided base maps,to which locality data collected in this study havebeen added, in order to illustrate Haageocereusdistribution in Peru, as well as in relation to cur-rent National Protected Areas. ArcView compu-ter software was modified at Kew’s GIS Unit toassess the IUCN (2001) Red List criteria, mainlyfor the application of criterion B of geographicrange, based on the “extent of occurrence” and“area of occupancy” and thus automatically gene-rating the IUCN rating and categories of threat

Bradleya 25/200748
Map 1. Study Area. Localities of Haageocereus visited in Peru = �.
N
100 0 100 200 Kilometers

Bradleya 25/2007 49
for each species (Willis et al., 2003). The use ofcriterion A (IUCN, 2001), based on the reductionof population, is alternatively applied when thereis evidence that population decline constitutesthe worst threat for the species and this has beenobserved in the past, present and is likely tooccur in the future.
History of the genusThe earliest publications possibly referring toHaageocereus, according to Ritter (1981) areCereus limensis Salm-Dyck (1845) that lacks apreserved type, any illustration and a meaning-ful description, and Cactus multangularisWilldenow (1809) that is even more uncertainand, according to Leuenberger (2004), should beavoided.
The first descriptions certainly referring toHaageocereus were published by Vaupel (1913),who described two species as Cereus decumbensand Cereus acranthus based on the collections ofA. Weberbauer held at the Berlin-DahlemHerbarium.
In 1920, the genus Binghamia Britton & Rosewas created to include Vaupel´s speciesCephalocereus melanostele and Cereus acran-thus. Nevertheless, to judge by the descriptionand photos, Britton & Rose had misidentified anew species (currently Haageocereus pseudome-lanostele) as C. melanostele (now Espostoamelanostele) and also did not notice that thename Binghamia had previously been assignedto a genus of algae (Agardh, 1894).
Werdermann & Backeberg (1931) describedas Cereus pseudomelanostele the species thatBritton & Rose had mistakenly referred toCephalocereus melanostele. Backeberg (1934a)created the genus Haageocereus to includeCereus pseudomelanostele, as well as C. acran-thus Vaupel and C. decumbens Vaupel(Backeberg, 1934b, 1937) and in later publica-tions (Backeberg, 1957, 1960) the genusHaageocereus was expanded by the inclusion ofnarrowly described taxa making a total of 49species names.
Akers (1947), an American cactus grower, cre-ated the genus Peruvocereus to include a group ofspecies he found in the central valleys of Peruand which he believed to be different fromHaageocereus pseudomelanostele (the type ofHaageocereus). Between 1947 and 1948, Akerspublished 10 species of Peruvocereus withdetailed descriptions and photographs, but didnot preserve their types. Some of these speciesnames are currently synonyms of Backeberg´s
H. pseudomelanostele, or have been recognized ashybrids (See Appendix 1: List of names ofHaageocereus (and Peruvocereus) to be possiblyreferred to ×Haagespostoa).
Rauh (1958) published Beitrag zur Kenntnisder peruanischen Kakteenvegetation, a remark-able work that included the 49 Haageocereustaxa that Backeberg & Rauh described with briefdiagnoses in 1957 as new species based on Rauh’scollections. The importance of Rauh´s monographreflects the collections he made in Peru in 1954and 1956, including photographs and informa-tion about the environment and vegetation asso-ciated with these cacti. Rauh’s types were kept atHeidelberg Botanic Garden and Herbarium(HEID), but unfortunately most have subse-quently been lost and the few exsiccata stillavailable have been transferred to the ZürichSucculent Collection Herbarium (ZSS).
Backeberg (1960) published a key toHaageocereus species and varieties that hearranged in 6 series (Reihe): Acranthi,Versicolores, Asetosi, Setosi, Decumbentes andRepentes. Backeberg´s classifications, as well asspecies concepts, were based on rather inconsis-tent vegetative characters. His key illustratesthe difficulty of defining clear morphological dif-ferences between the taxa he described.Nevertheless, Die Cactaceae (Backeberg,1958–1962) remains a much-consulted mono-graphic treatment for Peruvian cacti.
Ritter (1981) accepted 19 species ofHaageocereus, describing 7 new species thatincluded H. tenuis and H. lanugispinus.Unfortunately, he did not attempt to preparekeys to identify the species.
In the latest CITES Checklist (Hunt, 1999),Haageocereus is credited with 13 accepted speciesplus 8 provisionally accepted species and 3 accep-ted heterotypic subspecies. In the present study,this number has been further reduced to reflectthe lack of stable characters.
In relation to their conservation, severalimportant Haageocereus populations have suffe-red a dramatic reduction in numbers of individu-als and/or have disappeared in the last few deca-des (Ostolaza, 1995a). For instance, H. pseudo-melanostele (Werderm. & Backeb.) Backeb., canno longer be found at its type locality atCajamarquilla in the Rímac valley (Vaupel, 1913)because of housing developments. In the sameway, several populations of Haageocereus surro-unding Lima have declined in the last five years,especially in the Lurín and Rímac valleys. Inother parts of the country, these species remain

vulnerable and are currently under similarthreats.
MorphologyThe present morphological survey was based onthe study of Haageocereus sp. for the presentmonograph.
Currently, intergeneric relationships amongmembers of Trichocereeae and even the cladeBCT are unclear. For this reason, it is difficult toestablish a basal or sister taxon for Haageocereus(especially among its proposed allied generaEspostoa, Weberbauerocereus and Cleistocactus)in order to elucidate possible derived morpholog-ical characters in this genus. However, genesequence studies developed by Wallace (1997)found evidence supporting a monophyletic genusHarrisia (tribe Trichocereeae), and also that thesister group of Harrisia was the Bolivian endem-ic Samaipaticereus (S. corroanus). From this,Samaipaticereus could be hypothesized as abasal taxon or potential “ancestral morphotype”for Haageocereus and in this sense the characterspresent in Samaipaticereus will be hypothesizedas plesiomorphic and the derived characters pre-sent in Haageocereus will be hypothesized as apo-morphic.
There are several shared characters (poten-tial plesiomorphies) between Haageocereus andSamaipaticereus best represented by their flow-ers that are very alike, but also there are otherinteresting and contrasting features in the rest oftheir morphology.
S. corroanus are tree-like plants with erectbranches and a low number of ribs (4–6); trian-gular areoles; spines 5, 2.0–3.0 mm long, centralspine 5.0–10.0 mm long; flowers 4.5–5.0 cm long,narrowly funnelform, tube slightly curved andcovered by hairs and a few brown bristles emerg-ing from the axils of the bract-scales; outer peri-anth segments whitish-green, inner perianthsegments white; stamens numerous; stigma lobes7–10; nectar-chamber 1.0 cm long; fruit dehis-
cent, funicular pulp salmon-red; seeds broadlyovate, 1.3 × 1.0 mm, black, glossy, cuticle weaklystriated, hilum large and basal. According to thisbrief description and in comparison to the char-acters presented in Haageocereus, it is possible tohypothesize the possible plesiomorphies and apo-morphies of the latter genus (Table 1).
As already evidenced in pollination syn-dromes, shifts to a plesiomorphic state are con-sidered to represent a reversion, or a secondaryderived syndrome (Wallace, 2002). In the samemanner, trends in the fruit morphology ofHaageocereus, represented by the dehiscentfruits present in H. pseudomelanostele subsp.turbidus and the pinkish funicular pulp of H.tenuis, could also be hypothesized as a reversionfor the genus.
Habit and growth patterns Haageocereus presents terete stems, branchingmainly from the base in a prostrate, semi-pros-trate and erect manner (Figure 1). The plants areshrubby, the erect taxa up to 1.4–1.7 m, excep-tionally reaching 2.4 m. Lacking a lignified vas-cular cylinder, the tallest stems usually fall to theground in old age, and sometimes new branchesmay sprout from the fallen branches.
In some cases, erect-growing taxa may devel-op initially decumbent new branches that even-tually turn upwards and develop into erectstems. Haageocereus plants usually present 4–20branches per individual, but sometimes developinto very profusely-branched individuals, with upto 95 branches in H. acranthus subsp. acranthus.
H. decumbens, H. tenuis and H. lanugispinusall have prostrate branches with ascendingapices 5.0–10.0 cm above the ground. This char-acter is hypothesized as a secondary apomorphyfor the genus. Semi-prostrate to erect species arerepresented by H. repens, H. platinospinus andH. chilensis growing at first decumbent and latererect at ≥ 20.0 cm above the ground. In the caseof H. platinospinus that is a short bushy plant,
Bradleya 25/200750
Table 1. Hypothesized plesiomorphic and apomorphic characters of Haageocereus, based on outgroup compari-son with Samaipaticereus (Trichocereeae)
Plesiomorphies
Erect branchesRadial spine number 7–20Central spine presentFlowering areoles not clearly differentiatedSeed microrelief weakly striatedFlower tube straight to slightly curved
Apomorphies
Prostrate and semi-prostrate branchesRadial spine number (21–)56 Central spine absentFlowering areoles well-differentiatedSeed microrelief strongly striatedFlower tube well curved

Bradleya 25/2007 51
A
B
C
D
E
F
G
H
Figure 1. Characters for species recognition. Types of growth habit: A, erect (H. acranthus subsp. acranthus, N.Calderón 211); B, semi-decumbent (H. repens, N. Calderón 358); C, decumbent (H. tenuis, N. Calderón 419). Typesof flower tube curvature: D, straight (H. platinospinus, N. Calderón 444); E, slightly curved (H. platinospinus, N.Calderón 450); F, markedly curved (H. tenuis, N. Calderón 364). Areoles and spination: G, Areole with centralspine, radial spines and bristle-spines (H. pseudomelanostele subsp. pseudomelanostele, N. Calderón 474); H,Areole with central and radial spines covered by trichomes (H. tenuis, N. Calderón 364).

Bradleya 25/200752
the stems may be totally erect, somewhat curved,or semi-prostrate.
H. pseudomelanostele, H. acranthus and H.versicolor present upright branches. The first twospecies form part of the so-called “vegetation ofcolumnar cacti formation” of the Andean valleys,and the latter species is part of the “seasonallydry forest” in northern Peru.
Branch diameter varies from 1.2–1.4 cm (H. lanugispinus and H. tenuis) up to 7.5–8.0 cm(H. pseudomelanostele subsp. aureispinus and H. acranthus subsp. zonatus). Haageocereusspecies prove to have an extraordinary resistanceto dry conditions, as is reflected by their thick,swollen stems characteristic of very arid environments.
Rib number varies from 10 ribs (H. acranthussubsp. backebergii) to 24 ribs (H. pseudomelanos-tele subsp. pseudomelanostele and H. pseudome-lanostele subsp. aureispinus). Rib number is, inmost cases, inversely proportional to the ribwidth and it is an important key character.
In the species with lower numbers of ribs, theobservable stem colour is that of the epidermiswhen not obscured by the spines, being generallygreen for all the species, but brownish-green forthe southern populations of H. platinospinus andgreyish green for H. chilensis. The epidermis hasa waxy cuticle, which can be easily observed inthe species with few ribs and spines, like H. plati-nospinus and H. acranthus.
Areoles and spinationAreole shape is typically oval at maturity, but cir-cular when first developing at the stem apex. Theareoles vary in size from 1.5–1.6 mm diam. (H. lanugispinus and H. tenuis) up to 10.0–11.0mm diam. (H. acranthus). Areoles are spacedalong the stem ribs between 1.4–16.5 mm apart.They change in colour depending on age, beinggenerally yellow at first, turning brownish whenolder, but old areoles may darken because of pol-lution, especially in cacti growing close to citiesand busy roads.
Areoles are conspicuous in Haageocereus inspecies with lower numbers of ribs, like H. plati-nospinus, H. acranthus, H. chilensis andH. decumbens. These species have compact cush-ions of brownish yellow trichomes, sometimessubtended and marked by transverse rib-folds,this last feature being considered as a plesiomor-phic character because it is associated with theformation of “podaria” under the areoles (Zappi,1994). For other Haageocereus species, like H.tenuis or H. lanugispinus, areoles are not easy to
observe, presenting irregular white tufts of tri-chomes. Areoles may also be densely covered bynumerous spines, as in H. repens, H. pseudome-lanostele and H. versicolor and therefore, theirshape and hairiness may be difficult to observe.
Spines are comprised typically of numerousradials with 1–3 longer spines, called centralspines that are absent (a hypothesized apomor-phy) in H. lanugispinus. Central spines vary innumber from 1–2(–3), and in size from 0.3–15.0 cm long and 0.3–1.9 mm diameter at theirbases, H. chilensis, H. platinospinus and H.pseudomelanostele subsp. turbidus being thetaxa with the longest spines within the genus.Radial spines vary in number from 7–56, and insize from 2.0–15.4(–40.0) mm long and 0.2–0.7 mm diameter at base. There is variation inspine size within individuals of similar age, as inH. platinospinus, which can have central spinesin some individuals four-times the size of others.
As is the case with areoles, spines are yellowat first, or sometimes reddish in H. versicolor,becoming brownish or greyish with age. Thepresence of white and yellow bristle-spines canbe observed especially in H. pseudomelanostelesubsp. pseudomelanostele and also in H. repens.
The spines bear white trichomes only in H. lanugispinus and H. tenuis, this characterbeing rather rare within subfamily Cactoideaeand here hypothesized as an autoapomorphy,probably of high adaptative value, to enhance thedeposition of water droplets from the seasonalsea-fog in their coastal habitats.
Flower-bearing areoles In Haageocereus, flower-bearing areoles arerecognisable by having a few short white tri-chomes developing as the flower bud forms andfor most of the species these trichomes do notpersist for long. More conspicuous flowering are-oles can be seen in H. versicolor subsp. versicolor,H. versicolor subsp. pseudoversicolor and H. acranthus subsp. zonatus, which present aring-like “pseudocephalium” (a hypothesized apo-morphy), formed by many long white trichomesthat remain on the areoles after flowers andfruits have completed their development. As thischaracter is present only in mature individuals,care has to be taken when identifying youngplants of these species; on the other hand, oldindividuals show several of the characteristicring-like pseudocephalia along their erectbranches.

Bradleya 25/2007 53
Flowers and fruitsHaageocereus species present few, ephemeralflowers, developing terminally on the stem. Theyare crepuscular-nocturnal, but may remain openuntil the next morning. Flowers have a fun-nelform shape, generally between 6.0–8.5 cmlong, but they can be smaller, such as in H. pseudomelanostele subsp. pseudomelanosteleand H. lanugispinus (4.0–5.0 cm) or larger insome individuals of H. tenuis, H. acranthussubsp. acranthus, H. acranthus subsp. zonatusand H. pseudomelanostele subsp. aureispinus(9.7–11.5 cm), with a perianth-limb regularlysymmetrical that presents numerous perianthsegments or tepals that vary in colour, beinggreen to reddish outside and white to pinkish andreddish inside. The elongate receptacle tube maybe somewhat flattened or curved, externallybearing hair-spines and trichomes that emergefrom the bract-scale axils. Flower curvaturevaries from straight and slightly curved (H. acranthus, H. decumbens, H. pseudomelanos-tele, H. platinospinus and H. versicolor) to strongly curved (H. repens and H. tenuis), ahypothesized apomorphy (Figure 1). The peri-carpel encloses the ovary locule and bears fewbract-scales and trichomes externally; the nec-tar-chamber is tubular, somewhat swollen, pro-ducing sweet translucent nectar; and the flowerexhales a “sweet-pungent” smell. Stamens arenumerous (200–400), inserted in a single series;the stigma-lobes may be exerted or included.
Fruits are spherical to ovoid, between 2.0–6.5 cm long and 1.3–5.6 cm diameter, bright redwhen mature and easily seen from a long dis-tance. The flower remnants are persistent on topof the fruit. The pericarp is thin and mostly inde-hiscent, but in the case of H. pseudomelanostelesubsp. turbidus, fruits can split open when ripe.The funicular pulp is white and solid for most ofthe species, being translucent pink and mucilagi-nous in H. tenuis.
Seed morphology This study surveyed seeds of almost allHaageocereus taxa and has identified key charac-ters from the micro-morphology of the testa. Theterminology employed here is based on Barthlott& Hunt (2000), where 26 seed-characters are pro-posed to describe the seed diversity in the sub-family Cactoideae. These authors used generalfeatures of the morphology, testa appearance,individual testa-cells, anticlinal cell boundaries,periclinal wall sculpture, hilum-micropylarregion and other appendages of seeds.
Haageocereus seeds are mainly medium-sized(1.2–1.87 mm) that is the average size for sub-family Cactoideae, but can also be small(1.09–1.14 mm); typically mussel-shaped(Cereus-type), being broadly ovate (Plate 1.1) toovate in outline (Plate 4.6), calculated on thebasis of seed length: breadth ratio (1.10–1.99). Ingeneral, the testa has a glossy appearance, but inH. tenuis it is semi-matt.
Cuticular striations, as a result of cuticle fold-ing, are very strong in H. repens (Plate 5.3),H. tenuis (Plate 5.6) and H. versicolor (Plate 6.1),this character being hypothesized as an apomor-phy for these species. The blackish testa presentsa multi-cellular sculpture with interstitialcraters and low-domed convexities that are typi-cal for all Haageocereus (Plates 1–6). The pres-ence of par-convexities (Plate 1.5) was observedonly in H. acranthus subsp. zonatus towards theperipheral border of the seed. It should be notedthat par-convex structures are mostly associatedwith Notocacteae subtribe Notocactinae and theTrichocereeae subtribe Gymnocalyciinae.
The hilum-micropylar region (HMR) isoblique and large for all Haageocereus, but doesnot present any diversity of form allowing differ-entiation amongst species.
Reproductive biologyPollinationThe family Cactaceae is entirely zoophilous(Barthlott & Hunt, 1993) and more than one pol-linator can probably be found in species ofHaageocereus judging from flower morphology,where traits are found associated with bat, hum-mingbird and moth floral syndromes, althoughbats and hummingbirds have not been directlyobserved visiting Haageocereus flowers.
According to the pollination syndromes pro-posed by Faegri & Van der Pijl (1979), someassumptions can be made about the possible pol-linators of Haageocereus. The typical funnel-formwhite flowers of H. acranthus subsp. backebergii,mostly nocturnal with a sweet-pungent smell,are a good example of chiropterophily. On theother hand, pinkish to reddish flowers that openin the afternoon until next morning, with slightscent and also producing abundant nectar, can berecognized as hummingbird flowers that mightalso secondarily attract bats. Such flowers areseen in H. pseudomelanostele subsp. carmini-florus and H. acranthus subsp. acranthus.
Studies of the allied genus Weberbauerocereusin southern Peru (Sahley, 1996), confirm thepresence of bats and hummingbirds interacting

Bradleya 25/200754
Plate 1. SEMs of seeds. 1.1 H. acranthus subsp. acranthus. N. Calderón 211, side view. 1.2 H. acranthus subsp.backebergii. N. Calderón 111, side view. 1.3 H. acranthus subsp. acranthus. N. Calderón 211, detail of testa surface,microrelief without striations. 1.4 H. acranthus subsp. zonatus, N. Calderón 379, side view. 1.5 Ibid., detail ofperipheral region with par-convexities. 1.6 Ibid., detail of testa surface, microrelief weakly-striated.
1 4
2 5
3 6

Bradleya 25/2007 55
Plate 2. SEMs of seeds. 2.1 H. chilensis. F. Ritter 601, side view. 2.2 Ibid., detail of hilum-micropylar region(HMR). 2.3 Ibid., detail of testa surface, microrelief without striations. 2.4 H. decumbens. N. Calderón 414, sideview. 2.5 Ibid. 2.6 Ibid., detail of hilum-micropylar region (HMR).
1 4
2 5
3 6

Bradleya 25/200756
Plate 3. SEMs of seeds. 3.1 H. platinospinus. N. Calderón 404, side view. 3.2 Ibid., detail of hilum-micropylarregion (HMR). 3.3 Ibid., detail of testa surface, microrelief without striations. 3.4 H. pseudomelanostele subsp.pseudomelanostele. N. Calderón 361, side view. 3.5 Ibid., detail of hilum-micropylar region (HMR). 3.6 Ibid., detailof testa surface, microrelief without striations.
1 4
2 5
3 6

Bradleya 25/2007 57
Plate 4. SEMs of seeds. 4.1 H. pseudomelanostele subsp. carminiflorus. N. Calderón 298a, side view. 4.2 Ibid.,detail of hilum-micropylar region (HMR). 4.3 Ibid., detail of testa surface, microrelief without striations. 4.4 H.pseudomelanostele subsp. aureispinus. N. Calderón 372, side view. 4.5 Ibid., detail of testa surface. 4.6 H.pseudomelanostele subsp. turbidus. N. Calderón 276, side view.
1 4
2 5
3 6

Bradleya 25/200758
Plate 5. SEMs of seeds. 5.1 H. repens. G. Charles 254.01, side view. 5.2 Ibid., detail of hilum-micropylar region(HMR). 5.3 Ibid., detail of testa surface, microrelief with strong cuticular striations. 5.4 H. tenuis. N. Calderón371, side view. 5.5 Ibid., detail of hilum-micropylar region (HMR). 5.6 Ibid., detail of testa surface, microreliefwith strong cuticular striations.
1 4
2 5
3 6

Bradleya 25/2007 59
Plate 6. SEMs of seeds. 6.1 H. versicolor subsp. versicolor. N. Calderón 355, side view. 6.2 Ibid., detail of hilum-micropylar region (HMR). 6.3 Ibid., detail of testa surface, microrelief weakly striated. 6.4 H. versicolor subsp.pseudoversicolor. G. Charles 256.02, side view. 6.5 Ibid., detail of hilum-micropylar region (HMR). 6.6 Ibid., detailof testa surface, microrelief without striations.
1 4
2 5
3 6

Bradleya 25/200760
with the species W. weberbaueri. Because bat andhummingbird floral syndromes share severalcharacteristics, evolutionary transitions betweenthese syndromes within a plant lineage mayoccur relatively frequently (Helverson, 1993).
Attracted by the abundant pollen ofHaageocereus flowers, small coleoptera anddiptera have been found within the perianth,among stamens, but not inside the nectar-cham-ber that is closed by the filaments and thus inac-cessible for these visitors, who need especiallylong tongues to obtain the nectar.
Moth pollination could also be considered, dueto the nocturnal and relatively long tubularreceptacle of Haageocereus flowers and also fortheir sweet smell. In relation to this, there is onephotograph from Rauh (Backeberg, 1960) show-ing a moth visiting a Haageocereus flower.
Another remarkable feature of Haageocereusflowers is the marked curvature of the flowertube in some species, namely H. tenuis and H. repens. These plants have prostrate and semi-prostrate habit and it is probable that the angleof presentation of the flower may be related tothe right pollinator.
In relation to flower symmetry, Haageocereusflowers typically have a radially symmetricalperianth, discounting that the tubular receptaclemay be slightly to markedly curved and in a fewflowers of H. pseudomelanostele subsp. carmini-florus, a tendency towards a zygomorphic peri-anth has been observed.
The flowers of Espostoa melanostele, whenopen, exhibit a radial perianth resemblingHaageocereus. E. melanostele also presents noc-tural flowers, but developing from a lateralcephalium in contrast to Haageocereus. In thiscase, nocturnal pollinators would have a role intransferring pollen between these taxa and pro-ducing intergeneric hybrids. For instance, H. pseudomelanostele subsp. carminiflorus andH. pseudomelanostele subsp. aureispinus haveproduced intergeneric hybrids with Espostoamelanostele. These hybrids are recognized underthe genus ×Haagespostoa (See Appendix 1: Listof names of Haageocereus (and Peruvocereus)possibly referable to ×Haagespostoa) and suchplants seem to be stronger than their parents. Inthe valleys of Santa Eulalia and Pativilca, popu-lations of hybrids are very well represented andsometimes it is difficult to find the parentHaageocereus species.
There is no evidence of intraspecifichybridization in the few cases of sympatryamongst Haageocereus species.
DispersalFruits are mostly indehiscent, with one exceptionin H. pseudomelanostele subsp. turbidus thatopens by a lateral slit. The thin pericarp is red-dish, a character that makes the fruits easy tosee at a distance. Haageocereus fruits are moder-ately to very sweet and are also a source of liquid,which makes them very attractive to birds.
When fruits are damaged and the funicularpulp is exposed, the sweet pulp may attract antsthat may play a role as a disperser together withbirds (ornitho-myrmecochorus strategy), as itoccurs in other genera like Cereus (Barthlott &Hunt, 1993).
Haageocereus seeds present a striated cuticle,and testa cells with low-domed convexitiestowards the peripheral border of the seeds and,in one case, par-convexities. All these characterscould be associated with seed transportation byants, as has been suggested for Pilosocereusaureispinus (Zappi, 1994).
In a few cases, seed dispersal by wind mayaffect seeds that remain attached to the dryfruit’s pericarp.
Taxonomic treatmentDescriptions and keys include measurementsconnected by a multiplication sign (×) referring tothe length (or height) followed by the width (ordiameter) and, in the case of seeds, followed bythe thickness.
Haageocereus Backeb., Blätt. Kakt.-forsch.1934(6): unpaged (1934a); Rauh, Beitr. Kenntn.Peruan. Kakteenveg.: 368 (1958); Backeb., DieCact. 2: 1159 (1960); Krainz, Die Kakteen 16 (4):CVa, unpaged (1973). Type species: Haageocereuspseudomelanostele (Werderm. & Backeb.)Backeb. (1934a).Binghamia Britton & Rose, Cact. 2: 167 (1920),
pro parte, non Binghamia J. Agardh,Analecta Algologica 2: 63 (1894); ibid. 5: 158(1899).
Peruvocereus Akers, Cact. Succ. J. (US) 19: 67(1947).
Etymology of Haageocereus Backeb. (1934): FromHaage, the surname of the famous German fam-ily of cactus nurserymen and Cereus, from theLatin meaning waxen, in relation to its torch orcandle-like appearance.
Shrubby, branches terete, prostrate to erect,branching mainly at base; vascular cylinderweakly woody; tissues mostly mucilaginous; epi-

Bradleya 25/2007 61
dermis green, smooth or with a translucent waxycover; ribs 10–24, straight. Areoles with felt,spination variable. Spines mostly straight,opaque, rarely bearing trichomes. Flowering are-oles, slightly to strongly differentiated by persis-tent white trichomes, sometimes seen as a ring-like pseudocephalium. Flowers crepuscular-noc-turnal, appearing close to the apex of the stem,4.0–11.5 × 2.5–8.0 cm, perianth-limb regularlysymmetric; tube straight to markedly curved,green or reddish green, striated, covered bybract-scales and hair-spines emerging from theiraxils; outer perianth segments thick, greenish tored; inner segments delicate, white, pinkish orred; nectar-chamber tubular, slightly swollen,protected by the innermost stamens; stamens c. 200–400, anthers 1.6–6.0 × 0.4–2.0 mm, ± ver-rucose; style 34.7–78.0 × 0.8–2.5 mm, stigma-lobes 9–13, 2.3–8.0 mm, exserted or included inrelation to the anthers; ovary locule 3.6–12.0 ×3.5–11.0 mm, circular to elliptic in longitudinalsection. Fruit 2.0–6.5 × 1.3–5.6 cm, spherical toovoid, indehiscent or rarely dehiscent by a later-al slit, floral remnants persistent, blackening,erect or pendent, pericarp pinkish or red, coveredby few small bract-scales with axillary trichomes;funicular pulp solid to mucilaginous, mostlywhite or translucent pink. Seeds broadly ovate toovate in outline, small to medium-sized,1.09–1.87 × 0.79–1.36 × 0.62–1.04 mm, glossy tosemi-matt, blackish; border expanded aroundhilum; cells gradually smaller towards hilum andenlarged at the periphery, isodiametric, anticli-nal boundaries channelled, straight; cell junc-tions cratered forming ‘interstices’; relief convex,convexities low-domed and par-convex; microre-lief not striated to strongly striated; hilum large,0.34–0.86 mm, oblique, impressed, micropyleincluded, but separated by a sclerified band,hilum-micropylar region (HMR) oval.
Distribution: Along the western side of Peru andnorthern Chile, including arid coastal areas from50 m alt. to western Andean valleys at 3000 malt. (Pacific drainage). Northernmost limit inMorropón, Peru (79˚54’W, 5˚7’S) for the species H.versicolor; and the southernmost limit inCamiña, Chile (69˚25’W, 19˚18’S) for the speciesH. chilensis.
1. Haageocereus acranthus (Vaupel) Backeb.,Backeb. & F. M. Knuth, Kaktus-ABC: 207 (1936);Rauh, Beitr. Kenntn. Peruan. Kakteenveg.: 374(1958); Backeb., Die Cact. 2: 1176 (1960).
Binghamia acrantha (Vaupel) Britton & Rose,Cact. 2: 168 (1920). Cereus acranthus Vaupel inEngler Bot. Jahrb. Syst. 50 (111): 14 (1913). Type:Peru, Lima, Rímac valley, Santa Clara, Lima–LaOroya road, Loma formation, 400–600 m, 26October 1902, Weberbauer 1679 (B, photo!). H. olowinskianus Backeb., Blätt. Kakt.-forsch.
1937(5): unpaged (1937). H. acranthus subsp.olowinskianus (Backeb.) Ostolaza, Cact. Succ.J. (US) 67: 9 (1995a); Cact. Con. Init. 6: 8(1998a). Type locality: Peru, Lima (believednot to have been preserved). Lectotype (desig-nated here): Backeb., Blätt. Kakt.-forsch.1937(5): unpaged, photo (1937). Synon. nov.
H. olowinskianus var. repandus Rauh & Backeb.in Backeb. Descr. Cact. Nov. [1]: 24 (1956,publ. 1957). Type: Peru, Lima, desert areas,Pachacámac, 100 m, 1956, Rauh K177 (HEID,believed not to have been preserved). Neotype(designated here): Backeb., Die Cact. 2: 1188,fig. 1141 (1960). Synon. nov.
H. olowinskianus var. repandus subvar. erythran-thus Rauh & Backeb. in Backeb. Descr. Cact.Nov. [1]: 24 (1956, publ. 1957). Type: believednot to have been preserved. Neotype (desig-nated here): Backeb., Die Cact. 2: 1213, fig.1171, bottom right (1960). Synon. nov.
H. olowinskianus var. subintertextus Rauh &Backeb. in Backeb. Descr. Cact. Nov. [1]: 24(1956, publ. 1957). Type: Peru, Lima,Pachacámac, 1956, Rauh K177b (HEID,believed not to have been preserved). Neotype(designated here): Backeb., Die Cact. 2: 1189,fig. 1143 (1960). Synon. nov.
H. lachayensis Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 22 (1956, publ. 1957). Type:Peru, Lima, Chancay, Lomas de Lachay, 1954,Rauh K5 (HEID, believed not to have beenpreserved). Neotype (designated here):Backeb., Die Cact. 2: 1177, fig. 1130 (1960).
[H. limensis (Salm-Dyck) sensu Ritter, Kakt.Südam. 4: 1396 (1981), non Cereus limensisSalm-Dyck, Allg. Gartenz 13(45): 353 (1845b).Type: believed not to have been preserved.]
H. clavispinus Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 21 (1956, publ. 1957). H. acran-thus subsp. olowinskianus forma clavispinus(Rauh & Backeb.) Ostolaza, Cact. Succ. J.(US) 67: 10 (1995). Type: Peru, east of Lima,Lomas de Atocongo, 200 m, 1956, Rauh K44(ZSS iso. T23411!).
H. vulpes Ritter, Kakt. Südam. 4: 1423 (1981).Type: Peru, Lima, Chancay, Ritter FR1059 (Uholo!, ZZS seeds!). Synon. nov.
?H. olowinskianus var. rubriflorior Rauh &

Bradleya 25/200762
Backeb. in Backeb. Descr. Cact. Nov. [1]: 24(1956, publ. 1957). H. olowinskianus var.subintertextus subvar. rubriflorior Rauh &Backeb. in Rauh Beitr. Kenntn. Peruan.Kakteenveg.: 387 (1958), nom inval. (McNeillet al., 2006), based on H. olowinskianus var.rubriflorior. H. acranthus var. olowinskianusforma rubriflorior (Rauh & Backeb.)Ostolaza, Brit. Cact. Succ. J. 14(4): 167(1996). Type: Peru, Lima, Pachacámac, 1956,Rauh K177a (HEID, believed not to have beenpreserved).
Habit erect, branching at base, up to 1.55 mtall; branches 4.0–8.5 cm diam., epidermis green;ribs 10–15, 11.6–18.6 × 6.2–13.0 mm. Areoles6.0–11.2 × 5.0–8.3 mm, 4.8–16.5 mm apart, ovalto circular, felt yellow and grey. Spines, opaque,at first yellow and brown, later grey; centralspines 1–2(–3), 7.4–55.0 × 0.8–1.9 mm at base;radial spines (21–)25–50, 3.0–15.4 × 0.2–0.7 mmat base. Flowering areoles not differentiated tomarkedly woolly. Flowers 7.0–10.0 × 3.5–7.5 cm;pericarpel 9.0–16.5 × 12.0–19.4 mm; tube 4.0–6.5× 0.9–2.0 cm at base, widening towards apex to
2.25–3.0 cm diam., slightly curved, green, bear-ing hair-spines emerging from the bract-scaleaxils; outer perianth-segments 10–14, 18.0–29.0× 4.0–8.6 mm, greenish; inner perianth-segments10–24, 21.7–27.0 × 5.7–9.0 mm, white, pinkish orreddish; nectar-chamber 10.0–28.0 × 4.2–10.0mm, tubular; anthers 2.8–6.0 × 0.6–1.3 mm; style55.0–78.0 × 1.0–2.5 mm; stigma-lobes 10, 3.8–5.0mm; ovary locule 4.0–12.0 × 4.6–11.0 mm, cylin-dric to elliptic in longitudinal section. Fruit2.0–5.3 × 2.6–5.5 cm, spherical to ovoid. Seedsbroadly ovate, medium-size, 1.36–1.62 ×1.06–1.28 × 0.73–0.93 mm, glossy; relief (SEM)convex and sometimes par-convex towards bor-der; microrelief ± striated; hilum large, 0.59–075mm, oblique, forming an angle of 26°–44° withlong axis of seed.
Habitat and Distribution: Desert areas and rockyhillsides of valleys of Lima (100–2800 m) andarid valleys in Ica (863 m), (Map 2).
Conservation status: Vulnerable. VU[A4c]. Aninferred population size reduction of ≥ 30% overten years, including both past and future time
Key to the identification of Haageocereus species1. Branches prostrate, with ascending apices 5.0–10.0 cm above the ground (coastal deserts, 50–620 m) ........2
Branches upright, if semi-prostrate with part of branches erect ≥ 20.0 cm above the ground (coastal deserts,seasonally dry forest and west Andean valleys, 50–3000 m) ........................................................................4
2. Spines bearing trichomes, sometimes only visible with help of hand lens ....................................................3Spines without trichomes......................................................................................................3. H. decumbens
3. Trichomes on the spines visible with lens; central spine 1–2; flower (6.5–) 8.0–11.5cm long........8. H. tenuisTrichomes on the spines easily visible by the naked eye; central spine 0; flower 5.0 cm long. ..................................................................................................................................................................4. H. lanugispinus
4. Radial spines 7–20 (southern Peru and northern Chile, 1000–3000 m) ...................................................... 5Radial spines (21–) 25–56 (central and northern Peru, 50–2800 m) ............................................................6
5. Radial spines 7–10(–11), (10.0–)15.0–40.0 mm (southern Peru and northern Chile, 2000–3000 m) ..............................................................................................................................................................2. H. chilensis
Radial spines (12–) 14–20, 4.6–10.0 mm (southern Peru, 1000–2600 m) ..........................5. H. platinospinus6. Ribs 10–15; epidermis easily observed between spines ..........................................................1. H. acranthus
Ribs 16–24; epidermis difficult to observe, obscured by spines ....................................................................77. Flowering areoles well differentiated, with tufts of white wool, generally disposed in ring-like
pseudocephalia around the stem, persistent............................................................................9. H. versicolorFlowering areoles not clearly differentiated ................................................................................................8
8. Branches erect; flower-tube slightly curved; seeds with microrelief non-striated to weakly-striated (SEM) ....................................................................................................................................6. H. pseudomelanosteleBranches semi-prostrate; flower-tube markedly curved; seeds with microrelief strongly-striated (SEM) ..........................................................................................................................................................7. H. repens

Bradleya 25/2007 63
period, where the causes of its reduction have notceased and a decline is observed in the area ofoccupancy, extent of occurrence and the quality ofhabitats (particularly those close to the vicinityof Lima).
Comments: The specimen of Cereus acranthusVaupel, collected by Weberbauer in 1902, wasbelieved to have been destroyed during theSecond World War, together with other importantCactaceae collections, until rediscovered byLeuenberger (1978).
Britton & Rose (1920) included this specieswithin Binghamia; however, the use of thisgeneric name was abandoned as it alreadybelonged to an algal genus.
Backeberg (1936) recombined this species asH. acranthus (Vaupel) Backeb., but included inhis concept other taxonomic entities that will betreated in this study as H. acranthus subsp.backebergii.
Ritter (1981) believed this species was in factthe long lost Cereus limensis Salm-Dyck (1845b)and published the combination H. limensis(Salm-Dyck) Ritter. However, the scant descrip-tion and non-existent type of C. limensis Salm-Dyck prevent attribution of the name to the pre-sent species and it is considered a doubtful name(see Appendix 1: List of doubtful names attrib-uted to the genus Haageocereus).
H. acranthus presents several charactershypothesized as plesiomorphies, such as conspic-uous areoles (10.0–11.0 mm diam.) subtended bytransverse rib-folds associated with the forma-tion of podaria, erect growth pattern and someflowers (H. acranthus subsp. backebergii and H. acranthus subsp. zonatus) present the flower-
tube covered by bract-scales subtended by decur-rent podaria (Plates 8.3, 8.4).
The populations of H. acranthus are restrict-ed to the central coast and central-western val-leys in Peru, being most diversified in Lima. H.acranthus is subdivided into three subspecies: H.acranthus subsp. acranthus, H. acranthus subsp.zonatus and H. acranthus subsp. backebergii.
1a. Haageocereus acranthus subsp. acranthusHabit up to 1.4 m tall; branches 4.0–7.0 cm
diam.; ribs 12–14, 11.6–18.6 × 6.2–13.0 mm.Areoles 6.0–10.0 × 5.0 mm diam., 4.8–7.8 mmapart. Central spines 1–2, 15.0–50.0 × 1.0–1.5mm at base; radial spines 30–50, 5.0–14.0 ×0.2–0.5 mm at base. Flowering areoles not differ-entiated. Flowers 7.5–9.7 × 4.2–5.5 cm; peri-carpel 12.0–16.5 × 12.0–17.4 mm; tube 4.4–6.5 ×0.9 cm at base, widening towards apex to 2.25 cmdiam., bearing short brownish hair spines emerg-ing from the bract-scale axils; outer perianth-segments 12, 18.0–26.0 × 4.0–7.0 mm, greenish;inner perianth-segments 22, 22.0–27.0 × 5.7–9.0mm, white, pinkish or reddish; nectar-chamber10.0–28.0 × 6.0–10.0 mm, tubular; anthers2.8–5.4 × 0.6–0.9 mm; style 59.0–78.0 × 1.1–1.3mm; stigma lobes 10, 4.9 mm; ovary locule4.6–12.0 × 4.6–11.0 mm, cylindric to elliptic inlongitudinal section. Fruit 2.0–5.3 × 2.6–4.2 cm.Seeds 1.62 × 1.28 × 0.93 mm, glossy; relief (SEM)convex; microrelief without cuticular striations;hilum large, 0.63 mm, oblique, forming an angleof 64° with long axis of seed (Figure 2, Plates 1.1,1.3, 7.1–7.3).
Examined material: PERU. LIMA: Rímac valley,Santa Clara, Lima–La Oroya road, Loma forma-
Key to subspecies of Haageocereus acranthus1. Flowering areoles well differentiated, woolly, white, generally disposed in ring-like pseudo-
cephalia; flower-tube covered by abundant white trichomes; seed relief with few (2–8) par-convexstructures, testa-cell walls (SEM) with microrelief weakly and regularly striated (northern Lima:Huaura and Pativilca valleys, 1520–2600 m) ............................1c. H. acranthus subsp. zonatusFlowering areoles not clearly differentiated; flower-tube not covered by abundant white tri-chomes; seed relief without par-convex structures, testa-cell walls (SEM) without microrelief (cen-tral and southern Lima: Rímac, Lurín, Chillón and Santa Eulalia valleys, 100–2800 m; Ica: Icavalley, 863 m) ......................................................................................................................................2
2. Radial spines 30–50, covering rib edges; flower-tube covered by flat bract-scales (Lima: Lurín andCañete valleys and deserts, 100–230 m; Ica: Ica valley, 863 m) ..........................................................................................................................................................1a. H. acranthus subsp. acranthusRadial spines (21–) 25–37, not covering rib edges; flower-tube covered by bract-scales subtendedby decurrent podaria (Lima: Chillón, Rímac, Lurín and Cañete valleys, 1260–2800 m) ............................................................................................................1b. H. acranthus subsp. backebergii

Bradleya 25/200764
tion, 400–600 m, 26 Oct. 1902, Weberbauer 1679(B holo, photo); East of Lima, Lomas de Atocongo,200 m, 1956, Rauh K44 (ZSS T23411); CerrosCaracoles, km 55 Panamericana Sur, 100 m, 1Dec. 1980, C. Ostolaza 20 (USM); Lurín valley,Manchay, km 7 Lima–Cieneguilla road,76°51’57’’W, 12°8’26’’S, 200 m, Jan. 2001, N.Calderón 67 (Herb. B. G. La Molina); Cardal, 230m, 20 Feb. 2002, N. Calderón 211, 213 (Herb. B.G. La Molina); ICA: Ica valley, 75°35’56’’W,14°0’59’’S, 863 m, 2005, O. Whaley, photo.
Habitat and distribution: Desert areas and rockyhillsides of valleys on the coast of Lima (100–600 m) and arid valleys in Ica (863 m), (Map 2).
Phenology: Flowers: January; Fruits: February.
Conservation status: Endangered. EN[B1ab(ii,iii,iv)]. The extent of occurrence is estimated tobe 2435 km2, and is severely fragmented.Continuing decline was observed in the area ofoccupancy, the quality of habitats and the num-ber of subpopulations.
This subspecies grows in the proximity ofLima city, facing the constant and increasingthreat of habitat loss, which already caused thedisappearance of populations at Cajamarquillathat were illustrated by Weberbauer (1945) andSanta Clara in the Rímac valley. Less than 50individuals are protected at the ReservaNacional de Lachay (Lima). The negative impactof human expansion is also visible in the serious-ly deteriorated health of the cacti observed atPicapiedra and Cardal in the Lurín valley thatshow spines and epidermis accumulating dustand, in some cases, plastic residue, a saddeningsituation accentuated by the dry weather inLima, where rains are infrequent.
Comments: H. olowinskianus Backeb. was con-sidered by Ostolaza (1998a) as a subspecies of H.acranthus and combined it as H. acranthussubsp. olowinskianus. By comparing the type-material of H. acranthus with the available illus-tration (lectotype) of H. olowinskianus, it becameclear that this name is conspecific with H. acran-thus subsp. acranthus.
A number of varieties and subvarieties werecreated by Rauh & Backeberg (1957) whoattempted to describe the slight differences with-in the natural variation of H. olowinskianus,such as H. olowinskianus var. repandus (stemsprostrate at base), H. olowinskianus var. repan-dus subvar. erythranthus (flower reddish), H.
olowinskianus var. rubriflorior (flower reddish)and H. olowinskianus var. subintertextus (over-lapping spines), which are here treated as syn-onyms.
Rauh & Backeberg (1957) also described H. lachayensis (from “Lomas de Lachay”) and H. clavispinus Rauh & Backeb. (club-shapedspines) that do not exhibit significant differencesfrom H. acranthus subsp. acranthus. H. vulpesRitter (1981) refers to the same morphologicalcharacters of H. acranthus subsp. acranthus butfrom a different locality, in Chancay. Study of thetype-specimen as well as the SEM’s of seeds of H. vulpes confirms that this name is better con-sidered as a synonym.
H. acranthus subsp. acranthus is character-ized by erect columnar stems, relatively low ribnumber (12–14), thick greyish central spines(1.0–1.5 mm diam. at base), seed cuticle withoutstriations (SEM) and undifferentiated flower-bearing areoles. The inner perianth segments ofthe flower vary in colour from typically white tosometimes pinkish or reddish. This subspeciesgrows sympatrically with H. pseudomelanostele(Werderm. & Backeb.) Backeb. and in associationwith other plants in Lima (Calderón 2006).
1b. Haageocereus acranthus subsp. backebergii N. Calderón subsp. nov. Holotype:East and above Chosica, in the Rímac valley,1953, Ritter FR 145 (U 0008476!).H. limensis var. andicola Ritter, Kakt. Südamer.
4: 1397, diagn. lat. (1981). Type: as above.H. acranthus var. crassispinus Rauh & Backeb.
in Backeb. Descr. Cact. Nov. [1]: 22 (1956,publ. 1957); Die Cact. 2: 1177 (1960). Type:Peru, Ica, Cañete valley and Pisco valley,1000 m, 1954, Rauh K31 (HEID, believed notto have been preserved). Neotype: Backeb.,Die Cact. 2: 1182, fig. 1134 (1960). Synon.nov.
Habit up to 1.55 m tall, branching at base;branches straight, 5.1–7.6 cm diam.; ribs10–13(–14), 11.6–18.6 × 6.2–13.0 mm. Areoles8.0–11.2 × 5.0–8.1 mm, 7.0–16.5 mm apart.Central spines 1–2(–3), 7.4–28.7 × 1.0–1.9 mm atbase; radial spines (21–) 25–34, 3.0–6.8 (–11.3) ×0.3–0.7 mm at base. Flowering areoles not differ-entiated. Flowers 7.0–8.4 × 3.5–6.0 cm; peri-carpel 9.0–14.0 × 13.0–16.5 mm; tube 4.0–4.8 ×1.3–1.65 cm at base, widening towards apex to2.4–2.5 cm diam., bearing short brownish hairspines emerging from the bract-scale axils, thebract-scales subtended by decurrent podaria;

Bradleya 25/2007 65
Plate 7. 7.1 H. acranthus subsp. acranthus. N. Calderón s.n., Cardal in the Lurín valley, 2001. 7.2 Ibid., Picapiedrain the Lurín valley, 2001, in fruit. 7.3 Ibid., N. Calderón 213, Chillón valley, in bud and flower.
1
2 3

Bradleya 25/200766
Plate 8. 8.1 H. acranthus subsp. backebergii. N. Calderón 499, Rímac valley. 8.2 Ibid., N. Calderón s.n., cult. LaMolina Univ. Botanic Garden, 2005, in fruit. 8.3 Ibid., N. Calderón s.n., Chillón valley, in bud. 8.4 Ibid., N.Calderón 496, in bud.
1 2
3 4

Bradleya 25/2007 67
Plate 9. 9.1 H. acranthus subsp. zonatus. N. Calderón s.n., Pativilca valley, 2005. 9.2 Ibid. N. Calderón 362, inflower. 9.3 Ibid., N. Calderón s.n., Huaura valley, 2005, in fruit. 9.4 Ibid., N. Calderón s.n., Pativilca valley, 2005.
1 2
3 4

Bradleya 25/200768
Map 2. H. acranthus subsp. acranthus = �; H. acranthus subsp. backebergii = �; H. acranthus subsp. zonatus = .
N
100 0 100 200 Kilometers

Bradleya 25/2007 69
outer perianth-segments 10–12, 22.3–29.0 ×6.2–8.6 mm, green; inner perianth-segments10–13, 21.7–26.0 × 7.2–8.3 mm, white; nectar-chamber 14.0–16.0 × 4.2–10.0 mm, tubular;anthers 2.9–4.0 × 0.8–1.0 mm; style 55.0–62.0 ×1.4–2.2 mm; stigma-lobes 10, 3.8–5.0 mm; ovarylocule 4.0–8.0 × 7.0–9.5 mm, elliptic in longitudi-nal section. Fruit 2.3 × 2.9 cm. Seeds 1.36 × 1.06mm, 0.73 mm thick, glossy; relief (SEM) convex;microrelief without cuticular striations; hilumlarge, 0.59 mm, oblique, forming an angle of 42°with long axis of seed (Figure 3, Plates 1.2,8.1–8.4).
Examined material: PERU. LIMA: Rímac valley,east and above Chosica, 1953, Ritter FR 145 (Uholo 0008476); km 70 Lima–La Oroya road, 2200m, 14 April 1980, C. Ostolaza 47 (USM); km 54Lima–La Oroya road, 16 Feb. 2001, N. Calderón89, 92 (Herb. B. G. La Molina); km 55 Lima–LaOroya road, 76°31’34.8’’W, 11°54’11.5’’S, 1460 m,19 Aug. 2005, N. Calderón 496, 497, 498, 499, 500(Herb. B. G. La Molina); Santa Eulalia valley,Huinco, 1956, Ritter FR 145c (U holo 0008479);loc. cit. 1300 m, 6 March 2002, N. Calderón 240(Herb. B. G. La Molina); loc. cit., Barba Blanca,76°37’37.4’’W, 11°50’51.8’’S, 1400 m, 19 Aug.2005, N. Calderón 510 (Herb. B. G. La Molina);Chillón valley, surroundings of Canta, 2700–2800m, 3 July 1958, R. Ferreyra 12947 (USM); loc.cit., Umarcata hill, km 69 Lima–Canta road,76°46’54’’W, 11°37’23.4’’S, 1260 m, 8 March 2001,N. Calderón 110, 111a (Herb. B. G. La Molina);Lurín valley, Tinajas canyon, 76°37’40.7’’W,12°07’03.5’’S, 1755 m, 24 July 2005, N. Calderóns.n. (Herb. B. G. La Molina, photo), loc. cit.,Antioquía–Langas road, 76°28’10.3’’W,12°06’14.2’’S, 1804 m, 22 Oct. 2005, N. Calderóns.n. (Herb. B. G. La Molina, photo); Cañete valley,Cantera Baja, 75°56’42.6’’W, 12°45’24.9’’S, 1380m, C. Véliz, photo.
Habitat and distribution: Found in the Rímac,Chillón, Santa Eulalia, Lurín and Cañete valleys,all in Lima department (1260–2800 m) and Piscovalley in Ica department (1000 m), (Map 2).
Phenology: Flowers: February and August;Fruits: March and September.
Conservation status: Endangered. EN[B1ab(ii,iii,iv)]. The extent of occurrence is estimated tobe 2379 km2. It is known to exist at no more thanfive locations and continuing decline wasobserved in the area of occupancy, the quality of
habitats and the number of subpopulations, espe-cially in the Rímac and Santa Eulalia valleys.
Comments: Backeberg (1936) misapplied thename H. acranthus to this taxon in the narrowsense. However, it is clearly different from thetype of Vaupel’s species, necessitating the cre-ation of a new name for Backeberg’s taxon at therank of subspecies. Ritter (1981) also recognizedthat the taxon misdetermined by Backeberg(1936) needed a new name and created H. limen-sis var. andicola. H. acranthus subsp. backebergiiis based on Ritter’s H. limensis var. andicola,taking the opportunity to use a more appropriateepithet than Ritter’s relatively meaninglesschoice of ‘andicola’.
Rauh & Backeberg (1957) created H. acran-thus var. crassispinus based on the apparentlythicker and longer (to 3.0 cm) central spines ofthis cactus found in the valleys of Cañete (Lima)and Pisco (Ica). However, these characters arewithin the variation of H. acranthus subsp.backebergii and therefore H. acranthus var. cras-sispinus is here treated as a synonym.
H. acranthus subsp. backebergii is character-ized by its few and short radial spines (21–)25–34, 3.0–6.8 (–11.3) mm long, not covering theedges of the ribs. The flower tube is covered bybract scales subtended by podaria, bearing shortbrownish hair spines in their axils. This sub-species is sympatric with H. pseudomelanostelesubsp. aureispinus in the Chillón valley, and withH. pseudomelanostele subsp. carminiflorus in theSanta Eulalia and Lurín valleys, where it alsogrows among other plant communities (Calderón,2006).
1c . Haageocereus acranthus subsp. zonatus(Rauh & Backeb.) Ostolaza in Ostolaza et al.,Brit. Cact. Succ. J. 21 (2): 94 (2003). H. zonatusRauh & Backeb. in Backeb. Descr. Cact. Nov. [1]:22 (1956, publ. 1957); Die Cact. 2: 1180 (1959).H. limensis var. zonatus (Rauh & Backeb.) Ritter,Kakt. Südam. 4: 1400 (1981). Type: Peru, Lima,Huaura valley, Churín, 2400 m, 1956, Rauh K96(ZSS holo!). H. acranthus var. fortalezensis Rauh & Backeb.
in Backeb. Descr. Cact. Nov. [1]: 22 (1956,publ. 1957). Type: Peru, Lima, Fortaleza val-ley, 1400–1800 m, 1956, Rauh K51a (HEID,believed not to have been preserved). Neotype(designated here): Backeb., Die Cact. 2: 1184,fig. 1136 (1960). Synon. nov.
H. achaetus Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 25 (1956, publ. 1957); Backeb.,

Bradleya 25/200770
A
B
C
D
E
F
G HI
J
K
L
M
N
Figure 2. H. acranthus subsp. acranthus (A, N. Calderón 211; B, N. Calderón 513; C, E, G, H, N. Calderón 224; D,F, I, J, M–N, N. Calderón 216; K–L, N. Calderón 213); A, habit (scale = 50cm); B, stem (scale = 6cm); C–F, flower,side view and longitudinal section (scale = 2cm); G, outer perianth segment (scale = 5mm); H, inner perianth seg-ment (scale = 5mm); I, anther (scale = 1.5mm); J, stigma lobes (scale = 1.5mm); K–N, fruits, side view, and lon-gitudinal section (scale = 3cm).

Bradleya 25/2007 71
A
B
C
D
E
F
G
H I J K L M
NO
P
Figure 3. H. acranthus subsp. backebergii (A, N. Calderón 516; B–E, G–O, N. Calderón 496; F, P, N. Calderón 240);A, habit (scale = 50cm); B, stem (scale = 5cm); C, transverse section of stem (scale = 3cm); D–G, flower, side viewand longitudinal section (scale = 2cm); H–I, bract-scales (scale = 5mm); J–K, outer perianth segments (scale =5mm); L–M, inner perianth segments (scale = 5mm); N, anther (scale = 1mm); O, stigma lobes (scale = 5mm); P,fruit (scale = 3cm).

Bradleya 25/200772
Die Cact. 2: 1176 (1959). Type: Peru, Lima,Churín valley, 1200 m, 1956, Rauh K92(HEID, believed not to have been preserved).Neotype (designated here): Backeb., Die Cact.2: 1210, fig. 1167 (1960). Synon. nov.
H. deflexispinus Rauh & Backeb. in Backeb.Descr. Cact. Nov. [1]: 22 (1956, publ. 1957);Backeb., Die Cact. 2: 1179 (1959). H. acran-thus subsp. deflexispinus (Rauh & Backeb.)Ostolaza in Ostolaza et al., Brit. Cact. Succ. J.21 (2): 94 (2003). H. limensis var. deflexispi-nus (Rauh & Backeb.) Ritter, Kakt. Südam. 4:1399 (1981). Type: Peru, Lima, Churín valley,1200 m, rocky desert area, 1956, Rauh K103(HEID, believed not to have been preserved).Neotype (designated here): Backeb., Die Cact.2: 1185, fig. 1137 (1960). Synon. nov.
Habit up to 1.4 m tall, branching at base;branches straight, 5.0–8.5 cm diam.; ribs 12–15,12.0–18.0 mm. Areoles 6.8–10.0 × 5.3–8.3 mm,5.0–9.0 mm apart. Central spines 1–2, 12.0–55.0 ×0.8–1.6 mm at base; radial spines (21–)25–37,5.2–15.4 × 0.2–0.7 mm at base. Flowering areolesof mature branches woolly, white, generally dis-posed in ring-like pseudocephalia around thestems, persistent. Flowers 8.0–10.0 × 5.6–7.5 cm;pericarpel 13.0–15.0 × 13.0–19.4 mm; tube 5.0–5.4× 1.3–2.0 cm at base, widening towards apex to3.0 cm diam., bearing abundant long white tri-chomes and brownish yellow hair spines emergingfrom the bract scale axils, the bract-scales sub-tended by decurrent podaria; outer perianth-seg-ments 12–14, 20.0–27.5 × 4.3–4.4 mm, greenish;inner perianth-segments 22–24, 25.0–26.0 ×6.5–8.0 mm, white; nectar-chamber 17.0–18.0 ×7.0–8.5 mm, tubular; anthers 3.8–6.0 × 0.9–1.3mm; style 69.0–75.0 × 1.0–2.5 mm; stigma-lobes6.0 mm; ovary locule 7.0–9.2 × 8.0–10.0 mm, cylin-dric to elliptic in longitudinal section. Fruit4.0–4.8 × 4.2–5.5 cm, pericarp bearing few toabundant white trichomes. Seeds 1.45 × 1.11–1.19mm, 0.78–0.79 mm thick, glossy; relief (SEM)with par-convex structures; microrelief with cuti-cle weakly striated; hilum large, 0.63–0.75 mm,oblique, forming an angle of 44° with long axis ofseed (Figure 4, Plates 1.4–1.6, 9.1–9.4).
Examined material: PERU. LIMA: Peru, Lima,Huaura valley, Churín, 2400 m, 1956, Rauh K96(ZSS holo!); Churín–Andajes road, 2500 m, 10June 2002, N. Calderón 280, 281 (Herb. B. G. LaMolina); loc. cit., road towards Cochamarca,77°07’11.58’’W, 10°53’35.82’’S, 2500 m, 22 Nov.2003, N. Calderón 362 (Herb. B. G. La Molina);
loc. cit., 77°05’26.6’’W, 10°58’15.3’’S, 1520 m, 17Feb. 2004, N. Calderón 379 (Herb. B. G. LaMolina); loc. cit., road towards Paccho,76°58’40.1’’W, 10°53’38.8’’S, 1670 m, 18 Feb.2004, N. Calderón 385 (Herb. B. G. La Molina);loc. cit., 76°58’42.1’’W, 10°53’40.3’’S, 1752 m, 5March 2005, N. Calderón 425, 426, 429, 430, 437(Herb. B. G. La Molina).
Habitat and distribution: North of Lima, rockyhillsides of Huaura and Pativilca valleys,1520–2600 m.
Phenology: Flowers: December, January andFebruary; Fruits: January, February and March.
Conservation status: Endangered. EN[B1ab(ii,iii)]. The extent of occurrence is estimated tobe 1880 km2, and it is known to exist at no morethan five locations, where continuing decline inthe area of occupancy and the quality of habitatshas been observed.
Comments: This taxon was first described byRauh & Backeberg (1957) and its type is held atZSS. Rauh & Backeberg (1957) created H. acran-thus var. fortalezensis to describe a cactus with“decumbent” branches, 14–15 ribs and persistentflower-bearing areoles in the Fortaleza valley.From Backeberg’s illustration (1960) and recentfield studies it can be stated that this plant is notdecumbent, being in fact a synonym of H. acran-thus subsp. zonatus.
Rauh & Backeberg (1957) also created H.deflexispinus and H. achaetus, both lacking dis-tinctive flowering areoles, and for the lattername, as having extremely thick stems (to 15 cmdiam.). Nevertheless, the lack of flowering are-oles constitutes merely a juvenile stage, and it ispresumed that the quoted original stem mea-surement is mistaken (considering the thickeststem observed in Haageocereus is to 8.5 cmdiam.). For these reasons, H. deflexispinus andH. achaetus are treated here as synonyms.
H. acranthus subsp. zonatus is characterizedby woolly flowering areoles, disposed in ring-likepseudocephalia, a hypothesized apomorphy andby flowers and fruits bearing few to abundantwhite trichomes and/or hair spines. The seed’stesta cells present par-convexities towards theseed border, this character being unique withinthe genus. This subspecies grows sympatricallywith H. pseudomelanostele subsp. aureispinus inthe Huaura and Pativilca valleys and in associa-tion with other cacti (Calderón, 2006).

Bradleya 25/2007 73
2. Haageocereus chilensis Ritter ex D. Hunt,Cact. Syst. Init. 20: 19 (2005). Type: Chile, dept.Arica, below Chapiquiña, 1957, Ritter s.n. (Uholo!). H. chilensis Ritter, Winter [seed] Katalog 5: 9
(1958), nom. nud. H. fascicularis sensu Ritter,Kakt. Südam. 3: 1125 (1980), non Cactus fas-cicularis Meyen, Allg. Garten. 1: 211 (1833).
Habit semi-decumbent to erect, up to 0.5 mtall, branching at base; branches upright, 4.0–7.0cm diam., epidermis green to greyish; ribs 12–18,5.0–8.0 mm tall. Areoles 4.0–8.0 mm diam., 10.0mm apart, felt brownish at first, later grey. Spinesopaque, brown and grey; central spines 1–2,40.0–150.0 mm, ascending and descending; radialspines 7–10(–11), (10.0–)15.0–40.0 mm. Flowers7.0–8.5 cm; tube 2.5 × 1.0 cm at base, wideningtowards apex, green, bearing few hair spinesemerging from the bract-scale axils; outer peri-anth-segments reddish green; inner perianth-segments 25.0 × 6.0–8.0 mm, white; nec-tar-chamber 20.0 × 10.0 mm, tubular; style 55.0mm; stigma-lobes 10, 6.0 mm; ovary locule 10.0 ×4.0 mm. Fruit ovoid. Seeds ovate, medium-sized,1.87 × 1.13 mm, 1.03 mm thick, glossy; relief(SEM) convex; microrelief without cuticular stria-tions; hilum large, 0.86 mm, oblique, forming anangle of 39° with long axis of seed (Plates 2.1–2.3).
Examined material: CHILE. ARICA: BelowChapiquiña, 2000–3000 m, 1957, Ritter s.n. (Uholo); TARAPACÁ: Camiña, Sept. 1960, RitterFR601 (ZSS SR13517, seeds only); IQUIQUE: 63km from the Panamericana towards Mamiña,Quebrada Guataguata, small valley on the leftside of the road, 69°15’49.8’’W, 20°4’39’’S, 2400 m,2 Feb. 1997, Eggli & Leuenberger 2732 (ZSS18040); loc. cit., Quebrada Mamiña, 69°12’33.6’’W,20°4’19.2’’S, 2800 m, 24 Feb. 1997, Eggli &Leuenberger 2747 (ZSS 18046); loc. cit., hill north-east of the village of Mamiña, 69°25’24’’W,19°18’28.8’’S, 2450 m, 24 Feb. 1997, Eggli &Leuenberger 2788 (ZSS).
Habitat and distribution: Found in rocky hillsidesof Andean valleys in northern Chile, 2000–3000 mand also reported for southern Peru in Tacna (butnot confirmed), (Map 3).
Phenology: Flowers: February; Fruits: Februaryand March.
Conservation status: Ritter (1980) recorded thistaxon for the southernmost part of Peru, in
Tacna, but it has not been observed by the firstauthor in Peru. Populations reported byHoffmann & Walter (2004) for the region ofTarapacá in the Andean Cordillera are believedto be threatened.
Comments: The name H. chilensis first appearedin H. Winter´s seed catalogue (1958) as ‘100a FR601 Ritter sp. nov.’ without a description. Later,Ritter (1981) listed H. chilensis as a nomennudum under H. fascicularis (Meyen) Ritter,based on his collections FR 601 (U!) and FR 125.Because H. fascicularis is based on the old Cereusfascicularis Meyen (1833), a name without extantmaterial, illustration or a meaningful descrip-tion, that also had been taken as the type speciesof Weberbauerocereus, it is preferable not to usethis name (Hunt, 2005). The only specimen orig-inally labelled H. chilensis is Ritter FR s.n.(Utrecht Herbarium). Hunt (2005) cited thisspecimen as the holotype when validatingRitter’s catalogue name.
H. chilensis Ritter ex D. Hunt is characterizedby a low radial spine number of 7–10(–11), longradial spines, to 4.0 cm and very long centralspines, to 15.0 cm. The areoles are very conspicu-ous and the plant reaches to 0.5 m tall.
3. Haageocereus decumbens (Vaupel) Backeb.,Blätt. Kakt.-forsch. 1934 (6): unpaged, (1934);Backeb. & F. M. Knuth, Kaktus-ABC: 207 (1936);Krainz, Die Kakteen 1(11): CVa, unpaged (1965);Backeb., Die Cact. 2: 1237 (1960); Rauh, Beitr.Kenntn. Peruan. Kakteenveg.: 391 (1958).Binghamia decumbens (Vaupel) Werderm.,Kakteenkunde: 24 (1937c). Cereus decumbensVaupel, Engler Bot. Jahrb. Syst. 50, Beibl. 111: 18(1913). Type: Peru, Arequipa, Mollendo, sandycoast, 50–100 m, 7 Oct. 1902, Weberbauer 1550(B, photo!).H. decumbens var. spinosior Backeb., Cact. Succ.
J. (US) 23 (2): 47, 1951; [First mentioned:Backeb. & F. M. Knuth, Kaktus-ABC: 208(1936), nom. inval. (McNeill et al., 2006)]; H.decumbens f. spinosior (Backeb.) Krainz, DieKakt. CVa, unpaged (1965). Type locality:Peru, Arequipa, Mollendo (believed not tohave been preserved). Neotype (designatedhere): Backeb., Die Cact. 2: 1239, fig. 1203(1960). Synon. nov.
H. ambiguus var. reductus Rauh & Backeb. inBackeb. Descr. Cact. Nov. [1]: 25 (1956, publ.1957). Type: Peru, Arequipa, Atico, Pana -mericana km 725, 1956, Rauh K133 (HEID,believed not to have been preserved). Neotype

Bradleya 25/200774
A
B
C
D
E
FG H I J
K
L
Figure 4. H. acranthus subsp. zonatus (A–B, N. Calderón 437; C–J, N. Calderón 280; K–L, N. Calderón 384); A,habit (scale = 50cm); B, stem (scale = 5cm); C–E, flower, side view and longitudinal section (scale = 3cm); F,bract-scale (scale = 5mm); G, outer perianth segment (scale = 5mm); H, inner perianth segment (scale = 5mm);I–J, anthers (scale = 3mm); K–L, fruits, side view and longitudinal section (scale = 2cm).

Bradleya 25/2007 75
A
B
F
C
D E
Figure 5. H. decumbens (A, N. Calderón 451; B, N. Calderón 454; C–E, Ritter FR582; F, Ritter FR187); A, habit(scale = 10cm); B, stem (scale = 2cm); C, flower, (scale = 2cm); D, outer perianth segment (scale = 5mm); E, innerperianth segment (scale = 5mm); F, fruit (scale = 3cm.).

Bradleya 25/200776
(designated here): Backeb., Die Cact. 2: 1247,fig. 1212 (1960). Synon. nov.
H. mamillatus var. brevior Rauh & Backeb. inBackeb. Descr. Cact. Nov. [1]: 26 (1956, publ.1957). Type: Peru, Arequipa, Ocoña–Camaná,desert, 800 m, gypsic, 1956, Rauh K137 (ZSSholo!). Synon. nov.
H. australis f. nanus Ritter, Kakt. Südam. 3:1127 (1980). Type: Peru, Moquegua, Ilo, June1954, Ritter FR 126c (ZSS iso!). Synon. nov.
H. decumbens var. brevispinus Ritter, Kakt.Südam. 4: 1392 (1981). Type: Peru, Arequipa,northern Ocoña, coast, Ritter FR1024 (Uholo!). Synon. nov.
H. chalaensis Ritter, Kakt. Südam. 4: 1389(1981). Type: Peru, Arequipa, Chala, coast,1953, Ritter FR187 (U holo!); 9 September1954, Ritter FR187 (ZSS iso!). Synon. nov.
H. subtilispinus Ritter, Kakt. Südam. 4: 1419(1981). Type: Peru, Arequipa, Atico, 1954,Ritter FR582 (U holo!). Synon. nov.
The following are also probably referrable here:H. australis Backeb., Jahrb. Deutsch Kakt-Ges. 1:
104 (1936); Blätt. Kakt.-forsch. 1937(5):unpaged (1937). Type locality: South of Peru(believed not to have been preserved).
H. ambiguus Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 25 (1956, publ. 1957). Type:Peru, Arequipa, Atico, km 725 Panamericanahighway, 1956, Rauh K132 (HEID, believednot to have been preserved).
H. litoralis Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 26 (1956, publ. 1957). Type:Peru, Arequipa, hills near Atico, 1956, RauhK157 (HEID, believed not to have been pre-served).
H. mamillatus Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 25 (1956 publ. 1957). Type:Peru, Arequipa, Camaná, 400 m (km 165Arequipa–Camaná road), 1956, Rauh K139(HEID, believed not to have been preserved).
H. ocona-camanensis Rauh & Backeb. in Backeb.Descr. Cact. Nov. [1]: 26 (1956, publ. 1957).Type: Peru, Arequipa, sandy desertOcoña–Camaná, 200 m, 1956, Rauh K155(HEID, believed not to have been preserved).
Habit decumbent, branching at base; branch-es prostrate, 2.0–5.0 cm diam., 0.25–0.6(–1.0) mlong, with ascending apices 5.0–10.0 cm abovethe ground, epidermis green; ribs 14–20, 5.6–8.0× 2.3–3.9 mm. Areoles 2.5–6.0 × 1.5–3.5 mmdiam., 1.4–8.8 mm apart, oval to circular, feltbrownish grey and white. Spines opaque, lightbrown and grey; central spines 1–2, 7.0–28.0 ×
1.0–1.5 mm at base, ascending and descending;radial spines 19–33, 3.0–7.0 × 0.2–0.5 mm atbase. Flowers 6.0–6.5 cm; pericarpel 7.9–8.6 ×10.0–11.4 mm; tube 3.2–3.6 × 0.9–1.0 cm at base,widening towards apex to 1.6 cm diam., bearingfew hair spines emerging from the bract-scaleaxils; outer perianth-segments 15.0 × 4.0 mm;inner perianth-segments 10.0 × 5.0 mm, white;nectar chamber 18.6–20.0 × 6.4–7.1 mm, tubular,straight; anthers 3.2 × 1.1 mm; stigma-lobes 12;ovary locule elliptic, 3.6–4.0 × 5.7–6.8 mm. Fruits4.0 × 3.8 cm. Seeds broadly ovate, medium-size,1.41 × 1.15 × 0.86 mm, lustre glossy; relief (SEM)convex; microrelief without cuticular striations;hilum large, 0.78 mm, oblique, forming an angleof 55° with long axis of seed (Figure 5, Plates2.4–2.6, 10.1–10.3).
Examined material: CHILE. TARAPACA: Arica,Jun. 1954, Ritter FR 126b, (ZSS T4529); loc. cit.,Ritter, Kakt. Südamer. 3: 1228, fig. 1089 (1980).PERU. AREQUIPA: Mollendo, sandy soils,50–100 m, 7 Oct. 1902, Weberbauer 1550 (B, type);loc. cit., 70°52’44.8’’W, 17°00’9.6’’S, 150 m, 15March 2004, N. Calderón 418 (Herb. B. G. LaMolina); loc. cit., 1955, Ritter FR 126, (ZSST4528); Ocoña–Camaná, 800 m, gypsic, 1956,Rauh K137 (ZSS); Northern Ocoña, coast, RitterFR1024 (U); Atico, 1954, Ritter FR 582 (U); Chala,coast, 1953, Ritter FR 187 (U holo!); loc. cit., 9September 1954, Ritter FR 187 (ZSS); Camaná,km 955 Panamericana Sur, 72°37’36.7’’W,16°34’10.4’’S, 620 m, 16 March 2004, N. Calderón415, 416, 417 (Herb. B. G. La Molina);MOQUEGUA: Ilo, June 1954, Ritter FR 126c(ZSS); TACNA: Morro Sama, rocky hills facing thePacific ocean, 72°1’28.8’’W, 18°00’6.12’’S, 128 m,14 March 2004, N. Calderón 412, 413, 414 (Herb.B. G. La Molina); km 113 Costanera highway,towards Tacahuay hills, sandy area, 71°7’45.2’’W,17°48’50.5’’S, 84 m, 5 April 2005, N. Calderón 453,454, 455 (Herb. B. G. La Molina).
Habitat and distribution: Found in sandy androcky areas of the Pacific coastal desert in south-ern Peru (Arequipa and Tacna) 50–620 m, andnorthern Chile (Arica), (Map 3).
Phenology: Flowers: October; Fruits: November.
Conservation status: In Peru, Vulnerable:VU[B1ab(ii,iii,iv)]. The extent of occurrence isestimated to be 14842 km2. It is known to exist atno more than ten locations and continuing declinein the area of occupancy, the quality of habitats

Bradleya 25/2007 77
and the number of subpopulations has beenobserved. According to Hoffmann and Walter(2004), this species is believed to be threatened inChile. Populations of this species were observedgrowing on sandy and rocky areas close to the seain Arequipa, Moquegua and Tacna along thesouthern coast of Peru. In the case of MorroSama, in Tacna, housing development is increas-ing rapidly, resulting in habitat loss for thisspecies. The other populations, in Arequipa andMoquegua, are under indirect impact fromhuman activities where they are close to high-ways and towns.
Comments: This species was first published asCereus decumbens by Vaupel (1913). The type ofthis name, together with that of Cereus acran-thus, represent the two important Weberbauercollections of Haageocereus held currently at theBerlin Herbarium.
Backeberg (1936) and Backeberg & Rauh(1957) described a number of prostrate cacti grow-ing in diverse localities along the southern coastof Peru such as H. decumbens var. spinosior, H.ambiguus var. reductus and H. mamillatus var.brevior; however, the characters of these taxa arewithin the variation of H. decumbens, with pros-trate stems 0.2–0.9 m long × 4.0–5.0 cm diameter,ribs 15–18, and for these reasons these names arebetter placed as synonyms.
Ritter created a number of taxa which haveslight differences from H. decumbens, such as H.australis f. nanus (Ritter, 1980), based on smallerbranches 10 cm long, H. decumbens var. brevispi-nus (Ritter, 1981), with shorter spines 10.0–20.0mm and radial spines 3.0–5.0 mm, H. subtilispi-nus (1981), with awl-like spines and H. chalaensis(1981), with stems 0.5–1.0 m long, ribs 12–19 andcentral spines 2.0– (10.0) cm long. Since thesecharacters are insufficient to distinguish themfrom H. decumbens, these Ritter names are treat-ed here as synonyms.
Rauh & Backeberg (1957) described H.ambiguus, H. litoralis, H. mamillatus, and H.ocona-camanensis. These taxa, although lackingoriginal preserved material and meaningful illus-trations, their prostrate growth habit, slenderstems 2.0–4.0 cm diameter (except in H. litoralisto 8.0 cm) and proximity to the type locality sug-gest that they very likely belong as synonyms ofH. decumbens.
H. australis Backeb. (1936) lacks a type andmeaningful illustration although, because of itsdecumbent habit, stems 25.0 cm long, 14 ribs and28 radial spines, it seems plausible that H. aus-
tralis may well refer to H. decumbens.H. decumbens (Vaupel) Backeb. is recogniz-
able by its characteristic decumbent habit (henceits epithet), a possible apomorphy for the species.In contrast to the spines with trichomes and seedswith strong cuticular striations found in H.tenuis, H. decumbens lacks any of these charac-ters.
In Peru, H. decumbens grows with other cactilike Neoraimondia arequipensis (Meyen) Backeb.,but so far it has not been found sympatric withother Haageocereus. In Chile, this species growswith Eulychnia iquiquensis Britton & Rose andEriosyce iquiquensis (Ritter) Ferryman(Hoffmann & Walter, 2006).
4. Haageocereus lanugispinus Ritter, Kakt.Südamer. 4: 1395 (1981). Type: Peru, betweenLima and Ancash, North of Pativilca, 1957, RitterFR 583 (U holo!).Possibly referrable here:Pygmaeocereus densiaculeatus Backeb., Descr.
Cact. Nov. [3]: 12 (1963), nom. inval. (McNeillet al., 2006); Kakteenlex.: 377 (1966). Type:Peru? (believed not to have been preserved).
Habit decumbent, 0.1–0.2 m, branching atbase; branches 1.2–2.0 cm diam., epidermisgreen; ribs 12–15, 1.5–2.0 mm high. Areoles 1.5 ×0.8–1.0 mm, 2.0 mm apart, oval, brownish andwhite. Spines opaque, densely covered by whiteplumose trichomes; central spines 0; radial spines25–35, 3.0–5.0 × 0.25 mm diam. at base. Flowers5.0 cm, tube bearing white trichomes emergingfrom the bract-scale axils; inner perianth-seg-ments white; nectar-chamber 12.0 × 2.0 mm,tubular; stigma-lobes included.
Examined material: PERU. LIMA: North ofPativilca, 1957, Ritter 583 (U holo).
Habitat and distribution: North of Pativilca,between Lima and Ancash (Map 5).
Phenology: Not known.
Conservation status: Data Deficient DD. H.lanugispinus has not been found again after itsdiscovery by Ritter; amateurs have reported thisspecies but its locality remains a secret amongstthem and the conservation status remains uncer-tain. Recent attempts to find this species in thefield have not yielded any results.

Bradleya 25/200778
Map 3. H. decumbens = �; H. platinospinus = �; H. chilensis = �. (Chilean localities in yellow.)
N
100 0 100 200 Kilometers

Bradleya 25/2007 79
Map 4. H. pseudomelanostele subsp. pseudomelanostele = ; H. pseudomelanostele subsp. aureispinus = �; H. pseudomelanostele subsp. carminiflorus = �; H. pseudomelanostele subsp. turbidus = �.
N
100 0 100 200 Kilometers

Bradleya 25/200780
Comments: After Ritter (1981) described H.lanugispinus, it has not been found again andtherefore other biological aspects and the conser-vation status are still uncertain.
Pygmaeocereus densiaculeatus has beenregarded by Ritter (1981) and later by growers asa probable synonym of H. lanugispinus based onfew available records provided in the earlierBackeberg descriptions and illustration of P. den-siaculeatus (Backeberg, 1963, 1966). However,the absence of a type, locality and herbariumrecords for P. densiaculeatus, provide insufficientevidence to confirm or reject the proposal thatthese two epithets refer to the same taxon.
H. lanugispinus presents spines bearingwhite trichomes that give the cactus the woollyaspect indicated by its epithet. This character ishypothesized as an autoapomorphy of probablehigh adaptative value. Further fresh materialfrom the field is needed to prepare a completedescription of this species. However, due to thevery small size of the individuals, it is not sur-prising that this species has remained elusive forso long.
5. Haageocereus platinospinus (Werderm. &Backeb.) Backeb. in Backeb. & F. M. Knuth,Kaktus-ABC: 209 (1936); Rauh, Beitr. Kenntn.Peruan. Kakteenveg.: 391 (1958); Backeb., DieCact. 2: 1234 (1960). Cereus platinospinusWerderm. & Backeb. in Backeb., Neue Kakteen:76 (1931). Type locality: Peru, Arequipa, deserticareas (believed not to have been preserved).Lectotype (designated here): Backeb., NeueKakteen: 76, photo on right side (1931). H. pluriflorus Rauh & Backeb. in Backeb. Descr.
Cact. Nov. [1]: 23 (1956, publ. 1957); Backeb.,Die Cact. 2: 1185 (1960). Type: PERU,Arequipa, Majes valley, Hacienda Ongoro,1000 m, 1956, Rauh K151 (HEID, believednot to have been preserved). Neotype (desig-nated here): Backeb., Die Cact. 2: 1213, topfig. 1171 (1960). Synon. nov.
Habit semi-decumbent to erect, up to 0.5 mtall, branching at base; branches curved orupright, 4.0–6.8 cm diam., epidermis green tobrownish; ribs 12–16, 5.0–19.0 × 4.0–7.0 mm.Areoles 5.0–8.0 × 3.0–7.0 mm, 2.1–10.0 mmapart, oval to circular, felt white and grey. Spinesopaque, at first yellow and brown, later grey; cen-tral spines (0–) 1– (2), 13.5–90.0 × 0.6–1.0 mm atbase, ascending and descending; radial spines(12–)14–20, 4.6–10.0 × 0.3–0.6 mm. Flowers6.0–8.0 × 4.2–6.0 cm; pericarpel 9.0–12.0 ×
10.0–12.0 mm; tube 3.5–5.5 × 0.8–1.2 cm at base,widening towards apex to 2.0–2.4 cm diam.,slightly curved, green, bearing few brownish yel-low hair spines emerging from the bract-scaleaxils; outer perianth-segments 17–29, 11.6–28.0× 3.9–6.6 mm, reddish green; inner perianth-seg-ments 12–18, 18.5–33.7 × 6.8–12.6 mm, white;nectar-chamber 6.0–14.4 × 4.2–5.0 mm, tubular;anthers 2.2–5.6 × 0.8–1.4 mm; style 40.0–61.0 ×1.4–1.7 mm; stigma-lobes 9–10, 2.6–4.0 mm;ovary locule 7.0–9.3 × 4.0–6.2 mm, circular toelliptic in longitudinal section. Fruit 3.2–5.0 ×2.5–3.8 cm, spherical to ovoid. Seeds broadlyovate, medium-sized, 1.61–1.73 × 1.02–1.19 ×0.97–1.0 mm, glossy; relief (SEM) convex;microrelief without cuticular striations; hilumlarge, 0.76–0.81 mm, oblique, forming an angle of55° with long axis of seed (Figure 6, Plates3.1–3.3, 11.1–11.3).
Examined material: PERU. AREQUIPA: Yuravalley, hills surrounding town, 71°42’27.6’’W,16°14’53’’S, 2470 m, 9 March 2004, N. Calderón404 (Herb. B. G. La Molina); loc. cit., 71°42’22’’W,16°14’54.3’’S, 2525 m, 9 March 2004, N. Calderón401 (Herb. B. G. La Molina); loc. cit., roadtowards Huanca, 71°47’19.44’’W, 16°14’49.02’’S,2357 m, 17 March 2004, N. Calderón 469 (Herb.B. G. La Molina); loc. cit., 71°42’22.3’’W,16°13’21.9’’S, 2600 m, 1 April 2005, N. Calderón447, 450 (Herb. B. G. La Molina); Tiabaya,Guayrondo chico, sandy and rocky hills,71°36’14.8’’W, 16°27’29.7’’S, 2190 m, 10 March2004, N. Calderón 408, 409, 410 (Herb. B. G. LaMolina); TACNA: Tacna–Tarata road, arid hills,70°6’15.9’’W, 17°42’05.5’’S, 2450 m, 6 April 2005,N. Calderón 456, 457, 458 (Herb. B. G. LaMolina).
Habitat and distribution: Found on rocky hill-sides of valleys in southern Peru, being well rep-resented in Arequipa and Tacna, [1000–]2000–2650 m (Map 3).
Phenology: Flowers: February, March and April;Fruits: March and April.
Conservation status: Endangered. EN[B1ab(ii,iii)].The extent of occurrence is estimated to be 2469km2. It is known to exist at no more than fourlocations and continuing decline in the area ofoccupancy and the quality of habitats has beenobserved.
In most areas, populations grow healthily andabundantly, but in some places like Cerro Verde

Bradleya 25/2007 81
Map 5. H. versicolor subsp. versicolor = �; H. versicolor subsp. pseudoversicolor = �; H. repens = �;H. lanugispinus = �; H. tenuis = .
N
100 0 100 200 Kilometers

Bradleya 25/200782
(Arequipa), much of the land is disturbed by min-ing activities, including the extraction of rocksand other building materials.
Comments: The species was first described byWerdermann & Backeberg (1931) but type mate-rial was not preserved, therefore the photographpublished with the description is here designatedas the lectotype.
Rauh & Backeberg (1957) published H. pluri-florus because of its numerous flowers; however,this is not a constant character and is likely to belinked to the environment and for this reason H.pluriflorus is here recognized as a synonym.
H. platinospinus is characterized by its lowheight (up to 0.5 m), low radial spine number(10–20) and greyish spines (hence its epithet). H.platinospinus has conspicuous areoles and cen-tral spines and ribs may present transversalfolds of the epidermis around areoles, these char-acters being hypothesized as plesiomorphic.
6. Haageocereus pseudomelanostele(Werderm. & Backeb.) Backeb., Blätt. Kakt.-forsch. 1934 (6): unpaged (1934); Backeb. & F. M.Knuth, Kaktus-ABC: 209 (1936); Rauh, Beitr.Kenntn. Peruan. Kakteenveg.: 426 (1958);Backeb., Die Cact. 2: 1226 (1960). H. multangu-laris var. pseudomelanostele (Werderm. &Backeb.) Ritter, Kakt. Südamer. 4: 1406 (1981).Cereus pseudomelanostele Werderm. & Backeb.in Backeb., Neue Kakteen: 74 (1931); Werderm.,Fedde Rep. Spec. Nov. 30: 61 (1932). Type locali-ty: Peru, Lima, Rímac valley, by Cajamarquilla,on the Lima–La Oroya road (believed not to havebeen preserved). Lectotype (designated here):Backeb., Neue Kakteen: 75, photo (1931). H. chosicensis (Werderm. & Backeb.) Backeb.,
Backeb. & F. M. Knuth, Kaktus-ABC, 207(1936). Cereus chosicensis Werderm. &Backeb. in Backeb., Neue Kakteen: 74 (1931);Fedde Rep. Spec. Nov. 30: 60 (1932); Krainz,Die Kakteen 1 (11): CVa, unpaged (1964). Typelocality: Peru, Lima, by Chosica, on theLima–La Oroya road, (believed not to havebeen preserved). Neotype (designated here):Backeb., Die Cact. 2: 1210, fig. 1168 (1960).
H. horrens Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 22 (1956, publ. 1957). Type:Peru, Trujillo, Panamericana highway km720, in arid coastal hills, 1956, Rauh K68(ZSS iso!). Synon. nov.
H. horrens var. sphaerocarpus Rauh & Backeb. inBackeb. Descr. Cact. Nov. [1]: 22 (1956,publ.1957). Type: Peru, Fortaleza river, km
230, 1956, Rauh K48 (ZSS holo!). Synon. nov.H. multangularis sensu Ritter, Kakt. Südamer. 4:
1400 (1981), non Cactus multangularis Willd.1814.
H. pacalaensis Backeb., Backeb. & F. M. Knuth,Kaktus-ABC: 412 (1936); Backeb., Blätt. Kakt.-forsch. 1936(4): unpaged (1936); Die Cact. 2:1202 (1960); Cereus tapalcalaensis Backeb.,Kakteenfreund 2 (5): 54 (1933) nomen. Typelocality: north of Peru, Malabrigo, (believednot to have been preserved). Neotype (desig-nated here): Backeb., Die Cact. 2: 1204, fig.1161 (1960). Synon. nov.
H. pacalaensis var. longispinus (Rauh & Backeb.)Krainz, Die Kakteen C. Va. (1962b). H. lare-densis var. longispinus Rauh & Backeb. inBackeb. Die Cact. 2: 1195 (1960); Descr. Cact.Nov. [1]: 23 (1956, publ.1957). Type locality:Peru, Fortaleza valley, 500 m, (believed not tohave been preserved). Neotype (designatedhere): Backeb., Die Cact. 2: 1200, fig. 1157(1960). Synon. nov.
H. laredensis (Backeb.) Backeb. in Backeb. & F.M. Knuth, Kaktus-ABC: 208, 412 (1936). H.pacalaensis var. laredensis (Backeb.) Krainz,Die Kakteen C. Va. (1962). Cereus pseudome-lanostele var. laredensis Backeb.,Kakteenfreund 2: 54 (1933). Type locality:north of Peru, close to Laredo, (believed not tohave been preserved). Neotype (designatedhere): Backeb., Die Cact. 2: 1200, fig. 1156(1960). Synon. nov.
H. pseudomelanostele subsp. setosus (Akers)Ostolaza, Zonas Aridas 7: 206 (2003). H.pseudomelanostele var. setosus (Akers)Backeb., Cact. Succ. J. (US) 23: 47 (1951).Peruvocereus setosus Akers, Cact. Succ. J.(US) 19 (5): 68 (1947a). Type locality: Peru,south of Lima, Caracoles hills, (believed not tohave been preserved). Lectotype (designatedhere): Akers, Cact. Succ. J. (US) 19 (5): 67, fig.41 (1947a).
H. pseudomelanostele var. clavatus (Akers)Backeb., Rauh, Beitr. Kenntn. Peruan.Kakteenveg.: 429 (1958), nom. inval. (McNeillet al., 2006). H. clavatus (Akers) Cullmann,Kakt. and. Sukk. 8 (12): 180 (1957), nom.inval. (McNeill et al., 2006) given as a comb.nov., but without any indication of abasionym. Peruvocereus clavatus Akers, Cact.Succ. J. (US) 20 (4): 55 (1948a). Type locality:Peru, Lima, north of Lurín river valley(believed not to have been preserved).Lectotype (designated here): Akers, Cact.Succ. J. (US) 20 (4): 55, fig. 35 (1948a).

Bradleya 25/2007 83
H. pseudomelanostele var. chrysacanthus (Akers)Ritter ex Krainz, Kat. ZSS 2: 65 (1967). H.chrysacanthus (Akers) Cullmann, Kakt. and.Sukk. 8 (12): 180 (1957), Nom. inval. (McNeillet al., 2006). Peruvocereus chrysacanthusAkers, Cact. Succ. J. (US) 21 (2): 45 (1949).Type locality: North of Lima, km 226 ofPanamericana highway, (believed not to havebeen preserved). Lectotype (designated here):Akers, Cact. Succ. J. (US) 21 (2): 45, fig. 30(1949).
Peruvocereus setosus var. longicoma Akers, Cact.Succ. J. (US) 19 (6): 91 (1947b). Type locality:South of Lima, Caracoles hills, (believed notto have been preserved). Lectotype (designat-ed here): Akers, Cact. Succ. J. (US) 19 (6): 91,fig. 58 right (1947b). Synon. nov.
[?Cactus multangularis Willd., Enum. Pl. Suppl.:33 (1814). ?Cereus multangularis (Willd.)Haw., Suppl. Pl. Succ.: 75 (1819). Binghamiamelanostele sensu (Vaupel) Britton & Rose,Cact. 2: 167 (1921) non Cephalocereusmelanostele Vaupel (1913). Binghamia mul-tangularis (Willd.) Britton & Rose, Cact. 4:279 (1923). Peruvocereus multangularis(Willd.) Akers, Cact. Succ. J. (US) 22 (6): 174(1950). H. akersii Backeb. in Rauh, Beitr.Kenntn. Peruan. Kakteenveg.: 416 (1958),nom. inval. (McNeill et al., 2006) given as syn-onym of Peruvocereus multangularis. Type:believed not to have been preserved.].
Habit erect, up to 1.4 (–1.7) m tall, branchingat base; branches 4.0–8.0 cm diam., columnar,epidermis green; ribs 17–24, 6.0–11.0 × 2.0–3.5mm, straight. Areoles 4.0–7.2 × 3.0–5.1 mmdiam., 3.0–8.0 mm apart, oval to circular, feltwhite, yellow and grey. Spines opaque, yellow atfirst, later brownish and grey; central spines 0–2,11.0–100.0 × 0.6–1.3 mm at base, ascending anddescending; radial spines 21–56, 5.0–10.0 ×0.2–0.5 mm. Flowering areoles not differentiated.Flowers 5.0–10.0 × 2.5–4.25 cm; pericarpel 10.4 ×8.0–13.0 mm; tube 3.5–6.0 × 0.7–1.7 cm at base,widening towards apex to 1.5–2.7 cm diam.,slightly curved, green, bearing short brown andyellow hair spines emerging from the bract-scalesaxils; outer perianth-segments 12–20, 11.4–18.0× 3.5–5.3 mm, greenish; inner perianth-segments10–35, 10.7–18.0 × 3.8–6.6 mm, white; nectarchamber 12.0–27.0 × 3.9–8.6 mm, tubular;anthers 1.6–5.0 × 0.5–1.1 mm; style 42.0–57.0 ×1.0–2.4 mm; stigma-lobes 9–13, 3.6–6.0 mm;ovary locule 6.0–8.5 × 3.5–7.0 mm, circular toelliptic in longitudinal section. Fruit 3.0–6.5 ×
2.0–5.6 cm, spherical, pericarp red. Seeds broad-ly ovate, medium-sized, 1.26–1.40 × 0.89–1.03 ×0.69–0.75 mm, glossy; relief (SEM) convex;microrelief non-striated to weakly-striated;hilum large, 0.59–0.71 mm, oblique, forming anangle of 26°–54° with long axis of seed.
Habitat and distribution: This taxon can befound in isolated arid areas and valleys in Lima,extending its range to the northern departmentsof Ancash, Lambayeque and La Libertad,50–500(–780) m, and to the southern departmentof Ica, 1500–2000 m (Map 4).
Conservation status: Vulnerable. VU[A4c]. Aninferred population size reduction of ≥ 30% over tenyears, including both past and future time period,where the causes of its reduction have not ceasedand a decline is observed in the area of occupancy,extent of occurrence and the quality of habitats(particularly those close to the vicinity of Lima).
Comments: Britton & Rose (1920) misidentifiedthis taxon as Cephalocereus melanostele Vaupel,the basionym of Espostoa melanostele (Vaupel)Borg and having created the genus BinghamiaBritton & Rose, made the combinationBinghamia melanostele (Vaupel) Britton & Rose.Later, Britton & Rose (1923) considered thisspecies identical to Cactus multangularis Willd.(1814), based on a painting of a plant fromHaworth’s living cactus collection and replacedBinghamia melanostele (Vaupel) Britton & Rosewith Binghamia multangularis (Willd.) Britton& Rose. Later combinations have been madebased on Willdenow’s epithet ‘multangularis’.Nevertheless, from its scant description and non-existent type, it is not possible to be certainwhich taxon Cactus multangularis Willd. actual-ly refers to and it is better left as a doubtful name(See Appendix 2: List of doubtful names attrib-uted to the genus Haageocereus).
Werdermann & Backeberg (1931) did notaccept Binghamia Britton & Rose as a genus andpreferred to describe this species as Cereuspseudomelanostele, establishing a relationshipwith the earlier but misapplied epithetBinghamia melanostele (Vaupel) Britton & Rose.Later, Backeberg (1934) created the genusHaageocereus, making the combination H.pseudomelanostele (Werderm. & Backeb.)Backeb., the type species of the genus. As thegenus Binghamia had been previously occupiedby a genus of algae (Binghamia Agardh, 1894),the genus Haageocereus became widely used in

the following years. In the absence of a type, thepicture published with Cereus pseudomelanosteleWerderm. & Backeb. (1931) is taken as the lectotype.
H. chosicensis (Werderm. & Backeb.) Backeb.(1935) is considered synonymous with H.pseudomelanostele because it shares most of theimportant diagnostic characters with H.pseudomelanostele, such as erect growth habit,numerous radial spines (30) and 19 ribs.
Various taxa were described by Rauh &Backeberg for Haageocereus of central and north-ern localities of the Peruvian coast, such as H.pacalaensis Backeb. (1936), H. laredensisBackeb. (1936), H. laredensis var. longispinusRauh & Backeb. (1957), H. horrens Rauh &Backeb. (1957) and H. horrens var. sphaerocar-pus Rauh & Backeb. (1957). The slight differ-ences amongst these taxa are related to theirheight, varying from 0.8–1.7 m tall, branch diam-eter (6.0–)7.0–8.5–(10.0) cm, rib number from18–22, and numerous spines (40–45). However,these characters fit within the variation recog-nized for H. pseudomelanostele subsp. pseudome-lanostele. As a result of the study of the proto-logues and available types, field observations inthe type localities, further morphological andmicro-morphological studies on seeds (SEM) fromthe type populations, it was concluded that thenames H. pacalaensis, H. laredensis, H. lareden-sis var. longispinus, H. horrens, and H. horrensvar. sphaerocarpus are better placed as syn-onyms of H. pseudomelanostele.
Akers created the genus Peruvocereus (Akers,1947a) to describe a number of taxa, such as P.setosus Akers (1947a), P. setosus var. longicomaAkers (1947b), P. clavatus Akers (1948a) and P.chrysacanthus Akers (1949) that were mainlydistinguished by presenting numerous bristlespines, a variable character that could well rep-resent a form of H. pseudomelanostele, being bet-ter recognised as synonyms.
Ritter (1981) used Cereus multangularis Haw.(1819) to make the combination H. multangu-laris because he intended not to refer to Cactusmultangularis Willd. (1814), or to any of thepaintings subsequentely associated with thisname (See Appendix: List of doubtful namesattributed to the genus Haageocereus). Accordingto Ritter, Cereus multangularis Haw. belonged toa different entity than Cactus multangularisWilld. In doing so, Ritter’s combination excludesthe type and is rendered invalid by lacking a type(McNeill et al., 2006).
H. pseudomelanostele is characterized by sev-
eral plesiomorphic characters, such as erectstems, undifferentiated flower-bearing areolesand slightly curved flower tube. Hypotheticalapomorphic characters are represented by the17–24 ribs (the higher rib number for the genus)and 21–56 radial spines.
H. pseudomelanostele is subdivided in foursubspecies: H. pseudomelanostele subsp.pseudomelanostele, H. pseudomelanostele subsp.aureispinus, H. pseudomelanostele subsp.carminiflorus and H. pseudomelanostele subsp.turbidus.
6a. Haageocereus pseudomelanostele subsp.pseudomelanostele
Habit erect, up to 1.4 (–1.7) m tall; branchesstraight, 4.0–7.0 cm diam.; ribs 18–24, 9.0 mm.Areoles 6.0–7.0 × 4.0–5.0 mm, 6.0 mm apart,bearing long bristle-spines. Central spines 1–2,15.0–50.0 × 0.2–1.2 mm at base; radial spines35–55, 5.0–15.0 × 0.2–0.4 mm at base. Flowers4.0–7.5 cm; pericarpel 9.0–14.7 × 10.5–11.8 mm;tube 2.7–4.9 × 0.7–1.2 cm at base, wideningtowards apex to 1.75–2.1 cm diam.; outer peri-anth-segments 17–20, 10.7–15.3 × 4.0 mm, red-dish and green; inner perianth-segments 24–27,12.3–17.3.0 × 4.0–7.0 mm, pinkish, red or white;nectar-chamber 11.0–21.0 × 5.3–8.0 mm; anthers2.3–2.4 × 1.0 mm; style 34.0–53.0 × 1.2–1.4 mm;stigma lobes 10, 3.7–7.0 × 0.6 mm; ovary locule6.0 × 4.8–10.0 mm, elliptic in longitudinal sec-tion. Fruit 4.0–4.5 × 5.0 cm, spherical to ovoid.Seeds broadly ovate, small, 1.14–1.18 × 0.79–0.90× 0.62–0.70 mm, glossy; relief (SEM) convex;microrelief non-striated to weakly striated;hilum large 0.34–0.68 mm, oblique, forming anangle of 44–64° with long axis of seed (Figure 7,Figure 8: A–D; Plates 3.4–3.6, 12.1–12.4).
Examined material: PERU. LIMA: Rímac valley,Lima–La Oroya road, Cajamarquilla, 500 m, 1May 1980, C. Ostolaza 800065 (Herb. B. G. LaMolina); loc. cit., 19 Nov. 1959, Hoffmann s/n(USM); Fortaleza valley, road towardsChasquitambo km 40, 77°39’26.6’’W,10°21’55.9’’S, 660 m, 4 Aug. 2005, N. Calderón476 (Herb. B. G. La Molina); Pativilca valley,Panamericana Norte km 239, 77°51’6.0’’W,10°37’13.2’’S, 250 m, 5 Nov. 2005, N. Calderón338 (Herb. B. G. La Molina); loc. cit., roadtowards Ocros, 77°29’14.6’’W, 10°37’47.7’’S, 582m, 6 Aug. 2005, N. Calderón 484 (Herb. B. G. LaMolina); ANCASH: Casma–Huaraz road,78°04’54’’W, 09°30’33’’S, 520 m, Nov. 2004, N.Calderón 361 (Herb. B. G. La Molina, seeds);
Bradleya 25/200784

Bradleya 25/2007 85
LAMBAYEQUE: road towards Olmos, km 722Panamericana Norte, rocky hill, 79°33’53.8’’W,07°03’51.6’’S, 50 m, Nov. 2004, N. Calderón 341(Herb. B. G. La Molina); loc. cit., G. Charles597.01 (Herb. B. G. La Molina, seeds).
Habitat and distribution: This taxon can befound in very isolated arid areas and valleys inDepto Lima, extending its range to the northerndepartments of Ancash, Lambayeque and LaLibertad, 50–500(–780) m (Map 4).
Phenology: Flowers: December; Fruits: January.
Conservation status: Vulnerable. VU[B1ab(ii,iii,iv)].The extent of occurrence is 17304 km2. It isseverely fragmented and continuing decline wasobserved in the area of occupancy, the quality ofhabitats and the number of subpopulations.
This subspecies has severely decreased in thelocalities where it used to predominate with habi-tat loss even worse for the last remaining locali-ties surrounding Lima. For this reason, althoughit has an apparently wide distribution in centralPeru, its situation is not free of concern.
Comments: H. pseudomelanostele subsp.pseudomelanostele is characterized by areolesusually bearing long bristle spines, small seeds(1.14–1.18) (SEM) and flowers with inner peri-anth segments white to pinkish. This subspeciesgrows sympatrically with H. acranthus subsp.acranthus and other cacti (Calderón, 2006) invalleys surrounding Lima.
6b. Haageocereus pseudomelanostele subsp.aureispinus (Rauh & Backeb.) Ostolaza Cact.Cons Init. 6:9 (1998). H. aureispinus Rauh &Backeb. in Rauh Beitr. Kenntn. Peruan.Kakteenveg.: 404 (1958). H. pseudomelanostelevar. aureispinus (Rauh & Backeb.) Ostolaza, Brit.
Cact. Succ. J. 14 (4):170 (1996). Type: Peru,Lima, Chillón valley, Canta, 800–1000 m, 1956,Rauh K170 (HEID, believed not to have been pre-served). Neotype (designated here): Backeb., DieCact. 2: annex of photos, unpaged, pl. 86 (1960).H. acanthocladus Rauh & Backeb. in Backeb.
Descr. Cact. Nov. [1]: 23 (1956, publ. 1957);Rauh, Beitr. Kenntn. Peruan. Kakteenveg.:408 (1958); Backeb., Die Cact. 2: 1200 (1960);H. pseudomelanostele subsp. acanthocladus(Rauh & Backeb.) Ostolaza in Ostolaza et al.,Brit. Cact. Succ. J. 21 (2): 94 (2003). Type:Peru, Lima, Churín valley, Sayán, 900 m,1956, Rauh K90 (HEID, believed not to havebeen preserved). Neotype (designated here):Backeb., Die Cact. 2: appendix of photos,unpaged, pl. 85 (1960). Synon. nov.
H. pachystele Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 24 (1956, publ. 1957). Type:Peru, Lima, Churín valley, 900 m, 1956, RauhK91 (HEID, believed not to have been pre-served). Neotype (designated here): Backeb.,Die Cact. 2: 1236, fig. 1199 (1960). Synon.nov.
H. symmetros Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 24 (1956, publ. 1957). Type:Peru, Lima, Churín valley, 1200 m, 1956,Rauh K102 (HEID, believed not to have beenpreserved). Neotype (designated here):Backeb., Die Cact. 2: 1232, fig. 1195 (1960).Synon. nov.
H. viridiflorus (Akers) Backeb., Die Cact.2: 1195(1960). Peruvocereus viridiflorus Akers, Cact.Succ. J. (US) 19: 143 (1947e). Type locality:Peru, Chillón river, about 10 km above SantaRosa de Quives (believed not to have beenpreserved). Neotype (designated here): Akers,Cact. Succ. J. (US) 19: 143, fig. 95 (1947e).Synon. nov.
Key to subspecies of Haageocereus pseudomelanostele1. Areoles usually bearing long bristle-spines; inner perianth segments white, sometimes pinkish; seeds small,
1.14–1.18 mm length (SEM) (central and northern Peru: Lima, Ancash, Lambayeque and La Libertad,50–660 m) ......................................................................6a. H. pseudomelanostele subsp. pseudomelanosteleAreoles rarely bearing bristle-spines; inner perianth segments white or reddish; seeds medium sized,1.26–1.40 mm length (SEM) (central and southern Peru: Lima and Ica valleys, 515–2000 m) ..................2
2. Inner perianth segments reddish (central Lima, Lurín and Santa Eulalia valleys, 930–1750 m) ................................................................................................................6c. H. pseudomelanostele subsp. carminiflorus
Inner perianth segments white (northern Lima and south-east of Ica) ......................................................33. Fruits indehiscent (northern Lima: Chillón and Huaura valleys, 510–1980 m) ..........................................
..................................................................................................6b. H. pseudomelanostele subsp. aureispinusFruits dehiscent (south-east Ica: Ica valley, 950–2000 m) ..............6d. H. pseudomelanostele subsp. turbidus

Bradleya 25/200786
Plate 10. 10.1 H. decumbens. N. Calderón 451, Tacahuay at Tacna. 10.2 Ibid., N. Calderón 454. 10.3 Ibid., N.Calderón 451.
1
2
3

Bradleya 25/2007 87
Plate 11. 11.1 H. platinospinus. N. Calderón 450, Yura valley. 11.2 Ibid., N. Calderón 410, Tiabaya, in fruit. 11.3Ibid., N. Calderón 450, in flower.
1
2 3

Bradleya 25/200788
Plate 12. 12.1 H. pseudomelanostele subsp. pseudomelanostele. N. Calderón s.n., Chicama valley in La Libertad,2003. 12.2 Ibid., Picapiedra in the Lurín valley, 2001. 12.3 Ibid., Fortaleza valley, 2005. 12.4 Ibid., N. Calderón 338,Pativilca valley, in flower.
1
3
2
4

Bradleya 25/2007 89
The following are possibly referrable here:H. aureispinus var. fuscispinus Rauh & Backeb.
in Rauh, Beitr. Kenntn. Peruan. Kakteenveg.:407 (1958). Type: Peru, Lima, Canta (Chillónvalley), 800–1000 m, 1956, Rauh K170b(HEID, believed not to have been preserved).
H. rigidispinus Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 26 (1956, publ. 1957). H.aureispinus var. rigidispinus Rauh & Backeb,Rauh, Beitr. Kenntn. Peruan. Kakteenveg.:407 (1958), nom. inval. (McNeill et al., 2006)based on H. rigidispinus. H. aureispinus var.rigidispinus (Rauh & Backeb.) Rauh &Backeb. in Backeb. Die Cact. 2: 1198 (1960).Type: Peru, Lima, Chillón valley, Canta,800–1000 m, 1956, Rauh K170a (HEID,believed not to have been preserved).
Habit erect, up to 1.4 m tall; branches 5.8–7.5cm diam.; ribs 17–22(–24), 6.0–11.0 mm. Areoles4.0–7.2 × 3.0–5.1 mm diam., 3.0–8.0 mm apart.Central spines 0–2, 11.0–47.0 × 0.7–1.3 mm atbase; radial spines (21–)25–56, 5.0–10.0 ×0.2–0.4 mm at base, yellow, brown, brownish redand grey. Flowers 7.3–7.6(–10.0) × 3.2–3.5 cm;pericarpel 12.0 × 9.0–11.0 mm; tube 4.3 × 0.8–1.1cm at base, widening towards apex to 2.0–2.5 cmdiam.; outer perianth-segments 20, 13.4–16.2 ×3.5–5.3 mm, greenish; inner perianth-segments35, 12.0–18.0 × 5.5–6.6 mm, white; nectar-cham-ber 17.0–18.5 × 4.7–5.6 mm; anthers 2.7–4.2 ×0.5–1.1 mm; style 50.0–55.0 × 1.0 mm; stigma-lobes 11–13, 5.0–6.0 mm; ovary locule 8.0–8.5 ×5.0–7.0 mm, circular to elliptic in longitudinalsection. Fruit 4.5–6.5 × 4.2–5.6 cm, spherical,indehiscent. Seeds broadly oval, medium-size,1.26–1.29 × 0.98–1.03 × 0.74–0.75 mm, glossy;relief (SEM) convex; microrelief without cuticu-lar striations; hilum large, 0.63–0.71 mm,oblique, forming an angle of 42°–54° with longaxis of seed (Figure 8: E–G, K–O; Plates 4.4, 4.5,13.1–13.3).
Examined material: PERU. LIMA: Huaura val-ley, Sayán, 700 m, 9 June 2002, N. Calderón 250,251, 252 (Herb. B. G. La Molina); loc. cit., roadtowards Cochamarca, 77°05’26.6’’W, 10°58’15.3’’S, 1520 m, 17 Feb. 2004, N. Calderón 372,375 (Herb. B. G. La Molina); loc. cit., roadtowards Paccho, 76°58’38.5’’W, 10°53’40.9’’S,1720 m, 18 Feb. 2004, N. Calderón 387 (Herb. B.G. La Molina); loc. cit., 76°58’40.3’’W,10°53’42.5’’S, 1746m, 5 March 2005, N. Calderón428, 434, 435, 438, 439, 440, 442, 442.1 (Herb. B.G. La Molina); Chillón valley, Umarcata hill, km
69 Lima–Canta road, 76°46’54’’W, 11°37’23.4’’S,1260 m, 20 Sept. 2003, N. Calderón 333, 334, 335,336 (Herb. B. G. La Molina).Habitat and distribution: Found in Huaura andChillón valleys in central and northern Lima,515–1750 m (Map 4).
Phenology: Flowers: June and July; Fruits: June,July and August.
Conservation status: Endangered. EN[B1ab(ii,iii)]. The extent of occurrence is estimated tobe 2559 km2; it is known to exist at no more thanfour locations and continuing decline wasobserved in the area of occupancy and the quali-ty of habitats.
Comments: This taxon was described under vari-ous names by Rauh & Backeberg (1957, 1958).Type materials were not preserved, so pho-tographs published later by Backeberg (1960) arehere proposed as the neotypes for the namesinvolved (accepted and synonyms).
H. viridiflorus (Akers) Backeb. (1960) wasfirst described a decade earlier by Akers (1947e)under the genus Peruvocereus for a species grow-ing in the Chillón valley that was distinguishedby sometimes having a green to greenish-whiteflower, but being identical in all other respects toH. pseudomelanostele subsp. aureispinus it istreated here as a synonym.
Rauh & Backeberg described from theHuaura valley H. acanthocladus, H. pachysteleand H. symmetros, and from the Chillón valley,H. aureispinus var. fuscispinus and H. rigidispi-nus, referring to erect plants of 17–24 ribs, withareoles rarely bearing bristles but with numer-ous radial spines (21–) 25–56, flowering areolesnot clearly differentiated, bearing white flowers(inner perianth segments) and indehiscent fruits.All these characters fit well within the circum-scription of H. pseudomelanostele subsp.aureispinus. From the available illustrations andfield studies at the type localities, H. pachysteleand H. symmetros are synonyms of H. pseudome-lanostele subsp. acanthocladus. However, for thenames H. rigidispinus Rauh & Backeb. (1957)and H. aureispinus var. fuscispinus Rauh &Backeb. (1958) that do not have any illustrationavailable, it is difficult to be completely certainthat they are synonyms of H. pseudomelanostelesubsp. aureispinus.
H. pseudomelanostele subsp. aureispinus issympatric with H. acranthus subsp. backebergiiin the Chillón valley and with H. acranthus

Bradleya 25/200790
Plate 13. 13.1 Habitat of H. pseudomelanostele subsp. aureispinus. Paccho hills in the Huaura valley, 2005. 13.2Ibid., N. Calderón s.n., in fruit. 13.3 Ibid., N. Calderón 255, in flower.
2 3
1

Bradleya 25/2007 91
subsp. zonatus in the Huaura, Fortaleza andPativilca valleys.
Intergeneric hybrids between H. pseudome-lanostele subsp. aureispinus and Espostoamelanostele (×Haagespostoa) can be found in theHuaura valley, being more common towardsAncash department, where large populations ofhybrids have been seen in the Fortaleza andPativilca valleys.
6c . Haageocereus pseudomelanostele subsp.carminiflorus (Rauh & Backeb.) Ostolaza inHunt, Cact. Cons. Init. 6: 9 (1998a). H. pseudome-lanostele var. carminiflorus Rauh & Backeb. inBackeb. Descr. Cact. Nov. [1]: 21 (1956, publ.1957); ibid., Die Cact. 2: 1228 (1960); Rauh, Beitr.Kenntn. Peruan. Kakteenveg.: 428 (1958);Ostolaza, Brit. Cact. Succ. J. 14 (4): 167 (1996).Type locality: Peru, Lima, Santa Eulalia valley,1000 m, 1956, Rauh K20 (HEID, believed not tohave been preserved). Neotype (designated here):Backeb., Die Cact. 2: 1229, fig. 1192 (1960).
Habit erect, up to 1.4 m tall; branches 4.0–5.0cm diam.; ribs 18–23, 5.0–6.9 × 3.0–4.7 mm.Areoles 4.0–6.0 × 3.4–4.3 mm, 3.5–4.0 mm apart,oval to circular. Central spines 1–2, 18.0–40.0 ×0.6–1.1 mm at base; radial spines 40–50,5.0–10.0 × 0.2–0.5 mm at base. Flowers 6.3–8.8 ×2.5–4.25 cm; pericarpel 10.5–13.0 × 8.0–13.0 mm;tube 3.5–6.0 × 0.7–1.4 cm at base, wideningtowards apex to 1.5–2.0 cm diam.; outer peri-anth-segments 12–19, 11.4–18.0 × 3.8–5.1 mm,reddish; inner perianth-segments 10–16,10.7–16.5 × 3.8–5.1 mm, reddish; nectar-chamber12.0–27.0 × 3.9–8.6 mm; anthers 2.6–5.0 ×0.7–0.9 mm; style 42.0–57.0 × 1.1–2.4 mm; stigma-lobes 9–11, 3.6–5.4 × 0.6–0.8 mm; ovarylocule 6.0–7.5 × 3.5–5.5 mm, cylindric to ellipticin longitudinal section. Fruit 3.4–3.5 × 4.0–4.5 cm, spherical to ovoid, indehiscent, pericarpred. Seeds broadly ovate, medium-sized, 1.40 ×1.03 × 0.75 mm, glossy; relief (SEM) convex;microrelief without cuticular striations; hilumlarge, 0.66 mm, oblique, forming an angle of 26°with long axis of seed (Plates 4.1–4.3, 14.5, 14.6).
Examined material: PERU. LIMA: Lurín valley,Tinajas canyon, 76°39’50.3’’W, 12°07’05.2’’S, 1282m, 23 Jan. 2002, N. Calderón 175a (Herb. B.G.La Molina); loc. cit., 24 July 2005, N. Calderón470, 471 (Herb. B.G. La Molina); loc. cit.,76°37’9.7’’W, 12°07’33.9’’S, 1743 m, 24 July 2005,N. Calderón 472, 473, 475 (Herb. B.G. LaMolina); Santa Eulalia valley, Barba Blanca,
76°37’37.4’’W, 11°50’51.8’’S, 1400 m, 11 Feb.2003, N. Calderón 298a (Herb. B.G. La Molina);loc. cit., 19 Aug. 2005, N. Calderón 501.1, 501.2,509 (Herb. B. G. La Molina).
Habitat and distribution: Found in the Lurín andSanta Eulalia valleys, in central Lima, 930–1750m (Map 4).
Phenology: Flowers: July and November; Fruits:August and December.
Conservation status: Endangered: EN[B1ab(ii,iii)+2ab(ii,iii)]. The extent of occurrence is estimat-ed to be 186 km2, and the area of occupancy 66km2. It is known to exist at no more than twolocations, and continuing decline was observed inthe area of occupancy and the quality of habitats.
Comments: This subspecies was first described asH. pseudomelanostele var. carminiflorus by Rauh& Backeberg (1957) for a population growing inthe Santa Eulalia valley and, because type mate-rial was not preserved, a photograph publishedlater by Backeberg (1960) is here used to desig-nate a neotype.
H. pseudomelanostele subsp. carminiflorus ischaracterized by its reddish flowers, sometimeswith the perianth limb slightly zygomorphic. It issympatric with H. acranthus subsp. backebergiiand other cacti (Calderón, 2006) in the SantaEulalia and Lurín valleys, where it also occurswith Espostoa melanostele (Vaupel) Borg, form-ing the intergeneric hybrid ×Haagespostoa.
6d. Haageocereus pseudomelanostele subsp.turbidus (Rauh & Backeb.) Ostolaza in Hunt,Cact. Cons. Init. 6: 9 (1998a). H. turbidus Rauh &Backeb. in Backeb. Descr. Cact. Nov. [1]: 25(1956, publ. 1957); Die Cact. 2: 1217 (1960);Rauh, Beitr. Kenntn. Peruan. Kakteenveg.: 425(1958); H. multangularis var. turbidus (Rauh &Backeb.) Ritter, Kakt. Südamer. 4: 1416 (1981).H. pseudomelanostele var. turbidus (Rauh &Backeb.) Ostolaza, Brit. Cact. Succ. J.: 16 (3): 132(1998c). Type locality: Peru, Ica, Nazca valley,1200 m, 1956, Rauh K105 (HEID, believed not tohave been preserved). Neotype (designated here):Backeb., Die Cact. 2: 1223, fig. 1185 (1960).H. turbidus var. maculatus Rauh & Backeb. in
Backeb. Descr. Cact. Nov. [1]: 25 (1956, publ.1957). Type locality: Peru, Ica, Nazca valley,1200 m, 1956, Rauh K110 (HEID, believednot to have been preserved). Neotype (designated here): Backeb., Die Cact. 2: 1224,

Bradleya 25/200792
Plate 14. 14.1 Habitat of H. pseudomelanostele subsp. turbidus. Nazca desert in Ica, 2002. 14.2 Ibid., N. Calderón276, in fruit. 14.3 Ibid., N. Calderón 273, in flower. 14.4 Ibid., N. Calderón 276, in fruit. 14.5 H. pseudomelanostelesubsp. carminiflorus N. Calderón 470, in fruit. 14.6 Ibid., N. Calderón 475, in flower.
1
2
3
4
5
6

Bradleya 25/2007 93
fig. 1186 (1960).
Habit erect, up to 1.4 m tall; branches 5.0–8.0cm diam., columnar, epidermis green; ribs 19,7.0–11.1 × 2.0–3.5 mm, straight. Areoles 4.7–5.2× 3.9–4.8 mm diam., 4.6–5.4 mm apart, oval tocircular. Central spines 1–2, 50.0–100.0 × 0.7–1.0mm at base; radial spines 35–45, 7.5–9.0 × 0.20mm at base. Flowers 5.0–8.0 × 2.5 cm; pericarpel10.4 × 9.0 mm; tube 5.5 × 1.1–1.7 cm at base,widening towards apex to 2.7 cm; outer perianth-segments reddish-green; inner perianth-seg-ments 15.0 × 3.0 mm, white; nectar-chamber 15.2× 8.2 mm; anthers 1.6 × 0.54 mm; style 45.7 ×1.6–2.2 mm; stigma-lobes 10, 7.0–8.0 mm; ovarylocule 6.4 × 4.8 mm, cylindric to elliptic in longi-tudinal section. Fruit 3.0–4.0 × 2.0–3.0 cm,spherical, dehiscent, pericarp reddish. Seedsovate, medium-sized, 1.35 × 0.89 × 0.69 mmglossy; relief (SEM) convex; microrelief withoutcuticular striations; hilum large, 0.59 mm,oblique, forming an angle of 39° with long axis ofseed (Figure 8: G–L, M–N; Plates 4.6, 14.1–14.4).
Examined material: PERU. ICA: Nazca, km 30Puquio–Nazca road, 1500 m, 26 May 2002, N.Calderón 273, 276 (Herb. B. G. La Molina).
Habitat and distribution: Found in the rocky aridmountains of Nazca, 1200–2000 m (Map 4).
Phenology: Flowers: May; Fruits: May
Conservation status: Critically Endangered.CR[B2ab(iii)]. The area of occupancy is less than10 km2. This subspecies is known to exist at onlya single location, and continuing decline has beenobserved in the quality of habitat.
Comments: This subspecies was first described asH. turbidus by Rauh & Backeberg (1957) at theNazca valley. Unfortunately, type material wasnot preserved and a photograph published laterby Backeberg (1960) is here designated as theneotype.
H. turbidus var. maculatus was described byRauh and Backeberg (1957) but its type materialwas not preserved. However, H. turbidus var.maculatus occurs together with H. turbidus anddiffers in spine colour. Such a character wasobserved in the field and proved to be rather vari-able, not justifying recognition of the populationat a taxonomic level, therefore H. turbidus var.maculatus is here considered as a synonym.
H. pseudomelanostele subsp. turbidus is dis-
tinguished by presenting dehiscent fruits, aunique feature in the genus, also uncommon inrelated Trichocereeae, being hypothesized as asecondary apomorphy. This subspecies growstogether with Browningia candelaris Britton &Rose and very little other vegetation in this aridand rocky landscape.
7. Haageocereus repens Rauh & Backeb. inBackeb. Descr. Cact. Nov. [1]: 26 (1956, publ.1957); Die Cact. 2: 1241 (1960); Rauh, Beitr.Kenntn. Peruan. Kakteenveg.: 398 (1958). H.pacalaensis var. repens (Rauh & Backeb.) Krainz,Die Kakteen 1(12): CVa, unpaged (1962b). H.pacalaensis subsp. repens (Rauh & Backeb.)Ostolaza, Brit. Cact. Succ. J. 18 (3): 130 (2000).Type: Peru, La Libertad, sandy desert betweenCasma and Trujillo, 1956, Rauh K88 (HEID,believed not to have been preserved). Neotype(designated here): Backeb., Die Cact. 2: 1243, fig.1208 (1960).
Habit semi-decumbent, branching at base;branches, 4.5–5.4(–8.0) cm diam., 1.0–2.0 mlong., part of branches growing erect ≥ 20.0 cmabove the ground, epidermis green; ribs 19–20.Areoles 4.0–5.0 × 2.0–3.0 mm, 5.0–10.0 mmapart, oval, felt grey. Spines opaque, at first yel-low and brown, later grey; central spines 1(–2),15.0–30.0 × 0.4–0.6 mm at base, ascending anddescending; radial spines (25–)30–40, 5.0–10.0 ×0.2–0.3 mm at base. Flowering areoles not differ-entiated. Flowers 7.0 × 3.5 cm; tube markedlycurved, green, bearing few hair spines emergingfrom the bract-scale axils; outer perianth-seg-ments reddish-brown; inner perianth-segmentswhite; nectar-chamber 13.0 × 6.0 mm, tubular.Fruit ovoid. Seeds broadly ovate, medium-sized,1.48 × 1.04 × 0.77 mm, semi-matt; relief (SEM)convex; microrelief with strong cuticular stria-tions; hilum large, 0.64 mm, oblique, forming anangle of 42° with long axis of seed (Figure 9,Plates 5.1–5.3, 15.1–15.3).
Examined material: PERU. LA LIBERTAD:South of Trujillo, Panamericana highway km546, sandy desert, 78°56’40’’W, 8°13’42.7’’S, 130m, 18 Nov. 2003, N. Calderón 358 (Herb. B. G. LaMolina); loc. cit., G. Charles 254.01 (seeds, pri-vate collection).
Habitat and distribution: Found in sandy desertin northern Peru, 130 m (Map 5).

Bradleya 25/200794
A
D
E
F
G
H
B
I
L
J
K
M N
O P
QRS
TU
V
W
C
C
V
Figure 6. H. platinospinus (A, C, N. Calderón 444; B, D, E, J, N, P, S, T, N. Calderón 447; F–I, K–M, O, Q, R, U,N. Calderón 450; V–W, N. Calderón 443); A, habit (scale = 20cm); B, stem (scale = 3cm); C, transverse section ofstem (scale = 3cm); D–H, flower, side view and longitudinal section (scale = 2cm); I–L, bract-scales(scale = 5mm);M–N, outer perianth segment (scale = 5mm); O–P, inner perianth segment (scale = 5mm);Q–S, anthers (scale =2mm); T–U, stigma lobes (scale = 2mm); V–W, fruits, side view and longitudinal section (scale = 3cm).

Bradleya 25/2007 95
A
B
C
E
F
G
H
I
J
K L MN
O
P
QD
Figure 7. H. pseudomelanostele subsp. pseudomelanostele (A–D, N. Calderón 474; E–G, K–O, C. Ostolaza s.n.); H.pseudomelanostele subsp. carminiflorus (H, J, Q, N. Calderón 63; I, P, N. Calderón 175); A, habit (scale = 50cm);B, stem (scale = 5cm); C, transverse section of stem (scale = 2cm); D, areole (scale = 1cm); E–J, flower, side viewand longitudinal section (scale = 2cm); K, bract-scale (scale = 5mm); L, outer perianth segment (scale = 5mm);M, inner perianth segment (scale = 5mm); N, anther (scale = 1mm); O-P, stigma lobes (scale = 5mm); Q, fruit(scale = 3cm).

Bradleya 25/200796
A
B
C
D
E
F
G
H
I
J
K
L
M
NO
Figure 8. H. pseudomelanostele subsp. pseudomelanostele (A–C, N. Calderón 338; D, N. Calderón 232); H.pseudomelanostele subsp. aureispinus (E–G, K–O, N. Calderón 250); H. pseudomelanostele subsp. turbidus (G–L,N. Calderón 273; M–N, N. Calderón 276); A–L, flower, side view and longitudinal section (scale = 2cm); M–O,fruits (scale = 3cm).

Bradleya 25/2007 97
Phenology: Flowers: November; Fruits:November
Conservation status: Critically Endangered:CR[B2ab(ii,iii)]. The area of occupancy is esti-mated to be less than 3 km2, and it is known toexist only at a single locality and continuingdecline has been observed in the area of occupan-cy and in the quality of the habitat.
The habitat of H. repens is likely to disappearas a result of the agricultural pressure from theChavimochic irrigation project for growingasparagus and other crops.
Comments: The species was first described byRauh & Backeberg (1957) but its material waseither never preserved or subsequently lost. Aneotype is designated here based on an illustra-tion published slightly later by Backeberg (1960).
H. repens was recombined as H. pacalaensisvar. repens (Rauh & Backeb.) Krainz (1962b) andlater as H. pacalaensis subsp. repens (Rauh &Backeb.) Ostolaza (2000) because the growthhabit was considered the only difference betweenthe populations of H. repens Backeb. & Rauh(prostrate stems) and H. pacalaensis Rauh &Backeb. (erect stems).
H. repens was observed in its type localitygrowing semi prostrate, sometimes with fewstems creeping, and with flowers with a marked-ly curved tube. The study of the seeds has shownstrong secondary folding of the testa (SEM). Allthese characters contrast with the erect growinghabit, slightly curved flowers and seeds withoutstriations seen in H. pacalaensis Rauh &Backeb., suggesting that H. repens Rauh &Backeb. should be accepted at specific level. H.pacalaensis Rauh & Backeb., on the other hand,is being treated in this study as a synonym of H.pseudomelanostele (Werderm. & Backeb.)Backeb.
H. repens is characterized by a semi-prostraterather than a totally decumbent habit (hence itsepithet), markedly curved flowers and stronglystriated cuticle of the seed testa (SEM), and thesecharacters are hypothesized as apomorphic.
No other plants are found growing in thesame habitat and the closest populations of H.pseudomelanostele subsp. pseudomelanostele arefound 20 km north-east, in the Moche valley(dept. La Libertad).
8. Haageocereus tenuis Ritter, Kakt. Südamer.4: 1421 (1981); Ostolaza & Rauh, Kakt. and.Sukk. 41(2): 44 (1990). Type: PERU, Lima,
between Chancay and Huacho, nearPanamericana norte highway, July 1956, RitterFR126e (U holo!).H. australis f. subtilispinus Ritter, Kakt.
Südamer. 3: 1127 (1980). Type: Chile,Pisagua, Junín, 1954, Ritter FR126a (U holo!). Synon. nov.
Habit decumbent, branching at base andalong the main stems, each individual occupyingup to 2 m2 area; branches prostrate with ascend-ing apices up to 5.0–10.0 cm, 1.4–3.4 cm diam.,0.3–1.0 m long., epidermis green; ribs 12–15,1.5–2.5 × 4.0–5.0 mm, straight. Areoles 1.6–2.9 ×1.2–2.0 mm, 2.0–4.0 mm apart, oval to circular,felt white and brownish. Spines opaque, lightbrown at first, later greyish, covered by tri-chomes visible with lens; central spines 1–2,3.0–12.0 × 0.3–0.8 mm at base, ascending anddescending; radial spines 28–35, 2.0–8.5 ×0.25–0.4 mm at base. Flowers (6.5–)8.0–11.5 ×6.0–8.0 cm; pericarpel 12.0–14.0 × 6.0–10.0 mm;tube 5.0–7.0 × 0.8–1.1 cm at base, wideningtowards apex to 1.8–2.25 cm, curved to markedlycurved, reddish green, bearing short white tri-chomes emerging from the bract scale axils; outerperianth-segments 10, 6.0–24.0 × 3.0–5.0 mm,reddish green; inner perianth-segments 17,13.0–28.0 × 5.0–7.0 mm, white; nectar-chamber18.0–26.0 × 3.0–4.0 mm, tubular, curved; anthers2.5–3.0 × 0.4–0.8 mm; style 38.0–60.0 × 0.8–0.9mm; stigma-lobes 9–11, 3.5 mm; ovary locule 4.0× 8.0 mm, circular to elliptic in longitudinal sec-tion. Fruit 2.2–2.6 × 1.3–1.8 cm, spherical toovoid, funicular pulp translucent pink. Seedsbroadly ovate, medium-sized, 1.63 × 1.15 × 0.9mm, semi-matt; relief (SEM) convex; microreliefwith strong cuticular striations; hilum large, 0.83mm, oblique, forming an angle of 40–45° withlong axis of seed (Figure 10; Plates 5.4–5.6,16.1–16.3).
Examined material: PERU. LIMA: Halfwaybetween Chancay and Huacho, along thePanamericana norte highway, July 1956, RitterFR126e (U holo); Huaral, km 118 ofPanamericana norte highway, 77°27’12.5’’W,11°19’46’’S, 376 m, 16 Feb. 2004, N. Calderón364, 365, 369, 371 (Herb. B. G. La Molina); loc.cit., 4 March 2005, N. Calderón 419, 420 (Herb.B. G. La Molina); loc. cit., 19 Sept. 2001, N.Calderón 138 (Herb. B. G. La Molina). CHILE:Pisagua, Junín, 1954, Ritter FR126a (U holo).

Bradleya 25/200798
Plate 15. 15.1 H. repens. N. Calderón s.n, South of Trujillo, 2003. 15.2 Ibid., N. Calderón 358. 15.3 Ibid., N.Calderón 358, in flower.
2 3
1

Bradleya 25/2007 99
Habitat and distribution: Found on the Pacificcoastal desert: in Peru it is located in northernLima 274–380 m, and in Chile is reported forJunín, Pisagua (Map 5).
Observations of living plants of H. tenuis inPeru are based on populations found adjacent tothe Panamericana norte highway km 118, beinghighly probably the same location where Rittercollected the type. This area is a sandy expanse,where the soil is composed of sand and mollusc-shell fragments that receive the constant seabreeze. Another small group of H. tenuis is found1 km to the east of this location, on a small rockyhill.
This species may occur far to the south, on thecoast at Junín (Chile), according to Ritter’s col-lection 126a (U) that he described as H. australisf. subtilispinus (Ritter, 1980). This collection con-stitutes the first record of H. tenuis in Chile.
Phenology: Flowers: January and February;Fruits: February and March.
Conservation status: In Peru, CriticallyEndangered: CR[B2ab(ii,iii,v)]. The area of occu-pancy is estimated to be 3 km2 and populationsize to be of 252 individuals (Ceroni, unpubl.).This species is known to exist only at a singlelocation where continuing decline was observedin the area of occupancy, the quality of habitatand the number of mature individuals.
The most evident threat for this species is theproximity to the Panamericana norte highwaythat opens up a path where the wind blows plas-tic and paper residue (bottles, bags, etc.) into thedesert and even feathers from nearby chickenfarms, covering the stems of H. tenuis. Anotherrisk is the further urban expansion along thebusy highway. Currently, this species is beingsuccessfully conserved at La Molina UniversityBotanic Garden as result of ex situ conservationactivities that this institution undertakes.
The current status of this species in Chile isunknown and further fieldwork is necessary toclarify if this species still exists in that country.
Comments: H. tenuis was collected by Ritter in1956 during one of his various expeditions to thePeruvian south coast, although the formal publi-cation of the species only occurred 25 years later(Ritter, 1981). Rauh & Ostolaza (1990) publisheda more complete diagnosis with reproductivematerial in 1990, including photographs of spinesurfaces showing the presence of trichomes onthe spines, a key character that this species
shares with only H. lanugispinus.H. australis f. subtilispinus Ritter (1980) was
described from a locality on the coast at Junín(Chile). Study of the type and SEM of the seedsshowed that H. australis f. subtilispinus has tri-chomes on the spines and seeds with microreliefstrongly striated, in the same manner as H.tenuis. Since these characters are important fordiagnosis, H. australis f. subtilispinus is consid-ered to be a synonym of H. tenuis.
H. tenuis is characterized by prostrate indi-viduals making very conspicuous groups expand-ing on the ground up to 2 m2 and having tri-chomes (visible with lens) on the spines, this lat-ter character being hypothesized as an autoapo-morphy, probably of high adaptative value. Itsdecumbent habit, the curved flower-tube andseed microrelief with strong cuticular striationsare considered as probable apomorphic charac-ters. The pink funicular pulp of the fruit is hereconsidered as a probable secondary apomorphy.
The species epithet refers to its slender stems(1.4–3.4 cm diam.). In the field, these stems arefound partially covered by sand and mollusc shellfragments and, in some cases, by living snailcolonies that do not seem to cause damage to thecacti.
In Peru, H. tenuis lives in isolation from otherplants, but its typical flat, sandy habitat can bealso inhabited by seasonal herbs such asStenomesson coccineum (Amaryllidaceae). Whengrowing near rocky slopes, H. tenuis lives in closeproximity with H. pseudomelanostele (Werderm.& Backeb.) Backeb. and Cleistocactus acanthurus(Vaupel) Hunt.
9. Haageocereus versicolor (Werderm. &Backeb.) Backeb. in Backeb. & F. M. Knuth,Kaktus-ABC: 209 (1936); Backeb., Blätt. Kakt.-forsch. 1936(4): unpaged (1936); Krainz, DieKakteen 1(11): CVa, unpaged (1963); Rauh, Beitr.Kenntn. Peruan. Kakteenveg.: 399 (1958);Backeb., Die Cact. 2: 1188 (1960). Cereus versi-color Werderm. & Backeb. in Backeb., NeueKakteen: 81 (1931); Werderm., Fedde Rep. Spec.Nov. 30: 62 (1932). Type locality: Peru, Piura, byMorropón (believed not to have been preserved).Neotype (designated here): Peru, Piura,Morropón, 80°1’6.5’’W 5°12’57.4’’S, 129 m, 28May 2005, N. Calderón 465 (MOL neo!).H. icosagonoides Rauh & Backeb. in Backeb.
Descr. Cact. Nov. [1]: 23 (1956, publ. 1957);ibid., Die Cact. 2: 1186 (1960). Type: Peru,Lambayeque, Saña valley, 500 m, 1956, RauhK86 (HEID, believed not to have been pre-

Bradleya 25/2007100
served). Neotype (designated here): Backeb.,Die Cact. 2: 1215, fig. 1174 (1960). Synon.nov.
H. icosagonoides f. heteracanthus Ritter, Kakt.Südamer. 4: 1395 (1981). Type: Peru,Lambayeque, Saña valley, 500 m, RitterFR169a (U holo!). Synon. nov.
H. versicolor var. catacanthus Rauh & Backeb. inBackeb. Descr. Cact. Nov. [1]: 23 (1956, publ.1957). Type: Northern Peru, close toCanchaque, 100 m, 1956, Rauh K71a (HEID,believed not to have been preserved). Neotype(designated here): Rauh, Beitr. Kenntn.Peruan. Kakteenveg.: 400, fig. 184-II (1958).Synon. nov.
H. versicolor var. xanthacanthus (Werderm. &Backeb.) Backeb. in Backeb. & F. M. Knuth,Kaktus-ABC: 210 (1936). Cereus versicolorvar. xanthacanthus Werderm. & Backeb. inBackeb. Neue Kakteen: 81 (1931). H. versicolorvar. xanthacanthus Werderm. & Backeb. inBackeb. Blätt. Kakteenforsch. 1936-4: 3 (1936).Type locality: Peru, Piura, close to Serrán(believed not to have been preserved). Neotype(designated here): Backeb., Die Cact. 2: 1197,fig. 1153 (1960). Synon. nov.
The following may be referable here:H. versicolor var. humifusus (Werderm. &
Backeb.) Backeb. in Rauh, Beitr. Kenntn.Peruan. Kakteenveg.: 401 (1958); H. humifusus(Werderm. & Backeb.) Backeb. in Backeb. & F.M. Knuth, Kaktus ABC: 208 (1935). Cereusversicolor var. humifusus Werderm. & Backeb.in Backeb., Neue Kakteen: 81 (1931). Typelocality: Peru, Piura, close to Canchaque, 1500m (believed not to have been preserved).
H. versicolor var. lasiacanthus (Werderm. &Backeb.) Backeb. in Backeb., & F. M. Knuth,Kaktus-ABC: 210 (1935, publ. 1936). Cereusversicolor var. lasiacanthus Werderm. &Backeb. in Backeb. Neue Kakteen: 81 (1931).Type locality: Peru, Piura, close toCarrasquillo (believed not to have been pre-served).
H. versicolor var. aureispinus Backeb. in Backeb.& F. M. Knuth, Kaktus-ABC: 210 (1935, publ.1936); Cact. Succ. J. (US) 23: 47 (1951). Typelocality: Peru, Piura, close to Despoblado, coastof Sechura desert (believed not to have beenpreserved).
H. versicolor var. fuscus Backeb., Fedde Rep. Spec.Nov., 51: 62 (1942). Type locality: Peru, Piura,Salitrales-Talara (believed not to have beenpreserved).Habit erect, branching at base, up to 1.7(–2.4)
m tall; branches 2.0–8.0 cm diam., terete, epider-mis green; ribs 16–18 (–22), straight. Areoles ovalto circular, felt reddish at first, later white andyellow. Spines opaque, reddish at first, later yel-low and greyish; central spines 0–2, 10.0–25.0 ×0.5–0.9 mm at base, ascending and descending;radial spines 25–41, 4.0–10.0 × 0.25–10.0 mm atbase. Flowering areoles of mature branches wool-ly, white, generally disposed in ring-like pseudo-cephalia around the stems, persistent. Flowers5.0–9.0 cm, tube slightly curved, green, bearinglong curly white trichomes and short hair-spinesemerging from the bract scale axils; outer peri-anth-segments reddish green; inner perianth-segments white. Fruit 2.0–3.1 × 2.6–3.4 cm,spherical, pericarp greenish red. Seeds broadlyovate, small to medium sized, glossy; relief con-vex; microrelief non-striated to strongly striated;hilum large (SEM).
Habitat and distribution: Found in the seasonal-ly dry forest and valleys in northern Peru,120–1670 m. This is the northern limit for thegenus (Map 5).
Conservation status: Vulnerable: VU[B1ab(ii,iii)].The extent of occurrence is 8062 km2; it is knownto exist at no more than seven localities and con-tinuing decline has been observed in the area ofoccupancy and in the quality of habitats.
Comments: The species was first described asCereus versicolor by Werdermann & Backeberg(1931) but its type was not preserved. Later,Backeberg (1936) combined the species underHaageocereus, including a photograph in his pub-lication. Nevertheless, it is not clear whether thephotograph presented in 1936, also appearing inBackeberg’s work of 1960, actually belongs to thetypical species or to H. versicolor var. xanthacan-thus. Because of this possible confusion, a newspecimen (N. Calderón 465, MOL) has been cho-sen as a neotype.
H. icosagonoides was published by Rauh &Backeberg (1957) and because its type was notpreserved, a neotype is here proposed, based onthe illustration published later by Backeberg(1960). H. icosagonoides is distinguished by thelack of a well-defined central spine. From thesame locality of H. icosagonoides, Ritter (1981)published H. icosagonoides f. heteracanthusreferring to plants with well defined centralspines. It became obvious through field studythat this character is widely variable within apopulation and hence is insufficient to justify

Bradleya 25/2007 101
A
B
C
D
E
G H I J
K L
M
N
F
Figure 9. H. repens (A–N, N. Calderón 358); A, habit (scale = 20cm); B, stem (scale = 3cm); C, transverse sectionof stem (scale = 2cm); D–E, areoles (scale = 1cm); F, flower (2cm); G, bract scale (scale = 5mm); H, outer peri-anth segment (scale = 5mm); I–J, inner perianth segment (scale = 5mm); K, anther (scale = 1mm); L, stigma lobes(scale = 5mm); M–N, inmature fruits, side view and longitudinal section (scale = 1.5cm).

Bradleya 25/2007102
A
B
C
D
E
F G
H
IJ
K
L M
N
O
P
Q
Figure 10. H. tenuis (A–C, N–Q, N. Calderón 419; D–M, N. Calderón 364); A, habit (scale = 10cm); B, stem (scale= 2cm); C, transverse section of stem (scale = 3cm); D, areole (5mm); E, spine (scale = 5mm); F–H, flower, sideview and longitudinal section (scale = 2cm); I, bract-scale (scale = 5mm); J, outer perianth segment (scale = 5mm);K, inner perianth segment (scale = 5mm); L, anther (scale = 1mm); M, stigma lobes (scale = 5mm); N–Q, fruits,side view, top view, and longitudinal section (scale = 2cm).

Bradleya 25/2007 103
A
B
C
DE
F
G
H
I
J
KL
M
N
O
P
Figure 11. H. versicolor subsp. versicolor (A–B, N. Calderón 465; C–G, N. Calderón 467; H–P, N. Calderón 468);A, habit (scale = 50cm); B–C, stems (scale = 3cm); D–E, transverse sections of stem (scale = 2cm); F–G, areoles(scale = 2cm); H–J, flower, side view and longitudinal section (scale = 2cm); K–L, bract-scales (scale = 5mm); M,outer perianth segment (scale = 5mm); N, inner perianth segment (scale = 5mm); O, anther (scale = 2mm); P, stig-ma lobes (scale = 5mm).

Bradleya 25/2007104
Plate 16. 16.1 Habitat of H. tenuis, Huaral, north of Lima, 2005. 16.2 Ibid., H. tenuis. N. Calderón s.n., in fruit.16.3 Ibid., N. Calderón 364, in flower.
2 3
1

Bradleya 25/2007 105
recognition of taxa. Thus, H. icosagonoides andH. icosagonoides f. heteracanthus are treated assynonyms of H. versicolor.
H. versicolor var. catacanthus Rauh &Backebeberg (1957) was described for northernPeru, near Canchaque, and because its type wasnot preserved, an illustration published later byRauh (1958) is here designated as the neotype.The supposed difference from H. versicolor var.versicolor was its central spines being somewhatreddish, straight and descending. The same char-acter was also found in nearby populations of H.versicolor in the Piura valley and were notdeemed sufficient to justify separate taxonomicstatus; therefore, is it treated as a synonym.
H. versicolor var. xanthacanthus Werderm. &Backeb. (1936) was distinguished from the typi-cal variety by presenting few ribs (10–14) and1–2 “stout” central spines, and because the typematerial was not preserved, an illustration pub-lished later by Backeberg (1960) is here desig-nated as the neotype. Backeberg indicated thatH. versicolor var. xanthacanthus grows inDespoblado, Cajamarquilla valley and close toChilete at 500 m. According to recent field obser-vations in the Piura valley and close to Chilete,variation in the length of central spines exists (to40.0 mm long), and, although individuals withsuch a small number of ribs were not found, laterillustrations of this taxon published byBackeberg (1960) do not show individuals withless than 15 ribs. For this reason, C. versicolorvar. xanthacanthus is considered a synonym of H. versicolor.
Further names relating to H. versicolor, suchas H. versicolor var. humifusus (Werderm. &Backeb.) Backeb., H. versicolor var. lasiacanthus(Werderm. & Backeb.) Backeb., H. versicolor var.aureispinus Backeb., and H. versicolor var. fuscusBackeb., have no type material preserved andwere not illustrated. Nevertheless, the descrip-tions of these taxa show slight morphological dif-ferences in the colour, size and direction of thespines in comparison with H. versicolor. Forthese reasons, is probable that H. versicolor var.humifusus, H. versicolor var. lasiacanthus, H.
versicolor var. aureispinus and H. versicolor var.fuscus Backeb. are merely synonyms of H. versi-color, but in the absence of any authentic material to designate as neotypes, these namesremain as possible synonyms of H. versicolor.
H. versicolor shows “seasonal stem develop-ment” in its branches, where the new reddishareoles and spines contrast with the rest of thestem that has yellow spines. Its specific epithetrefers to this phenomenon. A distinctive andprobably apomorphic character of this species isthe ring-like pseudocephalium composed of thewoolly flower-bearing areoles observed in matureindividuals.
H. versicolor is here subdivided in two sub-species: H. versicolor subsp. versicolor and H. ver-sicolor subsp. pseudoversicolor.
9a. Haageocereus versicolor subsp. versicolorHabit up to 2.35 m tall; branches 2.0–3.5
(–5.0) cm diam.; ribs 16–18(–22), 2.2–8.8 mm,straight. Areoles 2.3–4.2 × 2.4–4.6 mm, 2.4–5.2mm apart. Central spines 0–2, 12.0–24.0(–40.0)mm; radial spines 4.0–9.0 mm. Flowers 5.8–6.8 ×3.1–5.0 cm; pericarpel 10.0–11.0 × 9.0–11.0 mm;tube 3.4–4.0 × 0.9–1.0 cm at base, wideningtowards apex to 1.8–2.0 cm diam., slightlycurved, green; outer perianth-segments 13–17,12.0–17.7 × 3.8–5.0 mm; inner perianth-seg-ments 14–22, 17.0–18.3 × 5.7–7.5 mm; nectar-chamber 15.0–19.0 × 5.0–7.1 mm, tubular;anthers 2.9–4.6 × 2.0 mm; style 34.7–40.0 ×1.1–2.5 mm; stigma-lobes 10–11, 2.3–5.9 mm;ovary locule 6.5–7.0 × 5.6–6.4 mm, circular toelliptic in longitudinal section. Fruit (only imma-ture fruit seen) 2.9–3.1 × 2.6–3.4 cm, spherical,pericarp green. Seeds broadly ovate, small, 1.09mm × 0.92 × 0.73 mm, glossy; relief (SEM) con-vex; microrelief with strong cuticular striations;hilum large, 0.68 mm, oblique, forming an angleof 65° with long axis of seed (Figure 11, Plates6.1–6.3, 17.1–17.3, 17.5).
Examined material: PERU. PIURA: Morropón,80°1’6.5’’W, 5°12’57.4’’S, 129 m, 28 May 2005, N.Calderón 465 (MOL, Neotype); ibid., N. Calderón
Key to subspecies of Haageocereus versicolorBranches slender, 2.0–3.5(–5.0) cm diam.; seeds small, microrelief strongly-striated (SEM) (Piura,
Lambayeque and La Libertad, 119–1670 m; Cajamarca: Chilete, 900 m)......................................................................................................................................................................9a. H. versicolor subsp. versicolor
Branches thick, 4.0–8.0 cm diam.; seeds medium sized, microrelief non-striated to weakly-striated (SEM)(Cajamarca: Jequetepeque valley, 250–620 m)................................ 9b. H. versicolor subsp. pseudoversicolor

Bradleya 25/2007106
460.1, 462, 463, 464.1, 466, 467.1, 468 (Herb. B.G. La Molina); ibid., Piura valley, road towardsHuancabamba, 79°47’19.3’’W, 5°25’10.7’’S, 180 m,7 Nov. 2003, N. Calderón 347 (Herb. B. G. LaMolina); LAMBAYEQUE: East of Olmos, 380 m,6 Nov. 2003, N. Calderón 342 (Herb. B. G. LaMolina); CAJAMARCA: road towards Chilete,78°50’10.8’’W, 7°13’38.4’’S, 950 m, 17 Nov. 2003,N. Calderón 351 (Herb. B. G. La Molina); LA LIB-ERTAD: Contumaza–Cascas road, 78°47’27’’W,7°26’10.8’’S, 1670 m, 18 Nov. 2003, N. Calderón355 (Herb. B. G. La Molina). LAMBAYEQUE:Saña valley, 500 m, Ritter FR169a (U holo).
Habitat and distribution: Found in the seasonal-ly dry forest and valleys in northern Peru,120–1670m. This is the northern limit for thegenus (Map 5).
Phenology: Flowers: November and May; Fruits:December and June.
Conservation status: Endangered EN[B1ab(ii,iii)]. The extent of occurrence is estimated tobe 3981 km2. It is known to exist at no more thansix localities and continuing decline observed inthe area of occupancy and the quality of habitats.
The most evident threat for this species is theloss of habitat due to the construction of newroads and the subsequent transit activities. Goatshepherding has also been observed, represent-ing a threat for this subspecies, as its branchesare easily eaten by these animals.
Comments: This subspecies lives together withArmatocereus cartwrightianus (Britton & Rose)Backeb., Neoraimondia arequipensis (Meyen)Backeb. and other deciduous plants typical of theseasonally dry forest, in a landscape dominatedby Prosopis sp. (Leguminosae) that may also actas nurse plants for the early stages of develop-ment of the cacti.
9b. Haageocereus versico lor subsp.pseudoversico lor (Rauh & Backeb.) N.Calderón comb. et stat. nov. H. pseudoversicolor Rauh & Backeb. in Backeb.
Descr. Cact. Nov. [1]: 23 (1956, publ. 1957);Rauh, Beitr. Kenntn. Peruan. Kakteenveg.: 401(1958); Backeb., Die Cact. 2: 1194 (1960). Type:Peru, Lambayeque, Saña valley, 100–200 m,1956, Rauh K85 (HEID, believed not to havebeen preserved). Neotype (designated here):Backeb., Die Cact. 2: 1199, fig. 1155 (1960).
Habit up to 1.7 m tall; branches 4.0–8.0 cmdiam.; ribs 18–19. Central spines 1–2,12.3–30.0 mm; radial spines 10.0 mm.Flowers 5.0 cm long.; inner perianth-seg-ments 15.0 × 6.0 mm; nectar-chamber12.0mm; ovary locule 10.0 × 4.0 mm. Fruit2.0–3.0 cm. Seeds broadly oval, medium-sized, 1.45 × 1.04 × 0.75 mm, glossy; reliefconvex; microrelief non-striated to weakly-striated; hilum large, 0.75 mm, oblique, form-ing an angle of 48° with long axis of seed(SEM) (Plates 6.4–6.6, 17.4).
Examined material: PERU. CAJAMARCA:Jequetepeque valley, 79°15’19.3’’W, 7°16’1.36’’S,250 m, 6 Nov. 2003, N. Calderón 340 (Herb. B. G.La Molina); ibid., Gallito Ciego (Irrigation pro-ject), 78°57’49.2’’W, 7°14’1.10.8’’S, 620 m, G.Charles 256.02 (seeds, private collection).
Habitat and distribution: Found in Jequetepequevalley at Cajamarca in northern Peru, 250–620 m (Map 5).
Phenology: Flowers: November; Fruits:December.
Conservation status: Endangered: EN[B2ab(iii)].The area of occupancy is estimated to be 22 km2
and is known to exist at no more than two locali-ties and continuing to decline in the quality of itshabitat.
Comments: H. versicolor subsp. pseudoversicolorwas first described by Rauh & Backeberg (1957)based on Rauh’s collection K85, 1956, but thismaterial was not preserved (or has been subse-quently lost). Therefore, an illustration publishedlater by Backeberg (1960) is here designated as aneotype for this subspecies.
The erect growing pattern, 18–19 ribs, centralspines 1–2, 12.3–30.0 mm long, conspicuous flow-ering areoles and white flower are all within thevariation accepted for the species, but, on theother hand, a few characters, like the stem diam-eter (to 8.0 cm) and the seed micromorphology(microrelief not striated to weakly striated) dis-tinguish this taxon from the typical subspecies.
Occurring geographically close to the typelocality of H. versicolor subsp. versicolor, thistaxon is not sympatric with other Haageocereusspecies. This subspecies lives in association withNeoraimondia arequipensis (Meyen) Backeb.,Melocactus peruvianus Vaupel and other plantssuch as Prosopis sp. (Leguminosae).

Bradleya 25/2007 107
PhytogeographyThe phytogeographic knowledge of Haageocereusis based on the endemism, species richness anddistribution patterns related to habitat condi-tions.
Haageocereus species are distributed on thePacific coast and the western slopes of the Andes,being mostly endemic to Peru, with two speciesalso occurring in northern Chile. The middleAndean region of northern Chile, Bolivia andPeru, has been proposed as the probable centre oforigin for the Cactaceae because a number of ple-siomorphic basal groups are endemic to this geo-graphic area (Wallace, 2002). The most recentgene-based phylogenetic analysis suggests thatat least the subfamilies Opuntioideae andCactoideae [plus Maihuenioideae] could havethis centre of origin, but the situation for thePereskioideae is more complex and its origin isapparently partly centred on the Caribbean(Edwards et al., 2005).
In terms of species richness, Haageocereushas diversified mostly in central Peru, where H.acranthus, H. lanugispinus, H. pseudomelanos-tele and H. tenuis are found. Occurring in thenorth are H. repens and H. versicolor and in thesouth, H. decumbens and H. platinospinus. Casesof sympatry are found in central Peru, involvingH. acranthus and H. pseudomelanostele andbetween H. pseudomelanostele subsp. pseudome-lanostele and H. tenuis.
Recent classifications of Peruvian vegetationand life zones have been published (ONERN,1995; INRENA, 1995; Brack et al., 2000).However, these classifications, although usefulfor some taxonomic groups, are not entirely accu-rate for describing cactus habitats because theyare not subdivided and, for the purposes of thisstudy, it is preferable to start with a broader con-cept of Peruvian natural regions. The three mainPeruvian regions are the coast, the Andes andthe tropical forest and Haageocereus is restrictedto the coast and the western Andes.
To describe Haageocereus habitats, terminolo-gy was taken and slightly modified fromWeberbauer (1945) and Pennington et al. (2004).Among the vegetation formations recognised byWeberbauer, Haageocereus occurs in “coastal ter-ritories of deserts and lomas”, “desert formationin the western Andes of southern Peru”, “colum-nar cacti formation in the western Andes ofsouthern Peru”, “desert formation of the westernbasins in central Peru”, “columnar cacti forma-tion of the western basins in central Peru”, and“western basins and inter-Andean valleys of
northern Peru”. Weberbauer (1945) also refers toa “piso del bosque pluviifolio” that corresponds towhat Pennington et al. (2004) term “seasonallydry forest”. From these definitions, five types ofhabitat are recognised as follows:
Along the Pacific coast, Haageocereus is foundbetween 50–900 m alt. in the following habitats:(1) The Xerophytic Deserts (XD), including arid
areas of sandy slopes and rocky hills close to thesea, where ephemeral herbs (Lomas vegetation)or xerophytic plants such as Tillandsia spp.(Bromeliaceae) may be present (Plates 15.1,16.1).(2) The Xerophytic Coastal Valleys (XCV) that isa transitional area between the desert and theAndes and includes rocky hillsides of the lowercourses of rivers draining to the Pacific, wherewoody shrubs such as Trixis cacaliodes(Asteraceae) are rather common as well as thexerophytic Tillandsia spp. and other cacti, suchas Mila caespitosa and Cleistocactus acanthurus(Plates 7.1, 12.1, 12.2).
In northern Peru, Haageocereus is found fromthe coastal plains, at around 120 m, to the Andes,at about 1800 m: (3) The Seasonally Dry Forest (DF), essentially awoody vegetation, consisting of mostly deciduoustrees and shrubs with or without a closed canopyand lacking a continuous grass layer, other cactibeing present such as Neoraimondia arequipen-sis subsp. gigantea and Armatocereus carwright-ianus. Vegetation with Prosopis spp. (Legum -inosae) is typical, as well as other trees suchBougainvillea spp. (Nyctaginaceae), Ceiba spp.(Bombacaceae), Jacquinia spp. (Theophr -astaceae), Loxopterygium (Ana cardiaceae) andother genera of Leguminosae and Bignoniaceae(Plate 17.1).
In the western Andes, Haageocereus is foundbetween 900–2800 m alt., forming part of the“Columnar cacti formation” (Weberbauer, 1945)in the following habitats:(4) The Xerophytic Formation of Central Andes(XCA) including steep mountains and rocky hills(“canyons”) close to the upper course of Pacificrivers, where xerophytic shrubs, such asCnidoscolus baciacanthus and Jatropha macran-tha (Euphorbiaceae) are typical, growing withherbs and sometimes small trees such as Schinusmolle (Anacardiaceae), Acacia macracantha(Leguminosae) and Carica mito (Caricaceae).Cactaceous communities are much more diversehere, with representatives of the generaArmatocereus, Austrocylindropuntia, Brown -ingia, Cleistocactus, Corryocactus, Espostoa,

Bradleya 25/2007108
Plate 17. 17.1 H. versicolor subsp. versicolor. N. Calderón s.n., Morropón, 2005. 17.2 Ibid., N. Calderón 465, inbud. 17.3 Ibid., in flower. 17.4 H. versicolor subsp. pseudoversicolor. N. Calderón 340, Jequetepeque valley. 17.5H. versicolor subsp. versicolor. N. Calderón 451, with immature fruits.
4
1
2
3
5

Bradleya 25/2007 109
Melocactus, Mila, Neoraimondia, Opuntia andWeberbauerocereus being present (Plate 13.1).(5) The Xerophytic Formation of Southern Andes(XSA), including rocky arid mountains andplateaux, where cactaceous communities ofBrown ingia, Corryocactus, Neoraimondia,Opuntioideae, Oreocereus and Weberbauero -cereus, predominate among the very reducedherbaceous vegetation and shrubs (Plate 11.1,14.1).
Taking into account their habitat spread, H.pseudomelanostele and H. acranthus have diver-sified with relatively more success than the othertaxa, being present in most of the habitats iden-tified for the genus (Table 2). In southern Peruthe genus is mostly represented by H. decum-bens, H. platinospinus and H. chilensis, whosehabitats are more closely related to those of H.acranthus and H. pseudomelanostele than tothose of the northern H. versicolor.
It is important to highlight the restricted dis-tribution of Haageocereus to the western slopes ofthe Andes, while other genera of tribeTrichocereeae such as Espostoa, Cleistocactus,Browningia and Weberbauerocereus that occa-sionally share the same habitat with variousspecies of Haageocereus are more expansive onthe eastern slopes of the Andes (including the so-called inter-Andean valleys). The formation ofthe Andean mountain chain has made possiblethe immense variety and complexity of vegeta-tion it comprises where plesiomorphic groups ofTrichocereeae occur to the east and west of theAndes, such as Rauhocereus (in Peru) andSamaipaticereus (in Bolivia). The latter genushas been taken as an example of a basal memberof Trichocereeae for comparison withHaageocereus, as described earlier. The presentdistribution of Haageocereus, west of the Andes,suggests that the diversification of Haageocereuscould have occurred after the formation of theAndes, contrasting to other more widespreadgenera, such as Espostoa, Cleistocactus,Browningia and Weberbauerocereus. Other exam-ples of genera restricted to the western side ofthe Andes are represented by Copiapoa in Chile,Mila in Peru and Eulychnia in Chile and Peru.
ConservationOverviewCacti, like many other plants, are seriouslythreatened by habitat destruction, whether forthe development of new agricultural land, forexpanding urban areas, or for other human activ-ities, such as road building and mining (Boyle &Anderson, 2002). In this context, Haageocereus isa good example of a seriously threatened genus,most of its localities being in the proximity ofgrowing cities and therefore, suffering constanthuman disturbance.
Conservation of cacti has been recognised asone of the urgent actions to be taken because ofthe ecological significance of this group that com-prises taxa of unique value, as judged by theirendemism, varied ecological adaptations andtheir relative importance as environmental com-ponents.
International efforts towards the conserva-tion of cactus species were established in 1984,when the Cactus and Succulent Specialist Groupof the Species Survival Commission (SSC), a partof IUCN, was created. In 1997 this SpecialistGroup published the Cactus and SucculentPlants Conservation Action Plan (Oldfield, 1997),including a compilation of information about suc-culent plant groups, conservation measures,regional accounts, and action proposals, includ-ing one for Peru. The proposal for Peru recom-mended Assessment of the in situ conservationrequirements of succulents, including: “assess-ment of the extent to which the protected areasystem of Peru protects the habitats of endemicsucculents, survey work being necessary to deter-mine the degree of threat to populations of par-ticular succulent species in the more mesicareas”.
Conservation measures listed by the Cactusand Succulent Specialist Group also includedinternational legislation (e.g. The Convention onBiological Diversity and The Berne Convention);trade controls by CITES authorities and in situand ex situ conservation actions.
While in situ and ex situ conservation mea-sures are considered to be of high importancewithin the present work, trade based on wild-col-lected plants is not one of the concerns forHaageocereus.
In 2002, at the Conference of the Parties tothe Convention on Biological Diversity (CBD),The Global Strategy for Plant Conservation waspresented. Targets of this Global Strategyinclude, for instance, that 60% of the world´sthreatened species be conserved in situ by 2010.

Therefore, in response to this aim, in situ conser-vation actions are also being presented in thisstudy.
Conservation assessmentsIn 1980, IUCN established criteria and cate-gories of threat for assessing extinction risks tospecies. The IUCN Red List criteria were pub-lished in 1994 and a revised version in 2001. Thislater version is being used in the present study toassess categories of threat for Haageocereus. Thecategories of threat are: Extinct (Ex), Extinct inthe Wild (EW), Critically Endangered (CR),Endangered (EN), Vulnerable (VU), NearThreatened (NT), Least Concern (LC), and DataDeficient (DD).
For listing as Critically Endangered,
Endangered or Vulnerable there is a range ofquantitative criteria; meeting any one of thesecriteria qualifies a taxon for listing at that levelof threat and each taxon should be evaluatedagainst as many of the criteria as practicable.
Haageocereus species have been evaluatedagainst the IUCN Red List criteria of “geograph-ic range” in the form of area of occupancy and/orextent of occurrence and, when appropriate,“reduction of subpopulations” of Haageocereus.The extent of occurrence and/or area of occupan-cy was stated in the treatment of each taxonabove. These estimates are based mainly on fieldstudies and herbarium records made during thelast five years.
Both bibliographic records and personalobservations in the field were fundamental for
Bradleya 25/2007110
Habitat Habitat Taxon Sympatric species
E / NE
Xerophytic E H. decumbens noneDeserts (XD) E H. repens none(XD) E H. tenuis H. pseudomelanostele subsp.
pseudomelanostele NE H. acranthus subsp. acranthus H. pseudomelanostele subsp.
(also in XCV) pseudomelanostele.NE H. pseudomelanostele subsp. H. acranthus subsp. acranthus or
pseudomelanostele (also in XCV) H. tenuis
Xerophytic NE H. acranthus subsp. acranthus H. pseudomelanostele subsp.Coastal (also in XD) pseudomelanosteleValleys NE H. pseudomelanostele subsp. H. acranthus subsp. acranthus(XCV) pseudomelanostele (also in XD)
Seasonally E H. versicolor subsp. versicolor noneDry Forest E H. versicolor subsp. none(DF) pseudoversicolor
Xerophytic E H. acranthus subsp. zonatus H. pseudomelanostele subsp.Formation, aureispinusCentral E H. acranthus subsp. backebergii H. pseudomelanostele subsp.Andes aureispinus or H.(XCA) pseudomelanostele subsp.
carminiflorusE H. pseudomelanostele subsp. H. acranthus subsp. zonatus or
aureispinus H. acranthus subsp. backebergiiE H. pseudomelanostele subsp. H. acranthus subsp. backebergii
carminiflorus
Xerophytic E H. pseudomelanostele subsp. noneFormation, turbidusSouthern E H. platinospinus noneAndes (XSA)
Table 2. Habitat types for Haageocereus including habitat preference codes: E = exclusive habitat forthe taxon; NE = non-exclusive habitat for the taxon.

Bradleya 25/2007 111
estimating the original distribution and thereduction of subpopulations for the species H.acranthus and H. pseudomelanostele. In general,Haageocereus localities are outside Peruviannational protected areas (Map 6) and recommen-dations towards the conservation of their habi-tats are being presented.
For estimating the area of occupancy at theonly known locality of H. tenuis, the results of arecent field survey (Ceroni, pers. comm., 2004)were incorporated. In the case of H. repens thatalso has only one known population, the area ofoccupancy was inferred from direct observation.For the other species of Haageocereus occurringin Peru that have more than one populationknown, their geographic range was estimated atthe GIS Unit (RBG, Kew) where the area of occu-pancy and extent of occurrence was calculatedautomatically, using a specially designed soft-ware program. (Map 7)
Most Haageocereus species are categorized asCR, EN or VU (Table 3), for taxa existing atbetween one and ten localities and also facingcontinuing decline in the area of occupancy andthe quality of habitat. Importantly, the loss ofhabitat, especially in arid areas, is a continuous-ly increasing threat.
Conservation actions and recommendationsModified habitats of Haageocereus, such as thoseby the coast (Xerophytic Deserts and XerophyticCoastal Valleys) are extremely difficult to recoverbased on the first author´s observations in thepast 5 years, but some alternative strategies caninclude ex situ conservation in botanical gardensand possible reintroduction. In the case of H.tenuis that is found only 13 km from the “ReservaNacional de Lachay”, it could be proposed toextend the extent of this protected area to includethe present species.
The Seasonally Dry Forest has been receivingattention from researchers during the last fewyears (Centro de Datos para la Conservación-Univ La Molina, pers. comm.), towards the cre-ation of a protected area. In this context, includ-ing some of the geographic range of H. versicoloris strongly recommended, as this species onlygrows in this vegetation type.
The Xerophytic Formations of the CentralAndes and Southern Andes are potentially excel-lent areas to be considered for in situ conserva-tion and thus to be included in the national sys-tem of protected areas. These environmentsinclude 6 taxa (Table 2) of Haageocereus andother important cacti. There are several reasons
to recommend this initiative:• There is no existing protected area coveringthese environments.• These formations have some relativelyuntouched land not yet under direct humaninfluence (e.g. farming, mining or other kind ofexploitation of the resources in these habitats).• These environments are included in the StatusSurvey and Conservation Action Plan-IUCN/SSC Cactus and Succulent SpecialistGroup (Oldfield, 1997) within the Subdivisions Iand II of the Andean Region of Peru (500–3500 malt.) for being extremely rich in succulent plantspecies and with a high level of endemism.• The conservation of part of these environmentswill enhance target vii of the Global Strategy forPlant Conservation (CBD, 2002) for conservingplant diversity, i.e. that 60 per cent of the world’sthreatened species be conserved in situ.
According to field observations in the last fiveyears, as well as ex situ conservation effortsundertaken at La Molina University BotanicGarden, specific actions and recommendationscan be listed (see Table 3).
ConclusionsHaageocereus has been regarded as a very com-plex genus in need of particular attention, and,although the present work recognizes only 9species and 6 heterotypic subspecies, there aremore than 100 names attributed to this genus.Most of these names reflect a very poor speciesconcept, based on very few and variable charac-ters. Type material is also poor, if not lacking.Extensive field work, morphological studies, aswell as the outcomes from recent studies in theCactaceae generally, have greatly contributed toa clarification of important biological, ecologicaland even possible evolutionary aspects of thegenus Haageocereus and its species.
The taxonomic and phytogeographic knowl-edge of Haageocereus is very important for mak-ing conservation assessments and the develop-ment of recommendations towards a coherentconservation action plan. In this context, it ispossible to present the following conclusions:(1) The delimitation of Haageocereus species inthis work led to the first interpretation of theirmorphological characters within the complextribe Trichocereeae. Morphological comparison ofHaageocereus with a basal member ofTrichocereeae, Samaipaticereus, represents thefirst attempt to elucidate the potential apomor-phies of this genus. (2) As in many Cactaceae, typification of

Bradleya 25/2007112
Map 6. Localities of Haageocereus = �, and national protected areas (in grey).
N
100 0 100 200 Kilometers
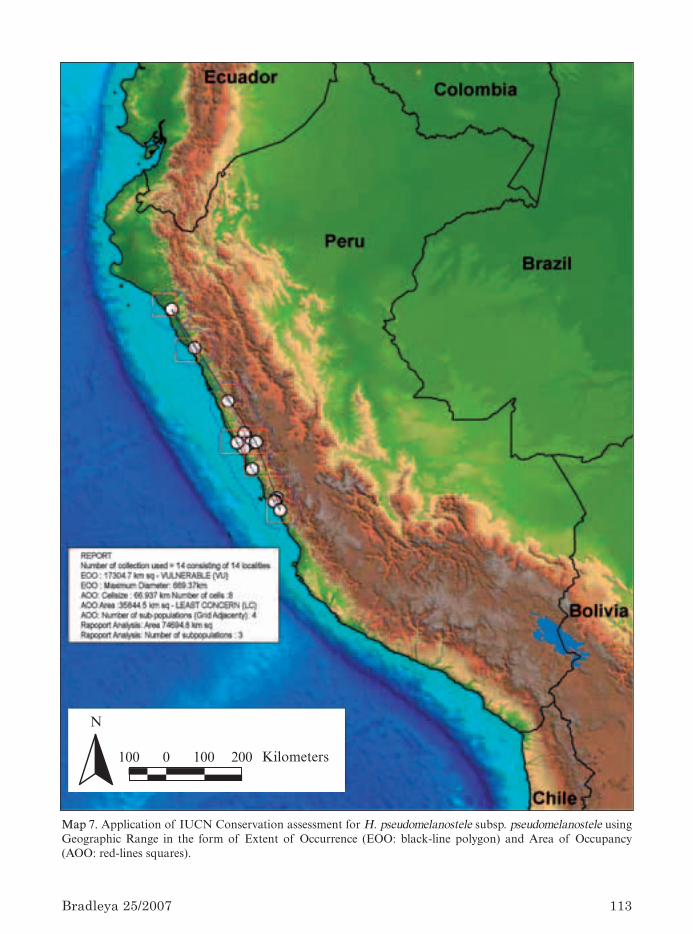
Bradleya 25/2007 113
Map 7. Application of IUCN Conservation assessment for H. pseudomelanostele subsp. pseudomelanostele usingGeographic Range in the form of Extent of Occurrence (EOO: black-line polygon) and Area of Occupancy(AOO: red-lines squares).
N
100 0 100 200 Kilometers

Bradleya 25/2007114
Haageocereus names, where types are unavail-able, is based mainly on original illustrations,published as part of the protologues or in laterpublications, except for H. versicolor, where aspecimen was designated as the neotype. Theherbarium records of Haageocereus speciesacquired for this work includes most of thePeruvian taxa recognised here (with the excepti-on of H. lanugispinus and H. chilensis), all heldat La Molina University (MOL).(3) The restricted distribution of Haageocereus tothe Pacific drainage of the Andes together withthe fact that other members of Trichocereeae,such as the allied genera Espostoa andCleistocactus, occur in inter-Andean valleys,leads to the hypothesis that Haageocereus couldhave originated after the formation of theAndean mountains and has been unable to radi-ate eastwards due to the altitude of the Andeanranges. In phytogeographic terms Haageocereuscould possibly be a “modern genus” within thetribe, although further research, especially agene-based phylogeny, is still needed to deter-mine its exact position in the tribe.(4) The current situation of the habitats ofHaageocereus is dramatic in terms of conservati-on, with all taxa listed under IUCN categories ofthreat. A list of recommendations is presented, asa prerequisite to the future development of moredetailed proposals to the appropriate Peruvianauthorities. Habitats of Haageocereus, especiallyin the north (Piura) and south (Arequipa) of Peruare particularly suitable for the creation of pro-tected areas, because of the biodiversity well pre-served in these beautiful landscapes.Universities are particularly encouraged to takepart in ex situ conservation activities, throughthe incorporation of native and threatened spe-cies in their botanic gardens. (5) The present study presents essential informa-tion about the taxonomy, phytogeography andconservation of Haageocereus and, at the sametime, provides research tools for their identifica-tion and further information towards a bettercomprehension of other Peruvian cacti. Furtherresearch in associated areas like reproductivebiology, ecology and phylogenetics is especiallyimportant for a better understanding of bothHaageocereus and other the Peruvian cacti.
AcknowledgementsThe present study was based on the Master the-sis of the first author that was greatly supportedby:The Kew Latin American Research Fellowships
(KLARF) Programme, specifically by the PranceFellowship in Neotropical Botany Award, and theBentham–Moxon Grants Programme, for fund-ing research at RBG Kew and other Europeaninstitutions.Dr Alan Paton (RBG Kew) and Dr Colin Clubbe(RBG Kew) for providing much help in the devel-opment of the present work. Dr Simon Owens(RBG Kew) and Dr Julie Hawkins (ReadingUniv.) for useful discussions and recommenda-tions.Dr Carlos Ostolaza (Peruvian Cactus &Succulent Society), Graham Charles (UK) andChris Pugh (UK) for providing much guidanceand support during field excursions.To the following companions when doing field-work in Peru: Viviana Castro, Pamela García,Milagros Sosa, Sidney Novoa, Michael Vega,Gustavo Dreyfus, Gisella Cruz, Fabiola Parra,Pamela Puppo, Antonio Saccsara, JohannaBustamante, Maura Jurado (all from La MolinaUniv., Lima), Mag. Fátima Cáceres (San AgustinUniv., Arequipa), Sonia Guillen, Gerardo Carpio(Centro Mallqui, Moquegua), José Pizarro (TacnaUniv., Tacna) and Mercedes García (RicardoPalma Univ., Lima).European botanists, who allowed visits to and orloans from herbaria in the institutions wherethey work and also gave valuable information onCactaceae: Dr Beat Leuenberger (B), Dr RobertBogt (B), Dr Urs Eggli (ZSS), Thomas Bolliger(ZSS), Dr Paul Maas (U), Dr Marion Janson-Jacobs (U), Dr Erik Simonis (U), Dr Roy Vickery(BM) and Dr Reto Nyffeler (Univ. ZurichHerbarium, Z).Dr Monica Shaffer-Fehre for translations fromGerman, Dr Hannah Banks for her help andtraining with use of the SEM, Dr Justin Moat forproducing distribution maps and help with con-servation assessments, Tania Durt and DrNicolas Hind for logistic support at all times (allfrom the Herbarium, RBG Kew).
ReferencesAGARDH, J. G. (1894). Analecta Algologica 2: 63.AGARDH, J. G. (1899). ibid. 5: 158.AKERS, J. (1947a). New genus and new species
from Peru. Cact. Succ. J. (US) 19 (5): 67–70.AKERS, J. (1947b). A new variety from Peru. ibid.
19 (6): 91.AKERS, J. (1947c). New species from Peru. ibid.
19 (7): 109–110.AKERS, J. (1947d). New species from Peru. ibid.
19 (8): 121–123.AKERS, J. (1947e). Newspecies from Peru. ibid. 19 (9): 143–144.

Bradleya 25/2007 115
AKERS, J. (1947f). New species from Peru. ibid. 19(10): 162–163.
AKERS, J. (1948a). New species from Peru. ibid.20 (4): 55–56.
AKERS, J. (1948b). A key to some of the Peruviangenera. ibid. 20 (9): 128–131.
AKERS, J. (1948c). New species from Peru. ibid.20 (10): 154–156.
AKERS, J. (1948d). New species from Peru. ibid.20 (12): 184–186.
AKERS, J. (1949). New species from Peru. ibid. 21(2): 45–46.
AKERS, J. (1950). From Peru – Peruvocereus mul-tangularis. ibid. 22 (6): 174–175.
ANDERSON, E. (2001). The Cactus Family. TimberPress, Portland, Oregon.
ANDERSON, M. & HEWITT, T. (2002). The Worldencyclopedia of Cacti & Succulents. HermesHouse Editors, New York.
APG (ANGIOSPERM PHYLOGENY GROUP). (1998). Anordinal classification for the families of flow-ering plants. Ann. Missouri Bot. Gard. 85:531–553.
Taxon IUCN Red List Proposed actions recommendedCategory [criteria]
H. acranthus VU[A4c] See below for subspecies.H. acranthus subsp. EN[B1ab(ii,iii,iv)] Less than 50 individuals are protected at the Reserva Nacional de acranthus Lachay (Lima). Enhance current ex situ conservation efforts and
establish ex situ gene banks to enable future re-introductions. Some ex situ conservation efforts have already been undertaken byLa Molina Univ.
H. acranthus subsp. EN[B1ab(ii,iii,iv)] Establish a protected area in the surroundings of Umarcata and backebergii Orovel hills in the Chillón valley (Lima). Conservation in other
localities in the Lurín valley is also recommended to maintain genetic diversity.
H. acranthus subsp. EN[B1ab(ii,iii)] Establish a protected area in the surroundings of Paccho in the zonatus Huaura valley (Lima). Conservation in other localities in the
Pativilca valley is recommended.H. decumbens VU[B1ab(ii,iii,iv)] Enhance current ex situ conservation and establish ex situ gene
banks to enable future reintroductions.H. platinospinus EN[B1ab(ii,iii)] Establish a protected area in the Yura valley (Arequipa).H. pseudomelanostele VU[A4c] See below for subspecies.H. pseudomelanostele VU[B1ab(ii,iii,iv)] Enhance current ex situ conservation. Some ex situ conservation subsp. efforts have been already undertaken by La Molina Univ. pseudomelanosteleH. pseudomelanostele EN[B1ab(ii,iii)] Establish a protected area in the surroundings of Paccho in the subsp. aureispinus Huaura valley (Lima) and also in the surroundings of Umarcata
and Orovel hills in the Chillón valley (Lima) to maintain genetic diversity.
H. pseudomelanostele EN[B1ab(ii,iii) Establish a protected area in the Tinajas canyon in the Lurinsubsp. carminiflorus + 2ab(ii,iii)] valley (Lima).
H. pseudomelanostele CR[B2ab(iii)] Establish a protected area in the Nazca valley. subsp. turbidusH. repens CR[B2ab(ii,iii)] Enhance current ex situ conservation. Some ex situ conservation
efforts have been undertaken by the Univ. of Trujillo. H. tenuis CR[B2ab(ii,iii,v)] Site recommended for being included at the Reserva Nacional de
Lachay (Lima). Some ex situ conservation efforts have been undertaken by La Molina Univ.
H. versicolor VU[B1ab(ii,iii)] See below for subspecies.H. versicolor subsp. EN[B1ab(ii,iii)] Establish a protected area in the surroundings of Morropónversicolor (Piura).H. versicolor subsp. EN[B2ab(iii)] Enhance current ex situ conservation activities.pseudoversicolor
Table 3. IUCN Red List categories for Haageocereus and proposed protected areas and recommendations.

APGII (ANGIOSPERM PHYLOGENY GROUP II).(2003). An update of the phylogeny groupclassification for the order and families offlowering plants. Bot. J. Linn. Soc. 141:399–436.
ARAKAKI, M. (2002). Systematics of the genusWeberbauerocereus Backbg. (Cactaceae).Master of Arts, University of Texas, unpub-lished.
BACKEBERG, C. (1931). Neue Kakteen. Trowitzsch& Sohn, Frankfurt.
BACKEBERG, C. (1932). Following in Dr. Rose’sFootsteps. Cact. Succ. J. (US) 3 (8): 130–132.
BACKEBERG, C. (1933a). Genus Binghamia. Cact.Succ. J. (US) 4 (12): 387–388.
BACKEBERG, C. (1933b). Kakteenfreund 2 (5): 54.BACKEBERG, C. (1934a). Haageocereus Bckbg. n.
g. Blätt. Kakt.-forsch. 1934 (6): unpaged.BACKEBERG, C. (1934b). Haageocereus decumbens
Bckbg. ibid. 1934 (6): unpaged.BACKEBERG, C. (1936a). Haageocereus pacalaen-
sis Bckbg. ibid 1936 (4): unpaged.BACKEBERG, C. (1936b). Haageocereus australis
Bckbg. n. sp. Jahrb. Deutsch Kakt-Ges. 1: 104.BACKEBERG, C. (1937). Haageocereus olowin-
skianus Bckbg. n. sp. Blätt. Kakt.-forsch. 1937(5): unpaged.
BACKEBERG, C. (1942). Neue Arten aus “StachligeWildnis”. Repert. Spec. Nov. 51: 61–65.
BACKEBERG, C. (1951). Some results of twentyyears of cactus research, Part 3. Cact. Succ. J.(US) 23 (2): 45–52.
BACKEBERG, C. (1957). Descr. Cact. Nov. [1]. Jena.BACKEBERG, C. (1958–1962). Die Cactaceae.
Handbuch der Kakteenkunde, Vols. 1–6, VEBGustav Fischer Verlag, Jena.
BACKEBERG, C. (1963). Descr. Cact. Nov. [3]. BACKEBERG, C. (1966). Das Kakteenlexicon:
Enumeratio diagnostica Cactacearum. VEBGustav Fischer Verlag, Jena.
BACKEBERG, C. (1977). Das Kakteenlexicon:Enumeratio diagnostica Cactacearum. G.Fischer Pub., Stuttgart.
BACKEBERG, C. & KNUTH, F. M. (1936). KaktusABC. Gyldendalske Boghandel-NordiskForlag, Copenhagen.
BARTHLOTT, W. & EHLER, N. (1977). Raster-Elektronenmikronenmikroskopie der epider-mis-oberflächen von spermatophyten. Trop.Subtrop. Pflanzen. 19: 367–467.
BARTHLOTT, W. & HUNT, D. R. (1993). Cactaceae.In: Kubitsky, K., The Families and Genera ofVascular Plants 2: 161–197. Springer Verlag,Berlin.
BARTHLOTT, W. & HUNT, D. R. (2000). Seed-diver-sity in the Cactaceae Subfam. Cactoideae.Succ. Pl. Res. 5: 1–173.
BARTHLOTT, W. & VOIT, G. (1979).Mikromorphologie der Samenschalen undTaxonomie der Cactaceae. Pl. Syst. Evol. 132:205–229.
BEHNKE, H. (1981). Sieve-element characters.Nord. J. Bot. 1: 381–400.
BERGER, A. (1905). A systematic revision of thegenus Cereus. Ann. Rep. Missouri Bot. Gard.16: 57–86.
BERGER, A. (1926). Die Entwicklungslinien derKakteen. Gustav Fischer Verlag, Jena.
BOKE, N. H. (1963). Anatomy and development ofthe flower and fruit of Pereskia pititache.Amer. J. Bot. 50: 843–858.
BOKE, N. H. (1964). The cactus gynoecium: A newinterpretation. Amer. J. Bot. 51: 598–610.
BOKE, N. H. (1966). Ontogeny and structure ofthe flower and fruit of Pereskia aculeata.Amer. J. Bot. 53: 534–542.
BOKE, N. H. (1968). Structure and development ofthe flower and fruit of Pereskia diaz-romeroana. Amer. J. Bot. 55: 1254–1260.
BOYLE, T. H. & ANDERSON, E. F. (2002).Biodiversity and Conservation. In: Nobel, P.S. Cacti: Biology and Uses. University ofCalifornia Press, Berkeley, pp. 125–141.
BRACK, A. & MENDIOLA, C. (2000). Ecología delPeru. PNUD, Ed. Bruño.
BRAKO, L. & ZARUCCHI, J. L. (1993). Catalogue ofthe Flowering Plants and Gymnosperms ofPeru. Missouri Botanical Garden, St. Louis,Missouri.
BRITTON, N. L. & ROSE, J. N. (1919–1923). TheCactaceae, Vols. 1–4. Carnegie Institution,Washington D.C.
BREGMAN, R. 1994. Borzicactinae: 9.Haageocereus maritimus. Succulenta. 73 (3):111–112.
BRUMMITT, R. K. & TAYLOR, N. P. (1990). To corrector not ot correct? Taxon 39: 298–306.
BUSTAMANTE, E. & BURQUEZ, A. (2005). Fenologíay biología reproductiva de las cactáceascolumnares. Cact. Suc. Mex. 50 (3): 68–88.
BUXBAUM, F. (1950–54). Morphology of Cacti. I–III. Abbey Garden Press, Pasadena.
BUXBAUM, F. (1958). The phylogenetic division ofthe subfamily Cereoideae, Cactaceae.Madroño 14 (6): 177–216.
BUXBAUM, F. (1962). Das Phylogenetische Systemder Cactaceae. In: Krainz, H. Die Kakteen1(8): System 1–6, unpaged.
Bradleya 25/2007116

Bradleya 25/2007 117
CÁCERES, A., GARCÍA, F., PONCE, E. & ANDRADE, R.(2000). “El Sancayo”, Corryocactus brevistylus(Schumam ex Vaupel) Britton & Rose. Quepo15: 37–42.
CALDERÓN, N. (2002). Diagnóstico poblacional delas cactáceas de Manchay, Prov. de Lima.Quepo 16: 40–45.
CALDERÓN, N. (2003). Estudio Taxonómico delgénero Haageocereus (Familia Cactaceae) enel departamento de Lima-Peru. Thesis for theaward of the Biologist degree. UniversidadNacional Agraria La Molina, Lima, unpublis-hed.
CALDERÓN, N. (2006). Haageocereus: Taxonomyfor the conservation of the genus in Peru. M.Phil. thesis, Open University, Milton Keynes,unpublished.
CALDERÓN, N., CERONI, A. & OSTOLAZA, C. (2004).Distribución y estado de conservación delgénero Haageocereus (Familia Cactaceae) enel departamento de Lima. Peru. EcologíaAplicada 3 (1–2): 17–22.
CÁRDENAS, M. (1952). New Bolivian cacti III.Cact. Succ. J. (US) 24 (5): 141–148.
CBD. (2002). Global Strategy for PlantConservation. The Secretariat of theConvention on Biological Diversity & BotanicGardens Conservation International, UK.
CROIZAT, L. (1942a). Is Binghamia valid? Cact.Succ. J. (US) 14 (9): 126–128.
CROIZAT, L. (1942b). Haageocereus Backeberg(Syn.: Binghamia Britton & Rose). Cact. Succ.J. (US) 14 (10–11): 145–148.
CRONQUIST, A. (1981). An integrated System ofClassification of Flowering Plants. ColumbiaUniversity Press, New York.
CULLMANN, W. (1957). The genera HaageocereusBackeberg und Peruvocereus Akers. Kakt.and. Sukk. 8 (12): 177–180.
CULLMANN, W. (1960). Haageocereus Backeb. –Peruvocereus Akers. Succulenta 39: 9–13.
CULLMANN, W. (1963). Die Gattung HaageocereusBackeb. Sukkulentenkunde VII/VIII: 17–31.
CULLMANN, W. (1973). Haageocereus Backeb.Stachelpost 9 (45): 61.
EDWARDS, E. J., NYFFELER, R. & DONOGHUE, J.(2005). Basal cactus phylogeny: Implicationsof Pereskia (Cactaceae) paraphyly for thetransition to the cactus life form. Amer. J. Bot.92 (7): 1177–1188.
EGGLI, U. (1985). Backeberg’s invalid cactusnames. Bradleya 3: 97–102.
EGGLI, U. (1993). Glossary of botanical terms withspecial reference to succulent plants includingGerman equivalents. British Cactus &
Succulent Society, Richmond, Surrey.EGGLI, U. (1995). Bibliography of succulent plant
periodicals. Fričiana 60: 1–144.EGGLI, U., MUNOZ SCHICK, M. & LEUENBERGER, B.
E. (1995). Cactaceae of South America: TheRitter Collections. Englera 16: 1–646.
EGGLI, U. & NEWTON, L. (2004). Etymological dic-tionary of succulent plant names. Springer-Verlag, Berlin.
EGGLI, U. & TAYLOR, N. (eds.) (1991). List ofCactaceae names from RepertoriumPlantarum Succulentarum (1950–1990).Royal Botanic Gardens, Kew, Richmond.
ENDLER J. & BUXBAUM, F. (1982). DiePflanzenfamilie der Kakteen. Lehrmeister-Bücherei 872. Albrecht Philler, Minden.
FAEGRI, K. & VAN DE PILJ, L. (1979). The princi-ples of pollination ecology. Pergamon Press,Oxford.
FARJON, A. & PAGE, C. N. (eds.) (1999). Conifers:Status Survey and Conservation Action Plan.IUCN/SSC Conifer Specialist Group, IUCNGland, Switzerland and Cambridge.
FLEMING, T. H. & VALIENTE-BANUET, A. (eds.)(2002). Columnar cacti and their mutualists:Evolution, Ecology and Conservation.University of Arizona Press, Phoenix.
FUNK, V. A. & BROOKS, D. R. (1990). Phylogeneticssystematics as the basis of comparative biolo-gy. Smithsonian contributions to Botany,Washington DC.
GIBSON, A. C. (1976). Vascular Organization inshoots of Cactaceae. 1. Development and mor-phology of primary vasculature inPereskioideae and Opuntioideae. Amer. J. Bot.63: 414–426.
HAWORTH, A. H. (1819). SupplementarumSucculentarum. J. Harding, London.
HELVERSON, O. V. (1993). Adaptations of flowersto the pollination by glossophagine bats. In:W. Barthlott et al. Plant-animal interactionsin tropical environments. Museum AlexanderKoenig, Bonn, pp. 41–59.
HERSCHKOVITZ, M. A. & ZIMMER, E. A. (1997). Onthe evolutionary origin of the cacti. Taxon 46:217–232.
HOFFMANN, A. E. (1989). Cactáceas en la flora sil-vestre de Chile. Ediciones de la FundaciónClaudio Gay, Santiago, Chile.
HOFFMANN, A. E. & WALTER, H. E. (2004).Cactáceas en la flora silvestre de Chile. 2ndEd. Ediciones de la Fundación Claudio Gay,Santiago, Chile.
HUNT, D. (1967). Cactaceae. In: Hutchinson, J.(ed.) The genera of flowering plants 2:

427–467. Oxford University Press, Oxford. HUNT, D. (1999). CITES Cactaceae Checklist. Ed.
2. Royal Botanic Gardens, Kew, Richmond. HUNT, D. (2002). Haageocereus. Cact. Syst. Init.
14: 17.HUNT, D. (ed.) (2004). Catalogue of nothogeneric
names. Cact. Syst. Init. 18: 22.HUNT, D. (2005). Haageocereus. Cact. Syst. Init.
20: 19.HUNT, D. & TAYLOR, N. P. (eds.) (1990). The gen-
era of Cactaceae: Towards a new consensus.Bradleya 8: 85–107.
HUNT, D., TAYLOR, N. & CHARLES, G. (2006). Thenew cactus lexicon: descriptions and illustrations of the cactus family. DH Books,Milborne Port.
IGM. (1995). Mapa Ecológico del Peru. Mapa edi-tado sobre el mapa físico político del Peru, 1:1 000 000.
ISAYAMA, V. (1988). Morfología polínica de algunasespecies de la flora del departamento deLambayeque. Zonas Aridas 5: 45–60.
INRENA. (1995). Guía explicativa del mapaforestal.
IUCN. (2001). Red List Categories and Criteria,version 3.1. Gland, Switzerland.
JEFFREY, C. (1976). Nomenclatura Biológica.Código Internacional de nomenclatura botá-nica, Código Internacional de nomenclaturazoológica. H. Blume, Madrid.
JUDD, W. S., et al. (2002). Plant Systematics: Aphylogenetic approach. Ed. 2. SinauerAssociates, USA.
KATTERMANN, F. (1994). Eriosyce (Cactaceae): Thegenus revised and amplified. Succ. Pl. Res. 1:1–176
KRAINZ, H. 1962a. Das Phylogenetische Systemder Cactaceae (by F. Buxbaum). Die Kakteen1(8): System 1–6, unpaged.
KRAINZ, H. (1962b). Haageocereus pacalaensis.Die Kakteen 1(12): CVa, unpaged.
KRAINZ, H. (1963). Haageocereus versicolor. ibid.1(11): CVa, unpaged. ibid.
KRAINZ, H. (1964). Haageocereus chosicensis.ibid. 1(11): CVa, unpaged. ibid.
KRAINZ, H. (1965). Haageocereus decumbens.ibid. 1(11): CVa, unpaged. ibid.
KRAINZ, H. (1973). Gattung Haageocereus. ibid.16(4): CVa, unpaged. ibid.
KITCHING, I. J., FOREY, P. L., HUMPHRIES, C.J.WILLIAMS, D.M. (1998). Cladistics: The theoryand practice of parsimony analysis. 2nd ed.Oxford University Press, Oxford.
LEUENBERGER, B. E. (1978). Type specimens ofCactaceae in the Berlin-Dahlem Herbarium.
Cact. Succ. J. Gr. Brit. 40 (4): 101–104.LEUENBERGER, B. E. (1979). Typen der Cactaceae
in der Naßpräparate-Sammlung des Herbars.Willdenowia 8: 625–635.
LEUENBERGER, B. E. (1986). Pereskia (Cactaceae).Mem. New York Bot. Gard. 41: 1–141.
LEUENBERGER, B. E. (2004). The Cactaceae of theWilldenow herbarium, and of Willdenow(1813). Willdenowia 34: 309–322.
LOT, A. & CHIANG, F. (1986). Manual de Herbario:Administración y manejo de colecciones, técni-cas de recolección y preparación de ejemplaresbotánicos. UNAM, México.
MAUSETH, J., KIESLING, R. & OSTOLAZA, C. (2002).A Cactus Odyssey. Journeys in the wilds ofBolivia, Peru and Argentina. Timber Press,Oregon.
MCNEILL, J., BARRIE, F.R., BURDET, H.M.,DEMOULIN, V., HAWKSWORTH, D.L., MARHOLD,K., NICOLSON, D.H., PRADO, J., SILVA, P.C.,SKOG, J.E., WIERSEMA, J.H. & TURLAND, N.J.(eds.) (2006). The International Code ofBotanical Nomenclature (Vienna Code).Regnum Vegetabile, 146.
MEYEN, I. (1833). Cactus Arten. Allg. Garten. 1:211.
MOSTACERO, J & MEJÍA, F. (1993). Taxonomía defanerógamas peruanas. CONCYTEC.
NOVOA, S. (2003). Los habitantes de los cactus.Quepo 17: 94–98.
NYFFELER, R. (2002). Phylogenetic relationships inthe cactus family (Cactaceae) based on evi-dence from trnK/ matK and trnL-trnFsequences. Amer. J. Bot. 89 (2): 312–326.
OLDFIELD, S. (Ed.) (1997). Cactus and SucculentPlants: Status and Conservation. Action Plan.IUCN/SSC Cactus and Succulent SpecialistGroup. IUCN, Gland, Switzerland.
ONERN. (1995). Guía explicativa del MapaEcológico del Peru.
OSTOLAZA, C. (1980). Las cactáceas y la conserva-ción de la flora. Boletín de Lima 7: 40–44.
OSTOLAZA, C. (1988). Los cactus de los alrededoresde la ciudad de Lima y su conservación. ZonasAridas 5: 7–18.
OSTOLAZA, C. (1995a). Cacti in the vicinity of Lima,Peru, and their conservation status. Cact.Succ. J. (US) 67: 6–13.
OSTOLAZA, C. (1995b). IOS Cambios recientes.Quepo 9.
OSTOLAZA, C. (1996). A Closer Look at theConservation Status of Cacti in the Vicinity ofLima, Peru. Brit. Cact. & Succ. J. 14 (4):158–174.
OSTOLAZA, C. (1998a). Nomenclatural adjustments
Bradleya 25/2007118

Bradleya 25/2007 119
in Peruvian Cactaceae. Cact. Con. Init. 6: 8–9.OSTOLAZA, C. (1998b). Conservation of cacti in
Lima, Peru. Brit. Cact. Succ. J. 16 (2): 63–66.OSTOLAZA, C. (1998c). The cacti of the Pisco, Ica
and Nazca valleys, Peru. Brit. Cac. Succ. J.16 (3): 127–136.
OSTOLAZA, C. (2000). Haageocereus tenuis andHaageocereus repens rescued Peruvian cacti.Brit. Cact. Succ J. 18 (3): 129–131.
OSTOLAZA, C. (2002). Cactus de Lima en peligrode extinción. Quepo 16: 56–58.
OSTOLAZA, C. (2003). Categorización yConservación de Cactáceas Peruanas. ZonasAridas 7: 199–210.
OSTOLAZA, C., CERONI, A., DREYFUS, G., CALDERÓN,N. & NOVOA, S. (2003). Cacti of the Huaurariver valley, Peru – a conservation survey.Brit. Cact. Succ. J. 21 (2): 87–97.
OSTOLAZA, C. & RAUH, W. (1990). Bemerkungenzur Kakteenvegetation der peruanischenKüstenwüste, insbesondere zu Haageocereustenuis Ritter, einer wenig bekannten Art.Kakt. and. Sukk. 41 (2): 40–44.
PENNINGTON, T. D., REYNEL, C & DAZA, A. (2004).Illustrated guide to the Trees of Peru. DavidHunt, Sherborne, Dorset.
PFEIFFER, L. (1837). Enumeratio DiagnosticaCactacearum. Ludovici Oehmigke, Berlin, pp.77–78.
PORSCH, O. 1939. Das Bestäubungsleben derKakteenblüte.
PRIMACK, R. (1998). Essentials of ConservationBiology. 2nd ed. Boston University, Boston.
RAUH, W. (1958). Beitrag Zur KenntnisPeruanischen Kakteenvegetation. Springer-Verlag, Heidelberg.
RAUH, W. & BACKEBERG, C. (1957). In: BackebergC., Descr. Cact. Nov. [1]. Jena.
RAUH, W. & BACKEBERG, C. (1958). In: Rauh, W.Beitr. Kenntn Peruan. Kakteenveg. (See Rauh,1958).
RETAMOZO, E. (2002). Formas polínicas de cactá-ceas en Arequipa. Quepo 16: 28–32.
RITTER, F. (1956–1958). H. Winter seed Katalog.Frantfurt A.M.
RITTER, F. (1980). Kakteen in Südamerika. 3.Privately published, Spangenberg.
RITTER, F. (1981). Kakteen in Südamerika. 4.Privately published, Spangenberg.
ROSS, R. (1982). Initiation of stamens, carpels,and receptacle in the Cactaceae. Amer. J. Bot.69: 369–379.
ROWLEY, G. D. (1980). Name that Succulent.Stanley Thornes Ltd., Cheltenham.
ROWLEY, G. D. (1982). Intergeneric hybrids in
Succulents. Nat. Cact. Succ. J. 37 (3): 76–80.ROWLEY, G. D. (1999). Salm-Dyck’s cactus pain-
tings. Bradleya 17: 1–26. SAHLEY, C. T. (1996). Bat and Hummingbird pol-
lination of an autotetraploid columnar cactus,Weberbauerocereus weberbaueri (Cactaceae).Amer. J. Bot. 83 (10): 1329–1336.
SALM-DYCK, J. F. V. (1845a). Cactaceae in HortoDyckensi Cultae. Crapelet, Paris.
SALM-DYCK, J. F. V. (1845b). Beschreibung einigerneuen Cacteen. In Otto, F. & Dietrich, A. Allg.Garten. 13 (45): 353–354.
SALM-DYCK, J. F. V. (1850). Cactaceae in HortoDyckensi Cultae. Henry & Cohen, Bonn.
SCHILL, R., BARTHLOTT, W. & EHLER, N. (1973).Cactus spines under the electron scanningmicroscope. Cact. Succ. J. (US) 45: 175–185.
SOULÉ, M. (1987). Viable Populations forConservation. Cambridge University Press,Cambridge.
STACE, C. A. (1989). Plant taxonomy and biosys-tematics. 2nd ed. Cambridge UniversityPress, Cambridge.
STEARN, W. T. (1966). Botanical Latin. HafnerPublishers, New York.
TAKHTAJAN, A. L. (1980). Outline of the classifica-tion of flowering plants (Magnoliophyta). Bot.Rev. 46 (3): 226–359.
TAYLOR, N. P. & ZAPPI, D. C. (1989). An alterna-tive view of generic delimitation and relation-ships in tribe Cereeae (Cactaceae). Bradleya7: 13–40.
TAYLOR, N. P. & ZAPPI, D. C. (2004). The Cacti ofEastern Brazil. Royal Botanic Gardens, Kew.Richmond, Surrey.
TERRAZAS, S. T. & MAUSETH, J. (2002). ShootAnatomy and Morphology. In Nobel P.S. (Ed.)Cacti, Biology and Uses. University ofCalifornia Press, Berkeley/Los Angeles/London, pp. 23–40.
VAUPEL, F. (1913). Cactaceae Andinae. In Engler,A. (Ed.) Beibl. Bot. Jahrb. 50 (111): 12–19.
VILCAPOMA, G. (2000). Frutas silvestres de laCuenca del río Chillón (Cactaceae). Quepo 15:59–67.
WALLACE, R. S. (1995). Molecular systematicstudy of the Cactaceae: Using chloroplastDNA variation to elucidate Cactus phylogeny.Bradleya 13: 1–12.
WALLACE, R. S. (1997). The Phylogenetic positionof Mediocactus hahnianus. Cact. Cons. Init. 4:11–12.
WALLACE, R. S. (2002). Phylogeny andSystematics of Columnar Cacti. In Fleming,T. H. & Valiente-Banuet, A. (Eds.) Columnar

Cacti and Their Mutualists: Evolution,Ecology and Conservation. The University ofArizona Press, Tucson, pp. 42–65.
WALLACE, R. S. & COTA, J. H. (1996). An intronloss in the chloroplast gene rpoC1 supports amonopheletic origin for the subfamilyCactoideae of the Cactaceae. Curr. Genet. 29:275–281.
WALLACE, R. S. & DICKIE, S. L. (2002). Taxonomicimplications of chloroplast DNA sequencevariation in subfam. Opuntioideae(Cactaceae). Succ. Pl. Res. 6: 9–24.
WALLACE, R. S. & GIBSON, A. C. (2002). Evolutionand Systematics. In Nobel, P. S. (Ed.) Cacti:Biology and Uses. University of CaliforniaPress, Berkeley/Los Angeles/London, p.p.1–21
WEBERBAUER, A. (1911). Die planzenwelt der peru-anischen Anden in ihren Graundzugendargestellt. W. Engelmann, Leipzig.
WEBERBAUER, A. (1945). El Mundo Vegetal de losAndes Peruanos. Estudio Fitogeográfico. 2nd.Ed. Estación Experimental Agrícola de LaMolina, Dirección de Agricultura. Ministeriode Agricultura, Lima.
WERDERMANN, E. (1932). Neue sukkulentePflanzen im Bot. Garten B.-Dahlem. FeddeRep. Spec. Nov. 30: 52–66.
WERDERMANN, E. (1937a). Neue sukkulentePflanzen in Bot. Garten B.-Dahlem. FeddeRep. Spec. Nov. 42: 4–6.
WERDERMANN, E. (1937b). Neue und kritischeKakteen aus den Sammelergebnissen derReise von Harry Blossfeld durch Sudamerika1936/1937.II. Binghamia climaxantha.Kakteenkunde 1937 (4): 51–54.
WERDERMANN, E. (1937c). Binghamia Br. et R.Haageocereus Bbg. – Pseudoespostoa Bbg.Kakteenkunde 1937 (2): 21–24.
WERDERMANN, E. & BACKEBERG, C. (1931). InBackeberg, C. Neue Kakteen. Trowitzsch &Sohn, Frankfurt.
WILLDENOW, C.L. (1809). Enumeratio plantarumhorti regii botanici berolinensis, continensdescriptiones omnium vegetabilium in hortodicto cultorum. Taberna Libraria ScholaeRealis Pub., Berlin.
WILLDENOW, C.L. (1814). Enumeratio plantarumHorti regii berolinensis. Supplementum postmortem autoris editum. Taberna LibrariaScholae Realis Pub., Berlin.
ZAPPI, D. C. (1992). Revisão Taxonômica dePilosocereus Byles & Rowley (Cactaceae) noBrasil. PhD. Thesis, Univ. São Paulo, unpublished.
ZAPPI, D. C. (1994). Pilosocereus (Cactaceae). Thegenus in Brazil. Succ. Pl. Res. 3: 1–160.
Appendix 1: List of names of Haageocereus(and Peruvocereus) possibly referring to×HaagespostoaThe following taxa are believed to be namesreferring to hybrids between Haageocereus andEspostoa.H. albisetatus (Akers) Backeb., Die Cact. 2: 1208
(1960); Rauh, Beitr. Kenntn. Peruan.Kakteenveg.: 414 (1958). H. albisetatus(Akers) Cullmann, Kakt. and. Sukk. 8 (12):180 (1957) Nom. inval. (McNeill et al., 2006),given as comb., but without any indication ofa basionym. ×Haagespostoa albisetata (Akers)Rowley, Nat. Cact. Succ. J. 37 (3): 76 (1982).Peruvocereus albisetatus Akers, Cact. Succ. J.(US) 20 (12): 184 (1948d). Type locality: Peru,Lima, Santa Eulalia valley, hills above valley(believed not to have been preserved). Parents: H. pseudomelanostele subsp.carminiflorus and Espostoa melanostele.
H. albispinus (Akers) Backeb., Rauh, Beitr.Kenntn. Peruan. Kakteenveg.: 414 (1958); inBackeb., Die Cact. 2: 1210 (1960). H. chosi-censis var. albispinus (Akers) Backeb., Cact.Succ. J. (US) 23 (2): 47 (1951). Peruvocereusalbispinus Akers, Cact. Succ. J. (US) 20 (10):154 (1948c). Type locality: Peru, Lima, SantaEulalia valley.Parents: H. pseudomelanostele subsp.carminiflorus and Espostoa melanostele.
H. albispinus var. floribundus (Akers) Backeb.,Die Cact. 2: 1210 (1960). Peruvocereus albis-pinus var. floribundus Akers, Cact. Succ. J.(US) 20 (10): 155 (1948c). Type locality: Peru,Lima, hills above Santa Eulalia river valley. Parents: H. pseudomelanostele subsp.carminiflorus and Espostoa melanostele.
H. climaxanthus (Werderm.) Croizat, Cact. Succ.J. (US) 14 (10–11): 148 (1942b). Binghamiaclimaxantha Werderm., Fedde Rep. Spec.Nov. 42: 4 (1937a). ×Haagespostoa climaxan-tha (Werderm.) Rowley, Nat. Cact. Succ. J. 37(3): 76 (1982). Type: Peru, Lima, by Chosica,1936, Blossfeld 84 (B, photo!).Parents: H. pseudomelanostele subsp.carminiflorus and Espostoa melanostele.
H. comosus Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 21 (1956, publ. 1957); Die Cact.2: 1230 (1960). Type: Peru, Lima, SantaEulalia valley, 1000 m, 1956, Rauh K27(HEID, believed not to have been preserved). Parents: H. pseudomelanostele subsp.
Bradleya 25/2007120

Bradleya 25/2007 121
carminiflorus and Espostoa melanostele.H. dichromus var. pallidior Rauh & Backeb. in
Backeb. Descr. Cact. Nov. [1]: 24 (1956,publ.1957); Die Cact. 2: 1214 (1960). Type:Peru, Lima, Churín valley, 1700 m, 1956,Rauh K99 (HEID, believed not to have beenpreserved).Parents: H. pseudomelanostele subsp.aureispinus and Espostoa melanostele.
H. divaricatispinus Rauh & Backeb. in Backeb.Descr. Cact. Nov. [1]: 26 (1956, publ. 1957);Die Cact. 2: 1231 (1960). Type: Peru, Lima,Lurín valley, 500–1200 m, 1956, Rauh K176(HEID, believed not to have been preserved). Parents: H. pseudomelanostele subsp.carminiflorus and Espostoa melanostele.
H. pseudomelanostele subsp. chryseus D. Hunt,Cact. Syst. Init. 14: 17 (2002). Type: PERU,Ancash, Huallanca, Ritter FR147a (U holo!).H. chryseus Ritter, Kakt. Südam. 4: 1390(1981). Type: Peru, Ancash, Santa valley,Huallanca, Ritter FR585 (U, syn) & FR147a.Nom inval. (McNeill et al., 2006), based ontwo syntypes: Ritter FR585 and RitterFR147a.Parents: H. pseudomelanostele subsp.aureispinus and Espostoa nana.
H. seticeps Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 21 (1956, publ. 1957); Die Cact.2: 1217 (1960). Type: Peru, Lima, SantaEulalia valley, 1000 m, 1956, Rauh K43(HEID, believed not to have been preserved).Parents: H. pseudomelanostele subsp.carminiflorus and Espostoa melanostele.
H. seticeps var. robustispinus Rauh & Backeb. inBackeb. Descr. Cact. Nov. [1]: 21 (1956, publ.1957); Die Cact. 2: 1217 (1960). Type: Peru,Lima, Santa Eulalia valley, 1000 m, 1956,Rauh K37 (HEID, believed not to have beenpreserved).Parents: H. pseudomelanostele subsp.carminiflorus and Espostoa melanostele.
H. zehnderi Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 23 (1956, publ.1957); Die Cact.2: 1225 (1960). Type: Peru, Ancash, Santa val-ley, Huallanca, 1300 m, 1956, Rauh K67 (ZSSholo!).Parents: H. pseudomelanostele subsp.aureispinus and Espostoa nana.
Peruvocereus albisetatus var. robustus Akers,Cact. Succ. J. (US) 20 (12): 186 (1948d). Typelocality: Peru, Lima, Santa Eulalia valley,hills above valley (believed not to have beenpreserved).Parents: H. pseudomelanostele subsp.
carminiflorus and Espostoa melanostele.Peruvocereus albicephalus Akers, Cact. Succ. J.
(US) 19 (10): 162 (1947f). Type locality: Peru,Lima, Santa Eulalia valley, hills above valley(believed not to have been preserved).Parents: H. pseudomelanostele subsp.carminiflorus and Espostoa melanostele.
Peruvocereus albicephalus var. armatus Akers,Cact. Succ. J. (US) 19 (10): 163 (1947f). Type:(believed not to have been preserved).Parents: H. pseudomelanostele subsp.carminiflorus and Espostoa melanostele.
Appendix 2: List of doubtful names attrib-uted to the genus HaageocereusCactus multangularis Willd., Enum. Pl. Suppl.:
33 (1813). Cereus multangularis (Willd.)Haw., Suppl. Pl. Succ.: 75 (1819). Binghamiamelanostele sensu (Vaupel) Britton & Rose,Cact. 2: 167 (1921) non Cephalocereusmelanostele Vaupel (1913). Binghamia mul-tangularis (Willd.) Britton & Rose, Cact. 4:279 (1923). Peruvocereus multangularis(Willd.) Akers, Cact. Succ. J. (US) 22 (6): 174(1950). H. akersii Backeb., Rauh, Beitr.Kenntn. Peruan. Kakteenveg.: 416 (1958),nom. inval. (McNeill et al., 2006) given as syn-onym of Peruvocereus multangularis. Type:believed not to have been preserved.According to the scant description and non-existent type of Cactus multangularis Willd.,it is not possible to determine which taxonthis name actually refers to. However, a laterpainting from Haworth’s cultivated plant ofCereus multangularis (Willd.) Haw. (dated1824 and reproduced by Britton & Rose, 1923,p. 279, Fig. 255) led to the proliferation oflater combinations using this specific epithetbecause of its priority as an earlier name.Another “controversial” painting of Cactusmultangularis Willd. (undated) is from Salm-Dyck (reproduced by Rowley, 1999) that wasalso published by Leuenberger (2004) and,from his point of view, this species could be asynonym of Weberbauerocereus johnsonii, butto avoid further confusion Cactus multangu-laris should be rejected.
Cereus limensis Salm-Dyck, Allg. Garten. 13 (45):353 (1845b). Type: believed not to have beenpreserved.Because of the scant description and non-exis-tent type it is not possible to attribute Cereuslimensis to any known species ofHaageocereus. However, Ritter made the com-bination H. limensis, nom. inval. (McNeill et

al., 2006) because he considered it to be con-specific with Cereus acranthus Vaupel (1913)and in this manner preserved Salm-Dyck’sold name. Existing doubts about the identityof Cereus limensis suggest that this name isbetter left as unidentifiable.
H. acranthus var. metachrous Rauh & Backeb. inBackeb. Descr. Cact. Nov. [1]: 22 (1956, publ.1957); Die Cact. 2: 1177 (1960). Type: Peru,Ica, Pisco valley, 2000 m, 1955, Rauh K162(HEID, believed not to have been preserved).There is insufficient evidence to attribute thisname to any known species of Haageocereus.
H. albispinus var. roseospinus (Akers) Backeb.,Die Cact. 2: 1211 (1960). Peruvocereus albispi-nus var. roseospinus Akers, Cact. Succ. J.(US) 20 (10): 156 (1948c). Type locality: Peru,Lima, hills above Santa Eulalia river valley.There is insufficient evidence to attribute thisname to any known species of Haageocereus.
H. aticensis Ritter, Katalog H. Winter, (unpaged),1958. Nom. Inval. (McNeill et al., 2006) pub-lished as provisional name (Eggli & Taylor,1991). According to Ritter this taxon is the same asH. subtilispinus Ritter (1981, p. 1419).However, there is insufficient evidence toattribute this name to any known species ofHaageocereus.
H. australis var. acinacispinus Rauh & Backeb.in Backeb. Desc. Cact. Nov.[1]: 25 (1956, publ.1957). Rauh, Beitr. Kenntn. Peruan.Kakteenveg.: 392 (1958). Type: Peru, south,km 697 Panamericana highway, rocky desert,1956, Rauh K131 (HEID, believed not to havebeen preserved).There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. chosicensis var. rubrispinus (Akers) Backeb.Cact. Succ. J. (US) 23 (2): 47 (1951). H.rubrospinus (Akers) Cullmann, Kakt. and.Sukk. 8 (12): 180 (1957), nom inval. (McNeillet al., 2006) based on Peruvocereus rubrispi-nus (Eggli & Taylor 1991). H. chosicensis fa.rubrispinus (Akers) Krainz, Die Kakt. C Va(1964). Peruvocereus rubrispinus Akers, Cact.Succ. J. (US) 19 (8): 121 (1947d). Type locali-ty: Peru, Lima, 300 m above Santa Eulaliariver, western ridge.There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. crassiareolatus Rauh & Backeb. in Backeb.Descr. Cact. Nov. [1]: 24 (1956, publ.1957); DieCact. 2: 1214 (1960). Type: Peru, Lima,Churín valley, 1200 m, 1956, Rauh K90b
(HEID, believed not to have been preserved). There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. crassiareolatus var. smaragdisepalus Rauh &Backeb. in Backeb. Descr. Cact. Nov. [1]: 24(1956, publ. 1957); Die Cact. 2: 1214 (1960).Type: Peru, Lima, Churín valley, 1200 m,1956, Rauh K94 (HEID, believed not to havebeen preserved).There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. dichromus Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 24 (1956, publ.1957); Die Cact.2: 1212 (1960). Type: Peru, Lima, Churín val-ley, 1200 m, 1956, Rauh K101 (HEID,believed not to have been preserved).Haageocereus chosicensis var. dichromus(Rauh & Backeb.) Ritter ex Krainz, Kat. ZSS2: 64 (1967).There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. elegans Ritter, Katalog H. Winter, (unpaged),(1957) nom. nud. H. versicolor var. elegansRitter, Katalog H. Winter, (unpaged), (1957)nom. nud.This is an invalid name that was later consid-ered by Ritter as synonym of H. icosagonoides(Kakt. Südam. 4: 1422, 1981). However, thereis insufficient evidence to link this name toany accepted species in this work.
H. fulvus Ritter, Kakt. Südam. 4: 1393 (1981).Type: Peru, Lima, Fortaleza valley, RitterFR584 (ZSS only seeds!), believed not to havebeen preserved.According to Ritter, this taxon and H. acran-thus var. fortalezensis Rauh & Backeb. aresynonymous. Nevertheless, there is insuffi-cient evidence to attribute this taxon to anyknown species of Haageocereus.
H. fulvus var. yautanensis Ritter, Kakt. Südam.4: 1393 (1981). Type: Peru, Ancash, Yaután,Ritter FR 1067 (U holo!, ZSS seeds!).The available evidence is insufficient toattribute this taxon to any known species ofHaageocereus.
H. icensis (Backeb. ex) Ritter, Kakt. Südam. 4:1394 (1981). Type: Peru, Ica, Canza Mine,1953, Ritter FR146 (ZSS iso!). The very poor type specimen and descriptionare not enough evidence to attribute thistaxon to any known species of Haageocereus.
H. longiareolatus Rauh & Backeb. in Backeb.Descr. Cact. Nov. [1]: 21 (1956, publ. 1957);Die Cact. 2: 1208 (1960). Type: Peru, Lima,Santa Eulalia valley, Chosica, 1000 m alt.,
Bradleya 25/2007122

Bradleya 25/2007 123
1956, Rauh K40 (HEID, believed not to havebeen preserved). There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. multangularis var. aureus Ritter, Kakt.Südam. 4: 1405 (1981). Based on Ritter 147d.Nom inval. (Art. 36.1, ICBN 2000) publishedas provisional name (Eggli & Taylor 1991).There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. pacalaensis var. montanus Ritter, Kakt.Südam. 4: 1417 (1981), nom. inval. (McNeillet al., 2006), published as provisional name,based on Ritter FR 294a.There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. peniculatus Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 21 (1956, publ.1957); Die Cact.2: 1214 (1960). Type: Peru, Lima, SantaEulalia valley, 1000 m, 1956, Rauh K40(HEID, believed not to have been preserved).According to Hunt (1999) this taxon isbelieved to be a synonym of H. albispinus(Akers) Backeb. (H. albispinus is included inthe list of names possibly referring to×Haagespostoa). However, there is insuffi-cient evidence to attribute this taxon to anyknown species of Haageocereus.
H. piliger Rauh & Backeb. in Backeb. Descr. Cact.Nov. [1]: 26 (1956, publ. 1957); Die Cact. 2:1233 (1960). Type locality: Peru, Lima,Pachacámac, 100 m, 1956, Rauh K178 (HEID,believed not to have been preserved).There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. pseudoacranthus Rauh & Backeb. in Backeb.Descr. Cact. Nov. [1]: 23 (1956, publ. 1957);Die Cact. 2: 1176 (1960). Type: Peru, Lima,Lurín valley, 1000 m, 1956, Rauh K181(HEID, believed not to have been preserved).There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. salmonoideus (Akers) Cullmann, Kakt. and.Sukk. 8 (12): 180 (1957) Nom inval. (McNeillet al., 2006) given as comb. nov. but withoutany indication of a basyonym (Eggli & Taylor1991). H. salmonoideus (Akers) Backeb. inRauh, Beitr. Kenntn. Peruan. Kakteenveg.:429 (1958). Peruvocereus salmonoideus Akers,Cact. Succ. J. (US) 19 (7): 109 (1947). Typelocality: Peru, Lima, Rímac canyon, 15 kmabove Chosica. There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. tenuispinus Rauh & Backeb. in Backeb. Descr.Cact. Nov. [1]: 22 (1956, publ. 1957). Type:Peru, Ancash, km 465 Panamericana high-way, 30 km north of Pativilca, desertic hills,1956, Rauh K89 (ZSS).There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. smaragdiflorus Rauh & Backeb. in Backeb.Descr. Cact. Nov. [1]: 21 (1956, publ. 1957);Die Cact. 2: 1209 (1960). Type: Peru, Lima,Santa Eulalia valley, 1000 m alt., 1956, RauhK40 (HEID, believed not to have been pre-served). There is insufficient evidence to attribute thistaxon to any known species of Haageocereus.
H. zangalensis Ritter, Kakt. Südam. 4: 1424(1981). Type: Peru, Cajamarca, Zangal–Chilete, 2000 m, Ritter FR1074 (U holo!).The incomplete description without flowers orfruits and very poor type prevent the attribu-tion of this taxon to any known species ofHaageocereus. Fieldwork carried out duringthis project failed to shed more light into theidentity of this name.
Pygmaeocereus densiaculeatus Backeb., Descr.Cact. Nov. [3]: 12 (1963). Type: believed not tohave been preserved.There is insufficient evidence to attribute thistaxon to any known species of Haageocereus,however Ritter (1981) believed it to belongwith H. lanugispinus.
Appendix 3: List of names of Haageocereusnow placed in other generaH. albus (Ritter) Rowley, Nat. Cact. Succ. J. 37
(3): 76, 1982. = Weberbauerocereus longicomus Ritter,in Hunt, D., CITES Cactaceae Checklist: 282(1999).
H. andinus Ritter, Backeberg’s Descr. & Erort.Taxon. nomenklat. Fragen.: 14, 1958. Nom.inval. (McNeill et al., 2006) (published as pro-visional name) (Eggli & Taylor, 1991). = Cleistocactus hystrix (Rauh & Backeb.)Ostolaza, as synonym in Hunt, D. CITESCactaceae Checklist: 171 (1999).
H. convergens Ritter, Katalog H. Winter,(unpaged), (1956). Nom inval. (McNeill et al.,2006) (1956, publ. 1957). (Eggli & Taylor,1991).= Cleistocactus acanthurus (Vaupel) Huntssp. acanthurus, as synonym in Hunt, D.,CITES Cactaceae Checklist: 170 (1999).
H. hystrix Ritter, Katalog H.Winter, (unpaged),(1958). Nom. inval. (McNeill et al., 2006) pub-

lished as provisional name (Eggli & Taylor1991).= Cleistocactus hystrix (Rauh & Backeb.)Ostolaza, as synonym in Hunt, D. CITESCactaceae Checklist: 171 (1999).
H. imperialis Ritter, Katalog H.Winter,(unpaged), (1958). Nom. inval. (McNeill et al.,2006) published as provisional name (Eggli &Taylor, 1991).= Cleistocactus acanthurus (Vaupel) Huntssp. acanthurus, as synonym in Hunt, D.,CITES Cactaceae Checklist: 170 (1999).
H. montanus Ritter, Katalog H.Winter,(unpaged), (1958). Nom inval. (McNeill et al.,2006) published as provisional name (Eggli &Taylor 1991).
= Cleistocactus hystrix (Rauh & Backeb.)Ostolaza, as synonym in Hunt, D. CITESCactaceae Checklist: 171 (1999).
H. pacaranensis Ritter, Katalog H.Winter,(unpaged), (1958).= Cleistocactus acanthurus (Vaupel) Huntssp. acanthurus, as synonym in Hunt, D.,CITES Cactaceae Checklist: 170 (1999).
H. paradoxus Rauh & Backeb., Descr. Cact. Nov.[1]: 21 (1956, publ. 1957); Die Cact. 2: 1242,(fig.) (1959). Type: Peru, Lima, Santa Eulaliavalley, 1000 m, 1956, Rauh K42 (HEID,believed not to have been preserved). = Cleistocactus acanthurus (Vaupel) Huntssp. acanthurus, as synonym in Hunt, D.,CITES Cactaceae Checklist: 170 (1999).
Bradleya 25/2007124

















![FYdgYdcYf]k Yf\ FYdgYj]f]k - Samagra](https://static.fdokumen.com/doc/165x107/631f358f13819e2fbb0fa1db/fydgydcyfk-yf-fydgyjfk-samagra.jpg)


