
Functional endothelial progenitor cells selectively recruit neurovascular protective...
13
REGENERATIVE MEDICINE Functional Endothelial Progenitor Cells Selectively Recruit Neurovascular Protective Monocyte-Derived F4/80 1 /Ly6c 1 Macrophages in a Mouse Model of Retinal Degeneration SHINICHI FUKUDA, a,b MASUMI NAGANO, a,c TOSHIHARU YAMASHITA, a KENICHI KIMURA, a IKKI TSUBOI, a GEORGINA SALAZAR, a SHINJI UENO, d MINEO KONDO, e TILO KUNATH, c TETSURO OSHIKA, b OSAMU OHNEDA a a Department of Regenerative Medicine and Stem Cell Biology, Graduate School of Comprehensive Human Science, and b Department of Ophthalmology, Graduate School of Comprehensive Human Science, University of Tsukuba, Tsukuba, Japan; c MRC Centre for Regenerative Medicine, Institute for Stem Cell Research, University of Edin- burgh, Edinburgh, United Kingdom; d Department of Ophthalmology, Graduate School of Medicine, University of Nagoya, Nagoya, Japan; e Department of Ophthalmology, Graduate School of Medicine, University of Mie, Tsu City, Mie, Japan Key words: Endothelial progenitor cell transplantation • Neurovascular protection • Monocyte derived macrophages • Residual microglia ABSTRACT Retinitis pigmentosa is a group of inherited eye disorders that result in profound vision loss with characteristic ret- inal neuronal degeneration and vasculature attenuation. In a mouse model of retinitis pigmentosa, endothelial progenitor cells (EPC) from bone marrow rescued the vasculature and photoreceptors. However, the mecha- nisms and cell types underlying these protective effects were uncertain. We divided EPC, which contribute to angiogenesis, into two subpopulations based on their alde- hyde dehydrogenase (ALDH) activity and observed that EPC with low ALDH activity (Alde-Low) had greater neuroprotection and vasoprotection capabilities after injection into the eyes of an rd1 mouse model of retinitis pigmentosa compared with EPC with high ALDH activity (Alde-High). Of note, Alde-Low EPC selectively recruited F4/80 1 /Ly6c 1 monocyte-derived macrophages from bone marrow into retina through CCL2 secretion. In addition, the mRNA levels of CCR2, the neurotrophic factors TGF-b1 and IGF-1, and the anti-inflammatory mediator interleukin-10 were higher in migrated F4/80 1 /Ly6c 1 monocyte-derived macrophages as compared with F4/ 80 1 /Ly6c 2 resident retinal microglial cells. These results suggest a novel therapeutic approach using EPC to recruit neuroprotective macrophages that delay the pro- gression of neural degenerative disease. STEM CELLS 2013;31:2149–2161 Disclosure of potential conflicts of interest is found at the end of this article. INTRODUCTION Retinitis pigmentosa with an incidence of 1 in 3,500 is a group of inherited eye disorders that result in night blindness, visual field loss, and arteriolar attenuation, and often progress to complete blindness [1]. More than 100 genes responsible for retinitis pigmentosa have been identified and most of these encode for photoreceptor-specific proteins [2]. Gene therapy [3,4] and injecting neurotrophic factors [5,6] have been explored as therapeutic options. In recent years, several stud- ies have tested a wide range of cell populations for intravi- treal or subretinal transplantation experiments, including retinal and neural progenitor cells (postmitotic rod precursors and embryonic stem cell-derived photoreceptor precursors), mesenchymal stromal cells, and hematopoietic stem cells [7– Author contributions: S.F.: conception and design, financial support, collection and/or assembly of data, data analysis and interpretation, manuscript writing, and final approval of manuscript; M.N.: conception and design, administrative support, provision of study material or patients, collection and/or assembly of data, data analysis and interpretation, manuscript writing, and final approval of manuscript; T.Y.: conception and design, provision of study material or patients, collection and/or assembly of data, data analysis and interpretation, and final approval of manuscript; K.K. and I.T.: collection and/or assembly of data, data analysis and interpretation, and final approval of manuscript; G.S.: collection and/or assembly of data, data analysis and interpretation, manuscript writing, and final approval of manu- script; S.U.: provision of study material or patients, collection and/or assembly of data, data analysis and interpretation, and final approval of manuscript; M.K.: provision of study material or patients, collection and/or assembly of data, data analysis and interpreta- tion, and final approval of manuscript; T.K.: data analysis and interpretation, manuscript writing, and final approval of manuscript; T.O.: conception and design, financial support, administrative support, and final approval of manuscript; O.O.: conception and design, finan- cial support, administrative support, provision of study material or patients, data analysis and interpretation, manuscript writing, and final approval of manuscript. Correspondence: Osamu Ohneda, M.D., Ph.D., Department of Regenerative Medicine and Stem Cell Biology, Graduate School of Com- prehensive Human Sciences, University of Tsukuba, 1-1-1 Tennodai, Tsukuba, Ibaraki 305–8575, Japan. Telephone: 181-29-853-2938; Fax: 181-29-853-2938; e-mail: [email protected] Received December 10, 2012; accepted for publication June 11, 2013; first published online in STEM CELLS EXPRESS July 10, 2013. V C AlphaMed Press 1066-5099/2013/$30.00/0 doi: 10.1002/stem.1469 STEM CELLS 2013;31:2149–2161 www.StemCells.com
Transcript of Functional endothelial progenitor cells selectively recruit neurovascular protective...

REGENERATIVE MEDICINE
Functional Endothelial Progenitor Cells Selectively Recruit
Neurovascular Protective Monocyte-Derived F4/801/Ly6c1
Macrophages in a Mouse Model of Retinal Degeneration
SHINICHI FUKUDA,a,b MASUMI NAGANO,a,c TOSHIHARU YAMASHITA,a KENICHI KIMURA,a IKKI TSUBOI,a GEORGINA
SALAZAR,a
SHINJI UENO,d
MINEO KONDO,e
TILO KUNATH,c
TETSURO OSHIKA,b
OSAMU OHNEDAa
aDepartment of Regenerative Medicine and Stem Cell Biology, Graduate School of Comprehensive Human Science,
and bDepartment of Ophthalmology, Graduate School of Comprehensive Human Science, University of Tsukuba,
Tsukuba, Japan; cMRC Centre for Regenerative Medicine, Institute for Stem Cell Research, University of Edin-
burgh, Edinburgh, United Kingdom; dDepartment of Ophthalmology, Graduate School of Medicine, University of
Nagoya, Nagoya, Japan; eDepartment of Ophthalmology, Graduate School of Medicine, University of Mie, Tsu
City, Mie, Japan
Key words: Endothelial progenitor cell transplantation • Neurovascular protection • Monocyte derived macrophages • Residual
microglia
ABSTRACT
Retinitis pigmentosa is a group of inherited eye disordersthat result in profound vision loss with characteristic ret-inal neuronal degeneration and vasculature attenuation.In a mouse model of retinitis pigmentosa, endothelialprogenitor cells (EPC) from bone marrow rescued thevasculature and photoreceptors. However, the mecha-nisms and cell types underlying these protective effectswere uncertain. We divided EPC, which contribute toangiogenesis, into two subpopulations based on their alde-hyde dehydrogenase (ALDH) activity and observed thatEPC with low ALDH activity (Alde-Low) had greaterneuroprotection and vasoprotection capabilities afterinjection into the eyes of an rd1 mouse model of retinitis
pigmentosa compared with EPC with high ALDH activity(Alde-High). Of note, Alde-Low EPC selectively recruitedF4/80
1/Ly6c
1monocyte-derived macrophages from bone
marrow into retina through CCL2 secretion. In addition,the mRNA levels of CCR2, the neurotrophic factorsTGF-b1 and IGF-1, and the anti-inflammatory mediatorinterleukin-10 were higher in migrated F4/80
1/Ly6c
1
monocyte-derived macrophages as compared with F4/801/Ly6c2 resident retinal microglial cells. These resultssuggest a novel therapeutic approach using EPC torecruit neuroprotective macrophages that delay the pro-gression of neural degenerative disease. STEM CELLS
2013;31:2149–2161
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Retinitis pigmentosa with an incidence of 1 in 3,500 is agroup of inherited eye disorders that result in night blindness,visual field loss, and arteriolar attenuation, and often progressto complete blindness [1]. More than 100 genes responsiblefor retinitis pigmentosa have been identified and most of these
encode for photoreceptor-specific proteins [2]. Gene therapy[3,4] and injecting neurotrophic factors [5,6] have beenexplored as therapeutic options. In recent years, several stud-ies have tested a wide range of cell populations for intravi-treal or subretinal transplantation experiments, includingretinal and neural progenitor cells (postmitotic rod precursorsand embryonic stem cell-derived photoreceptor precursors),mesenchymal stromal cells, and hematopoietic stem cells [7–
Author contributions: S.F.: conception and design, financial support, collection and/or assembly of data, data analysis and interpretation,manuscript writing, and final approval of manuscript; M.N.: conception and design, administrative support, provision of study materialor patients, collection and/or assembly of data, data analysis and interpretation, manuscript writing, and final approval of manuscript;T.Y.: conception and design, provision of study material or patients, collection and/or assembly of data, data analysis and interpretation,and final approval of manuscript; K.K. and I.T.: collection and/or assembly of data, data analysis and interpretation, and final approvalof manuscript; G.S.: collection and/or assembly of data, data analysis and interpretation, manuscript writing, and final approval of manu-script; S.U.: provision of study material or patients, collection and/or assembly of data, data analysis and interpretation, and finalapproval of manuscript; M.K.: provision of study material or patients, collection and/or assembly of data, data analysis and interpreta-tion, and final approval of manuscript; T.K.: data analysis and interpretation, manuscript writing, and final approval of manuscript; T.O.:conception and design, financial support, administrative support, and final approval of manuscript; O.O.: conception and design, finan-cial support, administrative support, provision of study material or patients, data analysis and interpretation, manuscript writing, and finalapproval of manuscript.
Correspondence: Osamu Ohneda, M.D., Ph.D., Department of Regenerative Medicine and Stem Cell Biology, Graduate School of Com-prehensive Human Sciences, University of Tsukuba, 1-1-1 Tennodai, Tsukuba, Ibaraki 305–8575, Japan. Telephone: 181-29-853-2938;Fax: 181-29-853-2938; e-mail: [email protected] Received December 10, 2012; accepted for publication June 11, 2013;first published online in STEM CELLS EXPRESS July 10, 2013. VC AlphaMed Press 1066-5099/2013/$30.00/0 doi: 10.1002/stem.1469
STEM CELLS 2013;31:2149–2161 www.StemCells.com

11]. However, despite these efforts to better understand andtreat retinitis pigmentosa, there are still no effective treat-ments to slow or reverse the progression of this disease [2,3].
Endothelial progenitor cell (EPC) have been used clini-cally to treat cardiovascular disease and severe limb ischemia[12–18]. However, the clinical applications of EPC transfu-sions are limited due to the small numbers of available cells[15]. In a recent study, aldehyde dehydrogenase 1 (ALDH)activity was reported to be an effective marker that specifi-cally identified “functional EPC” [19]. Functional EPCs arethose that have high proliferative and migratory capabilitiesand exhibit a significant capacity to regenerate ischemic tissue[19,20]. It was shown that EPC with low levels of ALDHexpression (Alde-Low EPC) migrated to an ischemic regionand decreased necrotic area in a mouse model of skin flapischemia after making an incision on the dorsal surface [19].In a rat model of acute cerebral infarction, Alde-Low EPCsaccumulated and migrated into the infarct site and subse-quently decreased the infarct volume [20]. ALDH activity isan effective marker for functional EPC because Alde-LowEPC are more functional than those with high ALDH activity(Alde-High EPC) [19].
Inherited retinal degeneration is characterized by retinalneuronal apoptosis and retinal vasculature attenuation.Recently, the therapeutic effects of EPC on central nervoussystem and peripheral nerves under hypoxic conditions suchas in stroke and diabetic neuropathy were investigated [21–23]. However, few reports have assessed the neuroprotectivecapability of EPC in nonhypoxic environments.
Lineage-negative hematopoietic stem cells that includedEPC could rescue retinal blood vessels and photoreceptors innonischemic neurodegenerative diseases, such as retinitis pig-mentosa [9]. However, it was unclear whether the increase inblood flow by revascularization improved neural damage orwhether EPC had any neurotrophic effects. Hematopoietic stemcells are multipotent stem cells that give rise to all blood celltypes. The lineage commitment of cells arising from hemato-poietic stem cells contributed was not confirmed. In this study,purified functional EPC isolated based on their low ALDHactivity showed neuroprotective and vasoprotective effects.
In this study, we report for the first time that Alde-LowEPC can induce the selective recruit of a distinct subset ofF4/801/Ly6c1 macrophages with high mRNA expression lev-els of chemokine C-C motif receptor 2 (CCR2), neurotrophicfactors, and anti-inflammatory mediators. These results sug-gested that intraocular injection of Alde-Low EPC effectivelyrecruited inflammation-resolving and neuroprotectivemonocyte-derived macrophages that were different from F4/801/Ly6c2 resident microglial cells.
MATERIALS AND METHODS
Mice
C57BL/6J and C3H/HeN (rd1) mice were purchased from JapanSLC, Inc. (Shizuoka, Japan). Mice were treated in accordancewith the NIH Guide for the Care and Use of Laboratory Animalsand the ARVO Statement for Use of Animals in Ophthalmic andVision Research. All experimental procedures were approved bythe University of Tsukuba.
Isolation of Alde-High EPC and Alde-Low EPC fromUmbilical Cord Blood
EPC was isolated from umbilical cord blood and cultured as previ-ously described [19]. In brief, mononuclear cells from human
umbilical cord blood were separated by density gradient centrifuga-tion after depleting hematopoietic cells. Then, CD452/CD311 cellswere sorted and cultured in Iscove’s Modified Dulbecco’s Medium(IMDM) with 10% fetal bovine serum (FBS) and basic fibroblastgrowth factor (b-FGF). More than approximately 7 days, adherentcells began to grow and rapidly evolved to form colonies withtightly compact morphologies. Subsequently, DiI-Ac-LDL-posi-tive/CD311 cells were sorted (DiI-Ac-LDL; Molecular Probes,Eugene, OR, http://probes.invitrogen.com). Culture of sorted cellswas continued in IMDM/10% FBS supplemented with b-FGF.These cells displayed endothelial cell (EC)-like morphology.
ALDH activity was analyzed with an Aldefluor reagent(StemCell Technologies, Vancouver, BC, Canada, http://www.stemcell.com). Then, EPCs were divided into two fractionsbased on their level of ALDH expression: Alde-High EPCs andAlde-Low EPCs [19]. EPC were infected with lenti-GFP (Sigma-Aldrich, St Louis, MO, http://www.sigmaaldrich.com) and greenfluorescent protein (GFP)-positive EPC was purified usingMoFLo (MoFlo XDP; Dako, Carpinteria, CA). As a control,human umbilical vein endothelial cells (HUVEC) was purchasedfrom Cambrex (Walkersville, MD) and maintained according tothe manufacturer’s instructions.
Intravitreal and Subretinal Injection of Cells
Cells were injected using a 33-gauge needle (Ito Co. Fuji, Japan,http://www.ito-ex.co.jp). To evaluate vasoprotective and neuro-protective effects, cells were injected at P6. The density ofinjected cells varied according to the technique used. For intravi-treal injections, approximately 1 3 105 cells per 0.5 mL of EPCand HUVEC was delivered. For subretinal injections, 0.5 3 105
cells per 0.2–0.3 mL of EPC and HUVEC, approximately 1 3104 cells per 0.2–0.3 mL of F4/801/Ly6c1 cells were used.Immunosuppression was induced with daily intraperitoneal injec-tion of cyclosporin-A (Wako, Osaka, Japan, http://www.wako-chem.co.jp/english) at 20 mg/kg body weight 2 days before theassay, as in previous reports [19,20].
Flow Cytometry Analysis of Retina and PeripheralBlood Cells
One week after injecting PBS or cells, the total retinal cell popu-lation was isolated for flow cytometry analysis (MoFlo XDP;Dako). Blood was collected from the retro-orbital sinus of 6- to8-week-old wild type (WT) mice 2 days after intravitreal injec-tion of PBS or cells. Mononuclear cells in peripheral blood wereisolated using Histopaque density gradient as described previ-ously [12]. The numbers of F4/801/Ly6c1 cells were evaluatedby flow cytometry. F4/801/Ly6c1 cells isolated from the periph-eral blood of WT mice were injected into the subretinal spaces ofrd1 mice at P6. The following monoclonal antibodies (mAbs)were used according to the manufacturers’ protocols: phycoery-thrin (PE) conjugated F4/80 antibodies (1:20, A3-1, AbD Serotec,Raleigh, NC); biotin-conjugated anti-Ly6c antibody (1:50, AL-21,BD Biosciences, San Diego, CA, http://www.bdbiosciences.com),and allophycocyanin (APC) conjugated streptavidin (1:50, BD).
Terminal Deoxynucleotidyl Transferase-Mediated Bio-tinylated UTP Nick End Labeling Assay
A terminal deoxynucleotidyl transferase-mediated biotinylatedUTP nick end labeling (TUNEL) assay kit with anti-fluoresceinisothiocyanate (FITC) conjugated horseradish peroxidase (HRP)(TaKaRa, Ohtsu, Japan, http://www.takara.co.jp) was used forcryostat retinal sections (7 mm) of P13 of an rd1 mouse [24]. Threedifferent areas (400 mm in width; unbiased sampling) in a singlesection that contained the optic nerve head nuclei were counted.
Evaluation of Vasoprotective Ability Effects Based onMeasured Lengths of the Retinal Vasculature
Two hundred microliters of tetramethylrhodamine isothiocyanate-conjugated Bandeiraea simplicifolia lectin (0.1 mg/mL; Sigma-
2150 EPC Selectively Recruited Protective Macrophage

Aldrich) was injected through a tail vein, after which a retinal flatmount were prepared. Four randomly chosen independent fields (600mm 3 600 mm) were used to quantify of the vasculature as previ-ously described [9]. The total vasculature length was measured.
Evaluation of Neuroprotective Ability by Measuringthe Thickness of Outer Nuclear Layer
Harvested eyes were immersion-fixed in 4% paraformaldehyde(PFA) and embedded in OCT compound (Tissue-Tek; SakuraFineTech Inc.). Cryostat sections (7 mm) were stained with hema-toxylin and eosin (H&E). The thickness of outer nuclear layer(ONL) was measured at four points that were decided upon bydividing the retina (from the nerve head to the peripheral retina)evenly into four parts (Fig. 2A).
Retinal Function Recording by Electroretinography
Alde-Low EPCs or PBS were randomly chosen and injected into theintravitreal spaces of six rd1 mice at P6, and electroretinography(ERG) was recorded at P17. ERG was recorded as described previ-ously [25]. Luminance was 1.0 log cd-s/m2 on a rod-desensitizingwhite adapting background of 1.3 log cd/m2. A total of 150 ERGrecordings were averaged using a computer-assisted signal averagingsystem (Power Lab; AD Instruments, Castle Hill, Australia). Theamplitude was measured based on the highest positive wave by aresearcher blinded to the experimental conditions used.
Immunohistochemistry
Whole eyes were harvested and fixed with 4% PFA. Whole-mount retina or cryostat sections (7 mm) were immunohisto-chemically stained. To stain microglial cells, anti-F4/80 (1:20,A3-1, AbD Serotec), anti-CD11b (1:50, Abcam, Cambridge, MA,http://www.abcam.com), anti-CX3CR1 (1:50, Abcam), and anti-Ly6C (1:50, ER-MP20, Abcam) antibodies were used, followedby Alexa 488 or 594 conjugated secondary antibodies (1:1,000,Invitrogen, Carlsbad, CA, http://www.invitrogen.com) andisolectin-IB-4 conjugated to Alexa Fluor 594 (1:200, Invitrogen).
Numbers of Migrated Macrophages
Migrated macrophages in the retinas of whole-mount preparationswere stained with an anti-F4/80 antibody and counted using aconfocal microscope (TCS SP5, Leica, Wetzlar, Germany). Fourrandomly chosen independent fields (800 mm 3 800 mm) wereused for quantification.
Systemic Depletion of Circulating Macrophages
Clodronate-loaded liposomes were provided by N. van Rooijen(Vrije Universiteit, Amsterdam, The Netherlands) and were intra-peritoneally injected (at an appropriate dilution for each experi-ment) at a volume of 0.1 mL per day [26,27].
Enzyme-Linked Immunosorbent Assay for CCL2
To measure paracrine activity, Alde-Low EPC, CCL2 overex-pressed Alde-Low EPC (Alde-Low CCL2 EPC), CCL2 downregu-lated Alde-Low EPC (Alde-Low CCL2kd EPC), Alde-High EPC,CCL2 overexpressed Alde-High EPC (Alde-High CCL2 EPC),and HUVEC were placed in basal medium with 2% serum andavoiding potentially contaminating supplemental growth factors.Samples were collected after 24 hours. The number of adherentcells was also quantified. Results were expressed as nanogramssecreted per 105 cells for each paracrine factor over a 24-hourperiod. The concentration of human CCL2 was determined usinghuman CCL2 enzyme-linked immunosorbent assay (ELISA) kits(R&D Systems, Minneapolis, MN, http://www.rndsystems.com).
Quantitative Reverse Transcriptase Polymerase ChainReaction Analysis
Total RNA was extracted and purified using an RNeasy Mini Kit(Qiagen, Valencia, CA, http://www1.qiagen.com). RNA was
reverse-transcribed to cDNA using ReverTra-Plus-TM (Toyobo,Osaka, Japan) and then used as template for reverse transcriptasepolymerase chain reaction (RT-PCR) analysis. The sequences ofthe PCR primers (sense and antisense, respectively) were 50-GAACTTTCTGCTGTCTTGGGTGCATTG-30 and 50-CTGCATGGTGATGTTGGACTCCTCAGT-30 for human vascular endothelialgrowth factor (VEGF), 50-CTGACTCACATAGGGTGCAGCAATCAG-30 and 50-AGGCTGGTTCCTATCTCCAGCATGGTA-30 forhuman Ang-1, 50-AACTCAGCTAAGGACCCCACTGTTGCT-30
and 50-TGAGTAAGCCTCATTCCCTTCCCAGTC-30 for humanAng-2, 50-TTGACGGGGTCCGGGAGAAGAGCGACC-30 and 50-TTGGAAGAAAAAGTATAGCTTTCTGCCCAGG-30 for humanbFGF, 50-CTTGATCTAAGGAGGCTGGAGATGT-30 and 50-CTTGTTCCTGCACTCCCTCTACTTG-30 for human insulin-likegrowth factor type 1 (IGF-1), 50-GATCCGCTCCTTTGATGATCTCCAACGC-30 and 50-ACTGCACGTTGCGGTTGTTGCAGCAGCC-30 for human platelet-derived growth factor b (PDGF-b), 50-CTTGTCATGCTGCTCCTCCTG-30 and 50-TGCGACTCCTCACATCTCTGC-30 for human epidermal growth factor (EGF), 50-CAGCAGAGCACACAAGCTTCTAGGACA-30 and 50-GTCCAGACAGAGCTCTCTTCCTCCATCAGAA-30 for human interleukin (IL)28,50-CCATATTCCTCGGACACCAC-30 and 50-TGTACTCCCGAACCCATTTC-30 for human CCL5, 50-GGCTGAGACTAACCCAGAAAGATCCAA-30 and 50-TGGGTTGTGGAGTGAGTGTTCAAGTCT-30 for human CCL2, 50-TCAAGTCTTCGGAGTTTGGGTTTGCTT-30 and 50-GTACTTGCGCTCAGGAGGAGCAATGAT-30 forhuman b-actin, 50-GCTCTTACTGACTGGCATGAG-30 and 50-CGCAGCTCTAGGAGCATGTG-30 for mouse IL-10, 50-AAAATTCGAGTGACAAGCCTGTAG-30 and 50-CCCTTGAAGAGAACCTGGGAGTAG-30 for mouse TNF-a, 50-TACCATGCCAACTTCTGTCTGGGA-30 and 50-TGTGTTGGTTGTAGAGGGCAAGGA-30
for mouse TGF-b1, 50-CTACAAAAGCAGCCCGCTCT-30 50-CTTCTGAGTCTTGGGCATGTCA-30 for mouse IGF-1, 50-TTGGAACCATCTTCCTGTCC-30 and 50-ACGCCCAGACTAATGGTGAC-30
for mouse CX3CR1, 50-ATCCACGGCATACTATCAACATC-30
and 50-CAAGGCTCACCATCATCGTAG-30 for mouse CCR2, 50-GTCGTACCACAGGCATTGTGATGGACT-30 and 50-CACCAGACAGCACTGTGTTGGCATAGA-30 for mouse b-actin. Reactionsfor b-actin mRNA were performed concurrently in the same plate asthose for the test of mRNAs, and results were normalized by the cor-responding amount of b-actin mRNA.
Small hairpin RNA (shRNA)
To downregulate CCL2 expression, we used a shRNA MISSIONlentiviral transduction system (NM_002982; clone:TRCN0000006279; sequence; 50-CCGGGATGTGAAACATTATGCCTTACTCGAGTAAGGCATAATGTTTCACATCTTTTT-30;Sigma-Aldrich) performed as specified in the manufacturer’s pro-tocol. cDNA levels were determined by quantitative RT-PCR, andprotein levels were determined by ELISA.
CCL2 Overexpression in Alde-Low EPC, Alde-HighEPC, and HUVEC
Human CCL2 was overexpressed in Alde-Low EPC, Alde-HighEPC, and HUVEC using murine stem cell virus, Phoenix-Ecopackaging cells (Clontech Laboratories, Palo Alto, CA, http://www.clontech.com), a FuGENE transfection kit (Roche, Basel,Switzerland, http://www.roche-applied-science.com), and PT67cells (Clontech Laboratories) as previously described [28]. Theexpressions of human CCL2 mRNA and protein were confirmedby quantitative RT-PCR and ELISA, respectively.
Statistical Analysis
Results are given as means 6 SDs. Results for two groups were com-pared using Student’s t tests. Paired t tests were used to compare theresults for the same group at different time points. One-way ANOVAwith Bonferroni multiple comparisons was used to compare morethan two groups. In the graphs, y-axis error bars indicate SDs. Allanalyses had been carried out in a masked fashion.
Fukuda, Nagano, Yamashita et al. 2151
www.StemCells.com
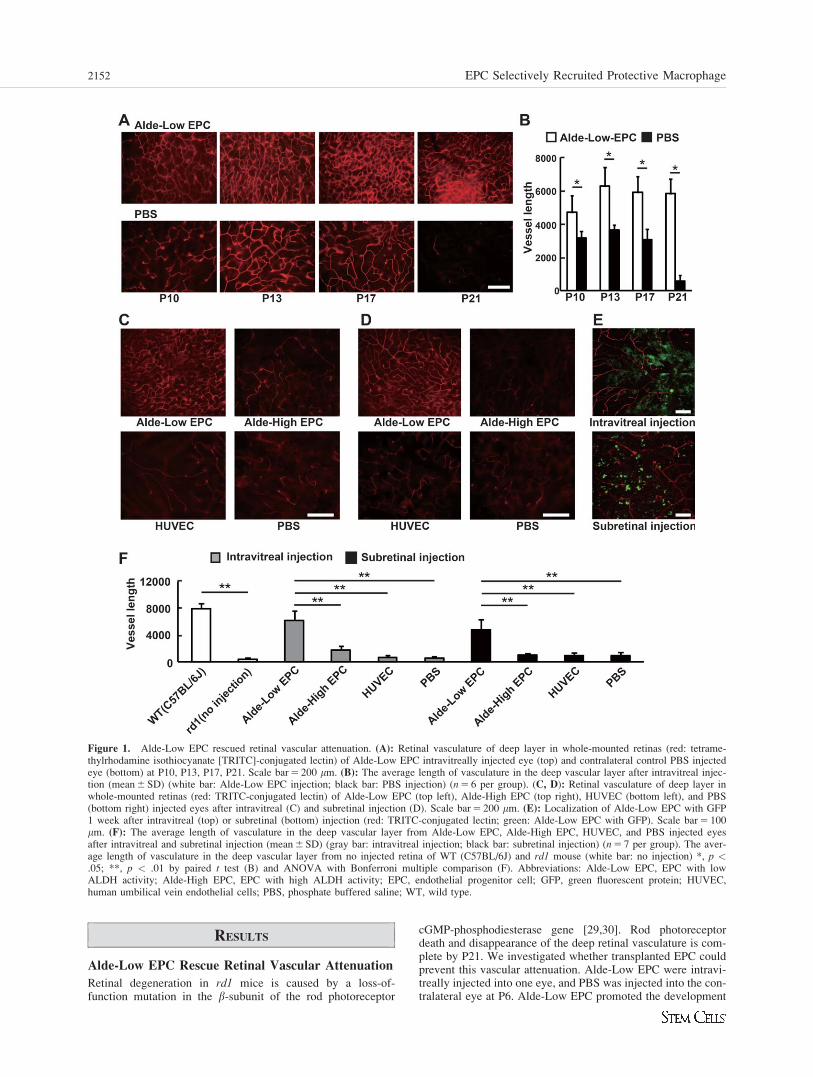
RESULTS
Alde-Low EPC Rescue Retinal Vascular Attenuation
Retinal degeneration in rd1 mice is caused by a loss-of-function mutation in the b-subunit of the rod photoreceptor
cGMP-phosphodiesterase gene [29,30]. Rod photoreceptordeath and disappearance of the deep retinal vasculature is com-plete by P21. We investigated whether transplanted EPC couldprevent this vascular attenuation. Alde-Low EPC were intravi-treally injected into one eye, and PBS was injected into the con-tralateral eye at P6. Alde-Low EPC promoted the development
Figure 1. Alde-Low EPC rescued retinal vascular attenuation. (A): Retinal vasculature of deep layer in whole-mounted retinas (red: tetrame-thylrhodamine isothiocyanate [TRITC]-conjugated lectin) of Alde-Low EPC intravitreally injected eye (top) and contralateral control PBS injectedeye (bottom) at P10, P13, P17, P21. Scale bar 5 200 mm. (B): The average length of vasculature in the deep vascular layer after intravitreal injec-tion (mean 6 SD) (white bar: Alde-Low EPC injection; black bar: PBS injection) (n 5 6 per group). (C, D): Retinal vasculature of deep layer inwhole-mounted retinas (red: TRITC-conjugated lectin) of Alde-Low EPC (top left), Alde-High EPC (top right), HUVEC (bottom left), and PBS(bottom right) injected eyes after intravitreal (C) and subretinal injection (D). Scale bar 5 200 mm. (E): Localization of Alde-Low EPC with GFP1 week after intravitreal (top) or subretinal (bottom) injection (red: TRITC-conjugated lectin; green: Alde-Low EPC with GFP). Scale bar 5 100mm. (F): The average length of vasculature in the deep vascular layer from Alde-Low EPC, Alde-High EPC, HUVEC, and PBS injected eyesafter intravitreal and subretinal injection (mean 6 SD) (gray bar: intravitreal injection; black bar: subretinal injection) (n 5 7 per group). The aver-age length of vasculature in the deep vascular layer from no injected retina of WT (C57BL/6J) and rd1 mouse (white bar: no injection) *, p <.05; **, p < .01 by paired t test (B) and ANOVA with Bonferroni multiple comparison (F). Abbreviations: Alde-Low EPC, EPC with lowALDH activity; Alde-High EPC, EPC with high ALDH activity; EPC, endothelial progenitor cell; GFP, green fluorescent protein; HUVEC,human umbilical vein endothelial cells; PBS, phosphate buffered saline; WT, wild type.
2152 EPC Selectively Recruited Protective Macrophage

of the deep vascular layer, and prevented vascular attenuation,as compared with PBS injection (Fig. 1A, 1B).
Next, Alde-Low EPC, Alde-High EPC, HUVEC, or PBSwere intravitreally [9] and subretinally [7,31] injected intord1 mouse eyes at P6. Injecting Alde-Low EPC resulted in apronounced rescue of the deep vascular layer as comparedwith the control (Fig. 1C–1F). At 1 week after intravitreal or
subretinal injection, a large number of Alde-Low EPC withGFP survived on the retinal surface, and in the intravitrealspace or subretinal space (Fig. 1E). However, Alde-Low EPCwere not incorporated in any vessels of the three retinal layers(data not shown). After longer time observation, the numberof Alde-Low EPC gradually decreased and had completelydisappeared by 6 weeks after injection.
Figure 2. Neuroprotective effect of Alde-Low EPC. (A): Retinal cross-sections of Alde-Low EPC injected eyes (top) and contralateral controlPBS injected eyes (bottom) at P10, P13, P17, P21. Scale bar 5 200 mm. (B): The ONL thickness after intravitreal injection (mean 6 SD) (whitebar: Alde-Low EPC injection; black bar: PBS injection) (n 5 15 per group). (C): Maximum amplitudes by electroretinography recording wereused to measure the retinal function of Alde-Low EPC injected eyes (top) and PBS injected eyes (bottom). Representative cases were shown(mean 6 SD) (white bar: Alde-Low EPC injection; black bar: PBS injection) (n 5 6 per group). (D): The ONL thickness at P17 of Alde-LowEPC (white bar), Alde-High EPC (gray bar), HUVEC (striped bar), and PBS (black bar) intravitreally injected eyes (mean 6 SD) (n 5 15 pergroup). (E, F): The number of TUNEL-positive cells in retina of Alde-Low EPC (top left), Alde-High EPC (top right), HUVEC (bottom left),and PBS (bottom right) injected eyes. (mean 6 SD) (n 5 5 per group). Scale bar 5 200 mm. (G): Retinal cross-sections of Alde-Low EPC subreti-nally injected eyes. Arrow indicates injected region of cells, and arrowheads indicate injected Alde-Low EPC with GFP. Scale bars 5 200 mm (Gbottom) and 1 mm (G top). (H): The ONL thickness at P17 of Alde-Low EPC or PBS injected eyes by intravitreally or subretinally (mean 6 SD)(white bar: Alde-Low EPC injection; black bar: PBS injection). (n 5 7 per group). *, p < .05; **, p < .01 by paired t test (B), Student’s t test(C) and ANOVA with Bonferroni multiple comparison (D, F, H). Abbreviations: Alde-Low EPC, EPC with low ALDH activity; Alde-High EPC,EPC with high ALDH activity; INL, inner nuclear layer; ONL, outer nuclear layer; PBS, phosphate buffered saline; TUNEL, terminal deoxynu-cleotidyl transferase-mediated biotinylated UTP nick end labeling.
Fukuda, Nagano, Yamashita et al. 2153
www.StemCells.com
Neuroprotective Effect of Alde-Low EPC
To determine whether Alde-Low EPC could protect the retinafrom neural degeneration, Alde-Low EPC were intravitreallyinjected into rd1 mice at P6. After the P13 stage, this injec-tion had retarded the degenerative process as compared withPBS injection (Fig. 2A, 2B).
Alde-Low EPC showed some functional rescue of the ret-ina at P17 based on ERG, although the signal amplitude wasconsiderably lower than normal, besides they also presentedan abnormal pattern (Fig. 2C). These signals needed to beaveraged using a strong flash for more than 100 times todetect these waveforms because their amplitudes were toosmall to be recorded. Because some ERG signals had ana-wave unlike the others, only b-waves were measured. Fur-ther analyses found that a greater number of photoreceptors
remained in the ONL at P17 when only Alde-Low EPC wereinjected (Fig. 2D). In addition, using a TUNEL assay[24,30,32] there were fewer apoptotic cells in the ONLs ofmice that were injected with Alde-Low EPC than in othermice (Fig. 2E, 2F). This indicated that Alde-Low EPC had aneuroprotective effect in the retina.
To eliminate the influence of the blood-retina barrier,Alde-Low EPC were directly injected into the subretinalspace, a site closer to the degenerated retina. When Alde-LowEPC were injected into this subretinal site, their neuroprotec-tive effect was localized around the injection site and nodirect integration into the ONL was apparent (Fig. 2G, 2H).There was a tendency for a greater ONL thickness afterinjecting Alde-Low EPC subretinally as compared with intra-vitreally, although this difference was not significant (Fig.
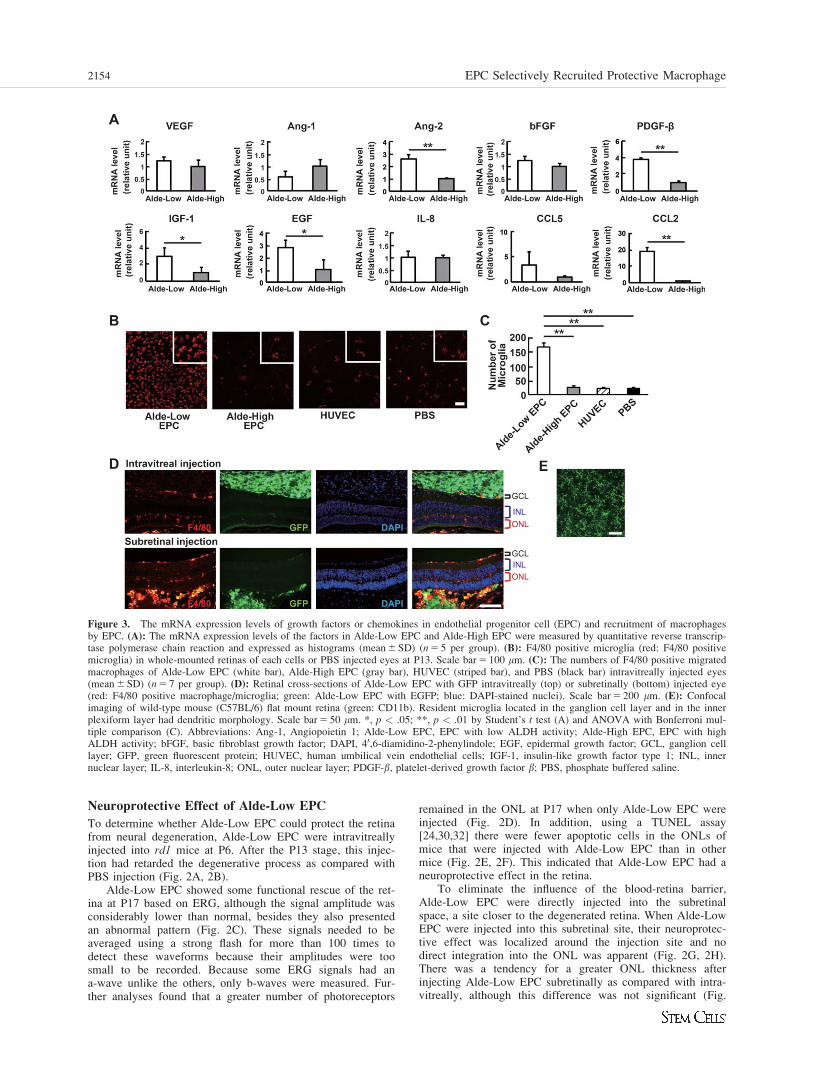
Figure 3. The mRNA expression levels of growth factors or chemokines in endothelial progenitor cell (EPC) and recruitment of macrophagesby EPC. (A): The mRNA expression levels of the factors in Alde-Low EPC and Alde-High EPC were measured by quantitative reverse transcrip-tase polymerase chain reaction and expressed as histograms (mean 6 SD) (n 5 5 per group). (B): F4/80 positive microglia (red: F4/80 positivemicroglia) in whole-mounted retinas of each cells or PBS injected eyes at P13. Scale bar 5 100 mm. (C): The numbers of F4/80 positive migratedmacrophages of Alde-Low EPC (white bar), Alde-High EPC (gray bar), HUVEC (striped bar), and PBS (black bar) intravitreally injected eyes(mean 6 SD) (n 5 7 per group). (D): Retinal cross-sections of Alde-Low EPC with GFP intravitreally (top) or subretinally (bottom) injected eye(red: F4/80 positive macrophage/microglia; green: Alde-Low EPC with EGFP; blue: DAPI-stained nuclei). Scale bar 5 200 mm. (E): Confocalimaging of wild-type mouse (C57BL/6) flat mount retina (green: CD11b). Resident microglia located in the ganglion cell layer and in the innerplexiform layer had dendritic morphology. Scale bar 5 50 mm. *, p < .05; **, p < .01 by Student’s t test (A) and ANOVA with Bonferroni mul-tiple comparison (C). Abbreviations: Ang-1, Angiopoietin 1; Alde-Low EPC, EPC with low ALDH activity; Alde-High EPC, EPC with highALDH activity; bFGF, basic fibroblast growth factor; DAPI, 40,6-diamidino-2-phenylindole; EGF, epidermal growth factor; GCL, ganglion celllayer; GFP, green fluorescent protein; HUVEC, human umbilical vein endothelial cells; IGF-1, insulin-like growth factor type 1; INL, innernuclear layer; IL-8, interleukin-8; ONL, outer nuclear layer; PDGF-b, platelet-derived growth factor b; PBS, phosphate buffered saline.
2154 EPC Selectively Recruited Protective Macrophage

2H). There was no recovery of degenerated photoreceptorsafter intravitreal or subretinal injection of Alde-Low EPCs atlater stages (data not shown).
Growth Factor and Chemokine Expressions by EPC
To investigate which growth factors [21–23] or chemokines[33] might have functional roles in these protective effects,we determined the mRNA levels of candidate factors in Alde-Low EPC and Alde-High EPC (Fig. 3A). Quantitative RT-PCR results showed that Ang-2, PDGF-b, IGF-1, EGF, andCCL2 mRNAs were expressed at significantly higher levels inAlde-Low EPC as compared with Alde-High EPC. In agree-ment with previous reports that showed that CCL2, alsoknown as monocyte chemotactic protein-1, recruited mono-cytes and macrophages, a greater number of macrophageswas observed in the retinas injected with Alde-Low EPC (Fig.3B, 3C).
Characterization of Migrated Macrophages
In a WT mouse, resident microglial cells located in the gan-glion cell layer and in the inner plexiform layer had dendriticmorphologies (Fig. 3E). When retinal degeneration occurs,resident and resting microglial cells migrate to the ONL andphagocytose apoptotic photoreceptors [34]. In this study,microglial cells were observed in the ONL of an rd1 mousewithout injecting cells or PBS and these microglial cellsmight have been involved in the phagocytosis of apoptoticphotoreceptors, as in a previous study [34]. The number ofF4/80 positive cells (a marker for macrophages/microglialcells) was the highest after injecting Alde-Low EPC (Fig.3B). F4/80-positive cells were observed in the ONL as wellas on the surface and in the inner retina after injecting Alde-Low EPC (Fig. 3D). Confocal microscope images of a retinalflat mount clearly showed that recruited F4/80-positive cellshad amoeboid-like shapes, which was different from theshapes of resident and resting microglial cells (Fig. 3B).
CCL2 Secreted by Alde-Low EPCs Induces Macro-phage Migration and Prevents Vascular Attenuationand Neural Degeneration
To investigate whether CCL2 played an important role, weused shRNA to reduce CCL2 expression in Alde-Low EPC(Alde-Low CCL2kd EPC; Fig. 4A, 4B). When Alde-LowCCL2kd EPC were injected intravitreally, the numbers ofmigrated macrophages decreased dramatically and resulted inphotoreceptor degeneration and a failure to rescue vascularattenuation (Fig. 4D, 4F–4H).
To determine how CCL2 was involved in the inhibitingvascular attenuation and neural degeneration, we overex-pressed CCL2 in Alde-High EPC and HUVEC (Alde-HighCCL2 EPC and HUVEC-CCL2 cells; Fig. 4A–4C). Afterintravitreal injection of Alde-High CCL2 EPC, the number ofmigrated macrophages was greater than after injecting Alde-High EPC (Fig. 4D, 4H). Although vascular attenuation andneural degeneration were retarded as compared with injectingAlde-High EPC alone, the same levels of protection were notobserved as when injecting Alde-Low EPC (Fig. 4F, 4G, 4H).This suggested that Alde-Low EPC-derived factors other thanCCL2 might have vascular and neuronal protective effects. Incontrast to the results of Alde-High CCL2 EPC injection, thenumber of macrophages did not increase after transplantingHUVEC-CCL2 cells. Vascular attenuation and neural degen-eration were not recovered by HUVEC-CCL2 cells, whichsuggested that CCL2 alone was not sufficient to attract thosemacrophages that might contribute to delaying retinal degen-eration (Fig. 4E–4H).
Macrophage Migration Induced by Alde-Low EPCsIs Involved in Protection Against Vascular Attenua-tion and Neural Degeneration
To investigate how macrophages contributed to these protec-tive effects in the retina, clodronate liposomes were injectedto eliminate migrated macrophages [26]. Peritoneal injectionof clodronate liposomes completely abolished macrophages’migration (Fig. 4E, 4H) and also disrupted the vasoprotectiveand neuroprotective effects (Fig. 4E–4G). These results indi-cated that macrophages migrating to the retina played animportant role in vasoprotection and neuroprotection ratherthan any direct effects by Alde-Low EPC. In contrast toinjecting Alde-Low EPC, injecting CCL2-transfected Alde-Low EPC (Alde-Low CCL2 EPC) failed to promote anyincrease in the numbers of migrated macrophages and did notsignificantly change retinal vascular attenuation and photore-ceptor degradation. This suggested that the number ofmigrated macrophages due to CCL2 was limited, and that theprotective effects may have depended on the numbers ofmigrated macrophages. Taken together, these results indicatedthat migrated macrophages, recruited by Alde-Low EPCs viatheir secretion of CCL2 and other unknown factors, played animportant role in delaying the progression of retinaldegeneration.
Migrated Macrophages Are Different from ResidentRetina Microglial Cells
To further investigate migrated macrophages, F4/80-positivecells were stained with other macrophage markers, CD11b andIsolectin-IB4. Macrophages were also stained with CX3CR1 (amyeloid lineage marker) [34–36] and Ly6c (a marker of bonemarrow [BM]-derived monocytes) [37] after the injectingAlde-Low EPC (Fig. 5A). Migrated Ly6c-positive cells inte-grated not only into the ONL but also onto the surface andinto the inner retinal layer (Fig. 5B). In contrast, residentmicroglial cells in the ONL were negative for Ly6c.
Intraperitoneal injection of clodronate liposomes clearlyshowed that these liposomes affected the recruitment of F4/801/Ly6c1 macrophages; however, this effect was notobserved for F4/801/Ly6c2 cells (Fig. 5C, 5D). F4/801/Ly6c2
cells were resident retinal microglial cells and had migrated tothe ONL to phagocytose apoptotic photoreceptors [27,38].Thus, we speculated that these F4/801/Ly6c1 cells weremigrating monocyte-derived macrophages. We then character-ized these macrophages by analyzing their mRNA expression.
Anti-inflammatory mediator IL-10 mRNA expression wasfound in migrated monocyte-derived macrophages (Ly6c1),whereas resident retinal microglial cells (Ly6c2) expressedinflammatory mediator TNF-a mRNA. The mRNA levels ofgrowth factors, including TGF-b1 and IGF-1, were higher inmigrated monocyte-derived macrophages (Ly6c1) than in resi-dent retinal microglial cells (Ly6c2) (Fig. 5E, 5F). Of note, themRNA level for CCR2, a chemokine receptor for CCL2, washigher in migrated monocyte-derived macrophages (Ly6c1)than in resident retinal microglial cells (Ly6c2). This suggestedthat transplanted Alde-Low EPCs secreted the chemokineCCL2 to attract monocyte-derived macrophages to the retina.
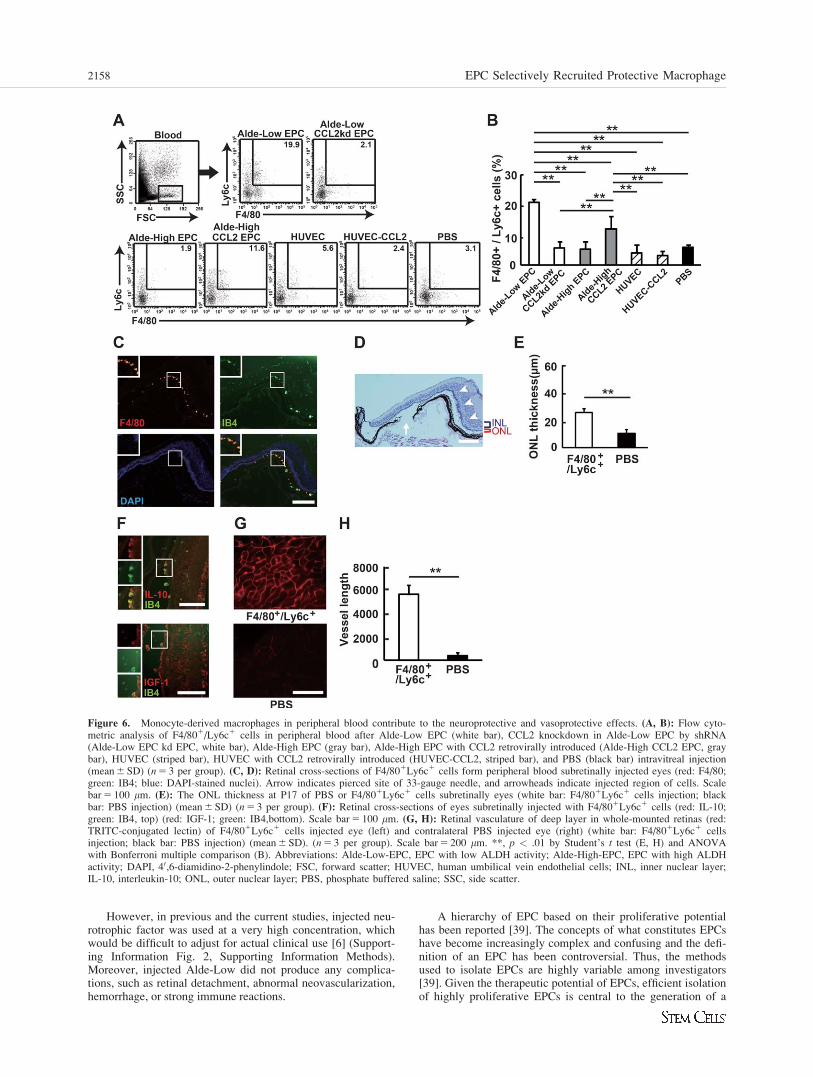
Monocyte-Derived Macrophages in Peripheral BloodContribute to the Neuroprotective andVasoprotective Effects
Mononuclear cells in the peripheral blood were isolated afterintravitreal injection of PBS or cells. Flow cytometry analysisshowed that there was a higher frequency of F4/801/Ly6c1
cells in the peripheral blood of Alde-Low EPC injected mice
Fukuda, Nagano, Yamashita et al. 2155
www.StemCells.com

as compared with other mice (Fig. 6A, 6B). This indicatedthat Alde-Low EPC induced the migration of F4/801/Ly6c1
cells from the BM into the peripheral blood.We then injected 1 3 104 F4/801/Ly6c1 cells isolated
from peripheral blood into the subretinal spaces of rd1 mice atP6 and analyzed the number of rescued cells. There were morerescued cells in the ONL as compared with subretinal injectionof PBS. However, the neuroprotective effects were localized tothe injection site (Fig. 6C–6E). Interestingly, injected F4/801/
IB41 monocyte-derived macrophages remained at the rescuedregion and expressed IL-10 and IGF-1 mRNAs (Fig. 6F).Finally, subretinal injection of F4/801/Ly6c1 cells clearly res-cued the deep vascular layer construct as compared with theinjecting PBS (Fig. 6G, 6H). However, this F4/802Ly6C1 pop-ulation showed no neuroprotective or vascular protective capa-bilities after injection. The F4/802Ly6C1 populationcomprised neutrophils, ECs, or T-cell subsets based on flowcytometry analysis (Supporting Information Fig. 1).
Figure 4. CCL2 released from Alde-Low EPC and migrated macrophages related to prevention of vascular attenuation and neural degeneration.(A, B): The mRNA expression levels (n 5 5 per group) and protein level (n 5 4 per group) of CCL2 in Alde-Low EPC, Alde-Low EPC withCCL2 retrovirally introduced (Alde-Low CCL2 EPC), CCL2 knockdown in Alde-Low EPC by shRNA (Alde-Low EPC kd EPC), Alde-HighEPC, Alde-High EPC with CCL2 retrovirally introduced (Alde-High CCL2 EPC), and HUVEC (mean 6 SD). (C): The mRNA expression levelsof CCL2 in HUVEC and HUVEC with CCL2 retrovirally introduced (HUVEC-CCL2) (mean 6 SD) (n 5 3 per group). (D, E): Hematoxylin andeosin-stained cross-sections image of each cells injected eyes at P17 (HE) (top). Scale bar 5 200 mm. TRITC-conjugated lectin stained deep vas-cular layer in whole-mounted retina of each cells injected at P21 (Lectin-TRITC) (middle, red: TRITC-conjugated lectin). Scale bar 5 200 mm.F4/80 positive microglia (F4/80) (bottom, red: F4/80 positive microglia) in whole-mounted retinas of each cells injected eyes at P13. Scalebar 5 100 mm. (E): The effects of clodronate liposome by peritoneal injection with Alde-Low EPC, Alde-Low CCL2 EPC, and Alde-High CCL2EPC intravitreal injection were assessed. (F): The ONL thickness, (G) the average length of vasculature in the deep vascular layer, (H) or thenumbers of migrated microglia of each cells injected eye expressed as histograms (mean 6 SD) (n 5 7 per group). *, p < .05; **, p < .01 byANOVA with Bonferroni multiple comparison (A–C, F–H). Abbreviations: Alde-Low-EPC, EPC with low ALDH activity; Alde-High-EPC, EPCwith high ALDH activity; EPC, endothelial progenitor; HUVEC, human umbilical vein endothelial cells; ONL, outer nuclear layer; TRITC, tetra-methylrhodamine isothiocyanate.
2156 EPC Selectively Recruited Protective Macrophage

DISCUSSION
In this study, we showed that Alde-Low EPC effectivelyrecruited monocyte-derived macrophages by secreting CCL2and these recruited cells exerted vasoprotective and neuropro-tective effects in the retina. This remarkable vascular protec-tive capability might be useful in neurodegenerative as well
as in neural ischemic diseases. Alde-Low EPCs were effectivein acute ischemic brain injury [20]. Alde-Low EPCs alsoshowed neuroprotective effects equivalent to that observedafter injecting neurotrophic factors, such as ciliary neurotro-phic factor (CNTF) (Supporting Information Fig. 2). CNTF isthe only known factor with evident neuroprotective effects inthe rd1 mouse [6]. Thus, intravitreal CNTF injection wasused as a positive control for neuroprotection.
Figure 5. Characterization of migrated macrophages. (A): Confocal imaging of Alde-Low EPC (top) and PBS (bottom) intravitreally injected flatmount retinas (left, green: CD11b) (left middle, green: IB4; red: F4/80) (right middle, green: IB4; red: Ly6c) (right, green: CX3CR1; red: F4/80).Scale bar 5 50 mm. (B): Retinal cross-sections intravitreally injected with Alde-Low EPC (top) and PBS (bottom) (red: Ly6c; green: IB4; blue:DAPI-stained nuclei). Scale bar 5 200 mm. (C, D): Flow cytometric analysis of F4/801/Ly6c1 cells in Alde-Low EPC (white bar), CCL2 knock-down in Alde-Low EPC by shRNA (Alde-Low EPC kd EPC, white bar), Alde-Low EPC with clodronate liposome (Alde-Low EPC 1 Liposome,white bar), Alde-High EPC (gray bar), Alde-High EPC with CCL2 retrovirally introduced (Alde-High CCL2 EPC, gray bar), HUVEC (striped bar),and PBS (black bar) intravitreally injected retina. (D): The ratio of F4/801/Ly6c1 cells in retina after cells or PBS injection (mean 6 SD) (n 5 3).(E): The mRNA expression levels of the factors in F4/801/Ly6c1 cells and F4/801/Ly6c2 cells were measured by quantitative reverse transcriptasepolymerase chain reaction and expressed as histograms (mean 6 SD) (n 5 3 per group). (F): Retinal cross-sections intravitreally injected with Alde-Low. Migrated macrophages were positive with IL-10 and IGF-1 (red: IL-10; green: Ly6c, left) (red: IGF-1; green:Ly6c, right). Scale bar 5 100mm. *, p < .05; **, p < .01 by Student’s t test (E) and ANOVA with Bonferroni multiple comparison (D). Abbreviations: Alde-Low-EPC, EPCwith low ALDH activity; CCR2, chemokine C-C motif receptor 2; DAPI, 40,6-diamidino-2-phenylindole; FSC, forward scatter; GCL, ganglion celllayer; HUVEC, human umbilical vein endothelial cells; IGF-1, insulin-like growth factor type 1; IL-10, interleukin-10; INL, inner nuclear layer;ONL, outer nuclear layer; SSC, side scatter; TGF-b, transforming growth factor-b; TNF-a, tumor necrosis factor-a.
Fukuda, Nagano, Yamashita et al. 2157
www.StemCells.com
However, in previous and the current studies, injected neu-rotrophic factor was used at a very high concentration, whichwould be difficult to adjust for actual clinical use [6] (Support-ing Information Fig. 2, Supporting Information Methods).Moreover, injected Alde-Low did not produce any complica-tions, such as retinal detachment, abnormal neovascularization,hemorrhage, or strong immune reactions.
A hierarchy of EPC based on their proliferative potentialhas been reported [39]. The concepts of what constitutes EPCshave become increasingly complex and confusing and the defi-nition of an EPC has been controversial. Thus, the methodsused to isolate EPCs are highly variable among investigators[39]. Given the therapeutic potential of EPCs, efficient isolationof highly proliferative EPCs is central to the generation of a
Figure 6. Monocyte-derived macrophages in peripheral blood contribute to the neuroprotective and vasoprotective effects. (A, B): Flow cyto-metric analysis of F4/801/Ly6c1 cells in peripheral blood after Alde-Low EPC (white bar), CCL2 knockdown in Alde-Low EPC by shRNA(Alde-Low EPC kd EPC, white bar), Alde-High EPC (gray bar), Alde-High EPC with CCL2 retrovirally introduced (Alde-High CCL2 EPC, graybar), HUVEC (striped bar), HUVEC with CCL2 retrovirally introduced (HUVEC-CCL2, striped bar), and PBS (black bar) intravitreal injection(mean 6 SD) (n 5 3 per group). (C, D): Retinal cross-sections of F4/801Ly6c1 cells form peripheral blood subretinally injected eyes (red: F4/80;green: IB4; blue: DAPI-stained nuclei). Arrow indicates pierced site of 33-gauge needle, and arrowheads indicate injected region of cells. Scalebar 5 100 mm. (E): The ONL thickness at P17 of PBS or F4/801Ly6c1 cells subretinally eyes (white bar: F4/801Ly6c1 cells injection; blackbar: PBS injection) (mean 6 SD) (n 5 3 per group). (F): Retinal cross-sections of eyes subretinally injected with F4/801Ly6c1 cells (red: IL-10;green: IB4, top) (red: IGF-1; green: IB4,bottom). Scale bar 5 100 mm. (G, H): Retinal vasculature of deep layer in whole-mounted retinas (red:TRITC-conjugated lectin) of F4/801Ly6c1 cells injected eye (left) and contralateral PBS injected eye (right) (white bar: F4/801Ly6c1 cellsinjection; black bar: PBS injection) (mean 6 SD). (n 5 3 per group). Scale bar 5 200 mm. **, p < .01 by Student’s t test (E, H) and ANOVAwith Bonferroni multiple comparison (B). Abbreviations: Alde-Low-EPC, EPC with low ALDH activity; Alde-High-EPC, EPC with high ALDHactivity; DAPI, 40,6-diamidino-2-phenylindole; FSC, forward scatter; HUVEC, human umbilical vein endothelial cells; INL, inner nuclear layer;IL-10, interleukin-10; ONL, outer nuclear layer; PBS, phosphate buffered saline; SSC, side scatter.
2158 EPC Selectively Recruited Protective Macrophage

reliable and safe cell-based therapy. We identified highly prolif-erative EPC based on their ALDH activity, which we referredto as “Alde-Low EPC.” By flow cytometry analysis, the fre-quency of Alde-Low EPC in umbilical cord blood was[mt]35% [12,19]. It has also been reported that transplantedEPC not only induced the recruitment of monocytes/macro-phages and promoted neovascularization but also accelerateddermal wound healing [15,19,40,41].
Importantly, injecting CCL2-transfected HUVEC did notaffect the migration of monocytes/macrophages and showed novasoprotective/neuroprotective effects in the retina (Fig. 4E),which suggested that CCL2 alone was not sufficient for inducingthe migration of monocytes/macrophages to the retina. In con-trast, injecting CCL2-transfected Alde-High EPC induced anincreased number of migrated monocytes/macrophages (Fig. 4D)which, in turn, had vasoprotective/neuroprotective effects in theretina. However, the protective effects achieved after transplant-ing CCL2-transfected Alde-High EPC were not as good as thatachieved when transplanting Alde-Low EPC (Fig. 4F, 4G), whichsuggested that some unknown factors might be involved in pro-viding effective protection. Transplanting CCL2-transfectedAlde-Low EPCs did not increase the numbers of migrated mono-cytes/macrophages as compared with injecting Alde-Low EPCalone (Fig. 4D), possibly because the amount of secreted CCL2needed for migration may have become saturated.
We also found that Alde-Low EPC recruited monocytes/macrophages (Fig. 3B) and resulted in the early recoveryfrom vessel occlusion [20]. Altogether, these findings suggestthat EPC may be effective in two distinct ways: in a directmanner in which EPCs themselves are incorporated into neo-vessels and in an indirect manner in which EPC induce themigration of monocytes/macrophages to impaired sites. We
found that Alde-Low EPC transplanted into the retina oper-ated in the indirect rather than the direct manner.
Clodronate liposomes reduce the recruitment of BM-derived macrophages into the peripheral blood but do notreduce the numbers of tissue-resident microglial cells [26,27].Clodronate liposomes completely abolished the vasoprotec-tive/neuroprotective effects of Alde-Low EPC transplantation(Fig. 4E), which suggested that migrated monocytes/macro-phages from the BM were primarily responsible for the pro-tection against retinal degeneration. It has been reported thatEPCs are different from CD452/CD141 “colony formingunit-ECs” that differentiate into macrophages [19,42]. There-fore, it was unlikely that transplanted EPC had differentiatedinto macrophages at the transplantation site.
Resident monocytes/macropahges play significant patho-genic roles in inflammatory diseases [43–46], including retini-tis pigmentosa [34]. However, these reports only investigatedresident monocytes/macropahges and did not assess circulat-ing monocytes or macrophages derived from the BM. Circu-lating monocytes/macrophages can be categorized into severalsubsets and there are two major monocyte/macrophage sub-sets: Ly-6c1/CX3CR1lo/CCR21 cells, which infiltrate injuredtissues; and Ly-6c2/CX3CR1hi/CCR22 cells, which primarilycontribute to tissue-resident macrophages [38]. In this study,migrated monocyte-derived macrophages (F4/801/Ly6c1
cells) expressed IGF-1, one of the endogenous neurotrophicfactors, to a greater extent than resident microglial cells (F4/801/Ly6c2 cells; Fig. 5E, 5F). Ly6c is expressed on mono-cytes in the BM and shortly after their migration into the cir-culation, which suggests that Ly6c might be a useful markerfor isolating protective monocytes/macrophages from theperipheral blood (Fig. 7E).
CCL2 and CCR2 are important for recruiting Ly6c1
monocytes from the BM, as CCL2 and CCR2 deficiencieshave been shown to disrupt Ly6c1 monocytes entry into theblood or inflammatory tissues [47,48]. CCL2/CCR2 signalingis also crucial for upregulating IGF-1 mRNA expression inmacrophages. Recently, disrupted recruitment of CCR21/Ly6chi/CD11c1 monocytes was implicated as a mechanismunderlying poor recovery in a model of spinal cord injury.Injecting large numbers of Ly6chi monocytes into the circula-tion enhanced recovery after spinal cord injury.
These monocytes express IL-10 and have an anti-inflammatory role [49], which suggests that migratingmonocyte-derived macrophages that express IL-10 could alsohave an anti-inflammatory role. Abolishing the endogenousmonocyte pool results in a reduced number of transcripts thatencode for anti-inflammatory mediators, such as TGF-b1/2and IL-10, and an increase in proinflammatory mediators[36]. As compared with resident microglial cells, migratedmonocyte-derived macrophages had higher mRNA expressionlevels for anti-inflammatory mediators and lower mRNAexpression levels for proinflammatory factors (Fig. 5E, 5F). Inretinitis pigmentosa, TNF-a expression is induced before theonset of photoreceptor cell death during retinal degeneration[34]. Taken together, these findings indicate that migratedmonocyte-derived cells are inflammation-resolving macro-phages in the retina (Fig. 7).
In this study, Cyclosporine-A was used for immunosup-pression, and we observed that Cyclosporine-A had no effecton retinal degeneration in rd1 mice neither on protection noron getting worse. Cyclosporine-A exerts its immunomodula-tory activities by its effects on T lymphocytes. However, pre-vious studies reported that Cyclosporin-A had effects onmacrophage, such as decreased IL-6 production [50]. Thus,Cyclosporine-A could possibly have had some effects onmicroglial cells or macrophages.
Figure 7. Schematic summarizing the study. Migrated monocyte-derived macrophages (F4/801/Ly6c1/CCR2highcells) recruited byAlde-Low EPC through CCL2 expressed neurotrophic factors andanti-inflammatory mediators such as IGF-1, TGF-b1, and IL-10.These migrated cells were associated with neuroprotection and vaso-protection. On the other hand, residual microglia (F4/801/Ly6c2/CCR2Low cells) expressed TNF-a and was associated with phagocyto-sis of apoptotic photoreceptors. Abbreviations: Alde-Low-EPC, EPCwith low aldehyde content; CCR2, chemokine C-C motif receptor;EPC, endothelial progenitor cell; GCL, ganglion cell layer; INL, innernuclear layer; IGF-1, insulin-like growth factor type 1; IL, interleukin;ONL, outer nuclear layer; TGF-b, transforming growth factor-b;TNF-a, tumor necrosis factor-a.
Fukuda, Nagano, Yamashita et al. 2159
www.StemCells.com

CONCLUSION
In conclusion, intraocular injection of Alde-Low EPC effec-tively recruited inflammation-resolving and neurovascular pro-tective monocyte-derived macrophages that delay theprogression of neural degenerative disease.
ACKNOWLEDGMENTS
This work was supported by Grants-in-Aid for ScientificResearch from the Japanese Ministry of Education, Culture,
Sports, Science, and Technology, Grant-in-Aid for JSPS Fel-lows, and a Research Grant from the Study Group on Choriore-tinal Degeneration and Optic Atrophy, the Ministry of Health,Labor and Welfare, Japan. This article was presented in part atAssociation for Research in Vision and Ophthalmology AnnualMeeting, May 2011 and 2012, Fort Lauderdale, FL.
DISCLOSURE OF POTENTIAL
CONFLICTS OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Hartong DT, Berson EL, Dryja TP. Retinitis pigmentosa. Lancet 2006;368:1795–1809.
2 Humphries P, Kenna P, Farrar GJ. On the molecular genetics of retini-tis pigmentosa. Science 1992;256:804–808.
3 Bramall AN, Wright AF, Jacobson SG et al. The genomic, biochemi-cal, and cellular responses of the retina in inherited photoreceptordegenerations and prospects for the treatment of these disorders. AnnuRev Neurosci 2010;33:441–472.
4 Cideciyan AV. Leber congenital amaurosis due to RPE65 mutationsand its treatment with gene therapy. Prog Retin Eye Res 2010;29:398–427.
5 Frasson M, Picaud S, Leveillard T et al. Glial cell line-derived neuro-trophic factor induces histologic and functional protection of rod pho-toreceptors in the rd/rd mouse. Invest Ophthalmol Vis Sci 1999;40:2724–2734.
6 LaVail MM, Yasumura D, Matthes MT et al. Protection of mousephotoreceptors by survival factors in retinal degenerations. Invest Oph-thalmol Vis Sci 1998;39:592–602.
7 Inoue Y, Iriyama A, Ueno S et al. Subretinal transplantation of bone mar-row mesenchymal stem cells delays retinal degeneration in the RCS ratmodel of retinal degeneration. Exp Eye Res 2007;85:234–241.
8 MacLaren RE, Pearson RA, MacNeil A et al. Retinal repair by trans-plantation of photoreceptor precursors. Nature 2006;444:203–207.
9 Otani A, Dorrell MI, Kinder K et al. Rescue of retinal degeneration byintravitreally injected adult bone marrow-derived lineage-negativehematopoietic stem cells. J Clin Invest 2004;114:765–774.
10 Meyer JS, Katz ML, Maruniak JA et al. Embryonic stem cell-derivedneural progenitors incorporate into degenerating retina and enhancesurvival of host photoreceptors. Stem Cells 2006;24:274–283.
11 Yang P, Seiler MJ, Aramant RB et al. Differential lineage restrictionof rat retinal progenitor cells in vitro and in vivo. J Neurosci Res2002;69:466–476.
12 Asahara T, Murohara T, Sullivan A et al. Isolation of putative progeni-tor endothelial cells for angiogenesis. Science 1997;275:964–967.
13 Gehling UM, Ergun S, Schumacher U et al. In vitro differentiation ofendothelial cells from AC133-positive progenitor cells. Blood 2000;95:3106–3112.
14 Peichev M, Naiyer AJ, Pereira D et al. Expression of VEGFR-2 andAC133 by circulating human CD34(1) cells identifies a population offunctional endothelial precursors. Blood 2000;95:952–958.
15 Rafii S, Lyden D. Therapeutic stem and progenitor cell transplantationfor organ vascularization and regeneration. Nat Med 2003;9:702–712.
16 Murohara T, Ikeda H, Duan J et al. Transplanted cord blood-derivedendothelial precursor cells augment postnatal neovascularization. JClin Invest 2000;105:1527–1536.
17 Shi Q, Rafii S, Wu MH et al. Evidence for circulating bone marrow-derived endothelial cells. Blood 1998;92:362–367.
18 Hristov M, Erl W, Weber PC. Endothelial progenitor cells: Mobiliza-tion, differentiation, and homing. Arterioscler Thromb Vasc Biol 2003;23:1185–1189.
19 Nagano M, Yamashita T, Hamada H et al. Identification of functionalendothelial progenitor cells suitable for the treatment of ischemic tis-sue using human umbilical cord blood. Blood 2007;110:151–160.
20 Nakamura K, Tsurushima H, Marushima A et al. A subpopulation ofendothelial progenitor cells with low aldehyde dehydrogenase activityattenuates acute ischemic brain injury in rats. Biochem Biophys ResCommun 2012;418:87–92.
21 Fan Y, Shen F, Frenzel T et al. Endothelial progenitor cell transplanta-tion improves long-term stroke outcome in mice. Ann Neurol 2010;67:488–497.
22 Ohta T, Kikuta K, Imamura H et al. Administration of ex vivo-expandedbone marrow-derived endothelial progenitor cells attenuates focal cere-bral ischemia-reperfusion injury in rats. Neurosurgery 2006;59:679–686.
23 Jeong JO, Kim MO, Kim H et al. Dual angiogenic and neurotrophiceffects of bone marrow-derived endothelial progenitor cells on diabeticneuropathy. Circulation 2009;119:699–708.
24 Gavrieli Y, Sherman Y, Ben-Sasson SA. Identification of programmedcell death in situ via specific labeling of nuclear DNA fragmentation.J Cell Biol 1992;119:493–501.
25 Ueno S, Kondo M, Miyata K et al. Physiological function of S-conesystem is not enhanced in rd7 mice. Exp Eye Res 2005;81:751–758.
26 van Rooijen N, Sanders A, van den Berg TK. Apoptosis of macro-phages induced by liposome-mediated intracellular delivery of clodro-nate and propamidine. J Immunol Methods 1996;193:93–99.
27 Sasahara M, Otani A, Oishi A et al. Activation of bone marrow-derived microglia promotes photoreceptor survival in inherited retinaldegeneration. Am J Pathol 2008;172:1693–1703.
28 Xu G, Nagano M, Kanezaki R et al. Frequent mutations in the GATA-1 gene in the transient myeloproliferative disorder of Down syndrome.Blood 2003;102:2960–2968.
29 Bowes C, Li T, Danciger M et al. Retinal degeneration in the rdmouse is caused by a defect in the beta subunit of rod cGMP-phospho-diesterase. Nature 1990;347:677–680.
30 Pittler SJ, Baehr W. Identification of a nonsense mutation in the rodphotoreceptor cGMP phosphodiesterase beta-subunit gene of the rdmouse. Proc Natl Acad Sci USA 1991;88:8322–8326.
31 Kaneko H, Nishiguchi KM, Nakamura M et al. Retardation of photore-ceptor degeneration in the detached retina of rd1 mouse. Invest Oph-thalmol Vis Sci 2008;49:781–787.
32 Chang B, Hawes NL, Hurd RE et al. Retinal degeneration mutants inthe mouse. Vision Res 2002;42:517–525.
33 Zhang Y, Ingram DA, Murphy MP et al. Release of proinflammatorymediators and expression of proinflammatory adhesion molecules byendothelial progenitor cells. Am J Physiol Heart Circ Physiol 2009;296:H1675–1682.
34 Zeng HY, Zhu XA, Zhang C et al. Identification of sequential eventsand factors associated with microglial activation, migration, and cyto-toxicity in retinal degeneration in rd mice. Invest Ophthalmol Vis Sci2005;46:2992–2999.
35 Harrison JK, Jiang Y, Chen S et al. Role for neuronally derived frac-talkine in mediating interactions between neurons and CX3CR1-expressing microglia. Proc Natl Acad Sci USA 1998;95:10896–10901.
36 London A, Itskovich E, Benhar I et al. Neuroprotection and progenitorcell renewal in the injured adult murine retina requires healingmonocyte-derived macrophages. J Exp Med 2011;208:23–39.
37 Leenen PJ, Melis M, Slieker WA et al. Murine macrophage precursorcharacterization. II. Monoclonal antibodies against macrophage precur-sor antigens. Eur J Immunol 1990;20:27–34.
38 Geissmann F, Jung S, Littman DR. Blood monocytes consist of twoprincipal subsets with distinct migratory properties. Immunity 2003;19:71–82.
39 Ingram DA, Mead LE, Tanaka H et al. Identification of a novel hierar-chy of endothelial progenitor cells using human peripheral and umbili-cal cord blood. Blood 2004;104:2752–2760.
40 van der Strate BW, Popa ER, Schipper M et al. Circulating humanCD341 progenitor cells modulate neovascularization and inflammationin a nude mouse model. J Mol Cell Cardiol 2007;42:1086–1097.
41 Suh W, Kim KL, Kim JM et al. Transplantation of endothelial progen-itor cells accelerates dermal wound healing with increased recruitmentof monocytes/macrophages and neovascularization. Stem Cells 2005;23:1571–1578.
42 Yoder MC, Mead LE, Prater D et al. Redefining endothelial progenitorcells via clonal analysis and hematopoietic stem/progenitor cell princi-pals. Blood 2007;109:1801–1809.
2160 EPC Selectively Recruited Protective Macrophage

43 Chen L, Wu W, Dentchev T et al. Light damage inducedchanges in mouse retinal gene expression. Exp Eye Res 2004;79:239–247.
44 Jo N, Wu GS, Rao NA. Upregulation of chemokine expression in theretinal vasculature in ischemia-reperfusion injury. Invest OphthalmolVis Sci 2003;44:4054–4060.
45 Nakazawa T, Hisatomi T, Nakazawa C et al. Monocyte chemoattrac-tant protein 1 mediates retinal detachment-induced photoreceptor apo-ptosis. Proc Natl Acad Sci USA 2007;104:2425–2430.
46 Abu el-Asrar AM, Van Damme J, Put W et al. Monocyte chemotacticprotein-1 in proliferative vitreoretinal disorders. Am J Ophthalmol1997;123:599–606.
47 Lu H, Huang D, Saederup N et al. Macrophages recruited via CCR2produce insulin-like growth factor-1 to repair acute skeletal muscleinjury. FASEB J 2011;25:358–369.
48 Lu H, Huang D, Ransohoff RM et al. Acute skeletal muscle injury:CCL2 expression by both monocytes and injured muscle is requiredfor repair. FASEB J 2011;25:3344–3355.
49 Shechter R, London A, Varol C et al. Infiltrating blood-derived macro-phages are vital cells playing an anti-inflammatory role in recoveryfrom spinal cord injury in mice. PLoS Med 2009;6:e1000113.
50 Garc�ıa JE, L�opez AM, de Cabo MR et al. Cyclosporin A decreaseshuman macrophage interleukin-6 synthesis at post-transcriptional level.Mediators Inflamm 1999;8:253–259.
See www.StemCells.com for supporting information available online.
Fukuda, Nagano, Yamashita et al. 2161




















