
Süreli Yayın Tasarımı için Temel Tipografi Bilgisi (Ders Notları, 2004)

HAYVAN FİZYOLOJİSİ
Version 0.5
DR UTKU GÜNER
Ders Notları

T RAKYA UN İVERS İ TES İ
Fen Fakültesi Biyoloji Bölümü
Kod adı: AŞURE 3+3 Vertex site
Trakya Üniversitesi Fen fakültesi
Biyoloji Bölümü Mayıs 2014 DERLEME
Power point sunumları için
https://yadi.sk/i/tpOjhN3_ceGui adresine gidin.

1
İ ç i n d e k i l e r
Temel Fizyolojik Yasalar ................................................................ 1
Termodinamik yasalar ..................................................................... 1
Avogadro hipotezi ............................................................................ 2
Boyle yasası .................................................................................... 4
Herry yasası .................................................................................... 5
Laplace yasası ................................................................................ 6
Fick Yasası ...................................................................................... 6
Frank-Starling hipotezi..................................................................... 7
Hücre Fizyolojisi ............................................................................... 8
Hücre ............................................................................................... 8
Hücrenin inorganik bileşikleri ......................................................... 10
Hücrede su .................................................................................... 10
Suyun fonksiyonu .......................................................................... 12
Hücrenin organik molekülleri ........................................................ 13
Proteinler ....................................................................................... 14
Proteinlerde Üç Boyutlu Yapısı ...................................................... 17
Karbohidratlar ................................................................................ 21
Glikozaminoglikanlar ..................................................................... 25
Lipidler ........................................................................................... 25
Enzimler ........................................................................................ 28
Vitaminler ...................................................................................... 31
Hücre Zarı .......................................................................................33
Fosfolipid yapısı ............................................................................ 34
Hücre membranın görevleri ........................................................... 36
Membran Proteinleri ...................................................................... 38
Membran proteinlerinin görevleri: .................................................. 41
Membran Karbohidratları ............................................................... 41
Kolesterol ...................................................................................... 43
Kolesterol İşlevi ............................................................................. 45
Membran bağlantıları..................................................................... 45

3
Bazal lamina .................................................................................. 48
Adhezyon Molekülleri: ................................................................... 49
Hücre dışı matriks(Ekstrasellüler matriks) ..................................... 51
Fosfolipidlerin geçirgenliği ............................................................. 52
Basit Difüzyon ............................................................................... 54
Filtrasyon ....................................................................................... 55
Taşıma proteinleri ile Kolaylaştırılmış diffüzyon ............................. 56
Kolaylaştırılmış difüzyon GLUT glikoz transporterları .................... 58
Kolaylaştırılmış difüzyon Aquaporin su kanal proteinleri ............... 58
Aktif- pasif Taşıma ......................................................................... 59
Simport-Antiport aktif taşıma yolları ............................................... 62
Primer aktif taşıma Na-K ATPaz .................................................... 63
Primer aktif taşıma Ca ATPaz ....................................................... 64
Sekonder Aktif Taşıma Na -Glikoz ve Amino asit koportu ............. 65
Ekzositoz ....................................................................................... 66
Endositoz ....................................................................................... 67
Membran reseptörleri .................................................................... 69
Sitoplazma ......................................................................................70
Sıvı kısım sitozol ........................................................................... 70
Endo plazmik Retikulum ................................................................ 71
Düz E.R. işlevi ............................................................................... 71
Granüllü ER İşlevi .......................................................................... 73
Ribozom ........................................................................................ 73
Golgi Aygıtı .................................................................................... 75
Lizozom ......................................................................................... 76
Lizozom membran hasarı .............................................................. 77
Mitokondri ve Enerji Üretimi ......................................................... 80
Kemoosmotik Atp sentezi ............................................................. 81
Hekzos Monofosfat Yolu (Pentoz Fosfat Yolu) .............................. 82
Hücre iskeleti Görevi ..................................................................... 82
İntermedial Filamentler .................................................................. 84
Mikrofilamentler ............................................................................. 85
Mikrotübüller .................................................................................. 85
Mikrotüpüllerin başlıca görevleri : .................................................. 86
Sentrozom ..................................................................................... 86
Kamçı ve Siler ............................................................................... 87
Hücre iskeleti ve hareket ............................................................... 87
Çekirdek (Nükleous) .......................................................................88
Sinir sistemi ....................................................................................91
Nöron ............................................................................................. 92

4
Gibbs Donan Dengesi ................................................................... 95
Refleks Olayı ................................................................................. 96
Sinir iletisi ...................................................................................... 97
Membran Dinlenme potansiyeli ..................................................... 98
İyon Kanalları ................................................................................ 98
Aksiyon Potansiyeli ..................................................................... 102
Depolarizasyon-Hiperpolarizasyon .............................................. 104
İmpulsun sinir boyunca yayılması ................................................ 104
Mutlak refrakter periyod: .............................................................. 106
Relatif (Nisbi) refrakter periyod: ................................................... 106
Reobaz (Eşik değer) ................................................................... 108
Ekdize Edici Post Sinaptik Potansiyel (EPSP) İnhibe edici post sinaptik potansiyel (IPSP): ........................................................................ 112
Sumasyon: .................................................................................. 116
Nörotransmiter maddeler ............................................................. 117
Asetilkolin .................................................................................... 120
Histamin ...................................................................................... 121
Dopamin: ..................................................................................... 121
Glutamik asit ve aspartik asit ....................................................... 122
MAO (Mono amino oksitaz) ......................................................... 123
COMT (Katekol-O-Metil Transferaz): ........................................... 123
Serotonin ..................................................................................... 124
Sinir sistemi .................................................................................. 125
Sinir sistemin evrimsel gelişimi .................................................... 125
Sinir Organizasyonu .................................................................... 129
Omurilik sinirleri ........................................................................... 132
Beyinden çıkan sinirler ................................................................ 133
Beyin Kısımları ............................................................................ 134
Retiküler Aktive edici sistem (Retiküler Formasyon) ................... 136
Bazal Ganglion ............................................................................ 137
Limbik sistem ............................................................................... 137
Hipotalamus ................................................................................ 138
Talamus ....................................................................................... 139
Sinir sistemi organizasyon ........................................................... 140
Sempatik Sinir Sistemi ................................................................. 141
Parasempatik Sistem................................................................... 143
Kolinerjik Ve Adrenerjik Etkiler .................................................... 144
Cerebellum (Küçük Beyin) Fonksiyonlari ..................................... 145
Büyük Beyin (Beyin korteksi) (Cortex cerebri) ............................. 146
Tek Hücrelilerde hareket ............................................................. 147

5
Çok hücrelilerde hareket .............................................................. 148
Kas ............................................................................................... 150
Kas tipileri .................................................................................... 150
Kas Yapısını Oluşturan Proteinler ............................................... 153
Aktin ............................................................................................ 154
Tropomiyozin: .............................................................................. 155
Troponin : .................................................................................... 155
Miyozin : ...................................................................................... 155
Sarkomer Yapısı .......................................................................... 156
Motor Ünite .................................................................................. 157
Nöromüsküler bağlantı (kavşak, uç plak) ..................................... 158
Kas Kasılmasi .............................................................................. 161
İskelet Kasında Gevşeme ............................................................ 163
Tetani .......................................................................................... 164
Merdiven Treppe Olayı ................................................................ 165
İskelet kası tipleri ......................................................................... 165
İskelet Kasında Enerji .................................................................. 166
Alaktik Anaerobik Sistem (Fosfatojen = ATP- Keratin fosfat): ..... 168
Glikoliz ......................................................................................... 170
Kasılma Tipleri ............................................................................. 171
Rigor Mortis ................................................................................. 171
Kalp kası ...................................................................................... 172
Düz Kas ....................................................................................... 175
Mandal Mekanizması................................................................... 175
Kalsiyumu Bağlıyan Protein -Kalmodulin ..................................... 179
Düz Kas Tipleri ............................................................................ 180
Viseral Düz Kas (Tek Birimli Düz Kas) ........................................ 180
Multiunit Düz Kas ........................................................................ 180
Reseptörler ................................................................................... 182
Duyu Organları ve Reseptörler .................................................... 184
Duyu Organı Sınıflaması ............................................................. 185
Deride sıcaklık algılanması .......................................................... 189
İki-Nokta Ayırımı .......................................................................... 190
Reseptörlerdeki Elektriksel ve İyonik Olaylar ............................... 190
Jeneratör Potansiyeller ................................................................ 190
Fotoreseptörler ............................................................................ 190
Omurgalı Göz Yapısı ................................................................... 191
Görme Olayı ................................................................................ 192
Görmenin Kimyası ....................................................................... 193
Çubuk Hücreleri ........................................................................... 195

6
Retina .......................................................................................... 195
Koni Hücreleri .............................................................................. 196
Bipolar Hücreler ........................................................................... 196
Ganglion Hücreleri ....................................................................... 196
Fotoreseptörler uyarı oluşumu ve bunun işlenmesi ..................... 197
Görme sinirleri ............................................................................. 198
Gece Körlüğü .............................................................................. 198
Koku ............................................................................................ 199
Tad duygusu ................................................................................ 201
İşitme Duyusu ve Denge ............................................................. 201
Tüy Hücreleri ............................................................................... 205
Ses özelikleri ............................................................................... 206
Kulak Zarı ve Kemiklerin İşlevleri ................................................ 207
İşitme Sinir Liflerindeki Aksiyon Potansiyelleri ............................. 208
Odiyometre .................................................................................. 210
Vestibüler Fonksiyon .................................................................. 210
Vestibular Apparat Krista Ampullaris ve Dinamik Denge ............. 211
Nistagmus ................................................................................... 211
Utrikular Reseptör Hücreleri Üzerine Yerçekiminin Etkisi ............ 212
Kalp ve damarlar ........................................................................... 213
Böcekte dolaşım sistemi .............................................................. 214
İnsanda Kalp ............................................................................... 215
Kalp Sesleri ................................................................................. 218
Kalp ileti sistemi ........................................................................... 219
Elektrokardiyogram (EKG) ........................................................... 227
Kalp atımı etkileyen faktörler ....................................................... 229
Stannius bağları .......................................................................... 231
Kan basınçı ................................................................................. 232
Dolaşım sistemi ........................................................................... 234
Sistemik Dolaşım ......................................................................... 235
Pulmonar dolaşım ....................................................................... 236
Kan damarları .............................................................................. 237
Kapiller kan damarları.................................................................. 238
Kapillerde madde taşıma yolları .................................................. 239
Kapillerin Fonksiyonları: .............................................................. 240
Kan akımı düzenlenmesi ............................................................. 241
Akut kontrol ................................................................................. 241
Kan Akımının Uzun Süreli Kontrolü ............................................. 243
İyonlar ve Diğer Kimyasal Faktörlerle Vasküler Kontrol ............... 247
Bazı Özgül Dokularda Kan Akımının Özel Kontrolü ve Endotel ... 247

7
Kapiller membrandan sıvı geçişi: Starling güçleri; ....................... 250
Lenf sistemi ................................................................................. 251
Kan Fizyolojisi ............................................................................... 254
Plazma Proteinleri ....................................................................... 254
Plazma Lipidleri ........................................................................... 258
Plazma Karbonhidratları .............................................................. 258
Plazmada Bulunan İnorganik Maddeler ....................................... 259
Kan hücreleri ............................................................................... 259
Eritrositler(akyuvarlar) ................................................................. 259
Eritrosit membran proteinleri ....................................................... 261
Hemoglobin Yapımı ..................................................................... 262
Demir Metabolizması ................................................................... 264
Alyuvarların Yıkımı ...................................................................... 266
Anemi .......................................................................................... 268
Polisitemi ..................................................................................... 269
Kan Grupları ................................................................................ 269
Eritroblastosis fetalis.................................................................... 270
Trombositler ................................................................................ 271
Kanın Pıhtılşaması(hemoztaz) .................................................... 273
Ekstrensek Yol ............................................................................ 277
İntrensek Yol ............................................................................... 277
Pıhtılaşma Faktörleri.................................................................... 277
Pıhtılaşmanın Sınırlandırılması ................................................... 278
Kan Pıhtısının Erimesi ................................................................. 280
Vitamin K eksikliği ....................................................................... 280
Hemofili ....................................................................................... 280
Tromboembolik Olaylar ............................................................... 281
Lökositler-Akyuvarlar Beyaz Kan Hücreleri ................................. 281
Nötrofil ......................................................................................... 281
Eozinofil ....................................................................................... 283
Bazofil .......................................................................................... 283
Lökositoz-lökopeni ....................................................................... 285
Lösemi ......................................................................................... 286
Vücudun Enfeksiyonlara Direnci .................................................. 286
Doğuştan gelen bağışıklık ........................................................... 287
Kazanılan bağışıklık .................................................................... 287
Aktif ve pasif bağışıklık ................................................................ 288
Antijen ve antikorlar ..................................................................... 289
Komplement sistem ..................................................................... 291
Toll-benzeri reseptörler (Toll-like receptors-TLR) ........................ 293

8
Hapten ......................................................................................... 293
Aşırı duyarlılık reaksiyonları ........................................................ 294
Tip I Hipersensitivite .................................................................... 294
Tip II Aşiri Duyarlilik ..................................................................... 296
Tip III Aşiri Duyarlilik .................................................................... 296
Tip IV Aşiri Duyarlilik.................................................................... 297
Püy(irin oluşumu) ........................................................................ 299
Sindirim Fizyolojisi ........................................................................ 300
Farklı besinlere göre sindrim sistemi ........................................... 301
Geviş getiren ve getirmeyen Otçullarda sindirim sistemi ............. 302
Sindirim kanalının yapısı ............................................................. 303
Müküs salgısı .............................................................................. 305
Tükrük Salgısı ............................................................................. 305
Özofagus Salgısı ......................................................................... 306
Mide............................................................................................. 306
Mide salgı hücreleri ..................................................................... 308
Pankreas ..................................................................................... 310
Gastroenteroendokrin Hormonlar ................................................ 311
Enterohepatik dolaşım ................................................................. 313
İnce Barsak ................................................................................. 314
Kalın Barsak ................................................................................ 315
Sindirim kanalındaki başlıca Sfinkterler ....................................... 316
Besinlerin Kimyasal sindirimi ......................................................... 317
Protein sindirimi ........................................................................... 317
Karbonhidrat sindirimi .................................................................. 321
Yağların sindirimi ......................................................................... 325
Karaciğer ..................................................................................... 326
Safra tuzlarının ............................................................................ 332
Homestazi ..................................................................................... 333
Geribildirim .................................................................................. 335
Refleks ve yerel homeostatik yanıtlar .......................................... 336
Isı dengesi ................................................................................... 337
Canlıların çevreye osmotik durumuna uyumu ............................. 343
Azotlu atık maddeler .................................................................... 345
Üre sentezi .................................................................................. 345
Ürik asit ....................................................................................... 347
Boşaltım organları ....................................................................... 348
Balıklarda Osmoregülasyon ........................................................ 349
Kuşlarda ozmoregülasyon: .......................................................... 351
Sıvı-Elektrolit Dengesi ................................................................. 352

9
İntrasellüler sıvı ........................................................................... 353
Extrasellüler sıvı .......................................................................... 354
Sıvı değişiminde etkili faktörler .................................................... 354
Osmotik Basınç ........................................................................... 355
Sodyum ve Potasyum.................................................................. 356
Asit-Baz dengesi ......................................................................... 356
İnsanda Azotlu atıklar(Üre, Ürik asit, Kreatin) .............................. 357
Amonyağın Taşınması................................................................. 358
Ekstrasellür ve İntrasellüer Tamponlar ........................................ 358
Boşaltım sistemi Böbrekler ............................................................ 360
Böbrek Yapısı .............................................................................. 361
Böbreğin Kanlanması .................................................................. 362
Nefron tipleri ................................................................................ 363
Nefron yapısı ............................................................................... 363
Filtrasyon yarıkları ve Podositlerin uzantıları ............................... 365
Bowman kapsülünde basınç bağlı filtrasyon ................................ 366
Glomerül filtrasyon Oranı ayarlanması ........................................ 367
Mezangiyal hücreler .................................................................... 369
Böbreklerde Geri emilim .............................................................. 370
Vesa recta ................................................................................... 374
Jukstaglomerüler Apparat ve Hormonal kontrol ........................... 374
Renin ........................................................................................... 376
ANP ............................................................................................. 378
ADH ............................................................................................. 379
İdrar oluşumu .............................................................................. 381
Boşaltım sistemi diğer kısımları İdrar Torbası ve İdrar Yolları ..... 382
Hemodiyaliz ................................................................................. 383
Solunum Fizyolojisi ....................................................................... 384
Direkt ve indirekt solunum ........................................................... 385
Trake solunumu ........................................................................... 387
Deri solunumu ............................................................................. 388
Solungaç solunumu ..................................................................... 388
Hava kesesi ve akçiğer solunumu ............................................... 389
Ters Akım Sistemi: ...................................................................... 391
Kuşlarda solunum ........................................................................ 392
İnsanda Solunum Sistemini ......................................................... 394
Soluk alıp verme .......................................................................... 395
Alveollerindeki yüzey gerilimi ...................................................... 396
Solunum pigmentleri .................................................................... 396
Akciğer ve Dokularda Gaz Değişimi ............................................ 397

10
Alveoller ....................................................................................... 400
Oksijenin taşınması ..................................................................... 401
Karbondioksit taşınması .............................................................. 403
Respiratuvar değişim oranı (RER, R) .......................................... 404
Solunum hızı kontrolü .................................................................. 404
Akciğer Hacim ve kapasiteleri ..................................................... 407
Metabolizma ................................................................................ 410
Su Altı (Dalma) Fizyolojisi ............................................................ 411
Şnorkel ile Solunum..................................................................... 412
Hücre haberleşme yolları .............................................................. 413
G-Proteinine Bağlı Reseptörleri ................................................... 418
Tirozin Kinaz Reseptörleri ........................................................... 421
İyon Kanalı Reseptörleri .............................................................. 422
İkincil haberciler ........................................................................... 423
IP3 ikincil haberci yolu ................................................................. 424
DAG ikincil haberci yolu ............................................................... 425
Kalsiyum ve kalmodülin ............................................................... 427
Protein Fosforilasyonu ................................................................. 429
Hücre İçi Reseptörler ................................................................... 430
Hormon ........................................................................................ 431
Hormon reseptörleri ..................................................................... 437
Hormon Etki Mekanizmaları ........................................................ 438
Endokrin Bezler ........................................................................... 440
Hipotamus ................................................................................... 442
Hipofiz Hipotalamus İlişkisi .......................................................... 442
Hipotalamus hormonları .............................................................. 443
Hipofiz hormonları(trofik hormonlar) ............................................ 443
Antidiüretik Hormon (ADH) .......................................................... 445
Oksitosin ...................................................................................... 445
Ön hipofiz hormonları .................................................................. 446
Pro-opiyomelanokortin ailesi hormonlar ...................................... 446
Glikoprotein ailesi hormonlar ....................................................... 447
Somatomammotropin ailesi hormonları ....................................... 448
Opiyomelanokortin ailesi ön hipofiz hormonları (POMC) ............. 449
Pineal bez (Epifiz) hormonu (melatonin) ...................................... 449
Böbrek hormonları (Eritropoietin, 1,25 kalsiferol) ........................ 450
Timus bezi hormonları ................................................................. 451
Kalpten salgılanan ANP hormunu ............................................... 451
Plasenta hormonları Koryonik gonadotropin (hCG) ..................... 452
Tiroid Bezi hormonları ................................................................. 453

11
Kalsitonin ..................................................................................... 455
Paratiroid Bezi hormonları ........................................................... 456
Böbrek Üstü Bezleri (Adrenal Bezler) hormonları ........................ 457
Glukokortikoidler .......................................................................... 459
Mineralokortikoid ......................................................................... 459
Gonadokortikoidler ...................................................................... 459
Pankreas hormonları ................................................................... 459
Glukagon ..................................................................................... 460
İnsülin .......................................................................................... 461
Diabetes Mellitus ......................................................................... 464
Gonadlar: Cinsiyet bezleri ........................................................... 464
Diğer Dokulardan salgılanan Hormonlar ...................................... 468
Kaynaklar ...................................................................................... 469
Indeks ........................................................................................... 471

H A Y V A N F İ Z Y O L O J İ S İ
1
Temel Fizyoloji Yasaları ve
Hücre Fizyolojisi
Temel Fizik ve kimya yasalarının bir kısmı fizyolojik önemi bakınundan incelenmiştir. Hücre fizyolojisinde özellikle hücre membrane yapı ve işlevi ile organeller hedeflenmiştir.
Temel Fizyolojik Yasalar
Termodinamik yasalar
1. yasa: evrendeki enerjiler yok olmaz sadece birbirine veya işe dönüşür. Evrendeki toplam enerji sabittir (istisnai olarak nükleer reaksiyonlarda kütle enerjiye dönüşür, ama evrendeki toplam kütleyi de enerji cinsinden kabul edersek, toplam enerji yine de sabit olur).
ŞEKİL 1.1 Termodimaniğin birinci yasası enerji ve entropi
Canlı sistemler enerjilerini temel olarak ya diğer canlılardan, güneşten yada kimyasal bağ enerjisinden elde eder. Enerji yok olmaması yalnızca farklı formlara dönüşmesi
Bölüm
1
Kalori (cal) sıcaklığı artırmak için gerekli enerjidir olarak tanımlanır. 1 gram suyu 14.5°C dereceden 15.5°C. yükseltmek için gereken enerjisidir. Kalorinin 1000 katı kilokalori(kcal) olarak tanımlanır. Diğer bir enerji birimi ise joule (J) dür. 1000 joule 1 kilo joule eşittir. 1 Kcal 4.184 kJ eşittir.
Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
2
ekositemde canlı metabolizmasında önemlidir. Tüm enerji döngüleri sırasında bir miktar enerji kaybolur yani entropi(düzensizlik) artar.
2. yasa: Hiçbir cihaz veya sistem aldığı ısıyı tamamen işe dönüştürecek şekilde çalışamaz(%100 verim olmaz). Ayrıca sadece ısıyı bir sıcaklıktan daha yüksek bir sıcaklığa transfer eden bir süreç mümkün değildir. Canlı sistemlerde enerji dönüşümü mekanik sistemlerden daha yüksek verimde olmasına rahmen hiçbir zaman yüzde yüze seviyesine gelemez. "Hiçbir enerji akışı, düzensizlikten düzene doğru olamaz." veya "Bütün sistemler, evrenin yapısı gereği düzensizliğe doğru gitmeye mahkûmdur." veya "Yapılar, her zaman düzenli bir halden, düzensiz bir hale doğru ilerlerler."Biyolojik yapılarda enerji dönüşümleri insan yapımı sistemlere daha yüksektir. En yüksek verimli enerji dönüşümü biyolümünasta gözlenir. Ateşböceği lüsiferin adlı maddenin ATP ile fosforilasyonu sonucu bir parıltı meydana getirir. Kimyasal enerji böylece ışık enerjisine dönüşür ve böcekler arasında iletişimi sağlayan ışık parlamaları meydana gelir. Az bir enerjide ısı olarak kaybolur(verim %98). 3. yasa: mükemmel kristallenmiş bütün maddelerin mutlak sıcaklıktaki (0 kelvin) entropileri sıfırdır. Entropi hesabının temelini oluşturan yasadır. Sıfır Kelvin üzerindeki tüm moleküllerin belirli bir kinetik enerjileri vardır. Bu enerji canlı sistemlerindeki kendi kendine olan enerji kullanılmayan tüm olay için gereklidir. 0. yasa (evet sıfırıncı): Eğer iki cisim üçüncü bir cisim ile ısısal denge halinde ise birbirleriyle de ısısal dengededir ve bu nedenle aynı sıcaklıktadır.
ŞEKİL 1.2 Termodimaniğin sıfıncı yasası ısısal denge
Sıcaklık ölçümünün temeli bu yasadır. Sıfırıncı olmasının sebebi ise diğer yasalardan sonra çıkmış olması ama diğer yasaların anlaşılabilmesi için gerekli olmasıdır. Canlı sistemdeki ısı dengesinde 0. yasa kullanılır. Canlılar ısı koruması için yada ısının diğer ortamlara transferinde bu yasa önemlidir. Avogadro hipotezi
Bütün maddeler; demir, kalsiyum ya da oksijen gibi saf kimyasal maddeler, kimyasal elementlerden oluşur. Bir element kimyasal olarak daha küçük parçalara ayrılamayan maddedir.
ŞEKİL 1.3 Avargadro hipotezi

H A Y V A N F İ Z Y O L O J İ S İ
3
Maddeler normal şartlarda gaz, katı yada sıvı halde olabilir. Vücudun yaklaşık % 98'i oksijen, karbon, hidrojen, nitrojen, kalsiyum ve fosfor olmak üzere sadece altı elementten meydana gelmiştir. Madde moleküler farklı atom ağırlığa sahip olmalarına rahmen molar olarak eşitlerse aynı sayıda atom içeririrler. Aynı sıcaklık, basınç ve hacim altında gazlar, aynı sayıda molekül içerirler. Buna göre, belirli bir hacimdeki gazın bulundurduğu molekül sayısı, gazın kütle ya da boyutundan bağımsızdır. Örnek olarak, aynı hacimdeki hidrojen ve nitrojen verilebilir. Buna göre, hidrojen de nitrojen de, aynı hacim, aynı basınç ve aynı sıcaklıkta aynı molekül sayısına sahiptir. Bu durum solunda alınan havaın birleşenlerin dağılımında önemlidir. Soluk havasında bulunan gazların dağılımında basınç ve sıcaklık önemli rol oynar buna karşın molekül ağırlıları gaz karşımda gazların dağılımı değiştirmez. Bu noktada gazların sıvılar içinde erimelerinde diğer faktörlerinde rolü olduğu unutmamalıdır. Avagadro kanunu matematiksel olarak şöyle gösterilebilir: V \ n= a. V kübik metre olarak hacim, n gazın mol sayısı, a da bir sabittir. Bu denklem, sadece homojen maddeler(tek bileşenli) için geçerlidir. Buna homojen sıvılar ve katılar da dahildir. Avogadro yasasının en önemli sonucu, yasanın ideal gaz sabitinin tüm gazlar için aynı olduğunu bulmasıdır.
P paskal olarak basınç, T de Kelvin olarak sıcaklıktır. Denkleme göre, tüm gazlar için bu sabit eşittir. Yani gazın boyutunun ya da kütlesinin bu sabitin değerini değiştirmez. Standart durumda, bir mol ideal gaz, 22.4 litre (dm3) yer kaplar. Bu değer, genellikle molar hacim olarak kullanılır. Bir moldeki molekül sayısı olan Avogadro sayısı, yaklaşık olarak mol başına 6.02×1023 parçadır. Avogadro yasası, toplam gaz yasasını oluşturan Boyle yasası, Charles yasası ve Gay-Lussac yasasıyla birlikte ideal gaz yasasını oluşturur. Farklı gazların karışımı olan havadaki her bir molekül farklı atom ağırlığına sahip olmasına karşın aynı hacim ve basınçta eşit sayıda molekülden oluşmuştur. Sıcaklığın ve basınçın değişmesi hava karışımdaki moleküllerin sayısını etkiler. Bu durum dalış fizyolojisi için önemlidir. Molekül ağırlığı, bir maddenin molekülünün yapısına katılan tüm atomların ağırlıklarının toplamıdır. Örneğin suyun (H2O) molekül ağırlığı; 2 x 1,008 + 16,00 = 18,016’dir.Molekül ağırlığının gram cinsinden ifadesi mol olarak tanımlanır.
Örneğin 1 mol (1000 mmol) su, 18,016 gram su demektir veya 18,016 gram su 1 mol’dür Gerçekte 1 mol (1000 mmol) suda Avogadro sayısı (6,023 x 1023) kadar su molekülü bulunur Ekivalan ağırlık (eşdeğer ağırlık), bir element veya bileşiğin 1 mol hidrojen ile birleşen veya onun yerine geçebilen miktarını ifade eder; moleküler ağırlığın valansa (değerlik) bölümüne eşittir. Ekivalan ağırlığın gram cinsinden ifadesi, ekivalan sayısı (Eq) olarak tanımlanır. 1/1000 Eq=1 mEq veya 1 Eq=1000 mEq Örneğin 1 ekivalan HCl, 36,46 gram HCl demektir veya 36,46 gram HCl, 1 ekivalan HCl’dir. Aynı şekilde 1 Eq (1000 mEq) kalsiyum(Ca), 40,08/2=20,04 gram kalsiyum demektir veya 20,04 gram kalsiyum 1 Eq (1000 mEq) kalsiyumdur. Valans (değerlik); bir asit için moleküldeki yer değiştirebilen H atomları sayısı, bir baz için moleküldeki yer değiştirebilen OH- iyonu sayısı ve bir tuz için moleküldeki
1/1000 mol=1 mmol veya 1 mol=1000 mmol.

H A Y V A N F İ Z Y O L O J İ S İ
4
(+) yüklü iyonların yerine geçebilecek H+ iyonu sayısı oksidan bir madde için reaksiyon sırasında alınıp verilen elektron sayısını gösterir.
Boyle yasası
Boyle yasasına göre, sıcaklıklar sabit tutulduğu sürece, belirli ölçüde alınan bir ideal gazın hacmiyle basıncının çarpımı sabittir. Matematiksel bir anlatımla: P x V = k P paskal olarak basınç, V kübik metre olarak hacim, k gaz sabiti (8.3145 J/(mol K).
Boyle yasası, genellikle, sadece hacim ya da basınç anlamında yapılan bir değişikliğin sonuçlarını önceden tahmin etmek için kullanılır. Belirli ölçüdeki herhangi bir gazın, sıcaklığın sabit tutulma şartıyla (bunun için soğutma ve ısıtma kullanılmalıdır), "önce" ve "sonraki" hacim-basınç ilişkisi aşağıdaki gibidir: �� Bu denklem, genellikle herhangi bir (basınç ya da hacim) "sonra" öğesinin bulunması için kullanılır. Örnek: Gazlar, sıkışabilirler. Gazlar Basınç farkı yönünde akarlar. Direnç artarsa akış azalır. “Hava”, bağımsız olarak difüze olan gazların bir karışımıdır. Her bir gazın karışım içinde miktarına bağlı bir “kısmî basıncı” (Pgaz) vardır. Boyle yasası akciğerlerin çalışmasında önemlidir. Akciğerlerde soluk verilmesi sırasında hacim artışı olur ve akciğer basıncı azalır bu durumda dıştaki hava akciğerlere dolar. Tersine soluk vermede akciğer hacim azalı ve artan basınçla akciğerlerdeki hava dışa atılır.
ŞEKİL 1.4 Hacim basınç arasındaki ilişki
Boyle kanuna göre sabit sıcaklık altında gazların hacimleri basınçlarıyla ters orantılıdır. Basıncın artışı ile dolaşımdaki ve dokulardaki gazların hacimleri ve oluşmuş gaz
P1 x V1 = P2 x V2 P2 = P1V1/V2
Sabit sıcaklıkta tutulan belirli bir miktar gazın, hacmi ile basıncının çarpımı
sabittir.

H A Y V A N F İ Z Y O L O J İ S İ
5
kabarcıklarının çapları küçülür. Bu etkiden gaz embolisi ve kabarcık oluşumuyla ilintili olan dekompresyon (vurgun) hastalığında faydalanılır. Su dalmak için yüzeyde hava alıp dalan bir dalgıç akciğer hacmi 6 lt kabul edilirse 10 metrede bu hacim 3 litreye düşer (her on metrede 1 atm basınç artışı olur). Dönüp hiç nefes vermeden su yüzeyine çıktığında akciğerindeki hava miktarı aynı olduğundan, akciğerin hacmi de ilk haline eşit olacaktır. Dalgıç su yüzüne çıkarken sıkıştırılmış olarak solunan bu fazla havanın boşalması için yeterli zaman tanımazsa akciğerlerde genişleyen gaz dokularda yırtılmalara yol açar. Fakat tüple dalış yapan (scuba- self contained underwater breathing apparatus) bir dalgıç için durum farklıdır. Bu sefer dalgıç, 10 metrede hacmi yarısına inmiş olan ciğerini tüpteki havayla doldurur. Şimdi akciğerlerde yüzeydekinin iki katı kadar hava vardır. Yine hiç nefes vermeden su yüzeyine geri döndüğünde, akciğerin içindeki hava ilk hacminin iki katına ulaşmak isteyecektir. Bu da akciğerin kapasitesini zorlayacağından, zarar görmesine sebep olabilir. Eğer dalgıç derinde soluduğu havayı su yüzeyine çıkarken geri vermezse genişleyen hava akciğer dokusunu zedeleyebilir. Yırtılan damarlardan dolaşıma katılan hava tıkanmaya (emboliye) yol açabilir. Dokularda ve merkezi sinir sisteminde çözünen azot derinde karar verme gibi fonksiyonları bozup dalgıçın yaşamını tehlikeye sokabilir. Herry yasası
Bir maddenin belli bir çözücünün belli bir miktarında, belli basınç ve sıcaklıkta çözünebilen en fazla miktarına o maddenin çözünürlüğü denir. Çözülmeyi bazı
faktörler etkiler: Belirli şartlar altında bir gazın sıvıdaki çözünürlüğü o gazın çözülme katsayısı ve basınçı ile doğru orantılıdır. Gazın “kısmi basıncı” ile çözülür doğru orantılı iken sıcaklık ile ters orantıdır. Çeşitli çözücülerdeki gaz çözünürlüğü değişir Örneğin Suda O2: 0.15 mmol/L (zayıf) iken CO2: 3.0 mmol/L (kuvvetli [x20]). Gazın erime katsayısı artıkça yada gazın basınçı artıkça daha fazla gaz sıvıde
erimiş hale geçer. Başka bir ifade ile bir sıvının içinde çözünebilecek gaz miktarı o gazın kısmi basıncıyla doğru orantılıdır.
Gazın basıncı arttıkça sıvıda eriyen gaz miktarıda artar. Buna örnek olarak kola şişesinde erimiş halde bulunan CO2 verilebilir. Kolada çözünen gaz CO2tir ve yüksek basınçlarda daha çok çözünür. İçecek kapağı açıldığında gaz çıkışı fark edilir. Basınç kalktığı için çözünmüş CO2 uzaklaşır ki bu da köpürme şeklinde görülür. Herry yasasının fizyolojik önemi solunum sisteminde ve vurgunda gözlenir. Dalgıçların zaman zaman yaşadıkları“vurgun” olayı da gazların çözünürlüğü ile ilgilidir. Dalgıçlar su altında basınçlı hava solurlar ve bu nedenle kanda daha fazla miktarda azot gazı çözülür. Dalgıç yüzeye çıktığında fazla çözünmüş azot kabarcıklar oluşturur. Bu kabarcıklar eklem ve damarlarda şiddetli ağrılara, felç ve hatta ölümlere neden olur. Herry yasasının canlılardaki diğer önemli etkisi sıcaklığa bağlı olarak çözülmenin(Oksijen sıvıdaki örneğin plazma içinde) çözülmesinin artmasıdır.
• Çözünen maddenin türü, • Çözücünün türü, • Sıcaklık, Basınç, • Ortak iyon etkisi, • Ortamın pH sı, • Yabancı iyonlar, • Kompleks oluşumu.

H A Y V A N F İ Z Y O L O J İ S İ
6
Laplace yasası
Kan damarı duvarı, kan basıncı ve kabın dışındaki çevre basıncı arasındaki farkın bir sonucu olarak gerilir. La Place yasası transmural basınç farkı ile gerginlik, yarıçapı ve damar duvarının kalınlığı arasındaki ilişkiyi açıklar. Tabii ki, daha yüksek bir basınç farkı, fazla gerilime neden olacaktır. Damar çapı daha büyük olan daha fazla bir gerilime sahiptir. Bu üç kural bir denklemin içine sonuçlanır: �
T duvarlarında gerilim olduğunda, P duvar boyunca olan direnç, R silindirin yarıçapıdır ve M duvarın kalınlığı basınç farkıdır. Kalp kan getiren venlerle kalp basınçı artar. Bu durumda kalp ölçüde gerilmiş olur ve yarıçapı (R) ventrikül artar. Bu nedenle çok daha büyük bir duvar tention (T)kalp kası tarafından geliştirilebilir olan kan ejeksiyon sırasında aynı basıncı ( P) oluşturmak için kullanılır. Böylece genişlemiş kalp, normal büyüklükte kalp kıyasla kan aynı miktarda pompalamak için daha fazla enerji gerektirir. Bir balon üflemediğinizi düşünün. Daha sert balonda, içindeki hava basıncı ile dış arasında daha yüksek bir basınç farkı vardır. Basınç farkı yükselir, bu balon lastik duvarlarında gerilim de artar ve bu da balon gerginliğe neden olur. Şimdi çok kalın kauçuktan yapılmış bir balon üfleme hayal edin. Daha fazla basınç farkı balon duvarlarında gerginliği aşmak için gereklidir.
ŞEKİL 1.5 Bir kabın cidarına yapılan basınç
Bir organın duvarındaki gerilim, organ içi basınç ve lümen çapı ile doğrudan ilişkilidir. (gerilim = basınç x çap). Kalbin duvar gerilimi arttığı takdirde beslenmesi bozulur, iskemi sonucunda yetmezlik görülebilir
Fick Yasası
Bir gazın diffüzyonu gazın solübilitesi ve membranın yüzey ölçümü ile doğru, membranın kalınlığı ile ters orantılıdır. Diffüzyon için membranın iki tarafında basınç farkı olmalıdır.
La place yasası Basınç altında kalan damar ve alveolerde basınça maruz kalan membranın şekline bağlı olarak farklı gerilim formülleri kullanılır.
T = (P x R ) / M

H A Y V A N F İ Z Y O L O J İ S İ
7
ŞEKİL 1.6 Gaz difüzyonun etkileyen faktörler.
Moleküler kütle taşınımı, difüzyon, konsantrasyon farklarının bulunduğu bir ortam içinde, yüksek konsantrasyon bölgelerinden düşük konsantrasyon bölgelerine yönelmiş moleküler kütle taşınımı ile karşılaşılır. Difüzyon adı verilen bu olayda, bir noktada birim yüzeyden birim zamanda geçen mol sayısına difüzyon akı yoğunluğu (Mdif ) denir. Fick yasasına göre, difüzyon akı yoğunluğu konsantrasyon gradyenti ile orantılıdır. Ortamın ve taşınan moleküllerin özelliklerine bağlı olan D parametresine difüzyon katsayısı adı verilir
Frank-Starling hipotezi
Frank-Starling mekanizması temel olarak, kalp kası doluş sırasında ne kadar çok gerilirse, kasılmanın kuvvetinin ve aortaya pompalanan kanın miktarının da o kadar büyük olacağı anlamına gelir. Fizyolojik sınırlar içerisinde diyastol esnasında kalp ne kadar kanla dolarsa(kalbe ne kadar gelirse), sistolde de o oranda fazla miktarda kan pompalanır. Yani kısaca kalbe ne kadar kan gelirse kalp o kadar kan pompalar. Frank-Starling yasasının altında yatan temel mekanizma kalp kası liflerinin gerildiklerinde kasılma güçlerini artırmaları yatmaktadır(örneğin lastiğin gerildikten sonra bırakılması gibi). Kalbe venöz dönüsteki herhangi bir artış diyastol sonu hacmi artırır, bu artış ventrikülleri genişletir, kalp kası liflerini gerer, atım hacmini sonuçta da Kalp output (kalp çıktısı) artırır. Atardamarın kapiller damar girişinde kan basıncı 40 mm Hg Osmotik basınç ise 25 mmHg iken ven ucunda kan basıncı 15 mm Hg Osmotik basınç 25 mmHg seviyesindedir. Arter damar ucundan başlayarak kapiller damarlar botunca su, iyonlar kısmende albümin kan proteinleri doku sıvısına(ekstrasellüler sıvıya) geçmeye başlar bu neden kapiller boyunca kanın osmotik basınçi 25 mm Hg 15 mmHg seviyesine iner buna nedenle kapiller damarların arteriol ucunda kan dan doku sıvına net geçiş varken kapillerin ven ucunda osmatik basınçın azalmasından dolayı sıvı geçişi doku sıvıdan kana doğru olur.
V gaz = A x D x (P1 – P2) / T

H A Y V A N F İ Z Y O L O J İ S İ
8
Hücre Fizyolojisi Canlılar, hücre adı verilen en küçük yapısal birimlerin, çok karmaşık fonksiyonları yerine getirebilecek şekilde bir araya gelmesiyle oluşmuşlardır. Hücre, gözle görülemeyecek kadar küçüktür ve çevresi yarı geçirgen bir zarla kuşatılmış durumdadır. Hücreye şeklini veren sitoplâsma gerekli yaşamsal öğeleri içeren, yarı sıvı bir maddedir. Aynı görevdeki hücrelerin kümelenmesi ile dokular; farklı dokuların belirli bir işlevi görmek üzere birleşmesiyle de organlar oluşur. Yapısal özellikleri farklı olan organların bir araya gelmesi sonucunda da belirli bir işlevsel bütünlük gösteren sistemler şekillenir. Canlıdaki tüm metabolik faaliyetler hücre içinde geçer. Hücrenin iki temel bölümü nükleus ve sitoplâzmadır. Nukleus, sitoplazmadan bir nükleus membranından ayrılmıştır; sitoplazma, kendini çevreleyen sıvıdan hücre membranıyla ayrılır. Hücre
Canlının, tüm canlılık işlevlerini yürütebilen temel birimine hücre denir. Hücreyi meydana getiren başlıca beş esas madde; su, elektrolitler, proteinler, lipidler ve karbonhidradlardır. Hücreyi meydana getiren bu maddelerin bütününe protoplazma denir. Bir hücrenin işlevleri organel adı verilen alt sistemler (hücre çekirdeği, mitokondri, endoplazmik retikulum, golgi cisimciği, ribozomlar vb.)tarafından bir iş bölümü halinde yürütülür. Organeller hücre içi sıvıdan zarlarla ayrılırlar. Hücre de dış ortamdan hücre zarı ile ayrılır. Protoplazmayı dıştan çevreleyen bu zar, canlı, esnek, seçici geçirgen özelliğine sahiptir. Hücrenin Fonksiyonel Özellikleri: Hücreler ortamdan ham materyali alırlar. Enerji üretirler: Bu enerji iç ortam dengesini sağlamak ve sentez reaksiyonlarını yürütmek için gereklidir. Kendi moleküllerini sentez ederler. Organize bir şekilde büyürler. Çevreden gelen uyarılara cevap verirler. Çoğalırlar (bazı istisnalar haricinde). Hücrenin Yapısal Özellikleri Kalıtsal bilgiler DNA içinde saklanır. Hücredeki genetik kod temelde aynı yapıdadır. Bilgi DNA dan proteinlere RNA aracılığı ile geçer. Proteinler ribozomlar tarafından yapılır. Proteinler hücrenin fonksiyon ve yapısını düzenlerler. Bütün hücreler seçici geçirgen bir zar olan plazma membranı ile çevrilmiştir.
ŞEKİL 1.7 Hücre ve organeleri
Çekirdek Hücre çekirdeği çekirdek zarı , sitoplazması çekirdekçik ve kromatin iplikten meydana gelir. Hücre bölüneceği zaman çekirdek zarı ve çekirdekçik eriyerek kaybolur. Hücre bölündükten sonra tekrar meydana gelir. Çekirdekçik rRNA’nın sentezlendiği yerdir ve Ribozomun oluşturulmasını sağlar. Çekirdek zarı Endoplazmik retikulumla bağlantılıdır. Porlar yardımıyla çekirdekten çıkan maddeler hücre sitoplazmasına geçer. Çekirdek 3 temel yönlendirme yapar. 1) Hücrenin en basit şekilde iki yeni hücre vermek üzere bölünmesini kapsayan hücresel çoğalması, 2) Hücrenin metabolik aktivitelerinin oluşumu sırasında hücrenin farklılaşmasını kontrol etmesi, 3) Hücrenin metabolik aktivitelerini düzenlemesidir
Bu bölüm için Power
point sunumu adresi:
https://yadi.sk/d/qsERoP
jIKTmpH

H A Y V A N F İ Z Y O L O J İ S İ
9
Hücre boyutu fonksiyon ve işlevine uygun biçimde büyük farklılıklar içeririr. Örneğin İnsan ovumu 200 mikron, Balık yumurtası 5 mm ve Tavuk yumurtası 30 mm boyutlarındadır. Buna karşın beyin hücrelerinin en küçüğü 4-5 mikron çapındadır. Hücre ve hücre içindeki organeller boyutları 20 um başlar, buna karşın makro moleküllerin boyutları 20 nanometre kadardır. İnsan vücudunda yer alan hücrelerin boyutları oldukça farklıdır. Örneğin en küçük hücrelerin boyutu 4-5 mikron kadar olduğu halde yumurta hücresi (ovum) 200 mikron çapındadır ve çıplak gözle bile görülebilir. Hücre boyutları açısından sözü edilmeye değer en ilginç örnek ise sinir hücreleridir. Uzantıları ile birlikte boyu 1.5 metreyi bulan tipleri vardır. İnsan vücudundaki hücreler kübik ve silindirik biçimden örümceksiye kadar değişen biçimler gösterirler. İnsanda Kan hücreleri hariç toplam hücre sayısı 1013–1014 kadardır. Merkezi sinir sistemi, retina, lens kristali hücre sayısı sabittir ve sonradan çoğalamaz. Hücreler çoğunlukla renksizdir, buna karşın deri ve gözdeki pigmentli hücreler vardır. Sıvı ortamda bulunan hücreler (akyuvarlar) küremsi biçimlidirler. Hücrenin şeklide fonksiyona uygun biçimde odukça farklıdır. Sinir hücreleri dalanma bir birine temas ederken, Epitel hücreleri yan yana gelerek fonksiyonlarına uygun yüzey alanları oluştururlar.
ŞEKİL 1.8 Hücre boyutu
1. Hücre, canlının tüm yaşam belirtilerini barındıran ve uygun ortamda
gelişen, bölünüp çoğalabilen en küçük yaşam birimidir.
2. Sitoplâzma inorganik maddeler ve karbon içeren organik moleküllerden
oluşmaktadır.
•••• İnorganik maddeler: Su, mineral iyonlar (anyon, katyon)
•••• Organik maddeler: protein, karbonhidrat, lipid, nükleik asitlerdir.

H A Y V A N F İ Z Y O L O J İ S İ
10
Atomlar molekülleri, moleküller makromolekülleri, makromoleküller makromoleküler kompleksleri oluşturmasıyla, dokuların en küçük yapı taşları olan ve yaşamın tüm özelliklerini sergileyen hücreler oluşmaktadır. Genel olarak tüm hücreler temelde aynı yapıya sahiptirler. Fakat kökene aldıkları dokuya ve dolayısıyla fonksiyonlara bağlı olarak bazı özelleşmeler gösterirler. İnorganik moleküllere, hücrede yaygın olarak bulunan ve önemli görevler üstlenen sodyum (Na+), potasyum (K+), kalsiyum (Ca2+), klor (Cl-), hidrojen (H+), oksijen (O2), tuz (NaCl) ve su (H2O) oluşur. Organik moleküller ise, karbohidratlara; basit şekerler, glikojen, lipidlere; yağ asidleri, trigliseridler, fosfolipid, steroidler, proteinlere, keratin, antikor, aktin, miyozin ve enzimler nükleik asidlere ise; deoksiribonükleik asid (DNA) ve ribonükleik asid (RNA). Protoplazma temel olarak beş maddeden oluşur: su, elektrolitler, proteinler, lipitler, karbonhidratlar oluşur.
Birleşik yada madde Hücredeki oranı(ağırlık) Su(H2O) 70 İnorganik iyonlar(Na, K, Mg, Ca, CI) 1 Küçük metabolitler 3 Proteinler 18 RNA 1.1 DNA 0.25 Fosfolipit ve diğer lipitler 5 Polisakkaritler 2 Hücrenin inorganik bileşikleri
Hücrenin temel sıvı ortamı sudur, birçok hücrenin %70-80’i sudan oluşmuştur (yağ hücreleri hariç). Hücre içindeki birçok kimyasal madde suda çözünmüş durumdadır, diğer maddeler parçacıklar halinde süspanse olmuştur. Kimyasal reaksiyonlar, erimiş kimyasallar arasında ya da süspanse haldeki zarsı yapı ve partiküllerin yüzeyiyle su arasındaki sınırda gerçekleşir.
ŞEKİL 1.9 Moleküllerin üç boyutlu ve iki boyutlu gösterimi.
Suyun önemi, H atomlarının kısmi (+)yük, O2’in de kısmi (-)yük taşımasından ileri gelmektedir. Bu su molekülüne dipol iki kuvvetli özelliği kazandırmaktadır. Hem birbirlerine hem de artı ve eksi yüklü diğer moleküllere bağlanabilmektedirler. Hücrede su
Canlı tüm metabolik olaylarıı su içerisinde gerçekleşir. Su ayrıca maddelerin difüzyonlarına veya bir yerden diğerine akışına olanak sağladığı için hücre içi taşımanın

H A Y V A N F İ Z Y O L O J İ S İ
11
gerçekleştirilmesini sağlar. Su aynı zamanda diğer kimyasal maddelerin davranışını belirleyen çok önemli bazı fiziksel özellikler taşır. Su molekülü aynı sayıda elektron ve proton taşıdığı için nötr bir moleküldür. Su yüksüz olmakla birlikte taşıdığı elektronların dağılımı dengesizdir ve bu nedenle molekülde bir kutuplaşma ortaya çıkar. Oksijen atomunun çekirdeği 8 proton taşır ve hidrojen atomlarının elektronlarını kendine doğru çeker. Bu yüzden su molekülünde oksijen atomunun bulunduğu bölge, hidrojen atomlarının bulunduğu bölgeye oranla biraz daha negatiflik kazanır. Buna karşın hidrojen atomlarının bulunduğu bölge hafifçe elektropozitiftir. Su katı madde molekülleri ile zayıf elektiriksel bağlar kurar. Bu özelliği nedeniyle su birçok katı madde için çözücü görevi yapar. Bunun tipik bir örneği NaCl’ün su içerisinde çözünmesidir. Zıt yüklü olan Na ve Cl atomları arasındaki kuvvetli çekim nedeniyle katı NaCl kristalleri oluşur. Buna karşın su molekülleri Na+ ve Cl- iyonlarına doğru çekilirler ve bu iyonların çevresi su molekülleri ile kaplanır. Sonuçta iyonların etrafında su moleküllerinden oluşan bir tabaka oluşur. Buna iyonların “hidrasyon kabuğu” adı verilir. Hücredeki toplam suyun % 5’i hücredeki diğer yüklü moleküllere bağlanır. Bu suya bağlı su denir. Serbest su ise hücredeki suyun %95’ini oluşturur. Serbest su hücrede diğer maddeler için iyi bir eritici ve sitoplazmanın kolloidal yapısı için iyi bir dağıtıcı ortam sağlar.
. Hücrenin su içeriği, hücrenin yaşı ve hücrenin metabolik etkinliği ile ilgilidir. Su miktarı embriyonik hücrelerde en fazla iken ergin ve yaşlı organizmalarda bu oran azdır. Ayrıca kemik minesi ve kemik dokuda su miktarı yok denecek kadar azdır. Su, yüksek erime ısısına sahiptir. 1 gram buzu eritmek için 0 °C'de 80 kalori gerekir. Erime ısısının yüksek olması suyun donmasını geciktirir; böylece biyolojik sistemler düşük sıcaklıklara dayanıklı olabilen özelliklerini kazanırlar.
Suyun Isınma (özgül) ısısı yüksektir. 1 g suyun sıcaklığını 1 °C arttırmak için 1 kalori'lik enerji gereklidir. Bu özgül ısı, amonyak dışındaki tüm maddelerinkinden yüksektir. Böylece su
• Tüm Biyokimyasal Reaksiyonlar Sulu Ortamda Gerçekleşir.
• İyi bir çözücüdür.
• Hidrolazlar gibi enzimler için substrat. • Isı düzenleyicidir.

H A Y V A N F İ Z Y O L O J İ S İ
12
sıcaklıklarda fazla artış olmadan daha fazla enerji depolayabilir ve böylece canlı sistemde sıcaklık ve metabolik olaylar daha kararlı olabilmektedir.
Suyun gizli buharlaşma ısısı yüksektir. 100 °C'de 1 g suyu 1 g su buharı haline dönüştürmek için 539 kaloriye ihtiyaç vardır. Gizli buharlaşma ısısının yüksekliği canlı sisteminin izotermal olmasında en önemli katkıya sahiptir. Suyun gizli buharlaşma ısısı, Hidrojen bağlarından dolayı yüksektir. Hidrojen bağlarının kırılması için yüksek enerjiye gerek vardır.
Suyun fonksiyonu
Suyun canlıdan ve sistemlerden bağımısz olarak en temel fonksiyonu iyi bir çözücü, taşıma sistemi , sıcaklı düzenleyicisi ve elektrolit dengeleyici olmasıdır.
1. Hücrelerin ihtiyacı olan maddeleri hücreye taşımak: su temel molekül olarak tüm suda erimiş maddeleri taşır.
2. Hücrelerin fonksiyonlarını yerine getirebilmesi için gerekli olan katı maddelerin çözünmesini sağlar, tüm maddeler(hirofobikler hariç) suda erir.
3. Hücrelerde metabolik faaliyetler sonucu oluşan atık maddeleri boşaltım organlarına (böbrek, akciğer, deri, sindirim kanalı) taşıyarak vücut dışına atılımını sağlar.
4. Su vücut ısısını dengede de tutar. 5. Vucud içindeki tüm sıvılar temel olarak kan üzerinden ayarlanır su kanın
hacmini dengeler bu yolla tüm sıvıların miktarını ve yoğunluğunu ayarlar. 6. Besinlerin sindirimine yardımcı olur hidroliz suyla gerçekleşir. 7. Beyin, omurilik gibi bazı organları dış etkenlerden korumak, suyun görevleridir. 8. Suyun ısı kapasitesinin yüksek oluşu vücut ısısının ayarlanmasını sağlar.
Buharlaşma enerjisi yüksek oluşu nedeniyle vücuttaki fazla ısı terleme ve buharlaşma ile dışarı atılır. Vücut ısısı dengelenir. Buharlaşma ısısı yüksek olduğu için su kaybı az olur. Vücuttaki suyun %20 sinin kaybı hayati tehlikeye neden olur.
9. Suyun sıvı halinin yoğunluğu katı halinin yoğunluğundan fazladır. Bu sayede su üstten donmaya başlar. Bu da suda yaşayan canlılar için hayati önem sağlar. Bu maddede açıklamam gereken bi konu var; su üstten donduğu için su dışındaki düşük ısıdan su daha çok etkilenmez. Yani su yüzeyindeki buz tabakası, suyu yorgan gibi örttüğünden dışarıda su -15 derecelerdeyken bile su o kadar soğumaz.
10. Su donarken dışarı ısı vererek izolasyon görevi yapar. 11. Su, vücuttaki boşluklara pasif difüzyonla geçer ve basınçla dengeyi sağlar. 12. Vücuda alınan besinlerin bir kısmının çözülmesini sağlayarak sindirimi
kolaylaştırır. Ayrıca suda bazı vitaminler çözünür, bunlar da metabolizmanın düzenlenmesinde yardımcıdır.
13. Vücutta iyonize olarak asit-baz dengesinin korunmasında rol oynar. Besin maddelerinin ve oksijenin hücrelere taşınmasını ve biyokimyasal reaksiyon ürünlerinin dışarı atılması için taşıyıcı olarak görev yapar

H A Y V A N F İ Z Y O L O J İ S İ
13
Hücrenin organik molekülleri
Hücrede lipit, protein ve karbohidrat olmak üzere temelde 3 organik molekül vardır. Yapısal olarak vücutta en fazla bulunan organik molekül kollejendir. Hücrede su dışında bulunan anyon (negatif yüklü iyonlar) ve katyon (pozitif yüklü iyonlar) hücre kitlesinin %1’ini oluşturur. Bunlar hücre metabolizmasında görev alır ve hücre işlevinde kritik roller oynarlar. Asit-baz dengesini korur ve osmotik basıncın ayarlanmasında önemlidir. Canlı vücudunda biyosentez reaksiyonları sonuncu oluşturulan maddelerdir.
Organik moleküllerin vücutta çeşitli görevleri vardır. Bunlar;
Karbonhidrat Protein Yağ (Lipid) Vitamin Nükleik asit
Element C, H, O C, H, O, N (S, P) C, H, O ----- C, H, O, N, P
Monomer Glikoz Amino asit (a.a.) 3 yağ asidi + gliserol ------ Nükleotid
Görevleri —Enerji vermek (Yapım ve onarım) —Hücre yapısına katılmak (Hc. Membranında) —Depo maddesi (Karaciğer)
—Yapım ve onarım —Yapı ham maddesi (Membranlar) —Düzenleyici rol (Hormonlar) —Enerji kaynağı (Açlık halinde)
—Yapım ve onarım (Hc memb. Yapısına katılır) —Depo maddesi —Isı izolasyonu —Enerji vermek —Basınç ve darbelerden koruma
—Düzenleyici Koenzimlerin yapısına katılırlar.
—Kalıtımdan sorumlu —Yönetici moleküller Protein sentezi
Örnek Nişasta, Glikojen Albumin, Glikoprotein Fosfolipidler, Steroidler
A, D, E, K vitaminleri
DNA, RNA
Enerji Miktarı
1gr 4.1 Kcal 1gr 4.3 Kcal 1gr 9.3 Kcal ---- -----
Bağlar Glikozit Peptid Ester ---- Fosfodiester
Enerji verici moleküller = karbonhidratlar, lipitler, proteinler.
Yapıcı-onarıcı moleküller = proteinler, lipitler, karbonhidratlar.
Düzenleyici moleküller = proteinler, vitaminler, lipitler.
Yönetici moleküller = nükleik asitler.

H A Y V A N F İ Z Y O L O J İ S İ
14
Proteinler
Proteinler anino asit polimerleridir. Karbon (C), hidrojen (H), oksijen (O), azot (N) bilişimindedir. Ayrıca yapılarında kükürt (S) ve fosfor (P)da bulunabilir. Amino asitler amfoter bileşikleridir. 20 çeşit amino asit bulunur, amino asitlerin her birinin radikal grubu farklıdır. İnsan vücudunda sekiz çeşit amino asit sentezlenemez(valin, lösin, isolösin, treonin, metionin, lisin, fenilalanin, triptofan). Bunlara temel (esansiyel) amino asit denir, besinlerle dışarıda alınır. Glisin hariç tüm aminoasitler alfa karbonuna 4 farklı grup bağlandığı için optikçe aktiftir veya şiral karbon taşırlar. Optikçe aktif aminoasitler polarize ışığı sola çeviren L formu Sağa çevirenler D formu optik izomer, stereoizomer veya enantiyomer denir. Proteinler hücre kütlesinin %10-20’sini oluşturur. Hücre proteinleri yapısal proteinler ve genellikle enzim olarak işlev gören globular proteinler olarak ikiye ayrılır. Hücredeki bu tip yapısal proteinler, genellikle uzun, ince filamentler halinde bulunur.
Bu tür intraselüler filamentlerin en bilinen kullanımı, tüm kasların kontraktil mekanizmasıdır. Filamentler ayrıca mikrotübüller halinde silia, sinir aksonu, mitotik iğcikler gibi “hücre iskeleti" organellerini de oluştururlar. Fibriler proteinler ekstrasellüler olarak özellikle kollejen ve bağ dokusunun elastin liflerinde, kan damarları, tendon, ligament vb. içinde bulunur. Globüler proteinler tamamen farklı tiptedir, genellikle tek protein moleküllerinden oluşmuştur ya da daha büyük oranda, fibriler yapıdan çok globüler yapıda bir araya gelmiş bir kaç protein molekülünden oluşurlar.
ŞEKİL 1.10 Polipeptid bağı ile meydana gelen di, poli ve peptid veprimer sekonder ve tersiyer yapı
• Hücre zarı yapısında yer alır ve dokuların yapısal bileşenlerini
oluştururlar. Ör: keratin
• Küçük moleküllerin taşınmasında ve depolanmasında görev alır.
Ör:hemoglobin tarafından O2 taşınması • Hücreler arası sinyal iletim işine katılırlar. Ör:hormonlar
• Enfeksiyonlara karşı koruma görevi üstlenirler. Ör:antikorlar
• Enzim yapısını oluştururlar. Ör: Renin

H A Y V A N F İ Z Y O L O J İ S İ
15
Bu proteinler genellikle hücrede enzim işlevi görür ve fibriler proteinlerin tersine hücre içi sıvıda erimiş durumdadır. Önemli bir bölümü de hücre içindeki membrana yapışık durumdadır. Bu enzimler hücredeki diğer maddelerle doğrudan ilişki içindedir ve kimyasal reaksiyonları katalizlerler.
Proteinler farklı şekiller sınıflandırılır. Protein yapı ve fonksiyon olarak çok farklı şekillerde olabilir.
1 - Katalizör Proteinler : Biyolojik sistemlerde hemen hemen tüm kimyasal reaksiyonlar enzim denen spesifik makromoleküllerle katalizlenirler. Bu reaksiyonların bazıları CO2’in hidrasyonundaki gibi oldukça basittir, diğer bazıları ise, kromozom replikasyonundaki gibi oldukça karmaşıktır. Enzimler çok büyük bir katalitik güç oluştururlar ve reaksiyonların hızını en az bir milyon kez arttırırlar. Yaklaşık 1000 kadar enzim karakterize edilmiş ve bunlardan bazıları kristalize edilmiştir. Pepsin, tripsin, kimotripsin, lipaz, amilaz ve ribonükleaz mide-bağırsak kanalının sindirim enzimleri olarak sırasıyla proteinleri, yağları, şekerleri ve nükleik asitleri parçalama yeteneğindedirler. Bilinen enzimlerin tümü proteindir. Böylece proteinler biyolojik sistemlerde kimyasal dönüşümlerin gerçekleşmesinde yekpare rol oynarlar. Hücre proteinlerinin en büyük kısmını enzimler oluşturur. Bunlar hücrede kısmen eriyebilir şekilde kısmen de yapıya bağlı halde bulunurlar. Memeli hayvan karaciğerinde bilinen enzimlerin sayısı o kadar fazladır ki hücre proteininin hemen hemen tümünü temsil ederler. Tek hücreli E.coli’de 2500 kadar enzim bulunduğu sanılmaktadır.
2 - Taşıyıcı ve Depolayıcı Proteinler: Bazı küçük moleküller ve iyonlar spesifik proteinlerle taşınırlar. Örneğin, hemoglobin eritrositlerde oksijeni kaslara taşır. Demir kan plazmasında transferrin ile taşınır ve karaciğerde farklı bir protein olan ferritin ile kompleks oluşturarak depolanır. Yumurtada ovalbumin, sütte kazein, mısırda zein ve bağday tohumunda gliadin amino asit deposu fonksiyonu gören besinsel proteinlerdir.
3 - Koordineli Hareketten Sorumlu Proteinler: Kasın en büyük kurucusu proteindir. Kas kasılması aktin ve miyozin denen iki cins protein flamentinin birbiri üzerinde kaymasıyla

H A Y V A N F İ Z Y O L O J İ S İ
16
birliktedir. Mikroskopik bakıda, kromozomların mitoz safhasındaki hareketleri ve spermlerin kamçıları ile hareketleri de proteinlerin kasılma hareketiyle meydana gelirler. Kirpiklerin ve flagella’ların hareketi dynein adlı protein ile mümkündür.
4 - Mekanik Destek Sağlayıcı Proteinler: Deri ve kemiğin yüksek taşıma gücü yapısında bulunan fibröz bir protein olan kollagen ile ilgilidir. Deri bu sayede gergin durur ve kemikler bu sayede uyumlu bir bağlantı sağlarlar. Fibronektin ve integrinler hücre dışı matriks proteinleri olarak hücrelerin matrikse bağlanmalarına aracılık ederler. Kolajen birbiri üzerine sarılmış 3 zincirden meydana gelir. Her bir zincir 1400 amino asit uzunluğundadır . 3 amino asidin tekrarlanması ile dengeli bir yapı oluşur. 3.amino asit gliserindir ve içerde kalır. Dışarıda ise proline ve hidroksiproline bulunur. Vücudumuzdaki tüm proteinlerin ¼’ü kolajen’dir. Kolajen, moleküler telleri şekillendirerek, tendonları ve boşlukları güçlendiren, cildin ve iç organların esnekliğini sağlayan ana yapısal proteindir. Kemikler ve dişler de kolajen’e eklenen mineral kristalleri ile oluşmuştur.
5 - İmmun Koruma Sağlayıcı Proteinler :Antibadiler(IgE, IgM, IgD, IgA vb.), bakteri, virus veya diğer organizma hücreleri gibi yabancı maddelerle birleşebilen yüksek derecede spesifik proteinlerdir. Organizmaya yabancı bir protein veya makromolekül (antijen) girdiğinde antikorlar ortaya çıkar ve antijeni bağlayarak antikor-antijen kompleksi oluştururlar. Bu reaksiyona “immun yanıt” denir. Bu durum sadece omurgalılar için söz konusudur.
6 - Sinir İmpluslarının Oluşumundan ve İletiminden Sorumlu Proteinler : Spesifik bir uyarıma sinir hücresinin cevabı reseptör proteinler ile sağlanır. Rodopsin, retinanın rod hücrelerinde bir reseptör proteindir. Reseptör proteinler asetilkolin gibi küçük spesifik moleküllerle tetik çekebilirler. Sinir impulslarının sinir hücreleriyle birleşme yerlerine ulaştırılmasında bunlar gereklidir.
7 - Büyüme ve Farklılaşmanın Kontrolünden SoRumlu Proteinler (Regülatör Proteinler): Genetik bilginin bir sıra dahilinde kontrol edilmesi hücrelerin düzenli büyümesi ve farklılaşması için zorunludur. Yüksek organizmalarda büyüme ve farklılaşma büyüme faktörü proteinler tarafından kontrol edilir. Örneğin sinir büyüme faktörü sinir ağının oluşumunu yönetir. Çok hücreli organizmalarda değişik hücrelerin fonksiyonları hormonlarla düzenlenir. Bu hormonlardan bazıları proteindirler (insulin, ACTH, GH, TSH vb). Gerçekten de proteinler hücrelerde enerji ve madde akışını kontrol eden alıcılar olarak hizmet ederler. Bir çok hormonal uyarıya hücrelerin cevabı G-proteinler olarak adlandırılan ve GTP bağlayan bir protein sınıfı üzerinden gerçekleşir.
8 - Ekzotik Proteinler: Sınıflamaya girmeyen bazı proteinler de vardır ki bunlara ekzotik proteinler denir. Örneğin Antarktika sularında yaşayan balıklarda antifriz proteinler soğuk ortamda yaşamak durumundaki canlıyı donmaya karşı korur. Bir Afrika bitkisinin kuvvetli şeker tadında bir proteini olan monellin insanların tüketimine sunulan, toksik olmayan ve yağlanmayı önleyen bir gıda tadlandırıcısıdır. Bazı canlılarda mevcut yapışma özelliğine sahip tutkal proteinler bu gruba örnek oluştururlar.

H A Y V A N F İ Z Y O L O J İ S İ
17
Protein tipi Örnekler Yapısal proteinler Regülatör proteinler Kontraktil proteinler Taşıyıcı proteinler Depo proteinleri Omurgalı kanındaki koruyucu proteinler Membran proteinleri Toksinler Enzimler
Kolajen ipek, virüs kılıfı, mikrotübüller İnsülin, Adrenokortikotropin, büyüme hormonları Aktin, miyozin, dyenin, knesin Hemoglobin, miyoglobin, transferin Yumurta akı, albümin Antikorlar Membran transport proteinleri, antijenler Besin zehirleri, Difteri toksini Sukraz, pepsin, tripsin
Proteinler sekonder ve tersiyer yapıları ile fonksiyonel hale gelir. Primer yapıdaki bazı amino asitler sekonder yapının değişimine yol açan bağlar ve etkileşimler yapar. Sekonder ve tersiyer yapıda en fazla rastlanan bağ ve etkileşimler iyonik etkileşimler, hidrojen bağları disülfit kovalent bağları vander waals etkileşimleridir. Kovalent bağlar iki atom arasında ortaklaşa kullanılan elektron çiftinden oluşan bağlardır. Negatif yüklü elektronun bağa katılan iki atom tarafından eşit kuvvette çekildiği kovalent bağa nonpolar kovalent bağ denir. Negatif yüklü elektronun bağa katılan iki atomdan birine daha yakın bulunduğu kovalent bağa polar kovalent bağ denir. Proteinlerde Üç Boyutlu Yapısı
Proteinler; belirli bir kimyasal yapıya ve molekül ağırlığa, genlerle belirlenen bir tek amino asit dizisine: iyi belirlenmiş üç boyutlu bir yapıya (konformasyon) sahiptir. Konformasyon, basit bağlar etrafında mümkün olan dönmeler sonucunda bağlarda kopma olmaksızın çok çeşitli pozisyonlarda bulunabilen grupların uzaysal düzenini ifade eder. Konformasyon; proteinlerde daha çok ikincil, üçüncül ve dördüncül yapıların beraberliğini ve bütünlüğünü izah eder. Proteinin görevi ile (taşıma, katalizleme vb.) sıkı ilişkilidir.
Proteinler yüksek yoğunluğa sahiptir(1,4 g/cm3). Bu neden proetinleri oluşturna amino asitler molekülde oldukça sık durumdadırlar. Peptid zincirlerinde yüksek yoğunluğa bağlı olarak iç kısımda hidrofob, az hidratize olan bir bölge oluşur. Yüklü gruplar hemen hemen yalnız yüzeyde bulunurlar ve sulu çözeltide bir hidrat zarfı ile çevrildiğinden molekül olduğundan biraz daha büyük görünür. Bu durum ultrasantrifüjde, sedimentasyonda ve jel kromatografisinde önemlidir. Bu tür proteinler su ve tuz çözeltilerinde çözünerek alınabilirler. Ancak biyolojik membranların sentezine katılan proteinler dış kısımlarında hidrofob gruplar taşırlar ve böylelikle membran lipidleriyle hidrofobik etkileşmeye girerler ve membranda sabitleşirler. Membran içinde yer almış olmaları birçok protein için (özellikle solunum zinciri enzimleri) fonksiyonel bakımdan önemlidir. Hidrofob yüzey nedeniyle böyle proteinler suda veya tuz çözeltilerinde çözünmezler, ancak deterjanlarla çözelti halinde alınabilirler.
Her protein biyolojik aktivitesiyle ilişkili olarak yüzeyinde bir veya birkaç spesifik bölge veya alan ihtiva eder. Bu bölge toplam yüzeye oranla çok küçük bir bölümü oluşturur. Böylece her enzim katalize ettiği reaksiyonunun substratıyla doğrudan ilişkiye girebilecek bir aktif bölgeye sahiptir. Taşıyıcı proteinler ilgili oldukları molekülleri dönüşümlü olarak bağlayacak bir bağlama bölgesi taşırlar. Örneğin, Hemoglobin 4 hem grubu taşır ve herbiri reverzibl olarak O2 ile birleşebilir. Hücre reseptörleri olarak hizmet eden proteinler spesifik substratları için bağlama bölgelerine sahiptirler. Bundan başka, protein hormonlar bir hücre yüzeyinde hormon reseptörleriyle doğrudan ilişki kurabilecek spesifik yüzeysel yapılar içerirler.
Proteinler hidrojen bağlarından zengindir. Proteinlerin üç boyutlu yapısını belirleyen güçler, protein omurgasındaki peptid atomlarının ve yan zincir (R) grupları arasında şekillenen

H A Y V A N F İ Z Y O L O J İ S İ
18
etkileşmeler sonucu meydana gelir. Proteinlerin aktif üç boyutlu yapılarını kazanmalarında çözücü sistemin (su ve içinde çözünmüş bulunan bileşiklerin) de önemli bir rolü vardır.
Biyolojik sistemlerde tüm bu geri-dönüşümlü moleküler etkileşimler başlıca 3 güçle sağlanır:
Hidrojen bağları ana zincirinin =NH ve –C=O grupları arasında alfa-sarmal ve beta yaprak tabaka yapıları oluşturur. Ayrıca, 20 temel amino asitten 11’inin yan zinciri de hidrojen bağı oluşumuna katılabilir.;
Bu gruplar pH değişimine bağlı olarak bazen alıcı ve bazen de verici olarak hizmet ederler.
Proteinlerin üç boyutlu yapılanmasına katılan bu güçler kovalan olmayan zayıf bağlardandır. Fakat hepsinin bir arada çalışmasıyla kooperatif bağlanma adı verilen ve birbirinin etkisini güçlendiren bir yapı ortaya çıkar. Bu şekilde proteinler ortam koşullarındaki aşırı değişmelerde bile konformasyonlarını muhafaza edebilirler. Hatta birçok protein biyolojik aktivitesini kaybetmeden kristalleştirilebilir. Bir molekülün özel üç boyutlu yapısı çeşitli fiziksel metotlarla (x ışını kristalografisi gibi) tayin edilebilir. Canlı hücrelerdeki konformasyon (ya da buradan izole edilmiş en yüksek biyolojik aktivitedeki bir proteinin konformasyonu) natif durum diye adlandırılır. Bir protein natif halde görevlerini en üst derecede sergiler. Yapısal olanlara kollagen, katalitik olanlara enzimler ve taşıyıcı olanlara ise hemoglobin örnek verilebilir. Protein moleküllerinin çoğu biyolojik aktivitelerini veya fonksiyonel kapasitelerini çok sınırlı bir ısı ve
Hidrojen bağı oluşturma yeteneği olan gruplar:
1. Triptofan (indol) ve argininin (guanidin) yan zincirleri (sadece hidrojen bağı vericisidirler). 2. Peptid grubuna ek olarak glutamin, asparajin, serin ve threonin yan zincirleri (hidrojen bağı vericisi ve alıcısı olarak hizmet edebilirler). 3. Lizin e-amino grubu, aspartik asit ve glutamik asit b- ve g-karboksil grubu, tirozin fenol grubu ve histidin imidazol grubu.
1. Hidrojen bağları 2. Elektrostatik bağlar (tuz bağı yada iyonik bağ) 3. van der Waals çekmeleri
Amfoterik, Amfiprotik, Amfipatik Amfoterikterimi hem asit hem de baz olarak davranabilen türler(atom, molekül veya iyon) için kullanılır. Amfiprotik ise proton, H+alabilen veya verebilen tür anlamına gelir . Suyu veya HCO3- gibi bir iyonu ele alırsak bunların hem amfoterik hem de amfiprotik olduğunu görürüz. Ama öte yandan tek başına metaller veya bazı metal/yarı metal oksitler (Al, Zn, Al2O3,PbO,SnO gibi..) amfoter özellik gösterdiği halde amfiprotik değillerdir. Amfipatik'e gelince hem hidrofobik hem de hidrofilik bölgelere sahip moleküller için kullanılır. Verilebilecek en basit örnek fosfolipidlerdir: fosfolipidlerde yağ asitlerinin bulunduğu kısım hidrofobiktir ve hücre zarının içi kısmında yer alırlar; fosfat grubunun bulunduğu kısım ise hidrofiliktir ve zarın dış kısmına bakar.

H A Y V A N F İ Z Y O L O J İ S İ
19
pH’da muhafaza ederler. Yüksek ısı veya pH’da protein molekülleri denatürasyon denen bir yapısal değişikliğe uğrarlar. Birincil yapı dışındaki diğer yapılar bozulur, kovalan bağlar dışındaki bağ yada çekmeler kopar. Proteinlerin çoğu 50-60oC’nin üzerinde denatüre olur. Bazıları 10-15 C’nin altında da denatüre olurlar. Denatürasyon proteinlerin biyolojik aktivitelerinde kayba neden olur (enzimlerin ısı ile inaktivasyonu, proteinlerin ısı ile koagülasyonu). Denatürasyon çok sayıda etken tarafından oluşturulabilir. Bunlar ısı ve ilaçlar (6 mol/L üre vb), deterjanlar (SDS vb) ve sülfidrilli ayıraçlardır (merkaptoetanol vb). Denatürasyona uğramış protein bazan sebep olucu etkenin ortadan kalkmasıyla yeniden natif hale dönebilir (renatürasyon). Denatüre edilen protein bir enzim ise renatürasyon ile katalitik aktivitesini yeniden kazanabilir. Renatürasyon biyolojik bir aktiviteyi restore edebilir, fakat asla natif proteinde mevcut olmayan bir aktiviteyi meydana getirmez.
ŞEKİL 1.11 Polar ve nonpolar kovalent bağlar.
İyonik (elektrovalent) bağlar ise atomlar, elektron kazanarak ya da kaybederek iyon adı verilen yüklü parçacıkları oluştururlar. Zıt yüklü iyonlar arasındaki çekim kuvveti sonucu olarak da iyonik bağlar oluşur Vander waals bağları elektriksel çekim kuvvetlerinin etkisi ile birbirlerine yaklaşan iki atom arasında, atomlar birbirlerine göre en kararlı oldukları uzaklıkta oluşur. Hidrojen bağları, aynı cins (hidrojen ile azot ve oksijen) arasında oluşur moleküller arasında, farklı cins moleküller arasında, bir molekül içinde oluşabilir.
ŞEKİL 1.11 Hidrojen bağları.
Kovalent bağ, iki atom arasında, bir veya daha fazla elektronun paylaşılmasıyla karakterize edilen kimyasal bağın bir tanımıdır. Genellikle bağ, ortaya çıkan molekülü bir arada tutan ortak çekim gücü olarak tanımlanabilir. Paylaşılan elektron ya da elektronlar, her iki çekirdek etrafında dolanacaklar, iki çekirdek arasındaki bölgede daha uzun süre bulundukları için bu bölgede (-) yüklü bir alan yaratacaklardır. Bu alan, her iki çekirdeğe bir çekme kuvveti uygulayarak bir bağ yaratır. Kovalent bağ, söz konusu atomların dış yörüngelerinin dolması ile meydana gelir. Bu tür bağlar, moleküller arası hidrojen bağından daima daha güçlü, iyonik bağ ile ise ya aynı güçte ya da daha güçlüdür. Bazı inorganik maddelerin hidrojen(H), amonyak (NH3), klor (Cl), su(H2O) ve azot(N) molekülleri ile tüm organik maddelerin molekülleri kovalent bağ ile bir arada tutulmaktadır.

H A Y V A N F İ Z Y O L O J İ S İ
20
Bir hidrojen (H) atomunun oksijen (O) ve azot (N) gibi bir elektronegatif atoma kovalent bağlanması halinde, elektronların oksijen ve azot atomuna hidrojenden daha yakın bulunmaları nedeniyle elektropozitif hale gelen hidrojenin başka bir elektronegatif atom tarafından çekilmesi sonucu meydana gelir. Hidrojen bağlarında, hidrojen bağı donörleri (vericileri) diye bilinen −−−−OH, ⟩⟩⟩⟩ NH, −−−−SH gruplarının hidrojen atomları, O, N, S gibi akseptör ( alıcı) atomların serbest elektronları ile etkileşirler.
ŞEKİL 1.12 Sekonder ve tersiyer yapıdaki bağ ve etkileşimler.
Hücrede sentezlenen protein her canlıda kendine özgüdür. Proteini oluşturan amino asitlerin çeşidi, sayısı ve dizilişleri hücre DNA’sı tarafından her canlıda farklı bir şekilde belirlenir. Proteinlerde amino asitlerin dizilişi genlerle kontrol edilir. Proteinler, yüksek sıcaklık, basınç veya kuvvetli asitlerle etkileştirilirse yapısı bozulur, aktifliklerini kaybeder. Bu olaya denatürasyon denir.
ŞEKİL 1.13 Amino asit çeşitleri
Proteinler her organizmada bulunan önemli bir makromolekül sınıfıdır. Proteinler, 20 farklı tip L-α-amino asitten meydana gelen polimerlerdir. Biyolojik fonksiyonlarını yerine getirebilmek için proteinler uzay içinde belli bir biçim alacak şekilde katlanırlar. Bu katlanmayı yönlendiren güçler, protein atomları arasındaki hidrojen bağı, iyonik etkileşimler, Van Der Waals güçleri ve hidrofobik istiflenme gibi, kovalent olmayan etkleşimlerdir. Proteinlerin işlevlerini moleküler düzeyde anlayabilmek için genelde onları üç boyutlu yapısının çözülmesi gerekir.

H A Y V A N F İ Z Y O L O J İ S İ
21
Proteinler başka birleşiklerle oluşturdukları bileşiklere iki gruba ayrılır 1-Basit proteinler: Hidrolize uğradıklarında yapı taşları olan amino asitlere ayrılırlar. Basit proteinlere örnek; albumin ve globulinler verilebilir. 2-Bileşik proteinler: Amino asitlere ek olarak protein özelliğinde olmayan başka grupları da içerirler. Bileşik proteinlerin hidrolizleri sonunda amino asitlerin yanı sıra başka bir organik bileşik de meydana gelir. Örneğin;
Canlı hücredeki reaksiyonların çoğu, beş genel kategoriden birine uyar:
Karbohidratlar
Karbohidrat terimi, orjinal olarak karbon, oksijen ve hidrojenden oluşmuş bir grup bileşiği belirtmektedir. Karbohidratlar, kimyasal yapı bakımından, polihidroksi aldehit, polihidroksi keton ya da hidrolizi sonucu bu tip bileşiklere ayrılan bileşiklerdir. Karbohidratlar yapısal olarak başlıca dört gruba ayrılabilirler. Bunlar sırasıyla, monosakkaritler, disakkaritler, polisakkaritler ve karbohidrat türevleridir.
Karbonhidratlar glikoprotein moleküllerinin bir parçası olmak dışında yapısal açıdan fazla işlevsel önem taşımazlar ama hücre beslenmesinde büyük rol oynarlar. İnsan hücrelerinin çoğunda karbonhidrat depoları fazla büyük değildir, genellikle toplam hücre kitlesinin %1’ini oluşturur, bu oran kas hücrelerinde %3, karaciğer hücrelerinde %6’dır. Çözünmüş glikoz şeklinde karbonhidrat ekstrasellüler sıvıda her zaman kullanılabilir durumdadır. Az miktarda karbon hidrat, hücrede, suda çözünmeyen glikoz polimeri olan glikojen halinde depolanmıştır, glikojen, gerekli olduğu durumlarda hızla hücrenin enerji gereksinimini karşılar.
• Nukleoprotein = protein + nukleik asit (Histon + DNA) • Glikoprotein = protein + polisakkarit • Lipoprotein = protein + yağ asitleri • Kromoprotein = protein + boya maddeleri (hemoglobin)
• Fonksiyonel grup transferi
• Oksidasyon-redüksiyon
• Bir veya daha fazla karbon atomu çevresindeki bağ yapısının yeniden düzenlenmesi
• CC bağlarını oluşturan veya yıkan reaksiyonlar
• Bir molekül su çıkışıyla iki molekülün kondensasyonu reaksiyonları

H A Y V A N F İ Z Y O L O J İ S İ
22
ŞEKİL 1.14 Sükroz molekülü meydana getiren fruktoz ve glikoz
Aldozlar ve Ketozlar: Karbonil bileşikleri ve önemi Karbonil grubu (C=O), sp2 melezleşmesi yapmış ve oksijenle çift bağ oluşturmuş bir karbon atomu taşır. Bu grubun geometrisini sp2 melezleşmesi yapmış olan karbon atomu belirler. Karbonil grubu sp2 karbonu dolayında düzlemseldir. Karbonla oksijen arasında bir sigma bağı, bir pi bağı vardır. Ayrıca oksijende iki ortaklanmamış elektron çifti bulunur. Karbonil grubu alkol ve eterdeki C-O bağından daha polardır. Bu durum, pi elektronlarının elektronegatif oksijen atomu tarafından, C-O bağındaki sigma elektronlarına göre daha kolay çekilmesine yorulabilir. Fonksiyonel grup olarak bir aldehid içeren monosakkaridlere aldozlar, keton grubu içerenlere de ketozlar denir. ör: gliseraldehid ve glukoz bir aldoz, dihidroksiaseton ve fruktoz ise ketozdur.
D-Glikoz bir aldohekzozdur. Dekstrorotatör olduğundan dekstroz adını da alır. Birçok önemli disakkarit ve polisakkaridin yapısına katılır. Üzümde bol miktada bulunduğundan üzüm şekeri de denir. Tatlı ve lezzetlidir.
D-Galaktoz doğada nadiren serbest halde bulunur. Laktozun yapı taşıdır. Ayrıca serebrositlerin, gangliositlerin ve glikoproteinlerin yapısında yer alır. L-Galaktoz yosunlardan elde edilen bir polisakkarit olan agar içerisinde bulunur. Tadı glikozdan daha azdır. Fermentasyon yönünden de glikoza kıyasla mayalar tarafından daha yavaş fermente edilir.
D-Fruktoz bir ketoheksoz olup levorotatör özelliğinden dolayı levuloz adını alır. Sakkarazun ve inulinin yapı taşıdır. Bitkilerde yaygındır. Balda da bulunur. Fötüs kanında, plasentada ve sperma sıvısında serbest halde fruktoz vardır. Bir trisakkarid olan rafinozun yapısına katılır.
D-Mannoz kısmen serbest ve kısmen de bağlı şekilde bitkilerde bulunur. Hayvansal organizmada gilkopepditlerin, kan grubu maddelerinin yapı taşı olarak sıkça rastlanır.
Tabiatta bulunan pentozlardan L-arabinoz, D-riboz ve D-ksiloz gibi aldozlar ve ketopentoz olan L-ksiluloz mevcuttur. Alfa-D-ksiluloz pyranoz halkası teşkil eder ve bu halka şekli glikozun 5 numaralı karbonuna bağlı hidroksimetil grubu hariç diğer bakımdan glikoza çok benzer.

H A Y V A N F İ Z Y O L O J İ S İ
23
D-Riboz ribonükleik asitlerin ve koenzim görevi yapan nükleotidlerin yapısında bulunur.
D-Ksiloz Ağaç zamklarında, proteoglikanlarda bulunur. Bu şekerin keto şeker karşıtı D-ksilüloz’dur. D-ksilüloz üronik asit yolunda önemli bir ara maddedir.
Nöyraminik asitler 9 karbon taşıyan( bu yüzden nonoz adını alır) D-şekerlerdir. Piruvik asitle mannozaminin birleşmesi sonucu oluşur. Hayvansal hücre çeperlerinin önemli yapı taşıdır. Lipidlerin polisakkaridlerin, glikoproteinlerin ve mukoproteinlerin yapısında olarak bakterilerde ve hayvan dokularında yaygın bir şekilde bulunur. Nöyraminik asitin N-açil türevleri sialik asitler olarak bilinir. Sialik asitler tükrük bezi salgısı ve diğer mukoza salgılarındaki glikoprotein yapısında bulunurlar. Ayrıca kan grubu maddelerin yapı taşıdır. Mümarik asit bakteri hücre duvarlarının önemli yapı taşıdır. Burada da amino grupları asetillenerek N-asitil mümarik asit oluştururlar. Silaik asit farklı canlıların membran yapılarında bulunur örneğin virüs, bakteri, protistler, Trichomonas cinsi (Trichomonas fetus), hayvan sakatatlarında bulunur. Özellikle bakterilerden, Vibrio cholerae, Bacillus sp kapsül yüzeyinde belirlenmiştir Viruslerin (myxovirus paramyxovirus) kan hücreleri bağlanması için reseptör görevi görür.
Glikozidler: Monosakkaridler, glikozid bağlarıyla bağlanarak daha büyük yapılar oluştururlar. İki monosakkarid içeren yapıya disakkarid denir. ör: laktoz (galaktoz + glukoz), maltoz ( glukoz + glukoz), sukroz (glukoz + fruktoz). 3–12 monosakkarid ünitesi içerenlere oligosakkarid, ör: glikoproteinlerde bulunur, 12'den fazla monosakkarid içerenlere de polisakkarid denir, ör: glikojen (homopolisakkarid) ve glikozaminoglikanlar (heteropolisakkarid).
Monosakkaritteki aldehit grubu veya keton grubu ile molekül içerisindeki bir alkolik hidroksil grubu arasında oluşan hemiasetal ve hemiketal bağ aynı zamanda gilkozidik bağdır. Eğer glikozidik bağ monosakkaritlerin D-glikoz ve D-fruktoz içinde meydana gelecek olursa her iki şekerin a-D ve b-D şekilleri meydana gelmektedir. Bu glikozidik bağ monosakkaridin kendi içerisinde meydana gelebildiği gibi iki monosakkarit arasında da meydana gelebilir. Bu takdirde disakkaritler oluşmaktadır. Eğer glikozidik bağ pek çok monosakkarit arasında oluşuyorsa bu durumda da polisakkaritler meydana gelmektedir. Metil alkol ile glikoz reaksiyona girerse bu iki yapı arasında glikozidik bir bağ oluşur ve metil glikozid meydana gelir. Bu şekilde oluşan esterlere de glikozidler denir. Bunların da a- ve b- şekilleri vardır. Tabiatta, hidrolize olduğunda şeker ve alkol veren, karbonhidrat olmayan artıklar ihtiva eden bir çok glikozid vardır. Bu glikozidlerin karbonhidrat olan bölümüne glycone, karbonhidrat olmayan bölümüne de aglycone denir. Glycone’un hidrofilik karakterinden dolayı glikozid, suda aglycone’dan daha fazla erir. Aglycone’lar, çiçek renkli maddelerindeki çeşitli flavonlar ve anthocyanin’ler dahil, ekseriye bitkilerde bulunan fenolik bileşiklerdir. Bir çok meyve ağaçlarının köklerindeki zehirli bir glikozid olan phlorizin’deki polyphenol ve kendisinden indigo boyasının elde edildiği glikozidde bulunan indoxyl aglycone’dırlar. Tıpta kullanılan glikozidlerden en önemlisi kalp glikozidi olarak bilinen dijitaldir.
Glikojen Hayvanlarda glikozun depo şeklidir. Dallıdır ve alfa glikozit bağı ile bağlanmıştır. Suda çözünür. İyot ile kahverengiye boyanır. En fazla karaciğer ve kaslarda bulunur, depo edilir. Hidroliz olunca glikoz ünitelerini verir. Amiloz’dan çok amilopektin’e benzer. Hem a-1,4 hem de a-1,6 glikozidik bağ taşır. Glikojende her bir indirgeyici olmayan son gruba karşılık 8-12 glikoz artığı bulunur. Molekül ağırlığı 270.000 ile 100 milyon arasında değişir.

H A Y V A N F İ Z Y O L O J İ S İ
24
Glikojen hayvan hücresinde nişasta granüllerinden çok daha küçük parçacıklar halinde bulunur. Glikojen su ile kolay karışır ve opalescent eriyikler meydana getirir. Dekstrinler nişastanın enzim veya asitle hidrolizi sonucu oluşur. Glikoz ünitelerinden kuruludur. Suda erir. Çocukların beslenmesinde kullanılır.
Karbonhidratlar, yağ asitlerine dönüşebilir ve asetil CoA ara metabolizmasının kavşak noktasındadır. Karbonhidratlar, bazı aminoasitlere çevrilebilirler. Karbonhidratların α keto aside dönüşmesinin ardından aminlenmesi gelir.
Bazı aminoasitler glukozun karbon atomlarından köken alabilirler. Aminoasitleirn metabolizmasının krebs siklusunun bir ara yapısına dönüştüğü durumlarda, bu özellik söz konusudur. Çünkü oksaloasetat, fosfoenol piruvatı verebilir. Bazı aminoasitler, lipidlere çevrilebilir. Lipidlere çevrilmeler asetil CoA nın ve gliseraldehit 3-fosfattan itibaren gerçekleşir. Şöyleki aminoasitlerin karbon iskeletinin daha ileri yıkılımında son ürün olarak sitrik asit siklusu ara maddeleri olan piruvat ve asetil CoA meydana gelmektedir. Oluşan asetil CoA, sitrik asit siklusuna katılmakta veya asetoasetat üzerinden yağlara dönüşmektedir. Krebs siklusu aracılığıyla yağ sentezlerinden itibaren, karbonhidratların net bir sentez olasılığı yoktur. Çünkü iki karbon atomu bir molekül asetil CoA tarafından taşınıyorsa, iki karbon atomu da CO2 biçiminde elimine edilir. Yağ asitleri yine bazı aminoasitlerin, aynı proseslerle prekürsörleri olabilmektedirler. 1- Gerek krebs siklusunun prosesi ki, bu proses α ketoglutarik asitten türeyen aminoasitler için çok daha geçerlidir. 2- Gerekse glioksilik asit siklusunun prosesi 3- Nükleotidlerin biyosentezi: Glukozdan riboz 5-fosfat, çeşitli aminoasitler (aspartik asit, glisin, glutamin) yine bazı aminoasitlerin metabolizmasından kaynaklanan tek karbonlu üniteler burada yer alırlar. Karbon iskeletlerinin metabolizması sırasında aminoasitlerden ortak son ürünler elde edilmektedir.
Piruvat → alanin Okzaloasetik asit → aspartik asit α ketoglutarik asit → glutamik asit 3-fosfogliserik asit → serin ve diğerleri
• Karbohidratlar C, H, O den oluşmuş organik moleküllerdir.
• Monosakkarit ve disakkarit yıkımı ile hücresel enerji sağlanır ve bunlar
diğer hücresel bileşenlerin sentezi için başlangıç materyali sağlar.
Ör:Asetil ko enzim A
• Polisakkaritler hücrenin depo formunu (nişasta glikojen) ve hücrelerin
yapısal bileşenini (selüloz, kitin) oluştururlar.
• Polisakkaritlerin daha kısa polimerleri hücrelerin komşularına adezyonu
ve proteinlerin uygun intraselüler hedeflere taşınmaları görevini üstlenirler.

H A Y V A N F İ Z Y O L O J İ S İ
25
� Glikozaminoglikanlar
Glikozaminoglikanlar az miktarda protein ve negatif yüklü heteropolisakkaritler zincirlerinden oluşan büyük zincir yapılarıdır. Temel olarak su tuz dengesinde fonksiyoneldirler. Hücrelerin arasında bulunan "dolgu" maddesi olan hayvansal ekstraselular matriksinin büyük bir bileşenidir. Bu moleküller ayrıca, katyonları (sodyum, potasyum ve kalsiyum gibi) ve suyu bağlamakla ve de matriks boyunca moleküllerin hareketini düzenlemekle ilişkilendirilirler. Matriks içindeki sinyalleme moleküllerinin ve proteinlerin stabilitesini sağlar. En öenmli glikozaminoglikanlar 'Hiyoluroik asit'' ,sinoviyal sıvıda, kemik ve kıkırdak dokuda, gözün iç kısımında ve bağ dokuda bulunur. ''Kondriotin sülfat'' ise kemik dokuda, kalp kapakçıklarında ve atardamar duvarlarında bulunur. Yer aldığı dokularda çeşitli fonksiyonlara sahiptir. ''Keratan sülfat'' ise insanda aort duvarında yaşlılığa paralel olarak artan bir mukopolisakkarittir. Kanın pıhtılaşmasını önleyen ''heparin'' bazofil lökositlerde ve karaciğerde bulunur. Bunlarda başka metabolizmada farklı işlevleride vardır. Kan gruplarının oluşumunda da fonksiyonu olan''Mukoid'',solunum borusunda, bağ dokuda, tükrükte, sümükte, rahim ağzında ve sinidirim bezlerinde varolan bir Glikozaminoglikanlar . Lipidler
Suda erimezler ancak yağ çözücülerde (alkol, eter, kloroform gibi organik eriticiler) erirler. Hücrelerin çoğundaki en önemli lipidler fosfolipidler ve kolesteroldür. Bu tip yağ toplam hücre kitlesinin %2’sini oluşturur. Fosfolipid ve kolesterolün suda erimemesi, hücre membranının ve hücre içi bölümleri birbirinden ayıran membran bariyerlerin oluşumunu sağlar.
ŞEKİL 1.15 Amphipatik molekülün su içinde konumlanması.
Alanin, glisin, serin, sistein, treonin, hidroksiprolin ve triptofan → piruvat Aspartik asit ve asparajin → okzalaoasetat Tirozin, fenilalanin ve aspartat → fumarik asit İzolösin ve metiyonin → süksinil CoA Glutamat, glutamin, histidin, prolin ve arjinin → α ketoglutarat İzolosin, losin ve triptofan → asetil CoA oluşturmaktadır.

H A Y V A N F İ Z Y O L O J İ S İ
26
Fosfosfingolipidlerde gliserol yerine sfingosin ve sfingosine bağlı bir yağ asidi yine fosforik asit ve X değişkeni bulunur. Sfingosin bir amino alkoldür. Sadece hayvansal hücrelerde bulunur. Fosfosfingolipidlere örnek olarak sfingomiyelin verilebilir. Fosfosfingolipidlerde bir yağ asidi bulunur. Glikolipidler gliserol ve sfingosin içermesine göre sınıflandırılırlar. Gliserol içerenlerde iki yağ asidi ve bir gliserol ile glukoz bulunur. Sfingosin içerenlerde ise bir yağ asidi ve sfingosin ile glukoz bulunur. Gliserol içeren glikolipidler bitkisel hücrelerde bulunduğu halde sfingosin içeren glikolipidler hayvansal hücrelerde bulunurlar. Sfingosin içeren glikolipidlere örnek olarak serebrosid ve gangliosid verilebilir. Serebrosidde tek glukoz molekülü bulunduğu halde gangliosidde çok sayıda glukoz molekülü bulunur. Zar yapısına giren bütün lipidler amfipatik moleküllerdir. Sulu bir ortama bırakıldıkları zaman hidrofilik kısımlar suya yönelirken hidrofobik kısımları birbirine bakar. Tek tabakalı, çift tabakalı sıralar halinde dizildikleri gibi dairesel şekilde de dizilebilirler. Bu özellikleri dolayısıyla hücre organellerinin zarlarını ve hücre zarını oluşturabilirler. Fosfolipidlerde yağ asitlerinin bulunduğu kısım hidrofobiktir ve hücre zarının içi kısmında yer alırlar; fosfat grubunun bulunduğu kısım ise hidrofiliktir ve zarın dış kısmına bakar.
ŞEKİL 1.16 Lipit yapısı doymuş ve doymamış trigiliserol.
Fosfolipidler genel olarak fosfodiester köprüsü ile diaçilgliserol ve sfingozin’e bağlanmış bir alkolden oluşan polar ve iyonik bileşiklerdir.
1-Fosfogliserit (Gliserofosfolipit) � � Fosfolipidlerin ana sınıfıdır. Hepsi fosfotidik asit (PA) içerir. Trigilesrid sentezi sırasında da oluşan bir ara ürün olup gliserolden türeyen en basit fosfolipiddir.
1.Fosfotidik asit + Alkol türevleri :
Fosfotidil etanolamin(Sefalin) : PA + Etanolamin
Fosfotidil gliserol : PA + Gliserol

H A Y V A N F İ Z Y O L O J İ S İ
27
Fosfotidil inozitol : PA + İnozitol
1.Membranlar üzerinde glikozil fosfotidil gliserol şeklinde proteinlere bağlanma noktası oluşturur.
2.Fosfotidil inozitol 3,4-difosfat (PIP2) şeklinde hücre içi ikincil haberci sistemine dahildir.
3.Membranlarda bulunan fosfotidil inozitol fosfolipaz A2 tarafından yıkılarak araşidonik asit meydana getirir.
Fosfotidil kolin (Lesitin) : PA + Kolin
1.Hücre zarında en bol bulunan fosfolipid fosfotidilkolindir.
2.Yüzey gerilimini düşüren sürfaktan’ın yapısında bulunur.
Fosfotidil serin : PA + Serin
Kardiyolipin : 2 molekül PA + Gliserol
Antijenik özelliğe sahip tek insan fosfogliserolüdür. İç mitokondri membranında yer alır
2.Plazmojenler (Eter fosfolipidler veya Alkil fosfolipidler)
YA gliserol molekülünün 1.C’u ile ester bağı yerine eter bağı yapar.
PAF (Trombosit agreve edici faktör) : 1-alkenil fosfotidal kolin (Alkil fosfolipid yapıda)
2-Seramid türevleri :����Gliserol yerine bir amino alkol olan sfingozin içerir (Seramid : Sfingozin + YA + 1 molekül H)
1.Sfingomyelin (Fosforil kolin + Seramid) Sinir liflerindeki myelin kılıfının önemli bir bileşenidir.
2.Glikolipidler (Glikosfingolipidler) (Karbohidrat + Seramid) (Fosfat grubu İÇERMEZLER)
Esas olarak plazma membranlarının dış yüzeyinde hücre yüzey karbohidratlarıyla birlikte bulunurlar. Hücresel etkileşimler, büyüme ve gelişmenin düzenlenmesinde rol oynarlar. Eritrosit yüzeyindeki kan grubu ag'leri, çeşitli embriyonik ag'ler ve bazı tümör ag'lerinin yapısında bulunurlar. Kolera ve difteri toksini ve bazı virüsler için hücre yüzey reseptörü olarak görev yaparlar . Antijeniteleri çok yüksektir
Galaktoserebrozit : Seramid + Galaktoz
Gangliozid : Seramid + Oligosakkarit + NANA (N-asetil-nöraminik asit)
Globosid : Seramid + Oligosakkarit
Glukoserebrozit : Seramid + Glukoz
Glikosfingolipidlerin sentezi

H A Y V A N F İ Z Y O L O J İ S İ
28
ER ve golgide glikozil transferazlar tarafından sentezlenirler. (Bu enzimler aynı zamanda glikoprotein sentezinde de görev alırlar) Sülfatid sentezinde de PAPS’dan (3-fosfoadenozin 5-fosfosulfat) sülfotransferaz enzimi aracılığıyla sülfat aktarımı olur.
Fosfolipidler ve kolestrol dışında hücrelerde çok miktarda nötral yağlar da denen trigliseritler de bulunur. Yağ hücrelerinde hücre kitlesinin %95’i trigliseritlerden oluşmuştur. Bu hücrelerdeki yağ, vücudun gereksinim duyduğu her durumda çözünerek enerji sağlayan besinlere dönüşen ana depolardır. Yağ moleküllerindeki karbon atomları arasında bulunan bağlara göre yağ asitleri iki gruba ayrılır.
1. Doymuş yağ asitleri: Karbon atomları arasında tek bulunduğu için bunlar H’e tamamen doymuşlardır. Örneğin, bütirik asit ve palmitik asit. Bu yağ asitlerini bulunduran yağlar genellikle oda sıcaklığında katıdır. Örneğin tereyağı doymamış yağ asidi taşır.
ŞEKİL 1.17 Doymuş ve doymamış yağ asitleri .
2. Doymamış yağ asitleri: Karbon atomları arasında, bazılarında çift bağ bulunabilir. Böyle durumlarda karbon atomları hidrojen ile tam doymamıştır. Örneğin, oleik asit ve linoleik asit doymamış yağ asitleridir. Bunları içeren yağlar da genellikle oda sıcaklığında sıvıdır. Örneğin, ayçiçeği, mısırözü ve zeytin yağ gibi. Enzimler
Enzimler biyolojik katalizörlerdir, yani biyokimyasal reaksiyonları hızlandıran biyolojik kökenli maddelerdir. Enzimlerin en önemli özellikleri: katalitik etkinlik, spesifiklik ve regülasyondur. Enzimler reaksiyonları yaklaşık 1 milyon kat hızlandırırlar. Bilinen en hızlı enzim karbonik anhidraz saniyede 105 CO2 molekülü bağlayabilir (dokulardan kana CO2 taşır). Reaksiyonu 107 kat hızlandırır.
Doymamış Yağ Asitleri Çift bağlı karbon atomları içerirler. Neredeyse tamamı sıvı haldedir. Sık karşılaşılan örnekleri linoleik asit, linolenik asit ve oleik asittir. Çift bağlarının konumuna göre cis veya trans şeklinde bulunabilirler. Böyle yağ asitlerine doymamış yağ asitleri denir. Doymuş Yağ Asitleri Yağ asitlerinin yapısında bulunan karbon atomları mümkün olan en fazla hidrojen ile bağ yapmışlarsa doymuş yağ asidi olarak adlandırılır. Doymuş yağ asitleri oda sıcaklığında katı halde bulunurlar ve hayvansal kaynaklıdırlar.

H A Y V A N F İ Z Y O L O J İ S İ
29
ŞEKİL 1.18 Emzim etkisi
Enzimler, spesifik kimyasal reaksiyonları hızlandırırlar; substratları için yüksek derecede spesifiteye sahiptirler; sulu çözeltilerde çok ılımlı sıcaklık ve pH durumları altında fonksiyon gösterirler. Enzimlerle katalize edilen tepkimeye katılan kimyasal moleküllere substrat adı verilir. Aktif merkez için, enzim-substrat bağlanmasını açıklayan iki model ileri sürülmüştür. Fisher’in anahtar-kilit modelinde, substrat ve enzimin aktif yerinin birbirine uyacak şekilde önceden belirlenmiş olduğu varsayılır. Koshland’ın uyum oluşturma modeline göre aktif merkez esnek yapıdadır; substrat varlığında, proteinin tersiyer yapısında oluşan bir değişiklikle, enzim substratını katalize uygun en doğru biçimde bağlayacak biçimsel bir değişikliğe uğrar.
ŞEKİL 1.19 ATP bağlı olarak konformasyon değiştiren hücre membran pompa proteinleri(Enzim).
Biyolojik reaksiyonlar besinlerin parçalanması, sinir uyarılarının gönderilmesi, kas kasılması gibi olaylarda gereklidir. Aksi durumda bu reaksiyonlar çok yavaş ilerler. Biyolojik ortamlarda reaksiyonlar çok hızlıdır ve enzim, substrat ve ürün için uygun koşulları sağlar. Enzimin substratı ile reaksiyon verdiği bölge aktif bölge (aktif merkez) ve aktif bölgeye bağlanan molekül de substrat olarak adlandırılır. Aktif merkezdeki amino asitler ve bunların grupları substrat ile bağlanır ve kimyasal dönüşümü gerçekleştirir.
Enzimler biyolojik yapıdaki çoğunlukla protein yapısında (RNA hariç) katalizör görevi yapan kimyasallardır. Enzimler reaksiyonu binlerce-milyonlarca kez hızlı olmasını sağlar. Tepkimeye giren maddelerin aktivasyonu düşürür. Enzimler, proteinlerden yapılmışlardır ve doğal olarak yalnız canlılar tarafından sentezlenirler. *Enzimler; etki ettiği maddeye SUBSTRAT ve bu maddeni sonuna "ase=az" eki getirilerek ya da katalizlediği tepkimenin çeşidine göre adlandırılırlar.

H A Y V A N F İ Z Y O L O J İ S İ
30
ŞEKİL 1.20 Enzim aktif merkezi
Bazı enzimler aktivite için, protein yapıyı oluşturan amino asit kalıntılarından başka kimyasal bileşen gerektirmezler. Bazı enzimler ise kofaktör diye adlandırılan bir ek kimyasal bileşen gerektirirler. Kofaktörü ile birlikte tam, katalitik olarak aktif bir enzim, holoenzim olarak adlandırılır. Holoenzimin bir protein kısmı bir de kofaktör kısmı vardır. Holoenzimin protein kısmı apoenzim veya apoprotein olarak adlandırılır. Holoenzimin kofaktör kısmı, bazı enzimler için Fe2+, Mg2+, Mn2+, Zn2+ gibi bir veya daha fazla inorganik iyon; bazı enzimler için ise koenzim denen bir organik veya metalloorganik kompleks bir moleküldür. Koenzimler, bazı enzimlerin aktiviteleri için gerekli olan ve kofaktör diye adlandırılan ek kimyasal bileşenlerin organik veya metalloorganik molekül yapısında olanlarıdır. Koenzim enzime çok sıkı bağlanmış olabildiği gibi, koenzim enzime çok gevşek olarak bağlanmış olabilir. Koenzimlerin enzim proteinine çok sıkı bir şekilde kovalent olarak bağlı olup enzim proteininden ayrılmayanları prostetik grup olarak adlandırılırlar
ŞEKİL 1.21 Enzim çalışması
Koenzimlerin enzim proteinine çok gevşek bir şekilde nonkovalent olarak bağlı olup enzim proteininden ayrılabilenleri kosubstrat olarak adlandırılırlar. Uluslararası Biyokimya ve Moleküler Biyoloji Birliği (IUBMB) tarafından önerilen ve benimsenen sistematik adlandırmada enzimler, altı sınıfa ayrılırlar, her sınıfın da katalizlenen reaksiyon tipine dayanan alt sınıfları vardır:

H A Y V A N F İ Z Y O L O J İ S İ
31
�
Enzimler kimyasal reaksiyonları gerçekleştirdiklerinde bazı faktörlerin etkisi altında kalırlar. Bunlar;
Isı: Her enzim reaksyonunun optimal bir ısı seviyesi vardır. İnsanda bu ısı 36,5 derecedir. Sıfır derecede enzimler canlılıkta kaybedilmeyebilir. Genel olarak enzimler 60 C derece de bozulurlar.
pH: (asitlik-bazlık oranı): Her reaksiyonun gerçekleşebilmesi ortamın pH'ını belirleyen belli oranda [H+] ve [OH-] iyonları konsantrasyonu olmasına bağlıdır.
Substrat konsantrasyonu: Ortamda reaksiyon hızını artırıcı yapılardan biride enzim ve substrat miktarıdır. Her ikisinin miktarı belirli oranlarda artırılırsa reaksiyon hızı sürekli artar.
Su: Enzim reaksiyonunun gerçekleşebilmesi için ortamda belirli oranda su olması gerekir. Çünkü moleküllerin birbirine çarparak reaksiyonu gerçekleştirebilmesi için hareketi sağlayacak sıvı bir ortamın olması gerekir. Tohumlarda su miktarı az olduğundan reaksiyonlarda minimal seviyede gerçekleşmektedir
Vitaminler
Canlı yaşamın sürdürülmesi için gerekli olan ve besin içerisinde doğal olarak bulunan basit yapılı bileşiklerdir. Vücut fonksiyonlarının düzenlenmesinde önemlidir. Vitaminler vücutta parçalanmazlar, yani vitaminler doğrudan enerji elde etmek için kullanılmazlar. Vücut, her vitaminden gerekli olan miktarın kan dolaşımında sürekli mevcut olmasını sağlar. Yağda eriyenler (A, D, E, K) yağ dokuda depolanır, suda eriyenler (B1, B2, B3, B5, B6, B12, C) fazlası vücuttan atılır her gün alınmalıdır.
Suda Çözünür Vitaminler Yağda Çözünür Vitaminler
Thiamin (B1), Riboflavin (B2), Niacin (B3), Pantothenic Acid (B5), Pyridoxal, Pyridoxamine, Pyridoxine (B6), Biotin, Cobalamin (B12), Folic Acid, Ascorbic Acid
Vitamin A, Vitamin D ,Vitamin E ,Vitamin K
Gözdeki pigment tabakası büyük miktarlarda A vitamini bulundurur. A vitamininin ışığa duyarlı pigmentlerin önemli bir ön maddesidir. Gece körlüğü, ciddi A vitamini eksikliğinde meydana gelir. Bunun basit nedeni uygun miktarda retinale dönüşecek yeterli A vitamini olmamasıdır. Bu yüzden, yapılabilecek rodopsin miktarı ciddi şekilde azalır.
Enzim sınıfları
1- Oksidoredüktazlar: elektron transferi (hidrid iyonları veya H atomları)
2- Transferazlar: Grup transfer reaksiyonları
3- Hidrolazlar: Hidroliz reaksiyonları (suya fonsiyonel grupların suya transferi)
4- Liyazlar: Çift bağlara grupların ilavesi veya grupların uzaklaştırılmasıyla çift bağların oluşturulması.
5- İzomerazlar: Molekül içine grupların transferi ile izomerik formların oluşturulması.
6- Ligazlar : ATP hidrolizi ile C-C, C-S, C-O, ve C-N bağlarının oluşumu.

H A Y V A N F İ Z Y O L O J İ S İ
32
ŞEKİL 1.21 Vitaminlerin yapısı
Trioyit bezi üzerinde bulunun paratriot bezlerinden salgılanan paratiroid hormonu böbreklerde D vitaminini aktive eder, aktive olmuş D vitamini de kalsiyum absorpsiyonunu artırır. C vitamini Askorbik asit, kollajenin yapısal bir bileşiği olan hidroksipirolinin oluşumundaki hidroksilasyon aşamasını hızlandıran prolil hidroksilazın aktivasyonu için gereklidir. Askorbik asit olmadan vücudun hemen hemen bütün dokularında yapılan kollajen lifleri kusurlu ve zayıftır. Bu nedenle, C vitamini, derialtı dokusu, kıkırdak, kemik ve dişlerde liflerin büyümesi ve dayanıklılığı için gereklidir. E vitaminin doymamış yağların oksidasyonunu engeller. B1 Tiyamin karbonhidratların ve birçok amino asit metabolizmaları için gereklidir. Riboflavin (B2 Vitamini), dokularda normal olarak fosforik asitle birleşerek iki koenzim olan, flavin mononükleotid (FMN) ve flavin adenin dinükleoid (FAD)’i oluşturur. Bunlar da mitokondrilerdeki önemli oksidatif sistemlerde hidrojen taşıyıcısı olarak görev yaparlar. Folik asit en önemli görevi, DNA sentezinde gerekli olan pürinlerin ve timinin sentezidir. Bu nedenle folik asit, B12 vitamini gibi hücresel genlerin replikasyonu için gereklidir. İnce barsağın ileum kısmında B12 vitaminin emilimi icin gerekli olan intrensek faktör, hidroklorik asit ile birlikle paryetal hücrelerden salgılanır. B12 vitaminin alınması için intrensek faktör ile bağlanması şartır. İntersek faktör B12 vitaminin düşük pH değerlerinde bozulmadan kalmasını sağlar barsağın ileum kısımda reseptöre bağlı pinositoz ile alınmasını gereklidir. İntersek faktör olmadan B12 alınması 1/50 oranında azalır ve kemik iliğinde eritrositler yeterince olgunlaşamaz ve pernisiyoz anemiye oluşur. B6 vitamini protein sentezi için gereklidir. B6 vitaminin en önemli rolü, amino asitlerin sentezindeki transaminasyon olayındaki koenzim görevidir. B6 vitamini Transminasyon amino transferazlar adı verilen enzimlerle çalışır. Bu vitaminin yokluğunda amino asitlerin sentezi çok azalır ve bu nedenle protein yapımı da normal devam edemez. K vitamini intestinal kanalda bakteriler tarafından surekli olarak sentezlenir. K vitamini protrombin, faktör VII, faktör IX, faktör X ve protein C gibi beş önemli pıhtılaşma faktörünün yapımı için gereklidir. Eksikliğinde pıhtılaşma süresi artar. Kumarin olan walfarin K vitamini ile yarışarak ve böylece K vitamininin etkinliğini önleyerek gösterir. Karaciğerde büyük miktarlarda D vitamini ve B12 vitamini de depo edilir. Ayrıca A vitamini eksikliğini on ay gibi uzun bir süre önlemeye yetecek kadar A vitamini karaciğerde depo edilebilir. D vitamini eksikliğini üç-dört ay önleyecek kadar, B1 2 vitamini ise en az bir yıl ya da daha uzun süre eksikliği onleyecek kadar depo edilebilir.

H A Y V A N F İ Z Y O L O J İ S İ
33
Hücre Zarı Hücre zarının bileşimi, hangi dokuda ne görev yaptığına göre değişir. Örneğin Sinir hücrelerinib üstünde yer alan myelin kılıfta lipit oranı yüzde 80 iken diğer hücrelerede yaklaşık olarak %55 protein, %25 fosfolipid, %13 kolesterol, %4 diğer lipidler ve %3 karbonhidratlardan oluşur. Bu maddeler birbirleriyle bağlı ya da bağımsız halde zarın yapısında bulunur. Özetle zarın yapısı fonksiyonuna göre farklılık gösterebilir.
Hücre zarına “plazma zarı" da denir. Hücreyi dış ortamdan ayıran, seçici geçirgen canlı yapıdır. Hücreyi çevreleyen birim zar ortalama olarak 75 Angström (75x10-7 mm) kalınlığındadır. Elektron mikroskobu çalışmaları, zarların lipoproteinlerden yapılmış mozaik şeklindeki fonksiyonel birimler olarak incelenmesinin daha uygun olacağını göstermektedir. Hücre zarı hücreye şekil vermekle kalmaz, besin maddelerinin ve artık maddelerin hücreye giriş çıkışını da ayarlar. Zar aynı zamanda hücrenin koruyucusudur.
İlk bilimsel model 1935 yılında Danielli ve Dawson tarafından ortaya atılmıştır. Bu model uzunca bir süre benimsendi ancak bu model hücre zarının işleyişini açıklayamadı. 1972 yılında Singer ve Nicolson'ın akıcı-mozaik zar modeli önermiştir. Bu modele göre zarın yapısında %65 protein, %33 lipit, %2 karbonhidrat bulunmaktaydı. Hücre zarı, gözenekli ve yarı geçirgen yapıya sahiptir. Esas yapı taşları lipid ve proteinlerdir. Her hücrenin protein, yağ ve karbonhidrat oranları birbirlerinden farklı olduğu için her hücre zarı, o hücreye özgüdür. Hücreye gelen bütün kimyasal maddeler ve elektriksel iletiler hücre zarı ile alınır. Hücre zarının yapısında protein, yağ ve karbonhidrat bulunur. Hücre zarının görevleri;
Yapı Sınıf Altsınıf Fonkisyonu
Lipidler Fosfolipid Gliserol-
Fosfolipitler İki yağ a. oluşan kuyruk ve gliserolü baş
Kolesterol Zarın sertliğine katkı sağlar
Proteinler
Periferal proteinler
Enzim ve sinyal algılama
İntegral proteinler
Kanal proteini Selektif iyon taşıma Taşyıcı protein Selektif madde taşıyıcısı
Karbon hidrat
Hücre dışı glikokaliks
• Hücre zarı, sitoplazmayı çevreleyerek hücreye şekil verir ve
dağılmasını engeller.
• Madde alış verişini düzenler.
• Ozmatik dengenin düzenlenmesinde görev alır.
• Salgı görevi vardır.
• Enzimleri taşıyıcı görevi vardır.
Bu bölüm için Power
point sunumu adresi:
https://yadi.sk/d/FSEQfN
WZKTmpm

H A Y V A N F İ Z Y O L O J İ S İ
34
Hücre zarının temel olarak 4 fonksiyonu vardır.
1. Uyarı iletimi ve alımı yapar.
2. Hücrelerin birbirlerini tanımalarını sağlar.
3- Madde geçisi denetler.
4- Hücrenin matriks(ESM) bağlanmasını sağlar
ŞEKİL 1.22 Hücre zarındaki lipit ve protein dağılımı
Fosfolipid yapısı
Fosfolipitlerin fosfat grubunu içeren polar başları hidrofilik (suyu seven), apolar kuyrukları ise hidrofobik (suyu sevmeyen) özellik gösterir. Bu yüzden sulu ortamda fosfolipitler hidrofobik kuyruklarını yanyana ve uçuca yerleştirerek, sadece hidrofilik başların suyla temas ettigi bir çifte tabaka oluştururlar.
ŞEKİL 1.23 Fosfolipit molekül ve şematik görünümü

H A Y V A N F İ Z Y O L O J İ S İ
35
Oluşan bu çifte tabaka esnek, bir sıvı kristal gibi akışkan, ve kısmen geçirgendir. Bu şekilde oluşan zar yapısının içinde yer alan fosfolipit ve diger moleküller, zarın düzlemi içinde yatay olarak serbestçe hareket edebilirler. Sıvı Mozaik Modeline göre hücre zarındaki lipitler adeta bir mozaik yapı oluştururlar ve bunun içinde yer alan protein ve diğer maddeler için bir çözücü olarak işlev görürler. Zarın oluşturduğu düzlemde bu moleküllerin difüzyon yoluyla yatay hareketi sayesine, hücre zarında çeşitli biyokimyasal reaksiyonlar gerçekleşir. Laboratuar ortamında fosfolipit kullanarak lipozom veya vesikül adı verilen, küresel şekilli, içleri boş, lipit çift tabakalı bir zarla çevrili kesecikler yapılabilir. Hücre zarı üzerinde iyon ve molekül pompaları bulunur. Bu pompalar hücrenin dışındaki bir çok maddeyi hücre içine transfer etmekle görevlidir. Bazı özelleşmiş proteinler ise zara homojen bir şekilde dağılarak çeşitli fonksiyonlar üstlenmiştir. Hücre zarının seçici geçirgen özelliği vardır.
Hücre zarının yapısı için akıcı mozaik zar modeli geçerlidir. Bu modele göre zar proteinleri dağınık haldedir ve tek tek proteinler çift katlı fosfolipit tabakaya gömülüdür.
ŞEKİL 1.24 Fosfolipit yapısı
Lipit ve fosfolipid molekülleri bir iskelet üzerine yerleşmiş polar ve bu nedenle hidrofilik bir baş ile polar olmayan ve bu nedenle hidrofobik iki karbon kuyruğundan oluşur. Lipid ve fosfolipidler sulu bir ortam içinde bulunduklarında, hidrofobik kısımları su molekülleri tarafından dışlanır. Bu nedenle lipid molekülleri ya misel adı verilen küresel biçimde yada çift tabaka (bilayer) biçimde kümelenirler. Fosfolipitler dört bileşenden oluşurlar; bir veya iki yağ asit grubu, negatif yüklü bir fosfat grubu, bir alkol grubu ve de bunları birbirine baglayan bir omurga. Gliserol omurgalı fosfolipitlere gliserofosfolipit veya fosfogliserit denir. Gliserol yerine bir amino alkol olan sfingozin içerir (Seramid : Sfingozin + YA + 1 molekül H) Sfingozin omurgalı tek bir fosfolipit vardır: sfingomiyelin. Seramid üzerine bağlanan sakkarit yapılar eritrositler kan grublarının oluşunu sağlar. Ayrıca Gangliosidler üzerinde oligosakarit gruplar(Sialik asit) taşıyan, hücre zarlarının yüzeylerinin dışına uzanan ve hücre tanıma ve hücreden hücreye iletişimde görev alır. Bu karbohidrat baş grupları bazı bakteriyel protein toksinleri örenğin kolera toksini gibi spesifik reseptör olarak yada hipofiz glikoprotein hormonları için reseptör görevi görür .
Fosfolipaz C hücre içi ikincil haberci (görevi hücre membranındaki fosfatidil inositolü yıkarak iki parçaya ayırmaktır. Bunlar inositol trifosfat ve diaçil gliserol sisteminde rol alan enzimdir. Clostridiyumun alfa toksininde bulunur. Fosfolipaz A2 pankreatik sıvıda bulunan ve gıdalarla alınan fosfolipitleri yıkan enzimdir. ayrıca hücre içinde de var olup, araşidonik asiti serbestlestirir. en önemli sentez blokerı steroidlerdir. bu bloğu lipokortin sentezini arttırarak yaparlar. Yılan ve arı zehirinde bol miktarda bulunur, membrandaki fosfolipitleri yıktıgı icin hücre yapısını bozup ödem geliştirir

H A Y V A N F İ Z Y O L O J İ S İ
36
Gangliosidler karsinogenez gibi dokuların gelişimi ve farklılaşması fonksiyonları gibi içindeki önemli rol oynadığını düşündürmektedir . Tümör oluşumu gangliozide yeni bir tamamlayıcı sentezini neden olabilir ve belirli bir gangliozide çok düşük konsantrasyonlarda ve farklılaşmasını teşvik ettiği bulunmuştur. Hücre zarlarının (membranlarının) ana bileşenleri fosfolipitler, kolesterol ve glikolipitlerdir. Fosfolipit, sentezi mitokondri ve sitoplazmada ER yapılır. Fosfogliserit sentezinin ilk aşamasında fosfatidatın kimyasal olarak aktive edilmesi, yani reaksiyona girmeye eğilimli hale gelmesi gerekir. Fosfolipitlerin sentezinde ya aktive edilmiş bir diasilgliserol ya da aktive edilmiş bir alkol gerekir. Fosfatidil serin ve fosfatidil inositol oluşumunda, bir alkol (serin veya inositol) hidroksili ile sitidin difosfodiasilgliserol (CDP-diacylglycerol) arasında bir fosfo ester bağlantısı oluşur. Fosfatidil etanolamin sentezinde, alkol önce ATP ile fosforlanır, sonra sitidin difosfat (CDP) ile reaksiyona girip aktive olur. Bu alkol ardından diasilgliserol ile reaksiyona girip son ürünü verir. Memelilerde fosfatidil kolin iki yoldan sentezlenebilir: ya fosfatidil etanolamin sentezine benzer bir seri reaksiyonla ya da fosfatidil etanolamin metil transferaz enziminin katalizlediği fosfatidil etanolaminin metilasyonu ile. Sfingomiyelinin sentezinde, sfingozinin amino ucu, uzun zimcirli Asetil-CoA ile birleşerek seramit oluşturur. Bunun ucundaki hidroksil grubuna fosfatidilkolin (diğer adıyla lesitin) eklenmesiyle sfingomiyelin meydana gelir
Hücre membranın görevleri
Lipid tabaka sadece yağda eriyen maddelere geçirgendir; Sterol yapısındaki hormonlar (Kortizol, Testestron ) ya da yüksüz küçük moleküller (O2, CO2, N2). Lipid tabaka içindeki proteinler yüzey boyunca hareket edebilir.
1. Hücre içindeki sitoplâzmayı hücre dışındaki dış ortamdan ayırır.
2. Hücreleri diğerlerinden ayırır.
3. Kimyasal reaksiyonların oluşabileceği belirgin bir yüzey oluşturur.
4. Hücreye materyal giriş-çıkışını düzenler.
5. Yapısında bulunan proteinler hücreye yapısal destek sağlarlar.
6. Bazı proteinler kimyasal reaksiyonları hızlandıracak enzim görevi yaparlar.
7. Reseptör görevi yaparlar.
8. Hücreye antijenik özellik kazandırırlar (hücrelerin birbirlerini tanımasını sağlarlar).
9. Hücre zarının en önemli özelliklerinden birisi seçici geçirgenlik özelliğine sahip olmasıdır. Bu; homeostazis için ve hücrelerin uyarılması için gereklidir.

H A Y V A N F İ Z Y O L O J İ S İ
37
İntegral proteinler- yapısal proteinler başlıca işlevleri:
Secici geçirgen olan Na iyon kanalları çapı 5 A olup Na daha küçük olan Li geçirirken Na daha büyük olan moleküllerin geçmesine izin vermez.
Taşıma yapan kanaların yanlzıca çapları değil aynı zamanda yükleri madde geçişi üzerinde etkidir. Bu yolla kanal proteinlerin belirli maddelere karşı geçirgen belirli maddelere karşı ise geçirgen olmaması sağlanır.
ŞEKİL 1.25 Na iyon kanalı geçirgenliğini belirleyen aralık
Membran hasarı genelde fosfolipazlara ve ATP azalmasına bağlıdır (Tüm hücre membranlarını etkiler) bu durumda membran geçirgenliği ve diğer fonksiyon hasar görür.
Bazı bakteriyel, viral ürünler, sitolitik lenfositler, litik kompleman komponentleri, fiziksel-kimyasal ajanlar plazma membranına zarar verebilir.
- pompa (Na-K ATPaz, SERCA, Na-Glikoz pompası SGLT)
-taşıyıcı protein (GLUT glikoz transportorları)
-iyon kanalı ( Na kanalları, K kanalları, Ca Kanalları)
-reseptör (Protein reseptörleri EPO)
-enzim ve antikor (ACE, Eritrosit antijen oligosakkaritleri)

H A Y V A N F İ Z Y O L O J İ S İ
38
Temel olarak hücre Membranların ortak özellikleri :
� Membran Proteinleri
Hücre zarı ya da plazma zarı hücrenin organellerini ve sıvı içeriğini saran ve hücreye yapısal bütünlük sağlayan ayrıca çok önemli işlevleri yürüten seçici geçirgen bir yapıdır. Her ne kadar biyolojik zarların temel yapısı lipidler tarafından oluşturuluyorsa da, hücrenin yaşamasıyla ilgili birçok özel fonksiyon proteinler tarafından gerçekleştirilir. Proteinler iki tiptir: integral proteinler zarıboydan boya kat ederler, periferik proteinler ise memranın yalnızca bir yüzeyine tutunmuşlardır. Zarda yer alan proteinlerin tipleri ve miktarları oldukça farklıdır. Örneğin; Sinir aksonlarını kuşatan myelin zarlarda, zarın toplam kütlesinin % 25 inden daha azı proteinken, mitokondri, kloroplast gibi enerji üreten yapılarda bu miktar % 75 kadardır. Olağan zarlarda ise bu miktar yaklaşık % 50 dolaylarındadır. Lipid molekülleri, protein molekülleriyle kıyaslandığında, hacım olarak daha küçük olduğundan bu % 50 lik oranda bile çok fazla sayıda lipid molekülü vardır. Genel olarak zarlarda 1 protein molekülüne karşılık yaklaşık 50 lipid molekülü yer alır.
1-Membranlar birkaç molekül kalanlığında ince tabaka yapısındadır. Birçok membranın kalınlığı, 60-100 A arasındadır. Membran kalınlığı fonksiyona göre değişir.
2. Esas yapılarını protein ve lipidler oluşturur. Membranlarda ayrıca protein ve liptlere bağlı karbonhidratlar da bulunur.
3. Membran lipitleri, hem hidrofilik hem de hedrofobik kısımları bulunan nispetin küçük moleküllerdir. Bu lipitler sulu bir ortamda kendiliklerinden kapalı iki tabakalı yapılar oluştururlar. Bu lipit tabakaları yüklü moleküllerin geçişi için bariyer görevi görürler.
4. Özgül proteinler membranın belirli fonksiyonlarına aracılık ederler. Membran proteinleri, pompa, geçit, reseptör, enerji transdüseri ve enzim olarak fonksiyon görürler. Bu proteinler lipit tabakaları arasında yer alırlar ki, bu liptler proteinlerin görev yapabilmeleri için uygun ortamı sağlarlar.
5. Membranlar nonkovalent yapılardır. Membranların yapısını oluşturan protein ve lipit molekülleri birçok nonkovalent bağlantılar kurmuşlardır.
6. Membranlar asimetriktir. Membranların iç ve dış yüzeyleri birbirlerinden farklıdır. Apostatik hücreler hücre içi proteinler Aneksin V dış tarafa döner.
7. Membranlar sıvı yapıdadır. Lipit yapılar membranın yüzeyine difüze olmuşlardır. Özgül birtakım bağlantılar yapmadıkları zaman proteinler de bu şekilde yerleşmişlerdir. Membranlar “proteinlerin ve liptlerin iki, boyutlu çözeltileri” olarak tanımlanabilir.

H A Y V A N F İ Z Y O L O J İ S İ
39
ŞEKİL 1.26 Hücre membrane yapısı
Hücre membranı temel görevleri:
• Hücre içi ortamın özgün bileşimini hücre dışı ortamdan ayırmak, Hücre içi ile hücre dışı ortamlar arasında seçici bir şekilde madde alışverişini sağlayarak hücrenin atıklarını hücre dışı ortama vermek, hücre dışından hücreye gerekli maddeleri almak ve hücre içi ortamın özgün yapısını korumaya yardımcı olmak,
• Komşu hücrelerle iletişimi ve madde alışverişini sağlamak,
• Hücreyi dış ortamdan ayırır. Hücreye şekil verir.
• Madde giriş-çıkşını düzenler. Aktif taşıma olayını düzenler.
• Hücrenin beslenmesine yardımcı olur.
• Komşu ve yabancı hücreyi bulur.
• Hücreyi alınacak hormonları tanır.
• Hücrenin yıpranan kısmını onarır.
• Metabolizma atıklarının dışarı atılmasını sağlayarak iç ortamı düzenler.
• Prokaryot hücreye sahip canlılarda zardaki solunum enzimleri sayesinde enerji üretimi sağlanır.

H A Y V A N F İ Z Y O L O J İ S İ
40
ŞEKİL 1.27 Hücre membrane proteinlerin fonksiyonu
Birçok zar proteini, lipid tabakada bir taraftan diğer tarafa kadar uzanır. Transmembran proteinler olarak adlandırılan bu proteinler amfipatik özelliktedir, yani hem hidrofilik hem de hidrofobik bölgelere sahiptirler. Hidrofobik kısımları lipid tabakanın hidrofobik iç kısmıyla, hidrofilik kısımları ise lipid tabakanın her iki yanında yer alan su tabakasıyla ilişkidedir. Bu proteinlerin bazılarının hidrofobikliği kendilerine kovalent olarak bağlanmış olan ve lipid tabakanın sitoplazmik yarısı içine gömülü bulunan yağ asiti zincirleri nedeniyle artabilir. Zarların sitoplazmik yarısına bağlanan bazı proteinler sadece yağ asiti zincirleri aracılığıyla lipid tabakaya tutunur. Bazı hücre yüzey proteinleri ise küçük bir fosfolipid olan fosfatidil inositole kovalent olarak bağlanarak lipid tabakanın dış yüzünde yer alırlar. Yağ asitlerine kovalent olarak tutunan proteinler zarda bulunan lipidlere kovalent olarak bağlanır. Bu proteinler üç tip lipid grubuyla bağlanırlar.
İntegrinler, İntegral proteinler
İnetgrin proteinler hücre yapışmasında sorum hücre ahdezyon molekül sınıfındadır. Bazı durumlarda hücre sinyal reseptörü olarakta görev alırlar buna karşın İntegral proteinler hücre membranında bulunan membranıı boydan boya geçen bütün proteinlerin ortak adıdır. İntegral proteinler reseptör, enzim, sinyal iletimi, taşıyıcı, hücresel yapışma (integrin) gibi zardan geçen her türlü proteinlerini içeren genel bir kategoridir.
ŞEKİL 1.28 Hücre membran bileşenleri yeri ve dağılımı

H A Y V A N F İ Z Y O L O J İ S İ
41
Membran proteinlerinin görevleri:
Membran proteinlerini tipi, sayısı hücrenin fonksiyona gör değişir. Tüm membran proteinleri temel olarak yaptıkları işlevler:
• Enzimatik görevleri vardır.
• Reseptör olarak görev yaparlar. Hormon, ilaç, bakteri ve virüsler için
reseptör oluştururlar. • Taşımada veya taşımaya yardımcı kanallar oluşturmada görev
almaktadırlar.
• Plazma membranında yer alan transmembran proteinler bir yandan hücre iskelet elemanları ile diğer yandan hücre dışında bulunan bazı ipliksi
proteinlerin bağlantı noktalarını oluştururlar.
• Hücre-hücre etkileşimini sağlarlar.
ŞEKİL 1.29 Hücre membran proteinlerinin görevleri
Membran Karbohidratları
Plazma zarının hücre dışı yüzeyi, zar lipit ve proteinlerinin bazılarına kovalent olarak bağlanmış az miktarlarda karbonhidrat içermektedirler. Glikoproteinler ya da glikolipitler şeklinde adlandırılırlar. İntegral proteinlerin çoğu glikoproteinlerdir ve zarın lipit moleküllerinin yaklaşık onda biri glikolipitlerdir. Bu moleküllerin “gliko” bölümleri hemen daima hücre yüzeyinden dışa doğru değişmez şekilde çıkıntı yapar(örneğin sialik asit).
Glikolipitlerr yalnızca hücre membranın dış kısmında bulunur. Glikolipitler lipit molekülleriyle oligosakkaritlerin oluşturduğu bileşiklerdir. Temel fonksiyonları hücre ile çevresi arasındaki iletişim düzenlenmesidir. Glikolipitlerin diğer önemli bir özelliği düşük pH yani asidik ortamda hücreyi korunaktır. Glikolipitler glikokaliks yapısına katılır.
Proteoglikan olarak adlandırılan karbonhidrat bileşiklerinin birçoğu, küçük protein çekirdeklerle birbirlerine tutunmuş ve hücrenin dış yüzeyine gevşek biçimde bağlanmış karbonhidrat maddeleridir. Bu nedenle hücrenin dış yüzeyi çoğu kez bütünüyle glikokaliks adı verilen gevşek bir karbonhidrat örtüyle kaplıdır.

H A Y V A N F İ Z Y O L O J İ S İ
42
Hücrenin dış yüzeyine tutunan karbonhidrat uçlarının (glikokaliks) birkaç önemli işlevi vardır:
Membran karbonhidratları hemen daima proteinler ya da lipidlerle kombine olarak glikoproteinler ve glikolipidler halinde bulunur. Integral proteinler çoğu glikoproteindir. Bu moleküllerin “gliko” bölümleri hemen daima hücre yüzeyinden dışa doğru asılarak çıkıntı yapar, karbonhidrat bileşiklerinin proteoglikan adı verilen birçoğu, küçük bir protein çekirdekle biri birine tutunmuş ve hücrenin dış yüzeyine gevşek biçimde bağlamış karbonhidrat maddelerdir. Bu nedenle hücrelerin dış yüzeyi glikokaliks adı verilen gevşek bir karbonhidrat örtüyle kaplıdır. Hücrenin dış yüzeyine tutunan karbonhidrat uçlarının birkaç önemli işlevi vardır: Glikolipid ve glikoproteinlerin gliko kısmının hücre yüzeyinde yaptığı karbonhidrat örtüye glikokaliks denir. Karbonhidrat uçlar negatif yüklüdür (hücre yüzeyi negatif yüklü olur).
(1) Glikokalilks hücrenin kimliğinin teşhis edilmesinde,
(2) Çoğu elektriksel olarak negatif yüklü oldukları için hücrenin dış yüzeyinin negatif yüklü olmasına neden olur ve diğer negatif yüklü maddeleri iterler.
(3) Bazı hücrelerin glikokaliksi diğer hücrelerin glikokaliksine bağlanır, böylece hücreler birbirlerine tutunmuş olurlar.
(4) Karbonhidratların çoğu insülin gibi hormonların bağlanması için reseptör görevi yapar, bağlanmadan sonra bu kombinasyon bağlı olan internal proteinlerin aktive olmasını sağlar, bu da bir dizi intraseiüler enzimin aktivasyonuna neden olur.
(5) Bazıları da bağışıklık reaksiyonlarına girer
Glikokaliks •Hücre örtüsüdür •Negatif elektrik yüküne sahiptir. •Antijen antikor birleşmesi için uygundur •Bir hücrenin başka bir canlıda antijen özelliğini göstermesine yol açar. •Kan grupları glikokaliksin antijenik özelliklerine göre belirlenir ŞEKİL 1.28 Hücre zar.
Glikoproteinler tarafından gerçekleştirilen bazı işlevler
Çatı molekülü Kollajenler
Kaydırıcı ve koruyucu ajan Müsinler
Taşıma molekülü Transferrin, serüloplazmin
İmmünolojik moleküller Ig, doku uyuşmazlık antijenleri Hormon hCG, TSH Enzim ALP Özgül kh’larla etkileşim Bazı lektinler

H A Y V A N F İ Z Y O L O J İ S İ
43
Hücre membranın üstündeki glikokaliksin işlevi;
—Eritrositlerdeki glikokaliks kan grubunun oluşumunda etkilidir.
—Difüzyon için seçici bir engeldir.
—Hücrelerin birbirini tanımasını sağlar
—Kontakt İnhibisyona neden olur —Hücreye antijenik özellik kazandırır
—Reseptör görevi yapar
Kolesterol
Kolesterol, hayvanların vücut dokularındaki hücre zarlarında bulunan ve kan plazmasında taşınan bir sterol, yani bir steroid ve alkol birleşimidir. Daha düşük miktarlarda bitkilerde de bulunur. İlk defa 1754'te safra taşlarında kolesterol bulunduğu için bu maddenin ismi Yunanca chole- (safra) ve steros (katı) sözcükleri ile kimyadaki -ol ekinden türetilmiştir.
ŞEKİL 1.30 Zar proteinleri.
Kolesterol, özellikle hayvansal gıdalarda bulunur ama vücuttaki kolesterolun ancak ufak bir kısmı gıda kaynaklıdır; çoğu vücut tarafından sentezlenir. Vücudun her hücresinde bulunmakla beraber, onun sentezlendiği veya hücre zarlarının daha çok olduğu organ ve dokularda, örneğin karaciğer, omurilik ve beyinde kolesterolun yoğunluğu daha yüksektir. Kolesterol kanda normalden fazla bulunması halinde damarlarda birikerek damar sertleşmesine (ateroskleroz) yol açar. Bazen de safra pigmentleri ile birleşerek safra taşlarının oluşumunda rol oynar.

H A Y V A N F İ Z Y O L O J İ S İ
44
ŞEKİL 1.31 LDL alınması ve hücre içi sindirimi
Kolesterol pek çok biyokimyasal reaksiyonda yer almasına rağmen özellikle lipoproteinlerin kolesterolü taşıma biçimleri ve kandaki kolesterol düzeyleriyle kalp hastalıkları arasındaki bağlantıdan dolayı bilinir. Vücut, kolesterolü kullanarak hormonlar (kortizol, üreme hormonları), D vitamini ve yağları sindiren safra asitlerini üretir. Bu işlemler için kanda çok az miktarda kolesterol bulunması yeterlidir.
ŞEKİL 1.32 Kolestrol yapısal formülü
Eğer kanda fazla miktarda kolesterol varsa kan damarlarında birikir ve sertleşmeye ve daralmaya (ateroskleroz veya arteriyoskleroz) yol açar. Aterosklerozda damar duvarında biriken tek madde kolesterol değildir; akyuvarlar, kan pıhtısı, kalsiyum gibi maddeler de birikir. Yüksek kan kolesterolünün zararlarından bahsedilirken söz konusu olan "kötü kolesterol", yani düşük yoğunluklu lipoprotein (İngilizce low density lipoproteins LDL) tarafından taşınan kolesterol düzeyidir. Yüksek yoğunluklu lipoprotein (İngilizce high density lipoproteins HDL) tarafından taşınan kolesterola "iyi kolesterol" denir.
Düşük yoğunluklu lipoprotein (İngilizce karşılığı olan Low Density Lipoprotein'den LDL olarak kısaltılır) kanda kolesterol taşıyan ve yoğunluğu 1,019-1,063 g/mL arasında olan lipoprotein sınıfına karşılık gelir. Apoprotein B100 reseptörlerine bağlanır. Karaciğerde üretilen çok düşük yoğunluklu lipoprotein (İngilizce Very Low Density Lipoprotein, VLDL) metabolizması sonucu oluşur. LDL tanecikleri 18-25 nm çapındadır, taşıdığı lipitlerin yanı sıra apolipoprotein B-100 (apoB-100) ve apoE proteinlerini içerir.

H A Y V A N F İ Z Y O L O J İ S İ
45
Kolesterol İşlevi
Kolesterol, D vitamini ve çeşitli steroid hormonlarının öncülüdür. Ayrıca safra asitleri de kolesterolden sentezlenir. Kolesterol hücre zarlarının (membranlarının) inşası ve bakımı için gereklidir.
Hücre zarının akışkanlığı: Kolesterol hücre membranında fosfolipidlerin hareketlerini kısıtlar, membranın mekanik olarak daha sağlam olmasına hizmet eder.
Yüksek sıcaklık durumunda zarın akıcılığı bozualmasını ve zarın bütünlüğünün kırılmasını engeller. Kolesterol içeren membranlar daha geniş sıcaklık aralığında akışkanlıklarını korurlar. Kolesterol, yağların sindirimine yarayan safranın sentezlenmesinde kullanılır. Ayrıca aralarında yağda çözünen vitaminlerin (A, D, E ve K vitaminleri gibi) metabolizmasında rolü önemlidir. Aldosteron, testosteron, östrojen ve projesteron gibi steroid hormonlarının ve kortizolun sentezlerinde yer alır. Başka araştırmalar kolesterolün sinir hücreleri arasındaki sinapslarda ve bağışıklık sistemi hücrelerinin işlevlerinde rol oynadığını gösterir. Hücre membranının yapısına etkisi sonucunda hücre sinyal iletimine ve membranlardaki iyon ve proton geçirgenliğine de etki eder.
Membran bağlantıları
ŞEKİL 1.33 Bağlantı komplekleri
• Sıcaklık ile doğru orantılıdır.
• Yağ asidi zincir uzunluğu ile ters orantılıdır.
• Yağ asidi doymamışlık derecesi ile doğru orantılıdır. • Kolesterol miktarı ile ters orantılıdır.

H A Y V A N F İ Z Y O L O J İ S İ
46
Tight juction – sıkı bağlantı: Sıkı Bağlantı ya da Zonula Occludens, vücudu dış etkilerden koruyan hücrelerde bulunan, hücrenin yan yüzeyinde yaptığı hücreler arası bağlantılardandır. Epitel hücrelerindeki kuvvetli bağlantılar bu tiptedir. Bu tip bağlantı da hücreler arası boşluk yok denecek azdır. Genellikle de yalıtma özellikleri fazladır. Beyin kan bariyeri, ince barsakta epitelyum hücrelerinde sıkı bağlantı vardır.
ŞEKİL 1.34 Hücreler arası bağlantı yapıları
En yüzeydeki hücreler arası bağlantı noktası. Okludin ve klaudin proteinlerini içerir. Bariyer yapılarında yer alır (kan-beyin, kan-timus, kan-testis, kan-gaz, kan-göz gibi). Mesane ve gastrointestinal sistemde de bulunur
• Gap junction – geçit bölgesi: Epitel hücrelerinin lateral yüzeyindeki morfolojik değişikliklerdir. Bu bölgeler epitel hücrelerinde olduğu gibi düz kas ve kalp kası hücrelerinde de bulunan bir sıkı bağlantı bölgesi olarak iki hücrenin elektriksel bağlanma bölgeleridir. Nexus(6 adet koneksin proetinleri birleşmesiyle konnekson) da delikler bulunduğu için delikli geçit bölgeleri olarakta bilinmektedir. Bu deliklerden iki hücre arasında molekül ağırlığı 1000 daltondan daha küçük moleküller geçer. Örneğin cAMP (siklilk Adenozin mono fosfat) molekülleri bir hücreden diğer hücreye geçerek hücre de bazı faaliyetleri başlatırlar.
Gap junction konnesin adlı protein subunitlerinden oluşur. 6 adet konnesin bir araya gelerek konnekson yapısını oluşturur. Her iki hücrede meydana gelen konneksonlar bir araya gelerek gap juction oluşturan Nexus meydana getirir. Sinsityum halinde çalışan dokulardaki hucreleri bir kanal yapısında birbirine bağlayan oluşumlardır. Normalde hücre mebranları hücre içini dişindan izole eder, gap junction'lar komşu hücre mebranlarını kat eden kanallar vasitasiyla bu hücreler arasi elektriksel uyari ve madde gecişine izin verir, böylece mesela uyarilan bir hücre komşu hücrelere bu uyariyi elektriksel olarak veya kimyasal ligandlar yoluyla iletebilir ve tüm doku uyarilmiş olur.

H A Y V A N F İ Z Y O L O J İ S İ
47
ŞEKİL 1.35 Gap junction connexin proteinlerin bir araya gelemsiyle oluşur.
Desmosoz aynı işlevi yürüten hücrelerin ortak hareket etmelerini ve birbirine yapışmalarını sağlayan sitoplazmik uzantılardır. Çoğunluk simetriktirler. Bu uzantılar küçük bölgeler halinde olabilir (düğme desmozom) ya da hücrenin etrafını çepeçevre sarar (kemer desmozom).
• Gap junctionlar kalp kası, uterus, osteosit, korneada bulunur.
• Hücreden hücreye elektriksel iletimi sağlayan bağlantı yapılarıdır.
• Bir hücrede 6 adet konneksin bir araya gelerek bir yarı kanal (konnekson) oluşturur.
• İki hücredeki konneksonlar yan yana gelince tam bir kanal oluşur (neksus)
• Böylece iki hücre arasında iyon ve hormonların geçişini sağlayan bir kanal oluşmuş olur

H A Y V A N F İ Z Y O L O J İ S İ
48
Mekanik etki altında kalan hücrelerde düğme desmozom daha fazladır. Esasında hücre bağlantıları, hücrelerin serbest yüzünden derinlere doğru farklı bölgeler gösterir.
�
ŞEKİL 1.36 Hücre zarının farklı bağlanma yapıları.
Bazal lamina
Bazal zar ya da bazal membran; epitel hücrelerinin bazal kısımlarını döşeyerek epiteli altındaki bağ dokudan ayıran, düz kas, iskelet kası, kalp kası, sinir hücreleri ve yağ hücrelerinin etrafını kaplayan, çeşitli fibrillerden ve aramaddeden oluşan ince tabakaya denir. Bazal lamina esnek, ince, ekstar sellüler matriksin özelleşmiş şeklidir.
ŞEKİL 1.37 İntersellüler matriks elemanları
Bazal lamima epitel hücrenin altında, ya da düz kas, iskelet kası hücrelerin etrafını kuşatır veya glomerulus, akciğer alveolleri gibi bölgelerde iki hücre tabakası arasında yer alır. Bazal lamina ve retiküler lamina olmak üzere iki katmanı vardır. Bazal zar ait olduğu hücreler tarafından sentezlenir. Epitel hücrelerinin bağ dokusuna bakan yüzeylerindeki hücre zarlarının altında uzanır. Kalınlığı 400–700 Ao arasında değişir. Glikoprotein
belt- kemer desmosom spot- düğme desmosom
Bazal lamina: Fibroblast, epitel, düz kas ve swann hücreleri tarafindan sentezlenen, epitel hücreleri, kas hücreleri, adipöz (yag doku) ve schwan hücreleri etrafinda bulunan tabakadir. Molekül olarak laminin, entactin (nidojen), fibronektin, tenasin, kollagen tip iv proteinleri (glikoprotein) ve heparan sülfat bulunur. laminin ve nidojen temel komponentleridir.

H A Y V A N F İ Z Y O L O J İ S İ
49
yapısındadır. Alttaki retiküler lifler ile süreklilik gösterir. Burada retiküler lifler yoğunlaşıp retiküler laminayı oluşturur. Bazal lamina ile alttaki retikulum telcikleri ağının oluşturduğu tabakaya bazal membran denilir. İki katmandan oluştuğu gözlemlenmiştir: Daha az yoğunlukta ve hücre zarına yakın olan katmana, lamina rara, (lamina lucida); fazla yoğun ve bağ dokusu tarafında yer alan katmana ise, lamina densa denilmektedir. Bazı dokularda bağ dokusu tarafında bulunan üçüncü katmana lamina reticularis denilir. Çok katlı epitelin altında iyi gelişmiş olarak bulunur.
Bazal lamina birbirine belli düzende tutunmuş olan 4 bileşenden oluşur.
�
Bazal lamina filtrasyonda, hücre polaritesinin oluşturulmasında, hücre metabolizması, farklılaşması ve gelişmesinde görev alır. Hasar gören hücrelerin yenilenmesinde ve hücre göçünde de bazal lamina görevi vardır. Pek çok kanser türünde bazal lamina tamamen kaybolur. Yaşlanma, diyabet, Hipertansiyonda bazal lamina kalınlığı artar ve dokular yeterince beslenemez.
Adhezyon Molekülleri:
Adheren (tutturucu, yapıştırıcı) bağlantılar epitel hücrelerinde, fibroblastlarda, kalp kası hücrelerindeki interkalar disklerde ve düz kas hücrelerinin arasında bulunur. Adezyon
Kadherinler: Kadherinler aynı tip hücreleri bağlarlar (homofilik). Selektinler :Selektinler farklı tip hücreleri bağlarlar (heterofilik).Selektinler endotel hücresinde, lökosit, trombositlerde bulunur Lökositlerin kapille damar endotel bağlanmasında görev alır. Hücre göçünde, hücre tipinin belirlenmesinde kadherinler görev yapar. Kadherinler kalsiyuma bağlı reseptör proteinlerdir.
İntegrinler: Epitel hücrelerini, altlarındaki bazal membrana bağlayan adezyon molekülüdür. Bazal membrandaki laminin ve fibronektine tutunurlar.
1-Laminin :Glikoprotein
2-Tip IV kollagen
3-Entaktin : Glikoprotein
4-Perlekan : Heparan sülfattan zengin bir proteoglikan

H A Y V A N F İ Z Y O L O J İ S İ
50
molekülleri, hücrelerin özgül olarak dokulara yönlenmelerinde, birbirlerini tanımalarında, embriyogenez, hücre büyümesi, hücre farklılaşması ve inflamasyon gibi olguların düzenlenmesinde görev alırlar. Adezyon kuvvet molekülleri bugün dört sınıfta incelenmektedirler: (1)integrinler, (2)selektinler(P-selektin, E-selektin L-Selektin, (3)immünglobulin süper-ailesine dahil adezyon kuvvet molekülleri ve (4)kaderinler. Bir de fonksiyonel olarak adezyon kuvvet görevi gören ama yukarıdaki gruplar içerisinde sınıflandırılamayan adezyon kuvvet molekülleri vardır. İntegrinler: İntegrinler, heterodimer transmembran proteinlerdir. Aktif ya da inaktif halde bulunabilen integrinlerin, biribirine kovalent olmayan bağlarla bağlı alfa ve beta (ß) alt üniteleri vardır. Artıca bu molekülün fonksiyonel aktivitesi için her iki alt ünite de gereklidir, ancak bağlanma özgüllüğünün alfa alt ünitesi ile ilişkili olduğu düğünülmektedir. İntegrinlerin yapıları ve fonksiyonları iki-değerli katyonlara bağımlıdır (Ca2+, Mg2+). İntegrinler arginin-glisin-asparagin (RGD) aminoasit dizilerine sahip moleküllere bağlanma özelliği gösterir. Bu diziler hüce-dışı matriks glikoproteinlerinde, bazı hücrelerin yüzeyinde ve bazı kompleman proteinlerinde bulunur. Sitoplazmik kısımları ile vinkulin, talin, aktin, alfaaktinin, tropomiyozin gibi hücre içi iskelet yapıları ile etkileşirler. Dolaşımdaki lökositlerin damar endoteline tutunup yapıştıktan sonra, inflamatuvar reaksiyonun bulunduğu alana göç etmelerinde rol alırlar. Hücre dışı sinyaller aracılığı ile haberleşmeyi sağlarlar. İntegrin adı, bu moleküllerin hücredışı matriks ve hücre iskeleti ile ilgili aktivitelere aracılık etmesinden (integre etmesi) kaynaklanır. Embriyolojik geliğim, hemostazis, trombosis, yara iyileşmesi, immün ve immün-olmayan savunma mekanizmaları gibi birçok fizyolojik olayda hücre-hücre ve hücre-matriks adezyonuna katılırlar. Kardiyovasküler sistemde hücre-hücre ilişkisi dinamik bir olgudur ve ince ayarlı bir düzenleme gerektirir. Fibrinojen (2 mg/ml) varlığına rağmen trombositler agrege olmaz, kan akımına rahmen lökositler inflamasyon alanına gidebilir. Bütün bu olaylarda integrin grubu hücre yüzey molekülleri rol oynar. İntegrinler, insan vücudunda bulunan hemen tüm hücrelerde eksprese olurlar. Aktif hale geçen bir hücre sitoplâzmasından sinyal iletildi İnde, integrinlerin hücre-dışında kalan kısmı şekilde işlevini göstererek kendi ligandına olan afinitesini arttırır. Bu işleme içeriden-dışa(inside-out) sinyal iletimi denir. Bu işlem adezyon molekülleri arasında bir tek integrinlerde görülür. İntegrinlerin ligandına başlanması ile bu kez dışardan-içeriye (outside-in) sinyal mekanizması çalışır; bu da hücre içerisinde apoptozisten proliferasyona kadar birçok işlevde etkili olur. İntegrinler, liganlarının aviditesi yönünden düşük ve yüksek afiniteli durumda olabilirler. İntegrinler farklı yollardan aktive edilebilirler. TCR kompleksi veya protein kinazı C (PKC) aktive eden forbol esterler aracılığıyla �içeriden dışarıya doğru sinyal iletimi sa lanabilir. CD2, CD44 veya CD43’e karşı monoklonal antikorlar da C11/CD18 aktivasyonuna neden olurlar. Mg2+ ve Mn2+ ve bazı integrin başlayan antikorlar da (MEM83, KIM127, KIM18) hücre içi sinyali olmaksızın integrin aktivasyonu yapabilirler. Kadherinler: Hücre yapışmasında ve farklılaşmasında önemli rol oynar. Kanser hücrelerinde kadherinler kaybolur bu neden kanser hücreleri başka dokulara atlayarak çoğalmalarına devam eder. Kadherinler ailesi adhezyon moleküllerin işlevi yapabilmesi için kalsiyum iyonana ihtiyaç duyarlar. 40 çeşitten fazla kadherin molekülü vardır. En tipik kahdrein olan E-kadherin bir birine komşu olan iki hücre arasında aynı sınıf kahderinler homofilik bağlarla bir birine bağlanır. N-kadherin tipi merkezi sinir sistemi, göz merceği, iskelet ve kalp kasında bulunur. Selektinler: Kadherinler gibi sleketin ailesi adhezyon moleküllerin işlevi yapabilmesi için kalsiyum iyonana ihtiyaç duyarlar. Dolaşımdaki lökositler dokulara göçünde (diyapedezinde)

H A Y V A N F İ Z Y O L O J İ S İ
51
görev alır. P-selektin tip endotel hücreleri ve tranbosit membranlarında, E-selektin tipi epitelyum hücreleri membralarında, L-Selektin tipi ise lökosit membran zarlarında bulunur. İmmünglobulin süper-ailesi: Bir hücredeki İg super ailesi molekülü diğer hücrede bulunan aynı sınıf molekül ile bağlanabilir(homotipik bağlanma) En tipik İg super ailesi molekülü ICAM-1(hücreler arası hücre yapışma molekülü 1) ICAM-2(hücreler arası hücre yapışma molekülü 2), VCAM( vasküler hücre yapışma molekülü) NCAM( Nöral hücre yapışma molekülü). Fibro nektinler: Fibro nektinlerin esas görevi hücreleri matrikse bağlamaktır. Fibronektinler iki polipeptid zincirinden meydana gelmiştir. C-terminal uçlarından birbirleriyle disülfit bağları ile bağlanmıştırlar. Uzunluğu 60–70 nm, kalınlığı ise 2–3 nm kadardır. Üzerinde çok sıkı katlanmış globüler bölgeler vardır. Bu bölgelerle matrikste özellikle kollagen tip I, II, III ve V ' e, heparan sülfata, fibrin, denature kollagen ve hücre zarlarındaki integrinlere bağlanırlar. Vücudun çeşitli bölgelerinde 20 çeşit fibronektin izole edilmiştir. Bütün bu fibronektinler tek bir fibronektin geninin çeşitli (splicing) diziliş gösteren kalıplardan oluşturulan RNA'lardan sentezlenmiştir. Fibronektinler hücre göçünün yapılmasında da etkin moleküllerdir. Bu moleküller 6 kolu bulunan tenasin proteini ile özellikle gelişmekte olan organlarda epitel hücrelerinin altında yer alan mezenşim hücrelerinin bol olduğu ortamda birleşirler.
Hücre dışı matriks(Ekstrasellüler matriks)
Hücre dışı matriks pek çok işlevsel protein ve polisakkaritten oluşur (Glikoprotein, Proteoglikan). İçinde bulunduğu veya temas halinde olduğu hücreler tarafından salgılanır. Örneğin konnektif dokuda bu matriks makromoleküllerini (ESM) salgılayan büyük hücreler fibroblastlardır. Kemik ve kartilaj dokularında ise bu hücreler kondroblast, osteoblastlardır. Kendilerini üreten hücrelerin yüzeyi ile yakın ilişki içindedirler. Hücrelerin ve dokuların bir arada tutulmasına yardım eder. Hücreler arası boşlukları doldurur. Destek (Kemik ve kıkırdak) görevi vardır. Hücre yapısı ve hareketi ile hücre gelişim ve farklılaşmasında önemlidir. Besin ve madde alışverişi ve hücre yaşamının devamlılığında gereklidir. Ayrıca hücrelerin migrasyonunu ve birbirleri ile olan ilişkilerini organize eder.
Hücrelerde, N-bağlı şekerler proteinlere bağlanarak protein çeşitliliğine neden olurlar. Bu glikoproteinler, hücre zarının yüzeyine yerleşerek hücrelere yeni kimliklerini kazandırır. Bununla beraber N-glikosilasyonlu proteinlerin şeker profilinin uç kısmında bulunan şekerler, örneğin; sialik asitler hücrelerin birbirleriyle ve çevreleriyle iletişiminde önemli rol oynar. Negatif yüklü sialik asitler universal bir molekül olarak hücre yüzeylerinin özelliklerinin şekillenmesine katkıda bulunurlar ve hücrelerin çevreleriyle arasında birçok ilişkinin kurulmasında çok önemli rol oynarlar. Çok çeşitli hücresel tanıma olayları, hücre farklılaşması, pozitif yüklü iyonların taşınması, reseptörleri maskeleyerek asıl reseptörün tanınmasının engellenmesi ve onları bozulmaya karşı koruması bunlardan bazılarıdır. Hücre içinde veya yüzey elemanlarına bağlı sialik asitler, hücresel tanıma, tutunma, farklılaşma, yaşlanma gibi fizyolojik olaylarda yüzey değişikliklerinden ve hücrelerde bilgi iletimi gibi işlevlerle, hücrede birçok moleküler ilişkinin şekillenmesine katkıda bulunurlar. Hücre dışı matriks farklı moleküllerden meydana gelir.

H A Y V A N F İ Z Y O L O J İ S İ
52
Glikozaminoglikanlar (GAG); Hidrate, jelimsi
Hyalüronik asit, Kondroitin sülfat, Dermatan sülfat, Heparin ve
heparin sülfat, Keratan sülfat
Fibröz proteinler
Yapısal olanlar:
Kollajen, (Gerilme gücü)
Elastin (Matrikse esneklik)
Adezif olanlar:
Hücrelerin matrikse uygun yerlere tutunmaları Fibronektin, laminin, tenasin, entaktin, vitronektin, İntegrin,
Selektin, Kahderin, VCAM, ICAM, v.b
Diğer ECM proteinleri:
Fibrillin, fibulin, nudelin, netrin, galaktin, glutaktin, vs. En fazla bulunan hücre dışı protein %25 ile kollejendir. Kollajen (bazen kolajen veya
kollojen olarak da anılır) hareket sisteminin yapı taşlarını, özellikle kemik, kıkırdak, lif ve eklemleri oluşturan proteindir. Bu madde Fibroblast, Osteoblast, Kondroblast, Retiküler hücre, Epitel hücre, Endotel hücresi, Düz kas hücresi, Schwann hücresi tarafından sentezlenir. Bu protein birbiri üzerine sarılmış üç alfa zincirinden meydana gelir. 19 tane değişik tipi tanımlanmış olup, tip I, tip II şeklinde isimlendirilir. Bu çeşitlilik moleküler yapıdan kaynaklanmaktadır. Kollajenin ana molekülü tropokollajendir. Tropokollajenler de hücrenin içinde üretilen prokollajenlerden oluşur.
Fosfolipidlerin geçirgenliği
Hücre zarı, seçici geçirgen bir yapıya sahiptir. Molekülün büyüklüğüne, yağda veya suda çözünmesine, polaritesine, ortamdaki yoğunluğuna veya türüne göre zar üzerinden madde taşınmasını dört farklı şekilde gerçekleştirir. Basit pasif difüzyon da gazlar gibi solutler ve su kendi elektrokimyasal gradiyentleri doğrultusunda membranlardan kolaylıkla geçerler. Lipit çift tabaka iyonlar için geçirgen değildir, iyonlar sadece özel kanallar sayesinde hücre membranından geçebilir. Fosfolipidler, fosfogliseridler ve sfingomyelin olmak üzere ikiye ayrılır.

H A Y V A N F İ Z Y O L O J İ S İ
53
Fosfolipid Sfingolipid
ŞEKİL 1.38 Fosfolipit molekülden geçen maddeler ve diffüzyon
Hücre zarınında pasif difüzyonun diğer bir tipi ise kanallar aracılığı ile olur ve genellikle iyonlar bu yolu kullanırlar. Elektrolitlerin lipid çözünürlüğü kötü olduğu, suyla hidrojen bağı kuramadığı için membranın çift katlı fosfolipid tabakasından difüzyonu yoktur. Bu sebeple iyonların membrandan geçiş katsayıları düşüktür ve membrandan geçebilmek için bir kanala (protein yapısındaki iyon kanalları) ihtiyaç duyarlar. Sinir hücrelerinde bulunan ligand kapılı ve voltaj kapılı kanallar bu şekildedir
Membrane Transportu kullanılan farklı yollar temel olarak ATP kullanması yada kullanılmasına göre sınıflandırır. Diffüzyon, kolaylaştırılmış diffüzyon fitrasyon, osmoz ATP kullanılmadan membranda madde taşıma yolarıdır. Primer ve sekonder aktif taşıma, Ekzositoz, Endositoz ise ATP kullanarak yapılan madde taşır.
Transport Tipi Konsantrasyon farkına göre yön Enerji ihtiyacı
Basit difüzyon Aynı yönde Hayır
Kanal ve porlar Aynı yönde Hayır
Pasif transport Aynı yönde Hayır
Aktif transport Karşı yönde Evet

H A Y V A N F İ Z Y O L O J İ S İ
54
Basit Difüzyon
Molekül ve elementlerin sahip oldukları kinetik enerji yani Brown hareketleri sonucu yüksek konsantrasyonlu bir bölgeden düsük konsantrasyonlu bölgeye moleküllerin (veya iyonların) membranı geçerek transportudur. Net hareket düsük konsantrasyon yönündedir. Yüksek konsantrasyonlu bölgedeki moleküllerin düsük konsantrasyonlu bölgeye doğru hareket etme ihtimali, düsük konsantrasyonlu bölgedeki moleküllerin yüksek konsantrasyonlu bölgeye hareket etme ihtimalinden daha büyüktür. Bu nedenle zaman içersinde konsantrasyon farkı yönünde moleküllerin hareketi olacaktır. Bu hareket her iki bölgedeki moleküllerin sayıları esit oluncaya kadar devam eder ve sonuçta her iki bölgedeki madde konsantrasyonu esit olur. Bu hareketin hızı tamamen konsantrasyon farkıyla orantılıdır. Hidrofobik maddeler lipid membranları hidrofilik maddelere göre daha kolay geçerler. Hidrofobik ortamda yüksek erime özelliğine sahip olan bu moleküller (örn. gliserol) plazma membranını suda eriyen maddelere göre daha kolay geçerler. Bu kuralın istisnası su dur. Su son derece polar olduğundan gliserol için söylenenden daha düsük bir diffüzyon hızına sahip olmalıdır. Aslında suyun diffüzyon hızı lipid yapay membranda gliserolun 7 katı kadardır. Tabii hücre membranında ise suyun diffüzyon hızı gliserolun 100 katıdır (aquaporinler etkisi). Membran taşıma proteinden ve enerjiden(ATP) bağımsızdır. Sadece gradiente bağlı olarak geçiş olmaktadır. Taşınacak madde konsantrasyon gradienti ne kadar fazla ve ne kadar hidrofobik ise geçiş o kadar hızlı olmaktadır. Taşınacak molekül büyüklüğü, ne kadar küçükse, membran kalınlığı, ne kadar ince ise ve geçiş yüzeyinin büyüklügü ne kadar fazla ise difüzyon hızı o kadar artar.
En hızlı difüzyon gazlarda meyadana gelir, sıvılarda daha yavaş olur. Difüzyon katılarda da meydana gelebilir; bu, atomlara birbirlerinin etrafında hareket edip yer değiştirme olanağını sağlayan kristal kafesi kusurlarında oluşur. Bununla birlikte bunun oranı o denli küçüktür ki sıkı temas halinde iki katı cisimde yıllar sonra bile görünür bir karışmaya tanık olunmaz. Difüzyon oranı, konsantrasyon gradieninin fonksiyonudur, İki alan arasında konsantrasyon farkı ne kadar büyük olursa, yüksek konsantrasyonlu alandan düşük konsantrasyonlusuna moleküller o kadar hızla yayılır (difüze olur).
Farklı maddelerin difüzyon katsayıları farklıdır.

H A Y V A N F İ Z Y O L O J İ S İ
55
ŞEKİL 1.39 Diffüzyon bağlı olarak konsantrasyon değişimleri .
Yağı seven Lipofilik (hidrofobik yada başka bir ifade ile sudan kaçan) maddeler membranda daha kolay çözüldükleri için daha hızlı geçiş gösterirler. Bu nedenle gliserol, üre, yağ asitleri membranları daha kolaygeçer. Ancak membranda en hızlı difüzyona uğrayan molekül su molekülüdür (Ozmoz). Bir maddenin çift katlı lipid tabakadaki hareket hızını belirleyen en önemli faktörlerden birisi o maddenin lipiddeki eriyebilirliğidir. Örneğin oksijen, karbon dioksid, azot ve alkolün lipidde çözünürlüğü yüksektir. Böylece bütün bu maddeler çift katlı lipid tabakada doğrudan çözülürler ve sudaki eriyiklerinde olduğu gibi difüzyonu uğrarlar, bu maddelerin membrandan difüzyon hızlan, onların lipidde eriyebilirliği ile doğru orantılıdır. Filtrasyon
Bir zarn iki kesimindeki hidrostatik basınç fark nedeniyle sıvının membranda bulunan porlardan (deliklerden) zarın bir bölümünden diğer bölümüne geçişlidir. Özellikle kılcal damarlarda meydana gelen bu olayda süzülen maddenin miktarı ile ortamda bulunan basınç farkı ve hücre zarının geçirgenliği etkilidir. Böbrek glomerullerinde ve kılcal damarlarında plazmanın süzülmesi de buna güzel bir örnektir.
Bir membranın iki yüzü arasındaki hidrostatik basınç farkı nedeniyle, basıncın yüksek olduğu taraftan az olduğu tarafa doğru sıvı ve beraberinde erimiş küçük moleküllerin geçişine filtrasyon (süzülme) denir.
ŞEKİL 1.40 Filtrasyon
Difüzyon, Geçişme veya Yayılma olarak da bilinir, maddelerin çok yoğun ortamdan az yoğun ortama göçü. Fiziksel kimyada ise moleküllerin kinetik enerjilerine bağlı olarak rastgele hareketlerine denir. Difüzyon, maddenin bütün hallerinde farklı hızda ve özellikte görülür.

H A Y V A N F İ Z Y O L O J İ S İ
56
Vücutta filtrasyona örnek kapillerlerdeki ve böbreklerdeki taşıma olayları gösterilebilir. Kapillerlerdeki olay basınç farkı nedeniyle su ve suda erimiş partiküllerin damar dışına çıkışıdır (dokular arası sıvıya geçişidir). Filtrasyonda proteinler gibi büyük moleküller damar dışına geçemez. Fitrasyonda hücreler arası geçiş tipinde parasellüler geçiş önemlidir.
Taşıma proteinleri ile Kolaylaştırılmış diffüzyon
Bir molekülün yüksek konsantrasyonda bulunan bölgeden(mebran ile ayrılmış iki bölgeden) düşük konsantrasyon tarafına, plazma membranında bulunan protein tasıyıcı ile taşınması olayına verilen isimdir. Bu olay pasif bir olaydır yani enerji kullanılması gerekmez. Taşıma proteinleri ile yapılan kolaylaştırılmış diffüzyon konsantrasyon gradyanı yönünde yapılır. Üç temel özelliğe sahiptir:
Glikoz, laktoz, amino asid, nükleotidler ve gliserol v.s. gibi benzer moleküllerin herbiri için spesifik, özel tasıyıcılar vardır. Glikoz tasıyıcısı D-glikozu tasıyabilir, fakat L-glikozu taşıyamaz. Taşınacak molekül bağlandığı zaman tasıyıcı molekül yapısal bir değisiklik geçirmek üzere etkilenir. Böylece küçük molekülü membranın öbür tarafına geçirir. Plazma membranını geçerken hareket hızı, basit diffüzyondan daha hızlıdır. Belki de bu hücre içinde kullanılan, normal olarak membrandan çok düsük hızda diffüze olan hidrofilik moleküllerin transportu için gelismis bir mekanizmadır. Herhangi bir hücrede bir molekül veya iyon için sınırlı sayıda tasıyıcı vardır. Bütün tasıyıcılar bağlandığı zaman, transport hızı maksimum olur. Bu yüzden olay satüre olabilir özelliğe sahiptir. Transport kinetiği bir basit enzim için tanımlanan proseslere benzer. Bir tarafta konsantrasyon büyük olduğu zaman daha çok tasıyıcı bağlanacak ve diğer tarafa net hareket meydana gelecek. Konsantrasyon farkı sıfır olduğunda ise tasıyıcılar hala faaliyet gösteriyor olsalar da sonuçta içeriye ve dısarıya aynı hızda madde taşınır. Bu yüzden net diffüzyon gözlenmez. İnsülin ve epidermik büyüme faktörü gibi hormonlar normal olarak gözlenen diffüzyon hızından daha fazlasının olmasını sağlarlar. Örneğin glikozun hücre içersine girişini insülin hormonu hızlandırır(GLUT glikoz taşıma proteinlerinin hücre zarına yerleşmesi, yeni GLUT sentezinin yapılmasına bağlı olarak). Kolaylaştırılmış diffüzyonda eloktrokimyasal gradient söz konudur. Yani taşınacak molekül çok olduğu yerden az olduğu yere doğru taşınır. Kolaylaştırılmış difüzyon enerjiye(ATP) ihtiyaç duyulmaz. Glikozun hücre membranından geçişi kolaylaştırılmış transport ile olmaktadır(GLUT). Suyun difüzyonunda da aquaporin adındaki suya özel taşıma kanalları görev yapar. Organizmadaki diğer örnekleri demirin incebarsaktan absorbsiyonu ve plesentadan glikozun geçişidir.
Band 3 proteini: Eritrositlerin başlıca görevi akciğerlerden dokulara oksijen, dokulardan akciğere karbondioksit taşımaktır. Karbondioksitin oksijen ile değişimi bir anyon değiştirici transport proteini sayesinde olur. İşte eritrositlerdeki bu anyon transport proteinleri Band 3 proteinlerdir. Band 3 proteini de bir (multipass) transmembran proteindir. Kolaylaştırılmış difüzyonla iyon değişimi yapar(klor shift).
1) Taşıma proteinleri bir partiküler molekül için spesifiktir.
2) Kolaylaştırılmış difüzyon basit diffüzyondan daha hızlıdır.
3) Kolaylaştırılmış difüzyona taşınan madde kendine has olan tüm taşıma proteinlerini doldurmuş olabilir. Satüre olabilir(doyma kinetiği, Tm değeri vardır).

H A Y V A N F İ Z Y O L O J İ S İ
57
ŞEKİL 1.41 Membran madde taşıma yolları.
Taşınacak madde molekülleri hücre membranlarını bir özel taşıyıcı denilen özel trasportor ile kompleks yaparak hücre membranını geçerler. Taşıma işlemi taşınacak moleküle özel olduğundan bir tip taşıma molekülü yalnız bir molekülün kolaylaştırılmış difüzyona izin verir. Öğreniğin benzer yapıda olmasına rahmen glikoz taşıma proteinleri(GLUT5) bir tipi yalnız fruktoz taşıması yapar.
ŞEKİL 1.42 Kolaylaştırılmış difüzyonda da gözlenen transfermaksium Michael-mental kinetiği
Çoğu protein kanalı bir veya daha fazla sayıdaki iyon veya molekülün transportu için yüksek düzeyde seçicidir. Bu, kanalın çapı, şekli ve iç yüzeyindeki elektriksel yük gibi niteliklerin sonucudur. Hücre membranından çok büyük miktarda difüze olan madde sudur. Örneğin, su alyuvar membranından her iki yönde düzenli olarak difüze olur. Saniyede alyuvar membranından her iki yönde difüze olan su miktarı yaklaşık olarak hücre hacminin 100 katma eşittir. Normal ol arak her iki yönde difüze olan suyun miktarı öyle hassas ayarlanmıştır ki suyun net hareketi görülmez. Bundan dolayı, hücrenin hacmi sabit kalır. Buna karşın belirli bazı koşullarda tıpkı öteki maddeler için geliştiği gibi, membramn iki tarafı arasında su için de bir konsantrasyon farkı gelişir. Bu koşullarda, membranda suyun net hareketinin yönüne bağlı olarak hücre şişer ya da büzülür. Suyun konsantrasyon farkından doğan bu net hareketine osmoz denir.
Kolaylaştırılmış difüzyon bir taşıyıcı aracılığı ile gerçekleşir: (1) Taşınacak madde taşıyıcı proteine bağlanınca, taşıyıcı proteinde şekil değişikliği olur ve içte kapalı olan hücre kanalının ucu açılır. (2) Molekül buradan içeri girmeye başlar. (3) Proteine zayıf bağlandığı için hücre içine yakın bir yere geldiğinde, ısıdan kaynaklanan hareketle protein, molekülden ayrılır ve molekül hücre içine girer.

H A Y V A N F İ Z Y O L O J İ S İ
58
Kolaylaştırılmış difüzyon GLUT glikoz transporterları
Glukoz taşıyıcılar GLUT hücrelerin yüzeyinde bulunan ve kolaylaştırılmış difüzyon ile glukoz taşıyan protein ailesidir. Farklı tipleri olan bu proteinler hemem hemem tüm hücrelerde vardır.
• GLUT-1: Merkezi sinir sistemi(beyin ve omurilikte) ve eritrositler, plasentada yer alır. Bazal glukoz transportundan sorumludur.
• GLUT-2: Karaciğeri pankreasi ince barsak ve böbrek proksimal tüplerinde glukoz yakalama ve salınmasın görev alır.
• GLUT-3: Glukoz affinitesi en yüksek olan temel olana beyin nöronlarında olan taşıma proteidir.
• GLUT-4: Daha çok periferik dokularda (yağ doku ,kalp ve iskelet kası) bulunan İNSÜLİN’le uyarılabilen tek transport proteinidir.
• GLUT-5: Sindirim sistemin’de ve böbrekte fruktoz taşıması yapar.
ŞEKİL 1.43 Glikoz kolaylaştırılmış diffüzyon proteinleri GLUT taşıma sıstemi
Farklı tip GLUT olması sayesinde bazı hücrelerin örneğin beyin omurlilik ve sinir hücrelerinin insülin gibi glikoz hücre içine alınmasında düzenleme yapan hormonlardan(insülin, glokagon) bağımsız olarak alınmasına izin veririr Kolaylaştırılmış difüzyon Aquaporin su kanal proteinleri
Hücre içinde ve dışında su molekülleri canlı için hayati öneme sahiptir. Su basit difüzyonla hücre zarından geçer ayrıca su kanalları olarak da bilinen, aquaporinler gözenekli yapıya sahip su için özelleşmiş taşıma proteinlerdir. Bazıları, aquaglyceroporins olarakta bilinir. Bu taşıma proteinleri aynı zamanda por büyüklüğüne bağlı olarak, örneğin, zar boyunca gliserol, CO2, amonyak(NH3) ve üre gibi diğer küçük yüksüz eriyikleri taşıayabilir. Örneğin, aquaporin 3 kanal 8–10 A por genişliğine sahiptir ve 150-200 Da molekül ağırlığına sahip hidrofilik moleküllerin geçişini sağlar. Aquaporinler porları protonlar gibi hücre membranı elektrokimyasal potansiyelinin korunmasında kritik önemi olan moleküllerin karşı geçirgen değildir.

H A Y V A N F İ Z Y O L O J İ S İ
59
ŞEKİL 1.45 Aquaporin membran taşıma proteinleri 7 heliks yapısı
Memelilerde aquaporins on üç bilinen türü vardır ve bunlardan altısı böbrekte bulunur. En çok çalışılan aquaporinler AQP1, AQP2, AQP3 ve AQP4 dir. Su fosfolipid çift-katlı membran tapakasını aquaporinler adı verilen özel su kanalları aracılığıyla veya difüzyon ile geçer. Şimdiye kadar ondan fazlası memelilerde aquaporinler tespit edilmiş, fakat daha birçok varlığı kanıtlanmamıştır. Aquaporinler iyonları ve diğer küçük moleküllerin geçişine izin vermez buna karşın özel su kanalları olarak görev yapar. Bazı aquaporinler - aquaglyceroporins olarak bilinen - su artı gliserol ve birkaç diğer küçük molekülleri taşıma görevi yapabilir.
• Aquaporin 1: AQP1 böbrekte bulunan, yaygın olarak ifade edilen su kanalı proteinleridir. Böbrekte proksimal tübüllerde, Henle kulpunda vasa rektatda inen bölümü içinde bazolateral ve apikal plazma zarlarında bulunur. Buna ek olarak, kırmızı kan hücreleri, vasküler endotelyum, gastrointestinal sistemde, ter bezleri ve akciğer bulunmaktadır. Aquaporin 1 (ADH) vazopressin tarafından denetlenir.
• Aquaporin 2: AQP2 böbreğin toplama kanalı ana hücrelerinin apikal hücre zarlarında ve hücre içinde bulunan hücre içi veziküller içinde bulunur. Bu aquaporin tipi peptit hormonu olan vazopresinle iki yolla düzenlenir: bir artış yoluyla da apikal plazma membranı apikal bölgeye kayması, AQP2 veziküllerin hareketi sayesinde kısa süreli düzenleme (dakika), ve uzun vadeli düzenleme (gün) AQP2 gen ekspresyonu. Bu kanalda mutasyonlar otozomal dominant ya da resesif olabilir. Şekersiz diyabet ile ilişkilidir. Genellikle bipolar bozukluğu tedavi etmek için kullanılan lityum, AQP2 genin ekspresyonunu azaltarak diabetes insipidus neden olabilir. AQP2 geninin ekspresyonu, gebelik ve konjestif kalp yetmezliği gibi su retansiyonu ile ilişkilidir koşullarında artar.
• Aquaporin 3 ve 4: Aquaporinler ana toplama kanalı hücrelerin bazolateral hücre zarında bulunan ve bu hücrelerinden su çıkışı için bir yol sağlar. AQP4 yapısal olarak ifade edilir, buna karşın böbrekte, AQP3 gen ifadesi, (ADH) vazopresin ile düzenlenir. AQP4 astrositler olarak ifade edilir ve merkezi sinir sistemine doğrudan uyarılması regüle edilir.
Aktif- pasif Taşıma
Lipid zar yapısı hidrofilik ve iyon yada büyük maddelerin geçisi izin vermez. Buna karşın hücrenin metabolizması için gerekli olan maddelerin uygun şekilde alınması gereklidir. Bunun yanında hücre içi ve dışı madde yoğunlukları difüzyon yada kolaylaştırılmış difüzyona izin vermiyecek durumda olabilir. Bu noktada hücre aktif taşıma yapar. Aktif taşıma ATP enerjisi

H A Y V A N F İ Z Y O L O J İ S İ
60
kullanarak madde taşınması yapılmasıdır. Şekerler ve amino asitler polar (kutuplu) moleküllerdir. Yağ çözülmezler yada yağı çözmezler. Ayrıca çapları 8 A°'dan büyük olduğu için normal yollardan hücre zarını geçemezler. Halbuki, bu iki maddenin de hücre zarını bolca geçmeleri gerekir. Çünkü hücrenin enerji üretimi ve protein sentezi yapabilmesi için bu iki maddeye İhtiyacı vardır. Öyleyse, başka bir mekanizmayla hücre zarını geçmeleri gerekmektedir. Yapılan deneyler, bu maddelerin zar yüzeyinde özel yerlere bağlandığım ve zarın dış yüzünden iç yüzüne (hücre içine) taşındığını göstermiştir. Bu tür taşınma şekline taşıyıcılar aracılığıyla taşınma denir. Çünkü hücre zarında bulunan taşıyıcı moleküller, taşımaya aracılık ederler. Bu yolla zarı geçen maddeler kimyasal özgüllükleri, rekabet ve doymuşluk yaratma özellikleriyle basit difüzyonla geçen maddelerden ayrılırlar.
•Sodyum-potasyum pompası, özellikle sinir hücrelerinde yaygındır. Sinir hücrelerinde, Na-K yoğunluklarındaki değişliklikler elektriksel yük meydana getirirler. Hücre zarı boyunca Na ve K’un sürekli diffüzyonu ve sızması, Na - K pompası ile gerçekleşir. Diğer önemli bir fonksiyonu hücrelerin su alarak patlamaları engelenmesidir.
•Kalsiyum pompası, kas kasılmasında kalsiyum iyonlarının taşınması için gereklidir. Kalsiyumun hücre içinde ikincil haberci olarak görev yapmasından dolayı hücre içi kalsiyum seviyesinin kontrolü için hem ER hem mitokondrilerde hem de sitoplazma membranında bol miktarda kalsiyum pompası bulunur. Bu pompalara SERCA(Sarco(Endo)plasmic Reticulum Ca++-ATPase. ) adı verilir(P sınıfı ATPaz).
• Sodyum bağı kotransportta, şeker ve aminoasitler aktif olarak taşınırken Na iyonları da pasif olarak eşllik eder. Kısaca Na yoğunluğa bağlı olarak membrandan taşınması sırasında buna bağlı olarak şeker ve aminoasitlerin Na bağlı olarak taşınır. Na farkı Na-K ATPaz tarafından sağlanır.
• Hidrojen bağlı kotransportta ise hidrojen iyonları bağlanırken, şekerler aktif olarak taşınır. Diğer bir hidrojen pompası yada proton pompası midede potasyum hidrojen pompasıdır.
ŞEKİL 1.46 Aktif taşıma sistemleri

H A Y V A N F İ Z Y O L O J İ S İ
61
Tüm aktif taşıma sistemlerinde bulunan temel özellikler:
1-Özgüllük: Aktif taşıyıcılarla taşınmada, taşıyıcıların bazı kimyasal gruplara özgü olması özelliğidir. Örneğin şeker taşıyan sistem, amino asitleri taşımaz. Bazı sistemler o kadar özgüldür ki aynı atom sayısı ve kimyasal grupları taşıyan iki molekülden birisinin molekül biçiminde küçük bir değişiklik olsa, bunu ayırt edebilmekte ve o molekülü taşımamaktadır. Aktif taşıma sistemleri taşınacak maddelere özeldir.
2-Doymuşluk(micheal mental enzim kinetiği) Basit difüzyonda, hücre dışında bulunan ve difüzyonla hücre içine girecek olan maddenin dıştaki yoğunluğu ne kadar artarsa, hücreye giren madde miktarı da o kadar artar. Aktif taşıyıcılarla olan taşınmadaysa maddenin hücreye girişi, dıştaki yoğunluğun bağlı değilidir. Örneğin midede hidrojen iyonu yoğunluğu mide lümeninde salgı yapan hücreye göre binlerce kat fazla olabilir buna rağmen aktif taşıma sistemleri ile hidrojen iyonları lümene gönderilebilir.
3-Rekabet: Birbirine benzer iki madde, aynı taşıyıcı sistemle taşınıyorsa birbiriyle rekabet eder. Örneğin amino asitlerden alanin ve glisin aynı taşıyıcıyı kullanırlar, yani aralarında rekabet vardır. Benzer olarak nötür amino asitler arasında aynı taşıma proteinine bağlanmak için rekabet oluşur. Rekabet taşıma sistemin bağlanacak kadar benzer maddeler arasında gözlenir.
4-Enerji kullanımı: Aktif taşıma sistemleri ister direk ATP kullansın(ATPaz) İsterse na bağlı sekonder aktif taşıama olduğu gibi Na yoğunluk farkından faydalansın(bunu içinde Na-K ATPaz gerekir) enerji yani ATP kullanılması gerekir.
Zarda bulunan taşıyıcı moleküllerle konsantrasyon farkından bağımsız olarak birim zamanda yalnızca bir molekül ya da iyonun sitosolden içeri ya da dışarı doğru taşınması üniport taşıma olarak tanımlanır. Memeli hücrelerinde kalsiyum taşınması buna bir örnektir(SERCA)
Aktif taşıma için taşınacak madde konsantrasyon farkı zorunluluğu yoktur. (madde konsantrasyon yönününe zıt olarak taşınabilir). Aktif taşıma da görev yapacak proteinler taşınacak molekülü membran içine özel proteinler(taşıma proteinleri) kullanarak taşır.
1) ATP-az’lar: Na+/K+ transport sistemi(Na-K ATPaz) dışında en önemli transport sistemi kas liflerinin sarkoplazmik retikulumun mebranında yer alan Ca+2-transport sistemidir (SERCA). Bu sistem kasın relaksasyon(gevşeme) zamanında etkili olur ve Ca2+’u hücre içindeki küçük vesiküler boşluklara kaldırır. Bunu hücre stoplazmasıyla vesikül içi arasındaki büyük kalsiyum konsantrasyonu farkına rağmen yapabilir (Ca2+ plazma = 10-7 mol/L, Ca+2 vezikül= 10-3 – 10-2 mol/L). Aktif taşıma yapılan transport ATP’nin parçalanmasından açığa çıkan metabolik enerji ile gerçeklesir. Bir ATP molekülünün hidrolizi ile 2 mol Ca2+ vesiküle alınır. SERCA ile klasiyum pompalanması için Mg2+’un varlığına ihtiyaç vardır. SERCA fonksiyonları idlerin varlığında fonkCa2+ ve Mg2+ tarafından stimüle edilir. Ca2+ / Mg2+ ATP-az , Na+/K+ ATP-az gibi sadece lipsiyon görür. Hücre membranlarından özellikle eritrositlerden izole edilmistir. Ca2+ / Mg2+ ATP-az ’a ek olarak mide, böbrek ve bezlerin hücre membranlarında ATP -az’lar da vardır. Bu ATP- az’lar HCO-
3 veya K+ tarafından aktive olurlar.

H A Y V A N F İ Z Y O L O J İ S İ
62
3) Aktif H+ transportu (Redox sistem): Mitokondriler hücre solunumunun gerçeklestiği yerlerdir. Redoks reaksiyonları mitokondirinin içinden dışına aktif H+ transportunu sağlar. Bu transport mitokondri membranının iki tarafı arasında bir elektrokimyasal potansiyel farkının oluşmasıyla sonuçlanır. Bu potansiyel proton geri akısı olarak ATP sentezini yönetir. Oksijenli solunum sonunda ATP sentezin en fazla yapıldığı basamak mitokondri zar arasında elektrokimyasal gradiye bağlı proton taşıması sırasındadır (Kemosmotik ATP sentezi). Mitokondri içindeki H+ iyonlarının dısa transportuyla dış fazın baslangıçtaki asitlik özelliği değişir. Membranın lipid fazı Triton 100 gibi deterjanlarla parçalanırsa asitlikte değişiklik olmaz. H+ iyonlarının transport hızı direkt olarak oksijen kullanımıyla orantılıdır. Bir oksijen atomu için 6H+ iyonu transfer edilir. ATP - az inhibitörler bunu etkilemezler. Fakat solunum zincirini etkileyen zehirler değistirebilir. H+ iyonlarının dışa tasınması bir membran potansiyeli meydana getirir(dış pozitif, iç negatif). Membran potansiyelini ölçmek için mitokondiriler çok küçüktür.
Simport-Antiport aktif taşıma yolları
Simporta iki madde aynı yönde taşınırken antiport bir birine zıt yönde iki madde taşınması yapılır. Simport–antiport taşıma yollu mekanizması pasif taşınmanın tersinedir. Taşınacak molekülün düşük konsantrasyondan yüksek konsantrasyona doğru, bir taşıyıcı molekül aracılığıyla taşınmasıdır. Yokuş yukarı taşınma diye tabir edilen bu sistemde, enerjiye ihtiyaç duyulur. Enerji ihtiyacı, ATP molekülünden sağlanır. Bu sistemde moleküller genellikle tek yönde taşınır. Bazı sistemlerde, taşıyıcı molekül taşınacak maddeyi, her iki yönde de taşıyabilir. Ancak, taşıyıcının taşınacak moleküle karşı affinitesi (birleşme isteği, meyli) bir tarafta yüksek, öteki taraftaysa düşüktür. Taşınmanın bir yöne doğru olması ve yoğunluğun bir tarafta yüksek tutulabilmesi için bu şarttır.
ŞEKİL 1.47 Uniport ve kotransport yolu.
Böbreklerde ve sindirim kanalında glikoz aktif rezorpsiyonu sekonder aktif transporta örnektir. Hücrenin diğer yüzeyinde(bazal membran tarfında) aktif Na+/K+ pompası vardır. Bu mekanizma Na+ iyonlarını hücrenin iç kısmından dış kısmına(kapiller damara) taşır. Bu yüzeyde ise glikoz için pasif bir transport mekanizması(GLUT) vardır. Sodyum-glikoz Co-transport mekanizması lümene ait hücre membranlarında(Epitel hücrelerin lümen yüzeyinde) bulunmuştur. Protein yapısındaki taşıma proteinleri inhibitör maddelerle durdurulabilir. Örneğin Oubain Na+/K+ ATP az pompasını inhibe edebilir. Bazal lamina kısmında bulunan (Kontraluminal tarafında) hücre membranınında eritrositlerde tanımlanana benzer sekilde glikoz transport mekanizması, floretin tarafından inhibe edilebilir. Luminal hücre membranlarının Na+-glikoz Co-transport mekanizması florizin tarafından inhibe edilir.

H A Y V A N F İ Z Y O L O J İ S İ
63
Sodyum iyonları, yeterli glikoz molekülleri varsa elektrokimyasal potansiyel gradyanı boyunca böbrek veya sindirim sistemi lümeninden hücreye girer. Sodyum, hücre dışında içerideki miktarından 10 kat fazladır. Potasyumsa, hücre içinde dışa göre 30-35 kat daha fazladır. Bu iyonların hücre içinde ve dışındaki yoğunluk farkı, aktif taşınmayla muhafaza edilir. Na-K pompası, bu sistemle çalışır. Taşıyıcı molekül, hücre içinden 3 mol sodyumu hücre dışına taşır. Arkasından da hücre dışından 2 mol potasyumu hücre içine taşır. Tüm bu olaylar için 1 mol ATP kullanılır.
Na bağlı glikoz taşıma proteini bir glikoz molekülü bir veya iki sodyum iyonuna bağlanır. Böylece intrasellüler glikoz konsantrasyonu, kan tarafındaki diğer ortamdakinden daha fazla oluncaya kadar artar. Glikozun kolaylaştırılmış difüzyonla taşınması mümkündür. Bu durumda glikoz molekülleri pasif mekanizma ile kana geçebilir(GLUT). Glikozun lümenden kan tarafına taşınması devamlıdır. Hücrenin her iki tarafındaki sıvı hacimleri içersinde mutlak olarak aynı bileşim olsa bile bu işlem sürer. Sodyum transportu azalırsa, mesela oubain ile o zaman glikoz transportu da azalır. Glikoz konsantrasyonu azalırsa, sodyumun aktif transportu da azalır. Sekonder aktif transport esas olarak tüm epitelyal dokularda bulunur. Böbrekte sekonder aktif transport mekanizması bikarbonat, protonların sekresyonu ile beraber glikoz, amino asit, monokarboksil ve dikarboksil asitler, safra asiti, fosfat, sülfat ve kalsiyum iyonlarının rezorpsiyonundan sorumludur. Glikozun sodyuma transpotunda SGLT denen transtport proteinleri bu taşımada görev alır. İntraselüler ortama giren sodyum daha sonra sodyum-potasyum ATPaz pompası ile hücre dışına atılmaktadır. Oral rehidratasyon(su kazandırma) sıvılarında sodyum ve glukoz beraber verilmektedir. Çünkü sodyum ve glikoz birbirlerinin emilimini incebarsak düzeyinde kolaylaştırırlar. Sodyum ve glikoz böbrek tübüllerinde de geri beraber emilir. Diabetik hastalarda filtratta glikoz fazla olması nedeniyle daha fazla olan glikozun geri emilimi için daha çok sodyum ve bunla beraber su emilir.
Primer aktif taşıma Na-K ATPaz
Sodyum-potasyum pompası (Na+- K+) sodyum iyonlarını hücre dışına taşırken, aynı anda potasyum iyonlarını da hücre dışından içine taşır. Bu pompa vücuttaki bütün hücrelerde vardır. Pompa, hücre içinde negatif bir elektiriksel potansiyelim oluşmasının yanı sıra, sodyum ve potasyum konsantrasyon farklarının korunmasından da sorumludur.
�
Sodyum-potasyum pompası:
o Sodyum-potasyum pompasında hücrenin içine doğru çıkıntı oluşturan tarafında sodyum iyonlarının bağlanması için üç reseptör bölgeye sahiptir.
o Dış tarafında potasyum iyonları için iki reseptör bölgeye sahiptir.
o Proteinin iç tarafında sodyumun bağlanma yerlerine yakın ya da ona komşu bir bölge ATP-az aktivitesi gösterir.

H A Y V A N F İ Z Y O L O J İ S İ
64
ŞEKİL 1.48 Na-K ATPaz.
Na-K ATPaz membran dinlenme potansiyelinin oluşturur. Hücreden ne iyon çıkışı(katyon çıkışı) sağlar. Na-K ATPaz çalışmaması sonunda hücre içi iyonların bağlı olarak su girişi ile hücre patlar. Primer aktif taşıma Ca ATPaz
Kalsiyum (Ca 2+ ) hücre içinde sekonder haberci olması gibi birçok önemli rolleri vardır. Serbest kalsiyum bezlerden salgılama için, çizgili kas hücrelerinde kasılma yada döllenmiş yumurta tetiklenmesinde görev yapabilir. Serbest kalsiyum hücre içinde düşük konsantrasyonları muhafaza edilmesi için kalsiyum pompaları çalışır. İskelet kas sarkoplazmik retikulumun zarında bulunan ATPaz ve sitozol içinde, hücre membranda bulunan kalsiyum pompaları kalsiyum hücre içinde Sarkoplazmik retikülüm yada ekstra sellüler sıvı içine pompalar. Bu pompalama işi primer aktif transport mekanizmasına örnektir. Kalsiyum iyonları normal olarak vücuttaki tüm hücrelerde intraselüler sıvıda gerçekten çok düşük konsantrasyonda, ekstrasellüler sıvının 1/10.000 oranında bulunur. Bu amaçla iki kalsiyum pompası çalışır. Bunlardan biri hücre membranındadır ve kalsiyumu hücreden dışarıya pompalar. Diğeri, kalsiyum iyonlarını, kas hücresinin sarkoplazmik retiküluma veya tüm diğer hücrelerde mitokondriler gibi bir veya birden fazla vesiküler organelin içine pompalar. SERCA, yada sarko/endoplasmik retikulum Ca2+-ATPaz, SR Ca2+-ATPaz, ATP varlığında sitoplazmadan endoplasmik retikulum kalsiyum iyonu pompalayan primer aktif taşıma yapan bir taşıma sistemdir.
Primer-aktif taşıma sistemleri: • Proximal tübülde Na+’un geri emilimi, • Distal tübülde Na’un geri emilimi ve • Kas sarkoplazmasında Ca2+’un geri emilimi aktif taşıma ile yapılır

H A Y V A N F İ Z Y O L O J İ S İ
65
ŞEKİL 1.49 Ca taşınmasın kullanılan yollar.
Sekonder Aktif Taşıma Na -Glikoz ve Amino asit koportu
Metabolik falliyetlerin düzenlenmesinde anahtar rol üstenen iki önemli molekül olan Glikoz ve aminoasitlerin çoğu hücrelerin içine büyük konsantrasyon gradyanlarına karşı taşınır. Yani hücre içinde yüksek konsantrasyonda olmalarına karşın ekstrasellüler sıvıdan ATP kullanılarak hücre içine pompalanırlar. Bu taşıma maddelerin taşıma mekanizması sekonder aktif taşımadır.
ŞEKİL 1.50 Sekonder aktif taşıma ile glikozun barsak epitelinde emilimi
Taşıyıcı proteinlerin, dış tarafta, biri sodyum, diğeri glikoz için olmak üzere iki bağlanma yerine sahiptir. Aynı zamanda sodyum iyonlarının konsantrasyonu dışarı da çok yüksek, içerde çok düşüktür. Bu taşınma için gerekli olan enerjiyi sağlar. Taşıyıcı proteinin spesifik bir özelliği glikoz molekülü bağlanıncaya kadar sodyum iyonlarının içeriye hareketine izin veren konumsal değişikliğinin ortaya çıkmamasıdır. SGLT taşıma proteinleri glikoz sodyum simportu ile lümenden glikoz alır. SLGT2, Sodyuma bağımlı glikoz cotransporters (ya da sodyum-glukoz bağlanmış taşıyıcı , SGLT ) ailesidir glükoz taşıyıcıdır.

H A Y V A N F İ Z Y O L O J İ S İ
66
Bağırsak mukoza ( ince barsak bölgesinin) ve nefronlarda proksimal tübül içinde bulunur. böbrek glikoz geri emiliminin katkıda bulunur. Böbreklerde glikozun % 100 kısmını geri emilir. Glomeruluslarda glikoz % 98 nefron boyunca SGLT2 ile yeniden absorbe edilir. Çok yüksek plazma glukoz konsantrasyonu (hiperglisemi durumunda ), glikoz idrarla atılır. (glikozüri) SGLT2 sodyum ile glikoz arasında 4900 kadar çalışırken SGLT1 bu oran 70 dir.Barsak hücresinden kana glikoz verilmesi, başka bir taşıma proteini (kolaylaştırılmış difüzyon yapan GLUT) tarafından yapılır.
ŞEKİL 1.51 Barsakta kullanılan kotransport yolu.
Ekzositoz
Proteinlerin ve büyük moleküllü maddelerin transportu için özel transport mekanizmaları vardır. Taşınacak olan madde membranın bir parçası ile paketlenir. Hücre içersine alınırsa endositoz veya önceden oluşmuş ve hücre duvarına temas ile icindekileri hücre dısına boşaltırsa ekzositoz adı verilir. Bu şekilde proteinler veya daha büyük partiküller, hücre parçaları veya bakteriler sindirim için hücre içersine alınır. Bunun yanında hücrede üretilen hormon ve enzimler de dıştaki ortama serbest bırakılırlar..
ŞEKİL 1.52 Endositoz ve Ekzositoz.
Bağlanma olayı canlı membran materyelinde lokal formasyonunu tetikler. Kesin olarak istirahat durumundaki lipid komposizyonu ve protein dağılımı farklılaşır. Yeni oluşan hücre membranının iki lipid tabakasında yüzey gerilimin değişmesinden dolayı membranın invaginasyonu görülür. Bu vesiküller hücre duvarından ayrıldıktan sonra lizozomlar tarafından sindirilir. Hücre içinde sentezlenen çeşitli maddelerin hücre dışına çıkartılması olayına

H A Y V A N F İ Z Y O L O J İ S İ
67
sekresyon denir. Ökaryotik hücrelerin birçoğu sekresyon yapabilirler ve sekresyon hemen her zaman ekzositoz yolu ile yapılır. Hücre dışına çıkartılan maddelerin üç yoldan birini izler. Bunlar ya ekzositoz oldukları hücrenin yüzeyine tutunurlar, ya ekstraselüler matrikste kalırlar ya da kan dolaşımına katılırlar.
ŞEKİL 1.53 Endositoz ve Ekzositoz.
Endositoz
Birçok hücrede endositoz sürekli olarak meydana gelir ve bu tip endositozda ekstraselüler matriks ve bunun içinde çözünmüş maddeler, veziküller içinde hücreye girerler. Buna sıvı-faz endositozu (pinositoz) denir. Başka bir tip endositoz ise ancak hücre endositoz için uyarıldığı zaman görülür. Buna reseptör aracılı endositoz denir. Pinositoz, çok büyük moleküllerin hücre içine girebilmesinin tek yoludur, örneğin proteinlerin çoğu bu yolla hücre içine alınır. Bu tür moleküllerin hücre zarıyla temaslarının artması pinositik vezikül oluşumunu artırır. Pinositotik veziküllerin oluşma hızı bu tür moleküller membranın dış yüzüne değince artar. Bu moleküler genellikle absorbe edilecek proteine- reseptöre bağlanır, reseptörler hücre membranının dış yüzeyinde kaplı (örtülü) çöküntü (ceplerde) yoğunlaşmış olarak bulunurlar. Bu ceplerin hücre içine bakan bölümünde Klathrin adı verilen bir fibriler protein aktin ve miyozin ve dyamin gibi kontraktil bir ağ oluşturur. Reseptöre bağlı endo sitozda görev alan üç tip membran bağlanma molekülü önemlidir.
• Klatrin kaplı veziküller : Hücre dışından endositoz ile alınan vezikülleri kaplarak lizozomlara yada , golgi aygıtına ve/veya GER taşınmasını sağlar.
• COP-1(Coated protein-1) kaplı vezikül: Golgi aygıtından çıkan vezikülleri kaplayarak endoplazmik retikuluma yada lizozoma taşınmasından sorumludur.
• COP-2(Coated protein-2) kaplı vezikül: Endoplazmik retikulumda yapılan veziküllerin golgiye taşınmasından sorumludur.

H A Y V A N F İ Z Y O L O J İ S İ
68
Reseptöre bağlı endositozla LDL, B12 vitamin alınması ve transferin ile demir aktarılması yapılabilir
ŞEKİL 1.54 Klatrin kaplı çukurlarla endositoz
Klatrin, hücre dışı veziküllerin belirli bir şekle kavuşması için rol oynayan temel proteinlerdir ve sitoplazmada küçük vezilküller oluşmasını kolaylaştırır. Protein molekülleri reseptörlerine bağlandığı zaman membıamn yüzey özellikleri, tüm cebin hücre içine doğru çökmesine neden olacak biçimde değişir. İçe çöken cebin çevresindeki proteinler, az miktarda ekstıaselüler sıvıyla birlikte reseptöre tutunan proteinlerin etrafının sarılmasını sağlar.
ŞEKİL 1.55 Ekzositoz ve endositoz.
Reseptörle sağlanan endositoz; LDL’lerin hücre içine alınmasında, epidermal büyüme faktörünün hücreye alınmasıda, sinir büyüme faktörünün hücreye alınmasında, difteri toksininin hücreye alınmasında, virüslerin hücreye alınmasında görev(HIV virüsünün T lenfoistlerin CD4 reseptörlerine bağlanması) alır.

H A Y V A N F İ Z Y O L O J İ S İ
69
Membran reseptörleri
Farklı hücre tipleri farklı tip membran reseptörleri vardır İyon Kanalları reseptörler ligant varlığında etkidir örneğin Asetilkolin, muscarinic ve nicotinic receptörler üzerinde etkilidir. GABA: Beyin ve spinal kord da nörotransmitter inhibitörüdür ve Glutamat: MSS üzerinde depolarizasyon, Na+, K+ and Ca2+ iyonlarına karşı geçirgenliğin artırılması sağlar. Enzimatik aktiviteye sahip reseptörler G protein dir bu reseptörler Beta-adrenergik reseptörler ve Alpha -adrenergik reseptörlerdir. Diğer grup membran reseptörleri ise Integrin reseptörleridir. Bu reseptörler hücre adesyon görev alır.
Pankreastan beta hücrelerinden salgılanan insülin, ve IGF-1 (insülin benzeri büyüme faktörü), farklı hücrelerden sentezlenen büyüme faktörleri (EGF, FGF, PDGF gibi growth faktörler) tirozin kinaza bağlı reseptörleri kullanırlar. İnsülin tirozin kinazın beta kuyruğunu aktive eder. Hipofizden salgılana büyüme faktörü ise JAK-STAT’ı sekonder haberci sistemeni kullanır.
ŞEKİL 1.56 Membranda buluna hormon reseptörleri
Hormon bağlanmış triozin kinaz bir seri protein üzerinde fosforilasyon şelalesi oluşturarak tek bir moleküle karşın 106 kadar değişime uğramış aktif madde sentezine yol açabilir. Bu yola çok küçük miktardaki haberleşme molekülleriyle(sinyal moelkülleri yada hormonlar) büyük miktarda etki yapmak mümkündür. Fosforilasyon şelalesi sinyal molekülü varken hızlı ve artılmış bir yanıt oluşumu imkânı verir. Benzer şekilde sinyal molekülünün( yada hormonun) ortamdan uzaklaşması, yıkılması durumunda hücresel yanıtın hızla durmasına olanak tanır.
Bir hormona veya nörotransmitter'e duyarlı reseptörlerin sayısını artırarak veya azaltarak hücreler bu moleküle olan duyarlılıklarını ayarlayabilirler. Bu bir geri besleme mekanizmasıdır. Bir uyarıya uzun süreli veya tekrarlı maruz kalmak çoğu zaman bir reseptörün o uyarana düşük tepki vermesine yol açar, buna reseptör duyarsızlaşması denir. Reseptör duyarsızlaşması (desensitizasyonu) liganda olan afinitenin azalmasına yol açar. Ligand-bağlı reseptörlerin duyarsızlaşması (desensitizasyonu) ilk kez nikotinik asetilkolin reseptörlerinde gösterildi. Duyarsızlaşma aşağıdaki şekillerde olabilir: Reseptör fosforilasyonu. Reseptör effektör moleküllerinin ayrışması.
İnsülin hücre dışında yer alan zar proteinlere bağlanır. İnsülin bağlanması Triozin kinaz bir araya gelmesine yol açar ve bu durumda oluşan dimer aktif hale gelir. Aktif forma gelen triozin kinaz fosforilazyon şelalesi meydana getirirek İkincil habercilerde etkisini gösteririr.

H A Y V A N F İ Z Y O L O J İ S İ
70
Sitoplazma Hücrenin esasının protoplazma adı verilen pelte kıvamında kütle oluşturur. Hayat bu kütle içerisinde meydana gelen fizikokimyasal reaksiyonlar sonucu devam eder. Hücrenin yapısına Proteinler, karbonhidratlar, yağlar en fazla miktarda ise su katılmaktadır. Ayrıca Na, Ca, K, P gibi mineraller, enzimler, vitaminler, hormonlar da katılır.
Hücrenin metaboliz faaliyetlerinin gerçekleştiği yerdir. Sitoplazma, farklı büyüklükte ve dağınık durumda partiktül ve organelle doludur. Partiküllerin içinde dağıldığı sitoplazmanın berrak sıvı kısmına denir. Bu sıvıda çözünmüş proteinler, glikoz ve elektrolitler (Ca, K, Mg) bulunur. Hücrenin önemli metabolik faaliyetleri (hücre hareketi kasılması, glikolitik evre, hücrenin bölünmesi ve enerji dönüşümleri gibi) sitoplazmik matrikste yer alır.
Sıvı kısım sitozol
Sitozolun büyük kısmını (%90) su oluşturur. Bu oran bazı canlılarda %98’e kadar yükselebileceği gibi, sporlarda ve tohumlarda %5-15’e kadar düşebilir. Sitozolda organik ve inorganik (kuru madde) maddelerin oranı %10–40 arasında değişir. Kuru maddelerin %90’ını organik,%10’unu da inorganik maddeler oluşturur. Sitozolda en çok bulunan kuru madde protein molekülleridir. Ayrıca sitozolda; yağ, vitamin, hormon, organik ve inorganik asitler bulunur. Sitozolda bulunan önemli inorganik maddeler Na, Ca, K, P, Mg, Fe’dir. Bu elementlerin hücredeki fonksiyonlarını şöyle özetleyebiliriz: Bazı biyo-moleküllerin yapısına girerler. Örneğin Fe hemoglobinin yapısına katılır. Osmotik basıncın oluşmasını yani hücrede belli bir yoğunluk oluşturarak, suyun hücreye girmesini sağlar. Düzenleyici olarak görev yaparlar. Yarı akışkan, yoğun bir maddedir. Hücre sudan yoğun olup suyun içine atıldığında dibe çöker. Yarı akışkan bir sıvıdır. Sitoplazmanın sıvı kısmına sitozol denir. Sitoplazmanın % 90’ını su oolluuşşttuurruurr..
• Sitoplazma protein, karbonhidrat, yağ, vitamin, mineral, enzim ve su oluşmuştur.
• GGöörreevvii:: Biyokimyasal reaksiyonlar için zemin oluşturmak
• Organellere yataklık etmek
• Organellerin hareketini sağlamak
Bu bölüm için Power
point sunumu adresi:
https://yadi.sk/d/IS-
qtjkIKTmpy

H A Y V A N F İ Z Y O L O J İ S İ
71
Sitoplazma yer alan proteinler globuler ve ipliksel yapıdadırlar. Bu proteinler hücrenin gereksinimine göre iplikçikler (mikroflaman) yada borucuklar oluştururlar.
• Mikroflamanlar: ipliksel ve globuler proteinlerden oluşur(aktin). • Mikrotübülüslar: globuler protein içerir(alfa ve beta tübülin).
Globuler proteinler lipidlerle işbirliği yaparak membranları oluştururlar. Bu tür lipopratein yapıdaki membranlara unit membran- elementer membran adı verilir. Endo plazmik Retikulum
Endoplazmik retikülüm agranüler ve granüler olmak üzere hücre sitoplazmasının her tarafında yaygındır ve kanalcıklar, veziküller ve keseler gibi yapılardan oluşmuştur. Intrasellüler kalsiyumun en yüksek olduğu organeller endplazmik retinakulum ve mitokondridir. Endoplazmik retikulum membranında sitokrom redüktaz ve glukoz fosfataz gibi enzimler bulunur. Granüllü endoplazmik retikulumun görevleri; kollajen, pıhtılaşma proteinleri, serum albunini ve immünglobulinler gibi hücre dışına gönderilecek maddelerin sentezidir. Granülsüz endoplazmik retikulumun steroid hormon sentezi, ilaç defoksikasyonu, kas hücrelerinde Kalsiyum regülasyonu gilokojen ve lipid metabolizmasında fonksiyonu vardır. Özellikle steroid sentezi yapan adrenal kortex hücrelerinde çok gelişmiştir. Endoplazmik retikulum lipid, protein (ribozomlar aracılığı ile) ve kompleks karbonhidratların yapım yeridir. Endoplazmik retikulum hücredeki toplam membranların yarısından fazlasını oluşturur. Endoplazmik retikulum iki membrandan oluşur, iki membran arasında kalan boşluğa endoplazmik retikulum lümeni denir. İki tip endoplazmik retikulum vardır.
• Granüllü Endoplazmik Retikulum: Üzerinde ribozomlar vardır. Sisterna denilen yassılaşmış keseler şeklindedir. Salgı ve zar proteinleri yapılır.
• Düz Endoplazmik Retikulum: Ribozomları yoktur, tüplerden oluşan bir ağ şeklindedir. Endokrin bezler ve yağ sentezinde görevlidir.
Ribozom ve diğer organellerde sentezlenen bazı maddeler ER’daki enzimlerle işlenir ve Golgiye aktarılır.
Düz E.R. işlevi
Farklı hücre tiplerinde düz ER değişik metabolik süreçlerde görev alır. Bunlar arasında lipid sentezi, karbonhidrat metabolizması ve ilaç ya da zehirlerin detoksifikasyonu bulunur. Düz ER de bulunan enzimler yağ, fosfolipid ve steroid gibi farklı lipidlerin sentezinde görev alırlar. Düz ER de üretilen steroidler ör: eşey hormonları. Karaciğer hücreleri düz ER nin karbonhidrat metabolizmasındaki önemlidir. Karaciğer hücreleri karbonhidratı bir polisakkarit olan glikojen şeklinde depolar. Glikojenin hidrolizi karaciğer hücrelerinden glukoz salınmasına yol açar. Bu olay kan şekerini düzenler. Ancak glukojen hidrolizinin ilk ürünü glukoz fosfattır. İyonik formdaki bu şeker hücreyi terk ederek kana karışamaz. Karaciğer hücrelerinin düz ER zarı içinde bulunan bir enzim, glukozdaki fosfatı uzaklaştırır ve serbest glukoz hücreyi terk eder.
Mikrocisimcikler : ER dan köken alırlar. Karaciğer epitellerinde ve böbrek kanallarında çok bulunurlar. Hidrojen peroksidin yapım(ürikaz) ve yıkımını sağlayan(katalaz) peroksidaz grubu enzimleri içerirler. Mikrocisimciklere aynı zamanda peroksizomlarda denir.

H A Y V A N F İ Z Y O L O J İ S İ
72
ŞEKİL 1.57 Golgi aygıtın hücredeki fonksiyonu
Düz endoplazmik retikülüm, karaciğer, adrenal korteks, kas, over, testis, plasenta, böbrek ve deride bulunur. Başlıca görevleri steroid sentezi, ilaç detoksifikasyonu, glikojen ve lipid metabolizması olan bu organel kas hücrelerinde triad ve diad yapısına katılarak kalsiyumun depolanması ve sitoplazmaya salınması olaylarında da rol oynar.
�Düz ER deki enzimler özellikle karaciğer hücrelerinin ilaç ve zehirleri detoksifiye etmelerini sağlar. Detoksifikasyon genellikle ilaçlara hidroksil gruplarının eklenmesi ile gerçekleştirilir. Hidroksil grupları ilaçları çözünür hale getirir dolayısıyla vücuttan atılmalarını kolaylaştırır. Kas taki ER sarkoplazmik retikulm zarı kalsiyum iyonlarını sitosolden sisternal alana pompalar(SERCA). Kas hücresi bir sinir hücresi tarafından uyarıldığında,
• Granülsüz endoplazmik retikülüm sitoplazmik yüzeyinde ribozomlar içermezler.
• Farklı hücre tiplerindeki düz ER değişik metabolik süreçlerde görev alırlar. Bunlar arsında
• lipid sentezi (fosfolipid, steroid)
• Karbonhidrat metabolizması
• İlaç ya da zehirlerin detoksifikasyonu bulunur.

H A Y V A N F İ Z Y O L O J İ S İ
73
sisternedeki Kalsiyum ER zarını geçerek tekrar sitosole çıkar kas hücresinin kasılmasını tetikler.
Granüllü ER İşlevi
Salgı proteinlerinin yapılmasını sağlar. Granüllü ER kendi zar fosfolipidlerini yapar; ER zarında yerleşmiş olan enzimler sitosolden gelen fosfolipid öncülerini biraraya getirirler. ER zarı genişler ve transport vezikülleri şeklinde iç-zar sisteminin diğer bileşenlerine aktarırlar.
Ribozom
Ribozom, ribozomal RNA (rRNA) ve proteinlerden oluşmuştur ve hücrenin protein sentez yerlerine verilen addır. Virüsler hariç tüm hücrelerde bulunur. Sitoplazmada serbest veya endoplazmik retikulum'a bağlı olarak bulunan 120-200 Å (angstrom) çapında yapılardır. Ribozomun yaklaşık %65 kadarı rRNA, geri kalan %35'lik bir kısmı ise ribozomal proteinlerden oluşur Ribozomlar protein sentezlerinin yapıldığı merkezlerdir.
ŞEKİL 1.58 tRNA
Protein sentezleneceği zaman DNA'nın yarım dizisi karşısında sentezlenen mRNA zinciri ribozomun 40S'lik küçük alt birimine bağlanır.
ŞEKİL 1.59 Ripozomda protein sentezi

H A Y V A N F İ Z Y O L O J İ S İ
74
Ribozomlar tek tek yahut gruplar halinde bulunurlar. Tek bulunanlara monomer ribozom, gruplar halinde bulunanlara ise polizom veya poliribozom denir. Bunlar ayrıca hücrenin tipi, gelişmesi ve fonksiyonuna göre ya endoplazmik retikulum'a bağlı veya sitoplazmada serbest olarak bulunurlar. Endoplazmik retikulum'a bağlı olanlar hücre dışına verilecek proteinleri (pankreas, sindirim enzimleri v.b) serbest ribozomlar ise hücrenin ihtiyaç duyduğu yapısal proteinleri sentezler.
ŞEKİL 1.60 DNA üzerindeki bilgilinin protein haline gelinceye kadar değişmesi
Tüm proteinler ribozomdan sentezlenmez. Bazı proteinler NRPS enzimiyle sentzelenir. Non-Ribozomal sentez mekanizması ilk olarak 1971 yılında Fritz Lipmann tarafından ortaya atılmış ve thiotemplat mekanizması olarak adlandırılmışır. Bu mekanizmada peptitler, yüksek moleküler ağırlıklı multienzimler olan Nonribozomal Peptit Sentetazlar (NRPS) aracılığı ile yağ asidi sentezine benzeyen bir mekanizma ile sentezlenirler. Amino asitler enzimler aracılığı ile yanyana gelip birleşirler , bu proteinsentezi çesidinde ribozomlar görev almaz. Bu enzimler domainden oluşmuş büyük bir multifonksiyonel enzimdir. NRPS ile sentezlene bazı proteinler antibiyotik yapısındadır.
ŞEKİL 1.59 NRPS ile protein sentezi

H A Y V A N F İ Z Y O L O J İ S İ
75
Golgi Aygıtı
Golgi kompleksi hem yapı hem de fonksiyon yönünden endoplazmik retikulum ile yakından ilişkilidir. Bu organel birbirine paralel bir dizi membranöz kanaldan oluşur ve salgı yapan hücrelerde iyi gelişmiştir. Golgi kompleksinin fonksiyonu endoplazmik retikulumda sentezlenen maddelere son şeklini vermek ve bu maddeleri bir membranla çevrelemektir. Ayrıca hücre zarının yenilenmesi ve yüzeyinin genişletilmesi görevini de üstlenir. Uzun zaman, pek önemli bir organel olmadığı gerekçesiyle, dikkate alınmayan Golgi aygıtı son zamanlarda hücre zarının özgüllüğünü saptamada önemli görev almaşı nedeniyle, dikkatleri üzerine çekti. Çünkü hücre zarının özgüllüğü karbonhidratlarla saptanmaktadır ve karbonhidratlar da Golgi aygıtı'nda sentezlenmektedir. Bazı karbonhidratların, proteinler gibi kalıtsal denetim altında sentezlendiğine ilişkin kanıtlar vardır. Kan grupları ve immunokimyasal incelemeler bunu göstermektedir. Protein glikosilasyonu Endoplazmik reikülümü da başlar Golgide devam eder. Zarların protein ve fosfolipidleri de değiştirilebilirler. Örneğin çeşitli golgi enzimleri glikoproteinlerin oligosakkarit kısımlarını değişikliğe uğratırlar.
ŞEKİL 1.61 Golgi aygıtı.
Oligosakkaritlerin proteinlere eklenmesi, granüllü Endoplazmik retikülüm üzerinde protein sentezlenirken gerçekleşir. Ortaya çıkan glikoprotein, Endoplazmik retikülünün geri kalan kısmı ve Golgi içinden geçtiği sırada değişikliğe uğratılır. Hücreler tarafından salgılanan birçok polisakkarit Golginin ürünüdür. Golginin büyüklüğü hücre tiplerine göre değişkenlik gösterir. Örneğin, sinir , plazma, salgı hücrelerinde büyük, kas ,epidermis lenfositler ve bazı tümör hücrelerinde küçüktür. Golgi Hücre içinde genellikle nukleusa yakın bulunur. ER gibi zarla çevrili yassı keselerden oluşur. Golgi lipit mermbran ile kaplı 3-20 adet düz üstüste yığılmış keseye benzer sisterna adı verilen yapılardan oluşur. Sayıları hücre tipine bağlı olarak bir kaç tane ile yüzlerce arasında değişir. Salgı hücrelerinde genellikle çok sayıda bulunur. Golgi özellikle hücrede

H A Y V A N F İ Z Y O L O J İ S İ
76
moleküler trafik’in yönlendirilmesinden sorumludur. Neredeyse tüm molelüller varlıklarının bir döneminde Golgi aygıtından geçer. ER’nin ürünleri Golgi’de değişime uğratılır, depolanır ve daha sonra hücre içinde ulaşacakları bölgeye gönderilir. Hücre bölmelerine ulaşım vesiküller yoluyla olur. Proteinler vesikül üzerinde uygun reseptörleri ile bağlandığında vesikül membranla kaplanır ve hücre içinde taşınır. Bu vesiküller proteinlerin salgılanacağı bölge olan hücre membranına gönderilmesi için de kullanılır. Şeçici salgılama için kullanılan büyük salgı vesikülleri de bulunmaktadır.
Golginin başlıca işlevleri:
1) ER’den gelen protein ve lipitleri gerekli yerlere ulaştırmak
2) bazı protein ve glikoproteinlerin değişime uğratılması
3) değişime uğratılan bu proteinleri vesiküller içinde paketleyerek hücrenin diğer bölmelerine taşımak veya hücreden dışarı salgılamak
Golgi aygıtı aynı zamanda belli bazı makromolekülleride kendisi üretir. Hücre tarafından salgılanan pek çok polisakkarit Golgi aygıtı ürünüdür. Örneğin hayvan hücrelerinin birarada kalmasını (yapışmasını) sağlayan yapışkan madde “hyaluronik asit” buna örnek verilebilir. Golginin bu üretimi cis ve trans uçlar arasında farklı sisternalarda özel bir enzim grubu tarafından yapılır. Moleküllerin transferinde Golgi’de fosforlama yoluyla yapılan etiketleme de yardımcı olabilir. Ayrıca Golgi’den tomurcuklanan transport vesikülleri membranlarında özgün organellerin yüzeylerinde “liman bölgesi” tarafından tanınan eksternal moleküllere de sahip olabilirler.
Lizozom
Hidrolitik enzimler içeren zar ile çevrili organeldir. Asit fosfataz, esteraz, sülfataz, katepsin ve beta-glukuronidaz gibi enzimler içerir. Lizozomlar lipoprotein yapısında bir membranla çevrilidir. Olgunlaşmasını tamamlamış lizozomlar yani henüz bir madde ile karşılaşmamış lizozomlara primer lizozom denir. Hücre dışından alınmış yabancı maddeyi içeren vezikülle (fagozom) birleşmiş olan lizozomlar ise sekonder lizozom olarak bilinir. Hücre içinde yaşlanmış ve işlevini yitirmiş organellerin unit zarla çevrilip bir primer lizozomla birleşmesiyle otofajik vakuoller oluşur. Uzun yıllar hücrede kalan otofajik vakuoller lipofuksin pigmenti içerirler. Primer lizozomlardan oluşan sekonder lizozomlara otofagozom denir. Lizozomlar özellikle fagositik aktivite gösteren makrofajlarda bol miktardadır. Lizozom membranı lizozomun hücreyi tümüyle sindirmesini önler. Hidrolitik enzimler ve lizozom zarı, granüllü ER tarafından yapılır. Daha ileri değişiklikler için Golgi aygıtına gönderilir. �
• Lizozom sialik asit, galaktoz, hyalüronik asit ve kondroitin sülfat, gibi maddelerin sentezi de yapılır

H A Y V A N F İ Z Y O L O J İ S İ
77
Lizozom enzimlerinin 4 çeşit etki tarzı vardır;
1) Akyuvar ve makrofajler doku harabiyeti olan yere hücum ederler. Burada lizozomlar hücre içine alınan partikülleri sindirirler.
2) Otofaji; yaşlanmış organellerin yıkımı ve açlık hali
3) Otoliz; hücrede tamir işlevi durunca ve ölüm hali. Putrifikasyon birdenbire oksijensiz kalan hücrede lizozom membranı yırtılır. Bazı hücre zehirleri aynı durumu yaratır. Hücrenin kendisi ve civar dokular parçalanır.
4) Tiroid bezi hücrelerinden hormon salınma mekanizması.
ŞEKİL 1.62 Troid hormonun sentezi ve kemik yıkımda lizozom
Lizozom enzimleri daha çok hafif asidik ortamlarda etkendir. Hücrede birçok işlevinin yanısıra, bozukluklannda bazı hastalıkların ortaya çıkmasına neden olurlar. Örneğin, soluduğumuz havadan alınan karbon parçacıkları, akciğerimizdeki fagositlerde yıllarca kalmasına karşın, silisyum dioksit, fagositlerin lizozomuna girer ve oranda bulunan enzimlerin etkisiyle, kristallerinin üzerinde silisik asit oluşur. Silisik asidin hidroksil grupları, hücre zarının yapısında bulunan fosfolipit ve proteinlerin bazı gruplarıyla çok sıkı hidrojen bağları kurar. Böylece hücre ve lizozom zarları zedelenir. Ayrıca silisyum dioksit taşıyan fagositler hücre dışına bir madde salgılarlar. Bu madde, özellikle akciğerdeki bağ dokunun bir çeşit fibröz dokuya dönüşerek esnekliğin yitirmesine neden olur. Bnezer olarak aspest kristalleri benzer durumu oluşturur, özellikle mezotelyum (vücut boşluğunu astarlayan zar) kanserlerini meydana getirir.
Lizozom membran hasarı
Yüksek seviyede alınan A vitamini lizozom membranı etkiler ve membranı bozar. Deney hayvanlarında kemik ve kıkırdak dokuda lizozom enzimleri serbest kalmakta, bu nedenle kemikler kolayca kırılmaktadır. A vitamini lizozom membranını zayıflatır, ancak kortizon ise daha stabil hale getirir. Kortizonun yangı olayını azaltması veya durdurması, lizozom membranı üzerinde olan etkisi ile izah edilebilir. Lizozomlar kendi hücresi içerisindeki bazı maddeleri ya da organelleri (çoğunluk işlevlerim bitirmiş ya da bozulmuş) de sindirir.

H A Y V A N F İ Z Y O L O J İ S İ
78
ŞEKİL 1.63 ROS hücre membranı üzerindeki etkisi
Sindirim kofullarının içinde ribozom ve mitokondrilere rastlanır. Fazla A vitamininin kemiklerdeki ve kıkırdaktaki lizozom enzimlerim serbest bıraktığı ve dolayısıyla kemikleri kırılır bir duruma geçirdiği; fakat yeterli miktarlarda da yaşlı hücreleri yok etmeyi sağladığı için genç kalmada yardımcı olduğu saptanmıştır.
ŞEKİL 1.64 A vitaminin yapısı.
Silika ve asbest tozlarının uzun süre solunum havası ile alınmaları sonucu silikozis ve asbestos hastalıkları meydana gelmektedir. Asbest partiküllerine maruz kalmada bazı tür kanserlerin görülmesi, lizozomlar ile bu tür kanserler arasında bir ilişki olabileceğini düşündürmektedir. Fagositlerin ölümü, kollajen fibrillerini sentezleyen ve salan fibroblastlarda başka bir reaksiyona sebep olarak fibröz doku yapımını kamçılamaktadır. Gut, Metabolik bir bozukluk nedeniyle mono-sodyum ürat kristallerinin oynak yerlerinde toplanması sonucu ortaya çıkan bir hastalıktır. Sodyum ürat kristalleri fagositler tarafından alınır. Bu hastalıkta lizozom enzimleri serbest kalmakta GUT hastalığında görülen ağrı ve yangı olayları lizozomlardan ileri gelmektedir. Kanda ürik asit miktarı artar, eklem yerlerinde ürik asit kristalleri toplanır. Bu kristaller plazma kininlerini aktive eder. Plazma kininleri ağrı meydana getiren kimyasal maddelerdir.

H A Y V A N F İ Z Y O L O J İ S İ
79
ŞEKİL 1.65 Lizozom ve işlevi.
Kızarıklık, Sıcaklık, Şişlik, Ağrı ile kendisini gösteriri, Yangılı doku ve hücrelerden kapiler permeabilitesini arttıran histamin, seratonin ve bradikinin gibi maddeler açığa çıkar. Bradikinin serum proteinlerinin enzim etkisi ile parçalanması sonucu meydana gelen bir peptiddir. Yangıdaki doku tahribi bradikinin şekillenmesi lizozom proteazları tarafından yapılmaktadır.
Aktivitesi azalan dokuda lizozomal aktivite artar; -gebelikten sonra uterusun gerilemesinde, sütten kesince meme bezinin küçülmesinde ve çalışmayan kasların atrofisinde lizozom görev alır. Peroksizomlar Çeşitli substratlardan O2’e H
+ aktararak, yan ürün olarak H2O2 oluşturan enzimler içerirler. Hücre için zehirli olabilecek maddeleri oksitleyerek zehirsiz hale getirirler. (örn. alkol) H2O2’ in fazlası hücre için toksiktir ve bunu katalaz enzimi parçalar. H2O2- Katalaz enzim sistemi; hücredeki zararlı molekülleri oksitleyip etkisiz kılar.
Lizozomların işlevi;- dokuların gerilemesi
—hücrelerin otolizi —hücresel sindirim işlevi —yumurtanın döllenmesi —corpus luteum dejenerasyonu

H A Y V A N F İ Z Y O L O J İ S İ
80
Mitokondri ve Enerji Üretimi
Hücrenin ihtiyaç duyduğu ATP’nin sentezlendiği yerdir. Mitokondri hücrede kendini bölünerek yenileyebilen tek organeldir. Mitokondri iç zarı kristalar oluşturmak üzere katlantılar yapmıştır. İç membran yarı geçirgen yapıdadır. İç zarda fosforilasyon enzimleri, elektron transport sistemi bulunur. Matrikste kalsiyum içeren yoğun matriks granülleri bulunur. Mitokondriumda A, C ve E vitaminleri bulunur. Mitokondride TCA enzimleri bulunur. (amorf matrikste) Membran vezikül ve vakuol gibi üç farklı komponentten oluşmuştur. Intraselüler su miktarının ayarlanmasında görev alırlar. Belirli hücre ürünlerinin sindiriminde görevlidir. Hücre solunumun yapıldığı merkezlerdir. Hücrenin ATP üretim yeridir. Yani enerji santralidir. Bu organelde birim zar ile çevrilidir. İçteki zar kıvrımlıdır. Bu kıvrımlara “krista” denir. Mitokondrinin kendine ait DNA, RNA, ETS (elektron taşıma sistemi) ve ribozomu vardır. ETS’ si olduğundan kendi enerjisini üretir.
Eritrositler haricinde tüm hücrelerde farklı sayıda mitokondri bulunur. Lenfosit ve epitelyal hücrelerde az sayıda mitokondri bulunurken, midenin parietel hücreleri, böbrek tübül hücreleri, karaciğer hücreleri ve adrenal korteks hücreleri gibi yüksek metabolik aktiviteye sahip hücrelerde bol miktarda bulunur. Örneğin bir karaciğer hücresinde (hepatosit) 2000–3000 adet mitokondri bulunabilir.
ŞEKİL 1.66 Mitokondri ve ATP sentezi.
Mitokondri DNA’ sı olduğundan kendini eşleyebilir. Ayrıca mitokondri oksijenli solunumun yapıldığı yerdir. Oksijenli solunum sayesinde ATP üretilir. Hücre için gerekli enerjinin (ATP) elde edildiği organeldir. Kendi DNA, RNA, ribozom ve proteinleri olan mitokondri, kendisi bölünerek yenilenebilen tek organeldir. Mitokondride iç ve dış olmak üzere çift ünit zar vardır. Fosforilasyon enzimleri, elektron transport işlemi iç zar (kristalar. yağ asitleri karnitin aracılığıyla buraya geçer) üzerinde yerleşmiştir. Bu nedenle yapısında en çok protein olan membran mitokondri iç zarıdır (Lipit/protein oranı en yüksek yapı miyelin kılıftır). Matrikste ise kalsiyum içeren yoğun matriks granülleri izlenir. TCA siklusu enzimlerinden sadece süksinat dehidrogenaz iç zarda yerleşmiştir. Mitokondrinin biyokimyasal markerı glutamat dehidrogenazdır (matriks enzimi). P53 geninin bax protein ürünü mitokondri iç zarında kanal oluşturur. Matrikste bulunan sitokrom C bu kanaldan sitoplazmaya çıkar. Sitokrom C apopitozisi başlatan enzim olan caspase’ı aktive eder. Caspase (C-cysteinn, asp-aspartat, ase) enzimi proteinleri sistein ve aspartat rezidülerinden parçalar. Apopitozis gerçekleşmiş olur.

H A Y V A N F İ Z Y O L O J İ S İ
81
ŞEKİL 1.67 Enerji üretimi
Kemoosmotik Atp sentezi
Kemiosmoz; iyonların, elektrokimyasal gradyanı azaltmak için seçici geçirgen bir zardan geçme hareketidir. Hücresel solunumdaki ATP sentezinin gerçekleşmesini sağlayan enerjinin büyük bir kısmı hidrojenlerin yaptığı bu hareketten karşılanır. İyonlar şekildeki gibi bir kanaldan geçirildiğinde, sahip oldukları potansiyel enerji, kimyasal reaksiyonları gerçekleştirmek için kullanılabilir. Hidrojen iyonları (protonlar) proton konsantrasyonun yüksek olduğu yerden düşük olduğu yere doğru difüzyon hareketi yaparlar.
ŞEKİL 1.67 ATP ve enerji.
ATP-sentaz kemiosmoz aracılığıyla ATP sentezleyen enzimdir. ATP-sentaz protonların zardan geçmelerine imkan vererek; protonların sahip oldukları kinetik enerjiyi, ATP fosforilasyonu yapmak için kullanır. Bu mekanizmayla ATP üretimi kloroplast ve mitokondrinin yanı sıra bazı bakteri türlerinde de görülür.

H A Y V A N F İ Z Y O L O J İ S İ
82
ŞEKİL 1.68 Farklı organellerde ATP sentetazi yapılması.
Hekzos Monofosfat Yolu (Pentoz Fosfat Yolu)
Glikolizin bir yan yoludur. Pentoz fosfat yolu sitoplazmasında gerçekleşir. Pentoz fosfat yolu sonunda direk olarak hiç ATP üretilmez veya tüketilmez. Buna karşın metabolik olarak önemli maddelerin sentezi yapılır.
Pentoz fosfat yolunda glukoz 6-fosfat, riboz 5-fosfata ve CO2'ye oksitlenirken NADPH üretilir. Bu beş karbonlu şeker ve onun türevleri, ATP(indirek olarak) CoA, NAD+, FAD, RNA ve DNA gibi hayati öneme sahip biyomoleküllerin bileşenleridir. Bu olayın net reaksiyon denklemi aşağıdadır:
Glukoz 6-fosfat + 2NADP+ + H2O ���� Riboz 5-fosfat + 2NADPH + 2H+ + CO2
Glukoz-6-P dehidrogenaz: Pentoz fosfat yolunun düzenleyici enzimidir. NADPH tarafından inhibe edilir. Glukoz-6-P DH aktivitesini maddeler 1.NADP (Pentoz fosfat yolunda rol alan kofaktör). Glukoz-6-P DH aktivitesini maddeler:NADPH , Eritrosit hücre direnciyle ilgili oksidasyon tipidir. Eritrositlerde bulunan glukoz-6-P DH aracılığıyla elde edilen NADPH eritrositlerde bulunan H2O2’nin indirgenmesinde rol oynayarak hemolize olmasını önler.
Hücre iskeleti Görevi
Ökaryotik hücrelerin çeşitli şekillere dönüşmesi ve hareketlerinin koordineli olarak yönlendirilmesi, sitoplazma içerisinde yaygın halde bulunan kompleks bir protein ağı tarafından sağlanmaktadır. İşte bu ağ hücre iskeleti adını alır. Hücrenin şeklinin değişmesi, bölünmesi ve çevre uyaranlarına cevap vermesi ile yeniden organize olabilen oldukça
���� Pentoz fosfat yolu amaçları:
1.Biyokimyasal reaksiyonlarda redüktan olarak görev yapan NADPH ihtiyacını karşılar.
2.Nükleotid sentezi için gerekli olan riboz fosfatları üretir.
3. 3 yada 7 karbonlu şekerlerin birbirine dönüşümünü ve glikolize bağlantısını sağlar.

H A Y V A N F İ Z Y O L O J İ S İ
83
dinamik bir yapıdır. Hücre iskeletinin yapısında 3 tip protein filamenti bulunur. Hücre iskelteinin görevi hücreye desteklik ve şekil vermektir. Hücre iskeletinin temel işlevi çeşitli şekillerdeki hücre hareketini meydana getirmektir. Bunun dışında hücre içinde motor proteinlerin organelleri taşıması için bir yol oluşturmak hücre iskeletinin diğer görevidir.
ŞEKİL 1.69 Hücre iskeleti elemanları.
Mikrotübüller ve mikrofilamentler küresel proteinlerden oluştukları halde ara filamentler fibrilsi proteinlerden oluşur. Mikrotübüller 240 A° çapında, mikrofilamentler 70 A°, ara filamentler ise 100 A° çapındadırlar. Mikrotübül ve mikrofilamentlerin tek alt birimi olduğu halde ara filamentlerin 5 farklı alt birimi vardır. Mikrotübüller ve mikrofilamentler polimerizasyon için enerjiye ihtiyaç duydukları halde ara filamentler enerji ihtiyacı duymazlar.

H A Y V A N F İ Z Y O L O J İ S İ
84
İntermedial Filamentler
Bunlar mikrofilmentlerden daha kalın, mikrotübüllerden ise daha incedir. Görevleri arasında, Hücre biçimlerini güçlendirme, organellerin hücre içerisindeki yerlerini sabitleme gibi görevleri vardır. Mikrofilamentler iskelet kasında: kas kasılmasında, Hücre korteksinde: membran hareketinde, hücrenin yer değiştirmesi, endositoz, ekzositoz Hücre yüzey farklılaşmasında: Mikrovillusların yapısında (villin, fimbrin) mitozda görev alır.
Vimentin, proteinleri ara filamant (intermedial filament) ailesinin bir üyesidir. Ara filamanlar, ökaryotik hücrelerde mikrotübüller ve aktin mikrofilamanlarıyla beraber, hücre iskeletine oluşturur. Çoğu ara filamanın yapısı kararlı olsa da, vimentin değişken bir yapıya sahiptir. Vimentin organellerin yerini sabitlemeye yarar, hücreye esneklik sağlar ve hücre içinde kolesterol taşınmasında rol oynar. Vimentin ve mikrotübüller arasında sıkı ilişkiler vardır.
İntermedial filamentler kontraktil olmayan, bulunduğu hücreye desteklik eden filamentler. Ara filamentler çapları 10-12 nm arasında değişen heterojen bir gruptur. Dayanıklı, devamlı fibriller olup elektron mikroskopta düz veya hafifçe bükülmüş şekilde görülürler.
Ara filamentlerin 5 ana grubu vardır:
1.Sitokeratinler (cytokeratin veya prekeratin) 2.Vimentin 3.Desmin (Skeletin), 4.Glial fibriler asidik protein (GFAP, Glial fibriller) ve 5.Nörofilamentler

H A Y V A N F İ Z Y O L O J İ S İ
85
Mikrofilamentler
Mikrofilamentler 7 nm çapında sert çubuklar halinde bulunur. Aktin moleküllerinden meydana gelmişlerdir. Bir mikrofilament aktin alt birimlerinden oluşmuş ve birbiri üzerine sarılmış iki zincirden meydana gelir. Tüm ökaryotik hücrede bulunurlar. İki önemli görevleri vardır, birincisi hücre hareketini gerçekleştirmek ikincisi ise hücre şeklini korumak. Mikro filamentler değişik uzunlukta olabilen kontraktil protein iplikciklerdir. Hemen bütün hücrelerde bulunurlarsa da özellikle kas hücrelerinde çok miktarda bulunurlar (myofilament). Kas hücrelerindeki kasılma fonksiyonu aktin (actin) ve miyozin (myosin) arasındaki ilişkiye bağlıdır. Aslında “mikrofilament” terimi özellikle 6-8 nm kalınlığında aktin için uygun bir terimdir. Miyozin 15 nm kalınlığındadır ve kasın kalın miyofilamenti diye isimlendirilir. Kalın bir miyozin filamentinde 500 kadar miyozin başı vardır ve bunlar saniyede 5-10 defa aktin bağlayıp bırakabilir. Kas hücreleri dışındaki bazı hücrelerde aktin total proteinin %10’unu teşkil edebilir. Kas hücrelerindeki aktinin yapısı stabil iken diğer hücrelerde aktin parçalanıp yeniden yapılabilir. Mikrovillus içindeki filamentler aktin yapısındadır. Bunlar apikal sitoplazmadaki terminal ağlara tutunur ve kontraksiyonla mikrovillusda kısalma veya bükülmeye yol açarlar. Birçok hücrede plazmalemma altında ince bir tabaka oluşturan mikrofilamentler hücre membranının lokal hareketinde (örneğin fagositozisde) rol oynarlar. Mikrofilamentlerin sitoplazmik komponentlerin hareket ve hücre içi yer değiştirmelerinde (sitoplazmik akıntı) rolü olduğuna da inanılır. Mitozda hücrenin ikiye bölünmesine de katkıda bulunurlar. Hücrenin hareketinden ve sitoplazma akıntılarından sorumludurlar. Ayrıca kasılma-gevşeme olayında, uyan ve madde iletiminde görev alırlar. Mikrofılamentler, miyofıbriller (kas telcikleri) ve nörofıbriller (sinir telcikleri) olmak üzere iki tiptir.
Mikrotübüller Tüm ökaryotik hücrelerde bulunurlar. İçleri boş olan tübül duvarı, tübilin adı verilen globüler proteinlerden meydana gelmişlerdir. Hücre iskeletini oluşturmakla birlikte hücre içeride organellerin taşınması, kamçı ve sil yapısının meydana getirilmesi, sentrozomları oluşturmaları gibi birçok görevleri vardır.
ŞEKİL 1.70 Kamçı hareketi .

H A Y V A N F İ Z Y O L O J İ S İ
86
Hücre iskeleti deformasyona karşı hücre şeklini korunması sağlar ve mekanik direnç kazandırır; Hücre iskeleti, Hücre dışı matriks (HDM) ile hücre arasında iletişim sağla böylece hücre ve hücrenin çevresini deforme ve hücreler göçü düzenlener. Hücre iskeleti hücre sinyal yollarında görev alabilir. Bu yolla hücre dışında madde alınması (endositoz ;)yada hücre bölünmesinde kromozomlarını hareketinde ve sitokinez - ana hücrenin iki kardeş hücreye bölünmesinde görev alabilir. Ayrıca, hücre zarından yapılan flajeller, kirpikler, lamellipodia ve podosomes oluşturulmasında görev yapar. Hücre iskeleti tarafından gerçekleştirilen diğer önemli bir işlev kas kasılmasıdır.
Dynein silya yapısı içerisinde hareketi sağlar. Kinezin ise aktif hareket yapan silyanın kollarının eski hâline gelmesini sağlar. Dynein genetik olarak eksikse Kartagener sendromu oluşur. Temel olarak dynein hareket ettirir. Kinezin eksi konumuna getirir. Dynein ve Kinezin Her ikisinin de ATPaz aktivitesi var. Dynein retrograt iletimden sorumlu – kutba doğru Kinesin antegrat iletimden sorumlu + kutba doğru Dynein silya yapısı içerisinde hareketi sağlar. Kinezin ise ise aktif hareket yapan silyanın kollarının eski haline gelmesini sağlar.
Mikrotüpüllerin başlıca görevleri :
1.Mitoz bölünmede: Sayıca artan mikrotubuluslar, hücrenin iki kutbu arasında yerleşerek iğ iplikçiklerini oluştururlar. Kromozomların kutuplara çekilmesinde katkıları vardır. Bölünmeden sonra mikrotubullerin çoğu kaybolur.
2. Yeni sil ve flagellum yapımı: Sentriol yeni silin gelişeceği yüzeye göç ederek sillerin başlangıç yeri olan bazal cisimleri oluşturur.
3. İletim: Sinir hücrelerinde akson boyunca ilerleyen nörotubuluslar hücre içi madde iletiminde görev alırlar.
4. Hücre iskeletini oluşturur. Hücre içyapısına desteklik ederek, belli bir biçim ve dirilikte kalmasını sağlar.
5. Hareket: Silya ve flagellumun yapısına girerek hareketlerine katkıda bulunur.
6. Salgı atılması ve intrasellüler organel taşınması: Sentezlenen salgı maddelerinin hücre yüzeyine doğru hareket ettirilmesinde rolleri vardır. Kromozomların iğ iplikçiklerine hareketi, melanositlerde melanin pigmentinin sitoplazmik uzantılara göçü ve ER’dan Golgi aparatına taşıyıcı veziküllerin ilerlemesi mikrotubul ağının fonksiyonuyla bağlantılıdır.
Sentrozom
Sentriol çifti, etrafını saran sentroplazma ile birlikte sentrozom adını alır. Her sentriol enine kesitte 9 fıbrilden, her fıbril 3 subfıbrilden (mikrotubulus) oluşur. Hücre bölünmesinde görev alan sentrioller bölünme sırasında çoğalarak birer çift halinde kutuplara giderler ve bu sırada aster (iğ) adı verilen iplikçiklerin oluşumunu ve sentromerleri aracılığıyla bu iplikçiklere tutunan kromozomların hücrenin kutuplarına çekilmelerini sağlarlar. Sentrozomlar sentriyol çiftlerinden meydana gelmişlerdir.
• Dynein aksonal iletide retrograt iletimden sorumludur (içi boş vezikülü taşır). • Kinezin aksonal iletide anterograt iletimden sorumludur (içi dolu vezikülü taşır).

H A Y V A N F İ Z Y O L O J İ S İ
87
ŞEKİL 1.71 Sentrozomların yapısı
Kamçı ve Siler
Siller ve kamçılar hücrelerin hareketini sağlarlar. Siller daha kısa ama çok sayıda hücre yüzeyinde bulunurlar. Memelilerin solunum organlarındaki hücrelerin silleri tozların tutulmasını sağlar. Kamçı ise hücrelerde bir yada iki tane olur ve sillerden daha uzundur. Sil ve kamçıların yapısında mikrotübüller vardır (9+2).
Hücre iskeleti ve hareket
Hücrede 2 hareket sistemi vardır (Motor Sistemleri ):
• Aktin-tabanlı motilite: miyosinler
• Tübülin tabanlı motilite: kinesinler ve dyneinler
Dynein: ATP enerjisini mekanik enerjiye harekete dönüştüren (aynı zamanda motor molekül olarak da adlandırılır), bir motor proteindir. Dynein genellikle hücre merkezine yönelen mikrotübülün arasında eksi sonuna doğru sitoskeletal mikrotübüllere boyunca "yürüme" ile çeşitli hücresel kargoları taşır. "eksi uç yönelen motorlar" olarak bilinir. Bu tür taşıma retrograd nakil olarak bilinir. Bunun aksine yönünde ise, mikrotübüle 'artı ucuna doğru hareket sağlayan motor proteinler kinesinlerdir, Kinesinler uça yönelik motorlar proteinler olarak adlandırılır. Dyenin farklı memeli hücre tiplerinde vardır, solunum yolu epitel hücrelerinde ve sperm hücrelerindeki hareketli kirpikler ya da flagella'yı bulunur. Kinesinler, siliyer / kamçılı ucuna doğru axonemal mikrotübül boyunca parçacıkların taşınmasını yapar Sitozolik dyneins geri sitozolüne doğru axonemal taşıma yapar. Memlilerde en azından 40 farklı kinesin geni belirlenmiştir.

H A Y V A N F İ Z Y O L O J İ S İ
88
Çekirdek (Nükleous) Çekirdek nükleus membranı, kromatin, çekirdekçik, nükleoplazma içerir. Nükleus membranı iki paralel ünit zar ve bunların arasında yer alan perinükleer sisternadan oluşmuştur. İnsanda en büyük nükleus spinal ganglion ve oosit de bulunur. Karaciğer, kıkırdak, epitel hücreleri, vesica urineria ve leydig hücrelerinde iki tane nukleus olabilir. İç ve dış membranlarının birleşme yerlerindeki porlar 500 Ao çapındadır. Porlar nükleus ve sitoplazma arasında madde alışverişini sağlar. Bazı bölgelerde nükleus dış membranı ER ile devam eder. Membrana nuklearis externa'da ribozomlarda bulunabilir. Çekirdek nükleer DNA taşır. DNA molekülü aşırı derecede kıvrılarak ve proteinlerle birleşerek kromatin adı verilen yapıyı oluşturur. Bu yapı hücre bölünmesine girerken daha da kısalır ve kendini eşleyerek kromatid adı verilen yapıya dönüşür. Bir metafaz kromozomu böylece kendini eşlemiş bir çift kromatid den meydana gelir. Bunlara kardeş kromatidlerde denir. Genetik açıdan birbirlerinin aynısıdırlar ve aynı DNA yani nükleotid sırasına sahiptirler.
ŞEKİL 1.72 DNA yapısı ve paketlenemsi.
DNA molekülü kromatin haline gelebilmesi için histon adındaki proteinlerce paketlenmesi gerekir. Histonlar, çoğunlukla ökaryotik hücrelerde bulunan, küçük molekül ağırlıklı, bazik proteinlerdir. Lizin ve arginin içeriklerine göre, H1 (H5 de denir), H2A, H2B, H3, H4 ve arkeal histonlar olmak üzere 6 tiptir. Kromatin yapısında DNA, DNA'nın iki katı kadar protein bulnur. Hücre çekirdeğindeki bu proteinler, asidik özellikte "histon olmayan" ve bazik özelikte "histon" tipi proteinlerdir. Bir farenin karaciğer hücresinde yapılan araştırmaya göre hücre çekirdeğindeki proteinlerin %11'i histon %19'u histon olmayan proteinlerden oluşmaktadır. Spermde histonlar "protamin" adını alır. Histonlar molekül ağırlığı küçük moleküllerdir. Ökaryotların tüm soma ve eşey hücrelerinde bulunan histonlar, genelde türler arasında benzer tiptedir. Histon protein tipleri elektroforez yöntemleriyle kolayca ayrılabilir. Çekirdekçik(nükleous) Ribozom sentezlemekle görevlidir. Protein sentezi çok olan hücrelerde, çekirdekçik sayısı da artmaktadır. Hücre bölünmesi sırasında eriyerek kaybolur, bölünme tamamlandıktan sonra tekrar oluşur. Nukleolus bir yapı değildir sadece görünür kısımdır. Çekirdek zarında, zarların birbirine temas ettiği bölgelerde 400–1000 Angstrom çapında annulus adı verilen porlar meydana gelmiştir. Böylece nukleus membranı aracılığı ile nukleoplazma ve sitoplazma sürekli ilişki halinde olup bu porlardan RNA molekülleri, polipeptidler, tuzlar, enzimler, koenzimler, ATP ve şekerler rahatlıkla geçebilirler.

H A Y V A N F İ Z Y O L O J İ S İ
89
Sitoplazmada bulunan birçok organel ve çekirdek zarı mikrotübülüsler aracılığı ile sürekli bağlantı halindedir. Nükleus membranı, mitoz bölünmenin profaz evresi sonunda kaybolur, bölünmenin tamamlanmasından sonra yeniden oluşur.
NÜKLEOTİD
Nükleik asitler, tüm hücrelerin nukleus ve sitoplazmalarmda (mitokondri ve bitkilerde kloroplast) bulunurlar ve;- DNA (Deoksiribonükleik asit) - RNA (Ribonükleik asit) olmak üzere iki tiptedirler. Her iki nükleik asit de nükleotidlerin polimerize olması ile meydana gelir ve yapılarında; • Şeker • Fosfat • Baz ünitelerini bulundururlar.
ŞEKİL 1.73 Pürin ve primdin bazları
DNA da 5 karbonlu şekerlerden (pentoz) deoksiriboz bulunur. Deoksiribozun 1. Karbonuna glikozid bağ ile tutunmuş bazlar pürin ve pirimidin olmak üzere iki tiptir. Pürin bazları adenin (A) ve guanin (G), Pirimidin bazları ise sitozin (C ) ve timin (T) dir. DNA da bulunan bazlar kantitatif olarak Edwin Chargaff tarafından incelenmiş olup adenin miktarının timine, guanin miktarının ise sitozine eşit olduğu görülmüştür. Chargaff kuralı olarak bilinen bu kural; kısaca A=T ve G=C olarak özetlenebilir. Adenin ile timin arasında iki, guanin ile sitozin arasında üç adet hidrojen (H) bağı bulunur. Deoksiribonükleik asidin çift iplikli yapısına Watson-Crick sarmalı (double-heliks) adı verilmektedir. 20 Angstrom çapındaki sarmalda bazlar 3.4 Angstrom aralıklarla sıralanmıştır ve sarmal her 10 bazda bir dönüş yapar. Sarmaldaki iplikler antiparalel olup bir iplik 5’—>3', diğer iplik ise 3'—>5' yönündedir. Şeker ve fosfat omurgasından oluşan iplikler birbirlerine hidrojen bağları ile zayıf olarak bağlanmışlardır. Omurga oluşturulurken şekerin 5 numaralı karbonuna bağlı fosfat grubunun hidroksili ile diğer nükleotiddeki şekerin 3' karbon atomuna bağlı hidroksil grubu arasında bir fosfodiester bağı oluşur.
Azotlu organik baz 5 C’lu Şeker Fosforik asidin fosfor grubu
H3PO4
-Riboz -Deoksiriboz
PÜRİN -Adenin (A) -Guanin (G)
PİRİMİDİN -Sitozin (S=G) -Timin (T) -Urasil (U)

H A Y V A N F İ Z Y O L O J İ S İ
90
ŞEKİL 1.74 Kodon ve antikodon eşleşmesi ve DNA yapısı

H A Y V A N F İ Z Y O L O J İ S İ
91
Uyarılabilir Dokular Sinir ve
Kas sistemi
Uyarılabilen, uyarı taşıyan, uyarıya karşı tepki veren dokular Uyarılabilir dokulardır. Reseptörler oluşturulan aksiyon potansiyeli Merkezi sinir sistemine ileten sinir sistemi ile MSS gelen bilgiler ile hareket eden kas sistemi bu bölümde yer almaktadır.
Sinir sistemi Tüm canlılar herhangi bir uyaran karşı cevap verme yeteneğine sahiptirler. Bu uyaranlar ya dış ortamdan gelir yada canlının kendi iç ortamında meydana gelebilir. Hayvanları bitkilerden ayıran en önemli özellik uyaranlara çok hızlı cevap vermesidir. Bu da sinir sistemi ile gerçekleşir. Sinir sistemi çevrenin, mekanik, termal, kimyasal ve elektriksel özelliklerini algılar ve bunlara karşı cevaplar oluşturur. Bu ilişkide sözü edilen çevre sinir sisteminin kendisi dışındaki her şeydir. Tek hücrelilerde bütün bu olaylar tek bir hücre içinde meydana gelirken, çok hücrelilerde bu fonksiyonların her birini yapmak için özelleşmiş hücre ve hücre grupları bulunmaktadır. Çok hücreli gelişmiş canlılarda bu 4 komponentten herbiri için özelleşmiş bir hücre ya da organ vardır. �
Bölüm
2
1-Uyarılabilme: Yeteneğine sahip Yani uyarı alabilen reseptörler bulunur. Ör: göz retinası reseptörleri
2- Reseptörde meydana gelen sinyali merkezlere ileten duyu sinirleri conductivite işini yapan özel hücrelerdir.
3-Sinir merkezleri, buraya getirilen sinyali değerlendirip canlının ne yapması gerektiğini saptar (correlation).
4-Cevabın gerçekleştirilmesi için merkezden gerekli sinyaller motor sinirler yoluyla iş görecek organa (effektör organa) götürülür. Özelleşmiş effektör hücreler reaksiyon (reaction) gösterirler.
Bu bölüm için Power
point sunumu adresi:
https://yadi.sk/i/gsPOvgs
4ccCN9

H A Y V A N F İ Z Y O L O J İ S İ
92
Bu organ kas, bez ve hatta sinir sisteminin diğer kısımlarıdır. Effektör organ gelen sinyale göre aktive ya da inhibe edilir. Örneğin, gözün retinasında ışık dalgalarından etkilenen reseptör hücreler vardır. Reseptörlerde meydana gelen sinyali ileten duyu sinirleri iletim işini yapan özel hücrelerdir. Uyarılabilir doku ve hücreler:
Nöron
Sinir hücresine nöron adı verilir. Evrimi biraz daha ilerlemiş sinir sisteminde uyarma ile meydana gelen sinyalin iletiminde iki ayrı iletici (conductor) hücre vardır. Birinci nöron reseptörden sinyali alır, belirli bir yere kadar götürür; ikinci nöron ile ağlantı kurar ve bu ikinci nöron sinyali effektör organa iletir. Burada birinci kondüktor nörona sensorik nöron (duyu nöronu) denir ki duyuları ileten nörondur; ikinci nörona motor nöron denir ki bir effektörü harekete geçiren nörondur. Bir omurgalı spinal motor nöronu fonksiyonel olarak özelleşmiş bölgeler içerir. Bilgi akımı dendritlerden akson ucuna doğrudur. Bazı nöronlar bilgiyi hücre gövdeleriyle alırlar. Alınan bilgi nörotransmiterlerle akson terminallerine taşınır ve böylece sinyal iletilmiş olur. Bir Nöron hücresinde nörofilamanlar, nörotubuluslar (nörofizin I ve II taşıma proteinleri) bulunur. Bu yapılan dyenin ve knesin aracılığı akson ucunda salgılanan nörotrasmiteri taşır.
ŞEKİL 2.1 Nöronda nörotransmitter madde taşınması
Bir nöronda temel olarak 3 kısım vardır :
• Sinir hücreleri
• Kas hücreleri (kalp, iskelet ve düz kas)
• Bazı endokrin (hipofiz, pankreas beta adacık hücreleri, adrenal bez medulla hücreleri) bez hücreleri
• Bağışıklık sistemine ait bazı hücreler (makrofaj ve nötrofiller)
• Üreme sistemine ait bazı hücreler (fertilize yumurta)
Hücre gövdesi
Dendrit
Akson

H A Y V A N F İ Z Y O L O J İ S İ
93
Gelişmiş canlılarda sinir sisteminde duyu ve motor nöronlar arasında association nöron(internöron=ara nöron) lar yer almıştır. Ara nöronlar sayesinde bir tek reseptörden alınarak duyu nöron ile iletilen sinyal, çeşitli motor nöronlara ve çeşitli effektör organlara doğru yönetilir. Burada ara nöronlar ve motor nöronlar beraberce nöronlar topluluğunu (ganglion) meydana getirmişlerdir ki bu sinir merkezlerinin ilk taslağıdır. Nöronlar MSS’ndeki hücrelerin yaklaşık %10’unu oluştururken, geriye kalanını nöroglia da denen (glia=tutkal) glial hücreler oluşturur. Bunula birlikte, nöronlar, glial hücrelerden çok daha fazla dallanma yaptıkları için, nöronlar beyin ve omurilik hacminin %50’sinden fazlasını işgal ederler. Glial hücreler nöronun soma, akson ve dentritlerinin etrafını çevreler ve nöronları fiziksel ve metabolik olarak desteklerler. Oligodendrositler, astrositler, mikroglialar ve epandimal hücreler olmak üzere 4 çeşit glial hücre vardır.
• Oligodendrositler: MSS’nde aksonları kaplayan miyelini yapar (PSS’nde bu işi yine
bir nöroglia hücresi olan Schwann hücresi yapıyor). • Astrositler: (1) sinaps etrafındaki sinirsel iletim maddelerini ve potasyum iyonlarını
uzaklaştırarak, MSS’ndeki hücre dışı ortam sıvısının içeriğinin düzenlenmesine yardım eder. (2) kapiller duvarını oluşturan hücreler arasındaki sıkı bağlantıların oluşumunu yani kan beyin bariyerinin oluşumunu uyarmaktır. (3) glikoz sağlayıp amonyağı uzaklaştırarak, nöronlara metabolik destek sağlar. (4) embriyonel gelişim sırasında nöronların son olarak yerleşecekleri yere göç etmesine rehberlik eder ve büyüme faktörleri salgılayarak nöronal gelişmeyi uyarırlar. (5) pek çok nöron benzeri özelliğe de sahip olup, iyon kanallarına belirli sinirsel iletim maddelerinin reseptörlerine ve bunları işlemek için enzimlere, zayıf bir elektriksel yanıt oluşturma yeteneğine sahiptirler.
• Mikroglialar: MSS’nde immün işlevleri gerçekleştiren makrofaj benzeri özelleşmiş bir hücredir.
• Epandimal hücreler: beyin ve omurilik içinde sıvı bulunan boşluklarda bulunurlar ve BOS oluşumunu ve akışını düzenlerler.
ŞEKİL 2.2 Nöronlara yardım(fizyolojik ve metabolik destek yapan) eden diğer hücreler.

H A Y V A N F İ Z Y O L O J İ S İ
94
Aksonlar iki tipe ayrılır: Miyelinli aksonlarda aksolemma üzerini miyelin denilen lipoid tabiatında kalınca bir örtü çevreler. Miyelin örtüsü akson boyunca belirli aralıklarla kesintiye uğrayarak boğumlar meydana getirir. Bu
boğum yerlerine ranvier boğum denir. Aksonun miyelin kılıfı, periferik sinir sisteminde düzenli aralığa sahip, dar ve halka şeklinde boğumlanmalar Bu boğumlar, “Ranvier Düğümü” İki düğüm arasında kalan bölge, “internodal bölge” Bu bölgede, sadece bir tek Schwann hücresi bulunur. Düğümler arası bölgelerin uzunluğu, fetüste ve bebekte azdır, yaşla birlikte artar Ranvier düğümlerinin bulunduğu bölgede, miyelin kılıfı kesintiye uğrar. Dolayısıyla, buralardan madde alış-verişi gerçekleşir (Miyelin, madde alış-verişini engelleyici bir kılıftır). Miyelinli sinir tellerinde uyarının hızlı iletilmesinin iki sebebi vardır: Miyelin kılıfın izole edici özelliği ve uyarının, Ranvier düğümleri boyunca atlayarak aksonda saltatorik şekilde iletilmesi. Miyelin kılıfının, sinir tellerinin yenilenmesinde de önemli rolü vardır. Bir sinir teli kesilince, kesilme yerinden çevreye doğru önce miyelin kılıfı meydana gelir. Yenilenen akson, bu kılıfı takip eder. Miyelin kılıfı, akson için klavuzluk yapar. Yenilenmenin gerçekleşebilmesi için, nöronun hücre gövdesinin sağlam olması şarttır. Hücre gövdesi zedelenmiş ve ölmüş olan bir nöronun aksonunda, yenilenme olayı da gerçekleşmez. Miyelin kılıfının uyarı iletiminde önemli rolüne karşılık, besin fizyolojisi bakımından bunun tersi sayılabilecek bir etkisi vardır. Miyelin kılıfında lipit bulunduğu için, besin maddelerinin aksona geçişi zor olur. Bu durum özellikle kalın miyelin kılıflar için daha da önemlidir. Gerek Ranvier düğümleri ve gerekse Schmidt – Lantermann yarıkları, miyelin kılıfının kesintiye uğradığı veya azaldığı bölgelerdeki madde alış-verişinin sağlandığı kısımlardır. Bu açıklıklar, aksonla çevresi arasındaki bağlantı bölgeleridir. Gaz, metabolitler ve besin maddeleri, aksoplazmaya bu yerler aracılığıyla girip çıkar.
ŞEKİL 2.3 Myelinli nöron
Elektrofizyolojik çalışmalarda, uyarının kalın miyelinli aksonlarda bir Ranvier düğümünden diğerine atlar. Bu tür uyarı “Saltatorik Akım” denir. Mitokondriyonların Ranvier düğümlerinde bol olarak bulunmasının sebebi taşınma olayları için ihtiyaç duyulan enerjinin bu organellerce karşılanmasıdır. Miyelin tabakasının üzerini nörolemma denen membran çevreler. Bu membranı schwan hücreleri meydana getirir. Schwan hücresi önce bir tabaka halinde aksonu sarar. Daha sonra daireler halinde birçok tabaka teşkil ederek miyelin tabakasını oluşturur. Nörolemmanın üzerinde İnce ve retiküler fibrillerden yapılmış bir zar bulunur buna endonerium denir
1-Miyelinli akson 2-Miyelinsiz akson

H A Y V A N F İ Z Y O L O J İ S İ
95
Gibbs Donan Dengesi
İyonlar membranın bir tarafından diğer tarafına aktarılabilir ve elektrik yüklerinin ayrılması ile membranın iki tarafı arasında elektriksel potansiyel farkı yaratabilirler. Bu iş için iyonların devamlı aktif transportu gereklidir (Na-K ATPaz). Ayrıca membranın bir tarafında membranı geçebilen ve geçemeyen iyonlar var ise, bir potansiyel farkı meydana gelir ve korunur. Eğer membranı geçebilen partiküller sadece pasif olarak hareket edebiliyorlarsa, meydana gelecek iyon konsantrasyonu dağılışına Gibbs-Donnan dengesi denir. Membran içinde (-) yüklü proteinler ve dışarıya göre 30 kat fazla K+ iyonu bulunur. Membran dışında ekstraselüler sıvıda 10 kat fazla Na+ iyonu ve 14 kat fazla Cl- iyonu vardır. Difüzyon ile Na+ içeri K+ dışarı çıkma eğilimindedir. Bu durum Na-K pompası ile dengelenmeye çalışılır. Eksi (-) yüklü proteinler membrandan dışarı çıkamazlar. Çünkü büyüktürler. Cl iyonları geçebilir, fakat sinirde mevcut potansiyel fark, Cl iyonlarının çoğunu dışarıda tutmaya yeterlidir. O halde dışarı çıkamayan protein anyonlar ile gerek olmadıkça içeri giremeyen Cl iyonları bulunmaktadır. Na-K iyonları sinir impulsu oluşumu açısından önemlidir. Membranın Na+ a karşı geçirgenliği azdır; ama yine de bir miktar Na+ konsanatrasyon gradiyentine göre içeri sızar. Fakat içeri sızan sodyum (Na+ ) pompa sistemi tarafından dışarı atılır. Böylece dışarda yüksek Na+ konsantrasyonu muhafaza edilir. K+ iyonları membranı kolayca geçebilir. İçerde K+ çok olduğundan süratle dışarı sızar. İçerisi potasyum (K+ ) kaybettiği için de içerde bulunan (-) yüklü proteinlerden dolayı membran içi dışarıya oranla -70 milivolt kadar bir potansiyel farkı gösterir. Fakat potasyum(K+’un) dışarı çıkışının da bir sınırı vardır. İçerdeki (-) yüklü organik anyonların, (+) yüklü K iyonlarını çekmesi, dışarıdaki (+) yüklü Na iyonların gene (+) yüklü olan K iyonlarını itmesi gibi nedenlerle dışardakinin 30 kat K+ içerde muhafaza edilir.
ŞEKİL 2.4 Nöron ve sinaps
• En basit refleks yayı (hem duysal bilgiyi alan hem de efektör hücreyi kontrol eden aynı nörondur).
• Monosinaptik refleks yayı (duysal bilgiyi alan nöron merkezi sinir sistemindeki bir motor nöron ile sinaps yapar).
• Polisinaptik refleks yayı (nöronal devirde birden fazla sinaps vardır).

H A Y V A N F İ Z Y O L O J İ S İ
96
Akson Dendirit Nöron gövdesinden bilgi alırlar. Nöron gövdesine bilgi getirirler Yüzeyleri düzdür. Yüzeyi düz değildir. Bir nöronda bir akson bulunur. Bir nöronda çok sayıda dendrid vardır. Hiç ribozom yoktur. Ribozomları vardır. Miyelinli olabilirler Miyelinleri yoktur. Nöron gövdesinden uzakta dallanırlar Nöron gövdesinin yakınında dallanırlar. Refleks Olayı
Refleks, genel anlamda bir uyarıya verilen yanıttır. Ya da bir reseptörün (duyu alıcısı) uyarılması ile tepki organında oluşturulan istek dışı aktivitedir. Ancak sinir sisteminin çalışmasının temel mekanizmasını refleks oluşturur. Yani canlılardaki birçok davranış refleks ile sağlanmaktadır. Diğer bir anlatımla hem isteğimizle hem de isteğimiz dışındaki sinirsel aktiviteler aynı mekanizma ile gerçekleşmektedir. Duyu organlarına alınan ani, hızlı ve güçlü uyarılara karşı kaslar, bezler gibi uyarılabilir organlarla tepki gösterilmesine refleks denir. Refleks davranışları bilinçsiz olarak yapılır ve omurilik tarafından yönetilir. Refleks olayı vücudu koruyan bir mekanizmadır.
ŞEKİL 2.5 Refleks arkı
En basit bir davranışın başlatılması için en az bir reseptör, bir getirici nöron (afferent),bir merkez (omurilik), b götürücü (efferent) nöron ve bir yanıt organı gereklidir. İşte bu yapıya refleks yayı (refleks arkı) denir. En basit refleks yayı (hem duysal bilgiyi alan hem de efektör hücreyi kontrol eden aynı nörondur). Monosinaptik refleks yayı (duysal bilgiyi alan nöron merkezi sinir sistemindeki bir motor nöron ile sinaps yapar). Polisinaptik refleks yayı (nöronal devirde birden fazla sinaps vardır).
Refleks Arkı Herhangi bir sinir işlevinin yapılabilmesi için gerekli bir reseptör, bir afferent nöron, bir refleks merkezi, bir efferent nöron ve bir de faaliyet organından kurulmuş anatomik bir yapıdır.

H A Y V A N F İ Z Y O L O J İ S İ
97
ŞEKİL 2.6 Refleks yayı tipleri
Sinir iletisi
Nöronun kendisine gelen uyarıya hücre zarı üzerinde bulunan elektriksel potansiyel farkında hızlı değişiklik ile 'yanıt' vermesi, ona ileti özelliği kazandırır. Hücre membranının her iki tarafında pozitif yüklü Na ve K iyonları ile negatif yüklü aminoasit ve proteinlerin eşit olmayan dağılımı Membranın potasyuma karşı geçici geçirgenliği bağlı gerçekleşir. Uyarı iki kaynaktan başlar : Dış kaynaklar ve Duyusal bilgi akışı, İç kaynaklar: Nöronların membran potansiyelleri aracılığı ile sahip olduğu spontan aktivite ile başlar. Aksiyon potansiyeli zarın bir yerinden başladığında bütün zara yayılır; durdurulamaz.
� Uyarı, sinir hücre zarı boyunca giden bir elektriksel dalgadır. Sinir hücre zarında oluşan potansiyeli farklı tiplerde olabilir.
• Lokal Potansiyel: Bir çok nöronda dendritler yoğun Ca2+ ve K+ kanalları ile daha az sayıda Na+ kanalı içerir. Bu bölge Aksiyon potansiyeli oluşturmaktan daha çok elektrotonik sinyalleri toplayarak hücre gövdesinde değerlendirilmesini sağlar. Lokal Potansiyeller dereceli potansiyellerdir. Uyarı ne kadar büyük ise Lokal potansiyel kadar büyük olur.
• Tetikleyici Potansiyel: Aksiyon Potansiyeli, aktif bir nöronda zar boyunca her tarafa yayılan bir elektriksel dalgalanmadır. Uyarı ile Na+ kapıları açılır; Lokal potansiyel oluşur (depolarizasyon). Eğer lokal potansiyel eşik değeri aşarsa aksiyon potansiyeli oluşur. Nöronun tetikleme bölgesi (trigger zone) ise, yoğun Na+ kanallarından dolayı AP oluşumu için en elverişli bölgedir. Akson üzerinde çok sayıda bulunan Na+ ve K+ kanalları AP'nin hızlıca iletilmesini sağlar. Akson ucu (presinaptik) bölge ise çok sayıda Ca2+ kanalı bulundurur.
Sinir iletimde gerçekleşen olaylar:
1. Depolarizasyon
2. Na kanallarının açılması( voltaja bağlı Na kanalları) 3. Na hücre içine girişi
4. K kanallarının açılması(voltaja bağlı K kanalları)
5. Repolarizasyon 6. Hiperpolarizasyon
7. Dinlenme potansiyeli (Na-K ATPaz ile geri pompalama).

H A Y V A N F İ Z Y O L O J İ S İ
98
Membran Dinlenme potansiyeli
Na+-K+ pompası ve nöronların birçoğunda bulunan Cl- pompası da istirahat membran potansiyeline katkıda bulunur. Pasif Na+ ve K+ akışı aktif iyon pompası ile dengelenir. Her siklusta: [3Na+], [2K+] Pompanın her siklusu bir molekül ATP'nin hidrolizi ile gerçekleşir. Membran bazı iyonların difüzyonunu engellediği için membranın iki tarafı arasında bir “elektriksel potansiyel” farklılığı oluşur. Uyarılmamış nöronda membranın iki tarafındaki şarj farkı ~70 mV ’dur. Bu, nöronun iç tarafının dış tarafa nazaran 70 mV daha az olduğu anlamına gelir. Dinlenme sırasında, nöronun dış tarafında nispeten daha fazla sodyum, iç tarafında ise potasyum iyonu fazladır. Membran dinlenme potansiyeli: Nöronda: -70 mV, Kalp ve iskelet kasında: -90 mV kadardır. Membran potansiyeli iyon hareketleri ile değişir. Difüzyon- konsantrasyon gradyanına doğru ya da zıt elektrik şarjına doğru hareket etmeye neden olur. Aktif transport-ATP kullanarak konsantrasyon gradyanına karşı hareket sağlar. İstirahat membran potansiyeli Hücre uyarılmadığı durumda, hücrenin içi ve dışına devam eden pasif iyon akımları dengelenir ve potansiyel sabit kalır.
ŞEKİL 2.7 aksiyon potansiyeli
Aksiyon potansiyeli, bir elektriksel aktivitenin başlamasıdır. Bu, bir uyarıcının nöron dinlenme potansiyelinin 0 mV’a doğru çıkmasına neden olduğu anlamına gelir. Depolarizasyon, -55 mV’a ulaştığızaman bir aksiyon potansiyeline neden olur. Bu eşik değeridir. Eğer nöron bu kritik eşik noktasına ulaşamazsa, aksiyon potansiyeli oluşmaz. Eşik değerinin aşılması durumunda da oluşacak aksiyon potansiyeli değişmez sabit seviyede kalır. Aynı şekilde her türlü nöronda oluşan aksiyon potansiyeli aynıdır. Bir sinir hücresinde büyük veya küçük aksiyon potansiyeli oluşmaz, bütün aksiyon potansiyelleri aynı büyüklüktedir(aynı elektriksel değişimi gösterir). Na+/K+ ATPaz pompası: 3 Na+’u hücre dışına atarken 2 K+’u hücre içine taşır: Hücre içinde [anyon] artar. İstirahat durumunda hücrenin iç tarafında negatif, dış tarafında pozitif iyonlar birikir. Uyarılmadığı durumda sadece K+ geçirgen olan bir hücrede, anyonların itmesiyle hücre dışına K+ difüzyonu istirahat membran potansiyelinin oluşmasını sağlar. İyon Kanalları
Nöron eşik değere ulaşmadıkça da aksiyon potansiyeli oluşmaz. Bütün bu durumların meydana gelmesi için nöron membranındaki özel kanallar fonksiyonunu yapması gerekir. Bu kanalların bir kısmı voltaj değişimi ile açılıp kapanırken bir kısmı ise sürekli açık konumdadır.
Nöronda ileti Bir nöronda eşik değeri üstündeki aksiyon potansiyeli oluşur. Meydana gelene iyon değişimi sonucu nöron zarı +30 mV kadar yükselir daha sonra repolarizasyonla ve hiperpolarizasyonla zar potansiyeli -70 mV geri gelir

H A Y V A N F İ Z Y O L O J İ S İ
99
• Seçici geçirgenlik kanallar:İyonların geçişini düzenleyerek sinir ve kas gibi hücrelerde membran potansiyelinin hızlı değişimi ve aksiyon potansiyelinin oluşumuna aracılık ederler. Her biri belli iyonların taşınması ile görevlidir. Bu seçicilik ise iyonun hacimsel büyüklüğüne bağlıdır. Sinir hücre membranlarında zarları belirlenmiş, K+ sızıntı kanallarından çok daha fazla Na+ sızıntı kanalları vardır. Bunun sonucu olarak membran sodyuma göre potasyuma daha geçirgendir. Bazı anorganik ve organik moleküller, çeşitli iyon kanalı aktivitesini modüle edebilir Bu maddelere kanallın çalışmasını engeleyebilir: Red tide durumunda dinoflagellate tarafından oluşturlan bir toksin olan Saxitoxin voltaj bağımlı sodyum kanalları bloklar. Önemli lokal anesteziklerden Lidokain ve Novokain sodyum iyon kanallarını bloke eder. Mamba yılanı Dendrotoxin potasyum kanallarının bloklar. Buthus tamulus (Doğu Hint akrep) tarafından üretilen Iberiyotoksin tarafından potasyum kanallarının bloklar.
ŞEKİL 2.7 Sinapsları inhibe eden bazı maddeler.
• İyon kanalları: Kanallar seçicidir; açılıp kapanma özellikleri ve oranları açısından farklı
özellikler gösterirler. Anyon-seçici kanallar da, katyon seçici kanallar da genellikle tek bir iyon için seçicidir. Bazı katyon-seçici kanallar Na+, K+, Ca2+ ve Mg2+ gibi katyonların tümünün geçişine izin verir. Aksiyon potansiyeli sırasında, sinir membranın depolarizasyonu ve repolarizasyonunda önemli rol oynayan voltaj kapılı sodyum kanallarıdır. Voltaj kapılı potasyum kanalları ise repolarizasyonun hızla yükselmesinde önemli rol oynar. Bu iki voltaj kapılı kanal Na-K pompası ve Na-K sızma kanallarına ek olarak görev yaparlar. Sürekli açık olan K+-sızıntı (leak) kanalları: K+ konsantrasyon gradyanı yönünde (elektrik gradyanın aksi yönünde) hücre dışına net çıkış yapar.

H A Y V A N F İ Z Y O L O J İ S İ
100
ŞEKİL 2.8 Voltaja bağlı iyon kanalı(Na iyonu) K kanalınında neden Na geçemediği.
Uyarılabilir hücre zarlarında bulunan tüm sodyum kanallarının küçük ayrıntılar dışında benzerlik gösterdiği anlaşılmıştır. Bu kanallar paralitik ve öldürücü etkileri olan, suda çözünebilen iki önemli zehir, tetrodotoxin (TTX) ve saxitoksin (STX) tarafından bloke edilmektedir. Bazı lokal anestezikler ise kanal içindeki farklı bağlanma yerlerine bağlanarakkanalı bloke etmektedirler. Kapı mekanizması dış ortamdaki iki değerlikli iyon konsantrasyonundan ve pH dan etkilenmektedir.
ŞEKİL 2.9 Na ve K sızma kanalları
• Ligand kapılı: Nörotransmitterin reseptöre bağlanmasıyla açılırlar. Ligand-kapılı kanallarda enerji, kimyasal ajanın proteine bağlanması sırasında ortaya çıkan kimyasal reaksiyondan sağlanır. Ligandın (nörotransmitter ya da kimyasal ajan olabilir) iyon kapısı üzerindeki bir reseptöre bağlanması ile (direct, short-lasting) veya ayrı bir reseptöre (second messenger) bağlanması ile (indirect, long-lasting) kapı açılır. Ligand-kapılı kanallar ligandın varlığı devam ettikçe refrakter döneme girebilir (kanalın duyarsızlaşması).Bazı protein kanal kapıları, diğer bir molekülün protein ile bağlanmasıyla açılır(Nörotransmiter, Parakrin maddeler ve NO). Kimyasal kapıların en önemli örneklerinden birisi asetilkolin kanalı üzerine asetikolin etkisidir asetikolin liganta bağlı Na kanalının kapısını açar.
1- Na sızma kanalı 2- K sızma kanalı 3- Ca sızma kanalı

H A Y V A N F İ Z Y O L O J İ S İ
101
ŞEKİL 2.10 Liganta bağlı Na kapısı.
ŞEKİL 2.11 Liganta bağlı kanala proteinleiri
• Voltaj kapılı kanallar: Hücre zarının içi ve dışı arasındaki elektrik yük farklılığına bağlı olarak açılırlar Voltaj-kapılı kanallar için açık ve kapalı durumlar arasındaki süre bir kaç ms ile dakika arasıdır. Bir kanalın kapalı durumdan açık duruma geçmesi enerji gerektirir. Voltaj-kapılı kanallarda enerji, kanal proteini üzerindeki duyarlı bir bölgenin (voltage-sensor) voltaj artışı sonucu yüklenerek membran üzerinde net bir serbest enerji değişimine neden olması ile sağlanır Voltaj-kapılı kanalların birçoğunda aktif açık olma durumunu sona erdiren bir refrakter (inaktivasyon) dönem görülür. İnaktivasyon durumu kanalı oluşturan proteinin alt ünitelerinden biri ya da bir bölgesi tarafından sağlanır. Na(m ve h) ve K(n) kanalları. Aktivasyon kapısını açan voltajın artması aynı şekilde inaktivasyon kapısını kapatır, inaktivasyon kapısı, aktivasyon kapısının açılmasından sonra saniyenin onbinde biri kadar kısa sürede kapanır. Yani inaktivasyon kapısını kapalı duruma getiren konformasyon değişikliği yavaş bir olay olduğu halde; aktivasyon kapısını açan konformasyon değişikliği hızlı bir olaydır. Bu nedenle, sodyum kanalı saniyenin onbinde biri kadar süre açık kaldıktan sonra kapanır ve sodyum iyonları artık membranın içine akamaz. Bu anda membran potansiyeli, membran dinlenme dönemine geri dönmeye başlayarak repolarizasyon gelişir. Sodyum kanal inaktivasyonun önemli bir özelliği,

H A Y V A N F İ Z Y O L O J İ S İ
102
inaktivasyon kapısının membran potansiyeli başlangıç membran dinlenme potansiyeline veya ona yakın bir değere dönünceye kadar tekrar açılmamasıdır. Bu nedenle, sodyum kanallarının sinir lifi repolarize olmadan tekrar açılması mümkün değildir. Kapının moleküler konumu hücre membranının elektriksel potansiyeline bağlıdır. Normal şartlarda kapalı olan kapı hücre zarının elektriksel değişimine göre açılır. Hücre membarınının içi tarafı negatif yükünü kaybettiği zaman bu kapılar aniden açılır ve çok büyük miktarda sodyumun, sodyum kanallarından geçmesine izin verir Nöronlarda sinyallerin doğuşundan sorumlu aksiyon potansiyelinin temel nedenidir. Na voltaj kapıları sinir hücrelerinin eşik değerinin belirlenmesine önemli rol oynar.
-
ŞEKİL 2.12 Kanal proteinleri ve voltaj kapılı iyon kanalı. • Mekanik kapılı kanallar: Bu kanal tipi gerilim ve basınca duyarlıdırlar. Hücre
iskeletinin(cytoskeleton) gerilmesi ile açılırlar. Mekanik olarak aktive edilen kanallarda enerji, doğrudan membran tarafından ya da sitoskeletal yapılar tarafından iletilir. Böbrek aferent arteriyolün içini döşeyen düz kas hücrelerinde, damar duvarı gerildiğinde (örn. luminal basınçta artışa bağlı) aktive olan Ca2+’a geçirgen mekanosensitif kanallar bulunur.
Aksiyon Potansiyeli
Zar potansiyelinde dinlenim seviyesinden geçici değişmeler, elektriksel sinyal oluşur. Böylesi değişiklikler sinir hücrelerinin en önemli bilgi işleme ve iletme yoludur. Bu sinyaller iki şekilde olur: dereceli potansiyeller ve aksiyon potansiyelleri. Dereceli potansiyeller 3–5 mV’a sahip, ilerledikçe azalan ve kısa mesafeli sinyal iletimlerinde önemli iken aksiyon potansiyelleri sinir ve kas zarlarının uzun mesafeli sinyalleridir. Aksiyon potansyeli, hücre zarında oluşan kısa süreli ve bilgi taşıyan değişikliklerdir. Depolarize, repolarize ve hiperpolarize terimleri zar potansiyelinin dinlenim potansiyeline göre değişiminin yönünü tanımlamak için kullanılır. -70 mV’daki dinlenim zar potansiyeline sahip hücre polarizedir. Zar potansiyeli dinlenim seviyesinde daha az negatif (0’a daha yakın) olduğu zaman zar depolarizedir. Aşma potansiyeli (overshoot) zar potansiyel polaritesinin yani hücre içinin dışına göre pozitif olmasını anlatır. Depolarize olmuş bir zar potansiyeli dinlenim seviyesine geri döndüğü zaman repolarize olmuştur. Potansiyel dinlenim seviyesinden daha da negatif olduğu zaman hiperpolarizedir.

H A Y V A N F İ Z Y O L O J İ S İ
103
Dereceli potansiyeller plazma zarının küçük bir bölgesine sınırlı zar potansiyel değişmeleridir Genellikle hücrenin çevre ortamında bazı özel değişmeler zarın özelleşmiş bir bölgesini etkilediği zaman oluşurlar. Dereceli potansiyel denmesinin nedeni potansiyelin büyüklüğünün değişebilir (derecelenebilir) olmasından dolayıdır. Yaptıkları işleve veya potansiyelin yerleşimine göre dereceli potansiyellere reseptör potansiyeli, sinaptik potansiyel ve prepotansiyel gibi isimler verilir. Dereceli potansiyeller birikerek birikerek aksiyon potansiyeli oluşumu için eşik değeri aşabilir. Aksiyon potansiyelleri dereceli potansiyellerden çok farklıdırlar. Bunlar zar potansiyellerindeki büyük değişimlerdir (-70mV’dan +30/+100 mV’a kadar değişebilir). Her aksiyon potansiyeli, normal negatif zar dinlenim potansiyelinden pozitif potansiyele birtakım iyon/iyonların zardan içeri girmesiyle ani bir değişme ile başlar ve hemen hemen aynı hızla (1–4 milisaniye) zardan birtakım iyon/iyonların çıkışıyla tekrar negatif potansiyele döner. Bu olay bir saniyede birkaç yüz kez tekrar tekrar oluşabilir. Bir sinir sinyali iletisi, aksiyon potansiyeli sinir lifi boyunca yayılarak sinir ucuna gelinceye kadar sürer. Sinir ve kas hücreleri gibi bazı endokrin, immün ve üreme hücreleri de aksiyon potansiyeli oluşturma yeteneği olan plazma zarlarına sahiptirler. Bu zarlara uyarılabilir zarlar denir ve onların aksiyon potansiyeli oluşturma yeteneği uyarılabilirlik olarak bilinir. Tüm hücreler dereceli potansiyeller oluşturma yeteneğine sahipken, sadece uyarılabilir hücreler aksiyon potansiyeli oluşturabilir. Uyarılabilen bir hücre (veya doku), fiziksel, kimyasal veya elektriksel bir uyaran ile depolarize edilebilir. Bunun için uyaran şiddetinin hücreye yeteri kadar pozitif yüklü iyon geçişine yol açması gerekir. Hücre içine anyon (+) yük girişiyle, membran potansiyeli ateşleme seviyesine kadar (-50; -60mV) değiştirilebilirse, Na+’un bütün voltaja bağlı kanalları açılacağından; Na+, hızla hücre içine akar ve bir aksiyon potansiyeli başlar.
ŞEKİL 2.13 Aksiyon potansiyeli sırasında Na K değişimi
Eğer verilen uyarı, eşik altı değerde ise ve membran potansiyelini ateşleme seviyesine çıkaracak pozitif yükün hücreye girişini sağlayamadı ise, istirahat membran potansiyeli hafifçe pozitif yönde değişir, örneğin -70 iken, -68 olur, ancak depolarizasyon yönündeki bu küçük potansiyel değişiklik bir aksiyon potansiyeline yol açamaz. Uyarılabilen bir hücre eşik ve eşiküstü bir uyarana tek tip bir aksiyon potansiyeli ile yanıt verir. Eşik altı uyaran ile bir aksiyon potansiyeli oluşmaz. Buna “Hep veya Hiç Yasası” denir.

H A Y V A N F İ Z Y O L O J İ S İ
104
ŞEKİL 2.14 Ya hep ya hiç kuralı
• Membran üzerinde belirli sayıda kanalın açılarak Na+ geçişini başlatması diğer Na+ kanallarının da hızla açılarak hücre içinde hızlı bir elektropozitif durum oluşturur ve aksiyon potansiyeli oluşur (Eişik değerin açılması: HEP)
• Membran üzerinde belirli sayıdan az Na+ kanalının açılması bu tetiklemeyi gerçekleştirmez ve aksiyon potansiyeli oluşmaz (Eşik değerin aşılamaması: HİÇ)
Vücudumuzda aksonlar ve iskelet kası hücreleri tek tek değil; binlercesi bir araya gelerek sinir demetlerini (siyatik sinir gibi) ve kas kütlelerini (gastroknemius kası gibi) oluşturur. Böyle bir sinir veya kas preparatı elektriksel olarak uyarıldığında, önce düşük eşiğe sahip olan liflerde aksiyon potansiyeli (kas ise kontraksiyon), uyaran şiddeti arttırıldıkça eşiği yüksek olan diğer liflerde aksiyon potansiyeli (kas ise kontraksiyon) oluşur. Bilgisayar ortamında(örneğin PowerLab) yazdırılan eğriler de çeşitli amplitütlerde ve çok sayıda spike potansiyeli (sivri potansiyel) olan eğriler şeklindedir. Buna ‘birleşik aksiyon potansiyelleri’ (CAP) denir. Uyaran şiddeti arttıkça, uyarılan lif sayısı da artar. Bir sinir preparatında bütün liflerde aksiyon potansiyeli oluşturabilen uyarana; maksimum uyaran şiddeti denir. Depolarizasyon-Hiperpolarizasyon
Hücre içersindeki ve dışarıdaki iyon yükü farkı - 60mV olunca tüm akson boyunca voltaj kapılı Na kanalları açılmaya başlar İçeri daha fazla Na girer ve aksiyon potansiyeli oluşur. İyonların voltaj-kapılı kanallardan hareketi sonucu ortaya çıkar Na+, bazı durumlarda Ca2+ iyon kanallarının bu iyonların hücre içine hareketine izin vermesi membranı depolarize eder. Membran repolarizasyonundan sorumlu en önemli iyon: K+ dur. Her aksiyon potansiyeli, normal dinlenmede negatif potansiyelden pozitif membran potansiyeline ani bir değişme ile başlar ve hemen hemen aynı hızla tekrar negatif potansiyele döner. Bir sinir sinyalinin iletisinde, aksiyon potansiyeli sinir lifi boyunca sinir ucuna gelinceye kadar yayılır. Hücre zarında bulunan; iç kısmı su ile dolu integral bir membran proteini ile onun yüzeyine tutunmuş karbonhidrat gruplarını içeren; hidrofilik yapıda olduğu için hücre zarının lipid kısmından geçiş yapamayan iyonların taşınması ile görevli kanallardır. İmpulsun sinir boyunca yayılması
İmpuls meydana geldikten sonraki evre impuls yayılmasıdır. Bu safhada Aksiyon potansiyeli akson tepesinden terminal düğüme kadar ilerler. İletim değeri hücrenin özel uzunluk sabiti ile orantılıdır. Genellikle bu değer silindirik yapılarda silindir çapının artmasıyla artar. Öyleyse geniş çaplı silindirlerde iletim daha hızlı olmaktadır.
Ya hep ya hiç Tek bir sinir fibrilinin, tek bir kas fibrilinin ve kalp kasının eşik şiddetten düşük uyarana cevap vermemesi, eşik şiddette uyarılması ve şiddet arttıkça aksiyon potansiyelinin yükselmemesi olayına ya hep -ya hiç prensibi denir

H A Y V A N F İ Z Y O L O J İ S İ
105
İmpulsun yayılmasına impulsun da etkisi vardır. İmpuls hızı ısının artmasıyla artar. Bu yüzden sıcakkanlı hayvanlarda iletim hızı soğukkanlı hayvanlara göre fazladır. Ortamın pH’sı, oksijen miktarı da impulsun hızı üzerinde etkilidir. Akson tepesinde membran bir aksiyon potansiyeli meydana getirmek üzere uyarılırsa potansiyelde meydana gelen bir değişiklik membranın bitişik bölgelerini etkileyecektir. Uç potansiyel noktasında aksonun içi dışarıya oranla (+) yüklüdür (+40mV).
ŞEKİL 2.15 Nöronda uyarının yayılması
Membranın iç tarafında (+) yüklü iyonların benzer yüklü iyonların birbirini itmesiyle aksiyon potansiyeli bölgesinden kovulurlar. Membranın dış yüzünü daha az pozitif yapan Na+ iyonları hücre içine hareket eder, K+ iyonları ise hücre Dışını daha fazla pozitifleştirmek için aksiyon potansiyeli bölgesine doğru membranın dış yüzüne hareket ederler. Bu iç ve dış elektrik akımı membrannın bitişik bölgelerini etkiler. Eğer bu değişiklik başlatıcı uyarıya (en az –50 mV ya da –60 mV) ulaşırsa membranın bitişik bölgesinde birinci aksiyon potansiyeline eşdeğer bir aksiyon potansiyeli gelişir. böyece impuls aksiyon potansiyeli şeklinde tüm akson boyunca yayılır. Omurgalı hayvanların periferal sinir sistemindeki nöronların aksonlarının etrafında miyelin kılıf olabilir ya da olmayabilir. Akson etrafındaki miyelin kılıf, beyin ve omurilik içindeki sinirlerin aksonlarında oligodendroglial hücreler tarafından oluşturulur. Beyin ve omuriliğin dışındaki yapılarda bulunan sinir hücrelerinin aksonları etrafında bulunan miyelin tabaka ise Schwan hücreleri tarafından oluşturulur. Bildiğiniz gibi miyelin kılıf akson boyunca 1 mm’lik uzunluklarla kesilir (Ranvier boğum). Miyelin kılıf çok iyi bir izalotördür. Miyelin kılıfla sarılı aksonda difüzyon olmaz. O yüzden miyelinli ve miyelinsiz aksonlarda impulsun iletilmesi farklı olur.
Nöronları aktsiyon potansiyeli Dendritlerden hücre gövdesine, Hücre gövdesinden aksona doğrudur. Uyarılan sinir zarından pozitif yük ile negatif yük değişimi gerçekleşir.

H A Y V A N F İ Z Y O L O J İ S İ
106
ŞEKİL 2.16 Myelinli nöronda ileti yayılması
Miyelinsiz sinirlerde impuls bütün sinir lifi boyunca yayılırken miyelinli sinirlerde aksiyon potansiyelinin iletilmesi sadece miyelin kılıfın olmadığı Ranvier boğumlarında olmaktadır. Dendritleri ve hücre gövdesinden uyarım alan sinir hücresi öncelikle bu sinyali kendi aksonu boyunca taşınır. Bu taşıma sırasında miyelin kılıf enerji tasarrufu bakımından önemli bir rol oynar. Çünkü bu kılıf boyunca hiçbir protein kanal bulunmaz. Dolayısıyla iletim, miyelin kılıflar arasında yer alan Ranvier boşlukları’nda gerçekleşir. Bir boşluktan diğerine atlayan elektriksel sinyal, iletimi hızlandırmış olur. Üstelik miyelin kılıf boyunca enerji harcayan sodyum/potasyum pompaları bulundurmayan hücre enerjisini de korur. Sinir sistemimizdeki pek çok sinir hücresi miyelin kılıf bulundur. Eğer ki hücreler gerektiğinden daha az miyelin kılıf bulunduruyorsa sinirsel iletim sağlıklı bir şekilde gerçekleşemiyor ve duyularda azalma ya da felç gibi belirtiler ortaya çıkabilir. Mutlak refrakter periyod:
Sinir membranın aksiyon potansiyeli oluşturan kısmı sodyum ve potasyum kanallarının konumu gereği ya hiç uyarı oluşturamaz ya da ancak yüksek uyarı karşı uyarı meydana getirebilir. Bu durumda nörolemna bir süre uyarılamaz; duyarsız kalır. (uyarılara cevap vermez) Buna “Refrakter periyod” denir. Depolarizasyon ve repolarizasyonun erken dönemlerini içerir. Depolarizasyonda m kapısı açık olduğundan repolarizasyonda da h kapısı kapalı olduğundan hücre uyarılamaz. Depolarizasyonun son dönemlerinde h kapıları açılınca Na+ kapılarının inaktivasyonu kaldırılır ve mutlak refrakter peryot biter. Relatif (Nisbi) refrakter periyod:
Bu periyotta uyaranın şiddetli olması durumunda aksiyon potansiyeli oluşturulabilir. Relatif refrakter dönem mutlak refrakter dönemin hemen sonunda başlar. Uyaran daha-şiddetli olmalıdır çünkü az miktarda aktif Na+ kanalı ve fazla miktarda aktif K+ kanalı vardır. Aksonda aksiyon potansiyeli komşu bölgeleri uyararak yayılır. Yayılım myelinli ve myelinsiz liflerde farklı özellikler taşır. Bir reseptöre etki eden uyarını şiddeti arttığında Reseptör veya jeneratör potansiyel şiddeti artar. İlgili afferent sinirde impuls iletim frekansında artar. İlgili duyusal merkez ulaşan impuls frekansı artar. Birim zamanda oluşan aksiyon potansiyeli sayısı artar ancak şiddetinde değişiklik olmaz.

H A Y V A N F İ Z Y O L O J İ S İ
107
ŞEKİL 2.17 Çizgili ve kalp kasında gözlenen refraktörü periyotlar
Bu periyotta uyaranın şiddetli olması durumunda aksiyon potansiyeli oluşturulabilir. Relatif refrakter dönem mutlak refrakter dönemin hemen sonunda başlar. Uyaran daha-şiddetli olmalıdır çünkü az miktarda aktif Na+ kanalı ve fazla miktarda aktif K+ kanalı vardır. Aksonda aksiyon potansiyeli komşu bölgeleri uyararak yayılır. Yayılım myelinli ve myelinsiz liflerde farklı özellikler taşır. Bir reseptöre etki eden uyarını şiddeti arttığında Reseptör veya jeneratör potansiyel şiddeti artar. İlgili afferent sinirde impuls iletim frekansında artar. İlgili duyusal merkez ulaşan impuls frekansı artar. Birim zamanda oluşan aksiyon potansiyeli sayısı artar ancak şiddetinde değişiklik olmaz. Terminal plakta Ca++ eksikliği veya Mg++ fazlalığı Ach salımını inhibe eder.
ŞEKİL 2.18 Refraktörü periyot

H A Y V A N F İ Z Y O L O J İ S İ
108
Reobaz (Eşik değer)
Bir sinirin deneysel olarak uyarılmasında genellikle elektrik akımı kullanılır. Elektrik akımının şiddetini ve süresini hassas olarak ayarlamak mümkündür. Etkili uyarmada uyarıcının (elektrik akımının) şiddeti önemli olduğu gibi devamı da önemlidir. Şiddetli bir uyarma çok kısa zamanda siniri uyarır.
ŞEKİL 2.19 Reobaz eşik değeri
Uyarıcının şiddetini azaltırsak, reaksiyon elde etmek için uyarmanın daha uzun devam etmesi gerekir. Bir siniri uyaran en zayıf etki şiddetine “reobaz” (eşik değer) denir. Rheobaz ‘ın uyarmayı yaratması için belirli bir süre devamı gerekir. Rheobaz’dan daha zayıf bir etki şiddeti, ne kadar uzun devam ederse etsin siniri veya kası uyarmaz. Rheobase’ın iki misli şiddet uygulanınca bunun dokuyu uyarabilmesi için gereken süreye kronakzi denir. Sinaps Bir nöronun diğerinin elektriksel ve kimyasal etkinliğini değiştirdiği, iki nöron arasındaki özelleşmiş anatomik bağlantılara sinaps denir. Çoğu sinapsta sinyal, bir nörondan diğer nörona nörotransmiterler tarafından taşınır. Bir nörondan salgılanan nörotransmiter, diğer nöronun membranı üzerinde bulunan özel bir protein reseptöre bağlanarak onun etkinliğini değiştirir. Sinapsa sinyali getiren nörona presinaptik nöron, sinyali sinapstan alıp uzaklaştıran nörona ise postsinaptik nöron denir.
ŞEKİL 2.20 Aetil kolin

H A Y V A N F İ Z Y O L O J İ S İ
109
Nörotransmiterler presinaptik nöronda veziküllerde depolanır. Bu transmiterlerin depolandığı veziküllerin sinaptik aralığa salınımında esas rol oynayan kalsiyumdur. Depolarizasyon presinaptik bölgeye ulastığı zaman, presinaptik bolgede bulunan voltaj bağımlı (N tipi) kalsiyum kanalı acılır ve presinaptik bölgeye kalsiyum girer. Magnezyum kalsiyumla yarıştığı icin, bu nedenle nörötransmitter salınımını azaltır. Akson terminaline giren kalsiyum Kalmodulin bağımlı protein kinazı aktifler (CaM-K II). Aktiflenen protein kinaz sinapsin I proteini fosforiller ve onu aktif hale getirir. Sinapsin ise, icerisinde nörotransmitter bulunan veziküllerin membrana yaklasmasını sağlar. Diğer taraftan kalsiyum, sinaptotagmin proteinine bağlanır. Bu kompleks sintaksin ve sinaptobrebin (SNARE protein) aktivasyonunu sağlayarak egzositoz ile vezikullerin sinapsa atılmasını sağlar. 1-Nöron kaynaklı kimyasal haberciler( asetil kolin, GABA vb) sinaptik vezikül içinde depolanır. 2-Sinaptik vezikül membranında bulunan vezikül tutucu proteinler bunlara karşılık gelen presinaptik membranda bulunan mebran tutucu proteinlerle bağlanır( bağlantı sianpsin filamentlerince yapılır. 3-Akson depolarizasyona bağlı olarak açılan kalsiyum kanallarına bağlı olarak kalsiyum artışı ve bunun sonunda veziküllerin ekzositozu meydana gelir. 4- Sinaptik aralığa salınan Nöron kaynaklı kimyasal haberciler postsianpti membrandaki reseptörlere(kolinerjik yada adrenerjik) bağlanır. 5- Nöron kaynaklı kimyasal haberciler sinaptik aralıkta enzimlerce parçalanır.(asetil kolin için asetil kolinesteraz). Norepinefrin gibi haberciler ise reseptör aracılı endositoz ile alınarak mitondriyal bir enzim olan MAO ile paraçalanır.
ŞEKİL 2.22 Elektriksel ve kimyasal sinaps tipleri
Elektriksel sinapslar : Az sayıda sinaps, iletileri doğrudan ileten elektriksel sinapslardır. Elektriği gap junctionlar aracılığı ile bir hücreden diğerine ileten direkt kanallardır. Bir çok sinapsta iki nöron arasındaki açıklık (aralık sinaptik yarık yaklaşık 20 nm’dir)

H A Y V A N F İ Z Y O L O J İ S İ
110
fazladır ve 2 nöron asla birbirleriyle temas etmez. Bazı durumlarda ise küçük protein tübüllerinden oluşan bağlantılar vardır(Konneksin proteini, Konnekson). Çok az durumda hücre membranları arasındaki mesafe çok dardır. Akım direkt olarak diğer hücreye atlar. Depolarizan uyarılar gönderirler, inhibitor etkinliğe ya da etkinlikteki uzun süreli değişikliklere izin vermezler. İki nöron arasında sitoplazmik bir devamlılık söz konusudur. Akım presinaptik hücreden postsinaptik hücreye zaman ve mesafe sabitelerine göre pasif bir akışla geçer. Elektriksel geçiş eğimlidir ve presinaptik hücredeki akım aksiyon potansiyeli eşiğinin altında olsa bile postsinaptik hücrede pasif depolarizasyon gerçekleşir. Elektriksel sinapslar reflekslerin hızlı koordine olması gereken omurgasız sinapslarında, Balıklarda Mauthner hücrelerinde(kaçmasını sağlayacak kuyruk fiskesini sağlayan) Korunmak için mor renkli mürekkep salan Aplysia’da gözlenir.
ŞEKİL 2.23 Elektriksel sinapsta aktarılan uyarının azalması
Kalsiyum iyonundaki küçük değişimler akımın ve küçük partiküllerin geçişini azaltır. Hücre içinde pH’ın düşmesi kanaldan akımın geçişini azaltabilir. Konneksin moleküllerinde ikinci haberci sistemler aracılığı ile gerçekleştirilen geriye dönüşümlü fosforilasyon kanal etkinliğini değiştirebilir. Hücreden hücreye akım geçişinde herhangi bir gecikme olmaz. Birçok hücrenin hızlı bir şekilde eşzamanlı uyarılmasını sağlar (örn. kalp kasları) Hücreler arasında metabolik uyarıları geçirebilir. Buna karşın elektriksel sinaps bazı dezavatajları vardır. Küçük bir presinaptik akson büyük bir postsinaptik hücreyi aktive edemez. Presinaptik aksondaki akımın miktarı daha büyük bir postsinaptik hücrede yeterince güçlü bir tepki oluşturmayabilir. Sinaptik uyarıda amplifikasyon oluşamaz. Presinaptik aksiyon potensiyali IPSP’ye dönüşemez. Uyarı kimyasal sinapslarda olduğu denli modifiye edilemez. Kimyasal sinaps: Elektriksel sinaps göre daha yavaştır. Farklı sinyal üretme, sinyalin artırma olanağı sağlar. Hem eksitatör, hem inhibitor etkinliğe aracılık eder. Kimyasal sinapsta akım tek yönlüdür. Kimyasal nörotransmitterler postsinaptik hücredeki iyon kanallarının açılmasını doğrudan ya da dolaylı etkileyerek kontrol ederler.
• İyonotropik reseptörler Hızlı, iyon akımı Milisaniye seviyesinde çalışan, nörotransmiter olarak Glutamat, GABA, Glisin kullanır.
• Metabotropik reseptörler: Yavaş, İkinci haberciler ile çalışan, Saniye seviyesinde etkili, nörotransmiter olarak Monoaminler, nöropeptidleri kullanır.

H A Y V A N F İ Z Y O L O J İ S İ
111
ŞEKİL 2.24 Sinaps Myelinli nöron
Bir postsinaptik nöron, hücre gövdesi ve dendritleri üzerinde binlerce bağlantıya sahip olabilir, böylece pek çok presinaptik nörondan gelen sinyaller bu nöronu etkileyebilir. Aksiyon potansiyeli presinaptik membrandan postsinaptik membrana, presinaptik membran tarafından salınan nörotransmitter denilen kimyasal maddelerin sinaptik yarıktan geçerek postsinaptik membrana ulaşmasıyla meydan gelir. Bu nörotransmitterler presinaptik nöronun akson ucunda bulunan veziküllerde taşınır. Aksiyon potansiyelinin akson son ucuna gelmesi veziküllerde bulunan nörotransmitterlerin sinaptik yarığa boşalmasına neden olur. Nörotransmitter madde sinaptik yarıktan geçer ve postsinaptik nöronun hücre membranı üzerindeki uygun reseptörlere (proteinlere) etki eder. Bu birleşme postsinaptik membranın permeabilitesini değiştirir. Daha sonra transmitter madde sinaptik yarık içindeki bir hidrolitik enzimle parçalanır ya da presinaptik nöron tarafından geri emilimle alınır ve sinaptik yarıktan uzaklaştırılır
ŞEKİL 2.24 Sinaps Myelinli nöron
Kimyasal Sinaps Sinaps, nöronların (sinir hücrelerinin) diğer nöronlara ya da kas veya salgı bezleri gibi nöron olmayan hücrelere mesaj iletmesine olanak tanıyan özelleşmiş bağlantı noktaları. Bir motor nöron ile kas hücresi arasındaki kimyasal sinaps, aynı zamanda neuromuscular junction nöromusküler bağlantı olarak adlandırılır.

H A Y V A N F İ Z Y O L O J İ S İ
112
Nörotransmiterler çift tabakalı lipit membranlı küçük veziküllerde depolanır. Aktivasyon öncesinde birçok vezikül, presinaptik membranda aktif bölgeler olarak bilinen salınım alanlarında tutunmuş iken diğerleri terminal içinde dağılmıştır. Aksiyon potansiyeli presinaptik membran terminaline ulaştığı zaman nörotransmiter salınımı başlar. Aksiyon potansiyeli uca geldiğinde depolanmış nörotransmitterlerin boşalmasını uyarır. Sinaptik veziküller uyarı ile serbest bırakılırlar. Hücre adezyon molekülleri, sinapsın hem pre- hem de postsinaptik yüzlerinde sinyal molekülleri olarak yardımcı rol alır. Stok olarak bulunan diğer sinaptik veziküller aktin filamanlarına tutunur. Vezikül yüzeyi ile bir kompleks oluşturan sinapsin I, sinaptik vezikülleri bir arada tutar. Sinapsin I fosforilize olduğunda, sinaptik veziküller nörotransmitterleri serbest bırakmaya hazırlık olarak aktif zona doğru serbestçe hareket ederler. Tersine sinapsin I’in defosforilasyonu olayı tersine gerçekleştirir.
Bir nörondan diğer bir nörona ya da bir nörondan effektör bir organa impulsun geçtiği yerlere sinaps denir. Sinaps, aksiyon potansiyeli ile taşınan bilginin geçtiği özel bağlantı yerleridir. Sinaps yapan 2 hücreden 1.si impulsu sinaps yerine getiren diğeri ise impulsu buradan alıp effektör organa götüren hücredir. İmpulsu sinaps bölgesine getiren nörona presinaptik nöron, buradan impulsu alan 2. nörona postsinaptik nöron denir. Tranmisyonu kimyasal madde aracılığı ile olanlara “kimyasal sinaps’’, transmisyonu aksiyon akımı ile olanlara “elektriksel sinaps’’ denmektedir. Kimayasal sinapta kullanılan haberci moleküller akson boyunca motor proteinler olan kynezin tarafında sinaps taşınırken boşalan haberci molekül vezikülleri nöron gövdesine dynenin motor proetini tarafında geri götürülür.
Ekdize Edici Post Sinaptik Potansiyel (EPSP) İnhibe edici post sinaptik potansiyel (IPSP):
Bir nöronun aksonu son ucuna doğru birçok kollara ayrılır ve bu kollar çeşitli nöronlarla sinaptik birleşme kurabilir. Aksonun son kolları çok incedirler fakat sinapsis yapacak son uçlar bir yumru halini almıştır. Yüzlerce ayrı yerden impuls alan tek bir nöron, sayısı yüzlere varan başka nöronlarla bağlantı kurduğu düşünülürse, bir reseptörden alınan impulsun santral sinir sisteminin bütün merkezleri ile bağlantı halinde olduğu anlaşılır. Fakat santral sinir sisteminde her gelen impulsa her yol açık değildir. Ancak gerekli yollar impulsa açılır gereksiz yollar kapatılır. O halde her sinapsiste bir impuls bir nörondan diğerine geçemez. İmpulsu bir nörondan diğerine geçiren sinapsise ‘’ekdize edici sinapsis’’ (impulsu geçiren sinapsis), geçirmeyene de ‘’inhibe edici sinapsis’’ (impulsu geçirmeyen sinapsis) denir Postsinaptik nöronun membran potansiyeli eksitatör (uyarıcı) sinapsta eşiğe daha yaklaşır (yani depolarize) ve inhibitör (baskılayıcı) sinapsta eşikten uzaklaşır (yani hiperpolarize) ya da mevcut düzeyinde sabitleştirilir.
ŞEKİL 2.25 EPSP ve IPSP bağlantısı

H A Y V A N F İ Z Y O L O J İ S İ
113
Herhangi bir anda postsinaptik hücrenin uyarılabilirlik düzeyi o anda aktif olan sinaps sayısına ve bunların eksitatör ya da inhibitör olmalarına ve sayılarına bağlıdır. Postsinaptik nöronun membranı eşik değere ulaşırsa, aksonu boyunca terminal dallarına doğru iletilen aksiyon potansiyeli oluşturur. Eksitatör bir sinapsta, nörotransmitere verilen postsinaptik yanıt membran potansiyelini eşiğe yaklaştıran bir depolarizasyondur. Böyle sinapslarda aktive olan reseptörün postsinaptik membran üzerindeki genel etkisi, postsinaptik membrandaki sodyum, potasyum ve diğer küçük pozitif yüklü iyonlara geçirgen olan iyon kanallarını açmasıdır. Bu iyonlar da, elektriksel ve kimyasal farklarına göre membrandan serbestçe geçerler. Uyarıcı postsinaptik potansiyel durumunda direnç değişimi tüm iyonlarda bir permeabilite artışına (özellikle K+, Na+, Cl-) bağlıdır. Oysaki inhibitor postsinaptik potansiyel durumunda permeabilite değişikliği sadece K+, Cl- ve çoğu kez sadece K+ daki bir değişikliği kapsar. İnhibitör sinapslarda, postsinaptik nörondaki potansiyel değişik inhibitör postsinaptik potansiyel (İPSP) denilen hiperpolarize edici dereceli potansiyeldir. İnhibitör bir sinapsın aktivasyonu, postsinaptik hücrenin eşiğe depolarize olma ve bir aksiyon potansiyeli oluşturma olasılığını azaltır.
Eksitasyon:
Hızlı ya da direkt etkili nörotransmitter sistemi içinde glutamat, asetilkolin (ACh), seratonin ve adenozin trifosfat (ATP), Gama γ-amino bütirik asit (GABA) ve glisin, yavaş yada indirekt etkili nörotransmitter sistemi içinde glutamat, GABA, ACh, norepinefrin, dopamin, seratonin, substans P, histamine, ATP, adenozin ve opioidler bilinmektedir. Görüldüğü gibi bir nörotransmitter hem yavaş hem de hızlı etkili olabilir. Glutamat, ACh, seratonin, ATP ve GABA hem hızlı hem de yavaş etkileri olan nörotransmitterlere örnektir. Örnek olarak ACh için; nikotinik ACh reseptörleri ile ilişkili olarak nöromüsküler kavşaktaki hızlı nörotransmisyon ve muskarinik ACh reseptörleri ile ilişkili yavaş etki olarak bronkokonstruksiyon, salivasyon ve gastrointestinal motilite verilebilir. Bu nörotransmitter sistemlerinin çoğu anestezik ilaçlar ile etkilenir. Eksitatör hızlı nörotransmitterler (glutamat, ACh, seratonin ve ATP) ligand-kapılı iyon kanallarını açarak öncelikle Sodyum ve kalsiyum geçişine izin verirler ve hücre daha kolay uyarılabilir hale gelir. İnhibitör hızlı nörotransmitterler (GABA ve glisin) ise klor (Cl-) geçişine izin vererek hücre içindeki negatif yükü arttırır ve hücrenin daha az uyarılabilir olmasına yol açar.

H A Y V A N F İ Z Y O L O J İ S İ
114
Aksiyon potansiyeli oluşturan nörotransmitter etki mekanizması: 1. Fazla sayıdaki pozitif elektrik yükünün, postsinaptik hücrenin içine akmasını sağlamak üzere sodyum kanallarının açılması. Bu durum, membran potansiyelini eksitasyon için gerekli olan eşik değerine doğru pozitif yönde artırır. Eksitasyona neden olmanın en fazla kullanılan şeklidir.
2. Klorür veya potasyum kanallarından biri veya her ikisi aracılığı ile baskılanmış ileti. Bu negatif yüklü klorür iyonlarının postsinaptik nöronun içine difüzyonlarını veya pozitif yüklü iyonların dışarı difüzyonlarını azaltır. Her iki durumda da içerdeki membran potansiyeli normalden daha pozitif hale gelir ki, bu da eksitasyondur.
3. Hücre aktivitesini uyarmak için hücrenin iç metabolizmasındaki çeşitli değişiklikler veya bazı durumlarda, eksitatör membran reseptörlerinin sayısında artış ya da inhibitor membran reseptörlerinin sayısında azalma olmasıdır.
ŞEKİL 2.26 Esitatör ve inhitör amino asitler
Serotonin direkt ya da indirekt etkili olabilen diğer bir eksitatör nörotransmitterdir. Trombositler, sindirim kanalı, beyin ve az miktarda da retinada tanımlanan G-proteinine bağlı adenil siklaz ve fosfolipaz C’yi değiştiren 7 farklı reseptörü vardır. Sindirim kanalı ve area postremada bulunan iyonotropik 5HT3 reseptörleri kusmadan sorumludur. 5HT3 reseptör antagonistleri antiemetik olarak yaygın kullanılmaktadırlar. 5HT6 reseptörlerinin de antidepresanlara afinitesi yüksektir. Serotonin geri alım inhibitörleri de sinaptik aralıkta serotonin konsantrasyonunu arttırır ve depresyonu azaltır. Ancak fazlalığının da panik atağa yol açabileceği bildirilmiştir.
İnhibisyon:
Aksiyon potansiyeli oluşmasına engel olan nörotransmitter etki mekanizması: 1. Reseptör molekülü aracılığıyla klorür iyon kanallarının açılması. Bu durum negatif yüklü klorür iyonlarının postsinaptik nöronun dışından içine doğru hızlı difüzyonuna izin verir, böylece negatif yükleri içeri taşır ve içerdeki negativiteyi artırır ki bu da inhibisyondur.

H A Y V A N F İ Z Y O L O J İ S İ
115
2. Reseptör aracılığıyla potasyum iyonlarının iletiminde artış. Bu artış, pozitif potasyum iyonlarının dışarı difüzyonunu sağlar ki, bu da yine inhibisyondur.
3. Hücresel metabolik fonksiyonları inhibe eden veya inhibitör sinaptik reseptörlerin sayılarını artıran veya eksitatör reseptörlerin sayısını azaltan reseptör enzimlerinin aktive edilmesi. GABA beyindeki ana inhibitör transmitterdir. İyonotropik GABAA ve GABAC (Cl- iyonu geçişine izin verir) ve metabotropik GABAB (G proteinlerine bağlı olup K+ kanallarında geçirgenliği arttırırken adenilil siklazı ve buna bağlı Ca+ akışını inhibe eder) bilinen üç GABA reseptörüdür. GABAB ve GABAC reseptörleri nikotinik ACh ve birçok glutamat reseptöründeki gibi bir kanal oluşturacak şekilde yan yana dizilmiş beş alt birimden oluşur. GABAA reseptörleri de beş alt birimden oluşur, ancak burada altı α, dört β, dört γ, bir δ ve bir ε bulunur. Bu çok sayıda farklı bağlanma yerleri nedeni ile GABAA çok sayıda ilaç için hedeftir.

H A Y V A N F İ Z Y O L O J İ S İ
116
Sumasyon:
Sumasyon bir sinir gelen tek bir sinaps üzerinde kısa sürede çok sayıda uyarı gelmesi yada bir sinire gelen çok sayıdaki EPSP bağlı olarak sinirin eşik değeri üzerine çıkarak aksiyon oluşturmasıdır. Tek bir kası inverte eden bir sinir hücresi uyarı oluşturduğunda kas hücresi tamamem kasılır. Buna karşın bir kas demetine bağlı bir sinir demeti uyarıldığında farklı eşik değerlerine bağlı kas lifleri kasılır.
ŞEKİL 2.26 Sinirde meydana gelen sumasyonun şekli
Hızlı, tekrarlayan uyaranlarla tek bir sarsı cevabına nazaran, kas gerimi veya kısalmasında meydana gelen artış, yani cevapların birikmesidir. İki çeşit sumasyon vardır: yada tek tek kas sarsılarının birbirine katılarak uyum içinde kuvvetli kas hareketleri oluşturması aynı anda bir kasa birden fazla sinirden uyarı gelmesi ile sumasyon oluşması.
• Temporal (dalga) sumasyon: tek bir motor birimin kasılma hızının artmasıdır.
• Spasiyal (çoklu lif) sumasyon: aynı anda kasılan motor birim sayısının artması oluşturması.
Sumasyon Hızlı, tekrarlayan uyaranlarla tek bir sarsı cevabına nazaran, kas gerimi veya kısalmasında meydana gelen artış, yani cevapların birikmesidir.

H A Y V A N F İ Z Y O L O J İ S İ
117
Nörotransmiter maddeler Nöronlar arasında veya bir nöron ile başka bir (tür) hücre arasında iletişimi sağlayan kimyasallara nörotransmitter (uyarıcılara tepki) denir. Sinir sistemi boyunca sinirsel sinyaller bu kimyasal taşıyıcılar yardımıyla iletilir. Sinir hücrelerinin taşıdığı sinyaller nöronlar üzerinde son derece hızlı ilerler. Bu hız sinir hücresini türüne göre 1 m/sn ile 12 m/sn arasında değişir. Bir maddenin nörotransmitter olarak sınıflandırılması için şu özellikleri taşımalıdır.
Bu kriterlere uyan iki nörotransmitter madde: asetilkolin ve noradrenalin (norepinefrin) dir. Bunlardan başka çok sayıdaki kimyasal madde vardır ki gerekli şartlardan bir kısmına fakat tümüne değil cevap vermektedir. Bunlar: Dopamin, adrenalin, serotonin, histamin, gaba aminobütirik asit (GABA), glutamik asit, aspartik asit ve glisin bu gruptandır.
Dopamin beyinde hareket ve davranışları idare eden sinirlerde transmitter madde görevi yapar. Adrenalin beyin nöronlarında bulunmuştur, Histamin sinir uçlarında bulunmuştur. GABA ve Glisin inhibe edici nörotransmitterdir.
ŞEKİL 2.27 Nörotransmiter maddeler
Sinir hücreleri arasındaki bağlantı ve sinyal aktarımını ise sinaps denilen ve iki sinir hücresi arasında bulunan bölgelerce sağlar. Bu bağlantı bölgelerinde sinyalin geldiği nörondan salgılanan nörotransmitterler, karşıdaki nöronun hücre yüzeyinde bulunan protein reseptörlerce algılanarak sinyalin bu hücreye aktarılmasını sağlarlar. Post-sinaptik
1) Bir kimyasal maddenin sentezlenmesi için gerekli enzimleri o nöron taşımalıdır.
2) Sinir uyarıldığı zaman bu maddeyi salmalı ve madde postsinaptik hücre membranındaki özgül bir reseptörle reaksiyon vererek biyolojik bir aktiviteye neden olmalıdır.
3) Görevini yaptıktan sonra, bu maddenin etkisini çabucak ortadan kaldıracak bir mekanizma bulunmalı.

H A Y V A N F İ Z Y O L O J İ S İ
118
protein reseptörlerle iletişim yolu çoğu sinir ileticisi için geçerli olmakla beraber gaz formunda bir sinir ileticisi olan nitrik oksit post-sinaptik membranı aşarak intrasellüler cGMP düzeyi üzerinden etkisini gösterir. Günümüzde ATP de sinir ileticisi olarak kabul edilmektedir ve etkilerinin bir kısmı için pürinerjik post-sinaptik reseptöre ihtiyaç duymaz. Sinapslarda iki hücre arasındaki mesafe son derece azdır (yaklaşık olarak 20 nm). Bu durum fizyolojik sıcaklıklarda ve ortam koşullarındaki difüzyon hızı ile birlikte ele alındığında, bir hücreden salınan nörotransmitter maddenin diğerine varış zamanının neredeyse anlık olacak şekilde çok kısa olmasına neden olur. Nörotransmitter madde etkilerine göre iki grupta toplanır. Bunlar eksite edici (uyarıcı) ve inhibe edici (engelleyici) maddelerdir. Sinir istemindeki sinyallerin işlenip bunların bilgiye dönüştüğü yer olan sinapslarda bu iki farklı grup nörotransmitter madde sayesinde bazı sinyaller artırılırken bazıları azaltılmış olur. Bu özellik, sinir dokuların sinyal işleme yetisinin temel bileşenlerinden biridir. Başlıca nörotransmiter maddeler :
Transmiter maddeler nöron gövdesinde senzetlenir ve taşıma proteinleri ile taşınır.
Nörotransmiter nöron gövdesinde üretilmesine rağmen aksondan salınması gerekir bu durumda nörotransmiter özel taşıma molekülleriyle vezikül içinde salgılanacakları akson ucuna taşınır. Akson ucundan salgılanma ile boşalan vezikül tekrar nöron gövdesine getirilmesi gerekir. Bu amaçla kinezin ve Dineinin adında iki taşıma molekülü görev yapar. Kinezinin taşıması başlıca hücre gövdesinden akson terminaline doğru (anterograd) gerçekleşir. Bu işlem beslenme için gerekli moleküllerin, enzimlerin, mitokondrilerin, içlerinde nörotransmiter olan veziküllerin ve diğer organellerin taşınmasında önemlidir. Dineinin hareketi karşı yöndedir (retrograd) ve yeniden kullanılacak membran vezikülleri, büyüme faktörleri ve nöronun morfolojisini, biyokimyasını ve bağlanabilirliğini etkileyen diğer kimyasallar bu yolla taşınır. Retrograd taşıma, tetanoz toksini, herpes simpleks, rabies ve çocuk felci virüslerin gibi merkezi sinir sistemi içine yayılan membranlı ajanların da kullandığı bir yoldur.
1. Asetilkolin (ACh), 2. Biyojenik aminler: a) Katekolaminler Dopamin, Norepinefrin, Epinefrin b) Serotonin (5-hidroksitriptamin) c) 5-HT Histamin 3. Amino asitler: a) Eksitatör amino asitler; örneğin, glutamat
b) inhibitor amino asitler, örneğin, gama-aminobütirik asit (GABA)
c) glisin 4. Nöropeptitler: endojen opiyatlar, oksitosin, taşikininler 5. Çeşitli: a) Gazlar; örneğin, nitrik oksit
b) Pürinler; örneğin, adenozin ve ATP

H A Y V A N F İ Z Y O L O J İ S İ
119
ŞEKİL 2.28 Sinir hücresinde nörotransmiter madde taşınması
Vesiküller nöron membranına mikrotübüller ve mikrofilamentler aracılığı ile gider. Ca2+ iyonları mikrotübül kontraksiyonunu aktive eder. Vinblastin ve sitokalasin gibi maddeler mikrotübül fonksiyonunu önleyerek transmitter salınmasını önlemektedir. Sinaptik boşluğa verilen nörotransmiterin işlevleri yaptıktan sonra(İyon kanalı aktivasyonu yada inaktivasyonu) hızla yıkılmaları gerekir Bu amaçla MAO ve COMT enzimleri görev yapar. Katekolamin grubu nörotransmitter maddeler iki enzim tarafından yıkılırlar. Bunlar, catechol-O-methyltransferase (COMT) ve mono amine oxidase (MAO) enzimleridir. MAO seratonin, noradrenalin, dopamin, adrenalin ve diğer bir sürü bileşikten amino grubunu ayırır. Veya bir kısım noradrenalin presinaptik nöron tarafından geri alınarak etkisi ortadan kaldırılır. Sinir ucunda dopamin ve adrenalin in miktarı belli bir seviyeye ulaşınca, tirozin den bu maddelerin sentezi durdurulur. Kas hareketlerinin integrasyonunu sağlayan nöronların aksonunda dopamin eksikliği Parkinson hastalığına neden olur. Pakinson hastalarına DOPA tedavi için verilir. DOPA, dopamin ön maddesidir ve kan beyin bariyerini dopaminden daha kolay geçer ve beyin hücrelerine ulaşır. Nöradrenalin taşıyan sinirlerin aksonları cerebellum ve cortex cerebri nin kas hareketlerinin koordinasyonu, uyanık bulunma heyecan gibi fonksiyonları idare eden bölgelerine ve hipotalamusa gider. Aksonda yapılan taşıma kuduz virusun taşınmasında önemlidir. Asetil kolin reseptörlerine bağlanarak sinir hücresine giren virus retrograd aksonal taşıma yolu ile nöron gövdesine dyanin tarafından taşınır. Bu sırada çoğalan kuduz virusu merkezi sinir sistemine kadar yayılır. MSS de anterograd aksonal taşıma yolu ile periferik sinirler ile tükürük bezlerine yayılan virus tükürük yolu ile tekrar bulaşır. Yara bölgesine giren Clostridyum tetani bir proteaz olan tetanus toksini üreterek retrograd aksonal taşıma ile MSS iletir. Spinal sinaptik bölgede inhibe edici nörotransmiterlerin salgılanmasını engeleyen toksin çene kaslarında kasılmaya solunum yetmezliğine neden olur.

H A Y V A N F İ Z Y O L O J İ S İ
120
ŞEKİL 2.30 G proteinie bağlı uyarının hücrede ifadesi
Sinap iletimi sonucunda meydana gelen olaylar: 1. Postsinaptik hücre membranı boyunca özel iyon kanallarının açılması. G proteinine yanıt olarak açılan bir potasyum kanalı aktif olabilir; bu kanal, ikinci haberci sistemi kullanmadan direkt aktive olan iyon kanallarının hızlı kapanmasının aksine genellikle uzun süre açık kalır. 2. Nöron hücresinde siklik adenozin mono fosfat (cAMP) veya siklik guanozin monofosfat (cGMP)'m aktivasyonu. Nöronlarda çok özel metabolik mekanizmayı ya siklik AMP’nin veya siklik GMP’nin kontrol edebilirler. Böylece hücre yapısının uzun süreli değişimi de dahil olmak üzere pek çok kimyasal sonucun alınmasına neden olur. Bu durum nöronun uzun-süreli uyarılabilirliğini değiştirir. 3. Bir veya daha fazla sayıda intraselüler enzimin aktivasyonu. G-proteini, intraselüler enzimlerden bir veya daha fazlasını direkt olarak aktive edebilir. Enzimler de, hücredeki pek çok spesifik kimyasal fonksiyonlardan herhangi birine sebep olabilirler. 4. Gen transkripsiyonunun aktivasyonu. Bu belki de postsinaptik nöronun ikinci haberci sistemlerininen önemlisidir. Gen transkripsiyonu, nöronda yeni proteinlerin oluşmasına neden olabilir ve bu proteinler de hücrenin metabolik mekanizmalarım veya yapısını değiştirebilirler. Özellikle uzun-süreli bellek işlemlerinde, uygun olarak aktive edilen nöronlarda yapısal değişiklikler oluştuğu bilinmektedir. Asetilkolin çizgili kas (iskelet kası) liflerindeki nikotinik reseptöre bağlanarak kas hücresine Sodyum (Na+) içeri girmesini ve voltaja duyarlı kalsiyum kanallarının açılması ile depolarize olan kas hücresinin kasılmasını sağlar, Asetilkolinin öğrenme ve hafıza (M1 res) ile de derin bir ilişkisi bulunur ve bu yüzden beyinde asetilkolin içeren nöronlar mevcuttur. Parasempatik sinir sisteminde gangliyon ve sinapslarda yer alan asetilkolin, parasempatik sinir sisteminin uyarılmasıyla beraber gerçekleşen kalp atışının yavaşlaması, tükürük salgısının artması, bronkokonstrüksiyon (bronş lümeneninin -boşluğunun- daralması), miyozis, mesane ve gıs hareketlerinin artması gibi etkilerin gerçekleşmesine neden olur. Asetilkolin belli nöronlarda kolin asetil transferaz enzimi tarafından, kolin ve asetil-CoA'dan sentez edilir. Sinaptik boşluğa salınarak görevini tamamladıktan sonra asetilkolin, asetilkolinesteraz enzimi yardımıyla kolin ve asetat'a (asetik asit tuzu) çevrilerek yıkılır. Asetilkolin; kolin asetiltransferaz enzimi tarafindan kolinin asetilasyonu sonucu sentezlenir. Asetil kaynağı, mitokondrilerde sentezlenen asetilkoenzim A’dır.

H A Y V A N F İ Z Y O L O J İ S İ
121
Kolin kaynağı ise, sinaptik aralıkta yıkılan Asetilkolin’den olusan kolindir. (Kolinin presinaptik aralığa geri alınım basamağı sentezde hız kısıtlayıcı basamaktır). Sonucta asetilkoenzim A ve kolinden, kolin asetiltransferaz enzimi tarafından Ach olusturulur. Hemikolinyum koline cok benzediği icin presinaptik bolgeye alımda kolinle yarısır ve kolin geri alınımını azaltır. Bu nedenle asetilkolin sentezini inhibe eder.
Histamin bulunduran nöronlara histaminerjik nöron denir. Histamin, mide mukozasında, hipofiz bezinin ön ve arka lobunda mast hücrelerinde de bulunur. Histaminin uyanıklık, cinsel davranış, bazı ön hipofiz hormonlarının salınımının düzenlenmesi, kan basıncı, su içme ve ağrı eşiği ile ilişkisi vardır. Histamin mastositlerde bazofillerin hücre içi keseciklerinde bulunur. Orada bir protein ve heparinden oluşan komplekse bağlıdır. Histamin özellikle ani aşırı duyarlılık hallerinde serbest hale geçer. Bu geçiş, bir antijen organizmaya yeniden girdiğinde mastosit zarına bağlı bir antikorla tepkimeye girişince ortaya çıkar.
ŞEKİL 2.31Histamin
Bazı fiziksel olgular (deri irkilmeleri, yanıklar) sırasında da histamin serbestleşir; bazı kimyasal etkenler de onu serbest hale geçirebilir. Histaminin histaminerjik alıcılara yapışması yoluyla ortaya çıkar. Histamin kılcal damarların genişlemesiyle beraber yerel geçirgenliğin artmasına (karıncalanma), bronşların ve bağırsakların büzüşmesine, mide, tükürük ve böbreküstü bezi özeği salgılarının artmasına neden olur. Yani vazodilatasyon ve mide mukus salgısını arttırır. Beyindeki yeri ve işlevi hakkındaki bilgiler daha azdır; uyku-uyanıklık devrelerine, atardamar basıncının kontrolüne ve vücut sıcaklığının ayarlanmasına da katılmış olabilir. Dopamin: Noradrenalin ve adrenalin öncü maddesidir. Memeli beyninde bulunan bir nörotransmitterdir. Uyku ile ilgilidir ve kas aktivitesinde rol oynar. Dopamin (DA), vücutta doğal olarak üretilen bir kimyasaldır. Beyinde, dopamin reseptörlerini aktive ederek nörotransmiter olarak görev yapar. Dopamin, ayrıca, hipotalamustan da salgılanır ve kana karışarak nörohormon görevi yapar.
ŞEKİL 2.32 Dopamnin Nörohormon olarak görevi hipofizin ön lobundan prolaktin salgılanmasını baskılamaktır. Sempatik sinir sistemindeki etkileri dolayısıyla ilaç olarak; kalp atışlarını hızlandırmak ve kan basıncını yükseltmek için kullanılır. Kan-beyin omurilik sıvısı bariyerini geçemediği için merkezi sinir sitemini doğrudan etkileyemez. Parkinson hastalarında ve Dopa-duyarlı distoni hastalarında, beyindeki dopamin miktarını artırmak için, dopamin sentezinde öncü molekül görevi üstlenebilen L-DOPA molekülü kullanılır, zira L-DOPA kan-beyin bariyerini aşabililir.
Dopamin Hipotalamustan da salgılanır ve kana karışarak nörohormon görevi yapar. Nörohormon olarak görevi hipofizin ön lobundan prolaktin salgılanmasını baskılamaktır.

H A Y V A N F İ Z Y O L O J İ S İ
122
ŞEKİL 2.34 Trozin amino asidinden epinefrin sentezlenmesi
Glutamik asit ve aspartik asit krustaselerde bulunan transmitter maddelerdir. Glutamik asit kas kontraksiyonu yaratır, aspartik asit ise kas ve reseptör hücreler üzerinde hem uyarıcı hem de inhibe edici rol oynar. Beyin üzerinde etkili olan ilaçların hepsi nörotransmitter sistemlerin aktivitesini inhibe etme veya aktive etme yoluyla etkili olurlar. Örneğin, kafein, nikotin, ve amfetaminler kimyasal sinapslarda biyojenik aminleri oluşturarak beyin aktivitelerini uyarırlar. LSD (halusinasyon yaratan madde veya evham: liserjik asit dietilamin) ise serotonini inhibe eder ve uykusuzluğa neden olur. Glutamat göz fotoresetörlerden kaışığın varlığında salgılana inhibe edici bir nörotransmiterdir.
ŞEKİL 2. 35 Glutamatat
Kimyasal olarak monoamin yapısına sahip bileşikler olan Epinefrin (adrenalin), Norepinefrin (noradrenalin) ve Dopamin topluca katekolaminler olarak adlandırılırlar. Katekolaminler hormon ve nörotransmitter olarak fonksiyon görürler. Bu bileşikler adrenal medullada ve sempatik sinirlerde sentezlenirler.

H A Y V A N F İ Z Y O L O J İ S İ
123
ŞEKİL 2.36 Norepinefrin ve epinefrin etki mekanizması
MAO (Mono amino oksitaz)
Katekolaminleri parçalayan 2 enzim bulunmaktadır: COMT (Katekol-O-Metil Transferaz) ve MAO (Mono Amin Oksidaz):
MAO Mitokondriyal bir enzimdir. MAOlar nörotransmitterlerin inaktivasyonu önemli rol oynar. MAO sitoplâzmada vezikül dışında nöradrenalini parçalayan enzimdir. MAO, izoenzimleri bulunan mitokondrilerde yerleşik halde ve adrenerjik, dopa-minerjik ve serotonerjik sinir sonlarıyla karaciğer ve barsak çeperinde bulunan bir enzimdir. Sinir sonlarında bulunan MAO veziküllerden sızan monoaminlerin oksidatif deaminasyonla yıkımına neden olur. Bir kısım monoaminde bu yıkımdan kurtularak sinaptik aralıktan reuptake yoluyla geri dönen monoaminleri de yıkarak veziküllere girmesini engeller. Böylece MAO sinir sonlarıyla monoaminleri baskı altında tutucu bir görev üstlenir. Karaciğer ve barsak çeperindeki MAO'nun ise besinlerle alınan tirozin ve feniletilamin gibi toksik monoaminlerin dolaşıma geçmelerine engel olmak gibi bir görevi vardır.
İnsanlarda MAO iki tür vardır: MAO-A ve MAO-B . Her ikisi de nöronlar ve astroglialda bulunur.
• MAO-A da karaciğer, akciğer, vasküler endotel, gastrointestinal sistem ve plasenta bulunur.
• MAO-B çoğunlukla kanda trombositlerde bulur.
MAO sentezinin az yada çok olması bağlı olarak şizofreni depresyon, dikkat eksikliği bozukluğu , madde bağımlılığı , migren, meydana gelebilir
COMT (Katekol-O-Metil Transferaz): Katekolaminleri nöron dışında parçalayan enzimdir. COMT, damar çeperindeki (en çok akciğerlerde) endotel hücrelerinde bol olarak bulunur ve katekolaminleri metilasyona ugratarak parçalar.

H A Y V A N F İ Z Y O L O J İ S İ
124
ŞEKİL 2.37 COMT ve MAO etkisi
Serotonin
Serotonin(5-hidroksitriptamin) vasküler düz kaslarda, gastrointestinal sistemde ve beyinde tanımlanmış ve bu bölgelerde önemli işlevleri bilinen biyolojik amindir. Esansiyel bir amino asit olan L-triptofandan triptofan hidroksilaz enzimi ile sentezlenmekte, etkilerini kendisine özgü serotonerjik reseptör (5-HTR) aileleriyle göstermektedir.
ŞEKİL 2.38 Serotonin
Serotonin sentezi özellikle gastrointestinal sistemdeki enterokromaffin hücrelerinde ve beyinde serotonerjik nöron gövdelerinin yoğun olduğu “raphe nuclei”de gerçekleşir. Plateletlerde % 8 oranında depolanmakta ve plateletten salıverilen derişimlerde lokal olarak vazokonstriktör etki göstermektedir. Serotonerjik reseptörler; depresyon, besin alımı ve obezite, saldırgan davranışlar, obsesif kompulsif bozukluklarla anksiyete bozuklukları, sirkadyan ritm ve uyku, analjezi, migren, bulantı ve kusma, madde bağımlılıkları ve seksüel bozuklukluklarla ilişkilendirilir. Sinapslardaki serotonin düzeyinin artması besin alımını azaltarak obeziteyi engeller, insomnia gibi uyku bozukluklarını önler ve bağımlılıkların kesilmesi sırasında gözlenen depresyonu ortadan kaldırır. Teröpatik amaçla; serotonin transporter’ının (SERT) selektif inhibitörleri (SSRI), hücre gövdesindeki fazla serotonini yıkan mono amino oksidaz (MAO)’ın inhibitörleri, triptofan ve hidroksitriptofan supplement’leri kullanılmaktadır. Ancak bunlardan birkaçının birlikte kullanılması serotonin düzeyini çok yükseltmekte ve

H A Y V A N F İ Z Y O L O J İ S İ
125
serotonin sendromu olarak bilinen klinik tabloya sebep olmaktadır. Beyin serotonin düzeylerinin farmakolojik yöntemler dışında artırılması bireyin ruhsal durumu ve pozitif sosyal davranışlarını olumlu yönde etkileyebilmektedir. Parlak gün ışığı, uyku düzeni, egzersiz ve triptofandan zengin diyet endojen serotonin düzeyini sağlamak yönünde önemli rol oynamaktadır.
Sinir sistemi Sinir sistemin evrimsel gelişimi
Canlığın gelişimi sırasında basit hücre benzeri yapılar birbirleriyle dahi iletişim kuramıyorlardı. Sadece belirli yüzeylere tutunma veya belli kimyasalların varlığında hücre yapısının içeri bükülerek kimyasalı içeriye alması şeklinde tepkimeleri gerçekleştirebiliyorlardı. Ancak sonrasında, bu yapılar daha da özelleşti ve biraz daha karmaşıklaştı. Farklı tip haberleşme kimyasalları birbirleriyle etkileşmeye başladı, daha fazla sayıda ve tipte glukoprotein özelleşti ve tüm bunlar evrimsel süreçte hücrelerin karmaşıklaşmasıyla daha da karışık bir hal aldı ve iyice özelleşti.
Koaservatların daha da karmaşıklaşması ve evrimleşmesi sonucu oluşan daha gelişkin hücrelerin zarları üzerinde çok daha fazla sayıda reseptör proteini görmekteyiz. Bu reseptörler, yukarıda da anlattığımız şekilde, sayısız kimyasalın tanımlanmasını ve hücre içerisinde ilgili değişimlerin gerçekleştirilmesini sağlarlar. Hücrenin tanıyabildiği tüm kimyasallar için özelleşmiş reseptörler bulunmaktadır. Tanınmayan kimyasalların yok edilmesi içinse bazı başka reseptör yapıları bulunmaktadır. Sonuçta her hücre, her kimyasalı tanıyamaz. Tanıyabilmesi için ancak ve ancak evrimsel süreçte o kimyasala karşı reseptör üretmesini gerektirecek ve üretebilenlerin avantajlı olacağı bir evrimsel geçmişe sahip olmalıdır. Bu sebeple, tanınmayan yapıların risk oluşturabilmesine karşı, daha genel yapılar evrimleşmiş ve bu kimyasalları yok etme görevini üstlenmiştir.
Bazı canlılarda uyarıyı alan farklılaşmış yapılar yoktur ancak, canlı madde (protoplazma) en ilkel şekli ile her çeşit uyarıyı alır ve tepki gösterir. Sinir sistemine sahip olmayan hayvanlarda bazı koordinasyon mekanizmaları vardır. Hücre zarında meydana gelen depolarizasyon dalgası da bir ileti olarak kabul edilmektedir. Örneğin, silli epitel hücrelerinde sil hareketi hücreden hücreye iletilmektedir. Bir hücrelilerden siliyatlar'da bazal granülleri birbirine bağlayan bir fibril sistemi vardır. Ektoplazma içinde ve yüzeye paralel yer alan bu fibriller hareket ettirici sinir sistemini oluşturur. Birçok tek hücreli ve serbest yüzen larvada, vücutlarının bir bölgesinden diğer bölgesine kendiliğinden hareketli ve çok ağır işleyen bir ileti sistemi vardır. Kendilerini bir yere tespit ederek yaşayan hayvansal organizmalarda ise çok az gelişmiş, yüzeysel ve kendiliğinden hareket yeteneği olmayan bir ileti sistemi vardır. Sinir sistemi farklı gruplarda farklı şekilde özelleşmiştir. Sölenterler, sinir sistemi açısından çok önemli bir geçiş noktasında bulunmaktadır. Çünkü sabit yaşamdan hareketli yaşama geçiş, organizmayı oluşturan hücreler arasında sıkı bir iletişimi gerektirmektedir. Sabit bir yaşam sürerken buna pek gerek yoktur, zira çevre, hareketli bir canlının çevresine göre oldukça sabittir. Bu sebeple hücreler arası iletişim yavaş olabilir ve hatta aksamalar sorun çıkarmaz. Bu yüzden sabit canlılarda hücreler arası iletişim sadece kimyasal olarak ve oldukça yavaş bir şekilde gerçekleşir. Ancak canlı hareketli olacak şekilde evrimleştikçe, hücreler arası iletişim
Bu bölüm için Power
point sunumu adresi:
https://yadi.sk/d/07N617
ISE9kHx

H A Y V A N F İ Z Y O L O J İ S İ
126
konusunda da özelleşmiş hücreler gerekecektir. İşte en başında bahsettiğimiz, hücreler arası iletişim konusundaki kimyasalları kullanım açısından özelleşen hücreler, sinir sisteminin temellerini atmıştır. Hydra’larda nöronların kutuplaşmaması nedeniyle uyarılar belirli bir doğrultuda iletilmezler. Vücut üzerinde bir noktanın uyarılması her yöne doğru ışınsal bir iletim doğurur. Asıl merkezi sinir sistemi ilk yassı solucanlarda görülür. Vücudun iki yanında uzunlamasına seyreden sinir kordonları ve vücudun sağ ve solunda simetrik olan bölgeleri idare eden kısımlar arasında bağlantıların kurulmuş olması, omuriliğin ilk taslağı olduğu, hayvanın baş tarafında sinir kordonlarının ucunda sinir hücreleri topluluğunun artması, beynin ilk taslağı olduğu kanısını uyandırır. Annelidlerde ip merdiven sinir sistemi bulunur. Deri-kas kılıfının temel peristaltik ritmi ventral sinir şeritten kaynaklanır. Her segmentin hareketi bir çift ganglion tarafından yönetilir. Her segmentin hareketi bitişik ön segmentin hareketiyle başlatılır. Bu nedenle başsız bir toprak solucanı hareket hareket edebilir. Ancak bu hareket düzensiz ve süreklidir. Çünkü hareketin koordinasyonundan baş bölgesindeki serebral ganglion sorumludur. Bu sinir yapısı omurgalıların omuriliğinin ilk taslağıdır.
Daha gelişmiş canlılarda beynin de giderek özelleştiğini, alt bölgelere ayrıldığını, kısaca beyin için kısımlaşmanın (compartmentalization) başladığını görürüz. Bu da son derece mantıklıdır, çünkü beynin tamamı, her işe koşamaz. Bu, aynı anda gelen sinyallerin birbirine karışmasına ve vücuda olan iletimlerinde aksamalara sebep olacaktır. Bu sebeple beynin bölgeleri, farklı işlevler konusunda özelleşir. Aynı zamanda artık beyinden vücuda giden sinyallerin de karmaşık bir yol izlemek yerine, ana bir hattan dağılmaya başladığını görürürz. Özellikle Eklembacaklılar (Arthropoda) şubesinden itibaren başlayan bu özel kordon, önceliklesinir ipliği (nerve cord) olarak evrimleşecek ve özelleşecektir. Bu yapı, basitçe, beyinden çıkan sinirlerin merkezi bir sistem üzerinden vücuda dağılması için özelleşen sinir hücrelerinden ibarettir. Sinir ipliği notokord adı verilen ilkin, kemiksi bir yapı ile korunmaktadır. Daha sonrasında sinir ipliği daha da özelleşecek ve karmaşıklaşacak, omurga tarafından korunacak ve omurilik (spinal cord) halini alacaktır. Omurga da notokorddan evrimleşmiş bir yapıdır. Eklembacaklılardan böceklere baktığımızda, beyin ve bu ilkin merkezi dağıtım sisteminin, yani nöral tübün çok ilkel versiyonlarını görmekteyiz. Beyin artık birçok parçadan oluşuyor olsa da, sinir ipliği halen tam olarak gelişmemiştir ve vücuda dağıtım halen karmaşık şekilde yapılmaktadır. Aşağıda böceklere ait sinir sistemi ile hidraların sinir sistemlerinin karşılaştırmasını görmekteyiz:
Beyni nukleusudur ve nukleus, hücrenin diğer organelleriyle birlikte hareket eder. Fakat, çok hücreli bir organizmada hücreler hareket, beslenme, savunma ve bunun gibi pek çok bölümlere ayrılmışlardır. Hücrelerin birbirleriyle haberleşebilmesi için özel bir sisteme ihtiyaç vardır. Küçük ve yavaş organizmalarda bu haberleşme kimyasal olabilir.

H A Y V A N F İ Z Y O L O J İ S İ
127
ŞEKİL 2.39 Farklı canlı gruplarında sinir sistemi organizasyonu
Organizmanın bir kısmındaki hücreler hormon veya transmiter denilen kimyasal haberciler salgılarlar ve böylece organizmanın öteki kısmının ne yapması gerektiğini belirlerler. Fakat organizma büyüdükçe, zaman bir problem olmaya başlar. Difüzyon süresi, difüzyonda kat edilen mesafenin karesiyle doğru orantılı olduğu için, mesafe arttıkça süre de anormal şekilde uzar. Cevap süresi önemli değilse, kimyasal maddeleri taşımak için bir dolaşım sistemi de varsa, bu tür haberleşme sistemi tatmin edici olabilir. Hormonal kontrol sisteminde bu tür haberleşme sistemi kullanılır. Bu tür haberleşme sistemi, yavaş olduğu gibi aynı zaman da hassas bir kontrol sağlayamaz. Ani bir korku, kana fazla miktarda adrenalin salınmasına sebep olur. Fakat bu olay, ayrım göstermeksizin bütün vücut üzerinde etkisini gösterir. Hızlı ve sınırlı bir etki için bu kimyasal habercilerin mümkün olan en yüksek hızda ve etkinin görülmesi istenen yerde salınması gerekir. Sinir sistemi bu özelliklere sahip bir organizasyondur. Nöron denilen sinir hücreleri, aksonları ile etki etmek istedikleri organlara kadar uzanırlar ve nörotransmiter denilen kimyasal maddelerle etkinin görülmesi istenen hücreleri uyarırlar. Omurgasızların çoğunda sinir sistemi merkezileşmeye doğru gider. Sinir hücrelerinin bir araya gelmesiyle gangliyonlar oluşur. Gangliyonlardan sinir şeritleri çıkar. Baş bölgesinde genellikle iki gangliyondan oluşmuş bir beyin gangliyonu (serebral gangliyon) bulunur. Omurgasız örneği olan sülükte, baş kısmında bulunan beyin, çok sayıda nöron ve ara bağlantılar içerir. Beynin dışındaki nöron gövdeleri gangliyonlarda gruplanmıştır. Segmentli vücuda sahip olan sülüğün her segmentinde birçok davranışı kontrol eden bir gangliyon vardır.

H A Y V A N F İ Z Y O L O J İ S İ
128
ŞEKİL 2.40 Planeryada sinir ve rseptörler
Yüksek yapılı omurgasız hayvanlarda sinir sistemi yapısal benzerliğinden dolayı ip merdiven sinir sistemi olarak isimlendirilir. Mürekkep balığının (Loligo) sinir sisteminde dev aksonlar içeren Stellat sinir 1 mm den fazla çapı olan dev aksonlara sahiptir.
ŞEKİL 2.41 Dev akson

H A Y V A N F İ Z Y O L O J İ S İ
129
Bu aksonlar, aksiyon Potansiyelilerini hızla mantoya ileterek, mantodaki bütün kasların aynı anda kasılmalarına neden olurlar. Manto kasları aniden kasıldığında, mürekkep balığını ileri doğru iten su fışkırması olayı gerçekleşir. Krustaselerde nöromusküler bağlantıda inhibisyon oluşturan bazı nöronlar, presinaptik uyarıcı nöronları inhibe ederek etkili olurlar. Omurgasız kas fibrilleri birkaç motor nörondan sinaptik uyarı alabilir. Bunlar hem uyarıcı hem de durdurucu sinir sonlanmaları olabilir. Kurbağa kasının sinirlerle bağlantısında her bir nöron birkaç kas fibrilini kuşatır ama bir kas fibrili tek bir nöron tarafından uyarılır.
ŞEKİL 2.42 Farklı gruplardaki sinir sistemi
Sinir Organizasyonu
Uyarılabilen ve oluşan uyarıyı iletebilen yeteneğe sahip özelleşmiş hücrelerden oluşur. Sistemlerin gerektiği gibi ve birbirleriyle uyum içinde çalışmasının düzenlenmesinde çok önemli rol oynayan dokudur. Sinir dokusu hücreleri, (mekanik, ışık, ısı, ses, koku, basınç gibi) çeşitli enerji tiplerine duyarlıdır ve bunları, “aksiyon potansiyeli” adı verilen, sinirlerin özel uyarı biçimine dönüştürür. Sinir dokusunu oluşturan hücreler, aksiyon potansiyeli oluşturup iletim işlevine sahip olan “nöron” ve bu nöronlara destek görevi yapan “glia” hücrelerinden meydana gelir. Nöronlar, hücre gövdesi (soma) ve burdan çıkan “dendrit” ve “akson” adı verilen uzantılardan oluşur. Her nöronda sadece bir adet bulunan akson, uyarıyı başka bir sinir hücresine ya da kasa götürürken, sayıları birden fazla olabilen dendritler ise, sinir hücresine gelen bilgilerin toplandığı uzantılardır. Böylece sinirsel iletim, tek yönde olur. Nöronlar arası iletim, “sinaps” adı verilen yapılar aracılığı ile gerçekleşir. Sinapslar arası iletim, “nörotransmitter” adı verilen özel kimyasal maddeler aracılığı ile sağlanır. Uzantılarına göre nöronlar, insanda yalancı tek uzantılı (pseudounipolar), iki uzantılı (bipolar) ve çok uzantılı (multipolar) olmak üzere sınıflandırılır. Glia hücrelerinin ise astrosit, oligodendrosit ve mikroglia olarak alt tipleri bulunmaktadır. İnsan sinir sistemi, insan vücudundaki en karmaşık yapıdır ve 100 milyardan fazla sinir hücresinin bir araya gelerek oluşturduğu bir sistemdir. Sinir sistemi anatomik olarak ikiye ayrılır; beyin ve omurilikten oluşan merkezi sinir sistemi ile sinir fibrilleri ve küçük sinir hücresi gruplarından oluşan periferik sinir sistemi. Merkezi sinir sistemi 100 milyardan fazla nörondan oluşmuştur. Gelen sinyaller dendritlerdeki veya hücre gövdesindeki sinapslar aracılığı ile nörona ulaşır. Bu hücreler, diğerleri ile elektriksel veya kimyasal sinyaller kullanarak haberleşirler. Nöronlar, iç organların işlevlerini koordine

H A Y V A N F İ Z Y O L O J İ S İ
130
ederek homeostazı sürdürdükleri gibi duygulanıma aracılık ederek, hareketleri kontrol ederek ve insan zihnindeki karmaşayı kodlayarak da homeostazı sürdürürler.
ŞEKİL 2.43 Farklı gruplardaki sinir sistemi
Sinir sistemi (1) beyin ve omurilik (medulla spinalisi) içeren merkezi sinir sistemi (MSS) ve (2) Vücudun kas, bez ve duyu organlarını beyin ve omuriliğe bağlayan sinirlerden ibaret olan periferik sinir sistemi’nden (PSS) oluşur.

H A Y V A N F İ Z Y O L O J İ S İ
131
Periferik sinir sisteminde Schwann hücreleri denen hücreler aksonun uzunluğu boyunca düzenli aralıklarla ayrı ayrı miyelin kılıfları oluştururlar. Komşu miyelin bölümleri arasındaki boşluklara Ranvier boğumu denir ve buralarda aksonun plazma membranı hücre dışı ortama maruz kalır. Ranvier boğumu, miyelinli nöronların aksonlarında miyelin maddesini oluşturan Schwann hücreleri arasında kalan boşluktur. Nöronun akson ucundan gelen uyarı (impuls) bu boğumlardan elektriksel sıçramalarla ilerlerler. Miyelin kılıfı akson boyunca elektrik sinyalin iletimini hızlandırır ve enerjinin korunmasını sağlar. Merkezi sinir sistemindeki nöron topluluklarına nukleus, Periferik sinir sistemindeki nöron topluluklarına ganglion, Merkezi sinir sistemindeki akson topluluklarına traktus, Periferik sinir sistemindeki akson topluluklarına sinir adı verilir.
ŞEKİL 2.44 MSS ve PSS akson ve sinir hücrelerinin isimlendirilmesi.

H A Y V A N F İ Z Y O L O J İ S İ
132
ŞEKİL 2.45 Omurilikte çıkan sinirler
Omurilik sinirleri
Reseptörlerden gelen bütün afferent sinirlerin uçları gri cevherin columna dorsalisine (posteror koluna ) girer. Bu sinirlerin hücre gövdeleri medulla spinalis dışında dorsal kök ganglionu içindedir. Anterior ve lateral kolondaki motor nöronların aksonları ventral kök (anterior kök) içinde omuriliğini terk eder. Medulla spinalis’in dorsal kökü içindeki sinirler afferent, ventral kökü içindekiler ise efferent’tir. Bu Bell-Magendie kanunu olarak bilinir. Medulla spinalis segmental bir organdır ve her segmentine gelen sinirler (afferent sinirler) topluca bir kök halinde posterior kolona girerler. Gene her segmentten çıkan aksonları (efferent sinirler) topluca bir kök halinde anterior kolunu terk ederler. Her segmentin biri sağda biri solda olmak üzere iki posterior ve iki anterior kökü vardır. Medulla spinalisin ön köklerinden motor, arka köklerinden duyu sinirleri çıkar. Servikal, brakial, lumbar ve sakral pleksusları oluşturur. Vücudun en uzun siniri Siyatik sinir sakral pleksusdan çıkar.

H A Y V A N F İ Z Y O L O J İ S İ
133
ŞEKİL 2.46 Omurilikte çıkan sinirler
Beyinden çıkan sinirler
Cranial sinirlerin merkezleri beyin köküdür. Buradan kafa Sinirleri (Kranial Sinirler) çıkar : Kranial Sinirler 12 adettir. Baş ve boyunun motor-duyu işlevleri kontrol eder. Beyin sapı kaynaklıdır. Kranial sinirlerin ilk ikisi dışında, nukleusları (merkezleri) beyin kökünde bulunurlar. Kranial sinirlerin numaraları vardır. Ör. 1.kranial sinir deyince N.olfactorius, 10. kranial sinir deyince N.vagus anlaşılır. İlk ikisi N.olfactorius (koku alma siniri) ve N.opticus (görme siniri) bu sinirlerin hücreleri Gövdeleri beyin kökünde değil, MSS dışındaki ganglionlarda olmayıp burunda koku soğanı içinde ve gözde retina içindedirler. N. Trochlearis ve N. Abducence gözün dış kaslarına gider her iki gözün aynı anda sağa, sola aşağı, yukarı hareketi bu sinirler ile koordine edilir. Hepsi motorik sinirlerdir. N.oculomotorius göz yuvarlağı dış kaslarına gider. N.trigeminus; yüz ve ağız boşluğunun ağrı, ısı, temas ve basınç duyuları bu sinir yoluyla merkezlere iletilir ve pons cerebri deki merkezlere bağlanırk. Üç kökten meydana gelmiştir; sensörük kök, motorik kök, mesencephalik köktür. N.facialis; motorik ve sensorik kısımları vardır. Örneğin, motorik olanlar, yüzün mimik hareketlerini yaptıran kaslara giderler. Sekroterik olanlar; yüzün ter bezi, gözyaşı bezi, burun mukozası bezine giderler. Sensorik olanlar; dilin lezzet (temas, basınç, ısı, ağrı) tomurcuklarından gelirler. N.vagus vejetatif fonksiyonların çok iyi bir ayarlayıcısıdır. Motorik kısmı: Bronş, özafagus, mide, barsak kaslarının hareketlerini arttırır. Kalp hareketlerini frenler. Sekretorik olanlar: Mide bezlerine ve pankreasa gider. N.vagus un uyarılması pankreastan zimojen granüllerinin salınmasına sebep olur.

H A Y V A N F İ Z Y O L O J İ S İ
134
ŞEKİL 2.47 Omurilik sinır bağlantıları ve kranial sinirler
Beyin Kısımları
Evrimi ilerlemiş bir omurgalı ve memelilerin kafatası içi sinir sistemi yapısı beyin (cerebrum) ve beyincikten (cerebellum) meydana gelir.
ŞEKİL 2.48 Beyin kısımları

H A Y V A N F İ Z Y O L O J İ S İ
135
ŞEKİL 2.49 Beyinin kesidi
Beyin kökü (Pons bulbus-arka beyin):
• Medulla oblangata. Medulla spinalisin hemen üzerindeki 3 cm.lik kısım. Piramidal yol lifleri burada çaprazlaşır. Solunum ve dolaşımı düzenleyen hayati merkezler bulunur. Uyku uyanıklık ritmini ayarlayan retiküler formasyon lifleri buradan köken alıp hipotalamusa gider.
• Pons cerebri, Pons ve serebellum (beyincik) oluşturur. Serebellum motor fonksiyonu düzenler, kasların koordine hareketini sağlar, kas tonüsü, gücü hareketlerdeki koordinasyon ve hareket hızından ve dengeden sorumludur. Bilinçsiz, otomatik hareketleri düzenler.
ŞEKİL 2.50 Beyinin kesidi
Mesancepholon (orta beyin) Çeşitli refleks ve koordinasyon merkezleri bu bölümdedir. Ekstrapiramidal sistemde rol oynayan çekirdeklerin bazıları burada bulunur

H A Y V A N F İ Z Y O L O J İ S İ
136
Diencephalon (Ara beyin)
• Thalamus Bilincin kapısıdır. Tüm afferent liflerden gelen uyarılar buraya uğrar, süzülür ve korteksin ilgili bölümlerine gönderilir. Derin ve yüzeyel duyular, koku duyusu hariç başa ait duyular, beyin ve beyinciğin motor uyarıları ekstrapiramidal lifler, talamusa uğrar.
• Hypothalamus Üçüncü ventrikülün önündedir. Otonomik fonksiyonları düzenler. Otonom sinir sisteminin üst düzey merkezidir. Ön hipofiz bezi aracılığı ile endokrin sistemi de kontrol altında tutar. Ekstrapiramidal ve motor fonksiyonlarla da bağlantılıdır.
Cortex cerebri (ön beyin)
Beynin iki hemisferi, korteksi oluşturan gri madde ekstrapiramidal sisteme ait bazal ganglionların bazıları limbik sistem burada bulunur.
Retiküler Aktive edici sistem (Retiküler Formasyon)
Beyin kökünde bulunan bir bölgedir. Beyin kökünde retikülar formasyon denilen bir bölge bulunmaktadır. Burada nöronların aksonları birbirleriyle ağ oluşturacak şekilde birleşmiştir. Retikülar formasyon bölgesine görme, koklama, işitme, lezzet alma reseptörlerinden, ağrı ve ısı duyuları sistemlerinden, kas ve tendonlardan, iç organlardaki duyu sinirlerinden afferent impulslar gelir. Ayrıca cerebellum, bazal ganglionlar, thalamus ve cortex cerebriden de impulslar alır. Retiküler formasyon bölgesinden çıkan (efferent) impulslar beyin korteksinin (cortex cerebri) hemen her bölgesine ve omuriliğe gider. korteks merkezlerine ne tür duyu impulslarının gittiğinden retiküler formasyonun haberi olur. Bunun üzerine retiküler formasyon hemen beyin korteks merkezlerine impulslar göndererek o bölgeleri uyarır. Uyarılmış korteks merkezleri gelen duyuları değerlendirir. Retiküler formasyon tarafından uyarılmamış korteks aldığı duyuları değerlendiremez. Retiküler formasyon aldığı her impulsa aynı şekilde reaksiyon göstermez. Önemli olan ve olmayanı ayırt eder. Örneğin, anne bebeğin hafif bir ağlaması ile uyanırken baba etkilenmez. Sessizlikte uyumaya alışmış bir kimse uyuyamaz. Retiküler formasyonu tahrib olmuş bir şahıs sinir sistemini aktive edici sistemden yoksun kaldığı için sürekli komaya girer (bitkisel hayat). Retiküler formasyonun aktiviteleri üzerine bazı hormonlar, kimyasal maddeler ve ilaçların etkisi vardır(adrenalin, asetilkolin, CO2, anestezikler gibi). Anestezikler ve uyku verici ilaçlar duyu impulslarının retiküler formasyon içinde akışını bloke ederler.
Vücudun birçok bölgesinden impulslar alan ve hemen hemen bütün merkezi sinir sistemine impulslar gönderen retikülar formasyon bölgesi bilinçaltı (idrak edilemeyen) birçok koordine hareketlerin yapılmasında, bütün sinir sisteminin ve vücudun uyanık tutulmasında rol oynar. Retiküler formasyonun mesensephalon ve pons bölgeleri retiküler aktive edici sistemlerdir. Bu bölgenin uyarılması beyin korteksinin uyarılmasına neden olur. Hayvan uyuyorsa uyanır. Beynin bu bölgesi tümörler, kanamalar gibi nedenlerle fonksiyon yapamaz hale gelirse, şahıs komaya girer, normal uyandırıcı etkilerle uyanmaz.

H A Y V A N F İ Z Y O L O J İ S İ
137
Bazal Ganglion
Bazal ganglion, beynin orta kısmında bulunan alın korteksi ve alt motor ve duyu bölgeleri arasındaki iletişim ve yönetimi sağlayan yapıların genel adıdır. Karmaşık motor hareketlerin uygulanması ve yönetilmesi bu yapılar aracılığıyla yürütülür. Aynı zamanda hareketlerin hızlarının kontrol edilmesi gibi yüksek motor denetim özelliğine de sahip bir bölgedir. Bazal ganglion korteks, talamus ve diğer bazı yapılarla sıkı bir bağlantı gösterir. Güncel teoriler bazal gangliona eylem seçiciliği görevini yükler: olası seçeneklerden uygun olanı bulmak. Bazal ganglionun birçok motor sistem üzerinde inhibitör etkiye sahip olduğu gösterilmiştir. Bu etkinin ortadan kalkması sistemin aktif hale geçmesiyle sonuçlanır. Bazal ganglionun davranış belirleme fonksiyonu beynin birçok bölgesinden gelen iletilerle biçimlenir. Bu merkezlerden biri de yönetim özellikleri belirgin olan prefrontal kortekstir. Parkinson hastalığında basal ganglionlarda dejenerasyon vardır. Parkinson hastalığındaki titreklik heyecan halinde artar. Heyecan halinde adrenal medulladan salınan adrenalin, retiküler formasyonu uyarır ve titrekliği arttırır.
Parkinson hastalığında titreklik uykuda neden kaybolur? Çünkü retiküler formasyon uykuda aktif değildir. Kas hareketlerinin yapılmasında, aralarında bağlantılar bulunan üç beyin bölgesinin –motor korteks, serebellum ve basal ganglionların rolü vardır. Elektrofizyolojik araştırma sonuçları kas hareketleri başlamadan önce bu bölgelerin aktive gösterdiğini kanıtlamıştır.
Herhangi bir kas hareketi başlamadan önce beyin korteksinin somato sensöri, görme, işitme bölgelerinden impulslar hem basal ganglionlara, hem de serebelluma ulaşır. Serebellum ve basal ganglionlar bu enformasyonu değerlendirerek ve gerektiği biçimde değiştirerek talamus yoluyla motor komplekse impulslar gönderirler. Bundan sonra motor korteks kaslara emirler gönderir. Basal ganglionlar ve serebellum fonksiyon bakımından birbirlerinin tamamlayıcılarıdırlar. Serebellum hızlı, basal ganglion yavaş hareketleri kontrol eder. Parkinson hastalığında basal ganglionlarda zedelenme vardır. Titreklik, kaslar istirahat halindeyken daha belirgindir. Serebellum zedelenmesinde ise kas titremeleri istekle yapılan kas hareketleri sırasında ortaya çıkar.
Limbik sistem
Limbik sistem beyin kökünün bir kısmını ve ara beynin etrafını bir halka gibi çevreler. Limbus halka anlamına gelmektedir. Limbik sistemin koku alma ve yeme, içme ile ilgili davranışlarda rolü vardır. Hipotalamus la birlikte, heyecan, korku, hiddet reaksiyonları seksüel davranışları meydana getirir. Amigdala zedelenirse, hayvan yenebilir ve yenemez şeyleri ayırd edemez.
ŞEKİL 2.52 Limbik sistem

H A Y V A N F İ Z Y O L O J İ S İ
138
Anksiyete, saldırganlık vb. duygusal yanıtlardan sorumludur Limbik sistemin bazı bölgeleri, hayvanı memnun edici ve etmeyici (ödüllendirici ve cezalandırıcı) duygularla ve bu duyguların (ödüllendirme ve cezalandırma) duygularla ve bu duyguların gerektirdiği davranışlarla ilgili merkezler taşır. Ör: Hipotalamus ‘un ventromedikal nukleuslarının uyarılması, hayvanı hayatından çok memnun (ödüllendirilmiş duruma) soktuğu görülür. Limbik sistemin diğer bazı bölgelerinin uyarılması ile hayvanın incitilmiş gibi bir duygu içine girdiği ve bu duygunun gerektirdiği davranışları gösterdiği tespit edilmiştir.
Hipotalamus
Heyecan, korku ve hiddetin meydana gelmesi: Duyular ya hoşa gider ya gitmez. Bu durum doyum ve nefret olarak da adlandırılır. Belirli limbik alanların elektriksel yolla uyarılması hayvanın hoşuna giderken veya doyum sağlarken diğer bölgelerin uyarılması terör, ağrı, korku, savunma ve kaçma reaksiyonlarını doğurur. Hipotalamus insan ya da hayvan vücudunda organ sistemlerinin fonksiyonlarında düzenleyici rol oynar ve homeostasis’i sağlar.
ŞEKİL 2.53 Pons ve hipotalamus
1-Somato motor ve Visero motor aktivitelerin koordinasyonu: Somato motor ve Visero motor aktivitelerin koordinasyonu: Hipotalamus, somatik ve viseral motor aktivitelerin birbirine uygun olarak yapılmasını sağlar. Örneğin, aktif bir somatik organa (kas) otonom sinirlerin kontrolü ile fazla kan gönderilir. Kanın aktif dokuların ihtiyaçlarını karşılaması için içindeki oksijen miktarını arttırması gerekir. Demek ki kas aktivitesinin (somato-motor) normal devamı için solunumun (visero-motor) ona göre ayarlanması gerekir.
2-Kardiyovasküler düzenleme: Kardiyovasküler düzenlenme: Hipotalamus boyunca çeşitli alanların uyarılması kardiyovasküler sistemde arteriel basıncın artıp azalması, kalp hızının azalıp çoğalması gibi nörojenik etkilere neden olur
3-Vücut temperatürünün düzenlenmesi: Vücut temperatürünün düzenlenmesi: Hipotalamusun ön bölgesi vücut ısısının düzenlenmesiyle ilgilidir. Bu alandan geçen kanın ısısındaki artış termosensitif nöronların aktivitesini arttırırken, vücut ısısının düşmesi de bunların aktivitelerini azaltır. Vazodilasyon, titreme, terleme, hızlı nefes alma gibi hareketler vücut ısısını sabit tutmaya yönelik hareketlerdir. Bu hareketler hipotalamus tarafından kontrol edilir.
Pons: Orta beyinin altında, medulla oblongata’nın anterior ucunda transvers sinir lifleri. Köprüye benzer. Medulla oblongata ile beyin hemisferlerini birleştirir. Solunum kontroluna katkıda bulunur. Apnöstikve pnömotaksik merkez buradadır. Yüzeysel lifler cerebellum’a bağlanır. Motor ve sensorik (duysal) derin lifler medulla oblongata’dan pons üzerinden orta beyine gider. Medulla oblongata: Yaklaşık 3 cm, pons ve omurilik arasında traktlar omurilik ile beyin arasında komünikasyonu sağlar. Vital merkezler: kan damarlarının otonomik inervasyonu için vazomotor merkez; kalbin otonomik sinir kontrolunu sağlayan kardiyak kontrol merkezi; pons ile birlikte solunumu kontrol eden solunum merkezi. Omurilik ile beyin arası haberleşme alanıdır. Kalp hızı, solunum ve diğer öksürük, yutma, kusma gibi refleksleri kontrol eder.

H A Y V A N F İ Z Y O L O J İ S İ
139
4-Vücut suyunun (su dengesinin) düzenlenmesi: Vücut suyunun düzenlenmesi: Hipotalamus vücut suyunu 2 şekilde düzenler. 1) Susama duygusu yaratarak canlının su içmesini sağlar. 2)İdrarla su atılımını kontrol eder. Susama merkezi denilen bölgedeki nöronlarda bulunan elektrolitler yoğun duruma geçtiğinde hayvanlarda su içme ihtiyacı doğar ve su içerek elektrolit yoğunluğu normale döner. Vücut sıvıları çok yoğunlaştığında, böbreklerden su atılımının kontrolünün yapıldığı alandaki nöronlar uyarılır ve bu nöronlardan çıkan sinir lifleri hipofize gelir ve antidiüretik hormon salgılanır. Bu hormon böbreklerde suyun geri emilimini sağlar böylece idrarla su kaybı azalır.
5-Üreme ve seksüel davranış: Üreme ve seksüel davranış: Hipotalamus erkek ve dişi bireyde seksüel aktiviteyi kontrol eder. Bazı hayvanlarda ovulasyonu kontrol eder. Hipotalamustaki paraventriküler çekirdek (hipotalamusun ön kısmı) uyarıldığında oksitoksin salgılaması artar, özellikle gebeliğin sonunda oksitoksin salgısının artması doğum sancılarının oluşmasına ve bebeğin doğmasının kolaylaşmasına yardımcı olur. Oksitoksin sütün memeden boşalmasında da yardımcı olur.
6-Gastrointestinal sistem ve beslenmenin düzenlenmesi: Beslenmenin düzenlenmesi: Hipotalamusta a) açlık (besin alma), b) tokluk merkezi diye 2 merkez vardır. Hipotalamus bu merkezlerin aktivitelerini kontrol eder. Besin alma merkezi uyarıldığında hayvan lüzumundan fazla yemek yer, bu merkez tahrip edildiğinde ise hiç iştah duymaz ve açlık sonucu ölüm meydana gelir. Tokluk merkezi uyarıldığında ise hayvan yemek yiyemez, bu merkez tahrip edildiğinde ise aşırı derecede yemek yer. Hipotalamustaki diğer bir merkez de gastrointestinal aktivitelerin tümünü kontrol eden, dudak yalama ve yutma gibi refleksleri kontrol eden merkezdir.
7-Ön hipofiz bezi aracılığı ile endokrin hormon salgılanmasının kontrolü: Endokrin sistemin kontrolü: Hipotalamusun bazı alanlarının uyarılması ön hipofiz bezinden hormon salgılanmasına neden olur. Hipofiz bezi vücudun hemen hemen bütün endokrin bezlerinin aktivitelerini düzenler
8-Uyku ve uyanıklığın ayarlanması: Uyku ve uyanıklığın ayarlanması: Uyku kişinin duysal veya diğer uyaranlarla uyarabileceği bir bilinçsizlik durumudur. Uykuyu, kişinin uyandırılamadığı bilinçsizlik durumu olan komadan farklı tutmak gerekir. Hipotalamus, uyku ve uyanıklığın düzenlenmesinde retiküler formasyon merkeziyle beraber çalışır.
9-Heyecan, korku ve hiddetin meydana gelmesindeki rolü.:Heyecan, korku ve hiddetin meydana gelmesi: Duyular ya hoşa gider ya gitmez. Bu durum doyum ve nefret olarak da adlandırılır. Belirli limbik alanların elektriksel yolla uyarılması hayvanın hoşuna giderken veya doyum sağlarken diğer bölgelerin uyarılması terör, ağrı, korku, savunma ve kaçma reaksiyonlarını doğurur.
Talamus
Diensephalonun diğer parçası olan thalamus, periferden gelen tüm impulsları alır, değerlendirir ve ihtiyaca göre beyin korteksinin çeşitli merkezlerini uyarır. Talamus fizyolojik aktivitelerin organizatörüdür. Talamus taki nöronlardan kök alan aksonlar cortex cerebri’nin ilgili duyu alanına ulaşır. Bu nedenledir ki “thalamus cortex cerebri’ye giden yolların kapısıdır”. Aralarında kompleks bağlantılar kurmuş olan kortex cerebri dış dünyayı idrak etmemizi ve dış ortamdaki değişikliklere karşı gerekli reaksiyonları gösterebilmemizi sağlayan esas yapılardır.

H A Y V A N F İ Z Y O L O J İ S İ
140
ŞEKİL 2.55 Talamus
Sinir sistemi organizasyon
Sinir sisteminin motorik fonksiyonları kontrol eden iki bölümü vardır: Vücutta kalp, kan damarı, düz kaslar ve bezlerin aktivitelerini kontrol eden otonom sinir sistemine(vegetative) sinir sistemi de denir. Otonom sinirler isteğimize uymayan (irademiz dışı) visseral aktiviteleri kontrol ederler ve efferent sinirlerdir.
1-Somotik motor sistem
2-Otonomik motor sistem
A-Sempatik Sistem: Sinirler omuriliğin ilk torasik segmentinden 3. lumbar segmentine kadar olan sinir köklerinden çıkar.
B-Parasempatik Sistem: Bu sistemin sinirlerinden bir kısmı bazı kranial sinirlerle birlikte beyinden çıkar; bir kısmı da omuriliğin 2. 3. ve 4. sakral segmentlerinden çıkar, ventral kökler içinde omuriliği terk eder. Başka bir deyimle cranial ve sacral bölümünden çıkar.
Thalamus Diencephalon’un en büyük alt bölümüdür. Üçüncü ventrikülün sağ ve solunda yer almıştır. Vücudun dışında neler olduğunu beynin bilmesini sağlar. Koku hariç, cerebruma giden bütün sensorik (duyusal- göz ve kulak) bilgilerin geçiş (gidiş ve geliş) bölgesidir. Uyanıklık halini ve uyarı olduğunda uykudan uyanışı düzenler.

H A Y V A N F İ Z Y O L O J İ S İ
141
Sempatik Sinir Sistemi
Sempatik sistem doku ve organlara gönderdiği sinyallerle genel olarak vücudun aktivitesini, enerji tüketimini artırıcı yönde hareket eder. Örneğin sempatik sinirler kalbin çalışma hızını ve atardamarlardaki kan basıncını artırır. Sempatik sistem aynı zamanda organizmanın korku, öfke, dehşet, heyecan ve şiddetli ağrı gibi stres yaratan durumlarda tepki oluşturmasını sağlar. Örneğin kas aktivitesi gerektiren stres durumlarında, hipotalamusun uyarılmasıyla sempatik sistem organizmada bazı değişikliklere yol açar. Bunlardan bazıları kandaki glikoz yoğunluğunun artışı, atardamarlarda kan basıncının yükselmesi ve kas gücünün artmasıdır. Bu değişiklikler organizmanın stresle baş etmesinde etkili olur. Sempatik sinir sistemi nöronları, medulla spinalisin torakal ve lumbal bölgesindeki ön ve arka boynuzunda bulunur. Sempatik nöronlardan çıkan sinir lifleri, spinal sinirin ön kökleri içerisinde medulla spinalisi terk ederek spinal sinirin yapısına katılırlar. Bu lifler, vertebraların her iki tarafında bulunan truncus sympathicusa gelirler. Burada bulunan ganglionlara gelen sinir liflerinin bazıları buradaki nöronlarla sinaps yaparlar. Bazıları ise doğrudan organa giderler. Spinal iliği ventral kök içinde terk ederek sempatik ganglionlara gelirler. Sempatik sinirlerin preganglionik aksonları sempatik ganglionlarda sona erer ve buradan çıkan postganglionik aksonlar organlara giderler. Diğer bazıları ise simpatik ganglionda sinaps yapmadan geçerler ve kolateral ganglionda sinaptik birleşme kurarlar; buradan postganglionik aksonlar çıkar organlara gider. Normal koşullarda, kalp atım sayısı, kan basıncı, bronşların genişlemesi gibi fizyolojik aktiviteye sahiptir. Dinlenme durumunda hayvanlarda sempatik sinirlerin hepsi aktif durumda değildir. Zorlanım veya tehlike durumunda sinirlerin çoğu veya tamamına yakını etkin hale geçer. İsteğimize uymayan hemen her organ (kalp kası, düz kas, bez hücreleri) hem simpatik hem de parasimpatik sinirler alırlar. Görünüşte bu iki tür sinirlerin etkileri birbirinin aksidir. Hakikatte bu sistemler birbirinin tamamlayıcısı ve mütekabilidirler. Organizmanın aktivite icabına göre birisinin etkisi çoğalırken, öbürünün etkisi azalır
• SAVAŞ ya da KAÇ tepkisi oluşur • Gözbebekleri genişler/büyür • Kalp hızlı atar, kalp kası güçlü pompalar ve kan basıncı artar • Dolaşımdaki kan önemli organlara çekilir, iskelet kaslarına ve kalp
kaslarına daha fazla kan verilir • Deri terler • Soluk yolları genişler ve solunum hızlanır • Kandaki şeker seviyesi artar • Sindirim sistemindeki ve idrar yollarındaki sfinkterler kapanır. • Gelen uyarı uzun süre devam ederse sistem yükü kaldırmakta zorlanır ve
sinaptik bağlantılarda ve Adrenal Bezlerde NE salgısı azalabilir/durabilir • Sempatomimetik etki: SSS nin tepkisini taklit eden etkidir. • Sempatolitik etki: SSS tepkisini kesen (bloke eden) etkidir. • Katekolaminler: SSS ne benzer etkiler oluşturan içten ve dıştan gelen
(endojen ve eksojen) maddelerdir.

H A Y V A N F İ Z Y O L O J İ S İ
142
ŞEKİL 2.56 Omurgadan çıkan sinirler

H A Y V A N F İ Z Y O L O J İ S İ
143
ŞEKİL 2.57 Parasempatik ve sempatik sinirlerin postganlion nörotransmiterleri
Parasempatik Sistem
İstirahat şartlarında hakim olan sistemdir. Ana siniri Nervus Vagus’tur. Medulla spinalisin kranial ve sakral bölgelerinden çıkar. Preganglionik lifleri uzun, postganglionik lifleri ise kısadır. Parasempatik sistemin hem preganglionik hem de postganglionik liflerinden Asetilkolin salgılanır. Parasempatik sistem kolinerjik reseptörler veya muskarinik reseptörler aracılığı ile etki gösterir. Örneğin kalbde parasempatik sistem kalb atım hızını azaltır. Parasempatik sistem kalp atım hızının azalması yanı sıra, atriyumların kasılma gücünde hafif bir azalmaya neden olabilir. Deri kan damarlarında genişleme ve koroner damarlarda daralma dışında sistemik damarlara etkisi yoktur. Otonomik sinir sisteminin bu kısmı kranial ve sakral sinirlerden kurulmuştur. Kranial bölümün sinirleri orta beyinden ve M.oblangata ‘dan çıkar. 3.orta beyinden 7. 9. ve 10 sinirler m.oblangata’dan çıkar. Sakral bölümün sinirleri 2.3.4. sakral sinirler içinde seyreder. Lateral zincir ganglionlarını sinapsis yapmadan geçer. Parasempatik sinirlerin morfolojik özelliği, bu sinirlerin preganglionik aksonları gidecekleri organın içinde veya çok yakınında bulunan ganglionlarda sonlanır. Bu ganglionlardan çıkan ve effektör organlara giden postganglionik aksonlar kısadır. Bu özelliği göstermeyen parasempatik sinirler de vardır. Ör:kan damarlarına giden parasempatikler. Akson uçlarından asetil kolin salan sinirler (kolinerjik sinirler) parasempatik sinirlerdir. Bu sinirlerin uyarılması birçok düz kasların membranını depolarize eder ve impuls frekansını arttırır, kas tonusu artar ve ritmik kontraksiyonların sayısı artar. Parasempatik sinirler barsakların düz kaslarını uyarır, sfinkter kaslarını gevşetir. Sempatik sinirler uyarılınca akson uçlarından noradrenalin salınır, bu nedenle bunlara adrenerjik sinirler denir. Sempatik sinirler sfinkterlerin sirküler kaslarını uyarır, kasılmaya sevk eder. Parasempatik aktivitenin yüzde 80 kısmı N. vagus yapar. N.vagus kesilecek olursa, veya atropine ile paralyze edilirse, kalp vuruşlarının dakikadaki sayısı bariz derecede artar. Vagus’un frenleyici etkisi M.oblangata’daki vagus nukleusuna sinus caroticus’tan arcus aorticus’tan ve sol kalpten gelen impulslara bağlıdır. Bu reseptörlerden impulslar azalırsa n. vagus ‘un etkisi de azalır; impuls artarsa n vagus’un etkisi de artar, yani kalp atımını frenler. Sürekli egzersiz yapanlarda kalbin atım hızının yavaş olması, kan basıncının egzersiz süresince artmasından dolayı n.vagus’un daimi suretle uyarılmasına ve bu etkinin adet haline gelmesine bağlıdır.
MSS kullanılan
nörotransmiterler Semaptik sinir sistemindeki pre ganlion tarafında Ach post ganglion tarafında ise NE kullanılırken Parasempatik ise nörotransmiter madde olarak Ach kulanılır.

H A Y V A N F İ Z Y O L O J İ S İ
144
Parasempatik postganglioner liflerinin yaptığı nöroefektör sinapsta muskarinik kolinerjik reseptörler vardır. Muskarinik reseptörler membranı depolarize eder. Reseptörler kalsiyum kanalını açan ya da G proteinini aktive eden reseptörler olabilir. Gp proteini fosfolipaz aracılığı ile fosfatidil inositol difosfattan inositol trifosfat (IP3) ve diasil gliserol (DAG) oluşmasına neden olur. DAG protein kinaz C’yi aktive eder. IP3 ise endoplazmik retikulumdan Ca++ salınmasına neden olur. Gi proteini kalpte K+ kanallarını açarak hiperpolarizasyon yapar bu da sinus nodu ve atriyoventriküler nodda inhibisyon yapar. Parasempatik stimülasyonun bazı etkileri şunlardır. Göz, pupilla ve silier kaslarda konstriksiyon Bezler nazolakrimal, parotis, submandibular ve gastrik sekresyonlarda artış, Kalp; kasılma gücünde azalma, kalp atım sayısında azalma, A-V iletide yavaşlama Akciğer; bronşlarda konstriksiyon, kan damarlarında dilatasyon ve sekresyonlarda artış Barsaklar; lümende peristaltizm ve tonus artışı, sfinkterde gevşeme safra yolları; kasılma gözlenir.
Parasempatik sinir sisteminin etkisi: � DİNLEN ve SİNDİR tepkisi oluşur. � Vücudun kendine gelmesini, dinlenme anında enerji dengesinin düzeltilmesini
sağlar � Sempatik uyarıların eski haline dönmesini sağlar � Kalbin yavaşlamasını, soluk yolunun ve gözbebeklerinin eski haline(çaplarına)
dönmesini sağlar � Tükürük ve barsak salgıları ile barsak harelketlerini artırır � Eğer kişinin korkusundan kaçmak ya da korkusunu yenmek için çıkış kapısı
yoksa: parasempatik etkiler artar; idrar ve dışkı üzerindeki kontrolü kaybolur � Parasempatomimetik etki: PSS tepkisini taklit eden etkidir � Parasempatolitik (Antikolinerjik) etki: PSS tepkisini kesen (bloke eden) etkidir. Kolinerjik Ve Adrenerjik Etkiler
Kolinerjik: somatik sinir sisteminin parasempatik postgangliyonundaki sinirlerden Ach olarak salgılanan uyarıları ileten hücrelerdir (nörotransmitter). Asetilkoline duyarlı alıcılara(reseptörlere) Kolinerjik Reseptör denir ve bu aynı zamanda PSS etkisini belirtir.
Adrenerjik: otonom sinir sisteminin genellikle sempatik postgangliyonlarından E (epinefrin/adrenalin) ya da NE (norepinefrin/noradrenalin) olarak salgılanan uyarıları ileten hücrelerdir. Çoğu sempatik postgangliyalardan NE, Adrenal medulladan ise hem E hem de NE salgılanmaktadır. Norepinefrine duyarlı alıcılara Adrenerjik Reseptörler denir ve bu aynı zamanda SSS etkisini belirtir
Adrenerjik alıcılar: α1 F düzkaslardadır, kasılmaya neden olur, α2 F arteriyollerdedir, vazokonstriksiyona neden olur, kan basıncını yükseltir β1 F koroner damarlardadır, vazodilatasyona neden olur; +inotropik, + dromotropik, +kronotropik etki yapar β2 F akciğerlerdedir, bronkodilatasyona neden olur, oksijenlenmeyi artırır

H A Y V A N F İ Z Y O L O J İ S İ
145
Asetilkolin(acetylcholine, Ach) iki alıcıyı etkiler: 1. Nikotinik Alıcılar (Nicotinic receptors): İskelet kaslarını uyarırlar. Uyarı olduğunda
pregangliyonik sinirlerden salgılanan Ach, nikotinik alıcılara gider 2. Muskarinik Alıcılar (Muscarinic receptors): Çizgisiz kasları, kalp kasını ve salgı
bezlerini uyarırlar. Postgangliyonik sinirlerden salgılanan Ach, muskarinik alıcılara gider. Üç tip muskarinik alıcı vardır: M1: sinir sisteminde bulunurlar; M2: kalptedir, sempatik uyarı sonrası kalbin eski haline dönmesini sağlar.
Kulakçık(atriyal) kaslara etki ederek kasılmayı azaltır, karıncık(ventrikül) kasına etkisi yoktur. Ayrıca SA (sinoatriyal) düğüm ile AV (atriyoventriküler) düğümü etkileyerek hızı azaltır;
M3: Vücudun birçok yerinde bulunur ve düz kasları etkiler (kan damarları, akciğerler, sindirim sistemi gibi) Vazokonstriksiyon (damarlarındaralması), bronkokonstriksiyon (solukyollarının daralması) ve barsak hareketlerinin yavaşlamasından sorumludur. Çeşitli salgı bezlerinde de bulunan M3 tükürük bezlerinde ve diğerlerinde salgının artmasını sağlar.
Cerebellum (Küçük Beyin) Fonksiyonlari Küçük beyni çıkartılmış hayvanlarda yürüyüş ve vücudun vaziyet almasında koordinasyon ve denge bozukluğu gözlenmiştir. Doktorlar sarhoş yürüyüşünü küçük beyin fonksiyon bozukluğu semptomu olarak kabul ettiler. Cerebellum büyük beyin motor nöronlarından tıpkı kaslara giden impulslar gibi impulslar almaktadır. Ayrıca küçük beyin kontraksiyon halindeki kaslardan impulslar alır. Bu şekilde küçük beyin, büyük beyin merkezlerinden verilen emre karşı kasların gösterdiği reaksiyonun kontrol edilmesi ve emrin icabına göre değiştirilmesi ile, kasların hareketlerini koordine ediyor. Motor korteksten kök alan impuls (1)kasa ulaşır(1a). Aynı zamanda motor korteksin kasa ne maksatla emir verdiği nucleus pontis (1b) yoluyla cerebellum’a bildirilir. Kas kasılınca, kas gerilme ve tendon golgi aparatından proprioceptive impulslar (2) pedinculus anterioris yoluyla neocerebellum’a ulaşırlar. Böylece cerebellum, cortex’in ne yaptırmak istediğinden ve kasın ne derece maksada uygun hareket ettiğinden haberdar olur. Eğer kasın hareketinde hata varsa bu cerebellum cortexlerinden çıkan (afferent) impulsların thalamus yoluyla(3) cerebral cortex e ve reticüler formasyon yoluyla (4) kaslara ulaşması ile hata derhal düzeltilir. Cerebellum iskelet kaslarının istek dışı hareketlerini ve kas koordinasyonunu da buna benzer biçimde fakat daha başka merkezler aracılığı ile Pürkinje hücreleri küçük beynin esas ünitesidir ve afferent impulsların sağlar. efferent impulslar haline çevrilmesi bu hücrelerde olmaktadır. Her bir pürkinje hücresi impulsunu uzun bir akson ile küçük beyinden dışarı diğer motor merkezlere gönderir. Küçük beyin temas, işitme, görme duyularını da koordine eder. Temas, görme ve işitme merkezlerinin küçük beynin ilgili merkezleri ile bağlantı kurduğu tespit edilmiştir. Ör: gözler kapalı yapılan yoga hareketi veya elektrikler ansızın kapandığında evin neresinde ne olduğunu bildiğimiz halde yavaş hareket ediyor olmamız. Çünkü görme merkezlerinden küçük beyne gelen impulslar yoluyla kas hareketlerinin daha iyi koordine edilmesi Yani küçük beyin ve bağlantıları, sinir sisteminin sensorik ve motorik yolları arasına yerleşmiş bir kontrol sistemidir. Küçük beyinden talamus’a, basal ganglionlara ve beyin köküne impuls götüren yollar tespit edilmiştir. Küçük beyin temas, işitme, görme duyularını da koordine eder. Temas, görme ve işitme merkezlerinin küçük beynin ilgili merkezleri ile bağlantı kurduğu tespit edilmiştir. Yani küçük beyin ve bağlantıları, sinir sisteminin sensorik ve motorik yolları arasına yerleşmiş bir kontrol sistemidir.

H A Y V A N F İ Z Y O L O J İ S İ
146
Büyük Beyin (Beyin korteksi) (Cortex cerebri)
Duygu, irade, hafıza, düşünce, zekâ, muhakeme ve yaratıcılık gibi ruhsal fonksiyonlarını Cortex cerebri yapar. Ayrıca iskelet kaslarının motor aktivitelerini idare eden motor merkezler ve duyuların idrak edilmesini sağlayan duyu merkezleri beyin korteksinde bulunurlar. Beyin korteksi h.sel yapı bakımından özellikler gösteren birçok alanlara ayrılmıştır. Bilinç zeka algılama vb. üst düzey görevler korteksin vazifesidir.
Beyin korteksinde çeşitli fonksiyonların yapılmasını sağlayan merkezler az çok belirli alanlarda yer alır. Fakat unutmamak gerekir ki korteksin her alanı öteki alanlarla beraber, assosiyasyon halinde, tam fonksiyon yapabilir.
ŞEKİL 2.58 Beyindeki duyusal alanlar
Hemen hemen bütün istemli hareketlerin düzenlenmesi beyin korteksi tarafından yapılmaktadır. Bu herbir kasın her kasılmasının korteks tarafından istendiği anlamına gelmez. Aksine, korteks cerebri alt beyin alanlarıyla ortak çalışır ve alt bölgeden gelen isteklere göre cevaplar verir. Korteks serebride görme, işitme, konuşma, bellek, yorum, el becerileriyle ilgili merkezler bulunur. Ayrıca, yutma, çiğneme olayları da korteks serebri tarafından başlatılır sonra refleks olarak devam eder. Korteks serebri 2 hemisfer (yarı küre) den meydana gelir. Her hemisferde (parietal, frontal, temporal, oksipital) 4 lop bulunur. Parietal loba vücudun somatik alanlarından duyular ulaşır. Bu duyuların geldiği yerlere somatik duysal alanlar denir. Buralardan motor alanlara bilgiler gider, motor alanlarda bu bilgilere göre cevap hazırlanır ve ilgili somatik bölgelerdeki kaslara gönderilir. Beyin sapından korteks serebriye giden sinir sinyalleri yok olursa, beyin işe yaramaz. Beyin sapı, mesensefalon ve serebrum arasında meydana gelen bir tümör nedeniyle baskıya uğrarsa hasta hayatının geri kalan kısmını koma halinde geçirir.
• Kortekste motor, duysal ve bağlantı bölgeleri bulunur. • Görme, işitme, tat ve koku merkezleri de burada bulunur

H A Y V A N F İ Z Y O L O J İ S İ
147
Efektör Yapılar
Endokrin ve sinir sistemin etkisi ile tepki veren yapılara effektör yapılar kısaca reseptörlerde alına bilgi efektörlerde tepkiye dönüştürülür. Effektörler sinir sisteminden ayrı özelleşmiş yapılardır. Çok hücreli hayvanlarda effektör organ ya bir kas veya bez hücreleridir.
ŞEKİL 2.59 Efektör yapılar
Tek Hücrelilerde hareket
Hücredeki sitoplazmanın ektoplazması (dış kısmı) jel (katı), endoplazma (iç kısım) ise sol (sıvı) haldedir. Ameboid hareket sitoplazmanın jel ve sol hallerine dönüşümü sonucunda meydan gelir. Bu hareketin moleküler düzeydeki temeli ise kas kontraksiyonuna benzer hafif bir kasılmaya dayanmaktadır. Endoplazmada mikzomiyozin denilen bir protein bulunur. Bu protein, Ca2+ ve ATP etkisiyle kasılır ve gevşer. Bu kasılıp gevşeme hareketlerinin sitoplazmanın sol ve jel durumuyla ilgili olduğu sanılmaktadır. Bu nedenle ameboid hareketin temelinde kas hareketlerinin yattığından söz edilir. Ameboid harekette hücre bir uyaranla karşılaştığında cevap olarak hücrenin bir ucunda ektoplazma dışarıya doğru uzayarak pseudopod oluşmaya başlar ve pseudopod durmadan uzar.
ŞEKİL 2.60 Efektör yapılar
Bu sırada anterior uçta ise mikzomiyozin kasılmış, endoplazma sol halinden jel haline dönüşmüştür. Posterior uçta ise mikzomiyozin gevşemiş, ektoplazma jel halinden sol haline geçmiştir yani bir türlü erimiştir. Bu durumda ektoplazma endoplazmaya dönüşür ve endoplazma pseudopod içine akar. Endoplazmanın kontraksiyonla jel hale, ektoplazmanın ise sol hale geçmesi eriyen bölümün pseudopod içine akmasını sağlayan bir kuvvet oluşturmaktadır. Pseudopod ileriye doğru uzarken anterior uçta endoplazma kasılarak jel haline

H A Y V A N F İ Z Y O L O J İ S İ
148
geçer ve endoplazma ekzoplazmanın özelliğini kazanır, eksoplazma halinde geri döner. Bu olay durmadan devam eder, sonunda ameboid (amipsi) hücre çevresini değiştirmiş olur. Başka bir teoriye göre; Psödopodlar aktin alt birimlerinin biraraya gelerek mikrofilamentleri kurması, mikrofilamentlerin ise geri dönüşümlü olarak ağsı yapıyı oluşturmasıyla genişler ve kasılır. Bu birlikteliklerin kurulup, bozulması sonucunda sitoplazma jel ve sol durumları arasında değişime uğrar. Yaygın olarak kabul edilen bir modele göre, hücrenin sürüklenen arka kısmında yer alan filamentler miyozin ile etkileşerek, kasılmaya neden olurlar. Bu kasılma bir diş macunu tüpünün sıkılmasına benzer şekilde, içteki sıvıyı aktin ağının gevşetilmiş olduğu psödopod içine doğru iter. Aktin tekrar ağ yapısı oluşturmak üzere biraraya gelene kadar, psödopod genişler. Sadece amipler değil hayvan vücudunda birçok hücre de sürünerek hareket eder.
Ameboid hareketin en gelişmiş şekli amiplerde görülür. Bu olay sonunda hücre tümüyle yerini değiştirir. Ameboid hareket besin maddelerine veya tehlikeye karşı yapılan bir harekettir. Bu hareketi başlatan faktörün hücre zarında potansiyel değişikliğe neden olan bazı kimyasal (kimotaktik) maddeler olduğu sanılmaktadır. Bu olaya kimotaksis olayı denmektedir. Bu hareketin çoğu kez kimyasal maddelerin az olduğu yerden çok olduğu yere doğru meydana geldiği görülmektedir. Buna pozitif kimotaksis, tersi olan olaya ise negatif kimotaksis hareket denir. Ameboid hücreler besin maddelerine karşı (+), tehlikeye karşı (-) kimotaksis gösterirler. Çok hücreli hayvanların birçoğunun vücutlarının iç veya dış yüzeylerinde bulunan siller de bu hayvanların ortam içinde yer değiştirmesinde rol oynarlar (Ctenophora, turbellaria’da larva hareketi). Çok hücrelilerin bir kısmında ise sillerin görevi farklıdır. Bunlarda siller kendilerine temas eden katı ve sıvı cisimlerin hareketini sağlarlar. Örneğin, yumurta hücresi yumurta kanalının iç yüzünü döşeyen sillerin vuruşlarıyla kanalda ilerler, midyelerin solungaç epitelinde bulunan siller solunum için gerekli su akımını meydana getirdiği gibi su içinde bulunan besinlerin yakalanmasını da sağlarlar
Çok hücrelilerde hareket
Çok hücreli hayvanların birçoğunun vücutlarının iç veya dış yüzeylerinde bulunan siller de bu hayvanların ortam içinde yer değiştirmesinde rol oynarlar (Ctenophora, turbellaria’da larva hareketi). Çok hücrelilerin bir kısmında ise sillerin görevi farklıdır. Bunlarda siller kendilerine temas eden katı ve sıvı cisimlerin hareketini sağlarlar. Örneğin, yumurta hücresi yumurta kanalının iç yüzünü döşeyen sillerin vuruşlarıyla kanalda ilerler, midyelerin solungaç epitelinde bulunan siller solunum için gerekli su akımını meydana getirdiği gibi su içinde bulunan besinlerin yakalanmasını da sağlarlar. Amipte besin alınması, memeli vücudundaki alyuvarların fagositoz olayı gibi görevlerle de yükümlüdürler. Fetusun gelişmesi sırasındaki embriyonik hücreler ve fibroblastların hareketleri de ameboid harekettir. Ameboid hareket tek hücrelilerden Rhizopodada, çok hücreli hayvanların gezici hücrelerinde örneğin, süngerlerin mezenşim tabakasında, sölentaratların eşey hücrelerinde (Hydra’nın yumurtaları) görülür. Silyum dıştan sil zarı denilen ve plazma zarının devamı olan bir zar ile çevrilir. İçinde sil matriksi bulunur.
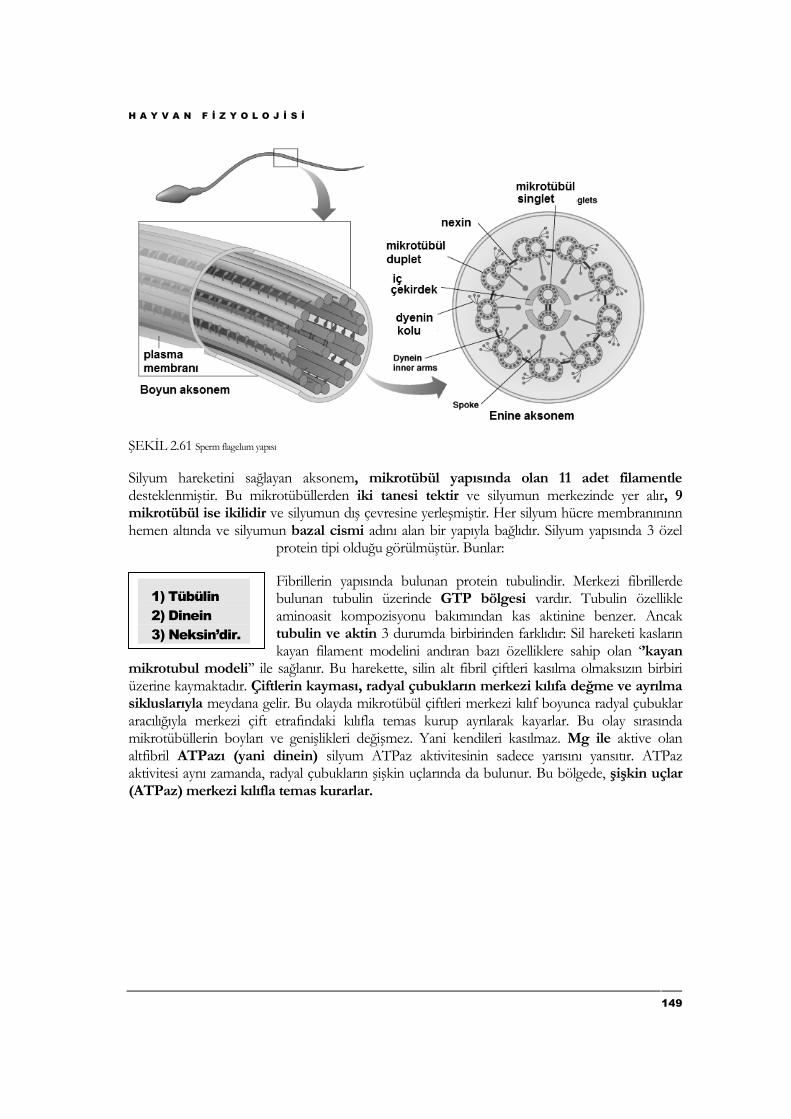
H A Y V A N F İ Z Y O L O J İ S İ
149
ŞEKİL 2.61 Sperm flagelum yapısı
Silyum hareketini sağlayan aksonem, mikrotübül yapısında olan 11 adet filamentle desteklenmiştir. Bu mikrotübüllerden iki tanesi tektir ve silyumun merkezinde yer alır, 9 mikrotübül ise ikilidir ve silyumun dış çevresine yerleşmiştir. Her silyum hücre membranınınn hemen altında ve silyumun bazal cismi adını alan bir yapıyla bağlıdır. Silyum yapısında 3 özel
protein tipi olduğu görülmüştür. Bunlar:
Fibrillerin yapısında bulunan protein tubulindir. Merkezi fibrillerde bulunan tubulin üzerinde GTP bölgesi vardır. Tubulin özellikle aminoasit kompozisyonu bakımından kas aktinine benzer. Ancak tubulin ve aktin 3 durumda birbirinden farklıdır: Sil hareketi kasların kayan filament modelini andıran bazı özelliklere sahip olan ‘’kayan
mikrotubul modeli’’ ile sağlanır. Bu harekette, silin alt fibril çiftleri kasılma olmaksızın birbiri üzerine kaymaktadır. Çiftlerin kayması, radyal çubukların merkezi kılıfa değme ve ayrılma sikluslarıyla meydana gelir. Bu olayda mikrotübül çiftleri merkezi kılıf boyunca radyal çubuklar aracılığıyla merkezi çift etrafındaki kılıfla temas kurup ayrılarak kayarlar. Bu olay sırasında mikrotübüllerin boyları ve genişlikleri değişmez. Yani kendileri kasılmaz. Mg ile aktive olan altfibril ATPazı (yani dinein) silyum ATPaz aktivitesinin sadece yarısını yansıtır. ATPaz aktivitesi aynı zamanda, radyal çubukların şişkin uçlarında da bulunur. Bu bölgede, şişkin uçlar (ATPaz) merkezi kılıfla temas kurarlar.
1) Tübülin
2) Dinein
3) Neksin’dir.

H A Y V A N F İ Z Y O L O J İ S İ
150
Kas Kas hücreleri, zar yüzeyleri boyunca aksiyon potansiyeli iletebilme
ve bu elektriksel değişikliği takiben mekanik olarak kasılma veya boylarını kısaltma yanıtı oluştururlar. Kasın bazı temel özellikleri vardır; Kontraktilite -kasılabilme, Eksitabilite-uyarılabilme, Estensibilite-
uzayabilme-gerilebilme, Elastisite-normal boyuna dönebilme. Kas hücrelerinin hayvanı hareket ettirebilmesi için mekaniksel bir dayanağa ihtiyaç vardır. Bu iskelet sistemidir. En ilkel iskelet sistemi hidrostatik iskeletir. Bu tür iskelet, annelidlerde özellikle toprak solucanlarında iyi gelişmiştir. Bu hayvanlarda, vücudun dış duvarı ve iç organlar arasındaki boşlukta (sölom) vücut sıvısı bulunur. Bu sıvı iskelet görevi yapmaktadır. Derinin altında birbirleriyle karşıt çalışan 2 kas tabakası bulunur. Bunlardan biri halka şeklinde dizilmiştir, diğeri ise boyuna olarak yer alır. Deri ve kasların oluşturduğu bu yapı “deri-kas’’ kılıfı adını alır. Halka kasların kasılması sölom sıvısında bir basınç meydana getirir. Halka kaslar kasıldığında, boyuna kaslar bu sıvı basıncıyla uzayıp gerilir ve hayvanın boyu uzar. Bundan sonra halka kasların gevşemesi ve boyuna kasların kasılması hayvanın boyunun kısalmasına neden olur. Kas tonusu: Kas kasılması ile oluşan gerimdir. Kaslar dinlenim halinde kısmen kasılı durumdadırlar, bu kası kasılmaya hazır tutmak içindir.
ŞEKİL 2.62 Karşılıklı kasılan kas grupları
Kas tipileri
Genel olarak iskelet kası, organizmayı dış ortamın değişikliklerine uydurmakla; düz kas organizmanın iç ortamındaki değişikliklerine reaksiyon göstermekle, kalp kası ise kalbin çalışmasını sağlamakla görevlidirler. Genel olarak omurgasızların düz kası çizgili kasa benzer fakat miyofilamentleri düzgün bir konum göstermez. Omurgasız hayvanlardan toprak solucanları, yumuşakçalar ve sölenterlerde kaslar çoğunlukla çizgili ve çizgisiz olma özelliklerine birlikte sahiptirler. Yine bazı balıkların bağırsaklarında düz ve çizgili kaslar birlikte bulunur.
• Kasılırlar • Hareket üretirler
• Güç üretirler
• Isı üretirler
• Destek görevi görürler
Bu bölüm için Power
point sunumu adresi:
https://yadi.sk/i/My7hAj
CMccCcT

H A Y V A N F İ Z Y O L O J İ S İ
151
İsekelet, Kalp Ve Düz Kas Arasındaki Farklar
İskelet kası Kalpkası Düz kas Çizgili Çizgili Çizgisiz Sinsisyum yok Sinsisyum yapmış Sinsisyum yapmış Küçük T boruları Büyük T boruları Genellikle güdük T boruları Sarkolazmik retikulum iyi gelişmiş ve Ca pompası hızlı etkili
Sarkolazmik retikulum var ve Ca pompası göreceli olarak hızlı etkili
Sarkolazmik retikulum gelişmemiş ve Ca pompası yavaş etkili
Plazmalemma bir çok hormon reseptöründen yoksun
Plazmalemmada çeşitli reseptörler var (α ve β adrenerjik)
Plazmalemmada çeşitli reseptörler var (α ve β adrenerjik)
Sinir uyarıları kasılmayı başlatır Kasılma intrensek bir ritme sahiptir Kasılma sinir uyarıları, hormonlar v.b. ile başlar. Hücre dışı sıvı Ca kasılmada önemsizdir.
Hücre dışı sıvı Ca kasılmada önemlidir
Hücre dışı sıvı Ca kasılmada önemlidir
Troponin sistemi var Troponin sistemi var Troponin sistemi yok Kaldesmon işe karışmaz Kaldesmon işe karışmaz Kaldesmon önemli bir düzenleyici proteindir. Çapraz köprüler çok hızlı döngülenir Çapraz köprüler çok hızlı
döngülenir Çapraz köprüler yavaş döngülenir ve uzamış kasılmaya ve daha az ATP tüketilmesine neden olur.
ŞEKİL 2.63 Kas tipleri
Kas Temel olarak düz ve çizgili kas varken istemli kontrol edilen çizgili kaslar iskelet kası istem dışı çizgili kaslar ise kalp kası olarak sınıflandırılır.
• İskelete kası: Çizgili kastır. İstemli çalışır. Somatik sinirlerle idare edilir. Yalnız sinir yoluyla uyarılır.
• Düz kas: Çizgili değildir. İstemsiz çalışır. Otonom sinirlerle idare edilir.
• Kalp kası: Çizgili kastır. İstemsiz çalışır. Otonom sinirlerle idare edilir. Otomatiktir (sinirleri olmasada kasılır.)

H A Y V A N F İ Z Y O L O J İ S İ
152
İskelet kasının hücresel organizasyonu ve yapısı myofibril-myoflament adı verine yapılardan oluşur. Kas lifleri myofibril adı verilen daha küçük lifler içerir, Myofibrillerde myoflamenta adı verilen ince ve kalın uzantılardan oluşur. Her kas lifi birkaç yüz ile birkaç bin arasında miyofibril içerir. Myofibriller iskelet kasının kasılma mekanizmasında görev alan fonksiyonel birimlerdir. Uzunlamasına incelendiklerinde, sarkomer adı verilen çok sayıda bölmelere ayrıldıkları görülür. Sarkomer kas hücresinde kasılma işini yapan en küçük birimdir. Miyofibril uzun proteinden oluşur aktin , miyosin , ve titin ve onları bir arada tutan diğer proteinler. sarkomerde myofibril uzunluğu boyunca ince filaman ve kalın filamentler tekrar halinde düzenlenir. İskelet kası başlıca hareket ve ısı üretimi ile ilgili olmasına rağmen, kas kütlesinin sürdürülmesi açlık sırasında glukoneogenez için protein rezervi sağlamada gereklidir. Kas aynı zamanda yemekten sonra glukoz ve trgliseridin kullanımının ana yeridir. GLUT4 ve lipoprotein lipaz aktivitesi sayesinde kas kandaki fazla enerji kaynaklarını uzaklaştırır.
ŞEKİL 2.64 Kas organizasyonu
İskelet kası, lif adı verilen, boyu 1 mm ile 30 cm, eni ise 10-100 mikron arasında değişen binlerce kas hücresinin bir araya gelmesi ile oluşmuştur. Kaslar, çoğu durumda yumuşak dokular ile çevrilmiştir(perimisyum, endomysium, epimysium). Kas hücreleri meydana getiren flamant yapısındaki, aktin ve miyozin bir birbirlerine kayarak daralır ve sarkomer uzunluğu ve hücrenin şekli değiştirir. Kaslar kuvvet ve hareketi üretmek için çalışır. Kas dokuları mezoderm embriyonik tabakasının köken alır. Kas oluşumuna miyogenesis denir.

H A Y V A N F İ Z Y O L O J İ S İ
153
ŞEKİL 2.64 Farklı kas tiplerinin kasılma süreleri.
Kasın işlevsel kasılma birimi olan sarkomerin kısalması aktin ve miyozin proteinlerine bağlıdır. Miyozinin baş kısmı ATP’yi hidrolize eden ATPaz aktivitesine sahiptir ve kalsiyum varlığında sarkomer kısalmasıyla sonuçlanır. Dinlenme durumunda ATP depoları kas kasılması sırasında büyük bir dalgalanma göstermez. Aktif olarak kasılan kas, kreatin fosfat mekiğiyle ADP’den ATP hızlı senteziyle anaerobik ve aerobik metabolizmanın ek ATP üretimine ihtiyaç duyar
Kas Yapısını Oluşturan Proteinler
Kas liflerinde endomisyumun hemen altında sarkolemma adı verilen kas hücre zarı bulunur Bu zar sarkoplazma adı verilen hücre plazmasını çevreler. Her lif sarkoplazma içerisinde asılı halde duran yüzlerce Myofibril’den (lifcik) oluşmuştur. Myofibriller, protein yapısındaki ince ve kalın myofilamentlerden oluşmuşlardır. Bunlardan ince olan ağırlıklı olarak Aktin olmak üzere Troponin ve tropmyozin moleküllerinden, kalın olan ise Myozin moleküllerinden oluşmuştur. Bu nedenle ince ve kalın flamentler sırasıyla aktin ve myozin flamentleri olarak da tanımlanırlar. Myozin flamentleri orta bölgeleri dışında, çapraz köprüler içerirler. Çapraz köprülerin başlarında myozin ATP’az enzimi yer alır. Kas hücresinde en küçük kasılabilen protein birimine sarkomer (iki Z çizgisi arasındadır) adı verilir. Koyu bant: A bandı (anizotropik. ışığı geçirmiyor) Açık bant: I bandı (izotropik). Kasılma sırasında boyu kısalanlar: H – I – Z bantlarıdır. A bandının ortasında açık boyanan H diski bulunur. İnce filaman: Aktin, troponin T-C-I ve tropomiyozinden oluşur. Kalın filaman miyozin moleküllerinden oluşur.

H A Y V A N F İ Z Y O L O J İ S İ
154
Kas Dokusunda Bulunan Diğer Proteinler
Titin Vücuttaki en büyük proteindir. Z çizgisinden M çizgisine uzanır. Kas gevşemesine katılır.
Desmin plazma ile miyofibriller arası bağlantı sağlar. Kalsinörin Sitoplazmada yer alır. kalmodulince düzenlenen protein fosfatazdır.
Kalp hipertrofisi ve yavaş ve hızlı çalışan kasları düzenlenmesinde önemlidir.
Nebulin Z çizgisinde yer alır ve aktin flamentlerinin uzunluğunu ve bir araya toplanmasını düzenler.
Distrofin Plazmalemmaya bağlanır ve eksikliğine duschenne tipi kas distrofisi oluşur.
αααα-aktinin Aktini Z çizgisine bağlar ve aktin flamentlerini kararlı hale getirir.
Aktin: Globüler aktin monomerleri (G-aktin) polimerleşerek F-aktin filamentlerini oluştururlar. Aktin filamenti de üç protein bileşeninden oluşmuş bir komplekstir: aktin, tropomiyozin ve troponin. Aktin molekülleri küçük yuvarlak moleküllerdir (G-aktin) ve yan yana dizilerek tek sıralı bir zincir kurarlar(F-aktin). İki düz aktin zinciri helezon biçimi birbirine sarılarak ince filamenti oluştururlar. Her aktin molekülünün ön yüzü, yanındaki molekülün arka yüzüne eklenmiştir. İnce filamentlerde aktinden başka tropomiyozin molekülleri vardır. İnce ve kalın filamentlerin kasılma esnasında sürgü gibi birbiri üzerinde kaymaları, miyozin başlarının aktinlerle yaptığı köprüler sağlamaktadır. Miyozin başları filamentin yarısında sağ tarafa doğru öbür yarısı ise sol tarafa doğru dönük bulunurlar. Bu nedenle bir sarkomerde ince filamentlerin bir yarısı sağa doğru, öteki yarısı sola doğru kayarak “Z’’ hatlarını birbirine yaklaştırır ve sarkomer boyunu kısaltır. Filamentlerin boylarında değişiklik olmaz.
�
• Myozin kalın filament(%56–50)
• Aktin, ince filament(%20–25)
• Tropomyozin (10–15)
• Troponin (%4–6) 3 kısımdan oluşur
� Troponin I
� Troponin T
� Troponin C

H A Y V A N F İ Z Y O L O J İ S İ
155
Tropomiyozin: Tropomiyozin aktin üzerindeki miyozinin bağlanacağı F-aktin noktalarını kapatarak kasın gevşek kalmasını sağlayan moleküldür. Tropomiyozinler uzun ince moleküllerdir; troponinler ise az çok globüler yapı gösterirler. İplik gibi olan tropomiyozin molekülü, yedi aktin molekülü boyunca uzanır. Kastaki bir ince filamentte 300–400 kadar aktin molekülü ve 40–60 kadar tropomiyozin molekülü vardır.
ŞEKİL 2.65 Troponin ve tropomyoszin yapısı Troponin : Üç alt birimden oluşur. Troponin I, istirahat halindeki kasta aktin ile sıkıca bağlıdır ve aktin miyozin birleşmesini önler. Aktine bağlandığı bölümdür. Troponin T, En iri alt üniteye TnT adı verilmiştir. Tropomiyozinle bağlıdır. Tropomiyozin miyozin başlarının aktine bağlanma yerlerini örter. Troponin-tropomiyozin kompleksi aktin ile miyozin arasındaki ilişkiyi önlediği için bunlara gevşetici protein denir. Tropomiyozine bağlandığı bölümdür. Troponin C, Kalsiyumun bağlandığı bölümdür (Dönme özeliği vardır. tropomiyozin de dönünce F-aktin noktaları açılır). Ca2+ bağlayarak kasılmaya yol açar. Kası kalsiyum iyonlarına bu derece hassas kılan, ince filamentlerdeki troponin ve tropomiyozindir. Serbest kalsiyum iyonları troponin C molekülüne bağlanır ve bu bağlanış tropomiyozin molekülünün pozisyonunu değiştirir. Kalsiyum iyonları bulunmadığı, daha doğrusu belirli bir miktardan az bulunduğu zaman, tropomiyozin molekülleri aktin üzerindeki miyozin başlarının bağlanacağı bölgeyi kapatmış durumdadır. Ca2+ iyonları serbest kalınca troponine bağlanır, tropomiyozin molekülünün pozisyonu değişir ve miyozin’in aktine bağlanacağı bölge serbest kalır ve miyozin başları aktin’e bağlanır Miyozin : Yapı bakımından çift başlı golf sopasına benzer. Baş ve kuyruk kısmı vardır. Gevşek kasta baş-kuyruk açısı 90 derecedir. Baş kısmı kasılma sırasında aktin üzerindeki F-aktin (miyozin başının ATPaz aktivitesini arttırır) noktalarına tutunur. Miyozin başının ATPaz aktivitesi vardır. Miyozin molekülünün kuyrukları demet halinde toplanarak filamentin gövdesini oluşturmaktadır; birçok baş ise gövdeden dışarı doğru sarkmıştır.

H A Y V A N F İ Z Y O L O J İ S İ
156
ŞEKİL 2.66 Miyozin yapısı
Ayrıca, her miyozin molekülünün sarmal kısmı başla beraber yana doğru uzanır ve şekilde görüldüğü gibi, başı vücuttan uzatan bir kol oluşturur. Dışarı doğru uzanan kollar ve başlara birlikte çapraz-köprü denir. Her çapraz köprünün menteşe denen, biri başın miyozin fılamentinden ayrıldığı, diğeri iki başın kolla birleştiği yer olmak üzere iki noktada bükülebilir olduğu düşünülmektedir. 90-50-45 derece şeklinde kasılır. Myozin büyük bir proteindir (480 kDa). Bir çift büyük ağır zincir (200 kDa) ve iki çift hafif zincirli (20 kDa) altı farklı polipeptitten oluşmaktadır. Ağır zincirler alfa kıvrımlı yapıda birbirlerine dolanarak halat benzeri uzun bir segment oluşturur, her bir zincirin N terminali büyük bir küresel baş oluşturur. Baş bölgesi aktin bağlayan molekül bölümüdür. Miyozin filamentleri miyozin moleküllerinin uç uca bağlanmaları ile oluşur ve kalın filamentler de bipolar bir yapı ile sonuçlanır. Sarkomer Yapısı
Sarkomer aktin ve miyozinden oluşur. Aktin ve miyozinin konumlanması sarkomerin bantlarını oluşturur. A bandı miyozin boyuna denk gelir. H bandı iki aktin arasındadır. M çizgisi H bandının ortasından geçer. I bandı iki miyozin arasındadır. Z çizgisi I bandının ortasındadır. Aktin ve miyozin flamentlerinin birlikte yer aldığı kısımlar daha koyu renk görülürler. Bu bölgeler A Bandı olarak isimlendirilirler. A bandının ortasında aktinin ulaşamadığı ve yalnızca miyozinden oluşan bir alan vardır. H Bölgesi olarak adlandırılan bu bölge I bandından daha koyu, A bandından ise daha açık renkte görülecektir. I bandı ortasında dikey olarak uzanan Z Çizgisi bulunur. Böylece, dinlenim durumundaki, iki Z çizgisi arasında H bölgesini saymazsak sırasıyla I-A-I bantları yer almış olur. Z çizgileri bir bir myofibrilden diğerine doğru uzanarak kas lifinin içindeki myofibrilleri birbirine bağlarlar. İki Z çizgisi arasında kalan bu bölgeye Sarkomer adı verilir. Sarkomer kas kasılmasında kısalma ve uzamanın gerçekleştiği bölümdür.

H A Y V A N F İ Z Y O L O J İ S İ
157
Şekil 2.67 Sarkomer Yapisi
Kas hücre iskeleti proteini olan Titin sarkomerdeki kalın filamentin bir hizzada tutulması işlemi için destek oluşmasında rol oynadığı düşünülmektedir. Kalın filamentte bulunan diğer ilave proteinler de (ör: miyomezin ve C-protein) kalın filamentin bipolar oluşumunda ve/veya biraraya gelişinde rol oynayabilir. Titin miyozini Z çizgisine bağlar. Diğer bir kas proteini olan Nebulin globuler aktin monomerlerini birbirine bağlayan proteindir. Alfa Aktinin Fibriler aktini Z çizgisine bağlar. Titin molekül ağırlığı en büyük olandır. Kalın filamanları (miyozin) her iki taraftan Z diskine tutturan elastik proteinlerdir. Z çizgisini iskelet kası hücre membranına bağlar. Başkar bir kas proteini olan Distrofin proteini aktini kas membranına bağlar ve hücre içi stabiliteyi oluşturur. Motor Ünite
Motor ünite bir motor nöron ve bu nöronun innerve ettiği kas liflerinden oluşur. Kas hücresinin zarına sarkolemma denir. Sarkolemmanın hücre içine doğru yaptığı transvers invaginasyonlara T (transvers) tübülü denmektedir. Görevleri aksiyon potansiyelinin hücre içine hızla yayılımını sağlamaktır. İskelet kasında T tübülünün her iki yanında sarkoplazmik retikulum sisternaları (sarnıç. Kalisyum depolanır. Kalsiyumla sıkıştıran protein kalsekestrindir) bulunur. Bu üçlü yapıya triad (iki sisterna + bir T tübülü) adı verilir. Miyofibriller de aktin-miyozin miyofilamanlarından oluşur.
ŞEKİL 268 Nöromusküler bağlantı

H A Y V A N F İ Z Y O L O J İ S İ
158
Bu dalların ucunda asetilkolin içeren vesiküller bulunur. Sinir uçları, kavşaktaki kalınlaşmış kas zarı bölümü olan motor son plaktaki çöküntülere yerleşir. Sinir ucunun altında, son plağın kas zarı kavşak kıvrımları yapar. Sinirle kalınlaşmış kas zarı arasındaki aralık sinapslardaki sinaptik yarığa karşılıktır. Bütün bu yapıya nöromüsküler bağlantı denir. Nöromüsküler bağlantı (kavşak, uç plak)
Bir motor nöron ile hedef kası arasındaki işlevsel bağlantı bir kimyasal sinaps olan uç plaktır. Uç plaklar, genellikle kasın tümü veya bir bölümünde band biçiminde uzanan kümeler oluşturur. Nöromüsküler kavşak büyük bir sinapstır; büyük,
saydam asetilkolin vezikülleri içerir. Bu sinaps, öyle tasarımlanmıştır ki, motor nörondaki her aksiyon potansiyeli, presinaptik sonlanmadan (uç ayak) gereğinden fazla asetilkolin serbestleyerek (güvenlik etkeni), postsinaptik membranda mutlaka aksiyon potansiyeli oluşturabilecek depolarizasyon yaratır. Serbestlenen asetilkolin hemen asetilkolinesteraz ile hidrolizlenerek, kas lifini yeniden, bir sonraki aksiyon potansiyeli için hep veya hiç tarzı yanıt verebilecek duruma hazırlar. Aynı motor nöronun inerve ettiği kas lifleri “sadık” ve senkronize biçimde motor nöronun her aksiyon potansiyeline yanıt verirler. Nöromüsküler kavşak postsinaptik membranı, eşik değere ulaşan depolarizasyona uğrayınca, aksiyon potansiyeli, sarkolemma yüzeyinde her yöne doğru görece yavaş (3–5 m/s) yayılır. Kas lifi, büyük çaplı, miyelinsiz sinir lifi ile elektriksel benzerlik gösterir; yani, yayılma, yüksek transmembran akımlar gerektirir. Bu akımlar, görece büyük potansiyel gradyanları yaratır. Kas kasılması sırasında voltaja bağımlı kalsiyum kanalı açılır. Kalsiyum egzositozla Ach boşaltır. Mg bunu engeller. Botoks sintaksin, sinaptobrevini bloklayarak Ach’in boşalmasını engeller. Ach nikonitik müsküler (Nm) reseptörüne bağlanır. Hücre içine sodyum girer. Sinir hücre membranında yer alan bir proetin Dihidropiridin(DHP) bunu algılar. DHP ile serkaplazmik retikulum membranı bulunan ryadoin reseptörleri arasında bağlantı sayesinde sarkoplazmik retikülümdan Kalsiyum salınır. Aksiyon potansiyeli T tübülüyle yayılır. Motor nöronun tek aksiyon potansiyeli, yüzlerce kas lifini senkronize harekete geçirebildiği için oluşan elektriksel sinyal kas dışından saptanabilir (ölçülebilir) düzeydedir. Ayrıca, asgari güçten daha fazlası gerektiğinde, birçok motor nöron, senkron olmayan aksiyon potansiyelleri üretir; ancak bunların arasında, örtüşerek, senkronize görünenleri de vardır. Sonuçta oluşan karmaşık örüntülü aksiyon potansiyelleri (tipik olarak 100 uM genlikli), elektromiyogram (EMG) adını verdiğimiz teknikle, deri yüzeyindeki elektrodlar arasından yazdırılabilir. Belirli kaslardan yazdırılan bu örüntülerin birbirlerine göre farklı zamanlama ve genlik özellikleri, her bir kası inerve eden motor nöronların “kümeleşme” (birlikte davranma) etkinliklerini yansıtır. Nöromüsküler bağlantıda meydana gelen olaylar şöyledir; motor nöronun ucuna ulaşan impuls bu ucun Ca2+’a karşı geçirgenliğini arttırır. Sinir uçlarına giren Ca2+ asetilkolin içeren vesiküllerin eksositozunda belirli bir artışa neden olur. Asetil kolin motor son plak zarının kavşak kıvrımlarında yoğunlaşmış nikotinik asetilkolin reseptörlerine doğru difüze olur. Asetilkolin bu reseptörlere bağlanması, zarın Na+ ve K+ iletkenliğini arttırır ve bunun sonucunda Na’un içeriye girişi son plak potansiyeli adı verilen depolarize edici bir potansiyel meydana getirir. Bu potansiyel değişikliğin meydana gelen akım kas zarının eşik değerine ulaşınca burayı depolarize eder yani impuls kasa iletilmiş olur.
1. Sızma kanalları (Na, K) 2. Voltaja bağımlı (Na, K, Ca)
3. Ligand kapılı (Ach, GABA)
4. Mekanik (Tüy hücreleri )

H A Y V A N F İ Z Y O L O J İ S İ
159
Nöromuskuler kavşakta başlıca 3 hücrenin görevi vardır: 1) Motor sinir terminali 2) İskelet Schwan hücresi 3) Kas hücresi
Motor nöron terminali Ach bakımından zengin veziküller içermektedir. Bu veziküllerin yoğun olarak toplandıkları bölgelere aktif bölge (active zone) adı verilir. Sinir terminali mitokondri açısından da zengin olup nörotransmiter sentezi, paketlenmesi ve serbestlenmesi için gerekli enerjiyi sağlarlar. Schwan hücreleri glia hücresi olup, aksonu omurilikten çıktığı noktadan sinir terminaline kadar sarar. Preterminal bölgede bir miyelin kılıf oluşturan Schwan hücreleri, terminal bölgede non-miyelin tabakaya dönüşür. Sinapsın karşı tarafındaki bölgesi olan kas hücresi ise sinir terminaline uyacak biçimde katlanmalar gösterir. Postsinaptik bölge Ach reseptörlerince zengindir ve bu bölge reseptörleri yerlerinde tutacak özel adhezyon ve sinyal proteinlerine sahiptir. Tersine, sinaptik olmayan bölgelerdeki hücre iskeleti yapısı farklı olup katlanma göstermez. Sinaps oluşumu, bir motor aksonun immatür kas lifine (miyotüp) ulaşması ile başlar. Miyotübün ve aksonun büyüme konisinin teması ile farklılaşma başlatılılır. Büyüme konisi bir sinir terminaline dönerken karşı taraftaki bölge ise non-sinaptik bölgeye göre farklılaşma gösterir. Gelişme devam ettikçe pre ve postsinaptik bölgelerin ultrastrüktürel yapıları farklılaşır. Nöromüsküler bağlantıda bazı maddeler ileti aktarımına engel olur.
• Kürar: Asetil kolin reseptörlerine bağlanır. Asetil kolinin post sinaptik reseptörlere bağlanmasına engel olur. Aneztezide kullanılır.
• Botulium toksini: Clostridium botulium bakterisi tarafından sentezelenen bir ekzotoksindir. Otonom sistemdeki kas paralizine yol açar.
• Myastenia gravis: Asetil kolin reseptörlerine karşı oluşturulan otoimmun bir hastalıktır. Otoantikorlar Asetil kolin reseptörlerine bağlanarak Asetil kolinin post sinaptik reseptörlere bağlanmasına engel olur. Sinir kas bağlantısı zayıflar.
• Organik fosforlu bileşikler: sinaptik boşlukta bulunan Asetil kolinini parçalayan Asetik kolin esteraz inhibe ederek kasta sürekli aksiyon potansiyeli oluşmasına ve tetaniye neden olur.

H A Y V A N F İ Z Y O L O J İ S İ
160
ŞEKİL 2.69 Nörumuskular bağlantı
Her miyofibril sarkomer denilen ve kasılma yeteneği olan parçalardan meydana gelmektedir. İskelet kaslarında uyarılar nöronlar tarafından oluşturulur. Diğer bir deyişle, iskelet kasları sinirsel impuls (uyarı) olmadıkça kasılamazlar. Kaslarda aksiyon potansiyeli oluşturup kasılmayı başlatan nöronlara "motor nöronlar’’ adı verilmektedir. İskelet kaslarının motor nöronlarındaki zedelenmeler bu kaslarda atrofi ve felce neden olur. Motor nöronlar bir iskelet kas lifi üzerinde sinir-kas kavşağı adı verilen özelleşmiş bir bölgede sonlanırlar. Motor sinirin ucuna gelen uyarı bu ucun Ca2+’a geçirgenliğini artırır ve Ca2+’un sinir hücresi içine girmesine neden olur. Hücre içindeki Ca2+ miktarının artması asetilkolin veziküllerinin ekzositozunu artırır. Asetilkolin sinir-kas bağlantısındaki boşluğu geçerek motor son plaktaki asetilkolin reseptörüne bağlanır. Asetilkolinin reseptörüne bağlanması zarın Na+ ve K+ geçirgenliğini değiştirir ve bunun sonucunda Na+ hücre içine girer, motor son plak potansiyeli meydana gelir. Bu lokal potansiyel komşu hücre zarını depolarize eder ve aksiyon potansiyeli başlar. Bu bölgelerin depolarizasyonundan sonra tübüllerin sisterna bölgelerindeki dihidropiridin reseptörleri tübüllerdeki uyarıyı, protein eşleşmesi yoluyla, intraselüler komşuluklarındaki sarkoplazmik retikulum ryanodin reseptörlerine ulaştırırlar. Ryanodin bir Güney Amerika bitkisi olan Ryania speciosa (Flacourtiaceae) adlı bitkide bulunmuş zehirli bir alkaloid olup insektisid olarak kullanılmaktadır. Bu bileşik çizgili kas ve kalp kasında bulunan önemli bir Ca++ kanalı olan ryanodin reseptörü üzerinde oldukça yüksek bir affiniteye sahiptir. Bu nedenle bu almaçlara ryanodin adı vermiştir. Sarkoplazmik retikulumdaki ryanodin reseptörleri (Ca++-release channels) aracılığı ile sarkoplazmaya, miyofibrillerin çevresine Ca++ salınır. Ortamda Ca++ artışı sonucunda miyofilament düzenleyici proteinlerden tropomiyozin, troponin ile aktinin bağlantısını keser ve aktin miyozin üzerinde, ATP’nin hidrolizasyonu ile oluşan ADP ve Pi bağlantısı aracılığı ile kayarak kontraksiyonu gerçekleştirir. Bu olaylara uyarılma-kasılma eşleşmesi adı verilir. Bu süreçler esnasında unutulmaması gereken nokta ise; oluşan aksiyon potansiyeli her iki yönde kas hücre zarı boyunca iletilir ve kısa bir latent dönemden sonra kas kasılması ile ilgili süreçler başlar.
Nöromüsküler
kavşak Nöromüsküler kavşak, yapısı ve fonksiyonları bakımından sinir hücreleri arasında iletimin sağlandığı sinapslara benzer.

H A Y V A N F İ Z Y O L O J İ S İ
161
Kas Kasılmasi
Kas istirahatte iken troponin TnT alt ünitesi tropomiyozine bağlıdır. Troponin TnI alt ünitesi aktine bağlıdır. İstirahatte halinde sarkomerde kalsiyum iyon konsantrasyonu düşüktür. Bu durumda troponin alt üniteleri arasında bağlar gevşektir. Kasa impuls (akisyon potansiyeli) ulaşıp kalsiyum iyonları serbest bırakılınca, TnC alt ünitesine bağlanırlar; troponin alt ünitelerinin birbirlerine olan bağlantıları kuvvetlenir ve tropomiyozini iki aktin zincirinin birleşme yerine doğru çekerler. Tropomiyozin istirahatteki yerinden ayırılınca, aktin üzerinde açık bıraktığı yere miyozin başları bağlanır. Bu bağlanma sonucu ATP parçalanması ve kas kontraksiyonu, olamaktadır. ATP hidrolizi magnesyuma(Mg) ihtiyaç gösterir. Hücrede, ATP ve ADP, Mg2+ iyonları ile kompleks yapmış olarak bulunmaktadır. ATP’nin katıldığı reaksiyonları hücre içi Mg2+
iyon konsantrasyonu etkiler. ATP hidrolizi sırasında iki türlü aktin-miyozin kompleksi meydana gelir ve birisi alçak enerjili, öteki yüksek enerjilidir. Yüksek enerjili aktif kompleks (yüklü miyozin-ATP kompleksi) aktin ile bağlanınca meydana geliyor. ATP parçalanır parçalanmaz bu kompleks alçak enerjili kompleks haline dönüşmektedir. Bu alçak enerjili komplekse Rigor kompleksi denir. Yeni bir ATP-Mg gelip eklenmedikçe akto-miyozin kompleksi çözülmez. Normal kasta ATP her zaman mevcuttur ve ATP alçak enerjili aktin-miyozin kompleksine eklenerek miyozini aktinden ayırır. Fakat canlı ölünce mevcut ATP kısa zamanda tükenir ve alçak enerjili komplekse, yahut rigor kompleksine, eklenecek ATP bulamayınca, aktin-miyozin kompleksi birbirinden ayrılmaz; kas daimi kasılmış kalır ve ölüm katılığı dediğimiz durum meydana gelir. Kas kasılmasında bir dizi sıralı olay gerçekleşir.
ŞEKİL 2.70 Kasta Uyarıya Bağlı Farklı Kasılma
Sarkomer kas kasılması esnasında %70’i kadar kısalabilir. Kısalan ana sarkomer bileşenleri kalın ve ince filamentlerdir.

H A Y V A N F İ Z Y O L O J İ S İ
162
�• Kastaki depolarizasyon Na ile, kasılma Ca ile olur.
Akson ucuna uyarı geldiğinde voltaja bağımlı kalsiyum kanallarından hücre içine kalsiyum iyonları girer. Kalsiyum iyonu asetilkolin veziküllerinin ekzositozuna neden olur. Asetil kolin nikotinik ACh reseptörüne tutunur. Hücre içine Na iyonları girer. Na iyonu çizgili kasta depolarizasyonu başlatır. Depolarizasyon membranda bulunan dihidropiridin (DHP) reseptörü tarafından algılanır. DHP reseptörü sarkoplazmik retikulumdaki kalsiyum kanalını (ryanodin) açar.
ŞEKİL 2.71 DHP ve Ryanodin reseptörler ile çizgili kas hücresinde Ca iyonu salımı.
1. Aksiyon potansiyeli motor sinir boyunca kas lifindeki sonlanmasına kadar yayılır.
2. Her sinir ucundan nörotransmiter olarak az miktarda asetilkolin (Ach) salgılanır.
3. Kas lifi membramnda lokal bir alanda etki gösteren asetilkolin, membrandaki asetilkolin kapılı kanalları açar.
4. Asetilkolin kanallarının açılması, kas lifi membranından çok miktarda sodyum iyonunun içeri girmesini sağlar. Bu olay kas lifinde aksiyon potansiyelini başlatır.
5. Aksiyon potansiyeli sinir membranında olduğu gibi kas lifi membranı boyunca da yayılır.
6. Aksiyon potansiyeli kas lifi membranın depolarize eder ve kas lifi içine doğru yayılarak, sarkoplazmik retikulum da depolanmış olan kalsiyum iyonlarının büyük miktarlarda miyofibrile serbestlemesine neden olur.
7. Kalsiyum iyonları, kasılma olayının esası olan filamentlerin kaymasını sağlayan, aktin ile miyozin filamentleri arasındaki çekici güçleri başlatır.
8. Sonra, saniyenin bölümleri içinde kalsiyum iyonları sarkoplazmik retikuluma geri pompalanır(SERCA). Yeni bir kas aksiyon potansiyeli gelinceye kadar burada depolanır: kalsiyum iyonlarının uzaklaştırılması kasılmanın sona ermesine neden olur.

H A Y V A N F İ Z Y O L O J İ S İ
163
Böylece kalsiyum intraselüler sıvıya salınır. Kalsiyum iyonları troponin C’ye bağlanır ve troponin molekülünde dönme hareketi oluşur. Troponin T’ye bağlı olan tropomiyozin de aktin üzerindeki yerinden ayrılır ve böylece miyozinin bağlanacağı bölgeler açığa çıkmış olur. Aktin bağlanma bölgeleri miyozin başını fosforiller. Ortamda magnezyum Mg varlığında miyozin başının ATPaz etkisi ortaya çıkar. İki Z çizgisi birbirine yaklaşır. H ve I bantlarının boyu kısalırken A bandının boyu değişmez. Bu şekilde aktin-miyozin kompleksi yeni bir ATP molekülü miyozin başına bağlanıncaya kadar değişmeden kalır. Buna ATP’nin yumuşatıcı ya da gevşetici etkisi denir. İskelet Kasında Gevşeme
T-tübülü ile terminal sisternalar arasında bir açıklık olmasına rağmen protein köprüleri bu açığı kapatır. Bu köprü proteinlerine ayaklar denir. Bu ayaklar terminal sisterna membranındaki Ca+2 salgı kanallarından ve aksiyon potansiyeline tepki olarak hücre içi Kalsiyum artışından sorumludur. Bu kanala ryanodin bağlandığından yaygın olarak bunlara ryanodin reseptörü (RYR) denir. Serkoplazmik retikülüm membanının bir kısmına gömülü olan RYR proteininin çoğu miyoplazma içindedir ve terminal sisterna ile T-tübülü arasındaki boşluğu doldurur. T-tübül membranında RYR’nin dihidropridin reseptörü (DHPR) denilen bir proteinle etkileşime girdiği düşünülmektedir. DHPR bir L-tipi voltaj kapılı Ca+ kanalıdır.
ŞEKİL 2.72 Miyofilament ve triad
ATP miyozini aktinden ayırır. Miyozin başının enerjisini arttırır. Ca’un geri sarkoplazmik retikuluma pompalanmasıyla gevşeme oluşur. Ca’un geri alınmasında bir ATP harcanır. Eğer gevşeme için gereken ATP olmazsa kas gevşeyemez. Kalsiyum SR’den salınımında görevli reseptör ryanodindir. Kas gevşemesi için kalsiyumu SR’ye pompalayan Ca pompasının adı ise SERCA’dır. Sarkoplazma içerisinde yer alan organellerden birisi de Sarkoplazmik Retikulum, uzunlamasına (longitudinal) tübüller ve bunların sonlandıkları sarnıç bölgelerinden oluşur. Uzunlamasına tübüller myofibrillere paralel olarak yerleşmişlerdir. Sarkoplazmik retikulum sarnıçları, hücre zarından lif içine doğru kıvrım yapmasıyla oluşan transvers
T tüpüller Sarkolemmanın kas lifleri içerisine yönelmesi ile oluşan transvers tubuller

H A Y V A N F İ Z Y O L O J İ S İ
164
tübüllerle (T tübüller) her iki yanda komşuluk yapar. Böylece T Tübül-Sarkoplazmik retikulum Sistemi ilişkisi sayesinde aksiyon potansiyeli lif içlerine kadar iletebilir. Bu ileti bir kalsiyum deposu olan sarkoplazmik retikulumdan Ca2+ iyonunun sarkoplazmaya salınmasına yol açarak kas kasılmasına neden olur. T-Tübül Sarkoplazmik Retikulum Sisteminin kas lifinde oluşturduğu hacim, antrenmanlı bireylerde normalin 3 katına kadar ulaşabilir. Transvers tübül’ ya da “T sistemi’’ denilen borucuklar, sarkolemmadan kasın içine doğru girerler ve ‘’Z’’ hattı boyunca yahut ‘’A’’ bandı ile ‘’I’’ bandı sınırı boyunca ilerlerler. Bu sistem sayesinde sarkolemma’nın depolarizasyon dalgası kasın içine, kontraktil elementlere kadar götürülür. Myofibriller arasında bulunan, kas filamentlerinde paralel biçimde yer almış, sarkomer boyunca devam eden sarkoplazmik retikulum bulunur. Z hatları hizzasında her iki kenarı kapalıdır ve transver tübüllerle (T sistemi ile) temas halindedir. Transvers tübülün iki tarafında yer alan sarkoplazmik retikulum kapalı uçları “terminal sisterna’’ adını alır. Transvers tübül ile terminal sisternaların uçlarının birleştiği yerde üçlü bir yapı meydana gelir. Buna ‘’Triad’’ denir. Trias ‘’Z’’ bandı hizzasında veya ‘’A’’ bandı ile ‘’I’’ bandı sınırı hizzasında bulunur. Tetani
Bir kasın uyarılma sonunda sitoplazmaya verilen kalsiyum iyonlarını sarkoplazmaya geri pompalanmadan yüksek sıklıkla gelen uyarı nedeniyle tekrar kasılması, kasın gevşeme yapmadan kasılmasıdır. Bu durumdaki bir kas gevşeme yapmadan kasılı kalır. Tetani durumu kasın gevşeme tapmadan ardı ardınca süerkli kasılmasıdır.
ŞEKİL 2.73 Merdiven ve tetanus olayı
Tetani olayı için kasın kasılma frekansının saniyede 100 kez fazla olması gerekir.
Tetani-Tetanus Tetanoz iskelet kas liflerinin toksin kaynaklı olarak uzun bir süre kasılı kalmasıdır. Gram-pozitif, zorunlu anaerobik basil bakteri olan Clostridium tetani kaynaklıdır.

H A Y V A N F İ Z Y O L O J İ S İ
165
ŞEKİL 2.74 Merdiven ve tetanus olayı
Tetanus, merdiven(treppe) olayları birbirini dönüşebilen olaylardır. Eşik değeri üzerinde kasılma yapan kas gelen uyarının frekansı ile kasın gevşemesi için gerekli olan (kalsiyum uzaklaştırılması ve ve ATP sentezi) olayların gerçekleşmesi arasındaki ilişkiye bağlıdır. Kalp kası tetani geçirmez çünkü kalp kasını tekrar uyarılması için gereken sürede kalp kası gevşemiştir. Başka bir ifade ile Mutlak Refrakter periyod süresi kalp kasında gevşemeye için verecek kadar uzundur.
Merdiven Treppe Olayı Bir kasa kası tetanize etmeyecek frekansta uyarı verilirse her bir kas sarsısında oluşturulan gerim giderek artar. Kasın kasılmasına neden olan uyarının sıklığının artmasına bağlı olarak kasta meydana gelen gerilim artışıdır. Birçok kasılmadan sonra uyarı başına tekdüze bir gerim oluşur. Buna merdiven (Treppe) olayı denmektedir. Başka bir deyişle iki uyarı arasındaki süre sarsı süresinin 1/3’ünden daha uzundur.
ŞEKİL 2.75 Merdiven(Treppe)olayı
Merdiven olayından sorumlu mekanizma her gevşeme sırasında kalsiyum iyonlarının tamamının sarkoplazmik retikuluma geri döndürülememesidir. Sonuç olarak troponin C’ye bağlı kalsiyum artacağı için kasılmaya dahil olan aktin miyozin sayısı da artar. İskelet kası tipleri
Hızlı lifler. (1) Daha büyük kasılma gücü için daha büyük lifler. (2) Kasılmayı başlatmak üzere hızlı kalsiyum serbestlemesi için geniş bir sarkoplazmik retikulum. (3) Glikolitik işlemle hızlı enerji sağlamak için çok miktarda glikolitik enzimler. (4) Oksidatif metabolizma ikincil önemde olduğu için daha az kan akımı. (5) Yine oksidatif metabolizma ikincil olduğundan daha az sayıda mitokondri.

H A Y V A N F İ Z Y O L O J İ S İ
166
Yavaş lifler. (1) Daha küçük lifler. (2) Daha küçük sinir lifleri ile inervasyon. (3) Daha fazla oksijen sağlamak için daha gelişkin kan damarı sistemi ve kapillerler. (4) Yüksek düzeydeki oksidatif metabolizmayı desteklemek için çok sayıda mitokondri. (5) Lifler bol miktarda, eritrositlerdeki hemoglobine benzer şekilde demir içeren bir protein olan miyoglobin içerir. Miyoglobin oksijenle birleşir, onu ihtiyaç oluncaya kadar depolar ve mitokondriye oksijen taşınmasını büyük miktarda hızlandırır. Miyoglobin nedeniyle yavaş kas kırmızımsı görünür ve kırmızı kas adı verilir. Hızlı kasta kırmızı miyoglobin yoktur ve beyaz kas adını alır. İskelet Kasında Enerji
Kasta dinlenme durumunda serbest yağ asitleri mitokondride beta okside edilip ATP’ye dönüştürülür. Bu ATP kreatin ile birleştirilir ve kreatin fosfat elde edilir. Egzersiz sırasında acil depo olarak kreatin fosfat kullanılır. Egzersiz devam ederse iskelet kası glikolizi enerji kaynağı olarak kullanır. Glikoliz sonucu laktat oluşur. Laktat dokularda aljeziktir (ağrı vericidir). Oluşan laktat kan yolu ile karaciğere gier burada laktat üzerinden tekrar glikoz sentezlenir(kori döngüsü).Ağır-uzun egzersizde oksidatif fosforilasyonla enerji elde edilir. Kan glikozu ve serbest yağ asitleri (maratoncu en çok bunu kullanır) kullanılır.

H A Y V A N F İ Z Y O L O J İ S İ
167
ŞEKİL 2.76 Cori döngüsü
ŞEKİL 2.77 Kasta enerji
Depo Oksijen: Vücutta depo oksijen; 0.5 litre akciğerlerdeki havada, 0.25 litre vücut sıvılarında erimiş olarak, 1 litre hemoglobinde ile birleşmiş olarak, 0.3 litre miyoglobinde bulunur (toplam; 2 litre). Özellikle akut hipoksilerde hemoglobindeki mevcut depo oksijen ancak 1-2 dakika yeter, bu süre dışında ek enerji kaynağı gerekir. Anaerobik glikoliz ile de birkaç dakikalık ek enerji sağlanır. Bu sırada glikoz pirüvat (O2 varlığında kas glikojeni fazla laktata çevrilmiyor, bir kısmı Krebs siklusuna girer; aerobik glikoliz) , sonra laktata çevrilir; laktat hücre dışına difüze olur. Bu durum yani hipoksi maksimal kas kasılmasının ilk dakikalarında ortaya çıkar. Kasta depo glikojen (ilk 10 saniyede fosfojen sistem yeterli) 10.saniye sonunda bu sistem devreye girer; 1-2 dakikalık kısa süreli ağır aktiviteler için depolar kullanılır. Glikojen

H A Y V A N F İ Z Y O L O J İ S İ
168
tükenince ağır aktivite sonlanır ve artık aerobik sistem (ki bu sistem anaerobikten yavaş enerji üretiyor; glikojen laktik anaerobik sistem (anaerobik glikoliz), aerobik glikolizden 2,.5-3 kat daha hızlı çalışır (mitokondriyal oksidatif sistem =aerobik sistem, daha yavaş ama kalıcıdır.)Kas meydana gelen laktik asir karaciğerde glikoza dönüştürülür. Kasta ise glikoz tekrar laktik asit haline gelir. Bu dönüşülere Cori döngüsü denir. Kaslar kimyasal enerjiyi mekanik enerjiye çevirirler. Kas enerjisinin kaynağı organik fosfat (PO4) bileşikleri olan ATP (Adenozin tri fosfat) ve CP'dir (Kreatin fosfat) . Kas aktivitesi veya genel vücut dokularının aktivitesi için gereken enerji 2 ana metabolik yol ile temin edilir.
- Anaerobik sistem (yol) - Aerobik sistem (yol)
Anaerobik sistem: Anaerobik deyimi enerji eldesinde oksijenin olaya karışmadığını ya da çok az karıştığını belirtir. Bu sistemde enerji iki şekilde elde edilir. Alaktik Anaerobik Sistem (Fosfatojen = ATP- Keratin fosfat): Terim, anaerobik ortamda elde edilen enerji esnasında yan ürün olarak laktik asitin oluşmadığını açıklar. Enerji, kaslarda hazır olarak bulunan ATP' den elde edilir. Tükenen ATP' yi Keratin fosfat bir fosfatını vererek yeniler.
ŞEKİL 2.78 Kreatin ve kreatin fosfat oluşumu .
Laktik Anaerobik Sistem: Laktik terimi, anaerobik ortamda(oksijen kullanmadan) elde edilen enerji esnasında yan ürün olarak laktik asitin oluşduğunu açıklar. Karbonhidratların bir özelliği O2 siz ortamda da enerji için kullanılabilmeleridir. Bu sistemde glikoz veya glikojenin glikolitik yol ile yıkımı olur. (hızı:2,5mol/dk). Bazı omurgasızlarda kreatin yerine arjinin aynı fonksiyonu yapmak için bulunur. Arjinin ATP ile dönüşümlü olarak fosfolanarak arjinin fosfat oluşturur.
-Hücrede fazla ATP sentezlenince bunun büyük kısmı Keratin fosfat ’ ye dönüştürülerek depolanır. ATP tükendiği anda bu depo kullanılır. -Fosfokreatin + ADP����-----���� ATP + kreatin şeklinde reversible ilişki; konsantrasyona göre sürekli iki yönlü çalışır. -Keratin fosfat’dan enerji transferinin önemli tarafı, bu olayın saniyenin küçük bir bölümünde gerçekleşmesidir. Bu olay özellikle ani bir kas kasılması sağlar. ATP ile beraber bu sisteme fosfatojen sistem denir ve her ikisi toplam 8 – 10 saniye maksimal kas gücü sağlar.

H A Y V A N F İ Z Y O L O J İ S İ
169
� ŞEKİL 2.79 Laktat sentezi.
Özellikle akut hipoksilerde hemoglobindeki mevcut depo oksijen ancak 1-2 dakika yeter, bu süre dışında ek enerji kaynağı gerekir. Anaerobik glikoliz ile de birkaç dakikalık ek enerji sağlanır. Bu sırada glikoz pirüvat (O2 varlığında kas glikojeni fazla laktata çevrilmiyor, bir kısmı Krebs siklusuna girer; aerobik glikoliz) , sonra laktata çevrilir; laktat hücre dışına difüze olur. Bu durum yani hipoksi maksimal kas kasılmasının ilk dakikalarında ortaya çıkar. Kasta depo glikojen (ilk 10 saniyede fosfojen sistem yeterli ) 10. saniye sonunda bu sistem devreye girer; 1-2 dakikalık kısa süreli ağır aktiviteler için depolar kullanılır. Glikojen tükenince ağır aktivite sonlanır ve artık aerobik sistem (ki bu sistem anaerobikten yavaş enerji üretiyor; glikojen laktik anaerobik sistem (anaerobik glikoliz), aerobik glikolizden 2,5-3 kat daha hızlı çalışır (mitokondriyal oksidatif sistem =aerobik sistem, daha yavaş ama kalıcıdır.) - Fosfatojen sistemin enerji üretim hızı ise anaerobik sistemin yaklaşık 2 katıdır. Ağır egzersizde glikojen deposu azalırken laktik asit konsantrasyonu artar. Egzersiz sonrası normal oksijen sağlanarak biriken laktik asit, tekrar glikoza çevrilir (çoğu karaciğerde cori döngüsü yada laktik asit döngüsü). Bir kısmı da (1/5) pirüvik aside çevrilerek siklik asit siklusunda kullanılır. 2.Aerobik Sistem: Kas kaynaklanan vücut ısısı CO2, Laktik asit, H+ İyonları gibi düzeylerini yükseltip, O2 düzeyinin azalışı sonucunda oluşan kimyasal düzenlemeler ile bölgesel damarlarda genişleme ve kan akımı artışı sağlanmaktadır. Bunun yanında İstirahat şartlarında total kalp debisinin, yani 1 dakikalık bir süre içerisinde perifere pompalanan kanın sadece %15-20 si gibi bir oranı iskelet kaslarına gönderilmektedir. Halbuki egzersizde egzersizin türü ve şiddetine bağlı olarak bu oran %80-85 gibi bir seviyeye ulaşmaktadır. Kasa gelen oksijen myoglobin tarafından depolanır. Besin maddelerinin mitokondrilerde oksidasyonu ile ATP sentezidir. Glikoz, yağ asitleri, aminoasitler, O2 ile birleşerek AMP (Adenozin mono fosfat) ve ADP (adenozin di fosfat)’ nin ATP’ye çevrilmesinde tüketilecek büyük miktarlardaki enerjiyi
- Glikoz + 2ATP (önce pürivat, sonra laktat oluşur) = 2 Laktik Asit + 4ATP, net kazanç 2 ATP - Glikojen + 1ATP , 2 Laktik Asit + 4ATP, (net kazanç 3ATP) - Glikozdan ATP üretiminin farkı; hücreye giren serbest glikozun parçalanmadan önce 1 mol ATP ile fosforilasyonudur. (+ 1 ATP’ de yıkım için kullanılır, tüketim=2 ATP). - Oysa glikojendeki glikoz zaten fosforiledir. Bu durumda hücredeki anaerobik koşullar için en önemli kaynak depo glikojendir.

H A Y V A N F İ Z Y O L O J İ S İ
170
serbestleştirirler. Örneğin: Depo glikojen tükenince yerine plazmadan glikoz alımı ile enerji sağlanır. Glikoz önce pirüvik aside dönüşür. Ortamda yeterli O2 varlığında pirüvik asit Krebs siklusuna girerek bir glikozdan 40 mol ATP elde edilir (2 ATP kullanılır net kazanç 38 ATP’dir). Besinler ve O2 olduğu sürece bu üretim sınırsızdır (O2 yetersiz ise pirüvat laktata dönüşür; anaerobik sistem). Karbonhidratların enerji için yetersiz olduğu veya kullanılmadığı koşullarda yağ asitleri, mitokondrilerde CO2 ve H2O’ya kadar yıkılır. Yağ asitleri oksidasyonu, serbest yağ asitlerinin kandan hücrelere alınmasıyla başlar. Mitokondride beta oksidasyon ile yağ asitleri asetil Co-A’ya yıkılır. Asetil CoA Krebss siklusuna girerek okside edilir.
ŞEKİL 2.80 Krebs döngüsü.
Oluşan ATP miktarı yağ asit zincirinin uzunluğuna bağlıdır (ör: palmitik asit; 129 ATP elde edilir). Yağ asitleri biter veya yetersiz olursa artık vücudun depo proteinleri yıkılır ve enerji elde edilir. Sonuçta üre meydana gelir, normal şartlar altında günlük fizyolojik bir protein yıkımı ve üre oluşumu vardır.
ŞEKİL 2.81 Cori döngüsü.
Kas hücresinde glikoliz sonunda meydana 3 karbonlu prüvat laktat haline getirilerek kan dolaşımı ile karaciğere taşınır. Karaçiğerde kandan alınan laktat prüvat haline getirilir. Karaçiğerde prüvat glukoneogenesiz ile tekrar glikoz haline getirilir. Bu yolla kas ve karaciğer arasında laktat taşımasıyla meydana gelen döngüye Cori döngüsü denir. Kana verilen laktat karaciğer dışında kalpte kas hücrelerince enerji kaynağı olarak kullanılır. Bu yolla eksersiz sırasında yoğun kas faaliyeti sonunda oluşan laktat kalp tarafında kullanılır.
Glikoliz
Glikozun pirüvik aside dönüşüm sürecidir. Bu süreç, hücrede bir çok safhada tamamlanır, yüksek enerji (ATP) oluşumu için her reaksiyon özel bir enzim tarafından katalizlenir. Bu süreç ya glikojenin parçalanması ile oluşan glikozu ya da kanda normal olarak bulunan glikozu

H A Y V A N F İ Z Y O L O J İ S İ
171
kullanır. Aerobik glikolizde pirüvik asit su ve CO2 ye indirgenir, Anaerobik glikoliz sonucu laktik asit oluşur. �
Kasılma Tipleri
Kasılma kasın kasılma sırasında boyunun kısalmasına, kasılma sırasındaki kuvvet değişimine göre farklı tiplere ayrılır.
Kas spazmı
Düz ve iskelet kasında gözlenen kasın sürekli olarak(uyarı ile) kasılması ve kasılı kalması durumudur. Spazm sırasında kas sürekli kasılır ve gevşemez. Örneğin bir kemiğin kırılmas durumda kemik çevresindeki kaslar yoğun açı uyarısıyla(omurilik üzerinden meydana gelen bir refleks ile) kasılır. Spazm başka bir nedeni ise kasın yetersiz dolaşıma bağlı olarak yeterşnce oksijen alamaması ve buna bağlı olarak oluşan ATP yetersizliğidir. Daha önce kısımlarda anlatıldığı gibi ATP kastaki miyoglobine bağlanması durumda kas gevşer. ATP eksikliği gevşemeye engele olur. Benzer şekilde kasın yüksek metabolik aktivasyonda metabolik atıkların artmasına neden olarak spazm oluşturabilir. Spazma neden olan diğer faktörler kalsiyum ve/veya mağnezyum eksikliğidir. Rigor Mortis
Ölen kişide ATP yokluğuna bağlı olarak oluşan bu katılığa rigor mortis (ölüm katılığı. 3. saatte görülür. 18. saatte çözülür) denir. Ölümden birkaç saat sonra bütün vücut kasları “rigor mortis" (ölüm katılığı) denen bir kontraktür durumuna girer; bunda aksiyon potansiyeli olmadığı halde kas kasılır ve katılaşır. Bu katılık, gevşeme sırasında çapraz köprülerin aktin filamentlerinden ayrılması için gerekli olan ATP’nin tamamen kaybedilmesinden kaynaklanır. Kasların gevşemesi sırasında kalsiyum iyonları aktif taşınma ile sarkoplazmik retikuluma geri alınır. Bu nedenle gevşeme sırasında da ATP tüketimi ve enerji sarfı vardır. Kaslar kas proteinleri yıkılana kadar katılıkta kalırlar. Yıkım 15–20 saat sonra lizozomlardan serbestleyen enzimlerin neden olduğu otolizle gerçekleşir.
• İzotonik kasılma: Kas kasılınca boyu kısalıp geliştirdiği kuvvet sabit kalıyorsa bu izotonik kontraksiyondur. • İzometrik kasılma: Kas kasılınca boyu değişmiyor, ancak oluşan kuvvet değişiyorsa bu izometrik kontraksiyondur. Duvarın itilmesi bu tip kasılmayla oluşur. • Oksotonik kasılma: Kasın hem geliştirdiği kuvvetin hem de boyunun değişmesidir.

H A Y V A N F İ Z Y O L O J İ S İ
172
Kalp kası
Kalp doku olarak Üç tabakadan oluşur; Epicardium: Kalbin yüzeyini kaplayan seröz membrandır, Myocardium: Kalp kasılmasından sorumlu kaslardan oluşan orta tabakadır, Endocardium: Kalbin iç boşluğunu kaplayan en içteki tabakadır. Kalp kasılmasında sorumlu olan kısmı myokardiumdur. Myokardiyom çizgili kas tipidir. Çizgili kas olmasına rahmen kalp kası ile çizgili kaslar arasında belirgin farklılar vardır. Kalp hücreleri miyozitler yapısal olarak iskelet kasınınkilere benzer kasılabilir diziler içerir İskelet ve kalp kasları arasındaki belirgin farklar düzenleyici protein troponin ile başlar.
A. Troponin :Troponin aktin ince flamanı ile ilişkili kalsiyuma duyarlı bir heterotrimerdir. Sarkoplazmik Ca2+ yoğunluğundaki artışlar troponinde üç boyutlu bir değişime yol açar. Değişim, ince flaman üzerindeki tropomiyozini yuvarlayarak miyozin bağlanma bölgelerinden uzaklaştırır ve böylece aktinmiyozin etkileşimini sağlar. İskelet kasında kasılma başlamadan önce troponin C iki Ca2+ iyonu bağlamak zorundadır. Kalp kasında kasılma için sadece bir Ca2+ iyonu gereklidir. B. İletişim (Komünikasyon) Yolları :İskelet kası lifleri istemli kontrol altındadır ve yalnızca bir alfa-motor nöron tarafından emir verildiğinde kasılır. Kalp kası sinir sistemi tarafından düzenlenir, fakat kasılmak üzere emir normalde miyokardiyumu bir hız düzenleyici (pacemaker) olarak işlev görmek üzere özelleşmiş olan bir bölgesinden (sinoatrial düğüm) kaynaklanır. Uyarı başlatan (pacemaker) hücreler organ içindeki tüm lifler dâhil olana kadar hücreden hücreye yayılan uyartıları periyodik olarak çıkarır. Bu, oluklu bağlantılar (gap junctions) ve ileri düzeyde hücresel dallanma ile mümkün olur. 1. Uyarı başlatan hücreler (pacemaker): Kardiyak uyarı başlatan hücreler sağ atriyum duvarı içinde üst kısımda yer alır. Bu bölgedeki hücreler kasılabilir değildir ve membranları “tuhaf akım” (funny current) ([lf] oluşturan iyon kanalları içerir ki bunlar spontan olarak depolarize olmalarına ve dakikada yaklaşık 100 kere aksiyon potansiyeli oluşturmalarına yol açar.
ŞEKİL 2.82 Kalp kasında aksiyon potansiyeli sırasındaki iyıon değişimi.
Bir kez başladığında bir aksiyon potansiyeli yenilenerek her iki atriyum ve sonunda ventrikülleri içerecek biçimde dağılır, bunu kasılma izler. Düğüm hücrelerinin hangi hızda depolarize olacakları ve uyarı başlatacakları OSS tarafından kalp hızını kontrol etmenin bir yolu olarak düzenlenir.

H A Y V A N F İ Z Y O L O J İ S İ
173
2. Oluklu bağlantılar: Oluklu bağlantılar bitişik hücreleri birbirine bağlar ve doğrudan elektriksel ve kimyasal iletişim için yollar sağlar. Bunlar, hücreleri birbirine mekanik olarak kaynaştırarak kas kasılması sırasında oluşturulan gerilime dayanmalarını sağlayan sarkolemmaya ait yapısal elemanlar da içeren (dezmozomlar ve fascia adherens), interkale diskler olarak bilinen özelleşmiş temas bölgelerinde yer alırlar. Bu, komşu iskelet kası lifleri arasında iletişim için yolların hiç bulunmaması ile zıtlık oluşturur. Kalp kasları iskelet kasları gibi çizgilidir. Kalp kas hücrelerin dallanmalar göstermesi iskelet kasına göre çok daha fazladır. Kalp kasının en belirgin özeliği belli bölgelerde özelleşmiş yapılar aracılığı ile birbirlerine bağlanmış olmasıdır (sinsisyal yapısı, gap junction, konekson) Kalp kaslarında bulunan bağlantı bölgeleri(gap junction konekson), bir hücredeki aksiyon potansiyelinin diğer bir hücreye kolayca geçer (interkalar diskler).Kendi uyarılarını kendisinin oluşturması ile ritmik kasılmalar yapar. Kalp kası bir otonomiye sahiptir. Kalp kası sinir sisteminden bağımsız bir organ değildir. Kalbin uyarı ve ileti sistemi ayrıdır. Kalp kası otonom sinir sisteminin gerek sempatik gerekse parasempatik bölümüne ait nöronlar ile bağlantıya sahiptir. Kalp kasının kasılma mekanizması iskelet kasına benzer ancak önemli bir fark, kalp kasının kasılma sırasında hücre içindeki kalsiyum iyonlarına ilaveten hücre dışından gelen kalsiyum iyonlarını da kullanmasıdır.
3. Hücresel Dallanma: Uyaran oluşturan hücreler tarafından bir kasılma emri verildiğinde bunun tüm kalp boyunca hızlı bir biçimde yayılması gerekir. İskelet kası lifleri uzun, ince ve dalsız iken kalp hücreleri geniş bir kapsamda dallanır. Dallanan hücreler ve oluklu bağlantıların kombinasyonu birbiriyle ileri düzeyde bağlantılı tek bir ünite (sinsisyum) olarak işlev gören hücresel bir ağ oluşturur. C. Transvers tübüller ve sarkoplazmik retikulum :Kalbin uyarı başlatan hücreleriyle oluşturulan aksiyon potansiyeli her kas lifinin merkezine transvers (T) tübüller ile aktarılır ve sonra sarkoplazmik retikuluma (SR) elektromekanik transdüksiyon (dönüştürme) için aktarılır. Her ne kadar SR kasılma için ihtiyaç duyulan Ca2+ hacmini temin etse de kalp kasında membran sistemi iskelet kasına nazaran daha az yaygınlaşmıştır. Kalp T tübüllerinde ve SR yapısında iskelet kasıyla karşılaştırıldığında iki dikkat çekici fark daha vardır. 1. Lokasyon: İskelet kasında T tübülleri kalın flamanların sonları boyunca uzanır (A-I birleşim yerinde), her sarkomer başına iki tanedir. Kalp kasında daha az tübül vardır ve Z hattı boyunca uzanırlar. T tübülleri kalp kasında daha geniştirler ve dallanmaları daha düşük kapsamlıdır. 2. Diadlar: T tübülleri aksiyon potansiyellerini SR’ye taşırlar. İskelet kasında tübüller triad olarak bilinen kavşak komplekslerinde iki SR sisternasıyla birleşirler. Kalp kasında, tübüller SR’nin tek bir uzantısıyla diad denilen benzer bir yapıda birleşirler. Kalp ve iskelet kası zarlarının özellikleri arasındaki en az iki temel farklılık, kalp kasında aksiyon potansiyelinin uzun sürmesinden ve platonun oluşmasından sorumludur. Birincisi, iskelet kasında aksiyon potansiyeli neredeyse tamamen,
(1) çok büyük miktarlarda sodyum iyonunun iskelet kası lifine girmesine izin veren çok sayıda hızlı sodyum kanalının aniden açılması ile meydana gelir. Bu kanallara “hızlı” kanallar adı verilmiştir, çünkü yalnızca birkaç 1/10.000 saniye süresince açık kalır ve sonra aniden kapanırlar. Kanallar kapandıktan sonra repolarizasyon başlar ve bir başka 1/10.000 saniye veya daha uzun bir süre içerisinde aksiyon potansiyeli son bulur.

H A Y V A N F İ Z Y O L O J İ S İ
174
(2) Diğer yandan, kalp kasında, aksiyon potansiyeli iki tür kanalın açılması ile meydana gelir:
(a) iskelet kasındakilerin aynısı olan hızlı sodyum kanalları,
(b) başka bir grup kanal olan yavaş kalsiyum kanalları ki bunlara kalsiyum-sodyum kanalları da denir.
Bu ikinci gruptaki kanallar daha yavaş açıldıkları için hızlı sodyum kanallarından farklıdır. Fakat daha da önemlisi, birkaç 1/10 saniye süresince açık kalırlar. Bu süre içerisinde çok büyük miktarlarda kalsiyum ve sodyum iyonu, bu kanallardan geçerek kalp kası lifinin içine akarlar. Bu akış, uzun süreli bir depolarizasyon sağlayarak aksiyon potansiyelindeki platoyu oluşturur. Dahası, aksiyon potansiyeli sırasında kasa giren kalsiyum iyonları, kas kasılması olayının uyarılmasına aracı olarak, önemli bir görev yaparlar. Kalp kası ve iskelet kası arasındaki bir başka farklılıktır. Kalp kası ve iskelet kası arasındaki ikinci temel işlevsel farklılık şudur: aksiyon potansiyelinin başlamasından hemen sonra iskelet kasında gözlenmeyen bir olay meydana gelir, kalp kası zarının potasyum geçirgenliği yaklaşık beşte birine kadar azalır. Potasyum geçirgenliğindeki bu azalma, aşırı miktarda kalsiyumun yukarıda belirtilen kalsiyum kanallarından hücre içine akmasına bağlı olabilir. Nedeni ne olursa olsun, aksiyon potansiyelinin platosu sırasında potasyum geçirgenliğinin azalması, potasyum iyonlarının hücre dışına akışını önemli derecede azaltarak potansiyelin dinlenim düzeyine dönmesini geciktirir. Yavaş kalsiyum-sodyum kanalları 0.2-0.3 saniyenin sonunda kapanınca, kalsiyum ve sodyum iyonlarının hücre içine akışı durur, zarın potasyum geçirgenliği hızla artar. Lif hızla potasyum kaybedince, zar potansiyeli dinlenim düzeyine geri döner, böylece aksiyon potansiyeli son bulur.
ŞEKİL 2.83 Kalpteki periyodik sinir uyarımı
Kalp kası laktik asidi piruvik aside çevirerek enerji için kullanma yeteneğine sahiptir. Bu olay buyuk olcude ağır egzersiz sırasında, iskelet kaslarından kana buyuk miktarda laktik asit serbestlendiği zaman gercekleşir. Daha sonra bunlar kalp tarafından enerji kaynağı olarak kullanılırlar.

H A Y V A N F İ Z Y O L O J İ S İ
175
Düz Kas
Düz kaslar sempatik ve parasempatik sinir sistemi ile uyarılmasına karşın hormonlarla da uyarılabilir. Tek nükleus taşıyan mekik şeklinde hücrelerden oluşurlar. Yavaş ancak güçlü kasılma yapabilirler. İskelet kaslarına göre çok az enerji harcarlar. Düz kas, kimyasal özellikleri iskelet kasındaki aktin ve miyozin filamentlerine benzeyen aktin ve miyozin filamentleri içerir. Aktin/miyozin oranı: düz kasta daha fazla (10:1), iskelet kasında (2:1). İskelet kasında kasılmanın kontrolü için gerekli olan, normal troponin kompleksini içermez, fakat kaldesmon (aktin ile ilişkili miyozin ATPaz inhibitörü) ve kalponin (Ca+2-kalmodulin bağımlı protein kinaz fosforilasyonuyla düzenlenen yaygın bir aktin ile ilişkili miyozin ATPaz inhibitörü) denen kasa özel iki düzenleyici proteinle bağlantılıdır, dolayısıyla kasılmanın kontrol mekanizması farklıdır. Düz kasta iskelet kasındaki aktin ve miyozin filamentlerinin çizgili yerleşimi yoktur. Dolayısıyla sarkomer yapısı da yok. Düz kasta Z çizgisi yoktur, onun yerine benzer göreve sahip olan yoğun cisimcikler (dens body) vardır. Miyozin flametlerin çoğu “yan kutup” adı verilen çapraz köprülere sahiptir, böylece bir yandaki köprüler döndüğünde o tarafa doğru bir kısalma sağlanırken diğer yandaki köprülerin dönmesi diğer yana doğru kısalma sağlar. Bu mekanizma miyozinin aktini bir uçta bir yana doğru çekerken aynı anda başka bir aktin flamentini başka bir bölümde zıt yöne doğru çekmesini sağlar. Bu düzenleme sayesinde iskelet kasında kasılmayla kasın boyu %30’dan az oranda kısalırken düz kasta bu oran %80’e çıkmaktadır. Mandal Mekanizması
Düz kas bir kez tam kasıldığında, kasın aktivasyon derecesi genellikle başlangıç seviyesinin çok altına düşürülebilir, ancak kas yine de tam kasılma gücünü sürdürebilir. Ayrıca, kasılmayı sürdürmek için tüketilen enerji genellikle çok azdır, bazen eşdeğer iskelet kasının sürekli kasılması için gerekli olan enerjinin 1/300’ükadardır. Buna “mandal” mekanizması denir. Aynı etki iskelet kasında da ortaya çıkar ancak düz kasa oranla çok daha azdır. Mandal mekanizmasının önemi, düz kasta az miktarda enerji kullanarak saatlerce tonik kasılmanın sürdürülebilmesini sağlamasıdır. Bunun için az miktarda sinirsel ya da hormonal uyarıya(epinefrin- norepinefrin) gereksinim vardır

H A Y V A N F İ Z Y O L O J İ S İ
176
ŞEKİL 2.84 Düz kas
Düz kasta parasempatik sisteme ait reseptörler bulunur. Düz kas kasılmak için ekstraselüler kalsiyuma bağımlıdır. Hücre içine Ca 2+ girişi kendi başına depolarizasyonu arttırıcı bir faktör olarak görev yapar. Bazı hücrelerde bulunan Ca 2+ duyarlı K+ kanalları ise Ca 2+ düzeyinin artışı ile depolarizasyonu azaltıcı yönde etkili olur. Bunun dışında Ca2+ , farklı hücrelerde kendi kanallını bloke etme, defosforilasyon, ikinci habercilerin uyarılması gibi süreçleri başlatabilir. Düz kasta T tübülleri yoktur. Kaveola denen sarkolemmanın hücre içine yaptığı invaginasyonlar triad’ın görevini görür. Düz kasta troponin de yoktur ve kalsiyum kalmoduline bağlanır. Ca-kalmodulin kompleksi de inaktif durumdaki miyozin hafif zincir kinazı aktive eder. Böylece miyozin ve aktin arasında çapraz bağlar kurulur. Kasılma dens bodylere iletilir. Dens bodyler iskelet kasındaki Z çizgisinin görevini yaparlar. Düz kasta aktin ve miyozin demetleri birbirine paralel değildir. Bu nedenle kasılma olduğunda düz kas boğum boğum kasılır. Bazı durumlarda bu çapraz bağlar açılmaz, buna letch fenomeni (mandal yapısı-kilitlenmiş köprü) denir. Bu olay düz kas kasılmasında ATP tasarrufu sağlamaktadır. Düz kasın gevşemesi için miyozinden fosfatın ayrılması gerekir. Bu ayırma işini miyozin fosfataz enzimi yapar. Düz kaslar ile ilgili olarak diğer genel özellikler ise şunlardır:
ŞEKİL 2.85 Düz kasın kalsiyum bağlı olan kasılma mekanizması
Düz kasta kasılma Bağ doku olarak epimisyum, perimisyum, endomisyum bulunur. Otonom sistem sinir hücreleri tarafından düz kas liflerinin çok yakınına salgılanan nörotransmitterler difüzyon ile geçer. Aktin filamentlerine göre, yaklaşık 10-15 kat daha az sayıda miyozin filamentleri bulunur. Çok sayıda aktin filamenti yoğun cisimciklere tutunmaktadır.

H A Y V A N F İ Z Y O L O J İ S İ
177
ŞEKİL 2.86 Düz kasın uyarılması ve kasılması
Düz kas işlevini kontrol eden ve birbiriyle yarışan sinyaller oldukça yoğundur (yüzlercedir). Bu sinyallerin bazıları kasılmayı güçlendirirken bazıları gevşemeye yol açar, fakat sonuçta bunların etkisi diğer kas tipindeki gibi Ca+2 üzerinde yoğunlaşır. Çizgili kas ve düz kas arasındaki temel fark, Ca+2 duyarlı basamağın troponin ve tropomiyozin aracılığıyla ince filamentten kalın filamente miyozin fosforilasyonu yoluyla taşınmasıdır. Kasılma sarkoplazmik Ca+2 konsantrasyonu arttığı zaman başlar. Düz kasta bu durum (Ca+2 artışı) üç potansiyel mekanizma ile gerçekleşir. Ca+2’un sarkolemmadan girişi, SR’dan CIRC ve SR’dan IP3 aracılığıyla serbestlenmesi ile gerçekleşir. Bu yolların hücre içi Ca+2 konsantrasyonu artışına katkı oranları düz kas tipine göre değişmektedir.
ŞEKİL 2.87 Düz kasın kasılmasındaki ikincil haberci molekülleri ve fonkisyonları

H A Y V A N F İ Z Y O L O J İ S İ
178
Düz kas kasılması sırasında sarkomer içine Ca2+ girince kalmoduline bağlanır. Kalsiyuma bağlana kalmodulin inaktif durumdaki myozin hafif zincir kinazı aktive eder. Myozin fosforlanır. Myozinin fosforilasyonu ile myozin ve aktin arasında çapraz bağlar kurulur. Kasılma dens bodylere iletilir. Bu da kasın boğum boğum kasılmasına neden olur. Bazı durumlarda bu çapraz bağlar açılmaz buna letch fenomeni (mandal yapısı-kilitlenmiş köprü) denir. Myozinden fosfatı fosfatazlar ayırarak relaksasyona neden olur. Düz kaslarda mitokondri sayısı azdır. Metabolik ihtiyaçlarını glikolizle karşılarlar. Düz kaslar kendileri aksiyon potansiyeli geliştirebilirler. Düz kaslar östrojen, progesteron gibi hormonlarlada uyarılabilir. Düz kaslarda plastisite özelliği vardır. Mesane düz kasları gibi. Düz kaslarda motor birim yoktur, aksiyon potansiyeli kalsiyumla oluşur.
• Düz kaslar östrojen, progesteron gibi hormonlarla da uyarılabilir.
• Düz kaslarda plastisite özelliği vardır. Lümenli organlardaki düz kaslar
lümen içindeki hacim arttığında buna adaptasyon için gevşeyebilirler. Buna plastisite denir. Mesaneye idrar dolması, midenin gıda ile dolması
plastisiteye örnektir.
• Düz kaslar hiperplazi ve hipertrofiye uğrayabilirler.

H A Y V A N F İ Z Y O L O J İ S İ
179
Miyozin fosfataz miyozinlerden fosfatı koparır. Kas gevşer. Düz kas üzerindeki en önemli parakrin hormonlardan biri NO(nitrit oksit)tir.
Kalsiyumu Bağlıyan Protein -Kalmodulin
Kalsiyumu bağlıyan proteinlerden bir kısmı, ikincil haberci Ca2+ icin intraselluler reseptör olarak hizmet eder. İlk kesfedilen Ca2+ reseptördür. Bu protein sadece çizgili kasta bulunan troponin C’ dir. Kalsiyum bağlayan diğer protein ise kalmodulindir. Bu protein normalde inaktiftir. Fakat Ca2+ ‘u bağladığı zaman aktif bir kompleks olur. Ca2+ -kalmodulin kompleksinin yalnız başına bulunan Ca2+’a göre çok sayıda protein ve enzimin aktivitesini etkilemekten sorumlu, düzenleyici bir ajan olduğu görünür. Kalmodulin omurgalıların düz kasında kasılmayı regüle edici bir kalsiyum reseptörudur. Kamodulin glikojen ve lipid parçalanmasında, hücre bölünmesinde ve sinir uyarılarına bağlı kimyasal ileticilerin serbest hale gelmesinde görev alır. Kalmodulin tıpkı omurgalıların çizgili kasında troponin C’nin etkisi gibi. Ca2+, ekstraselluler ortamda ilk, intraselluler ortamda ise ikinci haberci gibi davranır. Ca2+’un, kas kasılmasında, siliar aktivitenin olusmasında ve sinaptik salgının tetiklenmesinde, genel olarak ekzositoz olayında rolu vardır. Ayrıca oksidativ fosforilasyon, mikrotübül polimerizasyonu, amiboid hareket ve DNA replikasyonu gibi olaylar intraselluler Ca2+ konsantrasyonunun yukselmesiyle tetiklenir veya duzenlenir. Hücredeki kalsiyumun yukselmesi icin iki kaynak vardır. Bazı hücrelerde, kas lifleri gibi, Ca2+ uyarıya cevap sırasında intraselluler kaynaklardan salınır.
• L-arjinin --> L-sitrülin + NO , • NO solübl guanilat siklazı aktifler. • Arteriyolleri dilate eder.

H A Y V A N F İ Z Y O L O J İ S İ
180
Diğer bir sekilde uyarı Ca2+ icin ozel kanalların acılmasına sebep olur, iyonlar ekstraselluler ortamdan hücre icine girerler. Orneğin bu akıs memelilerin karaciğerinde ve tukruk bezlerinde epinefrinin α -adrenerjik reseptörleri aktive ettiği zaman meydana gelir . Kalmodulin cAMP’nin hem meydana gelmesini hem de ortadan kalkmasını etkiler. Aynı sekilde Ca2+ konsantrasyonunun hücre icinde artmasıyla Ca2+ mesajını gerekli yerlere iletir .Ayrıca Ca2+ ‘un hücre dısına pompalanmasını aktive ederek hücre içi konsantrasyonu düşürür ve sinyali sona erdirir. cAMP ve Ca2+ haberci sistemlerinin birbiri ile iliskilerini söyle özetliyebiliriz;
Düz Kas Tipleri
Viseral Düz Kas (Tek Birimli Düz Kas)
Düz kaslar siniri olmadan da çalışır. Barsakların kas tabakasını oluşturan düz kaslar ile Uterus gastrointestinal kanal ve üriner sistemdeki düz kaslar visseral tiptedir. Bu düz kas tipinde daha çok hücrelerarası bağlantılarla kasılmalarını gerçekleştirirler Gerilmeye karşı visseral düz kasın cevabı kontraksiyondur. Soğuk da düz kas kontraksiyonu artırır. Multiunit Düz Kas
Ayrı ayrı düz kas liflerinden oluşurlar. Her bir lif (genellikle) iskelet kasında olduğu gibi tek bir sinir sonlanması ile innerve edilir. Silyer kas (gözde), iristeki kas lifleri, piloerektör
1- Hücreyi dıstan etkiliyen bir haberci iki ayrı reseptoru etkileyebilir ve hucrede hem Ca2+ hem de cAMP yukselebilir. Burada koordine bir kontrol sistemi soz konusudur . 2- Bir haberci hücrede önce Ca2+ haberci sistemini aktive eder, sonra başka bir haberci cAMP haberci sistemini aktive ederek ilk habercinin olusturduğu cevabı kuvvetlendirebilir. 3- Hucre icinde Ca2+ miktarının artması bir hucresel reaksiyon doğurur; cAMP artması da meydana gelebilir ve Ca2+ ‘un meydana getirdiği reaksiyonu durdurabilir. Antagonistik bir etki mekanizması vardır. 4- Bazen hucre ici Ca2+ miktarının artması cAMP artmasına, cAMP artması da hucre ici Ca2+ miktarının artmasına neden olur.

H A Y V A N F İ Z Y O L O J İ S İ
181
kaslar, büyük bronşlar ve büyük arterlerde bulunurlar. İskelet kasındaki gibi nöromuskuler kavşak bulunmaz. Otonom sistem sinir hücreleri tarafından düz kas liflerinin çok yakınına salgılanan nörotransmitterler difüzyon ile membranı geçer. Bu düz kas hücreleri arasında özel bağlantı bölgeleri yoktur ve kasılmaları için sinirsel uyarı şarttır.

H A Y V A N F İ Z Y O L O J İ S İ
182
Duyular ve Reseptörler
Duyular(görme, duyma, dokunma, duyma, ağrı vb) İç ve dış çevre ile ilgili tüm bilgiler santral sinir sistemine çok çeşitli duysal reseptörler aracılığıyla ulaştırılırlar.
Reseptörler Reseptör terimi sadece duysal reseptörler için değil, aynı zamanda spesifik bir cevabı başlatmak üzere nörotransmitter, hormon ve diğer maddelere yüksek afinite ile bağlanan proteinleri tanımlamak için de kullanılır. Duysal reseptör nöronlarda aksiyon potansiyeli yaratan bir nöronun veya özelleşmiş bir hücrenin parçası olabilir.
Bölüm
3 Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
183
Bir reseptör sıklıkla çevresindeki nöral yapıda olmayan hücrelerle birlikte bir duyu organını oluşturur. Sinir sistemi çevrenin, mekanik, termal, kimyasal ve elektriksel özelliklerini algılar ve vücut parçalarının konumsal bilgi göre aksiyon potansiyeli oluşturur. Reseptörler farklı şekilde sınıflanabilir.
Reseptörler uyaran konuma göre
• Exteroceptors
• Enteroceptors = Visceroceptors • Proprioseptörlere
Uyarılma türüne(uyarılma enerjisinine) Göre Reseptörler
• Mekanoreseptörler
• Thermoreceptors • Kemoreseptörler
• Fotoreseptörler
• Nosiseptörleri (ağrı) Her reseptörün bir UYGUN UYARAN’ı Her duyusal sinyalin bir İŞARETLENMİŞ YOL’u vardır. Uyaran >>> reseptör potansiyeli oluşumu >>> aksiyon potansiyeli oluşumu >>> sinyalin (A.P’nin) merkeze taşınması >>> taşınırken bir yandan da modifikasyon >>> sinyal elenmeden kortekse bilince ulaşırsa >>> sensation (DUYUM); icabında reception (ALGI) Uyaran ve oluşan reseptör potansiyeli belirli bir değerin “eşik değer-şiddet” üzerinde olmalı ki sinyal oluşsun merkeze doğru taşınsın. Her genel reseptörde iki özel bölüm vardır:
• Transduser bölge: reseptör potansiyeli oluşum bölgesi • Spike bölgesi: Aksiyon potansiyeli oluşum bölgesi [Ve sonra da AP iletim bölgesi].
Özel reseptörlerde spike bölgesi yoktur. Bu yüzden özel reseptörlerde reseptör potansiyeli oluşur, aksiyon potansiyeli oluşmaz. Duyu organ reseptörleri belli bir enerji formuna karşı, bu enerji formuna yanıt veren diğer reseptörlerden daha düşük eşikte yanıt verecek şekilde uyum sağlamıştır. Bir reseptörün en fazla spesifik olduğu enerji şekline uygun uyarı adı verilir. Örneğin gözde koni ve basiller için uygun uyarı ışıktır. Reseptörler kendileri için uygun olan uyarının yanısıra diğer enerji şekillerine de yanıt verebilirler; ancak bu nonspesifik yanıtların eşiği daha yüksektir. Örneğin, göze uygulanan basınç koni ve basilleri uyarır ancak bu reseptörlerin basınca eşikleri derideki basınç reseptörlerinin eşik değerlerinden daha yüksektir. İç organlarda yer alan bazı reseptörlerin aldıkları duyular idrak edilemez. Sinus caroticus ve arcus aorticus taki baroreseptörlerin ve kemoreseptörlerin, akciğerdeki ve kaslardaki gerilme reseptörlerinin duyuları idrak edilemez. Bazı reseptörler afferent sinirlerin perifer uçlarıdır (ör: derideki reseptörler). Bazı reseptörler ise sinirsel olmayan özel reseptörlerdir ve afferent sinirin perifer ucu bu özel reseptör hücre ile sinaptik bağlantı halindedir (görme, işitme, tat, denge). Buna göre reseptörler duyarlı oldukları (hassas oldukları) enerji formundaki değişiklikleri sinir impulsu haline çeviren yapılardır.

H A Y V A N F İ Z Y O L O J İ S İ
184
ŞEKİL 3.1 Reseptörler tiplerinin bazıları.
İç ve dış çevre ile ilgili tüm bilgiler santral sinir sistemine çok çeşitli duysal reseptörler aracılığıyla ulaştırılırlar. Bu reseptörler çevrede çeşitli şekillerde bulunan enerjiyi, nöronlarda aksiyon potansiyeli sekline dönüştüren çevirenlerdir. Duyu Organları ve Reseptörler
Duyu organ reseptörleri belli bir enerji formuna karşı, bu enerji formuna yanıt veren diğer reseptörlerden daha düşük esikte yanıt verecek şekilde uyum sağlamıştır. Bir reseptörün en fazla spesifik olduğu enerji sekline uygun uyarı adı verilir Örneğin gözde koni ve basiller için uygun uyarı ışıktır. Reseptörler kendileri için uygun olan uyarının yansıra diğer enerji şekillerine de yanıt verebilirler; ancak bu nonspesifik yanıtların esiği daha yüksektir. Örneğin, göze uygulanan basınç koni ve basilleri uyarır ancak bu reseptörlerin basınca esikleri derideki basınç reseptörlerinin esik degerlerinden daha yüksektir Duyu Tipleri
Bilinçli olarak algılanabilen en az 11 tip duyu yer almaktadır. Buna ilaveten, bilinçli olarak algılamamızın dısında bilgiler gönderen duysal reseptörler de vardır. Örnegin kas iğcikleri kas uzunlugu hakkında bilgi verirken bazı reseptörler arteriyel kan basıncı, BOS'un pH' ı gibi degiskenler verirler. Bu duyular bilinçli olarak algılanmazlar buna karşın canlının homoztasinde önemli rol oynarlar. Duyuları sınıflandırma için kullanılan sınırlar tümüyle keyfidir ve genellikle hakkında bilgi oldukça değişken olabilmektedir. Örnegin koni ve basiller degisik dalga boylarındaki ısık uyaranlara maksimum yanıt verirler ve 3 ana renk için farklı koniler bulunmaktadır. Tatlı, tuzlu, eksi ve acı olmak üzere baslıca dört farklı tip tad duyusu vardır ve her biri farklı tip tad tomurcugu tarafından algılanır. Corti organındaki farklı grup tüy hücrelerinin uyarılması sayesinde farklı frekanslardaki ses dalgalarının algılanması mümkün olabilmektedir.
Fazik-Tonik Reseptör aynı hızda adapte olamaz. Bu nedenle, adaptasyon hızına göre reseptörler 2 gruba ayrılırlar Çabuk adapte olan (Fazik) reseptörler: Mekanoreseptörlerin bir kısmı. Böyle reseptörler sadece uyaran şiddeti değiştiğinde uyarılırlar bu yüzden sürekli sinyal yayamazlar. Yavaş adapte olan (Tonik) reseptörler: Fotoreseptörler, kemoreseptörler ve termoreseptörlerin genelikle yavaş adapte olduğu bilinmektedir. Uyaran bulunduğu sürece impulsları beyne göndermeye devam eder. Uyarıyı saatlerce iletebilirler.

H A Y V A N F İ Z Y O L O J İ S İ
185
ŞEKİL 3.2 Reseptörlere göre farklı tipteki iyon kanalları
Duyu Organı Sınıflaması
• Klasik olarak, özel duyular; – tad, koku, görme, işitme, dairesel ve çizgisel hızlanma'dır. Dokunma ile ilgili duyular deride bulunan reseptörler sayesinde algılanırlar • Viseral duyular ise, iç çevrenin algılanması ile ilgili olanlardır. Ağrı duyusu genellikle viseral duyu olarak sınıflandırılır. Reseptörlerin diger bir sınıflamaya göre ayrılması ise şöyledir:
(1) Teleseptörler (uzaklık algılayanlar) uzak mesafedeki olaylar ile ilgilidirler;
(2) Eksteroseptörler yakındaki dıs çevre ile ilgilidirler;
(3) Interoseptörler iç çevre ile ilgilidirler;
(4) Propriyoseptörler herhangi bir anda vücudun konumu ile ilgili bilgi verirler
Kemo reseptörler: Mekanik enerji bağlı olarak hücre zarındaki kanal proteinin konformasyon değiştirmesine bağlı olarak iyon geçirir. Mekonreseptörler ise hücre zarındaki gerilme enerjisini kanal proteini geçirgen konformasyona dönüşmesi için kullanılır. Kanal proteinlerinin aktif olması için tek foton bile yeterli olabilir. Bu etkiler ortak noktası normal durumda kapalı(yada açık ) olan kanal proteinin yapısal olarak değişmesi ve buna olarak geçirgenliğinin değişmesidir.

H A Y V A N F İ Z Y O L O J İ S İ
186
Duysal Sistemin Genel Özellikleri
Duyu Organı Sınıflaması • Nosiseptörler ağrı reseptörleridir
ŞEKİL 3.3 Serbest sinir uçaları
• Fotoreseptörler ısığa hassasiyet gösterirler. Işık enerjisine karşı hassatırlar. • Thermoreseptörler sıcaklık duyusu algılarlar. Soğuk ve sıcak algılayan iki tipi vardır • Mekanoreseptörler dokunma uyaranlarına cevap veren reseptörlerdir. Canlının farklı bölgelerine dağılmış farklı sayıda dokunma reseptörü bulunur. • Kemoreseptör içinde bulundukları ortamın kimyasal içeriğindeki değişimler ile uyarılan reseptörlerdir:
– Bunlar arasında tad ve koku reseptörleri, – Plazma O2 düzeyi, – pH ve – Osmolalite ile ilgili reseptörler
• Baroreseptörler basınca duyarlı reseptörlerdir (aortik ve karotik baroreseptörler.). Kan basınçı hakkında bilgiyi hipotalamusa aktarırlar. Yüzeyel Duyu Organları • 4 farklı tip yüzeyel duyu meycuttur: – Dokunma-bası, Soğuk, Sıcak ve Ağrı' dır • Deri, değişik tip duysal uç içerir, bunlar: – Çıplak sinir uçları, – Genis sinir uçları _ Merkel diskleri ve Ruffini sonlanmaları
ŞEKİL 3.4 Deride bulunan bazı reseptörler.
Serbest sinir uçları: Acı , ağrı reseptörleridir. Kıl kökü reseptör:kıl temas ve hareketi duyularını alır. Ruffin organı: Isı farkını ayırt eder. Kraus bulbusu soğukluğu ayırd eder. Merkel diskleri: sinir uçlarının üzerinde diskler ve diskler üzerinde tek sıra epidermis hücreleri bulunur (deri temas reseptörleri) Meissner korpüskülü deri papillaları içinde bulunur. Temas ve basınç algılar.

H A Y V A N F İ Z Y O L O J İ S İ
187
Kapsüllü sinir uçları _ Pacini cisimcigi, Meissner cisimcigi ve Krause soğanı Pacini korpüskülü: birçok lamelların birbiri üzerine konsantrik olarak sarılmasıyla şekillenmiştir. Duyu sinirinin miyelinli son ucu korpüskülün içine girer, son ranvier boğum korpüskül içindedir. Sinir aksonu düz bir şekilde korpüskülün ortasında yer alır. Korpüskülün mekaniksel olarak uyarılması korpüskülün ortasındaki miyelinsiz akson kısmında potansiyel değişikliği yaratır. Bu’’ impuls doğurucu potansiyel’’ yeteri kadar yüksek ise aksonun ilk ranvier boğumunda aksiyon potansiyeli yaratıyor. • Buna ilaveten duysal sinirler kıl folliküllerinin etrafında da bulunurlar. Yüzeyel Duyu Organları: Genis sinir sonlanmaları ve kapsüllü uçlar, dokunma uyaranlarına cevap veren mekanoreseptörler gibi fonksiyon görürler
• Meissner ve pacini cisimcikleri _ Hızlı Fazik, • Merkel diskleri ve Ruffini sonlanmaları _ Yavaş(tonik) adaptasyon gösterirler • Kıl folliküllerinin etrafında bulunan sinir uçları dokunmaya aracılık ederler ve kılların hareketleri dokunma duyusunu başlatır.

H A Y V A N F İ Z Y O L O J İ S İ
188
ŞEKİL 3.5 Duyuların beyin özerindeki alanları.

H A Y V A N F İ Z Y O L O J İ S İ
189
ŞEKİL 3.6 Derideki reseptörler
Dokunma ile ilgili reseptörler histolojik olarak özelleşme göstermemelerine rağmen fizyolojik olarak özelleşmiştirler. Bu nedenle, her bir sinir sonlanması sadece tek tip deri duyusu ile ilgili uyarıya neden olur. Her duyu organı belli bir enerji formunu duysal sinirler üzerinde aksiyon potansiyeller haline çevirecek şekilde özelleşmiştir. Her modalite beyne ayrı bir yolla iletilmekte ve alınan duyu ile bu duyunun ait olduğu vücut bölgesi beynin aktive edilen 131 özel bölgesi tarafından belirlenmektedir. Belli bir duyunun şiddetleri arasındaki ayırım ise duyu sinirlerindeki aksiyon potansiyel frekansı ile aktive edilen reseptör sayılarının değiştirilmesi gibi 2 şekilde ifade edilir. Bir duyu organının uyarılma şiddetindeki bir artış üretilen duyunun niteliğine pek az (veya hiç) etki yapar. Deri duyularına uygulanan bir diğer ilke noktasal temsildir. Deri, bir kılla uyarılırsa sadece dokunma reseptörlerinin üzerini örten deri noktalarında dokunma duyusu uyandırdığı, bu noktalar arasında kalan bölgelerin ise uyandırmadığı görülür. Aynı şekilde ağrı ve sıcaklık duyuları da sadece bu modalitelere ait duyu organlarının üzerinde yer alan derinin uyarılması ile uyandırılabilir Deride sıcaklık algılanması
Vücutta iki tür sıcaklık algılama organı bulunmakta olup bunlardan ilki vücut sıcaklığının üstünde olana ençok yanıt verenler, ikincisi vücut sıcaklığının altında olan sıcaklıklara ençok yanıt verenlerdir. Bunlardan ilkine sıcak, ikincisine ise soğuk duyu organları adı verilir. Soğuk noktaların sayısı sıcak noktaların 4-10 katıdır. Soğuk reseptörleri 10-38°C sıcaklıklara yanıt verirken sıcak reseptörleri 30-45°C sıcaklığa yanıt verir. Soğuğa ait afferentler Aδ ve C lifleri iken sıcağa ait afferentler C lifleridir. Bu afferentler bilgiyi lateral spinotalamik traktus ve talamik projeksiyonlarla postsentral girusa gönderir. Duyu organlarının subepiteliyal konumu nedeniyle yanıtları belirleyen şey deri altı dokuların sıcaklığıdır. Metal cisimler metalin deri sıcaklığını daha iyi iletmesi nedeniyle aynı sıcaklıktaki tahtadan daha soğuk olarak algılanır. 20°C altı ile 40°C üstü deri sıcaklıklarında herhangi bir uyumun olmaması fakat 20-40°C arasında uyum sağlanması nedeniyle bir sıcaklık değişikliğinin meydana getirdiği duyu giderek sönerek bir termal nötralliğe ulaşır. 45 ° üzerinde sıcaklıklarda doku harabiyeti görülmeye başlar ve alınan duyu ağrı duyusuna dönüşür.
Derideki reseptörler Mekanik reseptörlerin en önemlisi basınç duyusunu alan Pacini cisimciğidir. Pacini cisimciği deri altına ve iç organların duvarlarına yerleşmiştir. Basınç değişmelerini algılamamızı sağlar. Dokunma duyusunu alan reseptörler Meissner cisimciği ve Ruffini cisimciğidir. Bunlar parmak uçları ve dudaklarda yoğun olarak bulunur, cismin niteliğini algılamamızı sağlar. Deride dermis tabakasında bulunan Ruffini cisimciği ise sıcak duyusunu almamızı sağlar. Bu reseptörler çabuk yorulur. Deride bulunan kıl kökü reseptörleri de bir çeşit dokunma reseptörleridir. Hafif bir dokunma duyusunun kuvvetlendirilmesini sağlar. Serbest sinir uçları, en az özelleşmiş resepseptörlerdir. Derinin her tarafında ve diğer dokularda da bulunur ve ağrı duyusunu alırlar.

H A Y V A N F İ Z Y O L O J İ S İ
190
İki-Nokta Ayırımı
İki dokunma uyarısının birbirlerinden ayrı olarak algılanabilmeleri için aralarında bulunması gereken en küçük mesafeye 2-nokta eşiği adı verilir. Bunun büyüklüğü vücutta bölgeden bölgeye değişir ve en küçük olduğu yer dokunma reseptörlerinin en bol bulunduğu yerdir. Örneğin sırtta iki noktanın birbirinden ayrı olarak algılanabilmesi için gereken mesafe 65 mm iken parmaklarda birbirlerinden 3 mm kadar uzak olan iki nokta ayrı ayrı algılanabilir. Öte yandan 2-nokta eşiğinin büyüklüğü yaklaşık olarak tek bir duysal birimi besleyen deri alanının çapına eşittir. Reseptörlerdeki Elektriksel ve İyonik Olaylar
Görme, isitme, denge ve tad gibi karmasık duyu organlarında ayrı ayrı reseptör hücreler ve reseptörler ile afferent sinirler arasında sinaptik baglantılar bulunur. Ancak, dokunma ile ilgili pek çok duyu organında reseptörler özellesmis ve histolojik olarak degisime ugramıs duysal sinir lifi uçlarıdır. Dokunma reseptörü olan pacini cisimcigi ile ilgili, ayrıntılı çalısmalar yapılmıstır. Nisbeten büyük olmaları ve deney hayvanlarının mezanterinde bol miktarda bulunmaları nedeniyle izole edilerek mikroelektrodlarla çalısılabilmektedir. Her kapsül bir duysal sinir lifinin 2 µm çapında düz ve miyelinsiz ucundan yapılmıs olup bu uç bag dokusundan yapılmıs konsantrik lamellerle sarılmıstır ve bu yapı organa sogan görünümü verir. Duysal sinirin miyelin kılıfı cisimcigin içinde baslar. Ilk Ranvier bogumu da kapsül içinde yer alırken ikinci bogum genellikle sinir lifinin cisimcikten ayrıldıgı noktada yer alır. Jeneratör Potansiyeller
Pacini cisimcigine ufak bir basınç uygulandıgında, EPSP'ye benzeyen ilerlemeyen bir depolarizan potansiyel kaydedilir. Buna jeneratör potansiyel veya reseptör potansiyeli denir. Jeneratör potansiyelinin büyüklügü yaklasık 10 mV oldugunda duysal sinirde bir aksiyon potansiyeli doğar. Basınç daha fazla arttırıldığında jeneratör potansiyeli daha da büyür ve duysal sinir tekrar tekrar uyarı çıkarır. Araştırmalar, jeneratör potansiyelinin miyelinsiz sinir ucunda dogdugunu ortaya koymaktadır. Merkel disklerinde de jeneratör potansiyellerin baslangıç yeri sinir terminalleridir. Böylece reseptör mekanik enerjiyi elektriksel yanıta çevirmekte ve bu yanıtın büyüklügü uyarının siddeti ile orantılı olmaktadır. Daha sonra jeneratör potansiyel duysal siniri ilk Ranvier bogumunda depolarize etmektedir. Ateşlemenin belirli bir düzeyinde aksiyon potansiyeli olusmakta ve zar repolarize olmaktadır. Bunun sonucunda Ranvier boğumu reseptörün dereceli yanıtını aksiyon potansiyellerine dönüstürür. Fotoreseptörler
Işıktan etkilenen duyu alıcılarına fotoreseptör adı verilir. Bu tip alıcılarda genel olarak ışık kabul etme yeteneği vardır. Bunlar primer duyu hücreleridir. Fotoreseptörler ya çeşitli aydınlık derecelerini, ya çeşitli uzunluktaki ışık dalgalarını ayırt eder yani renkleri seçer, ya da her iki işi birlikte gerçekleştirirler. Çeşitli hayvan gruplarının görebildikleri dalga uzunlukları birbirinden farklıdır. Bazı hayvansal organizmalarda fotoreseptör bulunmadığı halde ışığa karşı duyarlılığa rastlanabilmektedir. otoreseptörler yapısal farklılıklarına karşılık temelde fonksiyon bakımından büyük benzerlik gösterirler. Bir hücreli hayvansal organizmaların çoğunda ışığa duyarlı bir yapı bulunmadığı halde, ışık değişmelerine yanıt verirler. Topraksolucanlarında da fotoreseptör bulunmaz, fakat tüm vücut ışığa duyarlılık gösterir. Hayvanlar aleminde lokalize olmuş fotoreseptörler çok çeşitlilik gösterir. Bunlar bazı kamçılı bir hücrelillerde stigma ve basit gözler şeklinde
Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
191
görülür. Eklembacaklılarda (örneğin böceklerde) bileşik göz olarak isimlendirilen gözler bulunur. Bu gözler basit gözlerin bir araya gelmesiyle şekillenmişlerdir ve bir tek görüntü oluşturacak şekilde yapılmışlardır. Morfolojik ve optik bakımından omurgalı gözlerinden tamamen farklıdırlar. Odak noktasına getirme mekanizması farklı optik prensiplere dayanır ve odak noktası daima sabittir. Çokhücreli omurgasız hayvanların birçoğunda basit gözler vardır. Ancak bunlardan Mürekkepbalıkları omurgalı gözleriyle karşılaştırılabilecek gözlere sahiptir. Göz kapağı ile kapatılabilen kornealı bir yüzey, gözü odalara ayıran bir mercek ve iris bunların göz yapısında yer almaktadır. Bu göz tipi, yapı bakımından omurgalı gözüne benzerliğinin yanında fonksiyon bakımından da benzemektedir. Omurgalı Göz Yapısı
Küresel bir yapı gösteren omurgalı gözü hayvan türüne göre çeşitli büyüklükte olabilmektedir. Örneğin insanda çapı 2.5 cm. kadardır. Kafatası kemiklerinden oluşan göz boşluğu içinde yer alır. Kuvvetli kaslarla bu yuvarlak içine yerleşmiş göz, göz kapakları ve göz yaşı bezi gibi yapılarla desteklenmiştir. Göz yuvarlağı üç temel tabakadan oluşur. Gözün en dış tabakası sklera (sert tabaka) dır. Bu tabaka ön kısımda bir çıkıntı oluşturarak korneayı meydana getirir. Skleradan sonra ortada koroid (damar tabaka) tabakası yer alır. Bu tabakada pigment maddeleri ile bol miktarda kan damarları bulunur. Koroid de gözün ön kısmındairis’i meydana getirir. Gözün en iç tabakası iki katlı olan retina (ağtabaka)dır.
ŞEKİL 3.7 Gözün yapısı
Bu katlardan dış çeper pigment epitelini, iç çeper ise görme epitelini oluşturur. Görme siniri, gözün üç tabakasını da delerek göze girer ve uçları retinanın iç çeperine yayılır. Görme sinirinin retinayı deldiği yerde görme hücreleri bulunmaz (kör nokta). Kör noktanın yanında sarıbenek adı verilen ve retinanın inceldiği bir bölge bulunur. Sarıbeneğin ortasında görme hücreleri sayısı artar. Bu nedenle görme olayının en net olduğu kısım burasıdır.

H A Y V A N F İ Z Y O L O J İ S İ
192
Görme Olayı
Göz hem ışık reseptörü, hem de uzaklık reseptörüdür. Değişik hayvan grupları değişik dalga boylarındaki ışığa karşı duyarlılık gösterir. Örneğin insanda infrakırmızı ışık enerjisinin düşük olması nedeniyle uyarı gerçekleşmezken, yılanlar infrakırmızı ışığa duyarlı reseptör taşırlar.
Göz algıladığı objeyi kornea (mercek) aracılığı ile ters (başaşağı) olarak retina üzerine ulaştırır. Ayrıca göze gelen ışık şiddeti pupillanın genişleyip daralması, skleranın saydam olmaması ve koroid, retina, iris pigmentlerinin absorbe edici özellikleriyle ayarlanır.
Duyusal retina, dört ana grup hücreden oluşur.
Çubuk ve koni olmak üzere iki tip fotoreseptör vardır. Çubuk hücrelerinin sayısı 110-125 milyon, koni hücrelerinin sayısı ise 6,3-6,6 milyon arasındadır.
ŞEKİL 3.8 Gözdeki retinada yer alan rod ve koni hücreleri
Fotoreseptör hücreleri görünür ışığı dalga boyuna yani rengine uygun olarak elektrik enerjisine çevirir. Bu uyarılar da retinanın en iç tabakasında yer alan ganglion hücreleri uzantıları tarafından oluşturulan optik sinir ile beyindeki görme merkezlerine ulaştırılır.
• Fotoreseptörler
• Bipolar hücreler • Ganglion hücreleri
• Destek hücreleri
• Fotoreseptörler

H A Y V A N F İ Z Y O L O J İ S İ
193
Görmenin Kimyası
Işık, çomakçık ve konilere çarptığında impuls meydana gelir. Her çomakçığın dış yüzeyi ince zarlı uzantılardan oluşmuştur. Bu zarla, kırmızımtırak görme pigmenti ''Rhodopsin'' belirli bir düzen içerisinde birleşmiştir. Diğer görme pigmentleri gibi rhodopsin de iki kısımdan meydana gelmiştir. Bir renkli kısım ''Cis - Retinal'' bir de protein kısmı ''Opsin'' vardır. Işığın absorbe edilmesiyle cisretinal izomerizasyona uğrayarak ''Trans-Retinal''a dönüşür. Transretinal ve opsin, ''Lumirhodopsin'' denen bir bileşik meydana getirirler. Fakat transretinal ve opsin geometrik yapıları dolayısıyla birbirlerine iyi uymazlar. Bu yüzden lumirhodopsin karasız bir maddedir. Dolayısıyla bu madde ''Metarhodopsin''e ve daha sonra da trans-retinal ile opsine parçalanır (renk solar). Bu durumda ışık enerjisine yalnız cis-trans izomerizasyonu için gereksinme vardır. Diğer değişiklikler ışığa gereksinme göstermezler. Renk pigmentlerine belirli bir düzen içerisinde bağlı olan duyu hücrelerinin zarlarının impulsları meydana getirir. Rhodopsin ışığa karşı olağanüstü duyarlıdır. Hesaplara göre tek bir ışık fotonu dahi rhodopsin molekülünü aktive edebilir ve bu da tüm bir çomakçığı harekete geçirir. Ama daha sonra göreceğimiz gibi aktive edilmiş tek bir çomakçık beyne gidecek bir impuls yaratamaz. Rhodopsinin tekrar sentez edilmesi için trans-retinalın, cis-retinala dönüşmesi gerekmektedir. Stereoizomerler moleküldeki atomların uzayda düzenlenişleri farklı olan izomerlerdir. Trans ve cis retinal stereoizomedir.
ŞEKİL 3.9 CIS-Retinal molekülünün ışığın etkisi iel tans-retinal oluşumu
Trans- Cis retinal değişimi işlemi enerjiye gereksinme gösterir. Ayrıca bu tepkimenin yürütülmesi için ''Retinal izomeraz'' denen bir enzim vardır. Cis-retinal, bu enzim aracılığıyla opsin ile kolayca birleşerek tekrar rhodopsini yapar. Bu parçalanma ve yeniden sentezlenme aralıksız sürer. Ama devamlı ışık alan bir gözde bu denge bozulacağından, o anda görmede zayıflamalar olacaktır. Karanlığa ve aydınlığa uyum yapabilmek için biraz zamanın geçmesi işte bu tepkimelerin tamamlanması içindir. Eğer bir görme hücresi devamlı ışık etkisi altında kalırsa, parçalanan rhodopsin, sentez edilenden az olacağından görme duyarlığı azalacaktır. Trans-retinal, retinolun (A vitamininin) bir aldehitidir. A vitamininin alkol dehidrogenaz enzimi ile katalizlenmesiyle meydana gelir. Tekrar sentezlemede bunun A vitaminine yavaş yavaş dönüşmesi söz konusudur. Bu tepkimeler retina tabakasında bulunan ışığa duyarlı hücreler içerisinde oluşur. Eğer göz uzun zaman karanlıkta kalırsa retinalın büyük bir kısmı rhodopsine dönüşeceğinden duyarlılık çok fazla artar (birkaç dakikada bin misline, bir saatte ise yüz bin misline ulaşır). Karanlık bir yerden aydınlığa çıktığımızda göz kamaşmasının nedeni bundandır. Kamaşmanın geçmesi için biraz beklemek yani rhodopsin miktarının azalmasını sağlamak gereklidir. Aydınlıktan karanlığa girdiğimizde de bunun tersi

H A Y V A N F İ Z Y O L O J İ S İ
194
olur. Biraz beklemekle rhodopsin miktarı artar ve bununla ilgili olarak duyarlılık yükselir. Göz saniyenin milyonda biri kadar süren bir ışığı; saniyenin onda biri kadar süren ışıklı bir görüntüyü algılayabilmektedir. Geceleri elektrik ışığını çok kısa bir süre yakıp söndürdükten sonra, yine çok kısa süren bir aydınlığın algılanması, meydana gelen lumirodopsinin parçalanmadan çomaklarda kalabildiği süreye denktir. Aslında retinal görüntünün, gerçek görüntü ortadan kalktıktan sonra da kısa bir süre devam etmesi, sinemadaki ve televizyondaki kesik kesik görüntülerin bir bütün olarak algılanmasını sağlamaktadır. Vitamin A, retinalın ilkel maddesidir. Çeşitli besinlerle vücuda alınır ve karaciğerde depo edilir. Azlığında gece körlükleri ortaya çıkar. Ayrıca sinir dokunun çalışması için gerekli olan nikotinamit, retinanın işlevleri için de gereklidir.
Koniler yalnız kuvvetli ışıklarda aktive edilebilirler ve renkleri görürler. Uzun süreden beri ışık spektrumunda yalnız üç rengin görüldüğü (mavi, yeşil ve kırmızı) kabul edilmektedir.
ŞEKİL 3.10 Farklı reseptörlerin optimum uyarılma dalga boyu
Diğer renkler bunların değişik karışımları ile algılanmaktadır. Çünkü yapılan gözlemlerde, yalnız üç tip renk körlüğü vardı; bu da üç çeşit renk gören koni tipinin olduğunu göstermektedir. Nitekim yeni ölenlerden izole edilen konilerde yalnız bu üç tip renk maddesine maksimum absorbsiyon gösterilmiştir. Konilerin ışığa duyarlı pigmentleri ''lodopsin''dir. Siyah beyaz görmeyi sağlayan (karanlıkta görme) rhodopsin konilerde pek az vardır, lodopsin retinal ve opsinin, rhodopsinden farklı olarak birleşmesiyle oluşmuştur. Üç koni tipinin arasındaki kimyasal farklar bugüne kadar açıklanamamıştır. Fakat omurgalılarda iki retinal ve birçok opsin tipinin olduğu ve bunların çeşitli kombinasyonlarıyla renk algılanmalarının ortaya çıktığı düşünülmektedir. Örneğin tatlı su balıklarının çomak hücrelerinde porfiropsin (bu hayvanlarda A vitaminin varlığını hatırlayınız!), koni hücrelerinde siyanopsin (keza kurbağa yavrularında ve tatlı su kaplumbağalarında da bu madde vardır) bulunur. Kurbağalar karaya geçtiklerinde, taşıdıkları porfiropsin, rhodopsine dönüşür.

H A Y V A N F İ Z Y O L O J İ S İ
195
ŞEKİL 3.11 Rodopsin değişime bağlı hiperpolarizayon oluşumu
Çubuk Hücreleri
İnce uzun hücrelerdir, retinada 100 milyon adet çubuk hücresi bulunur, bunların boyları yaklaşık 100-120 mikrondur. Çubuk hücreleri alaca karanlıkta görmemizi sağlarlar, bu hücreler renklere karşı duyarlı değildir renkleri grinin tonları olarak görmemizi sağlarlar. Dış kısım ışık uyarımının alındığı bölgedir, burada ışığa duyarlı bir pigment olan rodopsin bulunur. Bir çubuk hücresinin dış kısmında yaklaşık 600-1000 adet yatay yerleşimli disk vardır. Rodopsin bu disklerin zarları üzerinde yer alır. Birleştirici bir sap ile hücrenin iç kısmı ile iletişim sağlanır.
ŞEKİL 3.12 Rod reseptörlerinde hiperpolarizayon oluşumu
Retina
Retina ön tarafta hemen hemen siliyer cisimlere kadar yayılır; 10 tabakadan oluşmuştur ve görme reseptörleri olan basil ve koniler ile 4 tip nöron içerir. Bu nöronlar bipolar hücreler, gangliyon hücreleri, horizontal hücreler ve amakrin hücreler'dir . Retinanın reseptör tabakası koroidin karşısında yer aldığından ışık ışınları koni ve basillere ulaşmak için gangliyon hücresi ve bipolar hücre tabakalarmdan geçmek zorundadır.

H A Y V A N F İ Z Y O L O J İ S İ
196
Koroidin retinaya bitişik pigmentli tabakası, ışık ışmlarını, retinaya geri yansımalarını önlemek için absorbe eder. Böyle bir yansıma görsel imajların bulanmasma yol açacaktır. Retinanın nöral elemanları. Optik sinir göz küresinin arka kutbunun hafifçe üzeri ve 3 mm kadar medyalinde yer alan bir noktadan gözü terk ederken retina kan damarları bu noktadan göze girerler.Bu bölge oftalmoskopta optik disk olarak görülür. Disk üzerinde hiçbir görme reseptörü bulunmadığımdan bu nokta kördür (kör nokta) Gözün arka kutbunda sarı pigmentli bir nokta olan makula lutea vardır. Burası, konilerin yoğun şekilde bulunduğu, reseptörler üzerinde kan damarlarının olmadığı ve çok az sayıda hücre bulunan ince, basillerden yoksun retina bölümünü, yani fovea sentralis'i işaret eder. Fovea insanlarda iyi gelişmiştir. Görme keskinliğinin en fazla olduğu nokta burasıdır. Bir cisme dikkatle bakıldığında gözler, bu cisimden gelen ışık ışmlarını fovea üzerine düşürecek şekilde hareket ederler. Retinanın vitröz yüzeyine yakın yüzeyel tabakalarındaki arterler, arterioller ve venler oftalmoskopla görülebilir. Vücutta arterioIlerin gözle kolayca görülebildiği tek yer burası olduğundan, oftalmoskopik muayene diabetes mellitus, hipertansiyon ve kan damarlarını etkileyen diğer hastalıkların tanı ve değerlendirilmesinde büyük önem taşır. Retina damarları bipolar ve gangliyon hücrelerini sular, fakat reseptörler çoğu yerde koroiddeki kapiller pleksus tarafından beslenirler Retina ayrılmasının reseptör hücrelerini için bu derece zararlı olmasının nedeni budur.
Koni Hücreleri
Retina'da yaklaşık 3 milyon koni hücresi bulunur. Bunlar da uzun ince hücrelerdir, 65-75 mikron boyundadırlar. Yapıları hemen hemen çubuk hücrelerine benzer, yalnız dış kısımları koni şeklindedir. Koni hücreleri sarı nokta (fovea) denilen alanda yoğunlaşmıştır gündüz ışığında ve renkli görmemizi sağlarlar. Yüksek görme merkezi olan beyin korteksinin %90 kadar bir kısmı bu sarı noktadan gelen uyarıları işlemekle görevlidir.
Bipolar Hücreler
Bu hücreler, fotoreseptör hücrelerinden gelen uyarıları diğer hücrelere iletir ve sinyal integrasyonunda rol oynarlar. Bu hücrelerin bir alıcı kısmı bir de iletici bir kısmı vardır bu nedenle bipolar(iki kutuplu) olarak adlandırılır.
ŞEKİL 3.13 Rod ve koni hücrelerin retinadaki konumu
Ganglion Hücreleri
Ganglion hücreleri, retinanın iç kısmında yer alır ve gözbebeğinden gözün içine bakıldığında aksonal uzantıları ile oluşturdukları ince çizgiler görülebilir. Retinanın büyük bir kısmında tek bir katman oluştururlar ancak optik sinire yaklaşıldığında bu katmanların sayısı artar. Ganglion

H A Y V A N F İ Z Y O L O J İ S İ
197
hücreleri multipolardır –çok kutupludur-, dendritleri bipolar ve amakrin hücrelerin aksonları ile komşuluk içerisindedir. Ganglion hücrelerinin uzun aksonları vardır, bu aksonlar retina yüzeyine ulaşınca yaklaşık 90 derece açı yaparak optik sinirde toplanır ve gözün içini terk ederler. Ganglion hücreleri, fotoreseptör hücrelerce oluşturulan elektrik sinyallerini beyindeki görme merkezlerine taşırlar. Optik sinir işte bu ganglion hücrelerinin uzantılarının bir araya gelmesinden oluşmuştur. Horizontal hücreler, çubuk ve koni hücrelerinin terminal şişkinliklerine yakın yerleşimlidirler. Multipolar hücrelerdir ve görsel uyarımın integrasyonunda görev alırlar
Basiller ve konilerin her ikisi de bipolar hücrelerle yaptıkları sinapslarda glutamat serbestlerler. Ve, histolojik ve farmakolojik çalışmalar o bölgede tümü normalde inhibitör transmiter olarak işlev gören, gama-amino bütirik asit, glisin, dopamin, asetilkolin ve indolanıin gibi en azından sekiz çeşit transmiter maddesi serbestleyen bir çok tipte amakrin hücrelerin bulunduğunu göstermiştir.
Fotoreseptörler uyarı oluşumu ve bunun işlenmesi
Gözde temel olarak üç resim oluşumu (işlenmesi) olur. Birinci resim reseptör hücrelerince oluşturulur. İkinici resim bipolar hücrelerince, üçünçü resim ise ganglion hücrelerince meydana getirilir. Reseptör hücreleri ile bipolar hücreler arasında amakrin hücreleri, Bipolar hücrelerle gangliomlar arasında ise horizandal hücreler aracılık eder. Retinada aksiyon potansiyellerini başlatan potansiyel değişiklikleri ışığın basil ve konilerdeki ışığa duyarlı bileşiklere etkisi ile oluşturulur. Işık bu maddeler tarafından absorbe edildiğinde, yapıları değişir ve bu değişiklik nöral aktiviteyi başlatan bir dizi olayı tetikler. Göz, fotoreseptörlere ait reseptör potansiyelleri ile retinadaki diğer nöral elemanların çoğuna ait elektriksel yanıtlarının lokal, basamaklı potansiyeller olması ve oldukça uzak mesafelere iletilen hep veya hiç aksiyon potansiyellerinin sadece gangliyon hücrelerinde üretilmesi yönünden benzersiz bir organdır. Basil, koni ve horizontal hücrelerin yanıtları hiperpolarize, bipolar hücrelerin yanıtları ise hem hiperpolarize hem de depolarize edici türde iken amakrin hücreler gangliyon hücreleri tarafından üretilen ilerleyici dikenler için jeneratör potansiyeli görevi yapan depolarize edici potansiyeller ve dikenler üretir. Koni reseptör potansiyeli keskin bir başlama ve bitişe sahipken basil reseptör potansiyeli keskin bir başlama ve yavaş bir sonlanma gösterir. Reseptör potansiyellerinin genliği ile uyarı şiddeti arasındaki ilişkiyi gösteren eğriler koni ve basillerde birbirine benzerse de basillerin duyarlığı çok daha fazladır. Bu nedenle koniler için eşik altı düzeyde olan aydınlatmada basil yanıtları uyarı şiddetiyle orantılıdır. Öte yandan basil yanıtlarının azami olup artık değiştirilemediği aşırı aydınlatma düzeylerinde koni yanıtları uyarı yoğunluğu ile orantılıdır. Bu durum konilerin neden ışık şiddetinin zemin değeri üzerindeki değişikliklere yanıt verip mutlak aydınlatmayı yansıtmazken basillerin neden mutlak aydınlatmayı farkettiğini açıklar. Basil ve konilerin dış segmentindeki Na+ kanalları karanlıkta açık olduğundan iç seğmentten dış segmente akım akışı olur. Akım fotoreseptörlerin sinaptik uçlarına da akar . İç segmenttekil Na+ -K+ pompası iyonik dengeyi sürdürür. Karanlıkta sinaptik transmiter kararlı şekilde salınır. Işık dış segmente çarptığında Na+ kanallarından bazıları kapanır ve olayın sonucu hiperpolarize edici bir reseptör potansiyelidir. Bu hiperpolarizasyon sinaptik transmiter salınmasını azaltır ve bu da en sonunda gangliyon hücrelerinde aksiyon potansiyellerine yol açan bir sinyal üretir. Aksiyon potansiyelleri beyine iletilir. Tekrarlarsak retinada görsel bilgi işlenmesi bir anlamda 3 resmin oluşturulmasını kapsar. Işığın fotoreseptörlerdeki etkisiyle oluşan birinci resim bipolar hücrelerde ikinci resme dönüşür ve bu da daha sonra gangliyon hücresinde üçüncü resme çevrilir. Sinyal, ikinci resmin oluşumunda horizontal hücreler, üçüncünün oluşumunda amakrin hücreler tarafından değiştirilir.

H A Y V A N F İ Z Y O L O J İ S İ
198
Görme sinirleri
Gangliyon hücre aksonları, talamusun bir bölümü olan korpus genikulatum lateralede sonlanmak üzere optik sinir ve optik traktusda kaudal yönde ilerler. Retinanm nazal yarımlarından gelen lifler optik kiazmada çaprazlaşarak karşı tarafa geçer. Korpus genikulatumda, bir retinanm nazal yarımı ile diğer retinanın temporal yarımından gelen lifler, aksonları genikulokalkarin traktusu oluşturan hücrelerle sinaps yapar. Bu traktus serebral korteksin oksipital lobuna gider. Primer vizüel alıcı alan (görme korteksi, Brodmann‘ın 17. alanı) temel olarak fissura kalkarinanın kenarlarına yerleşmiştir. Gangliyon hücre aksonlarının dalları, optik traktustan görme reflekslerine aracılık eden bağlantıları oluşturdukları kollikulus superior ve orta beyinin pretektal bölgesine geçerler. Diğer aksonlar optik kiazmadan, endokrin değişiklikler ve aydınlıkkaranlık döngüsüyle ilgili diğer sirkadiyen ritimleri senkronize eden bağlantıları oluşturdukları hipotalamusun suprakiazmatik çekirdeğine direkt olarak geçerler. Görme uyarıları ile aktive edilen beyin alanları, maymunlarda radyoaktif 2-deoksiglükoz ile araştırılmıştır. Aktivasyon sadece oksipital lobda değil, aynı zamanda inferior temporal korteks, posteroinferior parietal korteks bölümleri ile frontal lobun bazı kısımlarında meydana gelir. Lateral genikülat cisme ek olarak aktive edilen subkortikal yapılar superior kollikulus, pulvinar, kaudat çekirdek, putamen, klaustrum ve amigdaleyi kapsar.
ŞEKİL 3.14 Görme bölgeleri
Gece Körlüğü
Gece körlüğü, ciddi A vitamini eksikliği olan herkeste meydana gelir. Bunun basit nedeni uygun miktarda retinale dönüşecek yeterli A vitamini olmamasıdır. Bu yüzden, yapılabilecek rodopsin miktarı ciddi şekilde azalır. Bu duruma gece körlüğü denir, çünkü gece ışık miktarı yeterli bir görüş için çok azdır, oysa gün ışığında aynı biçimde renk pigmentlerinde de azalma olduğu halde koniler yine de uyarılabilirler. Gece körlüğünün meydana gelebilmesi için, bir kişinin aylarca A vitamininden eksik bir diyetle beslenmesi gereklidir. Çünkü, büyük miktarda A vitamini normal olarak karaciğerde depo edilir ve gözler için kullanılabilir. Gece körlüğü meydana geldiğinde, bazen A vitamininin damar içine verilmesi ile 1 saatten kısa bir sürede geri döndürebilir.

H A Y V A N F İ Z Y O L O J İ S İ
199
Koku
Günlük hayatta sürekli olarak kullanılan bir duyu olan koku önemli bilgiler sağlar. Yiyeceklerin varlığı, alınan zevk ve tehlike hakkında bilgi verir. Yiyeceklerin sindirimi ve kullanımı için gerekecek olan fizyolojik değişilikler sağlar. İnsan genlerinde 1000 kadar koku reseptörü kodludur. İnsanlar, hayvanlardan büyük miktarda koku ayırt etme yetisine sahiptir. Koku moleküllerini ayırt etmede eğitimli olan koku uzmanları, 300–5000 farklı koku ayır edebilir..
ŞEKİL 3.15 Koku alma reseptörlerin işlevi
Koku duyusu komponentleri algı ve tanıma Adaptasyon, Hafıza ve Emosyon oluşur. Burun solunan havayı, Filtre eder, Isıtır ve Nemlendirir. Koku almak için suda ya da yağda eriyebilen bileşiklerin burun mukozasına teması gerekir. Yüksek oranda emilen kimyasallar, düşük koku oluşturma etkinliğine sahip etkin bir koku uyarımı için burun hava akımı gerekmektedir. Koku sirke gibi keskinse, trigeminal sinir uçları ile algılanır ve ağrı cevabına yol açar. Koku sinyallerinin, yemek alımı, ısı regülâsyonu, uyku siklusu, görme, işitme ve tat alma ile ilgili bölgelere iletilir.
ŞEKİL 3.16 Koku reseptörleri
Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
200
Koku algılama hücreleri: Dış dünya ile direkt ilişkidedir, bariyer yoktur Beyinle direkt iletişim kurar, Kokuyu ileten sinirler organizmadaki en ince ve yavaş sinirlerdir, diğer duyulara oranla algı biraz gecikir.
Her sinir hücresi yalnız bir tür koku resptörünü algılar. Koku duyusu sinirleri koku yakalamak için özelleşmiştir. Gerekli molekülleri ve iyonik ortamı sağlar. Çözünebilir, koku bağlayan proteinler bulunur. Molekül yoğunlaştırma, taşıma uzaklaştırma görevini yapar.
ŞEKİL 3.17 Koku alma reseptörlerinin Epitel dokudaki konumu işlevi
Koku duyusunun ilk basamağı burundaki koku duyu sinirlerinde başlar. Bu sinirler burun boşluğunun arkasında 5 cm2 lik bir alanı kaplayan özelleşmiş bir yapıda bulunmaktadır. Koku bilgisi beyindeki koku merkezine -ulaştığında, beyin daha önceki deneyimlerle belirlenmiş olan şifreleri çözerek kokunun tanınmasını sağlar. Beyine ulaşan koku bilgisi duygusal ve dürtüsel merkezlerle de iletişir. Kokunun davranış ve duygular üzerindeki etkileri bu bağlantılarla açıklanabilir. Reseptörün hissizleşmesi: Hoş olmayan bir kokuya sürekli maruz kalınırsa bir süre sonra fark edilmez olur. Temiz havada bir süre kaldıktan sonra aynı koku tekrar ayırt edilebilir. Beyin, insan yaşamında önemli kabul ettiği kokuları unutmaz. Özellikle çocukluk döneminde gençlik dönemlerinde alınan kokular unutulmaz. Kokular anıları canlandırabilir. Bazı türler feromon adı verilen türe özgü kimyasallar salarlar. Feromonlar kendi türlerinden olan diğer bireylerin cinsel ve toplumsal davranışı ile üreme fizyolojisi üzerinde çok önemli rol oynarlar. Feromonlar bazı türlerde (sığır domuz fare..vs). Puberte zamanını ayarlar. Erkek ve dişinin üreme ile ilgili davranışlarını etkiler. Dişinin yavrusuna karşı davranışlarında rol oynar. Kaynağı idrar ve bazı dış salgılardır.
3 çeşit hücre görev alır.
• destek hücreleri • bazal hücreler • koku algı sinirleri

H A Y V A N F İ Z Y O L O J İ S İ
201
Tad duygusu
Degustatörler 100’den fazla tad bileşenini ayırt edebilirler. Tad olarak algıladığımız lezzet çeşitleri, çiğneme ve yutma sırasında burun boşluğun arkasındaki olfaktör sisteme ulaşan uçucu moleküllerle; acı, tatlı, tuzlu, ekşi moleküllerin karmaşık karışımlarından oluşur. Temel olarak Tatlı, tuzlu, ekşi ve acı olmak üzere başlıca dört farklı tip tad duyusu vardır ve her biri farklı tip tad tomurcuğu tarafından algılanır. En arkanın tat duyusunu CN10, 1/3 arkanın CN9, 2/3 önün CN7 alır. Yutağın duyusunu vagus alır. Acı duyusu: K+ kanalları bloklanarak depolarizasyon oluşur. Şekerli maddeler: K+ kanalları kapanarak depolarizasyon oluşur. Ekşi maddeler: K+ kanalları kapanır ve depolarizasyon oluşur.Tuzlu maddeler: Na’un kendisi kanallardan girerek depolarizasyon oluşturur. Umami tat: Monosodyum glutamata bağlıdır. Acı duyusu en arkada, ekşi arka yanlarda, tuzlu ön yanlarda, tatlı önde algılanır.
Dilde bulunan sensörler Papillalar fonksiyon ve yapılarına göre 4 gruba ayrılır: 1. Filiform papilla (tat tomurcuğu içermez. En fazla bulunan papilladır. dokunmayla ilgilidir) 2. Fungiform papilla (tat tomurcukları içerir) 3. Sirkumvallat papilla (tat tomurcukları içerir. En az bulunan papilladır. Tat reseptöründen en zengin olandır) 4. Foliat papilla (insanda rudimenterdir) Bu tomurcuklar Ebner bezlerinin salgısı ile devamlı ıslak tutulurlar. Ebner bezi lingual lipaz salgılar. Fungiform papilla dilin ön kısmında yerleşmiştir. Yoğun tat tomurcuğu içerir. Bol miktarda kanlanması nedeniyle fungiform papilla dilin üzerindeki kırmızı noktalanmayı oluşturur. İşitme Duyusu ve Denge
İşitme ve denge gibi iki duyu modalitesinin reseptörleri kulağa yerleşmiştir. İşitme ve denge reseptörleri tüy hücreleri şeklindedir ve her yarım daire kanalında bir tane ve utrikul, sakkulus ve kohleada birer tane olmak üzere her içkulakta 6 grup tüy hücresi bulunmaktadır Dış ve Orta Kulak:
Dışkulak kepçesi ses dalgalarını dışkulak kanalına iletir. Dışkulak, orta kulak ve içkulağın kohleası işitme ile ilgili iken İçkulaktaki yarım daire kanalları, utrikulusve sakkulus ise denge ile ilgilidir. Yarım daire kanallarındaki reseptörler döngüsel hızlanmayı(rotasyonel akseleasyon) utrikuldaki reseptörler yatay yönde doğrusal hızlanmayı(lineer akselerasyon), sakkulustaki reseptörler dikey yönde hızlanmayı saptarlar. Dışkulak kanalıdışkulak deliğinden başlayarak içerdeki kulak zarı’na (timpan zarı) kadar ilerler. Orta kulak, östaki borusu (odituvar kanal) aracılığıyla nasofarinkse ve nasofarenks aracılığıyla dışa açılan, temporal kemik içinde yer almış içi hava ile dolu bir boşluktur. Östaki borusu genellikle kapalı ise de yutma, çiğneme ve esneme sırasında açılır ve kulak zarının iki tarafındaki basıncı dengede tutar. Orta kulakta çekiç, örs ve özengi gibi 3 tane işitme kemikçiği yerleşmiştir. Manubriyum (çekiş kemiğinin sapı) kulak zarının arkasına bağlanmıştır.
Bu bölüm için Power
point sunumu adresi:
Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
202
ŞEKİL 3.18Timpanel zar
Kulakta bulunan üç kemik duyma işlevi içim önemlidir. Çekicin başı orta kulak duvarına, kısa kolu örse bağlanır ve bu kısa kol daha sonra özenginin başı ile eklem yapar. Özenginin ayak tabanı annular bir bağla oval pencerenin duvarına bağlanmıştır. İki küçük iskelet kası olan tensör timpani ve stapedius da aynı şekilde orta kulağa yerleşmiştir. M. tensor timpani’nin kasılması çekiç kemiğinin uzun kolunu mediale doğru çeker ve kulak zarının titreşimini azaltır. M. stapediusun kasılması ise özenginin taban parçasını oval pencereden geri çeker.
İç Kulak (Tympanic Cavity)
İç kulak biri diğerinin içine yerleşmişiki parçadan oluşmuştur. Kemik labirent, temporal kemiğin petroz kısmında yer alan bir seri kanallar halindedir. Bu kanalların içkısmında bulunan zarsı labirent perilenf adı verilen bir sıvıyla çevrelenmiştir. Bu zarsı çatı aşağıyukarı kemiksi labirentin biçimine benzer. Zarsı labirent endolenf adı verilen bir sıvıyla dolu olup, endolenfa ve perilenfa ile dolu boşluklar arasında iletişim bulunmaz.
Kohlea İnsanda labirentin kohlea kısmı 35 mm boyda ve 2 ¾ kıvrım yapan sarmal bir tüptür. Baziler ve Reissner membranı kohleayı uzunluğu boyunca 3 odaya (skala) ayırır. Üstteki skala vestibulle alttaki skala timpani perilenfa içerir ve bunlar kohleanın apeksinde yer alan ve heliko trema adı verilen küçük bir delikle birbirine bağlanır. Skala vestibuli kohlea tabanında özenginin taban parçası ile kapatılmış olan oval pencerede sonlanır. Skala timpani, orta kulağın medialinde yer alan ve esnek sekonder timpanik membran ile kapatılmış olan yuvarlak pencerede sonlanır. Kohleanın orta odası olan skala media zarsı labirent ile devam eder, diğer iki odayla bağlantısı yoktur ve endolenfa içerir.
Korti organı bazal membran üzerine yerleşmiş ve işitme reseptörleri olan tüy hücrelerini içeren bir yapıdır. Kohlea, apeksten tabana doğru uzanmasının bir sonucu olarak spiral bir şekil gösterir. Tüy hücrelerinin uzantıları, Korti çubukları tarafından desteklenmiş, zara benzer katı bir yapı olan retiküler laminayı delerler. Bu tüy hücreleri 4 sıra halinde dizilmiş olup dıştüy hücrelerinin yaptığı 3 sıra Korti çubukları tarafından oluşturulmuş tünelin lateralinde, içtüy

H A Y V A N F İ Z Y O L O J İ S İ
203
hücreleri tarafından yapılmış bir sıra da bu tünelin medialinde yer alır. Her insan kohleasında 20 000 dış tüy hücresi ile 3500 içtüy hücresi vardır.
Bu tür sıkı kavşakların bulunmasından ötürü tüy hücrelerinin orta kulağın diğer kısımlarındaki düzenleniş biçimi de buna benzer, yani tüy hücrelerinin uzantıları endolenfa içine batmışken bu hücrelerin tabanları perilenfaya dalar. Tüy hücre sıralarıince, visköz fakat esnek bir tektorial zarile örtülü olup sadece dıştüy hücrelerinin tepesi bu zara gömülüdür. Tüy hücrelerinin tabanları çevresinde dallanmış olan afferent nöronların hücre gövdeleri kohleanın etrafına sarıldığı kemiksi koçan olan modiolus içinde yer alan spiral gangliona yerleşmiştir.
ŞEKİL 3.19 farklı seslerin farklı bölgelerde sinirleri uyarması
Bu afferent nöronların %90-95 içtüy hücrelerini innerve ederken sadece %5-10’nu, çok daha fazla sayıda olan dıştüy hücrelerini innerve eder ve her nöron bu dışhücrelerin birkaçını innerve etmektedir. Buna ek olarak işitme siniri içindeki efferent liflerin çoğu dıştüy hücrelerinde sonlanır. Tüy hücrelerini innerve eden nöronların aksonları vestibulo kohlear akustik sinirinodituvar (kohlear) dalını oluşturur ve medulla oblangatadaki dorsalve ventral kohlear çekirdeklerde sonlanır.
Her odituvar sinirdeki afferent ve efferent liflerin toplam sayısı yaklaşık 28 000’dir. Korti Organı Kohlea tüy hücreleri ile komşu falangial hücreler arasında sık ıkavşaklar vardır ve bu sıkı kavşaklar endolenfanın hücrelerin tabanına ulaşmasını önlerler. Bununla beraber baziler membran skala timpanideki perilenfaya görece geçirgendir ve sonuçolarak Korti organ tüneli ile tüy hücrelerinin tabanı perilenfaya dalmış haldedir.
Merkezi İşitme Yolları İşitme impulslarını taşımak üzere kohlear çekirdeklerden çıkan aksonlar çeşitli yollar aracılığıile işitme refleks merkezlerinin bulunduğu inferior kollikuliye ve talamustaki medial genikulat cisimcik yoluyla işitme korteksine ulaşır. Diğer aksonlar retiküler formasyona girerler. Her iki kulaktan gelen bilgi oliva süperiorlarda kavuşum gösterir ve daha

H A Y V A N F İ Z Y O L O J İ S İ
204
üst düzeylerde yer alan nöronların çoğu her iki taraftan gelen girdilere yanıt verir. İnsanlarda bu merkez lateral serebral fissürün tabanına yerleşmiştir ve normalde beyin yüzeyinde görülmez. Primer işitme alanına bitişik işitme asosiyasyon alanı geniş olup insulaya kadar uzanır. Olivokohlear banther odituvar sinir içinde efferent liflerin yaptığı belirgin bir bant olup hem ipsi lateral hem de kontralateral superior olivar kompleksten doğmaktadır. Primer olarak korti organındaki dıştüy hücrelerinin tabanları çevresinde sonlanmaktadır. Yarım Daire Kanalları Başın her iki yanındaki yarım daire kanalları uzaydaki her üçboyut düzlemine yerleşip birbirlerini dik bir konuş gösterir. Membranöz kanallar kemiksi kanalların içinde perilenfa içine asılı haldedir. Reseptör bir yapıolan krista ampullarisher membranöz kanalın genişlemişson kısmına (ampulla) yerleşmiştir. Her krista, ampullayı kapatan jelatimsi bir bölme (kupula) ile örtülmüştüy hücreleri ve destek hücreleri içerir. Tüy hücrelerin uzantılarıkupulaya gömülüiken bu hücrelerin tabanlarıvestibulokohlear sinirin vestibular parçasının afferent lifleri ile yakın temastadır. Utrikul ve Sakkulus Her membranöz labirentteki utrikulun tabanında bir otolitik organ (makula) vardır. Diğer bir makula yarıdik bir pozisyonda sakkulus duvarına yerleşmiştir. Makulalar destek hücreleri tüy hücrelerinden oluşmuştur ve kalsiyum karbonat kristallerinin (otolit) içine gömülü olduğu otolitik bir zar tarafından kuşatılmıştır. Otokoniaveya kulak tozu adıverilen otolitler insanda 3–19 µm boydadır ve endolenfadan daha yoğundur. Tüy hücrelerinin uzantıları zara gömülüdür. Tüy hücrelerinden gelen sinir lifleri vestibul okohlear sinir içinde kristalardan gelen liflerle birleşir. Sinir Yolları Her iki yanındaki makula ve kristaları besleyen 19 000 nöronun hücre gövdeleri vestibular ganglionda yerleşmiştir. Her vestibular sinir, aynı taraftaki vestibular ganglionun her 4 parçasında ve beyinciğin flokülonodüler lobunda sonlanır. Vestibular reseptörlerden kalkan impulsları talamus üzerinden serebral kortekse ulaştıran, anatomik olarak iyi tanımlanamamış başka yollar da bulunmaktadır
ŞEKİL 3.20 İşitme ve dengenin alınmasında temel yapılar.
İçkulaktaki tüy hücreleri ortak bir çatıya sahiptir. Böylece tüy uzantıları zar potansiyelinde yer değiştirme yönü ile orantılı potansiyel değişikliği doğuran bir mekanizma sağlamaktadır Afferent Sinir Liflerinde Aksiyon Potansiyellerinin Doğuşu Tüy hüclerinin uzantıları endolenfa içine uzanırken tabanları perilenfa içinde yüzer. Ayrıca görüldüğü kadarıyla stria vaskülariste özgün bir elektrojenik bir K+ pompası yer almış olup bu pompa skala medianın skala timpani ve skala vestibuliye göre elektriksel olarak pozitif olmasından sorumludur. Afferent Sinir Liflerinde Aksiyon Potansiyellerinin Doğuşu Eldeki kanıtlar her stereosiliumda yaklaşık bir kanal olacak şekilde stereosiliumların tepe uçlarında mekanosensitif kanallar bulunduğunu göstermektedir. En az 0.7 nm çapa sahip olan bu kanallar nispeten özgül olmayan katyon kanallarıolmakla beraber yüksek bir potasyum yoğunluğuna sahip endolenfa içinde

H A Y V A N F İ Z Y O L O J İ S İ
205
yüzmelerinden dolayı açık olmaları halinde K+ tüy hücresine girer ve depolarizasyon oluşturur Afferent Sinir Liflerinde Aksiyon Potansiyellerinin Doğuşu Burada Ca2+ da girer ve tüy hücresi ile temas eden afferent nöron veya nöronları depolarize eden sinaptik bir transmitter salınır. İkinci nöronlar vestibuler çekirdeklerden başlayıp vestibulospinal traktuslar içinde omurilik boyunca aşağı doğru iner ve medial longitüdinal fasikuluslar aracılığı ile göz hareketlerinin denetlenmesinden sorumlu kafa sinirlerinin motor çekirdeklerine ulaşmak üzere yukarıdoğru tırmanır. Bu hücreler destekleyici veya sustentaküler hücrelerinden yapılmış bir epitel içine gömülüdür. Hücrelerin tabanları afferent nöronlarla yakın temastadır. Hücrelerin apikal uçlarından çubuğa benzeyen 30-150 tane uzantı veya tüy çıkar. Kohleadaki durum hariç tutulursa bu uzantılardan bir tanesi, yani kinosilium hareketsiz fakat gerçek bir silia olup merkezde yer alan bir çift mikrotubuli etrafına daire şeklinde dizilmiş 9 çift mikrotubuliden yapılmıştır Kinosilium en büyük uzantılardan biri olup genişlemiş bir uca sahiptir.
Tüy Hücreleri
Ergin memelilerin kohleasındaki tüy hücrelerinden kinosilium ortadan kaybolmuştur. Bununla beraber stereosilia adı verilen diğer uzantılar tüm tüy hüclerinde bulunur. Stereosiliumların koçan kısmıbiribirine parelel aktin flamanlarından oluşmuştur. Her hücre üzerinde bu uzantılar bir çember tabana oturmuş koni şeklinde bir demete benzeyen düzgün bir çatı gösterir. Dik eksene göre bütün stereosiliumların boyları aynı iken periferden kinosiliuma doğru giden eksen boyunca stereosiliumların boyları giderek artar. Elektriksel Yanıtlar Tüy hücrelerinin zar potansiyeli –60 mV kadardır. Stereosilia ve kinosiliuma doğru itildiği zaman zar potansiyeli –50 mV’a kadar yükselir. Uzantıların yaptığı bu bant zıt yöne itildiğinde ise hücre hiperpolarize olur .
ŞEKİL 3.21 Kinosiller
Uzantıların bu eksene dik yönde yer değiştirmesi halinde zar potansiyelinde hiçbir değişiklik olmaz ve uzantıların bu iki uç arasında herhangi bir yöne doğru yer değiştirmesi, hareketin kinosiliuma veya kinosiliumdan uzağa doğru yönelme derecesi ile orantılı hiperpolarizasyon

H A Y V A N F İ Z Y O L O J İ S İ
206
veya depolarizasyona neden olur. Nedeni tam olarak bilinmemekle beraber bu düzenleme normal jeneratör potansiyel üretimi için gereklidir. Perilenfa temel olarak plazmadan oluşur. Plazmadaki mannitol ve sükrozun skala timpanideki perilenfaya geçişi skala vestibulideki perilenfaya geçişine göre daha yavaştır ve bu iki sıvı arasında küçük bir bileşim farkı varsa da her ikisi de hücre dışısıvıya benzer. Öte yandan endolenfa stria vaskülaris tarafından oluşturulur ve yüksek bir K+ ile düşük bir Na+ yoğunluğuna sahiptir. Stria vaskülaris hücreleri yüksek bir Na -K ATP’az yoğunluğuna sahiptirler. Bu uzantıların kinosiliuma doğru hareketi kanalların açık kalma süresini artırırken kinosiliumdan uzaklaşmaları bu kanalların açık kalma süresini azaltır. Transmitterin kimliği saptanmamışsa da böyle bir transmitter maddenin varlığıhakkında kesin kanıtlar bulunmaktadır.
Ses özelikleri
İnsanda duyulabilir ses frekans aralığı yaklaşık olarak saniyede 20-20 000 döngü(cps, Hz) arasında değişir. Ses Dalgaları Ses, dış ortamdaki moleküllerin longütudinal titreşimlerinin, yani moleküllerin sırayla yoğunlaşıp seyrekleşmesinin kulak zarına çarpması ile oluşan bir duyudur. Bu hareketlerin kulak zarı üzerine olan basınç değişikliği olarak çizilmesi bir dalga serisi verir ve dış ortamdaki bu hareketlere genelde ses dalgaları denir. Ses dalgaları deniz düzeyinde 20 0C sıcaklıkta havada yaklaşık 344 m/s (Saatte 770 mil) hızla hareket eder. Sesin hızı sıcaklık ve irtifa ile artar. İnsanın arasıra girdiği diğer ortamlarda ses dalgalarının aynı şekilde fakat farklı hızlarda iletildiği bulunmuştur. Örneğin 20 0C sıcaklıkta tatlısuda ses hızı 1450 m/sn olup bu hız tuzlu suda daha fazladır. Ses Dalgaları Bir sesin şiddetinin ses dalgasının genliği, bu sesin tınısının frekans ile ilişkili olduğu söylenir. Genlik ne kadar büyükse ses o kadar gürken frekans ne kadar fazla ise ses o kadar tizdir. Bununla beraber sesin tınısı frekans ek olarak diğer pek az anlaşılmış etmenler tarafından da belirlenmekte ve işitme eşiğinin bazı frekanslar için diğer frekanslara oranla daha düşük olmasından ötürü frekans sesin şiddetinide belirlemektedir. Yinelenen örüntülere sahip ses dalgaları, dalgaların tek başlarına karmaşık olmaları halinde dahi müzikal ses olarak algılanırken periyodik olmayan ve yinelenmeyen titreşimler gürültü duygusu verir. Müzikal seslerin çoğu sesin tizliğini belirleyen bir ana frekans ile bunun üzerine binmiş ve sesin özgün rengini veren armonik titreşimlerden (üst tonlar) yapılmıştır. Ses tınısındaki titreşimler aynı notayı çalmaları halinde dahi bizim değişik müzik gereçlerinin ayırt edebilmemize izin verir. Ses Dalgaları Bir ses dalgasının genliği kulak zarındaki en çok basınç değişikliği terimleri ile tanımlanabilirse de bağıl bir ölçek kullanmak daha uygundur.
ŞEKİL 3.22 Ses dalgaları
Diğer hayvanlar özellikle yarasa ve köpekler çok daha yüksek frekansları duyabilir. İnsan kulağının eşik düzeyi sesin tizliği ile değişmekte olup en yüksek duyarlılık 1000-4000 Hz

H A Y V A N F İ Z Y O L O J İ S İ
207
arasındadır. Konuşma sırasında ortalam erkek sesinin tizliği yaklaşık 120 Hz iken ortalama kadın sesinin tizliği yaklaşık 250 Hz’dir. Ortalama bir kişi tarafından ayırt edilebilen ses frekansları 2000 tane kadar iken eğitilmiş bir müzisyende bu sayıüst değerlere ulaşabilir. 1000-3000 Hz arasında en iyi frekans ayrımı yapılırken daha yüksek ve daha düşük frekanslarda ses ayrımı zayıflar. Ses Dalgaları Ses İletimi Kulak dışortamdaki ses dalgalarını işitme sinirlerindeki aksiyon potansiyellerine dönüştürür. Ses dalgaları kulak zarı ve kulak kemikçikleri tarafından özenginin taban parçasının hareketleri haline çevrilmektedir. Bu hareketler içkulak sıvısında dalgalanmalar yapar. Dalgaların Korti organı üzerine olan etkisi sınır liflerinde aksiyon potansiyelleri doğurur
Kulak Zarı ve Kemiklerin İşlevleri Kulak zarının dışyüzü üzerinde ses dalgalarının yaptığı basınç değişikliklerine yanıt olarak zar içe ve dışarı doğru hareket eder. Bundan dolayı zar ses kaynağının titreşimlerini taklit eden bir rezonatör gibi görev yapar. Ses dalgası durduğu zaman kulak zarının titreşmeside hemen derhal durur, yani kulak zarı hemen anında devreye giren kritik bir söndürme gücü’ne sahiptir. Özengi başının hareketleri, oval pencerenin arka kenarına içe ve dışa hareket edecek şekilde menteşelenmiş bir kapıya benzeyen taban parçasını ileri-geri sallar. Kulak Zarıve Kemiklerin İşlevleri İşitme kemikçikleri böylece bir kaldırgaç sistemi gibi fonsiyon görür ve bu yolla kulak zarının rezonatör titreşimlerini kohleanın skala vestibülisini dolduran perilanfaya karşıgelen özenginin hareketlerine dönüştürürler. Bununla beraber refleks reaksiyon zamanı 40–160 ms olduğundan silah atışı gibi kısa süre devam eden şiddetli uyarılara karşı koruyucu nitelik taşımaz Kemik ve Hava İletimi Ses dalgalarının kulak zarıve işitme kemikçikleri aracılığıile içkulaktaki sıvıya iletilmesine kemikçik iletimi denir. İçtüy hücrelerinin tüyleri olasılıkla tektoriyal membrana bağlanmamışsa da bunlar tektoriyal zar ile alttaki hücreleri arasında hareket eden sıvı tarafından belirgin şekilde bükülmektedir İç ve Dış Tüy Hücrelerinin İşlevleri İçtüy hücreleri işitme sinirlerinde aksiyon potansiyelleri üreten primer duyu hücreleridir ve muhtemelen sıvı hareketleri ile uyarılmaktadırlar. Kulak zarının hareketleri çekicin uzun koluna iletilir Çekiç kemiği, uzun ve kısa kollarının birleştiği yerden geçen bir eksen etrafında salındığından kısa kol böylece çekicin titreşmelerini örse iletir. Örs, bu titreşmeleri özenginin başına iletecek şekilde hareket eder. Bu sistem, çekiç ve örsün kaldıraç etkisinin gücü 1.3 kez daha fazla artırmasıve kulak zarı alanının örsün taban parçasının alanından çok daha geniş olması nedeniyle oval pencereye ulaşan ses basıncında artışa neden olur. Bu arda direnç nedeniyle ses enerjisinde kayıplar olursa da 3000 Hz altındaki frekanslarda kulak zarına çarpan ses enerjisinin %60’nın kohleadaki sıvıya iletildiği hesaplanmıştır. Timpanik Refleks Orta kulaktaki kaslar (tensor timpani ve stapedius) kasıldıklarızaman çekicin uzun kolunu içe, özenginin taban parçasını dışa doğru çekerler bu olay ses iletimini azaltır. Yüksek sesler genelde bu kaslarda bir refleks kasılma başlatır ve bu olaya timpanik refleks adı verilir. Bu refleks işitme reseptörlerinin aşırı uyarılmasına yol açan güçlü ses dalgalarını önleyerek koruyucu fonksiyon görür. Ses dalgaları aynı şekilde yuvarlak pencereyi kapatan ikinci timpatik zarda titreşimler başlatır. Normal işitme için önem taşımayan bu olaya hava iletimi adı verilir. İletimin üçüncü tipi olan kemik iletimi’nde kafatası kemiklerinin titreşimleri içkulaktaki sıvıya iletilir. Diyapozon veya diğer titreşen cisimlerin kafatasına direkt olarak uygulanması halinde önemli ölçüde kemik iletimi görülür. Bu yol çok güçlü seslerin iletiminde de rol oynar. İlerleyen Dalgalar Özengi kemiğinin taban parçasının hareketleri skala vestibüli içindeki perilanfada ilerleyen bir dalga serisini başlatır. Bu dalga kohleaya tırmanırken boyu bir doruğa yükselip daha sonra hızla düşer. Bu doruk nokta ile özengi arasındaki uzaklık dalgayı başlatan titreşimlerin frekans ile özengi arasındaki uzaklık dalgayıbaşlatan titreşimlerin frekansıile değişir

H A Y V A N F İ Z Y O L O J İ S İ
208
Yüksek tizlikte sesler kohlea tabanının yakınlarında doruk noktaya ulaşan dalgalar yaratırken pes sesler apeks yakınında doruğa ulaşan dalgalar üretirler. Skala vestibülinin kemik duvarlarıkatı ise de Reissner membranı esnektir. Baziler membran gerilim altında olmayıp üstelik skala vestibülideki dalgaların doruk noktaları tarafından skala timpani içine kolayca bastırılır. İlerleyen Dalgalar Skala timpanideki sıvının yer değiştiştirmesi yuvarlak pencereden havaya dağılır. Bundan dolayı ses baziler membranda bükülme meydana getirmekte ve bu bükülmenin doruk düzeyde görüldüğü nokta ses dalgasının frekansı tarafından saptanmaktadır. Korti organındaki tüy hücrelerinin tepeleri retiküler lamina tarafından gergin halde tutulur ve dıştüy hücrelerinin tepeleri retiküler hücrelerinin tüyleri tektoriyal zar içine gömülüdür. Özengi hareket ettiği zaman her iki zar aynı yönde hareket eder fakat farklıeksenler üzerinde döndüklerinden tüyleri büken ortak bir hareket görülür. Diğer taraftan dıştüy hücreleri, superior oliva komplekslerinden gelen kolinerjik efferent liflerle innerve edilmiştir. Bu hücreler hareketli olup depolarize olduklarında kısalır, hiperpolarize olduklarında uzarlar. Daha yüksek şiddette seslerde her akson geniş bir ses yelpazesine özellikle eşik uyarının görüldüğüf rekansın altında kalan frekanslarda karşı deşarj yapar.
İşitme Sinir Liflerindeki Aksiyon Potansiyelleri
Bir ses dalgası kulağa çarptığında algılanan tizliğin ana belirleyicisi Korti organının hangi bölgesinin azami düzeyde uyarıldığıdır. Kohleanın çeşitli bölgelerinden beyine giden yollar birbirlerinden ayrıdır. İşitme Sinir Liflerindeki Aksiyon Potansiyelleri 2000 Hz’den düşük ses frekanslarında sesin tizliğini algılamaya katılan ek bir etmen işitme sinirindeki aksiyon potansiyellerinin kalıbı olabilir. Bununla beraber bu yaylım ateş etkisi sınırlıdır; belli bir işitme sinir lifindeki aksiyon potansiyellerinin frekansı bir sesin tizliği yerine temel olarak şiddeti tarafından belirlenir İşitme Sinir Liflerindeki Aksiyon Potansiyelleri Bir sesin tizliği temel olarak ses dalgasının frekansına bağlı ise de sesin şiddeti de rol oynamaktadır. Bu tüy hücrelerinin pek az direkt alıcı özelliği bulunmakta ise de baziler membranın titreşim kalıplarını etkileyerek işitmeyi daha mükemmel hale getirirler. Bununla beraber bu kalıpların değiştirilmesinde kullanılan gerçek yöntem bilinmemektedir. İşitme Sinir Liflerindeki Aksiyon Potansiyelleri Tek bir işitme sinir lifindeki aksiyon potansiyellerinin frekansı uyarıcı sesin şidetiyle orantılıdır. Düşük şiddette seslerde her akson sadece tek bir ses frekansına karşı deşarj yapar ve bu frekans sinir lifinin kohleadan kaynaklandığı bölgeye bağlıolarak aksondan aksona değişir. Herhangi bir tondaki ses tarafından kurulmuşolan ilerleyici bir dalga baziler zar üzerinde belli bir noktada maksimum depresyon ve bunun sonucu olarak maksimum reseptör uyarılması yapar. Bu nokta ve özengi arasındaki mesafe sesin tizliği ile ters orantılıdır; pes tonlar kohlea apeksinde azami uyarıyaparken tiz sesler kohlea tabanında azami uyarıoluşturur. Frekans yeterince düşük olduğu zaman sinir lifleri bu ses dalgasının her döngüsüne bir impulsla yanıt vermeye başlarlar. Sesin şiddeti arttıkça pes sesler (5000 Hz altı) daha pesleşirken (4000 Hz üzeri) daha tizleşmektedir. Sesin devam ettiği sürede sesin tizliğini azda olsa etkiler. Bir sesin tizliğin algılabilmek için o sesin en az 0.01 saniye devam etmesi gerekir ve 0.01-0.1 saniye arasındaki sürelerde süre uzadıkça sesin tizliği artar. Son olarak belli bir frekanstaki armaniyi de kapsayan karma seslerin tizliği primer frekans kaybolsa (temel ses yitmesi) dahi hala aynı tizlikte algılanır. Medulla Oblongatadaki Nöronların İşitsel Yanıtları Kohlear çekirdeklerdeki ikinci nöronların ses uyarılarına verdikleri bireysel yanıtlar bireysel işitme sinir liflerindeki duruma benzer. En düşük şiddetteki seslerin frekansı birimden birime değişen bir yanıt uyandırır; ses şiddeti arttıkça yanıt verilen frekans yelpazesi giderek daha genişler. Birinci ve ikinci nöron yanıtları arasındaki en büyük fark medüller nöronlardan düşük frekans bölgesinden keskin bir “sınır çizgisi”bulunmasıdır. İkinci nöronların gösterdiği bu büyük özgürlük beyin sapında yer

H A Y V A N F İ Z Y O L O J İ S İ
209
alan bazı tür inhibitör süreçlere bağlı olabilirse de bunun nasıl gerçekleştirildiği bilinmemektedir. İşitme Korteksi Dorsal ve ventral kohlealar çekirdeklerden çıkan impulslar hem çaprazlaşan hem de çaprazlaşmayan karmaşık yollar içinde yukarı doğru tırmanırlar. Hayvanlardaki primer işitme korteksinde, sanki kohlea bunun üzerinde düz bir şerit halinde açılmış gibi ses tonlarının konuşlanmasında düzenli bir kalıp bulunur. İnsanda, pes sesler işitme korteksinde anterolateral, tiz sesler posteromedial olarak temsil edilir. Bununla birlikte kişiye ana sesi gitmiş karma bir ses dinletildiğinde uyarılan korteks bölümünün algılanan tizliğe karşılık gelmesinden ötürü işitme korteksinde per se olarak kodlanan şey sesin frekansıyerine tizliğidir. Yani saf frekansların ses tizliği haline işlenmesinin subkortikal bir düzeyde gerçekleşmesi zorunludur. İşitme Korteksi İşitme kortekslerindeki her nöron bir işitme uyarısının başlama, süre ve yinelenme hızı ile özellikle bu sesin geldiği yön gibi değişkenlere yanıt verir.
ŞEKİL 3.22 Ses yönün tayini
İşitme korteksinde bir çok nöron her iki kulaktan girdi alır ve bir uyarının bir kulağa erişme zamanıdiğer kulağa erişme zamanına göre sabit bir süre geciktiği zaman azami veya askari yanıt verirler Sağırlık Klinik sağırlık dış veya orta kulakta ses iletiminin bozulması(ileti sağırlığı) ya da tüy hücreleri veya sinir hücrelerinin hasarına (sinirsel sağırlık) bağlıolabilir. Streptomisinve gentamisingibi aminoglikozid antibiyotikler tüy hücrelerinin stereosiliumlarının mekanosensitif kanallarını tıkar ve hücrelerin yozlaşmasına neden olabilerek sinirsel sağırlık ve anormal vestibuler fonksiyona neden olur.
ŞEKİL 3.23 Ses algılanması
İşitmede Integrasyon ve Problemler Perde Yoğunluk Lokalizasyon Bütünleşme Medulla Thalamus Auditory cortex Sağırlık İletim Sensori nöral Sağırlık Uzun süreli gürültünün dıştüy

H A Y V A N F İ Z Y O L O J İ S İ
210
hücrelerinde yaptığı hasara işitme kaybı eşlik eder. Bu yönden bu nöronlar vizüel korteksteki bazı nöronlara benzerler Laboratuvardaki memeli hayvanlarda işitme korteksinin tahribi sağırlığa yol açmadığı gibi aynızamanda belli bir frekanstaki bir sese karşı geliştirilmiş koşullu yanıtlarıda ortadan kaldırmaz. Buna karşın işitme korteksi, ses özelliklerinin analizi ve sesin lokalize edilmesi ile beraber tonkalıplarının tanınması ile ilgilidir. Sesin Lokalizasyonu Yatay düzlem üzerinde yayılan bir sesin geldiği yeri saptamak, bu uyarının 2 kulağa geliş zamanları arasındaki farkın saptanmasına bağlıolup bu olay kaynağıdaha yakın olan tarafta sesin daha gür olacağı gerçeğine de bağlıdır. 20 µs kadar küçük olabilen ayırt edilebilir zaman farkının 3000 Hz altındaki frekanslarda en önemli etmen olduğu, 3000 Hz üzerindeki frekeanslarda ise ses şiddetindeki farkın en büyük önemi taşıdığı söylenmektedir. İleti sağırlığı nedenleri arasında dışkulak yolunun kulak kiri vaya yabancı bir cisimle kapanması, kulak kemikçiklerinin tahrip olmasıyinelenen orta kulak enfeksiyonlarından sonra kulak zarının kalınlaşmasıve özenginin oval pencereye bağlanmasında anormal bir katılık bulunmasıdır. Diğer nedenler arasında vestibulokohlear sinir ve serebellopontin açıtümörleri ve medullada vasküler harabiyet bulunmaktadır. İleti ve sinirsel sağırlık diyapozon ile yapılan bir grup basit testle birbirinden ayırt edilebilir.
Odiyometre
İşitme keskinliği genelde bir odiyometre ile ölçülür. Bu aygıt kulaklık aracılığıile deneye çeşitli frekanslarda saf tonlar gönderir. Her frekans için saptanan eşik şiddet bir grafik üzerine normal işitmenin yüzdesi olarak işaretlenir. Bu yolla sağırlığın derecesi nesnel olarak ölçüldüğügibi en fazla hasara uğramış ton ağırlığına ait bir tabloda elde edilir. Pencere (Fenestrasyon) Girişimleri İleti sağırlığının sık görülen bir şekli otoskleroza bağlı olup bu hastalıkta özenginin taban parçasının oval pencere ile bağlantısı anormal derecede katılaşmıştır. Bu hastalık bulunan kişilerde işitmede belli bir ölçüde düzelme sağlamak için hava iletimi kullanılabilir. Bu amaçla kemik labirentte zar ile örtülü bir delik oluşturulur ve sekonder kulak zarının titreşmesi ile oluşan dalgalar ileri dağıtılabilir. Bu “pencereleme” girişiminde horizontal yarım daire kanalına matkapla bir delik açılır ve bu delik deri ile örtülür
Vestibüler Fonksiyon
Döngüsel Hızlanmaya Verilen Yanıtlar Herhangi bir yarım daire kanalıdüzleminde döngüsel hızlanma bu kanalın kristasını uyarır. Bundan dolayı beyine ulaşan uyaran kalıbı dönüş düzlemi kadar dönüşyönü ile de değişir. Döngüsel Hızlanmaya Verilen Yanıtlar Doğrusal hızlanma olasılıkla kupulanın yerine değiştirmeyeceğinden kristaları uyarmaz. Vestübüler cekirdeklerden omurilik içinde aşağıya inen traktuslar temel olarak postür ayarlanması ile ilgilidirler; kafa sinir çekirdekleri için yukarı tırmanan bağlantılar ise geniş ölçüde göz hareketleri ile ilgilidirler.
Denge: Mekanoreseptör Integration Medulla Cerebellum Thalamus Cortex Denge ve Oriyantasyon Yolları Denge ve oriyantasyonla ilgili üçmodel vardır. Dönüş durduğunda hızın kesilmesi endolenfanın dönüş yönünde yer değiştirmesine neden olur ve kupula hızlanma sırasında yöneldiği yönün aksi tarafına deforme olur Döngüsel Hızlanmaya Verilen Yanıtlar Kupula 25–30 saniye sonra tekrar orta konumuna geri döner. Endolenfaya eylemsizliğinden dolayıdönüşyönünüaksi yönde yer değiştirir ve bu sıvı kupulayı iterek bunu deforme eder. Bu olay tüy hücrelerini uzantılarını eğer. Sabit bir dönüşhızına ulaşıldığında sıvıvücutla aynı hızda döner ve kupula dik konumuna geri döner. Kupulanın bir yöne doğru hareketi genelde bunun kristasından gelen sinir liflerinde artmışimpuls trafiğine neden olurken aksi yönde hareket genelde nöral

H A Y V A N F İ Z Y O L O J İ S İ
211
aktiviteyi inhibe eder. Dönme hareketi dönüşdüzeyine en yakın yarım daire kanallarında doruk uyarıya neden olur. Başın bir tarafındaki kanallar diğer taraftakilerin ayna hayali olduğundan endolenfa bir tarafta ampullaya doğru hareket ederken diğer taraftan ampulladan uzaklaşır. Bunun beraber elde bulunan kanıtlara göre labirentin bir bölümütahrip olduğunda diğer bölümler bunun fonksiyonlarını yüklenir. Bundan dolayıla birent fonsiyonlarının deneysel olarak lokalize edilmesi zordur.
Vestibular Apparat Krista Ampullaris ve Dinamik Denge
Nistagmus Dönüşün başlangıç ve bitişinde gözde görülen tipik sıçrama şeklinde ani harekete nistagmus denir. Görme uyarıları ile başlatılamamasına ve kör kişilerdede görülmesine karşı bu olay geçekte vücut dönerken gözün belirli noktalara tespitinin sürdürülmesini sağlayan bir reflekstir. Dönme başladığında gözler dönüş yönüne ters yönde yavaşça hareket ederek görsel tespiti sürdürür (vestibülo oküler refleks).Bu hareketin sınırına ulaşıldığında gözler hızla yeni bir tespit noktasına döner ve tekrar aksi yönde yavaşça harekete başlar. Hareketin yavaş kompenenti labirentlerden gelen uyarılarla başlatılırken hızlı konpenet beyin sapındaki bir merkez tarafından tetiklenir.
Nistagmus Nistagmus sıklıkla yatay ise de dönüş sırasında kafanın yana yatırılması halinde dikey veya kafanın öne eğilmesi halinde döngüsel de olabilir. Kulak enfeksiyonlarının tedavisi için kulak kanalı yıkanırken bu semptomların önlenmesi için, kullanılan sıvının vücut sıcaklığında olduğundan emin olmak önemlidir. Uzaysal Uyum Bireyin içinde bulunduğu uzaya uyumu büyük ölçüde vestibüler reseptörlerden alınan girdilere bağlı ise de görme ipuçlarıda önem taşır. Tanım olarak nistagmusta göz hareketinin yönü hızlı kompenentin yönüolarak tanımlanır. Dönüş sırasında hızlı kompenentin yönü dönüşyönüile aynı ise de dönüş durduğunda kupulanın dönüş yönünün aksi yönde hareket etmesi nedeni ile postrotatuvar nistagmus görülür. Beyin sapı lezyonları bulunan hastalarda dinlenme sırasında nistagmus görülür. Doğrusal Hızlanmaya Verilen Yanıtlar Memelilerde utrikul ve sakkulus makulalarıdoğrusal hızlanmaya yanıt verir. Genelde utrikül yatay, sakkulus dikey hızlanmayı yanıtlar. Otolitler endolenfadan daha yoğundur ve herhangi bir yönde hızlanma bunların zıt yönde yer değiştirmesine, tüy hücrelerinin uzantılarının bükülmesine ve sinir liflerinde aktivite dolmasına neden olur. Otolitler üzerine yerçekimi etkisi nedeniyle kafa hareket etmezken de makulalar tonik olarak deşarj yaparlar. Bu reseptörlerden doğan impulslar kafanın doğrulma refleksi ile diğer önemli postür düzenlemelerinden kısmen sorumludurlar.
ŞEKİL 3.24 Beyin konumunu algılanması

H A Y V A N F İ Z Y O L O J İ S İ
212
Doğrusal Hızlanmaya Verilen Yanıtlar Makulaların uyarılmasına verilen yanıtların çoğunun refleks tabiatında olmasına karşın vestübüler impulslar serebral kortekse de ulaşır. Bu impulslar olasılıkla hareketin bilinçli algılanmasından sorumludur ve uzayda oriyantasyon için gereken bilginin bir bölümünü sağlar. Aşırı vestibüler uyarıya eşlik ettiği bilinen bulantı, kan basıncıdeğişiklikleri, terleme, solgunluk ve kusma olasılıkla beyin sapındaki vestibüler bağlantılar üzerinde kurulu reflekslere bağlıdır. Vertigo (başdönmesi), gerçek bir dönme hareketi yokken kişinin dönüşduyusu almasıdır.
Utrikular Reseptör Hücreleri Üzerine Yerçekiminin Etkisi
Kalorik Uyarı Yarım daire kanallarıdışkulak yoluna vücut sıcaklığına göre daha sıcak veya daha soğuk su şırınga edilmesiyle uyarılabilirler. Sıcaklık farkıendolenfadan konveksiyon akışlarıbaşlatır ve sonuçta kupulanın hareketine neden olur. Bazen tanıamacıyla kullanılan bu kalorik stimülasyonyöntemi bir nistagmus, başdönmesi ve kusmaya neden olur. Vücudun çeşitli kısımlarının bağıl konumlarıhakkında bilgi sağlayan eklem kapsüllerindeki propriyoseptörlerden ve derideki eksteroseptörler ve özellikle dokunma ile basınçreseptörlerinden gelen impulslarla uygun bilgiler sağlanır. Bu dört girdi kortikal bir düzeyde birleştirilerek, kişinin uzaydaki konumuna ait sürekli bir resim haline getirilir.

H A Y V A N F İ Z Y O L O J İ S İ
213
Dolaşım fizyolojisi
Dolaşım sistemi kalp damar, kan doku hakkında bilgiler.
1. İlke: Vücuttaki bütün dokuların kan akımı, daima doku ihtiyaçlarına göre hassas biçimde kontrol edilir.
2. İlke: Kardiyak debi başlıca, lokal doku akımlarının tümü tarafından kontrol edilir.
3. İlke: Arteriyel basınç, genellikle lokal akım kontrolü ya da kalp debisi kontrolü mekanizmalarından bağımsız olarak düzenlenir.
Kalp ve damarlar Kalp benzeri yapılar omurgasızlarda bulunur. Omurgasızlarda kan yerine hemolenf pompa benzeri yapılarla hareketlendirilir. Odalara ayrılmış kalpler çoğunlukla omurgalılarda ve bazı yumuşakçalarda bulunur. Yumuşakçaların kalpleri bir ile iki kulakçık ile bir karıncıktan meydana gelmiştir. Kulakçıklar genellikle ince bir kas tabakasından yapılmıştır. Karıncık ise kuvvetli kontraksiyon yapan bir organdır, bu nedenle de oldukça kalın bir kas tabakasından yapılmıştır. Kafadanbacaklılarda ise kalp sistemi iyi gelişmemiştir ve bir perikard boşluğu içinde bulunur. Kan vücutta toplardamarlar aracılığı ile solungaçların tabanındaki solungaç kalblerine döner ve solungaçlardan, solungaç toplardamarları ile kalp sisteminin kulakçıklarına geçerler. Bu solungaç kalpleri ritmik kontraksiyon yeteneğine sahiptirler ve aynı zamanda midyelerin perikard bezerine karşılık gelen, yuvarlak bir kan bezi (solungaç dalağı) ile de ilişkilidir. Bu kalpler hem solungaçların ve hem de vücut dokularındaki damarların dirençlerine karşı koyabilirler. Balıklarda dolaşım sistemi kapalı olup, kalp bir kulakçık (atrium) ve bir karınıcık’a (ventrikulüse) sahiptir. Kan aortaya girmeden solungaç damarlarından geçer. İki kanalı olan kalb oksijenli ve oksijensiz kanı karışık olarak bulundurur. Kan solungaç kapillerinden geçtikten sonra yüksek bir basınca sahip olur. Kulakçığın kontraksiyonu ile çıkan kan, solungaçlardaki solunum kılcalları sistemine yayılır ve sonra tek bir damar ile toplanarak vucüt dokularında bulunan ikinci bir kılcal ağı sisteminden geçer. En son olarak da yine tek bir damar ile kulakçığa geri getirilir. Kanın karıncıktan kulakçığa dönmesine engel olmak için, ikisi arasında bir kapak bulunur. Fakat böyle bir sistemde, kılcallarda büyük bir direnç vardır. Bundan dolayı, kalbin kuvveti birinci kılcal sistemde hemen hemen tükenir ve ikinci kılcal ağı için çok az bir kuvvet kalır. Bu zararı ortadan kaldırmak için dolaşım diğer omurgalılarda biri küçük diğeri büyük olmak
Bölüm
4 Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
214
üzere ikiye ayrılmıştır. Böylece solunum ve vücut kılcal sistemleri, ayrı ayrı kalp karıncığından direkt olarak kanı alabilirler. Kurbağalarda kalp iki kulakçık ve bir karıncıktan meydana gelir. Bu nedenle vücuttan gelen kirli kan ile akciğerlerde temizlenip gelen kan karıncıkta birbiri ile karışır. Sürüngenlerde ise iki kulakçık ve birbiri ile tamamen ayrılmamış iki karıncıktan meydana gelen kalpte kirli ve temiz kan birbiri ile kısmen karışırlar.
ŞEKİL 4.1 Açık ve kapalı dolaşım
Böcekte dolaşım sistemi
Açık bir sistemde kanın hareketi, kapalı bir sistemdeki kadar hızlı ve verimli olmayacağından(kanın hidrostatik olarak kanallar yani damarlar yerine boşluklarda akmasından dolayı), böcekler gibi aktif ve nispeten yüksek metabolik hıza ve kusursuz bir iç düzenlemeye sahip hayvanların açık dolaşım sistemine sahip olmaları şaşırtıcı gelebilir. Buna karşın böcekler dokularına oksijen taşınması bakımından kana bağımlı değildir. Bu işlev çok dallı trake sistemi tarafından yerine getirilir. Bunun sonucu olarak, kanın çok hızlı ya da düzgün akması, böçekler için yaşamsal önemde değildir. Bu, bir canlının çeşidi sistemlerinin, onun ihtiyaçlarına uyum gösterirken karşılıklı etkileşimlerine güzel bir örnek oluşturur. Böceklerin dolaşım sistemi, diğer birçok eklembacaklınınkinden dahi daha basittir. Genellikle, bir böcekteki tek belirgin kan damarı ki kalp olarak kabul edilir, hayvanın toraksının ve abdomeninin üst tarafında, vücut boyunca uzanan bir damardır. Kalbin arka tarafında, bir dizi açıklık, ostiumlar, bulunur. Bunların herbiri, kanın sadece damar içine doğru hareketine izin veren kapakçıklar tarafından kontrol edilir. Kalp kasıldığı zaman kanı, açık olan ön ucundan dışarı, baş bölgesine iter. Tekrar gevşediği zaman kan, ostiumlar yoluyla kalbe dolar. Kan kalbin dışına çıktığında artık damarlar içinde değildir. Venler, kılcallar ya da arterler bulunmaz. Sadece, kapakçıklarıyla ve bazen bir arter olarak adlandırılıp ön ucu oluştu-ran kısa bir uzantısıyla birlikte kalbin kendisi bulunur. Kan, sadece böceğin iç organlarının arasındaki boşlukları doldurur. Böylece bu organlar kanla doğrudan temasta olurlar. Kalbin hareketi, kanın ön uçtan çıkıp vücut boşluklarında yavaşça ilerledikten sonra, arka uçtan yeniden kalbe girmesini sağlar. Hayvanın aktivitesi sırasında, vücut duvarındaki ve bağırsaktaki kasların karıştırma etkisiyle kanın hareketi hızlandırılır. Böylece, hayvanın organlarının hızlı besin sağlanması ve atıkların uzaklaştırılmasına en fazla ihtiyaç duydukları, koşma ya da uçma gibi hayvanın en aktif olduğu durumlarda, kan, aktivitenin kendisi nedeniyle, nispeten hızlı hareket eder.

H A Y V A N F İ Z Y O L O J İ S İ
215
ŞEKİL 4. 2 Omurgasızlarda kalp yapısı ve balıkta ve kurbağalarda ve kertenkelede dolaşım
İnsanda Kalp
Kalp sağda ve solda birer atrium ve ventrikül olmak üzere dört odacııktan oluşur. Kalpte iki adet atrioventriküler kapak, iki adet de büyük damar kapakları (semilunar kapak) olmak üzere 4 kapakçık bulunmaktadır. Sağdaki kulakçık ve karıncığı triküspit kapak (üç kapakçıktan oluşan); soldaki kulakçık ve karıncığı ise mitral kapak( iki kapakçıktan oluşan) ayırır. Kapakçıklar, kanın tek yönlü akmasını yani geriye dönüşünü engellemeye yarar. Kapaklar, kanın karıncıklara tek yönlü girişini sağlarken aynı zamanda tek yönlü çıkışını da sağlarlar.
Kalbin sol karıncığının bitimi ile kalpten çıkan ve insanın en büyük atardamarı olan aort damarının başlangıcı arasında aort kapağı vardır. Benzer olarak pulmoner kapak sağ karıncık ile pulmoner damar arasındadır. Kalbin sağ sistemine tüm vücuttan gelen kanı toplayan damarlar (vena cava inferior ve vena cava superior) açılır. Bu kan akciğer atardamarı (Pulmoner arter) ile sağ sistemden ayrılır. Akciğerlerden akciğer toplardamarları (pulmoner venler) ile dönen kan, sol kulakçık ve sol karıncığı dolaşarak aort damarları ile tüm vücuda pompalanır.
Vücuttaki bütün dokuların kan akımı, daima doku gereksinimlerine göre hassas biçimde kontrol edilir. Kardiyak debi başlıca, lokal doku akımlarının tümü tarafından kontrol edilir. Arteriyal basınç, genellikle lokal akım kontrolü ya da kalp debisi kontrolü mekanizmalarından bağımsız olarak düzenlenir. Dolaşımın temel fonksisyonu: O2 ve CO2 taşınması, sindirim sisteminden besinlerin alınıp gerekli yerelere taşınması, Fazla su ve metabolik artıkların böbreklere taşınması, vücut içinde açığa çıkan ısının vücut yüzeyine dağıtılması, endokrin salgıların hedef hücre ve organlara taşınmasıdır. Kısaca dolaşımdaki

H A Y V A N F İ Z Y O L O J İ S İ
216
kan damarlarıyla besin maddelerini, oksijen ve hormonları vücudun her tarafına taşır. Aynı zamanda hücre ve dokularda oluşan metabolizma artıklarını uzaklaştırır.
ŞEKİL 4.3 Kalp çalışma mekanizması
Kalp işlevini kasılma "sistol" gevşeme "diyastol" dönemleri ile gerçekleştirir. Atriyumlar ve ventriküller aynı anda kasılır ve gevşerler. Ventriküller, atriyumlardan 1/10 saniye sonra kasılırlar, bu sürede ventriküller atriyumlardan gelen kan ile dolar. Bu olay sürekli olarak tekrarlanır.
ŞEKİL 4.4 Kalpte kan akım yönleri

H A Y V A N F İ Z Y O L O J İ S İ
217
Kalbden çıkan (kalpten kan göntüren) damarlara arter adı verilir. Bunlar sürekli olarak dallanarak sayıca artarlar ancak çapları küçülür, arteriyel kapillar yataklarına ulaşırlar. Kapillar damarlar kan ile hücre ve dokular arasında madde değişiminin yapıldığı yerlerdir. Bu nedenle dolaşım sisteminin fonksiyonel ünitesi olarak kabul edilirler. Kanı kalbe geri getiren damarlara ise vena adı verilir. Kalp fonkisyonu ise kan basıncını oluşturur. Kan sirkülasyonunu yönlendirir. Kalp, sistemik ve pulmoner dolaşımı birbirinden ayırır.
Kalp dört odacıktan oluşur: üst kısımda iki kulakçık (sağ ve sol atrium) ve kulakçıkların altında iki karıncık (sol ve sağ ventrikül). Sol ve sağ kalp kısımlaım arasında septum bölme vardır. Bu bölme(septum) doğumdan sonra kapanır(Eksikliği kalp deliği olarak bilinir) Kulakçıklar ile karıncıklar arasında ve karıncıklarla buradan çıkan damarlar arasında kapaklar bulunur. Kapaklar, kanın tek yönlü akmasını, dolayısıyla kanın geri kaçışını engellemeye yarar. Kapaklar, kanın karıncıklara tek yönlü girişini sağlarken tek yönlü de çıkışını sağlarlar. Her kapak (2 yaprakçıktan oluşan mitral kapak hariç) 3 yaprakçıktan oluşur Bu dört kalp kapak şunlardır:
Kalb kanın iki taraflı pompalanması için özelleşmiş bir kan damarı yapısındadır. Vücuttan sağ kalbe dönen kan buradan akciğerlere pompalanır. Akciğerlerden sol kalbe dönen oksijenize kan buradan vücudun tüm organ ve dokularına pompalanır. Kalp ve akciğerler arasında irtibatı sağlayan damarlar akciğer dolaşımı’nı (pulmoner dolaşım), kalp ve diğer vücut bölgeleri arasındaki damarlar ise sistemik dolaşım’ı (periferik dolaşım) oluşturur.
Sistol; kalp odacıklarının kasılma dönemidir. Atriyumların (kulakçık) kasılması ile kan karıcıklara, ventriküllerin (karıncık) kasılması ile kan akciğerlere ve tüm vücuda gönderilir. Diyastol; atriyum ve venriküllerin gevşeme dönemidir, bu sürede kan ile dolarlar.
Kardiyak frekans; kalp atım hızıdır (nabız-kalp atım sayısı; KAS), bir dakika olarak değerlendirilir. Bir yaşında 120/130, normal bireylerde 70–75 arasındadır.
Atım hacmi; bir sistolde aorta ve akciğerlere gönderilen kan miktarıdır. Üst düzey dayanıklılık sporcusunda 110–120 ml, sporcu olmayanlarda 70 ml kadardır.
Kardiyak debi (kardiyak output); bir dakikada kalpten çıkan kan miktarıdır, 5–6 litredir. Kardiyak frekans ile atım hacminin çarpımıma eşittir.
• Triküspid kapak: Sağ kulakçık ile sağ karıncık arasında bulunur. Üç parçalıdır.
• Pulmoner kapak: Sağ karıncık ile pulmoner arter (akciğer arteri) arasındaki sağ karıncıkdan pompalanan kanın geri dönüşünü engelleyen üç adet yarım ay şeklindeki kapaklardır.
• Mitral kapak: Sol karıncık ve sol kulakçık arasında bulunur.
• Aort kapağı: Sol karıncık ile aort arasında bulunur. Bu kapaklar sol karıncıkdan pompalanan kanın geri dönüşünü engeller.

H A Y V A N F İ Z Y O L O J İ S İ
218
Taşikardi; kalp hızının artışını anlamına gelir, genellikle dakikada 100 atımdan daha büyük hızları tanımlar. Bradikardi; kalp hızının yavaşlaması anlamına gelir, genellikle dakikada 60 atımdan daha düşük hızları tanımlar.
ŞEKİL 4.5 Kalp kısımları
Kalp 3 tabakadan oluşur. Dıştan içe doğru perikart, miyokart ve endokart olarak adlandırılmaktadır. Dışta bulunan "perikart", kalbi dıştan saran fibro-seröz yapıda bir zardır. Bu zarın arasında sürtünmeyi azaltan bir sıvı(perikart sıvısı) bulunur.
ŞEKİL 4.6 Kalp tapakaları perikart sıvısı.
Kalp tapakalarında ortada bulunan "miyokart", kalbin kas tabakasıdır. Kalbin en kalın tabakası burasıdır. Pompalama görevi yapan karıncıklar, kulakçıklara göre özellikle sol karıncıkda daha kalın durumdadır. En iç kısım olan "endokart", tek katlı epitel hücrelerden oluşmuştur. Kalbin iç yüzeyini örten bu tabaka, içeriye doğru uzantılar vererek kalpteki dört kapağın temelini oluşturur.
Kalp Sesleri
Kalbin her iki tarafında, ritmik kasılmaları tek yönlü bir kan akışına dönüştüren ikişer kapakçık bulunur. Bu kapakçıklar, kanın geri akışına neden olabilecek bir basınç farkı oluştuğunda, anatomik yapıları gereği otomatik olarak kapanırlar. Kalbin kasılması(sistoli) ve gevşemesi(diastoli) sırasında kanın kapakçıklara çarpması, kapaçıkların kapanması, kasılma sırasında kanın arter ve aorta yoğun bir şekilde gönderilmesi sonunda kalp sesleri meydana gelir. Kalp sesleri stethoscop yada kardiyomikrofon ile duyulabilir.
Birinci kalp sesi Ventrikul kasıldığı zaman, ilk olarak A-V kapaklarının kapanması oluşur . Frekansı (perdesi, pilch) düşük, süresi nisbeten uzun olan bu titreşim 0,12 saniyea kadar sürer,

H A Y V A N F İ Z Y O L O J İ S İ
219
EKG birinci kalp sesi R dalgasına aynı zamana oluşur. Birinci kalp sesinin en iyi duyabileceği nokta sol beşinci intercostal aralıktır. İkinci kalp sesi birinci kalp sesine göre daha kısa süren yüksek tonda bir ses olup, diyastol sırasında aort ve pulmoler arterde yer alan semilunar kapakların kapanmasından meydana gelir. İkinci kalp ses 0,08 saniye kadar sürer EKG ikinci kalp sesi T dalgası ile aynı zamana gelir.
ŞEKİL 4.6 Kalp diyastoli ve sistoli sırasında meydana ses, basın ve elektriksel değişmeler.
Kalp ileti sistemi
Kalbin kasılarak kendisine gelen kanı bir pompa gibi göndermesi kalp üzerindeki özelleşmiş myokard hücre toplulukların oluşturdukları aksiyon potansiyeli otonom ve düzenli meydana gelmesiyle. Kalbin sinir ileti sisteminde özel hücre kümeleri, demetleri ve lifleri bulunmaktadır. Uyarı ve ileti sistemi, sinoatrial düğüm (SA), atrioventriküler düğüm (AV), atrioventriküler demet (his demeti) ve purkinje lifleri olmak üzere dört bölümden oluşmaktadır. Bunlardan ilk ikisi uyarı sisteminde(otonom aksiyon potansiyeli oluşturma), diğer ikisi ise ileti sisteminde yer almaktadır. Kalpte aksiyon potansiyellerinin kaynağı, normalde sinüs düğümü (sinoatriyal düğüm) dür. Sağ atriyumda bulunur; hücreleri kontraktil kas filamentlerini içermez ve küçük çaplıdır. Sinüs düğümü hücreleri atriyal kas hücreleri ile doğrudan bağlantılıdır. Sinüs düğümü hücresinin dinlenme potansiyeli, membranlarında sodyum ve kalsiyumu sızdıran kanalların olması nedeniyle -55 ila -60 mV arasındadır ve sabit değildir. Miyokardın kasılmadan sorunlu olan diğer hücrelerinde ise -85 ila -90 mV’dur. Dinlenme potansiyeli daha az negatif olduğundan hızlı sodyum kanalları kapalıdır. Bu nedenle yalnız yavaş kalsiyum sodyum kanalları açılabilir ve aksiyon potansiyeli oluşturabilir. Bu nedenle atriyal nodal aksiyon potansiyeli ventrikül kası aksiyon potansiyelinden yavaş gelişir. Ekstraselüler sıvıda sodyum iyon konsantrasyonunun yüksek olması ve membranda sodyum sızıntı kanallarının bulunması nedeni ile sodyum hücre içine girer(Pace maker dışında sızıntı

H A Y V A N F İ Z Y O L O J İ S İ
220
kanalları ile sodyuma bağlı depolarizasyon gözlenmez). Membran potansiyeli eşik voltaj olan yaklaşık -40 mV’a ulaşınca yavaş kalsiyum kanalları aktiflenerek aksiyon potansiyeli oluşur. Bu olaylar sonunda potasyum hücre içine girmesiyle pacemaker hücreleri ilk konum döner. Bu ritmik potansiyel değişikliklerine: “Prepotansiyel” ya da ''pacemaker'' potansiyeli denir. Pacemaker potansiyeline normalde yalnız SA ve AV düğümlerde rastlanır. İleti sisteminin diğer hücreleri ise latent karekterdedir; SA ve AV düğümler depresyona uğradığı ya da ileti bloğu meydana geldiği zaman ritmik deşarjlar yaratabilirler. Tuhaf akım “funny current” sinoatriyal düğüm (SA), atriyo-ventriküler düğüm (AV) ve iletim dokularında Purkinje liflerinde bulunur(Otonom Prepotansiyel” ya da ''pacemaker'' potansiyeli oluşturmayı sağlar).
ŞEKİL 4.7 Kalpteki ritmin kasılmayı sağlayan Ca ve K kanalları(pacemaker) kalp kas aksiyon potansiyeli
Tuhaf (normal olarak -40 mV ile -60 / -70 mV) akım AV ve SA düğümlerde bulunan de sodyum-potasyum sızma kanallarıdır. Bu durumda uyarı oluşturan kalp dokularının eşik potansiyelinin geçilmesi ve aksiyon potansiyeli oluşma ihtimalini artırır. Tuhaf akımı kanalarının başka bir sıra dışı özelliği cAMP aktivasyondur. Siklik adenozin monofosfat (cAMP) moleküller f-kanallara doğrudan bağlabilir ve bu kanalların açılmasını sağlar. cAMP bağlı olarak kalp atım hızının ayarlanaması otonomik regülasyonu sağlanır. Sempatik sinir uyarımı tuhaf kanalları ( f-kanallarına) bağlanan cAMP molekül seviyesini yükseltir. Bu durum kanallarla daha sık aksiyon potansiyeli oluştırmasına kalp atım hızının artışına yol açar. Tersine Parasempatik sinir stimülasyonu kalp atım hızını cAMP azaltarak yavaşlatır.

H A Y V A N F İ Z Y O L O J İ S İ
221
ŞEKİL 4.8 Kalp iletim demetleri
Kalpte sinirsel uyarı ilk olarak sinoatrial düğümden çıkar atriumları (kulakçıklar) dolaştıktan sonra atrioventriküler düğüme gelir ve burada biraz bekledikten sonra. Bir kalp atımı, aşağı inerek ventrikülleri (karıncıkları) uyarır kalbin sağ kulakçığının üst taraflarında bulunan ve sinoatrial (veya sinüs) düğüm adı verilen özelleşmiş bir hücre demetinden oluşan bölgenin elektriksel bir uyarı çıkarması ile başlar. Bu bölge kalbin doğal pili olarak bilinir (pacemaker) Sinüs düğümünden çıkan bu uyarı kalbin her iki kulakçığı boyunca ve aşağıya doğru yayılır ve kulakçıklar kasılarak içlerindeki kanı karıncıklara gönderirler. Daha sonra uyarı kulakçıklar ile karıncıklar arasında bulunan başka bir özel bölgeye; atrioventriküler (AV) düğüme gelir. AV nod dışında atriumlar ve ventriküller arasında doğrudan bir bağlantı olmaması, iletiyi geciktirir ve atrial kontraksiyonun ventriküler doluşa katkıda bulunmasını sağlar. Elektrik iletisi karıncıklara ulaştırılmadan önce atrioventriküler düğümde kısa bir süre bekletilir. Atriumlar ve ventriküller arasında bağ dokusu olduğundan impuls interkalar disklerle atriumdan ventriküle geçemez. İmpulsun atriumdan venriküle geçişi AV nodundan olur. Böylelikle kulakçıklarla karıncıklar aynı anda kasılmaz. Kulakçıkların kasılması bittikten sonra His-Purkinje sistemi adı verilen bir elektriksel ağ ile uyarı tüm karıncıklara yayılır ve kasılarak içlerindeki kanı akciğerlere ve aort yoluyla vücuda pompalarlar. Sinüs düğümü tekrar başka bir uyarı çıkararak yeni bir döngüyü başlatır. Normalde sinüs düğümünden dakikada 60-100 civarında uyarı çıkar. Bu da kalp hızını oluşturur. Kalp kasının, doğumdan ölüme kadar yorulmadan çalışır Kalp kasında uyarı oluşturan özelleşmiş hücreler de sodyum iyonları dışarıya pompalandıklarında zarın dinlenme potansiyeli tekrar kazanılmış olur. Fakat bunlardaki eşsiz durum şudur: Bir sinir ya da kas aracılığıyla yeniden uyarılmadan sodyum iyonları zardan içeriye sızar ve kendi kendine yeni bir uyarma meydana getirir. Bu sodyum iyonlarının içeriye ve dışarıya kendi kendine akması sinoatrial düğümün ritmik hareketlerini meydana getirir.
Sinoatriyal düğüm (SA düğümü veya sinüs düğümü), kalbin sağ kulakçığında yer alan vuru üretici (pacemaker) doku. Normal haldeki sinüs ritminin ortaya çıkışını sağlar. Sinoatriyal düğüm sağ kulakçık duvarında üst anatoplardamar girişine yakın bölgede yerleşik bir hücre grubudur. Bu hücreler kalpteki miyositlerin özelleşmiş biçimidir. Kalpte, sağ kulakçığın alt kesiminde kulakçıklararası bölmenin karıncıklar arası bölmeye geçiş yerinde yer alan, elektriksel etkinliğin karıncıklara geçmesini sağlayan uyarı üretim merkezi, AV düğüm, Aschoff-Tawara düğümü. Kulakçıklardan karıncıklara uyarının iletilmesini sağlar.

H A Y V A N F İ Z Y O L O J İ S İ
222
ŞEKİL 4.9 Kalp iletim demetleri
Pacemaker potansiyeline normalde yalnız SA ve AV düğümlerde rastlanır. İleti sisteminin diğer hücreleri ise latent karekterdedir; SA ve AV düğümler depresyona uğradığı ya da ileti bloğu meydana geldiği zaman ritmik deşarjlar yaratabilirler. SA düğümün kontrolü olmadığı durumlarda AV düğümü otonom olarak dakikada yaklaşık 50 atım yapacak aksiyon potansiyeli meydana getirir. Benezer olarak SA ve AV düğümün kontrolü olmadığında purkinje lifleri dakikada yaklaşık 30 atım yapacak aksiyon potansiyeli meydana getirir.
ŞEKİL 4.10 Kalpte ritim belirleyen merkezlerin önceliği.
Kalp kası uyarılması için sinirsel impulsa gereksinimi olmayan, kendi uyarılarını kendisi oluşturabilme özelliği olan bir kastır. Buna karşın kalp ihtiyaca göre atım gücü ve sayısı hormonal ve siniresl kontrol mekanizmaları denetlenir. Kalp kası otonom sinir sisteminin etkisi altındadır, ancak bu etki kalpteki uyarıları başlatma değil, kalbin kendiliğinden oluşturduğu kasılmayı düzenleyici niteliktedir. Parasempatik sinirler uyarılar atım hızı ve gücünü artırken Parasempatik(N. Vagus) kalp atım hızı ve gücünü azaltır.

H A Y V A N F İ Z Y O L O J İ S İ
223
ŞEKİL 4.11 Kalpi etkileyen sinirler ve eksersizdeki kalp atım sayısı
Bir kalp atımı, kalbin sağ kulakçığının üst bölümlerinde bulunan sinoatrial düğümün elektriksel bir uyarı çıkarmasıyla başlamaktadır. Bu düğümün özelliği eşit aralıklarla ve belirli bir hızda (dinlenme durumunda dakikada ortalama 60-80 kez) uyarı çıkarmasıdır. Bu bölge kalbin doğal pili olarak bilinmektedir (pacemaker). Pacemakerın otonom olarak uyarı oluşturmasının nedeni pacemaker hücrelerin potansyum ve kalsiyum karşı ritmik geçirgenlik göstermeleridir.
Faz İsim Olay iyon hareketi 0 Upstroke Hızlı Na kanallarının aktivasyonu (açılması) ve K+
permeabilitesinde azalma Na+ içeri
1 Erken hızlı repolarizasyon
Na+ kanallarının inaktivasyonu ve K+ permeabilitesinde geçici artış
K+ dışarı
2 Plato Yavaş Ca2+ kanallarının aktivasyonu Ca2+ içeri 3 Son repolarizasyon Ca2+ kanallarının inaktivasyonu ve K+ permeabilitesinin
artışı K+ dışarı
4 İstirahat potansiyeli veya diastolik repolarizasyon
Normal permeabilitenin restorasyonu (atrial ve ventriküler hücrelerde) Spontan olarak depolarize olan hücreler içine yavaş Na+ ve olasılıkla Ca2+ sızışı
K+ dışarı Na+ içeri, Ca2+ içeri?
Sinüs düğümünde (Sinoatrial) oluşmuş olan bu uyarı, kalbin her iki kulakçığı(atrium) boyunca, yine bu iş için özelleşmiş iletim yolları ile aşağıya doğru yayılıp bu uyarı ile birlikte kulakçıklar(atrium) kasılarak içlerindeki kanı karıncıklara(ventriküllere) gönderirler. Sonrasında uyarı, kulakçıklar ile karıncıklar arasında bulunan diğer bir özel bölgeye; atrioventriküler (AV) düğüme gelir.

H A Y V A N F İ Z Y O L O J İ S İ
224
ŞEKİL 4.12 Kalpteki ritmin kasılmayı ileten intekalat diskler
Elektrik iletisi karıncıklara ulaştırılmadan önce atrioventriküler düğümde 0.1 saniyelik gecikme kulakçıkların(atrium) karıncıklardan(ventrikülerden) önce kasılmasını sağlar. Böylelikle kulakçıklar ile karıncıkların aynı anda kasılması engellenir. Atrioventriküler düğümden geçen akım, His-Purkinje sistemi ile uyarı tüm karıncıklara yayılır ve karıncıklar kasıldıklarında içlerindeki kanı akciğerlere ve aort yoluyla vücuda pompalarlar. Böylelikle sinüs düğümü yeniden başka bir uyarı çıkarıp başka bir döngü başlatır. Sinoatrial düğüm dakikada ne kadar uyartı çıkartıyorsa (dinlenme durumunda ortalama 60–80 defa), kulakçıklar ve karıncıklar o sayıda sistol(kasılma) yaparlar. Bir kalp vuruşu karıncıkların sistolüdür. Kalp kasındaki uyarılma kasılma eşleşmesinin moleküler mekanizmasını inceleyecek olursak: “Uyarılma-kasılma eşleşmesi” terimi, aksiyon potansiyelinin kas miyofibrillerinin kasılmasını sağlamak için kullandığı mekanizmayı ifade eder. Bu mekanizma iskelet kası ile benzerdir. Kalp kasında bu mekanizmada da farklılıklar olup, bunların kalp kasının kasılma özellikleri üzerinde önemli etkileri vardır. İskelet kası için de geçerli olduğu gibi, bir aksiyon potansiyeli kalp kasının zarı üzerine ilerlerken aynı zamanda transvers (T) tübüllerin zarları boyunca kalp kası lifinin iç kısımlarına da yayılır. T tübüllerindeki aksiyon potansiyelleri longitüdinel sarkoplazmik tübüllerin zarlarını etkileyerek, Ca+2 iyonlarının sarkoplazmik retikulumdan kasın sarkoplazmasına serbestlenmesini sağlarlar. Bu Ca+2 iyonları, serbestlenmelerini izleyen birkaç 1/1000 saniye içerisinde miyofibrillerin içine doğru difüze olur; aktin ve miyozinin birbirleri üzerinde kaymalarını sağlayan kimyasal tepkimeleri katalizleyerek kas kasılmasına neden olurlar. Buraya kadar uyarılma-kasılma eşleşmesinin mekanizması iskelet kasındaki ile aynıdır, ancak çok farklı ikinci bir etki daha vardır. Sarkoplazmik retikulumun sisternalarından sarkoplazmaya serbestlenen kalsiyum iyonlarına ek olarak, aksiyon potansiyeli sırasında T tübüllerinden de sarkoplazmaya kalsiyum iyonlarının difüzyonu gerçekleşir. Bu da T tübülü zarı üzerindeki voltaj-bağımlı kalsiyum kanallarını açar . Hücre içine giren kalsiyum daha sonra sarkoplazmik retikulum zarı üzerindeki riyanodin reseptör kanalı da denilen kalsiyum serbestleyici kanalları(aktifleştirerek sarkoplazma içine kalsiyum serbestlenmesini tetikler. T tübüllerinden gelen bu sitoplazmik kalsiyum kalp kasını daha kuvvetli kasılmasını sağlar. Çünkü kalp kasının sarkoplazmik retikulumu, iskelet kasınınkine kıyasla daha az gelişmiştir ve tam bir kasılma sağlayacak

H A Y V A N F İ Z Y O L O J İ S İ
225
kadar kalsiyum içermez. Diğer yandan kalp kasındaki T tübüllerinin çapı iskelet kasındakilerin 5 katıdır. Bu da, hacminin 25 kat büyük olduğu anlamına gelir. Ayrıca, T tübüllerinin içinde büyük miktarda mükopolisakkarit bulunur ve bunlar elektronegatif yüklüdür ve bol miktarda kalsiyum iyonu bağlayarak aksiyon potansiyeli T tübülüne ulaştığı zaman bu kalsiyum iyonlarını kalp kası lifinin içine difüze olmaya hazır şekilde saklarlar. Kalp kasının kasılma kuvveti, büyük ölçüde, hücredışı sıvılardaki kalsiyum iyonlarının yoğunluğuna bağlıdır. Gerçekte kalsiyum içermeyen bir solüsyon içine yerleştirilen bir kalbin atımı kısa sürede durur. Bunun sebebi şudur: T tübüllerinin uçları hücre zarından geçerek kalp kasını çevreleyen hücredışı alana açıldığı için, kalp kasının interstisyumundaki aynı hücredışı sıvısı T tübüllerinde de dolaşır. Sonuç olarak, T tübül sisteminin içerdiği kalsiyum iyonlarının miktarı (kalp kası kasılmasını başlatmaya hazır kalsiyum iyonları) büyük ölçüde hücredışı sıvının kalsiyum iyonu yoğunluğuna bağlıdır. Bunun aksine, iskelet kasının kasılma kuvveti, hücredışı sıvıdaki kalsiyum yoğunluğunun orta dereceli değişimlerinden hemen hemen hiç etkilenmez. Çünkü iskelet kası kasılmasına neredeyse tamamen iskelet kası lifinin kendi içindeki sarkoplazmik retikulumdan serbestlenen kalsiyum iyonları neden olur. Kalbin aksiyon potansiyelindeki platonun sonunda, kalsiyum iyonlarının kas lifinin içine akışı aniden son bulur ve sarkoplazmadaki kalsiyum iyonları hızla hem sarkoplazmik retikuluma hem de T tübüllerine hücredışı boşluğa geri pompalanır. Kalp kası gevşemesi Ca2+’u sarkoplazmadan temizleyecek olan iki taşıyıcıya dayanır. 1-Sarkolemmadaki Na+- Ca2+ değiştirici sitoplazmada ekstrasellüler sıvıya kalsiyum pompalar, 2-Ca2+’u hücre dışına taşırken SERCA (Ca+2 -ATPaz pompası) Ca2+,u SR’ye döndürür. SERCA pompayı kısıtlayıcı işlev gören fosfolamban isimli, SR membranına gömülü (integral) küçük bir protein ile işbirliği yapar.

H A Y V A N F İ Z Y O L O J İ S İ
226
ŞEKİL 4.13 Kalp kasında kasılma mekanizması.
Fosfolamban fosforile edildiğinde pompayı inhibe etme özelliği azalır ki bu, pompayı daha hızlı çalıştırır. Bunun iki önemli sonucu olur: daha hızlı gevşeme süreleri ve bir sonraki kalp vurusunda salınmak üzere depolanmış Ca2+miktarında artış. Kalsiyum iyonları, ayrıca sodyum-kalsiyum değiştiricisi tarafından da hücreden uzaklaştırılır. Bu değişim sırasında hücreye giren sodyum daha sonra sodyum-potasyum ATPaz pompası tarafından hücre dışına taşınır. Sonuç olarak, yeni bir aksiyon potansiyeli oluşuncaya kadar kasılma durur. Kasılmanın Süresi. Kalp kası, aksiyon potansiyeli başladıktan birkaç milisaniye sonra kasılmaya başlar, aksiyon potansiyelinin son bulmasından birkaç milisaniye sonraya dek kasılmaya devam eder. Dolayısıyla, kalp kasında kasılmanın süresini plato da dahil olmak üzere aksiyon potansiyelinin süresi belirler. Bu süre, atriyum kasında yaklaşık 0,2 saniye, ventrikül kasında ise yaklaşık 0,3 saniyedir. Kalbin tetanik kasılma oluşturamaması, kalp kasının uzun bir mutlak refrakter döneme sahip olmasının bir sonucudur; refrakter dönem, aksiyon potansiyeli sırasında ve sonrasında uyarılmış durumdaki bir hücre zarının tekrar uyarılamadığı bir dönem olarak tanımlanır. Nöronlar ve iskelet kası liflerinde olduğu gibi, asıl mekanizma sodyum kanallarının inaktivasyonudur. İskelet kasının mutlak refrakter dönemi (1-2 ms) kasılma süresinden (20-100 ms) çok daha kısadır ve bu nedenle yapay uyarı ile birincisi tamamlanmadan ikinci bir kasılma oluşturulabilir (kasılmanın sumasyonu). Buna karşın, kalp kasında aksiyon

H A Y V A N F İ Z Y O L O J İ S İ
227
potansiyelinin uzamış depolarizasyon platosu nedeniyle, mutlak refrakter dönemi hemen hemen kasılma süresi (250 ms) kadar sürmektedir ve böylece kas bu sürede sumasyon oluşturabilecek şekilde tekrar uyarılamaz. İskelet kasından farklı olarak, ventrikül kasılmasının herhangi bir derecede sumasyon gösterme yeteneği yoktur ve bu çok iyi/gerekli bir şeydir. Kalp kasının uzamış tetanik kasılmalara uğrayabildiğini bir düşününüz, böyle bir kasılma döneminde ventriküller kan ile dolamazdı -çünkü dolma ancak ventrikül kasının gevşemesi ile oluşabilir. Ve bu durumda kalp bir pompa olarak görevini sürdüremezdi.
Elektrokardiyogram (EKG)
Kalpin sahip olduğu ileti sistemi ve bu ileti sistemine bağlı olarak kalp kasının ritmin kasılması EKG ile belirlebilir. Kalp kasındaki elektirik aktivite kaydına elektrokardiyogram (EKG) adı verilir. EKG nin temel bileşenleri, P dalgası, QRS kompleksi ve T dalgasıdır. P dalgası, sinoatriyal düğümde meydana gelen depolarizasyonu; PR aralığı SA düğümünden AV düğümüne elektiriksel sinyalin iletim süresini ifade eder. QRS kompleksi ventriküler depolarizasyonun sonucudur ve ventriküler kasılmanın başlangıcını gösterir. Bu dalgayı izleyen T dalgası ise ventriküler repolarizasyonun sonucudur ve ventriküler gevşemeyi ifade eder (atriyal repolarizasyon dalgası ise QRS kom pleksi tarafından maskelendiği için gözlenemez). EKG kalbin sadece elektiriksel aktivitesi hakkında bilgi veren bir kayıttır. Elektirik sinyallerinin üretilmesi ve iletilmesinde kesintiler olursa EKG değişir. Bu değişiklikler, kalp içinde olan değişikliklerin gözlenebilmesine yardımcı olur.
EKG temeli kalp kasındaki tüm hücrelerin aksiyon potansiyellerinin vücutta dalgalar halinde yayılmasından yararlanarak kalbin elektriksel aktivitesinin kaydedilmesidir. EKG kalbin mekanik aktivitesinin incelenmesi için uygun bir yöntem değildir. Kalp kası (myokard) kendi başına kasılma özelliğine sahiptir.
ŞEKİL 4.14 Kalp EKG alınmasında kullanılan elektrotların oluşturduğu üçgen(EINTHOVEN ÜÇGENİ)
Kalbin sinüsatriel düğümünden (SA) çıkan uyarılar özel bir iletim yoluyla kalp kası myokard hücrelerine oluşır. Dinlenme durumda polarize halde olan bu hücreler, gelen uyarı ile uyarılarak (depolarize olarak) kasılırlar ve boyları kısalır. Böylece kalp odacıklarını çevreleyen myokardın bütünü büzüşerek içindeki kanı büyük(sistemik) ve küçük(pulmoner) dolaşıma gönderir. Buna kalp kasılması (sistolü) denir. Myokard hücreleri çok kısa süren bu kasılma döneminden sonra hemen eski elektrik yüklerini kazanarak tekrar sakin (polarize) duruma geçerler. Bu olay

H A Y V A N F İ Z Y O L O J İ S İ
228
nabız sayısı kadar tekrarlanır. Nabız sayısı 60 olan kişide bu Depolarizasyon-Repolarizasyon olayı dakikada 60 defa tekrarlanır. Kalbin elektrik faaliyeti ile meydana gelen potansiyel değişiklikleri, kalp çevresindeki dokuların ve özellikle kanın yardımı ile bütün vücuda aynı anda yayılır. Vücudun çeşitli yerlerine konan iletici uçlar (elektrotlar) vasıtasıyla ortaya çıkan elektrik değişiklikleri yükseltilerek kaydedilir. Vücudun çeşitli noktaları arasındaki potansiyel farkları kaydedilir ve o bölgeye göre adlar verilir. Her bir değişik bölge için çizdirilen elektrokardiyogram eğrisine derivasyon denmektedir.
ŞEKİL 4.15 Kalpte EKG elektrokardiyogram
• P dalgası atriyum depolarizasyonunu gösterir. Sinoatriyal düğüm sağ atriyumda olduğu ve sinüs uyarısı da buradan çıktığı için ilk önce sağ atriyum depolarize olur. Bu nedenle P dalgasının ilk kısmı sağ atriyum, ikinci kısmı sol atriyum depolarizasyonunu gösterir. :
• QRS kompleksinin başındaki ilk negatif dalgadır. Her zaman bulunmayabilir. R: QRS kompleksindeki ilk pozitif dalgadır. S: QRS kompleksindeki ilk pozitif dalgadan (R) sonra gelen ilk negatif dalgaya denir. R': QRS kompleksinde, ilk pozitif dalgadan sonra gelen ikinci pozitif dalgadır. Q dalgası ventrikül repolarizasyonu nu gösterir. Bu safhada miyokard hücreleri yeniden negatif yüklerini alır ve tekrar depolarize olmaya hazırlanır.
• Normalde T dalgası QRS kompleksi ile aynı yönde dir. QRS'in pozitif olduğu derivasyonlarda T dalgasının da pozitif, QRS'in negatif olduğu durumlarda T dalgasının da negatif olması beklenir. aVR'de ise T dalgasının negatif olması beklenir.

H A Y V A N F İ Z Y O L O J İ S İ
229
Kalp atımı etkileyen faktörler
Kalp atımı etkileyen temel iki faktör vardır: (1) Kalbe dolan kanın hacmindeki değişikliklere cevap olarak kalbin pompalama işlevinin intrensek düzenlenmesi.
(2) Kalbin pompalama kuvvetinin ve hızının otonom sinir sistemi ile düzenlenmesi. Kalbin pompalama işlevinin intrensek düzenlenmesi Frank-Starling Mekanizması ile açıklanabilir. Kalbin bir dakikada pompaladığı kanın miktarını çoğu zaman, venlerden kalbe dolan kanın miktarı olan venöz dönüş belirler. Yani, vücuttaki her çevre doku, kendi kan akımını denetler ve tüm bölgesel kan akımları toplanarak venler yoluyla sağ atriyuma döner. Kalp ise gelen bu kanı kendiliğinden sistemik arterlere pompalar, böylece kan tekrar dolaşıma katılır. Kalbin gelen kanın hacminde meydana gelen değişikliklere karşı gösterdiği intrensek uyum sağlama yeteneğine, yüzyıl önce yaşamış iki büyük fizyolog, Otto Frank ve Ernest Starling’in anısına, kalbin Frank Starling mekanizması adı verilir. Frank-Starling mekanizması temel olarak, kalp kası dolma sırasında ne kadar çok gerilirse (sarkomer boyu ≈2,2µm), kasılma kuvvetinin ve aorta pompalanan kanın miktarının da o kadar büyük olacağı anlamına gelir. Diğer bir deyişle, fizyolojik sınırlar içerisinde kalp, venler aracılığıyla kendisine dönen kanın tamamını pompalar. Dinlenme halindeki bir kişide, kalp dakikada yalnızca 4-6 litre kan pompalar (atım hacmi x kalp hızı). Ağır egzersiz sırasında, kalbin bu miktarın dört ile yedi katını pompalaması gerekebilir. Kalbin pompaladığı hacmin düzenlenmesi başlıca iki yolla olur: Kuvvetli sempatik uyarılma (örn adrenalin ile), genç erişkin insanlarda dakikada 70 atım olan normal kalp hızını 180-200 hatta nadiren 250 atıma kadar artırabilir. Bunu aksiyon poansiyelinin dinlenim zar potansiyelini kısaltarak yapar. Ayrıca, sempatik uyarılma parasempatik uyarının kalp üzerine olan baskılayıcı etkisini azaltarak da kalp hızını artırır. Diğer taraftan, sempatik uyarılma kalp kasılmasının kuvvetini normalin neredeyse iki katına çıkararak pompalanan kanın hacmini ve fırlatma basıncını da artırırlar. Bu nedenle, sempatik uyarılma daha önce tartışılan Frank-Starling mekanizmasının kalp debisinde neden olabileceği artışa ek olarak, maksimum kalp debisini çoğu zaman iki üç katı kadar daha artırabilir. Bunun aksine, kalbe giden sempatik sinirler inhibe edilerek, kalbin pompalama gücü şu yolla orta derecede azaltılabilir: Kalbi besleyen sempatik sinir lifleri normal koşullarda, pompalama gücünü hiçbir sempatik uyarı olmaksızın gerçekleşecek olanın yaklaşık % 30 üzerinde tutacak şekilde, kalbe yavaş bir hızda ve sürekli uyarılar taşırlar. Dolayısıyla, sempatik sinir sisteminin etkisi normalin altına indiğinde, hem kalp hızı hem de ventrikülün kasılma kuvveti azalır ve kalbin pompalama gücü normalin % 30 kadar altına düşer . ► Termal etki: Sinus venosus bölgesinde sıcaklığın değişmesi kalp atım frekansını etkiler. Sıcaklık artarsa kalp atım frekansı yükselir, kasılma gücü azalır. Sıcaklık azalırsa kalp atım frekansı azalır, kasılma gücü artar. Termal etkinin myokard üzerindeki proteinlerin özellikle iyon kanalların üzerinde etkisi vardır. Vücut sıcaklığında artış, örneğin kişinin ateşi yükseldiği zaman gözlenen sıcaklık artışı, kalp hızının büyük oranda artmasına neden olur (1°C’lik artış kalp hızını 18 atım/dk artırır), hatta bazen normalin iki katına çıkarır. Düşük sıcaklık kalp hızını büyük oranda azaltır, öyle ki bir kişinin vücut sıcaklığı 15,5-21,1 °C arasında olup hipotermi nedeniyle ölüme yaklaştığı zaman, kalp hızı dakikada birkaç atıma kadar düşer. Bu etkiler olasılıkla, ısının kalp kası zarının kalp hızını belirleyen iyonlara geçirgenliğini artırarak, kendi kendine uyarılma sürecini hızlandırmasına bağlıdır. Sıcaklığın orta derecede artması, kalbin kasılma kuvvetini çoğu zaman (egzersizde olduğu

H A Y V A N F İ Z Y O L O J İ S İ
230
gibi) geçici olarak artırır, fakat sıcaklığın uzun süre yüksek kalması, kalbin metabolik sistemlerini tüketerek zamanla güçsüzlüğe neden olur
► Kimyasal uyarıcılar: Adrenalin kalp atım frekansını ve genliğini artarır. Adrenalin kalp kasılmaları β1 adrenerjik reseptörler üzerinden etkiler. Asetilkolin kalp atım sayısını azalır. Asetilkolin Muskarinik reseptörler yoluyla K+ geçirgenliğini arttırır.
ŞEKİL 4.16 Kalpte easetil koline bağlı yavaşlama mekanızması
Membranın eşik değere ulaşma süresi uzar. (Vagusun şiddetle uyarılması spontan deşarjları bir süre için duraklatır.) N. vagusun inhibitör etkisi kalbi durdurur. Uyarmaya devam edilirse, 15–20 san içinde ventriküllerin, Purkinje liflerinden doğan eksitasyonlarla tekrar faaliyete başladığı görülür. Bu olaya Vagustan kurtulma (vagal escape) denir. Bu anda ventriküller Purkinje liflerinin ritminde (15–40/dak) çalışır. Parasempatik sinirler tamamen çıkarıldığı zaman, normalde ortalama 70-80/dak. olan kalbin vurum sayısı 160/dak. kadar yükselir. Asetilkolin muskarinik ( M2 ) üzerindeki etkisi aracılığıyla kalp kasılmalarındaki frekans ve genliği kolinerjik reseptörlerle atırır. Kısaca kalp sempatik ve parasempatik uyarılara tepki verir.
ŞEKİL 4.17 Kalpte nör epinefrine bağlı kasılmanın ritminin ve gücünün artışı

H A Y V A N F İ Z Y O L O J İ S İ
231
► Ekstrasellüler sıvıdaki iyonlarının konsantrasyonu: hücre dışı Ca2+konsantrasyonunun arttırılması kasılma gücünü artırır. Hücre dışı K+
konsantrasyonu, seçici olmayan bir şekilde, dış porları Ca2+ ve Na+karşı bloklar. Böylece kasılmaların genliği azalır. Kalp diyastol (gevşeme) durumumda durur. K+ konsantrasyonu artışı (hiperkalemi), kalp hızının yavaşlamasına neden olur. Kalp aşırı genişleyerek diyastolde durur.
► Potasyum İyonlarının Etkisi. Hücredışı sıvılardaki aşırı potasyum, kalbin fazlasıyla geniş ve gevşek hale gelmesine ve kalp hızının yavaşlamasına neden olur. Büyük miktarlar, aynı zamanda kalp uyarısının A-V demeti yolu ile atriyumlardan ventriküllere iletilmesine de engel olabilir. Potasyum yoğunluğunun yalnızca 8-12 mEq/L’ye (normal değerin iki veya üç katı) yükselmesi, kalbi öylesine zayıf düşürebilir ve ritmini bozabilir ki, ölüme neden olabilir. Bu etkiler, kısmen hücredışı sıvılardaki yüksek potasyum yoğunluğunun kalp kası liflerinin dinlenim zar potansiyelini azaltmasına bağlıdır. Yani, yüksek hücredışı sıvı potasyum yoğunluğu, kısmen hücre zarını depolarize ederek zar potansiyelinin daha az negatif olmasına neden olur. Zar potansiyeli azalınca aksiyon potansiyelinin şiddeti de azalır, ki bu kalp kasılmasını giderek daha zayıf düşürür. ►Kalsiyum İyonlarının Etkisi. Artan kalsiyum iyonları, potasyum iyonlarının tam tersi etkiler yaparak, kalbi spastik kasılmaya doğru götürürler. Bunun nedeni, kalsiyum iyonlarının kalp kasılmasının başlatılmasında doğrudan etkili olmasıdır. Bunun aksine, kalsiyum iyonlarının eksikliği, yüksek potasyumun etkisine benzer şekilde, kalbin gevşemesine neden olur. Neyse ki, kalsiyum iyonlarının kan düzeyi normalde çok dar sınırlar içerisinde tutulur. Dolayısıyla, normalin dışındaki kalsiyum yoğunluklarının kalbe etkileri nadiren klinik önem kazanır.
► Digitalis, Digitalis purpurea yapraklarında elde edilen kardiotonik steroidleridir. Kalp atım gücünü özellikle kalp krizlerinde artıracak yönde etki gösterir. Etki mekanizması kalp kasılda myokard bulunan Na-K ATPaz ları inhibe ederek gösterir. Bunun sonuncunda myokarta hücre içinde daha fazla Na kalır. Hücre içi Na seviyesi artar. Na bağlı olarak kalp hücresindeki kalsiyum seviyesi artar bunu sonuncunda kalp daha güçlü kasılır.
Stannius bağları
Kalp kasılmalarının kendiliğinden ve belli bir bölgeden başladığına ait ilk deliller Stannius’un yaptığı deneylerle ortaya konduğu için bu deneylere stannius deneyleri denir. Kurbağa kalp kolay elde edilemesi nedeniyle eğitim amacıyla sıklıkla kullanılır. Kurbağada perikard kesesinin açılması kalpten daha fazla kan pompalanmasını sağlanır. Perikardın açılmasının kalp üzerinde etkisi Frank - Starling mekanizması açıklanır. Frank - Starling mekanizması kalpe gelen kan miktarı ile kalpten çıkan kan arasınadki ilişki ortaya koyar. Kısaca “ne kadar gelirse o kadar gider”.
Stannius bağları kurbağa kalbinde impuls iletimini göstermek için yapılana deneysel bir işlemdir. Kurbağada SA düğüm yok. Bunun yerine sinus venosus vardır. Sağ atriumla sinus venosus arasından bir ilmek atıldığında (1. stannius bağı), sinus venosus vurumlarına devam eder, atriumla ventrikül vuruşları durur (en az bir dakika kadar). Daha sonra atriumlarla ventrikül sinus venosustan ayrı bir ritimde çalışmaya başlayacaktır. Bu yeni ritmin impulslarını atriumun kas hücreleri yaratmaktadır. Atriumlarla ventrikül arasında 2. bir bağ atılırsa (2. stannius bağı), kısa bir duraklamadan sonra ventrikül kasılmaya başlar. Ancak sinus venosustan daha yavaş atar. Böylece 2 düğümle kalbin 3 ayrı bölgesi farklı bir şekilde çalışır.

H A Y V A N F İ Z Y O L O J İ S İ
232
ŞEKİL 4.18 Kurbağa kalbinde I ve II Stannius bağı
Memeli kalpinde Stannius bağı işlemi yapılamaz çünkü bağ kroner damarlarda kan akımı engeller.
Kan basınçı
Kalb-damar sisteminin değişik bölümlerinde kan basınçları farklıdır. Aortada yüksek bir basınç vardır (120 mm Hg sistolde, 80 mm Hg diastolde). Kanın vena kava’dan sağ atriuma dönerken hemen hemen basınçı 0 mm Hg’ kadar düşer. Kapilların; arteriyel yarımında 35 mm Hg, venöz yarımında ise 10 mm Hg kadar basınç vardır. Akciğerlerde ise düşük basınç bulunur; sistolik basınç 25 mm Hg, diastolik basınç 8 mm Hg dir. Böbrekler kalpten gelen kanın yüzde 21 kısmını alır küçük bir organ olan böbrekler en fazla kan alan organların başındadır. Bu nedenle böbreklerin fonksiyonel birimi olan nefronlara gelen afferent ateriollerdeki kan basınçı ortalama arter kan basınçının yüzde 60’na 60 mmHg kadardır. Kan basınçı kalbin konumuna göre de değişir. Beyin kalbe göre daha yukarıda olduğu için daha fazla güç gerektirirken (daha az basınç) kalbin altındaki dokulara kan pompalamak daha az enerji gerektirir( daha fazla basınç).

H A Y V A N F İ Z Y O L O J İ S İ
233
ŞEKİL 4.19 Korotkof sesleri
� Sistolik basınç kalp kasılıp kan arteryel sisteme fırlatıldığında oluşur. Kalp döngüsü boyunca en yüksek arteryel basınca sahiptir. Sistolik basınç, kalbin kanı sistemik dolaşıma fırlatması sırasında oluşur. Bu nedenle sistolik basınç debi ile ilişkilidir. Formül olarak debi ve periferik rezistansın çarpımına eşittir.(Debi x Periferik Rezistans)
� Diastolik basınç Kalbin gevşeyip kanın venler yoluyla kalbe döndügü sırada oluşur. Bu fazda aort kapağı kapalıdır. Diastolik basınç sadece periferik rezistansla ilişkilidir. Arter esnek kalın duvarları diastolik basınç oluşumu sağlar.
� Ortalama arter basınçı: Bir kardiyak siklus süresinde aorta ve proksimal arteriyel sistemdeki ortalama kan basıncının ölçütüdür (venöz basınç gözardı edilmiştir). Sistole göre diastol süresi daha uzun olduğundan (Ortalama Arter Basınçı) MAP, diastolik kan basıcına daha yakındır. Kalp Debisi periferik kan akım hızından daha yüksek oldukça MAP artar. Sistolik ve Diastolik basınçların ortalam arter basınçına etkisi farklıdır. Çünkü diyastol süresi sistol süresinin yaklaşık 2 katı kadardır. Bu nedenle diyastol basıncının ortalama basınca katkısı daha fazla olur.

H A Y V A N F İ Z Y O L O J İ S İ
234
ŞEKİL 4.20 Tansiyon ölçülmesi kan basınç değişimi
Damar sisteminde esas olarak rezistansı belirleyen damar arterioldür. Arteriol çapı küçüklükçe damar rezistansı artar ve dokuya giden kan akımı azalır. Ancak kan basıncı arttığı için diğer dokuların perfüzyonu artar. Arteriolde rezistans çok yüksek olduğu için kan basıncı aniden azalma gösterir. Arteriol sonrası kapiller sistemde basınç çok düşüktür. (0–4 mm-Hg) Kapiller sistemde akım süreklidir. Kılcal damarlarda sistol ve diyastol basınç farklılığı gözlenmez.
Dolaşım sistemi
Dolaşım kalp ile akçiğerler arasında ve vucud ile kalp arasında kanın sürekli olarak pompalanmasıdır. Arterler kalpte uzaklaşan kan arterioller ile daha küçük dallara ayrılır, fonksiyonel birim olan kapillerden dolaşımın en ince ve uzun kısmıdır, kan daha sonra venüller ve venlerle daha kalın damarlara ve en sonunda da kalp ulaştırılır. Kapiller damarlarda sızmış olan doku sıvısı ve geçer proteinler lenf sistemi ile toplananarak yine venler aracılığı ile dolaşıma katılır. Temel olarak iki dolaşım vardır, sistemik ve pulmoner.
MAP = DKB + (SKB - DKB ) / 3 Normal MAP 70-100 mm Hg.(DKB Diastolik kan basınçı, SKB Sistolik kan basınçı)

H A Y V A N F İ Z Y O L O J İ S İ
235
ŞEKİL 4.21 Vücuttaki başlıca bölmeler ve basınçları
Sistemik Dolaşım: Sistemik dolaşım kalp ile vücud arasında(akçiğerler hariç) olan dolaşımdır. Vena pulmonalislerle sol atriuma gelen arterial kan buradan sol ventriküle ulaşır. Aorta yolu ile dokulara pompalanır. Daha sonra venöz kan olarak vena cavalar ile sağ atriuma gelir.
Tablo kalpten pompalanan kanın dağılımı
Organ Miktar (ml/dakika) Yüzde(%)
Beyin 650 %13 Kalp 215 %4 Kas 1030 %20 Deri 430 %8 Böbrekler 950 %20 Abdominal 1200 %4 Diğer organlar 525 %10 Toplam 5000 %100 Memelilerde sol ventrikülde temiz kan bulunur. Buradan çıkan aort ikiye ayrılır. Biri üst bölgelere (başa) giden karotid arteri oluşturur, diğeri ise vücudun alt kısımlarına gider. Aort sırasıyla büyük arterler, küçük arterler, sistemlere giden arterioller sistemlerin içine giren kapilerlere değişir. Kapilerlerde kan ve vücut sıvıları arasında madde değişimi olur. Yani besin maddeleri, solunum gazlarının değişimi ve artık maddelerin değişimi kapiler duvarlarında olur. Kapilerlerin iki ucu vardır. Bu uçlardan biri arteriolle, diğer uç ise venül (en küçük ven) ile bağlantılıdır. Arterlerle vücuda dağılan temiz kan kirlendikten sonra venlerle sağ atriumdan kalbe döner. Kapilerlerde madde değişimi olduktan sonra kan, venül, küçük venler, büyük venlerde toplanarak vena cava inferiör ile sağ atriumdan kalbe döner, oradan da sağ ventriküle geçer. Karotid arter ile baş bölgesine giden temiz kan burada kirlendikten sonra vena cava superior ile sağ atriuma kirlenmiş olarak döner. Yani sağ atrium ve ventrikülde kirli kan bulunur.

H A Y V A N F İ Z Y O L O J İ S İ
236
ŞEKİL 4.22 Fetal ve normal dolaşım
Pulmonar dolaşım
Pulmonar dolaşım kalpin sağ atriumla başlayan ve sol ventrikülde sonlanan kalp ile akçiğerler arasındaki düşük basınçlı dolaşımdır. Sağ atriuma gelen venöz kan triküspid kapak yoluyla sağ ventriküle geçer. Arteria pulmonalis(akçiğer atardamarları) ile akciğerlere ulaşır. Akciğerlerde gaz değişimine uğradıktan sonra vena pulmonalislerle(akçiğer toplar sistemik dolaşıma katılacak temiz arterial kan olarak sol atriuma(sol kulakçığa) döner. Pulmoner arter ve dallarının: duvarları çok ince ve az sayıda düz kas bulunur, basınç düşüktür, bu nedenle damar içi basınç değişiklikleri pulmoner dolaşımda çok önemli etkiler yapar. Akciğer dolaşımında kanın çıkması gerekli en yüksek bölge apekslerdir. Buraya arter kanının çıkması için bu basınç yeterlidir, fazlasına gerek yoktur. pulmoner arter basıncı 6 lt/dak kan akımı için ortalama 15-18 mmHg(sistolik 25mmHg ,diyastolik 8 mmHg), basınç çok pulsatildir(rirmin dalgalanma gösterir). Akciğer damarlarındaki basıncın dağılımı büyük oranda akciğer volümüne bağlıdır. Pulmoner dolaşım temel fonksiyonu kanın dış ortamla gaz değişimin yapılmasıdır. Akciğer dolaşımı sağ ventrikülden venöz kanı taşıyan ana pulmoner arterle başlar, pulmoner arterler birbiri arkası bronşlar gibi dallanarak arterioller olarak terminal bronşiollere kadar bronşlara eşlik eder, sonra bölünerek alveol duvarlarında kapiller yatağı oluştururlar. Akciğer kapillerleri alveol duvarında gaz alışverişi için çok elverişli bir düzen sağlayan yoğun bir ağ oluştururlar. Buradan çıkan oksijenlenmiş kan lobüller arasında yol alan küçük pulmoner venlerde toplanır. Bunlarda birleşerek 4 ana ven halinde sol atriuma açılır. İlk bakışta bu dolaşım aortadan başlayan ve sağ atriumda sonlanan sistemik dolaşımın küçük bir örneğine benzetilebilir. Bu nedenle akciğer dolaşımına sıklıkla "küçük dolaşım" adı da verilir.

H A Y V A N F İ Z Y O L O J İ S İ
237
Kan damarları
Kan damarları dolaşım sisteminin organlarındandır. Görevi kanı vücudun farklı bölümlerine taşımak olan kan damarlarının farklı türleri vardır. Temel kan damarı tipleri atardamarlar (arter) ve toplardamarlardır (ven). Atardamarlar kanı kalpten alıp vücudun farklı bölümlerine taşırken, toplardamarlar vücudun farklı bölümlerinden kanı kalbe taşırlar. Bununla birlikte iki istisna mevcuttur: pulmoner arter kirli kan, pulmoner ven ise temiz kan taşır. Vücuttaki en büyük damar kanın kendisi aracılığıyla tüm vücuda doğru pompalandığı aort atardamarıdır.
ŞEKİL 4.23 Damarlar ve dolaşan kan oranı
Vücutta bulunan her organın en az bir tane temiz kanı kalpten getiren ve birden fazla kirli kanı kalbe götüren damarı vardır. İnsan vücudundaki damarların toplam uzunluğu 100.000 km kadardır. Kan damarı hsitolojik olarak üç tapakadan yapılmıştır:(İntima Media Ve Adventisya).
1. İntima: - Endotelium: Bazal membran üzerine oturmuş tek katlı yassı endotel hücreleri. Lamina elastica interna: Çok incedir.
2. Media: - 1–2 sıralı, sirküler seyirli düz kas tellerinden yapılmıştır. Çapı küçüldükçe kas tek sıralı hale dönüşür.
3.Adventisya : Çok ince bağ dokusudur, lamina elastica externa bulunmaz.
Arteriol’un çapı l0 mikronun altına düşerse prekapiller arteriol ya da metarteriyol adı verilir. Bunların duvarı sadece endotel hücreleri ve tek sıra düz kas tellerinden ibarettir.
Kan damarları aktif biçimde kanın taşınmasında yer almazlar (fark edilebilecek peristaltizme sahip değillerdir), fakat arterler - ve bir seviyeye kadar venler - kas tabakasının kasılması suretiyle kendi iç çaplarını kontrol edebilirler. Bu da organlara akan kan miktarını etkiler ve otonom sinir sistemi tarafından kontrol edilir. Ayrıca vazodilasyon ve vazkonstriksiyon termoregülasyon teknikleri olarak antagonistik biçimde (yani sıcaklıktaki değişikliğe karşı olarak) gerçekleşir. Kan tarafından taşınan en önemli besin kırmızı kan hücrelerineki hemoglobine bağlanarak taşınan oksijendir. Pulmoner arter dışındaki tüm arterlerde, hemoglobin yüksek oranda (%95-%100) oksijene doymuştur. Pulmoner ven dışındaki tüm venlerde ise, hemoglobin yaklaşık %70 seviyesinde doymamış hâle gelir. (Değerler pulmoner
• İntima
• media
• adventisya

H A Y V A N F İ Z Y O L O J İ S İ
238
dolaşımda terstir.) Vazokonstriksiyon kan damarlarının, duvarlarındaki vasküler düz kasın kasılmasıyla, konstriksiyonu yani kısılması, enine kesit alanının küçülmesidir ve vazokonstriktörler tarafından kontrol edilir. Bunlara parakrin etmenler ve nörotransmitterler dahildir. Benzeri bir mekanizmayla, tersi olan vazodilasyon da kan damarları tarafından gerçekleştirilebilir. Vazodilatörlerce kontrol edilen vazodilasyonda(damarların gevşemesi), iç çap genişletilir. En önemli vazodilatör nitrik oksittir(en güçlüsü ürotensin).
Kapiller kan damarları
Dolaşım en geniş ve uzun kısmı kapiller damarla olur. Dolasımın en önemli fonksiyonu besin maddelerinin dokulara tasınması ve hücresel atıkların uzaklastırılmasıdır. Gecirgenligi fazla, tek sıra endotel hücresinden meydana gelir. Kapiller çok ince yapılardır; (0.5 µm) Ortalama kapiller çapı: 4-9 µm olup 10 milyar kapillerin yüzey alanı: 500-900 m2 kadardır. İnsanda fonksiyonel hiçbir hücre kapillerden 20-30 µm den uzak olamaz. Kapiller dokulara göre özelleşmiştir. Beyinde endotel hücreleri arasındaki sıkı bağlantılar (tight junctions) sadece küçük moleküllerin beyin dokusuna geçişine izin verir. Karaciğerde tamamen tersi bir durum söz konusudur. Kapiller endotel hücreleri arasındaki yarıklar geniş bir açıklık gösterdiği için plazmada erimiş halde bulunan bütün maddeler (plazma proteinleri de dahil olmak üzere) kandan rahatlıkla karaciğer intertisyel aralığına geçebilir.
ŞEKİL 4.24 Kılcal damar ağının akım kontrolü ve basınç değişimi
Barsak kapillerlerdeki porlar ise kas ve karaciğer dokusu arasında bir yer almaktadır. Böbreğin glomerüler yumağında bulunan çok sayı da küçük oval pencereler; endotel hücresini ortadan penetre ederek büyük miktarda maddenin endotel hücreleri arasındaki yarıklardan geçmeden glomerülerden filtre olmasını sağlar.
ŞEKİL 4.25 Kılcal damarda madde taşıması

H A Y V A N F İ Z Y O L O J İ S İ
239
Kapillerlerin görevi sıvı, besin maddeleri, elektrolitler, hormonlar ve diğer maddelerin kan ile interstisyel sıvı arasında değişimini sağlamaktır. Bu göreve uygun olarak, kapiller çeperi çok incedir ve çok sayıdaki kapiller porlar su ve küçük moleküllü maddelere geçirgendir. Prekapiller sfinkter bölgesinin kas tabakası sinirsel inervasyonu yoktur. Prekapiller sfinkter çapı doku faktörlerinin miktarına baglı değişir. (Potasyum, Adenozin(ATP), Lokal CO2
miktarı , Hipoksi, Laktat miktarı gibi).
ŞEKİL 4.26 Damarda vazodilasyon.
Kapillerde madde taşıma yolları
Dolaşım sistemin yüzde 99 kısmı mikro dolaşım yapan kapiller dolaşıımdan oluşur.Kapiller dolaşım kan ile doku arasında madde alış verişin yapıldığı dolaşım kısmıdır.
ŞEKİL 4.27 Damarda vazodilasyonla basınç dengesi.
Kapiller kan damarı temel olarak 3 tiptir. 1.Çeperleri sürekli (penceresiz) kapillerler Sıvı ve yağda eriyen madde filtarsyonuna izin verir.
2.Çeperleri pencereli kapillerler Böbrek glomerulus, iç-dış salgı bezleri, pleksus koroideus, ince barsak villuslarında bulunur.

H A Y V A N F İ Z Y O L O J İ S İ
240
3.Kesintili çeperli kapillerler (sinüsoidler) Karaciğer, dalak, büyük moleküller (proteinler, lipit ve sakkaritler), eritrositlerde bulunur yüksek seviyede madde geçisi izin verir.
Maddelerin plazma ile interstisyel sıvı arasındaki en önemli geçiş yolu; difüzyondur. Hidrostatik basınç (P), Osmotik basınç (P) farkı ile gerçekleşir. Bir maddenin net difüzyon hızı,
membranın iki tarafı arasındaki konsantrasyon farkı ile doğru orantılıdır; difüzyon için %1 lik fark bile yeterlidir.
Kapillerin Fonksiyonları:
Kapillerlerin en önemli fonksiyonlarını üç alt başlık altında incelemek uygundur.
1. Geçirgenlik (Permeabilite): Kapillerler doku ile kan arasında oksijen, karbondioksit ve çeşitli metabolitleri geçişim bölgeleridir. Bu değişim gerçeğinin henüz tam olarak mekanizması aydınlatılmamış olsa da, üzerinde uzlaşılan bu işlemin üç olası mekanizmayla gerçekleştiğidir. Bunlar; a.Komşu endotel hücreleri arasındaki yarıklar, b.Pencereli kapillerdeki geçiş delikleri ve c. Endotel hücrelerini boydan boya kat eden pinositotik veziküllerdir.
2. Metabolik İşlevleri: Kapiller endotel hücreleri oldukça geniş bir işlevsel kapasiteye sahiptir. Anjiyotensin I’in anjiyotensin II’ye dönüşmesi, b. Bradikinin, seratonin, PG ler, nörepinefrin, trombin’in biyolojik olarak reaksiyon vermeyen bileşiklere dönüştürülmesi,c. Lipoproteinlerin, endotel hücrelerinin yüzeyindeki enzimler tarafından Trigliseritleri ve kolesterole parçalanması ve kapiller damarların örtüsü olan endotelin damar tonusu üzerinde etkisi olduğu düşünülen bir takım vazokonstriktif (Endotelin) veya vazodilatatör (NO) sentez ettiği bildirilmektedir.
3. Antitrombojen İşlevi: Endotel hücreleri döküldüklerinde, açıkta kalan subendotelyal bağ dokusu trombositlerin agregasyonuna sebep olur. Endotel hücreleri kanın subendotelyal bağ dokusuyla temasını önleyerek antitrombojenik etki gösterir.
Vazodilatör vazodilatasyona neden olan yani kan damarlarının genişlemesini sağlayan sinir veya (dış) ajanlara verilen isimdir. Vazodilatasyon damar duvarındaki çizgisiz kasın gevşemesiyle damarın genişlemesidir. Böylece, kan akışı için daha fazla yer açılır, kan basıncı (yani tansiyon) düşer. Vazodilatasyon işlemini kontrol eden kas ve sinirlere vazomotor denir. Vazodilatörler dokuların beslenmesini ve beyne giden kan akımını artırdıkları gibi, yüksek dozda kullanıldıklarında dolaşım yetersizliğine de yol
1. Filtrasyon 2. Difüzyon 3. Mikropinositoz

H A Y V A N F İ Z Y O L O J İ S İ
241
ŞEKİL 4.29 Damarda vazodilasyon.
Kan akımı düzenlenmesi
Dolaşımın en temel kurallarından biri her dokunun kendi kan akımını metabolik ihtiyaçlarına göre kendisinin belirlemesidir. Lokal kan akımların toplamı dolaşım hızı ve basınç ayarlnmasını sağlar. Dokudaki kan akımını belirleyen başlıca faktörler:
1. Oksijenin dokulara taşınması,
2. Glikoz, amino asitler, yağ asitleri gibi besin maddelerinin dokulara taşınması,
3. Karbondioksidin dokulardan uzaklaştırılması,
4. Hidrojen iyonlarının dokulardan uzaklaştırılması,
5. Artık maddeleri uzaklaştırmak,
6. Dokulardaki diğer iyonların uygun konsantrasyonlarda sürdürülmesi,
7. Çeşitli hormonların ve diğer moleküllerin farklı dokulara taşınması. Belirli organlar kan akımına özel gereksinim gösterirler. Örneğin, derinin kan akımı vücuttan ısı kaybını belirleyerek vücut sıcaklığının kontrolünü sağlar. Ayrıca yeterli miktarda plazmanın böbreklere taşınması sayesinde vücuttaki atık maddelerin böbreklerden atılması, vücut sıvı hacminin ve elektrolitlerin seviyelerinin düzenlenmesi sağlanır. Arteriyoller (direnç damarları) kardiyovasküler sistemin kilit noktalarında yerleşiktir ve bunun sonucu olarak çok sayıda denetim mekanizmasına tabiidirler. Kabul edilen iki genel kontrol mekanizması bulunmaktadır:
1- akut kontrol ve 2- uzun süreli kontrol.
Akut kontrol
Organ ve dokuların kendi arteriyol dirençlerini kendi kendilerine değiştirmesi, böylece kan akımlarını kendi kendine düzenleyen, sinir veya hormonlardan bağımsız bir mekanizmayı gösterir. Bu otokrin/parakrin ajanlar tarafından oluşturulan değişiklikleri kapsar.

H A Y V A N F İ Z Y O L O J İ S İ
242
Akut kontrol arteriyoller, metarteriyoller ve prekapiller sfinkterlerin yerel vazodilatasyon ve vazokonstriksiyonundaki hızlı değişikliklerle gerçekleşir ve yerel doku için gerekli kan akımını seri bir şekilde sağlamak üzere dakikalar veya saniyeler içinde gerçekleşir. Doku metabolizması, miyojenik yanıt, oksijen miktarı değişikliği, oksijen yokluğu teorisi, vazodilatör teoriler burada önemli role sahiptirler. Arteriyoller tarafından doku kan akımının kendi kendine düzenlemesi aktif hiperemi fenomeni, akım otoregülasyonu, reaktif hiperemi ve hasar için oluşan lokal cevabı içerir.
Aktif Hiperemi :Birçok organ ve dokunun metabolik aktiviteleri arttırıldığı zaman belirgin olarak artmış kan akımı (hiperemi) ortaya çıkar, bu durum, aktif hiperemi olarak isimlendirilir. Yani doku aktivitesi arttığı zaman görülen normal vazodilatasyon cevabıdır. Örneğin, kasın artmış aktivitesi ile doğru orantılı olarak egzersizde iskelet kaslarının kan akımı artar. Aktif hiperemi çoğu aktif organ veya dokuda doğrudan arteriyol dilatasyonun sonucudur. Aktif hiperemide arteriyol düz kas gevşemesine neden olan faktörler arteriyollerin etrafındaki hücre dışı sıvıda bulunan lokal kimyasal değişimlerdir. Bunlar, arteriyol yakınında bulunan hücrelerdeki artmış metabolik aktiviteden oluşur. Tutulan organlara ve artmış aktivitenin süresine bağlı olarak, farklı faktörlerin göreceli katkılarını kapsar. Belki en belirgin değişim, oksidatif fosforilasyonla ATP’nin üretiminde kullanılan oksijenin lokal yoğunluğunun azalması ile dokular daha aktif hale gelince oluşur. Metabolizma kan akımını arttırdığı zaman diğer kimyasal faktörlerin sayısı artar, Aktif hiperemiye aracılık eden kimyasal değişikliklerin bazıları:
a) Azalmış oksijen O2,
b) Artmış CO2 (oksidatif metabolizma son ürünü), H+ (laktik asitten), K+ (tekrarlayan aksiyon potansiyeli repolarizasyonunun birikiminden) ve adenozin (ATP’nin yıkım ürünü)
c) Yüksek moleküler ağırlıklı maddelerin artan yıkımları sonucunda osmolarite artışı,
d) Eikosonoid (zar fosfolipidlerinin yıkım ürünü) konsantrasyonu artışı,
e) Bradikinin (aktif bez hücrelerinden salınan kallikrein enziminin aktiflenmesi ile kininojen olarak adlandırılan dolaşan bir proteinden lokal olarak oluşan bir peptit) yapımının artması.
Tüm bu kimyasal faktörlerdeki lokal değişikliklerin, kontrol edilen deneysel koşullarda arteriyol dilatasyona neden olduğu gösterilmiştir ve bunların hepsi olasılıkla bir veya daha çok organda aktif hiperemi cevabına katkıda bulunur. Hücre dışı sıvıdaki bütün bu kimyasal değişimler lokal olarak arteriyol düz kasa etki ederek gevşemesine(vazodilasyona) neden olur. Sinir ve hormonları içermezler. En fazla iskelet kası, kalp kası ve bezlerde, dokularda gelişen aktif hipereminin, vücuttaki en geniş metabolik aktivite alanı göstermesi şaşırtıcı olmamalıdır. Bu oldukça etkilidir, bu yüzden, onların kan ihtiyacı esas olarak lokal olarak sağlanır.
Akım Otoregülasyonu :Otoregülasyon bir organın kan akımını perfüzyon basıncındaki değişikliklere rağmen içsel olarak sabit tutabilme yeteneğidir. Arteriyel basınç ani olarak artarsa, akım da artar. Metabolitler üretildiklerinden daha yüksek bir hızla uzaklaştırılırlar ve direnç damarları da refleks olarak kasılırlar. Aktif hiperemi sürecinde, doku veya organın

H A Y V A N F İ Z Y O L O J İ S İ
243
artmış metabolik aktivitesi lokal vazodilatasyona yol açan başlangıç bulgusudur. Yine de, bir doku veya organ, kan basıncında değişimin neden olduğu, kan miktarındaki bir değişimden zarar görürse, lokal olarak arteriyol direncine aracılık eden değişimler de meydana gelebilir. Dirençteki değişim, kan akımını, basınç değişikliğinin cephesinde neredeyse sabit tutacak şekilde koruma yönündedir ve bu yüzden akım otoregülasyonu olarak isimlendirilir. Örneğin, bir organda arteryel basınç düştüğü zaman, nispeten sabit akımın devamını sağlama eğilimi olarak arteriyol vazodilatasyona neden olan lokal kontrollerin, organa akım sağlayan arterdeki kısmi daralmadan ötürü olduğu söylenir. Akım otoregülasyonun mekanizması aktif hiperemi için tanımlanan aynı metabolik faktörleri içerir. Bu, her iki durum kan ihtiyacı ve hücre metabolik aktivitesinin düzeyi arasındaki başlangıç dengesizliğini yansıttığı için böyledir. Dikkat edilecek bir nokta; aktif hipereminin ve düşük arteryel basınca cevapta oluşan akım otoregülasyonunun vazodilatasyonu, lokal metabolik faktörleri içeren temel mekanizmalarındakinden farklı değildir, fakat değişmiş metabolizma veya değişmiş kan basıncı durumlarında bu mekanizmalar oynama gösterir. Akım otoregülasyonu arteryel basınçtaki azalma durumu ile sınırlı değildir. Çeşitli nedenlerden ötürü tersine durumlar meydana geldiği zaman, arteryel basınç artar: Başlangıçta, lokal vazodilatatör kimyasal faktörler üretildiklerinden daha hızlı bir şekilde basınçtaki artışla uzaklaştırıldıklarından ve aynı zamanda oksijenin lokal konsantrasyonu arttığından ötürü akım artar. Bu durum arteriyollerin daralmasına neden olur, böylece artmış basınç cephesinde nispeten sabit lokal akım sürdürülür.
Akım otoregülasyonundaki diğer bir mekanizma da miyojenik yanıttır. Bu yanıt, dolaşımda intraluminal basınç arttığı zaman refleks olarak direnç damarları kasılır. Kasılma, damar düz kas hücre zarlarında bulunan ve gerimle aktive olan Ca+2 kanalları üzerinden gerçekleşir ve arteriyel basınçta görülen dalgalanmalara karşı kapilleri korur. Örneğin postür değişiklikleri yerçekiminden dolayı alt ekstremine damar yatağında basıncın ani olarak 200 mmHg'nın üzerine çıkmasına engel olur. Tersine, arteryel basıncın azalmasının neden olduğu azalmış gerim, bu damar düz kaslarının tonusunun azalmasına neden olur.
Kan Akımının Uzun Süreli Kontrolü
Akut düzenlemeye ilave olarak, saatler, günler ve haftalar içerisinde uzun süreli yerel kan akımı kontrol mekanizmaları gelişmektedir. Uzun süreli kontrol, akut mekanizmalara göre tama çok daha yakın bir düzenleme sağlar. Örneğin, arter basıncı aniden 100 mmHg değerinden 150 mmHg değerine yükseldiğinde kan akımı da aniden % 100 oranında artar. Bunu izleyen 30 saniye ile 2 dakika içinde kan akımı düşerek orjinal kontrol değerinin ancak % 10-15 fazlası olacak düzeye iner. Eğer arter basıncı değeri sürekli bir şekilde 150 mmHg değerinde kalırsa birkaç hafta içinde dokulara giden kan akımının tamamen normal düzeyine doğru geri döndüğü görülecektir.
Reaktif Hiperemi :Bir organ veya dokunun kan ihtiyacı kısa bir süre tümüyle bloke olduktan sonra tekrar kanlandırılırsa dokuya giden kan akımı normale göre 4-7 kat artabilir. Reaktif hiperemi olarak bilinen bu fenomen aslında akım otoregülasyonunun aşırı bir şeklidir. Kan akımı olmadığı dönem sürecinde etkilenmiş organ veya dokudaki arteriyoller, daha önce tanımlanan lokal faktörlerden dolayı genişler. Şu halde, arter akımında tıkanıklık olur olmaz arteriyollerin genişlemesi ile büyük ölçüde kan akımı artarak tıkanıklık ortadan kaldırılır.
Yaralanmaya karşı lokal cevap: Doku harabiyeti, hücrelerden lokal olarak çeşitli maddelerin salınmasına neden olur. Bu maddeler arteriyoler düz kasta gevşeme yapar ve

H A Y V A N F İ Z Y O L O J İ S İ
244
haraplanan alanda vazodilatasyona neden olur. Histamin bu maddelerden birisidir ve vazodilatasyon dışındaki bir etkisi de kapiller geçirgenlikte yol açtığı artıştır. Sonuçta, doku hasarına karşı organizmanın genel cevabı olduğu bilinen inflamasyon ortaya çıkar. Bunun dışında doğrudan zedelenen arter, arteriyol ve venlerin ise güçlü bir şekilde kasıldığı görülür. Bu etkinin ortaya çıkışı büyük oranda zedelenmiş damar duvarına yapışan trombositlerden salınan serotonine bağlıdır. Buradan, uzun süreli düzenlemenin ortaya çıkması için gerekli zaman geçtiği takdirde arter basıncının 50 ile 250 mmHg değerleri arasında uzun süreli değişmesinin, yerel kan akımı üzerinde çok az etkili olduğu anlaşılmaktadır. Kan akımının uzun süreli düzenlenmesi özellikle bir dokunun metabolik ihtiyaçları değiştiği zaman önem kazanır. Yani, bir dokunun aktivitesi kronik olarak arttığında ihtiyaç duyduğu oksijen ve besin maddesi miktarı da artar. Bu durumda eğer dolaşım sisteminde bir patoloji yoksa ya da yanıt veremeyecek kadar yaşlı değilse arteriyol ve kapiller damarları genellikle birkaç hafta içinde dokunun gereksinimini karşılayacak şekilde artar. Yani uzun süreli kan akımı düzenlenmesinin mekanizması temel olarak doku damarlanmasının miktarını değiştirmektir. Örneğin, eğer doku metabolizması uzun süreli artarsa, damarlanma artar ve bu olay anjiyojenez olarak adlandırılır. Doku metabolizması azalırsa damarlanma azalır. Ayrıca atmosfer oksijeninin az olduğu yüksek irtifada yaşayan hayvanların dokularındada artmış damarlanma görülmektedir. Uzun süreli yerel kan akımı kontrol mekanizmalarından biri de kollateral dolaşımın gelişimidir. Bir arter veya ven tıkandığında, tıkalı olan yerin çevresinde yeni vasküler kanallar oluşmaya başlar ve etkilenen dokuya kısmen de olsa kanın tekrar gitmesini sağlar. Bu olaydaki ilk basamak tıkalı yerin altında ve üstünde kalan alanlar arasında bağlantıyı sağlayan çevre vasküler yatakta genişleme meydana gelmesidir. Bu dilatasyon ilk 1-2 dakika içerisinde görülür ve ilgili küçük damarların düz kaslarının metabolik nedenlerle gevşemesinden kaynaklanır. Kollateral damarların başlangıçta görülen bu genişlemesini takiben kan akımı, dokunun ihtiyaçları için gerekli olan akımın genellikle dörtte birinden daha azdır. Daha sonraki ilk saatlerde damarlarda daha fazla açılma meydana gelir ve 1 gün içinde doku ihtiyaçlarının yarısı, bunu takip eden birkaç gün içinde de tamamı karşılanabilir.
Sempatik sinirlerin arteriyolleri kontrolü vazodilatasyon oluşumu için de kullanılabilir(Nörepinefrin). Sempatik sinirler nadiren tümüyle sakin fakat bazı orta hızlarda deşarjları organdan organa değiştiği için, her zaman damarların intrensek tonusuna ek olarak biraz daha tonik kasılmaya neden olurlar. Dilatasyon bu bazal seviyenin altında sempatik aktivitenin hızının azalması ile sağlanabilir.
Deri, sempatik sinirlerin oynadığı rolün en iyi örneğini gösterir. Oda ısısında deri arteriyolleri sürekli sempatik deşarjın orta derecede etkisi altındadır. Uygun uyaran -örneğin soğuk, korku, veya kan kaybı- bu sempatik deşarjın refleks artışına neden olur ve sonra arteriyoller kasılır. Tersine, artmış vücut sıcaklığı refleks olarak derideki sempatik sinirleri inhibe eder, arteriyoller gevşer ve vücut ısısını yayana kadar deri kızarır. Kollateral damar büyümesi olaydan aylar sonra devam eder ve çoğu zaman tek ve büyük bir damar oluşturmak yerine birçok küçük kollateral damar oluşumu şeklinde kendini gösterir. Dinlenme halinde kan akımı genellikle normal iken doku aktivitesi arttığında yeni oluşan kanallar maksimum kan akımı sağlamakta ender olarak yeterlidir. Sonuç olarak, kollateral damarların gelişimi akut ve uzun süreli yerel kan akımı kontrol mekanizmalarının genel ilkelerini izler. Akut kontrol hızlı bir metabolik dilatasyona neden olurken, olay haftalar ve aylar içerisinde damarların büyümesi ve yeni damarlar oluşması ile devam eder. Kollateral kan damarlarının oluşumuna en önemli örnek koroner damarlardan birinin trombüs sonucunda tıkanmasından sonra görülmektedir. Altmış yaşına gelen hemen hemen bütün insanların en az bir küçük

H A Y V A N F İ Z Y O L O J İ S İ
245
koroner damarı kısmen veya tamamen tıkanmıştır. Yine birçok insan bu olayın farkında değildir; çünkü kollateral damarlar hızla gelişerek miyokard hasarı oluşmasını engellemektedir.
Sempatik Sinirler: Çoğu arteriyol sempatik postgangliyonik sinir liflerinden yoğun olarak beslenir. Bu sinirler esas olarak, damar düz kas alfa adrenerjik reseptörlerine bağlanarak vazokonstriksiyona neden olan norepinefrin salgılar. Tersine, ileti sistemini kapsayan kalp kasındaki norepinefrin reseptörlerini hatırlarsak, esas olarak beta adrenerjiktirler. Bu, kalpte norepinefrin aktivitesinin bloke edilmesi için beta adrenerjik antagonistlerin farmakolojik kullanımına izin verir fakat arteriyoller için değil ve tersine onlar için alfa adrenerjik antagonistler kullanılır.
Aktif hiperemi ve akım otoregülasyonunun tersine, sempatik sinirlerin kan damarları için esas fonksiyonu, lokal metabolik ihtiyacın düzenlenmesi ve kan akımı ile ilişkili değildir fakat tüm vücudun ihtiyacına hizmet eden reflekslerle ilişkilidir. Bu sinirleri kullanan en yaygın refleks tüm vücuttaki arteriyol direncini etkileyerek arter kan basıncını düzenlemektir Diğer refleksler özel bir fonksiyonu başarmak için kan akımını yeniden dağıtır (bir önceki örnekteki gibi, deriden ısı kaybının arttırılması).
Parasempatik Sinirler :Birkaç istisna ile, arteriyollerin önemli parasempatik innervasyonu çok azdır ya da yoktur. Diğer deyimle, kan damarlarının büyük çoğunluğu sempatik innervasyon alır fakat parasempatik uyarı almaz. Bu, çoğu dokuda görülen çift yönlü otonom innervasyon paterni ile terstir.
Non-kolinerjik, non-adrenerjik Otonom Sinirler
Otonom postgangliyonik sinirlerin, Ne asetilkolin ne de norepinefrin salgıladıkları için, kolinerjik olmayan, adrenerjik olmayan olarak sınıflandırılan bir grubu vardır. Yerine vazodilatör olan nitrik oksit ve olasılıkla diğer kolinerjik olmayan vazodilatör maddeleri salgılar. Bu sinirler kısmen, sindirim sisteminin, kan damarlarının kontrolünde önemli bir rol oynayan barsak sinir sisteminde göze çarparlar. Bu sinirler aynı zamanda diğer bazı yerlerdeki, örneğin ereksiyona aracılık ettikleri penisteki arteriyolleri innerve ederler.
Hormonlar: Antidiüretik hormon (ADH), anjiyotensin II (Ang-ll), atriyal natriüretik peptit (ANP) ve epinefrin gibi dolaşımdaki birçok hormon direnç damarlarının düzenlenmesinde görev alırlar.
1. Antidiüretik hormon (ADH): ADH aynı zamanda arjinin vazopressin diye de bilinir. Doku ozmolaritesi arttığı ya da kan volümü azaldığı zaman arka hipofizden salınır. Başlıca görevi böbrek su tutulumunu kontrol ederek hücredışı sıvı hacmini düzenlemektir fakat eğer dolaşımdaki miktarları yeterince yüksekse (örn hemorajide olduğu gibi), vazokonstriksiyona da sebep olabilir. ADH’ın damar düz kas hücreleri (DDKH) üzerine direkt etkisi ADH V1 reseptörleri üzerinden olur.
2. Anjiyotensin II: Anjiyotensin II güçlü bir vazokonstriktördür. Böbrek arter basıncı düştüğü zaman kan dolaşımında belirir. Sempatik sistemin aktive olması da Ang-ll salınımını tetikler. Anjiyotensin II damar düz kas hücreleri üzerine olan etkisini AT1A reseptörleri aracılığı ile gerçekleştirir.

H A Y V A N F İ Z Y O L O J İ S İ
246
3. ANP: Kalp atriyumları tarafından salgılanan bu hormon etkin bir vazodilatatördür. Bu hormon ve onun böbreklerdeki aktivitesi, arteriyollerin kontrolünde oynadığı yaygın fizyolojik rolü bulunmaktadır. Ayrıca böbreklerde Na+ reabsorbsiyonunu inhibe etmede etkilidir. ANP sekresyonu vücutta aşırı Na+ olduğunda artar buradaki uyaran atriyal gerimdeki artmadır.
4. Epinefrin: Epinefrin adrenal medullada üretilir ve SSS aktivasyonu süresince salınımı devam eder. Esas etkisi miyokart konraktilitesini ve kalp hızını artırmaktır ama aynı zamanda damar düz kas hücreleri üzerindeki α1-adrenerjik reseptörlerine bağlanarak SSS aracılı vazokonstriksiyonu başlatır. Bazı dolaşım bölgelerindeki (örn iskelet gibi) direnç damarları, epinefrin aracılı vazodilatasyon yapan β2-adrenerjik reseptörleri ifade eder. İskelet sistemi damar yatağında bulunan bu yolak "savaş ya da kaç' cevabında kaslara kan akımı artışını kolaylaştırıyor olabilir.
Endotelyal Kontrol :Endotelyal duvarın direnç damarları nitrik oksit (NO), prostaglandinler (PGs), endotelyum kaynaklı hiperpolarize edici faktör (EDHF) ve endotelinler (ETs) gibi çok sayıda vazoaktif bileşiğe aracılık görevi yaparlar.
Nitrik Oksit: NO hem arterlere hem ve venlere etki eden güçlü bir vazodilatördür. Aynı zamanda endotel kaynaklı gevşetici faktör (EDRF) olarak da bilinir. Hücre içi Ca+2 konsantrasyonu artışını takiben yapısal endotelyal NO sentaz (eNOS ya da tip II NOS) tarafından sentezlenir. NO, in vivo yarı ömrü 10 saniyeden daha az olan bir gazdır ki bu da yüksek ölçüde yerel etki gösterdiğinin belirtisidir. Endotel hücre zarından komşu damar düz kas hücreleri lerine doğru difüze olur ve guanil siklazı aktive eder. Artan siklik guanozin monofosfat (cGMP) seviyeleri, cGMP bağımlı protein kinazın fosforillenmesine sebep olur ve miyozin hafif zincir kinazı inhibe eder. Aynı zamanda SERCA (SR Ca+2 ATPaz) pompasını da fosforilleyip aktivitesini artırarak hücre içi Ca+2 konsantrasyonunun düşmesine neden olur. Sonuçta vazodilatasyona sebep olarak kan akışını artırır. NO birçok vazodilatörün etkisine aracılık eder. Yerel modülatörler, asetilkolin, substans P, adenozin trifosfat gibi nörotransmitterler, bradikinin, trombin, akımla indüklenen kayma stresi (shear stress) ve septik şoka sebebiyet veren bakteriyel endotoksinler bunlar arasında sayılabilir. Bu artmış “shear stres”e cevapta arter endoteli PGI2 salgılar, nitrik oksit miktarı artar ve ET-1 miktarı azalır. Bütün bu değişiklikler arter damar düz kasının gevşemesine ve arterin genişlemesine neden olur. Akımla oluşturulan bu arter vazodilatasyonu (arteriyol akım otoregülasyonundan ayrı düşünülmelidir) arterlerin yeniden modellenmesinde ve bazı koşullarda dokulara en iyi şekilde kan sağlanmasında önemli olabilir.
Prostaglandinler: Araşidonik asitten sentezlenen vazoaktif PG’ler endotel kaynaklıdır. PGE (PGE1, PGE2, PGE3) ve PGI2 (prostasiklin) birçok vasküler yatakta damar düz kas hücrelerinin gevşemesine neden olurken PGF (PGF1 PGF2α,PGF3α) ve tromboksan A2 vazokonstriktör olarak görev yaparlar.
Dolaşım Hiyerarşisi: Şimdiye kadar olan kısımda vasküler yataklarda dolaşımın düzenlenmesiyle ilgili mekanizmalar tanımlandı. Pratikte, an be an kontrol, MSS'nin tüm organizmanın ihtiyaçlarını karşılayacak şekilde hazırladığı miktara karşı, dokunun aktivitesini sürdürmesi için gerekli olan akım miktarın değerlendirilmesi ile gerçekleşir. Böylece arteriyel basıncı tehdit eden bir durum olduğunda MSS, kalp debisinin (KD) belli damar yataklarından mahrum edip, önemli organların kanlanmasının devamını sağlayabilir. Farklı organların kan akımına olan ihtiyaçları ve bu akımı sağlamaları, dolaşımsal hiyerarşiyi

H A Y V A N F İ Z Y O L O J İ S İ
247
doğurmuştur. Listenin başında beyin, miyokart ve iskelet kası (egzersiz sırasında) dolaşımları yer alır. Burada, merkezi kontrol mekanizmalarının hemen hemen hiç etkisi yoktur, baskın güç yerel kontrol mekanizmasıdır. Hiyerarşik düzende listenin en altındaki organlar ise bağırsaklar, böbrekler, deri gibi optimal koşullarda kanlanması sağlanan organlardır. Ancak eğer arteriyel basıncın korunması gerekiyorsa bu organların kan akımlarından kesintiye gidilir.
Endotel kaynaklı hiperpolarize edici faktör: EDHF, damar düz kas hücrelerin plazma zarlarındaki K+ kanallarının açılmasına sebep olur. Zarın hiperpolarize olması zar Ca+2 geçirgenliğini azaltarak hücre içi Ca+2 seviyesini düşürür ve vazodilatasyona sebep olur.
Endotelinler: Endotelinler, Anjiyotensin II, mekanik travma ve hipoksi gibi uyaranlara cevap olarak endotel hücreleri tarafından sentezlenen ve salınan bir peptid grubunun üyesidir. ET-1, damar düz kas hücre memembranları üzerindeki ETA reseptörlerine bağlanarak IP3 yoluyla hücre içi Ca+2 artışına tetikleyen güçlü bir vazokonstriktördür.
İyonlar ve Diğer Kimyasal Faktörlerle Vasküler Kontrol
Birçok farklı iyon ve kimyasal faktör kan damarlarında gevşeme veya kasılmaya yol açabilir. Bunların birçoğunun dolaşımın genel düzenlenmesinde küçük bir rolü olmakla beraber özel etkileri aşağıda anlatıldığı gibidir.
1. Ca+2 iyon konsantrasyonunda artma vazokonstriksiyona neden olur. Bu etki kalsiyumun düz kas kasılmasını uyarıcı özelliğinden kaynaklanır.
2. K+ iyon konsantrasyonunda artma vazodilatasyona neden olur. Bu etki potasyum iyonlarının düz kas kasılmasını inhibe etmesine bağlıdır.
3. Mg++ iyon konsantrasyonunda artma magnezyum iyonlarının düz kas kasılmasını inhibe edici etkisi nedeniyle güçlü bir vazodilatasyona neden olur.
4. H+ iyon konsantrasyonunun artması (pH da azalma) arteriyollerde vazodilatasyona neden olur. Bunun aksine, hafif derecede azalma arteriyollerde daralmaya yol açar.
5. Asetat ve sitrat gibi anyonlarınkan damarları üzerinde hafif derecede vazodilatasyona neden olan anlamlı etkileri bulunur.
6. CO2 konsantrasyonunda artma birçok dokuda orta derecede, beyinde ise belirgin vazodilatasyona neden olur. Karbondioksit beyindeki vazomotor merkeze etkili olması ile çok güçlü dolaylı bir etki meydana getirerek sempatik vazokonstriktör sistem aracılığı ile tüm vücutta belirgin vazokonstriksiyona neden olur.
Bazı Özgül Dokularda Kan Akımının Özel Kontrolü ve Endotel
Yerel kan akımının kontrolünde önemli olan genel mekanizmalar vücuttaki tüm dokularda geçerli olmakla beraber bazı özel alanlarda tamamen farklı mekanizmalar işlev görmektedir. Bu konular metin içinde ilgili organlar söz konusu olduğu zaman tartışılacaktır, fakat iki belirgin farklılığa değinmek yerinde olacaktır.

H A Y V A N F İ Z Y O L O J İ S İ
248
Endotel: Endotel damar duvarı ve dolşan kan arasında tek sıra endotel hücresinden oluşmuş fonksiyonel bir bariyerdir. İnsan dolaşım sisteminde 1000 m2 alana sahip yaklaşık 5 kalp ağırlığında vücudumuzun en büyük endokrin bezidir. Normal olarak endotel pıhtılaşmayı önleyici antikoagülan, fibrin parçalayıcı fibrinolitik, trombosit yapışmasını engeleyici antitromboist özelliktedir.
ŞEKİL 4.30 Endotelin foksiyonu.,Kısaltmalar: AGE'ler, ileri glikasyon son ürünleri; COX 2, siklooksijenaz 2; DAG, diasilgliserol; eNOS, endotelyal nitrik oksit sentaz; ET-1, endotelin; ET A , bir endotelin reseptörü; ET B , endotelin-reseptör; ICAM-1, adezyon molekülü 1; İs, interlökinler; MCP1, monosit kemotaktik protein-1; NO, nitrik oksit; OxLDL, okside LDL; NF KappaB, nükleer faktör KappaB; PGI 2 , prostasiklin; PGIS, prostasiklin sentez; PKC, protein kinaz ° C; PLC fosfolipaz ° C; ROS, reaktif oksijen türleri; TNF, tümör nekroz faktörü; TXA 2 , tromboksan 2 ; VCAM1, vasküler hücre yapışma molekülü
Lipoprotein ve eikozanoidlerin metabolizmasında, yüksek molekül ağırlığına sahip lipoproteinlerin uygun zaman ve dokulara geçişimde, kan kaynaklı sinyal moleküllerin iletimde, vasküler(damarların büyümesinde) büyüme, lökosit adhezyununda, bağışıklık oluşmasında katkısı ve görevi vardır.
Damar Gevşetici faktörler:
• Nitrik oksit • Prostasiklin • Brakinin
Damar Kasıcı faktörler • Endotelin • Tromboksan A2 • Prostaglandib H2 • Anjitensin II • Serbest radikallar

H A Y V A N F İ Z Y O L O J İ S İ
249
ŞEKİL 4.31 NO bağlı damarlarda genişleme (vazodilasyon)
Küçük moleküller Histamin, Serbest radikaller, EDRF=NO
Lipidler Prostaglandinler, PGI2, Prostasiklin, Lökotrienler, PAF(platelet aktivaysyon faktörü)
Proteinler
Endotelin-1, Büyüme faktörleri, Adhezyon molekülleri (ICAM, intraselüler ahdezyon molekülü, VCAM, vasküler adhezyon molekülü, PECAM, platelit endotel ahdezyun molekülü, E-selekti, P-selektin, Heparinsülfat, t-PA doku plazmojen aktivatörü, vWf won Willebrand faktörü,MHC-II , ACE
Endotelin hiperglisemi ve lipid metabolizması fonksiyonu üzerine Etkileri : Hiperglisemi, nitrik oksit ve PGI biyoyararlılığını azaltır. Vazokonstriktör Prostanoidler, Protein kinaz C ve endotelin sentezini arttırır. İltihap, yaralanma ya da yükselen LDL kolesterol sonunda endotel hücre aktivasyonu oluşur, Bunun sonunda vasküler endotelyuma yapışması ve monositlerin göçü gözlenir. İnfilamasyon olan yerde makrofajlar oluşur. Endotel hücreleri, düz kas hücreleri ve makrofajlar tarafından Reaktif azot ve oksijen türleri üretilir, LDL değişikliğinin katkısyla; okside LDL makrofaj yüzeyinde CD36 tarafından tanınır ve bağlanır. Kolesteril esterlerin birikmesi ve sitokinler makrofajların köpük hücrelerine dönüşmesine yol açar. Hiperglisemik nitrik oksit inaktiveasyonu AGE lerin geri dönüşümsüz oluşumuna neden olur. Aynı zamanda sitokinlerin fazla salgılanmasına yol açar. Reaktif oksijen türleri oluştumu, makrofajları ile ilgilidir ROS matriks proteinleri ve integrinler ile etkileşimlerini etkiler, hücre içi proteinlerin ile kovalent bağlar oluşturlar. Endotel ve makrofaj hücrelerinde gen ekspresyonu etkiler.
1. Böbreklerde kan akımının kontrolü tübüloglomerüler geribildirim adı verilen bir mekanizma ile olmaktadır. Buna göre, distal tübülün başlangıç kısımlarındaki sıvının içeriği, makula densa adı verilen bir tübül epitel yapısı tarafından saptanmaktadır. Makula densa, nefronda jukstaglomerüler aygıt hizasında distal tübülün aferent ve eferent arteriyollere komşu olduğu bölgede yerleşmiştir. Glomerüllerde, fazla miktarda sıvı kandan tübül sistemi içine filtre olduğunda makula densada oluşan geribildirim sinyalleri aferent arteriyollerde kasılmaya neden olur. Bu şekilde böbrek kan akımı ve glomerüler filtrasyon oranı normal düzeyine doğru geri döner.

H A Y V A N F İ Z Y O L O J İ S İ
250
2. Beyin kan akımının düzenlenmesinde etkili faktörler: Özellikle kimyasal faktörlere göre düşünülürse; beyinde kan akımının doku oksijenasyonuna göre ayarlanmasının yanı sıra karbondioksit ve hidrojen iyon konsantrasyonları da önemli bir rol oynamaktadır. Bunların herhangi birinde veya her ikisinde meydana gelen artış serebral damarları genişleterek biriken
karbondioksit ve hidrojen iyonlarının ortamdan uzaklaştırılmasını sağlar. Bu, beynin uyarılabilme düzeyinin karbondioksit ve hidrojen iyon konsantrasyonlarının hassas kontrolüne bağlı olması nedeniyle önemlidir.
3. Deride, kan akımı kontrolü vücut sıcaklığının düzenlenmesi ile yakın ilişkilidir. Kutanöz ve subkutanöz akım, merkezden vücut yüzeyine (sıcaklığın uzaklaştırıldığı yer) olan sıcaklık akımını ölçerek vücuttan sıcaklık kaybını düzenler. Deri kan akımı santral sinir sisteminin sempatik sinirleri ile kontrol edilir. Serin havada deri kan akımı 100 g doku başına sadece 3 ml/dak iken, gerekli durumlarda büyük değişiklikler gerçekleşebilir. Vücut sıcaklığı arttığında, deri kan akımı pek çok kat artabilir. Tüm vücuttaki kan akımı 7-8 L/dak’a ulaşabilir. Vücut sıcaklığı düştüğünde, deri kan akımı azalır ve çok düşük sıcaklıklarda sıfırın biraz üzerindedir. Ciddi damar daralması durumunda bile deri kan akımı genellikle bazal metabolik ihtiyaçları karşılamaya yeterlidir.
4. Koroner kan akımının kontrolü şu faktörlere bağlıdır:
Kalp frekansı ileri derecede arttığı zaman diyastol süresi kısalır. Koroner dolaşımın yüzde yetmişi diyastol süresinde olduğu için ileri derecedeki taşikardilerde koroner akım azalır. Ancak orta
derecede frekans artışında miyokard metabolizmasının yükselmesi koroner dolaşımı arttırır.
Kapiller membrandan sıvı geçişi: Starling güçleri;
Kapillerde kandan dokulara ve dokulardan kana madde geçisi farklı bir çok güç tarafından kontrol edilir. Temel olarak 4 temel osmtik(onkotik) güç kapiller damarlardan madde geçisini denetler. Bu güçlerin toplamı madde geçisin hangi yönde ne kadar olacağını belirler.
1. Kapiller Hidrostatik Basınç (Pk): Kapiller damarlardan sıvıyı kapiller damardan dışarı iten güçtür. Kalp pomplaması sonucu meydana gelen basınçtan kaynaklanır.
2. İnterstisyel sıvı basıncı (Pi)(+): Sıvıyı interstisyumdan yani doku tarfından kapiller damarın içine iten (-): ters yönde hareketlendiren güçtür.
3.Plazma kolloid osmotik (Onkotik) basıncı (πp) Kapiller damarın içine doğru sıvı osmozuna neden olan güçtür. Kan osmotik basınç artıkça plazma kolloid basınçında artar.
4. İnterstisyel sıvı kolloid osmotik (Onkotik) basıncı (ui) sıvının kapiller damardan dışarı osmozunu sağlayan güç. Doku osmtik basınçına bağlıdır.
1-Otoregülasyon,
2-Kimyasal faktörler,
3-Sinirsel faktörler.
1-Lokal kontrol,
2-Sinirsel kontrol,
3-Arteriyel kan basıncı,
4-Kalp frekansı.

H A Y V A N F İ Z Y O L O J İ S İ
251
�
Lenf sistemi
Kardiyovasküler sistem farklı olarak, lenf sistemi, kapalı bir sistem değildir. İnsan dolaşım sistemi ortalama günde 20 litrelik süzüntüyü yani plazması dokulara bırakır. Bu plazmanın yaklaşık olarak 17 litresi, kan damarları içine doku sıvısıı olarak terkrar alınır. Geriye kalan 3 litre ise, lenf sistemi tarafından dokudan geri alınır. Lenf sistemi kan dolaşımı gibi doku ve hücrelerdeki artık maddeleri toplar, fakat lenf sisteminin bu transport işlemi oldukça farklıdır. Kan dolaşımı arter, ven ve kapiller damarlardan oluşurken, lenf sistemi tek yönlü yol gibi sadece toplama işlemi yapar. Hücreler arasında kalan artık maddeleri lenf sistemi alarak ana lenf damarına (kanalına) ulaştırır, bu kanalda artık maddeleri toplardamarlara verir. İnterstiyel sıvıya geçen sıvının(plazma sıvısının) 1/10 u lenfatik kapillerler ile kana döner (2-3 lt/gün). Kılcal damarın venül kısımların bir miktar doku sıvısı kana geri emilir. Buna karşın doku sıvısıyla dokular arası sıvıya sızmış olan kan proteinleri doku sıvlarıyla lenf dolaşımı aracıyla tekrar kan dolaşımına döner. Lenf akımı belirleyen başlıca faktörler: �
Lenf dolaşımı doku sıvının interstisyel alandan kana akmasını sağlayan alternatif bir yol oluşturur. Lenf torasik kanal ile internal juguler ven-subklavian venin birleşim noktasında venöz sisteme boşalır. Lenf sistemi doku arasındaki maddelerin temizlenmesinde görev alır. Özellikle fazla sıvının ve doku sıvısındaki proteinleri lenfle uzaklaştırılır. Lenf akımı vücud aksarsa (kanın osmotik yapının değişmesi, lenf kapillerinin ve damarlarının tıkanması, doku osmotik basınçının değişmesi) dokularda ödem meydana gelir. Lenf sistemin diğer ana işlevi bağışıklık sistemi olarak görev yapmasıdır. Lenf sıvısı kan plazması çok benzer, lenf sıvısında lenfositler ve diğer beyaz kan hücreleri ile atık ürünler ve bakteri ve protein ve te hücre artıkları bulunur. Lenfatik kapillerler, kapiller endotel hücreleri bağ dokusuna bağlayıcı filamentler ile tutunur. İçinde geri akıma izin vermeyen valf sistemi vardır.
• Interstisyel sıvı basıncı • Lenfatik pompa aktivitesi ile belirlenir.
Sıvıyı dışarıya doğru iten ortalama güçler mmHg
Ortalama kapiller basınç 17.3
Negatif interstisyel serbest sıvı basıncı 3.0
İnterstisyel sıvı koiloid osmotik basıncı 8.0
DIŞARIYA İTEN GÜÇLERİN TOPLAMI 28.3
Sıvıyı içeriye doğru iten ortalama güçler
Plazma koiloid osmotik basıncı 28.0
İÇERİYE İTEN GÜÇLERİN TOPLAMI 28.0
Güçlerin toplamı
Dışarı 28.3
İçeri 28.0
NET DIŞARIYA İTEN GÜÇ 0.3

H A Y V A N F İ Z Y O L O J İ S İ
252
ŞEKİL 4.32 Lenf damarı
Protein konsantrasyonu 3-5 g/dl Besin maddelerinin absorpsiyonu (özellikle yağ) göre değişim gösteri. Lenfatik sistemin önemli bir fonksiyonu bakterilerin etkisizleştirilmesidir.
Interstisyel sıvı basıncını artıran faktörler; Kapiller basınç artışı, Plazma onkotik basıncının düsmesi, Interstisyel Sıvı protein artış,ı Kapiller permiabilite artışıdır Yalnızca 2 -3 mmHg olan interstisyel sıvı basıncı, atmosferik basıncın (0 mmHg) biraz üzerine çıkması ile
lenf akımı ~20 kat artırır.
ŞEKİL 4.33 Lenf kapilleri
Lenf akımını; Lenf damarları duvarındaki düz kasın sıvı ile gerilme sonucu otomatik olarak kasılması ve valfler arasındaki segmentin otomatik pompa gibi çalışarak sıvıyı ilerletmesi ile olur. Toplayıcı lenfatikler başlangıç lenfatiklerin birleşmesiyle meydana gelir. Başlangıç lenfatikleri kapiller damarın aynı işlevi görür(analogtur). Toplayıc kanallarda
Lenf sistemi Lenfatik sistem veya lenf sistemi lenf sıvısı, lenf damarları ve lenf düğümlerinden oluşan bir organ sistemidir. Dolaşım sisteminden bağımsız olarak çalışan lenfatik sistem bağışıklık sistemi içeriğini yine dolaşım sistemine boşaltır ve genel olarak bağışıklıkta rol alır. Lenf sistemi kan dolaşımı gibi doku ve hücrelerdeki artık maddeleri toplar. Kan dolaşımı atar ve toplar damarlardan oluşurken, lenf sistemi tek yönlü yol gibi sadece toplama işlemi yapar. Hücreler arasında kalan artık maddeleri lenf sistemi alarak ana lenf damarına (kanalına) ulaştırır, bu kanalda artık maddeleri toplar damarlara verir.
Lenf kapilleri Lenfatik sistem damarlarının başlangıcıdır. Değişen çapta lümene sahip az gelişmiş bazal membrana sahip son derece ince bir endotelyumla kaplanmıştır.

H A Y V A N F İ Z Y O L O J İ S İ
253
Lenfaniyonlara ve her 6–20 mm bir kapaçıklara sahiptir. Toplayıcı lenfatikler lenf düğümlerine açılır. Lenf düğümleri akyuvarlar hücrelerinden (lenfositler) üretirler Lenf düğümleri lenfleri filtre eder, içinde olası tüm mikroorganzimaları fagositozla yok eder.
Lenf dolaşımı lenf düğümlerinden sonra sistemik dolaşıma katılmak üzere santral kanallarla iki noktadan venlere bağlanır. Bacaklar ve barsakların lenfatikleri, sisterna şili ile birleşip duktus torasikusu oluşturur, Sol kol ve yüzün sol tarafı duktus torasikus lenf taşır. Sonunda sol taraftaki lenf akımı sol köprüçük altı toplardamarına (Sol Subklavian vene) dökülür. Sağ kol ve yüzün sağ tarafı sağ lenfatik kanalar ise sağ köprüçük altı toplardamarına(Sağ Subklavian vene ) dökülür. Lenf sistemin dokulardan yeterince sıvıyı almaması durumunda dokularda sıvısının artışı (ödem) meydana gelir.
�
Ödeme yol açan başlıca faktörler:
o Filtrasyon basıncının artması
o Arteriyollerin genişlemesi
o Venöz basıncın artması (yerçekimi, kalp yetersizliği, vena kapakcıklarının yetersizliği, venaların tıkanması, ekstrasellüler sıvının total hacminin artışı gibi)
o Kapiller çeperleri hizasında osmotik basınç gradyanının azalması
o Plazma protein düzeyinin düşmesi
o İnterstisyel alanda osmotik aktif metabolizma ürünlerinin birikmesi
o Kapiller permeabilitesinin artması
o Histamin ve benzeri maddeler
o Kininler ve Lenf akımının yetersizliği

H A Y V A N F İ Z Y O L O J İ S İ
254
Kan Fizyolojisi Kan doku özelleşmiş bir bağ doku elemanlarındandır. Her bağ dokusu gibi iki ana birleşeni vardır. Sıvı haldeki matriks plazma(yüzde 55) ve şekilli elemanlar ya da hücrelerinden(yüzde 45) meydana gelir. Kanın temel görevleri başlıcaları;
Kan hacmi vücut ağırlığının % 8’ilik kısmını oluşturur. Kan miktarı yaş, cinsiyet, vücut tipine bağlı olarak değişir. Erkeklerde 5-6 litre, Kadınlarda 4-5 litre kan bulunur. Buna karşın yenidoğanda vücuda göre daha fazla kan vardır. Kan hücreler(yüzde 45’lık kısmı) ve sıvı kısmından (Plazma yüzde 55’lik) kısım dan oluşur. Kan hacmi plazma kısmını gereğinde vücudun diğer kısımları ile tamamlanırken (ekstrasellüler sıvı ile) plazma aynı zamanda vucud içinde sıvı dağılımın (alınmasını ve verilmesinin) sağlar. Sindirim sistemi ile alınan tüm maddelerin hücrelere, hücrelerede oluşan tüm atık madddelerinde akçiğer, böbrekler, karaciğer gibi organlara taşınması kan ile yapılır.
ŞEKİL 4.33 Kan, şekil elemanlar ve plazma
Plazma Proteinleri
Kanın yüzde %55 kısmını oluşturan sıvı kısmı plazmada 3 tip protein vardır. Albüminler toplam proteinlerin %52-68’i, olşturur en önemli fonksiyonu taşıyıcı olmaları (Transferin demiri, Seroplasmin bakır) ve kan osmotik basınçını (280-300 mOsm) oluşturmalarıdır. Diğer bir protein protrombin (kan pıhtılaşmasından sorumlu olan prolitik bir proenzim) hemostazda görevlidir. Plazma proteinlerinin yaklaşık %60’ını oluşturur. Plazmadaki miktarı normalde 3.5- 4.5 g/dl’dir. Molekül ağırlıkları çok düşüktür (69.000
Hemotokrit Hematokrit, kırmızı kan hücrelerinin oluşturduğu hacmin, toplam kan hacmine oranıdır. Hematokritin normal değerleri yaş ve cinsiyete bağlı olarak değişmekte olup erişkin bir erkekte %42-52, kadında %36-46. aralığındadır. İnce uzun tüpe alınan kan sentrifüj makinasına konulur. Kanın içindeki ağır olan hücreler dibe çökerken daha hafif olan plazma üstte kalır. Plazma ve kan hücrelerinin arasındaki sınır değeri okunur
Taşıma Oksijen, besin maddeleri, atıklar, hormonlar, enzimler vs
Düzenleme Isı, pH, tuz ve sıvı dengesi, osmotik basınç, hormonal kontrol vs
Koruma Kan kaybına karşı (pıhtılaşma), enfeksiyonlara karşı (immün sistem) vs
Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
255
dalton). Yapıca küçük, sayıca çok olmasından dolayı albumin plazma protein osmotik basıncının %80’inden sorumludur. Kan ile doku sıvıları arasında osmotik dengenin ayarlanmasında önemli rol oynar. Plazma albumin miktarı düşerse bunun sonucunda kanda sıvı kaybı buna karşılık interstisyel boşlukta sıvı birikimi (periferal ödem) gözlenir. Osmotik rolü yanısıra albumin plazmada birçok maddeye bağlanarak onların çözülmesini sağlar ve böylelikle plazma transportunda da önemli görev alır. Albuminin bağlandığı maddelerden bazıları: İlaçlardan; barbituratlar ve penisilin, pigmentlerden; bilirubin ve ürobilin, hormonlardan; tiroksin ve diğer maddelerden de yağ asitleri ile safra asit tuzlarını sayabiliriz. Globülinler proteinler toplam proteinlerin %40’ı oluştur. Alfa 1-2, Beta 1-2 ve Ig ler tipleri(IgE, IgM, IgD vb) vardır, özelikle Antikorlar (immünite) olarak görev yaparlar. Globüler protein dağılımı : %4’ü alfa-1, %8’i alfa-2, %7’si beta-1, %4’ü beta-2 ve %17’si gamma globulindir. Alfa-1 globulinlerin çoğunluğu glikoprotein (karbonhidrata bağlanan protein) az bir miktarı ise lipoproteindir (lipide bağlanan protein). HDL (high density lipoprotein) yada iyi huylu lipoprotein) lipid transportunda görev alır ve kolesterolün arterlerin duvarlarına yapışmasını engeller. Alfa-1 grubunun diğer proteinleri tiroksin bağlayan globulin, kortizol bağlayan globulin (transkortin) ve vitamin B12 bağlayan globulin (transkobalamin) dir. Bunlar bağlandıkları maddelerin transportunu sağlarlar. Alfa-2 globulinler; haptoglobini (serbest hemoglobine bağlanan ve böbrekten atılmasını engelleyen protein) ve seruloplasmini (bakır içeren bir oksidaz enzimi) içerirler. Alfa-2 globulin yapısındaki diğer proteinler protrombin (kan pıhtılaşmasında görev alır), eritropoetin (eritrosit üretimini stimüle eden hormon) ve anjiotensinojen (kan basıncı, vücut sıvısı ve elektrolit balansının düzenlenmesinde görev alan bir hormon) dir. Beta globulinler (beta-1, beta-2) lipidlerin taşıyıcı proteinlerinin çoğunluğunu içerirler. Beta-1 lipoproteinleri, diğer adıyla kötü huylu kolesterol LDL (Low density lipoprotein B48 ve B100), kolesterolün arter duvarlarında çökmesine neden olurlar yani damar ve kalp hastalıklarında rol oynarlar. Beta globulinler tarafından taşınan diğer maddeler fosfolipidler, gliseridler, yağda çözünen vitaminler ile bakır ve demir metalleridir. Bakır ve demir transportunda rol oynayan transferrin bir beta globulindir. Kan proteinlerinin temel sentez yeri karaciğer olmasına rahmen Lenfositler(B lenfositler) gibi kan hücreleride plazma içinde protein sentezlerler. Kan proetinlerin kan pH sabit tutulmasını sağlar. Kan proteinleri komplement sistemde(C1-C11) vücudu istilacı organizmalara karşı korur. Kan proteinleri kanın pıhtlaşmasında(fibronojen) görev alır.
Gamma globulinler: Bağışıklık sisteminde antikor olarak görev alırlar. Antikorlar, vücuda giren yabancı maddelerle yani antijenlerle bileşikler kurabilen moleküllerdir. İmmunglobulinler olarak bilinen gama globulinler IgG, IgA, IgM, IgD ve IgE gruplarına ayrılırlar.
IgG: Sağlıklı bir insanda immunglobulinlerin % 75’ini oluşturur. G1, G2, G3 tipinde 3 formu vardır. Molekül ağırlıkları 51.000- 60.000 arasında değişir. İmmun sistemde pekçok antijene karşı sekonder yanıt şeklinde savunmada görev alırlar. Plasentadan geçerek yeni doğan bebeğe bağışıklık sağlar. Kompleman sisteminde klasik yolu aktive ederek antijenle savaşır. Tip 2 hipersentivede IgG görev alır.
IgA: IgA1 ve IgA2 olarak 2 tipi vardır. Molekül ağırlığı 52.000-56.000 arasındadır. Mukus salgısı yapan epitel hücrelerinin membranlarında ve sekresyon sıvılarında bulunur. IgA antikor reseptörünü taşıyan epitel hücreleri, sindirim kanalı, akciğer, meme dokusu, ürogenital sistem gibi dışa açılan sistemlerde yaygın bir savunma hattı oluştururlar. Mukus sıvısında bulunabilecek viral ve bakteriel antijenler,

H A Y V A N F İ Z Y O L O J İ S İ
256
bu reseptörlere bağlı IgA antikorlarınca yakalanıp yok edilirler. Antijen bilgisi, sistemde yer alan microfold (M) hücreleriyle alt tabakalardaki lenfoblastlara da iletilirler. Bu şekilde o antijene özgü IgA antikor sentezi artar. IgA lenf yoluyla diğer sistemlere taşınır. Amaç, gelebilecek antijen saldırısına karşı organlarda önlem almak, onları korumaktır.
IgM: Molekül ağırlığı 70.000 olup, en büyük antikor tipidir. Bağışıklık sisteminde, antijenik uyarıya karşı primer yanıt olarak oluşan ilk antikor grubudur. Kompleman fiksasyonunda etkilidir. T lenfositlerin başedemediği antijenleri opsoninleyerek savaşırlar. Antijen spektrumları çok geniştir.
IgE: Bazofil ve mast hücrelerinin uyarılmasıyla görevlidirler. Bu hücrelerin yüzeyindeki Fc reseptörlerine bağlanarak onları aktive ederler. Aşırı duyarlılık(hipersensitive reaksiyonaları) reaksiyonlarında ve enfeksiyonlara karşı korunmada çok önemlidirler. Tip I hipersensitive görev alır.
IgD: Serumda çok düşük düzeyde bulunan bir antikor tipidir. Fonksiyonu tam olarak bilinmemektedir. IgD reseptörlerinin B lenfosit yüzeyinde sıkça bulunması, antikor sentezinin kontrolünde etkili olabileceğini düşündürmektedir.
Fibrinojen: Plazma proteinlerinin % 0.3’nü oluşturan fibrinojen karaciğerde yapılır. Çözünebilir bir proteindir. Hemostazda trombinin etkisiyle çözünmez bir protein olan fibrine dönüşerek pıhtılaşmada önemli rol oynar
Kan protein farklı görevleri vardır. Birincisi: Hücre beslenmesi için amino asitlerin önemli rezerv kaynağı olarak hizmet ederler. Gerekirse; retiküloendotelyal sistem hücreleri (Karaçiğer, dalak, akciğer ve lenfatik dokulardaki makrofajlar) plazma proteinlerini içlerine alırlar, yıkıma uğratırlar ve amino asit komponentlerini kana salgılayarak diğer hücrelerin bunları yeni protein yapımında kullanmalarını sağlarlar.
Plazma proteinlerinin ikinci görevi: Diğer moleküllere taşıyıcı olarak hizmet etmeleridir. Küçük moleküllerin çoğu spesifik plazma proteinlerine bağlanarak barsaklardan yada depo edildikleri organlardan diğer dokulara kullanılmak üzere taşınırlar. Örneğin; demir kanda transferrin adı verilen transport proteinine bağlanarak taşınır. İlaçlar, pigmentler, hormonlar ve iyonların birçoğuda kanda plazma proteinlerine bağlanırlar.
Plazma proteinlerinin üçüncü görevi: Stabil kan pH'sının sağlanmasına yardımcı olmak üzere tampon etkisi göstermektedirler. Kanın normal pH'sı 7.35- 7.40 arasındadır. Proteinler amfoteriktirler (serbest NH2 ve COOH içerirler). Yani pH'ya bağlı olarak zayıf asit veya zayıf baz etkisi göstererek H+ yada OH- ile bağlanma yeteneğindedirler. Genellikle zayıf baz etkisi göstererek ortamdaki fazla H+ 'ı bağlarlar ve böylece kanı hafif alkali durumda tutmaya yardım ederler. Plazma proteinleri kanın tamponlama yeteneğinin % 15'sinden sorumludurlar.
Dördüncü görevi: Birçok plazma proteini kan pıhtılaşmasını sağlayacak şekilde spesifik yollarla interaksiyona girer. Kanın pıhtılaşması, vücudun vasküler yaralanmaya verdiği yanıttır ve kan kaybının önlenmesi ile mikroorganizmalara karşı korunmaya yardım eder.
Beşinci görevi: Plazma proteinleri suyun kan ve interstisyel sıvı arasında dağılımını yönlendiren kolloid osmotik (onkotik) basıncı oluştururlar.Normal plazmanın total osmotik basıncı yaklaşık 7.3 atm. (1 atm= 760 mmHg) ya da 5550 mm Hg.dir. Plazma ile aynı osmotik basınca sahip olan solüsyonlar izotonik olarak adlandırılır. İzotonik solüsyon (örneğin; % 0.9’ luk NaCl) içine yerleştirilen bir eritrosit ne su alır nede su kaybeder. Plazmadan daha yüksek osmotik basınca sahip olan solüsyonlar hipertonik, daha düşük osmotik basınca sahip olanlar ise hipotonik olarak adlandırılır. Hipertonik solüsyona yerleştirilen eritrosit su kaybedip büzülürken, hipotonik solüsyona konacak olursa

H A Y V A N F İ Z Y O L O J İ S İ
257
şişer, patlar ve hemolize uğrar. Plazma, interstisyel sıvı ve intrasellüler sıvı normalde osmotik dengede olduğu için homeostasis kritik olarak plazmanın osmotik basıncının regülasyonuna dayanır. Normal Ekstra sellüer sıvı osmotik basıncından herhangi bir sapma hücrelerin hacminde artmaya yada kayba, dolayısıyla hücrelerin fonksiyon yapmamasına ve ölümüne neden olur.
Plazmanın total osmotik basıncının yaklaşık % 99.5'i su ile birlikte kapiller membranlar boyunca rahatlıkla geçen moleküllere (örn: elektrolitler, üre, glikoz ve diğerleri) bağlıdır. İnterstisyel sıvı için de durum aynıdır. Plazmanın total osmotik basıncına plazma proteinlerinin katkısı yaklaşık % 0.5'dir ve plazmanın normal kolloid osmotik basıncı 28 mmHg'dır (bunun 25 mmHg'sı proteinlerden, yaklaşık 3 mmHg'sı ise (-) yüklü proteinlerin Na+ iyonunu tutmasından kaynaklanır) Plazma ve interstisyel sıvı arasında su dağılımının regülasyonunda onkotik basınç önemli bir rol oynar, çünkü plazma proteinleri kapiller membranlardan geçemezler. Kandan interstisyel sıvıya geçen az miktardaki protein interstisyel sıvı içinde toplanmaz çünkü bunlar lenfatik sistem tarafından uzaklaştırılır. Böylece kandan interstisyel sıvıya doğru bir protein konsantrasyonu gradyanı sağlanır ve osmotik olarak su ile küçük moleküllerin plazma ve interstisyel sıvı arasındaki hareketi yönlendirilir. Kısaca plazma proteinleri temel işlevleri:
• Viskoziteyi sağlamak
• Onkotik basıncın oluşmasını sağlamak
• Kan hacminin korunmasını sağlamak • Taşıma görevi
• Asit-baz dengesini korumak
• Beslenme-yapıtaşı • Pıhtılaşma, immün yanıt
Transferrin: Transferrin geriye dönüşümlü olarak demir, bakır, çinko, kobalt ve kalsiyum bağlayabilmektedir. Yalnız bu katyonlardan demir ve bakırın bağlanması fizyolojik önem taşımaktadır. Seruloplazmin (iki ferrik iyonu) Fe+2 iyonunun Fe+3 iyonuna yükseltgenerek transferrine bağlanmasını sağlamaktadır. Dolaşımdaki transferrin moleküllerinin yarı ömrü 7 gündür. Vücudun demir yetersizliğinde, dolaşımın transferrin düzeyi bir cevap olarak normal düzeyinin iki veya daha fazla katına yükselmektedir. Bu durumda transferrin apoproteini bağırsaktan demiri daha fazla çekebilmektedir. Bağırsaktan kazanılan demirle oluşan transferi- demir Tf-Fe+3 dolaşımla bu kez karaciğer, RES gibi demir depolarına ulaşır ve oralarda serbestleşen demir, ferritin veya hemosiderin sentezlerine katılır. Serbest halde toksik etkili olan demir, organizmada gerek duyulan dokulara taşınmak üzere transferrin ile birleştiğinde potansiyel toksisitesi azalmaktadır. Demir plazma içinde transferrine bağlı olarak ferrik iyonu (Fe+3) şeklinde taşınır. Transferrin spesifik hücre reseptörlerine bağlanınca demiri serbest bırakır. Ve bu kompleks içeri alınır. Demir ferritin veya hemosiderin olarak depolanır veya hem proteini sentezinde kullanılır.
Seruloplazmin(Cp): Tek bir polipeptid zincirinden oluşan ve bir α2 globulin olan seruloplazmin, molekül başına 6–8 demir atomu bağlamaktadır. Bakır oksidaz veya ferrooksidaz adlarını alan enzimdir.(kofaktörü bakır) Enzim olmasının yanında bakır vericisi fonksiyonuna sahiptir. Seruloplazmin iki değerlikli demir katyonunu (Fe+2) üç değerlisine (Fe+3) oksidasyonu katalizleyen (Ferrooksidaz) enzimdir. Ferrooksidaz, demirin transferine bağlanabilmesinden önce, hücre yüzeyinde ferro (Fe+2) forma

H A Y V A N F İ Z Y O L O J İ S İ
258
dönüşmesinde görev aldığından büyük öneme sahiptir. Enflamasyonda, ekzersizde, gebelik sırasında ve östrojen uygulamalarında serum seruloplazmin düzeyi yükselir. Uzun süre plazmanın major bakır taşıyıcı proteini olduğu düşünülen seruloplazminin fonksiyonu tartışmalıdır. Sitokrom oksidaz, tirozinaz, süperoksit dismutaz gibi birçok önemli enzimin metal kofaktörü olan bakır, erişkin bir kişinin vücudunda çoğunluğu kemik, karaciğer ve kasta olmak üzere 100–150 mg kadar bulunmaktadır. Plazmada bulunan bakırın %95 kadarını diyaliz ile ayrılmayan, seruloplazmine bağlı, % 5 kadarını ise diyaliz ile ayrılabilen ve albumin ile histidine gevşek olarak bağlı fraksiyonu oluşturmaktadır. Barsaktan karaciğere albumine bağlanarak taşınan bakır, karaciğerde seruloplazmin apoproteini ile birleşmekte ve moleküle sialik asit zincirleri eklendikten sonra dolaşıma verilmektedir. Seruloplazmin antioksidan olarak görev yapmaktadır. Organik bileşiklerin oksijen ile spontan oksidasyonu sonucu oluşan, yaşam için tehlike yaratan bileşikler, plazma ve dokularda bulunan antioksidanlar tarafından ortadan kaldırılmaktadırlar. Plazmada bulunan oksidan aktivitenin önemli bir bölümünü seruloplazmin ve transferrin göstermektedir. Bir akut faz proteini olan ve akut yanıtı sırasında konsantrasyonu % 50 artan seruloplazminin lipid peroksidasyonunu ve serbest radikal oluşumunu önlediği düşünülmektedir Böbreklerde, karaciğerde biriken bakır (siroza), beyinde biriken ise bazal ganglionlarda hasara, eritrositlerde biriken bakır hemolitik anemiye yol açmaktadır. Korneada biriken bakır, kornea çevresinde yeşil veya sarı renkli Kaiser-Fleisher halkası oluşturmaktadır. Wilson hastalığında serum seruloplazmin düzeyi tipik biçimde düşüktür. Buna karşılık iyon bakır düzeyi serumda yüksek bulunur. Bakırın üriner ekskresyonuda yüksektir.
Transkortin: Kanda dolaşan transkortizolün yaklaşık %90’ı plazma proteinlerine ve primer olarak da α- globuline bağlanmış bulunur. İşte kortizolün bu α-globuline bağlı biçimi kortikosteroid bağlayıcı globulin(CBG) veya transkortin olarak adlandırılır. Kortizolü yüksek afiniteyle bağlayan bu molekül 56.000 dalton ağırlığındadır. Aslında transkortin Androjen-bağlayan protein (sex hormon-bağlayan protein-SBP)vitamin D bağlayan protein(DBP), retinol bağlayan protein(RBP) gibi kabul edilir. Çünkü bütün bunlarda lipid veya benzeri ürün hormonunun özel taşıyıcısı durumundadır. Serumda yetişkinlerde normal seviyesi 70 mg/L kadardır. Gebelik, androjen veya östrojen uygulamalarında yükselir. Plazma Lipidleri
Yağlar sindirim sisteminden yağ asidi ve gliserol olarak emilir ve barsakta trigliseride dönüştükten sonra 0,5 mikron çapında partiküller halinde (şilomikron) kana geçer. Kolesterol ve fosfolipidler ise barsakta değişime uğramadan absorbe olur. Günlük diyetle fazla yağ alınırsa, plazmada yağ o kadar artabilir ki, süt gibi bir bulanıklık oluşur. Buna lipemi denir. Açlıkta da yağ mobilizasyonunun artması nedeniyle lipemi görülebilir. Kandaki iyonize yağ asitleri albuminle (esterleşmeyen yağ asidi), diğerleri ise gliserol ve kolesterol esterleri şeklinde taşınır. Hücrelerin artan enerji gereksinimini karşılamak üzere yağ tüketimi artınca kanda esterleşmemiş yağ asitleride çoğalır. Normal koşullarda bir mol albumin 3 yağ asidi taşıdığı halde gerektiğinde 30 yağ asidini taşıyabilir. Plazma Karbonhidratları
Glikoz %90–110 mg, laktik asit %4–10 mg düzeyindedir. Sindirim sisteminden karbon- hidratların % 80'i glikoz, gerisi fruktoz ve galaktoz halinde kana geçer. Fruktoz ve galaktoz da karaciğerde glikoza çevrilir. Yemeklerden sonra %140 mg'a çıkan kan şekeri, daha sonra %90 mg'a düşer. Kanda glikozun sabit bir düzeyde tutulması hormonlar(insülin, glukogon) yardımıyla olmaktadır. Azaldığında karaciğer ve

H A Y V A N F İ Z Y O L O J İ S İ
259
kaslardaki glikojen depolarından kana glikoz çekilir. Bütün kanda 5 g glikoz vardır, gerektiğinde organların kandan çektikleri glikoz saatte 7 g’ma yükselebilir. Kas çalışması sırasında kanda düzeyi yükselen laktik asitten karaciğer glikojen yapabilir. Plazmada Bulunan İnorganik Maddeler
Plazmada en çok bulunan iyonlar Na+ ve Cl- dur. K+ ve HCO3 daha azdır. Plazmadaki Ca2+’un 1/3’ü proteine bağlı, 2/3’ü de erimiş haldedir. Minerallerin miktarı; proteinlerle (%7-8 g) kıyaslanamıyacak kadar azsa da, kanın osmotik basıncını sabit tutmak gibi hayati bir rol oynarlar. Bunun nedeni moleküllerinin küçük, molekül sayılarının çok oluşudur. Bilindiği gibi: osmotik basınç moleküllerin türüne ve büyüklüğüne değil, sadece sayısına bağlıdır. Minerallerin yaşamın devamı bakımından başka rolleri de vardır. Kan hücreleri
Kanın şekilli elemanları santrifüjle çökeltilebilir ve kanın % 45’lik kısmını(hemotrokrik değeri) oluşturur. Eritrositlerin oluşturduğu hacmin, toplam kan hacmine oranına hematokrit denir ve erkeklerde % 40–50, kadınlarda % 35–45 arasındadır.(Vücut ağırlığının yüzde 8 kısmı kan hacmini verir.)
ŞEKİL 4.34 Kan bileşenleri
Kandaki eritrositler sayısı kadında 4.000.000–4.500.000/mm3 erkeklerde 4.500.000–5.000.000/mm3 arasındadır. Lökositler sayısı 7000 ile 11000/mm3 ve trombositlerin sayısı ise 150.000–450.000/mm3kadardır. Eritrositler(akyuvarlar)
Eritrositler kan hücrelerinin büyük kısmını oluştururlar(yüzde 99). Başlıca görevleri akciğerlerden dokulara oksijen götürmekle yükümlü olan hemoglobini taşımaktır. Bazı ilkel hayvan türlerinde hemoglobin eritrositler içinde değildir(Hemoeritrin toprak solucanı). Plazmada serbest protein halinde bulunur. Ancak, insanda hemoglobin kan plazmasına çıkar ve serbest dolaşmaya başlarsa o zaman bunun her geçişte yaklaşık % 3 kadarı kapiller membrandan doku aralıklarına ya da böbrekte glomerüler membrandan glomerüler filtrata sızar. Bu nedenle hemoglobin, dolaşımda kalabilmek için eritrosit içinde olmalıdır. Eritrositlerin basitçe hemoglobini taşımak ve korumaktan başka görevleride vardır. Örneğin; karbondioksit ve su arasındaki reaksiyonu katalize eden ve bu tersinir reaksiyonun hızını binlerce kez artıran karbonik anhidrazı çok miktarda içerirler. Böylece büyük miktarda karbondioksit, kandaki su ile reaksiyona
– Eritrositler % 99 – Lökositler <% 1 – Trombositler % 1

H A Y V A N F İ Z Y O L O J İ S İ
260
girerek, dokulardan akciğere bikarbonat iyonu (HCO3- ) halinde taşınır. Ayrıca eritrositler
içindeki hemoglobin çok iyi bir asit-baz tamponudur. Kanın şekilli elemanlarının(kanın yüzde 45 kısmının) % 99 unu alyuvarlar oluşturur. Alyuvarların şekilleri bikonkav disk şeklinde olup, çapları 6–8 mikrometredir(çoğunlukla 7 mikrometre). Alyuvarların çekirdeği yoktur (olgunlaşmış eritrositlerde). Sayıları cinsiyet, yaş ve yaşanılan yerin yüksekliğine göre değişebilmektedir. Eritrositler hemoglobin taşırlar. Eritrosit’in %34’ü Hemoglobin ’dir(her bir eritrositte 250 milyon hemoglobin). Hemoglobin miktarı erkeklerde 16 gr/dl– kadınlarda 14 gr/dl dir.
ŞEKİL 4.35 Eritrosit ve hemoglobin
Hemoglobin eritrositlere kırmızı rengini veren oksijenini zayıf bağlar kurarak taşıyan moleküldür ve eritrositlerin renkleri içerdikleri hemoglobin miktarına bağlıdır. Embriyonik yaşamın ilk birkaç haftasında vitellus kesesi, gebeliğin ikinci trimesteri sırasında dalak, lenf düğümleri ve karaciğer, gebeliğin son ayında ve doğumdan sonra kemik iliğinde oluşur. Beş yaşına kadar tüm kemikler, 20 yaşından sonra tibia ve humerusun proksimal bölümü dışında uzun kemiklerin ilikleri yağlanır ve alyuvar yapımı durur. 20 yaşından sonra vertebralar, sternum, kostalar ve iliyak kemik gibi yassı kemiklerin ilikleri, Yaş arttıkça bu kemiklerde de üretim düşer. Dolaşımdaki tüm hücreler kemik iliğindeki hemopoietik kök hücreden Hemositoblast kaynaklanır Bu kök hücreler birbiri ardına bölünerek dolaşımdaki hücreleri oluştururlar. Eritrosit Üretiminin düzenlenmesi doku oksijenasyonudur. Oksijenlenmeyi azaltan faktörler alyuvar yapımını artırır;
ŞEKİL 4.37 Kan Hücre Üretimi
Hemositoblast Miyeloyit ana hücresi. Kemik iliğinde bulunan ve her tip beyaz kan hücresi (monosit, granülosit, lenfosit) ile megakaryosit ve eritrositleri veren çok yönlü hücredir. Lenfoir ve myeloid seri ile kan hücreleri oluşturulur.
• Eritrosit 5.000.000 adet/mm3 • Toplam lökosit 7000 adet/mm3
Lökosit formülü: • Nötrofil 50–70% • Eozinofil 1–4% • Basofil 0.1% • Monosit 2–8% • Lenfosit 20–40% • Platelit 250,000 adet/mm3

H A Y V A N F İ Z Y O L O J İ S İ
261
Eritrosit yapımı için gerekli faktörler şunlardır: Eritropoietin, Fe, VitB12, Folik Asit. Alyuvar üretimini stimüle eden esas faktör eritropoietin hormonudur. Eritropoietin glikoprotein yapıdadır, % 90’ı böbreklerde, kalanı da karaciğer de yapılır. Eritropoietin aktivitesi için
sialik asid gereklidir. Eritropoietin kök hücreden proeritroblast oluşumunu artırır. Eritropoietin oksijenin az olduğu ortamda dakikalar içinde yükselmeye başlar, 24 saatte maksimuma ulaşır, ancak kanda yeni alyuvarlar 5 gün sonra görülür. Vitamin B12 ve folik asiti DNA sentezi için gereklidir. Yokluklarında DNA sentezi azalır, çekirdek olgunlaşması ve bölünme yetersiz olur. Olgunlaşma ve bölünmenin yetersiz oluşu makrosit adı verilen normalden büyük, hücre zarı dayanıksız, düzensiz, geniş, bikonkav olmayan, oval şekilli alyuvarlar oluşur. Vitamin B12 ve folik asit olgunlaşma yetersizliğine yol açar. Eritrosit membran proteinleri
GULT1: Eritrosit membranında yer alan glikoz kolaylaştırılmış difüzyon taşıma proteinleridir. Kolaylaştırılmış difüzyon mekanizmasıyla plazmada glikoz alımını düzenler GLUT1, düşük Km değerine sahiptir (enzim substrat ilgisi yüksektir Vmax/2).
Spektrin: Eritrositlerdeki protein moleküllerinin çoğu lipid tabakanın sitoplazmik yüzüne tutunmuş olan periferal zar proteinleridir. Bunların içinde de en bol bulunanı spektrin'dir. Spektrin yaklaşık 100 nm uzunlukta, uzun, ince, esnek, çubuk şeklinde olup zar proteinlerinin yaklaşık % 25 ini teşkil eder. Zarın altında yer alan sitoplazma iskeletinin esas elemanıdır. Spektrin iki büyük polipeptid zincirinden meydana gelmiştir. Bunlar: α-spektrin (~ 240 000 dalton) ve β-spektrin (~ 220 000 dalton) zincirlerdir. Esnek yapıdaki bu zincirler birbirlerine birkaç noktada nonkovalent olarak tutunmuştur. Spektrin heterodimerleri uç uca eklenerek 200 nm uzunlukta tetramerleri oluşturur, α ve β zincirler 106 amino asit uzunluğunda, çok sayıda tekrarlayan kısımlardan meydana gelmiştir.
Glikoforin A, B ve C: Eritrosit membranında yer alan önemli bir integral protein de glikoforindir. Tek bir polipeptid zincirinden oluşur. 16 tane oligosakkarit birimi içerir. %60’ ı karbonhidrattır. Glikoforin, kan grubu antijenleri taşımasına rağmen büyük ölçüde yapısal rol oynamaktadır. Glikoforin ailesi de transmembran proteinler ise de membranı boydan boya ancak bir kez kesen tek geçişli tiptedir. Glikoforin A, ana glikoforindir, ileri derecede glikoziledir. Eritrosit membranındaki sialik asitlerin ~ %90’ı bu proteine yerleşiktir. Glikoforin A, influenza virusu ve sıtma nedeni olan plazmodium falcifarum için bağlanma noktaları taşır.
– Düşük kan hacmi (kanama) – Anemi – Hemoglobin azlığı – Azalmış kan akımı – Akciğer hastalığı

H A Y V A N F İ Z Y O L O J İ S İ
262
ŞEKİL 4.38 Eritrosit membran yapısı.
Anyon değiş-tokuş proteini (Bant 3): Transmembran glikoproteindir. Karboksil ucu dış yüzeyde amino ucu stoplazmik yüzdedir. Çift katmanı en az on kez kesen çok geçişli bir membran protein örneğidir. Membranda büyük olasılıkla dimer halinde bulunur. İçinde klorun bikarbonatla değiştirilmesine izin veren bir tünel yapar. Dokularda oluşan karbondioksit eritrositlere bikarbonat halinde girer, akciğerde klorla yer değiştirir ve soluk havasında karbondioksit olarak atılır.
Hemoglobin Yapımı
Eritrositlerde bulunan ve kana kırmızı rengini veren hemoglobinin yapısında bulunan hem, önemli bir demir-porfirin bileşiğidir. Hemoglobinin en önemli özelliği oksijen ile gevşek bağlanmasıdır. Hemoglobin eritrositlerde bulunan ve oksijen taşıyan globuler bir proteindir. Hemoglobinin 1 gramı 1.34 ml oksijen bağlar ve 100 ml kan da 12–16 gr Hb bulunur. Oksijenin % 97 si hemoglobin ile % 3 ü plaznada çözünmüş olarak taşınır. Hemoglobinin hem bölümü mitokondride, globin bölümü ise endoplazmik retikulumda sentezlenir. Hem biyosentezi için temel prekürsör, glisin amino asididir. Hem sentezinde ilk reaksiyon ve son üç reaksiyon mitokondride, diğerleri sitoplazmada gerçekleşir. Hem grubu oksijeni bağlayan demir içerir. Hemositobalst hücresinin miyeloid farklılaşan hücrelerinden eritrositlere olgulaşan hücreler dönüşür bu hücreler hemoglobin sentezi yapar. Hemoglobin yapımı proeritroblastlarda başlar az miktarda retikülosit evresinde devam eder. Hemoglobin 4 hemoglobin zincirinin bir araya gelmesiyle oluşur.
Eritrosit membranından SDS poliakrilamit jel elektroforezi ile elde edilen proteinlerin ağırlıkları(dalton).

H A Y V A N F İ Z Y O L O J İ S İ
263
ŞEKİL 4.39 HEMOGLOBIN molekülü
Hemoglobinin alt tipleri vardır, yetişkinlerde en sık rastlanan hemoglobin A dır. Fetus hemoglobini erişkin insan hemoglobininden farklıdır. Fetusta hemoglobin sentezi proeritroblast, normoblast ve retikülositlerde olur. 12. haftadan önceki hemoglobin gower tip 1 ve gower tip 2 ve Hb Portland'dır. 2. ve 3. trimestır boyunca fetuste ana hemoglobinolarak HbF bulunurur. HbF iki alfa ve iki gama alt zincirinden oluşur. HbF'in oksijene aflinitesi erişkin tip hemoglobinden yani HbA'dan %50 daha fazladır.
ŞEKİL 4.40 Hemoglobin sentezi
Porfirin biyosentezi için ilk reaksiyon, mitokondride glisin ile süksinil-KoA’nın, α-amino-β-ketoadipat üzerinden δ-aminolevülinat (ALA) oluşturmak üzere birleşir. Hemin organik bölümü, sekiz adet glisin ve süksinil-CoA’dan oluşur. Bu ilk basamakta piridoksal fosfat (PLP, B6PO4), glisin ile Schiff bazı oluşturur. Glisinin α karbonunda oluşan karbanyon yapı, nükleofilik atak oluşumuna zemin hazırlar ve elektronsever molekül
Proforin Dört metiliden (-CH=) köprüsüyle birbirine bağlı dört pirol halkasından ibaret olan porfin halka sistemi ihtiva eden molekül sistemidir. Hemoglobin (1) Protoporfirin IX (2) globin, (3) Fe+ iyonundan meydana gelir
1. Hemoglobin Sentezi 2. 2 süksinil KoA + 2 glisin�pirol
3. 4 pirol � protoporfirin IX
4. Protoporfirin IX + Fe ++ � hem 5. Hem + globulin� hemoglobin (alfa veya beta)
6. 2 alfa+2 beta�Hemoglobin A

H A Y V A N F İ Z Y O L O J İ S İ
264
olan süksinil-CoAile birleşim sağlanır; δ-aminolevülinat (ALA) oluşur. Bu reaksiyonu katalizleyen enzim, ALA sentazdır. Mitokondride oluşan δ-aminolevülinat (ALA), sitoplazmaya çıkar. Sitoplazmada, 2 molekül δ-aminolevülinat (ALA), porfobilinojen (PBG) oluşturmak üzere kondense olur; reaksiyonu porfobilinojen sentaz (ALA dehidrataz) katalizler: ALA dehidrataz, Zn2+ gerektiren sitozolik bir enzimdir; sülfhidril grupları içerir; ağır metallerle inhibisyona çok hassastır. Kurşun ve kadiyum ile zehirlenmelerde ALA miktarı artar. Pb zehirlenmesimde ALA dehidrataz ve ferroşelataz aktiviteleri düşer. Eritrosit Zn protoporfirini yükselir. Porfobilinojen (PBG) molekülü, propiyonat ve asetat takıları içerir. Dört molekül porfobilinojen, bir seri enzimatik reaksiyon sonunda protoporfirin IX (Protoporfirin III) oluşturur. Mitokondride oluşan protoporfirin IX’a (Protoporfirin III) mitokondri içinde ferro halinde demir (Fe2+) katılmasıyla “hem” oluşur. 2,3-disfosfogliserat (2,3-DPG) eritrositlerdeki glikolitik yolda teşekkül eden önemli bir ara ürün olup bisfosfogliserat olarak da adlandırılmaktadır. Difosofliserat mutaz enzimi, 1,3-difosfogliseratı 2,3-difosfogliserat'a çevirir ve 2,3-difosfogliserat da 2,3-difosfogliserat fosfataz enzimi katalizörlüğünde 3-fosfogliserat'a hidrolizlenir Ayrıca, 2,3-DPG bir glikolitik yol enzimi olan monofosfogliserat mutaz enzimi için bir kofaktördür. Bazı kuşlarda 2,3-DPG bulunmamaktadır. Bunun yerine inozitol hekzofosfat bulunurki bunun hemoglobine olan ilgisi 2,3-DPG’ınkinden çok daha fazladır.
ŞEKİL 4.41 Hemoglobin molekülünde DPG etkisi
Demir Metabolizması
Demir hemoglobin, myoglobin ve sitokrom gibi yapıların sentezi için gereklidir. Vücuttaki toplam demir 4–5 gramdır (makro elementtir 1 gr fazla) ve bunun % 65 i hemoglobinde bulunur. Demir ince bağırsaklardan emilir. Plazmada bulunan apotransferrin ile birleşerek transferrini oluşturur ve plazmada transferrin olarak taşınır. Demir hücrelerde ise apoferritin ile birleşerek ferritin olarak depolanır. Ferritin depo demiridir. İnsan vücuda giren demir feritin yada farklı şekillerde depolanmasına

H A Y V A N F İ Z Y O L O J İ S İ
265
rahmen aktif olarak vücutan atılamaz. Demirin vücutan atılma yolu epitelyum dökülmesi, terleme, kan kaybıdır. Demir ayrıca hemosiderin olarak bilinen çözünmeyen formda da bulunur. İnsanda hergün yaklaşık 1 mg demir feçesle atılır.
ŞEKİL 4.42 Demir metabolizması
Kanamanın olduğu durumlarda kaybedilen demir miktar artar. Kadınlarda menstruel kanamalarda demir kaybı ortalama 2 mg /gün kadar çıkar. Demir emilimi yavaştır. Bu yüzden sindirimle alınan fazla miktradaki demirin ancak hemosiderin halde depolanır küçük bir kısmı kana geçebilir. Demir metabolizmasınında alınan miktar ile feçes ve terleme ve kadınlarda menstruel kana bağlı kayıp miktarın eşit olması gerekir.
ŞEKİL 4.43 Demirin hücre içine alınması
Hemosiderin: Hemoglobinden türeyen hemosiderin, altın sarısından- kahverengine kadar değişen renklerde görülen bir pigmenttir. Demirin hücre içinde birikme şekline örnektir. Kanamanın doğal sonucu hemosiderin pigmenti oluşur. Hücre içinde demir, apoferritin adı verilen proteine bağlı ferritin miçelleri şeklinde depolanır. Hücre ve doku içinde biriken demir histokimyasal olarak Berlin Mavisi denilen özel bir boya ile gösterilir. Makroskopik kanamalar veya yoğun vasküler konjesyonun neden olduğu mikroskopik kanamalar, demirin lokal artımını ve bunu takiben hemosiderini ortaya çıkarır. Buna en iyi örnek, zedelenmeden sonra görülen çürüktür (ekimoz). Çürükler, lokalize hemosiderozisin en iyi örneğidir. Kanama bölgesindeki eritrositlerin yıkımıyla ortaya çıkan kırmızı kan hücre artıkları, makrofajlar tarafından fagoside edilir. Hemoglobin içeriği lisosomlar tarafından katalize edilir ve hemosiderine dönüştürülür. Çürükte görülen renk değişikliği, bu dönüşümdeki aşamaları yansıtır.
Hepsidin Demir tüm canlı organizmalar için “esansiyel” bir elementtir. Son yıllarda bulunan peptid yapıda bir hormon olan hepsidin, demir metabolizmasını barsaklardaki demir emilimini, makrofajlardan demir çıkışını ve hepatik depolardan demir salınımını etkileyerek düzenler. Hepsidin hücrelerden demir çıkışını, ferroportine bağlanarak ve onun yıkımını uyararak engeller. Hepsidin sentezi, serum demirinin yükselişi ile artar, anemi ve hipoksi durumlarında azalır.
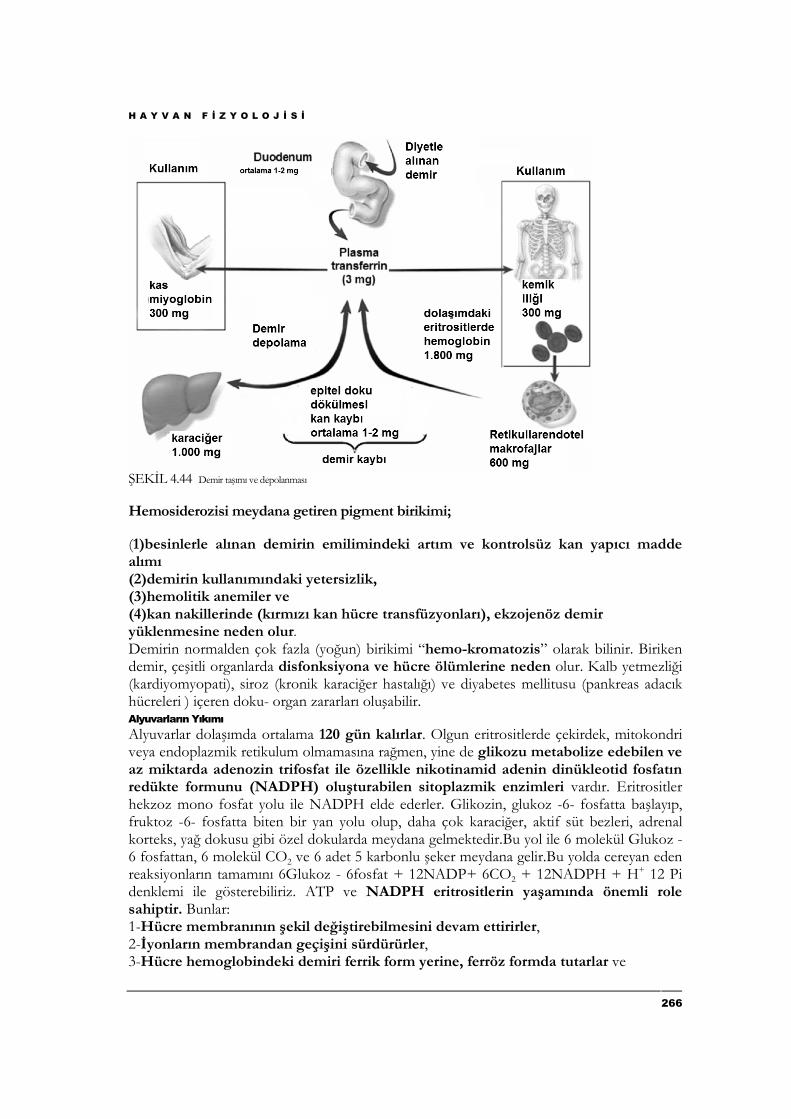
H A Y V A N F İ Z Y O L O J İ S İ
266
ŞEKİL 4.44 Demir taşımı ve depolanması
Hemosiderozisi meydana getiren pigment birikimi;
(1)besinlerle alınan demirin emilimindeki artım ve kontrolsüz kan yapıcı madde alımı (2)demirin kullanımındaki yetersizlik, (3)hemolitik anemiler ve (4)kan nakillerinde (kırmızı kan hücre transfüzyonları), ekzojenöz demir yüklenmesine neden olur. Demirin normalden çok fazla (yoğun) birikimi “hemo-kromatozis” olarak bilinir. Biriken demir, çeşitli organlarda disfonksiyona ve hücre ölümlerine neden olur. Kalb yetmezliği (kardiyomyopati), siroz (kronik karaciğer hastalığı) ve diyabetes mellitusu (pankreas adacık hücreleri ) içeren doku- organ zararları oluşabilir. Alyuvarların Yıkımı
Alyuvarlar dolaşımda ortalama 120 gün kalırlar. Olgun eritrositlerde çekirdek, mitokondri veya endoplazmik retikulum olmamasına rağmen, yine de glikozu metabolize edebilen ve az miktarda adenozin trifosfat ile özellikle nikotinamid adenin dinükleotid fosfatın redükte formunu (NADPH) oluşturabilen sitoplazmik enzimleri vardır. Eritrositler hekzoz mono fosfat yolu ile NADPH elde ederler. Glikozin, glukoz -6- fosfatta başlayıp, fruktoz -6- fosfatta biten bir yan yolu olup, daha çok karaciğer, aktif süt bezleri, adrenal korteks, yağ dokusu gibi özel dokularda meydana gelmektedir.Bu yol ile 6 molekül Glukoz - 6 fosfattan, 6 molekül CO2 ve 6 adet 5 karbonlu şeker meydana gelir.Bu yolda cereyan eden reaksiyonların tamamını 6Glukoz - 6fosfat + 12NADP+ 6CO2 + 12NADPH + H+ 12 Pi denklemi ile gösterebiliriz. ATP ve NADPH eritrositlerin yaşamında önemli role sahiptir. Bunlar: 1-Hücre membranının şekil değiştirebilmesini devam ettirirler, 2-İyonların membrandan geçişini sürdürürler, 3-Hücre hemoglobindeki demiri ferrik form yerine, ferröz formda tutarlar ve

H A Y V A N F İ Z Y O L O J İ S İ
267
4-Eritrositlerdeki proteinlerin oksidasyonunu önlerlersitoplâzmada taşır. Dönüştürülerek sindirim yolu ile atılır. Yaşlı alyuvar zarı kolay zedelenebilir hale gelir. Çoğunluğu dalak olmak üzere, karaciğer ve kemik iliğinden geçerken parçalanırlar. Bu nedenle dalağı uzaklaştırılması kandaki yaşlı eritrosit sayısını artırır. Dalak, karaciğer ve kemik iliğindeki makrofajlar tarafından fagosite edilirler ve içlerindeki hemoglobin açığa çıkar. Makrofajlar hemoglobinden demiri ayırarak kana verirler. Demir kanda transferrine bağlanır.
ŞEKİL 4.45 Demir metabolizması
Hemoglobinin geri kalan kısımları bilirubine çevrilip kana verilir ve daha sonra karaciğer tarafından safraya sekrete edilir. Kısaca yaşlı eritrositler dalak karaciğer ve kemik iliğinde sürekli uzaklaştırılır. Demir yeni eritrosi sentezi için kullanılırlarken hemoglobini oluşturulan globin proteinleri yıkılır. Bilirubin: Bilirubin, safrada bulunan ve safranın sarı-yeşil rengini veren başlıca pigmenttir. Kırmızı kan hücrelerinin mononükleer fagositik sistemde parçalanmasıyla (karaciğer- deki kupffer hücrelerinde) serbestleşen hemoglobinden türemiştir; fakat demir içermez(demir transferinle kan yapıcı dokulara ve karaciğer gönderilir). Organizmada normal yaşam sürelerini (100- 120 gün) tamamlayan bu eritrositlerin parçalanmasıyla konjuge olmamış (ankonjuge) bilirubin meydana gelir. Bu ankonjuge bilirubin, kan proteinlerine (albumin) bağlanarak karaciğer parankim hücrelerine (hepatosit) taşınır ve burada işlenerek konjuge bilirubine çevrilir. Bu işlem spesifik bir enzim (bilirubin uridindifosfat glukuronosil transferas) ile oluşur. Daha sonra safra aracılığıyla bağırsağa dökülür. Bağırsaktaki bakteriyel enzimlerin etkisiyle “urobilinojen”e dönüştürülür. Bu pigmentin bir bölümü (% 20) tekrar barsaktan geri emilerek (reabsorbe olarak), karaciğere döner. Bunun bir bölümü de idrarla atılır. Barsaktaki urobilinojenin geri kalan bölümü, daha ileri bir işlemle “ürobilin” (stercobilin)’e dönüşür. Dışkının bilinen rengini (sarı- kahverengi) veren bu maddedir. Konjuge bilirubin; suda çözünür, nontoksiktir ve idrarla atılır. Konjuge olmamış (Ankonjuge) bilirubin suda çözünmez, idrar ile atılmaz, toksiktir ve bilirubinin bilinen bütün toksik etkilerinin nedenidir. Kan plasmasında total bilirubinin normal miktarı 100 ml’de 0.3- 1 mg’dır. Kandaki bilirubin düzeyi (hem konjuge hem de ankonjuge) 2- 3 mg’ın üzerine çıktığında (bazı durumlarda 30- 40 lara

H A Y V A N F İ Z Y O L O J İ S İ
268
çıkabilir), deri ve sklerada sarı bir renk oluşur. Bu renk değişikliği, dokuların safra pigmenti birikimine bağlı olarak, sarıya boyanmasından ileri gelmektedir. Klinik olarak “sarılık” (ikter) diye tarif edilir ve meydana geliş biçimlerine göre şöyle incelenebilir. (1)yoğun eritrosit yıkımı (hemoliz artması), (2)hepatosellüler disfonksiyon ve (3)intrahepatik veya ekstrahepatik safra obstrüksiyonu ile safranın tutulması (kolestaz) sonucu sarılık ortaya çıkar. Anemi
Çeşitli nedenlerle kandaki alyuvar sayısının azalmasına anemi denir. Anemi farklı nedenlerle olabilir. Eritrositleri ömürleri sonunda yıkılır. Bu nedenle eritrosit kaybı (kan kaybı-kanama) ya da çok yavaş üretimi (B12 vitamini ve demir eksikliği ve sürekli olarak yaşlı eritrositlerin kandan uzaklaştırılmasına bağlı olarak) anemiye nedeni olabilir.
o Kan Kaybı Anemisi: Hızlı kan kayıplarından sonra kanın sıvı kısmı plazma 1-3 gün içerisinde yerine konur, fakat alyuvarlar hemen yerine konulamaz ve eritrositlerin kandaki konsantrasyonu azaılır. Hipoksiyaya bağlı eritropoetin üretimi ile yeni eritrosit üretimi artırılır. İkinci bir kanama olmazsa alyuvar konsantrasyonları 3–6 haftada normale döner.
o Aplastik Anemi: Aplazi bir organ veya oluşumun doğuştan yokluğu. Kemik iliği aplazisi yani fonksiyonel kemik iliginin yokluğu nedeniyle oluşan kansızlığa aplastik anemi adı verilir. Şiddetli nükleer bombardıman(yüksek enerjili ışınlar, Aşırı X ışını uygulamaları, endüstriyel kimyasal maddeler (kurşun, benzen, arsenik) aplastik anemiye neden olur.
o Megaloblastik Anemi Vit B12, folik asit ve intrensek faktör gibi alyuvar yapımında etkili faktörlerden herhangi birinin azalması kemik iliğinde alyuvar üretimini yavaşlatır. Sonuçta tuhaf, geniş şekilli, olgunlaşmamış megaloblastlar oluşur.
o Pernisiyöz Anemi Vitamin B12 nin yetersiz emilimi nedeniyle oluşan olgunlaşma kusuruna denir. Temel nedeni mide mukozasından salgılanan ve Vitamin B12 ye bağlanarak reseptöre bağlı pinositozla alınmasını sağlayan intrensek faktör olarak bilinen bir glikoproteinin yetersiz salgılanmasıdır.
o Hemolitik Anemi Doğuştan kazanılan çeşitli alyuvar anomalilikleri(örneğin hekzoz mono fosfat yolundaki bir enzim eksikliği G6FDH ) hücreleri frajil(kolay kırılır, narin) hale getirerek özellikle dalaktaki kapillerlerden geçerken kolaylıkla parçalanmalarına yol açar. Sonuçta alyuvarların yaşam süresi oldukça kısa olduğu için ciddi anemilere yol açabilir. Herediter sferositozis, orak hücre anemisi ve eritroblastozis fetalis hemolitik anemiye yol açan durumlardır.
o Herediter Sferositozis Alyuvarlar bikonkav disk yerine küreseldir ve çok küçüktürler. Esnek değildirler ve hafif bir sıkışmada bile kolayca parçalanırlar.
o Orak Hücre Anemisi Anormal tipteki hemoglobin (hemoglobin s) molekülü nedeniyle oluşur. Meydana gelen eritrositlerin uzun ve kırılgan yapıdadır. Hızla parçalarak anemiye yol açar.

H A Y V A N F İ Z Y O L O J İ S İ
269
Polisitemi
Polisitemi kandaki eritrosit sayısının artmasıdır. Sekonder polisitemi ve Polisitemia vera olarak bilinen iki türü vardır.
• Sekonder Polisitemi Yüksekliğe bağlı olarak basınç azalır. Daha düşük basınçta havanın yüzde 21 kısmını oluşturan oksijen miktarıda azalır (Oksijenin kısmı basınçının azalmasına bağlı olarak) Bu nedenle yüksek yerlerde daha az oksijen vardır. Fizyolojik polisitemi: 4-5 bin metrede yaşayanlarda görülür. Daha az oksijene maruz kalan organizmada kalp yetmezligi-oksijen taşıma yetersizliği durumlarında dokuların oksijensiz kalması ve buna bağlınarak eritrosit yapıcı organların aktivitesini(kemik iliği) artırır, buna sekonder polisitemi denir. Eritrosit sayısı 6–7 milyon/mm3 kadar çıkar.
• Polisitemia Vera-Eritremi Eritrosit üreten hücre serisinde gen anormalliği nedeniyle oluşur. Alyuvar sayısı 7–8 milyon/mm3 e çıkar, Hematokrit % 60–70, Kanın viskozitesi 3 katı kadar kadar artarak suyun 10 misline çıkar. Kısaca genetik bir bozukluğa bağlı olarak aşırı eritrosit üretimidir.
Kan Grupları
30 kadar eritrosit membran antijeni vardır bunlar içinde ABO ve Rh grupları kan nakilleri için önemlidir ABO kan grupları eritrosit membran yüzeyinde bulunan A ya da B antijenlerine (aglutinojen) göre gruplara ayrılır. Bu antijenler eritrosit membran yüzeyindeki glikoproteinlerdir (glikoforine [ceramid] farklı sakkaritler bağlayan farklı gilikosil transferaz olmasına bağlı olarak). Her kan grubunda antijen(A ve B) uygun olarak plazmada bu antijenler ile reaksiyona girebilen antikorlar (aglutininler) bulunur. Kanda eritrosit membran yüzeyinde A antijeni(glikoproetini) varsa yani kan A grubu ise, aynı kanda B antijenine ait (B grubu kana ait) antikorlar plazmada bulunur. Kandaki bir tip antijen (aglutinojen) diğer kan grubunun antikorları ile reaksiyona girer. Buna aglutinasyon reaksiyonu denir. O kan grubu olan kişilerdeki eritrositleredki glikoprotein yapıları çökelme özelliği göstermeyen H antijenidir.
ŞEKİL 4.46 AB kan antijenleriıni oluşturan oligosakkarit şeker grupları
Doğum sonunda bebekte plazmasına aglutinin bulunmaz ancak ilk aylardan beslenme ve barsakta yerleşen bakteriler karşı oluşturulan aglutininler eritrositlerde olmayan aglutinojenlere karşı üretilir. AB grubunda plazmada aglutinin üretimi yapılmaz iken O kan grubunda her iki tip aglutinin sentezi yapılır.

H A Y V A N F İ Z Y O L O J İ S İ
270
ŞEKİL 4.47 ABO kan antijenlerinin moleküler formülü
ABO kan grubu dışında kan transferlerinde önemli olan diğer bir kan grubu sınıfı Rh grublamasıdır. Rh antijenlerinin Rh faktörü olarak adlandırılan ve sık görülen 6 tipi vardır. Bunlar C, D, E, c, d ve e olarak adlandırılırlar. Tip D antijeni toplumda çok yaygındır ve diğer gruplara göre daha antijeniktir. Bu antijeni taşıyanlar Rh (+), taşımayanlar Rh (-) olarak değerlendirilir. Beyaz ırkın yaklaşık %85'i Rh (+) ve % 15'i Rh (-) grubudur.
Eritroblastosis fetalis
Eritroblastosis fetalis fetus ve yenidoğanın eritrositlerinin aglütinasyonu ve fagositozu ile kendini gösteren bir hastalıktır. Anne Rh (-), baba ise Rh (+) tir. Bebek babasından Rh (+) antijeni alır, anne ise bebeğin Rh antijeni ile karşılaştıktan sonra anti-Rh aglutininler geliştirir. Plasentadan geçen aglutininler fetusun kanında eritrosit aglutinasyonuna yol açar. Fetüsün Rh (+) alyuvarlarının Rh (–)annenin antikorları tarafından haraplanmasıyla oluşur. Bu antikorlar hücreleri frajil hale getirerek bebeğin ciddi anemi ile doğmasına yol açarlar. Rh faktörü yeni doğanda hemolitik bozukluğa yol açar. Fetüs RH+, anne RH- ise ve anne daha önce Rh+ fetüslü bir gebelik geçirmişse Rh faktörü fetüs için ölümcül olabilir. Rh bağlı aglutinasyon olması için daha önceden farklı grup Rh temas gereklidir. Maternal ve fetal kan grubu uyuşmazlığı sonucu annede oluşmuş olan antikorların, fetus’da neden olduğu bir hemolitik anemidir. Rh(-) bir annenin fetusu, babanın ki gibi Rh(+) olursa, anne ve onun bebeği arasında Rhesus (Rh) uyuşmazlığı meydana gelebilir. Anne; Rh antijeninden yoksun (Rh-) ise, fetusda mevcut olan Rh antijenlerine (Rh+) karşı antikorlar üretir. Rh(-) anne eritrositleri, Rh(+) fetus eritrositleri tarafından sensitize edilmiştir. Fetal eritrositler gebelik boyunca katılır. En büyük geçiş, doğum esnasında olur. Oluşan bu plasentadan sızarak annenin dolaşımına antikorlar, sonraki gebeliklerde plasenta yolu ile fetusa geçerek, fetusa ait kırmızı hücrelerin destrüksiyonuna (lizise, hemoliz) neden olur. Ortaya çıkan sendrom, “eritroblastosis fetalis” olarak bilinir. Yenidoğanın bu hemolitik hastalığında meydana gelen anemi, uterus içinde fetal ölüme yol açabilecek kadar şiddetli de olabilir. Anemiye reaksiyon olarak fetal kemik iliği, olgunlaşmamış eritrositleri (eritroblastları) fetusun dolaşımına katar. Eritroblastosis fetalis terimi; oluşan eritrosit destrüksiyo nunu kompanse etmek için, fetal dokulardaki kırmızı kan hücre prekürsörlerinin (hematopoesis) aşırı artmasını anlatır.

H A Y V A N F İ Z Y O L O J İ S İ
271
Rh uyuşmazlığının patogenezindeki sensitizasyonun önemi anlaşıldıktan sonra, bu hastalık belirgin bir şekilde kontrol altına alınmıştır. Rh sisteminin içerdiği pekçok antijenden yalnızca D antijeni, Rh uyuşmazlığının başlıca nedenidir. Rh(-) anneye, Rh(+) bebeğin doğumundan hemen sonra, anti- D globulin uygulanmaktadır. Anti- D antikorlar, doğum sırasında maternal dolaşıma sızan fetal eritrositlerdeki antijenik bölgeleri maskeleyerek, Rh antijenlerine karşı olan duyarlılığı engeller.
ŞEKİL 4.48 Kan uyuşmazlığı
Eritroblastosis fetalis; belirtilerine göre üç sendroma ayrılabilir. Şiddetli komplikasyonlar olmadan yaşam mümkün olan, yalnızca hafif anemiyle seyreden “yeni doğanda konjenital anemi” olarak adlandırılır. Şiddetli hemoliz vakalarında anemiye bariz sarılık eşlik eder, “ikterus gravis” sendromu oluşur. Dolaşım bozukluğundan, anazarka denilebilecek kadar şiddetli bir ödemin ortaya çıkışı, buna eşlik eden sarılık, “hidrops fetalis” olarak adlandırılan bir klinik tabloyu da ortaya çıkarabilir. Trombositler
Kan şekil elemanları olan trombositler eritrositlerden sonra kanda en fazla bulunan şekilli elemandır. Karaciğer ve böbrekler atarfıından üretilen glikoproetin yapısında olan bir hormon olan trombopoietin tarafından megakaryosit hücre sitoplazmalarından oluşurlar. Trombositler 1–4 µçapında(kandaki en küçük şekilli) eleman yuvarlak oval diskler şeklinde görülür. Trombositin üç farklı yapıdan oluşur. Hücre zarı, kanaliküler sistem ve granüller.
• Hücre zarı; trombosit fonksiyonunda önemli rol oynar. Hücre zarında çok miktarda fosfolipid ve daha az miktarda ise glikoprotein bulunmaktadır. Hücresel adhezyon molekül ailesinde selektin sınıfı (P-selektin trombosit zarında bulunur) özel sayesinde hasar görmüş damar çeperine tutunma özelliği vardır.

H A Y V A N F İ Z Y O L O J İ S İ
272
ŞEKİL 4.49 Trombositleri P selektin aracılığı ile monositleri aktive etmeleri ve hasarlı damar kısmını tutunmaları
• Glikoproteinler normal endotel yüzeyine değil de hasarlı bölgeye bağlanırlar. Glikoprotein-Ib(GP-Ib) trombositin kollajene bağlanmasını, Glikoprotein-IIb(GP-IIb) trombositin Von-Willabraund Faktör(vWF)’e bağlanmasını, Glikoprotein-IIIa(GP-IIIa) ise trombositin trombosite fibrinojen aracılığıyla bağlanmasını sağlar.
• Kanaliküler Sistem: Prostaglandin çevirici enzim ve kontraktil kalsiyum içerir. • Granüller: yoğun Granüller; Serotonin(vazodilatör), adenozin di fosfat(ADP),
Ca+2 içerirler. Trombositler kemik iliğinde myeloid serideki dev megakaryositlerden oluşur. Olgunlaşmaları 5 gündür. Ömürleri dolaşımda 8–10 gündür, nükleusu yoktur. Kanda miktarı 150.000-400.000/mm3 dir. Trombositlerin en önemli fonksiyonlarından biri hemoztaz ve pıhtı büzüşmesindeki rolleridir. Trombositlerin yapışkanlığı; trombositlerin hem birbirlerine yapışmasını sağlar hemde endotelyumdaki doku matriksinin kaplamasının çıkarılmasını sağlar. Aktive olan trombositler, içyapılarında çözülmeye uğrayarak ADP, tromboksan A2, von Willebrand faktörü (VWF), fibronektin, serotonin, faktör V, trombospondin, trombositlerden türeyen büyüme faktörü, heparinaz ve diğer faktörleri salıverirler. Tromboksan A2 ve ADP, diğer trombositlerin kollajene bağlı olanlara yapışmalarını sağlarlar; böylece gevşek ve geçici trombosit tıkacı (beyaz trombüs) oluşur. Bir glikoprotein olan von Willebrand faktörü (VWF), trombosit tıkacı oluşumunu kolaylaştırır.
ŞEKİL 4.50 Trombosit tıkacı oluşması mekanizmaları

H A Y V A N F İ Z Y O L O J İ S İ
273
Kanın Pıhtılşaması(hemoztaz)
Kan ve dokularda pıhtılaşmayı etkileyen birçok faktör bulunmaktadır. Bunların bir kısmı pıhtılşamayı aktive ederken prokoagülan (pıhtılaşmayı sağlayan) ve bir kısmıda pıhtılaşmayı engelen antikoagülanlar (pıhtılaşmayı inhibe eden) maddelerdir. Kan pıhtılaşması bu iki grup madde arasındaki dengeye bağlıdır. Normal kanda antikoagülanlar baskındır ve kan pıhtılaşmaz; fakat damar zedelendiğinde prokoagülanlar baskın hale gelir ve pıhtı oluşur. Damarın içindeki endotel bir şekilde hasar gördüğünde altındaki kollajen (bağ dokusu) açığa çıkar, aktive olan kan pulcukları kollajene bağlanır. Hasarlı bölge üzerine kan pulcukları kümelenir ve trombotik tıkaçı oluştururlar. Bunun (oluşan tıkacın) sonucu olarak da içerdikleri ettikleri granüllerin içeriğini ortama boşaltırlar. Ortama boşaltılan bazı maddeler yüzünden kan pulcukları birbirlerine bağlanırlar, yeni gelen kan pulcukları hasarlı yüzeye bağlanmış kan pulcukları bağlanır. Ayrıca granüllerin içeriği ortama boşaldığında ortaya çıkan serotonin salınımı damar duvarındaki düz kasların kasılmasına neden olarak hasalanmış bölümden kan akımını engeller. Bunun nedeni serotonininin vazokonstrüktör olmasıdır. Ayrıca agregasyon sırasında kan pulcuklarında yüksek oranda bulunan miyozin ve aktin filamentleri kasılarak oluşan tıkacı güçlendirirler. Kan pulcukları plazmada bulunan fibrinojene ilave olarak fibrinojen salgılar. Bunun sonucu olarak pıhtılaşma sırasında daha çok fibrinojen fibrine dönüşürek, daha çok (trombosit ve diğer) kan hücrelerinin tutunacağı fibröz ağ oluşturur. von Willebrand faktörü hasarlı damar duvarına yapışarak kan pulcuklarının buraya tutunmasını kolaylaştırır. von Willebrand faktörü eksikliği veya bozukluğunda koagülasyon bozuklukları görülür.
ŞEKİL 4.51 Pıhtılaşma mekanizmasında von willbrand faktörünün foksiyonu
Damar duvarı(endotel) prostasiklin (PGI2) isimli, kuvvetli bir trombosit agregasyon inhibitörü (engelleyici) sentezler. Böylece kan pulcukları tıkacı sadece hasarlı bölgede oluşur ve yayılamaz. Hasarlanmış kan damarlarının duvarında pıhtı oluşumu ve damar sistemi içindeki kanın sıvı durumda tutulurken kan kaybının önlenmesi işlemidir. Damar zedelenmesine ilk cevap (damar endotelinden salınan ADP ve serotonine bağlı) vazokonstriksiyon ve karşılıklı endotel yüzeylerinin bir araya gelmesidir. Sonraki olaylar

H A Y V A N F İ Z Y O L O J İ S İ
274
trombosit tıkaç oluşması ve pıhtılaşmadır. Trombositler, thrombostenin ler denilen kontraktil propteinler içerirler. Trombositler kasıldığı zaman pıhtının hacmini azaltırlar, damarın iki ucunu birbirine yaklaştırarak, hasarlı bölgenin hacmını azaltırlar ve daha kolay iyileşme sağlarlar.
Pıhtı oluşumda ise temel 3 olay vardır: 1-Damarın yırtılması ya da kanın kendisinin hasarlanmasına cevap olarak gelişen kimyasal reaksiyonlar sonucu, protrombin aktivatörü denen bir kompleksin oluşumu. 2-Protrombin aktivatörünün etkisiyle protrombinin trombine dönüşümü. 3-Trombinin fibrinojeni fibrin iplikçiklerine çevirmesi ve fibrin iplikçiklerinin trombositler, kan hücreleri ve plazmayı da içine alarak pıhtı oluşturması.
ŞEKİL 4.52 Trombosit tıkacı
Bir kanamada 3 hemostatik mekanizma çalışır; 1. Kan damarlarının kasılması (vazokonstriksiyon) 2. Trombositlerin bir araya toplanıp tıkaç oluşturması- trombosit tıkacı (aggregation) 3. Kanın pıhtılaşması (koagülasyon)

H A Y V A N F İ Z Y O L O J İ S İ
275
Pıhtılaşmanın son ürünü plazma proteini olan fibrinojenin fibrine çevrilmesidir. Bu reaksiyonda trombin adı verilen enzime gereksinim duyulur. Fibrin kan hücreleri ile birlikte pıhtıyı oluşturur. Pıhtılaşma ekstrensek ve intrensek yollar ile gerçekleşir. Trombosit Aktivatörler: Bilinen birçok kan pulcukları aktivatörü (etkinleştirici) vardır. Bunlardan bazıları:
� Trombositler zedelenmiş damarda açığa çıkan kollajene bağlanırlar ve sekretuar vezikül içeriklerini boşaltırlar.
a. Bunlar trombosit aktivasyonu ve agregasyonuna yardım ederler. b. Bu işlem endotel hücreleri tarafından salınan von Willebrand faktörü ve Trombositler tarafından salınan tromboksan A2 tarafından güçlendirilir. c. Fibrin agrege olan Trombositler arasında köprü oluşturur.
Trombosit tıkaç normal endotel boyunca ilerlemez çünkü somaki bölümden trombosit agregasyonunu inhibe eden prostasiklin ve nitrik oksit salınır. Bu maddeler trombositleri agregasyonu inhibe eder ve sınırlandırır.
Trombosit İnhibitörler
Zedelenmiş damarda fibrinojen fibrine dönüşür ve fibrin birbirine bağlanıp ağ oluşturur böylece kan katı bir jel haline dönüşür. Bu reaksiyon ve fibrin ağını stabilize eden faktör XIII’ün aktivasyonu trombin tarafından katalizlenir. Trombin protrombinden bir kaskad reaksiyonunun sonunda oluşur. a. Trombin Trombositleri ve birçok pıhtılaşma faktörünü aktive ederek kaskad reaksiyonlarına pozitif geribildirim yapar. b. Trombosit faktörü içeren ve birçok aktive plazma faktörü için bağlanma bölgesi olan aktive trombositler kaskad için gereklidir. Vücutta pıhtılaşma kaskadı genellikle doku faktörünün faktör Vlla ile kompleks yapmasıyla ekstrensek yol aracılığıyla başlar. Bu kompleks, faktör X’u aktive eder. Faktör X az miktarda protrombinin trombine dönüşümünü katalizler. Trombin; faktör XI, VIII ve Trombositleri aktive ederek intrensek yol aracılığıyla büyük miktarlarda trombin oluşmasını sağlar.
• Kollajen, özellikle von Willebrand faktörü ile beraber • Trombin • Tromboksan A2 • ADP • Konvulksin
• Prostasiklin • Nitrik oksit

H A Y V A N F İ Z Y O L O J İ S İ
276
ŞEKİL 4.53 Aroşidomik asit metabolitleri
Kanda K vitamine bağlı olarak üretilen pıhtılaşma faktörleri vardır. Bunlar Faktör VII, IX, X protrombin II, protein C ve S’dir. Trombositler pıhtılaşmada temel görevlerden birini görürler. Trombositler sadece yaralanan yerde tıkaç oluşturmakla kalmazlar, içlerindeki granüllerden salgıladıkları maddelerle hem damar büzüşmesine katkıda bulunur, hem diğer trombositlerin olay yerine gelmesini sağlar, hem iç pıhtılaşma yolunun başlaması için gerekli maddeleri ortama salar hem de pıhtılaşma faktörlerinin aktiflenmesi için gerekli fosfolipid yüzeyi oluştururlar. İç ve dış yolların birleşip ortak yolu oluşturmasından sonra meydana gelen trombin geriye dönerek trombositlerin bir araya gelmesine katkıda bulunurken aynı zamanda daha fazla trombin oluşmasına da yardımcı olur.
1964 yılında ortaya atılan kan pıhtılaşma kaskad sistemi(ekstersek ve intersek yol) günümüzde geçerli değilidir. Günümüzde kan pıhtılaşma kascad sistemi endotel zedelenmesimesine bağlı olarak subendotel hücrelerince üretilen doku faktörleri pıhtılaşmayı başlatır. Başka bir ifade ile kan pulcukları kollajen ile temas ettiklerinde aktive olurlar.
Araşidonik asit metabolitleri vazodilatasyona neden olan araşidonik asit metabolitleri Prostaglandinler ve lökotriyenlerdir. Araşidomik asit metabolizması sonucu açığa çıkan ürünler birçok biyolojik olayları etkiler. Her hücre yaralanması, fosfalipaz A2 yi aktive ederek araşidonik asit gibi 20 karbonlu poliansature yağ asitlerini yıkar. Bunun sonucunda yangı oluşur. Araşidomik asit poliansature bir yağ asididir ve hücre zarındaki fosfolipid'lerde önemli miktarlarda bulunur. Yangısal reaksiyon esnasında, nötrofil lizozomlarının, fosfolipaz'ların önemlidir. Lökotriyenler özellikle allerjik reaksiyonlarda indükleyici görev görür. Reaksiyon başladıktan sonra Araşidomik asit metabolizması iki temel yoldan birini seçer. Bunlar; Siklooksijenaz yolu ve Lipooksijenaz yolu'dur

H A Y V A N F İ Z Y O L O J İ S İ
277
Ekstrensek Yol
Damar duvarı ve çevresindeki dokuların travmaya uğramasıyla baslar. Zedelenmiş endotel hem trombositlerin yapışması için zemin görevi görür, hem de pıhtılaşmayı başlatan doku faktörünü (faktör 3) sentezler. Doku faktörü faktör VII ile birleşir. Doku faktörü-faktör VIIa kompleksi hem faktör X’u aktive eder. Bu aktivasyona yardımcı faktör Faktör V tir. Böylece ekstrensek yol aktive olmuş olur. Faktör X (protrombin aktivatörü) protrombini aktive edip trombin yapar. Trombin ise fibrinojeni fibrine dönüştürür. Faktör XIII ise fibrin monomerlerinin arasında çapraz kovalan bağlar kurarak fibrin molekülünü polimerize edip sağlamlaştırır. Ekstrensek yol protombin zamanı (PT) ile kontrol edilir. Normali 12–15 sn dir. İntrensek Yol
Kan içindeki yapıların hasarlanmasıyla başlar. Bu durumda çıkan trombosit fosfolipidlerinin serbestlenmesi (PF3), HMWK (Yüksek molekül ağırlıklı kininojen), prekallikrein faktör XII nin aktivasyonuna neden olur. Aynı faktörler Faktör XI de aktive edebilmektedir. Faktör XI, faktör IX u aktive eder. Faktör IX, VIII, PF3 beraber Faktör X u aktive ederler. Bundan sonraki kısım ekstrensek yoldaki ile aynıdır. intrensek yol a-PTT (aktive Parsiyel Tromboplastin zamanı) ile kontrol edilir. Normali 25-40 sn dir.
ŞEKİL 4.54 Pıhtılaşma yolu
Pıhtılaşma Faktörleri
Kanda pıhtılaşmayı hızlandıran bazı faktörler vardır, bunlara pıhtılaşma faktörleri denir. Bunlar faktör adı ile isimlendirilirler ve I den XIII e kadar numaralandırılırlar. Heparin ve antitrombin-heparin kofaktör kanamanın olmadığı durumlarda pıhtılaşmayı önler.

H A Y V A N F İ Z Y O L O J İ S İ
278
Kan damardan alınıp pıhtılaşmayı engelleyen bir madde katılmadan bir tübe bırakılırsa pıhtılaşır (ekstresek yol).
Pıhtılaşma Faktörleri
Faktör I : Fibrinojen
Faktör II : Protrombin
Faktör III : Doku faktörü(Doku tromboplastini)
Faktör IV : Kalsiyum
Faktör V : Proakselerin(labil faktör) Faktör VII : Serum protrombin konversiyon akseleratörü
Faktör VIII : Antihemofilik faktör A
Faktör IX : Plazma tromboplastin komponenti (Christmas faktör. Antihemofilik faktör B)
Faktör X : Stuart-Prower faktör Faktör XI : Plazma tromboplastin (Antihemofilik faktör C)
Faktör XII : Hageman faktör
Faktör XIII : Fibrin stabilize edici faktör Fibrinojen plazmada 100–700 mg/dl seviyesinde bulunur. Fibronojen karaciğerde sentezlenir. Trombin (proteaz yapısında) aktive olduğunda fibrinojen molekülünden fibrin monomerleri oluşumunu sağlar. Fibrin monomerleri birleşerek fibrin iplikçiklerini oluşturur. Protrombin plazma proteini (plazmada 15 mg/dl) α2-globulin Parçalandığında trombine dönüşür. Karaciğerde sentezlenmesi için K vitamini gerekir. Pıhtılaşmanın Sınırlandırılması
Doku faktörü TF (FIII) karaciğer ve diğer dokularda fagositlerce plazmadan uzaklaştırılır. Hepatositler aktif pıhtılaşma faktörlerini yıkar. Lokal kan dolaşımı pıhtılaşma ürünlerinin, uyarıcılarının yoğunluğunu, trombosit kümelerini azaltır. Trombin, fibrin tarafından absorbe edilir (Antitrombin I aktivitesi). Trombomodülin; Trombin bağlar; Protein C’yi aktive eder. FV ve FVIII inaktive olur, Protein S kofaktördür.
Karaciğer protrombin ve diğer pıhtılaşma faktörlerinin sentezi için K vitaminine ihtiyaç duyar. K vitamini eksikliğinde örneğin walfarin gibi K vitamini yerine geçen maddelerin varlığında pıhtılaşmaya geçikir yada hiç olmaz. Pıhtılaşma 3 olayla sınırlandırılır: (1) Doku faktörü faktör Vlla kompleksinin doku faktörü yolu inhibitörü tarafından inhibe edilmesi;
(2) Trombin tarafından aktive edilen protein C’nin faktör Va ve VIIIa’yı inhibe etmesi;
(3) Antitrombin III’ün trombin ve birçok pıhtılaşma faktörünü inhibe etmesi.
Aspirin prostaglandin H2 sentetaz-1 enzimi üzerindeki serin kısmına asetil grubu transfer ederek pihtılaşmayı engel olur.

H A Y V A N F İ Z Y O L O J İ S İ
279
Oluşan pıhtı fibrinolitik sistem tarafından eritilir. a. Plazminoje aktivatörleri aracılığyla plazminojenen oluşan plazmin fibrini sindirir. b. Doku plazminoen aktivatörleri edotel hücreleri tarafında sentezlenir ve pıhtıdaki fibrin tarafından aktif edilir.
Heparin :Tüm vücutta perikapiller bağ dokusuna yerleşmiş olan bazofilik mast hücreleri tarafından yapılır. Glokoamin-N-sülfirik asit ve glukuronik asit esterin kurulmuş yüksek negatif yüklü konjuge bir polisakkarittir. Antikoagülan etki gösterir. Pıhtılaşmaya engel olması trombonin fibronojen üzerindeki etkisini inhibe ederek gösterir. Tek başına antikoagülan etkinliği çok azdır. Antitrombin III ile birleşerek trombini uzaklaştırır. Heparin-antitrombin III kompleksi faktör XII, XI ve X’ u ortamdan uzaklaştırarak antikoagülan aktiviteyi arttırırlar. İntravenöz(damar içi) antikoagülan olarak kullanılır. 0.5- 1 mg/kg gibi düşük dozların enjeksiyonu yaklaşık 6 dk olan pıhtılaşma zamanını 30 dk üzerine çıkarır. Heparin etkisi ~ 1.5- 4 saat sürer, kan plazmasında bulunan heparinaz ile parçalanır.
ŞEKİL 4.55 Pıhtının yapımı ve yıkılması yolu
Bir kumarin olan warfarin kana verildiğinde, karaciğerde yapılan protrombin, faktör VII, IX ve X düzeyleri düşmeye başlar. Kanın pıhtılaşmasına engel olan (antikogülan etki gösteren) diğer önemli kimyasal maddeler ise EDTA (Etilen Daimin Tetra Asetikasit), Sodyum sitrat, Oksalat, Floruid tuzlar, Hirudin ve Dieumeroldur. Bu maddelerin etki mekanizması genellikle kalsiyumu bağlamalarıdır.

H A Y V A N F İ Z Y O L O J İ S İ
280
Kan Pıhtısının Erimesi (Fibrinoliz) Fibrinoliz fibrini parçalayarak etki gösteren protein eritici (proteolitik) enzimdir. Plazmin (Fibrinolizin) , Plazminojenden dönüşür. Fibrin, Fibrinojen (F I), F V, F VIII, protrombin, F XII yi parçalar. Fibrine bağlı olduğu sürece yıkılmaz. Plazminojen, uyarılınca plazmine dönüşür; plazmin fibrini sindirme özelliği nedeniyle fibrinolizin olarak da adlandırılan bir protein eritici enzimdir. Plazminojeni uyaran plazma etkeninin de uyarılmaya gereksinimi vardır.
ŞEKİL 4.56 Pıhtılnın yok edilmesi
Pıhtılaşmanın gerçekleşmesini sağlayan faktörler olduğu kadar aşırıya kaçmasını engelleyen faktörler de bir denge içinde görev yaparlar. Örneğin endotel hücreleri prostoglandin I2, nitrik oksit ve ADP adlı maddeleri salgılayarak trombositlerin biraraya gelmesini engellerlerken kollagen, fibronektin ve von Willebrand faktörünü salgılayarak pıhtılaşmayı başlatırlar. Ortaya çıkan fibrin tıkacı yine damar endotel hücrelerinden salgılanan doku plazminojen aktivatörü ile çözünmeye başlar ve aynı zamanda heparin ve trombomodülin de salgılanarak trombinin oluşumu engellenir. Bir başka engelleyici madde olan doku plazminojen aktivatör inhibitörü ise fibrinin çözülmesini engeller.
Vitamin K eksikliği
Normal yenidoğanda II, VII, IX ve X numaralı pıhtılaşma faktörleri erişkinlere göre daha düşük miktarlardadır. Yaşamın ilk günlerinde bu faktörlerin miktarı daha da düşerek 3. günde en düşük düzeylerine iner. Bunun nedeni doğumda vücutta depolanan K vitamini miktarının düşük olmasıdır. İnek sütündeki K vitamini anne sütüne göre 4 kat daha fazladır. Depoların düşük olması ve anne sütü ile beslenme eğer yeni doğana K vitamini takviyesi yapılmazsa hemorajik hastalık oluşur. Bu durum yaşamın 2.-4. günleri arasında mide ve barsaktan kanama, göbekten kanama veya iç organlara kanama şeklinde gözlenir. Hastalık K vitaminine cevap verdiği için tüm yenidoğanlara koruma amacıyla K vitamini verilmelidir. Hemofili
Hemofili çoğunlukla genetik geçiş gösteren, vücutta kanın pıhtılaşma sisteminde rol alan ve pıhtılaşma faktörleri olarak adlandırılan proteinlerin eksikliği veya yokluğu nedeniyle ortaya çıkan ve pıhtılaşma bozukluğu yaratan ve X kromozomundaki

H A Y V A N F İ Z Y O L O J İ S İ
281
çekinik bir gen ile taşınan bir tür kanın pıhtılaşamaması hastalığıdır. Faktör VIII eksikliğine (% 85 inde) bağlı kanama eğilimidir, bu tipine Hemofili A veya Klasik Hemofili denir. Vakaların % 15 inde Faktör IX (Hemofili B) eksikliği vardır. Hemofili yalnızca erkeklerde görülür, kadınlar taşıyıcı olabilirler. Tromboembolik Olaylar Trombüs: Kan damarında oluşan anormal pıhtı endotelin bozulması kan akımının yavaşlamasından kaynaklanır. Damar lümenleri ya da kalp boşlukları içerisindeyken katılaşan kan kitlesidir. Fibrin ağı ve şekilli kan elemanlarından oluşur. Emboli: Trombüsün dolaşması ve tıkamasıdır. Başka bir ifade ile Emboli, kanla taşınan yabancı bir cismin damarı tıkaması durumudur. Damardaki bir trombüsten kopan kan pıhtısının başka bir damarda tıkanmaya sebep olmasına ise tromboembolizm denilmektedir. Lökositler-Akyuvarlar Beyaz Kan Hücreleri
Lökositler tüm kanın şekilli elemanlarının % 1 ini oluştururlar. Sayıları; 7000–11000/mm3 tür. Lökositler ve lökositlerden kaynaklanan doku hücreleri vücudun enfeksiyonlara karşı koymasında rol alırlar. Vücudun savunma sisteminin hareketli birimleridir. Kemik iliğinde ve lenfoid dokularda oluşurlar. Dolaşımda 4–8 saat dokularda bulunanlar ise 4-5 gün kadar yaşarlar. Granülositler-büyük, loblu nükleuslu, fagositik hücrelerdir. Kan hücrelerinden olan lökositler, vücudun savunma sisteminin hareketli elemanlarıdır. Çeşitli yollarla vücuda giren mikroorganizmaları, ölü doku artıklarını, yabancı partikülleri ya fagositoz işlemi ile fagosite ederek ya da
ürettikleri antikorlarla ve duyarlı lenfositlerle harap ederek ortadan kaldırmaya çalışırlar.
ŞEKİL 4.57 Granülositler
ŞEKİL 4.60 Agranülositler
Granülositler Nötrofil (lökositlerin % 62 si, fagositoz yetenekleri vardır) Doku aralıklarına diapedez ile girer. Kemotaktik alanlara ameboid hareketle giderler. İnflamasyonlu doku bölgelerine kemotaksi ile çekilirler. Fagositoz işlemi ile mikroorganizmaları ve yabancı maddeleri sindirirler. Hasarlı doku bölgelerinden yer alan iltihaplı dejenere doku ürünleri, toksik maddeler komplement sistem ürünleri nötrofil ve makrofajları bu bölgeye
Granülosit Granülosit, lökositlerin (akyuvarların) bir bölümünü oluşturan çeşitli hücre tiplerine verilen isimdir. Bu ismi almalarının nedeni granülosit hücre tiplerinin sitoplazmalarında bulunan farklı boyama özelliklerine sahip granüllerdir. Bu hücrelere farklı şekillere sahip, çoğunlukla 3 loba ayrılmış biçimdeki, hücre çekirdekleri nedeniyle "polimorfonükleer lökosit" (PMN veya PML) de denir. Yaygın kullanımda polimorfonükleer lökosit tanımı çoğu kez nötrofil granülositini tanımlamak için kullanılır. Nötrofil granülositi granülosit tiplerinin içinde en çok sayıda bulunanıdır
Agranülosit Lenfositler, lenf sisteminde sıkça yer alan ve doğal öldürücü T hücrelerini içeren beyaz kan hücreleridir. Kan, B hücresi, T hücresi ve Doğal öldürücü hücre adı verilen üç tip lenfosit içerir. B hücresi, patojenin ortadan kaldırılması için gereken antikorları üretir. CD4+ (T helper) bağışıklık yanıtını yönetir. (HIV'de zarar görmüş olan bu hücredir) CD8+nanıdır
� Nötrofil � Eozinofil � Bazofil
� Lenfosit � Monosit

H A Y V A N F İ Z Y O L O J İ S İ
282
doğru çeker bu olaya kemotaksi denir. Nötrofilerin Ömürleri ortalama 4 ile 8 gün kadardır.
ŞEKİL 4.61 Yangı olayı
Nötrofiller ve diğer granülositler, kapillerler ve postkapiller venüllerin endotel hücreleri arasındaki bağlantı bölgelerinden geçerek bağ dokusuna ulaşırlar. Bu olaya diapedez adı verilir. Nötrofil granüllerinde elastaz ve proteinazlar bulunur Nötrofil membarıında komplemet sistem proteinlerinde C5a reseptörleri, L-selektin İntersellüler adezyon moleküllerinden ve ICAM- 1 ve ICAM-2 bulunur. İnflamasyonun ilk sonuçlarından birisi, yaralanan bölge ile diğer doku arasında “duvar örmesi"dir. İltihaplı bölgedeki doku aralıkları ve lenfatikler fibrinojen pıhtısı ile bloke olur ve sıvı aralıklarda güçlükle akar. Bu duvar örme olayı bakteri veya toksik ürünlerin yayılımını geciktirir. Madde Auzorofil granüller Fonksiyon
Asit hidrolazlar Alınan maddenin parçalanması Nötral hidrolazlar İltihaplı dokunun harabiyeti Lizozim Bakteriyel hücre duvarının dejenerasyonu Defensinler Oksijen bağımsız madde öldürme Myeloperoksidaz Oksijen bağımlı madde öldürme
ŞEKİL 4.62 Tromposit tıkacı ve pıhtılaşma için egereken faktörler

H A Y V A N F İ Z Y O L O J İ S İ
283
Madde Spesifik granüller Fonksiyon Lizozim Bakteriyel hücre duvarının dejenerasyonu Kobalamin bağlayan protein Bakteriyel kobalamin analoglarına bağlanma Apolaktoferrin Serbest demire bağlanma, Granülopoez kontrolü Kollejenaz Konnektif dokunun sindirimi Eozinofil;(lökositlerin % 2,3 ü, paraziter enfeksiyonlarda rol alırlar, allerjik reaksiyonlarda antijen-antikor kompleksini yok ederler, fagositoz yaparlar). Eozinofiller zayıf fagositlerdir. Paraziter enfeksiyonlu kişilerde çok miktarda üretilirler ve parazitli dokulara göçerler. Pişmemiş domuz etinde bulunan trişinella parazitinin kaslara yayılması ile oluşan trişinozis hastalığı eozinofiliye neden olur. Eozinofiller özellikle allerjik reaksiyonların olduğu dokularda örn; astımlı kişilerde akciğerlerin peribronşiyal dokularında ve allerjik cilt reaksiyonlarda deride toplanma eğilimindedirler. Eozinofilleri bağlıca etkileri (1 ) modifiye lizozomlar olan granüllerinden hidrolitik enzimleri salmak, (2) olasılıkla, oksijenin özellikle öldürücü olan yüksek derecede reaktif formlarını salmak(Radikal oksijen türleri ROS sentezi ve salımı) (3) granülleıinden büyük bazik protein diye adlandırılan yüksek derecede larvasidal polipeptidi salmak Bazofil; (lökositlerin % 0,4 ü, heparin, histamin, seratonin içerirler, allerjik reaksiyonlarda rol alırlar). Bazofiller, vücuttaki kapillerlerin çoğunun dışında yerleşik büyük mast hücrelerine benzer. Her ikisi de yağlı bir yemekten sonra heparin salarlar. Ayrıca, histamin ve az miktarda bradikinin ve serotonin de salarlar. Bradikininin deri ve gastrointestinal sistem kan akımının regulasyonunda rol oynar. Mast hücreleri ve bazofiller allerjik reaksiyonların bazı tiplerinde çok önemli rol oynarlar. Allerjik reaksiyona sebep olan antikor tipi IgE mast hücreleri ve bazofillere özel tutunma eğilimindedir. Bu bağlanma mast hücre ve bazofillerin içinde granüllerin salgılanmasına yol açar. Bu hücrelerde yer alan serotonin(vazodilatör), heparin(pıhtılaşma önleyici) bradikini alerjik reaksiyonların oluşumuna yol açar.
� Agranülositler Monositler; Lökositlerin % 5.3 ünü oluştururlar, dolaşımdaki fagositlerdir ve en büyük kan hücreleridir (15-20 mikrometre). Monositlerin kanda ömürleri 12-100 saat kadardır. Monositler dokulara geçerek makrofaj adını alırlar (dokularda makrofajlar farklı isimle adlandırılır, alveolar makrofajları, osteoklast, mikroglia, kuppfer hücreleri, histiyosit). Monosit makrofajlar yabancı materyalin sindirilmesinde, ölmüş hücrelerin temizlenmesinde, antijen sunumunda rol alırlar. Kemotaksiye neden olan faktörler İmmünoglobulinler sadece B tipi lenfositlerden yapılırken T hücreleri genel immün cevapta çeşitli alternatif roller oynarlar. B ve T hücrelerinin her ikisi de kemik iliğindeki kök hücrelerinden gelişir ve bunların farklılaşması daha sonra içerisine yerleştiği organ tarafından yapılır. B hücreleri kemik iliğinde olgunlaşır. T hücreleri ise timüs içerisinde olgunlaşır. T lenfositlerinin pek çok alt grupları bilinmektedir. T yardımcı hücreleri yüksek seviyede immünoglobulin üretimini harekete geçirebilmesi için B hücrelerini uyarır çoğu durumda Th hücrelerinin yardımı olmaksızın antikor yapımı çok az veya hiç olmaz. Sitotoksik hücreler yabancı hücreleri öldüren toksik maddeler salgılarlar ve Tdth (T-delayed-typed hipersensivite) hücresiyle birlikte hücresel immün cevaplarda primer rol oynar.

H A Y V A N F İ Z Y O L O J İ S İ
284
Perforin proteinleri hedef hücrede sitolitik etki gösteren hücrezarında delik açan bir proteindir. Sitotoksik T lenfositleri (CTL 'ler) ve NK hücreleri tarafından sentezlernir . Perforin hedef hücrenin zarı içine girer ve plazma membranı , bir deliğin oluşmasıına yol açar. Perforin bağlı litik zar deliğinin oluşması için MACPF bölgesine bağlanması gerekir. Bu bölge kolesterol taşıyan gram-pozitif bakteri membran zarlarında belirlenmiştir. Perforin non-spesifik bir hücre yokedicisidir. Sitotoksik T lenfositlerin etki mekanizması: Sitotoksik T lenfositleri antijen sunucu hücrelere bağlanır(makrofajlara). Sitotoksik T lenfositleri tarafında üretilen IL-2 ve antijen sunucu hücrelerce üretilen IL-1 Sitotoksik T lenfositlerin mitozla çoğalmalarına yol açar. Antijen sunucu hücreler makrofajlar içinde patojen örneğin virus bulunması durumda Sitotoksik T lenfositleri perforin salgılar. Salgılana perforinden hücre membranlarını korunmak için protektinde hücre membranlarında salgılanır. Salgılanan perforin hasarlı, enfekte, patojen makrofajların membran zarına yerleşerek osmotik şokla hücreyi öldürür. Başka bir öldürme mekanizması ise Sitotoksik T lenfositleri tarafında salgılana bir ligant olan Fas aracılığı ile enfekte makrofajın apopitozla ortdan kaldırılmasıdır. Doğal öldürücü T lenfositlerin NK hücre öldürme mekanizması:Tümör antijenlerinin varlığında doğal öldürücü T lenfositlerin interlökin-2(IL-2) makrofaj aktive edici faktör MAF ,TNF gibi sitokininler aracılığı aktive olurlar. T lenfositlerin NK hücrenden salgılanan perforin salgılanır. Salgılanan sitokininler makrofajların tümör bölgesine yok açar. Perforine bağlı osmotik hasarla tömür hücreleri yok edilir.
ŞEKİL 4.62 Perforin ile hücre yok edilmesi
Lenfositleri kemotaktik hareketien yol açan başlıca maddeler :
İki lenfosit alt grubu vardır; B hücreleri; kemik iliğinden köken alır ve lenfoid dokularda olgunlaşırlar, humoral bağışıklıktan sorumludurlar. İmmün sistem, bakteri ve diğer enfeksiyon ajanlarına karşı antikorlar geliştirir. Antikorlar immünoglobulin denilen gamma globulinlerdir. Antikorlar bakteri membranlarına tutunur ve bakteri
� Bazı bakteriyel toksinler � İltihaplı dokunun kendisinden açığa çıkan dejeneratif ürünler � İltihaplı dokuda aktive olan “kompleman kompleksinin” çeşitli reaksiyon ürünleri � İltihaplı bölgede plazma pıhtılaşmasının neden olduğu çeşitli reaksiyon ürünleri ve
diğer maddelerdir.

H A Y V A N F İ Z Y O L O J İ S İ
285
fagositoz için elverişli hale getirilir. Bunun için, antikor immün sistemin bir parçası olan kompleman şelalesinin C3 ürünü ile birleşir. C3 molekülü fagosit membranındaki reseptörlere tutunur, böylece fagositoz başlar. Tüm bu sürece opsonizasyon denir.
. ŞEKİL 4.63 Bağışıklık
– T hücreleri; timus bezinden köken alırlar ve hücresel bağışıklıktan sorumludurlar. Lökositoz-lökopeni
Lökosit sayısının 1 mm3 kanda 11000 üzerine çıkmasına lökositoz denir. Lökosit sayısının 1 mm3 kanda 7000 altına düşmesine lökopeni denir. Başka bir ifade ile kanda bulunan lökosit sayısının azalması Lökopeni durumudur. Lökositlerin ana görevi enfeksiyonlarla savaşmak olduğu için, lökosit sayısındaki düşüş enfeksiyon oluşumu riskini yükseltmektedir. Lökosit tiplerinin en yüksek sayıda bulunanı nötrofildir. Nötrofil sayısındaki azalmaya ise nötropeni denir. Nötropeni kavramı bazen lökopeni yerine kullanılır, bunun nedeni nötrofil sayımının enfeksiyon riskinin en önemli göstergesi oluşudur. Yine de, nötropeniyi lökopeninin bir altkümesi olarak değerlendirmek en doğrusudur. Kemoterapi, lösemi, miyelofibroz ve aplastik anemi düşük lökosit sayımlarına yol açabilir. Ayrıca, bazı yaygın ilaçlar da lökopeniye neden olabilir. Lökopeni tam kan sayımı yardımıyla tanımlanabilir.

H A Y V A N F İ Z Y O L O J İ S İ
286
ŞEKİL 4.65 Lökositoz ve lökopeni durumu
Lösemi
Bir kanser türü olan lösemi kontrolsüz akyuvar üretimidir. Kemik iliği veya lenfoid dokudaki hücrelerin kanseröz mutasyonu(pro-onkogenlerin onkogenler düşerek) nedeniyle oluşur. Kan dolaşımındaki anormal lökositlerin sayısı çok artar. Myeloid lösemide, kanseröz süreç arasıra, kısmi farklılaşmış hücreler üretir bunlar monositik lösemi olarak adlandırılırlar. Ancak daha sıklıkla lösemi hücreleri şekilsiz, değişikliğe uğramamış ve normal lokositlerin hiçbirine benzemeyen türdendir. İki tipi vardır; – Lenfoid lösemi; lenf düğümleri – Miyeloid lösemi; kemik iliği Löseminin ilk etkisi, lösemik hücrelerin metastatik gelişimidir. Kemik iliğindeki lösemik hücreler hızla çoğalabilir ve çevredeki kemiğe yayılarak, ağrıya ve kemik kırıklarına neden olurlar. Lösemik hücreler; dalak, lenf düğümleri, Karaciğer ve özellikle damar bölgelerine yayılarak buradaki metabolik elementleri kullanır ve sonuçta doku harabiyetine neden olur. Löseminin genel etkileri; enfeksiyonların gelişmesi ağır anemi ve trombositopeniye bağlı kanama eğilimidir. Vücuttaki en önemli etkisi; büyüyen kanserli hücrelerin metabolik maddeleri aşırı kullanarak açlığa ve ölüme neden olmasıdır. Vücudun Enfeksiyonlara Direnci
Bağışıklık sistemi, bir canlıdaki hastalıklara karşı koruma yapan, patojenleri ve tümör hücrelerini tanıyıp onları yok eden işleyişlerin toplamıdır. Sistem, canlı vücudunda geniş bir çeşitlilikte, virüslerden parazitik solucanlara, vücuda giren veya vücutla temasta bulunan her yabancı maddeye kadar tarama yapar ve onları, canlının sağlıklı vücut hücrelerinden ve dokularından ayırt eder. Bağışıklık sistemi, çok benzer özellikteki maddeleri bile birbirinden ayırabilir, örneğin; bir amino asidi farklı olan proteinleri bile birbirinden ayırabilecek özelliğe sahiptir. Bu ayrım, patojenlerin konak canlıdaki savunma sistemine rağmen enfeksiyon yapmaları için yeni yollar bulmalarına, bazı uyumlar sağlamalarına neden olacak kadar karmaşıktır. Bu mücadelede hayatta kalmak için patojenleri tanıyan ve onları etkisizleştiren bazı mekanizmalar gelişmiştir. Lökositler vücudun savunma sisteminin hareketli birimleridir. Yayılmacı

H A Y V A N F İ Z Y O L O J İ S İ
287
mikroorganizmaları fagositoz yolu ile harap ederek, Antikorlar ve duyarlı lenfositler oluşturarak yayılmacıyı harap ederler veya inaktive ederler.
Doğuştan gelen bağışıklık Bu tür bağışıklık aynı zamanda genetik bağışıklık olarak ta bilinir. Genetik olarak tanımlanmış özelliklerden meydana gelir. Bu tip bağışıklığın bir tipi tür bağışıklığı olup bir türün bütün üyelerinde mevcuttur. Örneğin; insanlar süs hayvanları, ev hayvanları ve bazı insana benzer bağışıklığa sahip olan hayvanlarda hastalıklara yol açan enfeksiyon yapan ajanlara karşı bağışıklığa sahiptirler. Doğal bağışıklıkta rol oynayan fagositer hücreler, memeli hücrelerinde bulunmayan belli moleküler kalıplara bağlanarak mikroorganizmaları tanırlar. Bu kalıplar mikroorganizmaların; nükleik asit, protein, lipid, karbonhidratlarında bulunabilir. Örneğin; çift-zincirli RNA, lipopolisakkaridler veya memeli dokularında bulunmayan ancak bazı bakteriler için tipik olan mannozdan zengin oligosakkaritler fagositer hücrelerin tanıdığı moleküllerdir. Doğal bağışıklık için gerekli olan reseptörler genom tarafından doğrudan kodlanır. Oysa antikorlar ve lenfosit reseptörlerini kodlayan genler sürekli yeniden-düzenlenmeye tabidirler. Doğal bağışıklıkta kullanılan bu reseptörler, yeniden düzenlenmeye uğramadıklarından özgüllük açısından sınırlı bir repartuara sahiptir. Ancak tanıdıkları bu kalıplar mutasyona uğramadığından verdikleri yanıt oldukça etkilidi. 1. Bakteri ve diğer saldırganların kan akyuvarları ve doku makrofaj sistemi hücreleri tarafından fagosite edilmesi. 2. Ağız yoluyla alınan organizmaların mide asit salgısı ve sindirim enzimleri ile haraplanması. 3. Derinin organizmaların istilasına karşı direnci. 4. Kanda yabancı organizma ve toksinlere bağlanıp, zarar veren kimyasal aracıların bulunması. Kazanılan bağışıklık
Doğuştan gelen bağışıklığın asine kazanılan bağışıklık kalıtsallık dışında herhangi bir şekilde temin edilmektedir. Bu ya doğal olarak veya yapay olarak kazanılır. Doğal olarak kazanılan bağışıklık genellikle bir özel hastalığı geçirmiş olmasıyla sağlanır. Hastalık sırasında bağışıklık sistemini; içeriye giren ajan üzerindeki antijen(Ag) olarak adlandırılan moleküllere karşı uyarılır; antikor (Ab) olarak isimlendirilen molekülleri üretir ve diğer aynı ajan tarafından oluşturulabilecek enfeksiyonlara karşı özel savunmaları başlatır. Bağışıklık aynı zamanda tabi olarak doğum sonrası salgılanan ilk sıvı ve süt emzirmeyle
Savunma mekanizmaları
Özgül olmayan savunma
mekanizmaları
Özgül savunma mekanizmaları
Lenfoistler Antikorlar Savunmanın Birinci hattı
1- Deri 2- Mükus salgılar
Savunmanın ikinci hattı 1- Fagositik lenfositler 2- Antimikrobiyal maddeler(komplenet sistem 3- Yangı tepkimeleri

H A Y V A N F İ Z Y O L O J İ S İ
288
yeni doğana veya plasenta ile fetus’e transfer edilen antikorla(IgG, IgA) sağlanır. Bunun aksine yapay olarak kazanılmış bağışıklık bağışıklık üretebilen serum veya aşının enjeksiyonuyla bir antijenin içeriye alınmasıyla temin edilebilir.
ŞEKİL 4.66 Hızlı ve yavaş bağışıklı yanıtlarının oluşturan hücreler
Aktif ve pasif bağışıklık
Bağışıklık ister kazanılmış olsun ister doğal olsun aktif veya pasif olabilir. Aktif bağışıklık; bir şahsın kendi bağışıklık sisteminin antikor veya diğer savunmaları bir enjeksiyon ajana karşı ürettiği zaman meydana gelir. Bu ömür boyu olurken antikorun kararlılığına bağlı olarak haftalar, aylar veya yıllar sürebilir. Doğal olarak kazanılan aktif bağışıklık birey enfeksiyon yapan ajana maruz kaldığı zaman üretilir. Yapay olarak kazanılan aktif bağışıklık şahsın canlı zayıflatılmış veya ölü organizmalar ve diğer toksinler ihtiva eden aşılara maruz kalmasıyla üretilir. Her iki aktif bağışıklık ta konağın kendi bağışıklık sistemi vücudu bir antijene karşı savunmak için özel olarak uyarır. Böylece bağışıklık sistemi genel olarak daha önce uyarılmış olduğu antijeni hatırlar ve aynı antijenle tekrar karşılaştığında onu engeller.
ŞEKİL 4.67 Bağışıklı tepkimeleri

H A Y V A N F İ Z Y O L O J İ S İ
289
Pasif bağışıklık; hazır yapılmış antikor vücut içerisine konularak sağlanır. Bu bağışıklık konağın bağışıklık sisteminin antikoru yapmamış olmasından dolayı pasif olarak adlandırılır. Doğal olarak kazanılmış pasif bağışıklık annenin bağışıklık sistemi tarafından yapılan antikor yavruya transfer edildiği zaman üretilir. Yeni anneler yavrularını emzirmeyi planlamasalar dahi antikorların(IgG) ilk süt ile yavruya geçebilmesi için birkaç gün emzirmelidirler. Yapay olarak kazanılmış pasif bağışıklık diğer bir konak tarafından yapılmış antikor yeni konak içerisine transfer edildiği zaman oluşur. Bu tip bağışıklıkta konağın bağışıklık sistemi etki için uyarılmaz. Hazır yapılmış antikorlar ve bağışıklık birkaç haftadan birkaç aya kadar etkisini sürdürür ve neticede konak tarafından tahrip edilir. Konağın bağışıklık sistemi yenisini yapamaz. Bağışıklık uyarısının oluşmasına sebep olan yabancı maddeler antijen olarak adlandırılır. Antijen uyarısı neticesinde bağışıklık sisteminin ürettiği özel proteinler antikorlar veya immünoglobulinler; özel hücreler ise aktiflenmiş T hücreleri olarak adlandırılır. Antikorları esas alan bağışıklık humoral bağışıklık (antikora bağımlı bağışıklık) ve aktiflenmiş T hücrelerini esas alan bağışıklığa da hücresel bağışıklık denir. Antijen ve antikorlar
Bağışıklık sisteminin başlaması antijenlerden tarafından harekete geçirilir. Antijen vücudun yabancı olarak algıladığı bir madde olup ve buna kaşı bağışıklık sistemi hareketi başlar. Antijenlerin çoğu karmaşık yapılı ve 10.000 den büyük moleküller ağırlığa sahip(8000 dalton altındaki antijenin maddelere hapten nedir.) büyük protein molekülüdür. Bazı antijenler polisakkarit ve çok azı glikoprotein (karbonhidrat ve protein) veya nükleoprotein (nükleik asit ve protein) dir. Proteinler polisakkaritlerden daha karmaşık yapıya sahip olduklarında genellikle daha büyük antijenik özelliğe sahiptirler. Büyük karmaşık proteinler antijenik özellik taşıyan ve antikorun bağlanmasını sağlayan pek çok antijenik determinant bölgeye sahiptirler. Antijen virüslerin bakteri ve diğer organizmaların bütün hücrelerinin yüzeyinde bulunurlar. Her hücrenin antijeninin gerçek kimyasal yapısı onun DNA sındaki genetik bilgi ile tanımlanır. Bakteriler kapsül, hücre duvarı ve hatta flagella üzerinde antijenlere sahip olabilirler. Pek çok organizma yüzeylerinde farklı antijenik determinantlara sahiptirler. Farklı antijenik determinantlara karşı tepkilerin olduğunu belirlemek etkili aşı yapmada önemlidir. Örneğin; kırmızı kan hücrelerinin yüzeyindeki antijenleri kan tipini belirler, diğer hücreler üzerindeki antijenler bir şahıstan alınan doku transplantasyonunun red edilip edilmeyeceğini belirler. Eğer bir antijen epitelyum yüzeyden içeriye geçerse bu hemen sonra bir fagositik hücre ile temasa girer. Makrofaj gibi hücreler içindeki sitoplazmik vesiküllerden litik maddeleri (proteaz, nükleaz, lipaz ve lizozim) salgılayarak antijeni öldürürler. Bakteriyel hücrenin parçalanması makrofaj içerisinde bakteriyel antijenlerin salınmasına yol açar. Bunlar içeriye direkt olarak alınan antijenle birlikte makrofaj tarafından işleme tabi tutulur ve antikor sentezinin erken basamaklarında kullanılırlar. Makrofajlar yabancı antijeni spesifik T ve B hücreleriyle temas edebilecek yüzeylerine yerleştirdiklerinden dolayı antijen sunan hücreler olarak iş görürler. Antijenin T ve B lenfosit hücreleri tarafından tanınması antikor üretiminin ilk basamağıdır.
İmmünglobulinler (= antikorlar), antijenin organizmaya girmesi ve immün sistemi uyarması sonucunda sentezlenen glikoprotein yapısında maddelerdir. Antikorlar globulin yapısında proteinlerdir ve immünolojik rolleri nedeniyle İmmünglobulin (= Ig) adı verilmiştir. Ig'ler kendisinin oluşumuna neden olan antijene özgüldür ve onunla özgül olarak (sadece o antijenle, başkalarıyla değil) birleşebilme ve reaksiyonlara yol açabilme özelliğindedir. Ig'ler bazı farklı özellikleri nedeniyle IgG, IgM, IgA, IgD ve IgE olmak üzere beş çeşide ayrılarak incelenirler. Ig iki kısa (L zinciri), iki uzun (H zinciri) olmak üzere dört polipeptid

H A Y V A N F İ Z Y O L O J İ S İ
290
zincirinden oluşur ve Y harfi şeklinde duruş gösterir. Y'nin kolları antijeni bağlar (= Fab kısmı), gövde kısmı ise çeşitli biyolojik aktivitelerde rol oynar (= Fc kısmı). Bazı Ig'lerde Y harfi şeklindeki birim yapıdan (=monomer) birkaç tane bulunabilir. IgG insanda en çok bulunan (Tüm Ig'lerin %75'i) temel Ig'dir. Plasentadan bebeğe geçebilen tek Ig'dir. Böylece bebekler anne antikorları sayesinde korunmuş olurlar. IgG'ler uzun ömürlüdür, infeksiyonlarda ve aşılamalarda yüksek miktarlarda oluşarak bağışıklıkta önemli rol oynarlar. IgM, pentamer yapıda, büyük Ig'dır. Antijen uyarımında ilk sentezlenir, kısa ömürlüdür. IgM intrauterin dönemde de sentezlenebilir. Yenidoğanda IgM varlığı ile konjenital infeksiyonlar araştırılabilir. M1 ve M2 olmak üzere 2 alt tipi vardır. Molekül ağırlığı en fazla olandır. Bu nedenle intravasküler yerleşimlidir. Bir antijene varilen birincil yanıtta üretilir. Komplemanı fikse eder (alternatif yol). Plasentayı geçmez. B hücreleri yüzeyinde antijen reseptörü vardır. Makrofaj ve nötrofillere bağlanmaz. En güçlü antibakteriyal etkiye sahiptir. Yarı ömrü 5-8 gündür. IgA hem serumda, hem de vücut salgılarında bulunabilen Ig'dir (Solunum, sindirim, ürogenital sistem salgıları, ter, tükrük, gözyaşı, süt gibi). Salgısal özelliği nedeniyle infeksiyonlardan korunmada önemlidir. IgD henüz tam incelenememiş bir Ig'dir. IgE allerjik reaksiyonlarda ve parazit infeksiyonlarında rol oynar. Bir antijenik uyarıma karşı özgül antikor sentezinin nasıl gerçekleştiği konusunda birçok görüş ileri sürülmüştür. Bunlardan en çok destek göreni doğal klon seçimi ve genetik temele dayanan görüşlerdir.
ŞEKİL 4.68 Antikor tipleri Kompelement sistemin etkisi
Bazı durumlarda hapten olarak adlandırılan küçük bir molekül(8000 daltondan küçük) eğer büyük bir protein molekülüne bağlanırsa antijen olarak rol oynar. Hapten proteinlerin yüzeylerinde antijenik determinant olarak iş görür. Bazen bunlar vücut proteinlere bağlanır ve immün uyarıyı harekete geçirir. Hapten veya vücut proteini yalnız başına antijen olarak iş göremezler fakat bunların kombinezyonu antijen olarak iş görür. Örneğin; penisilin molekülü hapten olarak iş görür. Protein molekülüne bağlanır ve alerjik reaksiyonu harekete geçirir. Bir yabancı maddeye karşı immün sisteminin en önemli uyarılarından biri antiantijen proteinlerinin ve antikorların üretimidir. Antikor bir protein olup antijene cevap olarak üretilir ve antijene seçici olarak bağlanma özelliğine sahiptir. Her antikor türü özel bir antijenik determinanta bağlanır. Bu tip bağlanma antijenin inaktivasyonuna katkıda bulunur veya bulunmaz. Antijen ve antikorun konsantrasyonu tartışıldığı zaman immünologlar sıklıla titerlerden söz ederler. Titer; verilen bir reaksiyonu üretmek için ihtiyaç duyulan bir maddenin miktarıdır. Örneğin bir antikor titeri özel miktarda antijene bağlanan ve onu nötralize etmek için gerekli antikor miktarıdır. Bağışıklık sistemi immün cevabı oluşturmak için değişik yollarda interaksiyon gösteren bir seri hücre ve organın faaliyetleri içermektedir.

H A Y V A N F İ Z Y O L O J İ S İ
291
Bağışıklık sisteminin organları vücut boyunca yerleşmişlerdir. İmmün cevapta rol oynayan anahtar hücre tipi bir beyaz kan hücresi çeşidi olan lenfosittir ve immün cevaba katkıda bulunan doku ise lenfoid doku olarak adlandırılır. Lenfositler kemik iliğindeki farklılaşmamış kök hücrelerinden gelişir.
Komplement sistem
Komplemant sistemin ana işlevi patojenleri opsonizasyonla fagositoz yapan hücreler makrofajlar ve nötrofiller tarafından ortadan kaldırılmasıdır. Komplemant, çoğunluğu enzim öncülü olan C1-C9 kadar isimlendirilmiş 11 adet plazma proteinleridir. Normalde bu proteinlerin tümü plazmada da bulunurlar.
�
“Komplement”, çoğunluğu enzim prekürsörü olan toplam 30 kadar proteini kapsayan genel bir tanımdır. Bu sistemde temel role sahip olanlar C1-C9, B ve D isimleri verilen toplam 11 proteindir. Normalde bu proteinlerin tümü plazma proteinleridir ve kapillerden dokuya sızan plazmada da bulunurlar.
ŞEKİL 4.69 Kompelement sistemin etkisi
Komplemant sistem fagositoz yapan makrofaj ve nötrofillerin patojenleri tanıması ve algılaması için gereklidir. Hücre dışı bakterilerle savaşan komplement sitem kuvveli bir savunma mekanizmasıdır. Komplent proteinleri prekürsörleri normalde inaktiftirler ve daha çok klasik yolla aktiflenebilirler: Kompleman sistemi birçoğu proteinaz olan 30’dan fazla proteinin bir kompleks serisi oluşturmasından meydana gelir. Enfeksiyona yanıtta aktive olan ilk sistemlerden birisidir. Kompleman proteinleri plazmada inaktif olarak dolaşır. Enfeksiyonlar bir seri proteolitik aktivasyon reaksiyonu ile kompleman sistemini aktive eder. Bu eşleşmiş proteolitik reaksiyonlar şiddeti gittikçe artan etkileri ile distal kompleman proteinlerinin aktivasyonuna sebep olurlar.
� Lizis � Aglütinasyon � Virüslerin nötralizasyonu � Kemotaksi � Mast hücresi ve bazofil aktivasyonu � İnflamatuvar etkiler

H A Y V A N F İ Z Y O L O J İ S İ
292
ŞEKİL 4.70 Komplement sistemindeki proteinlerin etkisi
Klasik yol antijen-antikor reaksiyonu ile uyarılır. Antijen antikor ile bağlanınca antikor yapısındaki “sabit” bölgede bulunan özgül bir reaktif bölgesi açığa çıkar ya da aktiflenir. Ardından, bu bölge kompleman sisteminin C1 proteinini bağlayarak C1 proenziminin aktivasyonu ile başlayan ardışık reaksiyonlar zincirini tetikler. Kompleman sistemi aktivasyonunun bu aşamasında birkaç antijen-antikor kombinasyonu yeterlidir. Aktiflenen C1 enzimi zincirin izleyen aşamaların da artan miktarlarda enzimleri aktifler ve küçük ölçekte başlayan aktivasyonla başlayıp "artmış” reaksiyon gelişir. Birçok son ürün oluşur ve bunların çoğu istilacı organizmanın ya da toksinin zararlı etkilerini engelleyici etkiler gösterirler.
1. Opsonizasyon ve fagositoz. Kompleman reaksiyon zincirinin bir ürünü olan C3b, nötrofil ve makrofajların fagositozunu uyarır ve onların antijen-antikor kompleksini bağlamış olan bakteriyi içlerine almalarını sağlar. Bu işlem opsonizasyon adını alır. Bu yokedilen bakteri sayısını yüzlerce kat artırabilir.
2. Lizis. Kompleman reaksiyon zincirinin en önemli ürünlerinden biri litik kompleks adını alan, kompleman faktörlerinden birçoğunun oluşturduğu bu kompleksin bakteri ya da diğer istilacı organizmaların membranlarmı direkt yırtma etkisi vardır.
3. Aglütinasyon. Kompleman ürünlerinin istilacı organizmanın yüzeyini değiştirerek birbirlerine yapışmalarını ve aglütinasyonunu kolaylaştırıcı etkileri vardır.
4. Viruslann nötralizasyonu. Kompleman enzimleri ve diğer kompleman ürünlerinin bazı virusların yapılarına saldırarak onları avirulan hale getirme özellikleri vardır.
5. Kemotaksi. C5a fragmanı nötrofil ve makrofajların kemotaksisini ve böylece çok sayıda fagositin antijenik ajanın bulunduğu bölgeye göçmesini sağlar.
6. Mast hücresi ve bazofil aktivasyonu. C3a, C4a ve C5a mast hücresi ve bazofılleri aktifleyerek bunların lokal sıvılara histamin, heparin ve diğer maddeleri salgılamalarını sağlarlar. Bu maddeler de yanıt olarak bölgesel kan akımını, dokulara sıvı ve plazma proteinlerinin sızmasını artırı ve antijenik ajanın hareketsizleştirilmesini ve inaktivasyonunu sağlarlar. Bu alerjik reaksiyonlra yo açabilir.

H A Y V A N F İ Z Y O L O J İ S İ
293
7. İnflamatuvar etkiler. Mast hücreleri ve bazofilleri uyararak oluşturdukları inflamatuvar etkilerinin yanısıra, diğer birçok kompleman ürününün de lokal inflamatuvar etkisi vardır. Bunlar, artmış olan kan akımını daha da artırarak kapillerlerden dokuya protein sızmasını ve proteinlerin doku aralıklarında pıhtılaşarak saldırgan organizmaların dokular arasından geçerek yayılmasını engellerler.
Toll-benzeri reseptörler (Toll-like receptors-TLR)
Çeşitli mikrobiyal ürünlere bağlanarak doğal bağışık yanıtı uyaran membran proteinleridir. Memelilerde 10 farklı TLR tanımlanmıştır. Bu reseptörler TLR makrofaj, dentrik hücreler, nötrofiller, NK hücreler, epitelyal ve endotelyal hücreler gibi doğal bağışık yanıtta rol alan çok sayıda hücrede vardır. TLR mikrobiyal yapılarla ilişkiye girdiğinde trankripsiyon faktörlerini aktifleşmesine neden olur. TLR uyarısına yanıt olarak açılan genler Bağışık yanıtta önemli rol oynayan çeşitli proteinleri kodlarlar. Gen ürünleri yanıt veren hücreye göre değişmekle birlikte bu proteinler enflamatuar sitokinler (TNF, IL-1, IL-2), endotelyal adezyon kuvvet molekülleri (E-selektin) ve mikroorganizmaların öldürülmesinde rol alan proteinler (uyarılabilir nitrik oksit sentaz) olabilir.
ŞEKİL 4.71 Bakteri yüzeyindeki yapıların tanımlanması(antikor antijen)
Hapten
Molekül ağırlığı 8.000’in altında olan maddeler nadiren antijen özelliği kazansalar da, düşük molekül ağırlıklı maddelere karşı bağışıklık gelişimi yine de özel bir yolla sağlanabilir: Hapten adını alan küçük molekül ağırlıklı bileşikler önce antijenik özellikli bir madde; örneğin bir protein ile birleşirlerse, bu bileşim bir immün yanıt oluşumu sağlar. Bu bileşime karşı gelişen antikorlar ya da aktif lenfositler çoğunlukla bileşimi oluşturan hapten ve proteine karşı ayrı ayrı da yanıt verirler. Böylece haptenle ikinci karşılaşmada antikor ve lenfositlerin bazıları organizmada yayılma ve hasar gerçekleşmeden hemen haptene yanıt verirler. Bu tür immün yanıt oluşturan haptenler genellikle düşük molekül ağırlıklı ilaçlar, tozda bulunan kimyasal maddeler, hayvan derisi artıkları (kepek), dökülen deri artıkları, endüstriyel kimyasal maddeler, zehirli sarmaşık toksini vb. olabilir.

H A Y V A N F İ Z Y O L O J İ S İ
294
ŞEKİL 4.72 B lenfositleri aktive eden hapten ve Th ile aktivasyon.
Antikor aracılığıyla aşırı duyarlılık reaksiyonunda, vücudun kendi hücreleri üzerinde yer alan antijenlere karşı gelişen bağışıklık yanıt sonrası antikorlar gelişmiştir. Hücreler üzerinde yer alan bu antikorlar içsel, diğer bir deyişle vucuda ait veya dışsal olabilir, dışsal antijenler virüs parçacıkları gibi hücre zarlarına bir infeksiyon sonrası yapışmış etkenler olabilir. Bu antijenlere karşı gelişmiş IgG ve IgM sınıfı antikorların söz konusu antijenlerle birleşmesi sonrası, klasik yol üzerinden kompleman sistemi aktivasyonu gerçekleşir. Kompleman aktivasyonu normalde, patojen maddeleri bağışıklık sistemine sunan hücrelerin ve dolayısıyla patojen taşıyan hücrelerin yok edilmesi amacını taşır. Reaksiyon bölgesinde, akut yangı etken maddeleri salgılanır ve hücre zarı eritici bileşenlerin varlığı ile hücreler parçalanır ve hücre ölümü gerçekleşir. Bu reaksiyonun oluşması ve etkinliği, saatler veya günler boyunca sürebilir. Bazı örnekleri, otoimmün hemolitik anemi, Goodpasture sendromu, pemfigus, pernisiyöz anemi, immün trombositopeni ve kan transfüzyonu reaksiyonları olarak sıralanabilir.
Aşırı duyarlılık reaksiyonları
Aşırı duyarlılık, normal bağışıklık sistemi tarafından üretilen aşırı, istenmeyen (zararlı, rahatsızlık üreten ve bazen ölümcül) reaksiyonlar anlamına gelir. Aşırı duyarlılık reaksiyonları konağın bir ön-duyarlı (bağışıklık) durumu gerektirir. Reaksiyon için alınan ilgili mekanizmaları ve süresine göre reaksiyonları dört tipe ayrılabilir, tip I, tip II, tip III ve tip IV.
Tip I Hipersensitivite
Tip I hipersensitivite aynı zamanda anafilaktik aşırı duyarlılık olarak da bilinir. Farklı doku ve organlardaki reaksiyonlar: deri (ürtiker ve egzama), gözler (konjonktivit), nazofarenks (burun akıntısı, burun iltihabı), bronkopulmoner dokular (astım) ve gastrointestinal sistem (gastroenterit) içerebilir. Erken tipte tip I reaksiyonlar IgE aracılık eder. Bu aşırı duyarlılık birincil hücreselleri mast hücresi ya da bazofildir. Tip I reaksiyonları, trombositler, nötrofiller ve eozinofiller tarafından yükseltilir ve / veya değiştirilir. Esas olarak mast hücreleri ve nötrofiller görev yapar. Reaksiyonunun mekanizması bazı antijenler (allerjen) karşılık olarak, tercih edilen bir IgE üretimini içerir.

H A Y V A N F İ Z Y O L O J İ S İ
295
ŞEKİL 4.73 Tip I duyarlılık reaksiyonları.
IgE mast hücreleri ve bazofiller üzerindeki reseptörüne çok yüksek bir afiniteye sahiptir. IgE Fc reseptörünün çapraz bağlanması, mast hücresi tetikleyici önemlidir. Mast hücresi degranülasyonunun çok önemli bir süreçtir. Mast hücrelerinden ve bunların etkileri salınan ajanlar Tip 1 önemlidir. Mast hücreleri, örneğin egzersiz, duygusal stres, kimyasal maddeler (örneğin, kalsiyum iyonoforlar, kodein, vb), anaflatoksik tetiklenebilir. (C4a, C3a, C5a, vb.) Aynı semptomlara neden olmasına rağmen, IgE alerjen etkileşimi olmağı bu reaksiyonlar, aşırı duyarlılık reaksiyonları değildir.
Tablo 1. Farmakolojik Aracıları
Ani hipersensitivite Aracı Önceden oluşturulmuş granüller aracılar
Histamin bronkokonstiriksiyon, mukus salgısı, vazodilatasyon, vasküler geçirgenlik
Triptazın proteoliz
Kininogenase kininler ve vazodilatasyon, vasküler geçirgenlik, ödem
ECF-A (Tetrapeptidler)
eozinofil ve nötrofilleri çekmezler
Yeni oluşan aracılar
Lökotrien B bazofil Cezbedici
Lökotrien C4 , D4 daha güçlü histamin gibi aynı ama 1000x
Prostaglandinler D 2 ödem ve ağrı
Paf trombosit agregasyonu ve heparin sürüm: mikrotrombi
Hipersensitivite Reaksiyonları, trombosit agregasyonu ve histamin salınmasını, heparin ve vazoaktif aminler neden olan PAF (trombosit aktivasyon faktörü) ortaya çıkar. Anafilaksi (ECF-A) ve nötrofil kemotaktik faktörler, eozinofil kemotaktik faktörü gibi çeşitli nekrozuna hidrolitik enzimler, sırasıyla serbest eozinofiller ve nötrofiller, çeker. Eozinofillerin bu rolü Eozinofiller, arylsulphatase, histaminase, fosfolipaz-Ge ve prostaglandin-E bırakarak lokal reaksiyon kontrol edebilir. Siklik nükleotid (cAMP ve cGMP seviyelerini) ani aşırı artışı duyarlılık reaksiyonu modülasyonunda önemli bir rol oynadığı görülmektedir. Bu maddelerin değişimi önemli ölçüde alerjik semptomlar

H A Y V A N F İ Z Y O L O J İ S İ
296
değiştirebilir. Hücre içi cAMP artırmak için kullanılan maddeler alerjik semptomları, özellikle bronşların genişlemesine ve rahat soluk alıp vermeye neden olur. Tersine, cAMP ya da cGMP azaltan ajanlar, bu alerjik durumları kötüleştirebilir.
Tablo 2 - alerjik semptomlar ve halkalı-nükleotidler arasındaki ilişki
cAMP düşürülmesi cAMP yükseklik
α-adrenerjik reseptör uyarılması (veya epinefrin-, fenil-epinefrin)
veya
β-adrenerjik reseptör bloke edici (propanolol)
β-adrenerjik reseptör stimülasyonu (epinefrin, isoproterenol)
α-adrenerjik reseptör bloke edici (fenoksibenzamin)
fosfodiesteraz inhibisyonu (teofilin)
siklik GMP-yükseklik reseptörlerine, histamin-2 ya da PGE bağlanmasının
γ-kolinerjik reseptör uyarılması (asetil kolin, carbacol)
Semptomların kötüleşmesine Belirtiler büyümesi
Acil aşırı duyarlılık için tanı testleri şüpheli alerjenlere karşı deri testleri (prick ve intradermal) yapılaması ve total IgE ölçümü ve spesifik IgE antikorları belirlenmesi gerekir. Toplam IgE ve spesifik IgE antikorları enzim immunoassay (ELİSA) bir modifikasyonu ile ölçülür. Semptomatik tedavi histamin reseptörlerini bloke antihistaminikler ile yapılır. Örneğin Kromolin sodyum, Ca+2 akışını engelleyerek, muhtemelen, mast hücre degranülasyonunu engellemektedir. Mast hücrelerine bağlanan IgE Fc kısımlarına karşı IgG antikorların kullanımı bazı maddeler alerji tedavisinde kullanılır. Bu maddeler Fc kısımları bloklayarak mast hücresi duyarlılığını azaltır. Hyposensitization (immunoterapi veya duyarsızlaştırma), polenler özellikle böcek zehirlerinin için, bir dereceye kadar alerji reaksiyonları tedaviside kullanılan bir yöntemidir. Hyposensitization mekanizması tam olarak belirlemememiştir, ancak belirtilerin azaltılmasında IgG engellemesi arasında bir ilişki vardır. Tip II Aşiri Duyarlilik
Tip II aşırı duyarlılık reaksiyonları da sitotoksik aşırı duyarlılık olarak da bilinir ve çeşitli organları ve dokuları etkileyebilir. Hücre zarlarına bağlanan çeşitli egzojenez kimyasal maddeler (haptenler), tip II aşırı-duyarlılığa yol açabildiği halde, antijenler, normal olarak endojen kökenlidir. Tip II aşırı duyarlılık reaksiyonları İlaca bağlı hemolitik anemi, granülositopeni ve trombositopeni gibi durumlarda gözelnir. Reaksiyon süresi saatler yada dakikalarla sınırlıdır. Tip II aşırı duyarlılık birincil olarak IgM veya IgG sınıfları ve tamamlayıcı antikorlar aracılığıyla meydana gelir.
Tip III Aşiri Duyarlilik
Tip III aşırı duyarlılık, aynı zamanda immün kompleks aşırı duyarlılık olarak da bilinir. Bu tip reaksiyonlar, akciğer (örneğin, aspergilloz), kan damarları (örneğin, poliarterit, genel (örneğin, serum hastalığı) ortaya çıkar. Tip III duyarlılıkta tek bir organ örneğin deri yada çok sayıda organ (örneğin, sistemik lupus eritematoz, Arthus reaksiyonunda),

H A Y V A N F İ Z Y O L O J İ S İ
297
böbrek (örneğin, lupus nefriti) kapsayabilir ), eklem (örn, romatoid artrit) etkilebilir. Tip III duyarlılıkta çözünür immün kompleksleriyle görev alır. IgM'de bulabilir. Ancak çoğunlukla IgG sınıfı etkilerim oluşumdan sorumludur. Antijen eksojen (kronik bakteriyel, viral veya parazitik enfeksiyonlar), ya da endojen (organa özgü olmayan oto-bağışıklık: örneğin, sistemik lupus eritematoz SLE) olabilir. Antijen çözünür ve ilgili organa bağlı değildir.
ŞEKİL 4.74 Tip III duyarlılık reaksiyonları .
Primer bileşenleri çözünür bağışıklık kompleksleri ve komplement sistem elemanları (C3a, 4a ve 5a) proteinleridir. Trombosit ve nötrofillerin hasarına neden olur. Tip III aşırı duyarlılık immünofloresan boyama belirlenebilir. Komplemant sistemdeki özel proteinlerin varlığı ya da yokluğu ve immün komplekslerin varlığı bu aşırı duyarlılık reaksiyonları için belirleyicidir. Tedavisinde anti-enflamatuar ajanları kullanılır.
Tip IV Aşiri Duyarlilik
Tip IV aynı zamanda hücre aracılı aşırı duyarlılık veya gecikmeli tip aşırı duyarlılık olarak da bilinir. Bu aşırı duyarlılık klasik örneği 48 saat antijen (PPD ya da eski tüberkülin) enjeksiyonundan sonra pik tüberkülin (Montoux) reaksiyonudur.

H A Y V A N F İ Z Y O L O J İ S İ
298
Tablo 3 - Geciken hipersensitivite reaksiyonları
Type Reaksiyon zamanı
Kilinik görünüm
'Histolojisi Antijen ve bağlanma bölgesi
İletişim 48–72 saat Egzama makrofajlar ve ardından lenfositleri; Epidermisin ödem
epidermal (organik kimyasallar, zehirli sarmaşık, ağır metaller)
Tüberkülin 48–72 saat Yerel endurasyon
Lenfositler, monositler, makrofajlar
Deri içi (tüberkülin, lepromin, vb )
granülom 21–28 gün Sertleştirme makrofajlar, epiteloid ve dev hücreler, fibrozis
Kalıcı antijen veya yabancı cisim varlığı (tüberküloz, lepra, vb )
Tip IV hipersensitivite enfeksiyonlar ve yabancı antijenlere bağlı olarak bir çok oto bağışıklık ve enfeksiyon hastalıkları (verem, cüzzam, blastomikoz, histoplazmoz, toksoplazmoz, layşmanyaz, vb) ve granülom patojenezinde rol oynamaktadır. Gecikmiş aşırı duyarlılık diğer biçimi lezyonlar örnek kontakt dermatit (zehirli sarmaşık ,kimyasallar, ağır metaller, vb) 'dir. Gecikmeli aşırı duyarlılık hasar mekanizmaları T lenfositler ve monositler ve / veya makrofajlar. Yardımcı T (TH1) hücreleri hasar kısmını neden monositler ve makrofajlar, sitotoksik T hücrelerini aktive ederek ve işle ve aktive sitokinler ise sitotoksik T hücreleri (Tc) zarar görebilir. Gecikmiş aşırı duyarlılık lezyonlar çoğunlukla monositler ve bir kaç T hücrelerini sınırlıdır. Gecikmeli aşırı duyarlılık reaksiyonu yer alan önemli lemfokinler monosit kemotaktik faktörü, interlökin–2, interferon-gama, TNF alfa / beta olarak sıralabilir. Tip IV hipersensitivite tedavisinde kortikosteroidler ve diğer bağışıklık bastırıcı maddeler kullanılır.
ŞEKİL 4.75 Tip IV duyarlılık reaksiyonları.

H A Y V A N F İ Z Y O L O J İ S İ
299
Tablo - aşırı duyarlılık reaksiyon türlerinin Karşılaştırılması
Özellikleri Tip-I (anafilaktik)
Tip-II (sitotoksik)
Tip -III (immün komp.)
Tip-IV (gecikmeli tip)
Antikor IgE IgG, IgM IgG, IgM Yok
Antijen Eksojen Hücre yüzeyi Çözünür Doku ve organ
Tepki süresi 15–30 dakika Dakika sonrası 3–8 saat 48–72 saat
Görünüm Parlama liziz nekroz
Eritem ve ödem, nekroz
Eritem ve endurasyon
Histoloji Bazofiller ve eozinofil
Antikor ve Komplement sistem
Komplement ve nötrofiller
Monositler ve lenfositler
Transfer şekli
Antikor Antikor Antikor T-hücrelerinin
Örnekler Alerjik astım, saman nezlesi
Eritroblastosis fetalis,
SLE, çiftçi akciğer hastalığı
Tüberkülin testi, zehirli sarmaşık, granülom
Püy(irin oluşumu)
Çok sayıda bakteri ve nekrotik dokuyu yutan nötrofil ve makrofajlar sonuçta ölür. Günler sonra, inflamasyonlu dokuda, içinde nekrotik doku parçaları, ölü nötrofiller, makrofajlar ve doku sıvısı bulunduran bir kavite oluşur. Bu karışım püy olarak adlandırılır. Enfeksiyon bastırıldıktan sonra püydeki ölü hücreler ve nekrotik doku günler içinde giderek otolize uğrar. Son ürünler çevredeki dokular tarafından absorbe edilir.

H A Y V A N F İ Z Y O L O J İ S İ
300
Sindirim sistemi fizyolojisi
Sindirim sistemi gastro intersistial kanal ve yardımcı organların hakkında bilgiler. Sindirim yeri, hormonal kontrolü, sindirim enzimleri yapı fonkisyonları, besin maddelerinin emilimi.
Sindirim Fizyolojisi Yaşam için gerekli enerjiyi hayvanlarda ancak besinlerdeki kimyasal maddelerden alabilir. Sindirim sisteminin görevi karbonhidrat, yağ, protein gibi başlıca besin maddeleriyle, su ve elektrolitlerin vücuda alınmasını sağlamaktır. Besin maddeleri kanaldan geçirilirken: Mekanik olarak parçalanır, Kimyasal olarak sindirilir, Basit moleküllerine ayrıştırılır, Gerekli ve yararlı olanlar emildikten sonra artıklar dışarı atılır. Sindirim besinlerin mekanik ve enzimatik olarak parçalanmalarıdır.
ŞEKİL 5.1 Farklı hayvan gruplarındaki sindirim sistemi
Sindirim sistemi aynı zamanda sindirim sistemi ve karaciğer, pankreas, safra kesesi ile mide-bağırsak (GI) oluşur. GI yolu anüs sonlanan ağzından başlayan uzun, bükümlü tüp şeklinde organize olmuştur. Gastrointestinal sistem oluşturan içi boş organların ağız, yemek borusu, mide, ince bağırsak, kalın bağırsak-rektum ve anüsten oluşur. Gıda ağızdan girer
Bölüm
5 Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
301
ve GI bölgesinin içi boş organları vasıtasıyla anüs kadar gider. Karaciğer, pankreas, safra kesesi ve sindirim sisteminine yardımcı organlardır. GI bakteriler, aynı zamanda sindirime yardımcı olur, bağırsak florasının veya mikrobiyomu besinleri sindiren enzimleri estra sellüer enzimler salgılar, çeşitli vitaminlerin üretimini sağlar(Vitamnin K). Sinir ve dolaşım sistemleri sindirim sürecinde rol oynar. Bu sistemler birlikte, sinirlerin düzenleyen, hormonlar salgılanmasını sağlar, bakterilerin, sindirim sisteminden kana geçişine engel olur(Karaciğer kuffer hücreleri).
�
ŞEKİL 5.2 Farklı hayvan gruplarındaki sindirim sistemi
Farklı besinlere göre sindrim sistemi
Etçilerde sindirim sistemi diğer memelilere göre sindirim borusu kısadır. (Sindirim sistemlerinin uzunluğu vücut uzunluğuna oranlandığında). Mide genellikle tek bölmeli ve kompleks bir yapısı yoktur. Çok bölmeli olması durumundaysa histolojik değil, fizyolojik bir farklılıktan söz edilir. Besinin niteliğine göre değişmekle birlikte, diğer memelilere oranla çok daha kısa ve düz bir kalınbağırsakları vardır. İlkel formlarda çekum gelişmiş değilken, diğerlerinde iyi gelişmiştir. Hepçilerde, Etçil memelilere göre sindirim borusu daha uzundur.(Sindirim sistemlerinin uzunluğu vücut uzunluğuna oranlandığında). Mide genellikle tek bölmeli ve kısmen kompleks bir yapıdadır. Temel besinin niteliğine göre değişmekle birlikte, etçil memelilere oranla daha uzun,
1- Besinin sindirim kanalı içinde yürütülmesi (GI motilite) 2- Sindirim sıvılarının salgılanması (GI sekresyon) 3- Sindirilen besin maddeleri, su ve elektrolitlerin emilmesi (GI absorbsiyon)

H A Y V A N F İ Z Y O L O J İ S İ
302
kaslı ve kısmen boğumlu bir kalınbağırsakları vardır. Marsupialia ve Primatlarda gelişmiş çekum bulunurken, diğerlerinde iyi gelişmemiştir.
Geviş getiren ve getirmeyen Otçullarda sindirim sistemi
Farklı canlı gruplarında sinirim yapan organ ve yapılar farklılık gösterir. Geviş getirenlerde Besinler önce, ağızda çiğnenir. Daha sonra besinler, işkembe ve börkeneğe geçer. Burada mekanik sindirime uğrar ve selüloz sindiren bakteriler, selülozu kimyasal sindirir. Geviş getirmeyen otçul memelilerde sindirim sistemi: At, eşek ve zebra bu gruba girer. Bu canlılara tek toynaklılar da denir. Bu canlıların mideleri, tek bölmelidir. İnce bağırsakları ise çok uzundur. Selüloz sindirimi kör bağırsakta gerçekleşir.
Otçul memelilerin diğer memelilere göre sindirim borusu daha uzundur.(Sindirim sistemlerinin uzunluğu vücut uzunluğuna oranlandığında) Mide genellikle çok bölmeli ve kompleks bir yapısı vardır. Çok bölmelilikte histolojik ve fizyolojik bir farklılıktan söz edilir. Temel besinin niteliğine göre değişmekle birlikte, diğer memelilere oranla çok daha uzun, kaslı ve boğumlu bir kalınbağırsakları vardır. Geviş getirenlerin mideleri çoğunlukla dört odadan oluşur:
Ayrıca işkembenin daha küçük bir ön odası vardır ki bu da ayrı bir mide olarak sayılırsa, midelerin sayısı beşe yükselir. Bazen işkembe'nin ve börkeneğin birbirlerine benzeyen görevlerinden dolayı birlikte tek bir bilimsel ad altında Reticulorumen olarak adlandırıldığına da rastlayabiliriz. Geviş getiren hayvanlar, otlanma anında bitkisel besinlerini ancak kabaca çiğner ve yutarlar. Bu yuttukları maddeler işkembeye gider. İşkembe ve diğer ön midelerinde çok sayıda bakteriler, protozoonlar ve maya mantarları gibi mikroorganizmalar vardır. Yutulan maddeler bu mikroorganizmalar ile iyice karışır. Bu mikroorganizmalar çoğu karbonhidratları işkembe duvarının alabildiği maddelere çevirebilirler. Fermentasyon denilen bu olayda çoğu diğer memelilerin sindiremedikleri bazı maddeler bile sindirilip enerji kaynağı olarak değerlendirilebilir (örneğin selüloz). Bu fermentasyonda meydana gelen gazlar börkeneğin içinde birikir ve ağız yoluyla dışarıya çıkar (geğirmek, garklamak).Yutulan maddeler daha çok parçalanmaları ve karışmaları için işkembe ile börkenek arasında ileriye geriye verilir. Bu işlem tamamlandıktan sonra bu hafif sindirilmiş maddeler börkenek ve işkembenin ön odasının kontraksiyonları ve yemek borusunun geriye doğru peristaltik hareketleri ile, küçük porsiyonlar halinde tekrar ağıza doğru taşınır. Ağızda maddeler geviş getirilerek daha çok parçalanır ve tekrar yutulur. Börkeneğin küçük ve büyük parçacıkları ayırabilme işlevi vardır; büyük ve kaba parçaları daima geri tutar ve ince parçaları kırkbayıra aktarır. Kırkbayır bu ince parçaları ezerek içindeki sıvıyı çıkarır. Bu şekilde katılaşan maddeler şirdene ulaşır. Şirdenin içinde asidin büyük bir kısmı notralize edilir. Maddede bulunan mikroorganizmaların
• Şirden (Abomasum) tek mideli memelilerin midesi gibidir. Bu mideye giden yolda yemek borusunun bölümleri olan:
• İşkembe (Rumen)
• Börkenek (Retikulum) ve
• Kırkbayır (Omasum) bulunur.

H A Y V A N F İ Z Y O L O J İ S İ
303
içindeki proteinler ve yağlar ince bağırsakta çıkarılır. Besinler bu kadar uzun süre midede tutulduğu, sürekli karıştırılıp katılaştırıldığı için geviş getirenlerin midelerinde bezoar taşları oluşur. Bu mide taşları, yutulmuş olan kıl ve bitki ipliklerinin zamanla birikip, yapışıp, sertleşmesi ile meydana gelir.
Sindirim sistemi besin maddelerini kan dolaşımına aktarılabilecek bileşiklere dönüştürür. Sığır, geyik, koyun, at ve tavşan gibi besin kaynağı olarak bitkilere bağımlı olan hayvan türlerine Herbivor denilir. Besin kaynağı olarak diğer hayvanların etlerine bağımlı olan kedi ve köpek hayvan türlerine Karnivor denir. Besin kaynağı olarak et ve bitkileri kullanabilen domuz ve kuş gibi canlılara ise Omnivor denir. Farklı hayvan türleri tükettikleri yemleri en iyi değerlendirebilecek şekilde adapte olmuş sindirim sistemlerine sahiptir. Herbivor, karnivor ve omnivorların sindirim sistemleri birbirlerinden farklılık gösterir. Ruminantlar besin madde ihtiyaçlarının büyük bir kısmının kaba yemlerden sağlamalarına imkân veren çok bölmeli bir sindirim sistemine (poligastrik) sahiptir. Sindirim kanalı dudaklardan anüse kadar uzanır. Sindirim sisteminde ağız, farinks, özafagus, mide ile ince ve kalın bağırsaklar yer alır. Yardımcı organlar arasında tükrük (salya) bezleri, karaciğer ve pankreas gibi organlar yer alır. Sindirim sisteminin uzunluğu ve yapısı (kompleksliği) türlere göre değişiklik gösterir. Herbivorlarda uzun ve kompleks yapıdadır.
Sindirim kanalının yapısı
Erişkin bir insanda yaklaşık 9 m uzunluğundaki sindirim kanalı fibromüsküler yapıda olup, yer yer işlevine uygun olarak farklılaşma gösterir. Sindirim kanalı ağız, farinks, özofagus, mide, ince ve kalın barsaklar ile anüsten oluşur. Sindirime yardımcı yapılar dişler, tükrük bezleri, pankreas, karaciğer, safra yolları ve safra kesesidir. Ağızdan ileumun distal ucuna kadar birçok yerde sindirim enzimlerinin salgılanması, Ağızdan anüse kadar bütün yolda sindirim kanalını yağlayan ve koruyan müküsün salgılanması.
ŞEKİL 5.3 Sinidirim sistemi modeli
• Ağız : Ağız boşluğundaki yapılar ve mekanik sindirim ve çiğneme görev alır.Ağızda tükürük müsin ( gıdaları bir arada bağlar) oluşur, tükürük amilaz (nişastayı maltoza sindirir ) ve mineral maddeler (yaklaşık nötr pH 7, dengeler.)
• Özofagus: Dairesel kaslar hareketi peristalsis ve gıda ve besin maddeleri aşağı doğru hareket ettirir. Yemek borusu, üst bölümü belirli bir yere kadar istemli kaslara sahiptir, Gıdanın ağzından mideye ulaşması için genellikle 4 ila 8 saniye süre yeterlidir.

H A Y V A N F İ Z Y O L O J İ S İ
304
• Mide: Asidik koşullara (optimum pH enzim pH 1-2,) sahiptir. Mekanik sindirim (gıda ona büyük bir yüzey alanı veren boyutta yıkmak amacıyla mide çalkalama hareketi yapar) yapılır. Gıda midede iken bu çalkalama ile mide sıvısı ile karışır. Midede gıda gastrin (kanda taşınan) üretilmesi için mide duvarına uyarır. Yağlı bir yemek enterogasterone yavaş çalkalama için sentezlenir. Aynı zamanda midenin asidik pH azaltılır. Mideden sentezlenen bağlıca maddeler : • Pepsinojen:- Şef bezlerinden proenzim olarak sentezlenir HCI ilave edildiği
zaman aktive olan bu zimojen enzim proteinlere etkiler Kendi dokuları otolitik olarak sindirmesini önlemek için İnaktif olarak sentezlenir ve daha sonra pepsin olarak aktif forma dönüşür. Görevi proteinlerin sindirimidir.
• Pro-renin: HCl sentezlendiğinde kaseinogen pıhtılaştırmasında ve çözeltilmesinde görev olan zimojen yapıdaki bir enzimdir. Bu yolla renin ve süt sindirmesi için yüzey alanını arttırır.
• Mukus: Sentezlenen mukus mideyi aşırı asidi karşı korur . Ayrıca aktif hale geçen enzimlerin mide içinde otoliz engeller. Bunun yanında sürtünmeyi azaltıcı etkisiyle yağlama maddesi olarak görev yapar.
• Hidroklorik asit (HCl) :Mide duvarında yer alan Paryetal bez hücrelerinden hidroklorik asit (HCl)salgılar.
ŞEKİL 5.4 Sinidirim kanalı
Mikrovilluslar ince bağırsağın iç yüzeyinde bulunurlar. Bu kadife tüyleri şekilndeki çıkıntılar yardımıyla sindirim son ürünleri kan ve lenf dolaşımına aktarılırlar. Lenf ve kan damarları tarafından zengin olan villi ve mikrovilliler yanlara doğru hareket ettikleri gibi uzama-kısalma yetenekleri de vardır. Villi hareketlerinin kontrolü sinir sistemi ve duodenum mukozasından salgılanan villikinin hormonu tarafından gerçekleştirilmektedir. Villiler bağırsak yüzeyinin olması gerekenden daha geniş olmasını sağlarlar Mikrovilliler villi üzerinde yer alan fırça benzeri küçük çıkıntılardır. Sindirim kanalının epitel tabakasında milyarlarca tek hücreli müköz bezler yer alır. Bunlara; müköz hücreler veya goblet hücreleri adı verilir. Sindirm kanalı epitel yüzey boyunca oyukcukluklarla (pit) kaplıdır. Pit ler ince barsakta Lieberkühn kriptaları (Entrokinaz) adını alır Mide ve duodenumun üst kısmında derin tübüler bezler bulunur(Şef ve periatel hücreler). Sindirim kanalı ile ilgili diğer bezler tükrük bezleri, pankreas ve karaciğerdir. Bezlerin Uyarılma İntestinal bezlerin uyarılması üç etkenden birisi ile gerçekleşir:
Mikrovillüs Hücrenin serbest yüzey farklılaşmalarından, özellikle emme görevi fazla olan hücrelerde, hücre dış yüzeyini arttırmak için, hücre zarının bir miktar sitoplazma ile meydana getirdiği parmak şeklindeki çıkıntılardır. Boyları yaklaşık, 0,6-0,8 mikron uzunluğunda, 0,08-0,1 mikron kalınlığındadır. Özellikle bağırsak epitelinde bulunan mikrovilluslar (çoğulu mikrovilli) yapılarında, makromolekülleri parçalayacak ve hücre içine taşıyacak enzimleri bulundururlar.
1- Dokunma uyarısı, 2- Kimyasal uyaranlar, 3- Barsak distansiyonu

H A Y V A N F İ Z Y O L O J İ S İ
305
Müküs salgısı
Müküs; su, elektrolitler ve birkaç glikoproteinden oluşan koyu bir salgıdır. Müküs besinlerin gastrointestinal kanal boyunca kolayca kaymalarını sağlar. Epiteli sıyrılmaya ve kimyasal etkilere karşı korur. Yapışkandır. Barsak duvarını kaplayan ve besin parçacıklarının mukoza ile doğrudan temasını önleyen yeterli bir kitlesi vardır. Sürtünmeye karşı direnci düşüktür. Feçes kitlelerini oluşturmak için feçes partiküllerinin birbirine yapışmasını sağlar. Gastrointestinal enzimlerin sindirimine karşı çok dirençlidir. Müküsteki mukopolisakkaritler az miktardaki asit veya alkalileri tampon edebilirler. Sindirim kanalı elektrolitler, su ve gıdaları sürekli olarak vücuda sağlar. Bunu gerçekleştirmek için
1) gıdaların sindirim kanalında hareketi, 2) sindirim salgılarının salgılanması ve besinin sindirimi, 3) sindirim ürünleri, su ve çeşitli elektrolitlerin emilimi, 4) emilen maddeleri uzaklaştırmak için gastrointestinal organlarda kanın dolaşımı 5) tüm bu fonksiyonların sinirsel ve hormonal mekanizmalarla kontrolü gerekmektedir.
Tükrük Salgısı
Tükürük insan ve diğer omurgalı ağızında sürekli olarak, mevcut olan renksiz, yarı şeffaf sıvıdır. Tükürük bezlerini tarafından salgılanan ağızda bulunan sulu bir maddedir. İnsan tükürük %99,5 su , % 0.5 diğer maddelerden, elektrolit , mukus, glikoproteinler, enzimler ve antibakteriyel bileşik olan IgA ve lisozim oluşur. Tükürük içinde bulunan enzimler, sindirim sürecinin başlatır (özellikle nişastar ve katı yağlarda). Bu sindirim enzimleri dişleri çürükten korumada görev alarak, diş çatlaklarına içinde tutulan gıda parçacıkları parçalar. Ayrıca, tükürük, yağlayıcı olarak görevi yapar, oral mukoza yüzeylerine kayganlık sağlar, gıda maddelerin ıslatılmasında ve yutmada görev alır.
ŞEKİL 5.5 Tükrük bezleri
Üç adet tükrük bezi (Parotid, Submandibuler, Sublingual) farklı yapıdaki tükrük sentezi yapar. Tükürüğün büyük bir kısmı submandibular tükrük bezi tarafından sentezlenir.
Tükrük bezleri Parotis bezi :Her iki kulak önünde, çene ekleminin yüzeyinde bulunur. Boyut olarak en büyük tükrük bezleridir. Sağ ve sol olmak üzere 2 adettir. Tükrük bezinin kanalı ağız içinde üstteki 2. azı dişlerinin hizasından ağız içine açılır. Bu bezi diğer tükrük bezlerinden ayıran en önemli özelliği yüzün hareketlerini sağlayan yüz sinirinin bu bez içinden geçmesidir. Bu cerrahide çok önem arz etmektedir. Cerrahi esnasında bu sinir bulunup korunması çok önemlidir. submandibüler ve sublingual bezler: Bu bezler çenenin alt tarafında sağda ve solda olmak üzere yerleşmişlerdir. Her iki bezden 2 adet bulunmaktadır. Tükrük bezleri tarafından yapılarak ağız boşluğuna boşaltılan salgılardır. İnsanda ağız boşluğuna açılan 3 çift tükrük bezi bulunur. Bunlar parotis, submaksiller ve sublingual bezlerdir. Yanağın iç kısmındaki küçük bukkal bezler az miktarda tükrük salgılarlar.

H A Y V A N F İ Z Y O L O J İ S İ
306
BEZ Histolojik tipi Sekresyon Toplam tükrüğün yüzdesi
Parotid Seröz Su %20 Submandibuler Karışık Kısmen visköz %70 Sublingual Müköz Visköz %5 Tükürük içinde su, bikarbonat, klor, müsin, müküs, Fosfat, Üre ve ürik asit bulunur. Bu maddelerin farklı görevleri vardır. İçindekiler Görevi Su Ağızı nemlendirir. Konuşmaya yardım eder. Tat alma ve sindirim işi
için çözücüdür. Bikarbonat Tükrüğün pH'sını 6.35-6.85 arasında (hafif asidik) tutmaya yarar Klor Tükrük amilazını aktifler Müsin Müküs oluşumuna katılan bir proteindir Müküs Besinleri yağlar, lokma oluşumunu sağlar. Yutmaya ve lokmadaki asit
ve bazları tamponlamaya yardım eder.
Fosfatlar Tükrüğün pH'sını düzenlemeye yardım eder. Üre, Ürik asit Tükrük yoluyla atılan artık maddelerdir. Sindirim fonksiyonları
yoktur Tükürük içinde farklı enzimler (pityalin) antimikrobiyal maddeler( IgA, Laktoferrin) farklı görev yapar. Enzim Görevi Pityalin Nişasta ve glikojeni maltoza parçalar IgA Bakteri ve viruslara karşı ilk immünolojik savunma Lizozim Bakterilerin duvarlarına saldırı Laktoferrin Demire bağlanır ve bakteriyostatik. Tiyosiyanat Antibakteriyel etki Prolinden zengin proteinler Dişlerin minesini korur
Özofagus Salgısı
Özofagus salgısı tamamen müküs karakterdedir ve görevi birinci derecede yutma için gerekli yağlamayı sağlamaktadır. Özofagusun baş kısmı ve mide ucundaki bezler birleşik müküs bezlerdir. Diğer taraflarında ise basit müküs bezler bulunur. Özafagus normal şartlarda mideden gelen sindirim enzimlerine ve düşük pH ‘a maruz kalmaz ancak mide özafagus arasındaki sifingterin yetersiz kalmasına bağlı olarak mide içeriği özofagusa kaçabilir. Bunu dışında kusma olayı sırasında özofagus müküs tarafından korunur. Mide
Yemek borusu ile barsakların ilk kısmı arasında bulunur ve bir bakıma sindirime hazırlanan gıdaların depo edildiği yer olarak görev yapar. Ağızda parçalanarak küçük bölümlere ayrılan ve tükrük ile yumuşatılan gıdalar, yemek borusu ile mideye aktarılır. Burada toplanan gıdalar bir süre daha belirli işlemlere tabi tutulur ve daha sonra belirli aralıklar ile barsağın birinci kısmına aktarılır. Midenin iki açıklığı vardır. Yemek borusunu mide ile birleştiren açıklığa kardia denir. Barsağın başlangıç kısmı ile birleştiren açıklığa ise pylor adı verilir. Pylorus sıkı oluşmuş sirküler kas liflerinden meydana gelmiştir. Dış görünüşüne göre mide üç kısma ayrılır. Bu bölümler; fundus, corpus ve pylor adını alır. Midenin ana bölümü korpus kısmıdır. Bu bölümün yukarıya doğru kubbe şeklinde yapmış olduğu çıkıntılı kısım diaphragma’nın altına sokulmuştur. Buraya fundus adı verilir. Gıdaların yutulması sırasında alınan
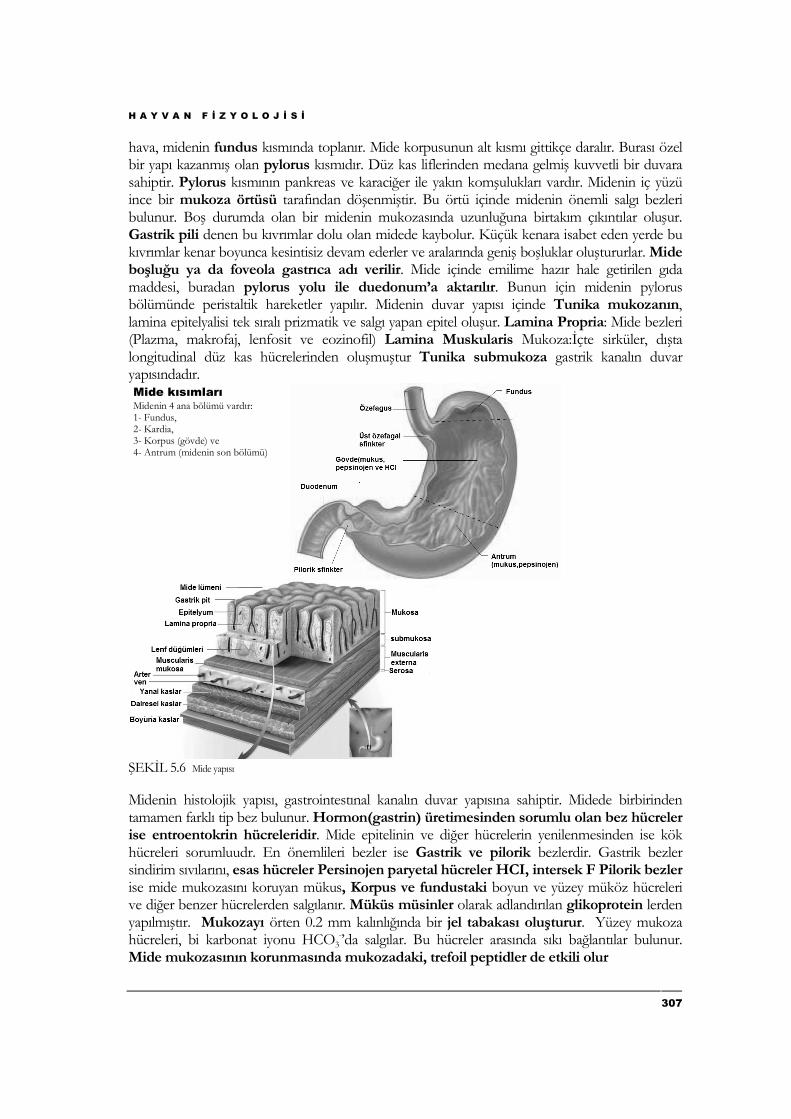
H A Y V A N F İ Z Y O L O J İ S İ
307
hava, midenin fundus kısmında toplanır. Mide korpusunun alt kısmı gittikçe daralır. Burası özel bir yapı kazanmış olan pylorus kısmıdır. Düz kas liflerinden medana gelmiş kuvvetli bir duvara sahiptir. Pylorus kısmının pankreas ve karaciğer ile yakın komşulukları vardır. Midenin iç yüzü ince bir mukoza örtüsü tarafından döşenmiştir. Bu örtü içinde midenin önemli salgı bezleri bulunur. Boş durumda olan bir midenin mukozasında uzunluğuna birtakım çıkıntılar oluşur. Gastrik pili denen bu kıvrımlar dolu olan midede kaybolur. Küçük kenara isabet eden yerde bu kıvrımlar kenar boyunca kesintisiz devam ederler ve aralarında geniş boşluklar oluştururlar. Mide boşluğu ya da foveola gastrıca adı verilir. Mide içinde emilime hazır hale getirilen gıda maddesi, buradan pylorus yolu ile duedonum’a aktarılır. Bunun için midenin pylorus bölümünde peristaltik hareketler yapılır. Midenin duvar yapısı içinde Tunika mukozanın, lamina epitelyalisi tek sıralı prizmatik ve salgı yapan epitel oluşur. Lamina Propria: Mide bezleri (Plazma, makrofaj, lenfosit ve eozinofil) Lamina Muskularis Mukoza:İçte sirküler, dışta longitudinal düz kas hücrelerinden oluşmuştur Tunika submukoza gastrik kanalın duvar yapısındadır.
ŞEKİL 5.6 Mide yapısı
Midenin histolojik yapısı, gastrointestınal kanalın duvar yapısına sahiptir. Midede birbirinden tamamen farklı tip bez bulunur. Hormon(gastrin) üretimesinden sorumlu olan bez hücreler ise entroentokrin hücreleridir. Mide epitelinin ve diğer hücrelerin yenilenmesinden ise kök hücreleri sorumluudr. En önemlileri bezler ise Gastrik ve pilorik bezlerdir. Gastrik bezler sindirim sıvılarını, esas hücreler Persinojen paryetal hücreler HCI, intersek F Pilorik bezler ise mide mukozasını koruyan mükus, Korpus ve fundustaki boyun ve yüzey müköz hücreleri ve diğer benzer hücrelerden salgılanır. Müküs müsinler olarak adlandırılan glikoprotein lerden yapılmıştır. Mukozayı örten 0.2 mm kalınlığında bir jel tabakası oluşturur. Yüzey mukoza hücreleri, bi karbonat iyonu HCO3
-’da salgılar. Bu hücreler arasında sıkı bağlantılar bulunur. Mide mukozasının korunmasında mukozadaki, trefoil peptidler de etkili olur
Mide kısımları Midenin 4 ana bölümü vardır: 1- Fundus, 2- Kardia, 3- Korpus (gövde) ve 4- Antrum (midenin son bölümü)

H A Y V A N F İ Z Y O L O J İ S İ
308
Mide salgı hücreleri
Midede bulunan pariyetal hücreler litrede 160 milimol hidroklorik asit (HCI) içeren, vücut sıvıları ile izotonik olan elektrolit bir solüsyon salgılarlar. Bu asit solüsyonun (HCl) pH'sı yaklaşık olarak 0.8 dir. Pariyetal hücreler bol miktarda H-K ATPaz ve oluşan asidi taşıyan intrasellülere kanaliküller içerirler. Mide suyunun sentezlenmesi için her litre mide sıvısı başına 1500 kalorinin üzerinde enerjiye gereksinim vardır bu neden çok sayıda mitokondri taşırlar. Midede HCI görevleri Pepsinojeni pepsine çevirmek ve pepsin için uygun değer pH'yı sağlamaktır. Bazı mineralleri redükte ederek (Ca2+, Fe2+) barsaktan emilimini kolaylaştırmak, Sütün kazeojenini kazein halinde çökeltmek ve mikropları öldürmek vardır.
ŞEKİL 5.7 Mide bezleri
Midedin pH düşürülmesi için hidrojenin mide lümenine transferi paryetal hücrelerin apikal zarındaki H+ - K+ ATPaz tarafından gerçekleştirilir. Bu aktif taşıma proteini H mide içine pompalarken K iyonunu pariyetal hücrelerin içine alır. Parasempatik uyarı(vagus sinir ile) ile (asetil kolin) ve gastrin hormonu (entroendokrin G hücrelerinden salgılanır) HCL salgılanmasını artırır. Histamin asetil kolinin ve gastrinin etkileri artırarark HCl salgılanmasını artıran diğer bir hormondur.
ŞEKİL 5.8 HCI sentezi
HCI asidin salgılanmasını sağlayan en önemli eleman mide pariyeal bezlerinde epitelyum hücrelerin cannaliculeri zarında bulunan bir H-K ATPaz veya "proton pompası" dır. Bu ATPaz magnezyum bağımlıdır.
Mide bezlerinden 1- Paryetal bezlerden HCI 2- Şef bezlerinden pepsinojen gastrik lipaz 3- Mukuz boyun bezlerinden mukus 4- D hücrelerinden somatistatin 5- G hücrelerinden gastrin

H A Y V A N F İ Z Y O L O J İ S İ
309
ŞEKİL 5.9 Hidrojen pompası
Asit salgılanmasını mekanizması:
• Hidrojen iyonları, parietal hücre içinde suyun ayrışmasından kaynaklanan oluştur. Bu işlemde karbonik anhidraz tarafından katalize edilen reaksiyonla karbon dioksit ve su birleştirilir. Sonuçta bikarboant ve hidrojen iyonları oluşur.
• Bikarbonat klorür karşılığında bazolateral membran üzerinden değiş tutuş yapılır (antiport) taşınır. Bunun sonunda "Alkali tide " olarak bilinen, kan pH hafif bir yükselti ile gözlenen bikarbonat çıkışı olur. Bu işlem, parietal hücre içerisinde hücre içi pH'ın muhafaza edilmesinde görev alır.
• Klorür ve potasyum iyonları sızma kanalları ile cannaliculus lümeni içine taşınır ve bu asit salgılanması için gereklidir.
• Hidrojen iyonu (proton) pompası tarafından potasyum karşılığında, lümenin içine, yani hücreden dışana pompalanır;
Besin mide lümenine geçtiğinde orada daha önce oluşmuş pepsin ve hidroklorik asitle temasa gelince proteinlerin sindirimi bağlar.
ŞEKİL 5.10 Pepsinojenin aktivasyonu
Şef hücrelerinden salgılanan pepsin ancak yüksek asit ortamda (pH 1,8–3,5) optimum etki gösterir. Mide Salgısındaki diğer enzimler; Gastrik lipaz, amilaz ve üreazdır. Gastrik Lipaz esasen bir tributinaz'dır. Tereyağındaki tributine etki eder. Gastrik amilaz nişasta

H A Y V A N F İ Z Y O L O J İ S İ
310
sindiriminde etkilidir. Gastrik jelatinaz etteki proteoglikanların bazılarının eritilmesine yardımcı olur. Gastrik üreaz mide bezlerinin salgısı olmayıp, midedeki bakterilerin etkisiyle oluşur.
ŞEKİL 5.11 Mide salgıları
Mide salgısı sinirsel ve hormonal mekanizmalarla düzenlenir. (1) Midenin dolu olup olmasına, besinlerin koku ve tatların alınmasına ve(2) sindirilmiş olan mide içeriğin bileşenlerine ile mide içeriğinin ince barsağa boşaltılmasıyla kontrol edilir. Mideye Parasempatik ve sempatik sinirler gelir. Sempatik sinirleri
gastrik ve epiploik arterlerin çevresinde çölyak pleksustan gelir. Parasempatik sinirler (asetil kolin kolinerjik reseptörlerle) ise vagustan gelir. Pankreas
Pankreas hem endokrin hem de ekzokrin bezdir. Ekzokrin pankreas asinar hücrelerden oluşur, sindirim enzimleri ve bikarbonatı pankreatik kanala salgılar ve bu da ince bağırsağa açılır. Endokrin pankreas ise langerhans adacıklarından oluşur ve hormon salgılar. Pankreas Ekzokrin salgısı pH değeri 8–8,5 seviyesindendir. Pankreastan günlük salgılanma miktarı : ~ 1500 ml /gün kadardır. Pankreas salgısında içinde bulunan temel katyonlar: Na+, K+, Ca+2, Mg++ iken temel anyonlar: HCO3-, Cl-, SO4
3-, HPO42-, albumin ve globulindür. Pankreas
salgısında çok farklı moleküllere etki eden farklı sindirim enzimleri vardır.
ŞEKİL 5.12 Barsakta aktive olan pankreaktik enzimler
Entrokinaz Bu enzim inca barsağın duedonum kısmında mikrovilluslarında bulunur. Pankreastan salgılanan bir zimojen olan tripsinojenin protein peptid zincirini kısaltarak aktif formu olan tripsine çevirir. Tripsin diğer pankreas zimojenlerini aktif forma çevirir. Tripsin
• Sefalik safha • Gastrik safha • İntestinal safha

H A Y V A N F İ Z Y O L O J İ S İ
311
Pankreastan salgılanan başlıca proteolitik enzimler; tripsinojen, kimotripsinojen prokarboksipeptidaz ve elastazdır. Pankreastan salgılanan tripsinojen enzimi barsak epiteli tarfından salgılanan enterokinaz tarafından aktif hale getirilir. Meydana gelen tripsin hem tripsinojeni hemde diğer proteolitik enzimleri aktifleştirir. Pankreas salgısı sinirsel ve hormonal mekanizmalarla düzenlenir.
ŞEKİL 5.13 Pankreas salgıları yapan hücreler ekzokrin ve endokrin bezler.
Midede sefalik safha başladığı zaman vagus sinirleri ile pankreasa da impulslar iletilir ve pankreas asinuslarına orta derecede bir enzim sekresyonu olur. İnce barsağın yukarı kısımlarında besinin bulunması başlıca iki hormonun sekretin ve kolestosistokinin salgılanmasına sebep olur ki bunlar pankreasın bol salgı yapmasını sağlarlar. Parasempatik sinirlerin sindirim kanalını uyarması bezlerin salgı miktarını artırır. Ancak intestinal bezler sempatik uyarımla inhibe olurlar. Pankreas taşıdığı çok sayıdaki zimojen enzimin aktif duruma geçerek kendi kendisini sindirmesini engel olan Pankreas Tripsin İnhibitörü (PTİ) ile otosindirim korunur. Gastroenteroendokrin Hormonlar
Sindirim kanalanın uygun zamanlarda uygun salgıları salgıması gerekir. Bu amaçla peptid yapısındaki hormonlar sindirim kanalalındaki hücrelerce sentezlenir. Sindirim kanalındaki başlıca hormonlar Gastrin hormunu, Kolesistokinin, Sekretin Gastrik ve motilindir.
ŞEKİL 5.14 Sindirimde görev yapan başlıca salgı hücreleri

H A Y V A N F İ Z Y O L O J İ S İ
312
• Gastrin hormunu; midedeki G hücrelerinden salgılanır. Mukoza gelişimini uyarır, mide asidi ve pepsin salgılanmasını uyarır, mide motilitesini artırır.
• Kolesistokinin hormonu; endokrin hücreler, üst GĠS, distal ileum ve kolondan salgılanır. Safra kesesinin kasılmasını artırır ve pankreasın enzim salgısını uyarır. Mide boşalmasını geciktirir, barsak motilitesini artırır.
• Sekretin hormonu; önce barsak S hücrelerinden salgılanır. Pankreas ve safra kesesinde bikarbonat sekresyonunu uyarır. Mide asidi salgısını azaltır.
• Gastrik inhibitör peptid (GIP), ince barsaktan doudenumdan salgılanır. Yağ ve glikoz varlığında mide salgısını inhibe eder. GİP serum glikoz seviyesi yüksek olduğunda insilün salgılanmasına neden olur.
• Vazoaktif intestinal polipeptid (VIP) • Ghrelin: Hipotalamus ve midede üretilir. Açlıkta artar, toklukta azalır. Leptinle
antagonist etkilidir. Tokluk etkisini melakortinin etkisini inhibe ederek sağlar. Uzun süreli beden ağırlığı regülasyonunda rol oynar.
• Motilin: açlık sırasında ince barsaktan salgılanır. Sinirim sistemi hareketlerini denetler
ŞEKİL 5.15 Sindirim hormonları ve mekanizmaları.
Safra kesesi boşaltılması Sekretinle pankreas salgısının uyarılmasıyla olur. Sekretin ince barsakların proksimal kısmının mukozasında prosekretin halinde bulunan ufak bir polipeptidir Kan dolaşımı ile pankreas salgıların salınmasına yol açar. Besinin barsakların proksimal kısmında bulunuşu buradan kolesistokinin salgılanmasına sebep olur. Sekretin gibi kolesistokininde kana geçerek safra salgısının salgılanmasına yol açar. Karaciğer hücreleri tarafından yapılan safra duktus sistikus yoluyla safra kesesine getirilerek biriktirilir ve konsantre edilir. Günlük safra salgısı 500-700 ml arasında olup, pH'sı 7,0-8,0 dır. Kesenin azami hacmi 40–70 ml. kadardır. Safra içindeki safra tuzları, safra asitleri, kolesterol, biluribin konsantre edilir. Su, sodyum ve klor kese mukozasından absorbe edilir.

H A Y V A N F İ Z Y O L O J İ S İ
313
ŞEKİL 5.16 Gastrin hormunu, Kolesistokinin, Sekretin mekanizmaları.
Enterohepatik dolaşım
Enterohepatik dolaşım safra tuzları ve diğer maddelerin yinelenen döngüyle karaciğerden atılması, bunların daha sonra bağırsak mukozasından geçerek tekrar karaciğer hücreleri tarafından alınması ve tekrar bağırsağa atılması sağlar. Safra salgısı 500–700 ml arasında olup yanlızca yüzde 5 kısmı atılır diğer kısmı elireke tekrar kullanılır. Safranın 2 işlevi vardır. Bunlardan birincisi, yağların sindirim ve emilimindeki işlevidir. Safra asitleri yağların pankreas lipazı tarafından parçalanabilecek küçük parçalara ayrılmasını sağlarlar. Ayrıca yağların sindirim ürünlerinin barsak mukozasından emilim ve taşınmasına yardım ederler. Safranın ikinci işlevi ise kolesterol ve bilirubin gibi yıkılım ürünlerinin atılmasını sağlamaktır. Bir kimyasal madde alınmasından sonrasında sindirim sistemi tarafından absorbe edilir ve hepatik portal sistem girer. Vücudun geri kalan ulaşmadan önce karaciğerden portal damar yoluyla geçer. Karaciğer, aktif ilacın(yada kimyasalın), sadece bir kısmını (az miktarda) dolaşım sisteminin geri gönderir, bazen kimyasal madde yada ilaç çok fazla metabolize olur. Bu duruma karaciğerden ilk geçiş etkisi (First pass effect) denir. Bu durumda, ilacın biyolojik olarak kullanılabilirliğini azalır ya da kimyasal maddenin etksisi sınırlandırılır.
ŞEKİL 5.17 Hepatik portal
Entrohepatik dolaşım yolu ile ilk geçişte elimasyona çok fazla oranda uğrayan ilaçların oral yolla değil diğer yollardan (parenteral) verilmesi tercih edilir. İlk geçişte eliminasyona uğrayan
Hepatik portal Portal ven kan akımı: 1000 – 1200 ml/dak. kadardır. Portal ven karaciğere gelen oksijenin %72’sini taşır.

H A Y V A N F İ Z Y O L O J İ S İ
314
ilaçlar genellikle çok fazla lipofiliktirler. Bu nedenle, emilimleri %100’e yakındır. Ancak, sistemik biyoyararlanımları %50’nin altındadır. Dolayısıyla, bu tip ilaçların oral ve parenteral dozları arasında da büyük farklar vardır.
İnce Barsak
Midede başlayan sindirim işlemi ince barsağın duodeunum kısmında devam eder. Barsak mukozasında ve pankreastik enzimlerce sindirilmiş olan besin maddeleri emilerek laınır. Dört-yedi metre uzunluğun sahip olan ince barsaklar 1-duodenum 2-jejunum 3- ileum kısımlarında oluşur. İki sfinter ince barsağı mide ve kalın barsaktan ayırı pilor sfinter ve ileoçekal sfinkter. Barsak epitel hücreleri günde yaklaşık 2000 ml salgı yaparlar. Salgı ekstrasellüler sıvı niteliğinde olup, pH'sı 6,5 – 7,5 arasındadır. İnce barsak salgılarında, tripsini aktive eden enterokinaz ve az miktarda da amilaz dan başka hemen hemen hiç enzim bulunmaz. Ancak mukozadaki epitel hücreleri çok miktarda sindirim emzimlerine sahiptir. Bunlar; peptidazlar, sükraz, maltaz, izomaltaz, laktaz ve barsak lipazıdır. Duodenumun ilk birkaç santimetrelik kısmında, başlıca pilor ile pankreas sıvıları ve safranın duodenuma boşaldığı yer olan vater papillası arasında Brunner bezleri bulunur (Mukus salgılar). Brunner bezlerinden müküs salgısına neden olan faktörler: Direkt dokunma uyarımları veya mukozayı tahriş edici maddeler, Aynı zamanda mide sekresyonunda da artışa yol açan vagal uyarı, İntestinal hormonlar, özellikle sekretin. Buna karşın sempatik uyarılar Brunner bezlerini kuvvetle inhibe eder. Barsak mukozasın üçüncü derece katlanmasıyla oluşan Liberkühn kriptaları entrokinaz salgılanmasından sorumludur. Liberkühn kriptaları basit tüpüler bezlerdir. İnce barsak epitelinde bulunan mikrovilluslar fırçamsı bir kenar oluşturarak emilim yüzeyini artırır. Bu yüzeyde laktaz, maltaz, entrokinaz, aminopeptidaz ve sükraz gibi çok çeşitli enzimler bulunur. Geniş bir alana sahip ince barsakların mikroorganzimalara karşı korunmasında Peyer plakları, Plazma hücresi ürünü olan Ig A ve Paneth hücreleri görev alır. Peyer plaktarı sindirim sistemin lenfoid dokusunu oluşturur. Az sayıda plazma hücresi taşır az özellikle proteazlarla savunma yapar. Plazma hücrelerince salgılana IgA ayrıca solunum epitelyum, süt ve tükrük içinde bulunur. Plazma hücreleri özellikle 3 tip lökositle birlikte bulunur eozinofiller, mast hücreleri ve makrofajlar. Paneth hücreleri TNF, lizozim ve defensin olarak bilinen proteinlerle savunma yapar.
ŞEKİL 5.18 Defensin ile baketri duvarı hasarı oluşturması
İnce bağırsakla kalın bağırsağın bağlantı kısmında bulunan körbağırsak (apendiks lenfoid bir doku) insanda çok küçülmüşken atlarda ve domuzlarda sellüloz sindirimi yapılabilmesi için çok gelişmiştir. Kemirici tavşanlarda ise çok uzun yapılı olan bu kısımda ek olarak B vitamini sentezlenir. Körbağırsaktaki salgının büyük bölümü müsin'dir. Müsin sindirim kanalını mekanik etkilerden korumasının yanısıra bağırsak duvarını bakteriyel aktiviteye

H A Y V A N F İ Z Y O L O J İ S İ
315
karşı korur. Aynı zamanda atık maddelerin birbirine yapışarak şekillenmesini sağlar. Sindirim ile ilgili enzim sekresyonu olmamasına karşın kalın bağırsaklara ulaşan içerikten mukoza hücreleri aracılığıyla su inorganik tuzlar bazı kısa zincirli yağ asitleri ve glukoz emilimi sağlanır. Kalın Barsak
Kalın barsak çekum, yukarı çıkan, transvers ve aşağı inen kolon, sigmoid kolon, rektum ve anüsten meydana gelir. Kalın barsaklarda yegâne önemli salgı müküs salgısıdır. Salgı miktarı 60 ml, pH 7,5 – 8,0 dir. Kalın barsakta müküs salgısı barsak duvarını sıyrılmalara karşı korur, feçes partiküllerini birbirine yapıştırır ve kitle oluşturan bir madde görevi yapar. Salgı artışı motilite artışı ile birlikte görülür. Kolon boyunca mineralokortikoidlerce (aldesteron) tarafında kontrol edilen Na ve CI emilimi vardır. Bu emilim peitel membranına yerleşmiş olan aktif taşıma proteinlerince yapılır.
ŞEKİL 5.19 Kalın barsak
Sindirim Kanaldaki Kontraksiyonlar 1-Tonik kontraksiyonlar Besinlerin karıştırılması ve peristaltik olarak itilme hızını, segment içindeki devamlı basıncın miktarını, sfinkterlerdeki tonik kontraksiyonlarda barsak içeriğinin hareketine karşı sfinkterlerin gösterdiği direncin derecesini belirler. 2-Ritmik kontraksiyonlar Gl yolda bazen 15–20/dk, bazen de 2–3/dk kez oluşurlar. Bunlar Sindirim yolunun son kısmı olan kalın barsaklar, besinin karıştırılması ve yürütülmesi gibi devirli görevlerin yerine getirilmesini sağlarlar. Kalınbarsaklarda su, Na+ ve diğer minerallerin absorbsiyonu yapılmaktadır. İleumdan kalınbarsaklara her gün 1000–2000 ml kimus geçer, bu kimusun 200-250 ml si kalın barsaklarda yarı katı feçese çevrilmektedir. Kalınbarsakların kuvvetli absorbsiyon yetenekleri vardır.

H A Y V A N F İ Z Y O L O J İ S İ
316
Sindirim kanalındaki başlıca Sfinkterler
ŞEKİL 5.20 Sindirim sistemi bulunan sfingterler
• Hipofaringiyal - Üst özofagus sfinkteri • Kardia - Gastro özofageal sfinkter • Pilor sfinkter • İleoçekal sfinkter • Anal sfinkterler

H A Y V A N F İ Z Y O L O J İ S İ
317
Besinlerin Kimyasal sindirimi Sindirim temel olarak iki basamakta yapılır:
1. Mekanik Sindirim: Besinlerin enzim kullanılmadan fıziksel olarak parçalanmasına mekanik sindirim denir. Ağızda dişler, mide ve bağırsaklarda ise kaslar yardımıyla gerçekleşir. Mekanik sindirimin amacı besinlerin sindirim yüzeyini artırarak enzimlerin etkisini hızlandırmaktır. Mekanik sindiri büyük molekülleri kücük moleküllere ayırmaktır.
2. Kimyasal Sindirim: Besinlerin su ve enzimler yardımıyla parçalanmasıdır. Ağızdan bağırsaklara doğru kimyasal sindirim artar. Kimyasal sindirim besinleri en küçük yapı taşına kadar ayırmaktır.
Protein sindirimi
Diyetteki proteinler sindirim sistemindeki proteolitik enzimler ve peptidazlar tarafından amino asitlerine parçalanırlar. Proteinler, ince barsaklardan emilemeyecek kadar büyüktürler bu sebeple amino asitlerine kadar parçalanmaları gerekir. Ancak çok az miktarda olmak üzere bazı küçük proteinler(2-3 peptidlik oligopeptidler)
ve peptitler barsak tarafından doğrudan emilebilirler(alerjik reaksiyona yola açar). Proteinaz ve peptidazlar, mide ve bağırsak mukoza hücrelerinde, pankreas hücrelerinde, oluştuğu organın kendisinin sindirilmemesi için aktif olmayan proenzim (zimojen) şeklinde oluşurlar ve salgılandıktan sonra parçalanma suretiyle aktif hale gelirler. Mide mukoza hücrelerinden salgılanan 42500 molekül ağırlıklı pepsinojen, HCl etkisiyle 35000 molekül ağırlıklı aktif pepsin haline dönüşür. Pepsin, optimal olarak 5’ten küçük pH’da etkili bir endopeptidazdır. Pepsin, seçimli olarak polipeptit zincirindeki tirozin, fenilalanin, glutamat ve aspartat gibi amino asitlerin oluşturduğu peptit bağlarını parçalar ve peptonları oluşturur. Tripsin, pankreastan 24500 molekül ağırlıklı inaktif tripsinojen şeklinde salgılanır. Tripsinojen, ince bağırsakta enteropeptidaz veya tripsin etkisiyle aktif tripsin haline dönüştürülür. Tripsin, bir endopeptidazdır; aktif merkezinde serin ve histidin bulunur.Tripsin, peptonlardaki arjinin ve lizin gibi bazik amino asitlerin oluşturduğu peptit bağlarını parçalar. Kimotripsin, pankreastan kimotripsinojen şeklinde salgılanır ve tripsin etkisiyle αααα-kimotripsin şekline dönüştürülür. αααα-kimotripsin, bir endopeptidazdır. αααα—kimotripsin, peptonlardaki fenilalanin, tirozin, lösin ve triptofanın oluşturduğu peptit bağlarını parçalar. Elastaz, proelastaz şeklinde salgılanır. Proelastaz da tripsin vasıtasıyla elastaz şekline dönüştürülür. Elastaz, elastindeki nötral alifatik zincirli amino asitlerin oluşturduğu peptit bağlarını parçalar. Mideden ince bağırsağa gelen polipeptitler, ince bağırsak lümeni içinde tripsin, kimotripsin, elastaz gibi Endopeptidazlar ve karboksipeptidaz A, karboksipeptidaz B, aminopeptidaz gibi Ekzopeptidazların etkisiyle oligopeptitlere, dipeptitlere ve amino asitlere parçalanırlar. Karboksipeptidaz B, prokarboksipeptidaz B şeklinde salgılanır. Prokarboksipeptidaz B de tripsin etkisiyle aktif olan karboksipeptidaz B şekline dönüştürülür. Bir ekzopeptidazdır; oligopeptitlerin karboksil ucunda arjinin ve lizin gibi bazik amino asitlerin oluşturduğu peptit bağlarını parçalar. Aminopeptidaz, oligopeptitlerin amino ucundaki peptit bağlarını ve dipeptitleri parçalar. Midede ısı ve HCl etkisiyle denatüre olan proteinler, midede ve ince bağırsaklarda proteinaz ve peptidazlar yardımıyla parçalanırlar.
Liziz biyo moeküllerdeki (polimeri oluşturan monomerlerin) C-C, C-O, yada C-N bağların kırılmasıyla parçalanması ve çift bağların oluşmasıdır.

H A Y V A N F İ Z Y O L O J İ S İ
318
ŞEKİL 5.21 Protein sindirimi
Bağırsaklara gelen proteinin iki kaynağı vardır. 70–100 g diyetle alınan 35–200 g endojen kaynaklı proteindir. Endojen kaynaklı proteinlerin kaynağı bağırsaktan salgılanan enzimler ve proteinler veya intestinal epitel hücrelerinin yıkılmasından ortaya çıkar. Polipeptid zincirini oluşturan aminoasitler arasındaki peptid bağının hidrolizi (proteoliz) zincirin her iki ucundaki ilk peptid Ektopeptidaz, iç kısımlarındaki peptid bağlarına etki eden Endopeptidazlarla olmaktadır. Protein sindirimi gastrik, pankreatik ve intestinal fazlara ayrılmaktadır.
ŞEKİL 5.22 Protein emilimdeki farklı yollar
Protein sindirimi proteinler sindirim kanalında proteolitik enzimlerle amino asitlere parçalanır . Emilim amino asitler yada küçük peptidler halinde gerçekleşir Di ve tripeptidler H iyonuna bağlı kotransport, Mono amino asitlerin Na bağlı transportu ve son olarak düşük sayılı polipeptidlerin transitozla barsak epiteline alınması.

H A Y V A N F İ Z Y O L O J İ S İ
319
Enzim Substrat Etki Proteolitik
• Tripsin • Kimotripsin • Karboksipeptidaz • Elastaz
Protein ve peptidler
Protein ve peptidlerin içindeki aminoasit bağarını kırarlar
Amilolitik • Amilaz
Polisakkaritler Polisakkaritlerin internal glukoz bağlarını kırar
Lipolitik • Lipaz • Fosfolipaz A2 • Kolesterol esteraz
Triasilgliseroller Lesitin Kolesterol esterleri
Triasilgliserolden iki yağ asidi parçalar Lesitinden bir yağ asidi ayırır. Kolesterol esterlerindeki ester bağlarını kırar
Nükleolitik • Ribonükleaz • Deoksiribonükleaz
Nükleik asitler
Nükleik asitleri ayırır
a- Gastrik Protein Sindirimi Mide mukozasında bulunan 30 milyona yakın salgı hücresinden salgılanan sıvı midede protein sindirimini başlatmaktadır. Mukoza hücrelerinden gastrin, esas hücrelerden pepsinojen, parietal hücrelerden ise 0,16 M HCL, 0,007 M KCL, intrinsik faktörve ATPaz içeren bir sıvı salgılanmaktadır. Denatürasyonla polipeptid zincirinin katlanmasının açılması sağlanır ve bu açılmada proteine proteazların etkilemesini kolaylar. Gastrin, parietal ve esas hücrelerden HCL ve pepsinojen salgılanmasını uyaran bir hormondur. Pepsinin, prekürsörleri ya pH 5’den daha düşük ortamda intramoleküler reaksiyon ile veya aktif pepsin ile aktive edilir. pH>2 olduğu durumlarda, serbestleşmiş peptid pepsine bağlı kalır ve oeosin aktivitesi için bir inhibitör gibi davranır. Bu inhibisyon pH nın 2’ye veya daha aşağı inmesi ile ortadan kalkar. Kolesistokinin (pankreozmin), polipeptid yapısında bir hormon olup pankreas zimojen granüllerinde bulunan proenzimlerin salıverilmesini sağlamaktadır. Yine duedonum ve jejunumdan salgılanan sekretin, pankreas kanalından su ve bikarbonat salgılanmasını uyarmaktadır. b- Pankreatik Protein Sindirimi İnce bağırsak salgısı içerisinde bulunan Enteropeptidaz (Enterokinaz) inaktif form tripsinojen (inaktif zimojen) molekülünün N ucundan bir hekzapeptidi uzaklaştırarak aktif tripsin oluşturmaktadır. Aktif tripsin aktive edildiği gibi diğer pankreatik zimojenleri Elastaz, Kimotripsin, Karboksipeptidaz A ve B’yi aktive eder. Elastaz, glisin ve alanin gibi küçük ve yüksüz yan grubu olan aminoasitlerin oluşturduğu peptid bağlarını hidroliz eder. Pankreas için potansiyel bir tehlike oluşturan proenzimlerin etkisine karşı pankreas kendini korumak için pankreatik tripsin inhibitör (PTI) adı verilen küçük bir protein sentezlemektedir. Bu protein, biyokimyada bilinen en güçlü kovalent olmayan bağ ile tripsinin aktif kısmına bağlanarak, tripsini aktive etmektedir. Pankreas kanalının tıkandığı durumlarda

H A Y V A N F İ Z Y O L O J İ S İ
320
pankreasta başlayabilen zimojen aktivasyonu, dokunun kendi proteininin sindirilmesine ve akut pankreatit diye isimlendirilen hastalık tablosunun ortaya çıkmasına yol açmaktadır. c- İntestinal Protein Sindirimi Aminoasitler en fazla jejunumdan olmak üzere ince bağırsağın her üç kısmından aktif taşıma ile lümenden bağırsak hücresine alınmaktadır. Aminoasit alımı için fırçamsı kenarın membran Na+ bağımlı Simport Sistemi, karşı yöne Na+ u taşıyan ATP bağımlı Na+ Pompası Sistemi ile fonksiyonel işbirliği içerisindedir. Bu, glukozun taşınmasında olduğu gibi indirekt aktif bir prosestir. Benzer şekilde fırçamsı kenarda di ve tripeptid transportu H+ bağımlı simportu ile enterosite taşınır. Kapiller taraftaki Na+ bağımlı transport sistemi iyon dengesini sağlar. Bu iki H+ bağımlı ve Na+ bağımlı sistem, di ve tripeptidlerin kolaylaştırılmış transportunu sağlayarak onlrı hepatik portal sisteme geçirir.
ŞEKİL 5.23 Amino asitlerin Na bağlı sekonder aktif taşıma mekanzimaları.
Bazı durumlarda bağırsak mukozasındaki defekt nedeniyle proteinler tamamen hidroliz olmadan emilmektedirler. Yabancı proteine veya oligopeptidlere karşı vücutta oluşması sonucunda alerjik reaksiyon ortaya çıkmaktadır. Non tropikal sprue (çöliak) hastalığında buğday proteini olan gluten hidroliz olmadan bağırsaktan emilerek organizmada alerjik yanıta yol açmaktadır. Hergün metabolize edilen 500 g aminoasitin 400 g kadarı yeniden protein sentezinde kullanılmakta, geri kalanı ise çeşitli katabolik yollara girmektedir. Besinlerle kg vücut ağırlığı başına 30–60 g protein, normal azot dengesinin sağlanması bir yetişkine (asgari 20 g) yetmekle beraber ortalama 100 g protein alımı sağlığa uygun bir beslenme olarak önerilmektedir. Aminoasit yapımında kullanılan ön maddelerden α ketoasitler: Bir aminoasidin karbon iskeletine uyan α ketoasit organizmada mevcutsa o aminoasit endojen olarak sentezlenir ve bunu amino transferazlar gerçekleştirir. İnce bağırsak lümenindeki amino asitlerin hemen hemen hepsi ince bağırsağın orta kısmından, intestinal villuslerin hücreleri tarafından emilirler ve emilen amino asitlerin çoğu portal kana geçerek karaciğere taşınırlar. Amino asitlerin emiliminde Na kotranstportu kullanılır. Bazı di ve tripeptidler H kotransportu İle alınabilir, Küçük peptidler transitozla hücre içine alınır. Sindirilen proteinlerden oluşan serbest amino grup asitlerin duodenumda kalış süreleri çok kısadır. Absorbe edilen amino asitler portal dolaşımla karaciğere taşınırlar. Bebeklerde peptitler de (globulinler) absorbe edilirken erişkinlerde sadece amino asitler bağırsaklardan emilebilir. Temel olarak amino asitlerin emilimi bağırsak epitelindeki ATP iel çalışan aktif transports sistemlerine( Na- Amino asit ATPaz ve H-Amino asit ATPaz)bağlıdır.

H A Y V A N F İ Z Y O L O J İ S İ
321
Taşıyıcı proteinin özelliğine göre en az dört tip amino asit taşıma sistemi vardır:
Di ve tripeptid üzerinde kana karışan oligopeptidler alerjik reaksiyonlara neden olur. Alınan tüm amino asitler karaciğer üzerinde sistemik dolaşıma katıldığı için karaciğer amino asit vücuda ihtiyaca göre proteinler halinde verir. Karbonhidrat sindirimi
Doğada hem αααα hem ββββ (1,4) endoglukozidazlar olduğu halde, ikinci grup insanda bulunmaz. Bu nedenle insanlar sellülozu sindiremez. Amilopektin ve glikojen de αααα(1,6) bağları içerdiğinden αααα-amilazın etkisiyle ancak daha küçük dallı oligosakkaridler (limit dekstrinler) açığa çıkar.
Ağız: Karbohidratların sindirimi ağızda başlar tükrük içinde bulunan alfa amilaz uzun polisakkarit zincirlerinin ağız içinde parçalamaya başlar. Parçalama işlemi nötür pH değerlerinde yapılır. Tükürük αααα-amilazı, amilopektin ve amilozdaki αααα(1→→→→4) glikozid bağlarını rastgele parçalayarak küçük moleküllü dekstrinlerin oluşumunu katalizler. αααα-amilaz, polisakkaritlerdeki iç bağları hidrolizler.
ŞEKİL 5.24 Niaşastanın alfa amilaz yıkımı.
Mide: Karbonhidrat sindirimi midede geçici olarak duraklar, çünkü yüksek asidite tükürük α-amilazını inaktive eder.
Pankreas: İnce barsaklarda sindirim sürer. Asidik mide içeriği ince barsaklara gelince, (g.i.) mukozadan sekretin salgılanır, bunun etkisiyle de pankreastan salınan bikarbonat pH'yı nötralize eder. Bir diğer sindirim hormon olan Kolestosistokinin etkisiyle salgılanan pankreatik αααα-amilaz sindirimi sürdürür. (yine αααα(1,4) bağlarını kırar.) Besin maddeleri mideden duodenuma geçtiğinde, karbonhidrat sindirimi, bikarbonat (HCO3
−−−−) ve pankreas αααα-amilazı içeren pankreas özsuyunun (Pankreas özsuyu, duodenuma günde 1,5 litre kadar
L-aminoasitlerin intestinal lümenden alınımı için; 1- Kısa ve polar yan zincirli aminoasitler (serin, Thr, Ala) için nötral aminoasit simporter 2- Aromatik veya hidrofilik yan zincirli aminoasitler (Phe, Tyr, Met, Val, Leu, İle) için nötral aminoasit simporter 3- İmino asit simporter (pro, OH-pro) 4- Bazik aminoasit simporter (Lys, Arg, Cys) 5- Asidik aminoasit simporter (Asp, Glu) 6- β aminoasit simporter (β Ala, Tau)

H A Y V A N F İ Z Y O L O J İ S İ
322
salgılanmaktadır.) etkisi ile devam eder. Pankreas αααα-amilazı, polisakkaritlerdeki αααα(1→→→→4) glikozid bağlarını hidrolize eder ve sonuçta maltoz, izomaltoz ve 3-8 glukozil kalıntısı içeren limit dekstrinler oluşur.
Barsaklar: Karbonhidrat sindiriminin son aşaması intestinal mukoza hücrelerince sentezlenen oligo ve disakkaridazlar ile gerçekleştirilir. Enzimler yoğun olarak jejunumun üst kısmındaki mukozal hücre yüzeylerinde bulunurlar.
Limit dekstrinlerdeki αααα(1→→→→6) glikozid bağlarının hidrolizi, ince bağırsak epitel hücrelerinin salgısı olan ince bağırsak salgısında bulunan ince bağırsak 1,6-glikozidazı etkisiyle olur. Limit dekstrinlerdeki dallı durumun ortadan kalkması ve αααα-amilazın tamamlayıcı rolüyle en sonunda trisakkaritler ve disakkaritler oluşur ki genellikle oluşan maltoz ve izomaltoz disakkaritleridir. Sindirim tamamlanınca glukoz, galaktoz ve fruktoz açığa çıkar.
Karbohidrat ve Monosakkaritleri Emilimi: Disakkaritler, ince bağırsak epitel hücresi zarında yerleşik uygun disakkaridazlar tarafından tutulurlar; geçiş sırasında hidrolizlenerek monosakkaritlere ayrılırlar ve böylece oluşan monosakkaritler ince bağırsak epitel hücresi içine ve oradan kana geçerler. Maltaz, izomaltaz, sakkaraz ve laktaz, ince bağırsak epitel hücrelerinin fırçamsı kenarında yerleşmiş olarak bulunan enzimlerdir. Duodenum ve jejunumun üst kısmı diyet şekerlerinin büyük kısmını absorbe eder.
ŞEKİL 5.25 Nono sakkaritlerin barsak lümeninden emilim mekanizmaları
İntestinal hücrelere glukoz alınması için insülin gerekmez. Ancak her şekerin emilme mekanizması farklıdır. Galaktoz ve glukoz mukozal hücrelere, hem aktif transportla (Na ile birlikte) hem de kolaylaştırılmış transportla (spesifik bir taşıyıcı ile) taşınırken, fruktoz, kolaylaştırılmış difüzyon ile emilir. Glukoz ve galaktozun mukoza hücrelerini terk etmesi ise, kolaylaştırılmış transport ve basit difüzyonla olurken, fruktoz muhtemelen pasif difüzyonla portal dolaşıma girer. Na bağımlı olanlar florhizin, bağımsız sistemler ise sitokalazin B ile inhibe olur.

H A Y V A N F İ Z Y O L O J İ S İ
323
ŞEKİL 5.26 Karbohidrat yıkımı
Karbonhidrat sindiriminde farklı dokulardan salgılanan enzimler görev alır: Lingula amilaz, Gastrik amilaz, Pankreaktik amilaz enzimleri görev yapar. Temel olarak uzun bir sakkarit zinciri önce daha küçük zincirlere en sonunda ise glikoz, fruktoz ve galaktoz haline getirilir.
ŞEKİL 5.27 Monosakkarit emilim mekanizması
• GLUT1 ve GLUT3, tüm memelilerde bazal glikoz alınması gerçekleştirir. GLUT1 ve GLUT3 hücrelerin metabolik faaliyetleri için gerekli olan glikozu sağlar.
• GLUT2, karaçiğer ve pankreatik beta hücrelerinde bulunur. Yüksek Km değerine sahip olduğundan ancak yüksek konsantrasyonlarda (15-20mM) bu hücrelerde çalışır. Böylece yüksek kan glikoz seviyelerinde karaçiğer ve pankreatik beta hücrelerin gereken uyarıyı ve aktivasyonu sağlar. Örneğin pankreasın beta hücrelerinden insulin sağlılanmasını sağlar. Benzer şekilde yüksek kan glikoz seviyelerinde karaciğer hücrelerinin glikoz alması sağlanır.
• GLUT4, Km değeri 5 mM olup insülin varlığında kas ve yağ dokusunun glikoz alınmasına olanak tanır. İnsülin ile kontrol edilir.
• GLUT5, Fruktoz transferi yapan taşıma proteini olup ince barsakta yer alır. Fruktoz barsak epitelinden özel bir protein (GLUT5) aracıyla kolaylaştırılmış diffüzyonla taşınır. Karbonhidratların sinidirim sonunda en fazla ortaya çıkan glikoz sinidirimi aktif taşıma mekanizmasıyla yapılır.
Karbonhidratların sindirimi Amilaz, bitkilerde bulunan nişastayı, bir disakkarit olan maltoza katalizleyen enzimdir. İnsanlarda, tükürük bezleri ve pankreasta salgılanır. Besinlerin çiğnenmesi sırasında nişastayı ve öteki büyük karbonhidrat moleküllerini daha basit maddelere parçalar. Alfa amilaz canlı organizmalarda çok bol bulunan bir enzimdir. İnsanın ve memeli hayvanlardan birçoğunun sindirim sisteminde, biri pityalin adıyla bilinen ve tükürük bezlerinde üretilen; öbürü amilopsin adıyla bilinen ve pankreas tarafından incebağırsağa salgılanan iki alfa amilaz önemli rol oynar.
GLUT Glukoz taşıyıcı 1 (ya da GLUT1) ayrıca, çözünen taşıyıcı alilesi 2 olarak da bilinir, kolaylaştırılmış glikoz taşıyıcı elemanı 1 (SLC2A1)uniporter(tek bir molekül tek yönde taşır), insanlarda SLC2A1 geni tarafından kodlanan bir proteindir. GLUT1 glukoz taşınmasını kolaylaştıran bu protein memeli hücrelerinin plazma zarlarında bulunur

H A Y V A N F İ Z Y O L O J İ S İ
324
ŞEKİL 5.28 GLUT tipleri ve Na bağlı glikoz taşınması
İnsülin hormonun yağ ve kas hücrelerinde glukoz alımını düzenlemesine olan etkisi son yıllarda aydınlatılmıştır. Son kanıtlar insülin etkisinin hücrede çoklu yollar olduğu göstermektedir. Hormona bağlı olan aktivasyonda, reseptör bağlı olarak, bir dizi tirozin fosforilasyonuna katalizörlük eder. Bunlardan biri, insülin reseptör substrat (IRS) protein ailesidir, protein kinaz C gibi protein kinazlann uyarılması ile sonuçlanan, fosfatidilinositol 3-kinaz yolağının aktivasyonunu başlatan lipid yıkının bulunduğu G proteini olan TC10, aktivasyonu sağlar. TC10 ise aktin filamentin(mikro filament) hücre iskeletinin değişikliklere yol açmasına adaptör protein CIP4 ile sağlar. Bu olaylar sonunda eksositoz gibi hücresel faaliyetler düzenlenebilir. Bu yollar kolaylaştırıcı glukoz transporter GLUT4 ile yağ ve kas hücrelerinde glikoz alını kontrol edilebilir. Glikoz barsak epitelinde sekonder aktif taşıma sistemiyle Sodyuma bağlı olarak taşınır. Glikozun emilimi SGLT1 proetienlerince kontransport ile yapılır. Barsak epiteli alınan monosakkaritlerin hepatik portal katılması ise farklı tipteki kolaylaştırılmış diffüzyon yapan proetinlerce yapılır.
ŞEKİL 5.29 Farklı dokulardaki farklı tipteli glikoz taşıma proteinleri.

H A Y V A N F İ Z Y O L O J İ S İ
325
Yağların sindirimi
Yağların sinidirimi özellikle barsaklarda yapılır. Buna karşın ağızda(lingual lipaz) ve midede (gastrik lipaz) vardır. Kolesterol ve kolesterol esterleri ağızda ve midede değişikliğe uğramadan ince bağırsağa gelirler. Mideden ince bağırsağa gelen kimus, asit reaksiyondadır; safra ve pankreas sıvısı tarafından nötralize edilir. Nötralize kimus içindeki lipidler üç basamakta sindirilirler. Yağlar organizmada yapı ve enerji maddesi olarak kullanılır. Yağların fiziksel sinirimi ince bağırsakta başlar ve tamamlanır. Ligual lipaz ağızda yetersiz sindirim yapar. Gastrik Lipaz esasen bir tributinaz'dır. Tereyağındaki tributine etki eder. Temel sindirim barsakta yapılır. Sekretinin uyarmasıyla pankreastan yağların sindirimi için lipaz enzimi salgılanır. Kolesistokinin hormonu ise kan yoluyla karaciğeri uyararak, oniki parmak bağırsağına safra sıvısının (öd sıvısı) salgılanmasını sağlar. Safra salgısı, yağları daha küçük parçalara ayırarak bir çeşit mekanik sindirim gerçekleştirir. Yağ damlacıklarının yüzeyini genişleterek lipaz enziminin etkinliğini arttırmış olur. Lipaz enzimi, su ile birlikte yağların, yağ asitleri ve gliserole parçalanmasını sağlar. Lipitler safra kesesinden salgılana safra tuzları ile pankreastik lipaz ile parçalanır. İnce bağırsak mukoza hücresinde yağ asitleri, koenzim A ile aktiflendikten sonra 2-monogliseridlerle esterleşirler ve tekrar trigliserid oluştururlar. Az miktarda emilen 1-monogliseridler de, pankreatik lipazdan farklı bir lipaz etkisiyle gliserole parçalandıktan sonra trigliserid sentezi için kullanılırlar. İnce bağırsak mukoza hücresinde 2-monogliseridlerden oluşan eksojen trigliseridler, az miktarda serbest kolesterol, kolesterol esteri ve fosfolipid ile biraraya gelirler; bir protein tabakasıyla da kaplanarak suda çözünebilir ve transport edilebilir şilomikronları oluştururlar. Şilomikronlar da lenf sistemi yoluyla dolaşıma katılırlar. Şilomikronlarda ağırlıkça % 2 oranında protein, %1 oranında serbest kolesterol, %3 oranında kolesterol esteri, %9 oranında fosfolipid, %85 oranında trigliserid bulunur. Şilomikronlar başlangıçta Apo B48 ve Apo A içerir. Daha sonra dolaşım sürecinde HDL ile etkileşme sonucunda Apo E ve lipoprotein lipazı aktive eden ApoCII apolipoproteinleri şilomikronlara katılır.
�
� Sekretinin uyarmasıyla pankreastan yağların sindirimi için lipaz enzimi salgılanır � Kolesistokinin hormonu ise kan yoluyla karaciğeri uyararak, oniki parmak
bağırsağına safra sıvısının (öd sıvısı) salgılanmasını sağlar. � Safra salgısı, yağları daha küçük parçalara ayırarak bir çeşit mekanik sindirim
gerçekleştirir. � Yağ damlacıklarının yüzeyini genişleterek lipaz enziminin etkinliğini arttırmış olur. � Lipaz enzimi, su ile birlikte yağların, yağ asitleri ve gliserole parçalanmasını sağlar

H A Y V A N F İ Z Y O L O J İ S İ
326
Lipaz adı Kısaltma Konum Bilgi
Pankreatik lipaz PNLIP sindirim sıvısı En iyi enzim etkinliğini elde etmek için pankreatik lipaz,
pankreas tarafından salgılanan, kolipaz adlı başka bir
proteine gerek duyar
Lizozomal lipaz LIPA Lizozomun içi Lizozomal asit lipaz veya kolesteril ester hidrolaz olarak
da bilinir
Hepatik lipaz LIPC endotel Hepatik lipaz, kandaki lipoproteinlerdeki lipitler üzerine
etkir, düşük yoğunluklu lipoproteinler (LDL) oluşturur .
Lipoprotein lipaz LPL veya
"LIPD"
endotel Lipoprotein lipaz,VLDL tarafından taşınan trigliseritleri
hidroliz ederek hücrelerin yağ asitleri elde etmelerini
sağlar.
Hormon duyarlı lipaz LIPE hücre içi
Gastrik lipaz/lingal lipaz LIPF sindirim sıvısı
Endotel lipaz LIPG endotel
Karaciğer
İnsandaki en büyük bez olan karaciğer(1,4 kg) hafifçe ayrılmış 4 adet loptan meydana gelir. Karaciğer kandaki birçok kimyasal maddenin miktarını düzenler ve safra bazı maddeleri safra ile atar. Safra ayrıca yağların sindirim ve emilimine yardımcı olur. Mide ve bağırsakları geşen tüm kan, karaciğer geçerek dolaşıma katılır. Karaciğer sindirim sisteminden gelen bu kanı işler ve dengeler ve vücut kullanımı için uygun hale getirir. Karaciğer aynı zamanda, ilaç gibi yapancı maddelerin metabolize edildiği en önemli organdır. Kısaca Karaciğer yaşam için temel organlardan biridir. Sindirim kanalından emilen besinlerin işlendiği ve diğer vücut kısımlarının yararlanması için bazılarının depolandığı bazılarının ise hemen dolaşıma verildiği bir organdır. Hem ekzokrin hemde endokrin çalışan bir bezdir ve 500’ün üzerinde farklı kimyasal reaksiyon gerçekleştir. Kan pıhtılaşma faktörleri dahil olmak üzere proteinlerini yapar, vitaminleri, şeker, yağlar ve başka besin maddeleri depolar, hormonlarını düzenler, yağ sindirerek için gerekli safrayı sağlar.

H A Y V A N F İ Z Y O L O J İ S İ
327
Karaciğer işlevleri fonkisyonları: • Detoksifikasyon karaciğer bulaşıcı organizmalar, alkol, ağır metaller, ilaçlar, kimyasallar,
zehirli yan ürünler ve kandan diğer zehirler detoxifiye eder. Bu fonksiyonu olmayan, insan vücudu kirliliği baş edemez.
• Sindirim - Karaciğer safra, sindirimi ve yağların absorbe etmek için gereken maddeleri üretir. Vücudu yardımcı sindirim maddesi safra yardımı ile yağ sindirimi , ek olarak Vitamin A, D, E ve K olmak bazı vitaminler, emilir.
• Sentez - Karaciğer enzimleri, hormonlar, kan proteinleri, pıhtılaşma faktörleri ve bağışıklık faktörleri gibi önemli proteinleri üretmektedir. Enerji ve hormonlar metabolizmasında önemli ve büyümeyi düzenleyen bir yapı taşı olan kolesterolü üretir.
• İşleme - Vucudumuza ağız yoluyla ve/veya nefes veya deri yoluyla giren hemen hemen her şey karaçiğer tarafında kontrol edilir. Vücudun biyokimyasal fabrika olarak da anılan, karaciğer, kan dolaşımındaki tüm maddeleri metabolize eder.
• Depolama - Karaciğer hücreleri vucudun diğer hücreleri için gerekli olan, demir, bazı vitaminler, mineraller ve glikojen gibi birçok önemli maddeyi depo ader.
Karaciğerin temel fonksiyonları: � Safra tuzlarıyla yağların emülsifye edilmesi � Bilirubin üretilmesi � Biyo moleküllerin depolanması � Gilikojen, yağ, vitaminlerin, bakır ve demir depolanması � Biyo moleküllerin bir birine dönüştürülmesi � Detoksifikasyon � Amonyumun üre haline getirilmesi � Fagositoz yapan Kupffer hücreleriyle bakteri ve yaşlı eritrositlerin yok edilmesi � Kanda bulunan bir çok proteinin sentez edilmesi Albuminler, fibrinogen,
globulinler, heparin, pıhtılaşma faktörleri(faktör I-XIII)
ŞEKİL 5.30 Karaciğerin yeri ve karaciğer lopüleri
Karaciğerin Karbonhidrat Metabolizması :Glikojenin depo edilmesiyle karaciğer glikozun fazlasını kandan alıp depo eder ve glikoz kan seviyesi düşmeye başladığı zaman da tekrar kana verir. Buna karaciğerin glikoz tamponlaına fonksiyonu adı verilir. Büyük miktarda karbonhidrat içeren bir yemekten sonra, karaciğeri çalışmayan kişide kan şekeri konsantrasyonu normal olana göre üç kat artış gösterir. Glikoneojenez de, kanda glikozun normal düzeyde kalmasına yardımcı olur. Glikoz konsantrasyonu normalin altına düşmeye

H A Y V A N F İ Z Y O L O J İ S İ
328
başladığı zaman önemli miktarda glikoneojenez gerçekleşir. Bu durumda büyük miktarda amino asidin glikoza çevrilmesi de kandaki glikoz konsantrasyonunun normale döndürülmesine katkıda bulunur
Karaciğerin Yağ Metabolizması : Enerji elde etmek üzere nötral yağlar ilk olarak gliserol ve yağ asitlerine ayrılır. Daha sonra yağ asitleri beta oksidasyonla iki karbonlu asetil köklerine ayrılır. Bunlar da asetilkoenzim A (asetil CoA)’yı oluştururlar. Asetil koenzim A, sitrik asit siklusuna girerek okside olur ve büyük miktarda enerji sağlar. Beta oksidasyon vücuttaki bütün hücrelerde yapılırsa da karaciğer hücrelerinde bu olay özellikle hızlıdır. Karaciğer oluşan asetil-CoA'nın hepsini kullanamaz. İki molekül asetil CoA’nm birleşmesiyle oluşan asetoasetik asit çok kolay erir ve karaciğer hücrelerinden ekstraseliiler sıvılara geçip, bütün vücuda taşınarak dokular tarafından absorbe edilir. Dokular da asetoasetik asidi tekrar asetil-CoA’ya çevirerek normal yoldan okside ederler. Bu nedenlerle, karaciğer yağ metabolizmasından büyük ölçüde sorumludur.
� Karaciğerin Protein Metabolizması: İmmunglobulinler ve erişkin hemoglobini hariç hemen hemen bütün proteinler karaciğerde sentezlenir. Karaciğerde sentezlenen en önemli protein albumindir. Karaciğer aynı zamanda koagulasyon proteinlerininde (Faktör VIII hariç, bütün pıhtılaşma faktörleri ) üretim yeridir. Amino asitlerin, enerji için kullanılmadan ya da karbonhidrat veya yağlara çevrilmeden önce deaminasyonu gerekir. Nonprotein azot metabolizmasının büyük bir kısmı karaciğerde meydana gelir. Üre karaciğerde sentez edilir. Çünkü arjinaz enzimi yalnızca karaciğerde mevcuttur. Yıkılan proteinlerden açığa çıkan aminoasitlerin çoğu tekrar protein yapımında kullanılmasına karşın total vücut protein nitrojeninin %0,3-0,4 civarı üreye çevrilerek atılır . Vücutta öteki dokularda, özellikle böbreklerde az miktarda deaminasyon olursa da, ekstrahepatik deaminasyon karaciğerdekine kıyasla çok önemsizdir. Karaciğer, üre oluşumuyla vücut sıvılarından amonyağı uzaklaştırır. Deaminasyon işlemlerinin ürünü olan büyük miktardaki amonyağa, barsaklarda bakterilerle sürekli olarak yapılıp kana absorbe edilen amonyak da katılır. Bu nedenle karaciğerin üre yapımı ile ilgili fonksiyonu kaybolduğunda, plazma amonyak konsantrasyonu hızla yükselir ve hepatik koma ile ölüm görülür. Gerçekten de, karaciğer kan akımı çok azaldığı zaman bile seyrek olarak, portal venle vana kava arasındaki şantlarda görülür- çok miktarda amonyak kanda birikerek toksik bir durum yaratır.
1- Diğer vücut fonksiyonları için enerji sağlayacak yağ asitlerinin büyük bir hızla oksidasyonu,
2- Lipo-proteinlerin çoğunun oluşumu, 3- Büyük miktarda kolesterol ve fosfolipid sentezi, 4- Büyük miktarlarda karbonhidrat ve proteinin yağa dönüşümü.
1- Glikojen depolama, 2- Galaktoz ve fruktozu glikoza çevirme, 3- Glikoneojenez, 4- Karbonhidrat metabolizmasının ara ürünlerinden bir çok önemli kimyasal maddelerin oluşturulması.

H A Y V A N F İ Z Y O L O J İ S İ
329
� Karaciğerin en önemli işlevlerinden biri de, bazı amino asitlerin sentezini yapması ve amino asitlerinden önemli kimyasal bileşikleri oluşturmasıdır. Örneğin, esansiyel olmayan amino asitlerin hepsi karaciğerde sentez edilebilir. Bu amaçla ilk olarak, yapılacak amino asitle aynı bileşimde keto asit (keto oksijen dışında) sentez edilir. Daha sonra amino kökü, uygun amino asitlerden bir çok traıısaminasyon aşamalarından sonra transfer edilerek keto oksijen grubunun yerine yerleştirilir.
ŞEKİL 5.31 Karaciğerde diğer dokulardan gelen azotlu atıkların üre olarak atılması
Karaciğerin Diğer Metabolik Fonksiyonları :
� Vitaminlerin Depo Edilmesi: Karaciğerin vitaminleri depo etme özelliği vardır. Hastaları tedavi etmede karaciğerin iyi bir vitamin kaynağı olduğu uzun süreden beri bilinmektedir. Karaciğerde en fazla depo edilen A vitaminidir. Karaciğerin Demiri Ferritin Şeklinde Depolaması. Vücutta, kandaki hemoglobinde bulunan demir dışında, demirin en büyük bölümü normalde karaciğerde ferritin şeklinde depo edilir. Karaciğer hücrelerinde, demirle az ya da çok miktarlarda birleşebilen bir protein olan apoferitin bol miktarlarda bulunur. Böylece, vücut sıvılarında demir miktarı arttığı zaman, apofertinle birleşerek ferritini oluşturur ve gerektiğinde başka bir yerde kullanılmak üzere hepatik hücrelerde saklanır. Dolaşımdaki vücut sıvılarında demir düşük bir düzeye indiğinde ferritin
1- Vitaminlerin Depo Edilmesi 2- Kan Pıhtılaşması ile Karaciğerin İlişkisi 3- Demir Depolanması 4- İlaçların, Hormonların ve Öteki Maddelerin Karaciğer Tarafından Atılması 5- Glikoliz 6- Pentoz fosfat yolu 7- Diğer 6 karbonlu şekerlerin glukoza dönüştürülmesi 8- Glukozun diğer monosakkaritlere ve yağa dönüştürülmesi
Karaciğerin amino asit metabolizması: 1- Amino asitlerin deaminasyonu, 2- Üre oluşumu ile amonyağın vücut sıvılarından uzaklaştırılması, 3- Plazma proteinlerinin oluşumu, 4- Vücuttaki metabolik olaylar için önemli amino asitlerin ve öteki maddelerin birbirine dönüştürülmesi

H A Y V A N F İ Z Y O L O J İ S İ
330
demiri serbestletir. Böylece, karaciğerdeki apoferitin-feritin sistemi bir demir deposu görevi yaptığı gibi, kan demirinin tamponu işlevini de yürütür. Kan Pıhtılaşması ile Karaciğerin İlişkisi. Karaciğerde yapılan ve koagiilasyon işleminde kullanılan maddeler fibrinojen, protrombin, akselerator globiilin, faktör MI ve bir çok diğer önemli koagülasyon faktörleridir. Karaciğerde protrombin, faktör VII, IX ve X’un oluşumundaki metabolik olaylar K vitaminini gerektirir. K vitamini yokluğunda bu maddelerin konsantrasyonu çok düştüğünden pıhtılaşma hemen hemen tamamen ortadan kalkar(walfarin K Vitamini yerine geçerek pıhtılaşmayı engeller). İlaçların, Hormonların ve Diğer Maddelerin Karaciğer Tarafından Uzaklaştırılması. Karaciğerdeki aktif kimyasal ortamın sulfonamid, penisilin, ampisilin ve eritromisin gibi çeşitli ilaçları zehirsizleştiıerek safra ile vücuttan uzaklaştırdığı iyi bilinmektedir. Aynı şekilde iç salgı bezlerinden salgılanan östrojen, kortizol, aldosteon gibi tüm steıoid hormonlar ve tiroksin de karaciğer tarafından ya kimyasal olarak değiştirilir ya da atılır. Böylece karaciğer haıabiyetinde, çok defa bu hormonlardan birinin ya da bir çoğunun vücut sıvılarında birikmesi, hormonal sistemin aşırı faaliyetine yol açar. Ayrıca, vücuttan kalsiyum da önce karaciğerden safraya sekresyona uğrar, daha sonra barsağa geçerek feçesle uzaklaştırılır.
ŞEKİL 5.32 Karaciğerdeki farklı metabolitlerin durumu
Etanol mideden emilir. Alınan etanol kan yolu ile yıkım için karaciğere gelir. Karaciğerde hepatositlerce asetaldehite ve asetata metabolize olur. Etanol yıkımında NADH bağımlı bir enzim olan alkol dehidrogenaz hepatosit endoplazmik retikülüm da görev alır. Bilirubinin Safra İle Atılması :Biluribin çok toksik bir maddedir. Safra yoluyla atılması Karaciğer ‘in en önemli fonksiyonlarındandır. Biluribin eritrosit yıkımı soununda hemoglobin yıkımından oluşur. Eritrositler 120 günlük yaşamları sonunda doku makrofajları tarafından fagosite edilir. Hemoglobin ilk olarak globin ve heme ayrılır ve hem halkası açılarak;

H A Y V A N F İ Z Y O L O J İ S İ
331
a- Serbest demir, kanda transferrinle taşınır b- Dört pirol çekirdeği düz bir zincir yaparak safra pigmentlerini oluşturur. Ürobilinojen Oluşumu : Barsaklara geçen bilirubinin yaklaşık yarısı bakteriler tarafından suda kolay eriyen ürobilinojene çevrilir. Ürobilinojenin bir kısmı barsak mukozasından kana absorbe edilir. Bunun büyük kısmı karaciğerden tekrar barsağa çıkarılır, yüzde 5'i de böbreklerden idrara atılır. Hava ile temas eden idrarda, ürobilinojen ürobiline oksitlenir.
ŞEKİL 5.33 Ürobilinojen oluşumu
Vücuttaki diğer organlar yalnız arteriyel sistemden kan sağladığı halde, Karaciğer iki sistemden kan alır:
Kan arterlerden kapillerlere, oradan da venlere akar. Venler de bu kanı kalbe taşırlar. Bunun bir istisnasını portal venler oluşturur. Portal venler, kanı ikinci bir organa taşırlar. Hepatik portal ven, sindirim sistemindeki organlardan aldığı kanı karaciğere taşır. Bağırsak duvarındaki kapiller besinleri absorbe ederler. Besin değeri yüksek olan bu zengin kan, superior mezenterik vende toplanır. Bu ven taşıdığı kanı (splenik ven ile birleşerek) hepatikportal vene boşaltır. Hepatik portal ven aynı zamanda inferior mezenterik ven aracılığıyla bağırsakların alt kısmından dönen ve splenik ven aracılığıyla dalaktan gelen kanı alır. Aldığı bu kanı, karaciğere taşır. Kan karaciğer kılcal damarları içinde akarken, karaciğer hücreleri besin maddelerini kapillerden uzaklaştırır. Kapillerdeki kan, hepatik venlere açılır. Bunlarda v. cava inferior’a akar.
• Kupffer’in yıldız hücreleri: Mononukleer fagositik sisteme dahil, monosit orijinli hücrelerdir, ilk kez tanıtan Kupffer’in adıyla anılırlar, kuvvetli fagositoz yaparlar. Komşu endotel hücreleri ile bağlantı yapmazlar, onlardan daha koyu ve büyüktür, yıldız şekillidir, sitoplazmik uzantıları vardır. Lizozom ve diğer organeller bakımından daha zengindir, peroksidaz reaksiyonu pozitiftir (endotel hücrelerinde negatif).
• İto hücreleri: Mezenşimal orijinli olan ito hücreleri, sitoplazmalarında yağ içerirler ve vitamin A’nın metabolizmasında ve depolanmasında rol oynarlar. Sağlıklı karaciğerde bu hücreler retinoidlerin alınması, depo edilmesi ve salınması, bazı ekstrasellüler matriks proteinlerinin ve proteoglikanların sentezi ve salgılanması, büyüme
1- A.hepatika yolu ile arteriyel sistemden 2- Vena porta yolu ile sindirimden gelen portal kanı alır.

H A Y V A N F İ Z Y O L O J İ S İ
332
faktörlerinin ve sitokinlerin salgılanması ve çeşitli düzenleyici maddelere (örn. prostaglandinler, trombaksan A2) yanıt olarak sinusoid çapının düzenlenmesi gibi işlevler görürler.
Safra tuzlarının
Safra tuzları karaciğer hücreleri tarafından kolesterolden oluşur ve safra tuzlarının sekresyonu sırasında günde 1 -2 gram kadar kolesterol de safraya sekrete edilir. Günde 1–2 litre kadar safra salgılanır. Safra ADEK ve kolesterol emilimi için gereklidir. Safranın başka bir görevi zararlı maddelerin vücudtan uzaklaştırılmasıdır. Safra tuzu oluşumu ve sekresyonunda bir yan ürün olduğu düşünülmektedir. Kolesterol saf suda hemen tamamen çözünmezdir. Safradaki safra tuzları ve lesitin kolesterole fiziksel olarak bağlanarak çözünür olan ultramikroskopik miçeller oluştururlar. Kolesterol kullanımı sonunda safra asitler oluşur. Gerçekten de, safra asitlerinin sentezi, memelilerde kolesterol katabolizmasının ana yoludur. Safra asidi sentezinde bulunan çeşitli enzimler , çoğu farklı birçok hücre tipinde etkin olmasına rağmen, bu enzimlerin tümüne sahip olan karaciğer biyosentez yapan tek organdır. Safra asitlerinin sentezi aşırı kolesterol atılımı için baskın mekanizmalarından biridir. Bununla birlikte, safra asitleri şeklinde kolesterol atılım kolesterol fazlalığı besin alımı telafi etmek için yeterli değildir. Safra asidi sentezi kolesterol katabolizmasının yolu oluşturmaktadır, ancak bu bileşikler, diyet kolesterolünün, lipidlerin, yağda çözünen vitaminler gibi karaciğer ulaştırılmas gereken maddeler için aracılık yapar. Safra asitlerinin sentezi 17 ayrı enzimler gerektirir. Bu enzimler hücre içi sitoplazmasında, endoplazmik retikulum (ER), mitokondri ve peroksisomlara gibi bölmeleri dağılmış halde bulunur.
ŞEKİL 5.34 Safra tuzunun yapısı , biliverdin ve ürobilinojen sentezi
Safra hemoglobin yıkım ürünü olan Billirubin ve biliverdinin vucuttan uzaklşatırılmasını sağlar. Safra karaciğer hepatositlerince sürekli olarak salgılanır. Karaciğer tarafından salgılana safranın safra kesesinde deponır ve konsantre edilir. Safra sindirim sırasında tekrar emilierek Entrohepatik dolaşımla karaciğere geri döner.
Safra Safranın bileşenleri:
• Su • Kolesterol • Lesitin (bir fosfolipit) • Safra pigmentleri (bilirubin ve
biliverden) • Safra tuzları (sodyum glikokolat ve
sodyum taurokolat)

H A Y V A N F İ Z Y O L O J İ S İ
333
Homestasi ve Boşaltım
sistemi fizyolojisi
Canlıdaki iç dengenin nasıl korunduğu, su, elektroit, atık madde, ozmoregülasyonun nasıl yapıldığıBoşaltım sistemi boşaltım maddeleri ve organları hakkında bilgiler.
Homestazi Hücre dışı sıvı içinde bulunan iyon ve besinler hücrelerin canlı kalmasını sağlar. Tüm hücreler aynı ortam içinde yaşarlar-hücre dışı sıvıda. Bu nedenle hücre dışı sıvı vücudun iç ortamını ya da yüz yıl önce, 19. yüzyıl büyük Fransız fizyoloğu Claude Bernard’ın ifade ettiği biçimiyle milieu interieur’u oluşturur. Hücreler bu iç ortamda yeterli derişimde oksijen, glikoz, farklı iyonlar, aminoasitler, yağlar ve diğer yapıtaşları bulunduğu sürece yaşar, büyür ve özel işlevlerini yapabilirler. Fizyologlar tarafından kullanılan homeostaz terimi, iç ortamın (hücredışı sıvı), yaklaşık olarak sabit koşullarda korunması anlamına gelir. Vücuttaki tüm organ ve dokular bu sabit koşulları korumaya yardım etmek üzere bir görev üstlenmiştir. Örneğin akciğerler, hücreler tarafından kullanılan oksijeni karşılamak üzere hücre dışı sıvıya sürekli oksijen sağlarlar; böbrekler iyon derişimini sabit tutar, sindirim sistemi besinleri sağlar. Uzun süre boyunca sabit kalan bir fizyolojik değişken gibi bir şey muhtemelen yoktur. Gerçekte bazı değişkenler gün boyunca bir ortalama değer etrafında çarpıcı değişiklikler gösterse de hala “dengede” kabul edilebilir. Çünkü homeostazis çeşitli denetlemelerle kontrol edilen, durağan olmayan, dinamik bir süreçtir. Örnek olarak gün boyunca kan glukoz düzeyindeki dalgalanmaları ele alacak olursak; yemekten sonra kan glukoz düzeyi neredeyse 2 katına çıkabilir. Normal düzeyden bu kadar büyük bir yükselme ‘sabit’ olarak kabul edilemez. Önemli olan glukoz yükseldikten sonra dengeleyici mekanizmaların hızla, ters yönde aşırıya kaçmadan (normal düzeyin altına inmeden) glukoz düzeylerini normal sınırlara çekmesidir. Glukozu normal düzeylere getirilmiş birinin akciğer hastalığından kaynaklanan kanda anormal yüksek karbondioksit düzeyi gibi ölümcül olabilecek bir sorunu olabilir. Yani sonuç olarak diyebiliriz ki bir kişinin bir değişken için homeostatik olabileceğini fakat başka bir değişken için olmayabileceğini fark etmek gerekir.
Bölüm
6 Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
334
Hayatın devamı için canlının iç ortamının değişmez tutulması şarttır. Bu sıvı ortam; hücreler arası sıvı= interstisyel sıvı= iç ortam olarak tanımlanır ve hücrelerin atmosferi gibidir. Hücreler yaşamları için her türlü maddeyi bu sıvı ortamdan alır ve metabolizma sonucu oluşturdukları ürünleri de bu ortama boşaltırlar. Hücrelerin yaşamlarını sürdürebilmeleri, iç ortamın sıcaklık ve kimyasal içerik yönünden (pH= Hidrojen iyonu konsantrasyonu ve ozmatik güç gibi) sürekli olarak değişmez tutulmasına bağlıdır. İç ortamın değişmez tutulması işi “homeostasis” olarak tanımlanır. Organizmada dolaşım, solunum, boşaltım ve sindirim gibi sistemler sürekli olarak homeostasisi sağlamak için çalışırlar.
Canlılarda bulunan bütün organ, sistem ve hücreler, mükemmel bir iş birliği, uyum ve denge içerisinde görev yaparlar. Bu yapıların böyle ahenkli bir şekilde görev yapmasıyla vücutta bir iç denge meydana gelir. Homeostaz(Homeostasi), hücre dışı gerçekleşen olaylar karşısında hücrenin kendi metabolizmasını korumasıdır. Organizmanın iç dengesinin belirli koşullarda sabit tutulmasıdır. homeostazi ile vücut sıcaklığı, kandaki su, madensel tuz, karbondioksit ve üre yoğunluğu kontrol altında tutulur. Hücrelerin yaşamlarının devamlılığının saglanması için iç ortamın sabit ya da degismez kosullarda tutulması gereklidir. Metabolik faaliyetler, yaşayan her hücrede devamlı görünürler ve ilgili iç çevreyi ya da vücudun durumunun sürekliliğinin (dengesini-düzenliliğini) korunması için dikkatlice ayarlanmalıdırlar. Vücudun iç durumunun dinginliği dış çevrede koşullar değişse bile iç dengenin sürekliliği ile bağlantılı bu otomatik koruma eğilimine homeostazis denir. Yasayan organizmaların bir takım fiziksel ve kimyasal özellikleri (ısı, pH, önemli moleküllerin kimyasal konsantrasyonları gibi) her zaman sabit, degismez kalmak zorundadır. Tüm fizyolojik ve kimyasal işlemler (metabolizma) su ile gerçekleşir. O nedenle, suyun olmadığı bir ortamda yaşamdan söz edilemez. Bu nedenle suyun korunması, alınması fazlasının atılması tüm metabolik reaksiyonlar için kritik öneme sahiptir. Suyun dengesi homeostaz için en önemli faktördür.
ŞEKİL 6.1 Negatif feed back mekanziması termostat örneği ve canlıdaki algoritması
Suyun vücudumuzda farklı birçok işleve sahiptir. “Maddelerin hücre içine ve dışına taşınmasını sağlar” Hücredeki faaliyetler için gerekli olan katı maddeleri çözer ¨ Vücut ısısını düzenler. Vücut sıvılarının fizyolojik ve kimyasal işlemlerinin devamlılığını sağlar. Homeostazis, canlı yapısını belli sınırlar içerisinde ve değişmez bir şekilde tutulmaya çalışılır. Vücudumuz, hücre düzeyinde gerekli ayarlama ve değişiklikleri yaparak, yani içten ve dıştan

H A Y V A N F İ Z Y O L O J İ S İ
335
gelen çeşitli etkilere karşı gerekli tepkileri göstererek bu dengeyi korumaya çalışır. Bu denge korunduğu sürece, yani homeostazis değişmez tutulduğu sürece, hayat sorunsuz olarak devam eder. Örneğin Vücudumuzun belli bir sıcaklığı vardır. Vücudumuzu oluşturan hücrelerin normal fonksiyonlarını sürdürebilmeleri için de bu sıcaklığın korunması gerekir. Hava sıcaklığı normal olduğu sürece, bu yönden bir sorun ortaya çıkmaz. Ancak çok soğuk bir ortamda bulunduğumuz zaman, vücut sıcaklığının dengelenmesi gerekir. Bunun için yüzeyel (deri) arterler daraltılır. Böylece, buralara fazla kan gitmesi engellenerek sıcaklık kaybı önlenir. Tersine çok sıcak ortamlarda vücut sıcaklığının düşürülmesini sağlamak için yüzeyel arterler genişletilir. Bu suretle deri bölgesine daha fazla kan gönderilerek, sıcaklığın deri yoluyla dışarıya verilmesi sağlanır. Bu mekanizma da yetmezse terleme olayı devreye girer. Bununla, sıcaklığın buharlaşma yoluyla azaltılması sağlanır. Bu örnekte de görüldüğü gibi vücudumuz, içten ve dıştan kaynaklanan bu gibi etkilere karşı gerekli cevabı vererek homeostazisi korumaya çalışmaktadır.
Geribildirim
Feedback (geribildirim) bildirimde yanıt, kendini oluşturan uyaranı azaltır. Kan glukoz düzeyinin düzenlenmesi, Kan O2-CO2 düzeylerinin düzenlenmesi, Vücut ısısının düzenlenmesi, Vücut sıvılarının asitliğinin düzenlenmesi buna örnek verilebilir. Pozitif Geribildirim ise bildirimde yanıt, kendini oluşturan uyaranı artırır. Bu durumda mekanizma sürekli artış yönünde kontrol sinyali oluşturur. Doğum sırasında uterus kasılmaları bebeğin başını uterus boynuna doğru yeterince güçlü biçimde itmeye başladığında gerilen uterus boynundan başlayan uyanlar uterus gövdesindeki kaslara iletilerek bu kasların daha güçlü kasılmasını sağlar. Uterus kasları kasılır ve duvarları gerilirken, uterustan kalkan sinyaller sinirler aracılığıyla arka hipofize iletilir. Hipofiz bezi uterus kasılmalarının güçlü bir uyarıcısı olan oksitosin hormonu salgılayarak yanıt verir. Oksitosine yanıt olarak uterus daha fazla kasıldıkça uterus duvarları daha fazla gerilir ve hipofize daha fazla sinyal gönderilir, bu, daha fazla oksitosin salınmasına neden olur. Kısır halka gibi görünen bu döngü bebek doğana kadar devam eder. Doğum olayında oksitoksin salgılanması uterus kaslarının kasılmasını artırır. Doğum boyunca bu olay artarak devam eder. Doğum sonunda ise pozitif geribildirim yol açan etki (doğum) yol olmasıyla mekanizma durum sonlanır.
Kanın pıhtılaşması pozitif geribildirimin işe yarayacak biçimde kullanılmasına bir örnektir. Bir kan damarı yırtıldığı ve bir pıhtı oluşmaya başladığı zaman, pıhtılaşma faktörleri adı verilen bir dizi enzim, pıhtının kendi içinde aktive edilir. Bu enzimlerden bazıları diğerleri üzerine etki ederek pıhtının hemen yakınındaki bölgede bulunan aktifleşmemiş enzimleri aktifler ve pıhtının büyümesini sağlar. Bu süreç damardaki delik kapanıp kanama duruncaya kadar devam eder.
Pozitif geribildirimin bir başka önemli kullanım yeri sinir sinyallerinin oluşumudur. Bir sinir lifi zarının uyarılması, zarda bulunan sodyum kanallarından hücre içine bir miktar sodyumun sızmasına neden olur. Lif içine giren sodyum zarın potansiyelini değiştirir, bu da daha fazla kanalın açılmasına, zar potansiyelinin daha da fazla değişmesine ve yine daha da fazla kanalın açılmasına yol açar. Böylece hafif bir sodyum sızıntısı hücre içine çok miktarda sodyum girişi patlamasına yol açar, bu da sinir aksiyon potansiyelini oluşturur. Oluşan aksiyon potansiyeli, lifte hem hücre içinde, hem de hücre dışında elektrik akımları oluşturur ve yeni aksiyon potansiyellerini başlatır. Olay, uyarı sinir lifinin tüm sonlanmalarına ulaşıncaya kadar devam eder.

H A Y V A N F İ Z Y O L O J İ S İ
336
Bazı durumlarda da pozitif geri bildirim ölüme yol açabilir. Örneğin; bazen pıhtılaşma mekanizması kendiliğinden işlemeye başlar ve istenmeyen pıhtıların oluşumuna neden olur. Çoğu akut kalp krizi, koroner arter içinde yer alan bir aterosklerotik plak üzerinde oluşmaya başlayan pıhtının arter tamamen tıkanıncaya kadar büyümesi nedeniyle ortaya çıkar.
İkinci olumsuz örnekte; kalp atım kontrolünden verilebilir. Normalde kalp dakikada 5 litre kan pompalar. Normalde 1 litre kan kaybından sonra negatif geribildirim yoluyla kalbin pompa etkinliği normale dönerken; eğer birdenbire 2 litre kan kaybedilirse vücutta kalan kan miktarı kalbin etkin bir pompa olarak çalışmasına yetmez. Sonuçta arteryel kan basıncı düşer ve koroner damarla kalbe ulaşan kan miktarı azalır. Bu, kalp kasının zayıflamasına ve pompa etkinliğinin daha da azalmasına neden olur. Koroner damarlara ulaşan kan daha da azalır, kalp biraz daha güçten düşer. Bu süreç tekrar tekrar kendini yineler ve ölümle sonuçlanır. Burada her bir geribildirim aşamasının kalbin daha da zayıflamasına neden olur.
ŞEKİL 6.2 Pozitif feed back mekanizması
Refleks ve yerel homeostatik yanıtlar
Duyu reseptörlerinin uyarılmasıyla efektör (yanıt) organda meydana gelen istemdışı hareketlere refleks denir. Belirli bir uyarana özgü istemsizdir, önceden tasarlanmamış ve öğrenilmemiş “yapıda bulunan” yanıtlardır. Tipik bir refleks arkının 5 bileşenden (reseptör, afferent yol, entegrasyon merkezi, efferent yol, efektör organ) oluşur. Reflekslere ek olarak yerel homeostatik yanıtlar denilen başka bir biyolojik yanıtlar grubu da homeostaz için büyük öneme sahiptir. Bunlar dış veya iç çevrede bir değişiklik ile (bu bir uyarandır) başlatılır ve uyarana karşılık veren bir net etki ile hücre aktivitesinde değişmeye neden olur.

H A Y V A N F İ Z Y O L O J İ S İ
337
ŞEKİL 6.3 Pozitif feed back mekanizması
Reflekse benzer şekilde yerel yanıt da, bir uyarandan türeyen olaylar zincirinin bir sonucudur. Refleksten farklı olarak bütün seri, sadece uyaran alanında meydana gelir. Örneğin, bir dokunun hücreleri metabolik olarak çok aktif olduğunda hücreler arası sıvıya bölgesel kan damarlarını genişleten maddeler salgılar. Oluşan kan akımı artışı o bölgeye ulaştırılan yakıt ve oksijen oranını artırır. Yerel yanıtların önemi her bir vücut alanına yerel kendini düzenleme mekanizmaları sağlamasıdır
Isı dengesi
Hayvanlardaki dolaşım sisteminin en önemli rollerinden biri sıcaklık düzenlemesidir. Kanın bir ısıtıcı ya da soğutucu sıvı rolü oynaması, büyük ölçüde, söz konusu organizmanın sıcakkanlı yani yüksek bir sıcaklığı, birçok enzim aktivitesi için optimuma yakın bir değerde koruyan ya da soğukkanlı olmasına bağlıdır. Daha sonra göreceğimiz gibi, kan kimyasının ve dokulardaki gaz alış verişinin büyük kısmı bir hayvanın fizyolojisinin bu özelliğine bağlıdır. Metabolik hız hücre metabolizması, karbohidratların, yağların ve proteinlerin oksidasyonuyla açığa çıkan enerjinin bir kısmını yakalayıp ATP’deki yüksek enerjili fosfat grupları içindeki enerjiye dönüştürür. Fakat bu işlemde, enerjinin kabaca yüzde 60 kadarı yitirilir yitirilen bu enerji ise metabolizmayı termodinamik açıdan çok elverişli kılar. Bu enerjinin büyük kısmı ısı olarak açığa çıkar. Hayvanların büyük çoğunluğu ve bitkilerin tümü, bu termal enerjinin büyük kısmını hızla çevrelerine vererek kaybederler. Böyle hayvanlar soğukkanlılar olarak bilinirler. Daha kesin terim-ler poikilotermik (“değişken sıcaklığa sahip”) ve ektotermik (dışarıdan ısıtılan) dir. Hayvanın sıcalığı büyük ölçüde dış kaynaklardan geldiği için, vücut sıcaklığı çevre sıcaklığına bağlı olarak dalgalanmalar gösterir. Vücut sıcaklığı, dinlenme durumunda, çevredeki ortamla, özellikle ortam suysa, hemen hemen aynıdır.
ŞEKİL 6.4 Isı bağlı dolaşım durması ilgisi

H A Y V A N F İ Z Y O L O J İ S İ
338
Bir organizmanın metabolizması, sıcaklığa yakından bağlıdır. Aktif organizmanın toleranslı olduğu dar sıcaklık sınırlarında, metabolik hız standart koşullar altında organizmanın oksijen kullanma hızıyla ve/ya da karbondioksit üretme hızıyla ölçülür- çok düzenli bir biçimde, artan sıcaklıkla birlikte artar, sıcaklık düşünce azalır. Metabolik hız ile sıcaklık arasındaki ilişki, genellikle Q10 adı verilen bir değerle ifade edilir. Bu değer, sıcaklıktaki her 10°C’lik yükselmede, hız artışını ölçer. Eğer hız, her 10°C’lik yükselmede iki katma çıkıyorsa Q10 2, üç katına çıkıyorsa Q10 3, vb. denir. Metabolik hızın en sık aldığı Q10 değeri 2’dir. Bir hayvanın 0°C’deki metabolik hızına X diyelim. Eğer bu hayvanın metabolik hızının Q10 değeri 2’yse, bu durumda 10°C’de bu değer 2X, 20°C’de 4X, 30°C’de 8X ve 40°C’de 16X olacaktır. Sıcaklık yükseldikçe hızın daha çabuk artmasına dikkat ediniz. bu tipte bir eksponansiyel artış, sıcaklık arttıkça eğimi dikleşen bir eğri verecektir. Bekleneceği gibi, ektotermik hayvanların aktivitesi, çevrelerindeki sıcaklık değişimlerinden kesin olarak etkilenir. Sıcaklık yükseldikçe (dar sınırlar içinde) daha aktif, sıcaklık düştükçe hareketsiz ve uyuşuk olurlar. Bu yüzden bu hayvanlar, etkin olarak işgal edebilecekleri habitatlarla sınırlı kalırlar, çünkü bu habitatlardaki sıcaklıkların insafına kalmışlardır. Kuşkusuz, birçoğu, iç sıcaklıklarını kısa bir süre için de olsa çevre sıcaklığının üzerine çıkartabilmek ve böylece sınırlarını bir dereceye kadar genişletmek için, güneş altında durmak gibi davranış adaptasyonları geliştirmişlerdir. Az sayıda hayvan, memeliler ve kuşlar, metabolizmalarının egzergonik tepkimeleri sırasında üretilen sıcaklıktan yararlanabilirler. Bunun için bazı mekanizmalar geliştirmişlerdir- yağ, kıl, tüy vb. ile yalıtılmış olmak gibi. Ne var ki, bu mekanizmalar çevreye sıcaklık kaybını yavaşlatırlar. Böyle hayvanlara genellikle sıcak kanlılar adı verilir. Biyologlar homeotermik (“sabit sıcaklığa sahip”) terimini kullanırlar. Vücut içinde sıcaklık üretilen hayvanlarda, vücut sıcaklığı oldukça yüksektir genellikle çevre sıcaklığından daha yüksek ve çevre sıcaklığı büyük dalgalanmalar gösterse bile, nispeten sabittir. Buna uygun olarak da endotermlerin metabolik hızları, yüksek bir düzeyde sabit olarak tutulabilir ve bunlar çok aktif olarak kalırlar. Böylece, bunlar, çevre sıcaklığına, ektotermlerden daha az bağımlıdırlar ve çok daha çeşitli habitatları, kendi çıkarları için kullanma şansına sahiptirler.
ŞEKİL 6.5 Terleme ve titreme olaylarıyla vucud sıcaklığını sabit tutulması.
Endotermikliğin bir diğer avantajı, homeotermlerin ateşlenebilmesidir. Ateşlenme, genellikle hastalıkların istenmeyen yan etkisi olarak düşünülür ve ateşi düşürmek için aspirin gibi ilaçların alınması önerilir. Halbuki ateşin (sınırlar içinde) bir uyum tepkisi olduğuna inanmak için iyi nedenler vardır. Örneğin eğer balıklar, amfibiler ya da

H A Y V A N F İ Z Y O L O J İ S İ
339
kertenkeleler gibi soğukkanlı hayvanlara yapay bir habitatta değişik sıcaklıklar sağlanırsa, bunlar sıcaklığın 38–39°C olduğu- homeotermlerin çoğunun korumaya çalıştığı sıcaklıkla hemen hemen aynı- bölgelerde toplanma eğilimi gösterirler. Eğer daha sonra bunlar bir bakteriyel hastalıkla enfekte edilirlerse, sıcaklık tercihleri 2-4°C yükselir. 34°, 36°, 38°, 40° ve 42°’lerde tutulan enfekte ektotermlerle yapılan deneyler göstermiştir ki, canlı kalma başarısı, en yüksek sıcaklıklarda en yüksektir. Bu da, hem homeoterminin hem de ateşin yararlı olduğuna işaret eder.
ŞEKİL 6.6 Kurtlarda ters akım sistemi ayaktan sıcaklık kaybının azaltılması ve vucud sıcaklığını sabit tutulması.
Hem endotermik, hem de ektotermik hayvanlarda ve bitkilerde normal metabolik hız, vücut büyüklüğüyle ters orantılıdır; organizma ne kadar küçükse, dokunun gram başına düşen nisbimetabolik hızı o kadar yüksektir. Bunun nedeni, endotermlere bakılarak kolaylıkla anlaşılabilir: Daha küçük hayvanlar, daha büyük bir yüzey/hacim oranına sahiptirler ve bunun sonucunda da birim zamanda çevreye nisbi sıcaklık kaybı daha büyüktür. Vücut yüzeyinden çevreye hızlı sıcaklık kaybına karşın, sabit bir yüksek vücut sıcaklığını korumak için, küçük bir hayvan, besini çok yüksek bir hızda okside etmek zorundadır. Tüketilen besinin nisbi miktarı ve sindirimi, solunum, vb. hızının, azalan büyüklükle birlikte artması nedeniyle, endotermlerin büyüklüğünde bir alt sınır vardır. Yaşayan en küçük memeliler sorekslerdir ve bunların ağırlığı sadece 4 gram civarındadır. Bunlar, her gün kendi ağırlıkları kadar yemek zorundadırlar ve eğer besinsiz kalırlarsa birkaç saat içinde açlıktan ölürler. Ektodermik hayvanlar ve bitkilerde, büyüklük ve nisbi metabolik hız arasındaki ters orantıyı açıklamak daha zordur. Soğukkanlı organizmalar metabolik sıcaklıklarını çevreye verdiklerinden ve sıcaklık kaybına normalde metabolizmada bir artışla tepki göstermediklerinden, daha büyük vücut ve bununla birlikte gelen daha küçük yüzey/hacim oranı, bir şekilde sıcaklık kaybını geciktirecek ve korunan sıcaklık, metabolizmayı hızlandıracaktır. Gerçekten de bitkilerde ve ektotermlerde daha büyük gövdenin neden daha düşük nisbi metabolik hızlarla korelasyon içinde olduğu hiçbir zaman tam olarak açıklanamamıştır. Bir olası faktör, artan büyüklüğün genellikle hayvanlarda iskelette ve diğer bağ dokularda, bitkilerde ise destek fibreleri de ve olgun ksilemlerde orantısız bir artışa yol açtığıdır; bir kızılağaç, bir papatyadan ve bir timsah, bir kertenkeleden çok daha fazla destek yapısı gerektirir. Bu dokular, metabolik olarak nispeten inaktif olduklarından, organizmanın tümünün birim ağırlığı başına düşen ortalama metabolik hız, bu az aktif; fakat gerekli yapısal dokuların oranı arttıkça düşer. Bu, bir embriyonun gelişim sürecinde görülür. Hemen tamamen metabolik olarak aktif hücrelerden oluşan erken embriyo, yüksek bir nisbi metabolik hıza sahipken, geç embriyoda daha az aktif doku tiplerinin oranı artar ve böylece nisbi metabolik hız düşer.

H A Y V A N F İ Z Y O L O J İ S İ
340
ŞEKİL 6.7 Isı denegesinin homotermlerde korunması
Isıtma Yeryüzünün birçok bölgesinde çevre sıcaklığı 39°C’den düşüktür. Bu, endotermlerin çoğunun kendilerini aktif olarak ısıtmak zorunda oldukları anlamına gelir. Eğer çevre sıcaklığı çok düşük değilse ve hayvan iyi yalıtılmışsa, dolaşım sisteminin sıcaklığı, üretim bölgelerinden vücudun diğer bölgelerine verimli olarak dağıtması koşuluyla, hayvanın kendi metabolizmasıyla üretilen sıcaklık fazlası, bu iş için yeterli olabilir. Eğer bazal sıcaklık üretimi yetersizse, birçok hayvan titreyerek tepki gösterir. Bu istemsiz izotonik egzersizin sonucu, kaslardan geçen kılcallardaki kanın ısıtılmasıdır; bu sıcaklık, dolaşım sistemi tarafından vücudun geri kalan kısmına taşınır. Aralarında sorekslerin, yarasaların ve sinek kuşlarının da bulunduğu bazı küçük hayvanlar, homoitermi lüksünü tam olarak kaldıramazlar ve büyük yüzey/hacim oranı nedeniyle, yüksek bir ısınma faturası ile karşı karşıya gelirler. Bunun yerine, bu hayvanlar, vücut sıcaklığının beslenme devreleri arasında azaldığı bir çeşit temporal hemoitermiyi tercih ederler. Hibernatörler, aynı şeyi, saatler yerine haftalar ölçeğinde yaparlar. Her iki durumda da sorun, hayvanın kendisini bu düşük sıcaklık uyuşukluğundan çıkarmasındadır. Bir ortak strateji, kılcallarca zengin bir merkezi ısıtıcıya sahip olmaktır. Örneğin yarasalar, “kahvengi yağ”, çok sayıda mitokondri (tipik rengin oluşmasına katkıda bulunur) ve gerekli oksidatif enzimler içeren yağ hücreleri depolarını kullanırlar. (Normal yağ hücreleri, kendi yağlarım metabolize etmezler. Bunun yerine, gerek duyulduğunda bunları dolaşıma salgılarlar ve hedef hücreler, bu yağları alıp kendileri için kullanırlar.) Uyuşuk durumdaki hayvan, aktif olma gereği duyunca, yağ oksidasyonuna başlar. Bu işlem, kanın ısıtılmasıyla sonuçlanır. Sıcaklığın korunması Çevre sıcaklığı, bir hayvanın vücut sıcaklığının çok altındaysa bir miktar sıcaklık kaybı kaçınılmaz olur. Yalıtım, – post, tüyler ve yağ- sıcaklık kaybını azaltabilir; fakat tamamen durduramaz. Aslında, böyle bir yalıtkan tabakanın varlığı sıcaklık fazlasının atılmasını gerektiren sıcak havalarda ya da ağır egzersiz sırasında farklı sorunlara yol açar. Sıcaklık kaybını en aza indirmede en ciddi sorun, Kuzey Kutbu’nda yaşayan hayvanların kol ve bacakları ile büyük yüzey/hacim oranına sahip diğer ekstremitelerinde yaşanır. Bu hayvanların çoğu uzaysal heterotermi gösterir. Yani, merkezdeki sıcaklık yüksek tutulurken, bacaklar ve kulaklar soğuğa bırakılır. Bu defa da ekstremiteleri canlı tutmak için, bunlarda dolaşan kauın sıcaklığını koruma sorunu ortaya çıkar. Strateji, yine, ters akım alış verişidir: Sıcak kan taşıyan arterler, soğuk kanla geri dönen venlerin yanında yer alırlar böylece arteriyel kanın sıcaklığının büyük kısmı venlere transfer olur. Soğutma Birçok hayvan, suyu buharlaştırarak kendilerini soğutur. İnsanlar için bu, terleme anlamına gelir. Kan, deriye yönlendirilerek sıcaklığı doğrudan havaya vermesi sağlanır ve bu sırada buharlaşmayla soğutma da katkı sağlar. Post ya da tüylerle yalıtılmış türlerde ise, buharlaşma çok daha bölgesel olmak zorundadır. Bu hayvanlarda sık olarak

H A Y V A N F İ Z Y O L O J İ S İ
341
kullanılan yüzey dildir. Köpeklerin hızlı hızlı soluması, buna, aşina olduğumuz bir örnek oluşturur. Şaşırtıcı görünebilir; fakat hızlı soluyan bir köpek havayı ağzı yoluyla değil, burun yoluyla alır. Solunum dokusu, alınan havaya su buharı vererek bunu ısıtır ve nemlendirir. Fakat dışarı verilen hava, burun yoluyla verilip sıcaklığın ve nemin büyük kısmı korunması gerekirken, ıslak bir yüzey olan dil üzerinden geçirilerek, ağız kenarlarındaki havayı da birlikte taşıyarak daha da fazla su buharlaştırılır. Sonuçta hem burunda hem de dilde bulunan kılcallardaki kan soğutulmuş olur. Birçok türde özellikle sıcak iklimlere uyum yapmış olanlarda, bu soğutulmuş kan, doğrudan vücuda döndürülmez. Bunun yerine, bir başka kılcal grubu olan carotid rete’yc (rete bir ağdır) akar. Bu kılcal ağı, beyine kan taşıyan arteriyel kılcallarla bir aradadır ve bu, kanı, ters akım alış verişi yoluyla soğutur”. Bu yolla, sıcaktan en kolay zarar gören organ olan beyin, tercihli olarak soğutulmuş olur.
ŞEKİL 6.8 Beyindeki sıcaklığın dengelenmesi için yapılan adaptasyonlar.
Isı dengesi korunmasında canlının vucud büyüklüğü ile yüzeyi arasındaki ilişki önemlidir. Hacimlerine göre daha geniş yüzey alanına sahip hayvanlar, daha az yüzey alanına sahip olanlara göre daha hızlı ısı kaybederler. Canlı vücudu sındığında ter bezleri ter atmak üzere uyarılırlar(insanda ve atta deri köpekte dil bu görevi yapar). Sıvı formda olan ter gaz haline dönüşür buharlaşır. Bunu yapmak için ısıya ihtiyaç duyar. Bu ısıyı da deri yüzeyinden sağlar. Bu durumda deri ısı kaybeder ve soğur. Sıcaklık kontrolüde vazodilasyon (damar genişlemesi) önemlidir. Kan ısı enerjisinin çoğunu taşımaktadır. Deri altında bol kapiller damar vardır. Vücut ısı artığında bu kapillere daha fazla kanla gider. Bu da kanın yüzeye daha yakın olması ve daha fazla ısı kaybetmesi anlamına gelir. Deri altındaki kapilerler kasılır ve daralır. Bu da kanın deri yüzeyine ulaşmasına engel olur ve ısı kaybı önlenir. Piloereksiyon (tüylerin diken diken olması) Tüyler deriye yakın alanda yüzeysel bir hava kitlesi oluşturur. Bu hava kitlesi vücut ısısı ile ısıtılır. Bu hava yalıtıcı bir katman oluşturur.
ŞEKİL 6.9 Termal denge için kullanılan yollar
� Radyasyon: Çevre sıcaklığından daha yüksek sıcaklığa sahip olan vücudun ısı kaybetmesidir
� Konveksiyon: Isınan havanın yükselmesi soğuduğunda ağırlaşarak aşağıya inmesi olayına konveksiyon adı verilir
� Kondüksiyon : Temas eden yüzeyler arasındaki ısı iletimi anlamındadır.
� Evaporasyon : Buharlaşma anlamındadır

H A Y V A N F İ Z Y O L O J İ S İ
342
Hipotermi, 37 °C olan normal vücut sıcaklığının, 35 °C'nin altına düşmesi halinde meydana gelen rahatsızlıktır. Genellikle yağmur, rüzgâr, kar veya soğuk suya maruz kalma gibi faktörler tarafından tetiklenir. İnsan vücudu bulunduğu ortamla ısı alışverişine girer ve bu alışverişten ortamın ısısına göre etkilenir. Ortam sıcak ise vücut ısınır, soğuk ise üşür. Örneğin; bir insan soğuk suya çıplak olarak girdiğinde vücudu suya hızla ısısını verir ve vücut ısı kaybeder. En sıcak havalarda bile çok uzun süre suda kalındığında üşüme olmasının nedeni budur. Ayrıca, birçok deniz kazasında (Titanik kazası gibi) çok kişi boğulmadan ziyade vücut ısısına bağlı ölüm, yani hipotermi yüzünden ölmüştür. Vücudun bu ısı alışveriş hızı ortamın yoğunluğuna göre de değişir. Örneğin, vücut denizde normal havadakine göre daha hızlı ısı kaybeder, çünkü deniz daha yoğundur. Bir insan cıva dolu bir havuza düşse 5-10 dakika sonra hipotermiden ölür, çünkü cıva çok yoğundur. Bir insanın ısı kaybı başladığında vücudun standart ısı seviyesi düşer. 32-35 derecede bilinç kaybı, daha aşağı düşüşlerde ölüm gerçekleşir.
ŞEKİL 6.10 Ters akım mekanizmasıyla sıcaklık kontrolü
Termoregülasyon vücut sıcaklığını kontrol eder ve sıcaklığın ideal aralıkta kalmasını sağlar (çekirdek sıcaklık 37°C). Ciltte bulunan ve sıcak ile soğuğu algılayan reseptörlere termoreseptör denir. Termoreseptörler cilt üzerinde çeşitli yerlerde bulunurlar ve sıcaklık soğukluk durumunu omuriliğe bağlı olan duyu lifler aracılığıyla hipotalamusta bulunan termoregülasyon merkezine iletir. Hipotalamus bu bilgileri "eşik aralık sıcaklıklarla" karşılaştırır. Termoreseptörler, sıcaklık değişimlerine yanıt verirler ve farklı sıcaklık düzeylerinde farklı yanıtlar verirler.
Konfor sıcaklık aralığı; vücutta üretilen ve atılan ısının dengeli veya dengeliye yakın durumda olduğu çevre sıcaklığı aralığı olarak tanımlanabilir. Çevre sıcaklığı, konfor aralığının altına düştüğünde hayvanın yem tüketimi artar, vücut yüzeyleri ile ekstremitelerdeki kan dolaşımı yavaşlar. Çevre sıcaklığının düşük olduğu durumlarda titreme başlar. Titreme, ısı üretiminin artırılmasını sağlayan koruyucu bir mekanizmadır. Kuşlarda titreme yerine tüylerin kabartılması gözlenir.
Memelilerde özelliklede insan vücud ısında meydana gelen değişmeler belirli değerler arasında kontrol edilebilir ancak vücud ısında oluşan ani ve büyük değişmelerde yeterince hızlı

H A Y V A N F İ Z Y O L O J İ S İ
343
yanıt verilemez. Eşik aralığının 4°C'ye çıkmasıyla vücut ısısının regülasyonu için gerekli olan vazodilatasyon ve vazokonstriksiyonda gecikmeler meydana gelir. 18.5 - 26.5 0C arasındaki çevre sıcaklıklarında deriye yakın kan damarları ile ekstremitelerdeki damarlar genişler. Böylece hayvanın vücut yüzeylerinin sıcaklığı artar ve su tüketimi ile solunumda artış görülür. Çevre sıcaklığı 32 0C nin üzerine çıktığında hayvanların yemden yararlanma düzeyinde gerileme, günlük canlı ağırlık artışında azalma, üreme performanslarında düşüş ve embriyonik ölümlerde artış ortaya çıkar. Belirtilen yüksek sıcaklıklarda hayvanlar aktivitelerini azaltmak yani daha az hareket etmek suretiyle vücuttaki ısı üretimini azaltmaya çalışırlar. Gölge alanlarda yatarak güneşten ve dolayısıyla yüksek sıcaklıktan korunmaya çalışırlar. Yüksek sıcaklıklarda hayvanların su tüketimi ve idrar atımları artmaktadır.
ŞEKİL 6.11 Hipotermi ve yüksek ısıda meydana gelen adaptif durumlar
• Termoregülasyon Merkezi - Vazodilatasyon - Terleme Vücut sıcaklığındaki artışın eşik aralığının üst sınırına erişmesi durumunda vazodilatasyon (damar genişlemesi) tetiklenir ve bu durum periferdeki daha soğuk olan kanın daha hızlı bir şekilde çekirdek sıcaklık ile karışmasını sağlar ve terleme ile vücudun serinlemesini sağlayarak sıcaklığın normal değerlerde kalmasını sağlar.
• Termoregülasyon Merkezi - Vazokonstriksiyon - Titreme Vücut sıcaklığındaki düşüşün eşik aralığının alt sınırına erişmesi durumunda vazokonstriksiyon (damar büzüşmesi) tetiklenir ve bu durum periferdeki daha soğuk olan kanın daha yavaş bir şekilde çekirdek sıcaklık ile karışmasını sağlar ve titreme ile enerji üreterek sıcaklığın normal değerlerde kalmasını sağlar.
Canlıların çevreye osmotik durumuna uyumu
Hücre çevresindeki sıvılara göre su alarak şişebilir yada su kaybederek büzülebiliri Hücresel Şişme olayı hücre içi sıvı ve iyon dengesinin bozulduğunda görülür. Hidropik değişme veya vakuoler dejenerasyon olarak da adlandırılan hücresel şişme, hücrede hemen her tip hasarın ilk göstergesi ekstrasellüler suyun, hücre içine geçmesi neticesi olan hücredeki büyüme “hücresel şişme” olarak bilinir. Hücre şişmesi, reversibl bir olaydır ve hafif hasarın (zedelenmenin) işaretidir.
Su alımı için iç osmotik basıncın dış ortamdan yüksek, hücre içinin hipertonik olması gerekir. Yani toplam çözünmüş madde derişimi daha yüksek olmalıdır. Bu durumda herbir

H A Y V A N F İ Z Y O L O J İ S İ
344
maddenin difüzyon basıncı farklı olacağından su moleküllerini geçiren zardan su kendi kinetik difüzyon dengesini sağlayıncaya kadar geçiş yapar. Hipertonik hücre turgor halindedir, sitoplazma çepere yapışık durumdadır. Çünkü osmotik basınç artışı çeperin karşı yöndeki basıncı ile dengelenmiştir. Hücre içinin izotonik osmotik basınca sahip olması halinde bir kısım suyunu kaybeder ve sitoplazmanın çeperden ayrılmaya başladığı görülür. Bu duruma sınır plazmoliz adı verilir ve izotonik osmotik basıncın ölçümünde kullanılır. Hücrenin iç osmotik basıncının dış basınçtan daha düşük olduğu hipertonisite durumunda sitoplazma çeperden ayrılarak ortaya toplanmaya başlar, hücre plazmolize olur.
• İzotonik: Hücre dışı ortam ile hücre içi ortamın ozmotik basınçlarının eşit olduğu ortama izotonik ortam denir. Ozmotik basınçları eşit olduğu içim suyun bir taraftan diğer tarafa (hacim değişikliğine neden olacak şekilde) ozmozu söz konusu olmayacaktır.
ŞEKİL 6.12 İzotonik, hipertonik ve hipertonik durumlar
• Hipertonik: Madde yoğunluğu hücre içi sıvı yoğunluğundan daha az olan çözeltilere denir. Saf su ozmotik çözeltiye örnektir. Hipotonik ortam; hücre sitoplazmasına göre, içinde çözünmüş madde miktarı az, su derişimi çok olan ortamlara denir. Bir hücre kendisinden daha az yoğun bir ortama konursa bulunduğu ortamdaki sıvıyı içine alarak şişer. Bu olaya deplazmoliz denir.
• • Hipotonik: Hücre dışı ortamın ozmotik basıncı hücre içi ozmotik basınçtan
yüksek olduğu ortamlara hipertonik ortam denir.

H A Y V A N F İ Z Y O L O J İ S İ
345
Azotlu atık maddeler
Canlılarda amino asit metabolizması sonunda amonyak ve amonyum iyonu meydana gelir. Meydana gelen amonyak düşük konsantrasyonlarda bile hücre için zararlıdır. Dolaşımla hızla karaciğeri getirilen amonyum iyonu glutamik asit ile birleştirilerek glutamin haline getirilir. Sentezlenen glutamin pürin sentezinde glukoz amin sentezinde kullanılır. Dolaşımda amonyağın taşınması Amonyak dokularda çok miktarda üretilmesine rağmen kanda çok az miktarda bulunur. 1.Üre : Atık maddeler içinde miktar olarak amonyağın en önemli atılım şekli karaciğerde üre kanda taşınarak böbrekler yoluyla atılır. 2.Glutamin: Kas, beyin ve çevre dokularda üretilen amonyağın depolanma ve transport şeklidir. Özellikle beyindeki amonyağın uzaklaştırılmasında ana mekanizmadır. Bu fonksiyonu nedeniyle beyin ve plazmada en fazla bulunan aa glutamindir. Glutamin böbrekler/sinidirim deaminize edilerek amonyak atılımı sağlanır. Amonyak insanda üre halinde getirilir. Üre yüksüzdür ve toksik etki yapmaz. Üre biyolojik zar ve membrandan kolaylıkla geçer. Üre sentezi Sitoplazmada ve mitokondride yapılır. Balıklarda azotlu atık amonyak halinde bol suyla birlikte solungaçlardan atılır. Sürüngen ve kuş gibi sınırlı suyun olduğu karasal canlılarda yüksek enerji maliyeti olmasına rahmen ürik asit halinde azot atık madde atımı yapılır. Ürik asidin diğer bir avantajı yumurta içinde büyümekte olan embriyonun yumurta içinde yoğunluğun değiştirmemesidir (Ürik asit yumurta içinde çözünmeyen kristaller halinde depolanır). Omurgalılarda boşaltım ürinlerinin miktarı ve tipi çeşitlik gösterir. Balıklarda deniz ve tatlı su olmasına bağlı olarak osmoregülasyon çeşitlilik gösterir. Hagfish glomerul tubuler yapı taşımaz iki yüklü iyonları atılımı yapar (Ca2+, Mg2+ , and SO4
2-) Bu ilkel balıkta osmoregülasyon düşük seviyededir. Hagfish te izoosmatik doku sıvısı olduğun regülasyon kolaylıkla yapılır. Tatlı Su balıklarında büyük glomeruslar dilue bol miktarda idrar oluşturur. Buna karşın deniz balıklarında az miktarda idrar oluşturan küçük glomeruslara sahiptir ve NH3 solungaçlarla atılır. Ampibialarda ve Reptillerde ise henle kulbu yoktur ve konsantre idrar yapamazlar buna karşın azotlu atık maddelere ürik asit halinde atılır. Memeli ve kuşlarda idrarı konsantre edebilir. Üre sentezi
Amino gruplarının en önemli kaynağı diyetle alınan proteinlerdir. Birçok aminoasid karaciğerde metabolize edilir. Bu yolla elde edilen amonyağın bir kısmı tekrar bazı biyosentetik reaksiyonlarda kullanılır. Fazla miktarı ise atılmak üzere üreye çevrilir ve böbrekler aracılığı ile direkt olarak atılır. Ekstrahepatik dokularda üretilen amonyak da karaciğere gelerek karaciğerde metabolize edilir. Azot metabolizmasında glutamat ve glutamin aminoasidleri merkezi bir rol oynar Azotlu Atık maddelerin temel kaynağı amino asitlerin yıkımıdır. Yıkılan amino asitlerin uzaklaştırılması iki basamaktan yapılır.
1-Transaminasyon: Amino gruplarının glutamatta toplanması Tüm aminotransferazların koenzimi Vitamin B6(priridoksal fosfat)’dir
a-Alanin aminotransferaz (ALT): Karaciğer, kalp, kas dokusu, böbrek ve beyinde bulunur. Bu dokulardan herhangi birinde oluşan hasarda kandaki AST düzeyi artmaktadır. Yani hem kas hastalıklarında hemde kalp krizinde bu enzimin kandaki düzeyi artmaktadır.

H A Y V A N F İ Z Y O L O J İ S İ
346
b-Aspartat amninotransferaz (AST): Serum Glutamik Oksaloasetik Transaminaz (SGOT) da denir: Karaciğer hücrelerinin içinde yer alan ve protein üretiminde kullanılan bir enzimdir. Karaciğer hücrelerinin parçalanmasıyla kana karışır ve miktarı yükselir. AST sadece karaciğerde değil kalp ve iskelet kaslarında da bulunur. AST yüksekliği karaciğer, kas veya kalp problemi olduğunu gösterir.
2-Oksidatif deaminasyon:Amino grupları transfer edilen transanimasyon reaksiyonlarının tersine oksidatif deaminasyon amino grupları serbest amonyak şeklinde açığa çıkar. Bu reaksiyon glutamat dehidrogenaz ile karaçiğerde yapılarak glutamat ile taşınan atık amino grupları üreye dönüştürülmesi yapılır.
Üre döngüsü karaciğerde gerçekleşir ve toksik bir madde olan amonyağın daha az toksik bir madde olan üreye çevrilmesini sağlar. Daha sonra üre kana verilerek böbreğe gönderilir, ordan da idrar ile vücuttan atılır. Üre döngüsü Mitokondri ve sitoplazma evresi olmak üzere iki evreden oluşur. Karaciğer hücrelerinin mitokondri matrikslerinde gerçekleşen bu evrede ilk olarak oksidatif deaminasyon sonucu oluşmuş olan amonyak, aktif karbondioksit ve 2 adet ATP molekülüne de alarak Karbomoil Fosfat Sentaz I enziminin yardımıyla Karbomoil Fosfat oluşturur. Burada dikkat edilecek nokta, bu enzim N-Asetil Glutamat olmadan işlev görememektedir. Daha sonra oluşan Karbomoil Fosfat, protein sentezinde kullanılmayan bir aminoasit olan Ornitin ile Ornitin Transkarbomoilaz enzimiyle reaksiyona girerek protein sentezine girmeyen diğer bir aminoasit olan Sitrülin'e çevrilir. Sitrülin ise sitoplazmaya taşınır. Ve bu noktada mitokondri evresi sonlanır.
ŞEKİL 6.13 Üre sentezi ve yapıldığı mitokondri ve sitolsol
Üre sentezine ilk giren amino grubu, karaciğer hücrelerinin mitokondrilerinde bulunan Glutamat Dehidrogenaz enziminin etkisiyle glutamatın oksidatif deaminasyonu ile ortaya çıkan serbest amonyaktan gelmektedir. Reaksiyon NAD+ gerektirmektedir. Bu reaksiyon sonucu ortaya çıkan amonyum iyonu (NH4
+), mitokondride solunum sonucu meydana gelen CO2 ve matrikste ATP bağımlı Karbamoil Fosfat Sentetaz 1 enziminin etkisiyle karbamoil fosfat meydana gelmektedir.
1-Karbamoil fosfat sentezi ve ornitin sitrülline dönüşümü (mitokondri) Bir glutamat türevi olan N-asetil glutamat, bu Karbamoil fosfat sentetazın allosterik bir aktivatörüdür. Karbamoil fosfat, ornitin karbamoil transferazın etkisi altında sitrüllini

H A Y V A N F İ Z Y O L O J İ S İ
347
vermek üzere ornitinin α aminli grubu ile etkileşir. Karbamoil fosfat karbon atomunu ve oluşacak üre molekülünün azot atomlarından birisini taşır.
2-Sitrüllinden arginine dönüşüm (sitozolde) Bu dönüşüm iki reaksiyon gerektirir. Birincide sitrüllin ATP varlığında Arjinosüksinik Asidi oluşturmak üzere aspartik asit ile yoğunlaşır. Arjinosüksinik asit, ikincide Arjinine Ve Fumarik Asite ayrılır. Aspartik asit oluşacak üre molekülünün ikinic atomunu taşır ve aspartik asit, fumarik asite dönüşmüştür. NH2 vericisi zorunlu aspartik asittir. Aspartik asitin aminli grubu glutamik asitten gelebildiği gibi (oksaloasetik asit üzerine transaminasyonla) ve glutamik asitin aminli grubunun kendiside, glutamik asitin çok aktif biçimde transaminasyon proseslerine katılmasından dolayı bir çok aminoasitten gelebilir.
3-Arjininin ornitine ve üreye dönüşümü Bu reaksiyon bir hidroliz reaksiyonudur ve Arjinaz tarafından katalizlenir. Karaciğer enzimi olan arjinaz, arjinini koparır ve üre ile ornitin meydana gelir. Burada meydana gelen ornitinin tekrar mitokondriye girmesi ile üre döngüsünün birinci turu tamamlanmış olur. Ornitin iknici bir tur başlatır. Memelilerin vücudunda protein maddelerinin yakılması sonucu meydana gelen amonyak, karaciğerde karbondioksitle üreye dönüşür. Kana geçen üre, idrarla dışarıya atılır. Üre ayrıca az miktarda ter, süt ve gözyaşında da bulunur. Yetişkin bir insan günde 25–30 gram üreyi idrarla atar. İnsan kanındaki üre miktarı normalde % 50 mg civarındadır. % 50 mg'ın üstü anormaldir. Fakat vücut yaşlandıkça, böbreklerin üreyi vücuttan atma kabiliyeti de her geçen yıl bir parça daha azalacaktır. 40 yaşından itibaren, her yıl böbreklerin süzme kabiliyeti % 1 oranında azalmaktadır. Bu yüzden 75–80 yaşındaki bir kişide kandaki üre miktarının % 65–75 mg bulunmasını normal olarak kabul etmek gerekir. Kandaki üre miktarının beklenen normal değerin üzerinde olması haline üremi adı verilir. Başlıca Amonyak kaynakları: 1.Amino asitlerin transaminasyon ve oksidatif deaminasyonu Kaslardan karaciğer'e en sık amonyum taşıyan amino asit alanin'dir 2.Glutamin Böbreklerde renal glutaminaz'ın etkisiyle amonyağa dönüşür ve NH4 olarak idrarla atılır (asit dengesinde önemlidir) intestinal glutaminaz'ın etkisiyle amonyağa dönüşür. 3.Bağırsaklarda bakterilerin etkisi Bağırsak lümenindeki üre bakterilerin etkisiyle amonyağa çevrilir. Oluşan amonyak emilerek portal ven aracılığıyla karaciğere gelir ve tekrar metabolize edilerek üreye çevrilir. 4.Aminler Amin oksidaz'ın etkisiyle amonyağa dönüşür. 5.Pürin ve pirimidinler Pürin pirimidin katabolizmasında halkalardaki amino grupları amonyak olarak salınırlar.
Ürik asit
Reptilia dan itibaren yumurtlayan hayvanların çoğunda, protein metabolizma artığı ürik aside çevrilerek vücuttan atılır. Bunun nedeni yumurtayı metabolizma artığı üre’nin osmotik basıncı arttırıcı etkisinden korunmak amacıyla, suda erimeyen forma ürik aside çevirmesinden dolayıdır. Yumurtlamayan yavrusunu doğuran hayvanlarda protein metabolizma artıkları üre haline çevrilir ve idrarla çıkarılır.

H A Y V A N F İ Z Y O L O J İ S İ
348
ŞEKİL 6.14 Farklı atık maddeler
Boşaltım organları
Yassı solucanlarda (planaria’da) borumsu boşaltım sistemi gelişmiştir. Dalbudak salmış ve vücudun uzun ekseni boyunca seyreden borular sistemi, birçok deliklerle vücudun dış yüzeyine açılırlar. Nefrium Annelidlerde görülür. Alev hücrelerinin aksine iki ucu açık tübüllerdir. İçteki uç silli bir huni (nefrostom) şeklindedir ve sölomla ilişkilidir. Diğer uç ise nefridiyopor denilen bir açıklıkla vücut dışına açılır. Her tübül etrafında zengin bir kapiler ağ bulunur. Böceklerde boşaltım organları malpighi tübülleridir. Sayıları 100-200 arasında değişir. Bu tübüllerin proksimal uçları sindirim kanalıyla diğer uçları ise hemolenf ile temastadır. Her tübül kaslı bir duvara sahiptir. Bu kasların peristaltik hareketleriyle artık maddeler (idrar) tübül lümeninden barsağa geçer. Hemolenfteki artık maddeler tübül hücreleri tarafından difüzyon ya da aktif transport ile alınırlar. Malpighi tübülleri sindirim kanalının çıkıntılarıdır ve açık kan sinusleri içinde hemolenf ile (kan ile) doğrudan temas halindedir.

H A Y V A N F İ Z Y O L O J İ S İ
349
ŞEKİL 6.15 Farklı canlılardaki boşaltım yapılarıı
Balıklarda Osmoregülasyon
Hagfish balıklarında izosmatik bir ortamda olamalarına rahmen glomerüler filtrasyon yapılır. Tüpüler Ekstrasyon (Ca2+, Mg2+ , SO4
2- ) atımı yoktur. Düşük seviyede osmoregülasyon vardır. Tatlısu Teleostlarında hiperosmotik bir ortamda yaşamaları nedeniyle büyük glomerüslarla, Dilue çok miktarda idrar atılması yapılır. Azotlu atıklar tatlısu balıklarında solungaçlarla atılır. Deniz Teleostları ise hiperosmotik bir ortamda yaşadıklarından az miktarda idrar oluşur, azotlu atıkların NH3 atılması solungaçlardan yapılır. Bol idrar oluşturan Tatlısu teleostlar büyük böbreklere ve bol glomeruslara sahipken deniz formları az idrar oluşturan küçük böbreklere ve az sayıda glomeruslara sahiptir.

H A Y V A N F İ Z Y O L O J İ S İ
350
ŞEKİL 6.16 Tatlısu ve deniz balıkların osmo regülasyon
Tatlı suda yaşayan bütün omurgalı ve omurgasız hayvanların vücut sıvıları, deniz suyundan çok daha düşük, fakat tatlı sudan daha yüksek osmotik basınca sahiptirler. Tatlı su kemikli balıklarda solungaçlarda mevcut özel hücreler, sudan gerekli tuz iyonlarını alarak kana aktarırlar. Su solungaçlardan vücut sıvılarına kana girer.
ŞEKİL 6.17 Balık, reptil ve memeli glomerus yapılarının karşılaştırılması ve balılardaki klor hücreleri ile aktif klor ve sodyum atımı.
Kana giriş yapan bu suyun vücuttan atılması böbreklerden çok sulu (vücut sıvılarına kıyasla hypoosmotik) idrar çıkarılması ile sağlanır. Denizde yaşayan kemikli balıkların vücut sıvıları, deniz suyuna kıyasla biraz hipotonik’tir. Bu nedenle, suda yaşamalarına rağmen, vücutlarından su kaybetme ve vücutlarına fazla tuz girmesi tehlikesi ile karşı karşıyadırlar.
ŞEKİL 6.18 Tatlısu ve deniz balıkların osmo regülasyon

H A Y V A N F İ Z Y O L O J İ S İ
351
Köpek balığı vatoz gibi kıkırdaklı balıklarda kanın yoğunluğu azotlu organik bir atık madde olan Trimetil amino oksit(TMAO) nedeniyle deniz suyuna göre daha yoğundur(Hiperosmotik). Bu nedenle hipertonik olan balık kanına ağız, sindirim, solungaçlarla deniz suyu girişi olur. Vatoz ve köpek balığı kemikli balıklardan farklı olarak deniz suyu içmez. Kıkırdaklı balıkların böbrekler temel fonksiyonu vücuda giren fazla suyun ve iyonların atılmasıdır.
ŞEKİL 6.19 Niçin deniz suyu içemeyiz.
Amphibian and reptillerde: Henle kıvrımı olmadığı için konsantre edilmiş idrar üretimi olmaz. Buna karşın boşaltım azotlu son ürün ürik asit olduğu için idrarla su atımı son derece azdır. Memeli ve kuşlarda: Konsantre edilmiş idrar üretilir. Justaglomeruler tip glomeruller hiper osmotik idrar oluşumu sağlar. Ortalama olarak, deniz suyunun tuzluluk değeri yaklaşık% 3,5 (35 g / L veya 599 mM ). Bu her kilogramı için deniz suyu (hacim olarak yaklaşık bir litre) yaklaşık 35 gram tuzları (özellikle sodyum ( Na+ ) ve klorür ( CI- ) iyonu anlamını gelir. İnsanda Boşaltımla böbrekler aktif olarak ancak 9 g /L (ağırlıkça% 0.9) atım yapabilir. Bu neden deniz suyu içildiğinde ancak ekstrasellüler sıvı ile seyreltilmesinden dolayı deniz suyu su kaypına yol açar. Kuşlarda ozmoregülasyon:
Deniz kuşları uzun süre denizde kaldıklarından deniz suyu kullanmak zorundadırlar. Vücutlarına aldıkları binde 35 lik tuzun atılması için tuz atma konusunda özellişmiş bir boşaltım organı kullanırlar Gaga üstünde bulunan nostik tuz bezleri yoğun halde tuz ters akım ilkesi kullanarak atar. Bu adaptasyon sayesinde deniz kuşları deniz suyunu kullanabilir.
Balıklarda Su dengesi korunması için geliştirilmiş adaptasyonlar şunlardır: 1) Deniz balıklarında vücut, suya karşı kısmen impermeable olan deri ile örtülmüştür. 2) Deniz balıkları balıklar su kaybını önlemek için devamlı olarak su içerler. 3) Solungaçlarında mevcut özel hücreler vasıtasıyla, vücuttan fazla tuzu atarlar 4)Nitrojen taşıyan maddelerin metabolizma artıkları, amonyak halinde solungaçlardan atılır; böylece idrarla fazla su çıkarılması önlenmiştir.

H A Y V A N F İ Z Y O L O J İ S İ
352
ŞEKİL 6.20 Deniz kuşlarında tuz atılımı yapan nostrik tuz bezleri ve çalışma ilkesi .
Sıvı-Elektrolit Dengesi
Hücre dışı sıvı içinde bulunan iyon ve besinler hücrelerin canlı kalmasını sağlar. Hücreler bu iç ortamda yeterli derişimde oksijen, glikoz, farklı iyonlar, aminoasitler, yağlar ve diğer yapıtaşları bulunduğu sürece yaşar, büyür ve fonksiyonlarını yapar. Hücresel düzeyde sıvı kaydırmaları ve böbreklerin idrarla vücudun gereksinimlerine göre su, elektrolit ve solüt atımını düzenlemesi sonucu korunan bu denge mekanizmasına sıvı-elektrolit dengesi denir.
Hücre dışı ve Hücre içi Sıvılar Arasındaki Farklar
• Hücredışı sıvı çok miktarda sodyum, klor ve bikarbonat iyonuyla birlikte hücreler için gerekli oksijen, glikoz, yağ asitleri ve aminoasitler gibi besinler içerir. Ayrıca hücrelerden atılmak üzere akciğerlere taşınan CO2 ve böbrekler yoluyla atılacak diğer hücresel artıklar da hücre dışı sıvı içinde yer alır.
• Hücre içi sıvı hücre dışı sıvıdan büyük ölçüde farklıdır, hücre dışı sıvıda bulunan sodyum ve klor iyonları yerine, büyük miktarda potasyum, magnezyum ve fosfat iyonları bulunur. Hücre zarlarından iyonların geçişini düzenleyen özel mekanizmalar bu farklılığı korur.
Su, canlılarda en çok bulunan moleküler maddedir ve insan vücudunda total vücut ağırlığının yaklaşık %60’ını oluşturur. Total vücut ağırlığındaki su oranı yaş, cinsiyet, obezite gibi faktörlerle değişiklik göstermektedir. Vücutta yağ miktarı arttıkça su oranı azalmaktadır: Yaşlılarda, kadınlarda ve obez kişilerde vücuttaki su oranı normalden azdır (%40). Yeni doğanlarda

H A Y V A N F İ Z Y O L O J İ S İ
353
ödemli hastalarda ise fazladır. (%80). Vücuda giren 2.2 litre su ek olarak metabolizma ile 0.3 litre su elde edilir(metabolik su). Toplam kayıp 2.5 litredir. Günde ortalama 2–3 lt su alınır. (0.3 lt’si Metabolik su) Su, barsaklardan süratle emilir ve hücre zarının geçirgenliği nedeniyle extrasellüler ve intrasellüler kompartmanlara difüzyonla dağılır.
Dokulardaki su en fazla su taşıyan doku kandır. Embriyo ilk haftalarında daha fazla su oranı vardır. Total vücut suyu (42 lt), iki büyük kompartmanda bulunur:
�
İntrasellüler sıvı
� Temel katyonu:Potasyum (K+) ve magnezyum(Mg++)’dur. � Temel anyonu: Proteinler ve Fosfat’ tır.
Diğerler iyonlar: Az miktarda Sodyum (Na++),Bikarbonat (HCO3) ve Klor(Cl¯)’dur. Kalsiyum (Ca+2 ) ise hemen hemen yok denecek kadar azdır. 1-İntravasküler sıvı(plazma):3,5 lt 2-İnterstisyel sıvı 3-Diğer sıvılar: (sindirim salgıları, idrar, ter, BOS, intraoküler sıvı vb) oluşur.
ŞEKİL 6.21 Vucud kompartamantları arası sıvı aktarımı
1- İntrasellüler sıvı (Hücre İçi Sıvı ) % 40 = 28 lt 2- Extrasellüler sıvı (Hücre Dışı sıvı) % 20 = 14 lt a) İnterstisyel sıvı (Hücreler arası sıvı) % 15 = 10.5 lt b) İntravasküler sıvı ( Plazma) % 5 = 3.5 lt _________________________________________________
Total vücut suyu % 60 = 42 lt
Günlük alınan suyun, • 0,3 lt’si Barsaklardan feçesle • 0,9lt’si Solunum sisteminde buharlaşma ile • 0,05 lt’si Deriden terlemeyle kaybedilir. • 1,5 lt’si ise Böbreklerden idrarla atılır.

H A Y V A N F İ Z Y O L O J İ S İ
354
Extrasellüler sıvı
Ekstrasellüler sıvı vücut ağırlığının yaklaşık % 20’si kadardır. • Temel katyonu: Sodyum (Na++) • Temel anyonu: Klor(Cl¯) ve Bikarbonat (HCO3)’tır.
Diğerleri: Az miktarda Kalsiyum (Ca+2 ), Potasyum (K+) ve magnezyum (Mg++)’dur. Extrasellüler sıvının osmotik basıncını büyük oranda sodyum sağlar.
ŞEKİL 6.22 Hücre içi ve dışı anyon ve katyonlar
Sıvı değişiminde etkili faktörler
1-Susuzluk Hissi: Susuzluk merkezi Hipotalamus dadır Susuzluk merkezinin çevresindeki ekstrasellüler sıvının osmolaritesinin artması, bu merkezi uyararak; Hipofizer antidiüretik sistemi etkiler ve ADH(antidiüritik hormon = vazopresin) salınımını uyarır. Uyaran faktörler:
• Osmoreseptör hücreler ile plazma arasındaki osmotik basınç farkı (Total osmotik basınç artışı ADH için uyarı oluşturmaz!)
• Emosyonel stres • Kanamalar
2-ADH (Antidiüretik Hormon) (Hipofiz arka lobu Nörohipofiz) Su değişimini etkileyen en önemli hormondur. ADH idrar hacim ve konsantrasyonunu düzenleyerek total vücut suyunun homeostazının kontrolünü sağlar. Hipotalamusta üretilerek arka hipofiz lobunda birikir ve buradan salgılanır. Böbreklerden su tutulumunu arttırır(distal ve toplar kanallarda aquariapor su kanallarının simule ederek su geri emilimi artırır). Uyaran faktörler: Korku, Ağrı, infeksiyonlar, Hipoksi…vb, Sıvı değişiminde etkili faktörler 3-Aldosteron :Böbrek üstü bezlerinden(korteks bölgesinden) salgılanır.
• Adrenal kortex’ten salınan güçlü bir mineralokortikoiddir • Böbreklerden Na ve su atılmasını azaltır, potasyum atılmasını arttırır. Buna bağlı
olarak su miktarını ve osmotik basıncı etkiler. • Uyaran faktörler: Plazmada K+ artışı , Renin-anjiotensin sistemi , ACTH
sekresyonu • Aşırı Aldosteron salınımı→Hipokalemi, kas zayıflığı • Aldosteronun azlığı→Hiperkalemi, kalp kontraksiyon zayıflığı, aritmi

H A Y V A N F İ Z Y O L O J İ S İ
355
Hücre Dışı Sıvı Hacminin Düzenlenmesi:
1- Hacim reseptörleri: Kan hacmi azaldığında kalpte ve kan damarlarında bulunan gerilim reseptörleri uyarılarak ADH salgılanmasını sağlar. Bu da idrarla sıvı itrahını azaltarak sıvı volümünü düzenler.
2- Baroreseptörler:ACİ arterinin bazı dallarında bulunan reseptörler kan basıncı düşünce ADH salgılanmasını sağlarlar. Bunun dışında kan basıncının düşmesi sempatik sistemi uyararak anjiotensin-aldesteron sistemini aktive eder. Bu da suyun ve sodyumun idrarla itrahını azaltarak plazma hacminin genişlemesini sağlar.
3- Osmoreseptörler: Beyinde bulunurlar. Osmotik basınç yükselince susama hissi sağlayarak kanda su miktarının çoğalmasına yardım eder.
Osmotik Basınç
Seçici geçirgenliğe sahip bir membrandan su moleküllerinin geçişi osmoza zıt yönde bir kuvvet uygulanarak engellenebilir. Osmozu engellemek için uygulanması gereken kuvvete osmotik basınç denir. Bir çözeltinin osmotik basıncı o sıvıdaki osmo-aktif partiküllerin yoğunluğu ile doğru orantılıdır. Örnek: 70.000 molekül ağırlıklı 1 mol albuminin osmotik etkisi, 180 molekül ağırlıklı 1 mol glikozunki ile aynıdır. Örnek: 1mol NaCl, Na ve Cl olmak üzere iki osmotik aktif partiküle sahiptir. Bu nedenle albumin veya glikoz molekülünün 2 katı osmotik etkiye sahiptir.
ŞEKİL 6.23 Osmotik basınç
Vücut sıvıları gibi seyreltik sıvılarda 1 kg ile 1 lt arasındaki fark küçük olduğundan bu iki terim hemen hemen eş anlamlı kullanılır. Genelde vücut sıvılarında osmotik aktiviteyi ifade etmek için osmol çok büyük bir birimdir. Bu nedenle çoğunlukla ( osmol’ün 1/1000 ‘i ) mOsmol birimi kullanılır. Bir litre sıvı içinde çözünmüş olan partiküllerin toplam miktarına “Osmolalite” denir. Osmolarite fiziksel bir kavram olup bunun biyolojik karşılığı “tonisite” dir. Vücutta kompartmanlar arasında efektif osmotik basıncı mebranlardan geçemeyen maddeler sağlar. Bunlar da plazmada bulunan proteinlerdir. Bu basınca “kolloid osmotik basınc” denir.
• Normal şartlar altında; Osmolalite = (2 x Na) + 10 • Üremi veya hiperglisemi gibi patolojik durumlarda; • Osmolalite = 2 x Na + (Glikoz / 18) + (Üre / 2.8) formülleri ile hesaplanır.
Osmolarite: 1 lt sıvıda çözünmüş 1 mol katı partiküle denir. ( 1 Osm/lt )
Osmolalite: 1 kg sıvıda çözünmüş 1 mol katı partiküle denir. ( 1 Osm/kg )

H A Y V A N F İ Z Y O L O J İ S İ
356
Sodyum ve Potasyum
Sodyum Ekstrasellüler sıvının temel katyonudur. Plazma hacmini düzenlemede, Asit-baz dengesinde, Sinir ve kas fonksiyonu görev alır. Hücrede Na/K ATPaz ile düzenlenilir. Hormonla olarak Aldosteron tarafından düzenlenir. Fazlalığı hipertansiyona yol açabilir. Tuz vücutta biriktikçe iki temel nedenle ekstrasellüler sıvıyı arttırmaktadır. 1. Vücutta fazla miktarda tuz bulunduğunda vücut sıvılarının osmolalitesi artmakta ve bu da susama merkezini (hipotlamusta buluanan Lateral Hipotalamik alan) uyararak ekstrasellüler tuz konsantrasyonunu normal seviyelere çekmek için kişinin fazla miktarda su içmesine yol açmaktadır. Böylece ekstrasellüler sıvı hacmi artmaktadır.
2. Ekstrasellüler sıvı osmolalitesindeki artış hipotalamik arka hipofizer salgı mekanizmasını da uyararak ADH salınımını arttırır. ADH idrar olarak atılmadan önce böbrek tübüler sıvısından yüksek miktarlarda suyun geri emilimine neden olur; böylece idrar miktarı azalırken ekstrasellüler sıvı hacmi artar. İdrarla su atımının kontrolü supraoptik çekirdekler tarafından yapılır; Vücut sıvıları yoğunlaşınca (idrar, ter ve solunumla atılan su buharı sonucu oluşan su kaybı) supraoptik çekirdekler uyarılır, uyarı arka hipofize gider ADH (vazopresin) salgılanır. Böbrek toplayıcı kanallarından su geri emilimi artar, idrarla su atılımı azalırken elektrolit atılımı devam ettiğinden vücut sıvılarının yoğunluğu normale döner. Asit-Baz dengesi
Hidrojen iyonu kosantrasyonun eksi logaritması olan pH bir solüsyonun içindeki hidrojen iyonu (H+) yoğunluğunu gösterir. Normal pH hücre içi enzimlerin aktivitesinin sürdürülmesi için zorunludur(7,4 - 7,3), Hücre içi ile hücre dışı pH sürekli olarak bir denge içindededir. Bu dengenin oluşumunda hem bazı iyon pompaları, hem de hücre içindeki tamponlar rol oynar. Normalde kan H+ konsantrasyonu 40 nmol/L düzeyindedir. Bu rakamın negatif logaritması olan pH hesaplanırsa 7.40 olarak bulunur. Fizyolojik koşullarda pH 0.04-0.05’lik oynamalar gösterebilir. Yaşamın mümkün olabildiği en düşük H+ konsantrasyonu 16 nmol/L (pH=7.8), en yüksek konsantrasyon ise 160 nmol/L (pH=6.8) dir.
ŞEKİL 6.24 Asit ve baz değişimi
• Potasyum intrasellüler sıvının temel katyonudur. • Vücutta bulunan potasyumun %95’i hücreler içinde yer almaktadır. • Normal koşullarda serum K+ konsantrasyonu: 3,5-5 mEq/lt • İntrasellüler (Hücre içi) K+ konsantrasyonu: 130-180 mEq/lt’dir. • Erişkin bir birey tarafından diyet yoluyla ve günde yaklaşık 1 mEq/kg kadar
Potasyum alınır.

H A Y V A N F İ Z Y O L O J İ S İ
357
Kanda hidrojen iyonlarının konsantrasyonu sabit olup pH 7.38-7.42 arasında tutulur. Kan pH ‘ının azalması asidoz, çoğalması alkaloza işaret eder. Asit Baz Dengesini Sağlayan Mekanizmalar:
• Kanda karbonik asidin bikarbonatlara oranı daima 1/20 olarak sabit tutulur. (H2CO3 / HCO3 = 1.35/27 = 1/20). Kanın pH’ını bu sabite tayin eder. Hücreden atılan CO2 kana geçer, bu madde kanda serbest kalabildiği gibi su ile birleşip karbonik asit oluşturabilir. Bu işlem reversibldir. (H2O + CO2 ���� H2CO3) Dokularda CO2 yapımının fazla olmasına bağlı olarak kanda CO2 ‘nin çoğalması karbonik asidin artmasına neden olur. Karbonik asidin artması, akciğer fonksiyonu normal olduğu taktirde, CO2 ‘nin atılması ile kolayca tampone edilebilir. Akciğerlerden CO2’nin atılımının kısıtlanması kanda karbonik asidin çoğalmasına neden olur. Vücutta değişik patolojiler esnasında meyda çıkan fosfat, sülfat, piruvat, laktat ve ketoasitler kandaki bikarbonatlar ile birleşip “sodyum asitleri” oluşturur ve böbrekle itrah edilirler. H2CO3 fazlası solunumun artması ile tampone edilir. (Doku asitleri + Na HCO3Na ���� Asitler + H2CO3) Organizmada metabolik denge yukarıda belirtilen mekanizmalar ile korunur. Ancak, bazı patolojilerle oluşan metabolitler idrarla veya solunumla atılamayacak kadar fazla olduğunda, solunumsal asidoz ya da alkaloz; metabolik asidoz veya alkaloz olarak 4 değişik tablo olarak ortaya çıkar.
• Karbonik asitler (H2CO3):Karbonhidrat ve yağ metabolizması sonucu her gün yaklaşık 15.000 mmol CO2 açığa çıkmaktadır. CO2 kendisi bir asit olmamasına karşın H2O ile reaksiyona girerek H2CO3’e dönüşmektedir.
• Nonkarbonik asitler: Protein metabolizması sonucu oluşmaktadır. Ana kaynak sülfür içeren sistin ve metiyonin gibi amino asitlerin yıkımı ile ortaya çıkan sülfirik asittir (H2SO4).
İnsanda Azotlu atıklar(Üre, Ürik asit, Kreatin)
Üre proteinlrin sindrimi oluşan amino asitlerin yıkımında ortaya çıkar amnonyum karaciğerde üreye dönüştürülür. Diğer atık madde olan ürik asit nükletotid katabolizması sonunda meyadana gelir. Kreatinin ise kreatinin fosfat katabolizmasında oluşur. Kandaki amonyak kaynakları:
• Amino asitler: en büyük kaynaktır. • Glutamin: böbreklerde enzimatik olarak glutaminden amonyak açığa çıkar. • Bağırsakta ürenin bakteriyel yıkımıyla amonyak oluşur. • Aminler: hormon veya nörotransmitter olan aminlerden açığa çıkar. • Pürin ve primidinler: yıkımları sonucunda halkalara bağlı olan amino grupları amonyak
olarak ayrılır .
ŞEKİL 6.25 Atık maddeler

H A Y V A N F İ Z Y O L O J İ S İ
358
Yüksek amonyak seviyesi beyne direk toksik etki gösterir. Hücredeki aktif taşıma yapan proteinleri etkiler, klor ve sodyum-potasyum pompalarını bozar. Ayrıca enerji metabolizmalarını bozar. Dokularda amino asitlerden açığa çıkan toksik amonyak uygun bir şekilde detoksifiye edileceği karaciğer taşınmalıdır. Amonyak perifer dokulardan karaciğere Glutamin ve Alanin şeklinde taşınır. Karaciğerde ise amonyak nontoksik üreye dönüştürülür.
Proteinlerin C ve H atomlarından da son ürün olarak karbondioksit (CO2) ve su (H2O) oluşmakla beraber, proteinlerde %16 oranında bulunan azot sadece proteinlere özgü olan atılım ürünlerinden amonyak ve üre oluşumuna yol açar. Protein yapısındaki C ve H den CO2 ve H2O oluşurken azot atomundan canlı organizma için toksik olan amonyak meydana gelmektedir. Amonyak ileri bir reaksiyona girerek toksik olmayan üreye dönüştürülerek vücuttan atılır. Amonyağın Taşınması
Özellikle MSS için toksik bir madde olan amonyak(Na-K pomplarını inhibe ederek) iki şekilde taşınmaktadır:
1-Amonyaktan glutamin sentez edilerek zararsız bir bileşik halinde vücutta dolaşması sağlanmaktadır. Karaciğer ve böbrekte bulunan glutamin sentetaz enzimi tarafından katalizlenen tepkimenin enerjisi ATP hidrolizi ile elde edilmektedir. Glutamin gerektiğinde sadece karaciğer ve böbrekte bulunan glutaminaz enzimi tarafından yeniden glutamik asit ve amonyağa parçalanmaktadır. Böylece amonyağın diğer dokularda serbest hale gelmesi önlenmektedir.
2-Diğer taşınma yolu glikoz-alanin döngüsüdür. Kas dokusunda transaminasyonla piruvattan oluşan alanin, kan yoluyla karaciğere taşınarak yeniden piruvata çevrilir. Karaciğerde transaminasyonla alaninden piruvat oluşurken α ketoglutarata aktarılmış olan amonyak, daha sonra glutamat dehidrogenaz ile serbest bırakılmaktadır. Böylece kas dokusundaki aminoasit azotu, üre sentezinde kullanılmak üzere karaciğere taşınmış olmaktadır. Aynı zamanda kas dokusunda glikoliz sonucu oluşan piruvat aynı yol ile alanin şeklinde karaciğere ulaşmaktadır. Yüksek organizmaların birçoğunda toksik olan ve pasif difüzyon ile uzaklaştırılamayan amonyaktan sentez edilen üre, azotlu son ürün olarak atılmaktadır. İnsanda amonyağın başlıca atılım şekli üredir. Bunun yanı sıra böbreklerde glutaminden serbest hale geçen amonyak, NH4
+ şeklinde klorür veya fosfat gibi anyonlarla eşleştirilerek atılmaktadır. Ekstrasellür ve İntrasellüer Tamponlar
Hemen her biyolojik reaksiyon optimum sabit pH değerlerinde gerçekleşir. pH değerindeki küçük bir değişme bile biyolojik bir işlemin gerçekleşmesinde büyük değişikliklere neden olur. Hücreler ve organizmalar özgül ve sabit bir pH’yı korurlar, böylece biyomoleküllerin en uygun iyonik durumda kalmaları sağlanır. Örneğin kan pH’sı 7.4, sitozol pH’sı ~ 7.0, mide pH’sı 1.5- 2.0, lizozom pH’sı ~ 5.0’dır ve bu pH sabitliği esas olarak biyolojik tamponlarla sağlanır. Tampon çözeltiler, bir zayıf asit ile eşlenik bazının veya bir zayıf baz ile tuzunun karışımından oluşan ve pH değişimlerine belirli ölçülerde direnç gösteren çözeltilerdir. Tampon çözeltiler, bir çözeltinin pH değerini belirli bir seviyede ve sabit tutmak gerektiği zaman kullanılır.

H A Y V A N F İ Z Y O L O J İ S İ
359
ŞEKİL 6.26 pH denegesinde böbrek ve akçiğerlerin rolü
Günlük oluşan H+ yükünün atılımı H+ iyonun tarafından ortamdan uzaklaştırılan HCO3 yeniden üretilmesi Filtre olan HCO3’ün tübüllerden geri emilir. Bunun yanında
ekstrasellüler ve inrasellüler sıvılarda farklı tampon sistemleri pH değişimlerine karşı koyar. Tampon sistemi genel olarak ortamdan Hidrojen iyonu vererek veya uzaklaştırarak bir dokuda veya olusyonda oluşabilecek pH değişiklikleri en aza indirgemeye çalışan sistemler
olarak tanımlanabilir.
Başlıca tamponlar H+ + HCO3����---------���� H2CO3 ����-------����CO2 + H2O
Metabolizma sonunda oluşan CO2 büyük bir kısmı ise alyuvarlara girer(%70). Alyuvarlarda karbonik anhidraz enziminin katalizlemesi sonucu CO2, su ile birleşerek karbonik asiti oluşturur. Karbonik asit (H2CO3), iyonlaşarak H+ ve HCO3
– (bikarbonat) iyonu meydana getirir. Bikarbonat plazmadaki en önemli tampon sistemi oluşturur. Bikarbona tamponun pK değeri 6.8 dir. Vücudun asit baz dengesinin iki önemli belirleyicisi, majör tampon sistemini oluşturan bikarbonat (HCO3
-) ve karbondioksit (CO2) tir. Böbrekler HCO3-, akciğerler CO2
konsantrasyonunun başlıca belirleyicileridir. Normal koşullarda kanda pH 7.35-7.45 arasıdır. Plazma HCO3
-düzeyinde azalma veya CO2’te artma asidemi, HCO3- düzeyinde artma
veya CO2’te azalma ise alkalemi tanımlanır. Normal ph değerinin 7.4 olduğu göz önüne alınırsa baz / asit oranının 20 olarak bulunur. Ayrıca böbreklerin HCO3
-, akciğerlerin ise CO2 konsantrasyonunun başlıca belirleyicileri olduğu göz önüne alındığında
olarak ifade edilebilir. 1-Kimyasal Tampon Sistemi (Saniyeler içerisinde etkisini gösterir) Bikarboant, Fosfat, protein ve hemoglobin tamponları 2-Solunum sistemi; Akciğerler yoluyla CO2 atılımının kontrolu (Dakikalar içerisinde etkisini gösterir)
• Bikarbonat tamponu • Fosfat tamponu • Proteinler • Hemoglobin
Proteinlere bağlı tampon sistemi

H A Y V A N F İ Z Y O L O J İ S İ
360
3-Üriner sistem; Böbrekler gerekirse geri emmek ve gerekirse salgılamak yoluyla H+ ve HCO3
- iyonlarının kandaki konsantrasyonlarını düzenleme yeteneğine sahiptir (Saatler, günler içerisinde etkisini gösterir).
Boşaltım sistemi Böbrekler Boşaltım sistemi bir çift böbrek ve üreter ile tek mesane (idrar torbası) ve tek üretradan oluşur. Boşaltım sistemin temel organı olan böbrekler bir çift halinde bel bölgesinde yağ doku ile çevrili olarak bulunur. Böbrek çevresindeki yağlar koru fonksiyonu yapar. Böbreklerde üretilen idrar, üreterden idrar torbasına doğru geçer, burada kısa süre için depolanır ve sonra üretra aracılığı ile dışarıya atılır. Her bir böbrekte 1 milyon adet fonksiyonel süzüntü birimi olan nefron, Her 24 saatte yaklaşık 1500 ml idrar oluşur. Temel foknksiyonu idar oluşturarak boşaltımı greçekleşmek olsada böbrekler birçok fonksiyonu birlikte görür.
• Böbrekler, vücudun sıvı ve elektrolit dengesini de düzenler ve aynı zamanda kan basıncının düzenlenmesinde görev alan Renin böbreklerde(%90) üretilir. Renin anjiotensi II adındaki hormonun üretilmesi için gereklidir.
• Eritrosit yapımını uyaran ve 30 kDa’luk bir büyüme faktörü glikoproteini olan Eritropoietin de böbreklerde üretilir. Triozin kinaz reseptörleri kullanan bu hormon eritrositlerin üretilmesi için gereklidir. Hipoksi ve/veya böbrek dolaşımında azalmasına bağlı olarak böbreklerce(%90) ve az miktada(%15-10) karaciğer tarafından sentezlenir.
• Eritropoietin aynı zamanda bir steroid ön hormon olan D3 vitaminini de, etkin biçimine hidroksile eder. Bu nedenle D Vitamin sentezi böbreklerde yapılır
• Bazı(protein) hormonların işlevleri tamaladıktan sonra yıkımı; kandan uzaklaştırılması böbrekler tarafında yapılır.
• Metabolik etki; böbrekler karaciğer kadar olmasa bazı maddlerin ihtiyaçı durumun sentezlenmesi diğer bileşiklerde üretilmesini yapabilir.
Böbrekler tarafından yıkılan hormonlar • İnsülin • Glukagon • Parathormon • Büyüme hormonu
Böbreklerce metabolize edilenler • Glukoneogenez • Lipid metabolizması
Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
361
ŞEKİL 6.27 Böbrek
Böbrek Yapısı
Böbrekler dış taraflarından düzensiz sıkı bağ dokusundan yapılı olan bir kapsül ile çevrilidir. Her bir böbreğin, iç bükey yapılı iç kenarında sinirlerin girdiği, kan ve lenf damarlarının girip çıktığı ve üreterin çıktığı yer olan hilusu ile dış bükey dış kenarı vardır. Böbrek dışta korteks ve içte medulla olmak üzere iki bölümde incelenebilir. Böbrekler; kırmızı-kahverenkte, parlak ve ince bir kapsülle örtülü, fasulye biçiminde iki organdır. Her böbreğin uzunluğu yaklaşık 10, genişliği 5 ve kalınlığı ise 3,75 cm.dir. Böbrekler karın bölgesinin arka kısmının iki tarafında, belin en üst kısmında, karın boşluğunun arkasında ve diyaframın altında bulunur. İnsanda böbrek medullası 10-18 adet konik ya da piramidal şekilli yapılar olan medullar piramitlerden oluşur. Her bir medullar piramidin tabanından kortekse uzanan birbirine paralel tübül demetleri olan medullar ışınlar çıkar.
ŞEKİL 6.28 Böbrek
Her nefron genişlemiş bir bölüm olan renal cisimcik (veya böbrek cisimciği), proksimal kıvrıntılı tübül, Henle kulbunun ince ve kalın kolları, distal kıvrıntılı tübül, toplayıcı tübül ve kanallardan oluşmaktadır. Her nefronun 2 ana bölümü vardır;

H A Y V A N F İ Z Y O L O J İ S İ
362
Böbreğin Kanlanması
Böbrekler kalpten gelen kanın yüzde 21 kısmını alır. Tüm kan yaklaşık her beş daikakada bir böbrekten geçmiş olur. Aortadan ayrılıp böbreğe gelen damar, böbrek arteri, hilustan böbreğe girer ve interlober, arkuat, interlobüler (radial) arterlere ve afferent arteriyollere ayrılır. Afferent arteriyoller, plazma proteinleri hariç, çok miktarda su ve maddenin filtre edilerek idrar yapımının başladığı glomerullardaki glomerular kapillerleri oluşturur.
ŞEKİL 6.29 Böbrek kanlanması
Erişkin bir kişide her iki böbreğe gelen kan(toplam kanın yüzde 21 kısmı), dakikada 1.21.3 litreyi bulur. Kanın tamamı her beş daikada bir böbreklerden geçmiş olur.
Damar adı Damar basınçları Başlangıç
Damar basınçları Son
Totalin %‟si Damar direnci
Böbrek arteri 100 100 ~%0
Arkuat ve interlobular arter ~100 85 ~%16
Afferent arteriol 85 60 ~%26
Glomerül kapillerleri 60 59 ~%1
Efferent arteriol 59 18 ~%43
Peritübüler kapiller 18 8 ~%10
Interlober ve interlobular venler 8 4 ~%4
Böbrek veni 4 ~4 ~%0
Böbreklerin kanlanması renal arterle olur. Her bir kortikal renal arter böbrek yüzeyine doğru olan seyri boyunca kendisine paralel seyreden ve herbiri bir glomerüle giden afferent arteriyolleri verir. Normalde glomerüle giren plazmanın sadece yaklaşık %20'si kapillerlerden Bowman kapsülüne filtre olur. Glomerüler kapillerler ise efferent arteriyol adı verilen diğer arteriyol sistemi ile birleşirler. Sonuçta kan her glomerülden bir efferent arteriyol ile ayrılır, hemen sonra ise ikinci bir kısım kapillerlere ayrılırlar. Bunlar ise peritubüler kapillerler olup yaygın olarak dağılmışlardır ve tübüllerin tüm bölümleri ile ilişkidedirler. Görevleri ise tubüler lümen ve
1-Glomerul ve Bowman kapsülünden oluşan Renal korpüskül 2-Proksimal tübül, ince bölüm ve distal tübülden oluşan Renal tübül.

H A Y V A N F İ Z Y O L O J İ S İ
363
kapillerler arasında solüt ve su geçişini sağlamaktır. Peritubüler kapillerler daha sonra yeniden birleşerek renal venülleri venüller birleşerek böbrekten ayrılan renal veni oluştururlar. Nefronda süzüntü oluşumu kan basıncı değişimlerinden en az etkilecek şekilde düzenlenir. Yani kan basıncı 80-180 mmHg arasında değişme göstersede Afferent arteriol kan basınç yaklaşık 60 mmHg sabit tutulur. Afferent arteriol basınç glomerüler filtrasyon oranı belirleyen en önemli değiken olduğun sabit kalması filtrasyon miktarının sabir kalmasını sağlar. Nefron tipleri
Böbreklerin fonksiyonel birimi olan nefronlar temel olarak iki tiptir. Kortikal nefronlar böbrekteki nefronların %80’ni oluştururlar. Bunların Henle kıvrımları uzun değildir, ince bölümleri ya çok kısadır ya da hiç yoktur. Diğer tip nefronlar Jukstamedullar nefronlar olarak bilir, korteksin medullaya komşu olan bölgesinde bulunurlar. Böbrekteki nefronların %20’sini oluşturur. Bunların Henle kıvrımları çok uzundur ve medullanın derinliklerine kadar inerler. Jukstamedullar nefron hipertonik idrar üretilmesi imkânı verir.
ŞEKİL 6.30 Kortikol ve justamedullar nefronlar
Jukstamedullar nefronlar sahip oldukları Henle kıvrımı yada kulpu, su tutma işleminde rol oynar; sadece böbreklerinde bu tür yapıları(Jukstamedullar nefronları) bulunan hayvanlar hipertonik idrar üretebilir ve vücut suyunu koruyabilirler(Kuş ve memelilerde bulunur). Sürüngenlerde jukstamedullar nefronlar gerek yoktur çünkü boşaltım ürünü çok az su ile atılabilen ürik asittir. Nefron yapısı
İnsanlarda her bir böbrekte 1.000.000 nefron birimi vardır. Her nefron renal korpüskül olarak adlandırılan filtrasyonda görevli bir komponent ile renal korpüskülden sonra devam eden tübülden oluşmuştur. Renal korpüskül birbiri ile ilişki içindeki kapillerlerin oluşturduğu kompak yumak şeklindeki glomerül (çoğulu glomeruli) ile glomerülün içine uzandığı balon benzeri içi boş bir kapsül olan Bowman kapsülü'nden oluşur.

H A Y V A N F İ Z Y O L O J İ S İ
364
ŞEKİL 6.31 Bowman kapsülü
Glomerül ve Bowman kapsülü arasındaki ilişkiyi gözümüzde canlandırmak için yumruğumuzu (glomerül) bir balonun (Bowman kapsülü) içine doğru bastırdığımızı düşünebiliriz. Bowman kapsülünün glomerül ile temas ettiği kısım içeri doğru çöker, ancak kapsülün zıt yüzeyi ile temas etmez. Buna göre, kapsülün iç kısmında hala bir boşluk kalmış olur ki buna üriner boşluk veya Bowman boşluğu denir. Glomerüler filtrat bu boşlukta birikir.
ŞEKİL 6.32 Nefron
Glomerüler ve Peritüpüller kapiller : Glomerüler kapiller, diğer kapiller ağlara göre daha yüksek hidrostatik basınca sahiptir. Ve tüm glomerül, Bowman kapsülü ile sarılıdır. Glomerüler kapillerden filtre olan sıvı, Bowman kapsülü içine ve sonra böbrek korteksinde yer alan proksimal tübül içine akar. Proksimal tübülden sonra medullanın derinliklerine inen Henle kulpunun çıkan kolu sonunda, nefron işlevinin kontrolünde önemli rol oynayan maküla densa bulunur. Peritüpüler kapiller proksimal, henle ve distal tüpülü çevresinde yer alan efferent arteiol ile venüller arasındadır. Peritüpüler kapiller geriemilim ve sekrasyonda görev alır. Affernet arteiol ile afferen arteiol arasında yer alan glomerus kapilleri bazı farklılıklara sahiptir :
1- Glomerüler kapiller iki arteiol damar arasında yer alan tek kapillerdir. 2- Glomerüler kapiller ortalama sistemik kan basıncının iki katı basınçta kan geçer(60
mmHg).
Bowman kapsülü, aslında böbrek tüplerinin merkezi glomerülü, yani kılcal damar yumağını dışardan sarmakta olan uzantısıdır. Bowman kapsülünün iki yaprağı vardır. Birincisi içte, kılcal damar oluşturan bütün damar kıvrımlarını örten bölümdür. Buna “Viseral yaprak” denilmektedir. İkincisi ise glomerül yumağını bir torba gibi dışardan saran bölümdür. Bu bölüme ise “Parietal yaprak” denilmektedir. Viseral yaprak ile parietal yaprak arasında bir boşluk bulunmaktadır. Bu boşluğa “İdrar boşluğu” denilmektedir. Parietal yaprak aslında viseral yaprağın bir uzantısıdır. Ancak, parietal yaprak “Skuamöz epitel” hücrelerinden yapılmış viseral yaprak ise “Podosit” denilen özel epitel hücrelerinden yapılmıştır

H A Y V A N F İ Z Y O L O J İ S İ
365
3- Glomerüler kapiller de iki hücre tapakası kapiller endotel ve kapsüller endotel ile filtrasyon yapılır. Yalnız süzülme olur geri emilim resoption olmaz. Sistemik kapiller e göre 100 kat geçirgen bir yapıdır.
ŞEKİL 6.33 Peritübüler kapillerrde kan basıncı ve böbrekte yoğun değişim
Bowman kapsülünün iki tabakası arasında, kapiller duvarından ve visseral tabakadan süzülen sıvının toplandığı idrar boşluğu bulunmaktadır. Her böbrek cisimciğinde, getirici (afferent) arteriyollerin girdiği ve götürücü (efferent) arteriyollerin çıktığı bir damar kutbu ve proksimal kıvrımlı tübüllerin başladığı bir idrar kutbu bulunur. Her böbrek cisimciğinin çapı yaklaşık 200 µm’dir ve kapiller bir yumak olan glomerulden oluşmuştur. Bu yumak, Bowman kapsülü olarak adlandırılan iki tabakalı epitelden oluşan bir kapsülle sarılmıştır. Kapsülün iç tabakası (visseral tabaka) glomerulün kapillerlerini dış taraftan sarar. Dış tabaka, böbrek cisimciğinin en dıştaki sınırını oluşturur ve Bowman kapsülünün pariyetal tabakası adını alır. Glomerular kapillerleden süzülen sıvı Bowman kapsülü içine ve daha sonra kapsülün devamı olan proksimal kıvrıntılı tübüle geçer. Bu iç tabakadaki hücrelerin gövdelerinden, birkaç birincil (primer) uzantı şekillenir ve bu hücreler ayaklı hücreler (podositler) adını alır. Her bir primer uzantı ayakçık (pedisel) denen glomerulün kapillerlerini saran çok sayıda ikincil (sekonder) uzantı oluşturur. İkincil uzantılar, 25 nm’lik sabit bir mesafede, bazal lamina ile doğrudan temas halindedirler. Ancak, podositlerin hücre gövdeleri ve birincil uzantıları bazal laminaya değmez.
Filtrasyon yarıkları ve Podositlerin uzantıları
ŞEKİL 6.35 Süzülme yapan podositler
• Glomerüler kapiller membran, diğer kapillerlerdenyüzlerce kat (300-600) daha • geçirgendir. • Glomerül kapiller membranın temel bariyeri bazal membrandır. Bazal
membrandaki proteoglikanlar güçlü (-)tir. Bu olay proteinlerin geçişini engeller ve – yüklü maddelerin nötral ve + yüklü olanlardan daha zor geçmesini sağlar.
• Glomerüler filtratın bileşimi şekilli elemanlar ve proteinler hariç plazmaya benzer. Ca++ ve yağ asitleri gibi proteinlere bağlı taşınan maddeler filter edilemezler, bu yüzden glomerüler ultrafiltratın içeriğinde bulunmazlar.
• Glomerüler filtrasyonu oluşturan temel güç kanın hidrostatik basıncıdır. Aort basıncı 40-50 mmHg‟ya düşürüldüğünde idrar oluşumu durur. Normal bir erişkinde dakikada oluşan glomerüler filtrat miktarı 125 ml‟dir.

H A Y V A N F İ Z Y O L O J İ S İ
366
Filtrasyon kısmın buluna glikoaminoglikanlar secici geçirdekli sağlayan negatif yüklü maddelerdir. Lamina rara, hücrelerin tutunmasına yarayabilecek olan fibronektin içerir. Lamina densa ise, negatif yüklü bir proteoglikan olan ve katyonik moleküllerin geçişini engelleyen heparan sülfat içeren bir matriks içinde tip IV kollajen ve lamininin oluşturduğu ağ şeklinde bir yapıdan oluşur. Yani glomerul bazal laminası, seçici bir makromoleküler filtre görevi üstlenmiştir. 3,50 nm’den daha büyük partiküller bazal laminadan geçemezken; su, iyonlar ve düşük molekül ağırlıklı bileşiklerin geçişine izin verir. Molekül ağırlığı albüminin molekül ağırlığından (69 kDa) fazla olan negatif yüklü proteinler ise membrandan eser miktarda geçerler. Özetle 3.5 nm küçük pozitif yada nötür moleküller filtrasyona süzünteye geçerken negatif yüklü ve 3.6 nm büyük moleküller süzülmeden efferent arteriolle geçer. Proteinlerin hemen hemen tümü, tüm hücreler efferent arteiole geçerken filtrasyona başta su olmak üzere glikoz, atık maddeler tüm yüksüz küçük moleküller geçer. Bowman kapsülünde basınç bağlı filtrasyon
Bowman kapsülün içinde yer alan glomerüler kapillerden filtrasyon ile ile ilk süzüntü meydana gelir. Filtrasyon, mekanik basınç etkisiyle(60 mmHg afferent arteriol kan basınçı nedeniyle) ile maddelerin membrandan geçidir. Filtrasyon için basınç farkı gereklidir. Böbrek kapsülünde basınç farkı 3 temel faktör tarafından kontrol edilir.
• Afferent arteiol basıncına (60 mmHg) • Affrent arteiol basıncına karşı koyan iki kuvvet vardır kolloid osmotik basınç (yada
onkotik basınç 32 mmHg) ve • bowman kapsül basınçı (18mmHg).
Net geçiş bowman kapsülü yönünde 10 mmHg basınçla olur. Bu basınç afferent arteiolden bowman kapsülüne madde geçişini sağlar. Tüm glomerüler kapiller boyunca madde geçisi bowman kapsülü içine doğrudur. Efferent arteiol tarafında basınç azalmasına rağmen yinede bowman kapsülüne doğru net fitrasyon olayı gözlenir.
ŞEKİL 6.36 Afrenet ve effrent arteriolde kan basınç değişimi
Glomerüler filtrasyon oranı değişimi temel olarak bu farktörlerin düzenlenmesine bağlıdır. İlke olarak glomerüler hidrostaik basınçının artması( afferen arteiol basınçın artması, efferent arteiol çapının azalması) GFO artırır. Benzer olarak bowman kapsül basınçının ve/veya glomerüler onkotik basınçın azalması GFO artırır.

H A Y V A N F İ Z Y O L O J İ S İ
367
Efferent arteriol tonus artarsa, glomerüler filtrasyon basıncı artar
Afferent arteriol tonus artarsa, glomerüler filtrasyon basıncı azalır
Kanın hidrostatik basıncına yanıt olarak glomerul süzüntüsü oluşur. Glomerul süzüntüsünün kimyasal bileşimi kan plazmasına benzer ancak, makromoleküller( plazma proteinleri, tüm şekilli elementler) glomerul duvarını geçemediği için hemen hiç protein(çok az miktarda geçiş olabilir) içermez. Glomerul süzüntüsüne geçebilen en büyük proteinin molekül ağırlığı 69 kDa civarındadır ve süzüntüde eser miktarda albümin görülür. Glomerul kapillerlerinin endotel hücreleri pencereli tiptedir ama diğer pencereli kapillerlerin açıklıklarını kaplayan ince perdeye sahip değillerdir. Endotel hücreleri ve podosit ayaklarının yanı sıra glomerul kapillerlerinin duvarlarına tutunan mezangiyal hücreleri vardır. Mezangiyal hücreler kasılabilen hücrelerdir ve anjiyotensin II reseptörlerine sahiptirler. Bu reseptörler etkinleştiğinde, glomerula gelen kan akımı azalır. Glomerül filtrasyon Oranı ayarlanması
Böbrekte afferent kan akımın miktarını (glomeruler filtrasyon oranı) sabit tutan mekanzimalara sahiptir:
Normalde GFH günde 180 litredir ve bunun 178,5 litresi tübülden geri emilir; 1,5 litre/gün sıvı idrarla atılır. Otoregülasyon yokluğunda kan basıncında kısmen küçük bir artış (100’den 125 mmHg’ya) GFH'de benzeri %25’lik artışa sebep olacaktır (180 litre/gün’den 225 litre/gün). Eğer tübül geriemilimi 178,5 litre/gün değerinde sabit kalırsa bu idrarın toplam miktarında 30 katından fazla artış yaparak idrar akımını 46,5 litre/gün’e çıkaracaktır (GFH ile tübül geriemilimi arasındaki fark). Toplam plazma miktarı sadece 3 litre olduğu için böyle bir değişiklik kan hacmini çok çabuk tüketecektir.
Glomerüler filtrasyona neden olan basıncı ~60 mmHg dır. (OAB ‘ nin % 60 ) Plazma onkotik P = ~32 mmHg (filtrasyona engel olan basınç) Renal interstisyel P = ~18 mmHg(filtrasyona engel olan basınç)

H A Y V A N F İ Z Y O L O J İ S İ
368
ŞEKİL 6.37 Nefronda hidrositaik basınç ve kolliod osmatik basınç bağlı filtrasyon
Fakat gerçekte arter basıncında böyle bir değişiklik iki nedenle idrar hacmini çok daha az etkiler: (1) böbrek otoregülasyonu GFH’de olacak değişiklikleri önler ve (2) böbrek tübüllerinde glomerülotübül dengesi denilen ve GFH arttığında tübüllerin geriemilim hızını artırmaya olanak veren bir ilave uyum mekanizması vardır. Bu özel kontrol mekanizmalarına karşın arteryel kan basıncı artışı böbrek su ve sodyum atılmasına önemli etki yapar; buna basınç diürezi veya basınç natriürezi denir. Bu mekanizma, vücut sıvı hacminin ve arteryel basıncın düzenlenmesinde çok önemlidir. 1. Miyojenik Yanıt: GFO ve böbrek kan akımının otoregülasyonundan iki mekanizma sorumludur: birinci mekanizma arteryal basınç değişiklerine yanıt verir. Diğeri tübüler sıvıdaki NaCl konsantrasyonu değişimine yanıt verir. Birlikte aferent arteriyoldeki tonusu ayarlarlar. Basınç algılama mekanizması ya da miyojenik mekanizma damarın düz kasının intrensek özelliği ile ilgilidir: Gerildiğinde kasılmaya eyilimlidir. Buna göre arteriyal basınç arttığında (90-180 mmHg) ve böbrek aferent arteriyolün içini döşeyen düz kas hücrelerinde, damar duvarı gerildiğinde (örn. luminal basınçta artışa bağlı) aktive olan Ca2+’a geçirgen mekanosensitif kanallar(mekanik kapalı kanallar) bulunur. Hücre içine Ca2+ girişi kas kasılmasını başlatır ve arteriyol refleks olarak daralır. Arteriyol dirençteki artış böbrek kan akımı ve basınçtaki artışı dengelediği için, GFO sabit kalır (yani, eğer ∆P/R oranı sabit tutulursa böbrek kan akımıda sabit kalır). 2. Tübüloglomerüler geribildirim: Tübüloglomerüler geribildirim (TGG, ing; TGF böbrek tübüllerinde sıvı akımını optimize edecek şekilde ayarlanmasını sağlayan mekanizmasıdır. Jukstaglomerüler aygıtın (JGA) aracılık ettiği bir otoregülasyondur. Renal Tübüler akım arıtışı GFR düşürür. Kan basıncı artışı, Na reabsorbsiyonunu azaltır. Makula densa, afferent arteriol ve glomerül kapiller permeabiliteyi etkiler. OAB ‘da azalır ve Makula densaya gelen NaCl renin-anjiotensin sistemini aktive eder. Anjiotensin2’nin efferent arteriolde vazokonstrüksiyona yol açar. Justaglomerüler hücrelerin afferent kan basıncına bağlı olarak salgıladığı renin enzimi ile GFO düzenlemsidir. Sonuç olarak, otoregülasyon arteriyel basınçtaki değişikliklere karşı böbrek kan akımının ve Glomerüler filtrasyon oranı cevabını yavaşlatır veya azaltır, ancak bu değişiklikleri tamamen önleyemez. B. Otonom sinir sistemi – Sempatik Sistem ile GFO korunması: Aferent ve eferent arteriyoller sempatik nöronlar tarafından uyarılırlar. Bununla beraber dolaşan kan hacmi normal iken, sempatik tonus minimumdur. Sempatik sinirlerden norepinefrin salınır ve dolaşımdaki

H A Y V A N F İ Z Y O L O J İ S İ
369
epinefrin adrenal medülla tarafından salgılanır. Başlıca aferent arteriyolde bulunan α-1 adrenoreseptörlere bağlananan bu maddeler vazokonstriksyona yol açar. Bu bağlanma böylece böbrek kan akımı ve Glomerüler filtrasyon oranı düşürür. Efektif dolaşım kan hacmindeki azalma veya korku ve ağrı gibi güçlü emosyonel uyarılar sempatik sinirleri aktive ederek, böbrek kan akımı ve Glomerüler filtrasyon oranı düşürür. Sempatik uyarılar (adrenerjik), omurganız T4-L1 arasından çıkar. β-1 reseptörleri (nörotransmiterleri nörepinefrin ve dopamin) jukstaglomerüller aparatı uyarır. α-1 Proksimal tübül Na reabsorbsionunu arttırır. α-2 Na reabsorbsiyonunu azaltır ve su atılımını artırır. D1 afferent ve efferent arteriolde vazodilasyon yapar. D2 arteriollerin vazodilasyon sağlar. Dopamin, Na reabsorbsiyonunu azaltır. C. Hormonlar / Parakrinler : Kan volümü artınca sağ atriyum basıncı artar. Atriyumdaki myositlerden ANP salgılanır. ANP vazodilatasyon yapar. ADH salınımını baskılar. Su atılımını RAAS ‘ı baskılar.
ŞEKİL 6.38 RAAS sistemi
Aldosteronun distal tübül ve toplayıcı tübüldeki etkisini antagonize eder. Endotelden Kaynaklanan Nitrik Oksit, Prostaglandinler ve Bradikinin böbrek damar direncini azalması ve son olarak Anjiyotensin II Efferent Arteriyolleri daralmasına yol açar.
Mezangiyal hücreler
Mezengiyal (Mesos, orta, angeion, damar) hücreler glomerul bazal membranı tarafından yakalanmış olan normal ve patolojik (immünkompleks) molekülleri uzaklaştırır. Belki de sitokinler ve prostaglandinler gibi kimyasal aracıları üretirler. Damar kutbunda yer alan ancak glomerulun dışında, jukstaglomerular aygıtın bir kısmını oluşturan glomerul dışı mezangiyal hücreler de vardır. Mezangiayal hücreler ile Glomerüler filtrasyon oranı düzenleyen mekanizması:
• Mezangiyal hücreler kasılabilen hücrelerdir ve anjiyotensin II reseptörlerine sahiptirler.
• Bu reseptörler etkinleştiğinde, glomerula gelen kan akımı azalır. • Mezangiyal hücrelerin, kalbin atriyum hücreleri tarafından üretilen atrial natriüretik
faktöre (ANP) ait reseptörleri de vardır. ANP, damar genişleticidir ve mezangiyal hücreleri gevşeterek muhtemelen kan akımını ve süzülme için bulunan etkin yüzey alanını artırır.
Damar kutbunda yer alan ancak glomerulun dışında, jukstaglomerular aygıtın bir kısmını oluşturan glomerul dışı mezangiyal hücreler de vardır. Mezangiyal hücrelerin başka birkaç işlevi daha vardır:
1- Glomerula yapısal destek verirler(mekanik destek), 2- Glomerus içindeki hücre dışı matriksi sentezlerler ve Endositoz gerçekleştirirler,

H A Y V A N F İ Z Y O L O J İ S İ
370
Böbreklerde Geri emilim
Nefronda glomerusun ardında proksimal, henle, distal tüpülde ve toplama kanalında geri emilim tapılır. Geri emilim aktif yada pasif olabilir. Aktif olarak geri emilen veya salgılanan solütlerin pek çoğunun taşınabilmesinde bir sınır vardır. Buna taşıma maksimumu (maksimum transport) denir. Bu maddeler proteinlere bağlı taşındıklarında taşıma proetinlerin sayısı taşıma kapasitesini belirler.
Glomerüler filtrasyon----� Tubüler reabsorbsiyon
Tubüler sekresyon-------�Ekskresyon
Bir maddenin geriemilebilmesi için önce : (1) tübül epitel zarından böbreğin hücrelerarası sıvısı içine ve sonra (2) peritübül kapiller zar-aracılığı ile kana geri taşınması gerekir. Solütlerin hücre zarlarından taşınması pasif mekanizmalarla, aktif taşıma mekanizmalarıyla veya endositozla olabilir. Memelilerde solüt hareketi hem aktif hem de pasif mekanizmalarla görülürken tüm su hareketi pasiftir Örneğin, çözünmüş maddeler veya su doğrudan hücre zarlarından geçebilecekleri gibi (transselüler yol), hücreler arasındaki bağlantı bölgelerinden de geçebilirler (paraselüler yol). Su ve çözünmüş maddeler tübül sıvısından hücrelerarası sıvıya geçtikten sonra, peritübüler kapiller duvardan kana ultrafiltrasyon (kütle akımı) ile geçerler. Kütle akımı olayı hidrostatik ve kolloid ozmotik kuvvetler ile oluşmaktadır. Peritübül kapillerleri, diğer birçok kapillerin venöz ucu gibi davranır; çünkü burası, sıvı ve maddelerin hücrelerarası alandan damar içine geriemilmesini sağlayan net güçlerin bulunduğu bölgedir.
ŞEKİL 6.39 Böbrekte geri emilim ve sekrasyon oranları ve NaCl farklı tüpüllerdeki emilim oranları
Proksimal tüpülde Na reabsorbsiyonunun %65 (Na-K ATPaz aktivitesi ile) yapılır. Ayrıca K, Ca, Mg reabsorbsiyonu (elektriksel gradiyent) yapılır. Tüm Glikoz, amino asit ve Fosfat reabsorbsiyonla geri alınır. Cl reabsorbsiyonu (pasif olarak veya K-Cl kotransportu ile) da proksimal tüpülde olur. Proksimal tüpülde kan pH dengesi için önemli olan HCO3
- reabsorbsiyonu %90 oranında olur. H+ sekresyonu (pH yüksektecek şekilde olur) da proksimal tüpülde olur. Proksimal tüpül membranın lümen tarafında Na reabsorbe edilirken H+ sekrete edilir. Proksimal tüpülde yapılan geri emilimde su kalsiyum ve potasyum paraselüler yolu kullanırken, suyun bir kısmı glikoz ve peptidler transellüler yolu kullanır.

H A Y V A N F İ Z Y O L O J İ S İ
371
1. Membranın bazolateral tarafındaki Na-K ATP az pompası tarafından oluşturulan elektrokimyasal fark ile sodyum, apikal membran da denen lüminal membrandan, hücreye doğru difüze olur. 2. Na, luminal membrandan tübüler hücre içine bazolateral membrandan sodyum-potasyum ATP az aracılığı ile konsantrasyon ve elektriksel farka zıt yönde taşınır. 3. Sodyum, su ve diğer maddeler, hidrostatik ve kolloid osmotik basınç farklarının yönlendirdiği ultrafiltrasyon denen pasif bir hareketle hücreler arasından, peritübiller kapiller içine geri emilirler.
ŞEKİL 6.40 Böbrekte proksimal tüpülde glikozun aktif taşınması
Henle kulbunda Na iyonlarının %25 kısmı reabsorbsiyonunun edilir Henlenin inen kalın kolu suya geçirgen değildir. Henle çıkan kolunda Na, Cl reabsorbsiyonu olur. (Na/K/2Cl transport sistemi) Henle kulbunda tübül sıvısındaki Cl konsantrasyonu hız belirleyici faktördür. Madde Mol. Wt. Filtrasyon oranı(suya göre oranı) Üre 60 1.00
Glikoz 180 1.00 Inulin 5,500 1.00 Myoglobin 17,000 0.75 Hemoglobin 64,000 0.03 Serum albumin 69,000 0.01 Henle kulbunda Magnezyum en fazla reabsorbe olduğu yerdir. Kalsiyum miktarı üzerine etkili olan Parathormon henle kulbunda Ca+2 reabsorbsionunu hızlandırır. Henle kulpunun diğer bölümlerinde su osmotik gradienti takip eder. Distal tüpülde Filtre edilen Na’un %5’i reabsorbe edilir. Kapiller tarafta enerji, Na-K ATPaz aktivitesinden kazanılır. Lümen tarafında Na reabsorbsiyonu, Na-Cl aktivitesi ile olur. Madde Günlül Filtrasyon İdrarla atılan Günlük reabsorsiyon oranı Su L 180 1.8 l %99 Sodyum Na g 630 3.2 g %99.5

H A Y V A N F İ Z Y O L O J İ S İ
372
Glikoz, g 180 0 g %100 Üre , g 54 30 g %44 Distal tübül, Parathormon ve D vitamininin etkilediği temel yerdir (Kalsiyum reabsorbsiyonu) Aldosteronun sodyum reabsorbsiyonuna etkisi vardır. Distal tübüldeki hücreler su, NaCl, bikarbonat iyonlarının geri emilmesinde ve K, Na ve NH4 iyonlarının süzüntüye eklenmesinde rol oynarlar. İdrar, asit özelliğini distal tübüllerde kazanır. Distal tüpülde yapılan aktif taşımada Na:K:2Cl ko-transporter görev alır Bu transpoterun inhibitörü olan madde Furosemide’dir.
ŞEKİL 6.41 Na:K:2Cl ko-transporter ve inhibitörü olan flurosemide
Distal tübüllerden geçen idrar, birbirlerine bağlanarak daha büyük, düz toplayıcı kanalları oluşturan toplayıcı tübüllere boşalır. Bu kanallar, medullar piramitlerin ucuna yaklaştıkça genişler. Glomeruslarda filtre edilen daha sonra sırasıyal proksimal henle ve distal tüpüllerde geri emilen süzünde en son Kortikal toplayıcı tübül, Medüller toplayıcı tübül yoğunlaştırır. Tübüllere çok miktarda bikarbonat iyonu fíltre edilir ve eğer bu idrarla atılırsa kandan baz alınmış olur. Diğer taraftan tübüler lümene tübüler epitel hücreleri tarafından çok miktarda hidrojen iyonu salgılanır ve böylece kandan asit uzaklaştırılmış olur. Eğer filtre edilen bikarbonat iyonlarından daha fazla hidrojen iyonları salgılanırsa ekstraselüler sıvılardan asit kaybı oluşur. Tersine olarak da, salgılanan hidrojen iyonundan daha fazla bikarbonat filtre edilirse baz kaybılacaktır. Böbrek bu yolla akçiğerle birlikte kanın pH değerinin(7,4-7,8 arasında) sabit kalmasında anahtar rol oynar. Toplayıcı kanalların epiteli, arka hipofiz(nörohipofizden) tarafından salgılanan ADH tepki verir. Eğer su alımı sınırlı ise, ADH (hipotamustan supraoptik çekirdeklerden sentezlenir ve hipofizden nörohipofizden salgılanır) antidiüretik hormon salgılanır ve toplayıcı kanalların epiteli glomerul süzüntüsünden emilip kan kapillerlerine aktarılan ve böylece vücutta tutulmuş olan suya geçirgen hale gelir.

H A Y V A N F İ Z Y O L O J İ S İ
373
ŞEKİL 6.42 Macula densa
Antidiüretik hormon varlığında ADH, lümen zarında yer alan zar içi tanecikler, su emilimi için kanallar oluşturabilecek şekilde toplanırlar (Aquaporin2). Bu görevi yapan aquaporinler hücrenin hacim ve ozmolaritesi düzenleyen taşıyıcı proetinlerdir. Hücre membranında suyun geçişi için bariyer ve geçiş noktası olarak görevi görür. Özel su kanalları (aquaporinler) ilk kez su geçirgenliği yüksek hücrelerde tanımlanmıştır. Memelilerde 13 tip olmak üzere canlılar aleminde 200’den fazla üye ile temsil edilen aquaporin ailesi aynı zamanda gliserol, üre, arsenit ve bazı iyonların taşınmasına da görev alır.
ŞEKİL 6.43 Aquaporinlerin ADH ile aktivasyonu

H A Y V A N F İ Z Y O L O J İ S İ
374
Diüretikler idrar miktarını artıran maddelerdir. Örneğin çay ve kahvede bulunan kafein ve teofilin Na -K ATPaz inhibitörü olarak böbreklerden su geri elimini azaltır ve bol idrar oluşumuna neden olur. Diğer diüretik madde olan alkol hipotalamusta supraoptik çekirdekten ADH sentezini azaltır. Buna bağlı olarak su geri emilimi (Distal tüp ve toplama kanallarındanki Aquaporinler üzerinden) azaltarak idrar mikarını artır.
Vesa recta
Vesa recta böbrek medüla kısmına inen ve henle kulbu çevresinde yer alan peritüpüler kapiller kan ağıdır. Vesa recta kan akımı azdır, total renal kan akımının sadece % 1 – 2 'sini oluşturur, bu yavaş kan akımı dokuların metabolik ihtiyaçlarını karşılamak için yeterlidir, fakat medulla interstisyumundan erimiş madde kaybını en aza indirmeye yardım eder. Vesa recta medüla bölgesindeki dokuların beslenmesi yanında medüla bölgesinin yüksek osmotik basınçta (1200 Osm) kalmasını sağlar. Vesa recta yüksek osmotik basınç sağlanması ve korunmasında ters akım ilkesininden yararlanır. Zıt yönde akan kapiller damarlar hem kendi aralarında hemde henle kulbu arasında etkin bir ters akım gerçekleşir.
ŞEKİL 6.44 Vesa recta
Vesa rectanın aşağı inen kolundan aşağı akan plazma, suyun kandan dışarı diffüzyonu ve erimiş maddelerin renal interstisyel sıvıdan kana doğru diffüzyonu nedeniyle daha hiperosmotik olur. Vasa rectanın çıkan kolunda, erimiş maddeler interstisyel sıvı içine, su da vaza rekta içine geri diffüzyona uğrar. Eğer ters akım mekanizması olmasaydı büyük miktarlarda erimiş madde medüladan vaza rektaya geçer, kaybedilebilirdi. Vaza rekta, ters akını değişim sistemi olarak çalışır ve(U şeklinde kapiller sahip olmasından dolayı) medulla intersitisyumundan erimiş maddelerin uzaklaştırılması bu sayede en aza indirilir. Ters akım sayesinde medulla intersitisyumun hiper osmolaritesi korunur.
Jukstaglomerüler Apparat ve Hormonal kontrol
Distal kıvrımlı tübülüsler kortekste izledikleri yol boyunca bir noktada kendi nefronlarına ait glomerülün damar kutbuna değerler. Bu değme noktasında afferent arteriol ve distal tübülus epiteli modifiye olur. Tübülus epitel hücreleri bu Juxtaglomerüler (Jg) Bölge’de silindirik hale gelir, kısalır, kalabalıklarır ve çekirdekleri biraraya toplanır. Mikroskopik kesitlerde nükleusların yakın yerleğimi nedeniyle daha koyu görünen bu distal tübülus segmentine Makula Densa denir. Afferent arteriolün, glomerüle girdiği yerde, tunica

H A Y V A N F İ Z Y O L O J İ S İ
375
mediasında modifiye granüllü düz kas hücreleri bulunmaktadır. Makula densaya çok yakın olan bu hücrelere Jukstaglomerüler Hücreler adı verilir. Jukstaglomerüler hücreler ile makula densa'nın ve glomerülün birbirine iliştiği alanda mezengial hücrelere benzeyen, onlarla devam ediyormuş gibi görünen, Ekstraglomerüler Mezengial Hücreler, Lacis Hücreleri, Nongranüler Hücreler Veya Polkissen (Kutup Yastığı) şeklinde değişik isimler alan bir grup hücre vardır. Açık renk boyanan bu hücreler matriks sentezeleme prostagladin ve sitokinin salgılama gibi görevleri vardır. Ayrıca bu hücreler mekanik destek ve kasılma ve glomerüler kan akışını kontrol gibi işlevleri görür. Bu hücreler(lacis hücreleri), Justa glomerüler hücreler ve makula densa beraberce Jukstaglomerüler Apparatus'u oluştururlar. JG apparatus küçük bir endokrin organdır.
ŞEKİL 6.45 Makula densa
Kan basıncının düşmesi ile renin-angiotensin-aldosteron düzeneği aktive olur. Böbrekte salgılanan reninin ana kaynağı Jukstaglomerüler hücrelerdir. Özelleşmiş Afferent ve Efferent arteriol (jukstaglomerüler hücreler), Henle kulpunun çıkan kalın kortikal segmentinin son kısmı (maküla densa) bir araya gelir. Bowman kampsülüne gelen affernet arteriyolün orta tabakasında (tunika media) değişmiş düz kas hücreleri bulunmaktadır. Distal kıvrıntılı tübüllerde bulunan makula densa, jukstaglomerular hücrelerin yer aldığı affrent arteriyol kısmına çok yakındır. İkisi birlikte jukstaglomerular aygıt adını alırlar. Bu hücrelere jukstaglomerular hücreler adı verilir. Jukstaglomerüler hücrelerde Renin enzimi bulunur. Renin salınımı; sempatik uyarı, afferent arteriol basıncı, makula densa’nın Cl akımından etkilenir. Jugstaglomerular hücreler aynı zamanda, eritrosit yapımını uyaran eritropoietin de üretirler. Makula densa Glomerüler Filtrasyon Oranı (GFO) ayarlayan önemli bir mekanizmaya sabittir Distal Tüp içinde Na ve Cl seviyesine göre makula dense Justaglomerüler hücrelerden renin salgılanmasını uyarır.

H A Y V A N F İ Z Y O L O J İ S İ
376
ŞEKİL 6.46 Maküla densada Na ve Cl İyonuna bağlı olarak Renin salgılanması mekanizması ve renin etki mekanizması
Makula densa temel olarak olarak iki işlevi vardır (1) afferent arteriol direncini azaltarak glomerüler hidrostatik basıncı artırır ve GFR’nin normale dönmesine yardım eder ve (2 ) reninin esas depolandığı yer olan afferent ve efferent arteriyollerin jukstaglomerüler hücrelerinden serbestlemesini artırır. Bu hücrelerden salgılanan renin, bir enzim görevi yaparak anjiyotensin II'ye dönüşecek olan anjiyotensin I yapımını artırır. Sonuçta anjiyotensin II efferent arteriyolleri daraltır, böylece glomerüler hidrostatik basıncı artırarak GFR’yi normale doğru çevirir. Renin
Makula densa hücreleri distal tübül içi sıvıdaki klorür iyon içeriğine ve su hacmine duyarlıdır, dolaşıma renin salgısını başlatan moleküler sinyaller üretirler. Başlıca üç etken renin salgılanmasını artırır: (1) Böbrekte jükstaglomerüler aparat hücrelerine giden sempatik sinirlerin uyarılması adrenejik reseptörler ile dopamin ve nörepinefrin aracılığı ile olur. Jukstaglomerüler hücrelerin beta-reseptörleri aracılığıyla renin salgılanmasını artırır. (2) Böbrek kan akımının azalması: kan miktarının yada basınçının azalması justaglomerüler hücrelerden Renin enzimin kana verilmesine neden olur. (3) Distal tüpüllerden geçen ultrafiltratın Na+ düzeyinin azalması. Justa glomerüler hücreler Distal tüpüldeki Na ve Cl seviyesine göre Renin salgılanmasını düzenler. JG(jukstaglomerular hücreleri) hücreler, anjiyotensinojen adı verilen plazma proteinini anjiyotensin I’e dönüştürecek olan renin enzimini üretirler. Anjiyotensin I, akciğer endotel hücrelerinde yüksek yoğunlukta bulunan dönüştürücü bir enzimin ACE (Kininaz II, peptidil dipeptidazkininaz II) etkisiyle iki aminoasitini kaybederek anjiyotensin II’ye (8 amino asitlik polipeptid) dönüşür. Ayrıca ACE enzim, bir plazma kinini olan bradikinini (vazodilatör bir madde) inaktive eder. Kininaz II adı verilmesinin nedeni de budur.
ŞEKİL 6.47 Anjitensin etkisi

H A Y V A N F İ Z Y O L O J İ S İ
377
Anjiotensin bilinen en güçlü vazokonstriktörlerden biridir. Özellikle arteriyolleri ve prekapiller sfinkterleri kasmak suretiyle total periferik damar rezistansını artırır. Kan basıncını yükseltir. Anjiotensin ve anjiotensin III, kan basıncını etkileyemeyecek kadar ufak dozlarda bile aldosteron sentezini ve sagılanmasını artırırlar. Böylece Na+ ve su dengesinin düzenlenmesinde rol oynarlar Böbreklerdeki aferent ve eferent arteriyoller, Anjiotensinin vazodilatör etkisine en duyarlı damar segmentidirler.
ŞEKİL 6.48 RAAS
Bir kanamadan sonra kan hacmi azaldığında (kan basıncında düşme olduğunda) renin salgısı artar. Susuzluk (sıvı kaybı) Renin salgılanmasını artırır. Anjiotensin bilinen en güçlü vazokonstriktörlerden biridir. Özellikle arteriyolleri ve prekapiller sfinkterleri kasmak suretiyle total periferik damar rezistansını artırır. Kan basıncını yükseltir. Venler ve venüller üzerindeki kasıcı etkisi zayıftır.
ŞEKİL 6.49 Anjotensinojenin aktif hale gelmesi
Juxtaglomerular Hücreler: Glomerul afterent arteriol media tabakasında Renin salgılayan miyoepitelyal hücrelere verilen isimdir.

H A Y V A N F İ Z Y O L O J İ S İ
378
Böbreklerdeki aferent ve eferent arteriyoller, kasıcı etkisine en duyarlı damar segmentidirler. Eferent arteriyoller bazı koşullarda(renal arter stenozu gibi) anjiotensine aferent arteriyollerden daha duyarlıdır. Kapiller permeabiliteyi artırır. Sonuçta dolaşan plazma hacmini azaltır. Dolaşımdaki Anjiotensin II 1-2 dakika içinde anjiotensinojenaz enzim ile ile yıkılarak yok edilir. Üretilen anjiyotensin II, hem arteriyolleri daraltır hem de böbrek üstü bezlerinde adrenal kortekste üretilen aldosteron hormonunu salgılanmasını arttırır
ŞEKİL 6.50 Vazopressin(ADH) bağlı olarak aquaporin protein üretimi ve distal tüpül ve toplama kanalında artışı.
Böbrek üstü bezlerinden(korteksinden) salgılanan Adosteron, böbrek tübül hücrelerine (en çok distal tübüllere) etki ederek glomerul süzüntüsünden sodyum ve klorür iyonlarının emilimini arttırır. Sodyum ve klorür iyonlarındaki bu artış, sıvı hacmini arttırarak (özellikle kan plazma hacmini), kan hacminde artışa, dolayısıyla kan basıncında yükselmeye neden olur. Kan hacmini azaltan başka nedenlere bağlı olarak (örn. sodyum kaybı, dehidratasyon) kan basıncının düşmesi de, renin anjiyotensin II-aldosteron sistemini devreye sokarak, kan -asıncının korunmasına katkıda bulunur. Ortalama Kan basıncı 70- 160 mmHg arasında geniş oynamalar olsa bile GFO çok az değişir. ANP
Kalbin atriyum hücreleri yüksek kan basınç(baro reseptörein etkisiyle) yanıt olarak olarak atrial natriüretik faktör(peptid) (ANP) üretilir. Böbreklerde mezangiyal hücrelerinde ANP reseptörleri de vardır. ANP, damar genişleticidir ve mezangiyal hücreleri gevşeterek muhtemelen kan akımını ve süzülme için bulunan etkin yüzey alanını artırır. Kan volümü artınca sağ atriyum basıncı artar. Atrium hücrelerinin gerilmesi ANP salgılanmasına neden olur. BNP, kalp, beyin ve diğer organlar tarafından sentezlenen peptid yapısında bir hormondur. Kalp, bu peptititgenellikle kalp yetmezliği tepki olarak, atriyal ve ventriküler diastolinde serbestler Ayrıca sinirsel uyarı ile bu hormon salgılanır. Natriüretik peptidlerin ana fizyolojik etkisi kan hacmi ve sistemik damar direncini azaltılması ve buna bağlı olarak kan basıncını azaltmaktır. Özellikle atriyal miyositlerde tarafından sentezlene ANP 28-amino asit taşıyan bir peptit olarak anjiotensin II uyarılması, endotelin ve sempatik

H A Y V A N F İ Z Y O L O J İ S İ
379
stimulasyon ( beta-adrenoseptörü aracılığıyla) sonucu salgılanır. ANP önce prepro-ANP, olarak sentezlenir ve daha sonra aktif ANP ve biyolojik olarak etkin bir peptit olarak salgılanır. İkinci bir natriüretik peptid ( beyin tipi natriüretik peptid , BNP ) (ilk olarak beyinde tespit edilmiştir, ) büyük ölçüde ventriküller tarafından sentezlenen bir 32 amino asit peptittir. BNP, ilk önce prepro-BN sentezlenir daha sonra aktif olan BNP dönüşür. ANP gibi, BNP aynı mekanizmalarla benzer fizyolojik eylemleri yapar.
ŞEKİL 6.51 Kan hacmi ve basıncı ayarlanması
Atriyumdaki myositlerden salgılanan ANP’nin temel işlevi vazodilatasyondur(damarların gevşemesi). ANP hormonu nörohipofziden ADH salgılanmasını baskılar. Bütün olaylar sonunda su atılımını artar. Ayrıca ANP Aldosteronun distal tübül ve toplayıcı tübüldeki etkisini azaltır. ANP sodyumun vücudtan atılımını artır. Tüm bu olaylar kan basınçının azalmasına bu bağlı olarak atriyumdaki myositlerin daha az ANP salgılamalarına yol açar. ANP kontrolü negatif feedback ile kontrol edilir. ADH
Antidiüretik Hormon (ADH) diğer adlarıi Vasopressin, ve Arginin Vasopressin (AVP)dir. İnsan dahil olmak üzere memelilerin büyük çoğunluğunda bulunan bir hormondur. ADH birincil görevi, böbreklerden su geri emilimini arttırmaktır. ADH, peptid yapısında bir hormondur. Hipotalamusta sentezlenip arka hipofizdeki keseciklerde (veziküllerde) depolanan bir preprohormon öncüsünden sentezlenir. ADH bir kısmı direkt olarak beyne salıverilirken büyük çoğunluğu arka hipofizde, kana salıverilmek üzere depolanır. Vasopressinin en önemli görevlerinden biri vücuttaki su tutulumunu düzenlemektir. Vücut susuz kaldığında (dehidrate olduğunda) ADH salınır ve böbreklerin suyu tutması sağlanır. Böylece hacmi düşük, yoğunluğu yüksek bir idrar oluşturulur. Bunun yanı sıra ADH, orta dereceli bir vazokonstriksiyona sebep olarak kan basıncını da artırır. Ayrıca ADH (özellikle beyinde) nörolojik etkilere sebep olduğu da bilinmektedir. Ayrıca Arginin Vasopressin (AVP)'ye çok benzer bir madde olan lizin vasopressin (LVP) de domuzlarda bulunan, Arginin Vasopressin (AVP) ile aynı görevlere sahip olan bir hormondur ve insanlarda tedavi amacıyla kullanımı yaygındır. Arginin Vasopressin (AVP), böbreklerin temel fonksiyonel birimi olan nefronlardaki distal kıvrımlı tübülün ve toplayıcı tübüllerin suya geçirgenliğini arttırır. Böylece su geri emilimini artırır ve daha yoğun bir idrar oluşmasını sağlar (antidiürez). Bu etki, distal tübüldeki epitel hücrelerinin apikal kısmındaki hücre zarına ilave su kanalları(Aquaporin-2) eklenmesiyle sağlanır. Aquaporin'ler suyun distal kıvrımlı tübül ve toplayıcı tübüllerden hücrelere geçişini sağlayan kanallardır.

H A Y V A N F İ Z Y O L O J İ S İ
380
ŞEKİL 6.52 ADH bağlı geriemilim değişimi
Distal kıvrımlı tübül ve toplayıcı tübül boyunca sıralanan hücrelerin bazolateral kısmında, ADH için aktif bir alan içeren g-protein bağımlı V2 Reseptörleri bulunur. Hücre zarının hücreye bakan kısmında V2 ile temas eden G-proteini(Gs), ATP'yi siklik AMP'ye (cAMP) dönüştüren adenilat siklazı tetikler. Daha sonra cAMP şelalesi, Aquaporin 2 su kanallarının hücre zarına eklenmesini, depo veziküllerinin ekzositozu yoluyla tetikler. Protein kinaz C (PKC) geninin transkripsiyonunu düzenleyen baskılayıcı protein, cAMP için bir yapışma alanı içerir. Bu alana cAMP bağlandığında, düzenleyici proteinin yapısı değişir ve genin operatör bölgesinden ayrılır. Böylece düzenleyici proteinin, Protein Kinaz C (PKC) geninin transkripsiyonu üzerinde baskılayıcı etkisi ortadan kalkmış olur. PKC transkribe olur ve ATP'den fosfat ayrılmasıyla enerji üretimini tetikleyerek, zarlarında Aquaporin-2 kanal proteinlerini içeren veziküllerin hücre zarıyla kaynaşmasını sağlar. Bu süreçte kalsiyum iyonları gereklidir.
ŞEKİL 6.53 Nefronda geri emilim

H A Y V A N F İ Z Y O L O J İ S İ
381
ADH ayrıca nefronlardaki toplayıcı tübüllerin meduller kısmının üreye geçirgenliğini artırarak, üre geri emilimini artırır (nefronun yapısını gösteren şekli inceleyiniz). Böylece üre toplayıcı kanallardan meduller bölüme geçer ve distal kıvrımlı tübül ile toplayıcı tübülün kortikal kısmındaki yükselen su geri emilimi sonucu oluşan yoğunluk farkı dengelenmeye çalışılır. İdrar oluşumu
İdrar, böbreklerden ürinasyon işlemiyle salgılanan ve üretra vasıtasıyla boşaltılan genellikle steril durumda, vücudun yan ürünü olan bir sıvıdır. Asidik kökenli olmayan bu sıvı, çözeltilerini barındıran memeli canlıların vücutlarındaki suyun muhtevasındaki mineral, inorganik ve diğer maddelerin dengesini sağlamak amacıyla idrar torbasından dışarıya çoğunlukla cinsel organlarda varolan mekanizmalar yardımıyla atılır. İdrar oluşumu, vücutta mineral ve diğer maddelerin dengesinin sağlanmasında etkilidir. Vücutta olması gerekenden fazla olan veya vücuda zararlı olan maddeler idrar yolu ile dışarı atılır. İdrar, içinde erimiş ya da süspansiyon durumunda bulunan birçok maddeyi uzaklaştırır. Aynı zamanda idrar (alınan besine bağlı olarak) pH 5.0 – 8.0 değerliği arasındadır. Böbreğin idrar oluşturması için bol sıvı filtirasyonuna ihtiyaç vardır. İdrar ile atılan sıvı iki amaca hizmet eder. Birincisi idrar içinde suyla birlikte atık maddelerin uzaklaştırılması sağlanır. İkincisi idrar temel bileşini olan vucut için fazla olan suyun en hızlı ve büyük uzaklaştırma yoludur. Bu yola başta kan olmak üzere diğer tüm sıvıların hacmin ayarlanması yapılır. Bunun yanında sempatik sinir uyarımı idrara üretimini tamamen engeleyebilir. Antidiüretik hormon (ADH) idrar olarak atılmadan önce böbrek tubular sıvısından yüksek miktarlarda suyun geri emilimine neden olur; böylece idrar miktarı azalırken ekstraselüler sıvı hacmi artar. Aldosteron böbrek tubullerinde su ve tuz geri emilimini artırmakta, böylece bu maddelerin idrarla atılmalarını önlerken ekstraselüler sıvı hacmini çoğaltmaktadır. Anjiotensin II böbrek kapillerinde kasılmaya yol açarak filtrasyon oluşumu azaltır bunu sonucu idrar miktarıda azalır. Kalpin pompaladığı kan miktarının farklı nedenler azalması idrar oluşumu azaltır. Azalmış Glomeruler Filtrasyonu, Renin-Anjiyotensin Sisteminin Aktivasyonu yol açar ve Su ve tuzun Böbrek Tubuluslerinde geri emilimi artar. Reabsorpsiyonu ve son olarak aldosteron Sekresyonu artırarak idrar miktarını etkiler.
ŞEKİL 6.54 İdrar oluşumu

H A Y V A N F İ Z Y O L O J İ S İ
382
İdrar hacmini, daha doğrusu idrar itrah hızını artıran ilaçlara diüretik ilaçlar adı verilir. Bu ilaçlar böbreklerde nefronları ve toplayıcı tübülleri etkilemek suretiyle su reabsorbsiyonunu azaltarak, çıkarılan idrarın hacmini artırırlar. Çoğu diüretik ilaç primer olarak sodyum iyonunun, böbrek tübüllerinde reabsorbsiyonunu azaltırlar. Diüretiklerin bu etkisine natriüretik etki denir. Sonuç olarak, net sodyum ve net su itrahını artırırlar. Temel olarak vücuttan sodyum iyonunun atılmasını artırmak için kullanılmaktadır. Bunun sonucunda su miktarıda azalır. Diüretik ilaçlar etki yerlerine göre şöyle sınıflandırılabilirler: 1) Primer etkileri proksimal tübül üzerine olanlar: Karbonik anhidraz inhibitörleri, osmotik etkili diüretikler, ksantin türevleri. 2) Primer etkileri Henle kıvrımı üzerine olanlar (kıvrım diüretikleri): Furosemid, bumetanid, etakrinik asid, civalı diüretikler. Bunlar en güçlü diüretiklerdir. 3) Primer etkileri distal tübül üzerine olanlar: Tiazid grubu diüretikler ve tiazid-benzeri diüretikler (Klortalidon, klopamid, mefrusid, metazalon)'dur. 4) Primer etkileri kortikal toplayıcı tübüller üzerine olanlar: Aldosteron antagonistleri (sprinolakton ve kanrenon), triamteren ve amilorid'dir. Bunlara potasyum tutucu diüretikler adı verilir. Boşaltım sistemi diğer kısımları İdrar Torbası ve İdrar Yolları
İdrar torbası ve idrar yolları, böbreklerde üretilen idrarı depolayarak dışarı taşır. Kaliksler, renal pelvis, üreter ve idrar torbasının histolojik yapısı aynıdır. İçten dışa doğru; tunika mukoza, tunika muskularis, tunika adventisya tabakalarından oluşur.
ŞEKİL 6.55 İdrarın böbreklerden sonraki yolu sinirsel kontrol.
Üretra, idrarı idrar torbasından dışarıya aktaran bir tüptür. Erkekte ejakülasyon sırasında sperm de buradan geçer. Üretra, kadınlarda tümüyle idrarla ilişkili bir organdır. Üreterlerin duvarları, idrar torbasına doğru gidildikçe kalınlaşır. Bu organların mukozası çok katlı değişken epitel ve gevşekten tıkıza doğru değişen bağ dokusunun yaptığı lamina propreadan oluşur. Bu organların lamina propreasının çevresi yoğun düz kas tabakası ile örülmüştür. Üretra, idrarı idrar torbasından dışarıya aktaran bir tüptür. Erkekte ejakülasyon sırasında sperm de buradan geçer. Üretra, kadınlarda tümüyle idrarla ilişkili bir organdır. Erkekte üretra dört bölümden oluşur. Bunlar prostat üretrası, membranöz üretra ve penis üterasıdır. Prostatın konumu idrar torbasına çok yakındır, üretranın başlangıç bölümü prostatın içinden geçer ve prostat salgısını taşıyan kanallar, üretranın prostatik bölümüne açılır. Prostatik üretra, çok katlı değişken epitel ile döşelidir. Kadınlardaki üretra, arada yalancı çok tabakalı silindirik epitel, yer yer çok katlı keratinsiz yassı epitelle döşeli olan, 45 cm. uzunluğunda bir tüptür. Lamina proprea Littre bezlerini içerir. Dişi üretrasının orta kısmı, dıştan istemli büzücü çizgili kas ile sarılmıştır.

H A Y V A N F İ Z Y O L O J İ S İ
383
Hemodiyaliz
Canlıdaki membranlar bazı maddelere karşı geçirgen iken bazı maddelere karşı geçirgen değildirler. Küçük ve yüksüz maddeler özellikle gazlara karşı membranlar geçirgen büyük ve yüklü moleküller membranda pasif yolla geçişleri yağı eritmeleri yada yağda erimelerine bağlı olarak farklılık gösterir. Bu sebepten daha küçük basit moleküllerin bir membrandan (zardan) geçmesine müsaade etmek suretiyle bunları daha büyük kompleks moleküllerden ayırmak mümkündür. Bu olaya “Dializ” denir. Protein ve tuz ihtiva eden bir çözeltiden tuzun ayrılması dialize güzel bir örnektir. Eğer böyle bir çözelti tuzu geçiren, fakat proteini geçirmeyen torba şeklindeki bir zara konup zar suya daldırılırsa tuz zardan dışarı çıkar, protein ise içeride kalır. Eğer dışarıdaki su sık sık değiştirilirse, nihayet zarın içindeki çözeltide bulunan tuzun hemen hepsi dışarı alınmış olur. Tuzlanan su yerine saf su kondukça tuzun dışarıya çıkışı devam eder. Çünkü, konsantrasyon farkı sıfıra erişemez. Dializde yön, çok yoğundan az yoğuna doğrudur. Hemodiyalizin temel prensibi ince bir membranla bağlı olan çok küçük kan kanallarından kanın sürekli akışıdır. Membranın diğer tarafında kandaki istenmeyen maddelerin difüzyonla geçtiği bir diyaliz sıvısı bulunur. İki ince selofan membran arasından kanın ve membranın dışında da diyaliz sıvısının aktığı bir tip yapay böbrek şeması görülmektedir. Selofan, plazmadaki proteinler hariç diğer maddelerin her iki yönde, plazmadan diyaliz sıvısına veya diyaliz sıvısından plazmaya geçişine imkân verecek düzeyde porlu yapıdadır. Eğer bir maddenin plazmadaki konsantrasyonu diyaliz sıvısındakinden fazla ise bu maddenin plazmadan diyaliz sıvısına net geçişi olur.
ŞEKİL 6.56 Hemodiyaliz
Diyaliz membranında solüt maddelerin geçişi çeşitli faktörlere bağlıdır, (1) iki solüsyon arasındaki solüt madde konsantrasyon farklılığı, (2) membranın solüt maddeye karşı geçirgenliği, (3) membranın yüzey alanı ve (4) membranda kan ve sıvının temasta kaldığı sürenin uzunluğu.
İnsan kan hücreleri bu tarzda araştırılmış ve ozmotik basınçlarının 0oC’de takriben 6.6 atmosfer olduğu bulunmuştur. % 0.9 NaCl ihtiva eden bir eriyik, 0oC’de takriben 6.6 atmosfer ozmotik basınca sahiptir. Bu çözeytiye “İzotonik tuz çözeltisi” veya “fizyolojik tuz çözeltisi” denir. Göz veya burun boşlukları gibi narin membranların ilaçla tedavisinde su yerine fizyolojik tuzlu su kullanılması herhangi bir ağrı duyulmasını önler. Aynı ilaç izotonik çözelti yerine su ile birlikte alınırsa ağrı hissi duyulur. Çünkü çözeltinin temas ettiği narin zararlar zarar görür. Laboratuvarlarda difteri ve tetanoz antitoksinleri, fazla elektrolitlerinden dialize arıtılır.

H A Y V A N F İ Z Y O L O J İ S İ
384
Gaz değişimi ve Solunm
sistemi fizyolojisi
Solunum sistemi gaz değişimi ve organları hakkında bilgiler.
Solunum Fizyolojisi Bir çok canlı yaşamlarını sürdürmek için (gelişebilmek ve üreyebilmek için) enerjiye enerji için besine besini canlının kullanbileceği enerji şekli olan ATP dönüştürmek oksijene ihtiyaç duyarlar. Enerji elde etmek için farklı birçok yol olmasına rahmen (glikoliz, fotosentez, kemosentez vb) Oksijenli solunum birim madde başına en fazla enerji (ATP) elde edilen katabolik yoldur. Moleküllerin yıkımı sonunda açığa çıkan kimyasal bağ enerjisinden yaralanarak ATP sentezler. Canlıların besin maddelerini daha az enerj taşıyan başka moleküllere yıkarak onlardan enerji elde etmelerine solunum denir. Birçok canlıda hücresel solunum gerçekleştirilirken oksijen kullanılmaktadır(örneğin kasılmakta olan kırmzı lifli sarkomer hücresinde). Ayrıca oluşan karbondioksit hücrelerden uzaklaştırılmak zorundadır. Bu olaylar bir hücreli canlılarda hücre yüzeyi ile gerçekleştirilirken çok hücrelilerde hem vücut yüzeyi ile hem de özel bir solunum sistemi ile gerçekleştirilir.
ŞEKİL 7.1 Hücre çevre arasında gaz alış verişi yollar ve gaz alış veriş destekleyen sistemler
Bölüm
7 Bu bölüm için Power
point sunumu adresi:

H A Y V A N F İ Z Y O L O J İ S İ
385
Özel bir solunum sistemi bulunduğu durumlarda solunum sistemiyle kan ve dolaşım sistemi, işbirliği halinde çalışarak solunum görevini gerçekleştirirler. Hücreler içinde oluşan oksijen ye karbon dioksit alışverişine, yani biyolojik yanma olayına iç solunum denir. İç solunumda gaz alışverişi doku sıvısı aracılığı ile kan ve vücut dokuları arasında gerçekleşir. Hücrelerde tüm yaşamsal olayların sürmesi için gerekli enerjiyi sağlayan iç solunumdur. İç solunum da, hücreye gelen enerji yüklü bileşiklerin, özellikle karbonhidrat ve yağların biyolojik yanması ve yüklü oldukları enerjinin yaşam olayları için serbest bir hale sokulması demektir.
Bu sorunlar, solunum için özelleşmiş doku, organ veya sistemlerle giderilmiştir. Solunum sistemlerinin her çeşiti, anlıların yukarıda belirtilen gereksinimlerini karşılayacak şekilde evrimleşmiştir. Direkt ve indirekt solunum
Bir hücrelilerde özel solunum sistemi yoktur. Metabolizma için ihtiyaç duyulan O2 hücre zarında difüzyonla alınır. Canlıda metabolizma sonucu oluşan CO2 yine hücre zarından difüzyonla dış ortama verilir. Paramecium ve hidralar gibi küçük akuatik organizmalar dış ortamla direkt temasta olduklarından, etraflarını çeviren suda bulunan O2 hücrelere girişi ve CO2’in hücrelerden çıkışı kolaylıkla olur. Organizmanın hücreleri ve onun çevresi arasındaki O2 ve CO2 değişimi şeklindeki gaz değişimine direkt solunum denir. Büyük ve kompleks yapıya sahip hayvanlar her hücrenin dış çevreyle direkt gaz değişimi yapması imkansız hale gelir ve gaz değişimi için özelleşmiş bir vücut yapısı gelişmeye başlar. Difüzyon olabilmesi için gaz değişimi yapacak vücut bölgelerinin ince membranlı, geçirgen özelliğe sahip olması, O2 ve CO2 ancak eridikten sonra diffüze oldukları için aynı zamanda iyi bir kan dolaşımına sahip olması gereklidir. Solunum yüzeyleri bu nedenle sürekli nemli tutulması gerekir. İndirekt solunum için balık, yengeç, istakoz gibi birçok aquatik hayvanda solungaçlar gelişmiştir. Yüksek omurgalılarda (reptil, kuş, memelilerde) ise akciğerler gelişmiştir. Kara omurgasızlarından bazıları (toprak solucanı) nemli derilerini kullanırlar. Bazıları (böcekler) dış çevre ile por denen açıklıkla irtibatta olan kanallarını (trake) kullanırlar. Büyük ve kompleks yapıya sahip hayvanlarda difüzyon olabilmesi için gaz değişimi yapacak vücut bölgelerine ihtiyacı vardır. Solunum yüzeyleri ya da solunum membranları denilen bu yapılarda meydana gelen solunuma indirekt solunum denir. İndirekt solunumda iç ve dış solunum fazları görülür
Canlılarda Solunumla İlgili Karşılaşılan Sorunlar 1.Yeterli genişlikte solunum yüzeyi (dış çevreyle gaz değiştirme yüzeyi ) 2.Gaz değişimi yapılan yüzeyden daha içeride yer alan vücudun iç hücrelerine O2 taşınması ve bu hücrelerden CO2’in uzaklaştırılması 3.Solunum yüzeyinin mekanik etkilerden korunması 4.Solunum yüzeyinin aşırı su kaybetmeden nemli tutulması

H A Y V A N F İ Z Y O L O J İ S İ
386
ŞEKİL 7.2 Farklı canlı gruplarındaki solunum yapıları
Küçük ve yassı vücutlu hayvanlarda, difüzyonla yapılan gaz değişimi, canlının hücresel aktivitesini sürdürebilmesi bakımından yeterlidir. Çünkü bunların tüm hücreleri arasındaki mesafe çok kısadır. Oysa daha iri hayvanlarda gaz değişiminin yapılabilmesi ve gazların taşınabilmesi için bir dolaşım sistemine ihtiyaç vardır. Bu sistem her hücrenin etrafında oksijence zengin karbondioksitçe fakir bir ortam sağlar. Canlı bunu, bu gazların değişiminin meydana geldiği deri ile gerçekleştirir. Oksijen difüzyonla deriyi geçtikten sonra deri altındaki kapilerlere girer ve kan yolu ile bütün vücuda yayılır. Halkalı solcanlarda epidermis tabakasından salgılanan kutikula tabakası, vücudun dış yüzeyini tamamen örter. Bu tabaka daima nemlidir ve solunumun vücut yüzeyi ile yapılmasında hayati rol oynar. Gaz değişimi vücut yüzeyi, solungaçlar veya Parapod'lar ile gerçekleştirilir.
Tek hücrelilerde solunum gazlarının hücreye giriş çıkışı, hücre yüzeyinden geçiş (difüzyon) ile sağlanır. Çok hücreli organizmalardan süngerler ve sölenterelerde de, özelleşmiş bir solunum sistemi yoktur. Bunlarda tek hücrelilerde olduğu gibi sudaki erimiş oksijeni vücut yüzeyleri ile alır, CO2 yi de aynı yolla suya bırakılır.

H A Y V A N F İ Z Y O L O J İ S İ
387
ŞEKİL 7.3 Örümceklerdeki kitapsı akçiğerler
Vücut dış yüzeyini örten deri gaz değişimini sağlar. Alınan oksijen iç dokulara difüzyonla ya da kanla taşınır. Toprak solucanlarının tek katlı epitel dokudan ibaret derilerinde bulunan Goblet hücreleri çıkardıkları mukoz salgıyla vücut yüzeyinin devamlı nemli kalmasını sağlarlar.
Trake solunumu
Eklembacaklıların böcekler grubunda trake solunumu görülür. Eklembacaklıların ve özellikle böceklerin büyük bir kısmında her segmentte ‘stigma’ denilenbir çift delikle dışarı açılan ve vücudun içerisine bir ağ gibi girmiş, içerisinde hava bulunan sisteme trake sistemi denir. Solungaçlar ve akciğerler sadece oksijeni rezorbe etmelerine karşın, bu sistemle oksijenin dokulara hatta hücrelere kadar ulaştırılması söz konusudur. Böceklerin karın halkalarından dışarı açılan stigmalardan alınan hava, trake boruları ile tüm vücuda yayılır. Trake boruları helezon şeklindeki kitin halkalarla desteklenmiştir. Trakelerin iç yüzü tek sıralı epitel ile döşenmiştir. Trakeler, içi sıvı dolu trakeol ile sonlanır. Trakeollerin ucu bir sıvıyla doludur.Bu sıvı ,dokular ile borucuklar içindeki O2 ve CO2 diffüzyonunu kolaylaştırır.Gaz alış verişi trakeollerle doku hücreleri arasında olur. Böcekler karın halkalarını sürekli hareket ettirerek stigmalardan hava giriş çıkışını kontrol ederler. Stigmaların açılıp kapanma özelliği su kaybını önleme ve gaz giriş çıkışını kontrol etmesi böceklerin bulunduğu ortama uyumunu (adaptasyon) kolaylaştırmıştır.
ŞEKİL 7.4 Trake solunumu
Örümcek ve akreplerde genellikle karın tarafında bulunan, stigma ile başlayan, borucukların vücut içinde bir kitabın yaprakları gibi dallanmasıyla oluşmuş yapılara kitapsı akçiğer denir. Bu yaprakların üzerinde kan ulaşımını sağlayan sistemler uzanır.

H A Y V A N F İ Z Y O L O J İ S İ
388
Deri solunumu
Terliksi hayvan oksijeni hücre yüzeyinin tamamı ile alır. Aynı şekilde basit yapılı tatlısu polibi gibi çok hücrelilerde veya bazı solucanlarda, deri ile vücuda difüzyonla oksijen alınır. Oksijen, yoğunluk farkı nedeniyle vücuda girerken, aynı nedenle CO2 vücuddan dışarıya verilir. Solunumun bu şeklinin etkinliği azdır, çünkü difüzyonla nakledilecek gazın iletileceği mesafe ancak 1 mm olabilir. Bu nedenle büyük organizmalarda deri solunumu çok azdır. Bunun dışında büyük hayvanların derisi oldukça sert ve gaz geçirgenliği düşüktür. Deri solunumu burada, ilave bir solunum şeklidir. Bununla birlikte bazı durumlarda omurgalı hayvanlarda önemli olabilir. Mesela bir kurbağa kışı hiç hareket etmeden geçirir. Suyun altında olduğundan kurbağa akciğer solunumu yapamaz. Metabolik olayların en düşük düzeyde tutulduğu bu dönemde, deri solunumu ile alınan O2 miktarı, metabolik olayların tamamının durmaması için yeterlidir. Deri solunumunun insan solunumundaki payı ise sadece % 1'dir.
ŞEKİL 7.5 Toprak solcanında deri solunumu
Kurbağa ve semenderlerin erginlerinde esas solunum organı akciğerlerdir. Nemli olan deri gerekli oksijenin %25 inin alınmasını sağlar. Memelilerde de kısmi deri solunumu vardır. Ancak alınan oksijenin oranı çok azdır. (% 1 kadar) Solungaç solunumu
Suda yaşayan hayvanlarda görülür. Kurbağa larvaları, deniz solucanları, bazı yumuşakçalar, kabuklular ve balıklarda bulunur. Solungaçlar suda çözünmüş oksijeni alacak şekilde özelleşmiş, yaprak veya tüy biçimindeki yapılardır. Balıklarda solungaçlar esas solunum organı olup, bazı türlerde deri de solunuma yardımcı olurlar. Solungaçlar, başın her iki yanında derinin içeri doğru kıvrılmasıyla oluşmuşlardır. Bir solungaçta içten dışa doğru; Solungaç dikenleri, Solungaç yayları, Solungaç flamentleri olmak üzere 3 bölüm vardır. Solungaç boşluğu operkulum ile örtülüdür.

H A Y V A N F İ Z Y O L O J İ S İ
389
ŞEKİL 7.6 Solungaç solunumu ile Kurbağalarda buccofarigial solunum
Solungaç solunumu yapan canlılarda içinde çözünmüş oksijen bulunan su,ağzından girip solungaçlardan geçerken gaz alış verişi olur. Bu canlılar suda %2-3 oranında bulunan oksijenden daha fazla yararlanmak için solungaçlarında lif şeklinde dallanmalar yaparak geniş yüzeyler oluşturur.
ŞEKİL 7.7 Solungaç solunumu ile akçiğer solunum karşılaştırılması
Ayrıca solungaç kılcallarında kanın akış yönü ile dışarıda ki suyun akış yönü birbirine zıttır. Bu nedenle solungaçlarda gaz difüzyonu da hızlıdır. Ters akım prensibi olarak bilinen bu mekanizma sayesinde solungaç solunumu yapan canlılar suda ki oksijenin yaklaşık %85’inden faydalanır. Bu sırada oksijen, solungaç epiteline, oradan da kılcallara difüzyonla geçer. Aynı şekilde, kılcallardaki kanda bulunan karbon dioksit, solungaç epiteline, oradan da suya geçer. Böylece oksijeni azalmış ve karbon dioksit içeriği artmış olan su, balığın ağzının kapanıp solungaç kapaklarının açılmasıyla dışarı verilir. Hava kesesi ve akçiğer solunumu
Akciğerlerin ilkel yapısını ilk olarak kemikli balıklarda vardır. Bazı tür balıkların hava kesesinin anterior kısmı özefagus ile bağlantıdır. Kurak mevsimlerde hava kesesi hayvanın oksijen elde etmesine yardım eder. Bu yapı ilkel bir akçiğer olarak düşülenebilir. Kalpten sağ ventrikülde gelen kan pulmoner arter ile akçiğerlere gelir. Akçiğerlerde alveoller üzerinde gaz değişimi yaparak pulmoner venle sol atriuma geri gelir. Pulmoner dolaşım ile canlının ihtiyacı olan gaz değişimi önce kan(özellikle eritreositlerdeki hemoglobinle)ile yapılr.

H A Y V A N F İ Z Y O L O J İ S İ
390
Tüm solunum organların ortak özellikleri: 1-Tüm solunum yüzeylerindeki gaz değişimi difüzyonla olur. 2-Tüm solunum yüzeyleri nemlidir. 3-Tüm omurgalılarda solunum yüzeylerinde bol kılcal damar bulunur. Kılcal damar daha fazla kan ile daha fazla yüzey teması sağlar. 4-Tüm solunum yüzeylerinin etrafı gaz difüzyonunu kolaylaştırmak için yassı hücrelerden yapılmıştır. 5-Karada yaşayan tüm canlıların solunum organları vücut içindedir. Islak olan solunum yüzeyleri sürekli su kaybeder. Bu nedenle daha az su kaybı için solunum organları vücut içine alınmıştır. 6-Tüm solunum yüzeylerinin büyüklüğü canlının enerji ihtiyacı ile doğru orantılıdır. Canlıların metabolik hızlarına uygun oksijen ihtiyacı ve karbondioksit çıkışı olur. Solunum organlarını metabolik hıza uygun gaz değişimi yapabilmelidir.

H A Y V A N F İ Z Y O L O J İ S İ
391
ŞEKİL 7.8 Fizyolist ve fizistom hava keseleri
Hava kesesi kemikli balıkların çoğunda karın boşluğunun üstünde böbreklerin altında bulunur. Kıkırdaklı balıklarda bulunmaz. Balık türlerine göre değişir. Hava kesesinin bir kanal ile sindirim kanalına bağlantılı olduğu balıklara fizostom balıklar(genellikle tatlı su balıkları), bağlantısı olmayan balıklara fizoklist balıklar (genellikle deniz balıkları) denir. Hava kesesi vücut hacminin %4-11 arasındadır ve atmosferde bulunan ancak oranları farklı olan oksijen, azot ve karbondioksit gazı karışımı içerirler. Hava kesesi balıklarda batmazlık sağlayarak, hidrostatik organı olarak görev yapar.
Ters Akım Sistemi:
Ters akım sistemleri birçok canlıda ve sistemde bulunmaktadır. Balıkların solungaçlarında, leyleklerin bacaklarında, memelilerin plasentasında, testisinde veya böbreklerinde ve akciğerlerde ters akım sistemi ile madde yada ısı iletimi yapılır. Bütün bu sistemlerin ortak özeliği madde ve enerji değişimin yüksek bir verimle ve hızlı gerçekleşmesidir. İnsan vücudunda böbrekte vesa recta, barsakta vilüslarda ters akım sistemine sahip kapiller kan damarı vardır. Ters akım sayesinde sürekli gaz alışverişi sağlayan bu sistem, ısıl düzenleme ve ileti bakımından önem taşır. Ters akım madde ve yada enerji bir birlerine göre relatif yoğunluğu yada enerjisi kullanılır.
Hava kesesi Balığın yoğunluğunu, suyun yoğunluğuna göre ayarlar. Balık suda batmadan durmak için, içindeki gazı artırarak keseyi şişirir. Yüzerken havasını azaltır. Bazı balıklarda yüzme kesesi ikiye ayrılmıştır. Yüzme kesesi solunum, hidrostatik görev, ses meydana getirme ve bazı uyartıları hissetmede de etkilidir.

H A Y V A N F İ Z Y O L O J İ S İ
392
Ters akım madde ve enerji değişimde etkinliği attırır. Aynı yönde akan sıvılar içinde madde ve enerji değişim yüzde %50 sınırın kadar yapılabilir (Konsantaston yada sıcaklığın yarı değerine kadar değişim). Ters akımla yalnızca yapılan bir değişimle madde ve enerji aktarımda verim çok yüksek seviyelere getirilmiş olur.
ŞEKİL 7.9 Ters akım
Kuşlarda solunum
Kuşların solunum sistemi, yüksek metabolizma hızına, uçma nedeniyle yüksek enerji harcanmasına ve büyük yükseltilerde yeterli oksijen sağlanmasına gereksinmesine adapte olmuştur.
ŞEKİL 7.10 Akçiğerin farklı omurgalı gruplarındaki durumu
Ters akımla etkin
solunum Balıklarda ters akım prensibi görülür. Solungaç kılcallarından akan kanın akım yönü ile solungaçlardan geçen suyun akım yönü birbirine zıttır.Böylece suyun oksijeninden maksimum derecede faydalanır.

H A Y V A N F İ Z Y O L O J İ S İ
393
Kuşlarda solunum sistemi, iki akciğer ile iki uzun hava kesesinden oluşur. Göğüs ve karın boşluklarının büyük bir bölümünü dolduran hava kesecikleri, kemiklerdeki akciğerlerden daha büyük bir hacim oluşturan yardımcı hava boşluklarına bağlıdır. Kuşların, iki ana bronşa ayrılan soluk borusu, taze havayı akciğerlerden iki karın keseciğine iletir. Öbür hava keseciklerine havayı, ikincil bronşlara taşırlar. Hava daha sonra akciğer dokusuna geçerek, damar bakımından son derece zengin çeperli küçük hava kanallarının oluşturduğu sık bir ağ halindeki hava kılcal damarlarında son bulan üçüncül bronşlar tarafından taşınır. Hava kılcal damarları, memelilerdeki alveoller gibi işlev görür: Her ikisi de gaz alışverişini gerçekleştirdiği yerdir. Oksijen karbondioksit değişimi yapmış hava; akciğerlerden ön hava keseciklerin içine dolar ve soluk borusu aracılığıyla dışarı atılır. Taze hava, hem soluk alma sırasında, hem de soluk verme sırasında hava kılcal damarlarından geçer. Soluk alma sırasında, solunum kasları göğüs – karın boşluğunu genişleterek, bütün hava keseciklerinin içindeki basıncı düşürür. Taze hava, ön kesecikler dışında, bütün hava keselerine girer. O sırada bir miktar hava akciğerlere girerken, akciğerlerdeki bayat hava da , ön keseciklere geçer. Soluk verme göğüs – karın boşluğundaki hava basıncını yükselten solunum kaslarıyla gerçekleştirilir. Kullanılan hava dışarı ve taze hava, hava keseciklerinden arka hava kesecikleri yoluyla akciğerlere iletilir. Kuşlarda bu sürece, başından sonuna kadar kas tabakası yerine, bağdokusundan yapılma ince bir zar olan diyafram yardımcı olur.
ŞEKİL 7.11 Kuşlarda solunumu
Diyafram, göğüs çeperine yapışan kaslara bağlanır ve bu kasların kasılmasıyla düzleşir. Kuşlarda diyafram, soluk alma sırasında akciğer hacmini azaltırken, soluk verme sırasında genişletir. Kuşların akciğerleri, memelilerinkine oranla daha katı olduğundan, soluk alma ile soluk verme sırasında, akciğer hacminde az bir değişme olur. Buna karşılık, hava kesecikleri solunum sırasında önemli miktarda şişer ve söner. Uçuş sırasında, kuş kanat çırparken soluk alma kanatlar yukarı kalkarken, soluk vermeyse kanatlar aşağı inerken olur. Kanatların inmesi göğüs kafesini bastırır ve bayatlamış havanın, ön keseciklerinden soluk borusu aracılığıyla dışarı atılmasını sağlar.

H A Y V A N F İ Z Y O L O J İ S İ
394
İnsanda Solunum Sistemini
Hava ile solunum yapan omurgalılarda ve insanda solunum sistemi akciğerler ile havayı onlara getiren tüplerden oluşur. Solunum sistemi burunla başlar.
ŞEKİL 7.12 İnsada solunum yolu
Burun içerisinde konhe adı verilen yapılar vardır. Bu sayede hava, nemlendirilir, ısıtılır ve 5µm üzeri partiküllerden temizlenir. Fariks solunum sistemi ile sindirim sistemini birbirinden ayıran bölümdür. Reflex ile kapanır. Farins’in üst bölümü (nazofarinx) yumuşak damakla ağız boşluğu ve burun boşluğunu birbirinden ayırır. Alt bölümü (laringofarinx) ise trakea ve özefagusla bağlantı yapar. Gırtlak (Larinx), soluk borusu (trakea) ve akciğer (pulmo) alt solunum yollarını oluşturur. Solunum yolu aynı zamanda ses organıdır. Yapısında birbirine kas ve zarlarla bağlı olan kıkırdaklar bulunur. Bu nedenle gırtlak devamlı açık ve hava geçişine izin verilir. Gırtlağın yapısında birçok kıkırdak olup bunlardan tek olan kıkırdaklar daha büyük ve önemlidir. Bunlar yukarıdan aşağı doğru şu şekildedir. Trakea yaklaşık 2,5 cm genişliğinde 10-12 cm boyundadır. Kıkırdak halkalardan yapılmıştır. Sayıları 16–20 arasında değişir. Trakea sağ ve sol 2 tane ana bronşa ayrılır. Bir bronş sağ bir bronş sol akciğere gider. Bronşların ince dallarına bronşiol denir. Akciğerler (Pulmo) Solunum sisteminin oksijen ve karbondioksit değişiminin yapıldığı yerdir. Akciğerler costalar tarafından korunan hafif süngerimsi yumuşak elastik ve hassas bir organdır. Akciğerin uç kısmına akciğer tepesi (apex pulmonis), aşağıda geniş olan bölümüne ise akciğer tabanı(basis pulmonis) denir. Solunum, atmosferden alınan oksijen ile vücuttaki karbondioksitin yer değiştirmesidir. İki kısım oluşur: Dış solunum (external solunum) akciğerlerde olur. Oksijen havadan kana geçer, kandaki karbondioksit dışarı verilir. İç solunum (internal solunum), kanla dokular arasında olur. Oksijen kılcal damarlardaki kandan dokuya girer, karbondioksit dokudan kana geçer. Akciğerin üzerini 2
� Burun � Yutak � Gırtlak � Soluk borusu � Trake � Trakeoitler � Alveoller

H A Y V A N F İ Z Y O L O J İ S İ
395
katlı plevra zarları örter. Akciğerin dış yüzeyini saran tabakasına visseral plevra, göğüs kafesinin iç yüzündeki tabakasına ise parietal plevra denir. 2 katlı zar arasında lenf sıvısı ve hava bulunur. Oksijen içeren hava iletici hava yollarıyla akciğerlere ulaşır. Farklı dokulardan sağ kalbe gelen kan sağ ventrikül tarafından akciğerlere pompalanır. Pulmoner kapillerde karbondioksit ve oksijen değişimi olur. Akciğerleri terk eden kanın oksijen içeriği yüksek, karbondioksit içeriği düşüktür. Sol ventrikül tarafından dokulara pompalanır. Soluk alıp verme
• İnspiryum (soluk alma) : İnspiratuar kaslara uyarı gider. Diyafragma (eksternal interkostal kaslar) kasılır. Göğüs duvarının genişlemesiyle toraksın hacmi artar. İntraplevral basınç daha da negatifleşir. Alveoler transmural basınç gradyenti artar. Alveoller genişler. Bu durumda alveoler geri çekimi artar. Alveoler basınç, alveol hacminin artmasıyla birlikte atmosferik basıncın altına düşer ve dışarıdan içeri doğru hava akımı oluşur. Hava akımı alveoler basınç ile atmosferik basınç arası denge oluşana kadar devam eder.
ŞEKİL 7.13 Akçiğerlerde gaz alış verişi için gerekli olan basınç değişimleri modeli
• Ekspiryum(soluk verme) İnspiratuar uyarı sona erer. İnspiratuar kaslar gevşer. Toraks hacmi azalır ve intraplevral basınç daha az negatif olur. Alveoler transmural basınç gradyenti azalır. Artan alveoler geri çekiminin etkisiyle alveoller inspiryum öncesi durumlarına geri dönerler. Alveoler hacim azalınca alveoler basınç atmosferik basınçtan daha yüksek hale gelir. Bunun sonucunda hava akımı oluşur. Hava, alveoler basınç ile atmosferik basınç dengelenene kadar dışarı doğru akar. Oksijen içeren hava iletici hava yollarıyla akciğerlere ulaşır. Farklı dokulardan sağ kalbe gelen kan sağ ventrikül tarafından akciğerlere pompalanır. Pulmoner kapillerde karbondioksit ve oksijen değişimi olur. Akciğerleri terk eden kanın oksijen içeriği yüksek, karbondioksit içeriği düşüktür. Sol ventrikül tarafından dokulara pompalanır.

H A Y V A N F İ Z Y O L O J İ S İ
396
Alveollerindeki yüzey gerilimi
Suda veya sulu bir çözeltide çözündüğünde yüzey gerilimini etkileyen (çoğunlukla azaltan) kimyasal bileşiklere Surface active agent ya da yüzey aktif madde denir. Alveolde tip II hücrelerince üretilen yüzey gerilimini düşüren maddelere sürfektan denir. Bu sayede alveollerin büzülmesi ve alveol içerisine kan sıvısının dolması önlenir. Lipit, protein ve karbonhidrat karışımı olan bu madde Lesitin ve Sifingomiyelindir .
ŞEKİL 7.14 Sülfaktan madde etkisi
Alveolerde yüzey gerilimi azaltan maddelerin temel olarak iki fonksiyonu vardır:
1-Çok ince olan alveoli (1 mikron)duvarı yüzey gerilimi( su molleküllerin hidrojen bağları nedeniyle bir birine bağlanma durumu) nedeniyle kollabe( büzülmesini) önlemek
2-Alveollerin iç yüzünde gerilimi azaltarak, kan plazmasının alveoli boşluğuna doğru sözülmesi gereğinden fazla sızmasınasını engelemek. Tip II alveollerden salgılınan sülfaktan madde eksikliğinde alveolar kapiller damardan gelen plazma sıvısı ile dolar.
Sülfaktan maddeler yüzey gerimini önemli derecede azaltan yüzey aktif bir ajandır. Sülfaktan maddeler alveol yüzeyinde bulunan Tip II alveolar epitel hücrelerinden salgılanırlar. Tip II alveolar tüm alveol alanındaki hücrelerin %10’u kadarını oluşturur. Sülfaktan Fosfolipidler, protein ve iyonlar içeren kompleks bir karışımdır. Bileşiminde dipalmitolfosfotidilkolin, surfaktan apoproteinleri ve Ca+2iyonları vardır. Sülfaktan maddelerin hidrofilik ve hidrofobik kısımları vardır. Sülfaktan madde eksikliğinde Hiyelin Membran veya Atelectasis denen bozukluk ortaya çıkar.
Mekanik olarak alveollerin birbirine bağımlığı vardır. Bir neden dolayı bie alveol kapanmaya başlarsa, komşu alveollerin duvarındaki stresi arttırır ve onlar bu kapanan alveolü açık tutmaya çalışırlar.
Solunum pigmentleri
Oksijen taşıyan başlıca solunum pigmentleri Hemoglobin, Hemosiyanin, Klorokruorin, Hemoeritrin dir. Solunum pigmentlerinin ortak özellikleri, proteinden yapılmış olmalarıdır. Bu pigmentlerin en önemli özellikleri oksijenle ve karbon dioksitle kolayca birleşip ayrılabilmeleridir. Oksijenle birleştikleri bölgelerde demir, bakır gibi metal iyonları bulunur. Solunum pigmentleri kanın oksijen taşıma kapasitesini artırır.

H A Y V A N F İ Z Y O L O J İ S İ
397
ŞEKİL 7.15 Solunum pigmentleri
Omurgalıların çoğunluğunda, klorokruorinler (demirli porfirinle bileşmiş bir pigment), hemoeritrinler (demirli, ama porfirinle bileşmiş olmayan pigment) ya da hemosiyanin (bakırlı bir solunum pigmenti) içeren dolaşım sıvıları bulunur. Bütün bu pigmentler, dolaşımdaki sıvının oksijen taşıma yeteneğini artırır. Pigment Renk Element Konum Hayvan 100 ml kanda O2 ml miktarı
Hemoglobin KırmızıDemir Alyuvarlar Memeli 25 Kuşlar 18,5 Sürüngenler 9 Kurbağa 12 Balık 9
Plazma Halkalı solcan 1,5 Yumşakça 2-8
Hemosiyanin Mavi Bakır Plazma Yumşakça 2-8 Klorokruorin Yeşil Demir Plazma Halkalı solcan 9 Hemoeritrin KırmızıDemir Kan hücre. Halkalı solcan 2 Genel olarak, omurgalılarda kan, son derece etkili bir oksijen taşıma aracısı olan ve bir proteine (globin) bağlı bir demir-porfirinden(heme) oluşan hemoglobin içerir. Bazı omurgasızlarda da hemoglobin bulunmakla birlikte, bu hemoglobin genellikle dolaşım ya da sölom sıvısında çözünmüş durumdadır. Yüksek derecede gelişmiş omurgasızlarda (derisi-dikenliler ve daha yüksek omurgasızlar) hemoglobin, özel kan hücreleri içinde bulunur. Omurgalıların tümünde, bu tür hücreler içinde hemoglobin vardır. Balıklarda solungaç bulunduğu halde dolaşım bu genel yapıya uyar. Akciğer ve Dokularda Gaz Değişimi
Akçiğerlerin temel fonkisyonu metabolik ihtiyacı karşılayacak oksijeni sağlamak, sistemik venöz kanla akciğere getirilen karbondioksiti uzaklaştırmak, normal kan gazı homeostazını sürdürmektir. Solunum sisteminin görevi, vücudun gereksinimine göre dış ortamla gaz alışverişini sağlamak, dolaşım sistemi aracılığıyla da solunumu düzenlemektir. Solunum dört evreden oluşur:
1-Ventilasyon 2- Diffüzyon 3- Perfüzyon 4- Solunumun düzenlenmesi

H A Y V A N F İ Z Y O L O J İ S İ
398
ŞEKİL 7.16 Alveoller
Alveollerin taze hava ile havalandırılmasından sonra, solunum sürecinin ikinci adımı, oksijenin alveollerden pulmoner kana ve karbon dioksidin zıt yönde difüzyonudur. Difüzyon moleküllerin solunum membranından her iki yöne olan rastlantısal hareketlerdir. Alveoller Akciğerlerin fonksiyonel birimleri olan alveoller, küçük ve içi hava dolu keseciklerdir. Alveollerde difüzyona uğrayan tüm gazlar geçmesi gereken tapakalar:
1. Alveolü kaplayan sıvı tabakası; bu tabaka, sıvının yüzey gerilimini azaltan
süıfaktanı da içerir.
2. İnce epiteliyal hücrelerden oluşan alveol epiteli 3. Epitel bazal membranı
4. Alveol epiteli ile kapiller membran arasında kalan ince bir interstisyel boşluk.
5. Birçok yerinde epitel bazal membranı ile kaynaşmış kapiller bazal membranı 6. Kapiller endotel membranı.
ŞEKİL 7.17 Alelerde gaz geçişinde katmanlar

H A Y V A N F İ Z Y O L O J İ S İ
399
Alveolerin görünüşü üzüm salkımına benzer. Çevresinde pulmoner kapiller ağı vardır. Pulmoner kapillerlerin ortalama çapı 5 mikrometre olduğundan eritrositlerin sıkışarak geçmesi gerekir. Bu nedenle eritrosit membranı genellikle kapiller duvarına değdiğinden oksijen ve karbondioksit kolaylıkla difüze olur.
ŞEKİL 7.18 Erirosit gaz değişimi ve gaz değişim
Akciğerlerden dokulara taşınan oksijenin %97’si eritrosit içinde hemoglobinle kimyasal bileşik halindedir. %3’ü ise plazma ve hücre sıvısında çözünmüş durumda taşınır. Dinlenim durumunda, arteriyel kanda hemoglobin %98 oranında doymuştur. Toplam O2 içeriği ~20 ml.dl-1’dir. Bu miktarın 0.3 ml’si çözünmüş olarak plazmada, 19.7 ml’si ise hb’ne bağlı olarak taşınır. Venöz kanda hemoglobin %75 oranında doymuştur ve toplam O2 içeriği ~15.2 ml.dl-1’dir. Bu miktarın 0.12 ml.dl-1’si çözünmüş olarak plazmada, 15.1 ml.dl-1’si hemoglobine bağlı olarak taşınır.

H A Y V A N F İ Z Y O L O J İ S İ
400
ŞEKİL 7.19 İç ve dış solunum
Alveoller
Bir alveol çapı 0.3 mm kadardır. Solunum yüzey kalınlığı 0,2-0,6 mikrometredir. Solunum alanı yaklaşık 70 m2 dir. Solunum sırasında anlık bulunan kan miktarı 60-140 ml’dir. Bu kan 70 m2 lik bir yüzeye yayılmaktadır. Alveol duvarında hasara duyarlı Tip 1 epitel ve bölünüp dönüşebilen Tip 2 granuler hücreler, bazal membran yer alır. Alveollerdeki Proteoglikan, elastik ve kollajen lif içeren interstisyumlar esneyebilme ve geri toparlanmada rol oynar. Normalde alveol duvarı ile kapiller duvarı arasında 0.5-1 mikron mesafe vardır ve gazların değişimi pasif difüzyonla olmaktadır. Alveollere gelen hava solunum yollaraını izler. Bir gazın sıvıda çözünen miktarı onun parsiyel basıncı ile doğru orantlıdır. Normal hava: %20.9 O2 , %0.03 CO2 bulunur. Atmosfer basınçı : 760mmHg deniz kenarı ( yüseklik arttıkça basınç Patm düşer.) kadardır. Bu neden solunum yoluyla alveolere gelen solunan havası 37 CO, %100 nemli ortamda, su buharı parsiyel basıncı: 47mmHg.kadar olur. Bu nedenle solunan hava PinsO2: (760-47)x0.21: 150mmHg kadar azalır. Alveol içi PalvO2: PinsO2 – P Alv CO2 :150 – PaCO2/0.8:150-50:100 mmHg.( alveol içi gaz eşitlenmesi formulü) . Alveol içinde 100 mmHg O2 ve 50 mmHg CO2 mevcutdur. Gazların difüzyon hızlarının farklı olmasından dolayı CO2, O2 den 20 kat fazla diffüze olabilir. Normalde PAlvO2 ile PaO2 arasında 10mmHg fark vardır. Bu fark venöz karışım ve normal intrapulmoner şantlar nedeniyle olmaktadır. Atmosfer havası (1 Atm=760 mmHg)
Alveol havası Ekspirasyon havası

H A Y V A N F İ Z Y O L O J İ S İ
401
% 78,62 N2 (597 mmHg) % 20,84 O2 (159 mmHg) % 0,04 CO2 (0,3 mmHg) % 0,50 H2O (3,7 mmHg)
% 74,9 N2 (569 mmHg) % 13,6 O2 (104 mmHg) % 5,3 CO2 (40 mmHg) % 6,2 H2O (47,0 mmHg)
% 74,5 N2 (566 mmHg) % 15,7 O2 (120 mmHg) % 3,6 CO2 (27 mmHg) % 6,2 H2O (47,0 mmHg)
Oksijenin taşınması
Hayvanların kanında farklı tipte O2 taşıyıcı solunum pigmentleri (Hemoglobin, Hemosiyanin, Klorokruorin, Hemoeritrin) bulunur. Doğum öncesi kanda bulunan Fetal hemoglobin (hb F)’ de 2,3 DPG gama polipeptid zincirleri ile zayıf bağlandığı için O2’ne ilgi daha fazladır İnsanda alveollerde oksijene doğmuş olan eritrositler oksijeni taşır. Oksijen kanda eritrosit içinde oksihemoglobin halinde bulunur. Hemoglobin kanda solunum organından dokulara oksijen, dokulardan solunum organına ise karbondioksit ve proton taşıyan proteindir. Oksijenle dolu olan hemoglobine “oksihemoglobin” denir. Bu, kana parlak kırmızı rengini verir. Dokulara oksijen getirdikten sonra bir miktar karbondioksiti alarak akciğerlere getirir. Buna da “karbaminohemoglobin” denir. Başlıca sentez yeri eritrosit üretimi sırasında kemik iliğidir. Oksijenin çok az bir kısmı kan plazmasında çözünmüş olarak taşınır. (% 2 kadar). Akciğerlerde kana geçen O2, alyuvarlardaki hemoglobinle birleşip oksihemoglobini oluşturur.
Hb + O2 ����HbO2 (Oksihemoglobin) Oksijen için difüzyon kapasitesi 21 ml/dak/mmHg’dir. Fonksiyonel açıdan solunum membranının iki tarafı arasında ortalama 11 mmHg oksijen basınç farkı vardır. Bu basıncın difüzyon kapasitesi ile çarpımı (11x21) solunum membranından bir dakikada difüzyona uğrayan 230 ml oksijen hacmini verir. Bu vücudun oksijen kullanma hızıdır. Doku kılcallarında hemoglobinden ayrılıp doku sıvısına, oradan da difüzyonla hücrelere geçer. Pulmoner kapillerlerin ortalama çapı 5 mikrometredir. Eritrositler sıkışarak geçer. Eritrosit membranı genellikle kapiller duvara değdiğinden oksijen ve karbon dioksitin difüzyon hızını yavaşlatan plazmadan geçmesi gerekmez. Hemoglobin-O2 disosiyasyon eğrisi: Oksijen basınçı (PO2) ile hemoglobinin O2 taşıma gücü (% doygunluğu) arasındaki ilişkiyi gösteren eğri “hemoglobin- O2 (HbO2) disosiyasyon eğrisi” olarak adlandırılır. Hemoglobin oksijen disosiyasyon eğrisi farklı faktörlere bağlıdır Egzersizde O2-Hb eğrisinin sağa kayması neden olan: Egzersiz yapan kaslarda CO2 ve asidik ürünlerin oluşumu, Kas kapillerlerinde H+ düzeyinin çoğalması, Kas ısısında yükselmesidir. “Bohr etkisi” Kanda PCO2 ve H
+ düzeyinin azalmasının akciğerlerde hemoglobinin O2’lenmesini arttırmasıdır. Hemoglobinin oksijene afinitesinin artması (Dissosiasyon eğrisinin sola kayması) yol açana başlıca faktörler: Alkalozis, Eritrosit içi 2, 3 - DPG’nin azalması, Isının azalması, Karboksihemoglobin artması, Methemoglobinemi ve pCO2’nin azalmasıyla meydana gelir. Hemoglobinin oksijene afinitesinin azalması: (Eğrinin sağa kayması) ise Asidoz, Eritrosit içi 2, 3 - DPG artması (Yüksek irtifa, Tiroid hormonları, Anemi, androjenler), Isının artması, Hemoglobinopatiler (Orak hücre anemisi) PCO2 nin artması sonucu olur.

H A Y V A N F İ Z Y O L O J İ S İ
402
ŞEKİL 7.20 2,3 Difosfogliserat
Eritrositlerde Hb molekülü sayısı kadar 2,3-DPG(2,3 difosfo glisarat) molekülü bulunmaktadır.
Her nefesle 350–500 ml taze hava gelir. %21 oksijen içerir. %5-6 karbondioksit içeren 350-500 ml hava ekspiryum ile atılır. Karbonik anhidrazın katalizlediği temel enzimatik olay karbondioksitin (CO2) hidrasyonudur.
CO2 + H2O --> HCO3- + H+
Kinetik çalışmalar bütün karbonik anhidraz izoenzimlerinde iki basamaklı bir mekanizmanın olduğunu göstermiştir. İlk basamakta çinkoya bağlı hidroksit iyonunun CO2’e nükleofilik saldırısı gerçekleşmektedir. İkinci basamakta ise çinkoya bağlı su molekülünün iyonlaşması ve protonun uzaklaştırılmasıyla aktif bölgenin tekrar oluşturulması olayları gerçekleşmektedir.
ŞEKİL 7.21 Dokuda gaz değişimi

H A Y V A N F İ Z Y O L O J İ S İ
403
Karbondioksit taşınması
Kanda karbondioksit üç farklı yollarla taşınır. Bikarbonat iyonu halinde (%70), Hb’ne bağlı Kanda karbondioksit olarak (%23) ve son olarak Plazmada erimiş halde (%7). Alyuvarlarda CO2 ve H2O’nun karbonik anhidrazın (CA) etkisi altında geri dönüşümlü birleşmeleri sonucu H2CO3 üzerinden oluşan HCO3
- şeklinde taşınır. CO2 hemoglobinle karbaminohemoglobin [HbCO2] bileşiğini oluşturmak üzere (geri dönüşümlü) birleşir. Karbondioksit kısmi basınçı PCO2’nin doku kapillerlerine göre daha düşük olduğu alveollerde Hemoglobinden CO2’i kolayca bırakır. Klor “kayması” Doku kapillerinde eritrosit içinde oluşan HCO3
-, taşıyıcı bir protein aracılığı ile plazmaya geçirilirken aynı miktarda klor iyonuda diğer yönde taşınır(bant 3 proteinleri). Pulmoner kapillerlerde olay ters yönde gerçekleşir (tersine klor kayması). Kısaca plazmada HCO3
− konsantrasyonu artınca klorür kayması diye tanımlanan olayla klorür iyonu eritrositlerin içine kaçar. Plazmada bikarbonat konsantrasyonu azalınca da klorür iyonu plazmaya geri döner. Eritrositlerdeki anyon(klor iyonu) transport proteinleri Band 3 proteinlerdir. Deoksihb’nin H+ affinitesi oksihb’den çok daha fazladır. Dokuda oluşan deoksihb H+ iyonlarının büyük çoğunluğunu bağlar (HbH). “Haldane etkisi” Dokularda, hemoglobinin O2 doygunluğunun (HbO2 miktarının) azalması ile kanda CO2 taşıma yeteneğinin artmasıdır.
ŞEKİL 7.22 Haldene etkisi
Halden etkisi: Pulmoner düzeyde(alveollerin yüzeyinde) hemoglobine O2 bağlanması, HbCO2 ayrışmasını kolaylaştırır. pH düştükçe, asitlik arttıkca HCO3'ler H2CO3 e dönüşür. H2CO3 ise karbonik anhidraz tarafından H2O ve CO2 olarak ayrışır. Bu sayede kandaki CO2 alveollere geçer.

H A Y V A N F İ Z Y O L O J İ S İ
404
ŞEKİL 7.23 Alveolerde gaz değişmi
Respiratuvar değişim oranı (RER, R) Ağızdan atılan CO2 miktarının, alınan O2 düzeyine oranlanmasına respiratuvar değişim oranı denir. (VCO2/VO2) Normalde 0.825 kabul edilen bu oran metabolizma değişiklikleri hakkında bilgi verir (örn: karbohidratlar için 1,0, yağlar için 0,7). Respiratuvar değişim oranı ölçme solunum için kullanılan yakıta ( karbonhidrat ya da yağ) bağlı olarak değiştiğinden vücut tarafından tedarik metabolize olan organik maddelerini tahmin etmek için de kullanılabilir. RER modern bir diyet ile istirahatte yaklaşık 0.8 olarak ölçülür. Çalışan kaslara CO2 üretimi artar ve yoğun egzersiz sırasında 1 aşabilir. Orta ya da yüksek yoğunlukta kas faaliyeti sırasında RQ tahmin RER'ye kullanarak aerobik egzersiz yada anaerobik egzersiz yapıldığı belirlebilir. RER değeri 0.70 yağ baskın yakıt kaynağını, 0.85 RER değeri ise yağ ve karbonhidrat olan bir karışımını göstermektedir ve 1.00 yukarıdasındaki bir değeri karbonhidrat baskın yakıt kaynağı olduğu bir göstergesidir . Solunum hızı kontrolü
Solunum hızı kandaki CO2 miktarına göre düzenlenir. CO2 artışı soluk alıp vermeyi hızlandırır. Çünkü CO2 kanın pH sını düşürür ve ortam asit hale gelir Bu da beyni uyarır. Soluk alış verişinin hızı ve şiddeti omurilik soğanındaki sinirler tarafından denetlenir. Soluk alma hızı ve derinliğine göre farklı tip solunumlar vardır. Eupnea: İstrahat halindeki solunum, Hyperpnea: Frekans ve derinliği artmış solunum, Polypnea: Yüzeysel, çabuk ve kesik kesik solunum, Apnea: Solunumun geçici olarak durması, Dyspnea: Solunum güçlüğü olarak tanımlanır.

H A Y V A N F İ Z Y O L O J İ S İ
405
ŞEKİL 7.24 Solunum kontrolü salanayan reseptörlerin yerleri
Solunumun istemli kontrolu: Serebral motor korteks bağlı merkezlerce yapılır. Limbik ön beyin ağrı, dokunma, sıcaklık, koku, görsel bilgi, heyecan ile etkilenir. Bu durumlara göre solunum ayarlanması sağlanır. Kortikobulbar ve kortikospinal yol ile doğrudan solunum kaslarına emreder. Bilinçle denetlenir istemlidir. Fizyopatolojik değişimlere uygun yanıtını oluşturulmasını sağlar. Aspirasyondan, zararlı gaz inhalasyonundan korunma sağlanır.
Solunumun otomatik kontrolu: Temel olarak beyin sapında yerleşik merkezlerce kontrol edilir. Nörotransmiter maddeler ile (P maddesi, Ach, glutamat, opioid) solunum değiştirilir. Bu maddelerin türüne göre uyarıcı, engelleyici ya da değiştirici etkiler oluşabilir. Apnöstik merkez: alt ponsta yer alır. Solunumu iç çekmeler şeklinde derin inspiryum neden olur. Pnömotaksik merkez: üst ponsta yer alır. İnhibitor olarak görev yapar.
Solunum kontrolünde görev alan reseptörler: Solunum sistemin kontrolün görev alan reseptörler hem santral sinir sisteminde hemde diğer organlarda konumlanmıştır.
� Santaral kemoreseptörler: arka hipotalamus, serebellum, beyin sapı çekirdekleri, locus ceruleus vücut sıvılarının kimyasal bileşenlerinden etkilenir.
BOS PH: 7.32 CO2↑, H+↑, O2↓
� Periferik kemoreseptörler: karotid ve aortik cisimcikler: hızlı etkidir. Glomus karotikum, iki taraflı olarak karotis arterlerin çatallanma bölgesinde bulunur. Glomus aortikumlar arkus aorta boyunca sıralanmıştır. Buradan kaynaklanan sinyaller vagus siniri ile aynı merkeze ulaşır. Santral kemoreseptörlerin aksine periferal kemoreseptörler arteriyel kanla doğrudan temas halindedir. Periferal kemoreseptörler en çok arteriyel PO2 'na hassastırlar. Arteriyel PO2 azaldığında kemoreseptör etkinlik önce yavaş artar, ancak PO2 'nın daha düşük değerlerinde artış hızlıdır. Arteriyel PO2 azaldığında aortik ve karotid cisme ait sinirler artan uyarıları beyin sapındaki solunum merkezlerine iletir. Solunum merkezleri bu reseptörlerin uyarılarına göre solunum sıklığını ve tidal hacmi arttırarak cevap verirler.
PaO2↓,CO↓,oksijen kullanılamaması> PCO2 ↑,PH↓
Solunum Hızını Etkileyen Faktörler 1.Sinir impusları 2.Kaburga kaslarının kasılıp gevşemesi 3.Diyaframın Kasılıp gevşemesi 4.Akciğerde ki basınç azlığı ve fazlalığı 5.Kanda ki CO2 konsantrasyonu (CO2 artarsa asitlik artar ve solunum hızlanır)

H A Y V A N F İ Z Y O L O J İ S İ
406
Solunum hızının arışı ve tidal volum artışına yol açar, vazokonstriksiyon, ADH salgılanmasına artış ↑, surrenal uyarı artışa, bradikardi neden olur. Bu reseptörlerin adaptasyon ve tolerans yoktur.
Sinir sistemi alveolar ventilasyon hızını en ağır egzersizde bile kanın arteryel oksijen (PO2) ve karbondioksit basıncı (PCO2) hiç değişmeyecek şekilde ayarlar. Solunum hızının vücudun ihtiyacına göre düzenlenmesini sağlayan solunum merkezi alt beyinde bir dizi nöron gruplarından oluşmaktadır. Ancak, periferden (akciğerler, kemoreseptörler) ve merkezi sinir sisteminin diğer bölgelerinden gelen uyarıların etkisiyle solunum merkezinin faaliyeti değiştirilebilir. Solunum merkezindeki solunum nöronları iki gruba ayrılırlar. Dorsal solunum grubu nöronları, medullanın dorsaline (arka, sırt tarafına) yerleşmiştir, başlıca inspirasyondan sorumludurlar. Ventral solunum grubu nöronları ise medullanın ventraline (ön tarafına) yerleşmiştir. Buradaki nöronlar vücudun ihtiyacına göre inspirasyon ve ekspirasyondan sorumlu olabilirler.
ŞEKİL 7.25 Solunum kontrolü
Solunumun kimyasal düzenlenmesi: Solunumun ana hedefi, vücut sıvılarında O2, CO2 ve H+ iyon düzeylerini uygun düzeyde devam ettirmektir. CO2 veya H
+ iyonlarının fazlalığı doğrudan solunum merkezini uyarır. Sonuçta solunum kaslarına giden hem inspirasyon hem de ekspirasyon sinyalleri önemli ölçüde artar. Öte yandan O2, solunum kontrolü açısından beyindeki solunum merkezleri üzerine önemli bir doğrudan etkiye sahip değildir. Bunun yerine hemen tamamıyla aort yayı ve karotid arterin çatallanma noktasında bulunan periferik kemoreseptörler üzerine etki eder ve bunlar solunumun kontrolü için uygun sinirsel sinyalleri merkeze iletirler. Solunum düzenlenmesinde önemli rolü olan periferik kemoreseptörler, glomus karotikum ve glomus aortikumda bulunur.

H A Y V A N F İ Z Y O L O J İ S İ
407
ŞEKİL 7.26 Solunum kontrolü salanayan reseptörlerin beyindeki yerleri
Solunum sinirsel kontrolü Solunum merkezi, beyin sapındaki medulla oblangata’dadır. Kan kimyasındaki değişiklikler, karotis ve aort cisimciklerindeki değişikliğe duyarlı algılayıcılar (reseptörler; glomus aortikum ile glomus karotikum) tarafından algılanarak solunum merkezi uyarılmaktadır. Kimyasal denge solunumu öyle düzenler ki, normal koşullarda PCO2 değişmez (değeri sabit tutulur); PO2 tehlike yaratabilecek seviyelere düşmüşse, yükseltilir. Bir dakikadaki solunum hacmi metabolizma faaliyetleri ile orantılıdır. Ancak, solunumla metabolizma arasındaki ilişki CO2 ile sağlanmaktadır. Medullada bulunan solunum merkezi, kanın pH’sı, PCO2 ve PO2 olmak üzere 3 etken tarafından yönlendirilmektedir. Bunlardaki azalma veya artmalar, solunumda önemli değişikliklere neden olmaktadır. Örnek: Fazla egzersiz yapıldığında vücutta CO2 miktarı artar vekan pH’sı 7.4 ün altına düşer, normalde 1/20 olan H2CO3 / NaHCO3 oranı asit olarak artar. Bu durumdaki birey normalden daha fazla soluyarak (hiperventilasyon), fazla CO2 miktarını atmaya çalışır. Solunumla dışarı atılan hava nemli olduğundan CO2, gerçekte H2CO3 (karbonik asit) olarak atılır; bu sayede vücut sıvılarındaki H+ iyonunun yoğunluğu düşmüş olur. H2CO3 / NaHCO3 oranında sodyumbikarbonat (NaHCO3, baz) lehine artma (veya asit kısmında azalma) olursa, PCO2 düşer. Bu durumda birey, normalden az solur (hipoventilasyon), böylece karbondiosidin az miktarda atılmasını sağlar, tutulan karbondioksit su ile birleşerek bikarbonat oluşturur ve H+ yoğunluğu yükselir. H+ dengesinin düzenlenmesinde akciğerler çok önemlidir; çünkü H+ iyonunun yoğunluğunda hızlı değişim gerektiğinde CO2 in diffüze olabileceği geniş bir alana sahiptir. Hipoksi, solunum merkezini doğrudan uyarmaz. Kandaki PO2 düşmesiyle, aorta kavisinde yer alan glomus aortikum ile karotis arterin ikiye ayrıldığı bölgede yer alan glomus karotikumdaki kemoreseptörler (kimyasal algılayıcılar) uyarılır, refleks olarak uyarılan sinirler vasıtasıyla medulladaki solunum merkezi uyarılır ve solunum hızlanır. Solunum hızı kandaki CO2 miktarına göre düzenlenir. CO2 artışı soluk alıp vermeyi hızlandırır. Çünkü CO2 kanın pH sını düşürür ve ortam asit hale gelir Bu da beyni uyarır (Pons ve medula). Soluk alış verişinin hızı ve şiddeti omurilik soğanındaki sinirler tarafından denetlenir. Akciğer Hacim ve kapasiteleri
Ventilasyon (havalandırma): Akciğerlere havanın girip çıkmasıdır. İnspiryum: nefes alma, Normal inspiryum sırasında ekstra torasik hava yollarında transmural pozitif basınç hissedilir ve daralma, içe çökme eğilimi olur. İntra torasik hava yollarında hissedilen transmural basınç negatif olup, genişleme olur. Hava yollarındaki çap değişikliği basınca ve hava yolu kompliansina bağlıdır.

H A Y V A N F İ Z Y O L O J İ S İ
408
Ekspiryum: nefes verme, Normal inspiryum sona erdiğinde elastik geri çekilmeye bağlı olarak plevra ve alveol üzerinde pozitif basınç hissedilir. İntratorasik hava yollarında çap azalırken, ekstratorasik hava yollarında genişleme olur. Solunum hızının ve deriğinin artmasına hiperventilasyon denir. Hiperventilasyon vücudun fazla oksijen almak ve artan karbondioksitten kurtulmasını sağlar. İstemli durumda yapılması halinde ise kan pH değeri yükselir(alkolozis). Bu durum solunum merkezleri etkiler. Solunum normal göre daha az sıklıkla ve yüzeysel olmasına hipoventilasyon denir. Geri solunum ise bir torba ile ekspirasyon havasının tekrar alınması durumda ortaya çıkar.
ŞEKİL 7.27 Farklı solunum tipleri hiperventilasyon
Solunum alınan ve verilen hava miktarı spriometer ile ölçülür. Bu ölçümle Solunum foksiyon Testleri (SFT) yapılmış olur. Solunum fonksiyon testleri (SFT), solunum sistemini etkileyen hastalıkların tanı, izlem ve tedaviye yanıtının değerlendirilmesinde yaygın olarak kullanılmaktadır. Akçiğerde ölçülen yada hesaplanan hacimler şunlardır: �
1. İnspirasyon kapasitesi: Soluk hacmi ile inspirasyon rezervinin toplamıdır. 3500 ml 2. Fonksiyonel rezidüel kapasite: Ekspirasyon rezervi ile rezidüel hacmin toplamıdır. 2300 ml. 3. Vital Kapasite: İnspirasyon rezervi, soluk hacmi ve ekspirasyon rezervlerinin toplamıdır. 4500 ml. 4. Total Akciğer Kapasitesi: Vital kapasite ile rezidüel hacmin toplamıdır. 5800 ml

H A Y V A N F İ Z Y O L O J İ S İ
409
Solunum temel fonkisyonu dokuların oksijenlenmesi ve metabolizma sonunda ortaya çıkan karbokdioksit dokulardan uzaklaştırılmasıdır. Dokulardaki oksijen seviyesinin azalmasına hipoksiya denir. Bazı durumda ise dokularda oksijen seviyesi sıfıra kadar düşer. Bu duruma ise anoksiya denir
ŞEKİL 7.28 Spriometre
Akciğer kapasiteleri ise: Maksimum ekspirasyon ; yavaş ve zorlanmadan yapılırsa statik volüm olarak yavaş vital kapasite (SVC), zorlu yapılırsa dinamik volüm olarak zorlu vital kapasite (FVC) adını alır. Sağlıklı kişilerde; SVC = FVC SVC - FVC = hava hapsi
ŞEKİL 7.29 Fev1 ve FVC
1. Soluk Hacmi (Tidal Volum): Normal solunum hareketi ile akciğerlere alınan veya akciğerlerden çıkarılan hava hacmidir. 500 ml 2. İnspirasyon Rezervi: Normal soluk hacminin üzerine alınabilen fazladan soluk hacmidir. 3000 ml. 3. Ekspirasyon Rezervi: Normal bir soluk vermeden sonra zorlu bir ekspirasyonla fazladan çıkarılabilen hava hacmidir. 1100 ml. 4. Rezidüel (Tortu) Hacim: Zorlu bir ekspirasyondan sonra akciğerlerde kalan hava hacmidir 1200 ml .

H A Y V A N F İ Z Y O L O J İ S İ
410
FVC manevrasının başlangıcından itibaren zorlu ekspirasyonun birinci saniyesinde atılan hava volümüdür. FEV1: Zorlu ekspirasyonun 1. saniyesinde atılan volümdür. Normalde volümlerin % 80’i ilk saniyede atılır. Büyük havayollarını yansıtır. Havayolları obstrüksiyonunda belirgin azalır. Fonksiyonel rezidüel kapasite (FRC) Normal bir ekspirasyonun sonunda akciğerlerde bulunan hava volümüdür (RV + ERV) FRC= ekspiryum sonu akciğer volümüne (EELV)
ŞEKİL 7.30 Spriometre sonuçları
Solunum sırasında alınan havanın tamanı kulanılamaz. Havanın bir kısmı bronşlar ve bronşiyol gibi hava yollarında kalır. Alveol gaz alış-verişinde kullanılmayan bu havaya “ölü aralık” denir 500 ml hava için ölü aralık 150 ml kadardır. Ölü aralık hacmi her bir vücut ağırlığı için( 1 kg için) 2ml olarak hesaplanabilir. Anatomik ölü aralık Vd ile vital volum Vt ile gösterilirse
Vd=0.33 x Vt
Formülü ölü aralık hesaplamak için kullanabilir.
Metabolizma
Metabolizma kavramı genel olarak şu üç farklı olayı kapsar; Bu durumda organizmadaki madde ve enerji dönüşümlerinin tümünün metabolizma kapsamına girdiğini söyleyebiliriz. Metabolizma iki alt birimden meydana gelir; anabolizma ve katabolizma.
• Anabolizma: Küçük moleküllerden büyük moleküllerin sentezi ve enerji depolanmasıdır (protein, yağ, karbonhidrat şeklinde).
• Katabolizma: Büyük moleküllerin (karbonhidrat, yağ, protein) küçük moleküllere dönüşmesi ve bu esnada enerji üretimidir.
• Vücut içi ve vücut dışı kaynaklardan enerji üretimi, • Fonksiyonel ve yapısal doku bileşenlerinin sentezi, • Oluşan metabolik atık maddenin uzaklaştırılması.

H A Y V A N F İ Z Y O L O J İ S İ
411
ŞEKİL 7.31 Enerj eldesi kullanımı
Organizmanın en önemli enerji kaynağı ATP şeklinde depolanan yüksek enerjili fosfat (PO4) bağlarından oluşan kimyasal bileşiktir. Adenozin – PO3 ~ PO3 ~ PO3 “ ~ “ yüksek enerjili fosfat bağlarının sembolüdür. Bu iki yüksek enerjili bağlarını her birinin ayrılmasıyla fizyolojik koşullarda 12.000 kal/mol (= 12 k.kal) Depo Oksijen: Vücutta depo oksijen; 0.5 litre akciğerlerdeki havada, 0.25 litre vücut sıvılarında erimiş olarak, 1 litre hemoglobinde ile birleşmiş olarak, 0.3 litre miyoglobinde bulunur (toplam; 2 litre).
Su Altı (Dalma) Fizyolojisi
Suya dalan bir kimse hem suyun hem de su üzerinde kalan atmosfer basıncının baskısı altında kalmaktadır. Su yüzeyinde basınç 1 atmosferdir. Derinlere inildikçe her 10 metrede 1 atmosfer basınç artışı olur. Diğer bir değişle yaklaşık 30 metre derinlikte 4 atmosferlik bir basınç vardır. Boyle yasasına göre Basınçla hacim arasında ters bir orantı mevcuttur, basınç iki kat arttığında hacim yarıya düşmektedir. Örneğin yüzeyde 6 litrelik akciğer total kapasitesine sahip bir şahsın akciğer kapasitesi 10 metrede (2 atmosfer basınçta) 3 litreye, 20m ise (3 atmosfer basınçta) 2 litreye düşmektedir. 30 m den daha derinlere inmek tehlikelidir. Çünkü kemik yapılar özellikle göğüs kafesi dış su basıncına direnç gösterirken içteki hava basıncı aynı kalacak , kan basıncının artması nedeniyle kan, damar dışına sızacak , akciğerde ödem ve kanamaya yol açabilecektir. Derinlere inildikçe solunum kaslarının gücü su basıncını aşmaya yetmediğinden solunan havanın basınçlı olması gerekir. Bu nedenle yeteri kadar uzun bir boru yada snorkel aracılığıyla suyun altında kalıp nefes alıp verebilmek olası değildir. Bu amaçla SCUBA (self- ontained underwater breathing apparatus) adı verilen tüp sistemleri geliştirilmiştir. SCUBA sisteminde dipteki su basıncını yenebilecek güçte basınçlı hava gerektirmektedir. Örneğin 20m derinlikte 3 atmosferlik bir basınçlı (3 x 760 = 2280 mmHg) hava gerekir. Derinliklere inerken veya çıkarken belli prensiplere uyulması gerekmektedir, aksi takdirde hava embolisi , akciğerleri kollobs olması , dekompresyon (vurgun) vb.. patolojiler oluşabilmektedir. Dekompresyon derinliklerde kanda erimiş olarak bulunan nitrojenin kurallara uyulmaksızın ani yüzeye çıkılması durumlarında hacmin genişlemesi nedeniyle venöz ve arteryel kan damarlarının tıkanması nedeniyle oluşur. Belirtileri baş ağrısı sersemlik bilinç kaybı, kaslarda uyuşma, felç ve ölümdür. Tedavi için kişi yeniden aynı derinlik seviyelerine indirilerek uygun basınç koşullarında bekletilerek yavaş yavaş yüzeye çıkartılır. Yada vurgun yiyen şahıs rekomprasyon (basınç) odasında tutularak hacmi genişleyen nitrojen kabarcıklarının yeniden erimiş nitrojen duruma geçmesi sağlanır.

H A Y V A N F İ Z Y O L O J İ S İ
412
Şnorkel ile Solunum
Snorkel denilen bir boru yardımıyla yüzeydeki havanın solunması yöntemi soluk borusunun uzamasını, dolayısıyla ‘ölü boşluk’ hacmini arttırmaktadır. Buda CO2 Birikimine neden olmaktadır. Bu nedenle ara sıra kuvvetli nefes verme (inspirasyonla) ile ölü boşluktaki hava dışarı atılıp oksijenle zengin atmosfer havası solunmaya çalışılmalıdır. Su soğukluğu derinliklere inildikçe artar, bu nedenle vücut ısısı düşer, kalbin atım sayısı düşer, bradikardi oluşur. Ayrıca derinlerde orta kulak basıncı dengelemesi kulak zarında çökme ve yırtılmalar görüle bilir. Hiperventilasyonla suda kalış süresini arttırmak olasıdır. Hızlı nefes alıp vermekle CO2 atılımı artar. Solunumsal alkoloz oluşur. Su altında CO2 yavaş yükselmesi ve O2 azalması nedeniyle solunum merkezinin uyarılması gecikir. Buda bilinç kaybı nedeniyle boğulmalara yol açabileceğinden denenmemelidir.

H A Y V A N F İ Z Y O L O J İ S İ
413
Hücre haberleşme
mekanizmaları ve Endokrin
Sistem Fizyolojisi
Hücreler arası haberleşme mekanizmaları ile endokrin sistem yapısı organlar ve hormonlar hakkında bilgiler.
Hücre haberleşme yolları Hücre haberleşme molekülleri belirli dokulardaki hücrelerde sentezlenir. Bu hücrelerin sentezledikleri maddelerin etkileri özel hedef organlara özeldir. Genelde haberleşme amacıyla sentezlendikleri doku yada hücrelerden dolaşımla hedef hücrelere taşırlar. Ancak salgılandığı yerde yanındaki dokularda da etkili olabilir(parakrin haberleşme gastrin, sekretin). Hücreler arası haberleşmede farklı yollar vardır. Sinyal molekülleri: proteinler, küçük peptid, amino asid, nukleotid, steroid, Vitamin D3, retinoid, yağ asidi türevleri, çözünmüş gazlar (Nitrik oksid, CO ) olabilir.
Hücreler arası iletişimde 4 metod vardır:
� Gap junctions/Plazmodezma: Bitişik-komşu- hücreler arası direkt iletişim
� Temasa bağımlı iletişim: Komşu membran molekülleri arasındaki etkileşim
� Lokal iletişim: Otokrin ve Parakrin sinyal iletimi
� Uzak mesafeli iletişim:
-Kimyasallar (hormonlar) ile yavaş.
-Elektriksel (nörotransmitter) ileti hızlı
Bölüm
8 Bu bölüm için Power
point sunumu adresi:

414
Genel olarak olarak bir hücre diğer bir hücre ile 5 farklı yolla haberleşebilir.
ŞEKİL 8.1 Haberleşme yolları
İnsan genom projesi’nin verilerine göre, insan genomundaki yaklaşık 32000 genin %20’si hücre iletişiminde görev alan proteinleri kodlar. Sentezlenen sinyal molekülü ya sentezlendiği hücrenin yüzeyinde bağlı kalır veya bu hücreden salınır. Sinyal moleküllerinin birçoğu sinyal hücresi tarafından ekzositozla hücre dışına(polipeptid ve amin yapısındakiler) salgılanır. Diğerleri hücre zarından difüzyonla salınır. Hedef hücre sinyal molekülünü tanıyıp tutan (bağlayan) reseptör taşır. Sinyal molekülü reseptöre bağlanınca hedef hücre, metabolizmasında veya gen ekspresyonunda (anlatımında) değişiklikler yapar (hücresel yanıt). Görevini yapmış olan molekül (örneğin hormon) ortamdan hızla uzaklaştırılır yada yıkılır. İstenen yanıt sağlandığında sinyal sonlandırılır.
Etki Etki Yeri Örnek Endokrin Uzak hedef dokuya Ön hipofiz hormonları
Parakrin Komşu hedef dokuya Somatostatinin pankreas hücrelerine etkisi
Otokrin Salgılandığı hücreye Somatostatinin kendi salgısına etkisi
Jukstakrin Doğrudan bitişik hücreye Epidermal büyüme faktörünün komşu hücreye etkisi
Ekzokrin GIS lümenine salınıp farklı uzaklıklardaki mukoza hücrelerine
Gastrinin mide asit salgısına etkisi
Nöroendokrin Nöronlardan salınıp uzaktaki dokuya
Norepinefrinin kalbe etkisi
1. Nöral: Sinaptik bağlantılarda kimyasallar (nörotransmitterler) salgılanırlar ve lokal olarak hücre fonksiyonlarını kontrol ederler(Asetil kolin, nörepinefrin). 2. Endokrin: Bezler veya özelleşmiş hücreler tarafından kana kimyasal maddeler (hormonlar) salgılanır ve bu maddeler vücudun başka bir bölgesindeki hücrelerin fonksiyonlarını etkilerler(İnsülin glokogon, TSH). 3. Nöroendokrin: Nöronlar dolaşıma çeşitli maddeler (nörohormonlar) salgılarlar ve bu yolla vücudun başka bir bölgesindeki hücrelerin fonksiyonlarını etkilerler(ADH ve Oksitoksin). 4. Parakrin: Hücreler tarafından ekstraselüler sıvıya difüze olan ve komşu hücreleri etkileyen maddeler salgılarlar(NO, Prostogladin). 5. Otokrin: Hücreler hücre yüzeyindeki reseptörlerine bağlanarak aynı hücrenin fonksiyonunu etkileyen maddeler salgılarla(Nöral büyüme faktörü NGF)

415
ŞEKİL 8.2 Nörokrin, parakrin, nedokrin haberleşme modelleri
Hücrelerarası iletim tipleri hücre dışı sıvıya bir kimyasal habercinin salgılanmasını gerektirir. Bununla birlikte böyle bir salgılamayı gerektirmeyen iki önemli hücrelerarası kimyasal iletişim tipi de vardır. Gedikli bağlantılar (gap junction neksuslar aracılığı ile) ile meydana gelen birinci tipte, kimyasallar bir hücreden komşu hücreye, hücre dışı sıvıya girmeden geçer. İkinci tipte, kimyasal haberci gerçekte kendisini yapan hücreden salıverilmez, ancak hücrenin hücre membranında yerleşmiştir. Hücre habere yanıt verebilecek başka bir hücre tipi ile karşılaştığında membrana bağlı haberciler aracılığıyla iki hücre bağlanır. Bu tip iletişim (=jukstakrin) özellikle dokuların büyüme ve farklılaşmasında, ayrıca vücudu mikroplara ve diğer ajanlara karşı savunan hücrelerin işlevlerinde de önemlidir.
ŞEKİL 8.3 Haberleşme yolları
Hücreler arası haberleşme moleküllerin yanıt veren reseptör üç yolla etkilerini oluşturur. Bunlar G-proteinine bağlı reseptörler, tirozin kinaz reseptörleri ve iyon kanalı reseptörleridir.

416
Hücre içi haberleşmenin büyük bir yüzdesi, fosforilasyon/defosforilasyon reaksiyon çiftiyle gerçekleştirilir. Fosfat grubunu proteinlere ekleyen enzimlere proteinkinazlar adı verilir. ATP’den bir fosfat grubunu taşımak yolu ile, diğer proteinleri fosforile eden herhangi bir enzime verilen isimdir. Proteinkinazların yaklaşık 1000 kadar çeşidi vardır ve kinaz ailesi olarak tanımlanırlar. Proteinlere eklenmiş fosfat gruplarını uzaklaştıran enzimlere de fosfatazlar ismi verilmektedir. Fosfat grubuyla karşılaşan ve çoğu kez kendisi de bir enzim olan proteinde konformasyon (üç boyutlu) farklılaşması ve/veya etkinlik değişikliği oluşur. Birçok farklı protein kinaz vardır ve her biri yalnızca belirli bir proteinleri fosforile ederler.
ŞEKİL 8.4 Farklı haberleşme yolları
Hücre haberleşme için kullanılan haberci molekül aynı zamanda Ligant denir. Ligant moleküleri etki alanlarına göre 4 başlıkta toplanır. Parakrin Hücrenin yakın çevresindeki hücreler etkilenir. Sinaptik haberleşmede vücudun birbirinden çok uzaktaki kısımları arasında sinyal iletimi yapılıt.(Sinir hücresi uzantısı ile hedef hücreye) Endokrin Sinyal molekülleri (hormon) kan dolaşımına salınır (tüm vücuttaki hedef hücreler ) hedef hücreye bağlanarak etkisi gösterir. Otokrin Bağışıklık sistemi hücrelerinde (bazı T lenfosit tipleri kendi çoğalmasını uyaran büyüme faktörü sentezler. Erken gelişim evresinde “komünite etkisi” ve Kanser hücrelerinde görülür.
Tüm sinyal yollarında ortak üç olay gerçekleşir:
1. Sinyal alma: Sinyal alma, hedef hücrenin hücre dışından gelen bir sinyali algılamasıdır. Kimyasal bir sinyal, sinyal molekülü (ligant) hücresel bir proteine (reseptör) bağlandığı zaman algılanır. Protein hormonlar gibi hidrofilik ve büyük moleküllü sinyal moleküllerinin reseptörü genellikle hücrenin yüzeyinde bulunur.
2. Sinyal aktarımı: Sinyal aktarımı aşaması sinyali, özgül hücresel yanıt ortaya çıkaracak bir forma dönüştürür. Örneğin bir karaciğer hücresinin hücre zarındaki reseptör proteinin dıştarafına epinefrinin bağlanması, çeşitli basamaklar aracılığı ile glikojen fosforilazın aktivasyonuna yol açar. Sinyal aktarımı bazen tek bir basamakta gerçekleşmekle birlikte, çoğu

417
zaman çok sayıda farklı molekülde bir dizi değişikliğin olmasını gerektirir. Bu basamaklar sinyal aktarım yolu olarak adlandırılır. Bu yolda yer alan moleküllere genellikle aktarım molekülleri adı verilir.
ŞEKİL 8.5 Sekonder haberleşme moleküllerin amplikasyonu
3. Cevap: Hücre haberleşmesinin üçüncü aşamasında, aktarılan sinyal özgül bir hücresel yanıtı tetikler. Özetle tim hücreler arası haberlşemede:
1. Sinyal üreten hücre tarafından sinyal molekülünün sentezlenmesi 2. Sinyal üreten hücre tarafından sinyal molekülünün salınması. 3. Sinyal molekülünün hedef hücreye taşınması 4. Sinyalin hedef hücrede özgül reseptör protein tarafından tutulması 5. Hücre içi sinyal transdüksiyon yolunu tetiklemesi 6. Hücre metabolizmasında veya gen ekspresyonunda değişiklikler (hücresel
yanıt). 7. Sinyalin sönümlenmesi, çoğunlukla hücresel yanıtın sonlandırılması ligatın
yıkımı. Peptit/protein yapılı ligatın büyük bölümü reseptör aracılı endositoz ile hücre içine alındıktan sonra lizozomlarda hidroliz edilmektedir. Oksitosin ve anjiotensin gibi küçük molekül ağırlıklı bazı peptitler proteolizi plazmada olur. Katekolaminler, steroidler ve tiroid hormonları, özgül enzimatik değişiklikler ile inaktive edilmektedirler(MAO, COMT).

418
ŞEKİL 8.6 Sinyal moleküllerine bağlı olarak hücrede meydana faklı metabolik olayların genel yapısı
Hücreye, hücre dışı sıvıdan ulaşan ve kendi özgül reseptörüne bağlanan hücreler arası kimyasal haberciye birinci haberci denilirken; birinci habercilerin reseptörleri etkinleştirmesi sonucunda sitoplazmaya giren veya burada oluşan maddelere de ikinci haberci denilir.
G-Proteinine Bağlı Reseptörleri
Hücre reseptörleri olarak görev yapan önemli bir hücre içi proteidir. Plazma zarı iç yüzeyinde (sitozolik) yer alan ve G protein adı verilen heterotrimetrik (3 farklı alt birim içeren) proteinler ailesinden bir protein bileşiği ile bağlı olarak bulunur. Birinci habercinin reseptöre bağlanmasıyla üç boyutlu değişim tetiklenir ve G proteininin α alt birimine GTP bağlanır ve α alt birimi, diğer iki birimden ayrılır. Bu ayrışma, etkinleştirilmiş α alt birimin, bir iyon kanalı veya enzim gibi başka bir plazma zar proteinine bağlanmasını sağlar. Bu iyon kanalları veya enzimlere, hücre yanıtının oluşması için gerekli olan sonraki adımlara arıcılık ettiklerinden ötürü, plazma zarı efektör proteinleri adı verilir. G-proteinine bağlı reseptör, G proteini adı verilen bir protein yardımıyla iş gören hücre zarı reseptörüdür. Tüm G-proteinine bağlı reseptör bazı ortak özelliğe sahiptir(1) Ekstrasellüler ligant bağlıyıcı bölge(2) yedi adet transmembran heliks içeren integrin protein (3) G proteini ile etkileşime giren intrasellüler bölge. Mayalardaki çiftleşme faktörleri, epinefrin ve birçok diğer hormon ve nörotransmitterler gibi çok farklı sinyal molekülleri G-proteinine bağlı reseptörleri kullanılır. Bu reseptörlerin sinyal moleküllerini tanıyan bağlanma bölgeleri farklı olup, hücre içindeki farklı G proteinlerini tanır. Buna rağmen, G-proteinine bağlı reseptör proteinleri yapısal olarak birbirlerine benzer. Bunların hepsi, zarı kat eden yedi tane α heliks içerir.

419
ŞEKİL 8.7 G proteini
Zarın sitoplazmik tarafına gevşekçe bağlanmış olan G proteini, üzerine bağlı guanin nükleotidin tipine (GDP ya da GTP) göre açılıp, kapanan bir elektrik düğmesi gibi iş görür. (GTP yani guanozin trifosfat ATP’ye benzer.) GDP bağlı olduğunda inaktif olan G protein, GTP bağlı iken aktiftir. Bir sinyal molekülü G-proteinine bağlı reseptörün hücre dışındaki yüzeyine bağlandığı zaman, aktive olan reseptörün konformasyonu, inaktif haldeki özgül G proteine bağlanacak şekilde değişikliğe uğrar (1) ve bir GTP’nin GDP ile yer değiştirmesine neden olur. Bu durum G proteinini aktive eder (2) ve G proteini daha sonra genellikle bir enzim olan başka bir proteine bağlanarak, onun aktivitesini değiştirir (3). Eğer aktive edilen protein bir enzim ise metabolik yoldaki bir sonraki basamağı tetikler (4). Enzim ve G proteinindeki değişiklikler geçicidir; çünkü G proteini aynı zamanda GTPaz enzimi gibi davranır ve kısa süre sonra kendisine bağlı GTP’yi GDP’ye hidroliz eder. Böylece tekrar inaktif hale gelen G proteini, enzimi serbest bırakır; enzim de başlangıçtaki durumuna geri döner. G proteininin GTPaz aktivitesi, hücre dışı sinyal molekülünün ortadan kalkması durumunda, sinyal aktarım yolunun hızla durdurulmasını sağlar. G proteinler, plazma zarının efektör proteinleri, ikinci haberciler ve protein kinazlarla ilgili bazı çok önemli temel öğelerde G proteinlerle düzenlenen iki çok önemli efektör protein olan adenilat siklaz ve fosfolipaz C enzimleri ile yapmaktadır.
ŞEKİL 8.8 G proteini etkisi
G-proteini-Reseptör sistemleri son derece yaygın olup, çok çeşitli işlevler görür. G proteinleri embriyonik gelişimde önemli oldukları genetik çalışmalarla gösterilmiştir. Örneğin belirli bir G proteininden yoksun olan fare embriyoları normal kan damarları geliştiremez ve uterus

420
içinde ölür. G proteinleri duyu algılanmasında da iş görür. Örneğin insanlardaki görme ve koklama bu tip proteinlere bağlı olaylardır. Modern organizmalar ile prokaryotlar ve tek hücreli ökaryotlardaki G proteinleri ve G-proteinine bağlı reseptörler arasındaki yapısal benzerlikler, G proteinlerinin ve G-proteinine bağlı reseptörlerin çok erken dönemde ve muhtemelen ilk mikroorganizmaların algılama reseptörleri olarak evrimleştikleri fikrini vermektedir. G-proteinlerinin bakteriyel enfeksiyonların da dahil olduğu birçok insan hastalığında iş gördüğü keşfedilmiştir. Kolera, boğmaca ve botulizm (Clostridium botulinum’un ürettiği “Botulin” adlı zehir bulunan besinleri yiyen insanlarda gelişen ve felçlere yol açan zehirlenme tablosu) etmeni olan çeşitli bakteriler, G-proteinlerinin işlevini aksatan toksinler üreterek, kişiyi hastalandırır. Bu gibi enfeksiyonların ve diğer hastalıkların tedavisinde kullanılan ilaçlar genellikle deneme-yanılma yoluyla bulunmuştur. Farmakologlar bugün kullanımda olan tüm ilaçların yaklaşık %60'ının G-proteini ile ilgili yolları etkileyerek, bu hastalıklarda tedavi edici olduklarını gözlemlemiştir.
ŞEKİL 8.9 G proteini bağlı iyon kanalı etkisi
Gs======�adenilat siklaz aktive eder. Sitozolde cAMP miktarını attır.
Gi======�adenilat siklaz inhibe eder. Sitozolde cAMP miktarını azaltırır.
Gq======�fosfolipaz C’yi aktive eder. DAG ile Protein Kinaz C aktivasyonu, IP3 ile sitozolde kalsiyum seviyesinin artışı.
Gs adenilat siklaz Aktivasyonu
Beta-adrenoreseptörleri ;Serotonin 5-HT 4 , 5-HT 6ve 5-HT 7 ; Dopamin D 1benzeri reseptörler, histamin Hı 2
Kalp hızı, Düz kas gevşeme, nöronal aktiviteyi stimüle
Gi adenilat siklaz inhibe
Asetilkolin Anjiotensibojen II somatostatin
nöronal aktiviteyi, vazokondriksiyon
Gq fosfolipaz C Aktivasyonu
α 1 adrenoreseptörleri , muskarinik M 1 , E 3arasındadır ve M, 5 ,
[ 7 ]Histamin, H 1 , serotonin 5-HT 2 reseptör
Düz kas kasılması, Ca +2 çıkışı

421
Tirozin Kinaz Reseptörleri
Hücre haberleşmesinde görev alan hücre membran reseptör proteinidir. Tirozin kinaz, protein fosforilasyonunu sağlayarak, bir ligandın reseptörle etkileşmesini, liganta bağlı hücre hücre içi biyolojik aktivitelerin devamlılığını ve hücrenin çoğalmasını sağlayan protein kinaz ailesine mensup bir enzimdir. Bu reseptör, hücre zarında bulunan ve enzim aktivitesi taşıyan temel reseptör sınıflarından birisidir. Bu reseptör proteininin sitoplazmaya dönük olan kısmı tirozin kinaz olarak adlandırılan bir enzim olarak iş görür. Tirozin kinaz, substrat protein üzerindeki tirozin amino asidine ATP’den fosfat grupları aktarımını katalizler. Dolayısıyla, tirozin kinaz reseptörleri proteinlerdeki tirozinlere fosfatlar bağlayan zar reseptörleridir.
ŞEKİL 8.10 Triozin kinazlar
Tirozin kinaz reseptörlerinin birçoğu sinyal molekülü bağlanmadan önce bağımsız polipeptitler halindedir. Bunların her biri hücre dışında bir bağlanma bölgesine, zarı kat eden bir α helikse ve birkaç tirozin içeren hücre içi bir kuyruğa sahiptir. Bu tip bir reseptöre bir sinyal molekülünün bağlanması, proteinin sitoplazmik tarafını doğrudan aktive etmeye yetecek ölçüde konformasyonal değişikliğe neden olmaz. Bunun yerine, reseptör aktivasyonu iki basamakta gerçekleşir: 1. Ligandın bağlanması iki reseptör polipeptidin dimer(iki polipeptitten oluşan proteine dimer denir) oluşturacak şekilde bir araya gelmesine neden olur. 2. Bu bir araya geliş her iki polipeptidin tirozin kinaz aktivitesi gösteren kısımlarını aktive eder ve bunların her biri diğer polipeptidin kuyruğundaki tirozinlere fosfat ekler. Kısaca, sinyal molekülünün bir tirozin kinaz ve reseptör üzerindeki etkisi, polipeptidlerin bir araya gelmesi (agregasyon) ve reseptörün fosforile edilmesidir. Reseptör protein tümüyle aktive olunca, hücre içindeki özgül aktarım proteinleri tarafından tanınır. Bu proteinlerin her biri fosforlanmış haldeki özgül bir tirozine bağlanır ve aktif forma dönüşecek şekilde yapısal değişikliğe uğrar (aktarım proteini tirozin kinaz tarafından fosforile edilmiş ya da edilmemiş olabilir). Bir adet tirozin kinaz reseptör dimeri aynı anda on ya da daha fazla sayıdaki farklı hücre içi proteini aktive edebilir. (Fosforilasyon şelalesi) 3. Tirozin kinaz çok sayıda farklı aktarım yolunu ve hücresel cevabı tetikler. Birden fazla biyolojik yanıt gereken durumlarda örneğin hücre çoğalması, büyüme gibi durumlarda kontrolü gereken farklı metabolik yolların kontrolu triozin kinaz reseptörleri tarafından gerçekleştirilebilir.

422
4.Tek bir ligant bağlanma olayının çok sayıda yolu tetikleyebilme yeteneği, bu tip reseptörlerle G-proteine bağlı reseptörler arasındaki en önemli farktır. Ligant olmaksızın bir araya gelen anormal tirozin-kinaz reseptörleri çeşitli kanserlere neden olur. Büyüme faktörü reseptörü genellikle bir tirozin kinaz reseptörüdür. Hayvan hücreleri arasındaki haberleşmede görev alan kimyasal sinyaller arasında, hücreleri büyüme ve bölünme yönünde uyaran yerel düzenleyiciler olan büyüme faktörleri de bulunur. Hücre bölünmesi protein sentezi, kromozom duplikasyonu (ikilenmesi) ve hücre iskeleti elemanlarının yeniden düzenlenişi gibi farklı hücre kısımlarındaki çeşitli etkinlikleri içerir. Bütün bu etkinliklerin düzenli ve eşgüdümlü bir halde gerçekleşmesi için hücreye yardımcı olmak üzere özelleşmiş olan reseptör, aynı anda birden fazla sinyal aktarım yolunu tetikleme yeteneğindedir. İyon Kanalı Reseptörleri
Hücre haberleşmesinde görev alan diğer önemli grup reseptörler iyon kanalı proteinleridir. Hücre membranlarında her durumda açık olan pasif kanallarla birlikte, elektriksel veya cevredeki kimyasal değisikliklerle açılıp kapanan aktif kanallar bulunur. Membran sukun potansiyeli ve pasif membran direnci (rm) pasif kanallar nedeniyle meydana gelir. Birim yuzeydeki pasif kanal sayısı membranın sızıntı iletkenliğini olusturur. Pasif kanallardan genellikle K+ ve Cl- iyonları geçebilmektedir. Aktif kanallara kapılı kanallar adı da verilir. Çünkü bu kanallar belirli uyaranlarla açılıp kapanan kapı ile kontrol edilirler. Sinyalleri alan bazı zar reseptörleri ligant varlığında açılıp kapanan iyon kanallarıdır (ligant kapılı iyon kanalları). Bu kanallar kimyasal bir sinyale karşı cevap olarak açılan ya da kapanan, hücre zarında yer alan ve proteinlerle çevrili olan porlardır. Bu porların açılıp kapanması, Na+ ya da Ca+2 gibi iyonların geçişine izin verir ya da bunların geçişini engeller.
ŞEKİL 8.11 Ligant bağlı kanal
İyon kanaları integral proteinlerden oluşur. İyon kanallarının çeperlerinde polar gruplar bulunur. Bu tür kanallar bir yerinde geçirgen olduğu iyonun boyutlarına kadar daralır ve proteinlerden meydana gelen bir kapı(voltaja bağlı, liganta bağlı, mekano kapılı) ile kontrol edilir. Kanallar yarı açık ya da kapalı halde bulunurlar. Kapının açılıp kapanması icin gerekli şekil değisiklikleri yüklü veya dipolar yapıdaki voltaj sensorunu etkileyen elektriksel kuvvetlerle veya nörötransmiter molekullerinin bağlanmasından doğan kimyasal kuvvetlerle yönetilir. Aksiyon potansiyelinin yayılması ile ilgili en önemli kanallar Na+ kanalları, K+ kanalları ve Ca2+ kanallarıdır. Bir hücrede aynı iyona geçirgen farklı özelliklerde kanallar da bulunabilmektedir. Bir nörönun farklı bölgelerinde kanalların dağılımı farklılıklar gösterebilir. Böylece nörönun farklı bölgeleri farklı görevler üstlenebilmektedir. Örneğin Ca2+ kanalları sinir sonlama yerlerinde çok yoğundur. Bu kanallardan Ca2+ girişi ile nörötransmiter salınımı gerceklesir.

423
ŞEKİL 8.12 Potasyum iyon kanalı
Kanal proteininde ortaya çıkan biçim değişikliği hemen belirli bir iyonun hücre içi derişiminin değişmesine yol açar. Bu değişiklik genellikle hücrenin işleyişini doğrudan doğruya etkiler. Örneğin, sinir hücreleri arasındaki sinapslarda ortaya çıkan bu tip bir değişiklik, elektriksel bir sinyali tetikler. Bu elektriksel sinyal, alıcı hücre boyunca ilerler. Ligant varlığında açılıp kapanan iyon kanalları sinir sisteminde çok önemlidir. İkincil haberciler
Hücre yüzeyindeki reseptörlerin birincil habercilerle (hormon, nörotransmitter gibi) uyarılması sonucu hücre içinde üretilen, miktarları artan ya da azalan ve hücrede belli bir cevabın verilmesini sağlayan siklik adenozin monofosfat (cAMP), siklik guanozin monofosfat (cGMP), diasilgliserol, inozitol trifosfat (IP3) ve kalsiyum iyonları (Ca2+) gibi bileşiklerdir.
Adenilat siklaz-cAMP yolunda reseptörün, birinci habercinin bağlanması sonucu etkinleşmesi, bu örnekte Gs olarak bilinen (s=uyarıcı) protein ile olmaktadır. Bu da Gs’nin α alt birimi adenilat siklaz enziminin etkinleşmesini sağlar. Böylece ATP cAMP’ye (3’-5’ adenozin monofosfat) dönüşür. cAMP burada ikinci haberci olarak işlev görür. Hücrenin birinci haberciye karşı asıl yanıtını oluşturacak olaylar dizisini tetiklemek üzere, hücre içinde yayılır. cAMP fosfodiesteraz enzimi ile AMP’ye dönüştürülerek etkisi sonlandırılır. Bazı hastalıklarda cAMP düzeyleri aşırı artmıştır. Örneğin kolera hastalığında Gs aktivasyonu çok fazla olduğundan su emilimi azalır ve aşırı sulu ishal görülür. cAMP’nin çok arttığı diğer patolojik durumda Mc Cune-Albright Sendromudur. Ayrıca çay, kahve gibi içeceklerdeki kafein ve teofilin fosfodiesterazı inhibe ederek cAMP artışına yol açarak diüretik etki yaparlar. cAMP’nin hücre içindeki asıl görevi: sitoplazmik yerleşimli olan cAMP-bağımlı protein kinaz (yani protein kinaz A) denen enzime bağlanmak ve onu etkinleştirmektir. cAMP-bağımlı protein kinazlar - tıpkı diğer protein kinazlar gibi - diğer proteinlere (sıklıkla enzimleri) bir fosfat grubu ekleyerek fosforile ederler ve hücrenin yanıtını (salgı, kasılma vs) ortaya çıkarırlar. Adenilat siklazın aktivasyonu ile çağlayan bir etki görülmektedir. Örneğin karaciğerde adrenalin hormonu 108 glikoz
Potasyum iyon kanalı en dar yeri 3 Å çapında olup potasyum iyonun geçmesine izin verirken diğer maddelerin geçmesine izin vermez(daha küçük çapa sahip Lityum hariç)

424
molekülü oluşmasını sağlayabilir. Ayrıca cAMP ile etkinleştirilmiş protein kinaz A, hücre çekirdeğine difüze olabilir, birinci proteini fosforile edip, onun belirli genlerin özgül düzenleyici bölgelerine bağlanmasına yol açabilir. Böylesi genlere cAMP’ye yanıt veren genler denilmektedir. cAMP bağımlı protein kinazlar birçok proteini fosforilleyerek bunların uyarılmasına veya baskılanmasına yol açar. Bu sistemi kullanan hormonlar ise şunlardır: Sekretin, Somatostatin, Vazopressin (V2 reseptörü, epitel hücreleri), glukagon, kalsitonin, LH, FSH, HCG, PTH, ACTH, MSH, ADH, CRH, TSH ve katekolaminler (β reseptörlerini)dir .
Fosfolipaz C, diaçilgliserol ve inozitol trifosfat sisteminde(IP3) ligandı tarafından uyarılmış reseptörün eşleştiği ilgili G proteininin (Gq da denilir) α alt birimi, fosfolipaz A2 adı verilen bir plazma zarı efektör enzimini etkinleştirir. Bu enzim plazma zarı fosfolipidi olan fosfotidilinozitol bifosfatın (PIP2)���� diaçilgliserol (DAG) ve inozitol trifosfata (IP3) yıkılımını katalizler. Sonra da IP3 ve DAG, ikisi de ikincil haberci olarak ancak birbirinden çok farklı biçimde işlev görürler. DAG hücre membranına bağlı kalırken, IP3 hücre sitoplazmasında görev alır. IP3 ikincil haberci yolu
Hücre membranında bulunan fosfolipidlerin yağ asitlerin Fosfolipaz A2 tarafından yıkılmasıyla ikincil haberci olarak görev yapan farklı kimyasal maddelerin sentezi yapılır. Fofolipaz A2 aktivasyonu ligantın bağlanmasıyla G-proetin ligant kompleksi tarafından yapılır. Fosfolipid yağ asidi(arişodomik asit) yıkım ürünü olan IP3 ikinci mesajcı olarak görev yapar. IP3 endoplazmik retikulum veya akrozom üzerindeki reseptörlere bağlanarak hücre içi depolardan kalsiyumun açığa çıkmasına neden olurlar. Çıkan kalsiyum kalmoduline bağlanarak taşınır. DAG ise yine membrana bağlı bulunan proteni kinaz C'yi (PKC) uyarır. Protein kinaz C'nin aktif olarak görev yapabilmesi için kalsiyuma ihtiyaç vardır. İşte IP3'ün açığa çıkardığı kalsiyum bu rolü yerine getirir. PKC de proteinlerin treonin-serin aminoasitlerini fosforiller. Fosforillene proteinler ise ilgili fonksiyonları yerine getirir. IP3 hücre içi Ca2+ depolarından kalsiyum salımını yapar. Hücre içi kalsiyum yoğunluğun artışı hücrede hedeflene etkiyi sağlar Hücre içi Ca2+ konsantrasyonu, aktif olarak sitosolik Ca2+ hücreden sürekli olarak atıldığı için yani aktif Ca2+ pompalanmasının bir sonucu olarak son derece düşük bir seviyede (yaklaşık 0.1 uM) muhafaza edilmektedir. Ca2+ plazma zarından aktif olarak atılması yanında, hücre içi Ca2+ deposu olarak görev yapan, endoplazmik retikulum, içine de pompalanır. IP3 ligand-kapılı Ca2+ kanallarını reseptörlerine bağlanarak endoplazmik retikulumdan sitozole Ca2+serbest şekilde hareket etmesine neden olur. Bunun bir sonucu olarak, sitosolik Ca2+ seviyesi (Ca2+ 1uM), protein kinazlar ve fosfatazlar hedef proteinlerin modifikasyonunda kullanılır. Bunun yanında protein kinase C ailesinin bazı üyeleri, aktivasyon için Ca2+yanında diasilgliserol gerektirir, bu nedenle, bu protein kinazlarda, PIP2 sinyal verme yolunun, her iki kol ile birlikte düzenlenir.

425
ŞEKİL 8.13 Membran lipidlerinin etkisi
DAG ikincil haberci yolu
PIP2 hidrolizi(fosfolipaz A2 ile) ile üretilen diasilgliserol, hücre büyümesi ve farklılaşmasının kontrolünde önemli bir rol oynar. Bir çok protein kinaz C, farklı protein ailesine ait olan serin / treonin kinazları aktive eder. Protein kinaz C'nin bu rolü iyi bir örneği hayvanlarda tümör büyümesini desteklemek için yoğun bir şekilde araştırılmıştır. Forbol esterleri tümör arttırıcı aktivitesi diaçilgliserol analogları olarak hareket eden protein kinaz C uyarabilme yeteneğine dayanır. Protein kinaz C, sonra da transkripsiyon faktörü fosforilasyon, gen ekspresyonunda değişiklikler, hücre çoğalması ve uyarılması gibi farklı protein kinazlar kaskalarını aktive ederek etksisini gösterir.

426
Protein kinaz C (PKC) aktive ek olarak, diasilgliserol hücredeki farklı işlevleri vardır: • prostaglandinler için bir kaynak olarak kullanılılar • endokanabinoid 2-araşidonoilgliserolün ön maddesi olarak • trpC (Geçici Reseptör Potansiyeli kanonik) katyon kanalları, TRPC3 / 6/7 bir
subfamilyası aktivatörü olarak. DAG, topluca protein kinaz C adı verilen belirli bir protein kinaz sınıfına etkilidir. Bu enzimlerin etkinleşmesiyle çok sayıda diğer protein de fosforillenir ve hücresel yanıt oluşur. IP3 DAG’ın aksine ikincil haberci rolünü bir protein kinazı doğrudan etkinleştirme yoluyla yapmaz. Tersine, sitozole girdikten sonra, IP3 mitokondri ve endoplazmik retikulumun (ER) dış zarındaki kalsiyum kanallarına bağlanır ve onları açar. Mitokondri ve ER içindeki kalsiyum yoğunluğu sitozole göre çok yüksek olduğundan, kalsiyum bu organellerden dışarıya yayılır ve sitozolik kalsiyum yoğunluğu artar. Bu yüksek kalsiyum düzeyi, hücrenin birincil habercisine vereceği asıl yanıt (kas kasılması, salgılama vs) oluşana dek ortaya çıkacak olaylar dizisini başlatır. Ayrıca kalsiyumun etkilerinden biri de bazı protein kinaz C (C kalsiyum anlamındadır) biçimlerini etkinleştirmektir.
ŞEKİL 8.14 Hücresel yanııt
Fosfolipaz C ikinci haberci sistemini kullanan bazı hormonlarda şunlardır: Anjiyotensin II (damar düz kası), Katekolaminler (α reseptörleri), GnRH, GHRH, Oksitosin, TRH, Vazopresin (V1 reseptör, damar düz kası) dir. Kalsiyum iyonu (Ca+2), hem kimyasal (birinci haberci) hem de elektriksel uyaranlara hücrenin oluşturduğu çok çeşitli yanıtlarda ikinci haberci olarak işlev görmektedir. Buradaki ilginç bir mekanizma: voltaj kapılı kanallar üzerinden etkili elektriksel uyaranlara karşı da ikinci haberci gibi etkileyebilmesidir. Bu kanallardan içeri giren kalsiyum, belirli hücrelerde ER üzerindeki kanallara bağlanır ve bunları açar. Böylelikle bol miktarda kalsiyum sitozole salınır. Buna “kalsiyumla oluşturulan kalsiyum salınımı” adı verilir. Böylece, hücre ve sinyale (birinci haberci veya elektriksel dürtü) bağımlı olarak ER’dan kalsiyum salınımına yol açan temel ikinci haberci, ya IP3 ya da kalsiyumun kendisi olacaktır.

427
Kalsiyum ve kalmodülin
Ca2+ 'nın etkilerinin birçoğu Ca2+ bağlamayan kalmodulin ile aktive edilir. Bağlayıcı protein kalmodulin, (0.5 uM sitosolik Ca2+ konsantrasyonu seviyesinde) Ca2+ bağlanır. Kalmodulin (CaM) (kalsiyum ile modüle edilmiş bir protein için kullanılan bir kısaltma) tüm ökaryotik hücrelerde eksprese edilen bir kalsiyum-bağlayıcı proteindir. CaM, enflamasyon, metabolizma, apoptoz, kas kasılması, hücre içi hareket, kısa vadeli ve uzun süreli bellek, sinir büyüme ve bağışıklık yanıtı olarak süreçleri aracılık eder. Bu sayede daha sonra, protein kinazlar gibi, hedef proteinlerin etkileyen farklı hücresel fonksiyonları yapabilir. Ca2+ / kalmodulin yapısı hücresel proteinlerin düzenlenmesinde görev alabilir. Böyle bir Ca2+/ kalmodulin bağımlı protein kinazın örneği, miyosin hafif zincirlerin birimini fosforilleyerek, aktin-miyosin kısalma yapan Hafif zincir kinazındır. Ca2+/ kalmodulin tarafından etkin hale gelen diğer protein kinazlar metabolik enzimler, iyon kanalları ve transkripsiyon faktörleridir Bu enzimler fosforile olmuş CaM kinaz ailesinin üyelerini içerir. CaM kinazın bir formu, sinir sisteminde nörotransmitter sentez ve salımını regülasyonunda görev alır. Buna ek olarak CaM, kinazlar, transkripsiyon faktörlerini fosforile ederek gen ekspresyonunu düzenler. Ca2+ /kalmodülin ile adenilat siklaz ve fosfodiesterazların düzenleme, cAMP ile Ca2+ kanallarının regülasyonu ve her iki protein kinaz tip olan A ve Ca2 +/ kalmodulin bağımlı protein kinazlar tarafından, hedef bir dizi protein fosforilasyonunu kapsar. cAMP ve Ca2+ sinyal yolları geniş etkisi nedeniyle birçok hücresel yanıtları düzenlemede koordineli çalışırlar. Artmış sitozolik kalsiyum yoğunluğunun etkilerinin ortak paydası, çeşitli sitozolik proteinlere bağlanma kapasitesidir. Böylece proteinlerin 3 boyutlu yapısını ve işlevlerini değiştirmiş olur. Bu proteinlerin en önemlilerinden birisi, hemen tüm hücrelerde bulunan kalmodulindir. Kalsiyum ile bağlanınca, kalmodulin biçim değiştirir ve kalsiyum-kalmodulinin, çok çeşitli enzimlerin ve çoğu proteinlerin uyarılmasını veya baskılanmasını sağlar. Kalmodüline bağımlı protein kinazların uyarılması veya kenetlenmesi, fosforilasyon üzerinden, hücrenin birinci haberciye karşı geliştirdiği asıl yanıtında rol alan proteinlerin etkinleştirilmesi veya baskılanmasına neden olur. Kalsiyuma bağlanmak yoluyla etkilenen tek hücre içi protein kalmodülin değildir. Örneğin, kalsiyum, troponin C denen ve çizgili kas tiplerinde bulunan proteine bağlanarak, kasılma sürecini başlatır.
ŞEKİL 8.15 Kalmodüline bağlı protein kinaz katif hale gelmesi.

428
Hücrede Ca2+ son derece yaygın bir ikinci haberci moleküldür. Bu nedenle endoplazmik retikulum içinde tutulan Ca2+ IP3'e aracılığıyla sitoplamaya serbest Ca2+ bırakılarak, hücre içi Ca2+ konsantrasyonu artabilir. Alternatif bir yolda, plazma membranında bulunan Ca2+ kanalları aracılığıyla hücre dışı Ca2+ girişini yapılmasıdır. Birçok hücrede, IP3'ün üretiminden kaynaklanan hücre içi Ca2+ 'nın geçici artış, hücre dışı Ca2+ girdisi kaynaklanan daha uzun süreli bir artış takip etmektedir. Hücre dışı Ca2+ 'nın girişi plazma membranında voltaj kontrollü Ca2+ kanalları) membran depolarizasyonuna ile sinir iletimi ile yapılabilir. Hücre içi Ca2+ 'in artışı riyanodin reseptörleri ile Ca2+ kanallarını aktive edilmesine neden olabilir Hücre içi Ca2+ miktarının nöronlarda artışı sinir sisteminde kimyasal sinyallerin elektriksel sinyallere dönüştürülmesinde önemli bir rol oynar, yani sinir ileticilerin(nörotransmiter maddelerin) salıverilmesini tetikler. Kas hücrelerinde, Ca2+ membran potansiyeli değişikliklere yanıt olarak riyanodin reseptörlerinin açılmasına ve bunun sonunda Sarkoplazmik retikulum, saklanana Ca2+ sitoplazamaya verilmesine neden olur. Depolanmış Ca2+ bu durumda, kas kasılmasını tetiklemek için sitosolik Ca2+ miktarında büyük artışa, yol açar. Hücreler farklı etki ve fonksiyonları olan kalsiyum hücre içi seviyeleri kontrol edilmesi için çeşitli mekanizmalar kullanır. Hücrede Kalsiyum Bağlayici Proteinler;
• Kalmodulin; 4 adet kalsiyum bağlar. Hücre içi protein kinaz aktivasyonunda rol alir.
• Kalsekestrin; 50'ye yakin kalsiyum bağlayici bölge içerir. Sarkoplazmik retikulum içerisinde en fazla bulunan CaBP'dir.
• Kalretusilin; Yine sarkoplazmik retikulum kalsiyum bağlayıcıdır. Ancak kalretusilinin T lenfosit baskilama, GER den salınanacak proteinlerin katlanmasi (şaperon), perforinlerin Por oluşturmasını inhibisyonu gibi etkileri vardir.
• V Kalbindin; Sindirim kanali ve böbreklerden kalsiyum emiliminde ayrica nöronal fonksiyonlarında görev alan bir proteindir.
• S-100 protein ailesi; Kalsiyumun hem depolanmasi hem de hücre içi sinyalizasyonunda rol alan 20’den fazla değişik proteinden oluşan bir ailedir.
• Kalmirin; 2 adet kalsiyum bağlar • Anneksin; Sadece ökaryotlarda bulunur(Anneksin V). • Aralar-1 ve Sitrin; mitokondiri içinde kalsiyum bağlayan asp/glu taşıcısıdır. • Kalneksin; kalmodulinin prototipi kabul edilir
Protein Kinaz - C: Protein kinaz C ('PKC' EC 2.7.11.13) yaklaşık 10 izozimlerinin oluşan protein kinazlar ailesi proteinidir. Tüm Protein Kinaz C'ler yapısı düzenleyici etki alanı ve bir menteşe bölgesi ile birlikte kırıklı bir katalitik etki bölgesinden oluşur. Protein kinaz C Fonksiyon: Protein Kinaz C aktivitesinin- düz kas kasılmasında, spermlerin boşaltılmasında, mide suyu salgısın üretilmeside, trombosit agregasyonunda, CSF salgılamasını H + salgılanmasında , Na emiliminde ve bronkokonstriksiyonda önemlidir. CaM kinazlar: Ca2+ / kalmodulin bağımlı protein kinazları ya da diğer adıyla CAM kinazlar (EC 2.7.11.17), spesifik serin / treonin amino asitlerine fosfor grubu ekleyen protein kinazın etkisi öncelikle Ca2+ / kalmodülin ile düzenlenir.

429
CaM kinaz iki tipi vardır:
• Özel CaM kinazlar. Örneğin kasların kasılmasına neden miyozini fosforile eden miyozin hafif zincir kinaz (MLCK) 'dir.
• Çok fonksiyonlu CaM kinazlar. Nörotransmitter salgılanmasının,
transkripsiyon faktörü düzenlenmesi ve glikojen metabolizma gibi bir çok işlemle, rol oynar. CaM kinaz II olarak adlandırılır. Beyindeki proteinlerin % 1- % 2 kısmı CaM kinaz II proteidir.
CaM kinaz bir N-terminal katalitik alanı, düzenleyici bir etki alanı ve diğer bir üniteden oluşur. Miyoazin kinaz, düz kas kasılma mekanizmasında önemlidir. Düz kas içine kalsiyum akını sarkoplazmik retikulumdan yapılamaz, hücre dışı sıvıdan, düz kas liflerine kalsiyum girişi olmak zorundadır. Bu durumda ilk olarak, kalsiyum kalmoduline bağlanır. Bu durum myozinin aktin filament bağlanrak ve kontraksiyon(kasılmasına) (CrossBridge döngüsü boyunca) neden olur. Bu amaçla miyosin hafif zincir fosforile edilir. Miyoazin fosforalması MLCK, ile yapılır. Düz kas, çizgili kasta olduğu gibi, troponin gibi kompleks olmadığı için Miyoazin kinaz mekanizması, düz kas kasılması düzenlenmesi için ana yoldur.
ŞEKİL 8.16 Triozin kinaz G proteinleri ve iyon kanalları ile hücre haberleşmesi.
Protein Fosforilasyonu
Hücre zarındaki bir reseptöre, ona özgü bir sinyal molekülün bağlanması, sinyal aktarım yolundaki ilk basamağı tetikler ve hücrenin belirli bir yanıt vermesine yol açar. Sinyal ile aktive olmuş reseptör bir başka molekülü aktive eder. Bu molekül de başka bir molekülü aktive eder; bu süreç, hücresel yanıtı oluşturacak molekül aktive oluncaya kadar sürer. Aktarım molekülleri denen bu moleküller çoğunlukla proteinlerdir. Hücre haberleşmesindeki temel işleyiş aslında proteinlerin etkileşmesidir. Protein etkileşmesi proteindeki konformasyonal değişikliktir.

430
Konformasyonal değişiklik ise çoğunlukla fosforilasyon aracılığı ile ortaya çıkarılır. Gerçekten de protein fosforilasyonu, protein aktivitesini düzenlemek için kullanılan çok yaygın bir hücresel mekanizmadır. ATP’den bir proteine fosfat grupları aktaran bir enzimin genel adı protein kinaz’dır. Reseptör olan tirozin kinazların aksine, sitoplazmik protein kinazların çoğu kendi üzerlerine etki etmeyip, diğer substrat proteinler üzerine etki eder. Bunların birçoğu kendi substratlarını serin ya da treonin amino asitlerinden birisinden fosforile eder. Bu gibi serin/treonin kinazlar hayvan, bitki ve funguslardaki sinyal yollarında çok yaygın olarak bulunur. Sinyal aktarımındaki protein etkileşimleri “fosforilasyon şelalesi (silsilesi-kaskadı)” olarak nitelendirilir. Fosforilasyon şelaleleri bir sinyalin (ve dolayısıyla yanıtın) çoğaltılmasını sağlar. Sinyalin çoğaltılması sonucunda, örneğin karaciğer ya da kas hücresinin yüzeyindeki reseptörlere bağlanmış olan az sayıdaki epinefrin molekülü, glikojenden yüz milyonlarca glukoz molekülü oluşturulmasına yol açar. Hücre İçi Reseptörler
Sinyal reseptörlerinin tümü zar proteini değildir. Bunların bir kısmı hedef hücrelerin sitozolü ya da çekirdeği içinde çözünmüş durumdadır. Bir sinyal molekülünün böyle bir reseptöre ulaşabilmesi için, hedef hücrenin hücre zarından geçebilmesi gerekir; bunun için de hidrofobik olmalıdır. Hücre içi reseptöre sahip tüm uyarıcılar hücre zarında kolaylık(basit difüzyonla) geçebilmelidir.
ŞEKİL 8.17 Hücre içi reseptörler
Hidrofobik sinyal molekülleri arasında steroit hormonlar ve tiroit hormonları bulunur. Hücre içi reseptöre sahip bir başka kimyasal haberci, bir gaz olan nitrik oksit (NO)’tir. Çok küçük olan bu molekül zar fosfolipitleri arasından kolayca geçebilir. Testosteron steroit hormonlara bir örnektir. Testis hücrelerinden salgılanan bu hormon kan yoluyla taşınır ve tüm vücuttaki hücrelerin içine girer. Bu hormon hedef hücrelerin sitozolündeki reseptör proteine (testosteron reseptörü) bağlanarak, onu aktive eder. Hormona bağlanarak aktif forma dönüşmüş olan reseptör bu haliyle çekirdeğe girer ve erkek cinsiyet özelliklerini kontrol eden özgül genleri etkin hale getirir: Etkin hale gelen gene ait elçi RNA (mRNA) çekirdekten çıkar ve sitoplazmadaki ribozomlar tarafından özgül proteine çevrilir. Aktive edilmiş testosteron reseptörü bazı özgül genleri kontrol eden bir transkripsiyon faktörüdür. (Transkripsiyon faktörleri adı verilen özgül proteinler belirli bir hücrede, belirli bir anda hangi genlerin etkin hale getirileceğini -yani hangi genlerin mRNA oluşturmak üzere transkribe olacağını- kontrol

431
eder.) Hücre içi reseptörlerin birçoğu sinyal molekülleri onlara ulaşmadan önce çekirdeğin içindedir (örneğin östrojen reseptörleri). Hücre içi reseptör proteinlerin çoğu yapısal olarak benzerdir. Bu benzerlik bu proteinler arasında evrimsel akrabalık olduğunu gösterir.
ŞEKİL 8.18 Steroin hormonun etki mekanizması
Hormon
Endokrin sistem sinir sistemi ile birlikte homeostasiyi korur. İç ortamın istikrarı korumak için uzakta hedef hücreler üzerinde etkili özel kimyasallar (hormonların) kan yolu ile taşınması gerekir. Hormon bezleri (=endokrin bezler) tarafından hücreler arası sıvıya salgılandıktan sonra kan ile uzak dokulara taşınıp, kendileri için özgün reseptörler üzerinden hedef doku (= target doku) veya organların çalışmasını düzenleyen kimyasal habercilerdir. Dolaşıma salgılana bir hormon vücudun bütün bölgelerine gidebilir. Ancak hormonlar etkilerini sadece belirli hücrelerde ve dokularda gösterir. Bu hücrelere ve organlara hedef hücre veya organ denir. Hormonların salgılanmasında temel olarak iki kontrol mekanızması vardır, 1) sinir sistemi ile 2) negatif ve pozitif feedback mekanizmalar ile kontrol. Hormonlar etkilerini hücre membranı yüzeyindeki, yada sitoplazmanın veya çekirdeğin içindeki reseptörleri aracılığıyla gösterir. Hormonal sekresyonlar genellikle negatif feedback sistemler ile normal seviyede tutulur.
Hormonlar, amin, peptid, steroid, lipid yapısında olabilir.
• Amin yapısındaki hormonlar: Ör: adrenalin, dopamin.
• Peptid yapısındaki hormonlar: Sinyal iletim molekülleridir. Ör: insulin, prolaktin, endorfin.
• steroid yapısındaki hormonlar: küçük molekül ağırlıklıdır. Ör: Testesteron, östrojen, progesteron.

432
• Lipid yapısındaki hormonlar: Kısa süreli lokal etkiye sahip lipid türevi biyomoleküllerdir. Ör:Prostoglandin.
ŞEKİL 8.19 Hormon kanla taşınması
Hormonlar kan dolaşımına verilen uzak hedef dokuda çok düşük konsantrasyonlarda (10-9-10-12
molar seviyesinde) etki gösteren kimyasal habercilerdir. Çok az miktarda sentezlenen hormonlar genel olarak üç ana sınıfa ayrılırlar: (1) Protein ve polipeptidler: Ön ve arka hipofiz bezi hormonları, pankreastan (insülin ve glukagon), paratiroid bezinden (paratiroid hormon) salgılanan hormonlar ve diğerleri. (2) Steroidler: Adrenal korteksten (kortizol ve aldosteron), överlerden (östrojen ve progesteron),testislerden (testosteron) ve plasentadan (östrojen ve progesteron) salgılanırlar. (3) Trozin amino asidi türevleri : Tiroid bezinden (tiroksin ve triiyodotironin) ve adrenal medulladan (epinefrin ve norepinefrin) salgılanırlar. Polisakkarid veya nukleik asit yapısında hormon bilinmemektedir.

433
ŞEKİL 8.20 Sterol yapısındaki hormonlar
Hormonlar kimyasal yapılarına göre 3 grupta incelenirler. Yağda eriyebilen steroid kaynaklı, steroid hormonlar, Amino asit kaynaklı hormonlar ve Suda eriyen protein yapılı hormonlar. Vücuttaki hormonların çoğu poli peptid ve protein yapısındadır. Bu hormonlar üç adet aminoasitten oluşabilecek kadar küçük (tirotropin-salgılatıcı hormon) veya 200 aminoasitten oluşacak kadar uzun (büyüme hormonu ve prolaktin) olabilirler. Genelde, 100 veya daha
Daha ayrıntılı bakıldığında hücre haberleşme moleküllerin yapılarına göre: � Peptitler ve proteinler: Hipotalamus, hipofiz, paratiroit, pankreas, mide-bağırsak sistemi
ve bazı plasenta hormonları � Steroidler: Adrenal korteks ve gonadlardan salgılanan hormonlar ile bazı plasenta
hormonları � Amino asit türevi hormonlar: Adrenal medülla hormonları: Katekolaminler Tiroit hormonları � Eikozanoidler: Çoklu doymamış yağ asitlerinden elde edilirler. Prostaglandinler
(PG), prostasiklinler ,Tromboksan A2, lökotrienler . � Retinoidler Retanol � NO Nitrik oksit

434
fazla sayıda aminoasitten oluşan polipeptidler protein olarak adlandırılır ve 100 ’den daha az sayıda aminoasitten oluşanlar oligopeptid olarak bilinirler. Protein ve peptid hormonlar, diğer proteinlerde olduğu gibi endokrin hücrelerin granüllü endoplazmik retikulum(GER) tarafından sentezlenirler. Peptid yada proetin yapısında olan hormonlar Önce belirli bir genin mRNA halinde ifadesi olarak preprohormone adı verilen bir protein öncüsünün olarak üretilir. Preprohormones sıklıkla, post-translasyonel karbonhidrat içerirler ve ER (glikosilasyonla) tarafında değiştirilirler. Preprohormones sinyal sekansı prohormon oluşturmak üzere kaldırılır Bu işlem golgi tarafından gerçekleşir. Prohormon aktif hormon olarak işlenir ve salgı vesiküleri halinde paketlenir.
• Polipeptid ve Protein Yapısındaki Hormonlar Gerektikleri Süreye Kadar Sekretuvar Veziküllerde Depolanırlar. Peptid / protein hormonları Salgı keseleri harekete geçiren bir sinyal ile plazma zarına gelir. Sonra ekzositozla ve kan içine salgılanır. Bazı durumda prohormon salgılanan ve aktif hormonuna dönüştürülmesi hücre dışı sıvıda olur. Örneğin, anjiyotensin I böbrek ve akciğer tarafından salgılanan enzimlerle tarafından Anjiotensin II dönüştürülmesi ve aktif hale geçmesi. Protein yapısında olan başlıca hormonlar hipotalamus, hipofiz, paratiroit, pankreas, mide-bağırsak sistemi ve bazı plasenta hormonlarıdır.
• Steroid Hormonlar Tüm steroid hormonları kolesterol türetilen ve siklopentil halka
yapısı ve yan zincir bakımından farklılık vardır. Steroid hormonları depolanmazlar. Tüm steroid hormonları lipidte çözünür. Steroid hormonların kimyasal yapısı kolesterole benzer ve pekçok durumda kolesterolden sentezlenirler. Lipidde çözünürler ve üç sikloheksil halkasından ve bir siklopentil halkasından oluşurlar(Strean yapısı). Endokrin hücrelerde çok az miktarda hormon depolanmakla birlikte, sitoplazma vakuollerinde depolanan büyük miktardaki kolesterol esterleri uyarı sonucu steroid sentezi için hızla mobilize olur.

435
ŞEKİL 8.21 Steroid hormone sentezi ve steroid hormonlar
Steroid sentezleyen hücrelerdeki kolesterolün büyük kısmı plazmadan gelir ancak kolesterol sentezi hücrelerde de gerçekleşir. Steroidleıin lipidde çözünürlükleri oldukça fazladır. Bu nedenle, salgılanır salgılanmaz hücre membranından kolayca difüze olurlar, interstisyel sıvıya oradan da kana geçerler. Steroid hormonları suda çözülmez olmaları nedeniyle steroid hormonları kendilerine has bağlanma bölgeleri taşıyan kan globülinleri ile kompleks yaparak taşınır. Steroidler hedef hücrede aktif steroid dönüştürülebilir.
ŞEKİL 8.22 Farklı hedef hücrelerde steroidlerin farklı etkileri .
• Amin Yapısındaki Hormonlar Tirozinden amino asidinden sentezlenir. Tirozinden kaynaklanan iki grup hormon, tiroid ve adrenal medulla hormonlarının her ikisi de salgı hücrelerinin sitoplazmasmdaki enzimlerin etkisiyle oluşurlar. Amin yapıdaki hormonlar ya da amin türevi hormonlar katekolaminler, triptofan türevleri ve tirozin türevi hormonlardır. Amin yapısındaki proteinler suda çözünürler. Bu hormonlar salgı ve veziküllerde depolanırlar. Suda eriyen hormonlar (amino asit ve protein yapılı hormonlar) hücresel olayları membrana yerleşmiş reseptörleri aracılığıyla düzenlerler. Bu mekanizmaya göre hormon (1. haberci) hücre yüzeyindeki reseptörüne bağlanınca siklik AMP (cAMP)(ikinci haberci) oluşur. İkinci haberci olan cAMP hücre içine diffüze olarak hücrenin farklı fonksiyonlarının gerçekleştirilmesini sağlar.
• Gaz fazında bir parakrin haberci: NO Nitrik Oksit : periferal vasküler düz kas
üretilen güçlü bir gevşetici olarak 1979 tarif edilmiştir. Bir sinyal molekülü olarak tüm vücut tarafından kullanılır. Vücut sistemine bağlı olarak farklı işlevler hizmet verir: yani nörotransmitter, damar genişletici, bakterisit olarak görev yapar. Ayrıca ekosistemde çevre Kirleticisi olarak bilinir. Bilinen ilk gaz biyolojik haberci molekülüdür. Nitrik oksit, bir azot atomu ve bir oksijen atomundan oluşan bir iki atomlu serbest radikaldir . Lipitlerde Çözülebilir ve hücre zarlarının kolayca geçebilir. Biyolojik membranları hızla geçer (26 cm/sn hızıyla). Düz kaslarda ve diğer hücrelerde bulunan

436
guanilil siklazı, Fe2+ grubuna bağlanarak aktive eder. Bunun sonucu olarak sGMP düzeyini artırır. sGMP miyozin hafif zincirinin defosforilasyonuna neden olarak düz kas gevşemesi yapar. NO’in antitrombositik, antimitojenik, sitotoksik vd etkilerine sGMP’nin aracılık ettiği anlaşılmıştır. Diğer taraftan NO, ETZ’nin son kompleksi olan sitokrom oksidazı (kompleks IV), fizyolojik konsantrasyonlarda, inhibe eder. Kısa sürede bozulur (genelikle birkaç saniye içinde). Nitrik Oksit Sentezi hücre içinde yapılır. Nitrik oksit L-argininden sentezlenir. Bu tepkime, nitrik oksit sentaz(NOS ile 1294 AA) enzimi tarafından katalize edilir. NOS bir flavoproteindir. NADPH ve O2 ye bağımlı oksijenasyonu katalize eder. Bu enzimin kofaktörler(NADPH; FAD; FMN; tetrahidrobiopterin, hem, kalmodün gereklidir). NO çabuk yıkılır. Eliminasyon yarıömrü 6-50 sn dir. NO super oksit anyonu (serbest oksijen radikalleri) etkileşip peroksinitrit oluşur. Bu olay; Makrofajlar, polimorfonükleer lökositler, mezanjial hücreler ve endotel hücrelerinde gerçekleşir. Peroksinitrit; direkt veya indirekt olarak lipit peroksidasyonunu aktive eder. Dolayısıyla hem infeksiyon etkeni hücrelerde hemde vücut hücrelerinde sitotoksik etki oluşturur. Bu etkiler ortama superoksid dismutaz katılması NO nun inaktivasyonunu yavaşlatır (serbest oksijen radikallerini nötralize ederek). Hücre içindeki NOS Glutamat NMDA nörotransmitter reseptörleri bağlanır ve Ca2+ kanalları açarak hücre içine kalsiyum girişine neden olur. Ayrıca NO, K+ kanallarını da açarak hiperpolarizasyona neden olabilir. NO, sonuç olarak intrasellüler Ca2+ düzeyini düşürür. NO’ nun metabolizması genellikle ortamda bulunan süperoksid (O2
-) anyonları tarafından düzenlenir. NO, tiyollü (-SH) bileşiklere bağlanabilir. Nitekim S-nitrotoziyollerin NO’nun hücresel depoları olduğu ileri sürülmektedir. Ayrıca kanda NO, albumin proteinindeki tiyollü aminoasitlere bağlanarak bir yerden başka bir yere taşınabilir.
NOS aktivasyonuna bağlı olarak kalmodülin aktivasyon ve buna bağlı etki gözlenir. NO sentezi, endotel hücreleri, akciğer hücreleri ve nöronal hücrelerce yapılır. İnsan vücudunda Sinir sisteminde, dolaşım sisteminde, kas sistemi, bağışıklık sisteminde, sindirim sisteminde, nitrik oksit görev alır. Farklı dokularda tanımlanmış NOS enziminin de 3 farklı izoformu vardır. 1. Sinir ve bazı dokularda (akciğer, pankreas, mide ve uterus) bulunan nöronal NOS (nNOS) 2. İmmünolojik uyaranlarla indüklenen ve bütün çekirdekli hücrelerde bulunan indüklenebilir NOS (iNOS) 3. Endotel hücrelerde bulunan endotelyal NOS (eNOS) nNOS ve eNOS’a yapısal NOS (yNOS) adı verilmektedir. Bir nörotransmitter olarak nitrik oksit : NO bir nörotransmitter sinyal molekülü olarak işlev görür. Düz kas kasılması, adaptif gevşeme ve lokal vazodilasyonun NO sinyalleri yapılır. Ayrıca Nitrik oksit, uzun süreli hafızada bir rol oynadığına inanılmaktadır. Önerilen Hafıza mekanizma nöronların uzun süreli potensiyalizasyon (bellek) kolaylaştıran bir retrograd haberci Ca / kalmodulin ilgili sentez mekanizması NOS I aktive olan Guanil siklaz aktive olur. Bu NO tarafından uyarılan sinir aksiyon potansiyellerinin bir döngü başlatır. İnme sonrası en beyin hasarı nedeni olarak aşırı NO salınımı olur.

437
ŞEKİL 8.23 NO bağlı düz kas gevşemesi.
NO bir nörotransmitter olarak vücut hizmet verebilir, fakat, vücut içinde yaygın olarak kullanılan diğer nörotransmiterler arasında farklılıklar vardır. NO sabit miktarda sentezlenmez yanlız talep üzerine sentezlenir. NO veziküllerinde depolamaz buna karşın hücrelerin dışına yayılır ve Ekzositoz tarafından ortama verilir. Reseptörler yüzeye bağlanmaz bunun yerine, sitoplazma girer, hedef hücre sitoplazmasında etki gösterir ve hücre içi guanil siklazı etkiler. Nöro transmiterlere benzer bazı özelliklere sahiptir. Presinaptik terminal bulunur. Dolaşım Sistemi Nitrik Oksit damar genişletici olarak hizmet vermektedir . Yüksek kan akış hızına yanıt olarak salınan ve sinyal molekülleri (ACH ve bradikinin) son derece lokalize ve etkileri kısa olan NO sentezini, inhibe eder. NO aynı zamanda EDRF (endotel kaynaklı gevşeme faktörü) olarakta bilinir. Ca +2 NOS aktive damar lümeni salınır. NO vasküler endotel hücrelerinde NOS III sentezlenir . Bu durum, guanil siklazı aktive olarak cGMP'nin üretimine neden olur. cGMP bir artış Ca +2 pompalarının çalışarak klasiyumu hücre dışına atmasına , böylece hücreye Ca +2 konsantrasyonunun azaltılması, son olarak düz kas gevşemesi neden olur. Bağışıklık Sistemininde Nitrik Oksit görev alır NOS II konak savunma reaksiyonlarında kullanılır. NO sentezini katalize NOS II aktivasyonu hücreye Ca2+ bağımsızdır. NO sentezi, özellikle makrofajlar olur. NO Viral replikasyonun güçlü bir inhibitörüdür. NO, antibakteril ajandır. Diş eti yakın gıda ekstre nitrat oluşturulur. NO vücut için zararlı olabilir ağız içindeki bakterileri öldürür. Sindirim Sisteminde Nitrik Oksit daptif gevşemede kullanılır. NO midenin dolmasına yanıt olarak mide gerilmesi teşvik etmektedir. Mide dolu olunca, gerilim reseptörleri NO salan nöronlar aracılığıyla düz kas gevşemesi tetikleyebilir. Hormon reseptörleri
Hormon reseptörlerin hücre zarından geçebilmesine göre reseptörler ya hücre zarının dış kısmında yada hücre içinde bulunabilir. Hücre zarında reseptörü olan tüm hormonların etkileri ikincil haberciler ile gerçekleşirken hormona bağlı olarak metabolik bir faaliyet kontrol edilir. Buna karşın reseptörleri hücre sitoplazmasında ya çekirdek olan hormonlarıın temel işelvi DNA transkipsiyonun yada DNA mRNA sentezin düzenlemesidir.
1. Hormon reseptörleri hücre membranının içinde veya yüzeyinde bulunabilir. Membran reseptörleri daha çok protein, peptid ve katekolamin (epinefrin ve norepinefrin) hormonlarına özgüldür.

438
2. Hormon reseptörleri hücre sitoplazmasında bulabilir. Çeşitli steroid hormonların reseptörlerinin hemen hemen hepsi sitoplâzmada bulunur.
3. Hormon reseptörlerin hücre çekirdeğinde bulunabilir. Tiroid hormonlarının reseptörleri çekirdekte bulunur. Bu reseptörlerin bir veya birden fazla kromozom ile direkt ilişkide olacak şekilde yerleştiğine inanılmaktadır.
Hormon Etki Mekanizmaları
Suda eriyen hormonlar (amino asit ve protein yapılı hormonlar) hücresel olayları membrana yerleşmiş reseptörleri aracılığıyla düzenlerler. Bu mekanizmaya göre hormon (1. haberci) hücre yüzeyindeki reseptörüne bağlanınca siklik AMP (cAMP)(ikinci haberci) oluşur. İkinci haberci olan cAMP hücre içine diffüze olarak hücrenin farklı fonksiyonlarının gerçekleştirilmesini sağlar. Yağda eriyen hormonlar steroid yapılı hormonlardır ve kolesterolden sentezlenirler. Kortizol, progesteron, östrojen, testeron ve tiroksin hareketli reseptör mekanizması ile etki gösterirler, bu hormonların özel reseptörleri çekirdektedir. Bu mekanizma ile çalışan hormonlar hücre fonksiyonlarını etkileyen proteinleri sentezleyerek görevlerini yerine getirirler. Hücre yüzeyi reseptörlerine bağlanan hormonlar:
1. Adenilat siklaz aktivasyonu(Gs) veya inaktivasyonu(Gi) yapan hormonlar (cAMP) 2. Guanilat siklaz aktivasyonu yapan hormonlar(cGTP) 3. Fosfolipaz C aktivasyonu yapan(Gq) ve/veya sitozolik Ca2+ konsantrasyonunu
artıran hormonlar 4. Tirozinkinaz aktivasyonu yapan hormonlar
ŞEKİL 8.14 Hücre içi ikincil haberciler
Adenilat siklaz mekanizması kullanan hormonlar (cAMP) Glukagon, Kalsitonin, LH, HCG, FSH, PTH, ACTH, MSH, ADH, CRH, TSH, HCG, Katekolaminler, İnositol trifosfat (IP3) mekanizması kullanan hormonlar Gq proteini fosfolipaz C’yi aktive eder. Bu da membran fosfolipidlerini parçalayarak diaçil gliserol (DAG) ve IP3 olusturur. IP3 endoplazmik retikulumdan kalsiyum salınımına neden olur. DAG protein kinaz C’yi aktive eder. Protein kinaz C bazı proteinleri fosforilleyerek hormonun fizyolojik etkilerini ortaya çıkarır. IP3 sistemini kullanan hormonlar: Oksitosin, GnRH, TRH, ADH, GHRH, Anjiotensin 2, Katekolaminler (Alfa 1 Reseptörü), İyon kanalı, G proteinleridir.

439
ŞEKİL 8.24 IP3 ikinci haberci sistemin
Biyolojik sinyalizasyonda diasilgliserol ikincil mesajcı olarak çalışır. Fosfolipaz C (PLC) enzimi tarafından fosfatidil inositol-bisfosfat (PIP2) adlı fosfolipitin hidroliz ürünüdür. Hücre zarına bağlı olan bu enzim aynı reaksiyon ile inositol trifosfat (IP3) üretir. İnositol trifosfat sitozola dağılmasına karşın, diasilgliserol (DAG), hidrofobik olması nedeniyle plazma zarında kalır. IP3 düz endoplazmik retikulumdan kalsiyum iyonlarının salınmasına neden olur, DAG ise protein kinaz C (PKC)'nin fizyolojik aktivatörüdür. Membranda DAG oluşumu, PKC'nin hücre zarından sitozola yer değiştirmesini (translokasyonunu) kolaylaştırır. Tirozinkinazlar Hormon Reseptörleri: Hormonlar etkilerini hücre membranı yüzeyindeki, yada sitoplazmanın veya çekirdeğin içindeki reseptörleri aracılığıyla gösterir. Tek geçişli alfa heliks transmembran proteinlerdir. Sitozolik bölge protein tirozin kinaz etkinliği gösterir. Reseptör protein tirozin kinazlar substratlarını tirozin kalıntılarında fosforille ederek etkimlik gösterir. Çoğu tek bir polipeptid yapısındaki hormonlarla aktive olur. Triozinkinaz reseptörü kullanan başlıca hormonlar İnsülin, İGF-1 ve büyüme Faktörleridir. Steroyit ve Tiroyit Hormonların Etki Mekanizması: Steroyit hormon hücre sitoplazmasına diffüze olur ve buradaki spesifik reseptörüne bağlanır. Bunun sonucunda reseptörde DNA’ya bağlanan domain ortaya çıkar. (reseptör steroid hormon ortamda yok iken HSP-90 (Isı soku proteini) ile örtülüdür. Hormon reseptöre bağlandığında HSP aktif domain kısmından ayrılır) Hormon-reseptör kompleksi çekirdeğe geçer ve çekirdekte spesifik DNA bölgesinin regülatör bölgesi ile etkileşir. Transkripsiyon olur ve mRNA sentezlenir, bu mRNA sitoplazmada fizyolojik etkiyi yapacak proteine translasyone olur. Steroid hormonların ve progesteronun reseptörü sitoplazmada bulunur. Östrojen, androjen ve tiroid hormon reseptörleri ise çekirdekte DNA üzerinde bulunurlar.

440
Bir hücrenin hücre dışı bir sinyale normal bir yanıt vermesi için, başlangıçtaki sinyali ortadan kaldırıp sinyal aktarım yolunu kapatacak mekanizmalara sahip olması gerekir. Protein kinazların etkileri hücredeki protein fosfatazlar tarafından durdurulur. Protein fosfatazlar proteinlerdeki fosfat gruplarını uzaklaştırır. Herhangi bir anda fosforilasyonla düzenlenen bir proteinin aktivitesi, hücredeki aktif kinaz molekülleri ile aktif fosfataz molekülleri arasındaki dengeye bağlıdır. Hücre dışı sinyal molekülü olmadığı zaman, aktif fosfataz molekülleri çoğunluktadır. Dolayısıyla, sinyal yolu kapanır ve hücresel yanıt sona erer. Sinyal-reseptör sisteminin endositozla hücre içine alınması veya olduğu yerde inaktive edilmesiyle de sinyal sonlandırılır. Hormonların görevlerini yaptıktan sonra yok edilmeli gereklidir. Peptit/protein yapılı hormonların büyük bölümü reseptör aracılı endositoz ile hücre içine alındıktan sonra lizozomlarda hidroliz edilmektedir. Oksitosin ve anjiotensin gibi küçük molekül ağırlıklı bazı peptit hormonların proteolizi plazmada olur. Katekolaminler, steroidler ve tiroid hormonları, özgül enzimatik değişiklikler ile inaktive edilmektedirler.
ŞEKİL 8.25 Nöral ve hormonal kontrol mekanizmaları
Endokrin Bezler
Endokrin bezler vücud içinde farklı doku ve organlarda yer alırlar. Tüm endokrin bezler salgılarını kan bırakır. Kanda ya tek başlarına yada özel taşıma proetinlerine bağlı olarak hormonlar taşınır. Endokrin sistem ve sinir sistemi ile birlikte bir takım mesajlar iletir, Hedef organlar üzerinde kontrol sağlar ve homeostazda rol oynar.

441
ŞEKİL 8.26 Endokrin bezler
Hormonlar, kanalsız endokrin bezlerde üretilen ve kana salınarak hedef hücrelerde metabolik faaliyetleri düzenleyen steroid veya protein yapılı kimyasal mesajcı moleküllerdir. Metabolizmanın düzenlenmesi için düşük düzeyleri (pikomol - nmol) yeterlidir. Bir hücreden diğerine mesaj taşıyan sinyal molekülleridir. Hücre düzeyinde hormon etkisi hormonun reseptörü ile bağlanması ile başlar. Hormonlar sentezlendikleri yere göre tanımlabilir. Tüm hormonların sentezini düzenleyen hipotalamus ve çok sayıda farklı etkiye sahip hipofiz en fazla hormon üreten endokrin bezlerdir.
� Hormon salgılayan dokulara göre: 1. Hipotalamus hormonları 2. Hipofiz hormonları 3. Gonadotrop hormonlar üreme dokuları(testisler ovaryum salgılanan hormonlar) 4. Doku hormonları: Sindirim kanalında yapılırlar (mide, ince bağırsak).Yapıldıkları yer ile etki gösterdikleri yerler birbirine yakın veya uzaktır. Gastrin, Pankreosymin, Sekretin, Gip,Vip
o Hipotalamus hormonları
o Hipofiz hormonları
o Ön lop hormonları
o Orta lop hormonu
o Arka lop hormonları
o Tiroit hormonları T3, T4
o Paratiroit hormonu parathormon o Pankreas kormonları İnsülin glokugon
o Böbrek üstü bezi hormonları
o Adrenal korteks hormonları Aldesteron
o Adrenal medülla hormonları epinefrin,
o Cinsiyet bezleri hormonları
o Erkek cinsiyet hormonları
o Dişi cinsiyet hormonları
o Gastrointestinal, diğer doku hormonları (Gastrin, pankreosymin, sekretin, Gip, Vip)
o Hipofiz bezi-Hipotalamus
o Tiroid bezi
o Paratiroid bezi
o Böbrek üstü bezleri
o Gonadlar-cinsiyet bezleri
o Pankreas
o Diğerleri(Kalp, Plasenta)

442
5. Mediatör hormonlar: Organlarda veya kanda yapılırlar. Yapıldıkları yer ile etki gösterdikleri yerler birbirine yakın veya uzaktır Angiotensinogen, angiotensin, kinine, histamin, seratonin, prostaglandinler, neurotransmitter. 6. Glandüler hormonlar: Endokrin bezlerde yapılırlar (hypophyse ön lobu, böbrek üstü bezi, tiroit bezi, epiphyse, thymus). Dolaşıma salgılana bir hormon vücudun bütün bölgelerine gidebilir. Ancak hormonlar etkilerini sadece belirli hücrelerde ve dokularda gösterir. Bu hücrelere ve organlara hedef hücre veya organ denir. Hipotamus
Hipotalamus, beyinde talamusun altında bulunan ve üçüncü ventrikülün tabanını oluşturan önbeyin bölgesidir. Ektodermden gelişir. Hipotalamusta çok sayıda küçük nukleus vardır. Dört grupta toplanır: 1-Ön: Supraoptik ve paraventriküler nükleuslar 2-Orta: Tuber sinerum etrafındaki nükleuslar 3-Yan: Lateral nükleuslar 4-Posterior hipotalamik nükleuslar
ŞEKİL 8.27 Hipof,z hormonları üretim ve taşınması
Hipotalamusun en önemli görevlerinden birisi hipofiz bezi aracılığı ile beyin ve endokrin sistem arasındaki bağlantıyı sağlamaktır. Tüm omurgalılarda bulunur. İnsanda, kabaca bir badem şeklindedir. Memelilerde beyin merkezleri arasında ilinti sağlar. Vücut sıcaklığı mekanizmasını, sempatik sinir sistemini ve hipofizin çalışmasını denetler. Susama, acıkma hislerinin merkezi olup vücut ısısını ve kan basıncını ayarlar. Ayrıca ürettiği RF maddesi ile hipofizi uyarır. İç denge hipotalamus ile korunur. Karbonhidrat-yağ-protein metabolizmasını dengeler. Hipofiz, alt uç kısmında küçük bir yuvarlak durumundadır. Hipofiz arka lobunun salgıladığı antidiüretik hormon ile oksitosin denen madde hipotalamusta yapılıp hipofize aktarılmaktadır. Duyguların fiziksel temeli de hipotalamus tarafından oluşturulmaktadır. Hipofiz Hipotalamus İlişkisi
Hipofiz bezi kan damarları ve sinir lifleri ile hipotalamusla bağlantılıdır. Sinir sistemi ile endokrin sistem arasındaki direkt bir bağlantıdır. Hipotalamus ile adenohipfiz arasındaki bağlantı

443
hipotalamik-hipofiziel portal sistem adı verilen damar sistemi ile sağlanır. Hipotalamus ile nörohipofiz arasındaki bağlantı ise sinirler aracılığıyla gerçekleştirilir. Hipofiz Bezi, Beyinde hipotalamusun hemen altında bulunur. İki ayrı lobtan oluşmuştur; Anterior lob (adenohipofiz) ve Posterior lob (nörohipofiz). Adenohipofiz gerçek bir endokrin bezdir ve salgı hücreleri ihtiva eder. Nörohipofiz ise hipotalamustan köken alan pek çok sinir ucunun sonlandığı bölümdür.
ŞEKİL 8.28 Hipofiz ile hipotalamus arasındaki bağlantıi ve ekzokrin, endokrin kanal farklığı
Hipotalamus hormonları
Peptiderjik nöronlardan salgılanan, adenohipofiz hormonlarının sekresyonunu düzenleyen hormonlardır Hipotalamus hormonları( ön hipofizi düzenlerler). Hipofizer hormonların salınımı en az bir hipotalamik hormonun kontrolü altındadır. Hipotalamik hormonlar, hipofiz sapındaki hipotalamik – hipofizer sistem kapillerinin çevresinde sonlanan, hipotalamik sinir uçlarından salınır ve hipotalamus ile ön lobu birbirine bağlayan özgül bir portal sistem yoluyla ön loba ulaşır. Tümü peptid ve protein yapılı olan hipotalamik hormonlar ritmik biçimde salınırlar. Hipotalamik hormonların dolaşımdaki miktarları hipotalamik-hipofiz-hedef bez sisteminin etkileşimiyle düzenlenir. Örneğin GnRH, LH ve FSH salınmasını uyarırken dolaşımda yer alan ve hipotalamusa ulaşan gonad hormonlarının düzeylerinin artması GnRH’da negatif feed-back ile inhibisyona yol açar.
• CRH (Kortikotropin Salıcı Hormon) : Hipotalamustan salınır, hipofizden ACTH salgısını düzenler, hedef bezden ise hidrokortizon salgısını düzenler.
• TRH (Tirotropin Salıcı Hormon): Hipotalamustan salınır, hipofizden TSH salgısını düzenler, hedef bezden ise T3 ve T4 salgısını düzenler.
• GnRH ( Gonadotropin Salıcı Hormon): Hipotalamustan salınır, hipofizden LH, FSH salgısını düzenler, hedef bezden ise androjen, östrojen, progestin salgısını düzenler.
• GHRH (Büyüme Hormonu Salıcı Hormon): Hipotalamustan salınır, hipofizden GH salgısını düzenler, hedef bezden ise IGF-1 salgısını düzenler.
• GHRIH/SRIH (Büyüme Hormonu Salınımını İnhibe Eden Hormon; Somatostatin; Somatotropin Salınımını İnhibe Edici Hormon): Hipotalamustan salınır, hipofizden GH salgısını düzenler, hedef bezden ise IGF-1 ile T3-T4 salgısını düzenler
Hipofiz hormonları(trofik hormonlar)
Hipofiz veya diğer adıyla Pitürter bez bir fasülye tanesi büyüklüğünde yaklaşık 0,5 gram ağırlığında bir endokrin bezdir. Beyin tabanında, hipotalamusun altında bir çıkıntı şeklinde
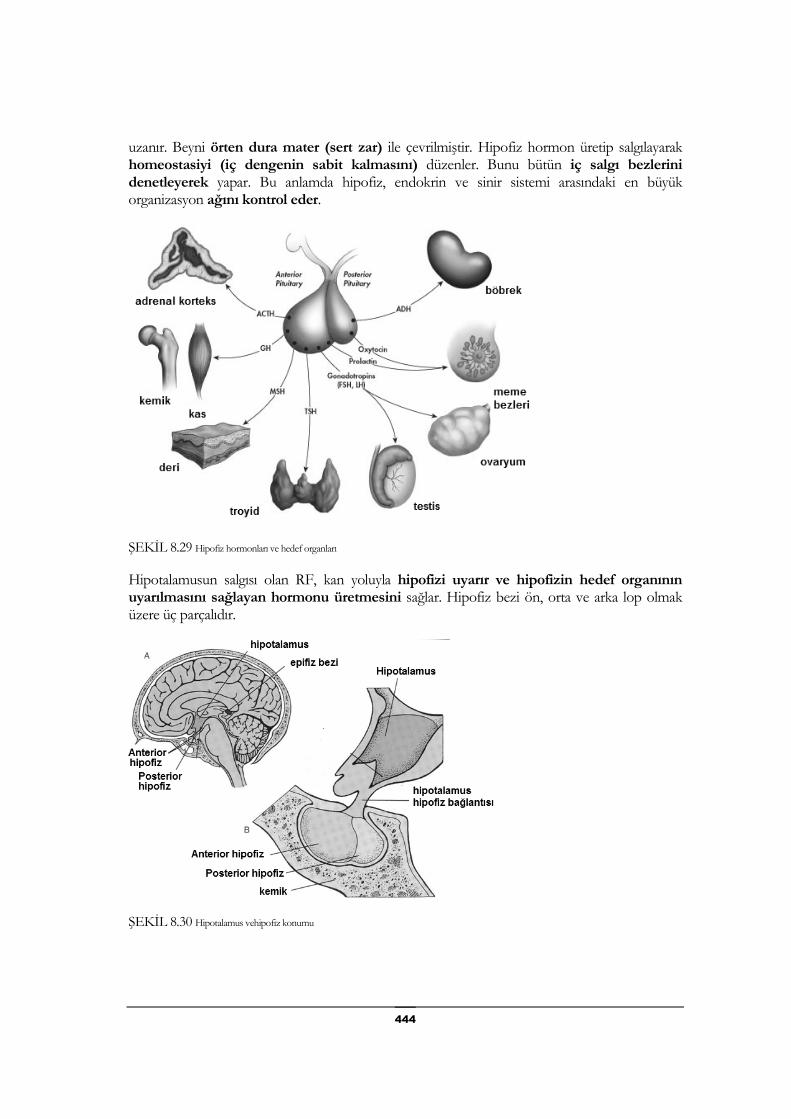
444
uzanır. Beyni örten dura mater (sert zar) ile çevrilmiştir. Hipofiz hormon üretip salgılayarak homeostasiyi (iç dengenin sabit kalmasını) düzenler. Bunu bütün iç salgı bezlerini denetleyerek yapar. Bu anlamda hipofiz, endokrin ve sinir sistemi arasındaki en büyük organizasyon ağını kontrol eder.
ŞEKİL 8.29 Hipofiz hormonları ve hedef organları
Hipotalamusun salgısı olan RF, kan yoluyla hipofizi uyarır ve hipofizin hedef organının uyarılmasını sağlayan hormonu üretmesini sağlar. Hipofiz bezi ön, orta ve arka lop olmak üzere üç parçalıdır.
ŞEKİL 8.30 Hipotalamus vehipofiz konumu

445
Hipofizzin ön lobu epitel doku, arka lobu sinir hücrelerinden oluşmuştur. Ön hipofizden (Adenohipofizden) salgılanan hormonların salgılanmasını düzenleyen salgılatıcı (releasing) ve salgıyı durdurucu (inhibiting) hormonlar salgılar. Bu hormonlar kan yoluyla hipofize gelir. Antidiüretik hormon (ADH) ve oksitosin hormonlarını sentezler, depolar ve nörohipofizden salgılatır. Bu hormonlar sinirin aksonu aracılığıyla (ADH için nörofizin II, Oksitoksin için nörofizin I ile kinezin taşıma proteini ile) hipofize gelir.
Antidiüretik Hormon (ADH)
Beynin hipotalamus bölümündeki Supraoptik çekirdekte özel sinir hücreleri tarafından sentez edilir. Bu hücrelerin uzantıları “Hipofiz Sapı” içinde ilerledikten sonra nörohipofize gelirler ve hipotalamustan taşıdıkları ADH’ yi buradaki özel hücrelere verirler. Nörohipofizdeki bu özel -hücrelere “Pituisit” denir. Pituisitler, ADH’ı depo ederler ve emir aldıklarında bu hormonu kan dolaşımına verirler. Antidiüretik hormon salgılandığı zaman böbreklerde hazırlanan idrar miktarındabir düşme olur ve bunun sonucu olarak da vücuttaki su miktarı artar. ADH diğer adıyla vazopressinin ana görevi vücut sıvı dengesinin düzenlenmesine katkıda bulunmaktır. ADH nın hedef organı böbreklerdir. Böbreklerden su geri emilimini artırarak, idrar yoluyla su atılımını azaltır.
ŞEKİL 8.31 ADH moleküler yapısıi
Oksitosin
Beyinde hipotalamusta paraventriküler çekirdekte sentezlene hormonudur. Arka hipofizden (nörohipofizden) salınır. Özellikle doğum esnasındaki ve doğum sonrasındaki rolü önemlidir. Doğum esnasında serviks ve uterusun gerilmesi ile çok miktarlarda salınır, rahim kaslarının kasılmasını uyarır ve doğumu kolaylaştırır. Doğumdan sonra ise meme başı uyarısı ile sütün salınımını sağlayarak emzirmeye yardımcı olur. Oksitosin salgılanmasındaki yetersizlik sosyopati, psikopati, narsisizm ve genel manipülasyon eğilimi ile ilişkili bulunmuştur.

446
ŞEKİL 8.32 Oksitoksin ve ADH hipotalamustan hipofize taşınarak sagılanması
Ön hipofiz hormonları
Pro-opiyomelanokortin ailesi hormonlar
Pro-Opiyomelanokortin (POMC) Peptid Ailesi: POMC ailesi hormonlar (ACTH, LPH, MSH) hormon gibi davranan veya nörotransmiter veya nöromodülatör (endorfinler) gibi davranan peptidlerden oluşur. POMC yaklaşık 285 amino asitlik bir peptid zinciri olarak sentezlenir ve hipofizin farklı yerlerinde farklı şekilde işlenir.
Kortikotropin (ACTH): Adrenokortikotropik hormon (ACTH) hipofizin ön lobunda üretilen polipeptid yapıda bir hormondur. 39 aa’li tek zincirli polipeptiddir. Adrenal kortekste etksini gösterir. Büyüme fonksiyonunu düzenler. Kolesterolün pregnanolona dönüşümünü arttırarak steroid yapımını uyarır. Plazma membranındaki reseptöre bağlanarak adenil siklaz aktivitesi ve cAMP yapımını arttırır. Hipotalamus-hipofiz-adrenal aksın önemli bir öğesidir. Genelde biyolojik strese karşılık olarak üretilir. (hipotalamustan CRH ,kortikotropin releasing hormon ya da kortikotropin salgılattırıcı hormon ile birlikte) En önemli etkisi, adrenal korteksten kortikosteroid hormonların ve kortizolün salgılanmasını uyarıcı etkisidir Melanosit stimüle edici hormon (MSH): Derideki melanositlerin melanin sentezini stimüle ederek cildin koyulaşmasını ve güneşte koruyuculuk yapmasını sağlar.

447
ŞEKİL 8.33 Kurbağada melanosit uyarıcı hormona bağlı renk değişim mekanizması
ββββ-endorfin: Hipofîz bezinde oluşan, beyin, pankreas ve plasentada bulunan, morfin reseptörlerine bağlanarak davranış değişikliğine yol açan, otuz bir amino asitlik bir polipeptitir. Ağrı duymayı bastırma yani analjezik etkisi vardır. Ancak endorfinlerin analjezik etkisi morfinden yaklaşık 30 kat daha fazladır. Bu nedenle bunlara ‘iç morfin’ anlamında endorfin denmektedir. Yoğun fiziksel aktiviteler endorfin düzeyini yükseltir.
Glikoprotein ailesi hormonlar
Tirotropin (TSH): Tiroid Uyarıcı Hormon veya Tiroid Simule Edici Hormon hipofiz bezinin ön lobundan(adenohipofizden) üretilen ve salgılanan, Tiroid bezinin çalışmasını düzenleyen peptid yapıda bir hormondur.

448
ŞEKİL 8.34 Troid sentezi ve kontrolü proteini
Gonadotropinler Hipofizin ön lobundan salgılanan ve gonadların faaliyetini uyaran çeşitli hormonlar. Folikül uyarıcı hormon (FSH), lütein hormonu (LH), plâsenta tarafından çıkarılan koryon gonadotropini gibi.
Luteinizan hormon (LH): Beynin ön hipofiz bölgesinden salgılanır. Erkekte : sperm yapımı ve testesteron salgılanmasını sağlar, kadında : kadınlık hormonlarının üretimi ve yumurta olgunlaşmasını sağlar. Follikül stimüle edici hormon (FSH) erkekte Testilerin tubuli konkorti seminiferi epitelini olgunlaştırarak spermatogenezin oluşmasını sağlar. Kadında FSH sürekli salgılanmaz, gelişen folikülden salgılanan östrojenin düzeyiyle ilişkilidir. Östrojen kanda yükselince negatif feedback ile FSH düşer. Siklusun birinci günü FSH yüksek östrojen düşüktür, 14. günde östrojen yüksek FSH düşüktür. FSH siklusun 25. günü yeni bir siklusu düzenlemek için tekrar yükselir.
Somatomammotropin ailesi hormonları
Somatotrop hormon (Büyüme hormonu, GH): Ön hipofizden salgılanan, peptit yapılı, insanlarda ve hayvanlarda büyüme, hücre üretimi ve yenilenmesini uyaran hormondur. Ön hipofizin somatotropik (asidofilik) hücrelerinde 191 aminoasitlik tek bir polipeptit zincir şeklinde üretilmektedir. GH sentez ve salınımı, hipotalamustan salgılanan GH-releasing hormon (Gh-RH) tarafından

449
kontrol altında tutulmaktadır. Somatostatin, GH salınımını azaltır. Bunun yanında insülin, glukagon, TSH, FSH, ACTH gibi hormonlar da salınımını baskılamaktadır. Egzersiz, stres ve uykunun derin döneminde artış gösterir. GH dokuları doğrudan etkilemez. Etkilerini somatomedin denilen peptitler, özellikle somatomedin C (İnsülin-like growth faktör-I; IGF-I) aracılığı ile gösterir. GH kıkırdak yapımını arttırmakta ve uzun kemiklerde büyümeyi sağlamaktadır. Bu yüzden çocukluk döneminde büyük önem arz etmektedir. Eksikliği büyümede yetersizliğe yol açar ve değişik tipte cücelikler görülür. Büyüme hormonu aşırı salınımı (genellikle hipofiz tümörüne bağlı) uzun kemik uçlarındaki epifiz plaklarının kapanmasından önce orantılı olarak aşırı büyümeye (gigantizm), epifiz plaklarının kapanmasından sonra ise akromegali hastalığına neden olur.
ŞEKİL 8.35 GH Büyüme hormona ve bağlı olarak karaciğerde salgılana IGF-1 faktörü.
Prolaktin (PRL): Prolaktinin kadınlarda iki görevi vardır; Östrojen (dişi cinsiyet hormonu) ile birlikte gebelikte meme bezlerinde meme kanallarının gelişimini uyarır. Doğumdan sonra meme dokusunda süt üretimini uyarır.
Opiyomelanokortin ailesi ön hipofiz hormonları (POMC)
Pineal bez (Epifiz) hormonu (melatonin)
Pineal bez, beyin yarımkürelerinin arasında, diencephalonun tavanında yerleşmiş, konik-çam kozalağı şeklinde küçük bir organdır. 7x5x4 mm boyutlarında. 100–180 mg ağırlığındadır. Pineal bez, karmaşık bir polinöronal yol izleyerek-retina üzerine düşen çevresel ışığa ait bilgileri alıpbuna cevap vermektedir. Işıkla ilgili sinyalleri endokrin sinyallere dönüştüren nöroendokrin transduser olarak kabul edilir. Epifiz bezindeki pinealositler tarafından salgılanan melatonin ve seratonin adenohipofiz, nörohipofiz, endokrin pankreas, adrenal korteks, adrenal medulla, paratiroid ve gonadlarüzennde genellikle inhibitor etki yapar. Karanlık pineal bezde aktivite artırıcı rol oynarken, .aydınlık azaltıcı rol oynamaktadır. Pineal bezdeki aktivite artışı, etkilediği iç salgılı bezlerde aktivite azalmasına neden olur.

450
ŞEKİL 8.36 Epifiz bezinin ışık ile kontrol edilmesine bağlı melatonin sentezi
Pineal bezin ayrıca uyku periodu, vücut ısısının ayarlanması, metabolizma, immün sistem, tümör büyümesi (inhibisyon), lokomotor aktivite, beyin transmitter metabolizması vb. daha birçok fonksiyonda rol oynadığı ileri sürülmektedir.
Melatonin epifiz bezinin pineolasit adı verilen hücrelerinden salgılanır. Biyoritmi (sirkadyan ritm) belirler ya da biyoritm üzerinde etkilidir. Hayvanlarda fizyolojik ve davranışsal özelliklerin çoğu ritmik sikluslar içinde dalgalanmalar gösterir. Bu siklusların çoğu sirkadyan ritmi izlerken bazıları daha kısa bir ritim (ultradian ritim) izler. Sirkadyan ritim Üç özelliğe sahiptir: Ritmin osilasyonları yaklaşık 24 saatte bir tekrarlanır. Ritim gözden beyne gelen ışık sinyalleri ile kendini yıl içerisindeki gece gündüz farklılıklarına adapte edebilir. Bu salınım lokomotor aktiviteden uyku-uyanıklığa kadar bir çok davranışı kontrol eder. Memelilerde sirkadyan ritim suprakiazmatik nukleus tarafından kontrol edilir. Pineolasit hücreleri ışığa duyarlıdır. Elektromanyetik dalga yoğunluğu arttıkça melatonin salgılanması azalır. Melatonin bir tür etanoamiddir. Melatonin, kişiden kişiye değişse de yaklaşık olarak 23:00 ile 05:00 saatleri arasında salgılanan bir hormondur. Hormonun temel görevi vücudun biyolojik saatini koruyup ritmini ayarlamaktır. Bunun haricinde melatoninin güçlü salgılanmasının kansere karşı koruyucu etkisi vardır. Bu nedenle lösemi ve diğer kansere yakalananların kesinlikle karanlık ortamlarda yatırılmaları istenmektedir. Yapılan son araştırmalara göre hormonun yaşlanmayı geciktirici etkisi de vardır. Böbrek hormonları (Eritropoietin, 1,25 kalsiferol)
Eritropoetin: Eritropoetin (ya da EPO) eritrositler (alyuvar) için sitokin görevi gören bir glikoprotein hormondur. Hematopoetin ya da hemopoetin olarak da adlandırılır. Böbreklerde, renal kortekste tübülüsler arasındaki dar interstisyel aralıkta yer alan fibroblast benzeri hücrelerce üretilir ve eritrosit üretiminin kontrolünden sorumludur. Böbrek tarafından üretilen glikoproetin yapısındaki bir hormondur. Böbrekler kemik iliğinde eritrosit üretimini kontrol eden eritropoetin hormonun yüzde 90 kısmı üretir. Eritropoetin hormonu salgılanması hipoksi (anemi, arteryel hipoksi, yetersiz renal kan akımı vs) uyaranıyla başladığı bilinmektedir. Eritropoetin kemik iliğini uyararak eritrosit üretimini artırır. Azalmış oksijene cevap larak EPO seviyesi artar. EPO, kemik iliğini RBC yapımı için uyarır ve artışına neden olur. Birçok anemi tipinde normalden yüksektir. Eritropoetin, memeli hücre kültüründe rekombinant DNA teknolojisi ile üretilerek tedavi amaçlı olarak da kullanılmaktadır. Böbrek yetersizliğine ya da kanser kemoterapisine bağlı anemilerin (kansızlık) tedavisinde yeri

451
vardır. Ayrıca bisiklet yarışı, triatlon ve maraton koşusu gibi dayanaklılık gerektiren sporlarda doping amacı ile suistimal edilmektedir.
Böbrekte Eritropoietin dışında, 1,25(OH)2D3 (1,25-dihidroksikolekalsiferol), Prekallikreinler, Prostaglandinler Renin enzimi sentezini yapar. Vitamin D’ nin dolaşımdaki öncülünü (1,25 – hidroksikolekalsiferol) de aktif vitamine dönüştürürler. Jukstaglomerular hücreler, afferent arteriyolün (nadiren efferent arteriyol) tunica media tabakasında bulunan değişikliğe uğramış düz kas hücreleridir. Çekirdekleri oval yerine yuvarlaktır. Sitoplazmalarında bir proteolitik enzim olan renin granülleri içerirler. Ayrıca bu hücrelerde anjiyotensin-dönüştürücü enzim (ACE), anjiyotensin I ve anjiyotensin II de bulunur.
ŞEKİL 8.37 Böbreklerin hormonal fonksiyonu
Timus bezi hormonları
Timüs organı, tiroid bezinin altında, göğüs boşluğunda ve soluk borusunun önünde bulunur. Timüs bezi bağ dokusundan yapılmış ince bir kapsülle çevrilmiştir. Kapsül, diğer lenfoit organlarda olduğu gibi bezin içine girerek onu bölmelere ayırır. Timüs bezinin bölmelerinde, retiküler hücreler ve lenfositler bulunur. Kan, lenf damarları ve sinirler bağ doku bölmeleri boyunca uzanır. T lenfositlerinin oluşumu ile ilgili hormon ve maddeler salgılar.
ŞEKİL 8.38 Timus bezi ve trioid bezlerinin konumlarıı
Kalpten salgılanan ANP hormunu
Kalp antrium kısmında yer alana bazı kas hücreleri ANP adı verilen bir hormon sentezler, depolar ve salgılar. ANP, Kulakçık kaslarının gerilmesi, kan hacmi ve basıncının artması ve aşırı tuz alınması sonucu kalbin kulakçık kasları tarafından yapılan ve salgılanan, kan hacminin ayarlanmasında, böbreklerle sodyum çıkarılmasında, idrar söktürücü, damar genişletici ve kan basıncının düşürülmesinde önemli rol alan peptit yapısında bir

452
hormondur. ANP güçlü bir damar genişleticidir ve kalp kası hücreleri tarafından salgılanan bir protein (polipeptid) hormondur. Vücut su, sodyum, potasyum ve yağ dokusu homeostatik kontrolüne katılır. Yüksek kan hacmine yanıt olarak kalp (atriyal miyositler) üst odaları (kulakçıklar) kas hücreleri tarafından salınır. ANP kan basıncı düşürür ayrıca dolaşım sistemi üzerinde, su, sodyum ve yağ yükleri azaltır. Son zamanlarda natriüretik peptidlerin beyin, akciğer ve böbrek olmak üzere çeşitli dokularda da sentezdiği bulunuştur. Böbrekte sentez edilen natriüretik peptid ANP’ye çok benzer olup, renal natriüretik peptid veya ürodilatin olarak adlandırılır. Ürodilatin idrarda bulunur, ancak plazmada yoktur. Olasılıkla distal kortikal nefronda sentez edilmektedir. ANP natriüretik etkisini hem GFR’yi artırarak hem de sodyum transporterlerin etkisini azaltarak, yani sodyum geri emilimini baskılayarak yapar. ANP’nin toplayıcı kanallardaki en belirgin etkisini luminal plazma membranı sodyum kanallarının açılma zamanını kısaltarak sodyum geri emilimini etkileyerek yapar. Hem sodyum taşıyıcıları ( Na+-H+ değiştirici ve Na+-fosfat transporter) hem de pompalar (Na-K-ATPaz) proksimal tübülüste ANP’nin hedef proteinleridir.
ŞEKİL 8.39 ANP, BNP ve CNP etkisi
Plasenta hormonları Koryonik gonadotropin (hCG)
Plasenta gebelikte fetüsün beslenmesi için gelişen özelleşmiş bir organdır. Koryonik gonadotropin (hCG), plasentada sentez edilir, hamileliğin ilk 4-6 haftasında korpus luteumun devamlılığından sorumludur. Plasenta östrojen, progesteron ve human koryonik gonadotropin hormonlarını salgılar. Bu hormonlar gebeliğin devamlılığına yardım ederler. Bu hormonların hedef organları overler, meme bezleri ve uterustur.
Şekil 8.40 Blastula tarafından salgılana hCG hormunu etkisi.

453
Tiroid Bezi hormonları
Tiroid bezi boyunda trakeanın önünde iki lob halinde bulunur. Larinksin hemen altında, trakeanın önünde iki tarafta yer alan ve erişkinde 15-20 gram ağırlığında olan tiroid bezi, vücuttaki en büyük endokrin bezlerden biridir. Tiroid bezi, T4 ve T3 olarak adlandırılan tiroksin ve triiyodotironin hormonlarını salgılar. Bu hormonlar vücutta metabolizma hızını artırmada önemli rol oynarlar.
ŞEKİL 8.41 Troid bezi
Triod salgılanması için gerekli olan iyot folikül hücrelerince kandan aktif taşıma ile kolloidler içine pompalanır. TSH hormunu bu pompalama işlemi ile hormon üretimi artırır. Tiroid hormonlarının oluşumunda gerekli ilk adım daha sonra direkt olarak tirozin amino asidiyle birleşebilecek iyodür iyonlarının ya yeni oluşan iyot ( 1°) ya da I3şeklinde okside iyoda dönüşümüdür, iylodinin bu oksidasyonu hidrojen peroksitle birlikte, iyodürleri okside edebilen güçlü bir sistem oluşturan, peroksidaz enzimiyle sağlanır. Tiroid hücrelerinin apikal yüzeyi kolloidin ufak bir bölümünü içine alan yalancı ayaklar gönderir. Bunlar tiroid hücresinin apeksinden içeri giren pinositik vezikülleri oluşturur. Daha sonra lizozomlar bu veziküllerle birleşerek kolloidle karışmış şekilde lizozom sindirim enzimlerini kapsayan, sindirim veziküllerini oluştururlar. Bu enzimlerden proteinazlar tiroglobulin molekülünü sindirir ve tiroksinle triiyodotironini salgılar. Bunlar daha sonra difüzyonla tiroid hücresinin tabanından onu çevreleyen kapillerlere geçer. Böylece tiroid hormonları kana verilmiş olur. Tiroid salgısının tam yokluğu, genellikle bazal metabolizma hızının normalin yüzde 40-50'si kadar düşmesine ve tiroid salgısının aşırı fazlalığı bazal metabolizma hızının normalin yüzde 60-100’ü kadar artmasına yol açar.

454
ŞEKİL 8.42 T3 ve T4 sentezi
Tiroid salgısı esas olarak ön hipofiz bezi tarafından salgılanan tiroid stimüle edici hormon (TSH) tarafından kontrol edilir. Tiroid bezinin folliküler hücrelerinden; Tiroksin (tyroxine)(T4), Tri-iodotironin (triiodothyronine) (T3), Tiroid bezinin parafolliküler hücrelerinden; Kalsitonin hormonları salgılanır.
ŞEKİL 8.43 T3 ve T4 hormunui
Tiroid hormonları TSH tarafından feedback mekanizmalar ile düzenlenir. T4 kanda düşükse, soğuk, stres ve gebelik gibi durumlar var ise ki bu durumlarda enerji ihtiyacı artar, TSH salgısı uyarılır, o da tiroid bezini uyarır. Tiroid Bezi Salgısı, % 90 ı tiroksindir (T4). T4 ve T3 tirozin amino asitine iyot bağlanmasıyla oluşur. Hemen hemen tüm hücrelerde reaksiyonların hızını artırarak, vücut metabolizma hızını yükseltirler. Tiroid Hormonlarının Etkileri: Vücudun pek çok hücresinde hücresel reaksiyonları hızlandırır. Bazal metabolizma hızı artar, büyüme hızlanır, hücresel farklaşma ve protein sentezi uyarılır. karbonhidrat ve yağ metabolizması uyarılır (kalorijenik etki).Tiroid bezi aktivitesinin artmasına hipertiroidi, azalmasına ise hipotiroidi denir.

455
Kalsitonin
Kalsitonin kan kalsiyum düzeylerini düşüren bir hormondur. Kemik hücresinde kemik yıkımını azaltır, kemikten kana kalsiyum geçişini azaltır, kandan kemiğe kalsiyum geçişini artırır, kan kalsiyumunu düşürür. Bu hormon tiroid bezinin crista neuralis kökenli C hücreleri tarafından üretilir. 32 amino asitlik tek bir polipepdir zincirden oluşur. Karboksil ucundaki prolin bir amid grubu ile bloke edilmiştir ve biyolojik aktivite için mutlaka gereklidir. Kalsitonin salgısının kalsiyum ile kontrolü PTH için söylenenlerin aksi yöndedir. Kalsitonin hipokalsemik etkisini primer olarak, osteoklastların aktivitelerini bastırarak meydana getirir. Buna bağlı olarak kemiklerden, kana kalsiyum geçişi de inhibe edilmiş olur. Ayrıca, kemik oluşumunda ve kalsiyum tutulmasında uyarıcı etkisi vardır. Ancak bu etki çok kuvvetli değildir. Kalsitoninin salınmasını, paratiroid hormonunun salınmasını bastırıcı özelliği olan, hiperkalsemi uyarır. Yani, hücre dışı sıvıda yüksek oranda kalsiyum bulunması sonucu, parafolikülar hücreler uyarılırlar ve kalsitonin salınır.
• Kalsitonin esas olarak kan plazmasındaki kalsiyumun yoğunluğunu azaltmada fonksiyonel olan bir hormondur. Ayrıca, fosfat, sodyum, potasyum ve magnezyum iyonlarının idrarla atılmalarını uyarır.
• Diüretik etkisi vardır. Etki, proksimal tübül hücrelerinde cAMP miktarının artmasıyla meydana gelir. Böylece, kalsitonin bu bölgede su ve elektrolitlerin reabsorbsiyonunun azalmasına neden olur.
• Özetle, kalsitonin kemik rezorbsiyonunu yavaşlatır ve böbrekten kalsiyum atılmasını arttırır. Bu olayların her ikisi de, hipokalsemiye neden olur. Hipokalsemik etki süt verme, gebelik, açlık, kol ve bacaklarda uzun süre devam eden hareketsizlik, hibernasyon gibi olaylarda iskeletteki kemiklerde rezorbsiyonun en alt düzeye inmesi gerektiği zamanlarda, fonksiyonel yönden önem taşır.
Kalsitonin salgısı hiperkalsemi ile uyarılır ve hipokalsemi ile inhibe olur. Ayrıca gastrin ve kolesistokinin de kalsitonin salgısını uyarırlar. Kemik yapımının hızlı olduğu durumlarda kalsitonin plazma kalsiyumunu hızla düşürür. Kalsitonin primer hipokalsemik etkisini osteoklast aktivitesini inhibe ederek ve kemik rezorbsiyonunu önleyerek gösterir. Büyüme ve laktasyon gibi stres durumlarında kalsitonin kemiği koruduğu kabul edilir. Kalsitoninin osteoklastlar üzerindeki etkisi bir glikoprotein olan spesifik reseptör aracılığı ile olur. Reseptör adenilat siklaz üzerinden hücre içi cAMP düzeylerini artırarak etki gösterir. Hiperkalsemi, yani kan kalsiyum düzeyinin yüksekliği klinik bulgular olmaksızın seyreder. Tedavi edilmez ise böbrek taşlarının, böbrek yetmezliğinin, osteoporoz, kemik kistleri, pankreatit ve koma oluşur. Hiperkalseminin en önemli nedeni hiperparatiroidizmdir. Bunun dışında hiperkalsemi meme kanseri ve diğer adenokarsinomlarda da sık gözlenen bir bulgudur. Hipokalsemi genellikle paratiroid yetmezliği sonucu ortaya çıkar. Hastalarda plazma kalsiyum düzeylerindeki düşklüğe karşın fosfat düzeyleri yüksektir. Düşük kalsiyum titremelere ve tetaniye neden olur. Kramplar ve ölüm gelişir. Kalsiyum ve fosfat (PO4)x metabolizması, birbirleriyle çok yakın bir ilişki içindedir. Hem kalsiyum hem de fosfat dengesinin düzenlenmesinde, büyük ölçüde dolaşımdaki paratiroid hormonu (PTH), vitamin D ve bunlar kadar olmasa da kalsitonin hormonunun etkileri vardır. Kalsiyum; kemik ve dişlerin şekillenmesi, kasların kasılması, kanın pıhtılaşması, sinir uyarıla- rının iletisi ve hormon salınması gibi, pekçok fizyolojik olayda anahtar rol oynar. Bu

456
nedenle kalsiyum dengesinin korunması kritik önem taşır. Vücuttaki kalsiyum depoları (iskelet sistemi) ve plazma kalsiyum konsantrasyonunun korunması; besinlerle kalsiyum alımına, gastroin- testinal kanaldan kalsiyum emilimine ve böbreklerden kalsiyum atılımına bağlıdır. Dengeli bir beslenmeyle günde yaklaşık 1000 mg kalsiyum alınır. Bu da sütün 1 litresindeki miktara eşittir. Kalsiyumun esas atılımı dışkı ve idrar ile olmaktadır. Bunun yanısıra, barsaktan geri emilim de olmaktadır. D vitamini, kalsiyumun barsaklardan emilimini arttırır. Böbreklerde aktif vitamin D sentezi arttırılarak, barsaktan kalsiyum emilimi arttırılır. Böbreklerde bir hasar mevcutsa, D vitamini etkisinin büyük bir bölümünü kaybeder ve barsak emilimi de azalır.
Paratiroid Bezi hormonları
Paratiroid bezler, herbir tiroid lobunun arka kenarı üzerinde yerleşmiş, mercimek şeklinde, toplam 4 adet iç salgılı bezdir. Konumlarına göre üst ve alt paratiroid bezler olarak adlandırılırlar. Paratiroid bezler, gevşek bir kapsülle sınırlanmış olup, parankimi sinuzoidal kapillerler arasında yer alan epitel hücre kordonlarından yapılıdır. Hücre kordonlarında, esas ve oksifil hücrelerbulunur. Esas hücreler kan kalsiyum düzeyini artıran parathormonu salgılarlar. Parathormon yaşam için mutlak gerekli olan bir hormondur. Parathomonun etkili olabilmesi için uygun miktarda D vitamini alınması ile böbreklerde üretilen dihidroksivitamin D3′e ihtiyaç vardır.
Paratiroid bezler tiroid bezinin arkasında bulunan küçük bezlerdir. Parathormon (paratiroid hormon) (PTH) salgılarlar. PTH kalsitoninin tersi etkilere sahiptir. Kan kalsiyum düzeylerini artırıp, fosfat düzeylerini azaltır. Kan kalsiyum miktarı azaldığı zaman PTH artar. Kan kalsiyum düzeylerini artırıp, fosfat düzeylerini azaltır.
ŞEKİL 8.44 Parathormon
Paratiroid hormonu; kalsiyum ve fosfat’ın barsaklardan reabsorbsiyonunu, böbreklerden atılmalarını ve ekstrasellüler sıvı ile kemikler arasındaki değişimleri düzenleyen bir hormondur. Paratiroid salgılığı (bezi) aktivitesinin artması, kemikten kalsiyum tuzlarının hızla rezorbsiyonuna yol açarak, ekstrasellüler sıvıda hiperkalsemi oluşturur. Bunu osteoklast aktivasyonu ile kemik rezorbsiyonu yani kalsiyumun mobilizasyonu arttırarak yapar. Bunun aksine, paratiroid salgılıklarının hipofonksiyonu, hipokalsemiye neden olur.

457
D vitamini, kemik rezobsiyonu (yıkımı) ve kemik depolanması (yapımı) yani remodelasyon üzerinde önemli etkilere sahiptir. Aşırı miktarda vitamin D fazlalığında, kemiklerde re- zorbsiyon oluşur. D vitamini eksikliğinde, paratiroid hormonunun kemik rezorbsiyonu üzerine olan etkisi büyük ölçüde azalır. Kan kalsiyum miktarı azaldığı zaman PTH artar. Paratiroid bezler tarafından salgılanan paratiroid hormon (PTH) disülfit bağı içermeyen 84 amino asitlik tek bir polipepdit zinciri olarak salgılanır. Bazik amino asitlerin yapıya yüksek miktarda katılması proteine bazik bir karakter kazandırır. İnsülin ve pek çok pepdit hormon gibi prehormon olarak sentezlenir. PTH plazma kalsiyum düzeylerini artırdığı için düşük kalsiyum düzeyi ile (hipokalsemi) sentezlenmesi uyarılır. Yüksek kalsiyum düzeyi ile de sentez inhibe olur. Ayrıca D vitamininin en aktif formu olan 1-25 dihidroksikolekalsiferol PTH sentezini inhibe eder. Beta adrenerjik ajanlar ve düşük plazma magnezyum düzeyleri de PTH salgısını artırır. PTH hücre zarındaki spesifik reseptörü ile etki gösterir. PTH adenilat siklazı aktive eder. Yükselen hücre içi cAMP düzeyleri sonuçta böbrek tübüllerinden kalsiyumun geri emilimini artırır. Böbreklerden kalsiyum atılımı azaltılırken fosfat atılımı artılrılarak plazma fosfat düzeyleri düşer. Bu durum 25 hidroksikolekalsiferolün hidroksilasyonunu uyararak D vitaminin aktif formunun yapımını uyarır. Parathormonun böbrekler üzerine olan etkisinin toplam sonucu, plazma kalsiyum düzeyinin artması, plazma fosfat düzeyinin düşmesi ve D vitamini aktivitesinin artmasıdır. D vitamini kemiklere ve barsaklara etki ederek plazma kalsiyum düzeyini yükseltir. Böbrek Üstü Bezleri (Adrenal Bezler) hormonları
Böbreküstü bezleri, anatomik ve fizyolojik yönden dışta korteks (Cortex), içte medulla olmak üzere iki kısımdan yapılıdır. Korteks, glukokortikoidler (Kortizol ve kortikosteron), minerelokortikoidler (Aldosteron) ile seks hormonları (özellikle androjenler) salgılar. Böbrek üsütü bezlerinin Medulla kısmı vücudumuzun en büyük paraganglionu niteliğinde olup sempatik uyarı ile adrenalin ve noradrenalin salgılar. Böbreküstü bezi, yaşam için zorunlu olan bir bezdir. Özellikle ekstrasellüler sıvının su ve elektrolit dengesini ayarlayan aldesteron hormonu ayrı bir öneme sahiptir.
ŞEKİL 8.45 Böbrek üstü bezlerinin insanda ve diğer memelilerdeki konumu
Böbrek üstübezleri diğer adıyla adrenal bezler böbreğin üst ucunda lokalize olmuşlardır. Her bir adrenal bez iki farklı bezin bileşiminden oluşmuştur. Medülla (iç bölümü), adrenal medulla kortex (dış bölüm, medullanın çevresi), adrenal kortex. Medulla ve kortex farklı hedef organları olan farklı hormonlar sentezler.
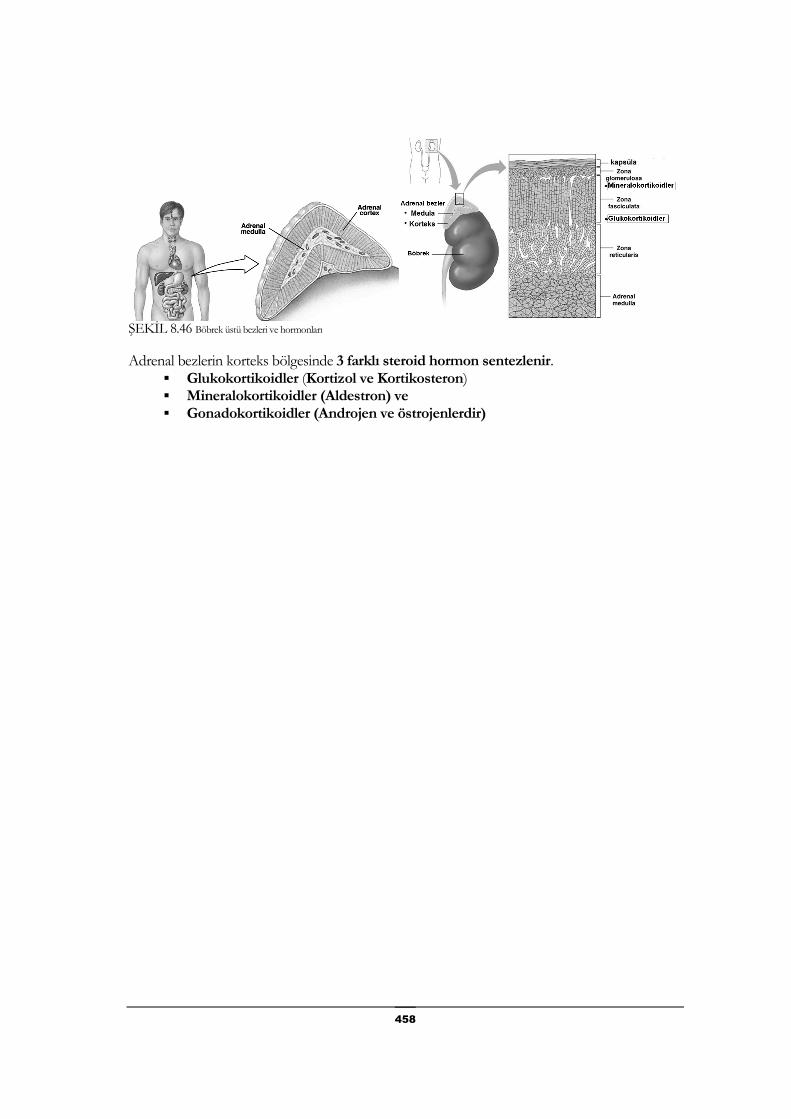
458
ŞEKİL 8.46 Böbrek üstü bezleri ve hormonları
Adrenal bezlerin korteks bölgesinde 3 farklı steroid hormon sentezlenir. � Glukokortikoidler (Kortizol ve Kortikosteron) � Mineralokortikoidler (Aldestron) ve � Gonadokortikoidler (Androjen ve östrojenlerdir)

459
Glukokortikoidler
Glukokortikoidler hormonlar 2 tiptir; Kortizol ve Kortikosteron. Bu hormonların etki dereceleri farklıdır. Glukokortikoid etkinin % 95 i kortizole aittir. Glukortikoidler karbonhidrat, protein ve yağ metabolizması için gereklidir. Glukoneogenezi (karbonhidrat dışındaki kaynaklardan karaciğerde glikoz sentezlenmesi) uyarır. Allerjik reaksiyonları ve inflamatuvar cevapları baskılar. Bu özelliği nedeniyle inflamtuvar giderilmesi, alerjik reaksyonların engelenmesinde kortizol ve türevleri ilaç olarak kullanılmaktadır.
Mineralokortikoid
En önemli mineralokortikoid aldosterondur. Aldosteron sodyum, potasyum geri emilimi aracılığıyla mineral dengesini kontrol eder. Mineralokortikoid aktivite; Ter, tükrük, idrar ve mide salgısı gibi sıvılarda sodyum geri emilimini dolayısıyla da su geri emilimini artırmaktır. Aldesteronun en önemli görevi böbrek distal tüpülerde Na ve Cl emilimi düzenlenmesidir. Buna bağlı olarakta suyun miktarı düzenlenir.
Gonadokortikoidler
Gonadokortikoidler cinsiyet organlarını etkilerler, fakat etkileri hafiftir. Bu hormonlar androjenler ve östrojenlerdir. Adrenal Medulla: Stres durumunda uyumu sağlayan Epinefrin (adrenalin) ve norepinefrin (noradrenalin) salgılanır. Medullanın ana salgısı epinefrindir, az miktarda norepinefrin salgılanır. Bu iki hormon sempatik sinir sistemini stimüle eder. Epinefrin ve norepinefrin sempatik sinir sitemine benzer etki yaparlar. Hormonal etki daha uzun sürer. Medulla hormonları stres hormonlarıdır.
Gastrointestinal hormonlar
Gastrin mide mukozasından salınır ve midenin asit salgılamasını uyarır. Sekretin duedonum mukozasından salınır ve pankreastan bikarbonattan zengin sıvı salgılatarak mideden gelen içeriğin asiditesinin nötralize edilmesini sağlar. Kolesistokinin de duedonumdan salgılanır ve safra kesesinin kasılmasını uyarır. Safra keseside yağ sindiriminde görev alan safrayı salgılanmasına neden olur.
Pankreas hormonları
Pankreas sindirim kanalına enzim salgılayan ekzokrin bir bez ve kana hormon salgılayan endokrin bir organ olarak işlev görür. Hormonlar pankreasın endokrin bölümündeki pankreatik adacıklarda sentezlenir, depolanır ve salgılanır. Endokrin pankreas dokusunu, gevşek bağ dokusundan yapılı zemin maddesine gömülü çok sayıdaki hücreler oluşturur. Etrafını çeviren ince bir bağ dokusu kapsülü, endokrin dokuyu, çevresindeki ekzokrin dokudan ayırır.
ŞEKİL 8.47 Pankreas hormonları
• Beta hücreleri (% 68) • Alfa hücreleri (% 20) • Delta hücreleri (% 10) • PP (pankreatik polipeptid
hücreleri (%2) • Serotonin hücreleri (nadir)

460
Pankreastaki bu endokrin yapılara, ilk tanımlayan araştırıcının adına atıfla, Langerhans adacıkları denir. Langerhans adacıklarının yaygın kan damarı ve sinir donanımları vardır. Kan damarlarının yaygın oluşu, adacık hücrelerinden boşalan salgı materyalinin (hormonların) çok çabuk olarak dolaşıma geçmelerini sağlar. Sinir uçları, adacıktaki hücrelerle çok yakın ilişkidedirler ve adacık hormonlarının sentez ve boşalmalarını kontrol etmede fonksiyoneldirler. İnsan pankreası, eksokrin doku arasına serpiştirilmiş yaklaşık 2.000.000 kadar Langerhans adacığı içerir. Bu kadar sayıdaki Langerhans adacığı, total pankreas dokusunun %1-2' ini oluşturur. Langerhans adacığında dört farklı tip hücre bulunur. Bunlar A, B, D ve PP hücreleridir. A (alfa hücreleri), glukagon, B (beta hücreleri) insulin, D (delta hücreleri) somatostatin, PP (F hücreleri) pankreas polipeptidlerini salarlar. .
ŞEKİL 8.48 Pankreas insülin salgılan mekanizması
Glukagon
Glikojenin glikoza çevrilmesini (glikojenoliz) uyararak kan glikozunu yükseltir. Yağ dokusundan yağ asidi ve gliserol salınımını uyararak enerji için yağ kullanımını teşvik eder. Bu nedenlerle glukagon hiperglisemik bir hormondur. Glukagon pankreasta langerhans adacıklarının a-hücrelerinde sentez edilir. Bütün memelilerde aynı olan 29 amino asitlik bir pepdittir. Düşük kan glikoz konsantrasyonuna cevap olarak salınır.
ŞEKİL 8.49 Kanda şeker ayarlanması

461
Glukagonun primer hedefi karaciğerdir. Öncül etkisi karaciğerde cAMP seviyesini artırmaktır. Normalde hipoglisemiye yanıt olarak gelişen glukagon artışı ile birlikte insülin salgısında da bir azalma olur. Stres hormonları olan epinefrin, GH ve kortizol glukagon salgısını direkt olarak artırırlar. Vazopressin ve beta-endorfin’de glukagon salgısını uyarır. Somatostatin ise inhibe eder. Glukagon hedef hücre yüzeyindeki spesifik reseptörlerine bağlanarak etki gösterir. Adenilat siklaz üzerinden gelişen cAMP artışı, protein kinaz-A’yı uyararak pek çok enzimin aktivitesinde değişikliklere neden olacak bir seri fosforilasyona neden olur. Karaciğerde glukagon glikojen fosforilazın fosforilasyonunu uyarır ve glikojenin, glikoz–1-fosfat ve glikoza yıkılmasını artırır. Aynı esnada glikojen sentetaz fosforilasyon ile inaktive edilir ve glikojen sentezi durur. Bunlardan glukagon, glikoneogenik yollara da etki eder. Fruktoz–2,6-bifosfatın düzeylerinin azaltılması ile glikoneogenezisin aktive olurken ve glikoliz inhibe edilir. Sonuçta karaciğerde hem glikogenoliz ve hem de glikoneogenezis uyarılarak glikoz yapımı artırılır. Glukagon karaciğerde aynı zamanda piruvat kinazın inhibisyonunu meydana getirir ve fosfoenol piruvatın birikmesine neden olur. Piruvat seviyesi azalır. Hem fosfoenolpiruvattan sentezi bloke edilmiş, hemde piruvat karboksilaz ve fosfoenol piruvat karboksikinaz reaksiyonları aracılığı ile PEP’a dönüştürülmesi devam etmektedir. Fosfoenol piruvatın birikmesi glukoneogenezizi uyarır. Glukagon adipoz dokuda cAMP düzeyini de artırır ki bu etkisi triacylgliserolün mobilizasyonun uyarılması yönündedir. Glukagon lipojenezi inhibe ederken ketojenezi artırır. Glikolizin inhibe edilmesi sonucu AsetilCoA yapımı inhibe edildiğinden yağ asitlerinin özellikle keton cisimlerine yıkımı artar. Yağ asitlerinin sitoplazmadan mitekondriye transferinde anahtar enzim olan karnitin açil transferaz–I enzim (CAT-I) yükesk glukagon ile maksimum düzeyde uyarılır. Bu durum mitokondriyal yağ asidi metabolizmasını hızlandırır.
Diabetes mellitus veya açlık gibi insülin yetmezliği durumlarında yağ asitlerinin fazla ve okzaloasetik asidin yetersizliğine bağlı olarak mitokondrialar AsetilCoA ile dolar ve ketoasidoz(ketonlara bağlı kan pH düşümesi) gelişir. İnsülin
İnsülin glukagonun tersi etkilere sahiptir. Dokulara glikoz girişini artırarak kan glikoz düzeylerini düşürür. Glikozun glikojene çevrilmesini (glikogenez) uyararak depo edilmesini sağlar. Polipeptid hormon olan insulin bütün türlerde birbirlerine disülfid köprüleri ile bağlı olan iki amino asit zincirinden kuruludur. Aktif insulin 21 amino asit kapsayan bir A zinciri ile 30 amino asit kapsayan bir B zinciri ihtiva eder. İnsulinin ön habercisi olan proinsulin aktif insulinde bulunmayan 33 amino asit daha kapsar. Bu birleştirici zincir “B” zincirinin COOH ucuna ve “A” zincirinin NH2 ucuna bağlıdır. Proinsulin endoplazmik retikulumun üzerinde sentezlendikten sonra golgi cihazında kıvrılarak disülfid köprüleri ile bağlanır. Disülfid bağlarının alkali veya indirgeyici bir madde ile kırılması insulini aktive eder. Çeşitli hayvan kaynaklarından gözlenen insulinin yapısı çeşitlidir.

462
ŞEKİL 8.50 İnsülin sentez basamakları
Ancak domuz pankreasının ki insan insulinine en çok benzeyenidir. İki insulin sadece B zincirinin terminalindeki amino asitlerin farklı oluşu ile ayrılır. İnsulin hedef dokusunun plazma membranı üzerinde özel reseptöre kuvvetle bağlanır. Membran reseptörü bir glikoproteindir. Böylece insulin fonksiyonunu hücreye girmeden yapar. İnsulinin hedef hücre membranına bağlanması c-AMP’da bir artış meydana getirmemesi bakımından epinefrin ve glukagondan farklıdır. İnsülin hücre içerisindeki etkisini reseptörünün tirozin kinaz aktivitesi ile hem kendisini, hem de başka proteinleri tiroziner üzerinden fosforillemesi ile gösterir. B-altbirimdeki tirozin kinaz, insülin bağlı olmadığı zaman a-altbirim tarafından inhibe edilir. Bu inhibüsyon insülinin bağlanması ile ortadan kalkar. İnsulin aslında c-AMP’de bir azalmaya sebep olmaktadır. İnsulinin yağ hücresi membranındaki reseptöre bağlandığı zaman c-AMP’de oluşan azalma ile birlikte siklik Guamozin monofosfat miktarında yükselmenin meydana geldiği tespit edilmiştir. Regülatör bu iki nükleotid (c-AMP, cGMP) karşılıklı zıt bir münasebet halinde çalışırlar. Bu münasebete “ying-yang” hipotezi denilir.

463
ŞEKİL 8.51 İnsülin etkisi
İnsulin, glikoz kullanılmasını, glikojen depolanmasının, yağ sentezinin, amino asit alımının ve protein sentezinin artmasına sebep olarak metabolizmada önemli rol oynayan ve karaciğer, kas, yağ dokuları üzerine etki eder. Glikoneogenezi inhibe eder. Karaciğerde albumin gibi pek çok proteinin sentezini artırır. Bazı hücrelerin büyümesi için gereklidir. Bütün insülin etkileri aynı insülin konsantrasyonlarında oluşmaz. Hatta bazen aynı hücerlerdeki değişik etkiler bile farklı insülin konsantrasyonları ile oluşmaktadır. Insulin, glikozun aktif transport ile hücre içine girmesini hızlandırır. İnsülin yokluğunda sadece beyin ve eritrositler glikozu enerji kaynağı olarak kullanmaktadır. Insülin varlığında ise glikoz hemen bütün hücreler için enerji kaynağıdır. Bunun için enerji kaynağına ATP’ye gerek duyulur. Glikoz taşıyıcı ile matriks içinde birleşiyor ve G-C kompleksini oluşturuyor. Membran içinde insulin bu kompleksin hareketini artırıyor. Hücre içine geçerken glikoz veriliyor. Taşıyıcı molekül membran matriksinde kalıyor. (Taşıyıcı molekül diğer glikozları taşımak için dönüyor). Hücre içindeki glikozun dışarı çıkmasıda yine aynı mekanizmayı izliyor. İnsulin yokluğunda taşıyıcı mekanizma zayıflıyor. İnsulin hücrelere glikoz taşınmasının ardından glikolizisi hızlandırarak, glikogenezisi hızlandırarak ve glukoneogenezisi yavaşlatarak etki eder. İnsulin piruvatkinaz, fosfofruktokinaz ve glukokinaz sentezini artırarak glikolizisi uyarır. Benzer şekilde, insulin glukoneogenezisi kontrol eden enzimleri bloke eder. Bunlar: Pruvat karboksilaz, fosfoenol piruvat karboksikinaz, fruktoz 1,6 difosfotazdır. İnsulin, glikolizisi uyarmakta, glukokinaz aktivitesini artırmakta ve böylece G-6-P miktarını artırmaktadır. G-6-P’da glikojen sentezi için gerekli glikojen sentetazı uyarır. İnsulinin de glikojen sentetaz üzerine uyarıcı etkisi vardır. Böylece glikojen sentezi de artar. Glikoz glikogen şeklinde depolanır. Adipoz dokuda insulin lipid sentezini uyarma işini, yağ asidi sentezi için gerekli Asetil-CoA ve NADPH yapımını artırarak sağlar. Adipoz dokuda insulin, glukagon ve epinefrin tarafından başlatılan yağ asitlerinin serbest bırakılışını azaltır. İnsulinin bu etkisi alfa-gliserofosfatın teşekkül ettiği glikolizisin yolu üzerindeki rolü ile ilgili olabilir. Böylece insulin etkisiyle yağ asitlerinin triacylgliserol şeklinde

464
depolanması kolaylaşır. İnsulin, glukagon ve epinefrin etkisiyle artan c-AMP artışını durdurarak lipolizisi azaltır. Böylece lipolizisden sorumlu olan lipazlar defosforile edilerek inaktif hale geçirilir. Yağ asitlerinin adipoz dokudan serbest bırakılmalarının insulin tarafından azaltılması çok önemlidir. Çünkü, çeşitli durumlarda fazla miktarlarda yağ asitlerinin açığa çıkması, glikolizisin çeşitli basamaklarda durdurulması ve glukoneogenezisin uyarılması ile glikoz metabolizmasının inhibe edilmesine yardım eder. İnsulin protein sentezini direkt olarak artırır. İnsulin ribozom seviyesinde etki ederek bu organellerin mRNA’dan haber tercümesi kapasitesini artırır. DNA sentezini hızlandırır ve dolayısıyla büyüme ve gelişmeyi etkiler. Glikoz, insulin sekresyonunu ve biyosentezini uyarır. Glikozdan başka, mannoz ve çok az olarak fruktoz insulin sentezini ve salgılanmasını uyarırlar. Insülin salgısı için hem cAMP ve hem de kalsiyum gereklidir. cAMP; glikoneogenezisi aktive eder ve artan glikoz, insulin salgılanması uyarılır. Glikogenalizisi artırır. Glukagon, sekretin, kolesistokinin, epinefrin, somatotropin gibi maddeler de (adenil siklaz) aktivitesini artırarak insulin sekresyonunu artırırlar. Hipoglisemik etkenler; Ağızdan alındığı zaman etkili olan birçok hipoglisemik ilaç vardır. (Tolbutamide). Bunlar şeker hastalığının hiperglisemisini kontrol için yararlıdır. Bu ilaçlar cAMP’nin kırılmasında rol alan difosfo esteraz enzimini inhibe ederek etki sağlarlar. Diabetes mellitus tedavisinde kullanılan sulfonilüreler ve biguanidler de pankreastan insülin salgısını artırırlar. Kafein, teofilin de aynı mekanizma ile insulin sekresyonunu uyarırlar. Norepinefrin gibi a-adrenerjik nörotransimitterler, a2-adrenerjik reseptör aracılığı ile salgıyı inhibe ederler. Langerhans adacıklarının delta hücrelerinden salgılanan somatostatin de insülin salgısının güçlü inhibütörüdür.
Diabetes Mellitus
Pankreas beta hücrelerince yeterli miktarda insülin üretilemediği zaman diabetes mellitus adı verilen hastalık ortaya çıkar. Hiperglisemi: kan glikozunda yükselme ile kendini gösterir. Fazla insülin hipoglisemiye yol açar. İnsülin ve glukagonun her ikisi de kan glikozu tarafından kontrol edilir.
Gonadlar: Cinsiyet bezleri
Gonadlar kadında overler erkekte ise testislerdir. Gonadlar üreme fonksiyonlarını kontrol eden hormonlar salgılarlar. Erkeklerde ana cinsiyet hormonu testesteron, kadınlarda ise östrojen, progesteron ve relaxindir. Testesteron, testislerin intersititiel hücrelerinden salgılanır. Erkekte FSH ve LH ile birlikte spermatogenezi (sperm üretimi) uyarır. Erkek cinsiyet organlarının gelişimi ve devamlılığı için gereklidir. Testesteron ikincil cinsiyet özelliklerinin gelişimini uyarır; Yüzde ve pubik bölgede kıllanma Larinkste genişleme Sesin kalınlaşması ve Kaslılığın artması yol açar.
1. Tip I diabet, insülin sekresyonu yokluğuna bağlıdır. İnsüline bağımlı diabet adını da alır. 2. Tip II diabet, hedef dokuların insülini metabolik etkilerine duyarlılıklarının azalmasına bağlı olarak gelişir ve insüline bağımlı olmayan diabet adını alır. Bu azalmış duyarlılığa insülin rezistansı da denir.
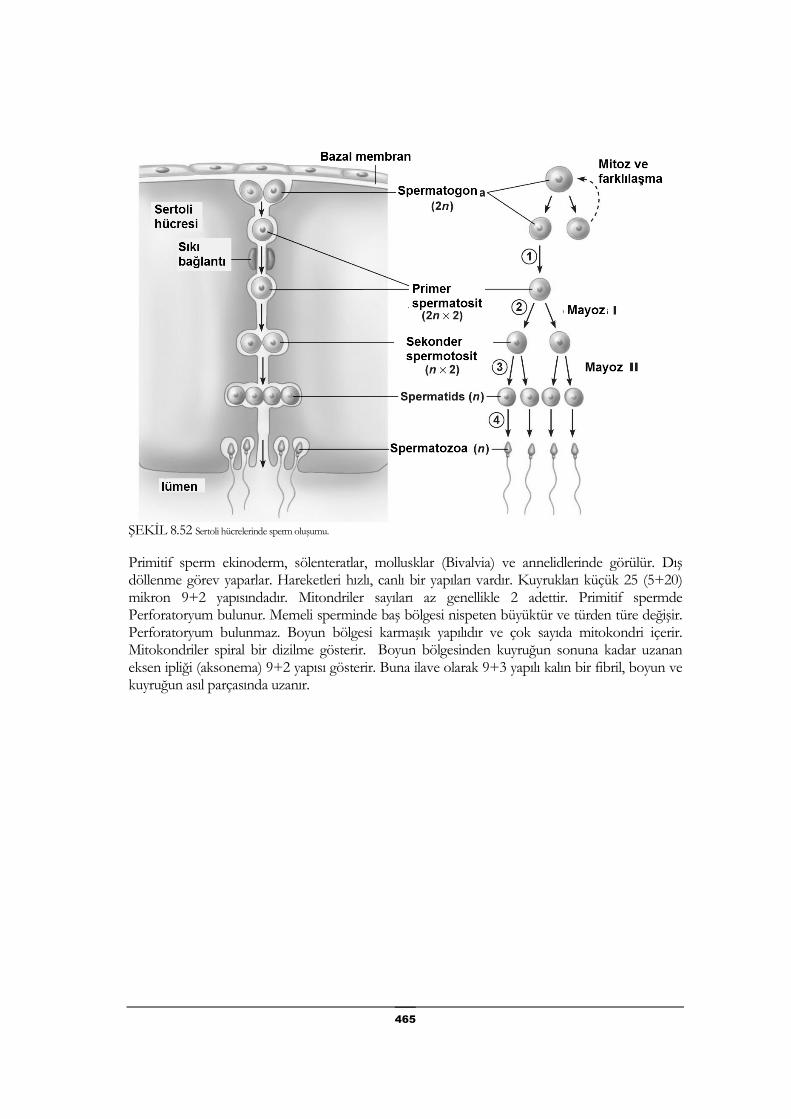
465
ŞEKİL 8.52 Sertoli hücrelerinde sperm oluşumu.
Primitif sperm ekinoderm, sölenteratlar, mollusklar (Bivalvia) ve annelidlerinde görülür. Dış döllenme görev yaparlar. Hareketleri hızlı, canlı bir yapıları vardır. Kuyrukları küçük 25 (5+20) mikron 9+2 yapısındadır. Mitondriler sayıları az genellikle 2 adettir. Primitif spermde Perforatoryum bulunur. Memeli sperminde baş bölgesi nispeten büyüktür ve türden türe değişir. Perforatoryum bulunmaz. Boyun bölgesi karmaşık yapılıdır ve çok sayıda mitokondri içerir. Mitokondriler spiral bir dizilme gösterir. Boyun bölgesinden kuyruğun sonuna kadar uzanan eksen ipliği (aksonema) 9+2 yapısı gösterir. Buna ilave olarak 9+3 yapılı kalın bir fibril, boyun ve kuyruğun asıl parçasında uzanır.

466
ŞEKİL 8.53 Testislerin yapısı

467
Kadınlarda overler salgılanan Östrojenler kadınlarda menstruel siklus, meme bezlerinin gelişimi ve ikincil cinsiyet özelliklerinin düzenlenmesine katkıda bulunur. Dişilerde hipofizden (adenohipofizden) salgılanan Progesteronda meme bezlerinin gelişimini ve menstruel siklusu düzenler, ayrıca gebelikte plasenta oluşumuna katkıda bulunur.
ŞEKİL 8.35 Sperm yapısı
ŞEKİL 8.54 Ovulasyon

468
Diğer Dokulardan salgılanan Hormonlar
�
ANP: Güçlü bir damar genişletici olan ANP , ve protein ( polipeptid ) yapısında olan kalp kası hücreleri tarafından bir hormondur. Su , sodyum , potasyum ve yağ ( yağ dokusu )homeostatik kontrol de görev alır . ANP sentez yeri kalp kulakçıklarıdır(Atriumlardır). ANP yüksek kan akımına karşı(basınçına) salgılanır.
ŞEKİL 8.55 ANP ve Diğer RAAS sistemi ile kan basınçıı ve hacminin ayarlanması
ANP böylece kan basıncının düşürülmesi, Dolaşım sisteminde su, sodyum ve yağ yüklerini azaltılmasına hizmet eder. ANP tam tersi işlevi böbrek üstü bezlerince salgılanan aldosteron tarafından sodyum tutulmasıyla yapılır.
Leptin: Yağ dokusu tarafından sentezlenen ve salgılanan leptin hormonu , enerji alımı ve enerji harcanması arasındaki dengeyi düzenleyerek bir tür anti obezite faktörü olarak fonksiyon görür. Bunun yanı sıra leptin hormonun üreme, sindirim sistemi fonksiyonların düzenlenmesi, sempatik sinir sistemi aktivasyonunun düzenlenmesi, kemik yoğunluğunun belirlenmesi,vücut ısısının düzenlenmesi ve beyin gelişimi gibi birçok fonksiyonda görevi olduğu bilinmektedir. Leptin hormonu kanda serbest ve proteine bağlı formda bulunmaktadır. Leptin’in aktivitesinden sorumlu olan kısım serbest formudur. Yapılan son çalışmalarda obez bireylerde serumdaki leptin’in büyük kısmının serbest formda olduğu tespit edilmiştir. Hem yağları yakan hem tokluk duygumuzu oluşturarak açlığımızı bastırma, doyduğumuzu beynimize iletme gibi çok önemli görevleri olan Leptin hormonu ancak 4-5 saatlik uzun öğün aralarında salgılanır . Leptin, 4-5 saat içinde bir şey yenmediği takdirde depo edilmiş olan yağları yakarak enerji ihtiyacımızı karşılamamızı sağlar. Uzun öğün aralarında yağ yakan Leptin Hormonu en çok gece saat 02 ile 05 arasında salgılanır. Akşam yemeklerimizi geç saatlerde yemezsek , gece yarısı leptin hormonumuz en yüksek düzeyde salgılanır.
• Kalpten� ANP
• Yağ dokusundan � Leptin
• Deriden � cholecalciferol (vit. D öncüsü)
• Plasentadan � Progesterone

469
Cholecalciferol : Cholecalciferol bir D3 vitaminidir. D vitamini vücuttaki kalsiyum işleyişi için kalsiyum emilmesi için önemlidir. Cholecalciferol D vitamini eksikliği tedavi etmek veya deri ya da kemiklerin D vitamin eksiklerine bağlı durumları neden olduğu birçok durumu önlemek için kullanılır. D vitamini ikili bir kökeni vardır: Gıda tarafından sağlanan öncül maddesi ve güneş ışınlarının (ultraviyole ışığı veya deri yoluyla vücut tarafından sentezlenen fonksiyonel D3 vitamin. D vitamini esas işlevi fosfor ve kalsiyumun bağırsakta emilmesini düzenlenmesidir ve karşılaştırın kapasitesini artırmak için:
1) Kemik, kıkırdak ve dişlerin dahil Kalsiyum minerallinin dokuların, optimal mineralizasyon sağlamak;
2) Kalsiyum ve fosfor plazma konsantrasyonlarını korumak.
ŞEKİL 8.57 Cholecalciferol molekülü.
Kaynaklar � Abdurrahman Aktümsek: Anatomi ve Fizyoloji insan Biyolojisi, 4. Baskı, Nobel Yayın
Dağıtım, 2009.
� Ali Yavuz Kılıç, Melih Zeytinoğlu: Hayvan Fizyolojisi, 11. Baskı, Anadolu Üniversitesi yayınları, 2009
� Andaç, S.O.: Hücre Fizyolojisi. Hacettepe Universitesi Yayınları. A 20. 1977.
� Barrett, K. E.; Barman, S. M.; Boitano, S.; Brooks, H. Ganong’s Review of Medical Physiology. McGraw-Hill Medical: 2012; Vol. 24th edition.
� Berne & Levy Physiology, Eds: Koeppen BN and Stanton BA; 6th Ed., 2010
� Boron, W. F.; Boulpaep, E. L. Medical Physiology. 2012; Vol. 2nd edition.
� Demir ve demir Metabolizması, Güncel Melike Sezgin Evim, Birol Baytan, Adalet Meral Güneş, Pediatri Dergisi ,102: 63-69.
� Guyton, A. C.; Hall., J. E. Tıbbi Fizyoloji. Nobel Tıp Kitabevleri: İstanbul, 2013; Vol. 12. baskı.

470
� http://bcs.whfreeman.com/thelifewire/content/chp00/00020.html
� http://faculty.ccri.edu/kamontgomery/physiology%20cell.htm
� http://faculty.dccc.edu/~swindig/DCCC-Unit-VI-Nervous-System.pdf
� http://highered.mcgraw-hill.com/sites/0072495855/student_view0/
� http://highered.mcgraw-hill.com/sites/dl/free/
� http://library.thinkquest.org/C0126536/main.php?currentchap=1¤tsect=neuron.htm
� http://people.eku.edu/ritchisong/301notes1.htm
� http://www.biochemj.org/csb/frame.htm
� http://www.lionden.com/cell_animations.htm
� http://www.medibul.com/files.php?p=pp&cid=31
� http://www.sinancanan.net/bilimsel/mss.htm
� Insulin Signaling and the Regulation of Glucose Transport, Mol Med. 2004 Jul-Dec; 10(7-12): 65–71. doi: 10.2119/2005-00029.Saltiel, PMCID: PMC1431367
� Koeppen, B. M.; Stanton, B. A. Berne & Levy Physiology. Mosby: 2009; Vol. 6th ed.
� Noyan, Ahmet. Fizyoloji: Ders kitabı. Anadolu Üniv., 1980.
� Preston, R. R.; Wilson, T. E. Physiology (Lippincott's Illustrated Reviews Series). Lippincott Williams & Wilkins: Philadelphia, PA, 2013; Vol. 1.
� Review of Medical Physiology, Ed. Ganong WF, 23rd Ed., 2011.
� Samson, W. E.; Scher, A. M. Mechanism of S-T segment alteration during acute myocardial injury. Circulation research 1960, 8, 780-7.
� Sefik DURSUN, hücre Membran Biyofiziği
� Vander İnsan Fizyolojisi, Eds: Widmaier EP, Raff H ve Strang KT; Çev Ed: Demirgören S, 2010.
� Vardar, S. A.; Kaymak, K.; Altun, A. Kalp kasının kasılmasında kalsiyum ve sarkoplazmik retikulumun rolü. T Klin Tıp Bilimleri 2002, 22, 630-634.
� Widmaier, E. P.; Raff, H.; Strang, K. Vander's Human Physiology: The Mechanisms of Body Function. McGraw-Hill Science/Engineering/Math: 2013; Vol. 13th ed.

471
� Principles of neural science ER Kandel, JH Schwartz, TM Jessell - 2000 - McGraw-Hill/Appleton & Lange
� http://pathmicro.med.sc.edu/mobile/m.immuno-17.htm
Indeks background, 26 border, 27 bullet, 26
color, 27 drawing, 27 drop cap, 26
footer, 28
frame, 27 graphic, 27 group, 27
header, 28 help, 28 link, 29
margins, 26
normal view, 26 number, 28 picture, 27, 29
print, 26 re-size, 27 section break, 26
shading, 26 style, 26, 26, 27 symbol, 26
Table of Contents, 28 template, 28, 29 ungroup, 27
Wingdings, 26
Copyright © 2022 FDOKUMEN




















