
Factors influencing axillary shoot proliferation and ...
10
Summary We developed procedures for in vitro cloning of Cedrus atlantica Manetti and C. libani A. Rich explants from juvenile and mature plants. Explant size was one determinant of the frequency of axillary bud break in both species. Shoot tips and nodal explants mainly developed calli, whereas bud sprouting occurred in defoliated microcuttings cultured on a modified Murashige and Skoog medium without growth regu- lators. Isolation and continuous subculture of sprouted buds on the same medium allowed cloning of microcuttings from C. at- lantica and C. libani seedlings and bicentennial C. libani trees, thus providing a desirable alternative for multiplying mature trees that have demonstrated superior characteristics. We also report adventitious bud differentiation from isolated embryos of C. atlantica. Neither auxin treatments nor other methods tested, including infection with Agrobacterium rhizogenes, were effective in inducing root initiation. Keywords: Cedrus atlantica, Cedrus libani, isolated embryos, microcuttings, tissue culture. Introduction Deforestation due to human activity, accompanied by deple- tion of forest tree genetic resources, is occurring in many parts of the world. Thus, there is an urgent need to conserve forest ecosystems for both environmental and economic reasons (Fen- ning and Gershenzon 2002). To maintain and sustain forest trees, conventional breeding and silvicultural approaches have been exploited for propagation and improvement, but tree- breeding efforts are restricted to the most valuable and fast- est-growing species. Such methods are limited because trees are generally slow-growing, long-lived, sexually self-incom- patible and highly heterozygous. Plant tissue culture offers an alternative approach for rapidly propagating and improving forest stock, as well as a means to overcome the impediments associated with conventional tree breeding (Altman 2003, Giri et al. 2004). However, other features inherent to the biology of forest trees—such as the recalcitrance of tissues from mature trees to in vitro manipulation—continue to challenge re- searchers developing tissue culture protocols for commercial tree production (Merkle and Dean 2000). Cedar forests are of great importance in the Mediterranean area. In Morocco, Cedrus atlantica Manetti is the main source of timber, fuel wood and other valuable products (Benchek- roun 1994). In Turkey and Lebanon, timber from Cedrus libani A. Rich has been used for centuries in construction, sculpture and crafts (Khuri et al. 2000, Talhouk et al. 2001). In addition, both species are cultivated worldwide as ornamental trees (Toth 1980, Hapla et al. 2000, Brunetti et al. 2001). Ce- dars have been found difficult to propagate asexually and have been propagated by grafting only under experimental condi- tions (Siniscalco 1994). However, tissue culture techniques may give rise to methods that allow efficient propagation of cedar species. A few reports on in vitro propagation of cedars are available (Bhatnagar et al. 1983, Abourouh and Najim 1990, Piola and Rohr 1996, Piola et al. 1998, 1999, Khuri et al. 2000) but none of them provide clear protocols yielding repro- ducible results that would provide a basis for commercial pro- duction. Our objective was to develop methods for the in vitro propagation of C. libani and C. atlantica from juvenile and adult trees. Materials and methods Juvenile explants Shoot apices (0.2–0.4 cm), nodal segments (with 2–3 axillary buds) and defoliated shoots (1–2 cm in length, hereafter re- ferred to as microcuttings) from C. libani and C. atlantica seedlings were used as initial explants. Seedlings were raised from seeds (from Chiltern Seeds, Ulverston, England, or Inter- semillas, Quart de Poblet, Spain) that had been soaked (1–3 min) in 1% NaClO and stratified for 1 month in darkness at 4 °C in plastic trays containing a sterile mixture of peat moss and perlite (1:1, v/v), moistened with half-strength Hoagland and Arnon (1950) nutrient solution with 0.1 g l –1 8-hydroxy- quinolin as fungicide. Subsequently, trays were transferred to a growth chamber at 25 ± 1 °C, with a 16-h photoperiod of 70 μmol m –2 s –1 and 70% relative humidity. After 2 to 3 months, stems (1–3 cm in length) were excised from the Tree Physiology 25, 477–486 © 2005 Heron Publishing—Victoria, Canada Factors influencing axillary shoot proliferation and adventitious budding in cedar BEGOÑA RENAU-MORATA, 1 JAVIER OLLERO, 1 ISABEL ARRILLAGA 1 and JUAN SEGURA 1,2 1 Departamento de Biología Vegetal, Facultad de Farmacia, Universidad de Valencia, Av. Vicent Andrés Estellés s/n, 46100-Burjasot (Valencia), Spain 2 Corresponding author ([email protected]) Received July 29, 2004; accepted October 2, 2004; published online February 1, 2005 Downloaded from https://academic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Factors influencing axillary shoot proliferation and ...

Summary We developed procedures for in vitro cloning ofCedrus atlantica Manetti and C. libani A. Rich explants fromjuvenile and mature plants. Explant size was one determinantof the frequency of axillary bud break in both species. Shoottips and nodal explants mainly developed calli, whereas budsprouting occurred in defoliated microcuttings cultured on amodified Murashige and Skoog medium without growth regu-lators. Isolation and continuous subculture of sprouted buds onthe same medium allowed cloning of microcuttings from C. at-lantica and C. libani seedlings and bicentennial C. libani trees,thus providing a desirable alternative for multiplying maturetrees that have demonstrated superior characteristics. We alsoreport adventitious bud differentiation from isolated embryosof C. atlantica. Neither auxin treatments nor other methodstested, including infection with Agrobacterium rhizogenes,were effective in inducing root initiation.
Keywords: Cedrus atlantica, Cedrus libani, isolated embryos,microcuttings, tissue culture.
Introduction
Deforestation due to human activity, accompanied by deple-tion of forest tree genetic resources, is occurring in many partsof the world. Thus, there is an urgent need to conserve forestecosystems for both environmental and economic reasons (Fen-ning and Gershenzon 2002). To maintain and sustain foresttrees, conventional breeding and silvicultural approaches havebeen exploited for propagation and improvement, but tree-breeding efforts are restricted to the most valuable and fast-est-growing species. Such methods are limited because treesare generally slow-growing, long-lived, sexually self-incom-patible and highly heterozygous. Plant tissue culture offers analternative approach for rapidly propagating and improvingforest stock, as well as a means to overcome the impedimentsassociated with conventional tree breeding (Altman 2003, Giriet al. 2004). However, other features inherent to the biology offorest trees—such as the recalcitrance of tissues from maturetrees to in vitro manipulation—continue to challenge re-searchers developing tissue culture protocols for commercial
tree production (Merkle and Dean 2000).Cedar forests are of great importance in the Mediterranean
area. In Morocco, Cedrus atlantica Manetti is the main sourceof timber, fuel wood and other valuable products (Benchek-roun 1994). In Turkey and Lebanon, timber from Cedruslibani A. Rich has been used for centuries in construction,sculpture and crafts (Khuri et al. 2000, Talhouk et al. 2001). Inaddition, both species are cultivated worldwide as ornamentaltrees (Toth 1980, Hapla et al. 2000, Brunetti et al. 2001). Ce-dars have been found difficult to propagate asexually and havebeen propagated by grafting only under experimental condi-tions (Siniscalco 1994). However, tissue culture techniquesmay give rise to methods that allow efficient propagation ofcedar species. A few reports on in vitro propagation of cedarsare available (Bhatnagar et al. 1983, Abourouh and Najim1990, Piola and Rohr 1996, Piola et al. 1998, 1999, Khuri et al.2000) but none of them provide clear protocols yielding repro-ducible results that would provide a basis for commercial pro-duction. Our objective was to develop methods for the in vitropropagation of C. libani and C. atlantica from juvenile andadult trees.
Materials and methods
Juvenile explants
Shoot apices (0.2–0.4 cm), nodal segments (with 2–3 axillarybuds) and defoliated shoots (1–2 cm in length, hereafter re-ferred to as microcuttings) from C. libani and C. atlanticaseedlings were used as initial explants. Seedlings were raisedfrom seeds (from Chiltern Seeds, Ulverston, England, or Inter-semillas, Quart de Poblet, Spain) that had been soaked(1–3 min) in 1% NaClO and stratified for 1 month in darknessat 4 °C in plastic trays containing a sterile mixture of peat mossand perlite (1:1, v/v), moistened with half-strength Hoaglandand Arnon (1950) nutrient solution with 0.1 g l–1 8-hydroxy-quinolin as fungicide. Subsequently, trays were transferred toa growth chamber at 25 ± 1 °C, with a 16-h photoperiod of70 µmol m–2 s–1 and 70% relative humidity. After 2 to3 months, stems (1–3 cm in length) were excised from the
Tree Physiology 25, 477–486© 2005 Heron Publishing—Victoria, Canada
Factors influencing axillary shoot proliferation and adventitiousbudding in cedar
BEGOÑA RENAU-MORATA,1 JAVIER OLLERO,1 ISABEL ARRILLAGA1 and JUANSEGURA1,2
1 Departamento de Biología Vegetal, Facultad de Farmacia, Universidad de Valencia, Av. Vicent Andrés Estelléss/n, 46100-Burjasot (Valencia),Spain
2 Corresponding author ([email protected])
Received July 29, 2004; accepted October 2, 2004; published online February 1, 2005
Dow
nloaded from https://academ
ic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022

seedlings and surface sterilized in 3% NaClO with 0.1%Tween-20 for 15 min or 0.1% HgCl2 for 3 min, followed byfour 5-min washes in sterile distilled water. Needles were re-moved with a razor blade at 1–2 mm above the needle–nodejunction, and these initial explants were cultured on differentnutrient media.
Freshly isolated C. atlantica embryos were used as primaryexplants in some experiments. Seeds were imbibed for 4 h insterile distilled water, surface-sterilized for 30 min in 70% eth-anol and rinsed in sterile distilled water; the mature embryoswere then dissected out of the seedcoat and female game-tophyte. Only undamaged embryos that were firm and white topale yellow in color were cultured.
Mature explants
Actively growing shoots (3–5 cm long) were collected fromseveral trees growing outdoors, including trees in a 20–30-year-old forest plantation of C. libani and C. atlantica locatedin Arbúcies, Girona, Spain; 15–20-year-old C. libani trees in anatural forest located in El Shouf, Lebanon; and 200-year-oldC. libani trees growing at the Reading University campus,Reading, U.K. All material was collected in the spring (May toearly June). After the leaves were removed, stems were im-mersed in 1% NaClO for 20 min, rinsed for 8 h in running tapwater, submerged for 15 min in an aqueous solution of 1%benomyl, and kept at 4–6 °C for 1–10 days. Shoots were thensurface sterilized with 0.1% HgCl2 as described for the juve-nile explants. Initial explants, meristematic domes plus one ortwo pairs of leaf primordia (0.6–1.0 mm), apical shoots (6–7 mm) and microcuttings were isolated under sterile condi-tions and cultured on different nutrient media. Unless other-wise stated, initial explants were chosen at random from dif-ferent donor plants.
Media and culture conditions
The basal media tested were: Murashige and Skoog (1962)medium, with macronutrients reduced to a half (MS/2) or aquarter (MS/4) of their original concentration; modified SHmedium (MSH), with Schenk and Hildebrandt (1972) macro-nutrients and MS micronutrients and vitamins; modified BFmedium (MBF), with Boulay and Franclet (1977) salts andMS vitamins; MSBN/2 medium, with half-strength MS saltsand half-strength Bourgin and Nitsch (1967) vitamins andSkoog (1944) amino acids; MSBN/4 (half-strength MSBN/2medium); and WPM medium (Lloyd and McCown 1980). Allmedia contained 3% sucrose except for MSBN/2 which con-tained 2% sucrose. Culture media were supplemented withgrowth regulators (indoleacetic acid (IAA), naphthaleneaceticacid (NAA), benzyladenine (BA), zeatin (Z) and thidiazuron(TDZ)) as specified. In some experiments, conifer-derived ac-tivated charcoal (Sigma, St. Louis, MO), calcium gluconate,triacontanol, paclobutrazol, pectimorf (a mix of oligopectinsfrom citric cell walls, kindly provided by Dr. Juan CarlosCabrera, INCA, Cuba), coumarin or phosphoric acid wereused. Several agar brands were tested: Difco-Bacto (0.7%;Sparks, MD), Sigma phytagel (0.2%), Sigma A-1296 (1%)and Pronadisa (0.8%; Torrejón de Ardoz, Madrid, Spain). The
pH was adjusted to 5.8 before autoclaving for 20 min at 120 °C(105 Pa). Except for TDZ and Z which were sterilized by filtra-tion, growth regulators and ancillary compounds were addedto the medium before autoclaving.
Culture vessels included 15 × 100-mm petri dishes and 150× 25-mm glass tubes covered with polypropylene closures(Bellco, Vineland, NJ), each containing 25 ml of agar-solidi-fied nutrient medium; Sigma jars with 100 ml of medium andglass jars with 50 ml of medium, both covered with sun-capclosures (Sigma); and 100-ml pots. In some experiments,explants were cultured, with continuous agitation (110 rpm),in 125-ml Erlenmeyer flasks (containing 25 ml of liquid me-dium) capped with Bellco stainless steel closures.
Unless otherwise specified, cultures were kept in growthchambers at 26 ± 2 °C and a 16-h photoperiod provided by flu-orescent lamps (70 µmol m–2 s–1 irradiance at culture level).
Axillary shoot proliferation from juvenile explants
Nutrient media, explant type, growth regulators and agarbrands Shoot apices and three types of nodal explants (upper(containing buds of the first verticil, immediately below theapex); central (second verticil); and basal (third verticil, imme-diately above the cotyledonary node)) were initially dipped for3 or 12 h in a sterile aqueous solution of 100 µM TDZ or BA,and then transferred individually to glass tubes containingagar-solidified MSH, MBF or MSBN/2 nutrient medium withor without BA (2.2 or 9 µM), 0.45 µM TDZ, 10–14 µM IAA orcombinations of the three types of growth regulators. Controlexplants were maintained for 3 or 12 h in sterile distilled water.The following brands of agar were tested: Difco-Bacto, Phyta-gel, Pronadisa and A-1296. Explant-derived organogenic calliwere isolated, transferred to their respective media withoutgrowth regulators and kept at 26 or 30 °C. At least 13 apical ornodal explants were cultured for each treatment. Total culturetime was 40 days.
Explant size, incubation temperature and cytokinin type Ina first experiment, shoot apices and microcuttings were cul-tured on medium with and without 9 µM BA and maintained at30 °C. In a second series of experiments, both entire and decap-itated microcuttings were cultured on medium without growthregulators or supplemented with 2.2 µM BA or 2.2 µM Z or0.45 µM TDZ. Cultures were kept at 26 or 30 °C. In all experi-ments, microcuttings were placed vertically in glass tubes con-taining MSBN/2 medium solidified with A-1296 agar. In addi-tion, microcuttings from C. libani proliferating cultures wereisolated and cultured on the same medium with or without2.2 µM Z and kept at 26 °C. Between 10 and 14 replicationswere prepared for each treatment. Cultures were examined forsurvival and sprouting percentages and mean number of axill-ary shoots per sprouted microcutting. Culture time was 40–60 days.
Axillary shoot proliferation from mature explants
In a first experiment, meristematic domes of C. atlantica andC. libani trees from Arbúcies, Spain, were cultured for 30 dayson Difco-Bacto agar-solidified MSH or MS/2 medium supple-
478 RENAU-MORATA, OLLERO, ARRILLAGA AND SEGURA
TREE PHYSIOLOGY VOLUME 25, 2005
Dow
nloaded from https://academ
ic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022

mented with BA (0, 2.2, 4.4 or 8.8 µM), alone or in combina-tion with NAA (0, 0.5, or 5.4 µM). Subsequently, explantswere transferred to their respective media without growth reg-ulators for another 30 days. Twenty explants were cultured pertreatment.
In a second experiment, shoot apices from C. libani treesgrowing in Arbúcies were cultured for 45 days in glass tubescontaining A-1296 agar-solidified MSBN/2 medium supple-mented with BA (0, 0.44, 2.22 and 4.44 µM), NAA (0 and0.05 µM), or combinations of both types of growth regulators.Subsequently, explants were transferred to their respectivemedia without growth regulators for another 45 days. Twenty-six explants were cultured per treatment.
In a third experiment, microcuttings from C. libani treesgrowing in El Shouf, Lebanon, were cultured in glass tubescontaining A-1296 agar-solidified MSBN/2 medium withoutgrowth regulators. To obtain enough material for further stud-ies, developing axillary shoots were routinely isolated fromthe elongated microcuttings and subcultured on the same me-dium. Shoots isolated from these proliferating cultures werefirst cultured for 60 days in glass tubes containing WPM me-dium or MSBN/2 medium supplemented with 0 or 0.45 µMTDZ or 2.22 µM Z. Subsequently, explants were transferred totheir respective media without growth regulators. At least 10microcuttings were cultured per treatment.
To test the effect of genotype, 50 to 140 microcuttings fromsix 200-year-old C. libani trees (genotypes AL1 to AL6),growing in Reading, U.K., were cultured on A-1296 agar-so-lidified MSBN/2 medium without growth regulators. Axillaryshoots that formed on these explants were subcultured on thesame medium to promote their elongation and the prolifera-tion of new buds. This procedure was repeated about every2 months for 2 years.
In all experiments, cultures were examined for survival andsprouting percentages and the mean number of axillary shootsper sprouted explant.
Adventitious bud differentiation from explants of C. atlantica
In a first experiment, excised needles from 3-month-old C. li-bani and C. atlantica seedlings were cultured (adaxial surfaceto the medium) in petri dishes containing MSH medium withor without 0.5 µM BA. Each treatment contained 20 replicates(20 dishes with five explants each) and culture time was60 days.
In a second experiment, freshly isolated C. atlantica em-bryos were cultured in petri dishes containing MSBN/2 me-dium supplemented with BA or Z (0, 2.2, 4.4, 6.6 or 9.0 µM).After 60 days, embryos were transferred to the same mediumwith or without 4.4 µM Z. Proliferating buds were isolated andtransferred to fresh medium without growth regulators. Em-bryos were also cultured for 60 days on MSBN/2 or WPM me-dium supplemented with Z (4.4, 9.0 or 18.0 µM), and thensubcultured as described previously. Each treatment containedfive replicates (five dishes with five embryos each).
In all experiments, media were solidified with A-1296 agarand cultures were examined for percent of explants with callusor buds and number of adventitious buds per explant.
Rooting
Both axillary and adventitious shoots (longer than 1 cm) iso-lated from C. atlantica and C. libani proliferating cultures,previously maintained for 1 month on MSBN/2 withoutgrowth regulators, were used for rooting experiments. Unlessotherwise stated, shoots with trimmed basal needles were cul-tured in glass tubes containing MBN/2 medium solidified withA-1296 agar and cultures were maintained in a 16-h photo-period.
In a first series of experiments, shoots were either culturedfor 7, 15 or 30 days on medium containing IBA or IAA (0, 0.3,2.5, 5, 10, 25 or 50 µM), or dipped in IBA (2.5 or 5 mM) in thepresence or absence of 1 mM NAA for 1 s and 5 min, respec-tively. Alternatively, the basal ends of the shoots were treatedwith rooting powders (2.5 mM IBA in talcum or RAIFORT SFfrom RIBA Quimiagra SL, Granollers (Barcelona), Spain, acommercial formulation containing 53.7 mM NAA and fungi-cides). Sterile support systems (filter paper bridges on liquidmedium, a mixture (1:1) of peat moss and perlite or vermicu-lite) were also tested. When appropriate, light was reducedfrom the rooting zone by covering the bases of vessels withaluminum foil. In all cases, shoots were transferred to the re-spective auxin-free support system for a 90-day culture period.
In a second series of experiments, the effects of the additionof several ancillary compounds to media containing IBA orNAA (0, 6 and 12 µM) were tested: (1) 0.25 mg l–1 paclo-butrazol, an inhibitor of gibberellin biosynthesis; (2) 3 or6 mM Ca2+ (as calcium gluconate); (3) 10 µM coumarin; (4) 2,5, 10 and 20 µg l–1 triacontanol; (5) 1, 3, 5, 7 and 10 mg l–1
pectimorf; (6) 15 and 60 g l–1 sucrose; (7) 150 or 300 mg l–1
H3PO4 and 500 mg l–1 glucose; and (8) 0.5 and 1% conifer-de-rived activated charcoal. Shoots were cultured for 30 days intheir respective media and subsequently transferred to basalmedium for 60 days. Cultures including coumarin were kept at26 and 20 °C.
In a last experiment, shoots were infected with Agrobac-terium rhizogenes LBA9402 strain. Basal shoot infection wasperformed either by dipping the basal end of the explants di-rectly into the bacterial suspension or after the shoot had beenwounded with a blade. In some cases, the bacterial suspensionwas injected into the basal cut surface of the shoots. After in-fection, explants were blot-dried between sterile filter paperand placed on medium with or without auxins (10 µM IAA orIBA), and incubated at 28 ± 1 °C in darkness for 24 h. Subse-quently, the explants were transferred to medium containing200 mg l–1 cefotaxime.
In all experiments, rooting percentage and the number ofroots per rooted shoot were recorded. Each treatment con-tained 10–12 replicates.
Statistical analysis
Significance of treatment effects was assessed by analysis ofvariance employing a completely random design. Percentagedata were subjected to arcsine transformation before analysis.Variation among treatment means was analyzed by Tukey’s(1953) procedure. All experiments were conducted at least
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
SHOOT PROLIFERATION IN CEDAR CULTURES 479
Dow
nloaded from https://academ
ic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022

twice. A hierarchic analysis of variance (nested ANOVA,Sokal and Rohlf 1995) design was also used to estimate vari-ance components for the morphogenic parameters recorded,partitioning the variation among seed lots and among seed-lings within lots. All analyses were performed with the Super-ANOVA program (Abacus Concepts, Berkeley, CA).
Results
Axillary shoot proliferation from juvenile explants
Nutrient medium, growth regulators and agar brands Fewerthan 5% of apical and nodal explants of C. atlantica and C. li-bani sprouted during 40 days of culture, although most ex-plants formed a callus. The frequency of sprouting was unaf-fected by pulse treatments with BA or TDZ , nutrient medium(MSH, MBF or MSBN/2), the presence of BA in the culturemedium or the agar brand (Difco-Bacto, Phytagel, Pronadisaor A-1296) (data not shown). Agar A-1296, which has beenused previously for C. libani cultures (Piola and Rohr 1996),was selected for subsequent experiments. Maximum callus for-mation (80–90%) was observed when apical explants werefirst dipped in BA or TDZ solution and then transferred toBA-supplemented medium. Although some calli underwentnecrosis, others differentiated adventitious buds and needles.These organogenic responses were mainly observed in apicalexplants from C. atlantica and C. libani growing in the pres-ence of 9 µM BA. Organogenic calli were subcultured on theirrespective basal media with or without activated charcoal at ei-ther 26 °C or at 30 °C (Piola and Rohr 1996), but the treatmentswere without significant effect on bud elongation.
Explant size, temperature and growth regulators Shoot api-ces and microcuttings, isolated from 3-month-old C. libani andC. atlantica seedlings, were cultured at 30 °C on MSBN/2 me-dium with or without 9 µM BA. Most of the shoot apices grownin the presence of BA produced a callus. Culture survival per-centages (85–100%) were not significantly affected by explantsize in either species; however, axillary bud proliferation oc-
curred only when microcuttings were used as primary explants(Figure 1). In both species, sprouting percentages and meannumber of axillary shoots per explant were significantly in-creased in microcuttings grown on medium without BA (80%without BA versus 60% with BA, P = 0.05; 3.0 versus 1.5, P =0.05).
In another experiment, entire or decapitated C. libani andC. atlantica microcuttings were cultured on MSBN/2 mediumwith or without BA or Z and kept for 45 days at 30 or 26 °C. InC. libani cultures, survival and sprouting percentages rangedfrom 85 to 100%, and these responses were not significantlyaffected by incubation temperature or the BA and Z treatments(P > 0.05) (data not shown). However, decapitation of C. li-bani microcuttings significantly reduced the mean number ofshoots formed per explant (3.0 versus 4.1, P = 0.05; Table 1).Basal MSBN/2 medium or Z was more effective than basalmedium + BA in promoting axillary shoot proliferation fromcultured C. libani microcuttings (4.0 or 4.5 versus 2.1 shootsper explant, respectively; P = 0.05, Table 1).
In C. atlantica cultures (Table 2), an incubation temperatureof 30 °C and explant decapitation both reduced microcuttingsurvival percentages (69.4 versus 87.8%, P = 0.05; 87.8 versus93.1%, P = 0.05; respectively). Incubation temperature, de-capitation and growth regulators all significantly influencedsprouting percentages, the best results being obtained at 26 °C(82.3 versus 66.7%, P = 0.05), with entire explants (84.9 ver-sus 64.1%, P = 0.05) and basal or Z-supplemented medium(83.8 or 77.1% versus 62.7%, P = 0.05). The detrimental effectof incubation at 30 °C versus at 26 °C was particularly evidentin decapitated microcuttings. The mean number of axillaryshoots formed per cultured explant was higher in entire micro-cuttings (2.6 versus 1.3, P = 0.05) and the presence of BA re-duced this response (1.2 versus 2.4 and 2.1 shoots per explantin Z-supplemented medium or basal medium, respectively;P = 0.05). Significant interactions between explant type andincubation temperature or presence of cytokinin in the me-dium were also evident, with the highest mean number ofshoots per explant obtained when entire microcuttings were
480 RENAU-MORATA, OLLERO, ARRILLAGA AND SEGURA
TREE PHYSIOLOGY VOLUME 25, 2005
Figure 1. Axillary shoot proliferationfrom C. atlantica (A) and C. libani (B)microcuttings cultured on MSBN/2without growth regulators.
Dow
nloaded from https://academ
ic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022
cultured at 30 °C in either basal medium or Z-supplementedmedium (Table 2).
Based on these results, entire microcuttings of C. atlanticaand C. libani were cultured at both 26 and 30 °C in the pres-ence of Z or TDZ (Table 3). In both species, survival percent-ages were lower when microcuttings grown on cytokinin- sup-plemented medium were kept at 30 °C. This negative effect ofthe higher temperature was especially evident in C. atlanticamicrocuttings cultured in the presence of TDZ (40%). Sprout-ing percentages ranged from 80 to 100% and neither incuba-tion temperature nor cytokinin type significantly influencedthis response (P > 0.05, data not shown). In contrast, meannumber of shoots formed per explant was higher at 30 °C thanat 26 °C (4.1 versus 2.9, P = 0.05), and Z was generally moreeffective than TDZ (4.1 versus 2.7, P = 0.05). Under the cul-ture conditions tested, shoot proliferation rates were higher inC. libani than in C. atlantica microcuttings (4.7 versus 2.4, P =
0.05). The maximum number of shoots formed per explant(7.0) was obtained when C. libani microcuttings were culturedon basal medium at 30 °C. This temperature effect was less ev-ident in microcuttings isolated from shoot-proliferating cul-tures. In all experiments, and irrespective of the treatments, thesprouted buds reached a final length of 1 to 2 cm.
These experiments were performed with seedlings from dif-ferent seed stocks obtained from two commercial companies.To determine the variance attributable to the seed lots and tothe individuals within each lot, the results were subjected to anested ANOVA, which indicated that there was no significanteffect of seed lot on survival or sprouting percentage. In con-trast, variations in shoot yield were due to differences amongseed lots (23 and 35% in C. atlantica and C. libani, respec-tively) and among seedlings within seed lots (77 and 65% forC. atlantica and C. libani, respectively).
Axillary shoot proliferation from explants of mature origin
None of the culture conditions tested promoted axillary shootproliferation from apical shoot meristems of mature trees ofC. atlantica or C. libani. Most of the responding explants(60–70%) formed calli. Neither the culture medium (SH orMS/2) nor growth regulators (BA or NAA, or both) affectedthis morphogenic response. Meristem-derived calli were main-tained in culture for almost 6 months, but attempts to induceorganogenesis failed.
Based on the data obtained with seedlings, shoot apices(0.6 cm) from 30-year-old C. libani were surface sterilizedwith either NaClO or HgCl2 and cultured on MSBN/2 mediumwith or without BA or NAA or both. Although the effective-ness of the two sterilizing agents was similar (an 80% yield ofaxenic explants), NaClO caused generalized chlorosis in someexplants. Survival percentage was 50% and neither the steriliz-ing agent nor the presence of growth regulators significantlyaffected this result. Explant elongation and subsequent ax-illary bud sprouting was observed only in cultures establishedon basal medium. Under these conditions, 15% of the explantselongated and showed bud sprouting (1–3 buds per explant)within 2–3 months of culture. Explants grown in the presenceof BA with or without NAA formed calli.
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
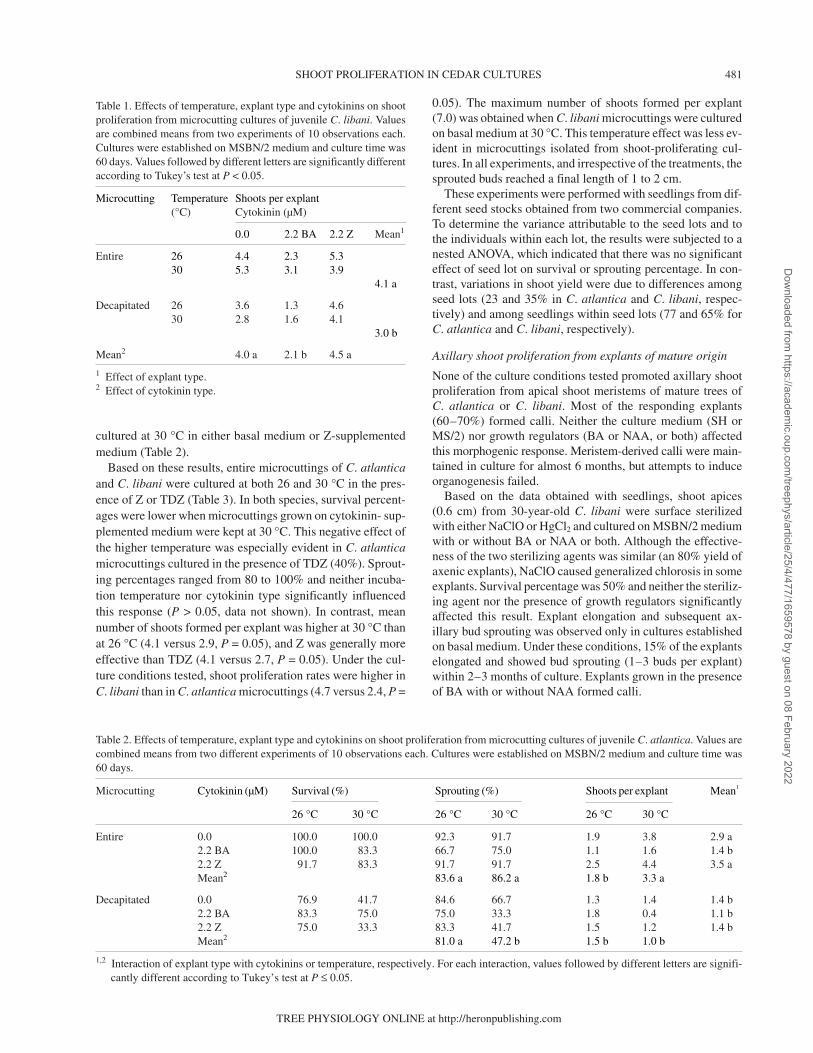
SHOOT PROLIFERATION IN CEDAR CULTURES 481
Table 2. Effects of temperature, explant type and cytokinins on shoot proliferation from microcutting cultures of juvenile C. atlantica. Values arecombined means from two different experiments of 10 observations each. Cultures were established on MSBN/2 medium and culture time was60 days.
Microcutting Cytokinin (µM) Survival (%) Sprouting (%) Shoots per explant Mean1
26 °C 30 °C 26 °C 30 °C 26 °C 30 °C
Entire 0.0 100.0 100.0 92.3 91.7 1.9 3.8 2.9 a2.2 BA 100.0 83.3 66.7 75.0 1.1 1.6 1.4 b2.2 Z 91.7 83.3 91.7 91.7 2.5 4.4 3.5 aMean2 83.6 a 86.2 a 1.8 b 3.3 a
Decapitated 0.0 76.9 41.7 84.6 66.7 1.3 1.4 1.4 b2.2 BA 83.3 75.0 75.0 33.3 1.8 0.4 1.1 b2.2 Z 75.0 33.3 83.3 41.7 1.5 1.2 1.4 bMean2 81.0 a 47.2 b 1.5 b 1.0 b
1,2 Interaction of explant type with cytokinins or temperature, respectively. For each interaction, values followed by different letters are signifi-cantly different according to Tukey’s test at P ≤ 0.05.
Table 1. Effects of temperature, explant type and cytokinins on shootproliferation from microcutting cultures of juvenile C. libani. Valuesare combined means from two experiments of 10 observations each.Cultures were established on MSBN/2 medium and culture time was60 days. Values followed by different letters are significantly differentaccording to Tukey’s test at P < 0.05.
Microcutting Temperature Shoots per explant(°C) Cytokinin (µM)
0.0 2.2 BA 2.2 Z Mean1
Entire 26 4.4 2.3 5.330 5.3 3.1 3.9
4.1 a
Decapitated 26 3.6 1.3 4.630 2.8 1.6 4.1
3.0 b
Mean2 4.0 a 2.1 b 4.5 a
1 Effect of explant type.2 Effect of cytokinin type.
Dow
nloaded from https://academ
ic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022
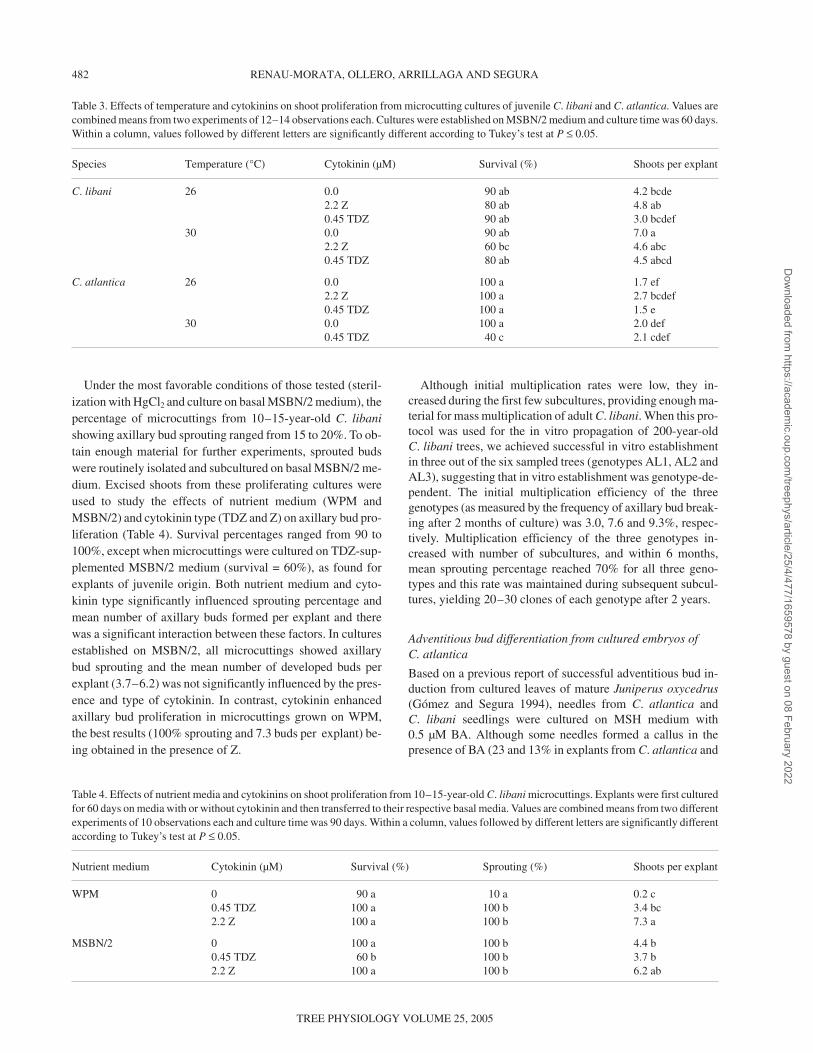
Under the most favorable conditions of those tested (steril-ization with HgCl2 and culture on basal MSBN/2 medium), thepercentage of microcuttings from 10–15-year-old C. libanishowing axillary bud sprouting ranged from 15 to 20%. To ob-tain enough material for further experiments, sprouted budswere routinely isolated and subcultured on basal MSBN/2 me-dium. Excised shoots from these proliferating cultures wereused to study the effects of nutrient medium (WPM andMSBN/2) and cytokinin type (TDZ and Z) on axillary bud pro-liferation (Table 4). Survival percentages ranged from 90 to100%, except when microcuttings were cultured on TDZ-sup-plemented MSBN/2 medium (survival = 60%), as found forexplants of juvenile origin. Both nutrient medium and cyto-kinin type significantly influenced sprouting percentage andmean number of axillary buds formed per explant and therewas a significant interaction between these factors. In culturesestablished on MSBN/2, all microcuttings showed axillarybud sprouting and the mean number of developed buds perexplant (3.7–6.2) was not significantly influenced by the pres-ence and type of cytokinin. In contrast, cytokinin enhancedaxillary bud proliferation in microcuttings grown on WPM,the best results (100% sprouting and 7.3 buds per explant) be-ing obtained in the presence of Z.
Although initial multiplication rates were low, they in-creased during the first few subcultures, providing enough ma-terial for mass multiplication of adult C. libani. When this pro-tocol was used for the in vitro propagation of 200-year-oldC. libani trees, we achieved successful in vitro establishmentin three out of the six sampled trees (genotypes AL1, AL2 andAL3), suggesting that in vitro establishment was genotype-de-pendent. The initial multiplication efficiency of the threegenotypes (as measured by the frequency of axillary bud break-ing after 2 months of culture) was 3.0, 7.6 and 9.3%, respec-tively. Multiplication efficiency of the three genotypes in-creased with number of subcultures, and within 6 months,mean sprouting percentage reached 70% for all three geno-types and this rate was maintained during subsequent subcul-tures, yielding 20–30 clones of each genotype after 2 years.
Adventitious bud differentiation from cultured embryos ofC. atlantica
Based on a previous report of successful adventitious bud in-duction from cultured leaves of mature Juniperus oxycedrus(Gómez and Segura 1994), needles from C. atlantica andC. libani seedlings were cultured on MSH medium with0.5 µM BA. Although some needles formed a callus in thepresence of BA (23 and 13% in explants from C. atlantica and
482 RENAU-MORATA, OLLERO, ARRILLAGA AND SEGURA
TREE PHYSIOLOGY VOLUME 25, 2005
Table 3. Effects of temperature and cytokinins on shoot proliferation from microcutting cultures of juvenile C. libani and C. atlantica. Values arecombined means from two experiments of 12–14 observations each. Cultures were established on MSBN/2 medium and culture time was 60 days.Within a column, values followed by different letters are significantly different according to Tukey’s test at P ≤ 0.05.
Species Temperature (°C) Cytokinin (µM) Survival (%) Shoots per explant
C. libani 26 0.0 90 ab 4.2 bcde2.2 Z 80 ab 4.8 ab0.45 TDZ 90 ab 3.0 bcdef
30 0.0 90 ab 7.0 a2.2 Z 60 bc 4.6 abc0.45 TDZ 80 ab 4.5 abcd
C. atlantica 26 0.0 100 a 1.7 ef2.2 Z 100 a 2.7 bcdef0.45 TDZ 100 a 1.5 e
30 0.0 100 a 2.0 def0.45 TDZ 40 c 2.1 cdef
Table 4. Effects of nutrient media and cytokinins on shoot proliferation from 10–15-year-old C. libani microcuttings. Explants were first culturedfor 60 days on media with or without cytokinin and then transferred to their respective basal media. Values are combined means from two differentexperiments of 10 observations each and culture time was 90 days. Within a column, values followed by different letters are significantly differentaccording to Tukey’s test at P ≤ 0.05.
Nutrient medium Cytokinin (µM) Survival (%) Sprouting (%) Shoots per explant
WPM 0 90 a 10 a 0.2 c0.45 TDZ 100 a 100 b 3.4 bc2.2 Z 100 a 100 b 7.3 a
MSBN/2 0 100 a 100 b 4.4 b0.45 TDZ 60 b 100 b 3.7 b2.2 Z 100 a 100 b 6.2 ab
Dow
nloaded from https://academ
ic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022

C. libani, respectively), none of the cultured explants differen-tiated adventitious buds.
In subsequent experiments, mature C. atlantica embryoswere cultured on MSBN/2 medium with or without Z and BA.Within the first week of culture, embryos elongated and thecotyledons and hypocotyls became green. Embryos on basalmedium produced no callus or adventitious buds, developingas normal seedlings. In the presence of cytokinins, hypocotylsand cotyledons in contact with the medium proliferated quick-ly, producing callus. Usually the radicles did not show a re-sponse either in color or in cell proliferation. The first adventi-tious buds were directly induced from the upper surface of thehypocotyls (Figure 2A) after 20–30 days of culture. Furtherbud differentiation occurred on the surface of the previouslyinduced calli. Indirect needle primordia differentiation wasalso observed, especially when embryos were cultured in thepresence of BA. After 50–60 days, the entire upper surface ofthe responding embryos was covered with adventitious budsand needles, although the latter did not develop into shoots.
Table 5 summarizes the bud differentiation process in cul-tured C. atlantica embryos. The bud-forming capacity of theexplants depended on cytokinin type and concentration, thebest results being obtained when the embryos were cultured inthe presence of 9 µM Z (47% of caulogenic explants and4.2 adventitious buds per embryo), whereas 9 µM BA induceda smaller response (13% and 0.3, respectively). Embryo sub-culture to medium with 4.4 mM Z enhanced the developmentof adventitious buds (Figure 2B). Further shoot elongation wasachieved following excision and transfer of these shoots tohormone-free medium. Transfer of embryos bearing adventi-tious buds to basal medium did not promote bud elongation(data not shown).
Although WPM provided a greater percentage of embryoswith buds than MSBN/2 medium (49.0 versus 40.0%, P =0.05), the mean number of buds formed per embryo in thepresence of 9 µM Z was similar in the two media (3.1). Irre-spective of the nutrient medium, 18 µM Z enhanced acicularprimordia differentiation and reduced adventitious bud differ-entiation.
Rooting
Rooting experiments were carried out with axillary and adven-titious shoots isolated from proliferating cedar cultures of ju-
venile and adult origin. None of the experimental variablestested promoted rooting.
Discussion
We observed in vitro axillary bud proliferation of explantsfrom juvenile and adult C. atlantica and C. libani trees. In bothspecies, explant size was an important factor affecting axillarybud proliferation in in vitro culture. Piola and Rohr (1996) re-ported that axillary and apical buds from in-vitro-grown mi-crocuttings of C. libani sprout at 30 °C but not at 24 °C. Subse-quently, Piola et al. (1998) demonstrated that needle removal,but not microcutting decapitation, substituted for the highertemperature requirement to break bud dormancy. Abscisicacid (ABA) concentrations in microcuttings with dormantbuds were higher than in microcuttings bearing sprouted buds,leading these authors to suggest that ABA accumulation inneedles caused bud dormancy of C. libani microcuttings at24 °C. Therefore, we always used defoliated microcuttings, to
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
SHOOT PROLIFERATION IN CEDAR CULTURES 483
Figure 2. Induction and developmentof adventitious buds from cultured em-bryos of C. atlantica. Adventitious buddifferentiation on MSBN/2 mediumwith 9 µM Z (left) and adventitiousbud development on MSBN/2 mediumwith 4.4 µM Z (right). Bar = 0.3 cm.
Table 5. Effects of cytokinin type and concentration on the differentia-tion of adventitious buds from embryos of C. atlantica. Values arecombined means of results from two experiments of 20 observationseach. Cultures were established on MSBN/2 medium and culture timewas 60 days. Within a column, values followed by different letters aresignificantly different according to Tukey’s test at P ≤ 0.05.
Cytokinin Concentration Explants with Buds per(µM) buds (%) explant
Z 0.0 0.0 0.02.2 13.3 0.84.4 33.3 1.36.6 33.3 0.89.0 46.7 4.2Mean 25.3 a 1.4 a
BA 0.0 0.0 0.02.2 6.7 0.14.4 6.7 0.16.6 13.3 0.19.0 13.3 0.3Mean 8.0 b 0.1 b
Dow
nloaded from https://academ
ic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022

exclude a possible ABA influence in our experiments. The useof defoliated microcuttings makes it difficult to compare ourresults with those reported by Piola and Rohr (1996) for juve-nile C. libani microcuttings; nevertheless, we found that an in-cubation temperature of 30 °C enhanced shoot yield in juve-nile microcuttings of both C. atlantica and C. libani. Piola andRohr (1996) found that a 3-h pulse with 0.1 mM BA inducedthe highest proliferation rate (4.5 buds per explant) in C. libanimicrocuttings. We observed similar results for C. libani micro-cuttings grown in the presence of 2.2 µM BA; however, BAnegatively affected bud sprouting in C. atlantica microcut-tings.
Nested ANOVA attributed most of the variability in shootyield from cultured microcuttings of juvenile origin to singleseedlings (77 and 65% for C. atlantica and C. libani, respec-tively). This strong genotypic effect was even more evident incultured explants of bicentennial C. libani trees, where onlythree out of the six genotypes were successfully established invitro. A similar genotypic effect on the bud-forming capacityof explants was observed by Tang et al. (2001). In some coni-fers, variation in morphogenic responses was evident not onlyamong embryos from different seed lots but also among em-bryos obtained from controlled crossings (von Arnold andEriksson 1982, 1986, Shen and von Arnold 1982). We ob-served differences in the initial in vitro performance of thethree genotypes, but these differences decreased with increas-ing number of subcultures.
Usually, in vitro manipulation of woody plants is more diffi-cult with mature explants than with juvenile explants, andCedrus was no exception. A comparison between cedar ex-plants of juvenile origin and those of adult origin showed thatmicrocuttings were the preferred explant for axillary shootproliferation in juvenile and adult cultures. Although this re-sponse did not require the presence of cytokinin in the culturemedium, Z increased axillary shoot yield in explants of matureorigin. Hormone-free nutrient media have been successfullyemployed for culture establishment of other mature Pinaceaeincluding Pseudotsuga menziesii (Mirb.) Franco and Pinuslambertiana Dougl. (Gupta and Durzan 1985), although thepresence of growth regulators favored axillary bud sproutingin shoot apices and microcuttings isolated from 20-year-oldLarix occidentalis Nutt. trees (Chesick et al. 1990).
Shoot proliferation rates in cultures of adult C. libani weregenerally greater on basal MSBN/2 medium than on basalWPM. Although the influence of nutrient medium on in vitromorphogenesis is well documented, its effect remains one ofthe most empirical aspects of plant cell culture (Preece 1995).Compared with basal MSBN/2 medium, basal WPM hashigher concentrations of calcium, sucrose, micronutrients andorganic supplements, and contains biotine and folic acid (bothabsent in MSBN/2 medium). A reduced amount of nutrientsstimulates axillary branching in some plants (Karhu 1997) andlow calcium availability can affect apical meristem integrity(McCown and Sellmer 1987), thus favoring axillary bud de-velopment; however, we are unable to offer an explanation forthe differential effect of MSBN/2 and WPM media on thebasis of our experimental data.
Adventitious organogenesis offers higher potential forshoot production than axillary bud proliferation, and is the pre-ferred method for coniferous micropropagation because conif-erous buds are generally induced directly on the explant(Thorpe et al. 1991). With a few exceptions, however, tech-niques for adventitious budding of coniferous explants ob-tained from trees in the adult growth phase are still limited, be-cause the caulogenic potential of isolated organs is affected bythe ontogenic age of the tissues (von Aderkas and Bonga 2000,Giri et al. 2004). In our experiments, callus derived from shootapices differentiated adventitious buds in the presence of cyto-kinin, but only when explants of juvenile origin were used andwe were unable to promote their elongation. In contrast,Hosseyni et al. (1999) reported the induction of adventitiousorganogenesis (buds and needles) from winter buds of 10–15-year-old C. libani cultured on WPM with BA or kinetin. Thisdifference between studies may be associated with tree age,because our trees were older.
We successfully generated adventitious shoots from C. at-lantica embryos, which opens up the possibility of mass prop-agation of this species. Our regeneration system included: (1)adventitious budding in the presence of 9.0 µM Z; (2) bud de-velopment on medium containing 4.4 µM Z; and (3) shootelongation on cytokinin-free medium. Zeatine or benzyla-denine alone was enough to stimulate both direct and indirectadventitious budding, which is in agreement with previousfindings in other conifer species (Thorpe et al. 1991); however,the presence of Z improved the bud-forming capacity of theexplants, and the process of bud development required transferof C. atlantica embryos to a medium with cytokinin. In con-trast, in other conifers, bud development depends on transferof explants to hormone-free medium (Gómez and Segura1994, Mata et al. 2001, Villalobos-Amador et al. 2002, Sches-tibratov et al. 2003).
Successful micropropagation of many woody species is fre-quently limited by their reluctance to form adventitious roots,and C. libani and C. atlantica were no exception. In many spe-cies, rooting can be achieved by adding auxins to the culturemedium or by using different support substrates (Hartmann etal. 1997). Activated charcoal, a high sucrose concentration,variations in environmental temperature conditions and lightreduction in the rooting zone improve auxin-induced rootingin many species, including conifers (George 1993). A varietyof ancillary compounds such as paclobutrazol, phosphoricacid, calcium, triacontanol, oligosaccharins and coumarin(George 1993, Haissig and Davis 1994, Tantos et al. 2001,Nandi et al. 2002) can improve rooting capacity in recalcitrantspecies. In some conifer species, infection with A. rhizogenesenhances rhizogenesis (McAfee et al. 1993, Tzfira et al. 1996,Mihaljevic et al. 1998, Villalobos-Amador et al. 2002). In ourexperiments, all of these variables failed to induce rooting inshoots of axillary or adventitious origin of C. libani and C. at-lantica. Nicholson (1984) and Nandi et al. (2002) were able toinduce rooting in C. deodara (D. Don) G. Don and rooting hasbeen reported in C. libani and C. atlantica (Dirr and Heuser1987) cuttings, although in the last two species, rooting per-centages were minimal and rooting was observed only when
484 RENAU-MORATA, OLLERO, ARRILLAGA AND SEGURA
TREE PHYSIOLOGY VOLUME 25, 2005
Dow
nloaded from https://academ
ic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022

cuttings were sampled at the end of the winter season. Hos-seyni et al. (1999) also reported rooting (29–33%) from ad-ventitious shoots of C. libani cultured on MS/4 or WPM/2media with 1.4 µM IAA but not with 1.4 µM IBA; both hor-mones, however, were ineffective in promoting rooting inshoots generated from winter buds of 10–15-year-old C. li-bani trees.
In conclusion, we have developed an efficient method topromote axillary bud proliferation from cedar microcuttingsof juvenile and adult origin. Our regeneration system (budsprouting in defoliated microcuttings cultured on basal MSBN/2medium at 26 or 30 °C, and isolation and continuous subcul-ture of sprouted buds onto the same medium) provides a meansof obtaining multiple shoots from a single microcutting fromcedar trees old enough to have demonstrated their superiorcharacteristics. Our regeneration method may be useful forclonal propagation of elite genotypes provided that a protocolfor the successful rooting of proliferating shoots is developed.The phenotypic characteristics of the putative clones weresimilar to those of seedlings grown from seeds. We also devel-oped a protocol for the successful regeneration of adventitiousshoots from mature embryos of C. atlantica.
Acknowledgments
The authors thank the European Union, Contract Number ERBIC18-CT97-0177, and Generalitat Valenciana (Grupos 03/102) for financialsupport.
References
Abourouh, M. and L. Najim. 1990. Culture in vitro de fragments decotyledons des plantules de Cedrus atlantica Manetti. Saussurea21:75–80.
Altman, A. 2003. From plant tissue culture to biotechnology: scien-tific revolutions, abiotic stress tolerance and forestry. In Vitro Cell.Dev. Biol. Plant 39:75–84.
Benchekroun, F. 1994. The economy of the Moroccan cedar forestand its impact on the development of local collectivities. Ann.Rech. Fort. Maroc 27:713–724.
Bhatnagar, S.P., M.N. Singh and N. Kapur. 1983. Preliminary investi-gations on organ differentiation in tissue cultures of Cedrus deo-dara and Pinus roxburghii. Indian J. Exp. Biol. 21:524–526.
Boulay, M. and A. Franclet. 1977. Recherches sur la propagationvégétative du Douglas Pseudotsuga menziesii (Mirb.) Franco. Po-ssibilités d’obtention de plantes viables à partir de la culture in vitrode bourgeons de pieds-mères juvéniles. CR Acad. Sci. 284D:1405–1407.
Bourgin, J.P. and J.P. Nitsch. 1967. Obtention de Nicotiana haploidesà partir d’étamines cultivées in vitro. Ann. Physiol. Vég. 9:337–338.
Brunetti, M., E.L. De Capua, N. Macchioni and S. Monachello. 2001.Natural durability, physical and mechanical properties of Atlas ce-dar (Cedrus atlantica Manetti) wood from Southern Italy. Ann.For. Sci. 58:607–613.
Chesick, E.E., D.E. Bilderback and G.M. Blake. 1990. In vitro multi-ple bud formation by 20-year-old western larch buds and stems.HortScience 25:114–116.
Dirr, M.A. and C.W. Heuser, Jr. 1987. The reference manual of woodyplant propagation: from seed to tissue culture. Varsity Press, Ath-ens, GA, 239 p.
Fenning, T.M. and J. Gershenzon. 2002. Where will the wood comefrom? Plantation forests and the role of biotechnology. TrendsBiotechnol. 20:291–296.
George, E.F. 1993. Plant propagation by tissue culture. 2nd Edn.Exegetics, Edington, U.K., 1361 p.
Giri, C.C., B. Shyamkumar and C. Anjaneyulu. 2004. Progress in tis-sue culture, genetic transformation and applications of biotechnol-ogy to trees: an overview. Trees 18:115–135.
Gómez, M.P. and J. Segura. 1994. Factors controlling adventitiousbud induction and plant regeneration in mature Juniperus oxy-cedrus leaves cultured in vitro. In Vitro Cell. Dev. Biol. Plant 30:210–218.
Gupta, P.K. and D.J. Durzan. 1985. Shoot multiplication from maturetrees of Douglas-fir (Pseudotsuga menziesii) and sugar pine (Pinuslambertiana). Plant Cell Rep. 4:177–179.
Haissig, B.E. and T.D. Davis. 1994. A historical evaluation of adven-titious rooting research to 1993. In Biology of Adventitious RootFormation. Eds. T.D. Davis and B.E. Haissig. Plenum Press, NewYork, pp 275–331.
Hapla, F., J.V. Oliver-Villanueva and J.M. González-Molina. 2000.Effect of silvicultural management on wood quality and timber uti-lisation of Cedrus atlantica in the European mediterranean area.Holz als Roh-Werkstoff 58:1–8.
Hartmann, H.T., D.E. Kester, F.T. Davies and R.L. Geneve. 1997.Plant propagation: principles and practices. Prentice Hall, London,770 p.
Hoagland, D.R. and D.I. Arnon. 1950. The water culture method forgrowing plants without soil. Circular 347, California AgriculturalExperiment Station, Berkeley, CA, 32 p.
Hosseyni, N., A. Ozan and Z. Kaya. 1999. Basal nutrient and hor-monal requirements for direct organogenesis of Cedrus libaniA. Rich. In Plant Biotechnology and In Vitro Biology in the 21stCentury. Eds. A. Altman, M. Ziv and S. Izhar. IX InternationalCongress on Plant Tissue and Cell Culture, Jerusalem. Kluwer Ac-ademic Publishers, Dordrecht, pp 53–56.
Karhu, S.T. 1997. Axillary shoot proliferation of blue honeysuckle.Plant Cell Tissue Organ Cult. 48:195–201.
Khuri, S., M.R. Shmoury, R. Baalbaki, M. Maunder and S.N. Tal-houk. 2000. Conservation of the Cedrus libani populations in Leb-anon: history, current status and experimental application ofsomatic embryogenesis. Biodivers. Conserv. 9:1261–1273.
Lloyd, G.B. and B.H. McCown. 1980. Comercially-feasible micro-propagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Proc. Int. Plant Propagation Soc. 30: 421–421.
Mata, M., V.M. Chávez and R. Bye. 2001. In vitro regeneration ofplantlets from immature zygotic embryos of Picea chihuahuanaMartínez, an endemic Mexican endangered species. In Vitro Cell.Dev. Plants 37:73–78.
McAfee, B.J., E.E. White, L.E. Pelcher and M.S. Lapp. 1993. Rootinduction in pine (Pinus) and larch (Larix) spp. using Agrobac-terium rhizogenes. Plant Cell Tissue Organ Cult. 34:53–62.
McCown, B.H. and J.C. Sellmer. 1987. General media and vesselssuitable for woody plant culture. In Cell and Tissue Culture in For-estry. Eds. J.M. Bonga and J. Durzan. Vol. 1. Marinus Nijhoff Pub-lishers, Dordrecht, pp 4–16.
Merkle, S.A.and J.F.D. Dean. 2000. Forest tree biotechnology. Curr.Opin. Biotechnol. 11:298–302.
Mihaljevic, S., V. Katavic and S. Jeleska. 1998. Root formation inmicropropagated shoots of Sequoia sempervirens using Agrobac-terium. Plant Sci. 141:73–80.
Murashige, T. and F. Skoog. 1962. A revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiol. Plant. 15:473–497.
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
SHOOT PROLIFERATION IN CEDAR CULTURES 485
Dow
nloaded from https://academ
ic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022

Nandi, S.K, S. Tamta and L.M.S. Palni. 2002. Adventitious root for-mation in young shoots of Cedrus deodora. Biol. Plant. 45:473–476.
Nicholson, R. 1984. Propagation notes on Cedrus deodara Shalimarand Calocedrus decurrens. Plant Propagator 30:5–6.
Piola, F. and R. Rohr. 1996. A method to overcome seed and axillarybud dormancy to improve Cedrus libani micropropagation. PlantTissue Cult. Biotechnol. 2:199–201.
Piola, F., P. Label, P. Vergne, P. von Aderkas and R. Rohr. 1998. Ef-fects of endogenous ABA levels and temperature on cedar (Cedruslibani Loudon) bud dormancy in vitro. Plant Cell Rep. 18:279–283.
Piola, F., R. Rohr and P. Heizmann. 1999. Rapid detection of geneticvariation within and among in vitro propagated cedar (Cedruslibani Loudon) clones. Plant Sci. 141:159–163.
Preece, J.E. 1995. Can nutrient salts partially substitute for plantgrowth regulators? Plant Tissue Cult. Biotechnol. 1:26–37.
Schenk, R.V. and A.C. Hildebrandt. 1972. Medium and techniquesfor induction and growth of monocotyledonous and dicotyledo-nous plant cell cultures. Can. J. Bot. 50:199–204.
Schestibratov, K.A., R.V. Mikhailov and S.V. Dolgov. 2003. Plantletregeneration from subculturable nodular callus of Pinus radiata.Plant Cell Tissue Organ Cult. 72:139–146.
Shen, X. and S. von Arnold. 1982. In vitro formation of adventitiousplantlets from embryos of Pinus sylvestris L. Sci. Silvae Sin. 18:405–408.
Siniscalco, C. 1994. Grafting propagation of cedar selected plants(Cedrus atlantica). Ann. Rech. For. Maroc 27:475–486.
Skoog, F. 1944. Growth and organ formation in tobacco tissue cul-tures. Am. J. Bot. 31:19–24.
Sokal, R.R. and F.J. Rohlf. 1995. Biometry. 3rd Edn. W.H. Freeman,New York, 887 p.
Talhouk, S.N., R. Zurayk and S. Khuri. 2001. Conservation of the co-niferous forests of Lebanon: past, present and future prospects.Oryx 35:206–215.
Tang, W., R. Whetten and R. Sederoff. 2001. Genotypic control ofhigh-frequency adventitious shoot regeneration via somatic or-ganogenesis in loblolly pine. Plant Sci. 161:267–272.
Tantos, A., A. Mészáros, T. Farkas, J. Szalai and G. Horváth. 2001.Triacontanol-supported micropropagation of wood plants. PlantCell Rep. 20:16–21.
Thorpe, T.A., I.S. Harry and P.P. Kumar. 1991. Application of micro-propagation to forestry. In Micropropagation: Technology and Ap-plication. Eds. P.C. Debergh and R.H. Zimmerman. KluwerAcademic, Dordrecht, pp 311–336.
Toth, J. 1980. Le cèdre dans les pays du pourtour mediterranéen etdans deux autres pays et son importance forestière. Forêt méditerr.2:23–30.
Tukey, J.W. 1953. Some selected quick and easy methods of statisticalanalysis. Trans. N.Y. Acad. Sci. Series II 16:88–97.
Tzfira, T., O. Yarnitzky, A. Vainstein and A. Altman. 1996. Agrobac-terium rhizogenes-mediated DNA transfer in Pinus halepensisMill. Plant Cell Rep. 16:26–31.
Villalobos-Amador, E., G. Rodríguez-Hernández and E. Pérez-Molphe-Blach. 2002. Organogenesis and Agrobacterium rhizo-genes-induced rooting in Pinus maximartinezii Rzedowsky andP. Pinceana Gordon. Plant Cell Rep. 20:779–785.
von Aderkas, P. and J.M. Bonga. 2000. Influencing micropropagationand somatic embryogenesis in mature trees by manipulation ofphase change, stress and culture environment. Tree Physiol. 20:921–928.
von Arnold, S. and T. Eriksson. 1982. Production of adventitiousplants from spruce and pine. In Symposium of Clonal Forestry. No.32. Swedish University of Agricultural Science. Dept. Forest Ge-netics, Uppsala, pp 7–13.
von Arnold, S. and T. Eriksson. 1986. Norway spruce (Picea abies).In Biotechnology in Agriculture and Forestry. Vol. 5. Ed. Y.P.S. Bajaj.Springer-Verlag, Berlin, pp 291–310.
486 RENAU-MORATA, OLLERO, ARRILLAGA AND SEGURA
TREE PHYSIOLOGY VOLUME 25, 2005
Dow
nloaded from https://academ
ic.oup.com/treephys/article/25/4/477/1659578 by guest on 08 February 2022




















