
Four ways towards tropical herbivore megadiversity
19
REVIEW AND SYNTHESIS Four ways towards tropical herbivore megadiversity Thomas M. Lewinsohn 1 * and Tomas Roslin 2 1 Laborato ´ rio Interac ¸o ˜ es Insetos- Plantas, Depto. Zoologia, IB, UNICAMP, Campinas 13083-970, Sa ˜ o Paulo, Brazil 2 Metapopulation Research Group, Department of Biological and Environmental Sciences, FI-00014 University of Helsinki, Helsinki, Finland *Correspondence: E-mail [email protected] Abstract Most multicellular species alive are tropical arthropods associated with plants. Hence, the host-specificity of these species, and their diversity at different scales, are keys to understanding the assembly structure of global biodiversity. We present a comprehensive scheme in which tropical herbivore megadiversity can be partitioned into the following components: (A) more host plant species per se, (B) more arthropod species per plant species, (C) higher host specificity of herbivores, or (D) higher species turnover (beta diversity) in the tropics than in the temperate zone. We scrutinize recent studies addressing each component and identify methodological differences among them. We find substantial support for the importance of component A, more tropical host species. A meta-analysis of published results reveals intermediate to high correlations between plant and herbivore diversity, accounting for up to 60% of the variation in insect species richness. Support for other factors is mixed, with studies too scarce and approaches too uneven to allow for quantitative summaries. More research on individual components is unlikely to resolve their relative contribution to overall herbivore diversity. Instead, we call for the adoption of more coherent methods that avoid pitfalls for larger-scale comparisons, for studies assessing different components together rather than singly, and for studies that investigate herbivore beta-diversity (component D) in a more comprehensive perspective. Keywords Beta-diversity, diversity structure, herbivory, host specialization, plant–animal interac- tions, rarity, species richness, tropics. Ecology Letters (2008) 11: 398–416 A SPECIAL FONDNESS FOR TROPICAL BEETLES – AND OTHER PLANT-EATING BUGS Recent decades have seen a vigorous debate regarding the probable number of extant species on Earth. In 1982, Terry Erwin stirred this up by suggesting that terrestrial species richness may surpass presently known species by a factor of 15, largely because of undescribed tropical arthropods. ErwinÕs estimate escaped from academia into public awareness. A major Non-Governmental Organization dis- tributed a bumper sticker stating ÔSave Tropical Forests – 30 million insects ca not all be wrongÕ. Nonetheless, this figure is still controversial. As pointed out by Stuart Pimm (2001), our best approximations of the number of species on Earth are far less precise than current estimates of the size of the universe. We do know that most multicellular species alive are tropical arthropods associated with plants (Price 2002), and that the regional diversity of most such taxa is highest in the tropics (with aphids and sawflies as notable exceptions; Dixon et al. 1987; Gaston & Williams 1996). Hence the diversity of herbivorous arthropods at different scales is central to understanding how global biodiversity is put together. Furthermore, herbivorous arthropods maintain many important ecosystem processes (Didham et al. 1996) and form sizeable parts of terrestrial food webs (Lewis et al. 2002; Morris et al. 2004). Given the likely contribution of a large proportion of species to different ecosystem functions (Kinzig et al. 2002; Hector & Bagchi 2007), and the manifold links between species diversity, overall biomass, resource utilization and the stability of important ecosystem services (Balvanera et al. 2006; Cardinale et al. 2006), the dynamics of terrestrial ecosystems seem critically linked to arthropod diversity. A central element in ErwinÕs (1982) estimate was the number of insect species unique to each tree species and, by extension, the expected overlap among the faunas associated with different hosts in tropical forests. Reevaluation of Ecology Letters, (2008) 11: 398–416 doi: 10.1111/j.1461-0248.2008.01155.x Ó 2008 Blackwell Publishing Ltd/CNRS
Transcript of Four ways towards tropical herbivore megadiversity

R E V I E W A N DS Y N T H E S I S Four ways towards tropical herbivore megadiversity
Thomas M. Lewinsohn1* and
Tomas Roslin2
1Laboratorio Interacoes Insetos-
Plantas, Depto. Zoologia, IB,
UNICAMP, Campinas 13083-970,
Sao Paulo, Brazil2Metapopulation Research
Group, Department of
Biological and Environmental
Sciences, FI-00014 University of
Helsinki, Helsinki, Finland
*Correspondence: E-mail
Abstract
Most multicellular species alive are tropical arthropods associated with plants. Hence, the
host-specificity of these species, and their diversity at different scales, are keys to
understanding the assembly structure of global biodiversity. We present a comprehensive
scheme in which tropical herbivore megadiversity can be partitioned into the following
components: (A) more host plant species per se, (B) more arthropod species per plant
species, (C) higher host specificity of herbivores, or (D) higher species turnover (beta
diversity) in the tropics than in the temperate zone. We scrutinize recent studies
addressing each component and identify methodological differences among them. We
find substantial support for the importance of component A, more tropical host species.
A meta-analysis of published results reveals intermediate to high correlations between
plant and herbivore diversity, accounting for up to 60% of the variation in insect species
richness. Support for other factors is mixed, with studies too scarce and approaches too
uneven to allow for quantitative summaries. More research on individual components is
unlikely to resolve their relative contribution to overall herbivore diversity. Instead, we
call for the adoption of more coherent methods that avoid pitfalls for larger-scale
comparisons, for studies assessing different components together rather than singly, and
for studies that investigate herbivore beta-diversity (component D) in a more
comprehensive perspective.
Keywords
Beta-diversity, diversity structure, herbivory, host specialization, plant–animal interac-
tions, rarity, species richness, tropics.
Ecology Letters (2008) 11: 398–416
A S P E C I A L F O N D N E S S F O R T R O P I C A L B E E T L E S –
A N D O T H E R P L A N T - E A T I N G B U G S
Recent decades have seen a vigorous debate regarding the
probable number of extant species on Earth. In 1982, Terry
Erwin stirred this up by suggesting that terrestrial species
richness may surpass presently known species by a factor of
15, largely because of undescribed tropical arthropods.
Erwin�s estimate escaped from academia into public
awareness. A major Non-Governmental Organization dis-
tributed a bumper sticker stating �Save Tropical Forests –
30 million insects ca not all be wrong�. Nonetheless, this
figure is still controversial. As pointed out by Stuart Pimm
(2001), our best approximations of the number of species
on Earth are far less precise than current estimates of the
size of the universe.
We do know that most multicellular species alive are
tropical arthropods associated with plants (Price 2002), and
that the regional diversity of most such taxa is highest in the
tropics (with aphids and sawflies as notable exceptions;
Dixon et al. 1987; Gaston & Williams 1996). Hence the
diversity of herbivorous arthropods at different scales is
central to understanding how global biodiversity is put
together. Furthermore, herbivorous arthropods maintain
many important ecosystem processes (Didham et al. 1996)
and form sizeable parts of terrestrial food webs (Lewis et al.
2002; Morris et al. 2004). Given the likely contribution of a
large proportion of species to different ecosystem functions
(Kinzig et al. 2002; Hector & Bagchi 2007), and the
manifold links between species diversity, overall biomass,
resource utilization and the stability of important ecosystem
services (Balvanera et al. 2006; Cardinale et al. 2006), the
dynamics of terrestrial ecosystems seem critically linked to
arthropod diversity.
A central element in Erwin�s (1982) estimate was the
number of insect species unique to each tree species and, by
extension, the expected overlap among the faunas associated
with different hosts in tropical forests. Reevaluation of
Ecology Letters, (2008) 11: 398–416 doi: 10.1111/j.1461-0248.2008.01155.x
� 2008 Blackwell Publishing Ltd/CNRS

overlap coefficients in light of other studies has produced
total figures much below the original 30 million, of the
order of 6–8 million species (Thomas 1990; Ødegaard
2000a and references therein). Hence, the dissimilarity of
faunas associated with different plant species remains of
special interest to current ecology (Ødegaard 2000a;
Novotny et al. 2006; Dyer et al. 2007). It is an effective
quantity, as it can be directly related to ecological processes
and verified by experiments. For a group as poorly
described as arthropods, this measure may also be more
amenable to extrapolations than, for instance, taxonomic
discovery rates (Bebber et al. 2007).
In this appraisal, we scrutinize current ideas about
patterns underlying the higher diversity of herbivores in
the tropics than in the temperate zone. We propose that
recent notions can be represented within a general
conceptual framework, allowing us to apportion variation
in herbivore diversity into four primary components and
their combinations. We identify the relevant components,
scrutinize the most recent assessments of the role of each,
point out important differences among influential studies
and assess our present understanding of patterns and
explanations of tropical insect biodiversity.
F O U R T I E R S O F D I V E R S I T Y
The general idea of dissecting species diversity into
individual components was conceived in studies of
vegetational change over habitat gradients (Whittaker
1967). We will partition the diversity of herbivorous
arthropods into structural elements of herbivore-plant
associations. In this context, we will not discuss the
ultimate ecological or evolutionary processes generating
high tropical biodiversity, as they have been reviewed
repeatedly (Rosenzweig 1995; Gaston 2000; Hill & Hill
2001; Ricklefs 2004; Mittelbach et al. 2007). Instead, we
propose to focus on the resulting, measurable and specific
building blocks of insect–plant interactions. To this effect,
we put forward that current hypotheses for the higher
diversity of herbivores in the tropics than in the temperate
zone can be usefully grouped into four basic components:
(A) more host plant species per se, (B) more arthropod
species per plant species, (C) higher host specialization, or
(D) higher herbivore beta-diversity (Fig. 1). Clearly, these
components are not mutually exclusive: they may combine
in different ways to produce observed differences in total
diversity. Thus, they offer a comprehensive framework
where general differences in diversity can be partitioned
into individual components A–D, and their relative
contribution thereby conveniently compared. We will first
characterize the components individually, then evaluate the
empirical evidence regarding the magnitude of each one in
turn.
More host plant species per se
If the herbivore assemblages of two different regions have
similar patterns of host association, regional differences in
Figure 1 Four seminal components of tropical arthropod megadiversity. Herbivore diversity is depicted schematically as small circles linked
to their host plants by lines. Each symbol represents a different species. Herbivore species may have multiple hosts as indicated by crossing
lines. For each component, the reference assemblage is shown on the left, the more diverse assemblage on the right. Relevant differences are
highlighted by darkened symbols. Components contributing to increased herbivore diversity from the temperate to the tropical zone are: (A)
more host plant species per se; (B) more herbivore species per plant species; (C) less overlap or similarity of herbivores among hosts,
equivalent to higher host specialization; (D) higher turnover of herbivore species on the same hosts among localities.
Review and Synthesis Diversity of herbivore communities 399
� 2008 Blackwell Publishing Ltd/CNRS

floristic diversity will also produce differences in herbivore
diversity (Fig. 1A). The basic component of herbivore
diversity is therefore directly derived from host plant
diversity. Tropical forests are clearly characterized by
outstanding floristic diversity as compared to their temper-
ate counterparts (Gentry 1988; Richards 1996), a pattern
variously ascribed to a number of factors ranging from
current solar energy to Pleistocene history (Brown &
Lomolino 1998; Hill & Hill 2001). In fact, Erwin�s (1982)
estimate of extant insects was derived from the number of
species per host plant species examined, multiplied by the
number of plant species available (Ødegaard 2000a).
Arguably, a linear relation between plant and herbivore
diversity may be regarded as an ecological null pattern
(Gotelli & Graves 1996), so that relevant studies should
focus on deviations from this pattern. Such deviations may
then fruitfully be partitioned into those attributable to B, C
and ⁄or D (Fig. 1).
More herbivore species per plant species
A second way of increasing diversity involves the packing
of more herbivore species on each host species (Fig. 1B).
The general notion of herbivores being more tightly
packed on each tropical plant species can be traced to
several lines of thought which presume that tropical
conditions favour fundamental processes driving species
evolution. This view was formed by early naturalists like
Darwin (1845) and Wallace (1878) who, faced with the lack
of strong winters and the assumed lack of ice ages in the
tropics, believed that this stability would offer time to
build up both more and more highly specialized species
here than in the temperate zone (but for a critique, see
Vazquez & Stevens 2004). Evolution may indeed proceed
at a higher pace in the tropics, given higher temperatures
and faster generation times (Wright et al. 2006; Mittelbach
et al. 2007). As a corollary, Klopfer & MacArthur (1961)
suggested that the main difference between the tropics and
the temperate zone may reside not in the number of
niches available, but in an increase in the similarity of
coexisting species, as shown by reduced character dis-
placement (but see Hubbell & Foster 1986; Terborgh
1992). The extent to which these causal explanations apply
is beyond the scope of the present review. However,
empirical data tend to suggest that communities of
herbivorous insects on plants may not be saturated. Rather
than converging on a set number of insect species per
plant species, local communities tend to vary in proportion
to their regional species pool (Cornell 1985; Cornell &
Lawton 1992; Caley & Schluter 1997; Gaston 2000).
Hence, even without latitudinal differences in basic niche
width, local pools of herbivore species might vary in size
due to factors affecting species richness at larger spatial
scales, including geological and climatic dynamics (Gaston
2000; Hill & Hill 2001).
Another factor which may increase the number of
herbivore species per host species is the higher structural
complexity in tropical habitats. On average, tropical
vegetation in general and tropical forests in particular will
contain more microhabitats per unit ground area than their
temperate counterparts (MacArthur 1972; Proctor 1986).
Hence, each tropical plant species may offer more complex
resources for herbivore species to share or partition.
Thus, several distinct processes may drive Component B.
As the focus of this review is not on evaluating causes but
on their outcomes, our chief interest is whether they
produce a joint, measurable imprint of Component B.
Higher host specialization
Increased diversity in the tropics may also be due to a
smaller overlap of herbivore species among individual host
plant species (Fig. 1C). Like component B, component C
derives from the general notion that in the tropics
organisms are more specialized, with narrower and more
tightly packed niches (Dobzhansky 1950; Levins 1968;
MacArthur 1972). The key distinction is that component C
assumes that the herbivores divide the available set of plant
species more finely amongst themselves, whereas compo-
nent B derives from more herbivore species sharing any
given host. Nevertheless, separating the diversity compo-
nent attributable to host specialization from that of total
host plant diversity (component A, above) is not straight-
forward, as a flora richer in species will also tend to be richer
in genera and families (though not so among continents,
Prance 1994). Hence a comparison of the similarity of
herbivore faunas of two plant species in different families in
one area against that of two congeneric plant species in
another area is difficult to interpret. Some sort of
phylogenetic control will therefore be desirable (Irschick
et al. 2005; Ødegaard et al. 2005) and, whenever possible,
this should be extended to evaluate the phylogenetic �spread�of potential hosts in a local community (Weiblen et al. 2006).
Higher beta-diversity
The number of species in any region will be a function of
how many species we find locally (alpha-diversity) and the
differences we find in species composition among sites
(beta-diversity). Therefore, as a fourth and last alternative,
high diversity can also be due to higher turnover of species
among sites. Note that, for host-associated insects, some
authors also use beta-diversity to designate the turnover of
herbivores among host species (Novotny et al. 2007a;
Murakami et al. 2008). In our scheme, such among-host
turnover comes under component C, so that here
400 T. M. Lewinsohn and T. Roslin Review and Synthesis
� 2008 Blackwell Publishing Ltd/CNRS

beta-diversity is restricted to the spatial turnover of
herbivores on a given host (Fig. 1D).
Latitudinal patterns of beta-diversity were first sought in
plants. Here, initial work suggested low beta-diversity in
neotropical rainforests, despite high total diversity (Pitman
et al. 1999, 2001; Condit et al. 2002). For herbivores, two
ecological patterns may lead us to expect high species
turnover at low latitudes: first, the latitudinal increase in
species richness seems to accelerate towards the equator
(Rosenzweig 1995; but see Gaston et al. 2007) and second,
the species� latitudinal span allegedly decreases (Rapoport�srule; Stevens 1989). In combination, these two effects
should result in higher rates of species spatial turnover at
lower latitudes. Yet the generality of Rapoport�s rule is still
open to debate (Gaston et al. 1998; Kerr 1999; Arita et al.
2005). As an alternative, Novotny & Weiblen (2005) predict
low turnover rates at low latitudes, for two other reasons
combined: many herbivores are specialized on plant genera
and families rather than species (see below; Novotny et al.
2002), and many species-rich plant genera are widely
distributed across large areas of tropical forests (Gentry
1990; Pitman et al. 1999; Condit et al. 2002). Finally, some
recent studies (e.g. Weiser et al. 2007) argue against any
simple and large-scale relation between range size, species
richness and latitude. Thus, as for other components, there
are several theoretical reasons to examine the extent of
Component D, but conflicting predictions on what to
expect.
To end this overview, we note that the four components
are to be interpreted sensu lato. They therefore encompass
several factors which have sometimes been offered as
alternatives to a simple plant-based scheme. For example,
higher herbivore diversity in tropical forests has been
variously ascribed to greater overall plant biomass or to
higher net primary production (Wright 1983). Where such
explanations will fit in our proposed conceptual framework
will depend on their key feature: thus, if it is the individual
size of plants that allows accommodating more herbivore
species per plant, this comes explicitly under component B.
If �higher biomass� implies �more plant species per unit area�,the pattern falls into component A. Such implications are
further examined in the next section, where we turn to
empirical evidence for components A–D.
T H E E V I D E N C E
Most studies so far have addressed components of diversity
one at a time, and even a quick glimpse through the
literature reveals that different components have attracted
very different levels of interest so far (Fig. 2; see also
Appendix S1 in Supplementary Material). Few studies have
examined several components at the same time, or at least
controlled for others while assessing one. This limits the
direct comparison of the relative contribution of each
component, as well as the interpretation of empirical results.
To illustrate the wide array of study systems and approaches
that have been used to date, we have compiled a set of
representative studies in Table 1, including the sampling
design adopted and the main conclusions that concern
individual components.
More host plants
The general increase in diversity from the poles towards the
equator has been established for a very wide range of taxa,
including plants and insects (Hillebrand 2004). As Gaston
(1992) points out, there are no herbivorous insects where
there are no plants – and plenty of both in the tropics.
Therefore, on a crude scale, a general positive relationship
between numbers of arthropod species and numbers of
plant species is obligatory. Nevertheless, for component A
to be regarded as an important contributor to herbivore
diversity, this large-scale association between the diversity of
plants and the diversity of herbivorous arthropods should
clearly be observable across multiple spatial scales as well as
for multiple taxa. Moreover, as different studies have
examined the general relation between insect and plant
diversity by very different techniques, we should also
examine how methodological choices affect the results. To
these ends, we performed a meta-analysis (Cooper &
Hedges 1994) of effect sizes reported in studies addressing
component A. For other components, approaches and data
are too disparate to allow an equivalent exercise.
To examine whether the diversity of herbivorous insect
species will typically increase with the diversity of plants (the
core of component A), we compiled 31 published studies
reporting 80 correlations between the species richness of
herbivorous insects and plants. Search procedures, exact
criteria for inclusion and details on methodology are given
in Appendix S2 (in Supplementary Material), along with the
list of included studies. For each study, we extracted two
descriptors of the study system and four variables delimiting
how the authors dealt with methodological aspects
(Table 2). As our analysis dealt with correlations, we chose
Fisher�s z-transform as measure of effect size (Cooper &
Hedges 1994). To facilitate interpretation, correlation
coefficients were back-transformed [r = tanh(z)] with their
95% CL.
Our meta-analysis of the literature suggests that compo-
nent A makes a key contribution to high diversity of insect
herbivores in areas with high plant diversity. The overall
average correlation coefficient was high (r = 0.56) and
significantly different from 0 (95% CL: 0.519–0.597) across
a broad range of scales and taxonomic resolution. Contrary
to Wolters et al. (2006), we found no difference in the
average correlation effect size among studies conducted in
Review and Synthesis Diversity of herbivore communities 401
� 2008 Blackwell Publishing Ltd/CNRS

the tropical and temperate zones (P = 0.16), nor among
effect sizes from different arthropod taxa (P = 0.28), which
suggests a general pattern. Nevertheless, high heterogeneity
among individual values (Q = 408.8, d.f. = 79;
P < 0.00001) indicates that the pattern detected is strongly
affected by how the study was conducted. In particular,
sampling effects seemed a pervasive factor. The correlation
effect size was lower for studies focusing on smaller areas
(P = 0.006; Fig. 3a), and studies inferring local species
richness from limited sample size (P = 0.001; Fig. 3b).
Studies building on many records generated over long time
periods (literature compilations) consistently reported the
highest correlations (P = 0.002; Fig. 3c, see also Fig. 3b).
Furthermore, perhaps not surprisingly, studies where the
herbivores were actually observed on the vegetation
revealed a clearer link between herbivore and plant species
richness than studies where the insects were trapped off the
plant (P = 0.001; Fig. 3d).
Figure 2 Numbers of studies addressing
each individual Component A–D, as broken
down by their study region (abcissas:
Temperate [TEMP], Tropical [TROP] or
Comparative [COMP; i.e. studies spanning
both climatic regions]), and also by: (a)
spatial extent of study (local, regional or
continental); (b) habitats explored (forest,
open habitats or a mixture of both), and (c)
principal feeding mode of herbivores
(endophages, ectophages or both). The
studies included in each category are listed
in Appendix S1. Probability shown above
columns are exact values from the Fisher–
Freeman–Halton exact test (two-tailed) for
the Temperate-Tropical comparison (as
these are exact values, no d.f. apply).
Approximate limits for spatial categories are:
local (< 100 km2); regional (< 100 000 km2);
continental (> 500 000 km2). Open habitats
include savannahs, meadows, agricultural
and cleared areas. Endophages are borers,
miners and gallers whose larvae develop
inside tissues of living plants; ectophages are
especially leaf-chewers and sap-suckers.
402 T. M. Lewinsohn and T. Roslin Review and Synthesis
� 2008 Blackwell Publishing Ltd/CNRS

Tab
le1
Rep
rese
nta
tive
stu
die
sth
atev
alu
ate
ind
ivid
ual
com
po
nen
tsco
ntr
ibu
tin
gto
the
tota
ld
iver
sity
of
her
biv
oro
us
inse
cts
(Fig
.1).
Co
lum
n�M
eth
od�i
nd
icat
esth
eap
pro
ach
use
d
toco
llect
sam
ple
san
des
tab
lish
the
ho
stp
lan
tar
ray
per
her
biv
ore
spec
ies.
In�M
ain
con
clu
sio
n�l
ette
rsA
–D
ind
icat
ere
sult
sre
levan
tfo
rd
isti
nct
div
ersi
tyco
mp
on
ents
:A
,m
ore
ho
st
pla
nts
;B
,m
ore
her
biv
ore
spec
ies
per
ho
st;
C,
hig
her
spec
ializ
atio
n;
D,
hig
her
bet
a-d
iver
sity
;()
)in
dic
ates
con
trar
yev
iden
ce.
Ind
ivid
ual
stu
die
sar
eso
rted
acco
rdin
gto
the
com
po
nen
t(s)
that
they
pri
mar
ilyre
late
to.
Ref
eren
ceL
oca
tio
n,
hab
itat
Sp
atia
lex
ten
t
of
stu
dy
Tem
po
ral
exte
nt
of
stu
dy
Tar
get
pla
nts
Tar
get
her
biv
ore
sM
eth
od
Mai
nco
ncl
usi
on
Mu
rdo
ch
etal
.(1
972)
US
A,
Mic
hig
an,
old
fiel
ds
Lo
cal,
3p
lots
,
0.3
ha
tota
l
3ye
ars,
sum
mer
sam
ple
s
Her
bs
(40
spp
.)H
om
.(7
5sp
p.)
Net
swee
pin
g;h
ost
asso
ciat
ion
sn
ot
asse
ssed
A:
loca
lh
erb
ivo
reri
chn
ess
incr
ease
sw
ith
pla
nt
rich
nes
s
Axm
ach
er
etal
.(2
004)
Tan
zan
ia.
tro
pic
al
mo
nta
ne
fore
st,
c.2100
ma.
s.l.
Lo
cal,
2.2
km
span
,13
plo
ts,
2.6
ha
tota
l
2ye
ars,
rain
y
seas
on
s
All
vas
cula
r
pla
nts
(93
spp
.)
Lep
.,G
eom
etri
dae
(114
spp
.)
UV
ligh
ttr
aps;
ho
st
asso
ciat
ion
sn
ot
asse
ssed
A()
):lo
cal
her
biv
ore
rich
nes
sd
ecre
ases
wh
erea
sp
lan
td
iver
sity
incr
ease
sfr
om
clea
red
tom
atu
refo
rest
Erw
in(1
982)
Pan
ama,
LT
FL
oca
l,19
tree
s,
500
ha
1-t
ime
fogg
ing
Tre
e(1
sp.)
Co
l.(6
82
sup
po
sed
her
biv
ore
s)
Fo
ggin
g;h
ost
asso
ciat
ion
and
spec
ializ
atio
n
assu
med
B:
hig
hn
um
ber
of
her
biv
ore
so
na
tro
pic
al
tree
Jan
zen
(1988)
Co
sta
Ric
a,d
ry
tro
pic
alfo
rest
Reg
ion
al,
11000
ha
10
year
s,
con
tin
uo
us
Tre
es,
also
oth
er
gro
wth
form
s
(725
spp
.)
Lep
.,m
ost
ly
foliv
ore
s
(3140
spp
.)
Han
d-p
ickin
g,re
arin
g
reco
rds;
oth
erm
eth
od
s
B()
):lo
wh
erb
ivo
re
rich
nes
sp
erh
ost
C:
hig
hh
ost
spec
ifici
ty
War
d&
Sp
ald
ing
(1993)
Bri
tain
,al
lh
abit
ats
Co
nti
nen
tal
>50
year
sA
llvas
cula
r
pla
nts
(127
fam
ilies
)
All
her
biv
oro
us
inse
cts
and
mit
es
(6933
spp
.)
Co
mp
ilati
on
of
pu
blis
hed
reco
rds,
all
met
ho
ds
B:
her
biv
ore
rich
nes
s
hig
her
on
larg
erh
ost
fam
ilies
,an
dth
ose
wit
h
tree
s,lo
wer
on
aqu
atic
fam
ilies
C:
end
op
hag
esm
ore
spec
ializ
ed(a
th
ost
fam
ilyle
vel
)th
an
ecto
ph
ages
Fre
nze
l&
Bra
nd
l
(2003)
Po
lan
dR
egio
nal
;m
ost
dat
afr
om
exp
erim
enta
l
gard
ens,
span
no
tgi
ven
3–
5ye
ars,
NS
S*
Her
bs,
nat
ive
(41),
alie
n(3
6)
Co
l.,L
ep.,
oth
ers
(172
spp
.)
Han
dp
ickin
g,re
arin
g;p
lus
com
pile
dre
cord
s
B:
no
dif
fere
nce
bet
wee
nn
ativ
ean
d
alie
nh
ost
s
C:
end
op
hag
esm
ore
spec
ializ
edth
an
ecto
ph
ages
No
vo
tny
etal
.(2
006)
Cen
tral
Eu
rop
e
(48–
49�N
),
low
lan
dm
ixed
oak
fore
st;
vs.
Pap
ua
New
Gu
inea
(5�S
),
LT
F
Reg
ion
al,1
8si
tes
intw
ore
gio
ns,
20
and
120-k
m
span
;vs.
3si
tes
in20-k
msp
an
3vs.
6ye
ars,
‡1
year
for
each
ho
st
Tre
es,
two
ph
ylo
gen
eti-
cally
mat
ched
sets
(20
vs.
19
spp
.)
Lep
.,C
ol.,
Hym
.,
Ort
h.
foliv
ore
s
(850
spp
.)
Han
dp
ickin
g;re
arin
gp
lus
feed
ing
tria
ls
B()
):h
erb
ivo
reri
chn
ess
per
ho
stn
oh
igh
erin
the
tro
pic
sth
anin
the
tem
per
ate
zon
e
C()
):tr
op
ical
her
biv
ore
s
no
mo
reh
ost
spec
ific
than
tem
per
ate
on
es
Review and Synthesis Diversity of herbivore communities 403
� 2008 Blackwell Publishing Ltd/CNRS
Tab
le1
(Con
tinu
ed)
Ref
eren
ceL
oca
tio
n,
hab
itat
Sp
atia
lex
ten
t
of
stu
dy
Tem
po
ral
exte
nt
of
stu
dy
Tar
get
pla
nts
Tar
get
her
biv
ore
sM
eth
od
Mai
nco
ncl
usi
on
Co
rnel
l
(1985)
US
A,
Cal
ifo
rnia
,
var
iou
sh
abit
ats
Reg
ion
al,
c.
900-k
m
span
2ye
ars,
sum
mer
cen
suse
s
Que
rcus
(9sp
p.)
Hym
.ga
ll-m
aker
s
(max
39
spp
.p
er
ho
st,
tota
ln
ot
given
)
Vis
ual
cen
sus
of
galls
B:
no
evid
ence
of
loca
l
satu
rati
on
;
D:
con
stan
tra
tio
of
loca
l
tore
gio
nal
rich
nes
s
An
dre
w
&H
ugh
es
(2004,
2005)
Au
stra
lia,
Eco
ast,
26�–
35�S
;o
pen
low
lan
deu
caly
pt
fore
st
Reg
ion
al,
1150-k
msp
an
2ye
ars,
8
sam
ple
s
Aca
cia
(1fo
cal
+
8co
occ
urr
ing
spp
.)
Co
l.,H
em(1
94
spp
.)
Sh
ort
-dis
tan
cefo
ggin
gB
()):
Hem
rich
nes
s
dec
reas
esw
ith
lati
tud
e;
Co
l.n
och
ange
.
D(±
):b
eta
div
ersi
ty
dec
reas
esw
ith
lati
tud
e
(lo
cal
rich
nes
sco
nst
ant,
regi
on
alri
chn
ess
dec
reas
es);
bu
tsi
mila
rity
hig
her
atlo
wer
lati
tud
es
Zw
olf
er
(1987)
vs.
Lew
inso
hn
(1991)
Eu
rop
ean
dN
Tu
rkey
(41–
51�N
),
mo
stly
op
en
hab
itat
s;vs.
SE
Bra
zil
(17–
24�S
),co
asta
l
du
nes
tom
on
tan
e
mea
do
ws
Co
nti
nen
tal,
77
area
s,2700-k
m
span
vs.
regi
on
al,
11
loca
tio
ns,
720-k
msp
an
>10
year
s,
NS
Svs.
2ye
ars,
3sa
mp
les
in
dif
fere
nt
seas
on
s
Ast
erac
eae,
her
bs
tosh
rub
s
(37
spp
.)vs.
her
bs
totr
eele
ts
(70
spp
.)
Dip
t.,
Lep
.,C
ol.,
flo
wer
hea
d
end
op
hag
es(7
7vs.
86
spp
.)
Han
dco
llect
ing;
on
ly
rear
ing
reco
rds
use
d
B:
mo
resp
ecie
so
fin
sect
s
per
pla
nt
inB
razi
lth
an
Eu
rop
e
D:
hig
hb
eta-
div
ersi
tyin
bo
thre
gio
ns
Bas
set
&
No
vo
tny
(1999)
Pap
ua
New
Gu
inea
,
LT
Fan
d
coas
tal
site
s
17
·31
km
span
,p
lus
nea
rsh
ore
isla
nd
s
1–
2ye
ars,
var
iab
le
Fic
us(1
5sp
p.)
Co
l.,L
ep.,
Ort
h.
foliv
ore
san
d
Ho
m.
sap
-su
cker
s
(779
spp
.)
Han
dp
ickin
g;fe
edin
g
tria
lsan
dre
arin
g
B:
ver
yh
igh
rich
nes
sp
er
ho
stp
lan
t
D:
few
erin
sect
spp
.o
n
coas
tal
ho
sts
Maw
dsl
ey
&S
tork
(1997)
Bru
nei
,L
TF
loca
l,1-k
msp
an1-t
ime
fogg
ing
tree
s(5
spp
.)C
ol.
adu
lts
(859
spp
.,in
clu
din
g
oth
erfe
edin
g
mo
des
)
Fo
ggin
g;h
ost
asso
ciat
ion
infe
rred
fro
mco
n-
vs.
het
ero
spec
ific
inci
den
ce
C()
):h
ost
spec
ifici
tylo
w:
10%
of
Co
l.sp
ecie
sin
can
op
yes
tim
ated
as
ho
st-s
pec
ific
Ød
egaa
rd
(2000b
)
Pan
ama.
Can
op
y
of
dry
LT
F
Lo
cal,
on
e
site
,0.8
ha
1ye
ar,
mo
nth
ly
(day
⁄nig
ht)
Tre
esan
dlia
nas
(50
spp
.)
Co
l.ad
ult
s,le
af
chew
ers
(286
spp
.)
Bea
tin
g,h
and
pic
kin
gfr
om
cran
e;h
ost
asso
ciat
ion
fro
mo
bse
rvat
ion
s,tr
ials
C:
ho
stsp
ecifi
city
hig
h
No
vo
tny
etal
.
(2002)
Pap
ua
New
Gu
inea
,
LT
F
Lo
cal,
4si
tes,
17
·31
km
span
6ye
ars,
atle
ast
mo
nth
ly
Tre
esan
dsh
rub
s
(51
spp
.)
Co
l.,L
ep.,
Ort
h.,
foliv
ore
s(9
35
spp
.)
Han
dp
ickin
g;fe
edin
gtr
ials
and
rear
ing
C()
):lo
wfe
edin
g
spec
ializ
atio
n
Dye
ret
al.
(2007)
Can
ada
toB
razi
l,8
area
s(5
3�N
–16�S
).
Fo
rest
s,ce
rrad
o
and
oth
erh
abit
ats.
Lo
cal
to
con
tin
enta
l,
1600–
80
000
000
ha*
5–
20
year
s,
NS
S*
Tre
es(2
area
s);
or
all
gro
wth
form
s(1
27–
653
spp
.)*
Lep
.,m
ost
ly
bu
tter
flie
s(1
4–
281
spp
.)*
Han
d-p
ickin
g,b
eati
ng
and
oth
erre
cord
s;re
arin
g
reco
rds
on
ly
C:tr
op
ical
her
biv
ore
sm
ore
ho
stsp
ecifi
cth
an
tem
per
ate
on
es
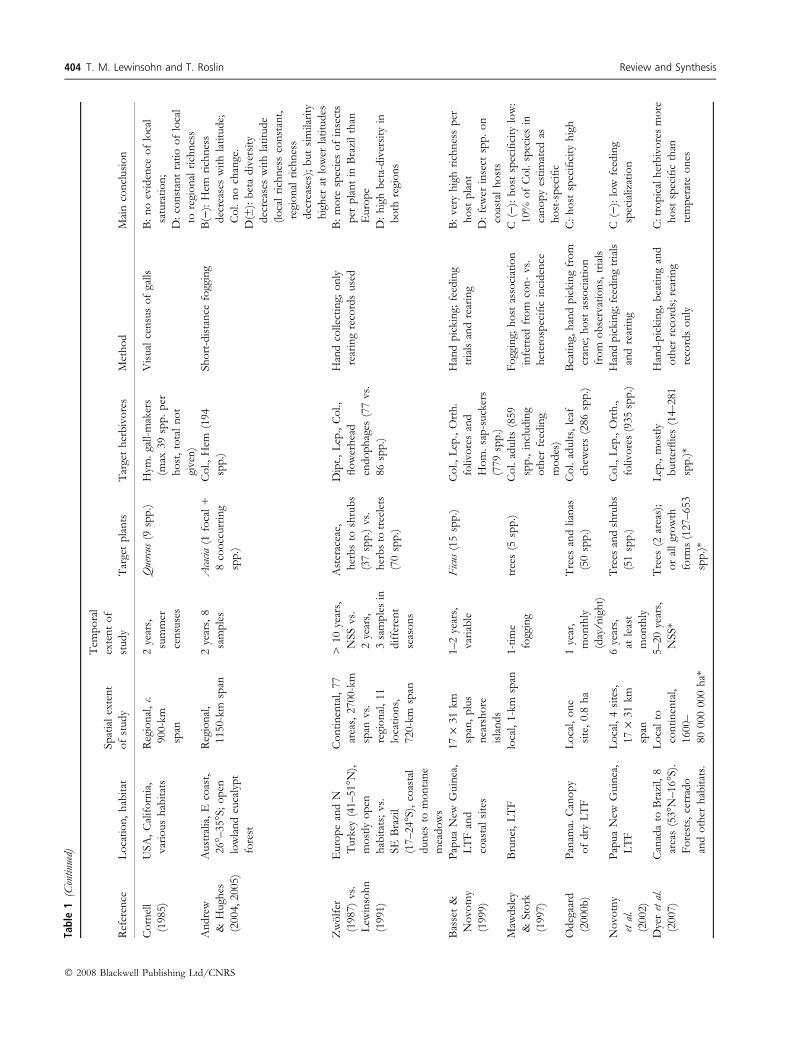
404 T. M. Lewinsohn and T. Roslin Review and Synthesis
� 2008 Blackwell Publishing Ltd/CNRS
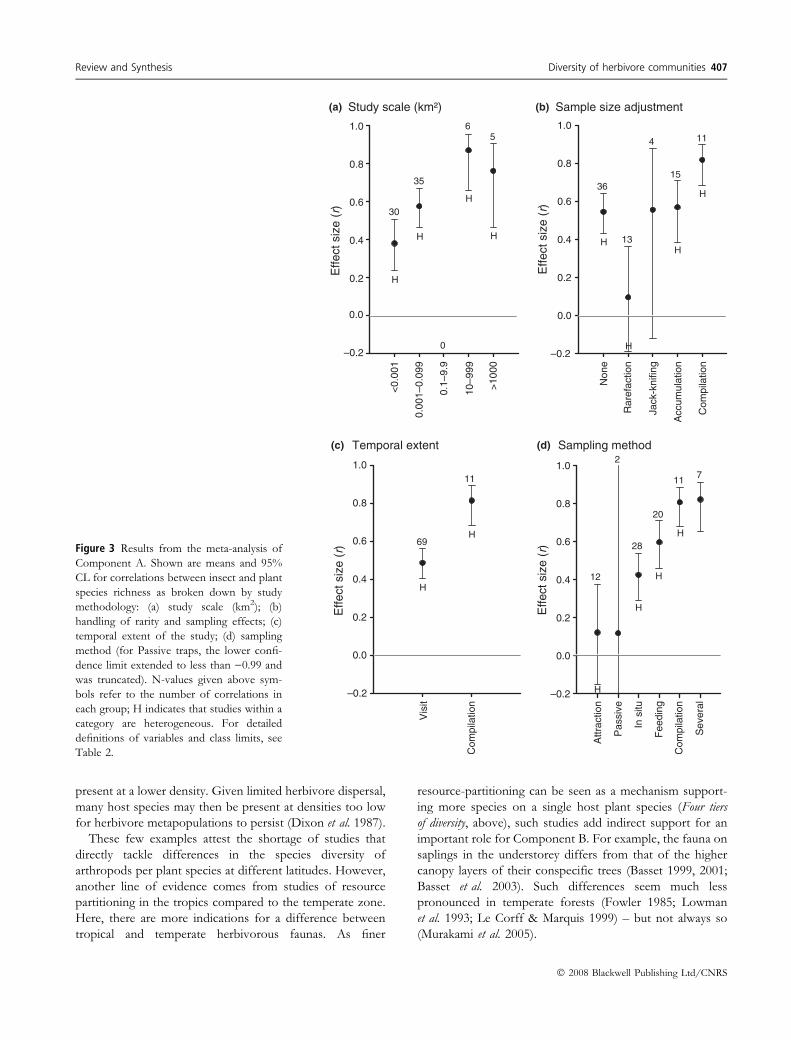
Heterogeneity remained high within groups even after
subdividing studies by methodology (Fig. 3), preventing the
estimation of a single, joint effect size (Cooper & Hedges
1994). Nevertheless, we believe that the most dependable
values derive from studies building on long-term samples
from relatively large areas. For such studies, the correlation
values typically range around 0.8 (Fig. 3a,c), which closely
matches the overall figure reported by Wolters et al. (2006)
for correlations between auto- and heterotroph species
richness. This suggests that c. 60% of variation in the species
richness of herbivorous insects is explained by variation in
the number of plant species.
We conclude that studies that sample herbivorous insects
with appropriate methods and sample sizes typically find an
intermediate to high correspondence between the species
diversity of plants and insects. Therefore, there is consid-
erable support for component A as a prime contributor to
the megadiversity of insects in the diverse flora of the
tropics. Nevertheless, the observed pattern leaves sub-
stantial scope for contribution by other components. This
impression is reinforced by the fact that significant
heterogeneity remains within almost every group formed
in our meta-analysis (Fig. 3), highlighting the unexplained
variation in herbivore diversity due to factors beyond total
plant diversity.
More herbivore species per plant species
Most of what we know about numbers of insects that
actually feed on individual plant species come from studies
which combine in situ feeding observations, experimental
feeding tests, and ⁄or rearing of immature stages (Zwolfer
1987; Marquis 1991; Basset et al. 2003). To date, these
studies offer little direct support for any significant
difference between tropical and temperate regions in terms
of the general number of herbivore species feeding on any
single plant species (Janzen 1988; Basset & Novotny 1999).
Nevertheless, compared to the fair number of assessments
of herbivore richness on plant species in a given locality or
region (Fig. 2), few studies have actually attempted a direct
comparison of herbivore richness among plants in tropical
and temperate sites. Of those that do, almost every one has
adopted a different approach. Four studies exemplify the
range of approaches taken (Table 1).
Andrew & Hughes (2004, 2005) compared insect
assemblages on a single plant species over a 10� latitudinal
range on Australia; they found a slight decrease in
hemipteran richness, but none in beetles (Table 1).
At a much larger spatial scale, Lawton et al. (1993)
examined another species, the bracken fern (Pteridium
aquilinum complex; Thomson 2005). This plant offers a
unique opportunity to compare what is virtually the same
host species among different continents on which it isTab
le1
(Con
tinu
ed)
Ref
eren
ceL
oca
tio
n,
hab
itat
Sp
atia
lex
ten
t
of
stu
dy
Tem
po
ral
exte
nt
of
stu
dy
Tar
get
pla
nts
Tar
get
her
biv
ore
sM
eth
od
Mai
nco
ncl
usi
on
Flo
ren
&
Lin
sen
mai
r
(1998)
Bo
rneo
,L
TF
Lo
cal,
1lo
calit
y,
scal
e?
2ye
ars,
var
iab
le
sch
edu
les
Lo
wer
-str
atu
m
tree
s(3
spp
.)
Co
l.ad
ult
s
(688
spp
.)
Fo
ggin
g,h
ost
asso
ciat
ion
assu
med
D:
hig
hh
erb
ivo
re
bet
a-d
iver
sity
amo
ng
bo
th
con
-an
dh
eter
osp
ecifi
c
tree
ind
ivid
ual
s
Ød
egaa
rd
(2006)
Pan
ama.
Can
op
y
of
mo
ist
and
dry
LT
F
Lo
cal,
2si
tes
c.80
km
apar
t,
0.8
–0.9
ha
each
1ye
ar,
mo
nth
ly
(day
⁄nig
ht)
Tre
esan
dlia
nas
(102
spp
.)
Co
l.,fo
livo
res
(520
spp
.)
As
inØ
deg
aard
2000b
D:
hig
hh
erb
ivo
re
bet
a-d
iver
sity
amo
ng
two
site
s(c
limat
e?)
No
vo
tny
etal
.(2
007a)
Pap
ua
New
Gu
inea
,
LT
F
Reg
ion
al,
8si
tes
in500-k
m
span
5ye
ars,
3m
on
ths
per
site
Tre
es(2
6sp
p.
in
4ge
ner
a)fo
r
Lep
.
Lep
.fo
livo
res
(370
spp
.),
Co
l.w
oo
d-b
ore
rs
(86
spp
.),
Dip
t.
(46
spp
.)
Lep
.h
and
-pic
ked
on
folia
ge,
rear
ed;
Co
l.
ind
ead
wo
od
bai
t;D
ipt.
insc
ent
trap
s(h
ost
sfr
om
oth
erst
ud
y)
D()
):lo
wh
erb
ivo
re
bet
a-d
iver
sity
in
ho
mo
gen
eou
sh
abit
at
Hab
itat
:L
TF
–lo
wla
nd
tro
pic
alfo
rest
.T
emp
ora
lex
ten
to
fsa
mp
ling:
NS
S–
no
sam
plin
gsc
hed
ule
.T
arge
th
erb
ivo
res:
Co
l.–
Co
leo
pte
ra,
Ort
h.
–O
rth
op
tera
+P
has
mat
od
ea,
Dip
t.–
Dip
tera
,H
om
.–
Ho
mo
pte
ra,
Hym
.–
Hym
eno
pte
ra,
Lep
.–
Lep
ido
pte
ra.
*Det
ails
on
ind
ivid
ual
dat
ase
tsar
egi
ven
inD
yer
etal
.(2
007,
Tab
le1
and
Su
pp
lem
enta
ryO
nlin
eM
ater
ial)
.
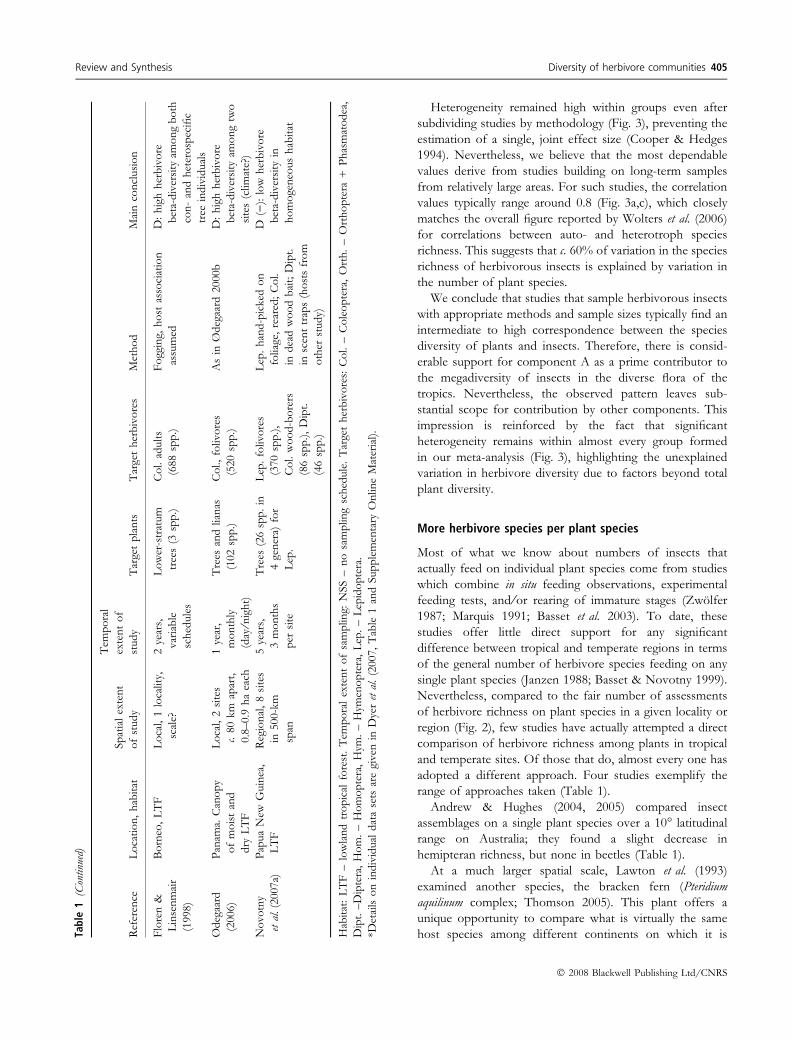
Review and Synthesis Diversity of herbivore communities 405
� 2008 Blackwell Publishing Ltd/CNRS

native. Here, the herbivore assemblage on bracken seems to
reflect idiosyncratic features of each regional biota, and no
latitudinal trend in diversity is detectable (Lawton et al.
1993).
Within an entire host family, Lewinsohn (1991) con-
fronted his own data with those of Zwolfer (1987) to
compare local species richness of flowerhead-feeders on
Asteraceae in the neotropics with that in Europe (Table 1).
Here, species diversity in non-forest ecosystems seems
higher in the neotropics, after adjusting for differences in
sampling effort. However, this comparison is complicated
by the fact that the commonest Asteraceae tribes are
different between the continents, and there is no way to
control for this.
Finally, Dixon et al. (1987); reanalysed by Gaston 1992)
used yet another approach, and focused on the level of the
full flora. With data for 23 countries, they showed that the
ratio of aphid to plant species declines with the number of
plant species per unit area, whereas the ratio of butterfly to
plant species is unrelated to plant diversity. Hence, for
butterflies, species diversity increases in direct proportion to
plant diversity, without any change in species numbers per
plant species. This demonstrates a significant contribution
of Component A (above) but not of Component B.
However, for aphids, tree species at lower latitudes will
actually sustain fewer insect species, an inverse effect of
component B. This may result from a dilution effect: in a
more diverse vegetation, individual host species will be
Table 2 Study descriptors extracted for the meta-analysis of Component A. For each study, we scored two descriptors of the study system
(Climate and Arthropod group) and four variables delimiting how the authors dealt with methodological aspects (Sampling method, Study
scale, Temporal extent and handling of rarity)1. The table defines each variable and classes formed within them.
Variable Classes
Climate – climatic belt where study conducted Temperate ⁄ tropical
Arthropod group – insect order or (where taxonomic
affliation not offered) feeding mode
Coleoptera ⁄Diptera ⁄Heteroptera ⁄Homoptera ⁄Hymenoptera ⁄Lepidoptera ⁄Orthoptera ⁄Thysanoptera ⁄Gallers ⁄Herbivores2 ⁄All3
Sampling method – technique used to generate the insect
material; separates between cases where insects caught or
observed on the plants and methods that build on insect
activity off the plant
Attraction4 ⁄Passive5 ⁄In situ6 ⁄Feeding7 ⁄Compilation8 ⁄Several9
Study scale – scale of individual plots within which plant and
arthropod diversity was measured.10
< 0.001 km2 ⁄0.001–0.099 km2 ⁄0.1–9.9 km2 ⁄10–999 km2 ⁄> 1000 km2
Temporal extent – time span over which the plots were
sampled
Visit11 ⁄Compilation12
Handling of rarity – whether and how author(s) dealt with
rarity and potential sampling problems in the insect material
None13 ⁄Rarefaction ⁄Exclusion14 ⁄Compilation15 ⁄Accumulation16
1Variables are not completely independent, because �compilation� enters as a separate class in three variables. This is justified because
literature compilations differ both from other collection methods (they usually combine data collected by several methods) and from short-
term sampling (they usually sum records obtained over long time periods), and avoid some sampling effects associated with short-term
sampling (they usually build on huge numbers of individual records).2A subset of insects singled out as herbivores by the author(s).3Groups together studies including all insects in the region, regardless of diet, but likely including a non-trivial fraction of herbivores among
them. Studies including at least three different herbivorous insect orders were also attributed to class All.4Trapping methods based on actively attracting the insect to light or baits.5Trapping methods where actively moving individuals are caught off the plant (e.g. pitfall and window traps).6Methods where the insects are observed in, on or near the vegetation (incl. sweep netting, drop traps and line transects).7Methods where the insects are actually observed directly feeding on the plant, e.g. visual observation of gallers.8Studies extracting their data from published catalogues of herbivorous insects and plants.9Studies including several of the methods defined above.10Where sizes varied, the study was classified on the basis of the average size of plots.11Plots sampled by the authors during one or a few field seasons.12Data reflect long-term compilations of the fauna and flora of the study areas.13Author used raw species counts.14Rare species excluded.15Data reflect long-term compilations of the fauna and flora of the study areas.16Species accumulation curve explicitly examined, deemed by author(s) to reach an asymptote.
406 T. M. Lewinsohn and T. Roslin Review and Synthesis
� 2008 Blackwell Publishing Ltd/CNRS
present at a lower density. Given limited herbivore dispersal,
many host species may then be present at densities too low
for herbivore metapopulations to persist (Dixon et al. 1987).
These few examples attest the shortage of studies that
directly tackle differences in the species diversity of
arthropods per plant species at different latitudes. However,
another line of evidence comes from studies of resource
partitioning in the tropics compared to the temperate zone.
Here, there are more indications for a difference between
tropical and temperate herbivorous faunas. As finer
resource-partitioning can be seen as a mechanism support-
ing more species on a single host plant species (Four tiers
of diversity, above), such studies add indirect support for an
important role for Component B. For example, the fauna on
saplings in the understorey differs from that of the higher
canopy layers of their conspecific trees (Basset 1999, 2001;
Basset et al. 2003). Such differences seem much less
pronounced in temperate forests (Fowler 1985; Lowman
et al. 1993; Le Corff & Marquis 1999) – but not always so
(Murakami et al. 2005).
Figure 3 Results from the meta-analysis of
Component A. Shown are means and 95%
CL for correlations between insect and plant
species richness as broken down by study
methodology: (a) study scale (km2); (b)
handling of rarity and sampling effects; (c)
temporal extent of the study; (d) sampling
method (for Passive traps, the lower confi-
dence limit extended to less than )0.99 and
was truncated). N-values given above sym-
bols refer to the number of correlations in
each group; H indicates that studies within a
category are heterogeneous. For detailed
definitions of variables and class limits, see
Table 2.
Review and Synthesis Diversity of herbivore communities 407
� 2008 Blackwell Publishing Ltd/CNRS

To conclude, at present the studies directly comparing
species numbers on tropical and temperate plant species are
relatively few, and their results too varied to support any
generalization. A larger and quicker-growing literature does
reveal differences in the level to which tropical resources are
partitioned among species, suggesting that after all, more
species may be able to share the same tropical plant species
by slicing it in more ways.
Higher host specialization
Arguably, there is more theory and data regarding the host
plant spectra used by herbivore species than on other
aspects of insect-plant associations (Lewinsohn et al. 2005;
Ødegaard 2006). Nevertheless, initial empirical approaches
suffered from methodological problems (see below, It ain�tnecessarily so). More recently, the knowledge of herbivore host
specificity has gradually been improving with studies based
on feeding observations or trials and on rearing records
(Table 1; Janzen 1988; Marquis 1991; Basset & Novotny
1999; Novotny & Basset 2005; Novotny et al. 2004). Here,
high specialization, where strict monophages represent a
large share of the species, has been found in some guilds
(e.g. seed predators, Janzen 1980). In other cases, genera (or
genus groups within families), appear to be the commonest
host span of herbivores (Barone 1998; Novotny et al. 2002),
even those belonging to more specialized guilds (Lewinsohn
1991).
Given this variation in host specificity among taxa, the
evidence for general differences between the tropics and the
temperate zone is ambiguous at best. Overall, the original
expectation of higher specialization in tropical organisms
has been challenged by empirical comparisons of various
organisms (Price 1991); for example, tropical wood-boring
beetles were said to be less, not more, specialized than their
temperate counterparts (Beaver 1979). But as for compo-
nent B, studies actually combining primary data from
different latitudes have been few (Fig. 2). Among the few
that do, Benson (1978) showed that the effective host span
of heliconiine butterflies increases with latitude; in this case,
component C does contribute to an increase in tropical
diversity.
Two other, more recent studies take opposite approaches
to investigate host specialization and its consequences for
herbivore diversity structure. Dyer et al. (2007) examined
specialization patterns over a large geographical range by
aggregating an extensive set of different insect-host studies
(Table 1). By contrast, Novotny et al. (2006) based their
study on a direct comparison of host specificity among
insects on phylogenetically matched sets of hosts in tropical
and temperate local communities (Table 1). Interestingly,
these two studies arrive at completely opposite conclusions.
Whereas Novotny et al. (2002, 2006) assert that tropical
arthropods are no more host-specific than temperate ones,
Dyer et al. (2007) conclude that host specificity is demon-
strably higher in the tropics. Hence, the few comparative
assessments of host specialization in different regions are
Figure 4 Fraction of studies addressing diversity components B
and C by different approaches in different climatic regions
(abcissas: Temperate [TEMP], Tropical [TROP] or Comparative
[COMP; i.e. studies spanning both climatic regions]). (a) Source of
data on feeding association. Here, publication refers to compilation
of published data; mass to data collected by mass sampling such as
fogging, trapping and similar methods; and field to field observa-
tions of actual feeding and ⁄or feeding tests and rearing performed
in the laboratory (b) Growth forms of focal plant taxa. Here,
herb ⁄ shrub refer to studies of herbs, shrubs and vines; tree to studies
focusing on trees including woody shrubs above 3-m height; and all
to studies including many growth forms in the vegetation.
Probabilities shown above columns are exact values from the
Fisher–Freeman–Halton Exact test (two-tailed) for comparisons
among all three climatic regions (as these are exact values, no d.f.
apply). For further details see Appendix S1.
408 T. M. Lewinsohn and T. Roslin Review and Synthesis
� 2008 Blackwell Publishing Ltd/CNRS

fairly evenly balanced in favour and against its contribution
to herbivore diversity in the tropics. Nevertheless, apparent
discrepancies in results may at least partly reflect discrepant
approaches – an issue to which we return below (It ain�tnecessarily so).
Higher beta-diversity
Patterns of species turnover are hard to address both under
temperate and tropical conditions, as they require extensive
– or at least similar – sampling at repeated sites. In the
tropics, the task is often complicated by challenging
logistical conditions. Yet, several studies have addressed
patterns of beta-diversity within each region (Fig. 2), but by
very different approaches. Two contrasting patterns seem
to emerge: the majority of studies have examined sites
differing in their physical conditions, for instance along an
altitudinal gradient (Novotny et al. 2005a) or sites differing
in disturbance or climate (Beck et al. 2002; Brehm et al.
2003; Ødegaard 2006). Studies in this category tend to
show considerable species turnover. This pattern contrasts
with more mixed findings from the very few studies
comparing sites lacking major physical differences. For
such sites in the tropics, the local species pool can typically
represent a large proportion of the regional species pool,
restricting the margin for species turnover among sites.
Such a pattern has been observed among both fruit flies
and butterflies (Orr & Hauser 1996; Novotny et al. 2005b).
In contrast, one neotropical study (Lewinsohn 1991) found
substantial turnover among similar montane sites in the
species composition of flower-head feeding insects
(Table 1). Yet, a more recent study sampling replicate sites
within a 500-km span reports little turnover across a large
expanse of lowland rainforest (Novotny et al. 2007a). The
authors suggest that from an insect perspective, rainforests
may typically consist of relatively large tracts of homoge-
neous habitat, where beta-diversity will contribute little to
overall arthropod diversity. However, the larger-scale
floristic uniformity of tropical rainforests is not supported
by other studies (Tuomisto et al. 1995; Schulman et al.
2007), and hence the results of Novotny et al. (2007a) will
require replication in other regions before their generality
can be asserted.
To conclude our overview of the evidence for Compo-
nents A–D, the bulk of current evidence indicates that
component A makes a major contribution to tropical
megadiversity, whereas evidence regarding the relative
contribution of B–D seems less easy to generalize. For B,
there is emerging consensus that tropical herbivores
partition their resources more finely than temperate ones,
and ⁄or that there are more and more diverse resources to
share. For components C and D, authors disagree on the
magnitude of individual components, and the data collected
to date do not allow major generalizations. Why do studies
diverge to this extent, and how can current obstacles be
overcome?
I T A I N ’ T N E C E S S A R I L Y S O
Unifying current ideas about factors underlying herbivore
diversity in a general framework not only serves to bring
together disparate patterns into a theoretical whole, but also
pinpoints biases in the evidence collected so far. Our review
of the literature shows that such biases represent critical
impediments for the development of the field. While studies
published to date offer substantial insights into how diverse
herbivore assemblages are put together, they do not allow
any fair comparison of the relative contribution of
components A–D. Most importantly, straightforward con-
trasts are compromised by issues related to sampling
methods, sampling effects, taxonomic biases and spatio-
temporal coverage. As illustrated by Table 1 and Figs 2–4,
different studies have approached them in very different
ways, with likely consequences for the results reported.
Sampling method
Some of the most extensive data on the species richness of
tropical arthropods on individual hosts (Table 1, Fig. 4a)
were obtained by mass-sampling the forest canopy by
insecticide fogging (reviewed in Stork et al. 1997). Yet this
approach is really unsuitable for dissecting insect-host
associations. Insects encountered on a given host do not
necessarily feed on it, and the fraction actually doing so is
hard to establish.
The earlier dependence on fogging data may have
influenced our view on the host specificity of tropical
insects (Fig. 4a; Ødegaard 2000a; Basset et al. 2003). Such
data are rife with pitfalls which also beset other mass-
collecting methods, such as vacuuming or net-sweeping. In
particular, much of the data suggesting high turnover of
insect species among host tree species originally lacked
proof of feeding association (Table 1). Two approaches
have been used to address this problem: first, compare
species turnover rates among heterospecific and conspecific
trees (Mawdsley & Stork 1997); second, estimate non-
sampled associations by fitting observations to a binomial
model (Ødegaard 2000a). Both approaches converged on
c. 10% as an estimate of the fraction of herbivore species
that are effectively restricted to an average host. The extent
to which this figure differs between areas with different
herbivore diversity, or among host plant assemblages that
differ in diversity, growth forms and taxonomic or
phylogenetic breadth, remain open questions.
Nevertheless, to fog or not may not be the only
methodological decision with bearings on likely results. As
Review and Synthesis Diversity of herbivore communities 409
� 2008 Blackwell Publishing Ltd/CNRS

shown by the preceding meta-analysis of the literature on
Component A, different sampling techniques tend to show
very different levels of association between species numbers
for herbivores and hosts. The tightest associations are
detected when insects are actually sampled on the vegetation
(or sampled by several techniques combined), whereas
methods that depend on the insects actively moving into
sampling devices report more diffuse associations (Fig. 3d).
As naturalists have known for hundreds of years, how you
seek largely determines what you get.
Rarity and sampling effects
Rarity is the worst nightmare of most diversity studies, and
decisions on how to deal with it often have profound effects
on the results that are obtained. Here, our major concern is
that rarity may affect tropical and temperate studies
differently, and thereby compromise comparisons across
regions.
In the vast majority of biotic communities, most species
are relatively rare, and especially so in species-rich tropical
communities (Price et al. 1995; Basset & Novotny 1999;
Novotny & Basset 2000). Rare species are statistically
elusive, hence researchers face tough choices: sample the
community exhaustively (which may be logistically impos-
sible), truncate data and concentrate on commoner species
(which may bias results), or extra- or intrapolate the data to
estimate real patterns (which again will depend on the
validity of necessary assumptions).
Regarding plant rarity, many authors have simply focused
on common trees (Table 1, Fig. 4b; Erwin 1982; Basset &
Novotny 1999; Barone 2000). For instance, Novotny et al.
(2006); Table 1) chose to sample a fixed number of more
abundant tree species for a phylogenetically controlled
comparison of herbivore richness and host specialization
among tropical and temperate sites. This option may entail a
pitfall: given that more plant species were left out of the
tropical than the temperate survey, and that most of those
are rare hosts, the results of Novotny et al. will only hold for
their entire communities if insect richness and specialization
are not affected by host abundance (Norton & Didham
2007; Novotny et al. 2007b).
Rarefaction techniques, commonly used to make differ-
ent-sized samples comparable (Gotelli & Colwell 2001) do
not fully solve the sampling problem of rare species. In the
meta-analysis of Component A, effect sizes of plant vs.
herbivore species richness were typically even lower for
rarefied samples than for uncorrected values, whereas long-
term data compilations ranked the highest (Fig. 3b).
Likewise, the estimated effect size was higher for long-term
studies than for single samples, likely due to the incom-
pleteness of any short-term sample (Fig. 3c). As each
methodological option comes with a potential bias, the
choice of study organisms and the frequency with which
methods are employed can also affect our general percep-
tion of the literature (Figs 2 and 4a). But do the rare species
most susceptible to sampling effects actually influence the
general structure of insect-plant associations? Recent
research suggests that they do.
Whereas a large fraction of the rare species may be
�tourists� that occur only incidentally on the plant, feeding
trials and rearings show that many rare species are indeed
associated with the studied hosts (Novotny & Basset
2000). Hence, infrequent interactions form a substantial
part of plant-herbivore assemblages (Price et al. 1995;
Novotny & Basset 2000). The simplest alternative in
dealing with such samples is to truncate data and only
consider herbivore species that are represented above an
arbitrary abundance or frequency threshold (Frenzel &
Brandl 1998; Novotny et al. 2007a), but again, in tropical
communities there is a much higher penalty for excluding
species that are statistically inconvenient because of their
rarity. Also, quite worryingly, the consequences of incom-
plete sampling may differ among generalist and specialist
insects (Frenzel & Brandl 1998), so that exclusion of rarer
species will produce biases in comparisons of host
specificity and beta-diversity among data sets and regions.
For beta-diversity, which is fundamentally a reflection of
the range size and local incidence of individual species, the
results are typically established by the commonest species.
Any relation between local abundance and regional
distribution (Hanski et al. 1993) will then threaten the
scope for generalizations.
To sum up, the rarity of a major proportion of both
herbivore and plant species offer one of the chief challenges
to studies of diverse herbivore-plant assemblages. New
procedures may help to overcome at least the statistical
obstacles (Cunningham & Lindenmayer 2005 and compan-
ion papers). For establishing feeding association, we see no
viable alternatives to experimental feeding trials and ⁄or
direct feeding and rearing records.
Taxonomic biases
Data collected from temperate and tropical locations are
often incommensurable. Studies which reach seemingly
different conclusions have often centred on ecologically
distinct taxa (Table 1). In herbivores, what you choose is
partly what you get, as different feeding guilds are known to
exhibit different levels of host specificity. Endophages tend
to be more specialized than ectophages (Gaston et al. 1992;
Ward & Spalding 1993). Thus, of the studies in Table 1,
those focusing on endophages tend to show higher host-
specificity than studies on ectophages, regardless of biome.
Overall, studies to date are biased towards folivores and
other ectophages (Table 1; Fig. 2c) which likely colours our
410 T. M. Lewinsohn and T. Roslin Review and Synthesis
� 2008 Blackwell Publishing Ltd/CNRS

perception of host specificity in general, and its role for
tropical diversity.
The choice of host plant species to study may also
influence results. So far, much of the work conducted in the
tropics is focused on forest trees (Table 1, Figs 2b and 4b;
for exceptions, see Lewinsohn 1991; Marquis 1991),
whereas temperate studies address either herbs (Brassica-
ceae, Frenzel & Brandl 1998; Asteraceae, Zwolfer 1987;
Table 1), shrubs and trees (oaks, Cornell 1985; Moran &
Southwood 1982; Table 1) or a range of growth forms
(Rosaceae, Leather 1986). Yet we now know that distribu-
tional patterns may differ among trees and other growth
forms. Compared to what appears to be low beta-diversity
in trees (Pitman et al. 1999, 2001; Condit et al. 2002),
pteridophytes and Melastomataceae show substantial turn-
over with both distance and habitat heterogeneity (Tuom-
isto et al. 1995, 2003). While some of these differences may
be attributable to sampling effects (Ruokolainen et al. 2002;
Tuomisto et al. 2003), others are likely true. If the insects
track their hosts, we may then expect different patterns
among insects associated with different kinds of hosts.
From the perspective of insects, there may be more to
plant growth form than potential differences in the hosts�distributional patterns: growth form is intimately linked to
structural complexity, which has been shown to influence the
size and composition of insect assemblages associated with
different hosts (Lawton 1983). Nonetheless, interest in
different growth forms has been highly uneven so far,
especially for studies on herbivore specialization (Fig. 4b).
Of the few extant studies, some do suggest differences in
host specificity among taxa associated with different growth
forms. For example, high numbers of beetles feeding on
lianas tend to show relatively high host specificity (Ødegaard
2000a). Hence, several considerations indicate that surveying
multiple components of the vegetation is another key priority
for future work on arthropods (Ødegaard 2000b, 2006).
Finally, differential focus on distinct growth forms may
entail yet another sampling issue. Given that there are more
studies of smaller-sized plants (e.g. herbs and shrubs) in
temperate than in tropical regions (Fig. 4b), tropical samples
may actually comprise more biomass or larger surfaces of
foliage. While some studies have standardized the area of
foliage examined (Barone 2000; Novotny et al. 2006) or
inspection time (Basset & Novotny 1999), most have not.
Though comparisons of host species with similar growth
forms are most desirable, robust comparisons will at least
require adjustment to equal weight or surface.
Spatiotemporal coverage
Studies of herbivore-plant associations have a very different
history in the tropics than in the temperate zone. As a result,
the very type of host plant records available to ecologists
will often differ between latitudes: whereas many temperate
studies analyse host records gathered over large areas and
long time periods (Strong et al. 1984), tropical data will often
come from the vicinity of biological stations or from
intensive efforts focused on relatively small sites (Janzen
1988; Novotny et al. 2002; Table 1, Fig. 2a). Comparisons
among regions are often based on catalogue data collected
over large areas and many years (Fig. 2a). Yet, the choice of
regional extent will likely affect the patterns observed. This
was illustrated by the meta-analysis of data pertinent to
Component A (Fig. 3b), but also by two studies addressing
Component C. As summarized above (see The evidence), Dyer
et al. (2007) and Novotny et al. (2006) take opposite
approaches to examine Component C – and arrive at
opposite conclusions. By aggregating diverse studies of
insect-host associations at local and regional scales, Dyer
et al. (2007) achieve one of the largest data sets ever on
numbers of hosts per insect. Inevitably, they then pay the
price of using heterogeneous data collected over different
time and space scales, and with different aims and sampling
procedures (Table 1). This constrains their conclusions,
because effects of latitude and scale cannot be easily
untangled. By adopting standardized samples and consistent
criteria for analysis, Novotny et al. (2006) are less exposed to
such methodological noise, but their strict approach limits
the number of observations that can be achieved. Their
temperate-tropical comparison relate to two pairs of local
surveys – which also constrains the generality of their
findings.
The contradiction of the results of these two major
studies highlights that current results are contingent on
methodological choices, and that the stalemate of opposing
conclusions will only be overcome by a number of studies
that comprise sets of sufficiently comparable local surveys
replicated in several regions. Only such data will allow us to
assess the relative contributions of Components A–D to
large-scale patterns in herbivore diversity. Such data are also
required to analyse the interaction structure of plants and
herbivores at different scales and its variation across space
and among habitats. This provides a unique link between
the diversity components of our conceptual scheme, and the
structural and functional attributes of interactive plant-
animal assemblages (Lewinsohn et al. 2006).
T O W A R D S I M P R O V E D B R O A D S C A L E
C O M P A R I S O N S
Modern ecologists have worked hard to disclose the factors
underlying tropical megadiversity of herbivores: over the
last decade or so, the density and quality of evidence has
made a quantum leap. This progress is epitomized by the
latest study by Novotny et al. (2007a), based on over 30
person-years spent under remote field conditions. Such
Review and Synthesis Diversity of herbivore communities 411
� 2008 Blackwell Publishing Ltd/CNRS

efforts also incorporate novel methods to investigate
challenging tropical systems. Compilation of the huge data
sets recently published would never haven been possible
without the engagement of local parataxonomists (Basset
et al. 2004; Janzen 2004) and the creative use of recently
produced phylogenies (Novotny et al. 2002, 2006; Weiblen
et al. 2006). Even so, recent progress is still insufficient to
settle disputes on the contribution of various components
to large-scale patterns of insect herbivore diversity. Very
significantly, our review reveals that while new information
is being steadily added both in the tropics and in the
temperate zone, much of it is incommensurable due to
both methodological disparities (different target taxa,
different spatial scales, different ways to handle rarity,
etc.) and to an overall diversity of approaches (especially by
investigating factors singly without controlling for others).
The bottom line of this review is hence that more evidence
of previous kinds will do little to resolve remaining
questions.
Further advances in this field will require new initiatives.
Considering the methodological biases highlighted in this
paper, it is clear that as long as we keep confronting
temperate acorns with tropical breadfruit, we will be
stumped by the same problems of incomparability over and
over again. Given the limitations and challenges that
tropical studies face, researchers were forced to devise ways
of overcoming these obstacles, and it may now be more
profitable to adopt similar protocols in temperate regions
for broad-scale collaborations (Lewinsohn et al. 2005).
What we really need to implement are common and strict
suites of methods across both tropical and temperate sites
(Novotny et al. 2006). But in addition to attempting to
implement the same procedures at several sites across the
globe, it is also important to recognize that some factors
are truly hard to control for. Breadfruit is breadfruit and
acorns are acorns, and whole forests cannot be trans-
planted in a reciprocal approach. As tropical and temperate
forests differ in terms of several factors at once (such as
climate, structural complexity, taxonomic and phylogenetic
diversity of plants), significant effort should be put not
only into detailed dissections of individual associations, but
also into valuable descriptive studies that document
differences between tropical and temperate forests, and
their implications for the assembly of arthropod-plant
communities (for such an ambitious initiative, see Basset
et al. 2007).
Our survey of the literature also highlights that some
components of diversity have received much less attention
than others (Fig. 2). In particular, we argue that stronger and
more comprehensive approaches to quantify patterns of
beta-diversity are needed. The remarkably low species
turnover among distant sites within uniform forest (Nov-
otny et al. 2007a) is related to a very particular definition of
beta-diversity: to species turnover among aggregate samples
from similar sampling localities. The observed pattern
reflects the wide distributional range of some common
species, but it reveals little about local species incidence on
individual host trees, i.e. about patterns that derive from
processes at the population level. More generally, although
the wide distributional range of common species (Hanski
et al. 1993) may inform us of dispersal over evolutionary
time (as reflected in the range sizes of different species), it
reveals little about dispersal in ecological time (with respect
to the contemporary flow of individuals among extant
populations). Novotny et al. (2006, 2007b; see also Novotny
& Weiblen 2005) argue that low beta-diversity in homoge-
neous environments implies that dispersal limitation is not a
key constraint for herbivorous insects. However, only direct
studies of insect dispersal may resolve this issue (Roslin &
Kotze 2005), but these are critically lacking for tropical
regions.
Patterns of low beta-diversity now observed for the first
time in arthropods seem akin to those previously reported
for tropical trees (Pitman et al. 1999, 2001; Condit et al.
2002). Here, a first paradigm of low beta-diversity was later
challenged by results from other taxa and growth forms, and
from the comprehension of species turnover among sites
differing in edaphic conditions (Tuomisto et al. 1995, 2003).
We therefore suggest that additional work on smaller-scale
patterns, on taxa that include other growth forms as well as
trees, may greatly enrich the emerging view on beta-diversity
in tropical as well as temperate arthropods.
Finally, we advocate studies that strive to address
different components of herbivore diversity together rather
than singly. This overview of current knowledge showed
that most studies to date have focused on one component in
a single locality or region. It is high time to collaborate on
more comprehensive study designs, replicated over and
across larger and different regions.
A C K N O W L E D G E M E N T S
We thank Lee Dyer, Angela Moles, Vojtech Novotny, Kalle
Ruokolainen, Hanna Tuomisto and two anonymous referees
for insightful comments on earlier drafts of the manuscript,
and for providing advance copies of papers in press. We
also wish to acknowledge the contribution of many
colleagues, with whom we have discussed these ideas over
the years. Yves Basset and Mar Cabeza-Jaimejuan are
thanked in particular for contributing valuable pointers to
additional literature. Umberto Kubota helped prepare
Figures 1, 2 and 4. Financial support by the Academy of
Finland (grant numbers 111704 and 213457), Fapesp (the
Sao Paulo Research Foundation, grant 04 ⁄15482-1) and
CNPq (the National Research Council of Brazil, grant
306049 ⁄2004) is gratefully acknowledged.
412 T. M. Lewinsohn and T. Roslin Review and Synthesis
� 2008 Blackwell Publishing Ltd/CNRS

R E F E R E N C E S
Andrew, N.R. & Hughes, L. (2004). Species diversity and structure
of phytophagous beetle assemblages along a latitudinal gradient:
predicting the potential impacts of climate change. Ecol. Ento-
mol., 29, 527–542.
Andrew, N.R. & Hughes, L. (2005). Diversity and assemblage
structure of phytophagous Hemiptera along a latitudinal gradi-
ent: predicting the potential impacts of climate change. Global
Ecol. Biogeogr., 14, 249–262.
Arita, H.T., Rodriguez, P. & Vazquez-Dominguez, E. (2005).
Continental and regional ranges of North American mammals:
Rapoport�s rule in real and null worlds. J. Biogeogr., 32, 961–971.
Axmacher, J.C., Tunte, H., Schrumpf, M., Muller-Hohenstein, K.,
Lyaruu, H.V.M. & Fiedler, K. (2004). Diverging diversity pat-
terns of vascular plants and geometrid moths during forest
regeneration on Mt Kilimanjaro, Tanzania. J. Biogeogr., 31, 895–
904.
Balvanera, P., Pfisterer, A.B., Buchmann, N., He, J.S., Nakashizuka,
T., Raffaelli, D. et al. (2006). Quantifying the evidence for bio-
diversity effects on ecosystem functioning and services. Ecol.
Lett., 9, 1146–1156.
Barone, J.A. (1998). Host-specificity of folivorous insects in a
moist tropical forest. J. Anim. Ecol., 67, 400–409.
Barone, J.A. (2000). Comparison of herbivores and herbivory in
the canopy and understory for two tropical tree species. Biotro-
pica, 32, 307–317.
Basset, Y. (1999). Diversity and abundance of insect herbivores
foraging on seedlings in a rainforest in Guyana. Ecol. Entomol.,
24, 245–259.
Basset, Y. (2001). Communities of insect herbivores foraging on
saplings versus mature trees of Pourouma bicolor (Cecropiaceae) in
Panama. Oecologia, 129, 253–260.
Basset, Y. & Novotny, V. (1999). Species richness of insect her-
bivore communities on Ficus in Papua New Guinea. Biol. J. Linn.
Soc., 67, 477–499.
Basset, Y., Novotny, V., Miller, S.E. & Kitching, R.L. (eds) (2003).
Arthropods of Tropical Forests: Spatio-Temporal Dynamics and Resource
Use in the Canopy. Cambridge University Press, Cambridge, UK.
Basset, Y., Novotny, V., Miller, S.E., Weiblen, G.D., Missa, O. &
Stewart, A.J.A. (2004). Conservation and biological monitoring
of tropical forests: the role of parataxonomists. J. Appl. Ecol., 41,
163–174.
Basset, Y., Corbara, B., Barrios, H., Cuenoud, P., Leponce, M.,
Aberlenc, H.-P. et al. (2007). IBISCA-Panama, a large-scale study
of arthropod beta-diversity and vertical stratification in a low-
land rainforest: rationale, study sites and field methodology. Bull.
Inst. R. Sci. Nat. Belg. Entomol., 77, 39–70.
Beaver, R.A. (1979). Host specificity of temperate and tropical
animals. Nature, 281, 139–141.
Bebber, D.P., Marriott, F.H.C., Gaston, K.J., Harris, S.A. &
Scotland, R.W. (2007). Predicting unknown species numbers
using discovery curves. Proc. R. Soc. London B. Biol. Sci., 274,
1651–1658.
Beck, J., Schulze, C.H., Linsenmair, K.E. & Fiedler, K. (2002).
From forest to farmland: diversity of geometrid moths along
two habitat gradients on Borneo. J. Trop. Ecol., 18, 33–51.
Benson, W.W. (1978). Resource partitioning in passion vine but-
terflies. Evolution, 32, 493–518.
Brehm, G., Homeier, J. & Fiedler, K. (2003). Beta diversity of
geometrid moths (Lepidoptera: Geometridae) in an Andean
montane rainforest. Divers. Distrib., 9, 351–366.
Brown, J. & Lomolino, M. (1998). Biogeography, 2nd edn. Sinauer,
Sunderland, MA.
Caley, M.J. & Schluter, D. (1997). The relationship between local
and regional diversity. Ecology, 78, 70–80.
Cardinale, B.J., Srivastava, D.S., Emmett Duffy, J., Wright, J.P.,
Downing, A.L., Sankaran, M. et al. (2006). Effects of biodiversity
on the functioning of trophic groups and ecosystems. Nature,
443, 989–992.
Condit, R., Pitman, N., Leigh, E.G., Chave, J., Terborgh, J., Foster,
R.B. et al. (2002). Beta-diversity in tropical forest trees. Science,
295, 666–669.
Cooper, H. & Hedges, L.V. (eds) (1994). The Handbook of Research
Synthesis. Russell Sage Foundation, New York.
Cornell, H.V. (1985). Local and regional species richness of cyni-
pine gall wasps on California oaks. Ecology, 66, 1247–1260.
Cornell, H.V. & Lawton, J.H. (1992). Species interactions, local
and regional processes, and limits to the richness of ecological
communities – a theoretical perspective. J. Anim. Ecol., 61,
1–12.
Cunningham, R.B. & Lindenmayer, D.B. (2005). Modeling count
data of rare species: some statistical issues. Ecology, 86, 1135–
1142.
Darwin, C. (1845). The Voyage of the Beagle. Wordsworth, Ware,
Herts.
Didham, R.K., Ghazoul, J., Stork, N.E. & Davis, A.J. (1996).
Insects in fragmented forests: a functional approach. Trends Ecol.
Evol., 11, 255–260.
Dixon, A.F.G., Kindlmann, P., Leps, J. & Holman, J. (1987). Why
there are so few species of aphids, especially in the tropics. Am.
Nat., 129, 580–592.
Dobzhansky, T. (1950). Evolution in the tropics. Amer. Sci., 38,
209–221.
Dyer, L.A., Singer, M.S., Lill, J.T., Stireman, J.O., Gentry, G.L.,
Marquis, R.J. et al. (2007). Host specificity of Lepidoptera in
tropical and temperate forests. Nature, 448, 696–699.
Erwin, T.L. (1982). Tropical forests: their richness in Coleoptera
and other arthropod species. Coleopt. Bull., 36, 74–75.
Floren, A. & Linsenmair, K.E. (1998). Non-equilibrium commu-
nities of Coleoptera in trees in a lowland rain forest of Borneo.
Ecotropica, 4, 55–67.
Fowler, S.V. (1985). Differences in insect species richness and
faunal composition of birch seedlings, saplings and trees: the
importance of plant architecture. Ecol. Entomol., 10, 159–169.
Frenzel, M. & Brandl, R. (1998). Diversity and composition of
phytophagous insect guilds on Brassicaceae. Oecologia, 113, 391–
399.
Frenzel, M. & Brandl, R. (2003). Diversity and abundance patterns
of phytophagous insect communities on alien and native host
plants in the Brassicaceae. Ecography, 26, 723–730.
Gaston, K.J. (1992). Regional number of insects and plant species.
Funct. Ecol., 6, 243–247.
Gaston, K.J. (2000). Global patterns in biodiversity. Nature, 405,
220–227.
Gaston, K.J. & Williams, P.H. (1996). Spatial patterns in taxonomic
diversity. In: Biodiversity: A Biology of Numbers and Difference
(ed Gaston, K.J.). Blackwell Science, Oxford, pp. 202–229.
Review and Synthesis Diversity of herbivore communities 413
� 2008 Blackwell Publishing Ltd/CNRS

Gaston, K.J., Reavey, D. & Valladares, G.R. (1992). Intimacy and
fidelity – internal and external feeding by the British Microlep-
idoptera. Ecol. Entomol., 17, 86–88.
Gaston, K.J., Blackburn, T.M. & Spicer, J.I. (1998). Rapoport�srule: time for an epitaph? Trends Ecol. Evol., 13, 70–74.
Gaston, K.J., Davies, R.G., Orme, C.D.L., Olson, V.A., Thomas,
G.H., Ding, T.S. et al. (2007). Spatial turnover in the global
avifauna. Proc. R. Soc. London B. Biol. Sci., 274, 1567–1574.
Gentry, A.H. (1988). Changes in plant community diversity and
floristic composition on environmental and geographical gradi-
ents. Ann. Miss. Bot. Gard., 75, 1–34.
Gentry, A.H. (ed) (1990). Four Neotropical Rainforests. Yale Univer-
sity Press, New Haven.
Gotelli, N.J. & Colwell, R.K. (2001). Quantifying biodiversity:
procedures and pitfalls in the measurement and comparison of
species richness. Ecol. Lett., 4, 379–391.
Gotelli, N.J. & Graves, G.R. (1996). Null Models in Ecology.
Smithsonian Institution Press, Washington.
Hanski, I., Kouki, J. & Halkka, A. (1993). Three explanations of the
positive relationship between distribution and abundance of
species. In: Species Diversity in Ecological Communities (eds Ricklefs,
R.E. & Schluter, D.). University of Chicago Press, Chicago,
pp. 108–116.
Hector, A. & Bagchi, R. (2007). Biodiversity and ecosystem mul-
tifunctionality. Nature, 448, 188–U6.
Hill, J.L. & Hill, R.A. (2001). Why are tropical rain forests so
species rich? Classifying, reviewing and evaluating theories. Prog.
Phys. Geogr., 25, 326–354.
Hillebrand, H. (2004). On the generality of the latitudinal diversity
gradient. Am. Nat., 163, 192–211.
Hubbell, S.P. & Foster, R.B. (1986). Commonness and rarity in a
neotropical forest: implications for tropical tree conservation.
In: Conservation Biology (ed Soule, M.E.). Sinauer, Sunderland,
pp. 205–231.
Irschick, D., Dyer, L. & Sherry, T.W. (2005). Phylogenetic meth-
odologies for studying specialization. Oikos, 110, 404–408.
Janzen, D.H. (1980). Specificity of seed-attacking beetles in a Costa
Rican deciduous forest. J. Ecol., 68, 929–952.
Janzen, D.H. (1988). Ecological characterization of a Costa Rican
dry forest caterpillar fauna. Biotropica, 20, 120–135.
Janzen, D.H. (2004). Setting up tropical biodiversity for conser-
vation through non-damaging use: participation by parataxon-
omists. J. Appl. Ecol., 41, 181–187.
Kerr, J.T. (1999). Weak links: �Rapoport�s rule� and large-scale
species richness patterns. Global Ecol. Biogeogr., 8, 47–54.
Kinzig, A.P., Pacala, S.W. & Tilman, D. (eds) (2002). The Functional
Consequences of Biodiversity : Empirical Progress and Theoretical
Extensions. Princeton University Press, Princeton, NJ.
Klopfer, P.H. & MacArthur, R.H. (1961). On the causes of tropical
species diversity: niche overlap. Am. Nat., 95, 223–226.
Lawton, J.H. (1983). Plant architecture and the diversity of phy-
tophagous insects. Ann. Rev. Entomol., 28, 23–39.
Lawton, J.H., Lewinsohn, T.M. & Compton, S.G. (1993). Pat-
terns of diversity for the insect herbivores on bracken. In:
Species Diversity in Ecological Communities (eds Ricklefs, R.E. &
Schluter, D.). University of Chicago Press, Chicago, pp. 178–
184.
Le Corff, J. & Marquis, R.J. (1999). Differences between under-
storey and canopy in herbivore community composition and leaf
quality for two oak species in Missouri. Ecol. Entomol., 24, 46–58.
Leather, S.R. (1986). Insect species richness of the British Rosa-
ceae: the importance of host range, plant architecture, age of
establishment, taxonomic isolation and species-area relation-
ships. J. Anim. Ecol., 55, 841–860.
Levins, R. (1968). Evolution in Changing Environments; Some Theoretical
Explorations. Princeton University Press, Princeton, NJ.
Lewinsohn, T.M. (1991). Insects in flower heads of Asteraceae in
Southeast Brazil: a case study on tropical species richness. In:
Plant–Animal Interactions: Evolutionary Ecology in Tropical and Tem-
perate Regions (eds Price, P.W., Lewinsohn, T.M., Fernandes,
G.W. & Benson, W.W.). John Wiley, New York, pp. 525–559.
Lewinsohn, T.M., Novotny, V. & Basset, Y. (2005). Insects on
plants: diversity of herbivore assemblages revisited. Annu. Rev.
Ecol. Evol. Syst., 36, 597–620.
Lewinsohn, T.M., Prado, P.I., Jordano, P., Bascompte, J. & Olesen,
J.M. (2006). Structure in plant–animal interaction assemblages.
Oikos, 113, 174–184.
Lewis, O.T., Memmott, J., Lasalle, J., Lyal, C.H.C., Whitefoord, C.
& Godfray, H.C.J. (2002). Structure of a diverse tropical forest
insect-parasitoid community. J. Anim. Ecol., 71, 855–873.
Lowman, M.D., Taylor, P. & Block, N. (1993). Vertical stratifica-
tion of small mammals and insects in the canopy of a temperate
deciduous forest: a reversal of tropical forest distribution? Selb-
yana, 14, 25.
MacArthur, R. H. (1972). Geographical Ecology; Patterns in the Distri-
bution of Species. Harper & Row, New York.
Marquis, R.J. (1991). Herbivore fauna of Piper (Piperaceae) in a
Costa Rican wet forest: diversity, specificity and impact. In:
Plant–Animal Interactions: Evolutionary Ecology in Tropical and Tem-
perate Regions (eds Price, P.W., Lewinsohn, T.M., Fernandes,
G.W. & Benson, W.W.). John Wiley, New York, pp. 179–208.
Mawdsley, N.A. & Stork, N.E. (1997). Host-specificity and the
effective specialization of tropical canopy beetles. In: Canopy
Arthropods (eds Stork, N.E., Adis, J. & Didham, R.K.). Chapman
& Hall, London, pp. 104–130.
Mittelbach, G.G., Schemske, D.W., Cornell, H.V., Allen, A.P.,
Brown, J.M., Bush, M.B. et al. (2007). Evolution and the latitu-
dinal diversity gradient: speciation, extinction and biogeography.
Ecol. Lett., 10, 315–331.
Moran, V.C. & Southwood, T.R.E. (1982). The guild composition
of arthropod communities in trees. J. Anim. Ecol., 51, 289–306.
Morris, R.J., Lewis, O.T. & Godfray, H.C.J. (2004). Experimental
evidence for apparent competition in a tropical forest food web.
Nature, 428, 310–313.
Murakami, M., Yoshida, K., Hara, H. & Toda, M.J. (2005). Spatio-
temporal variation in Lepidopteran larval assemblages associated
with oak, Quercus crispula: the importance of leaf quality. Ecol.
Entomol., 30, 521–531.
Murakami, M., Ichie, T. & Hirao, T. (2008). Beta-diversity of
lepidopteran larval communities in a Japanese temperate
forest: effects of phenology and tree species. Ecol. Res., 23, 179–
187.
Murdoch, W.W., Evans, F.C. & Peterson, C.H. (1972). Diversity
and pattern in plants and insects. Ecology, 53, 819–829.
Norton, D.A. & Didham, R.K. (2007). Comment on ‘‘Why are
there so many species of herbivorous insects in tropical rain-
forests?’’, Science, 315, 1666b.
Novotny, V. & Basset, Y. (2000). Rare species in communities of
tropical insect herbivores: pondering the mystery of singletons.
Oikos, 89, 564–572.
414 T. M. Lewinsohn and T. Roslin Review and Synthesis
� 2008 Blackwell Publishing Ltd/CNRS

Novotny, V. & Basset, Y. (2005). Host specificity of insect her-
bivores in tropical forests. Proc. R. Soc. London B. Biol. Sci., 272,
1083–1090.
Novotny, V. & Weiblen, G.D. (2005). From communities to
continents: beta diversity of herbivorous insects. Ann. Zool.
Fenn., 42, 463–475.
Novotny, V., Basset, Y., Miller, S.E., Weiblen, G.D., Bremer, B.,
Cizek, L. et al. (2002). Low host specificity of herbivorous
insects in a tropical forest. Nature, 416, 841–844.
Novotny, V., Miller, S., Leps, J., Basset, Y., Bito, D., Janda, M. et al.
(2004). No tree an island: the plant-caterpillar food web of a
secondary rain forest in New Guinea. Ecol. Lett., 7, 1090–1100.
Novotny, V., Miller, S.E., Basset, Y., Cizek, L., Darrow, K., Kaupa,
B. et al. (2005a). An altitudinal comparison of caterpillar (Lepi-
doptera) assemblages on Ficus trees in Papua New Guinea.
J. Biogeogr., 32, 1303–1314.
Novotny, V., Clarke, A.R., Drew, R.A.I., Balagawi, S. & Clifford, B.
(2005b). Host specialization and species richness of fruit flies
(Diptera : Tephritidae) in a New Guinea rain forest. J. Trop.
Ecol., 21, 67–77.
Novotny, V., Drozd, P., Miller, S.E., Kulfan, M., Janda, M., Basset,
Y. et al. (2006). Why are there so many species of herbivorous
insects in tropical rainforests? Science, 313, 1115–1118.
Novotny, V., Miller, S.E., Hulcr, J., Drew, R.A.I., Basset, Y., Janda,
M. et al. (2007a). Low beta diversity of herbivorous insects in
tropical forests. Nature, 448, 692–695.
Novotny, V., Drozd, P., Miller, S.E., Kulfan, M., Janda, M., Basset,
Y. et al. (2007b). Response to comment on ‘‘Why are there so
many species of herbivorous insects in tropical rainforests?’’
Science, 315, 1666c-.
Ødegaard, F. (2000a). How many species of arthropods? Erwin�sestimate revised. Biol. J. Linn. Soc., 71, 583–597.
Ødegaard, F. (2000b). The relative importance of trees versus
lianas as hosts for phytophagous beetles (Coleoptera) in tropical
forests. J. Biogeogr., 27, 283–296.
Ødegaard, F. (2006). Host specificity, alpha- and beta-diversity of
phytophagous beetles in two tropical forests in Panama. Biodiv.
Conserv., 15, 83–105.
Ødegaard, F., Diserud, O.H. & Ostbye, K. (2005). The importance
of plant relatedness for host utilization among phytophagous
insects. Ecol. Lett., 8, 612–617.
Orr, A.G. & Hauser, C.L. (1996). Temporal and spatial patterns of
butterfly diversity in a lowland tropical rainforest. In: Tropical
Rainforest Research – Current Issues. Proceedings of the Conference held in
Bandar Seri Begawan, April 1993 (eds Edwards, D.S., Booth, W.E.
& Choy, S.C.). Kluwer, Dordrecht, pp. 125–138.
Pimm, S.L. (2001). The World According to Pimm : A Scientist Audits the
Earth. McGraw-Hill, New York.
Pitman, N.C.A., Terborgh, J., Silman, M.R. & Nuez, P. (1999). Tree
species distributions in an upper Amazonian forest. Ecology, 80,
2651–2661.
Pitman, N.C.A., Terborgh, J.W., Silman, M.R., Nunez, P., Neill,
D.A., Ceron, C.E. et al. (2001). Dominance and distribution of
tree species in upper Amazonian terra firme forests. Ecology, 82,
2101–2117.
Prance, G.T. (1994). A comparison of the efficacy of higher taxa
and species numbers in the assessment of biodiversity in the
Neotropics. Phil. Trans. R. Soc. London B. Biol. Sci., 345, 89–99.
Price, P.W. (1991). Patterns in communities along latitudinal gra-
dients. In: Plant–Animal Interactions: Evolutionary Ecology in Tropical
and Temperate Regions (eds Price, P.W., Lewinsohn, T.M.,
Fernandes, G.W. & Benson, W.W.). John Wiley, New York, pp.
51–69.
Price, P.W. (2002). Resource-driven terrestrial interaction webs.
Ecol. Res., 17, 241–247.
Price, P.W., Diniz, I.R., Morais, H.C. & Marques, E.S.A. (1995).
The abundance of insect herbivore species in the tropics: the
high local richness of rare species. Biotropica, 27, 468–478.
Proctor, J. (1986). Tropical rain-forest – structure and function.
Prog. Phys. Geogr., 10, 383–400.
Richards, P.W. (1996). The Tropical Rain Forest : An Ecological Study,
2nd edn. Cambridge University Press, Cambridge; New York.
Ricklefs, R.E. (2004). A comprehensive framework for global
patterns in biodiversity. Ecol. Lett., 7, 1–15.
Rosenzweig, M.L. (1995). Species Diversity in Space and Time. Cam-
bridge University Press, Cambridge.
Roslin, T. & Kotze, D.J. (2005). Insects and plants in space. Ann.
Zool. Fenn., 42, 291–294.
Ruokolainen, K., Tuomisto, H., Vormisto, J. & Pitman, N. (2002).
Two biases in estimating range sizes of Amazonian plant species.
J. Trop. Ecol., 18, 935–942.
Schulman, L., Ruokolainen, K., Junikka, L., Saaksjarvi, I.E., Salo,
M., Juvonen, S.K. et al. (2007). Amazonian biodiversity
and protected areas: do they meet? Biodiv. Conserv., 16, 3011–
3051.
Stevens, G.C. (1989). The latitudinal gradient in geographical range:
how so many species coexist in the tropics. Am. Nat., 133, 240–
256.
Stork, N.E., Adis, J. & Didham, R.K. (eds) (1997). Canopy Arthro-
pods. Chapman & Hall, London.
Strong, D.R. Jr, Lawton, J.H. & Southwood, T.R.E. (1984). Insects
on Plants: Community Patterns and Mechanisms. Blackwell, Oxford.
Terborgh, J. (1992). Maintenance of diversity in tropical forests.
Biotropica, 24, 283–292.
Thomas, C.D. (1990). Fewer species. Nature, 347, 237.
Thomson, J.A. (2005). Taxonomy and Evolutionary Relationships in
Bracken: A Worldwide Perspective on the Genus Pteridium for 2005.
International Bracken Group, Liverpool.
Tuomisto, H., Ruokolainen, K., Kalliola, R., Linna, A., Danjoy, W.
& Rodriguez, Z. (1995). Dissecting Amazonian biodiversity.
Science, 269, 63–66.
Tuomisto, H., Ruokolainen, K. & Yli-Halla, M. (2003). Dispersal,
environment, and floristic variation of western Amazonian for-
ests. Science, 299, 241–244.
Vazquez, D. & Stevens, R. (2004). The latitudinal gradient in niche
breadth: concepts and evidence. Am. Nat., 164, E1–E19.
Wallace, A.R. (1878). Tropical Nature and Other Essays. Macmillan
and Co., London,.
Ward, L.K. & Spalding, D.F. (1993). Phytophagous British insects
and mites and their food-plant families – total numbers and
polyphagy. Biol. J. Linn. Soc., 49, 257–276.
Weiblen, G.D., Webb, C.O., Novotny, V., Basset, Y. & Miller, S.E.
(2006). Phylogenetic dispersion of host use in a tropical insect
herbivore community. Ecology, 87, S62–S75.
Weiser, M.D., Enquist, B.J., Boyle, B., Killeen, T.J., Jorgensen,
P.M., Fonseca, G. et al. (2007). Latitudinal patterns of range size
and species richness of New World woody plants. Global Ecol.
Biogeogr., 16, 679–688.
Whittaker, R.H. (1967). Gradient analysis of vegetation. Biol. Rev.,
42, 207–264.
Review and Synthesis Diversity of herbivore communities 415
� 2008 Blackwell Publishing Ltd/CNRS

Wolters, V., Bengtsson, J. & Zaitsev, A.S. (2006). Relationship
among the species richness of different taxa. Ecology, 87, 1886–
1895.
Wright, D.H. (1983). Species-energy theory – an extension of
species-area theory. Oikos, 41, 496–506.
Wright, S., Keeling, J. & Gillman, L. (2006). The road from Santa
Rosalia: a faster tempo of evolution in tropical climates. Proc.
Natl Acad. Sci. USA, 103, 7718–7722.
Zwolfer, H. (1987). Species richness, species packing, and evolu-
tion in insect-plant systems. Ecol. Stud., 61, 301–319.
S U P P L E M E N T A R Y M A T E R I A L
The following material is available for this article:
Appendix S1 Literature included in Figures 2 and 4: search
procedure and studies selected.
Appendix S2 A meta-analysis of Component A: data sources,
studies included and calculations implemented.
This material is available as part of the online article
from: http://www.blackwell-synergy.com/doi/full/10.1111/
j.1461-0248.2008.01155.x.
Please note: Blackwell Publishing are not responsible for the
content or functionality of any supplementary materials
supplied by the authors. Any queries (other than missing
material) should be directed to the corresponding author for
the article.
Editor, Jonathan Chase
Manuscript received 24 November 2007
First decision made 15 December 2007
Manuscript accepted 21 December 2007
416 T. M. Lewinsohn and T. Roslin Review and Synthesis
� 2008 Blackwell Publishing Ltd/CNRS




















