
conservation of the critically endangered negros bleeding - AWS
Upload
independentCategory
view
2download
0
BIODIVERSITYRESEARCH
From exploitation to conservation:habitat models using whaling datapredict distribution patterns and threatexposure of an endangered whaleLeigh G. Torres1*, Tim D. Smith2, Phil Sutton1, Alison MacDiarmid1,
John Bannister3 and Tomio Miyashita4
1National Institute of Water and
Atmospheric Research Ltd., Hataitai,
Wellington, 6021, New Zealand, 2World
Whaling History, Redding, CA, USA, 3The
Western Australian Museum, Perth, WA,
6000, Australia, 4National Research Institute
of Far Seas Fisheries, Shimizu, Shizuoka,
424, Japan
*Correspondence: Leigh G. Torres, National
Institute of Water and Atmospheric Research,
Ltd (NIWA), 301 Evans Bay Parade, Greta
Point, Hataitai, Wellington 6021, New
Zealand.
E-mail: [email protected]
ABSTRACT
Aim Sufficient data to describe spatial distributions of rare and threatened
populations are typically difficult to obtain. For example, there are minimal
modern offshore sightings of the endangered southern right whale, limiting our
knowledge of foraging grounds and habitat use patterns. Using historical
exploitation data of southern right whales (SRW), we aim to better understand
their seasonal offshore distribution patterns in relation to broad-scale oceano-
graphy, and to predict their exposure to shipping traffic and response to global
climate change.
Location Australasian region between 130° W and 100° E, and 30° S and
55° S.
Methods We model 19th century whaling data with boosted regression trees
to determine functional responses of whale distribution relative to environmen-
tal factors. Habitat suitability maps are generated and we validate these predic-
tions with independent historical and recent sightings. We identify areas of
increased risk of ship-strike by integrating predicted whale distribution maps
with shipping traffic patterns. We implement predicted ocean temperatures for
the 2090–2100 decade in our models to predict changes in whale distribution
due to climate change.
Results Temperature in the upper 200 m, distance from the subtropical front,
mixed layer depth, chlorophyll concentration and distance from ridges are the
most consistent and influential predictors of whale distribution. Validation tests
of predicted distributions determined generally high predictive capacity. We
identify two areas of increased risk of vessel strikes and predict substantial
shifts in habitat suitability and availability due to climate change.
Main conclusions Our results represent the first quantitative description of
the offshore foraging habitat of SRW. Conservation applications include identi-
fying areas and causes of threats to SRW, generating effective mitigation strate-
gies, and directing population monitoring and research efforts. Our study
demonstrates the benefits of incorporating unconventional datasets such as
historical exploitation data into species distribution models to inform manage-
ment and help combat biodiversity loss.
Keywords
Boosted regression trees, distribution patterns, global climate change, habitat
use, historical data, rare species, southern right whale, species distribution
models.
DOI: 10.1111/ddi.120691138 http://wileyonlinelibrary.com/journal/ddi ª 2013 John Wiley & Sons Ltd
Diversity and Distributions, (Diversity Distrib.) (2013) 19, 1138–1152A
Jou
rnal
of
Cons
erva
tion
Bio
geog
raph
yD
iver
sity
and
Dis
trib
utio
ns

INTRODUCTION
Biodiversity loss from marine ecosystems is an extreme prob-
lem with widespread impacts (Worm et al., 2006; Estes et al.,
2011). Knowledge of species distribution patterns is essential
for assessing threats to biodiversity that range from fine-scale
risks such as fisheries (Thrush & Dayton, 2002) to broad-
scale habitat alterations like climate change (Trathan et al.,
2007). Ecological-niche modelling has become an integral
part of species conservation, including identification of
important habitats, reserve planning, species reintroduction
programs and ecosystem restoration (Hirzel et al., 2006).
Based on the assumption that the distribution of encounters
reflects a species’ environmental preferences (Guisan &
Zimmermann, 2000; Hirzel et al., 2006), a variety of statisti-
cal techniques are used to associate distribution patterns with
habitat variability to identify a species’ realized environmen-
tal niche (Elith et al., 2006).
For remnant populations with historically broad distribu-
tion patterns prior to exploitation, it is often difficult or
impossible to collect comprehensive distribution data across
their current and historical ranges. However, data derived
from historical sources, such as harvest or museum records,
can be used to describe historical distribution patterns (Elith
& Leathwick, 2007; Gregr, 2011). The accuracy of distribu-
tion models produced from historical datasets for modern
times depends upon the conservation of a species’ ecological
niche over time (Peterson et al., 1999).
Whaling in the South Pacific decimated southern right
whale (Eubalaena australis) populations (Jackson et al.,
2008). An estimated 40,000 southern right whales (SRW)
were killed in waters around New Zealand and east Australia
(Carroll et al., 2011a) and an unknown, but likely compara-
ble, number off southern Australia. The current population
estimate of SRW off New Zealand is just 908 (Carroll et al.,
2011b) and off southern Australia is c. 3500 (Bannister,
2011). To ensure protection of these remnant populations
from present and future threats, a reliable understanding of
SRW distribution and habitat use patterns is essential.
Southern right whales have a strong migration cycle
between coastal wintering grounds used for calving and off-
shore foraging areas. These whales are capital breeders that
use previously accumulated energy stores to nurse calves in
winter, and thus need to encounter dense aggregations of
their preferred prey items (copepods and euphausiids) within
their dive limits while foraging offshore (c. 300 m; Gregr &
Coyle, 2009). Although the wintering grounds of SRW in
Australia and New Zealand are reasonably well defined (Ban-
nister, 2001; Patenaude & Baker, 2001), little is known about
their offshore and migratory habitat preferences (Ohsumi &
Kasamatsu, 1986; Bannister et al., 1999; Richards, 2002).
Despite the end of commercial whaling, SRW are not free
from anthropogenic threats. SRW in the Australasian region
are prone to ship strikes, with a single mortality negatively
affecting population vitality in such a small population
(Kemper et al., 2008). In addition, SRW may be subject to
oceanic changes due to climate change. The distribution
patterns of North Atlantic right whales (Eubalaena glacialis)
and their copepod prey are affected by climate change
(Greene & Pershing, 2004) and the breeding success of SRW
in the South Atlantic shows sensitivity to climate variation
(Leaper et al., 2006).
Historical whaling data have been used to examine diet
from stomach contents (Tormosov et al., 1998), describe
general distribution patterns (Townsend, 1935; Richards,
2002), generate habitat characterizations or models based on
presence-only data (Jaquet et al., 1996; Gregr & Trites, 2001;
Gregr, 2011) and estimate historical population sizes (Smith,
2001). Here, we use historical whaling data, including pres-
ence and absence data from the Australasian region to exam-
ine ecological responses of SRW to environmental variability.
The dataset derives from 19th century offshore whaling
vessels, allowing our study to focus on offshore habitat use
patterns where SRW spend a majority of their time and
where their distribution patterns are least understood. We
use boosted regression trees (BRT), a machine-learning tech-
nique recently applied to species distribution modelling
(Elith et al., 2006; Leathwick et al., 2006), to define func-
tional habitat use patterns of SRW. We use these models to
predict the distribution patterns of whales both today, which
are compared with shipping traffic density, and in the year
2100, as a result of climate change. Based on the resolution
of our datasets, we expect our models to be applicable at
meso and large scales for predicting how whales respond to
habitat variation such as temperature and density gradients
that may aggregate prey (Gregr & Coyle, 2009).
METHODS
Historical whaling data
We obtained daily observations from American whaling
vessels within our study region (130° W to 100° E, and
30° S to 55° S) extracted from voyage logbooks by Maury
(1852) and recently by the Census of Marine Life (www.
coml.org/) using methods described in Smith et al. (2012).
Whalers recorded their location daily and indicated when
whales were encountered. These data amounted to 24,733
daily records of vessel location over the period 1825–88
(Fig. 1). Mean vessel location error was estimated to be
0.22° latitude (c. 24 km) and 0.54° longitude (c. 34 km;
Smith et al., 2012). Therefore, we applied a spatial resolution
of 25 km 9 25 km grid cells to all analyses.
We grouped data by season to generate predictive models
that reflect seasonal changes in whale behaviour and environ-
ment. Spring (September, October, November): whales leave
calving grounds for foraging habitat, ocean temperatures
begin to warm, spring productivity blooms occur. Summer
(December, January, February): most whales occupy foraging
habitat, ocean temperatures are at a maximum. Autumn
(March, April, May): whales may begin to migrate towards
calving habitats, ocean temperatures begin to cool. No winter
Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd 1139
Habitat models based on historical data

habitat models were generated because our historical dataset
did not include sufficient observations in those months. We
also divided our study area into two regions that grouped
prominent habitat features based on geomorphology and
currents. We split the study area into eastern and western
regions along a natural gap in encounters at 155° E (Fig. 1).
A total of 1792 encounters with SRW (presence) were mod-
elled in the two regions, across three seasons (Table 1). No
model was generated in the western region during autumn
because of limited presence data (n = 11). The southern
boundary in both regions was limited to the extent of data
points, confining predictive models to only areas with data
(eastern: 55° S; western: 53° S).
Our historical whaling data reflect the inconsistent spatial
and temporal distribution of whaling effort through time
(Smith et al., 2012). To determine the need to group data
temporally for habitat models, we examined our data for
changes in the spatial distribution of SRW encounters over
time (see Appendix S1 in Supporting Information).
Habitat modelling
We generated predictive models of SRW probability of
occurrence using BRT. Our response variable was binomial
presence/absence data composed of daily encounters with
SRW and days with no encounters. The numerical
(a)
(b)
(c)
Figure 1 Seasonal spatial distribution of historical whaling data illustrating absence (grey dots) and southern right whale presence
(black dots) locations. Spring (a), summer (b), autumn (c). Black boxes demarcate boundaries of western and eastern regions in each
plot. North arrow and scale bar in (a) applies to all plots. The Chatham Rise, Macquarie Ridge (labelled in a), and Louisville Ridge
(labelled in c) are illustrated on plots; numerous presence and absence points fall under these features. Inadequate presence data in
western region during autumn restricted analyses. Maps in Mercator projection, datum wgs1984.
1140 Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd
L. G. Torres et al.
dominance of absence data relative to presence data in all
region–season combinations (Table 1) may be due to rela-
tively low prevalence (even prior to declines in SRW abun-
dance), and to false-absences (caused by dive behaviour, sea
state and weather conditions that decrease surface visibility).
Absence data are also biased by inconsistent observer effort
across the region. Therefore, we down-weighted absence data
in each region–season model so that presence and absence
data carried equal weight, a method that has been shown to
increase model accuracy (Table 1; Barbet-Massin et al.,
2012). The following equation was applied:
# presence =# absence ¼ weight applied to each absence point
Boosted regression trees is an ensemble method that inter-
prets complex relationships between species and their envi-
ronment (Elith et al., 2006; Leathwick et al., 2006). The first
of two algorithms implemented in BRT partitions the
response variable into groups with similar characteristics
using regression or classification trees. Boosting is the second
algorithm where trees are fitted iteratively; emphasizing
observations that poorly fit the existing collection of trees.
Boosting combines these trees to minimize misclassification
errors and improve predictive performance over a single tree
model (Leathwick et al., 2006). Boosting is optimized by the
learning rate (lr) that considers residual variation during tree
building, and tree complexity (tc) that estimates interactions
between predictor variables. Model fit and predictive perfor-
mance are balanced to reduce overfitting by jointly optimiz-
ing the number of trees (nt), lr, and tc (Elith et al., 2008).
We used the ‘GBM’ package (version 1.6-3.1, Ridgeway, 2007)
implemented in R (R Development Core Team) plus custom
code available online (Elith et al., 2008) to generate BRT
models of SRW distribution.
We generated multiple BRT models for each region–
season using different sets of predictor variables, and varying
the lr (range: 0.05–0.0005) and tc (range: 1–4) to fit models
with at least 1000 trees as recommended by Elith et al. (2008).
The relative importance of a predictor variable is determined
by its contribution to the model as measured by the number of
times it is selected for tree splitting (Elith et al., 2008). Our
first step to determine an ‘optimal’ model removed all vari-
ables that contributed < 3%. Then, we assessed response–pre-
dictor relationships and removed predictor variables that
produced ecologically unrealistic relationships. Fitted-func-
tions produced by BRT show the effect of each predictor vari-
able on the response while controlling for the average effect of
all other variables in the model (Buston & Elith, 2011). Finally,
we compared two performance metrics: area under the receiver
operating curve (AUC) and cross-validation per cent deviance
explained (CVdev). AUC is widely used to evaluate binomial
models (Fawcett, 2006) by measuring the ability to discrimi-
nate between areas with species presence or absence. AUC
ranges from 0 to 1: 1 = perfect discrimination, < 0.5 = worse
than random, > 0.7 is considered ‘useful’ (Swets, 1988).
Although AUC has limitations for measuring model perfor-
mance (Austin, 2007; Lobo et al., 2008), we use it as a relative
metric of model performance because it provides a single value
that is easy to interpret. CVdev is estimated by a cross-valida-
tion procedure run during the modelling process that with-
holds a subset of data at each tree. CVdev indicates how well
the model predicted the subsets of withheld data (Buston &
Elith, 2011).
Environmental layers
We assigned each presence and absence location values
from 23 environmental layers based on location and season
when relevant (Table 2). We derived mean, range and stan-
dard deviation values of depth and slope from the General
Bathymetric Chart of the World (http://www.gebco.net/).
We generated layers of these bathymetry statistics using a
12.5-km radius neighbourhood to correspond with the
assumed spatial accuracy of the whaling data
(25 km 9 25 km grid cells), and applied a buffer zone
around the boundary of the study region to minimize edge
effects. We generated a regional geomorphology map that
grouped areas of the seafloor into defined features based on
bathymetric parameters (See Appendix S2). We merged this
geomorphic map with a global layer of seamount locations
(Yesson et al., 2011) to produce a final geomorphology
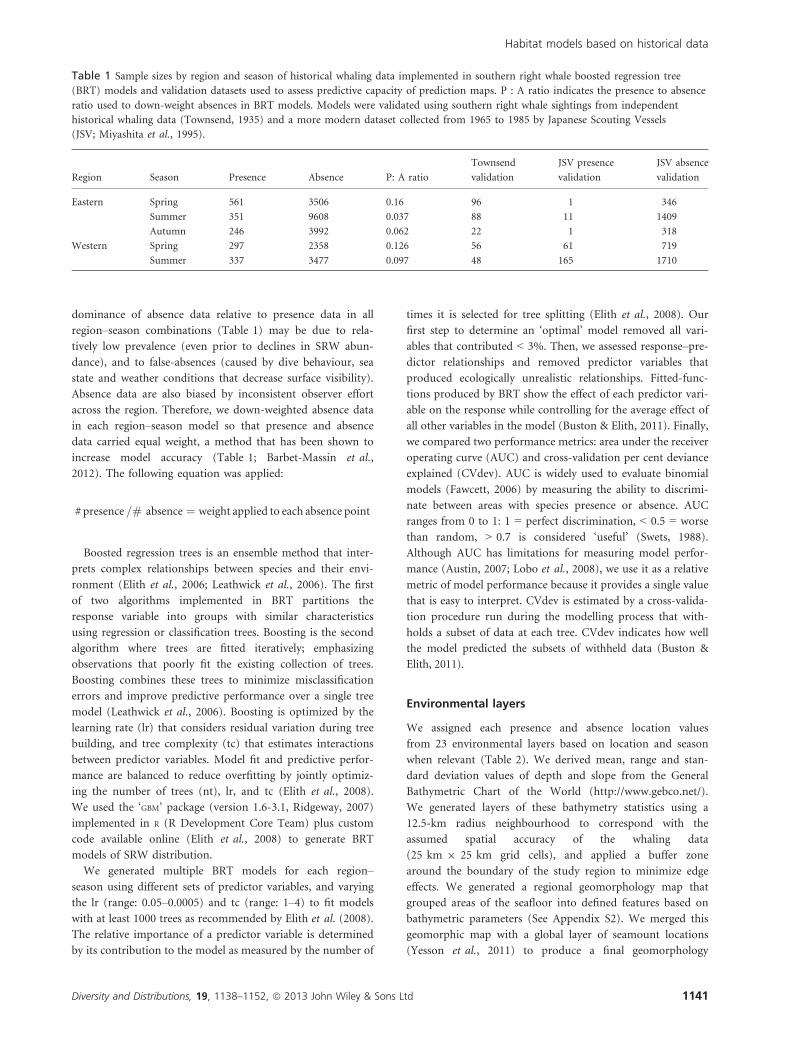
Table 1 Sample sizes by region and season of historical whaling data implemented in southern right whale boosted regression tree
(BRT) models and validation datasets used to assess predictive capacity of prediction maps. P : A ratio indicates the presence to absence
ratio used to down-weight absences in BRT models. Models were validated using southern right whale sightings from independent
historical whaling data (Townsend, 1935) and a more modern dataset collected from 1965 to 1985 by Japanese Scouting Vessels
(JSV; Miyashita et al., 1995).
Region Season Presence Absence P: A ratio
Townsend
validation
JSV presence
validation
JSV absence
validation
Eastern Spring 561 3506 0.16 96 1 346
Summer 351 9608 0.037 88 11 1409
Autumn 246 3992 0.062 22 1 318
Western Spring 297 2358 0.126 56 61 719
Summer 337 3477 0.097 48 165 1710
Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd 1141
Habitat models based on historical data

map of the Australasian region with the following feature
classes: shelf, slope, rise, plain, valley, trench, trough, basin,
hills(s), mountains(s), ridges(s), plateau, seamount. We used
ARCGIS (v9.3; ESRI, Redlands, CA, USA) to calculate grids
of distance from slope, trench, ridge, seamount, shelf, land,
valley and trough.
We derived values of dynamic environmental variables
from seasonal oceanographic and remote sensing climatolo-
gies. We recognized the temporal difference between the
whaling data and these modern oceanographic data. How-
ever, no large-scale change in the basic oceanographic struc-
ture of the region has been observed within the last
200 years (Hendy et al., 2002; Nott et al., 2007). Thus, we
assume that spatial and temporal environmental variability
has not changed substantially between time periods and that
the ecological relationships between whales and environmen-
tal variability are temporally consistent.
CSIRO Atlas of Regional Seas (CARS) is a global gridded
climatology of oceanographic properties in 0.5° grid cells
and average seasonal cycles that integrates all available ocean-
ographic profile data (Dunn & Ridgeway, 2002). While this
mapping may not resolve small-scale features to which
whales may respond, these oceanographic data are appropri-
ate for our study, which aimed to model and predict large-
scale distribution patterns of whales relative to general
habitat structure (i.e., fronts), not short-term oceanographic
variations. CARS provided layers of ocean temperatures, cur-
rents and mixed layer depth. Temperatures were extracted at
multiple depths: 0 m (surface), mean between 0 and 100 m,
100 m, mean between 0 and 200 m, and 200 m. Standard
Mapped Images of SeaWifs monthly climatology (http://
oceandata.sci.gsfc.nasa.gov/) were used to evaluate variability
in surface chlorophyll.
The Subtropical Front is a dominant feature in the South-
ern Ocean (Stramma et al., 1995) and has been anecdotally
associated with the foraging grounds of SRW (Bannister,
2001; Richards, 2002). We defined the Subtropical Front by
the position of the 12 °C isotherm in the mean 0–100 m
temperature layer during summer and autumn and the 10 °Cisotherm during spring. These values accurately located the
front and are consistent with the temperature values quoted
by Chiswell (1994) and Heath (1985).
We recognized high correlation between some of the
environmental layers implemented in the models (i.e., tem-
perature layers). When model results produced high contri-
butions (> 3%) by correlated variables we ran iterative
models with only one variable from each set of correlated
variables and compared performance measures between
models. The model with the best performance measures was
considered optimal, regardless of whether it did or did not
include correlated variables.
Model predictions and validations
We generated predictive maps of SRW distribution based on
optimal BRT models by implementing layers of each predic-
tor variable, gridded to 0.5° resolution. Each grid cell was
assigned a predicted value of habitat suitability (0–1) accord-
ing to the model’s fitted-functions and habitat characteristics
in each grid cell.
Predictor variable Acronym Derived from
Geomorphic class (categorical) geo_class Geomorphology layer
Chlorophyll a concentration (mg m�3) chla SeaWifs monthly climatology
Temperature at 0 m (°C) t0 CARS
Mean temperature between 0 and 100 m (°C) t0_100 CARS
Temperature at 100 m (°C) t100 CARS
Mean temperature between 0 and 200 m (°C) t0_200 CARS
Temperature at 200 m (°C) t200 CARS
Mixed layer depth (m) mld CARS
Distance from the subtropical front (km) dist_stf CARS
Distance from slope (m) dist_slope Geomorphology layer
Distance from trench (m) dist_trench Geomorphology layer
Distance from ridge (m) dist_ridge Geomorphology layer
Distance from seamount (m) dist_seamount Geomorphology layer
Distance from shelf (m) dist_shelf Geomorphology layer
Distance from land (m) dist_land Geomorphology layer
Distance from valley (m) dist_valley Geomorphology layer
Distance from trough (m) dist_trough Geomorphology layer
Mean depth (m) depth_mean GEBCO 08, at 25 km2
Standard deviation of depth (m) depth_std GEBCO 08, at 25 km2
Range of depth (m) depth_range GEBCO 08, at 25 km2
Mean slope (degrees) slope_mean GEBCO 08, at 25 km2
Standard deviation of slope (degrees) slope_std GEBCO 08, at 25 km2
Range of slope (degrees) slope_range GEBCO 08, at 25 km2
Table 2 Predictor variables implemented
in boosted regression tree habitat models
of southern right whale distribution.
Geomorphic classes include shelf, slope,
rise, plain, valley, trench, trough, basin,
hill(s), mountains(s), ridge(s), plateau
and seamount (See Appendix S2). CARS
implies CSIRO Atlas of Regional Seas.
GEBCO 08 implies General Bathymetric
Chart of the World 2008.
1142 Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd
L. G. Torres et al.

Few recent offshore sightings of SRW have been reported
in our study region (See Appendix S3). Therefore, we vali-
dated each prediction map using two datasets where each
presence and absence (or pseudo-absence, see below) point
sampled the appropriate prediction layer to obtain values of
habitat suitability. We used these values to determine each
map’s predictive capacity by calculating the AUC, sensitivity
(true positive rate), and 1�specificity (false positive rate).
First, we used data collected by Townsend (1935) from
other whaling logbooks to evaluate the representativeness of
our modelled dataset. We did not include these data in the
modelling process because absence data were not collected.
The Townsend data included 310 encounters with SRW
over the same spatial extent and similar time period (1831–
1907) as the modelled whaling data (Table 1). For each
region-season prediction map, we randomized 10,000
validation tests using all the presence locations in that
region-season against subsampled, randomly placed pseudo-
absences in equal number to the presence data. Second, we
used survey data collected by Japanese Scouting Vessels
(JSV) from 1965 to 1985 (Miyashita et al., 1995) to deter-
mine if models derived from historical data are consistent
over time. These data consisted of vessel noon positions
when SRW were (presence) and were not (absence) encoun-
tered. Few SRW were encountered in the eastern region.
Absence days greatly exceeded presence in all regions
(Table 1, Fig. 3). To minimize the effect of this disparity,
for each season, we randomized 10,000 validation tests
using all presence locations against a subsample of JSV
absence points of equal size.
A global map of commercial shipping traffic (Halpern
et al., 2008) was used to identify areas of collision risk with
SRW. This map represents ship density per 1 km2 grid cells
and ranges from 0 to 1158, which is the maximum number
of ship tracks per grid cell globally. We spatially calculated a
risk index of vessel collision with an SRW using Raster
Calculator (ARCGIS, v9.3; ESRI) and the formula:
SRW habitat suitability prediction � ship densityð Þ=1158ð Þ� 100
These values reflect the predicted probability of whale
presence and the relative level of ship traffic density globally
to provide a comparative risk index of collision between the
regions and seasons modelled. Similar to a spatial assessment
between north Atlantic right whale habitat and shipping traf-
fic (Ward-Geiger et al., 2005), we assume no seasonality to
shipping traffic. To assess the impact of global climate
change on SRW distribution patterns, we generated predic-
tive maps by replacing modern layers of ocean temperature
with predicted temperature layers for the period January
2090–December 2100. We derived predicted ocean tempera-
ture data from the ‘business as usual’ emissions scenario of
the CM2.1 climate model (Delworth et al., 2006; Gnanadesi-
kan et al., 2006; See Appendix S4).
RESULTS
Although the distribution of whaling effort changed over
time (Smith et al., 2012), we detected no distributional shift
of SRW (see Appendix S1) and generated habitat models for
each region-season without partitioning whaling data by
temporal period.
Predictor variables were removed from the model if they
had minimal contributions (< 3%) or the associated fitted-
functions were not ecologically meaningful. For instance,
distance from slope contributed to some initial models but
the fitted-function described increasing whale presence with
increasing distance (beyond 1000 km). While this relation-
ship may be true spatially, whales are unlikely to respond
ecologically at such a distance. Other variables removed from
models for lack of ecological realism were distance from
seamount, distance from shelf, distance from trench and
distance from land.
Optimal models in each region-season were also deter-
mined based on performance measures calculated during the
BRT routine (mean AUC = 0.846; mean CVdev = 0.357;
Table 3). In optimal models, complexity(tc) was two or
three, number of trees (nt) ranged from 1250 to 1750, and
the learning rate (lr) ranged from 0.00125 to 0.005. These
models included similar combinations of predictor variables
(Table 3). The mean temperature between 0 and 200 m
(t0_200) had high contributions to all five models (mean
contribution: 42.5%). Mixed layer depth (mld) was also
included in every model, but with minor contributions
(mean 7.6%). All models but eastern-spring included dis-
tance from the Subtropical Front (dist_stf; mean contribu-
tion: 28.1%). Chlorophyll concentration (chla) contributed
to three models (mean contribution: 14.5%). Mean depth
(depth_mean) was included in three models (mean contribu-
tion: 12.5%). Distance from ridge (dist_ridge) contributed to
all seasonal models in the eastern region (mean contribution:
13.6%). Temperature at the surface (t0) only contributed to
the western-spring model (15.2%), in addition to a large
contribution by t0_200.
Although fitted functions are not perfect representations
of species–environment relationships, they provide a useful
basis for interpretation (Elith et al., 2008) from which we
can surmise SRW habitat preferences (Fig. 2). Habitat use by
SRW appears to be strongly influenced by t0_200; a prefer-
ence for habitats where t0_200 is between 9 and 11 °C is
indicated. The maximum temperature at which SRW pres-
ence was predicted to occur was 14 °C in the eastern-spring
when whales were at their northern limit. The minimum
preferred temperature could not be determined due to a lack
of sufficient data from habitats with temperatures < 9 °C.Fitted-functions for all models imply that the presence of
SRW was most associated with habitat where the mld was
< 30 m, except in the western-summer model where habitats
with mld between 30 and 50 m were preferred. Relationships
with dist_stf derived from all models indicate a preference
for areas close to the Subtropical Front. For instance, SRW
Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd 1143
Habitat models based on historical data

presence was predicted to decline beyond 200 km (eastern-
summer) or 100 km (eastern-autumn) north of the front
and suggested distinct peaks at the front (dist_stf = 0). Rela-
tionships with dist_ridge indicated increased SRW presence
with proximity to the ridge. A maximum contribution
(27.8%) was made to the eastern-spring model where high-
suitability habitat surrounded the Louisville Ridge (Fig. 3a).
In autumn, whale presence declined dramatically beyond
250 km from the ridge; here the ridge of consequence is the
Chatham Rise that limits the northern extent of the Subtrop-
ical Front (Chiswell, 1994). During summer, as whales
transition between spring and autumn distributions, high-
suitability habitat encompassed the southern portion of
the Louisville Ridge, south of Chatham Rise, and
southwest of mainland New Zealand near the Macquarie
Ridge (Fig. 3a–c).
Except for the western-spring model, validation tests using
the Townsend historical data indicated high predictive capac-
ity for all models (mean AUC = 0.820, mean sensitiv-
ity = 0.56, mean 1�specificity = 0.05; Table 3). In all
models, a majority of SRW encounters coincided with areas
predicted to have high habitat suitability (stars in Fig. 3).
The validation test of the western-spring model indicated a
relatively low true positive rate (sensitivity = 0.41) but main-
tained a low false-positive rate (1�specificity = 0.03); com-
bining this led to relatively low overall discrimination
(AUC = 0.66).
Validation of the western-spring model with the modern
JSV data determined high predictive capacity (Table 3).
Despite the vast majority of SRW encounters occurring
within the east–west band of predicted high suitability
(Fig. 3j), validation of the western-summer model performed
poorly (AUC = 0.52). Limited JSV presence data in the east-
ern region restricted adequate tests, yet the validation of the
eastern-summer model based on 11 SRW encounters pro-
duced an AUC of 0.70.
The spatial calculation of collision risk between ships and
SRW implies that the majority of habitat predicted as high
suitability is without risk (Fig. 4a–e). Results suggest that
SRW only face relatively substantial risk (> 12%) in a few
areas. During summer and autumn, high SRW habitat suit-
ability south of the Chatham Rise overlaps with relatively
dense ship traffic (Appendix S4). In summer, this area has
the highest risk index of vessel collision (> 16%) and high
Table 3 Model parameters and validation results of southern right whale boosted regression tree habitat models.
Region/Season
Variables
(% contribution)
Tree
complexity
Learning
rate
Number
of trees AUC CVdev
Validation AUC
Townsend (sensitivity;
1�specificity)
Validation AUC
JSV (sensitivity;
1�specificity)
Eastern/Spring dist_ridge (27.8) 3 0.005 1300 0.835 0.306 0.84 (0.56; 0.01) NA
t0_200 (24.1)
depth_mean (21.7)
chla (15.5)
mld (10.8)
Eastern/Summer t0_200 (56.3) 3 0.0025 1250 0.876 0.407 0.85 (0.65; 0.07) 0.70* (0.27; 0.13)
dist_stf (27.6)
dist_ridge (7.6)
depth_mean (5.5)
mld (3.0)
Eastern/Autumn dist_stf (37.3) 2 0.00125 1750 0.928 0.587 0.89 (0.55; 0.01) NA
t0_200 (32.5)
chla (20.7)
dist_ridge (5.5)
mld (4.0)
Western/Spring t0_200 (45.8) 3 0.0025 1500 0.796 0.226 0.66 (0.41; 0.03) 0.77 (0.44; 0.16)
t0 (15.2)
dist_stf (14.8)
depth_mean (10.4)
chla (7.2)
mld (6.7)
Western/Summer t0_200 (53.8) 2 0.005 1700 0.797 0.232 0.86 (0.65; 0.15) 0.52 (0.56; 0.55)
dist_stf (32.6)
mld (13.6)
AUC, Area under the receiver operator curve; CVdev, Cross-validation per cent deviance explained; Townsend, Townsend (1935) historical whal-
ing data; JSV, Japanese Scouting Vessel data (Miyashita et al., 1995); dist_ridge, distance from ridge; t0_200, mean temperature between surface
(0 m) and 200 m; depth_mean, mean depth; chla, chlorophyll a concentration; mld, mixed layer depth; dist_stf, distance from the Subtropical
Front; t0, temperature at surface (0 m).
*AUC value calculated with only 11 presence locations.
1144 Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd
L. G. Torres et al.

collision risk is maintained during autumn. In the western
region during spring, an area of moderate to high habitat
suitability around 38° S and 135° E overlaps a very dense
shipping lane causing relatively high-risk indices of collision.
Minimal shipping traffic crosses high suitability habitat pre-
dicted by the eastern-spring and western-summer models,
leading to scattered grid cells with low to moderate values of
risk.
With predicted temperature layers for the 2090–2100 dec-
ade incorporated into predictions of SRW distribution, habi-
tat shifted southward in all region-season combinations and
the amount of high suitability habitat (yellow and red grid
cells in Fig. 4f–j) decreased, with the exception of the wes-
tern-spring prediction. Suitable habitat along the Louisville
Ridge in spring virtually disappeared and was compressed to
a thin band around 46° S (Fig. 4f). During summer in the
eastern region, the predicted distribution of SRW shifted
southward and contracted to between 47° and 50° S
(Fig. 4g). The suitability of habitat south of Chatham Rise in
autumn declined substantially with only a few grid cells
maintaining high (red) suitability (Fig. 4h). During spring in
the western region, SRW habitat shifted south and expanded
Figure 2 Fitted-functions produced by boosted regression trees of southern right whale distribution indicating the marginal effect on
whale presence/absence (y-axes) by each habitat predictor variable. Contribution of each variable given in parentheses. Y-axes are on a
logit scale and are standardized between fitted-function plots for each region-season. Rug plots show distribution of data across that
variable, in deciles, and are used as a measure of confidence in the shape of the fitted-function. Dist_ridge = distance from ridge (m);
t0_200 = mean temperature between 0 and 200 m (°C); Depth_mean = mean depth (m); Chla = chlorophyll a concentration
(mg m�3); mld = mixed layer depth (m); Dist_stf = distance from the subtropical front (km) with sign denoting south (�) or north
(+) of the front; t0 = temperature at the surface (°C).
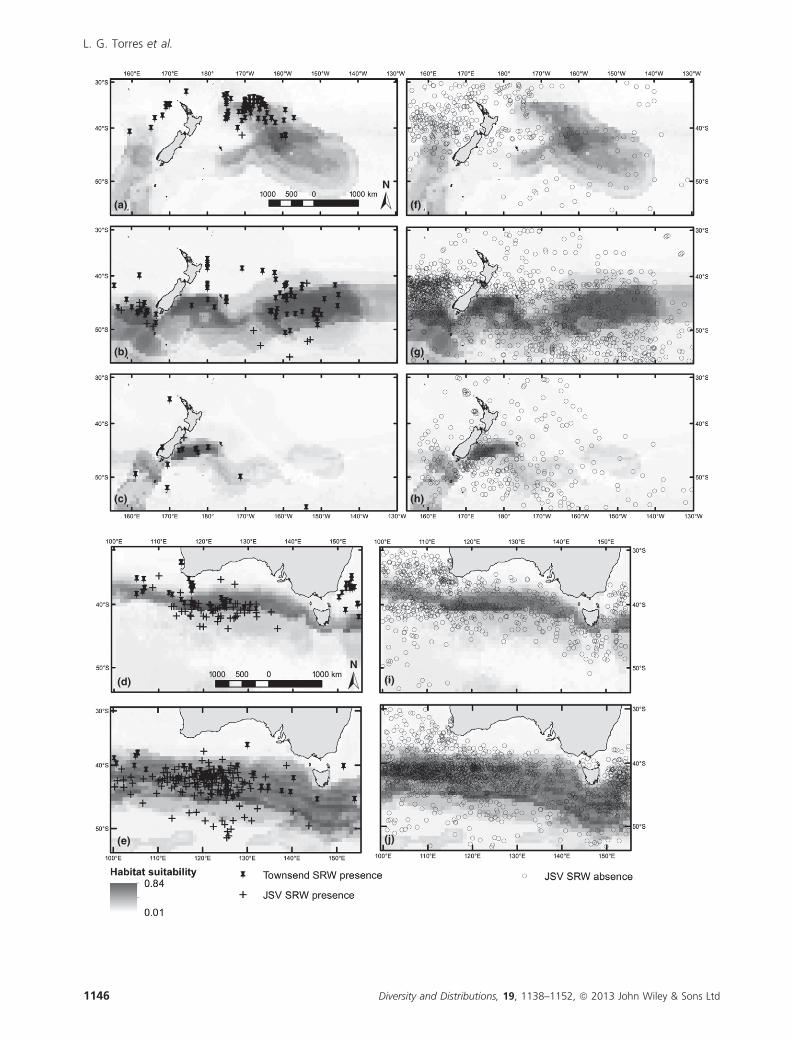
Figure 3 Predicted habitat suitability of southern right whales produced by boosted regression tree models. (a–e) Townsend (1935)
data (stars) and Japanese Scouting Vessel (crosses) presence locations used in validation tests overlaid. (f–j) Japanese Scouting Vessel
absence locations (circles) overlaid. Eastern region, spring (a,f); eastern region, summer (b,g); eastern region, autumn (c,h); western
region, spring (d,i); western region, summer (e,j). Grey scale of habitat suitability is standardized across all maps. North arrow and scale
bar in first map for each region apply to other regional plots. Note that symbols may overlap at this scale causing apparent, but false,
discrepancies with sample sizes presented in Table 1. The Louisville Ridge, Chatham Rise and Macquarie Ridge are shown in eastern
maps (a–c; f–h). Maps in Mercator projection, datum wgs1984.
Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd 1145
Habitat models based on historical data
(a)
N
(b)
(c)
(d)
(e)
(f)
(g)
(h)
(i)
(j)
N
1146 Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd
L. G. Torres et al.

150°E140°E130°E120°E110°E100°E
30°S
40°S
50°S
130°W140°W150°W160°W170°W180°170°E160°E30°S
40°S
50°S
30°S
40°S
50°S
130°W140°W150°W160°W170°W180°170°E160°E
30°S
40°S
50°S
150°E140°E130°E120°E110°E100°E
40°S
50°S
1000 0 1000 km500
1000 0 1000 km500
Risk index of vessel collision0.15% – 4%5% – 8%9% – 11%12% – 15%16% – 19%
130°W140°W150°W160°W170°W180°170°E160°E
40°S
50°S
30°S
40°S
50°S
130°W140°W150°W160°W170°W180°170°E160°E
30°S
40°S
50°S
150°E140°E130°E120°E110°E100°E30°S
40°S
50°S
150°E140°E130°E120°E110°E100°E
30°S
40°S
50°S
SRW habitat suitability
Value0.85
0.01
(a)
(b)
(c)
(f)
(g)
(h)
(d)
(e)
(i)
(j)
Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd 1147
Habitat models based on historical data

to an area between 40 and 45° S (Fig. 4i). Suitable habitat in
the western-summer prediction reduced considerably, losing
its cohesive structure, and decreasing in suitability (Fig. 4j).
DISCUSSION
Our southern right whale habitat models incorporate histori-
cal whaling data and modern environmental datasets to
describe habitat use characteristics, predict current and
future distribution patterns, and assess threats to this vulner-
able population. Validation results using independent histori-
cal whaling observations suggest that models are able to
proficiently predict historical SRW foraging distribution pat-
terns. Furthermore, the majority of recent SRW sightings
overlap predicted high suitability habitat indicating an
adequate ability to predict present-day SRW foraging habitat
at these broad spatial and temporal scales. These results
support our assumptions of (1) environmental stability
between the time periods of our datasets and (2) temporally
consistent ecological relationships between whales and their
environment.
Apparent exceptions in these affirmative validation tests
are worth further consideration. Poor model fit may explain
the low scores in the western-spring Townsend and western-
summer JSV validation tests. Many SRW encounters in the
Townsend western-spring dataset occurred outside predicted
foraging grounds and likely represent migratory whales leav-
ing calving grounds in southeast and southwest Australia
(Bannister, 2001). Our models did not describe migration
corridors causing low validation scores here. Strong valida-
tion of the western-spring prediction using JSV data suggests
that this model can adequately predict modern SRW distri-
butions with relatively high discrimination. Although JSV
presence data in the western-summer area overlaps well with
predicted high-suitability habitat, dominance of absence data
also within this high-suitability habitat may attribute to the
very low AUC (0.52). Low SRW prevalence led to high JSV
search effort with few encounters. Absence does not necessar-
ily indicate unsuitable habitat, just non-occupancy during
times of observational effort.
Mean water temperature between the surface and 200 m
(t0_200) was a consistently influential predictor of SRW dis-
tribution across all models and, as a result, dramatic changes
in SRW distribution were predicted for the 2090–2100
decade due to climate change effects on ocean temperatures.
Overall, the preferred offshore foraging habitat of SRW in
the Australasian region was characterized by t0_200 between
10 and 13 °C, mixed layer depth (mld) < 40 m, chlorophyll
concentration above 0.2 mg m�3, and being near the
Subtropical Front or close to the Louisville Ridge in spring.
These results represent the first quantitative description of
the offshore habitat of SRW.
At a regional to basin-wide scale, these broad oceano-
graphic features and patterns function to increase prey avail-
ability and density for whales by concentrating zooplankton
along vertical (i.e., thermoclines) and horizontal (i.e.,
currents) boundaries (Bakun, 1996; Gregr & Coyle, 2009).
Stomach samples of harvested SRW suggest a diet dominated
by copepods north of 50° S (Tormosov et al., 1998). We
extend this basic information by hypothesizing that SRW in
our study region prey on Neocalanus tonsus, a large-bodied
copepod common between 20 and 50° S and abundant in
the top 200 m during spring and summer (Voronina et al.,
1988). This hypothesis is based on similar habitat use pat-
terns of SRW as predicted by our models and N. tonsus as
documented in the literature.
Kawamura (1974) documented large swarms of N. tonsus
southwest of Australia in December, with maximum concen-
trations just north of the Subtropical Front and in surface
water of temperature ranging 13–14 °C. These patterns are
coincident with those derived by our western models for
SRW. In the eastern region, a longitudinal transect at
158° W during summer documented N. tonsus as the great-
est zooplankton species by mass and, at certain depths, by
quantity (up to 98%; Voronina et al., 1988). Neocalanus
tonsus was distributed from the Subtropical Front into
subtropical waters, aggregated in vast patches within the
upper mixed layer, and was depth-limited by the thermo-
cline. The top of the seasonal thermocline and the bottom
of the mixed layer are at approximately the same depth; this
link may explain the inclusion of mld in all our models.
Foraging dive depths of northern right whales are positively
correlated with depth of the bottom mixed layer, above
which prey is concentrated allowing more efficient foraging
dives (Baumgartner & Mate, 2003). Similarly, SRW may
select habitats with relatively shallow mld to promote effi-
cient foraging by spending less time transiting and more
time feeding. While temperature, mixed layer depth and
distance from the Subtropical Front do not determine the
fine-scale dynamics of zooplankton distribution (Bakun,
1996), the seasonal foraging distributions of SRW at scales
of 10s to 100s of km are likely to respond to these broad-
scale oceanic features that concentrate prey (Gregr & Coyle,
2009). Our hypothesis of SRW feeding on N. tonsus could
Figure 4 Predicted impacts from anthropogenic threats on southern right whale (SRW) habitat, by region and season. (a–e) Predictedrisk index of collision between ships and southern right whales based on spatial calculation (see ‘Methods’) between predicted whale
habitat suitability and ship traffic density layer (Appendix S4; Halpern et al., 2008). (f–j) Predicted SRW habitat suitability in the 2090–2100 decade based on changes in ocean temperatures due to global climate change. Colour scales of habitat suitability and collision risk
are standardized across all prediction maps. North arrow and scale bar in first map for each region apply to other regional plots.
Eastern region, spring (a,f); eastern region, summer (b,g); eastern region, autumn (c,h); western region, spring (d,i); western region,
summer (e,j). Maps in Mercator projection, datum wgs1984.
1148 Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd
L. G. Torres et al.

be tested by sampling near feeding SRW or by stable isotope
comparisons.
The calculation of relative collision risk between shipping
traffic and SRW based on present-day predicted distribution
patterns revealed generally lower risk across the majority of
high- suitability habitat and two relatively discrete areas of
higher risk. These areas are the southern slope of the Chatham
Rise in summer and autumn, and near 38° S and 135° E in
the western region during spring. Although the methods we
have used in our study are not directly comparable with those
used in the North Atlantic to identify areas of higher risk of
ship strikes for northern right whales (Ward-Geiger et al.,
2005), it appears that the threat of ship-strike to SRW in the
Australasian region is less than that for North Atlantic right
whales (Knowlton & Brown, 2007). However, SRW leaving
coastal wintering grounds along the south Australian coast are
likely to encounter heavy vessel traffic when they cross dense
shipping lanes to access foraging grounds. Results from our
analysis of vessel collision risk can guide research into SRW
ship-strikes and, if needed, inform mitigation measures.
Predicted effects of global climate change include increased
ocean temperatures (Smith et al., 1999; Levitus et al., 2000)
and changes to current structures (Mitchell & Hulme, 1999).
Zooplankton distributions have been shown to be sensitive
to ocean temperatures (Rutherford et al., 1999; Beaugrand
et al., 2002) and changes in location and strength of ocean
boundaries that concentrate zooplankton (Bakun, 1996).
Therefore, climate change could affect the distribution or
foraging efficiency of zooplankton-feeding organisms such as
right whales. Our integration of predicted ocean tempera-
tures for the 2090–2100 decade and the functional relation-
ships modelled between SRW and ocean temperature,
predicts substantial changes in the availability and location
of suitable SRW habitat. Areas of high habitat suitability
shifted south in all region–season combinations, including a
shift of more than 1000 km in the eastern region during
spring. Although our results suggest decreased availability of
suitable habitat for SRW, actual changes are likely to be
more complex because (1) we have not accounted for
changes in the distribution or strength of the mixed layer
depth, primary productivity or the Subtropical Front, and
(2) the unknown ecological responses of copepod prey and
their predators. Yet, if oceans continue to warm due to
global climate change as predicted, our work indicates that
SRW will face dramatic temperature changes in areas with
previously high habitat suitability. Although it is unknown if
the narrow temperature range in the upper 200 m preferred
by SRW is due to physiological limits of whales or their prey,
or both, this temperature sensitivity makes SRW particularly
vulnerable to the impacts of climate change, which may be
exacerbated due to matrilineal fidelity to feeding grounds
(Valenzuela et al., 2009).
Despite minimal recent offshore sightings of SRW, our
study described the locations and environmental characteris-
tics of SRW foraging grounds for the first time. In addition,
based on coincident distribution and habitat use patterns of
N. tonsus and SRW, we are the first to hypothesize the
main copepod prey species of SRW in the region. Without
the novel use of a historical dataset, the offshore habitat
use patterns of this endangered species in this area would
remain unknown. Furthermore, our assessment of threats to
this SRW population identified discrete locations of
increased risk of vessel collision and predicted large-scale
areas of habitat loss due to climate change. Our results can
aid conservation management of SRW by directing popula-
tion monitoring efforts, generating effective regulation strat-
egies, identifying critical habitats threatened by other
activities and guiding future research efforts. These pop-
ulations of SRW are currently increasing (Bannister, 2011;
Carroll et al., 2011b) and may recolonize former calving
and foraging grounds. Therefore, best management policies
will not only protect SRW from current threats but also
look ahead to protect these vulnerable populations from
future threats by minimizing degradation and exploitation
of critical habitats.
ACKNOWLEDGEMENTS
This project was jointly funded by the Australian Marine
Mammal Centre and the National Institute of Water and
Atmospheric Research, Ltd. (NIWA). We thank the
following people from NIWA for their advice, time
and insight: S. Bardsley, T. Compton, S. Moremede, S.
Mikaloff-Fletcher, J. Grieve, H. Neil. We also thank R.
Reeves, J. Lund and E. Josephson for help with historical
data collection, and three anonymous reviewers for help-
ful comments on this manuscript.
REFERENCES
Austin, M. (2007) Species distribution models and ecological
theory: a critical assessment and some possible new
approaches. Ecological Modelling, 200, 1–19.
Bakun, A. (1996) Patterns in the ocean: ocean processes and
marine population dynamics. California Sea Grant College
System, La Jolla, CA.
Bannister, J. (2001) Status of southern right whales (Eubalaena
australis) off Australia. Journal of Cetacean Research and
Management, 2, 103–110.
Bannister, J. (2011) Population trend in right whales off
southern Australia 1993–2010. Southern right whale
assessment workshop. International Whaling Commission,
Buenos Aires, Argentina.
Bannister, J.L., Pastene, L.A. & Burnell, S.R. (1999) First
record of movement of a southern right whale (Eubalaena
australis) between warm water breeding grounds and the
Antarctic Ocean, south of 60S. Marine Mammal Science,
15, 1337–1342.
Barbet-Massin, M., Jiguet, F., Albert, C.H. & Thuiller, W.
(2012) Selecting pseudo-absences for species distribution
models: how, where and how many? Methods in Ecology
and Evolution, 3, 327–338.
Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd 1149
Habitat models based on historical data

Baumgartner, M.F. & Mate, B.R. (2003) Summertime forag-
ing ecology of North Atlantic right whales. Marine Ecology-
Progress Series, 264, 123–135.
Beaugrand, G., Reid, P.C., Iba~nez, F., Lindley, J.A. &
Edwards, M. (2002) Reorganization of North Atlantic
marine copepod biodiversity and climate. Science, 296,
1692–1694.
Buston, P.M. & Elith, J. (2011) Determinants of reproductive
success in dominant pairs of clownfish: a boosted regres-
sion tree analysis. Journal of Animal Ecology, 80, 528–538.
Carroll, E., Jackson, J.A., Paton, D. & Smith, T.D. (2011a)
Estimating nineteenth and twentieth century right whale
catches and removals around east Australia and New Zea-
land. Report to the Ministry of Fisheries, p. 13. Wellington.
Carroll, E.L., Patenaude, N.J., Childerhouse, S.J., Kraus, S.D.,
Fewster, R.M. & Baker, C.S. (2011b) Abundance of the
New Zealand subantarctic southern right whale population
estimated from photo-identification and genotype mark-
recapture. Marine Biology, 158, 2565–2575.
Chiswell, S.M. (1994) Variability in sea surface temperature
around New Zealand from AVHRR images. New Zealand
Journal of Marine and Freshwater Research, 28, 179–192.
Delworth, T.L., Broccoli, A.J., Rosati, A. et al. (2006) GFDL’s
CM2 global coupled climate models. Part I: formulation
and simulation characteristics. Journal of Climate, 19, 643–
674.
Dunn, J.R. & Ridgeway, K.R. (2002) Mapping ocean proper-
ties in regions of complex topography. Deep Sea Research I:
Oceanographic Research, 49, 591–604.
Elith, J. & Leathwick, J. (2007) Predicting species distribu-
tions from museum and herbarium records using multire-
sponse models fitted with multivariate adaptive regression
splines. Diversity and Distributions, 13, 265–275.
Elith, J., Graham, C.H., Anderson, R.P. et al. (2006) Novel
methods improve prediction of species’ distributions from
occurrence data. Ecography, 29, 129–151.
Elith, J., Leathwick, J.R. & Hastie, T. (2008) A working guide
to boosted regression trees. Journal of Animal Ecology, 77,
802–813.
Estes, J.A., Terborgh, J., Brashares, J.S. et al. (2011) Trophic
downgrading of planet earth. Science, 333, 301–306.
Fawcett, T. (2006) An introduction to ROC analysis. Pattern
Recognition Letters, 27, 861–874.
Gnanadesikan, A., Dixon, K.W., Griffies, S.M. et al. (2006)
GFDL’s CM2 global coupled climate models. Part II: the
baseline ocean simulation. Journal of Climate, 19, 675–697.
Greene, C.H. & Pershing, A.J. (2004) Climate and the con-
servation biology of North Atlantic right whales: the right
whale at the wrong time? Frontiers in Ecology and the Envi-
ronment, 2, 29–34.
Gregr, E.J. (2011) Insights into North Pacific right whale
Eubalaena japonica habitat from historic whaling records.
Endangered Species Research, 15, 223–239.
Gregr, E.J. & Coyle, K.O. (2009) The biogeography of the
North Pacific right whale (Eubalaena japonica). Progress in
Oceanography, 80, 188–198.
Gregr, E.J. & Trites, A.W. (2001) Predictions of critical habi-
tat for five whale species in the waters of coastal British
Columbia. Canadian Journal of Fish and Aquatic Science,
58, 1265–1285.
Guisan, A. & Zimmermann, N.E. (2000) Predictive habitat
distribution models in ecology. Ecological Modelling, 135,
147–186.
Halpern, B.S., Walbridge, S., Selkoe, K.A., Kappel, C.V., Mic-
heli, F., D’Agrosa, C., Bruno, J.F., Casey, K.S., Ebert, C.,
Fox, H.E., Fujita, R., Heinemann, D., Lenihan, H.S., Ma-
din, E.M.P., Perry, M.T., Selig, E.R., Spalding, M., Steneck,
R. & Watson, R. (2008) A global map of human impact
on marine ecosystems. Science, 319, 948–952.
Heath, R.A. (1985) A review of the physical oceanography of
the seas around New Zealand – 1982. New Zealand Journal
of Marine and Freshwater Research, 19, 79–124.
Hendy, E.J., Gagan, M.K., Alibert, C.A., McCulloch, M.T.,
Lough, J.M. & Isdale, P.J. (2002) Abrupt decrease in tropi-
cal Pacific sea surface salinity at end of little ice age.
Science, 295, 1511–1514.
Hirzel, A.H., Le Lay, G., Helfer, V., Randin, C. & Guisan, A.
(2006) Evaluating the ability of habitat suitability models
to predict species presences. Ecological Modelling, 199, 142–
152.
Jackson, J.A., Patenaude, N.J., Carroll, E.L. & Baker, C.S.
(2008) How few whales were there after whaling? Inference
from contemporary mtDNA diversity. Molecular Ecology,
17, 236–251.
Jaquet, N., Whitehead, H. & Lewis, M. (1996) Coherence
between 19th century sperm whale distributions and satel-
lite-derived pigments in the tropical Pacific. Marine Ecology
Progress Series, 145, 1–10.
Kawamura, A. (1974) Food and feeding ecology in the south-
ern sei whale. Scientific report of whales research institute.
pp. 25–144. Tokyo.
Kemper, C., Coughran, D., Warneke, R., Pirzl, R., Watson,
M., Gales, R. & Gibbs, S. (2008) Southern right whale
(Eubalaena australis) mortalities and human interactions in
Australia, 1950–2006. Journal of Cetacean Research and
Management, 10, 1–8.
Knowlton, A.R. & Brown, M.W. (2007) Running the gauntlet:
right whales and vessel strikes. Harvard University Press,
Boston, MA.
Leaper, R., Cooke, J., Trathan, P.N., Reid, K., Rowntree, V.J.
& Payne, R.S. (2006) Global climate change drives south-
ern right whale (Eubalaena australis) population dynamics.
Biology Letters, 2, 289–292.
Leathwick, J.R., Elith, J., Francis, M.P., Hastie, T. & Taylor,
P. (2006) Variation in demersal fish species richness in the
oceans surrounding New Zealand: an analysis using
boosted regression trees. Marine Ecology-Progress Series,
321, 267–281.
Levitus, S., Antonov, J.I., Boyer, T.P. & Stephens, C. (2000)
Warming of the world ocean. Science, 287, 2225–2229.
Lobo, J.M., Jim�enez-Valverde, A. & Real, R. (2008) AUC:
a misleading measure of the performance of predictive
1150 Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd
L. G. Torres et al.

distribution models. Global Ecology and Biogeography, 17,
145–151.
Maury, M.F. (1852) Whales chart of the world. Series F
(wind and current charts) sheet 1. (1852), U.S. Naval Obser-
vatory, Washington, DC.
Mitchell, T.D. & Hulme, M. (1999) Predicting regional cli-
mate change: living with uncertainty. Progress in Physical
Geography, 23, 57–78.
Miyashita, T., Kato, H. & Kasuya, T. (1995) Worldwide map
of cetacean distribution based on Japanese sighting data.
National Research Institute of Far Seas Fisheries, Shizuoka,
Japan.
Nott, J., Haig, J., Neil, H. & Gillieson, D. (2007) Greater
frequency variability of landfalling tropical cyclones at
centennial compared to seasonal and decadal scales. Earth
and Planetary Science Letters, 255, 367–372.
Ohsumi, S. & Kasamatsu, F. (1986) Recent off-shore distri-
bution of the southern right whale in summer. Report to
the International Whaling Commission (Special Issue 10).
pp. 177–184.
Patenaude, N. & Baker, C.S. (2001) Population status and
habitat use of southern right whales in the sub-Antarctic
Auckland Islands of New Zealand. Journal of Cetacean
Research and Management, 2, 111–116.
Peterson, A.T., Sober�on, J. & S�anchez-Cordero, V. (1999)
Conservatism of ecological niches in evolutionary time.
Science, 285, 1265–1267.
Richards, R. (2002) Southern right whales: a reassessment of
their former distribution and migration routes in New
Zealand waters, including on the Kermadec grounds. Jour-
nal of the Royal Society of New Zealand, 32, 355–377.
Ridgeway, G. (2007) Generalized boosted regression models.
V1.6-3.1. March 2011.
Rutherford, S., D’Hondt, S. & Prell, W. (1999) Environmen-
tal controls on the geographic distribution of zooplankton
diversity. Nature, 400, 749–753.
Smith, T.D. (2001) Examining cetacean ecology using histor-
ical fishery data. The exploited seas: new direction for marine
environmental history (ed. by P. Holm, T.D. Smith and D.J.
Starkey), pp. 207–214. Memorial University of Newfound-
land, Newfoundland.
Smith, R.C., Ainley, D., Baker, K., Domack, E., Emslie, S.,
Fraser, B., Kennett, J., Leventer, A., Mosley-Thompson, E.,
Stammerjohn, S. & Vernet, M. (1999) Marine ecosystem
sensitivity to climate change. BioScience, 49, 393–404.
Smith, T.D., Reeves, R.R., Josephson, E.A. & Lund, J.N.
(2012) Spatial and seasonal distribution of American whal-
ing and whales in the age of sail. PLoS ONE, 7, e34905.
Stramma, L., Peterson, R.G. & Tomczak, M. (1995) The
South Pacific current. Journal of Physical Oceanography, 25,
77–91.
Swets, J.A. (1988) Measuring the accuracy of diagnostic
systems. Science, 240, 1285–1293.
Thrush, S.F. & Dayton, P.K. (2002) Disturbance to marine
benthic habitats by trawling and dredging: implications for
marine biodiversity. Annual Review of Ecology and
Systematics, 33, 449–473.
Tormosov, D.D., Mikhaliev, Y.A., Best, P.B., Zemsky, V.A.,
Sekiguchi, K. & Brownell, R.L. (1998) Soviet catches of
southern right whales Eubalaena australis, 1951–1971.
Biological data and conservation implications. Biological
Conservation, 86, 185–197.
Townsend, C.H. (1935) The distribution of certain whales as
shown by logbook records of American whaleships. Zoolog-
ica (NY), 19, 1–50.
Trathan, P.A., Forcada, J. & Murphy, E.J. (2007) Environ-
mental forcing and southern ocean marine predator popu-
lations: effects of climate change and variability.
Philosophical Transactions of the Royal Society B: Biological
Sciences, 362, 2351–2365.
Valenzuela, L.O., Sironi, M., Rowntree, V.J. & Seger, J.
(2009) Isotopic and genetic evidence for culturally inher-
ited site fidelity to feeding grounds in southern right
whales (Eubalaena australis). Molecular Ecology, 18, 782–
791.
Voronina, N.M., Kolosova, Y.G. & Flint, M.V. (1988) Distri-
bution and biomass of abundant mesoplankton species.
Pacific subantarctic ecosystems (ed. by M.E. Vinogradov and
M.V. Flint), pp. 197–210. Nauka Press, Moscow.
Ward-Geiger, L.I., Silber, G.K., Baumstark, R.D. & Pulfer,
T.L. (2005) Characterization of ship traffic in right whale
critical habitat. Coastal Management, 33, 263–278.
Worm, B., Barbier, E.B., Beaumont, N., Duffy, J.E., Folke,
C., Halpern, B.S., Jackson, J.B.C., Lotze, H.K., Micheli, F.,
Palumbi, S.R., Sala, E., Selkoe, K.A., Stachowicz, J.J. &
Watson, R. (2006) Impacts of biodiversity loss on ocean
ecosystem services. Science, 314, 787–790.
Yesson, C., Clark, M.R., Taylor, M.L. & Rogers, A.D. (2011)
The global distribution of seamounts based on 30 arc
seconds bathymetry data. Deep Sea Research Part I: Oceano-
graphic Research Papers, 58, 442–453.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the
online version of this article:
Appendix S1 Temporal distribution of whaling effort.
Appendix S2 Methods, definitions and illustration of the
geomorphic map.
Appendix S3 Description of recent southern right whale
sightings in the region.
Appendix S4 Details on the shipping layer and implementa-
tion of the CM2.1 model.
Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd 1151
Habitat models based on historical data

BIOSKETCH
Leigh G. Torres is a marine spatial ecologist with research
focused on the distribution and habitat use patterns of
marine mammals and seabirds in relation to environmental
variability and anthropogenic impacts. Leigh is particularly
interested in how behaviour patterns vary with habitat, how
spatial scale impacts predator-prey interactions, what envi-
ronmental cues species use to find prey, how species spatially
and behaviourally reduce inter- and intra-specific competi-
tion, and how foraging strategies change relative to prey
availability and spatio-temporal variability.
Author contributions: L.G.T. and A.M. conceived the idea;
T.D.S. contributed historical whaling data and assisted with
manuscript preparation; P.S. provided dynamic environmen-
tal data; T.M. contributed Japanese Scouting Vessel (JSV)
data; L.G.T. performed analyses and led the writing; all
authors discussed the results and commented on earlier
versions of the manuscript.
Editor: Janet Franklin
1152 Diversity and Distributions, 19, 1138–1152, ª 2013 John Wiley & Sons Ltd
L. G. Torres et al.
Copyright © 2022 FDOKUMEN




















