
Robust Resource Provisioning in Time-Varying ... - Semantic Scholar

ORIGINAL PAPER
Fledging success of little auks in the high Arctic: do provisioningrates and the quality of foraging grounds matter?
Johanna E. H. Hovinen • Katarzyna Wojczulanis-Jakubas •
Dariusz Jakubas • Haakon Hop • Jørgen Berge •
Dorota Kidawa • Nina J. Karnovsky • Harald Steen
Received: 20 April 2013 / Revised: 3 February 2014 / Accepted: 4 February 2014
� Springer-Verlag Berlin Heidelberg 2014
Abstract Long-lived birds often face a dilemma between
self-maintenance and reproduction. In order to maximize
fitness, some seabird parents alternate short trips to collect
food for offspring with long trips for self-feeding (bimodal
foraging strategy). In this study, we examined whether
temporal and spatial variation in the quality of foraging
grounds affect provisioning and fledging success of a long-
lived, bimodal forager, the little auk (Alle alle), the most
abundant seabird species in the Arctic ecosystem. We
predicted that an increase in sea surface temperature (SST),
with an associated decrease in the preferred Arctic zoo-
plankton prey, would increase foraging trip durations,
decrease chick provisioning rates and decrease chick
fledging success. Chick provisioning and survival were
observed during three consecutive years (2008–2010) at
two colonies with variable foraging conditions in Spits-
bergen: Isfjorden and Magdalenefjorden. We found that a
change in SST (range 1.6–5.4 �C) did not influence trip
durations or provisioning rates. SST was, however,
negatively correlated with the number of prey items
delivered to a chick. Furthermore, provisioning rates did
not influence chick’s probability to fledge; instead, SST
was also negatively correlated with fledging probability.
This was likely related to the prey availability and quality
in the little auk’s foraging grounds. Our findings suggest
that predicted warmer climate in the Arctic will negatively
influence the ability of parents to provide their chicks, and
consequently, the fledging prospects of little auk chicks.
Keywords Bimodal foraging � Fledging success �Alle alle � Calanus � Svalbard
Introduction
Under stressful conditions, for example when there is a
food shortage, long-lived species are predicted to prioritize
self-maintenance, and thus survival, at the cost of current
reproduction (Stearns 1989; Zera and Harshman 2001).
Seabirds are typically long-lived species (Lack 1968).
Their breeding efforts require a lot of energy since they
need to regularly return to a nest site to feed offspring
(Ricklefs 1983). In order to maximize the energy delivered
to offspring, and also maintain their own body condition
efficiently, some seabirds have adopted a bimodal foraging
strategy of interspersed short and long foraging trips
(Weimerskirch et al. 1994; Ropert-Coudert et al. 2004;
Congdon et al. 2005; Welcker et al. 2012). Parents self-
feed and store energy during long trips (LTs), and subse-
quently spend this energy gathering food for their chicks
during short trips (STs) (Grandeiro et al. 1998; We-
imerskirch et al. 2003; Brown et al. 2012; Welcker et al.
2012). If there is not sufficient amount of food close to
breeding sites or if food availability or quality deteriorates,
J. E. H. Hovinen (&) � H. Hop � H. Steen
Norwegian Polar Institute, Fram Centre, Tromsø, Norway
e-mail: [email protected]
J. E. H. Hovinen
University Centre in Svalbard, Longyearbyen, Norway
K. Wojczulanis-Jakubas � D. Jakubas � D. Kidawa
Department of Vertebrate Ecology and Zoology, University of
Gdansk, Gdansk, Poland
J. Berge
Faculty of Biosciences, Fisheries and Economics, UiT-The
Arctic University of Norway, Tromsø, Norway
N. J. Karnovsky
Department of Biology, Pomona College, Claremont, CA, USA
123
Polar Biol
DOI 10.1007/s00300-014-1466-1

bimodal foragers may commute longer distances and/or
spend more time on foraging (Duriez et al. 2000). This may
reduce the chick provisioning rate (e.g. Duriez et al. 2000;
Davoren and Montevecchi 2003) and hence decrease the
amount of prey and energy delivered to chicks, and nega-
tively influence survival of chicks (Schaffner 1990).
The bimodal foraging strategy has been documented in a
number of procellariiform seabird species (Chaurand and
Weimerskirch 1994; Weimerskirch et al. 2001), and
recently also in the little auk (Alle alle), a small (*160 g),
long-lived, zooplanktivorous alcid (Steen et al. 2007;
Welcker et al. 2009a, b, 2012; Wojczulanis-Jakubas et al.
2010; Brown et al. 2012). The little auk is probably the
most abundant seabird species breeding in the high Arctic
(Stempniewicz 2001; Barrett et al. 2006). It is a monoga-
mous colonial breeder, which lays a single-egg clutch in
rocky crevices in talus slopes, incubates an egg for
*29 days and provisions the hatched chick until it fledges
*20–31 days later (Stempniewicz 1981; Harding et al.
2004; Wojczulanis-Jakubas and Jakubas 2012). Both sexes
incubate the egg and feed the chick, but prior to fledging
only males are left to take care of the chick (Harding et al.
2004; Wojczulanis-Jakubas and Jakubas 2012). Each par-
ent alternates LTs interspersed with 3–5 STs daily to pro-
vide adequate energy for themselves and for the chick
during the intensive and short summer breeding period
(Steen et al. 2007; Welcker et al. 2009a, b; Wojczulanis-
Jakubas et al. 2010; Welcker et al. 2012). The main prey of
little auks during summer is lipid-rich Calanus copepods
that account for up to 90 % of their diet (Karnovsky et al.
2003; Kwasniewski et al. 2010). Little auks are major
harvesters of these copepods, and thus have an important
role in the nutrient cycling of the Arctic marine ecosystem
(Barrett et al. 2006). Arctic, cold water associated Calanus
glacialis and Calanus hyperboreus contain *25 % more
lipid (mg-1 dry weight) than the smaller, Atlantic (boreal)
counterpart Calanus finmarchicus, and are thus the most
preferred calanoid copepod prey by the little auks (Wes-
lawski et al. 1999; Scott et al. 2000; Karnovsky et al. 2003;
Jakubas et al. 2007, 2011). The quality of the little auk’s
foraging ground is largely influenced by the distribution of
warm Atlantic (T [ 3 �C, salinity [ 34.9 PSU) and cold
Arctic (T \ 0 �C, salinity: 34.3–34.8 PSU) water masses
(Saloranta and Svendsen 2001) that regulate the distribu-
tion and abundance of these three Calanus species (Willis
et al. 2006; Daase and Eiane 2007; Blachowiak-Samolyk
et al. 2008; Stempniewicz et al. 2013).
In the present study, we investigated the bimodal for-
aging strategy of parent little auks and the fledging success
of their chicks at the west coast of Spitsbergen, Svalbard.
Inter-annual variation in the inflow and temperature of
Atlantic water into this area can be high (Walczowski et al.
2012), and some of the previous little auk studies have
suggested that resulting changes in the abundance of
associated zooplankton prey may lead to substantial vari-
ation in the little auk foraging effort (Karnovsky et al.
2003; Jakubas et al. 2007, 2011; Kwasniewski et al. 2010).
Increase in the amount of Atlantic zooplankton relative to
Arctic zooplankton may force little auks to spend more
time in search of their preferred Arctic prey (Jakubas et al.
2007, 2011; Kwasniewski et al. 2010; Gremillet et al.
2012). Consequently, little auks may increase the duration
of their LTs and decrease provisioning rates of their chicks
(Welcker et al. 2009a) that in turn could result in lower
amount of food and energy received by the chicks (Ricklefs
1983). This may cause decreased chick survival and
fledging success (Welcker et al. 2009b).
We wanted to further investigate the above-mentioned
suggestions and thereby (1) to test whether varying
oceanographic conditions with their associated zooplank-
ton prey affect foraging trip durations (we considered both
STs and LTs) and chick provisioning rates of little auks,
and then (2) to examine whether variations in these
oceanographic conditions, foraging trips and provisioning
rates influence fledging success of little auk chicks. We
assessed the differences in oceanographic conditions with
ocean surface temperature (SST)—SST has been shown to
be strongly linked to species composition of local zoo-
plankton communities in our study area, with large, lipid-
rich copepods inhabiting cold water (Karnovsky et al.
2010). We predicted that long-lived auks with low fecun-
dity prioritize self-maintenance over that of their chicks,
and that (1) LT durations are longer when SST is higher;
(2) chick provisioning rates are lower when SST is higher;
(3) fledging success of chicks is lower when LT durations
are longer and provisioning rates lower.
Materials and methods
Study area
We collected data during the chick rearing phase (July–
August) in three consecutive years (2008–2010) at two
colonies in the Svalbard archipelago: Isfjorden (78�120N,
15�200E) and Magdalenefjorden (79�350N, 11�050E)
(Fig. 1). Both fjords as well as the adjacent shelf sea area
outside the fjords are regularly supplied with both Atlantic
water from the West Spitsbergen Current and Arctic water
from the South Cape Current (Nilsen et al. 2008, Kwas-
niewski et al. 2010; Trudnowska et al. 2012; Jakubas et al.
2013). Furthermore, the nearby marginal ice zone at the
north-west off Spitsbergen (Fig. 1) provides a good quality
foraging ground for little auks breeding in Magdalenef-
jorden (Kwasniewski et al. 2010; Jakubas et al. 2011, 2012,
2013).
Polar Biol
123

We used SST (�C) to characterize oceanographic con-
ditions and the quality of little auk’s foraging ground. We
estimated mean SST for the chick rearing period (July) of
each study year for an area of *5,500 km2 adjacent to
each of the two colonies (Fig. 1). These areas were chosen
such that they covered the maximum foraging ranges of
little auks observed in our study area (*150 km; Jakubas
et al. 2013). We acquired monthly means of SST data at
4 km 9 4 km horizontal resolution from the moderate-
resolution imaging spectroradiometer (MODIS/aqua;
http://disc.sci.gsfc.nasa.gov/giovanni) and computed the
final July means by averaging all monthly data points
available within the selected areas mentioned above.
Foraging trip duration and chick feeding rate
We estimated foraging trip durations and chick feeding
rates by observing marked adults at both colonies. Birds
were caught during late incubation or early chick rearing
period with a mist net or noose carpet, and marked with a
stainless steel ring and a unique combination of three
plastic colour rings. The observation area at both colonies
was chosen such that it ensured the presence of sufficient
number of birds with visible nest entrances as well as
minimized disturbance.
We carried out observations non-stop for 48 h, from 15
to 26 July in Isfjorden and 27 July to 3 August in Mag-
dalenefjorden, when the chicks were 8–17 days old
(Table 1). During 48-h watches, two observers registered
time of departure and arrival of all the marked birds in the
observation areas. Only birds that returned to colony with
food for their chick (noticeable by their bulging gular
pouch below the beak) were registered. We defined for-
aging trip duration as the time between departing the col-
ony with an empty gular pouch and returning to the colony
with a filled gular pouch. We logged a feeding event when
a bird left the nest with an empty gular pouch after entering
it with a full gular pouch. During the study, we monitored a
total of 114 birds at the two colonies (Table 1).
Chick food samples
We collected 8, 27 and 15 gular pouch samples in Isfjor-
den, and 18, 59 and 30 samples in Magdalenefjorden (in
2008, 2009 and 2010, respectively). Data on the diets from
Magdalenefjorden are also presented in Jakubas et al.
(2011) and Kidawa et al. (submitted). We caught birds
during the 2nd and 3rd week of chick rearing with a mist
net or noose carpet, and procured samples by gently
scooping out the content from the gular pouch. We sam-
pled each bird only once in order to avoid malnutrition of
their chick and a pseudoreplication problem. Samples were
transferred to 4 % formaldehyde solution in the field. Prior
to identification of prey species, samples were rinsed with
fresh water and all big prey items ([ 1 cm) were picked up
and identified. The rest of gular pouch samples were split
(with a simple box splitter; Motoda 1985) into subsamples
that contained approximately 150–300 individuals of Cal-
anus spp. The copepodite stage of Calanus spp. was
determined, and prosome length (PL, mm) measured from
the tip of the cephalosome to the distal lateral end of the
last thoracic segment (Hirche et al. 1994). Identification of
the species was based on the stage and PL (Weydmann and
Kwasniewski 2008). The remaining prey items in subs-
amples were also calculated and identified to species or
taxon. Total number of prey in a gular pouch was extrap-
olated from the subsample.
Chick fledging success
To estimate fledging success of the chicks, we monitored
nests at both colonies. Nests containing egg were located
and marked during late incubation. They were monitored
Fig. 1 Little auk colonies (black dots) in Isfjorden and Magdalenef-
jorden, main water currents transporting Atlantic water (West
Spitsbergen Current) and Arctic water (South Cape Current), average
location of the marginal ice zone at the northwest off Spitsbergen
during the study period, and areas (squares; *5,500 km2 adjacent to
each of the colonies) for which sea surface temperature (SST �C) was
extracted
Polar Biol
123
every 2–3 days during the hatching period in order to
detect hatch date. During the chick rearing period, nests
were monitored every 4–5 days in order to record chick
survival. Frequency of nest checks was increased to every
2–3 days when chicks started to fledge. The presence of an
egg or a chick in the nest was confirmed with a use of light-
scopes (Moritex Europe Ltd, Cambridge, UK) or flash-
lights. We considered chick fledged if it disappeared from
the nest at the age of C20 days post-hatching (Harding
et al. 2004; Wojczulanis-Jakubas and Jakubas 2012).
During the study, we monitored a total of 321 chicks at the
two colonies (Table 1). Due to problems with capturing
sufficient number of birds from particular nests, the chicks
monitored and parents registered during the trip observa-
tions were from different family units.
Statistical analyses
To classify single trips as either ST or LT, we determined
an unbiased data-driven cut-off value separately for each
colony and year according to the procedure by Welcker
et al. (2009a). The bimodal distribution of foraging trip
durations of the little auk can be considered as two separate
log-normal distributions of STs and LTs. The cut-off value
was determined as the minimum sum of variances of both
trip types given their log-normal distributions. Derived cut-
off values varied slightly between the two study sites and
years (Isfjorden: 5–6 h, Magdalenefjorden: 6–8 h). We
calculated daily chick feeding rate as the number of times a
foraging bird returned to the colony and entered the nest
with a filled gular pouch within the 48-h period, divided by
two. The number of prey items delivered to a chick by one
parent during a day was estimated by multiplying the
average number of prey items in one food load (i.e. gular
pouch) with the daily chick feeding rate.
To examine inter-annual and inter-colony variation in
SST data (including all monthly data points available), we
applied an analysis of variance (ANOVA) with study year
and colony as explanatory variables. In all the other models
(see below), we assessed the potential effect of SST on trip
durations, chick provisioning rates and chick fledging
success by first including only year and colony as
explanatory variables into models. Then, we ran additional
models where the year effect was replaced with SST. We
also assessed the effect of other potential explanatory
variables by replacing the year effect with them.
Foraging trip durations between the years and colonies
were compared by fitting linear mixed effects (LMEs)
models with colony and year as fixed effects, and separate
models were run for STs and LTs. Additional LMEs for
both trip types were run to test whether SST, chick age
(day) and LT duration (in ST model) influence trip dura-
tions. Since trip durations and chick ages were estimated
from different parents and nests, respectively, we used a
mean chick age for each colony and for each year (all nests
combined; Table 1). Due to repeated trip duration esti-
mates of the same individuals, ‘‘individual’’ was included
as a random effect into all of these LME models.
We compared daily chick feeding rates between the
years and colonies by applying generalized linear mixed
effect (GLME) model with Poisson error distribution and
with ‘‘individual’’ as a random effect. We fitted additional
GLMEs to test whether SST, LT and ST durations, or chick
age influence daily feeding rates.
We used logistic regression (LR) with binomial error
distribution to test for difference in fledging success
between the years and colonies. Fledging success was
expressed as 0 if the chick survived \20 days (unfledged
chick) and 1 if the chick disappeared from the nest
C20 days post-hatching (fledged chick). We ran additional
and separate LRs to test the effect of SST, LT and ST
durations, and daily chick feeding rates on fledging suc-
cess. In addition, we fitted generalized linear models
(GLM) with Poisson error distribution to test whether
colony and year, or SST and daily chick feeding rates
influence chick’s fledging age. Mean values of each of
these explanatory variables for each year and colony were
used due to lack of real parent–chick pairs.
We log transformed data when needed to meet the
assumption of normality. All results are reported as mean
values with associated standard errors. We performed
model selection by using the Akaike information criterion
adjusted for small sample size (AICc). We excluded all
models with DAICc [ 2 as they have very limited support
Table 1 Mean hatching date
and number of hatched chicks,
and number of parent birds
observed and age of the chicks
(day) during 48-h non-stop
observations (dates shown) in
Isfjorden (I) and
Magdalenefjorden (M)
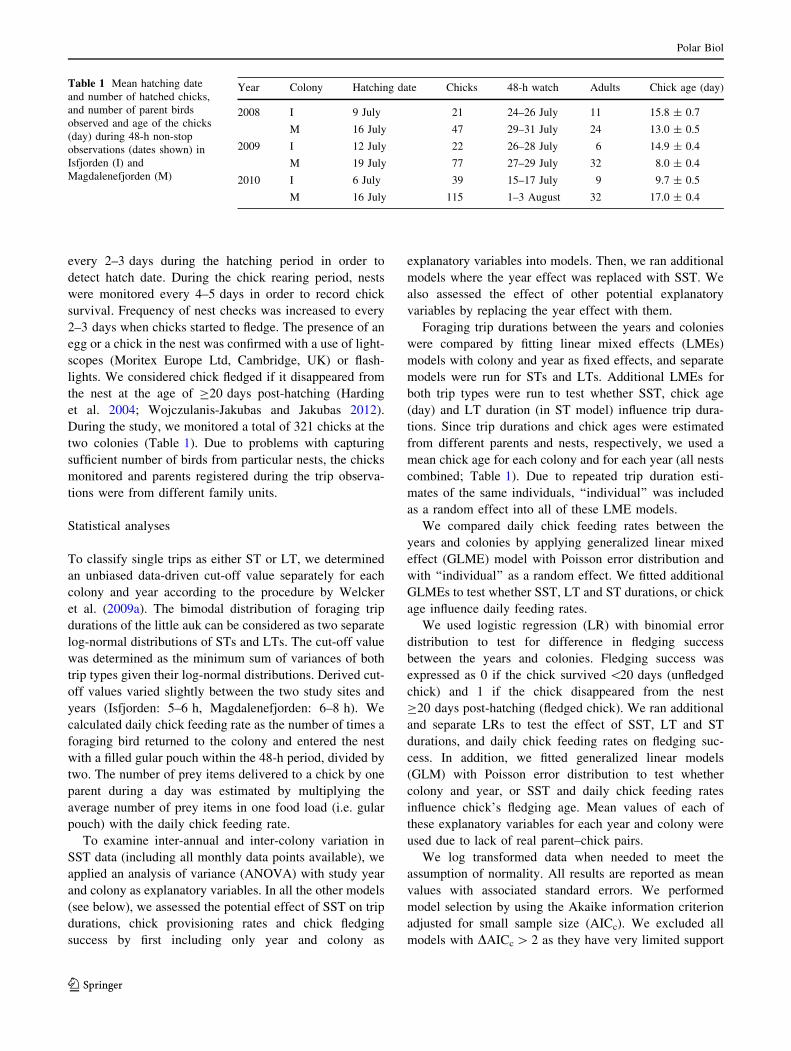
Year Colony Hatching date Chicks 48-h watch Adults Chick age (day)
2008 I 9 July 21 24–26 July 11 15.8 ± 0.7
M 16 July 47 29–31 July 24 13.0 ± 0.5
2009 I 12 July 22 26–28 July 6 14.9 ± 0.4
M 19 July 77 27–29 July 32 8.0 ± 0.4
2010 I 6 July 39 15–17 July 9 9.7 ± 0.5
M 16 July 115 1–3 August 32 17.0 ± 0.4
Polar Biol
123
(Burnham and Anderson 2002). We report all selected
models with their AICc weights and evidence ratios. AICc
weight is the likelihood of the model given the data and the
set of candidate models. Evidence ratio is the relative
weight of the model against the best model. Statistical
analyses were performed in R 2.15.0 (R Development Core
Team 2012).
Results
Sea surface temperature, chick age and trip duration
Model selection suggested that oceanographic conditions
and thereby the quality of little auk’s foraging ground
differed between the two colonies, and varied throughout
the study period (Table 2); SST was higher in the Isfjorden
area (range 4.70–5.38 �C) than in the Magdalenefjorden
area (range 1.59–2.36 �C), indicating a higher presence of
Atlantic water masses in the former compared to the latter
area (Fig. 2a). Top selected LME models describing ST
duration included an interaction between colony and chick
age or SST (both models had similar support; Table 2).
These interactions were, however, ambiguous, because
generally ST duration was similar (*2.2 h) at both colo-
nies, regardless of the differences in chick age or SST
(Fig. 2b). Only exception was 1 year in Isfjorden when STs
were on average shorter (1.7 ± 0.2 h; Fig. 2b). Top
selected LME model describing LT duration included only
the year effect (Table 2): LT duration was shortest during
2009 (Isfjorden: 12.5 ± 1.6 h, Magdalenefjorden:
11.8 ± 0.5 h) and longer (up to 16 h) during 2008 and
2010 at both colonies (Fig. 2c). Neither SST nor chick age
influenced LT duration as models including these effects
were not among the selected models (Table 2).
Daily chick feeding rate
There was strong evidence from model selection that only
LT duration controlled the daily chick feeding rate
(Table 2), with decreased number of food loads delivered
to a chick when LT duration increased (Fig. 3). Feeding
rate was similar at both colonies (range 4.1–4.8 food loads
chick-1 days-1; Fig. 3). Basically no support was shown
for the influence of ST duration on daily feeding rates
(Table 2). Also, intra-annual differences in feeding rate
between the two colonies, and effect of SST and chick age
on feeding rates, were not supported by model selection—
as GLME models including these effects were not among
the selected models (Table 2).
Food delivered to a chick
The average number of prey items delivered to a chick by
one parent during a day was lower in Isfjorden (\6,300)
than in Magdalenefjorden ([7,400) throughout the study
Table 2 Model selection results for the models used in the statistical analyses
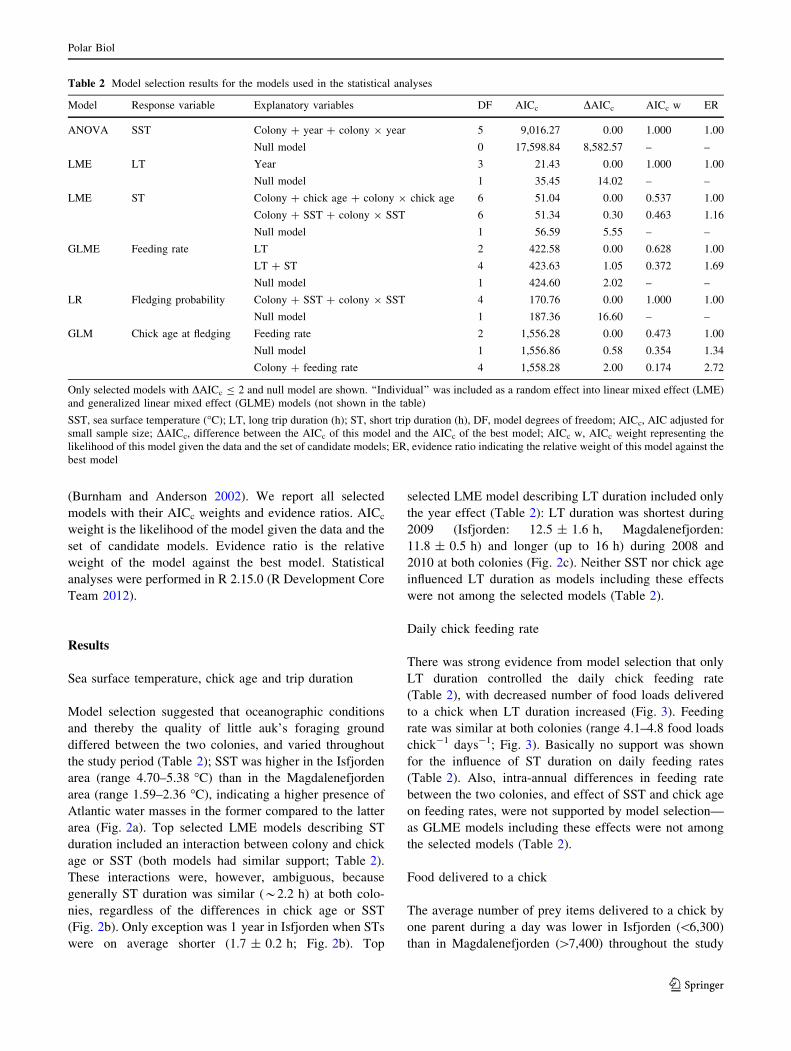
Model Response variable Explanatory variables DF AICc DAICc AICc w ER
ANOVA SST Colony ? year ? colony 9 year 5 9,016.27 0.00 1.000 1.00
Null model 0 17,598.84 8,582.57 – –
LME LT Year 3 21.43 0.00 1.000 1.00
Null model 1 35.45 14.02 – –
LME ST Colony ? chick age ? colony 9 chick age 6 51.04 0.00 0.537 1.00
Colony ? SST ? colony 9 SST 6 51.34 0.30 0.463 1.16
Null model 1 56.59 5.55 – –
GLME Feeding rate LT 2 422.58 0.00 0.628 1.00
LT ? ST 4 423.63 1.05 0.372 1.69
Null model 1 424.60 2.02 – –
LR Fledging probability Colony ? SST ? colony 9 SST 4 170.76 0.00 1.000 1.00
Null model 1 187.36 16.60 – –
GLM Chick age at fledging Feeding rate 2 1,556.28 0.00 0.473 1.00
Null model 1 1,556.86 0.58 0.354 1.34
Colony ? feeding rate 4 1,558.28 2.00 0.174 2.72
Only selected models with DAICc B 2 and null model are shown. ‘‘Individual’’ was included as a random effect into linear mixed effect (LME)
and generalized linear mixed effect (GLME) models (not shown in the table)
SST, sea surface temperature (�C); LT, long trip duration (h); ST, short trip duration (h), DF, model degrees of freedom; AICc, AIC adjusted for
small sample size; DAICc, difference between the AICc of this model and the AICc of the best model; AICc w, AICc weight representing the
likelihood of this model given the data and the set of candidate models; ER, evidence ratio indicating the relative weight of this model against the
best model
Polar Biol
123
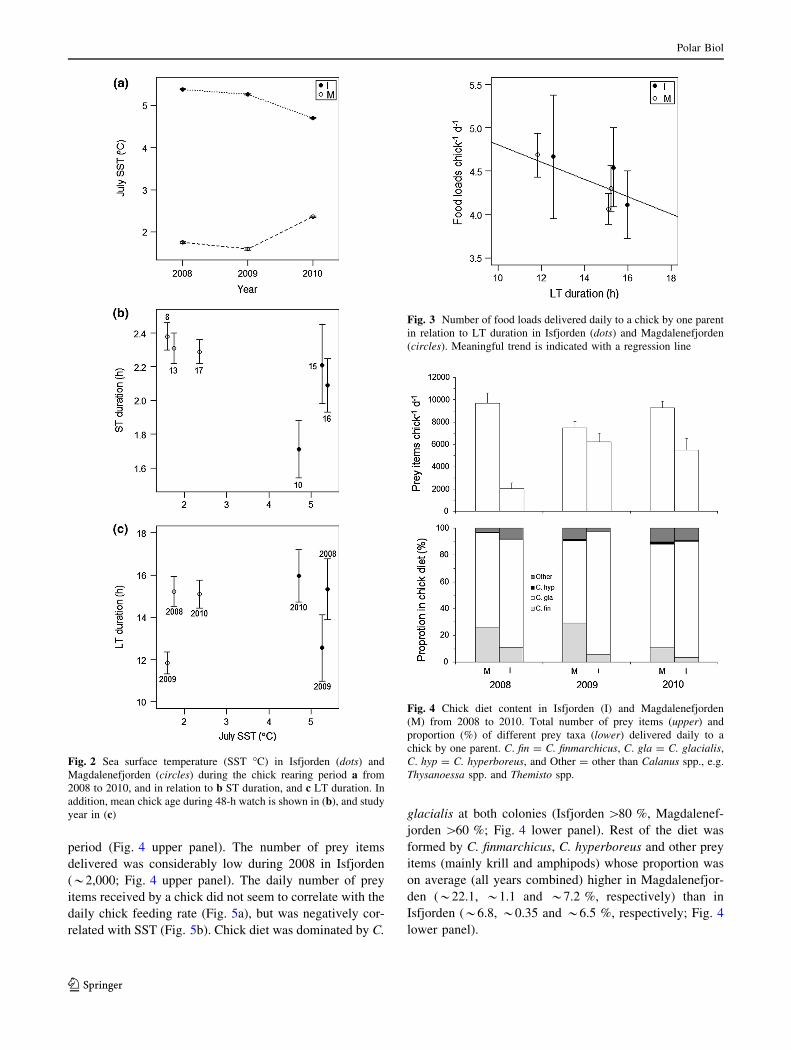
period (Fig. 4 upper panel). The number of prey items
delivered was considerably low during 2008 in Isfjorden
(*2,000; Fig. 4 upper panel). The daily number of prey
items received by a chick did not seem to correlate with the
daily chick feeding rate (Fig. 5a), but was negatively cor-
related with SST (Fig. 5b). Chick diet was dominated by C.
glacialis at both colonies (Isfjorden [80 %, Magdalenef-
jorden [60 %; Fig. 4 lower panel). Rest of the diet was
formed by C. finmarchicus, C. hyperboreus and other prey
items (mainly krill and amphipods) whose proportion was
on average (all years combined) higher in Magdalenefjor-
den (*22.1, *1.1 and *7.2 %, respectively) than in
Isfjorden (*6.8, *0.35 and *6.5 %, respectively; Fig. 4
lower panel).
Fig. 2 Sea surface temperature (SST �C) in Isfjorden (dots) and
Magdalenefjorden (circles) during the chick rearing period a from
2008 to 2010, and in relation to b ST duration, and c LT duration. In
addition, mean chick age during 48-h watch is shown in (b), and study
year in (c)
Fig. 3 Number of food loads delivered daily to a chick by one parent
in relation to LT duration in Isfjorden (dots) and Magdalenefjorden
(circles). Meaningful trend is indicated with a regression line
Fig. 4 Chick diet content in Isfjorden (I) and Magdalenefjorden
(M) from 2008 to 2010. Total number of prey items (upper) and
proportion (%) of different prey taxa (lower) delivered daily to a
chick by one parent. C. fin = C. finmarchicus, C. gla = C. glacialis,
C. hyp = C. hyperboreus, and Other = other than Calanus spp., e.g.
Thysanoessa spp. and Themisto spp.
Polar Biol
123
Chick fledging success and age at fledging
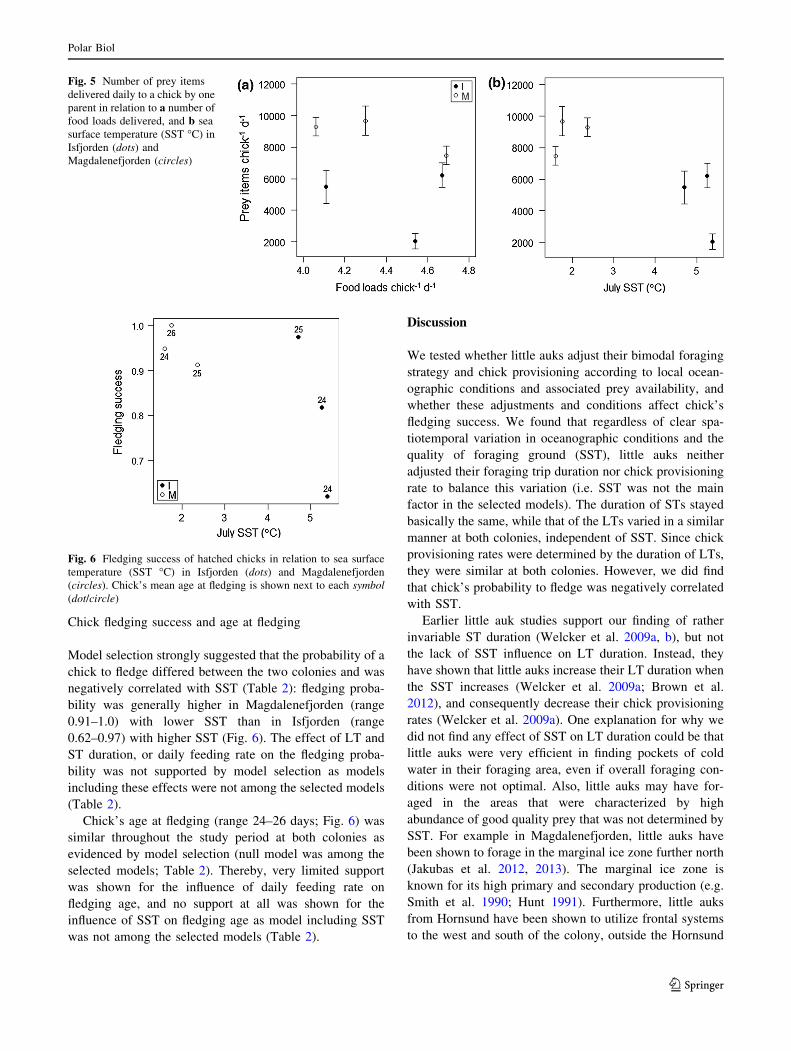
Model selection strongly suggested that the probability of a
chick to fledge differed between the two colonies and was
negatively correlated with SST (Table 2): fledging proba-
bility was generally higher in Magdalenefjorden (range
0.91–1.0) with lower SST than in Isfjorden (range
0.62–0.97) with higher SST (Fig. 6). The effect of LT and
ST duration, or daily feeding rate on the fledging proba-
bility was not supported by model selection as models
including these effects were not among the selected models
(Table 2).
Chick’s age at fledging (range 24–26 days; Fig. 6) was
similar throughout the study period at both colonies as
evidenced by model selection (null model was among the
selected models; Table 2). Thereby, very limited support
was shown for the influence of daily feeding rate on
fledging age, and no support at all was shown for the
influence of SST on fledging age as model including SST
was not among the selected models (Table 2).
Discussion
We tested whether little auks adjust their bimodal foraging
strategy and chick provisioning according to local ocean-
ographic conditions and associated prey availability, and
whether these adjustments and conditions affect chick’s
fledging success. We found that regardless of clear spa-
tiotemporal variation in oceanographic conditions and the
quality of foraging ground (SST), little auks neither
adjusted their foraging trip duration nor chick provisioning
rate to balance this variation (i.e. SST was not the main
factor in the selected models). The duration of STs stayed
basically the same, while that of the LTs varied in a similar
manner at both colonies, independent of SST. Since chick
provisioning rates were determined by the duration of LTs,
they were similar at both colonies. However, we did find
that chick’s probability to fledge was negatively correlated
with SST.
Earlier little auk studies support our finding of rather
invariable ST duration (Welcker et al. 2009a, b), but not
the lack of SST influence on LT duration. Instead, they
have shown that little auks increase their LT duration when
the SST increases (Welcker et al. 2009a; Brown et al.
2012), and consequently decrease their chick provisioning
rates (Welcker et al. 2009a). One explanation for why we
did not find any effect of SST on LT duration could be that
little auks were very efficient in finding pockets of cold
water in their foraging area, even if overall foraging con-
ditions were not optimal. Also, little auks may have for-
aged in the areas that were characterized by high
abundance of good quality prey that was not determined by
SST. For example in Magdalenefjorden, little auks have
been shown to forage in the marginal ice zone further north
(Jakubas et al. 2012, 2013). The marginal ice zone is
known for its high primary and secondary production (e.g.
Smith et al. 1990; Hunt 1991). Furthermore, little auks
from Hornsund have been shown to utilize frontal systems
to the west and south of the colony, outside the Hornsund
Fig. 5 Number of prey items
delivered daily to a chick by one
parent in relation to a number of
food loads delivered, and b sea
surface temperature (SST �C) in
Isfjorden (dots) and
Magdalenefjorden (circles)
Fig. 6 Fledging success of hatched chicks in relation to sea surface
temperature (SST �C) in Isfjorden (dots) and Magdalenefjorden
(circles). Chick’s mean age at fledging is shown next to each symbol
(dot/circle)
Polar Biol
123

fjord, despite the presence of Arctic water masses close to
the colony (Brown et al. 2012; Jakubas et al. 2013). Frontal
areas are often characterized by elevated stocks of phyto-
and zooplankton (Fauchald 2009). Thus, during LTs, little
auks from Magdalenefjorden may have foraged along the
marginal ice zone in the north, and little auks from Isf-
jorden may have sought frontal systems outside the fjord in
the shelf zone where these fronts normally occur (Saloranta
and Svendsen 2001; Walkusz et al. 2009). As the Isfjorden
colony is located deep inside the fjord (Fig. 1), the distance
to the fronts may have been comparable to the distance
from Magdalenefjorden colony to the marginal ice zone.
This could explain the similar duration of LTs at both
colonies. The shorter duration of LTs in 2009 may have
been due to the more southern location of the marginal ice
zone or more drift ice in the fjords (Ice Information Portal;
http://polarview.met.no) and closer location of the fronts.
Much shorter and less variable duration of STs likely
indicates that little auks collected food for their chicks
closer to the colonies than during LTs. This is further
supported by the content of chick meals: C. glacialis,
which is especially abundant on the shelf and fjord waters
of western and northern Spitsbergen (Blachowiak-Samolyk
et al. 2008; Søreide et al. 2008), dominated chick diet at
both colonies. The use of different foraging grounds during
LTs and STs has been previously suggested by Steen et al.
(2007), Brown et al. (2012) and Jakubas et al. (2012).
Furthermore, similar chick provisioning rates at both
colonies were likely due to similarity of distance to util-
izable foraging grounds during LTs—as LT durations,
which influenced the provisioning rates, were similar.
There were, however, differences in the number of prey
items delivered daily to a chick between the colonies. This
number did not correlate with the provisioning rates, but
seemed to be higher when SST was lower: Magdalenef-
jorden chicks received on average 4,200 more prey items
on a daily basis than Isfjorden chicks (all years combined).
This also implies that despite the higher percentage of C.
glacialis in the food portions delivered to chicks in Isf-
jorden, chicks there received generally less C. glacialis
than chicks in Magdalenefjorden. Also, the proportion of
other prey items (C. finmarchicus, C. hyperboreus, and krill
and amphipods) was higher in Magdalenefjorden. Thereby,
it is probable that foraging grounds used to collect food for
the chicks were offering more prey of high quality in the
Magdalenefjorden area than in the Isfjorden area.
Other seabird studies have shown that changes in the
chick provisioning rate and in the content of chick meals
can affect chick’s condition, survival and probability to
fledge (e.g. Schaffner 1990; Brekke and Gabrielsen 1994;
Golet et al. 2000; Davoren and Montevecchi 2003; Kadin
et al. 2012). In our case, we found that chick fledging
success was not affected by provisioning rates, but that
SST had a strong effect on fledging. This effect was likely
related to the prey availability and quality in the little auk’s
foraging grounds. In the Magdalenefjorden area where SST
was lower, chicks received generally more prey of good
quality than chicks in the Isfjorden area where SST was
higher—despite the similar delivery rate of chick meals at
the two colonies. During 1 year, however, fledging success
in the Isfjorden colony was at the similar level with that of
the Magdalenefjorden colony: In that 1 year, SST was
lower (4.70 �C) than during the other two years in the
Isfjorden area (Fig. 6), and Isfjorden chicks received more
good quality C. hyperboreus (biggest species with highest
lipid reserves among the three Calanus species; Scott et al.
2000), and krill and amphipods compared to these other
two years (Fig. 4). This may have increased the fledging
success. Furthermore, there were no differences in chick’s
fledging age. This is in contrast to the recent study by
Jakubas et al. (2013) who found that chicks fledged at an
older age when conditions in foraging grounds were not
optimal.
To conclude, the results of our study suggest that vari-
ation in foraging trips or provisioning rates was not related
to SST. This contrasts the previous studies that found a
negative relationship between SST, trip durations and
provisioning rates (Welcker et al. 2009a; Brown et al.
2012). Search efficiency and access to foraging grounds
known for elevated prey stocks (e.g. the marginal ice zone
and fronts) may have had a greater influence on trip
durations and provisioning rates. Also, little auks seemed
to prioritize self-maintenance over that of their chicks
when foraging conditions so demanded: little auks adjusted
the duration of LTs but not that of STs and provisioned
their chicks at lower rate when duration of LTs increased.
However, these foraging adjustments did not influence
chick’s probability to fledge. Instead, SST had a strong
effect on fledging probability, and this was likely related to
prey availability and quality in the little auk’s foraging
grounds. Our study is a first one to report such a detri-
mental effect of a change in foraging conditions on the
little auk chicks. It is very likely that predicted, warmer
climate in the Arctic (IPCC 2013) and consequential
changes in the little auk’s foraging areas (e.g. change in the
ratio of Arctic vs. Atlantic water masses and disappearance
of sea ice; Falk-Petersen et al. 2007; Stempniewicz et al.
2007) will negatively influence the ability of parents to
provide their chicks, and consequently, the fledging pros-
pects of little auk chicks even more in future.
Acknowledgments This work was supported by a grant from
Norway through the Norwegian Financial Mechanisms Project No.
PNRF-234-AI-1/07(ALKEKONGE) and conducted under the per-
mission of the Governor of Svalbard and Norwegian Animal Research
Authority. We thank Rafał Boehnke for analysing chick diet data
from Magdalenefjorden and Mateusz Barcikowski, Anika Beiersdorf,
Polar Biol
123

Eirik Grønningsæter, Aino Luukkonen, Atle Coward Markussen,
Adam Nawrot, Jan Samołyk, Lech Stempniewicz, Tobias Stal and
Mikko Vihtakari for their invaluable help in the field.
References
Barrett RT, Chapdelaine G, Anker-Nilssen T, Mosbech A, Montevecchi
WA, Reid JB, Veit RR (2006) Seabird numbers and prey
consumption in the North Atlantic. ICES J Mar Sci 63:1145–1158
Blachowiak-Samolyk K, Søreide JE, Kwasniewski S, Sundfjord A,
Hop H, Falk-Petersen S, Hegseth EN (2008) Hydrodynamic
control of mesozooplankton abundance and biomass in northern
Svalbard waters (79.81�N). Deep-Sea Res II 55:2210–2224
Brekke B, Gabrielsen GW (1994) Assimilation efficiency of adult
kittiwakes and Brunnich’s guillemots fed capelin and arctic cod.
Polar Biol 14:279–284
Brown ZW, Welcker J, Harding AMA, Walkusz W, Karnovsky NJ
(2012) Divergent diving behaviour during short and long trips of
a bimodal forager, the little auk Alle alle. J Avian Biol
43:215–226
Burnham KP, Anderson DR (2002) Model selection and multimodel
inference: a practical information-theoretic approach. Springer,
New York
Chaurand T, Weimerskirch H (1994) The regular alternation of short
and long foraging trips in the blue petrel Halobaena caerulea: a
previously undescribed strategy of food provisioning in a pelagic
seabird. J Anim Ecol 63:275–282
Congdon BC, Krockenberger AK, Smithers BV (2005) Dual-foraging
and coordinated provisioning in a tropical Procellariiform, the
wedge-tailed shearwater. Mar Ecol Prog Ser 301:293–301
Daase M, Eiane K (2007) Mesozooplankton distribution in northern
Svalbard waters in relation to hydrography. Polar Biol
30:969–981
Davoren GK, Montevecchi WA (2003) Consequences of foraging trip
duration on provisioning behaviour and fledging condition of
common murres Uria aalge. J Avian Biol 34:44–53
Duriez O, Weimerskirch H, Fritz H (2000) Regulation of chick
provisioning in the thin-billed prion: an interannual comparison
and manipulation of parents. Can J Zool 78:1275–1283
Falk-Petersen S, Pavlov V, Timofeev S, Sargent JR (2007) Climate
variability and possible effects on arctic food chains: the role of
Calanus. In: Ørbæk JB, Kallenborn R, Tombre I, Hegseth EN,
Falk-Petersen S, Hoel AH (eds) Arctic-Alpine ecosystems and
people in a changing environment. Springer, Berlin, pp 147–166
Fauchald P (2009) Spatial interaction between seabirds and prey:
review and synthesis. Mar Ecol Prog Ser 463(391):139–151
Golet GH, Kuletz KJ, Roby DD, Irons DB (2000) Adult prey choice
affects chick growth and reproductive success in pigeon
guillemots. Auk 117:82–91
Grandeiro JP, Nunes M, Silva MC, Furness RW (1998) Flexible
foraging strategy of Cory’s shearwater Calonectris diomedea,
during chick rearing period. Anim Behav 56:1169–1176
Gremillet D, Welcker J, Karnovsky NJ, Walkusz W, Hall ME, Fort J,
Brown ZW, Speakman JR, Harding AMA (2012) Little auks
buffer the impact of current Arctic climate change. Mar Ecol
Prog Ser 454:197–206
Harding AMA, van Pelt TI, Lifjeld JT, Mehlum F (2004) Sex
differences in little auk Alle alle parental care: transition from
biparental to paternal-only care. Ibis 146:642–651
Hirche HJ, Hagen W, Mumm N, Richter C (1994) The northeast
water polynya, Greenland Sea. III. Mesozooplankton and
makrozooplankton distribution and production of dominant
herbivorous copepods during spring. Polar Biol 14:491–503
Hunt GL (1991) Occurrence of polar seabirds at sea in relation to prey
concentrations and oceanographic factors. Polar Res 10:553–559
IPCC (2013) Summary for Policymakers. In: Stocker TF, Qin D,
Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia
Y, Bex V, Midgley PM (eds) Climate Change 2013: The
physical science basis. Contribution of Working Group I to the
fifth assessment report of the intergovernmental panel on climate
change, Cambridge University Press, Cambridge, UK and New
York, NY, USA
Jakubas D, Wojczulanis-Jakubas K, Walkusz W (2007) Response of
dovekie to changes in food availability. Waterbirds 30:421–428
Jakubas D, Gluchowska M, Wojczulanis-Jakubas K, Karnovsky NJ,
Keslinka L, Kidawa D, Walkusz W, Boehnke R, Cisek M,
Kwasniewski S, Stempniewicz L (2011) Foraging effort does not
influence body condition and stress level in little auks. Mar Ecol
Prog Ser 432:277–290
Jakubas D, Iliszko L, Wojczulanis-Jakubas K, Stempniewicz L (2012)
Foraging by little auks in the distant marginal sea ice zone during
the chick-rearing period. Polar Biol 35:73–81
Jakubas D, Trudnowska E, Wojczulanis-Jakubas K, Iliszko L, Kidawa
D, Darecki M, Blachowiak Samolyk K, Stempniewicz L (2013)
Foraging closer to the colony leads to faster growth in little auks.
Mar Ecol Prog Ser 489:263–278
Kadin M, Osterblom H, Hentati-Sundberg J, Olsson O (2012)
Contrasting effects of food quality and quantity on a marine
top predator. Mar Ecol Prog Ser 444:239–249
Karnovsky NJ, Kwasniewski S, Weslawski JM, Walkusz W, Bes-
zczynska-Moller A (2003) Foraging behaviour of little auks in a
heterogeneous environment. Mar Ecol Prog Ser 253:289–303
Karnovsky NJ, Harding AMA, Walkusz W, Kwasniewski S,
Goszczko I, Wiktor J Jr, Routti H, Bailey A, McFadden L,
Brown ZW, Beaugrand G, Gremillet D (2010) Foraging
distributions of little auks Alle alle across the Greenland Sea:
implications of present and future Arctic climate change. Mar
Ecol Prog Ser 415:283–293
Kwasniewski S, Gluchowska M, Jakubas D, Wojczulanis-Jakubas K,
Walkusz W, Karnovsky N, Blachowiak-Samolyk K, Cisek M
(2010) The impact of different hydrographic conditions and
zooplankton communities on provisioning little auks along the
West coast of Spitsbergen. Prog Oceanogr 87:72–82
Lack D (1968) Ecological adaptations for breeding in birds. Methuen,
London
Motoda S (1985) Devices of simple plankton apparatus—VII. Bull
Mar Sci 37:776–777
Nilsen F, Cottier F, Skogseth R, Mattsson S (2008) Fjord-shelf
exchanges controlled by ice and brine production: the interan-
nual variation of Atlantic Water in Isfjorden, Svalbard. Cont
Shelf Res 28:1838–1853
R Development Core Team (2012) R: a language and environment for
statistical computing. R Foundation for Statistical Computing,
Vienna
Ricklefs RE (1983) Some considerations on the reproductive
energetics of pelagic seabirds. Stud Avian Biol 8:84–94
Ropert-Coudert Y, Wilson RP, Daunt F, Kato A (2004) Patterns of
energy acquisition by a central place forager: benefits of
alternating short and long foraging trips. Behav Ecol 15:824–830
Saloranta TM, Svendsen H (2001) Across the Arctic front west of
Spitsbergen: high-resolution CTD sections from 1998–2000.
Polar Res 20:177–184
Schaffner FC (1990) Food provisioning by white-tailed tropicbird: effect
on the development pattern of the chick. Ecology 71:375–390
Scott CL, Kwasniewski S, Falk-Petersen S, Sargent JR (2000) Lipids
and life strategies of Calanus finmarchicus, Calanus glacialis
and Calanus hyperboreus in late autumn, Kongsfjorden, Sval-
bard. Polar Biol 23:510–516
Polar Biol
123

Smith SL, Smith WO, Codispoti LA, Wilson DL (1990) Biological
observations in the marginal ice zone of the east Greenland Sea.
J Mar Res 43:693–717
Søreide JE, Falk-Petersen S, Hegseth EN, Hop H, Carroll ML,
Hobson KA, Blachowiak-Samolyk K (2008) Seasonal feeding
strategies of Calanus in the high-Arctic Svalbard region. Deep-
Sea Res II 55:2225–2244
Stearns SC (1989) Trade-offs in life history evolution. Funct Ecol
3:259–268
Steen H, Vogedes D, Broms F, Falk-Petersen S, Berge J (2007) Little
auks (Alle alle) breeding in a High Arctic fjord system: bimodal
foraging strategies as a response to poor food quality? Polar Res
26:118–125
Stempniewicz L (1981) Breeding biology of the little auk Plautus alle
in the Hornsund region, Spitsbergen. Acta Ornithol 18:1–26
Stempniewicz L (2001) Alle alle little auk. The Journal of the Birds of
Western Palearctic, BWP Update, vol 3. Oxford University
Press, Oxford, pp 175–201
Stempniewicz L, Blachowiak-Samolyk K, Weslawski JM (2007)
Impact of climate change on zooplankton communities, seabird
populations and arctic terrestrial ecosystem—A scenario. Deep-
Sea Res II 54:2934–2945
Stempniewicz L, Darecki M, Trudnowska E, Blachowiak-Samolyk K,
Boehnke R, Jakubas D, Keslinka-Nawrot L, Kidawa D, Sagan S,
Wojczulanis-Jakubas K (2013) Visual prey availability and
distribution of foraging little auks (Alle alle) in the shelf waters
of West Spitsbergen. Polar Biol 36:949–955
Trudnowska E, Szczucka J, Hoppe L, Boehnke R, Hop H,
Blachowiak-Samolyk K (2012) Multidimensional zooplankton
observations on the northern West Spitsbergen Shelf. J Mar Syst
98–99:18–25
Walczowski W, Piechura J, Goszczko I, Wieczorek P (2012) Changes
in Atlantic water properties: an important factor in the European
Arctic marine climate. ICES J Mar Sci 69:864–869
Walkusz W, Kwasniewski S, Falk-Petersen S, Hop H, Tverberg V,
Wieczorek P, Weslawski JM (2009) Seasonal and spatial
changes in the zooplankton community of Kongsfjorden, Sval-
bard. Polar Res 28:254–281
Weimerskirch H, Chastel O, Ackermann L, Chaurand T, Cuenot-
Chaillet F, Hindermeyer X, Judas J (1994) Alternate long and
short foraging trips in pelagic seabird parents. Anim Behav
47:472–476
Weimerskirch H, Chastel O, Cherel Y, Henden J-A, Tveraa T (2001)
Nest attendance and foraging movements of northern fulmars
rearing chicks at Bjørnøya Barents Sea. Polar Biol 24:83–88
Weimerskirch H, Ancel A, Caloin M, Zahariev A, Spagiari J, Kersten
M, Chastel O (2003) Foraging efficiency and adjustment of
energy expenditure in a pelagic seabird provisioning its chick.
Anim Behav 72:500–508
Welcker J, Harding AMA, Karnovsky NJ, Steen H, Strøm H,
Gabrielsen GW (2009a) Flexibility in the bimodal foraging
strategy of a high Arctic alcid, the little auk Alle alle. J Avian
Biol 40:388–399
Welcker J, Steen H, Harding AMA, Gabrielsen GW (2009b) Sex-
specific provisioning behaviour in a monomorphic seabird with a
bimodal foraging strategy. Ibis 151:502–513
Welcker J, Beiersdorf A, Varpe Ø, Steen H (2012) Mass fluctuations
suggest different functions of bimodal foraging trips in a central-
place forager. Behav Ecol 23:1372–1378
Weslawski JM, Koszteyn J, Kwasniewski S, Stempniewicz L,
Malinga M (1999) Summer food resources of the little auk, Alle
alle (L.) in the European Arctic seas. Pol Polar Res 20:387–403
Weydmann A, Kwasniewski S (2008) Distribution of Calanus
populations in a glaciated fjord in the Arctic (Hornsund,
Spitsbergen)—the interplay between biological and physical
factors. Polar Biol 31:1023–1035
Willis K, Cottier F, Kwasniewski S, Wold A, Falk-Petersen S (2006)
The influence of advection on zooplankton community compo-
sition in an Arctic fjord (Kongsfjorden, Svalbard). J Mar Syst
61:39–54
Wojczulanis-Jakubas K, Jakubas D (2012) When and why does my
mother leave me? The question of brood desertion in the
Dovekie (Alle alle). Auk 129:632–637
Wojczulanis-Jakubas K, Jakubas D, Karnovsky NJ, Walkusz W
(2010) Foraging strategy of little auks under divergent conditions
on feeding grounds. Polar Res 29:22–29
Zera AJ, Harshman LG (2001) The physiology of life history trade-
offs in animals. Annu Rev Ecol Syst 32:95–126
Polar Biol
123
Copyright © 2022 FDOKUMEN




















