
a set design for bertolt brecht's - the caucasian chalk circle
Upload
independentCategory
view
3download
0
This article was downloaded by:[Füst, George][Füst, George]
On: 11 July 2007Access Details: [subscription number 780552888]Publisher: Informa HealthcareInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Immunological InvestigationsA Journal of Molecular and Cellular ImmunologyPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713597256
Frequency of Carriers of 8.1 Ancestral Haplotype and itsFragments in Two Caucasian PopulationsPetra Kiszel a; Margit Kovács a; Csaba Szalai b; Yan Yang c; Éva Pozsonyi d;Bernadett Blaskó a; Judit Laki a; Zoltán Prohászka ae; Ádám Fazakas a; PálPánczél a; Nóra Hosszúfalusi a; Katalin Rajczy d; Yee-Ling Wu c; Erwin K. Chungc; Bi Zhou c; Carol A. Blanchong c; Ágnes Vatay a; C. Yung Yu ca Third Department of Internal Medicine, Semmelweis University. Budapest.Hungaryb Central Laboratory of Human Genetics, Semmelweis University. Budapest.Hungaryc Center for Molecular and Human Genetics, Columbus Children's ResearchInstitute and College of Medicine and Public Health, The Ohio State University.
Columbus, Ohio. USAd Department of Immunogenetics, National Medical Center. Budapest. Hungarye Research Group of Metabolism and Atherosclerosis, Semmelweis University and Hungarian Academy of Sciences.Budapest. Hungary
Online Publication Date: 01 May 2007To cite this Article: Kiszel, Petra, Kovács, Margit, Szalai, Csaba, Yang, Yan, Pozsonyi, Éva, Blaskó, Bernadett, Laki,Judit, Prohászka, Zoltán, Fazakas, Ádám, Pánczél, Pál, Hosszúfalusi, Nóra, Rajczy, Katalin, Wu, Yee-Ling, Chung,Erwin K., Zhou, Bi, Blanchong, Carol A., Vatay, Ágnes, Yu, C. Yung and Füst, G. , (2007) 'Frequency of Carriers of 8.1Ancestral Haplotype and its Fragments in Two Caucasian Populations', Immunological Investigations, 36:3, 307 - 319To link to this article: DOI: 10.1080/08820130701241404URL: http://dx.doi.org/10.1080/08820130701241404
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.
© Taylor and Francis 2007

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
Immunological Investigations, 36:307–319, 2007Copyright © Informa HealthcareISSN: 0882-0139 print / 1532-4311 onlineDOI: 10.1080/08820130701241404
LIMM0882-01391532-4311Immunological Investigations, Vol. 36, No. 3, Mar 2007: pp. 0–0Immunological InvestigationsFrequency of Carriers of 8.1 Ancestral Haplotype and its Fragments in Two Caucasian PopulationsHSP70-2G and 8.1 Ancestral HaplotypeP. Kiszel et al.
Petra Kiszel,1 Margit Kovács,1 Csaba Szalai,3 Yan Yang,4 Éva Pozsonyi,5 Bernadett Blaskó,1 Judit Laki,1 Zoltán Prohászka,1,2 Ádám Fazakas,1 Pál Pánczél,1 Nóra Hosszúfalusi,1 Katalin Rajczy,5 Yee-Ling Wu,4 Erwin K. Chung,4 Bi Zhou,4 Carol A. Blanchong,4 Ágnes Vatay,1 C. Yung Yu,4 and G. Füst1,2
1Third Department of Internal Medicine, Semmelweis University, Budapest, Hungary2Research Group of Metabolism and Atherosclerosis, Semmelweis University andHungarian Academy of Sciences, Budapest, Hungary3Central Laboratory of Human Genetics, Semmelweis University, Budapest, Hungary4Center for Molecular and Human Genetics, Columbus Children’s Research Instituteand College of Medicine and Public Health, The Ohio State University, Columbus,Ohio, USA5Department of Immunogenetics, National Medical Center, Budapest, Hungary
Within the human MHC region larger stretches of conserved DNA, called conservedancestral haplotypes exist. However, many MHC haplotypes contain only fragments ofan ancestral haplotype. Little is known, however, on relative distribution of the ances-tral haplotypes to their fragments. Therefore we determined the frequency of carriersof the whole ancestral haplotype 8.1 (AH8.1) and its fragments in 127 healthy Hungarianpeople, 101 healthy Ohioian females, and in nine Hungarian families. The HLA-DQ2,HLA-DR3(17), RAGE -429C allele, the mono-S-C4B genotype, the HSP70-2 1267Gallele and the TNF -308A (TNF2) allele were used as markers of the AH8.1. Frequencyof carriers of the whole AH8.1 and its fragments was similar in the both populations.18% of the subjects carried the whole AH8.1 in at least one chromosome, while 17–20%,36–39%, and 24–29%, respectively carried two or three constituents of the haplotype,only one constituent or none of them. Similar results were obtained in the familystudy. In addition, marked differences were found in the relationship of the constitu-ents’ alleles to the whole AH8.1. In both populations, 29%, 50–59%, 52–56% and 76–96%,respectively of the carriers of HSP70-2 1267G, RAGE-429C, TNF2, and mono-S
Address correspondence to Dr. G. Füst, 3rd Department of Medicine, SemmelweisUniversity, Kútvölgyiút 4, Budapest H-1125, Hungary; E-mail: [email protected]

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
308 P. Kiszel et al.
carriers carried the whole 8.1 haplotype. These findings may have important implica-tions for studies of the disease associations with different MHC ancestral haplotypes.
Keywords hsp70, Heat shock proteins, MHC, HLA, Linkage Disequilibrium.
INTRODUCTION
Blocks of conserved DNA sequences maintained as genetically fixed haplo-types are present throughout the human genome (Gabriel et al., 2002;Stumpf, 2002). Within the human MHC region – located in chromosome 6 (6p21.3) larger stretches of conserved DNA, called conserved extended haplotypesor ancestral haplotypes exist (Yunis et al., 2003). The haplotypes are built upfrom genotypes of four major blocks (HLA Cw-B, TNF cluster, complementgene cluster and HLA-DQ-DR), but specific alleles located within regionsintervening within these blocks also could be the constituents of these ances-tral haplotypes (Yunis et al., 2003).
Many well-characterized ancestral haplotypes were defined within theMHC region. One of them whose frequency is the highest in the Caucasianpopulation is the 8.1. ancestral haplotype (AH8.1). Within the 4 major blocksmost characteristic constituents of the AH8.1 are HLA B8, TNF2 (TNF-308Aallele), mono-S-RCCX and HLA-DQ2-DR3(17) genotypes.
RCCX modules within the complement regions are formed by polygenicand modular variations of complement components C4A and C4B genestogether with their neighboring genes. These modules may be present in one(monomodular, two (bimodular), or three (trimodular) copies in each chromo-some (Blanchong et al., 2000; Chung et al., 2002; Yu et al., 2003) . The C4genes could be short (S) or long (L). Mono-S designate a monomodular RCCXcontaining a short C4B gene.
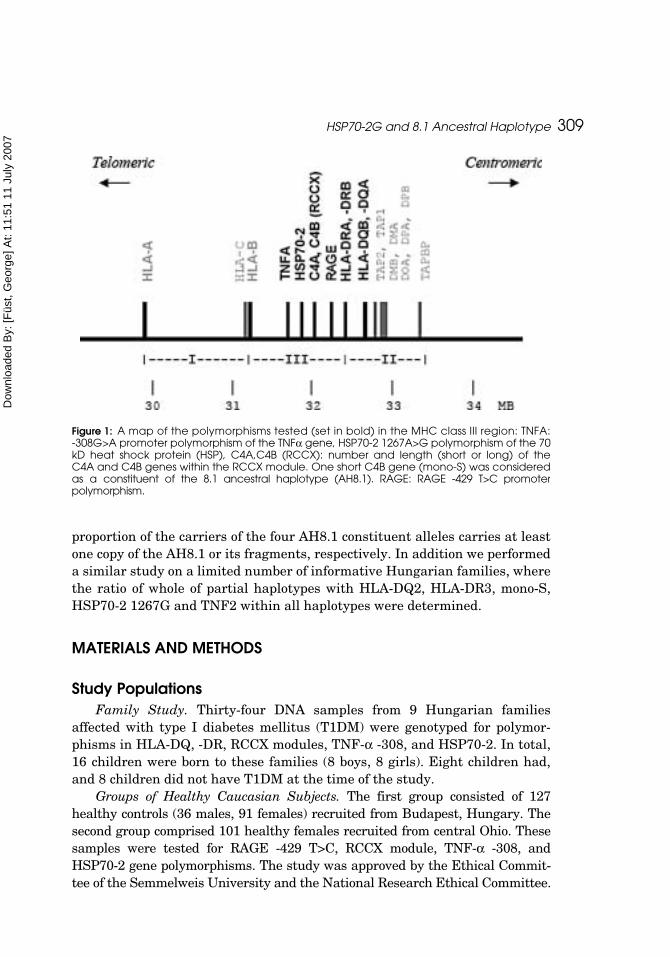
In addition to the genotypes listed here, some alleles in the interveningregions such as the RAGE -429C allele between the MHCII region and the com-plement cluster (Laki et al., 2006; Price et al., 1999) or the HSP70-2 1267Gallele located between the complement and TNF clusters (Caplen et al., 1990;Corzo et al., 1995; Price et al., 1999) (Figure 1) were found to be strongly linkedto the AH8.1. Therefore, they can be considered as constituents of AH8.1.
There are abundant data on the frequency and disease association of theAH8.1. However, many MHC haplotypes are not extended ancestral haplotypesbut contain only fragments or only a single small block of the ancestral haplo-type. It is also the case with the AH8.1. Little is known, however, on relativedistribution of the ancestral haplotypes to their fragments (Yunis et al., 2003).
Earlier we reported on a strong association of the mono-S with the TNF2allele of tumor necrosis factor-α (i.e., the 308A allele) (Vatay et al., 2003). Inthe meantime we also determined the RAGE -429T>C, and the HSP70-2 1267A>G SNPs in the same samples. The data allowed the determination of which
Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
HSP70-2G and 8.1 Ancestral Haplotype 309
proportion of the carriers of the four AH8.1 constituent alleles carries at leastone copy of the AH8.1 or its fragments, respectively. In addition we performeda similar study on a limited number of informative Hungarian families, wherethe ratio of whole of partial haplotypes with HLA-DQ2, HLA-DR3, mono-S,HSP70-2 1267G and TNF2 within all haplotypes were determined.
MATERIALS AND METHODS
Study PopulationsFamily Study. Thirty-four DNA samples from 9 Hungarian families
affected with type I diabetes mellitus (T1DM) were genotyped for polymor-phisms in HLA-DQ, -DR, RCCX modules, TNF-α -308, and HSP70-2. In total,16 children were born to these families (8 boys, 8 girls). Eight children had,and 8 children did not have T1DM at the time of the study.
Groups of Healthy Caucasian Subjects. The first group consisted of 127healthy controls (36 males, 91 females) recruited from Budapest, Hungary. Thesecond group comprised 101 healthy females recruited from central Ohio. Thesesamples were tested for RAGE -429 T>C, RCCX module, TNF-α -308, andHSP70-2 gene polymorphisms. The study was approved by the Ethical Commit-tee of the Semmelweis University and the National Research Ethical Committee.
Figure 1: A map of the polymorphisms tested (set in bold) in the MHC class III region: TNFA:-308G>A promoter polymorphism of the TNFα gene, HSP70-2 1267A>G polymorphism of the 70kD heat shock protein (HSP), C4A,C4B (RCCX): number and length (short or long) of theC4A and C4B genes within the RCCX module. One short C4B gene (mono-S) was consideredas a constituent of the 8.1 ancestral haplotype (AH8.1). RAGE: RAGE -429 T>C promoterpolymorphism.

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
310 P. Kiszel et al.
Isolation of the Genomic DNA. Genomic DNA was isolated from theblood samples of the Ohio group using the Puregene DNA isolation kit (GentraSystems, Inc., Minneapolis, MN, USA). In the case of the Hungarian healthygroup and families total genomic DNA was extracted from white blood cellsusing the method of Miller et al. (1988). There was no difference in the qualityof the DNA preparations extracted by the two methods.
HLA Typing. HLA-DQ and HLA-DR alleles were detected by GenovisionDR and DQ kits using PCR-SSP method.
Methods Used for Studying the RCCX Modules. Five micrograms of genomicDNA was digested with appropriate restriction enzymes in each reaction.Restriction enzymes included TaqI, PshAI (New England Biolabs) and PvuII(Invitrogen). PshAI was used for overnight digestion at 25°C, followed by anadditional 1 hour of 37°C incubation with PvuII. TaqI digest was performedovernight at 65°C. DNA fragments were resolved by electrophoresis in a 0.8%agarose gels, Southern blotted to a nylon membrane (Hybond-N+ membrane,Amersham Life Science), hybridized with appropriate [α32P] dCTP-labeledprobes for RP and C4, CYP21, and TNX, and analyzed by autoradiography(Blanchong et al., 2000).
Determination of the –308 G>A Polymorphism for TNFAThe TNFA –308 G>A polymorphism was determined by polymerase chain
reaction-sequence-specific primer (PCR-SSP) method using the primers sug-gested by Day et al. (1998). The PCR products were digested at 37°C withNcoI restriction enzyme and resolved by agarose gel electrophoresis.
Determination of the 1267 A>G Polymorphism for HSP70-2HSP70-2 PstI restriction fragment length polymorphism at position
1267 was characterized by a PCR-RFLP strategy using the following sense5’-CATCGACTTCTACACGTCCA-3’ and antisense 5’-CAAAGTCCTTGAGTC-CCAAC-3’ primers. First, a denaturation step was conducted for 3 min at95°C followed by 35 cycles of a 20-sec denaturation at 95°C, a 20-secondannealing at 60°C and a 60-second extension at 72°C. A final extension wasperformed for 6 minutes at 72°C before PstI digestion of 15 μl of the amplifiedDNA product. The DNA-lacking polymorphic PstI site within the HSP70-2gene generates a product of 1117 bp after restriction (A- allele), whereas thePstI site produced two fragments of 936 and 181 bp (G-allele). (Schroeder etal., 1999; Vargas-Alarcon et al., 2002).
Primer Design and PCR Conditions for RAGEGenomic DNA was extracted according to the method of Miller and co-
workers (Miller et al., 1988). Forward and reverse primers (forward: 5’ GGG

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
HSP70-2G and 8.1 Ancestral Haplotype 311
GGC AGT TCT CTC CTC-3’ and reverse: 5’ –TCA GAG CCC CCG ATC CTATTT-3’) were obtained from literature (Hudson et al., 2001).
Optimalized PCR consisted of a 50-μl reaction volume containing 50 pmolof forward and reverse, 100–500 ng of template DNA, 200 μmol/l of each deox-ynucleotide phosphate, 2 mmol/l MgCl2, 1 U Taq DNA polymerase (Promega)and a reaction buffer containing 20 mmol/l Tris-HCl, 50 mmol/l KCl(Promega). Amplification consisted of 35 cycles of 94°C for 1 minute, a primer-annealing step of 56°C for 1 minute, and primer extension at 72°C for 2 min-utes, followed by a final extension for 10 minutes at 72°C.
Restriction Fragment Length Polymorphism for RAGEFor polymorphism -429 PCR products were digested with AluI, (2 μl of
digestion mix containing 0.2 μl AluI and 2 μl Gibco-BRL buffer no. 1 per sam-ples were added to 10μl PCR product; incubation lasted for 6 hours at 37°C.Digestion products were run on 3% agarose gel.
Statistical AnalysisStatistical analysis was performed by Statistica 6.0 software <www.
statsoft.com> and Arlequin 2.000 software <http://anthro.unige.ch/arlequin>,[email protected]) was used for calculation of linkage disequilibrium.Frequencies were compared by the chi-square test. p values less than 0.05were considered significant.
RESULTS
Haplotypes of Marker Alleles of the AH8.1 in Nine Type 1 Diabetes Mellitus FamiliesNine Hungarian families affected with type 1 diabetes mellitus (T1DM)
informative for the inheritance of the 1267 A>G SNP of the HSP70-2 gene aswell as HLA class II gene haplotypes for DQ and DR, the presence andabsence of monomodular RCCX structure with short C4B gene, together withthe 308 G>A SNP of the TNFA gene in the MHC class III region were tested.Results of haplotype segregation analyses are shown in Table 1. Of the 36paternal and maternal Chromosome 6 from the 9 families, 18 different MHChaplotypes with DQ and DR alleles, the presence or absence of monomodularshort RCCX, the 1267 A>G SNP of HSP70-2, and the 308 G>A SNP of TNFAwere found (Table 1).
The most common haplotype is DQ2 - DR17 - S - 1267G - TNF2, where Sstands for the monomodular RCCX with a short C4B gene, 1267G for the1267G allele in HSP70-2, and TNF2 for the 308A allele of the TNFA gene. All

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
312 P. Kiszel et al.
constituents of this haplotype are markers of the AH8.1. It appears 7 timesamong the parents (Table 1). In addition, 6 variant or recombinant haplotypesrelated to AH8.1 that is containing 2–4 constituents of the AH8.1 are observedin 7 parents. It is of interest to note that all AH8.1 haplotypes and 2 of the 3variants were transmitted to the diabetic children. Five other haplotypespresent in 7 parents contained only one AH8.1 markers while none of themwas present in 6 haplotypes of 15 parents (Table 1).
It is noteworthy that DQ5-DR1-L-TNF1 may be linked to 1267G or 1267Aof HSP70-2. The haplotype with 1267G was present in 3 parents but nottransmitted to their diabetic children. The haplotype linked to 1267A waspresent in 4 parents and was transmitted to the diabetic children twice.
All but one of the diabetic children but only 3/8 of the healthy siblingsinherited the HLA-DQ2—DR17,mono-S,HSP70-2G,TNF2 haplotype.
Allelic Frequencies of Four AH8.1 Markers Alleles in Two Distinct Caucasian PopulationsThe RAGE -429 T>C, HSP70-2 1267A>G SNP, and the TNFα -308 G>A
SNPs were determined in 127 healthy Hungarian subjects, and 101 healthy
Table 1: Human MHC class II and class III haplotypes with different constituents (set in bold) of the 8.1 ancestral haplotype (AH8.1) in nine Hungarian families with type 1 diabetic children.
A. Caucasian MHC ancestral haplotype 8.1 Number (frequency)
1. DQ2 - DR17 - S - 1267G - TNF2 7 (0.19)B. Recombinants or variants of AH8.1 with 2 to 4
constituents2. DQ2 - DR17 – L - 1267G - TNF2 1 (0.028)3. DQ2 - DR17- S - 1267G - TNF1 1 (0.028)4. DQ2 - DR17- L - 1267G - TNF1 1 (0.028)6. DQ2 - DR7 - L - 1267G - TNF1 2 (0.056)8. DQ5 - DR1 - S - 1267G - TNF1 1 (0.028)5. DQ2 - DR17 - L- 1267A - TNF1 1 (0.028)C. Haplotypes with only one AH8.1 constituent7. DQ5 - DR1 - L - 1267G - TNF1 2 (0.056)9. DQ7 - DR13 - L - 1267G - TNF1 1 (0.028)10. DQ8 - DR4 - L -1267G - TNF1 2 (0.056)12 DQ5 - DR10 - L -1267A - TNF2 1 (0.028)14. DQ6 - DR13 - L -1267A - TNF2 1 (0.028)D. Haplotypes with none of the AH8.1 constituents11. DQ5- DR1 - L -1267A - TNF1 4 (0.111)13. DQ5 - DR16 - L -1267A - TNF1 2 (0.056)15. DQ6 - DR15 - L -1267A - TNF1 1 (0.028)16. DQ7 - DR11 - L -1267A - TNF1 4 (0.111)17. DQ8- DR4 - L -1267A - TNF1 3 (0.083)18. DQ9 - DR7 - S -1267A - TNF1 1 (0.028)
Abbreviations used: S, monomodular RCCX with short C4 gene; L, RCCX module(s) long C4gene in first module; 1267G, −1267G in HSP70-2; 1267A, −1267A in HSP70-2; TNF2, −308A inTNFA; TNF1, −308G in TNFA. Alleles characteristic for AH8.1 are set in bold.

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
HSP70-2G and 8.1 Ancestral Haplotype 313
Ohioan females, Out of these individuals the occurrence of the mono-S-C4B-RCCX module was also tested in 117, and 80 subjects, respectively (Table 2).All observed genotype frequencies were in Hardy-Weinberg equilibrium.
There were only slight differences in the frequencies of all alleles testedbetween the two populations. In both populations the HSP70-2 1267G allele istwice or more as frequent as the TNF2 allele, or the RAGE -429C allele, whichare 1.8-2.4 times more frequent than the mono-S RCCX modules. Thus thefrequency of the HSP70-2 1267G allele is 3.5–4-fold more frequent than thosefor mono-S RCCX.
Carrier Frequency of the AH8.1 and its FragmentsThe frequencies of the subjects carrying all the four AH8.1 alleles (upper
row) 2 or 3 of these alleles, only one or none of them are summarized in Table 3.
Table 2: Frequencies of the 8.1 AH constituent alleles tested in two Caucasian populations.
Allele frequencyHealthy Hungarian subjects
(HH)N=127Healthy Ohioian females
(HO)N=101
HSP70-2 1267G 0.390 0.366TNF -308 A 0.185 0.188Mono-S-RCCX 0.089 0.106RAGE -429C 0.175 0.182
Table 3: Frequency of carriers of the 8.1 ancestral haplotype and its fragments in two different Caucasian populations.
TNF2 HSP70-2G Mono-S RAGE-429C Hungarians Ohioian females
+ + + + 0.179 0.180
+ + + − 0.000 0.028− + + + 0.000 0.014+ + − + 0.019 0.028
+ + − − 0.075 0.056− + + − 0.000 0.000− − + + 0.009 0.000+ − + + 0.000 0.000+ − + − 0.000 0.014+ − − + 0.000 0.014− + − + 0.066 0.042
+ − − − 0.047 0.028− + − − 0.283 0.278− − + − 0.000 0.000− − − + 0.028 0.083
− − − − 0.292 0.250

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
314 P. Kiszel et al.
In both populations most mono-S carriers (76–96%) carried the other threealleles as well. By contrast it was the case only in 29% of the HSP70-2 1267Gcarriers. The RAGE and TNF SNPs were in between (50–59, and 52–56%,respectively). Haplotype of 2–3 markers were present in 4–24%, 26–27%, 27–31%and 29–40% of the mono-S, HSP70-2 G, RAGE -429C, and TNF2 carriers,respectively while none, 44–45%, 10–25%, and 8–15% of the same groups car-ried only one AH8.1 allele.
Strength of the Linkage Between AH8.1 Fragments and theHSP70-2 1267G AlleleFinally, linkage disequilibrium between HSP70-2 on the one hand and dif-
ferent AH8.1 fragments were tested in two different healthy Caucasian popu-lations (Table 4). LD coefficients (D’ values) were calculated with the Arlequinsoftware. Since the linkage was very strong and D’ values approached orreached 1 in many case, we used another approach as well. Odds ratios of thecarriers vs non-carriers of the given haplotype to carry the HSP70-2 G allelein both chromosomes was calculated as well (Table 4).
Again the results were similar, albeit not identical, in both populations.As a general tendency both D’ values and odds ratios were relatively low whenlinkage of one AH8.1 marker to the HSP70-2 G allele was tested. In the case ofthe TNF2 and RAGE -429C allele respectively in the Hungarian and Ohioianpopulation, odds ratios were not significant. When, however, the linkage oftwo-three marker haplotypes to the HSP70-2G allele was calculated, high andsignificant D’ values and odds ratios were obtained.
DISCUSSION
We determined the percentage of carriers of the whole AH8.1 and its frag-ments in two Caucasian populations. The repeated presence of the HLA-DQ2,HLA-DR3(17) in the class II region, together with the monomodular shortRCCX containing a single C4B gene but no C4A gene (mono-S) genotype, the1267G allele of HSP70-2 and TNF2 in the IDDM families is convincing andpowerful evidence for carrying the AH8.1. In accordance with the literaturedata AH8.1 was found to be frequent: 18% of the subjects carried this AH in atleast one chromosome in both populations tested. Similar results wereobtained in the family study. These findings are consistent with the haplotypefrequencies of AH8.1 reported for the Caucasians in the literature (Bartonet al., 2003; Price et al., 1999).
It is worthwhile to point out that the frequency of the 1267G allele ofHSP70-2 is about twice as frequent as the TNF2 allele, and four times as fre-quent as mono-S in the Caucasian populations. In addition, when the relation-ship of the constituents’ alleles/genotypes to the whole AH8.1 was assessed,

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
315
Tab
le 4
:St
ren
gth
of
the
lin
kag
e b
etw
ee
n t
he
HSP
70-G
1267
G a
llele
an
d a
llele
s o
r ha
plo
typ
es
with
kn
ow
n c
on
stitu
en
ts o
f th
e 8
.1
an
ce
stra
l ha
plo
typ
e in
th
ree
diff
ere
nt
Ca
uc
asia
n p
op
ula
tion
s.
Hun
ga
rians
(n=
125)
Ohi
oia
n fe
ma
les
(n=1
01)
Ha
plo
typ
eD
’ (p
) va
lue
s)O
R (9
5% C
I) (
p)
D’ (
p)
valu
es)
OR
(95%
CI)
(p
)
TNF2
-0.
674
(<0.
0001
)2.
32 (
0.88
–6.1
4) (
0.11
9)0.
668
(<0.
0001
)4.
86 (
1.61
–14.
67)
(0.0
048)
RA
GE-
429C
-0.
8223
(0.
0011
)5.
43 (
1.99
–14.
8) (
0.00
08)
0.51
5 (<
0.00
01)
2.74
(0.
94–7
.98)
(0.
087)
RC
CX
-mo
no
-S-C
41.
000
(<0.
0001
)3.
90 (
1.34
–11.
4) (
0.02
3)0.
910
(<0.
0001
)6.
87 (
1.84
–25.
69)
(0.0
049)
TNF2
- R
AG
E-42
9C1.
000
(<0.
0001
)5.
87 (
2.09
–16.
49)
(0.0
011)
0.92
8 (<
0.00
01)
6.95
5 (2
.21–
21.8
8) (
0.00
12)
TNF2
- R
CC
X-m
on
o-S
-C4-
0.91
99 (
<0.
0001
)4.
28 (
1.45
–11.
62)
(0.0
103)
0.66
78 (
<0.
0001
)4.
86 (1
.611
–14.
67) (
0.00
48)
RA
GE-
429C
- R
CC
X-m
on
o-S
-C4
1.00
0 (<
0.00
01)
4.1
1(1.
39–1
2.14
) (0
.021
8)1.
000
(<0.
0001
)10
.60
(2.6
3–42
.67)
(0.
0011
)TN
F2-
RA
GE-
429C
- R
CC
X-m
on
o-S
-C4
1.00
0 (<
0.00
01)
4.55
(1.
52–1
3.60
) (0
.008
4)1.
000
(<0.
0001
)12
.60
(3.0
3–52
.37
(0.0
006)
OR
= o
dd
s ra
tio t
o c
arr
y th
e H
SP70
-2-1
267G
alle
le.

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
316 P. Kiszel et al.
marked differences came out. In both populations, less than one-third of theHSP70-2 1267G carriers carried the whole 8.1 haplotype, another one-thirdcarried its fragments while almost half of them carried the HSP70-2 1267Gallele alone. By contrast, the same ratios for the mono-S genotype are 76–96%,4–24%, and 0%. These ratios for the TNF2 and RAGE-429C carriers were inbetween. As it is expected, the linkage between a haplotype with 2–3 AH8.1markers and the HSP70-2 1267G allele was stronger than that between thisallele and a haplotype with only a single AH8.1 marker allele (Table 4).
Data from the family study also reveals the presence of multiple recombi-nant MHC haplotypes in the families affected with type 1 diabetes. For exam-ple, out of the 11 MHC haplotypes with DQ2, HLA-DR3(17) seven segregatedwith mono-S RCCX, HSP70-2 1267G and TNF2, which are classic constituentsof the AH8.1. The other four, however, have variants with long C4 gene, TNF1allele, or 1267A of HSP70-2. The dichotomic nature of these variants suggestthat they were most likely derived from genetic recombinations. That is, thisraises the possibility that haplotype fragments with only one or two of the fourdesignated markers may have arisen independently of an ancestral chromosome.
Data obtained in a small number of families indicate that the HLA-DQ2,-DR17,mono-S,HSP70-2G,TNF2 (AH8.1) haplotype is transmitted more fre-quently to the diabetic children than to the healthy siblings. This observation,however, needs to be confirmed in a larger family study.
Our present findings may have important implications. In examining thegenetic risk factors of carriers with AH8.1 associated with autoimmune, infec-tious, inflammatory or other diseases (Barton et al., 2005; Candore et al.,2002; Cruz et al., 2006; Lio et al., 2001; Murphy et al., 1983; Price et al., 1999;Schroeder et al., 1998), it would be informative and relevant if the landmarkmarkers in the class III region, the RCCX modules and C4 polygenic varia-tions, the SNPs for HSP70-2 and TNFA are analyzed systematically in addi-tion to the conventional markers for class II and the class I alleles. Ahaplotype analysis for the whole AH8.1 and its fragments is warranted. Itcannot be excluded that some diseases are not associated with the wholeAH8.1 but only with fragments. For example, in a recent work (Laki et al.,2006) we found significantly higher maximal hemoglobinA1C concentration inpatients with type 1 diabetes who carried the RAGE -429C and HLA-DQ2alleles. This trait, however, was not linked to the whole AH8.1.
On the other hand, the relatively low proportion of the whole AH8.1 carri-ers among subjects who carried the HSP70-2 1267G allele indicate that theassociations between the HSP70-2 1267G allele and some diseases such asinfections due to Shigella flexneri (Mantis et al., 1996) septic shock (Schroederet al., 1999; Weingartmann et al., 1999), inflammation in asthma (Bertorelliet al., 1998), atopic keratoconjunctivitis (Berra et al., 1994), and experimentalosteoarthritis (Takahashi et al., 1997) may not be characteristic for the carri-ers of the whole AH8.1.

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
HSP70-2G and 8.1 Ancestral Haplotype 317
ACKNOWLEDGMENTS
This work was supported by the ATHERNET (QLG1-CT-2002–90393) grant ofthe EC and the OTKA T049266 grant of Hungarian Research Fund (GF), andby the U.S. National Institute of Health grants 5R01 AR050078 and 1P01DK55546 (CYY).
REFERENCES
Barton, J. C., Bertoli, L. F., Acton, R. T. (2003). HLA-A and -B alleles and haplotypes in240 index patients with common variable immunodeficiency and selective IgG sub-class deficiency in central Alabama. BMC Med. Genet. 4:3.
Barton, J. C., Wiener, H. W., Acton, R. T., Go, R. C. (2005). Total blood lymphocytecounts in hemochromatosis probands with HFE C282Y homozygosity: relationshipto severity of iron overload and HLA-A and -B alleles and haplotypes. BMC BloodDisord. 5:5.
Berra, A., Dutt, J. E., Nouri, M., Foster, C. S. (1994). Heat shock protein expression inhuman conjunctiva. Invest. Ophthalmol. Vis. Sci. 35:352–357.
Bertorelli, G., Bocchino, V., Zhuo, X., Chetta, A., Del Donno, M., Foresi, A., Testi, R.,Olivieri, D. (1998). Heat shock protein 70 upregulation is related to HLA-DRexpression in bronchial asthma. Effects of inhaled glucocorticoids. Clin. Exp.Allergy 28:551–560.
Blanchong, C. A., Zhou, B., Rupert, K. L., Chung, E. K., Jones, K. N., Sotos, J. F., Zipf,W. B., Rennebohm, R. M., Yung, Y. C. (2000). Deficiencies of human complementcomponent C4A and C4B and heterozygosity in length variants of RP-C4-CYP21-TNX (RCCX) modules in caucasians. The load of RCCX genetic diversity on majorhistocompatibility complex-associated disease. J. Exp. Med. 191:2183–2196.
Candore, G., Lio, D., Colonna, R. G., Caruso, C. (2002). Pathogenesis of autoimmunediseases associated with 8.1 ancestral haplotype: effect of multiple gene interac-tions. Autoimmun. Rev. 1:29–35.
Caplen, N. J., Patel, A., Millward, A., Campbell, R. D., Ratanachaiyavong, S., Wong,F. S., Demaine, A. G. (1990). Complement C4 and heat shock protein 70 (HSP70)genotypes and type I diabetes mellitus. Immunogenetics 32:427–430.
Chung, E. K., Yang, Y., Rupert, K. L., Jones, K. N., Rennebohm, R. M., Blanchong,C. A., Yu, C. Y. (2002). Determining the one, two, three, or four long and short lociof human complement C4 in a major histocompatibility complex haplotype encod-ing C4A or C4B proteins. Am.J.Hum.Genet. 71:810–822.
Corzo, D., Yunis, J. J., Salazar, M., Lieberman, J. A., Howard, A., Awdeh, Z., Alper,C. A., Yunis, E. J. (1995). The major histocompatibility complex region marked byHSP70-1 and HSP70-2 variants is associated with clozapine-induced agranulocy-tosis in two different ethnic groups. Blood 86:3835–3840.
Cruz, E., Vieira, J., Almeida, S., Lacerda, R., Gartner, A., Cardoso, C. S., Alves, H.,Porto, G. (2006). A study of 82 extended HLA haplotypes in HFE-C282Y homozy-gous hemochromatosis subjects: relationship to the genetic control of CD8+ T-lym-phocyte numbers and severity of iron overload. BMC Med Genet 7:16.
Day, C. P., Grove, J., Daly, A. K., Stewart, M. W., Avery, P. J., Walker, M. (1998)Tumour necrosis factor-alpha gene promoter polymorphism and decreased insulinresistance. Diabetologia 41:430–434.

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
318 P. Kiszel et al.
Gabriel, S. B., Schaffner, S. F., Nguyen, H., Moore, J. M., Roy, J., Blumenstiel, B.,Higgins, J., DeFelice, M., Lochner, A., Faggart, M., Liu-Cordero, S. N., Rotimi, C.,Adeyemo, A., Cooper, R., Ward, R., Lander, E. S., Daly, M. J., Altshuler, D. (2002).The structure of haplotype blocks in the human genome. Science 296:2225–2229.
Hudson, B. I., Stickland, M. H., Futers, T. S., Grant, P. J. (2001). Effects of novel poly-morphisms in the RAGE gene on transcriptional regulation and their associationwith diabetic retinopathy. Diabetes 50:1505–1511.
Laki, J., Kiszel, P., Vatay, Á., Blaskó, B., Kovács, M., Körner, A., Madácsy, L.,Blatniczky, L., Almássy, Z., Szalai, C., Rajczy, K., Pozsonyi, É., Karádi, I., Fazakas, Á.,Hosszúfalusi, N., Pánczél, P., Arason, G. J., Wu, Y.-L., Zhou, B., Yang, Y., Yu, C.-Y.,Fust, G. (2006). The HLA 8.1 ancestral haplotype is strongly linked to the C alleleof -429T>C promoter polymorphism of receptor of the advanced glycation endprod-uct (RAGE) gene. Haplotype-independent association of the -429 C allele with highhemoglobinA1C levels in diabetic patients. Mol. Immunol. 43:in press.
Lio, D., Candore, G., Colombo, A., Colonna, R. G., Gervasi, F., Marino, V., Scola, L.,Caruso, C. (2001). A genetically determined high setting of TNF-alpha influencesimmunologic parameters of HLA-B8, DR3 positive subjects: implications forautoimmunity. Hum. Immunol. 62:705–713.
Mantis, N., Prevost, M. C., Sansonetti, P. (1996). Analysis of epithelial cell stressresponse during infection by Shigella flexneri. Infect.Immun. 64:2474–2482.
Miller, S. A., Dykes, D. D., Polesky, H. F. (1988). A simple salting out procedure forextracting DNA from human nucleated cells. Nucleic Acids Res. 16:1215.
Murphy, C. C., Go, R. C., Acton, R. T., Barger, B. O., Roseman, J. M. (1983). Geneticanalysis of multiply-affected families of insulin-dependent diabetes mellitus(IDDM) probands. Hum. Hered. 33:344–356.
Price, P., Witt, C., Allcock, R., Sayer, D., Garlepp, M., Kok, C. C., French, M., Mallal, S.,Christiansen, F. (1999). The genetic basis for the association of the 8.1 ancestralhaplotype (A1, B8, DR3) with multiple immunopathological diseases. Immu-nol.Rev. 167:257–274.
Schroeder, H. W., Jr., Zhu, Z. B., March, R. E., Campbell, R. D., Berney, S. M.,Nedospasov, S. A., Turetskaya, R. L., Atkinson, T. P., Go, R. C., Cooper, M. D., Vol-anakis, J. E. (1998). Susceptibility locus for IgA deficiency and common variableimmunodeficiency in the HLA-DR3, -B8, -A1 haplotypes. Mol. Med. 4:72–86.
Schroeder, S., Reck, M., Hoeft, A., Stuber, F. (1999). Analysis of two human leukocyteantigen-linked polymorphic heat shock protein 70 genes in patients with severesepsis. Crit. Care Med. 27:1265–1270.
Stumpf, M. P. (2002). Haplotype diversity and the block structure of linkage disequilib-rium. Trends Genet. 18:226–228.
Takahashi, K., Kubo, T., Goomer, R. S., Amiel, D., Kobayashi, K., Imanishi, J.,Teshima, R., Hirasawa, Y. (1997). Analysis of heat shock proteins and cytokinesexpressed during early stages of osteoarthritis in a mouse model. OsteoarthritisCartilage 5:321–329.
Vargas-Alarcon, G., Londono, J. D., Hernandez-Pacheco, G., Gamboa, R., Castillo, E.,Pacheco-Tena, C., Cardiel, M. H., Granados, J., Burgos-Vargas, R. (2002). Heatshock protein 70 gene polymorphisms in Mexican patients with spondyloarthropa-thies. Ann. Rheum. Dis. 61:48–51.
Vatay, A., Yang, Y., Chung, E. K., Zhou, B., Blanchong, C. A., Kovacs, M., Karadi, I.,Fust, G., Romics, L. L., Varga, L., Yu, C. Y., Szalai, C. (2003). Relationshipbetween complement components C4A and C4B diversities and two TNFA

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
HSP70-2G and 8.1 Ancestral Haplotype 319
promoter polymorphisms in two healthy Caucasian populations. Hum. Immunol.64:543–552.
Weingartmann, G., Oehler, R., Derkits, S., Oismuller, C., Fugger, R., Roth, E. (1999).HSP70 expression in granulocytes and lymphocytes of patients with polytrauma:comparison with plasma glutamine. Clin.Nutr. 18:121–124.
Yu, C. Y., Chung, E. K., Yang, Y., Blanchong, C. A., Jacobsen, N., Saxena, K., Yang, Z.,Miller, W., Varga, L., Fust, G. (2003). Dancing with complement C4 and the RP-C4-CYP21-TNX (RCCX) modules of the major histocompatibility complex.Prog.Nucleic Acid Res.Mol.Biol. 75:217–292.
Yunis, E. J., Larsen, C. E., Fernandez-Vina, M., Awdeh, Z. L., Romero, T., Hansen, J.A., Alper, C. A. (2003). Inheritable variable sizes of DNA stretches in the humanMHC: conserved extended haplotypes and their fragments or blocks. Tissue Anti-gens 62:1–20.

Dow
nloa
ded
By:
[Füs
t, G
eorg
e] A
t: 11
:51
11 J
uly
2007
Copyright © 2022 FDOKUMEN




















