
Pliocene and Pleistocene archeological sites west of Lake Turkana, Kenya

Functional morphology and palaeobiology of the Pliocene rodent Actenomys (Caviomorpha: Octodontidae): the evolution to a subterranean mode of life
MARIA ELENA F E R N ~ D E Z ] , ALDO IVAN VASSALLO'* AND MARCEL0 ZARATE3
'LhCLA EEA Bariloche, Casilla de Correo 277 (8400) Rio &ego, Argentina, 'Departamento de Biologia, Fucultad de Ciencias Exactas y JVuturales, Universidud Nacional de M a r del Plata, C o J V ~ c E ~ Casilla de Correo 1245 (7600) M a r del PEata, Argentina, 'CONICE7; ~ r ~ ~ / C ~ r C ~ C a d l a de Correo 330 (5500) Mendoza, Argentina
Recezwd 21 Jnnuagl 1999; nrceptedjr pbhratzon 16 October 1999
The Pliocenc caviomorph rodent Actenomys has long becn recognized as an early fossorial representative of the subfamily CtenomJ-inae (Octodontidae), which includes one living genus, C'tenomys (tuco-tucos), and several species widely distributed in South America. To assess the degree of specialization for digging in Artenomys, we performed morphological comparisons with other octodontid genera (Octodon, Spa/acopus, and Ctenomts) of known mode of life and behaiiour. As a whole, our results indicate that, in terms of morphological specializations for digging, Bctenomys occupies an intermediate position between Octodon, a generalized semi- fossorial rodent which forages above ground, and Ctenomy.c-Spalucopu.c, two highly spccialized subterranean forms. The position of the deltoid process (humerus) and the length of the olecranon process (ulna)-two traits which affect the out-forces exerted by sc\-cral forelimb muscles-were in Actenomys intermediate between non-subterranean and subterranean taxa. The skull, particularly the rostrum, appears to be strong, and sagittal and nuchal crests are well marked. The zygomatic arches are as flared as those of the generalized Octodon. Notably, the upper and lower incisors of Actenomys were extremely procumbent, as in many highly specialized chisel-tooth digging species. Based upon thc stratigraphic and sedimentological analysis of the palaeosoils containing its fossil remains, we suggest that Actenomys lived in an environment of moderate to hig primary productivity. The texture of the palaeosoils indicate that theywere hard and highly cohesive. This situation, which contrasts with that observed in living Ctenomyinae, has relevant implications for burrowing cost. The integration of morphological and palaeoenvironmental data allowed testing of hypotheses about the palaeobiological attributes of this ancestral ctenomyine.
0 2000 The Linnean Society of Londuti
ADDITIONAL KEY WORDS:-Ctcnomyinae - Ctenomys ~ subterranean rodents ~ evolu- tionary morphology - palaeoenvironment - adaptation.
Corresponding author. E-mail: [email protected]
002.1 -1.066/00/09007 1 + 20 $35.00/0 71
0 2000 The 1,innean Societv of London

Introduction . . . . . . . . . . . . . . . . . . . . . . . 72 Slatcrial and methods . . . . . . . . . . . . . . . . . . . 73 Slorpiiological analysis . . . . . . . . . . . . . . . . . . . 7 :i t-'alarocn~.ironmental analysis . . . . . . . . . . . . . . . . . I ,>
5 lorpliological analysis . . . . . . . . . . . . . . . . . . Palaeoenvironmental analysis . . . . . . . . . . . . . . . . 80
irig potcntialiries of Ar/~nony,s . . . . . . . . . . . . . . 81 nstruction of ;Irte77omy habitat . . . . . . . . . . . . . .
Iriregrating morphological and palarornvironmental information . . . . 8,5 :\~kno\\.ledKcniciits . . . . . . . . . . . . . . . . . . . . 87 Kcfcrcnce~ . . . . . . . . . . . . . . . . . . . . . . . 88
-_ Re5ulth . . . . . . . . . . . . . . . . . . . . . . . . 75 - -
I D
Di5cussion . . . . . . . . . . . . . . . . . . . . . . . 80
8 3
\\ithiti the order Rodentia, six families (Geomyidae, Cricetidae, Spalacidae, Rhizomyidae. Octodontidae and Bathyergidae) contain se\,eral genera and more than 120 living subterranean species, which are extensively distributed within both the Old and New World. The open country environments-savannas, grasslands, dcscrts-which etnerged during the mid-to-late Cenozoic cycles of increasing aridity, prmided a new set of evolutionary challenges and opportunities for mammalian diversification, chiefly accomplished by cursorial herbivores, along with fossorial hcrbivorcs and insectivores. The repeated Miocene-Pleistocene adaptive radiation of unrelated rodent families into the ecological fossorial zone has been causally relatcd to the evolution of an open country biota (Nevo, 1979).
'i'he subfamily Ctenomyinae (Reig, 1958), which has long been recognized as a late Illiocenc offshoot of the family Octodontidae (Patterson & Pascual, 1968) includes one living genus, Ctenomys (tuco-tucos), which includes more than 5.5 rc~ognized species with a highly specialized subterranean mode of life (Keig ct al., 1 CIYO). Tuco-tucos are all homogeneous in their main fossorial adaptations, with specific adjustments to particular ecological conditions (Vassallo, 1998). Ctenomyinac ;dso includes several extinct paucispecific genera (Actenomys, Praectenomys, Eucelophorus, Partrctenomlis, Megactenomys; see Verzi, Montalvo & Vucetich, 199 l), which seem to rcpresent different alternative adaptations and, presumably, different commitments t o the underground ecological niche. In particular, the close association between Actcnomys (Burnieister, 1888) remains and fossil burrows in coastal cliffs of Bucnos =2ires province, Argentina helps to justify the hypothesis about fossorial habits in this extinct octodontid rodent.
XI1 suhtcrranean rodent species have a unique, extraordinary structure--thc burrow system which connects them to their environment (see Hickman, 1978, 1990: Andersen, E988; Altuna, 1991; Brett, 1991; Antinuchi & Busch, 1992). It pro\ides access to plant material, refuge, and particular ecophysiological conditions such as high relative humidity, small temperature variation, low 0, tcnsion, and high CO, tension (McNab, 1966). Many species of mammals, including epigeous rodents, utilize burrpws for refuge, food storage, and/or pup nursing (Carter Sr Encarnayiio, 1983; Agren, Zhou & Zhong, 1989; Carter & Kosas, 1997; Hodara, Suarcz & Kravetz, 1997). Semi-fossorial and fossorial rodents often haw extensivr

and very elaborate burrows, with several chambers and openings (Llanos & Crespo. 1952; YaAez & Jaksic, 1978; Scheck & Fleharty, 1980; Pearson, 1984; Ojeta et al., 1996). However, only those species that exclusively feed on plant material that they can obtain without leaving the strict proximity of their burrows are ecologically termed ‘subterranean rodents’. Hence, the acquisition of food always has a con- comitant foraging cost, determined mainly by the cost of excavation. The energetic cost of excavating burrows is 360-3400 times as great as moving the same distance across the surface (Vleck, 1979), and the need to loosen and move resistant material requires the capacity to produce and transmit considerable force during long-term activity periods. Out-forces exerted by digging tools (claws, incisors) increase as the ratio of in-lever arm to out-lever arm increases. Hence, the enlarged muscles of diggers have their origins and insertions relatively far from the joints they turn, and the distance from the pivot to the tip of the diggmg tool is short (see review in Hildebrand, 1985, 1988).
An analysis of early ctenomyine diversity is in progress (Reig & Quintana, 1992; Quintana, 1994), still requiring the morphological and palaeobiological analysis of some taxa. The aim of this study is to integrate the functional morphological analysis of the digging apparatus-structure and digging potentialities of the forelimbs and skull-of the extinct genus Actenomys, within the framework of Pliocene palaeo- environmental conditions. The palaeoenvironmental reconstruction is based on the morphological and sedimentological characteristics of the palaeosoils which permit inference of primary plant productivity and soil hardness. Morphological comparisons with other octodontid species of known mode of life, habitat, and behaviour, both subterranean and fossorial, were performed in order to characterize Artenomyr palaeobiology .
RlATEKIAL AND hIETHOIX
Morphological anaLysis
To assess the degree of specialization for digging of Actenomys, a selected group of musculoskeletal characteristics of the forelimb and skull were studied. These characteristics were chosen because they reflect the biomechanical potentialities of the structures commonly used for digging in several fossorial and subterranean mammals (see Hildebrand, 1985; 1988). Actenomys was compared with the following living octodontid taxa of known ecology, behaviour, and mode of digging: Octodon bridgesi, Spalacopus cyanus, Ctenomys australis, and C. talarum. Octodon is a generalized semi-fossorial form which digs burrows for sheltering and rearing, and spends a substantial portion of its life foraging above ground (Yafiez & Jaksic, 1978; Meserve, Martin & Rodriguez, 1984). The chisel-tooth digger S. cyanus, the scratch and chisel- tooth digger C. talarum, and the scratch digger C. austral& are fully subterranean forms (Reig, 1970; Reig et al., 1990; Vassallo, 1998). Spalacopus (subfamily Octodontinae) and Ctenomyr (subfamily Ctenomyinae) have experienced parallel evolution in their adaptations to their subterranean niches. Nonetheless, in addition to an overall similarity, they show noticeable differences associated with ‘chisel tooth’ and ‘scratch’ digging modes (Reig, 1970; Vassallo, 1998).
The specimens of Actenomys belong to the palaeontological and mastozoological
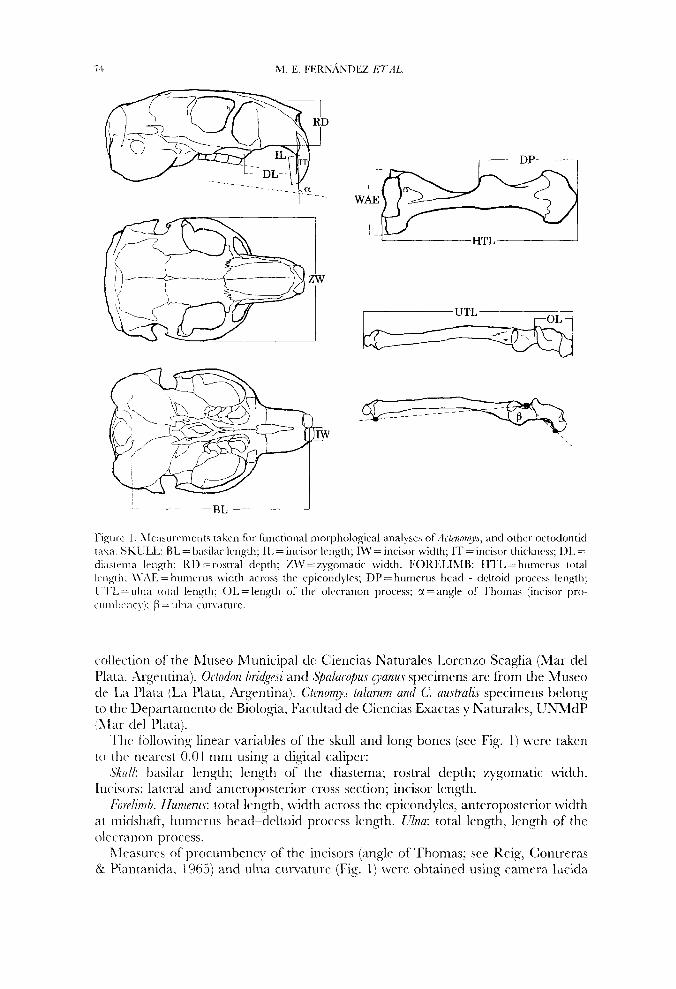
ligure I . LIe;isurrnierits takrii for liirictiotial inorpholosjcal analyses ofActenorny.c, and othcr octodontid i a w . SKULL: RL = hasilar length: 11, =incisor Iriigth; IW = incisor width; IT = incisor thickncss; 111, =
diastema length: KD = rostral depth; ZM' = zygomatic width. FOKE1,ISIB: HTL = humcrus total Iciigth; \\;\E = hunirrus \vidth across thr epicondylcs; DP = humctus hcad - dcltoid pro 1:'I.L = ulna tor;tl l e n ~ t h ; 01, =length of the olccranon procrss; CI = angle of l'homas (incisor pro- ~~i iniI)ci ic> \: /?I = uliia curvature.
collection of the hIuseo hlunicipal dc Ciencias Naturalcs Lorcnzo Scaglia (Mar clel Plata. Argentina). Octodon /iri4qe.~i and S)alacopus vanus specimens arc from the Musco de LA Plata (La Plata, Argcntina). Ctenomys talarum and C. australis specimens belong to the Departarncnto dc Biologia, Facultad de Ciencias Exactas y Naturales, UNMdP (Afar clel Plata).
The follo\.\-ing linear \,ariatiles of the skull arid long bones (see Fig. 1) were taken t o thc nearest 0.01 mni using a digital caliper:
L%u/k basilar length; length of the diastema; rostral depth; zygomatic width. Incisors: lateral arid anteroposterior cross section; incisor length.
Fordimh. Hiarwus: total length, width across the cpicondyles, anteropostcrior width at midshaft, hunicrus hcad-deltoid process length. Ulna: total length, lcngth of thc o h m m n process.
LIeasures of procumbcncy of the incisors (angle of 'Thomas; see Keig, Contreras & l'iantanida. E 965) and ulna curvature (Fig. 1) were obtained using camera lucida

E UNC? IONAL MOKI’HOI.OGY AN11 I’ALAEORIOLOGY 0 1 * AL7F,V0,441S
drawings. The position of incisor roots relative to the cheek teeth, and the degree of development of the teres major process (scapula) were assessed.
The functional/adaptive significance of each specific character with respect to burrowing will be provided in detail in the Discussion.
For the purpose of statistical comparisons, means and confidence intervals (a = 0.05) for linear variables, proportions and angles were calculated. T-tests were applied in all cases after checking for normality. Watson & Williams’ test for angles (Zar, 1984: 450) was used for the analysis of incisor procumbency and ulna curvature.
The body mass of Actenomys was estimated using the following allometric equation, calculated by Casinos, Quintana & Viladiu (1993) with extant species of the genus Ctenomys:
7 i
y=4.89 x0.3 r=0.97
where y is the total length of the humerus, x is body mass, and r is the correlation coefficient.
Palaeoenuironmenta I anahsis
The geological study was performed in a stratigraphic section (Los Acantilados, Playa San Carlos, 38” 05’ 30”s; 57” 35’ 0O”W) located at the coastal sea-cliffs 20 km south of Mar del Plata (Buenos Aires, Argentina). This exposure was selected because of the great abundance of Actenomys fossil burrows throughout the Pliocene section (Zarate, 1989). Following the Paleopedology Manual (Catt, 1990), the field morphological description of palaeosoils included colour, structure, texture, horizon boundaries, cutans, and CaCO? accumulations. Samples for grain size analysis which were collected aproximately every 30 cm, were treated by sieve and pipette analysis (Carver, 197 1) using the Udden-Wentworth grain size scale.
RESULTS
Morphologzcal anahsic.
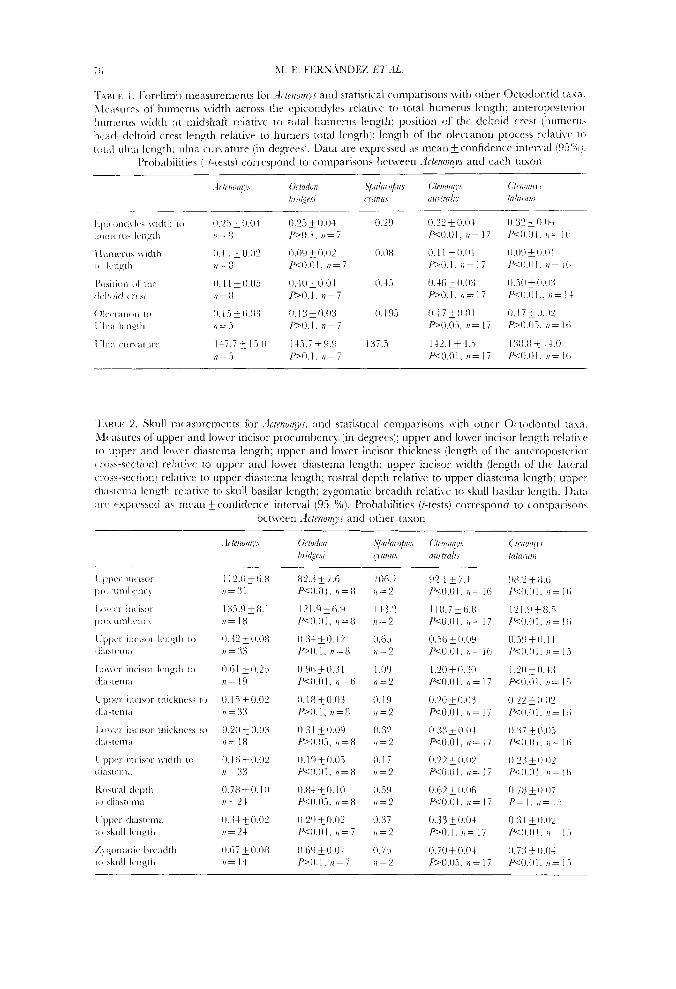
Tables 1 and 2 provide comparative information on several musculoskeletal features that may reflect the degree of specialization for digging. The width of the humerus across the epicondyles showed significant differences between Actenomys and the subterranean taxa, and no appreciable differences between the former and the generalized Octodon (Table 1, Fig. 2). In contrast, the location of the deltoid process-the insertion site of the M. deltoideus and M. pectoralk-onto the humerus was, in Actenomys, intermediate between those of Octodon and the subterranean species C. australir (Table 1, Fig. 2). The robustness of the Actenomys humerus, as reflected by the quotient anteroposterior width at midshaft/humerus total length, was statistically comparable to that of C. australis, and greater than those of C. talarum and Octodon (Table 1). The length of the olecranon process of the ulna-the in-lever arm of the M. triceps-showed no significant differences between Actenomys and both Octodon and Ctenomys-Spalacopus, i.e. Actenomys showed intermediate values between generalized
l - i e r . ~ I . k'orelimh ineasurrments for drl~nony.s and statistical comparisons with other Octodoiitid tam. \lc;isurcs of hunirrus \\.idth across the cpicond! Ics relatiw t o total humerus length: aiiteroposrrrior I i ~ i m r t ~ i ~ bvidtli a t midshaft rclati\r to tittal liuriicrus Icngth; position or thr drltoid crmt (liirmcru~ hrxd drltoid crest lrngth relative to humrrs total length): length of the olecranon pro< i o t ' i l uhi,i length: uhia cur\ ature (in degrrrs). Data are cxprrssed as mean & confidrncc inrer\.;d KL5"ci .
Prolt~ihilitk-s I /-tests) corrrspond to comparisoris Iwnveen A4c/eiro7rq and rach taxon
0 .2 i i l l . ( l 4 l i = 8
11. I I * 0.112 II = 8
0. k+ * 0.0.5 li = 8
11. 1 5 * 0.0:3
1 4 7 . i i 13.0
li = .I
l i = :I I
'I:WLL 2. Skull nirasuremrnts for .-lrftmory>s. mcl statistical comparisuns \vith other Oc.rodontid t i i u .
lkasurcs of upper and lo\i-cr incisor procumbency (in de<qecs); upper and lo~vcr incisor Irngth rrlati\.e to upper and 1oivt.r diastema length; upper and lo\vrr incisor thickness (length of the aiitrropostcrior
c t i c t n ) ralari~-r to upper and lotv-er diastema Icngth; upprr incisor tvidth (length of the l a t c m l cross-section; rrlativc to upprr diastema length; rostra1 depth relative to upper diasteina Icngtli; upprr riia\tcm,i length relativr to skull tmsilar length; zygomatic breadth rehtivc t o skull Imsilar Ivngth. Data ' I I T r x p r r w d A\ rncati confidciic.e i n t e n d (95 '?/o j. Probabilities (&tests) corrrspond to ( omparisoiis
bct.ri-een .-ictrnorgr.s and other taxon
I I2.O+li.8 II = 3 I
1 :i5.0 * 8. I 1 i = 18
11.32 iO.08 I I = 33
n.cii i 1 1 . 2 5 11= 19
0.1 i & 0.02 11 = '33
0.20 +o.oc I )= 18
0. I6 * 0.02 iI = 3 Y 11.78 i 0.10 1 1 = 2 k
0.s4 FO.02 i l = 24
11.67 *l l . I lR I ! = I I
IOtT.7 li = 2
1-1-3.2 11=2
O.ti3 11 = 2
1 ,IN I/ = 2
0. 19 I / = 2
0.32 i i = 2
0. I 7 11 = 2
ll..59 i i = 2
0.37 i i = 2
0.75 I I = 2
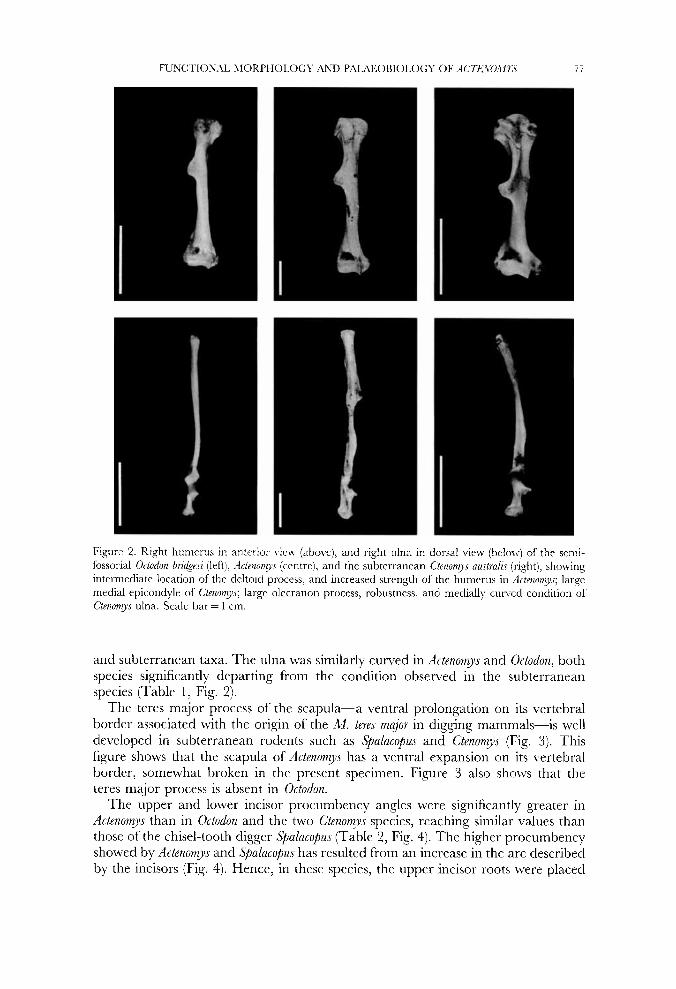
Figure 2. Right humerus in anterior view (above), and right ulna in dorsal \:iew (below) of the semi- fossorial O h d o n bridgrei (left), Actenomys (centre), and the subterranean C'tenorys austra1i.c (right), showing intermediate location of the deltoid process, and increased strength of the humerus in Bct~nolryc; large medial epicondyle of Ctenomyc; large olccranon process, robustness, and medially cuncd condition of Ctenomjs ulna. Scale bar = 1 cm.
and subterranean taxa. The ulna was similarly curved in Actenomys and Octodon, both species significantly departing from the condition observed in the subterranean species (Table 1, Fig. 2).
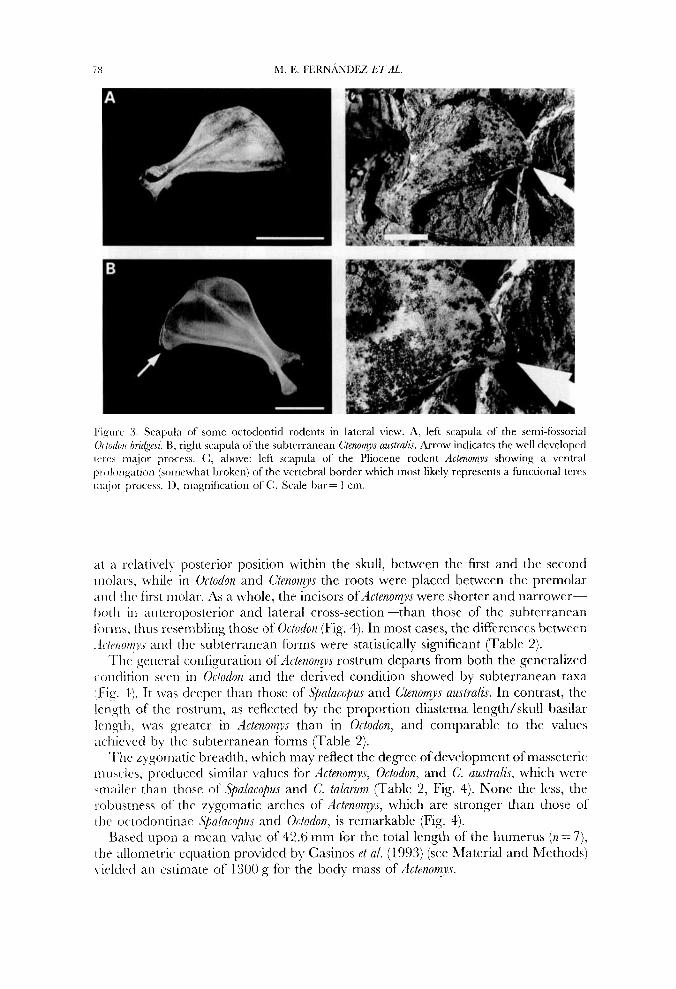
The teres major process of the scapula-a ventral prolongation on its vertebral border associated with the origin of the A4. teres major in digging mammals-is well developed in subterranean rodents such as Spalacopus and Ctenomjis (Fig. 3). This figure shows that the scapula of Actenomys has a ventral expansion on its vertebral border, somewhat broken in the present specimen. Figure 3 also shows that thc teres major process is absent in Octodon.
The upper and lower incisor procumbency angles were significantly greater in Actenomys than in Octodon and the two Ctenomys species, reaching similar values than those of the chisel-tooth digger Spalacopus (Table 2, Fig. 4). The higher procumbency showed by ActenomyJ and Spalacopus has resulted from an increase in the arc described by the incisors (Fig. 4). Hence, in these species, the upper incisor roots were placed
;ti hl. E. FERNANDEZ ILTAL.
l'ieure 3 . Scauula of some octodontid rodents in lateral virw. A. left scauula of the scmi-fossorial 1
Ododotr bridgebi. B, right scapula of the subtrrranean Ctenornys awtralir. Arrow indicates thc well dcveloped [('rf's major process. (:, atiove: lcft scapula of the Pliocene rodcnt Actenomys showing a vrntral p i elongation (somrwhat broken) of thc vertebral border which most likely represents a functional tcrcs i>iaior process. D, magnification of C. Scale bar= 1 cm.
at ii relatively posterior position within the skull, between the first and the second molars, while in Octodon and Ctenomys the roots were placed between the prcmolar anti tlir first niolar. As a whole, the incisors ofdctenomys were shorter arid narrower ~
1)otI i in anteropostcrior and lateral cross-section-than those of the subterranean limns, thus rrsembling those of Octudotz (Fig. 4). In most cases, the differences between . ic/cnom)~,r arid the subterranean forms were statistically significant (Table 2).
The general configuration of Actmongs rostrum departs from both the generalized condition seen in Octodon and the derived condition showed by subterranean taxa 1 Fig. 4). It \vas deeper than those of Spalacopus and Ctenomys australis. In contrast, the Icngth of the rostrum, as reflected by thc proportion diastema length/skull basilar Iciigth, \\as greater in Actenomys than in Octodon, and comparable to the values nchic\d by thc subterranean forms (Table 2j.
'The qgomatic breadth, which may reflect the degree of development of masseteric iiiusclcs, produced similar values for Actenomys, Octodon, and C. australis, which were mid1t.r than those of Spalacofm and 0". talarum (Table 2 , Fig. 4). None the less, the ro1)ustiiess of the zygomatic archcs of Actenomys, which are stronger than those of t Iic octodoiitinac Sjia1acopu.r and Oclodon, is remarkablc (Fig. 4).
Based upon a mcan value of 42.6 mm for the total length of the humerus (n = 7), the allometric cquation provided by Casinos ct al. (1993) (see Material and Methods) viclded an cstiniate of 1300 g for the body mass of Acten0my.r.

79 FUNCTIONAL MOKPHOLOGY AND PAIAEOBIOLOGY OF A CTFNOMYS
Figure 4. Skull of some octodontid taxa in lateral (A D), and dorsal (E-H) \iews. A & E, Octodon br&esi. B & F, Achomys. C & G, Spahapus qanus. D & H, Ctenomys australis. Note the high incisor procumbeney of both Aclenmys and the chisel-tooth digp- Spahopw, flaring (Spalaoopus) and strong (Chmys, A c h m z s ) zygornatic arches. Scale bar= 1 cm.

1 1 t. I'FKN \\L)1:7 br -1L
- 6-6 25
6.25-6.55 w 2 2 T! 6.85-7.30 c - 7.30-7.80 E, & 7.80-8.50
8.50-8.70 Y 2 8.90-10.2 t 3 10.2-10.55
& 10.55-11.4 0 20 40 60 80 100
Percentage of clay, loam and sand
I.'igurc. i. G r a i i i - s k of thc Plioccnr deposits containing burrobvs and fossil remains of d d m o i r y ~ . Pla) a >.in (:ado\. Partido tlr Gcncral Puryrrrdbn. Bucnos Arcs.
PalaeoenzdronmPntal anabsis
The studied section, Los Acantilados, is a 11 meter thick scquence composed o f silty to cla)-e)- silty deposits of volcanoclastic composition, deeply modified ti!.
pcdogrnesis. Stratigraphically, the section belongs to the Playa Sari Carlos Al- IoFormation (Zaratc, 1989) which approximately corresponds to the lower part of thc (:hapadmala1 Formation (Kraglievich, 1952). The age of this unit is betwecn 4 and S h I ~ - r (Ziirate et al., in press).
Tlic three major palaeosoil levels exposed at Los Acantilados consist of thick Kt horizons Lvhich grade into more massive deposits interpreted as overthickened Iiorizons. Nunicrous remains ofActenomys, mostly found in the burrows, wcrc collected at this sitr.
Thc palaeosoils were predominantly non calcareous; the presence of nodules of C:aC:O, 'tosca'- around the palaeocave walls, and some calcareous accumulations in R t horizons around fossil root channels are related to groundwater circulation. 'l'hr occurrenc.c of well developed Bt horizons with continuous and thick cutans suggcsts that the illu\iation was probably very strong. A great abundancc of hioturbations, c a w and root microchannels was observed all along the profile (for ;i niorc detailed description of thr palaeosoils see Fernandez, 1997).
Tht- predominant clay minerals are smectites (Carnilion, pers. comm.). 'I'hc pu~ulonietric analysis showed that thc clay + loam fraction, i.e. particles <0.062 mm. \\.as abundant -more than 60% of the total sedimen-in all samples (Fig. 5). This pcm'cntagc might underestimate the amount of fine material due to the partick aggrcptioii oxides reported for these sediments (Mormeneo, Carnilion Oi Iasi. I !Nfj'j.
In this section \vc assess the available morphological and stratigraphic e\idencc tc 1 ascertain the degrcc of specialization for digging in dctmomys, also providiiig a

probable picture of its ecolo\q and behaiiour. \Ye hope that the limitations of inferring function (hehaviour) from morphological data only (Lauder, 1995) are attenuated by our integration of morphological and palaeoenvironmental data.
Diging potentialities of Actenomys
On the basis of the current knowledge concerning living rodent taxa, it appears that there is a diverse array of morphological specialization for digging, ranging from mcrely semi-fossorial, burrowing species which, though occupying elaborate burrows, spend a considerable portion of their life above ground (i.e. Octodun sp., qmpanoctomjs barrerae, Ortonys mimax, Spermophilus sp., Qnomj~s hduvicianus) to highly specialized Subterranean species (Spalax sp., Bathiergus sp.) which are virtually unable to live above ground. The need to regularly break and transport resistant materials requires the use of digging tools, such as incisors, claws, head, or a combination of these. Moreover, an effective production and transmission of force demands a greater development of associated muscles, and a change in the relationship between in- lever to out-lever arms to maximize forces (Hildebrand, 1985). Departing from a generalized epigeous form, these structural and mechanical modifications lead to a general strengthening of the musculoskcletal compoiients of the skull, forelimbs, and pectoral girdle.
As a whole, our results indicate that Aitctenomq,s occupics an intermediate position between Octodun, a generalized seini-fossorial rodent, and Ctenomq,s-S~alacopus, two highly specialized subterranean forms. The ctenomyincs are considered a late Miocene branch of the family Octodontidae with striking burrowing adaptations (Patterson & Pascual, 1968; Reig et nl., 1990). In this context, the forelimb and skull characters studied in Octodon could be considered as plesiomorphic -as compared to those of Ctenonys and/or the octodontine Spalacopus-not departing from fossil octodontines such as dbalosia which lack burrowing adaptations iReig & Quintana, 199 1).
Forelimb structure The long bones of diggers are subject to loads imposed by both the action of
strong extensor and flexor muscles, and the soil resistance. Digging species have shorter and thicker long bones than epigeous forms which provide the strength to resist force without breakage (Casinos et al., 1993). The humerus of Actenomys was at least as sturdy as those of subterranean taxa (Table 1, Fig. 2). A flaring medial epicondyle greatly increases the area of origin of carpal and digital flexors, also indicating the degree of development of these muscles. In respect of this, Actenomys showed no substantial differences with Octodon, both genera differing from the condition seen in Subterranean taxa. The deltoid process is the site of insertion of the M. deltoideus, a flexor muscle of the forearm. In subterranean rodents, a well developed deltoid process concurs with a distal position of this process. l h i s last attribute enlarges the in-lever arm of hl. deltoideus, thus increasing out-forces exerted by this muscle. Notably, the deltoid process of Actenomys showed an inter- mediate position between non-subterranean and subterranean taxa (Table 1 , Figs 2 and 6). A similar trend was found in the structure of the ulna. The olecranon process is the site of insertion, and the in-lever arm of extensor muscles of the forearm, such
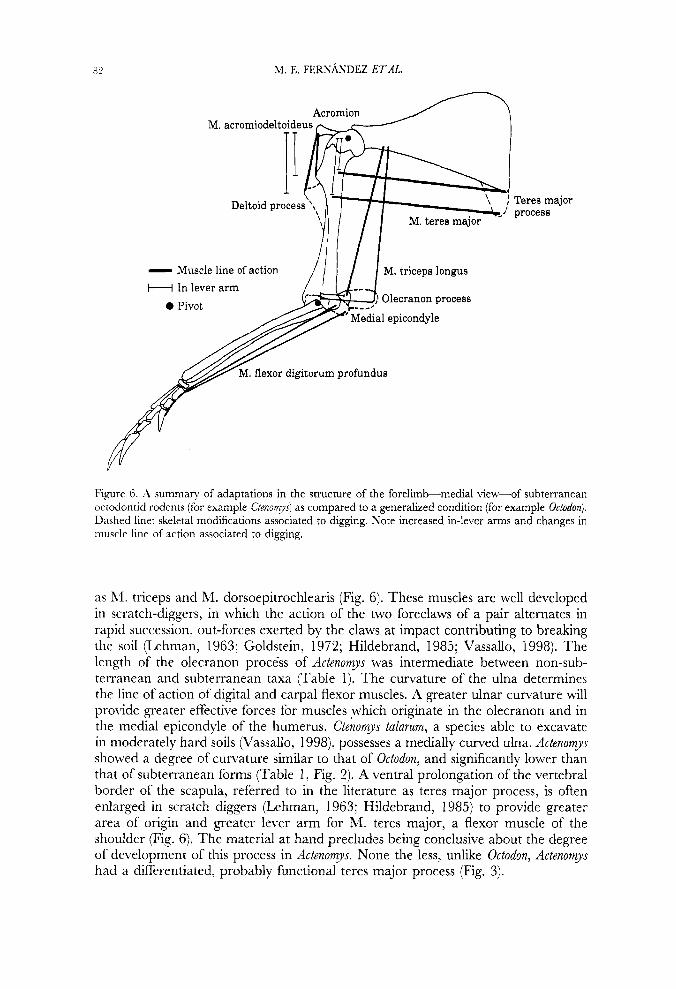
82 M. E. FERNANDEZ ETAL.
Acromion A
\ / Teresmajor Deltoid process ', M. teres major
- Muscle line of action M. triceps longus
Olecranon process
Medial epicondyle
Figure 6. A summary of adaptations in the structure of the forelimb-medial view-of subterranean octodontid rodents (for example Ctenomys) as compared to a generalized condition (for example O C ~ Q ~ O F Z ) . Dashed line: skeletal modifications associated to digging. Note increased in-lever arms and changes in muscle line of action associated to digging.
as hl. triceps and M. dorsoepitrochlearis (Fig. 6). These muscles are well developed in scratch-diggers, in which the action of the two foreclaws of a pair alternates in rapid succession, out-forces exerted by the claws at impact contributing to breaking the soil (Lehnian, 1963; Goldstein, 1972; Hildebrand, 1985; Vassallo, 1998). The length of the olecranon process of Actenomys was intermediate between non-sub- terranean and subterranean taxa (Table 1). The curvature of the ulna determines the line of action of digital and carpal flexor muscles. A greater ulnar curvature will provide greater effective forces for muscles which originate in the olecranon and in the medial epicondyle of the humerus. Ctenomys talarum, a species able to excavate in moderately hard soils (Vassallo, 1998), possesses a medially curved ulna. Actenomys showed a degree of curvature similar to that of Octodon, and significantly lower than that of subterranean forms (Table 1, Fig. 2). A ventral prolongation of the vertebral border of the scapula, referred to in the literature as teres major process, is often enlarged in scratch diggers (Lehman, 1963; Hildebrand, 1985) to provide greater area of origin and greater lever arm for M. teres major, a flexor muscle of the shoulder (Fig. 6). The material at hand precludes being conclusive about the degree of development of this process in Actenomys. None the less, unlike Octodon, Actenornys had a differentiated, probably functional teres major process (Fig. 3).

Skull rtructure The skull of chisel tooth diggers is strongly constructed. Incisors, as digging tools,
are usually procumbent thus providing an adequate angle of attack against the soil. The jaw musculature is well developed to provide greater out-forces at the incisor tips. Consequently, the zygomatic arches are powerful and flaring to provide broader sites of origin for masseter muscles. Also, the sagittal and nuchal crest, where M. temporalis originates, are well marked.
As in the structure of the forelimbs, the skull of Actenomys shows a disparate set of functional morphological features that reinforces the idea that this species was ecologically different from both semi-fossorial (Octodon) and subterranean forms (Cten- omyJ-Spalacopus). Although the skull of Actenomys, particularly the rostrum, appears to be strong, and sagittal and nuchal crests are well marked, the zygomatic arches are as flaring as those of the generalized Octodon (Table 2). Further, the masseteric crest of the lower jaw is not as flaring as in the subterranean Ctenomys. Hence, we can infer that the masseter muscle complex of Actenomys was moderately well developed.
Upper and lower incisors were extremely procumbent, as many highly specialized chisel-tooth digging species (Table 2, Fig. 4). The upper incisor roots were placed at a relatively posterior position within the skull, between molars 1 and 2, while in Ctenomys australis, a predominantly scratch digger, they were placed between the premolar and the molar 1. As noted by Landry (1957, quoted in Lessa, 1990) this is needed to resist the loads caused by the soil reaction forces on the incisor tips, which are located anteriorly in procumbent forms. The incisors of Actenomys were proportionally narrow, both in lateral and antero posterior cross section. Hencc, as digging tools, they probably exert more pressure against the soil than do broad incisors. This could have been a crucial factor in loosening hard soils such as those containing Actenolny~ burrows. Justo, Bozzolo & Dc Santis (1995) showed that the structure of the external enamel layer of the incisors of Actenomyr allows their use as digging tools.
Another attribute of dctenomys incisors is that they were markedly shortened, as compared with both subterranean and non subterranean octodontids (Fig. 4). However, as noted previously, they show a greater degree of procumbency. Because the combination of these two features respectively implies a reduction of the out- lever arm of jaw masseter muscler (Lessa, 1990) thus increasing out-forces for breaking the soil, and the maintenance of a good angle of attack against the soil, dctenomys incisors probably had the potential for an effectivc chisel-tooth digging behaviour. The action of sliding the mandible forward on the skull to engage the incisors is accentuated in chisel-tooth diggers (Hildebrand, 1985). As noted by Verzi (1 994), this action could be facilitated in Actenomys by the presumably anteriorly directed fibres of the masseter medialis muscle. This author also pointed out that ,4ctenom>ls has a high zygomatic index which indicates the presence of non-reduced eyeballs. Although this trait could be interpreted as denoting significant epigeous habits (Pearson, 1984), it should be noted that Ctenomys, a specialized subterranean form, has non-reduced eyes and good vision (Vassallo, pers. obs.).
Reconstruction of Actenomys habitat
The features described for Los Acantilados section allow us to infer that Actenomys lived in an environment with moderate to high primary productivity. Given the

closc association hetrveen water supply (precipitation) and plant primary producti\%! in present ecosystems (Barbour, Burk 8r Pitts, 1987; Ricklefi, 1993). the a11~n-c characterization is based upon traits \vliich denote good water a\-ailahilit)- throughout the. !.car. These traits are, mainly, the thickness of B horizons and the al~undancc. of illu\.iation features, both indicating excessive moisture in tlie soil during long periods of‘ tirnr (Rettallack, 1988; 199 1). ‘ h r lack of CaCO’, accumulationc rrlated t o soil formation suggests an en\.ironmcnt dominated leaching proces gr;idual houndarics between the different horizons and palacosoils indicate a relativcl!- lo\\ rate of scdimrntation on the land surhce: likelj. covered by enough \.cgctation to hamper drastic - hydric and/or aeolian--erosion.
’lhc evidence of good water availability coupled avith a high densit)- of grass root t r ; ic~s permits tlie inference that L4c.teriom_~1.r lived in a tall grass prairie, or an e to \\ i t h rcotonal \.rgetation as a sa\rannah, both characterized by high plant produ .l’his picturc. is in agreement lvith the em-ironmental reconstruction propc)secl 11) Pasciial ( 1 984; ~vhich is based upon fossil Lvtebratc assemblages. For the I’liocetic. this au thor suggests a park-sa\minah en\ironment similar to the present Cliaco region. although much ivetter. 1 his inference is supported ti>- the high hiomass of .r-azing hcrbi\.orous forms, and tlie presence of some key taxa such as anteaters
l~~~~mecoptiagidac) and cariamid birds. This vision contrasts \.\-it11 that o f C h i l i & ‘l’onni ( 1993) It-liich, based upon the remarkable number of mammal caves of di\.erst. size found in the Plioccrie deposits exposed along the sea-cliffi south of Alar clcl Plata. proposes arid and/or cold conditions for tlie Pliocene. Under arid cn\.iroiinicntal conditions, aridisoils are the dominant soils whereas the Pliocene palaeosoils indicatc a niorc humid enxironment and do not shon. morphological features attrihutahl(- to aridisoils. Although the evolutionary origin of fossorial bchaviour in mammals has been associated with the expansion of an open country biota from tlic hliocenc (Nc\-o, 1 Y79), the sul~erranean and burrower mammals inhabit a great rarigc of’ ciivirc)nriicnts anti climate conditions (e.g. 5liller, 1964; IlcKah: 15166: Best, 1973: \\‘alkcr. 197.7; Nevo ei al., 1979; Kedford Sr Eisenbcrg, 1992). Even fully subtcrrancan rodents she\\. 1iight.r population deiisities i n more producti\,e en\i-orinients t h i i t i
arid and cold oncs (Patton 8r Rrylski, 1987; Hadly: 1997). The texture o f the palaeosoils \\.here Llctenoryis li\.ed indicate that they \\.ere hard
and highly cohcsi\-e (Fig. 5). This situation contrasts with that o h s e n d in li\.ing spccics of tlie subfamily Ctenomyinae, \vliich prefer friable to moderately liarcl wils (Coiitreras, 1973) \\pith variable but lo\v 11roportions of loam and clay (hlalizia rl nl.. 109 1 j . T l i ~ soil hardness imposes rele\,ant bioenergetic constraints iii tcrms of’ Im-roiving cost. surely shaping the ecological niche occupied by A-ictenonii:y (see bclou.;.
‘l’hc burroivs of .Jctp?m!j:r are very ahndaiit in the coastal cliffs of Partido dc ( h r x d Puc) rredon, Buerios ilires province. outcropping at different Ic\.cls in th(, stratigraphic sequence corresponding to the Pliocene (Fig. 7). ‘1 set of\vell-preser\-eti clmients suggests that the hurroi3vs ivere possibly \w-y elaborate, 1 3 cm in diametrr and up to 5 m in length. Some tunnels cnded in a chamber and had lateral t i ra ic l ie~ i(;cnise. 1989). Some attributes of A4ctenonp burro\vs resemble tliosc studied in the li\.ing suhterrancan species Ctemn1:r talamni (Scognamillo, 1993). For esaniplc. a great proportion of the branching angles of d c t e m y s aiid C‘t~nony s liurroi\-s \\-err nc.arly orthogonal to the originating main tunnel. Howc\.cr. a recent study 011
~ ) , ~ / ~ ~ ~ ~ / ~ u f f o ~ r / i , , ~ bm.i.me, a srmi-fossorial octodontid rodent \shich forages a l i o \ ~ ground. sIio\\.ed that 1.11~ lateral branches of an excavated burrow s!’stcm \+-ere also ncarl!. a t riglit angles (sre fig. 4 in yjeda et a/.> 1996). Tunnel expansions. \i-Iiich posihly
r 7
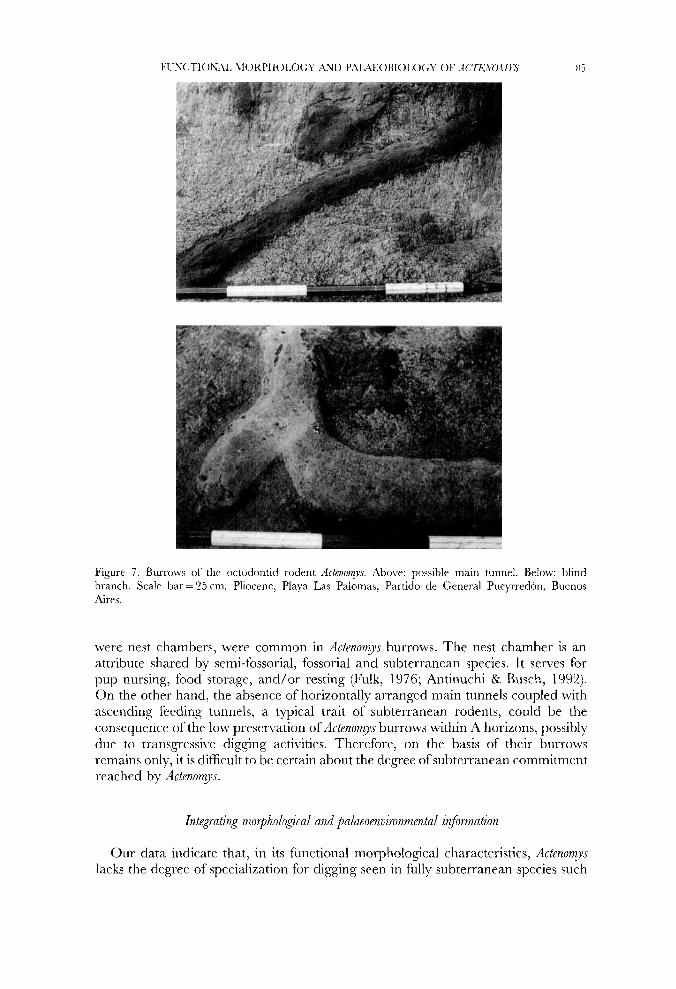
Figure 7. Burrows of thc octodontid rodent Actenonys. Above: possible main tunnel. Below: blind branch. Scale bar = 25 cm. Pliocene, Playa Las Palomas, Partido de General Pueyrredbn, Buenos Aires.
were nest chambers, were common in Actenomys burrows. The nest chamber is an attribute shared by semi-fossorial, fossorial and subterranean species. It serves for pup nursing, food storage, and/or resting (Fulk, 1976; Antinuchi & Busch, 1992). On the other hand, the absence of horizontally arranged main tunnels coupled with ascending feeding tunnels, a typical trait of subterranean rodents, could be the consequence of the low preservation of ActenomyJ burrows within A horizons, possibly due to transgressi\re digging activities. Therefore, on the basis of their burrows remains only, it is difficult to be ccrtain about the degree of subterranean commitment reached by Actenom_ys.
Integrating morphological and palaeoenuironmental information
Our data indicate that, in its functional morphological characteristics, Actenomjjs lacks the degree of specialization for digging seen in fully subterranean species such
Family Octodontidae Octodon Octomys I ;:::rial Orfrdontomys ~mpanoc tomys (surface dwelling Aconaemys (fossonal) Spalampus (subterranean) Ctvnomys (subterrancan~
galleries in rocks and roots)
I Eucehhorus I hn
1
2
5
1:
, I (subte;ranean)
Parnctenonivs l subterranean^ I Abalusia
(lacks fossorial adaptations1 1 praectenomys I (subterranean,) , ~
I . , I
I
1
I , _/'
I.
/ _ ,_ - - - ' - Subfamily Ctenomyinae .~ . , ._ Subfamily'. . . . . .
Octodontinae . I.._
, - A I Octodontinae-Ctenoinylnae , dichotomy
, - -
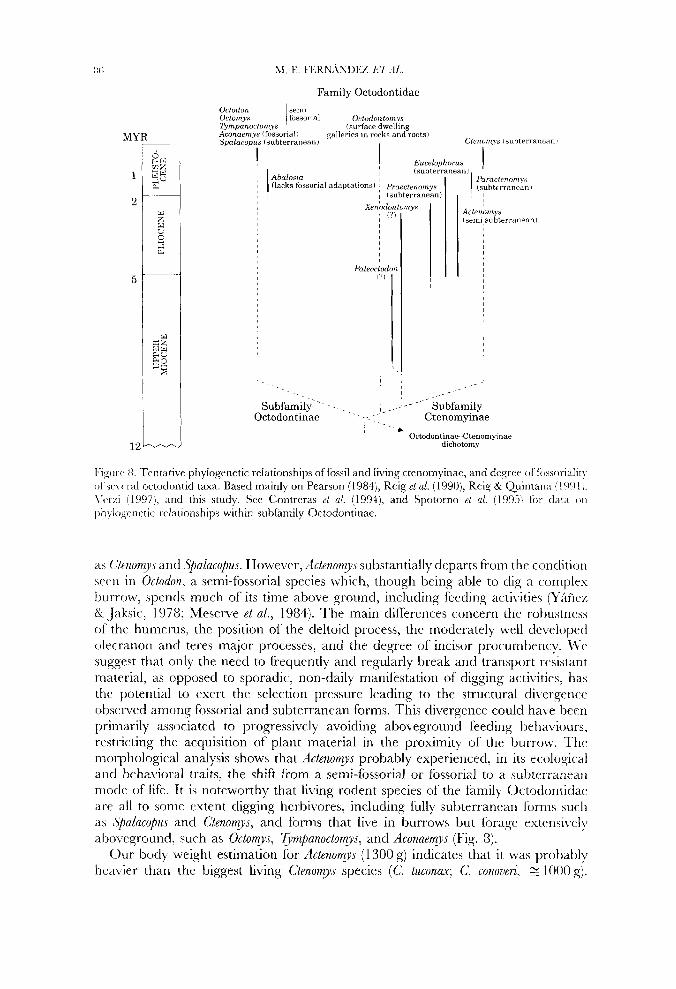
I-iyurc 8. Tcntative phylogenetic relationships of fossil and living ctcnomyinac, and dcaree oi'fiissorialir~ ol'.;(~\crul octodontid taxa. Based mainly on Pearson (19843, Reig et al. (1990), Rrig Cyr Quiiitana ;I991 ,. 1't.rxi I 1997). arid this study. See Contreras et a/. (1994), and Spotorno ~t a/. (19'35) for c l a ~ o i l
~h\.log(c:netic. rclationships within subfamily Octodontinae.
a s C'/enorrv., and +akucopu.r. However, Actenomys substantially departs from thr condition secn in Octodon, a semi-fossorial species which, though being able to dig a complex l>urrow, spends much of its time above ground, including feeding activities (YBficz 8r <Jaksic, 197U; hicscrve et al., 1984). The main differences concern thc rohus~ncss of' thc humcrus, the position of the deltoid process, the moderately well developed olecranon and teres major processes, and the degree of incisor procumhency. \ 2 ' c , suggest that only the need to frequently and re,gularly break and transport resistant material, as opposed to sporadic, non-daily manifestation of digging activities, has thc potential to exert the selection pressure leading to the structural divergencc ohscrved among fossorial and subterranean forms. This divergence could have hccn priniarily associated to progressively avoiding aboveground feeding hehaviours. rcstricting the acquisition of plant material in the proximity of the burrow. 7'hc morphological analysis shows that Actenomys probably experienced, in its ecological and behavioral traits, the shift from a semi-fossorial or fossorial to a suhterranran niodc of lifc. It is noteworthy that living rodent species of the family Octodontidac are all to somc extent digging hcrhivores, including fully subterranean forms such as LSpalacupu.s and C'tenomys, and forms that live in burrows but forage extensi\:cl)- atxnyqound, such as Octomys, 7ympanoctomys, and Aconaemys (Fig. 8).
Our body weight estimation for ilctenomys (1 300 g) indicates that it was probab1~- hcavier than the biggest living Ctenomy.r species (C. tuconax; C. conoueri, 2 1000 g).

FUNCTIONAL MORPHOLOGY AND PAALAEOBIOLOGY OF ACTENOMYS 87
Vleck (1 979, 198 1) showed that the energy cost of burrowing is directly proportional to the mass of soil removed and thus to the cross-sectional area of the burrow. As the cross-sectional area is proportional to the square of radius, burrowing cost is about proportional to the square root of body mass. This attribute, coupled with the data on palaeosoil hardness and granulometry, is of importance when inferring bioenergetic constraints on ecology and behaviour. Within living fully subterranean rodent taxa, the biggest representatives live in very friable, often sandy soils which demand a lower cost of burrowing (Miller, 1964; Walker, 1975; Nevo, 1979; Malizia et al., 1991). Our vision of Actenomys as a ‘transitional’ form to a subterranean mode of life fits with available information on body mass and soil hardness. Because of the cohesiveness of the clayey silty palaeosoils of its habitat, Actenomys must have been seriously limited to behave as a fully subterranean rodent, i.e. a rodent in which food acquisition always has an associated cost of excavation.
Another fossil ctenomyine, Eucelophoms, coexisted with Actenomys throughout nearly all the Pliocene. Eucelophorus, which has been characterized as a highly specialized chisel-tooth digger because of its strong, procumbent incisors (Reig & Quintana, 1992) is more frequent in the fossil record after the extinction of Actenomys, from upper Pliocene-lower Pleistocene (see Reig & Quintana, 1992 for a detailed chron- ostratigraphic record of Plio-Pleistocene taxa). The functional morphological evidence suggests Eucelophoms was possibly a fully subterranean form. However, any hypothesis about competitive replacement should be approached with caution because the species were probably ecologically segregated.
In conclusion, the mode of life ofActenomys had certain unique aspects, undoubtedly related to regional environmental conditions, and differing from that of both semi- fossorial and fully subterranean rodents. This interpretation agrees with the notion that species actively construct their ecological niches, rather than occupying pre- existing ones (Odling-Smee, Laland & Feldman, 1996). Our data support Reig’s thesis that the Pliocene-early Pleistocene ctenomyine cladogenesis give rise to different genera which represent different alternative adaptations to excavation in burrowing rodents (Reig, 1989; Reig at al., 1990). On the other hand, the middle Pleistocene- Recent diversification of Ctenomys species may have been due to stochastic processes of fixation of chromosomal rearrangements (Ortells, 1995, but see Cook & Lessa, 1998, for a critical review of rates of speciation in Ctenomys) “once an optimal burrowing adaptation was attained” (Reig & Quintana, 1992). However, the lifestyles of early ctenomyines such as Actenomys should no longer be considered as suboptimal approaches to a subterranean mode of life, more or less adapted to a unique underground niche. While lacking the degree of morphological specializations seen in fossil and living fully subterranean rodents, Actenomys was distinctively adapted to a particular mode of life and habitat conditions.
ACKNOWLEDGEMENTS
We express our thanks to Orlando Scaglia, Alejandro Dondas (Museo Municipal de Ciencias Naturales ‘Lorenzo Scaglia’, Mar del Plata, Argentina), Diego Verzi and Hugo Lbpez (Museo de La Plata, La Plata, Argentina) for permiting access to the collections. Enrique Lessa made valuable comments that improved the manu- script. The photographic work was done by Mr Abel Beruti, Department of

I'hotograph> . University of Xlar dcl Plata. This rcsearch \\-as supported hy thc rni\-c.rsiiy of hlai. dcl Plata and CONICET.

F L x c : ' r i m . - x i i ( )RPH( )LOGY .m) I~.;\WEOBIOLOGY 01: A LTE \QIIIS 8 9
Hildebrand M. 1988. dnabsiJ OJ' I'ertPbrate Structure. 3rd edition. New York: .John !Viley PC Sons, Inc. Hodara K, Suarez OV, Kravetz FO. 1997. Nesting and digging beha\.ior in two rodent species
(Akodon azarae and Caloirys Iaucha) undcr laboratory and field conditions. zeitschnj&r Saugetierkunde
Just0 ER, Bozzolo LE, De Santis LJM. 1995. AIicrostructurc of thc enamcl of the incisors of somc ctrnomyid and octodontid rodrnts (Kodentia, Chiomorpha). .tfastozoolo@ w,Veotropical 2: 43-5 1.
Kraglievich JL. 1952. El pcrlil geolbgico de Chapadnialal y hliramar, provincia de Buenos Aires, resumen preliminar. Retirta del 2\Iuseo de C imias .laturale.~ -tlar del Plata 1: 8-37,
Landry SO. 1957. Factors affecting the procumbency of upper incisors. 30urnal of ~ t h n m a l o g y 38: 223-234.
Lauder GV. 1995. On the inference of function from structurr. In: l'homason JJ, eds. Functional niorpholoa in tiertebrate pale onto log>^. Cambridge: Cambridge University Press, 1-1 8.
Lehmann WH. 1963. The forelimb architecture of some fossorial rodents. 3ournal of Morphology 113: 59-76.
Lessa EP. 1990. hlorphological evolution of subterranean mammals: integrating structural, functional, and ecological perspectives. In: Nevo E, Reig OA, eds. EL'olution of subterranean mammals at the oganisnlal and molecular lei'elr. Sew York: Allan Liss, 21 1-230.
Llanos AC, Crespo JA. 1952. Ecologia de la vizcacha (Lapostomus maxiniu.r marimus Blainv.) en cl nordeste de la provincia dc Entre Rios. Rezirta de Inaestigaciones Agricola.\ 6: 289-378.
Malizia AI, Vassallo AI, Busch C. 1991. Population and habitat characteristics of two sympatric species of (:tmong,c (Rodcntia: Octodontidar). Actu 7lzerioIogica 36: 87-94.
McNab BK. 1966. The metabolism of fossorial rodents: a study of con\wgrnce. Ecolo,> 47: 7 12-733. Meserve PL, Martin RE, Rodriguez J. 1984. Comparative rcology of the caviomorph rodent
Ortodon degus in two Chilean hlcditrrranean-type communities. Rrr i~ ta (:hilena de Historia Aatural 57: 79-89.
Miller RS. 1964. Ecology and distribution of pocket gophers (Gcomyidae) in Colorado. EcoloB 45:
Mormeneo ML, Camilion MC, Iasi RR. 1996. Composicibn de 10s agregados no dispersables de
Nevo E. 1979. Adaptivc convergence and divcrgcnce of subtcrrauean mammals. Annual Revlea of
Nevo E, Guttman R, Haber M, Erez E. 1979. Habitat selcction in emlving mole rats. Oecologza
Odling-Smee FJ, Laland KN, Feldman MW. 1996. Niche construction. American ,Vuturalict 147: 641 648.
Ojeda RA, Gonnet JM, Borghi CE, Giannoni SM, Campos CM, Diaz GB. 1996. Ecological obscnations of thc rcd vizcacha rat, 7ympanactomys barreme, in dcsert habitats of Argentina. A\la.rtozoologia .<Veotropiral 3: 183- 192.
Ortells MO. 1995. Phylogcnetic analysis of G-banded karyotypes among the South American subterranean rodrnts of genus Ctenomys (Caviomorpha: Octodontidae), with special reference to chromosomal evolution and speciation. Biological Journal o f the LinnPan Society 54: 43-70.
Pascual R. 1984. Late Tertiary mammals of southern South America as indicators of climatic deterioration. In: Rabassa J: ed. Quaternary of South AmPrica and Antarctic Peninsula 2: 1-30,
Patterson B, Pascual R. 1968. The fossil mammal fauna of South America. Quarterb Reziew ofBiolo~ 43: 409-43 1.
Patton JL, Brylski PV. 1987. Pockct gophcrs in alfalfa fields: causes and consequences of habitat- related body size variation. dnzerican .&furahst 130 493-506.
Pearson OP. 1984. Taxonomy and natural history of some fossorial rodents of Patagonia, southcrn Argentina. Journal of ~ o o l o ~ q 202: 225 -237.
Quintana CA. 1994. Sistematica y anatomia funcional del roedor ctenomyinae Pruectenomy.5 (Cavio- morpha: Octodontidae) del Plioceno dc Bolivia. Rrcirta Zcnica de YPFB 15: 175 -185.
Redford KH, Eisenberg JF. 1992. L21ammal~i of the uVeotmpicJ. Chicago: The University of Chicago Press.
Reig OA. 1958. Notas para una actualizacibn del conocimiento dc la fauna de la Formacion Chapadmalal. Lista Faunistica prcliminar. dcta Geoldgica Lilloana 2: 241--253.
Reig OA. 1970. Ecological notes on thc fossorial octodont rodcnt Spalacops cyanu.r (h.Iolina). Journal o f421am?nalo~ 51: 592 60 I .
Reig OA. 1989. Karyotypic repatterning as a triggering hctor in cascs of explosive speciation. In:
62: 23-29.
256-2 7 2.
sedimcntos pampcanos. Rerisia drl Aluseo d p la Plata (nueixa Aerie) 9: 135-148.
E c o l o ~ and Sq'stematic 10: 269-308.
43: 125-138.

90 M. E. F E R N ~ D E Z ETAL.
Fontdevilla A, ed. Evolutionary dynamics of transient, unstable populations. Berlin: Springer-Verlag,
Reig OA, Busch C, Contreras JR, Ortells MO. 1990. An overview of evolution, systematics, population biology, cytogenetics, molecular biology, and speciation in Ctenomys. In: Kevo E, Reig OA, eds. Evolution ofsubtenanean mammals at the oqanismal and molecular levels. New York Allan Liss, 7 1-96.
Reig OA, Contreras JR, Piantanida M. 1965. Contribucion a la elucidacion de las entidades del g h e r o Ctenomys (Rodentia, Octodontidae) 1. Relaciones de parentesco entre muestras de ocho poblaciones de tuco-tucos inferidas del estudio estadistico de variables del fenotipo, y su correlacion con las caracteristicas del cariotipo. Contribuciones Cientgcas, Facultad de Ciencim Exactas y Naturakr, Serie ~ o o l o g h 2: 301-352.
Reig OA, Quintana CA. 1991. A new genus of fossil octodontine rodent from the early Pleistocene of Argentina. Journal of mamma lo^ 72: 292-299.
Reig OA, Quintana CA. 1992. Fossil ctenomyine rodents of the genus Eucelophoms (Caviomorpha: Octodontidae) from the Pliocene and early Pleistocene of Argentina. Ameghiniana 29: 363-380.
Retallack GJ. 1988. Field recognition of paleosols. Geological Sociep ofAmerica, Special Paper 216: 1-20. Retallack GJ. 1991. Soils ofthe Past. An Introduction to Paleopedology. New York: Harper Collins.
Ricklefs RE. 1993. The Economy ofJVature. New York WH Freeman. Scheck SH, Fleharty ED. 1980. Subterranean behavior of the adult thirteen-lined ground squirrel
(Spennophilus tridecemlineatus), The American Midland Naturalist 103: 19 1-1 95. Scognamillo D. 1993. Estructura de las cuevas de Actenomys (Rodentia: Octodontidae) de la
aloformacion Playa San Carlos, Plioceno Tardio (Barranca de Los Lobos, Pdo. de Gral. Pueyrredon): significado paleoecologico y estratigrafico. Unpublished Tesis de Grado, Universidad Nacional de Mar del Plata.
Spotorno AE, Walker LI, Contreras LC, Torres JC, Fernandez-Donoso R, Barrios MS, Pincheira J. 1995. Chromosome divergence of Octodon lunatus and Abrocoma bennetti and the origins of Octodontoidea (Rodentia: Histricognathi). Revista Chilena de Historia Natural 68: 227-239.
Vassallo AI. 1998. Functional morphology, comparative behaviour, and adaptation in two sympatric subterranean rodents genus Ctenomys (Caviomorpha: Octodontidae). Journal o f < o o l o ~ 244: 41 5-427.
Verzi D. 1994. Origen y evolucion de 10s Ctenomyinae (Rodentia: Octodontidae): un analisis de la morfologia craneodentaria. Unpublished Tesis Doctoral, Universidad Nacional de La Plata.
Verzi D. 1997. Origin and evolution of the ctenomyinae rodents (Caviomorpha, Octodontidae). Abstracts of thr 7th International Theriological Congress: 365.
Verzi D, Montalvo C, Vucetich MG. 1991. Nuevos restos de Xenodontomys szmpsonz Kraglievich y la sistematica de 10s mas antiguos Ctenomyinae (Rodentia: Octodontidae). Ameghiniana 28: 325-33 1 .
Vleck D. 1979. The energy cost of burrowing by the pocket gopher llomomys bottae. Physiolopzcal
Vleck D. 1981. Burrow structure and foraging costs in the fossorial rodent, Thomomys bottae. Oecologia
Walker EP. 1975. Mammals ofthe World, 3rd ed. Baltimore: Johns Hopkins. YGez J, Jaksic F. 1978. Historia natural de Octodon degus (Molina) (Rodentia: Octodontidae). Museo
Zar JH. 1984. Biostatistical anahsis. 2nd ed. Englewood Cliffs: Prentice Hall. Zkate MA. 1989. Estratigrafia y geologia del Cenozoic0 tardio en 10s acantilados marinos com-
prendidos entre Playa San Carlos y Arroyo Chapadmalal, Pdo. de Gral. Pueyrredon, provincia de Buenos Aires. Unpublished Tesis Doctoral, Universidad Nacional de La Plata.
Zrirate MA, Schultz P, King J, Hames W. 1998. Geocronologia y paleoambientes del Plioceno de Chapadmalal (Bs. As.) e implicancias bioestratigraficas. VII Congreso Agentino de Paleontologia y RioestratigTaJa (in press).
246-289.
<OO~OQ 52: 1'22-136.
(Berlin) 49: 391-396.
Nmional de Historia Natural, Publicacidn Ocasional 27: 3-1 1.
Copyright © 2022 FDOKUMEN




















