
Face recognition as a function of social attention in non-human primates: an ERP study
12
ELSEVIER Cognitive Brain Research 2 (1994) l-12 COGNITIVE BRAIN RESEARCH Research Report Face recognition as a function of social attention in non-human primates: an ERP study Jaime A. Pineda *, Gillian Sebestyen, Carlos Nava Department of Cognitir,e Science 0515, Unir~ersir):of Cakforniu, San Diego, La Jolla, CA 9209.W53.5, USA Accepted 28 December 1993 Abstract Elpidural event related potentials (ERPs) were recorded from four squirrel monkeys (Saimiri sciureus) during the presentation of pictoral stimuli that comprised real human and monkey faces. Subjects viewed tachistoscopically presented stimuli belonging to four different categories: familiar and unfamiliar human faces, and familiar and unfamiliar monkey faces. Familiar faces were subcategorized into top, middle and bottom according to the perceived individual’s dominance ranking in a social hierarchy, as rateJ by human judges observing the group’s social behavior. Waveform peak components to monkey and human faces showed similarities in their spatial distribution. However, larger amplitude Nl and N2 components were elicited in response to monkey compared to human faces, particularly over lateral temporo-parietal sites. A similar trend was observed for the P3 component, with maximal differences along midline electrode sites. Responses to familiar and unfamiliar monkey faces showed larger Nls to familiar monkey faces and larger P3s to unfamiliar monkey faces. Nl and P3 components elicited by human faces showed no significant differences between conditions. N2 amplitudes were larger over posterior sites for top-ranked monkeys and larger over frontal sites for middle-and bottom-ranked monkeys. Top-ranked human faces elicited the largest N2 components, midtlle-ranked faces the next largest, and bottom-ranked faces the smallest. Nl, N2, and P3 latencies were similarly sensitive to the ranking of human but not monkey faces. These data suggest that non-human primates exhibit evoked potentials to conspecific and non-conspecific faces that are similar in morphology but different in function. Larger amplitude responses to monkey faces suggests increased processing for that category of stimuli. Additionally, monkey ERPs reflect familiarity with compecifics but not with human faces. Finally, the social status of the perceived individual, or at least the perceived threat posed by an individual, affects the latencies and magnitudes of ERP components produced by the viewer. These data are consistent with social attention hypotheses which propose that higher status (i.e. more dominant or socially meaningful) members of a gr0L.p reccivc more attention than lower status individuals. Key words: Monkey; Social status; Hierarchy; Event related potential; Nl; N2; P3 1. Introduction The ability to recognize faces quickly and efficiently is important for communication among humans be- cauxe it provides large amounts of information about the perceived individual in a complex social situation. Information can be gained regarding the identity, gen- der. emotional state, age, expression, and intent of the person [l&19]. Since a number of behavioral studies havl2 shown that non-human primates live in communi- ties in the wild involving very intricate social structures [1,15,37], it would be expected that face perception and * Glrresponding author. Fax: (1) (619) 534-l 128. 0926.6410/94/$07.00 0 1994 Elsevier Science B.V. All rights reserved SSD’ 0926-6410(94)00002-C recognition would also play an important role in their social interactions [ 12,341. Indeed, non-verbal mimic expressions and gestures are prominent communicative signals that regulate monkey social communication. In particular, one aspect of non-verbal communication that is critical to the interactions among non-human primates is recognition of the social status of the per- ceived individual [7,21]. It has been noted by several investigators that when viewing other conspecifics, monkeys elicit responses appropriate to the perceived individual’s social status [11,20]. This type of social attention is essential for dyadic interactions among group members, as well as for the coordination of group activities [391. Non-hu- man primates will also respond to pictoral stimuli as if
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Face recognition as a function of social attention in non-human primates: an ERP study

ELSEVIER Cognitive Brain Research 2 (1994) l-12
COGNITIVE BRAIN
RESEARCH
Research Report
Face recognition as a function of social attention in non-human primates: an ERP study
Jaime A. Pineda *, Gillian Sebestyen, Carlos Nava Department of Cognitir,e Science 0515, Unir~ersir): of Cakforniu, San Diego, La Jolla, CA 9209.W53.5, USA
Accepted 28 December 1993
Abstract
Elpidural event related potentials (ERPs) were recorded from four squirrel monkeys (Saimiri sciureus) during the presentation of pictoral stimuli that comprised real human and monkey faces. Subjects viewed tachistoscopically presented stimuli belonging to four different categories: familiar and unfamiliar human faces, and familiar and unfamiliar monkey faces. Familiar faces were subcategorized into top, middle and bottom according to the perceived individual’s dominance ranking in a social hierarchy, as rateJ by human judges observing the group’s social behavior. Waveform peak components to monkey and human faces showed similarities in their spatial distribution. However, larger amplitude Nl and N2 components were elicited in response to monkey compared to human faces, particularly over lateral temporo-parietal sites. A similar trend was observed for the P3 component, with maximal differences along midline electrode sites. Responses to familiar and unfamiliar monkey faces showed larger Nls to familiar monkey faces and larger P3s to unfamiliar monkey faces. Nl and P3 components elicited by human faces showed no significant differences between conditions. N2 amplitudes were larger over posterior sites for top-ranked monkeys and larger over frontal sites for middle-and bottom-ranked monkeys. Top-ranked human faces elicited the largest N2 components, midtlle-ranked faces the next largest, and bottom-ranked faces the smallest. Nl, N2, and P3 latencies were similarly sensitive to the ranking of human but not monkey faces. These data suggest that non-human primates exhibit evoked potentials to conspecific and non-conspecific faces that are similar in morphology but different in function. Larger amplitude responses to monkey faces suggests increased processing for that category of stimuli. Additionally, monkey ERPs reflect familiarity with compecifics but not with human faces. Finally, the social status of the perceived individual, or at least the perceived threat posed by an individual, affects the latencies and magnitudes of ERP components produced by the viewer. These data are consistent with social attention hypotheses which propose that higher status (i.e. more dominant or socially meaningful) members of a gr0L.p reccivc more attention than lower status individuals.
Key words: Monkey; Social status; Hierarchy; Event related potential; Nl; N2; P3
1. Introduction
The ability to recognize faces quickly and efficiently is important for communication among humans be- cauxe it provides large amounts of information about the perceived individual in a complex social situation. Information can be gained regarding the identity, gen- der. emotional state, age, expression, and intent of the person [l&19]. Since a number of behavioral studies havl2 shown that non-human primates live in communi- ties in the wild involving very intricate social structures [1,15,37], it would be expected that face perception and
* Glrresponding author. Fax: (1) (619) 534-l 128.
0926.6410/94/$07.00 0 1994 Elsevier Science B.V. All rights reserved SSD’ 0926-6410(94)00002-C
recognition would also play an important role in their social interactions [ 12,341. Indeed, non-verbal mimic expressions and gestures are prominent communicative signals that regulate monkey social communication. In particular, one aspect of non-verbal communication that is critical to the interactions among non-human primates is recognition of the social status of the per- ceived individual [7,21].
It has been noted by several investigators that when viewing other conspecifics, monkeys elicit responses appropriate to the perceived individual’s social status [11,20]. This type of social attention is essential for dyadic interactions among group members, as well as for the coordination of group activities [391. Non-hu- man primates will also respond to pictoral stimuli as if

to the real object [ 11,20,34]. Chimpanzees, for example, have been shown to recognize photographs of humans with whom they share a strong social relationship [lo].
Several studies have argued that high status mem- bers in non-human primate social groups are typically the center of attention [14,23]. However, a number of human and animal studies have failed to find an associ- ation between dominance status and social attention [39,40]. In general, these type of studies suffer from a lack of a clear operational definition of social atten- tion, a lack of support for the notion that social atten- tion refers to a single concept, and the lack of a rigorous method for evaluating the behavior. Hence, it would bc useful within this framework to examine the relationship between social status and a more clearly defined dimension of social attention, such as face recognition. Furthermore, a technique to record brain electrical activity, such as event-related potentials (ERPs), that would be sensitive to the temporal dynam- ics of these processes would be useful.
Previous studies have shown that specific aspects of face processing can be characterized by ERPs in hu- mans [3-5,241 and monkeys [31,32]. These studies indi- cate that ERP indices of face perception and recogni- tion can reflect the analysis of familiar and unfamiliar conspecific faces [3 1,321. They also indicate that human and monkey ERPs to faces are typically largest near the superior temporal sulcus (STS) in temporal cortex [9,24,31]. This is consistent with single cell studies of face recognition in non-human primates which support the hypothesis that specific cell populations in the superior temporal polysensory area (STP) of STS and in the inferior temporal cortex (IT) respond singularly to the configurational aspect of faces [6,13,17,18,22,25]. Cells in these regions respond independently of species, head orientations, facial components, changes in size, contrast. color, or viewing distance [30,33]. Moreover, cells respond to real faces as well as to photographed ones [171. Recent evidence argues that cells which respond to identity may be segregated in the TEm subdivision of IT, apart from cells which respond to expression, which are found primarily in the TPO subdivision of STP [22]. This segregation of function- ally distinct groups of cells, in combination with other cells that arc primarily sensitive to facial features, such as noses and eyes, give support to the idea of a face recognition network in temporal cortex [17]. Such a specialized detection and recognition system would be significant in a monkey’s ability to extract meaning from faces.
The goal of this study is to examine whether face recognition in non-human primates, as an aspect of social attention, can be characterized electrophysiolog- ically and whether this process is related to or affected by the social status of the perceived individual. To date, no ERP studies have addressed these issues. The
main questions to be answered are: (1) are there ERP differences in monkey responses to human versus mon- key faces? (2) are there differences in their responses to familiar versus unfamiliar faces, i.e. whether con- specifics or not‘? and (3) does the social status of the perceived individual affect how they are perceived?
2. Materials and methods
2 1. Subjects
Subjects were six adult. male squirrel monkeys (Samiri sc~urrw.~).
Their ages ranged from 7715 years and weighed hetween 700~1000 g. All monkeys had taken part in previous ERP studies and had prior interaction as a social unit. Only four of the monkeys had implanted electrodes to measure the electroencephalogram (EEG). The re- maining two monkeys were used in the behavioral part of the study to determine social rankings.
Monkeys had been implanted with an array of electrodes for at least one year prior to this study (see Pineda and Nava [31] for details). In summary, monkeys were anesthetized using ketaminr hydrochloride (SO mg/kg Cm.) and atropine (0.05 mg/kg i.m.). Anesthesia was maintained with isoflurane tO.S-1.5%) mixed with oxygen (1~1.5 I/min). while the animal was on a heating blanket and artificially respirated (12-15 breaths/mink
Subjects were placed in a Kopf stereotaxic apparatus, and using aseptic surgical procedures the skin overlying the dorsal aspect of the skull was resected, the underlying muscles retracted, and an array of small stainless steel screws (00 gauge, 1.5 mm length) were threaded into burr holes in the skull. Wire leads from these electrodes were attached to a multi-pin connector, and electrodes, leads and connec- tor were fixed to the skull with dental acrylic. Two small metal cylinders (5.7 cm long, 0.8 cm in diameter) were attached for later use in fixing the head during experimental sessions.
2.3. Experimental paradigm
As shown in Fig. I. stimuli consisted of 140 color slides of real human and squirrel monkey faces presented upright. The hack- ground was darkened so that only faces were visible. Slides consisted of 35 unique, unfamiliar human faces, 3.5 non-unique, familiar hu- man faces (i.e. multiple presentations of faces belonging to lab memhers who interacted with the monkeys on a daily basis), 35 unique. unfamiliar, squirrel monkey faces, and 35 non-unique, famil- iar. squirrel monkey faces (i.e. multiple representations of roommate faces). Human and monkey familiar faces were further subdivided into 10 slides of the top-ranked individual in a social hierarchy, IS slides of the middle-ranked individuals, and 10 slides of the hottom- ranked individual.
Slides were presented pseudorandomly for 500 ms using a com- puter controlled Kodak Ektagraphic Projection Tachistoscope with automatic shutter. They were hack-projected at a slight angle from a distance of 64 cm onto a translucent screen in front of the subject. Subjects viewed the images through a glass window from within an acoustically isolated chamber at a distance of 34 cm. Images sub- tended a visual angle of approximately 9.5” X 9.5”. A 4-s interstimulus interval (1st) occurred between slides. ERPs were recorded in one session for each subject. Lights were turned off inside the chamber and in the outer room during the experiment.

J.A. Pineda et 01. / Cogniti~3e Brain Research 2 (1994) l-12 3
PARADIGM TOTAL SLIDES
140
70 ' '70 (HUMAN) (MONKEY)
/\
(UNF! MILIAR) (FAMILIAR) (UNFAMILIAR) (FAMILIAR)
/I\ 10 15 10
/I\ 10 15 10
ITOP) (MIDDLE) (BOTTOM) (TOP) (MIDDLE) (BOTTOM)
Fig. I, Schematic of the structure of the paradigm in which 140 total slide\ were presented with 70 human and 70 monkey faces. These two categories of faces were further subdivided into 35 unfamiliar and 35 familiar faces. Additionally, familiar faces were subdivided into lop, middle. and bottom ranked individuals according to a social dominance hierarchy that was determined from individual behavioral prof les.
2.4. Social hierarchy
Ranking the six monkey subjects on a social hierarchy was based on observations conducted during ten sessions over the course of a month. All subjects were placed in a large cage consisting of two contiguous, large (5’ X 3’ X 3’) cages that contained branches, toys, ropes, and a water bottle. Subjects were easily recognized by their actic,ns and identifiable markings. Natural behaviors, including domi- nantc gestures, signs of aggression or submission, displays of author- ity, ,Inti-social behaviors, vocalizations, and patterns of interaction amo.lg the group were noted by an observer standing approximately 5 fe~:t in front of the cage. Observations were made as to which monkey initiated an action and who was the target of that action.
111 separate sessions, the observer manipulated the environment by placing small quantities of food in the playground. Observations wert made as to the level of competition for the food, which monkeys obtained the food, and the order in which it was obtained. In a few instances, to clarify the relationship among monkeys in the middle of the hierarchy, pairs of monkeys were placed in a single cage to more closely observe their interactions.
Iamiliar human faces were categorized in terms of a hierarchy based on the monkeys’ ‘fear’ reaction to the perceived individual. These reactions included cowering. avoidance of direct eye contact, mov ng to the back of the cage, and baring of teeth. The person when the subjects found to be most threatening (JP) was coded as top, the least threatening person. who elicited the least reaction, was coded as bottom (GS), while the remaining faces were coded in the middle rank. Such a structure is not analogous to the monkey dominance hierarchy, but it was important to maintain a balanced paradigm.
2.5. I:EG recording protocol
Monkeys were first acclimated to a specially designed primate chai: and to the experimental setup. The chair was designed for limited movement and included a restrictive collar that kept the
monkey limbs away from their heads. Once seated in the chair. monkey subjects were placed inside an acoustically shielded, sound attenuating chamber. The chair was positioned so that the monkey faced the screen. A cable was attached to the connector on the monkey‘s head to relay signals to the EEG polygraph. Electrode aitea recorded were analogous to those described hy the IO-20 Interna- tional Electrode Placement System. This system for electrode place- ment allows direct comparison of monkey and human ERP data. Signals were recorded from four midline (Fz, Cz, Pz, Oz) and four lateral (T3, T4, P3, P4) sites. Electrode> placed in the bony orhit dorsolateral to hoth eyes were used to monitor blinks and other eye movements. All sites were referenced to an electrode implanted approximately 1-2 mm below the inion.
Eight channels of EEG data were amplified by a Grass model 7D polygraph using 7PSB pre-amplifiers with handpass limits of 0.15 Hz and 35 Hz. EEG activity time-locked to stimulus presentation wah digitized on-line for 100 ms prior to and 900 ms following stimulus onset at a sampling rate of 256 Hz. Data were stored on a computer disk for later analysis.
2.6. Data rrnalysls
ERP averages were obtained based on species (human and mon- key faces), familiarity (familiar and unfamiliar human faces and familiar and unfamiliar monkey faces), and social ranking (top, middle, and bottom ranked human and monkey familiar faces). Grand averages across subjects were calculated for all of these conditions. An artifact rejection program was used to reject EEG trials in which the voltage exceeded i 75 PV (i.e. where blinks or eye movements occurred) or when amplifier blocking resulted from such movements. The following latency windows (in ms) were obtained from visual inspection of the grand averages and utilized for measur- ing the largest positive (P) or negative (N) peak within the interval: NI (60-200); N2 (200-400); P3 (250-600). Latencies (ms) and ampli- tudes (PV) were scored for all peaks, while area (PV-ms) was only obtained for P3. The data were automatically scored, although for approximately 10% of the data records, manual scoring was required. The largest peak amplitude within each window was measured relative to the mean voltage in the IO0 ms prestimulus interval. Latencies, amplitudes, and areas were analyzed using repeated-men- sure analyses of variance (ANOVA) with factors of species (2), familiarity (2), ranking (3). and electrode placement (8).
3. Results
3. I. Monkey dominance rankings
A schematic of the social hierarchy developed from behavioral observations is shown in Fig. 2. This domi- nance hierarchy was the result of observations and analysis by the second author (G.S.) with verification by the other authors. The results indicate that there were at least three levels of dominance in the hierar- chy. The top level was occupied by Grandpa, the oldest (approximately 15 years), unimplanted, and clearly the most dominant squirrel monkey in the colony. He is the largest animal, shows the most aggressive behavior, is rarely challenged, and is typically the first to eat. The middle ranked animals consisted of Peter, Kive, Chomp and Ziggy. Differences in behavior among these four monkeys were subtle, inconsistent, and it was unclear

J.A. Pirleda et 01. / Cognitirv Bruin Re.warch 2 (19941 I-12
HIERARCHY 3.2. Human IX monkey fuces
GRANDPA
/\ PETER KIVE
ZIGGY CHOMP
\/ UELI
Fig. 2. Schematic of the social hierarchy observed among six labora- tory monkeys as judged by three raters. The lack of a clear relation- ship among some of the middle ranked monkeys (e.g. between Peter and Kive) suggested a two-path rather than a linear relationship. Grandpa was clearly the most dominant, while Ueli was clearly the most submissive. These relationships were constant throughout the study.
as to what the exact relationship was among them. For this reason, the linear dominance hierarchy that is typically expected to exist has been drawn with two paths. The lowest level was occupied by Ueli, an im- planted monkey, approximately 11 years old. He was typically the least aggressive, the last to obtain food, and the most submissive of all the animals. The social rankings determined from these observations remained stable throughout the duration of the study.
Grand average ERPs to human and monkey faces are shown in Fig. 3. The waveforms include an Nl-P2- N2-P3 complex that was most clearly visible at the vertex (Cz> site. Nl (mean latency of 103 ms to both human and monkey faces) was masked by a large positivity over frontal cortex, while exhibiting maximum amplitude over temporal sites. N2 (mean latencies: 1X2 ms for monkey faces; 194 ms for human faces) was largest over frontal cortex while P3 (mean latencies: 307 for monkey faces; 290 for human faces) achieved maximum amplitude over posterior midline sites. P3 latencies decreased from front to back by a total of 30 ms.
The similarity in the spatial distribution of Nl am- plitude responses to human and monkey faces is illus- trated in the left column of Fig. 4. Table 1 shows the mean amplitudes and latencies of all the measured components. Increasingly larger Nl responses to hu- man and monkey faces occurred from front to back along midline sites (electrode placement, F,,z, = 13.77, P < 0.001) with no between category differences ob- served. Monkey facts elicited larger amplitude Nl components at lateral sites (species x electrode placc- ment, F,,*, = 4.00, P < 0.01). Larger N2 components were also elicited primarily over lateral electrodes in response to monkey compared to human faces (species x electrode placement, F,,z, = 2.81, P < 0.05). Like Nl, the topography of N2 amplitude responses was similar for both categories of faces (see right column of Fig. 4). That is, N2 decreased in amplitude from front to back along midline sites (electrode placement, F,,?, = 8.22, P < 0.001) with no between category differences. No
Monkey
Fig. 3. Grand average ERPs from four adult squirrel monkeys to upright human and monkey faces. Waveforms are presented from three midline (Fz. Cz, Pz) and two lateral temporal (T3. T4) electrode sites. Note the similarities in waveforms and peak latencies and the enhanced response to monkey faces.

J.A. Pineda et al. / Cognitille Brain Research 2 (1994) I-12
20 -
-60 -
-80 -
Nl N2 20 7
0 rJl
z - -20 - +
9 _ -
“a -40 -
I
-60 -
-80 - 0 Monkey 0 Human
-100 ’ ’ ’ ’ ’ ’ ’ ’ -100 ’ ’ ’ ’ ’ ’ ’ ’ Fz Cz Pz Oz T3 T4 P3 P4 Fz Cz Pz Oz T3 T4 P3 P4
Electrode Sites Electrode Sites Fig. 4. Distribution of Nl (left column) and N2 (right column) amplitudes (pV) in response to human and monkey faces. Note the similarities in the ,?patial distribution of Nl and N2 amplitude responses to the two categories of faces.
statistically significant differences in Nl or N2 laten- tie! to human and monkey faces were observed. Like- wisl:, no statistically significant differences were ob- tained for P3 amplitude or area, although amplitude responses to monkey faces were larger than those for human faces by 8 PV at Cz and 10 PV at Pz (see Fig. 3). Area responses also showed the largest difference (1800 PV-ms) at Pz.
3.3. Familiar us. unfamiliar faces
The grand average ERPs to familiar and unfamiliar human and monkey faces are shown in Fig. 5. Table 2
shows the mean amplitudes and latencies for all com- ponents as a function of familiarity. Nl amplitudes were marginally different in response to familiar and unfamiliar monkey faces (familiarity, F,,, = 6.34, P = 0.086). Larger responses occurred to familiar (-38 pV> compared to unfamiliar (- 24 pV> faces. No statis- tical differences were observed in response to human faces, although the ERP grand average data showed an opposite trend, that is, larger Nl responses to unfamil- iar human faces. N2 amplitudes and Nl latencies also did not show any statistically significant differences between familiar and unfamiliar conditions for either human or monkey faces. In contrast, N2 latencies were
Table 1 Me; II latencies (ms) and amplitudes (PV) for Nl, N2, and P3 in response to human and monkey faces
FZ CZ PZ OZ T3 T4
Nl L ltency
Human 98 85 108 108 104 101 Monkey 94 102 107 102 98 104
Amplitude Human 0.0 - 11.2 - 19.3 - 12.0 - 17.6 - 52.6 Monkey 12.0 ~ 12.5 ~ 26.2 - 16.8 - 22.8 - 77.2
N2 L itency
Human 196 188 195 192 206 186 Monkey 188 188 185 175 182 176
Amplitude Human - 6.0 - 4.6 - 1.2 1.4 - 14.2 - 13.9 Monkey -21.9 - 10.9 - 5.8 - 2.2 - 34.1 - 26.7
P3 L itency
Human 284 284 279 265 300 300 Monkey 304 290 280 265 320 322
Amplitude Human 10.0 13.9 21.6 23.6 9.4 10.1 Monkey 12.4 21.7 31.6 28.5 11.8 11.4
P3 P4
104 118 102 115
- 53.8 - 14.1 -6X.7 ~ 30.0
1x9 204 182 176
- 15.3 ~ lY.5 -27.1 - 39.9
308 301 356 323
10.8 12.4 12.3 13.5

at least 13-20 ms faster to unfamiliar faces at lateral electrode sites (human: familiarity x electrode place- ment, F,,, = 2.51, P < 0.05; and marginally significant for monl&y: familiarity x electrode placement, F,,z, =
2.20, P < 0.08). Larger amplitude P3s were observed in response to unfamiliar (21.7 FV) compared to familiar ( 17.1 FV) monkey faces (familiarity, F,,, = 17.11, P < 0.05). No other statistically significant differences in P3 amplitude, latency, or area occurred.
3.4. Social rank&s
Grand average ERPs to top, middle, and bottom ranked human and monkey faces are shown in Fig. 6,
*rahlc ?
while mean latencies and amplitudes for all compo- nents in these conditions are shown in Table 3. There was a main effect of ranking on Nl latencies to human faces (ranking, FL,<, = 6.90, P < 0.05). That is, bottom ranked human faces elicited faster latencies (88 ms) than either middle (99 ms) or top (99 ms) ranked faces. No statistically significant differences in ranking were observed for Nl amplitude or latency in response to monkey faces, nor was there an effect on Nl amplitude to human faces. N2 amplitudes showed a marginally significant main effect of ranking in responses to hu- man faces, with top-ranked faces eliciting larger ( - 3 1.1 PV) responses than middle ( - 7.1 PV) or bottom (4.2 PV) ranked faces (ranking, (F,,, = 3.87, P = 0.08). As
Mean Iatencies (ms) and amplitudes (WV) for Nl, N2, and P3 in res~onsr to familiar and unfamiliar human and monkev faces
PZ 02 T3 T4 P3 P4
Nl
Latency
fluman
Familiar
Unfamiliar
Monkey
Familiar
Unfamiliar
Amplitude
Human
Familiar
Unfamiliar
Monkey
Familiar
Unfamiliar
N?
Latrncy
Human
Familiar
Unfamiliar
Monhq
Familiar
Unfamiliar
Amplitude
Human
Familiar
Unfamiliar
Monkey
Familiar
Unfamiliar
I’.?
Latency
Human
Familiar
Unfamiliar
Monkey
Familiar
Unfamiliar
Amplitude
I luman
Familiar
Unfamiliar
Monkey
Fnmillar
Untamiliar
FZ c/!
97 YX
YX X4
YO IO I 03 104
6.9 ~ X.6
~ 0.3 ~ IS.9
I I .o ~ IX.7
Y.5 ~ IO.9
IYY I75
194 IXX
lxx I90
IX7 IX5
~ I.9 I .h
~ 14.6 - 13.5
- 26.3 - 13.3
- IO.3 - IO.1
271 2x2 275 28’)
30x 297
304 300
13.7 17.7
6.X 10.x
I I.3 23.7
16.0 23.2
107
III
I IO
IIX
96
I06
IO2
IO3
I OS
IO3
-4Y.l
~ S7.0
~ X2.7
~ 4Y.O
204
100
IYX
172
~ 10.7
- IX.2
~ 32.0
~ 2S.2
324
303
345
323
17.2
X. I
I I.h
15.x
I02
I06
102
IO.?
-4x.x
~ 60.0
-7x.4
-61.7
211
I74
203
I74
~ 12.0
~ 24.5
~ 35.0
72.5
307
310
361
364
14.3
7.7
10.2
17.x
I14
11x
101
I03
100
OX
I04
Oh I 10
I Oh
- 15.2 _ 23.5
-x.7
- Il.6
- IS.6 _ 21.x
- IO.6 _ 73.2
-21.x
- IS.5
-35.2
- IS.‘)
- 44.0 -21.1
IX4
lY2
I OX
17x
21.1
IXX IX8
I90
IX’)
17s
I77
17.3
IX4
177
177
176
2.3 6.0
- 12.8
-21.4
I.‘)
- 0.3
~ 12.1
~ 29.2
- 13.6
- 2.0
- 6.5 ~ 49.5
I.2 - 26.7
- 45.5
- 35.4
264 31X
27.5 301
295 28X
322
312
2x1
30s
so0 2x0
323
326
345
322
245
22.6
22.h 2Y.O
14.2
7.6
16.4
I I.5
3.73 32.3
2h.O
35.0
Y.h
19.X
IO.‘)
13.5

J.A. Pineda et al. /Cognitive Brain Research 2 (1994) I-12
Monkey Human
_ _. (jfl/zam;,;ar 5;s _
200 400 600 800
mscc
Fig. 5. Grand average ERPs from four monkey subjects in response to familiar and unfamiliar monkey (left column) and human (right column) face-. Waveforms were recorded from four midline (Fz, Cz, Pz, Oz) and four lateral (T3. T4, P3, P4) electrode sites but only three midline sites are ,,hown.
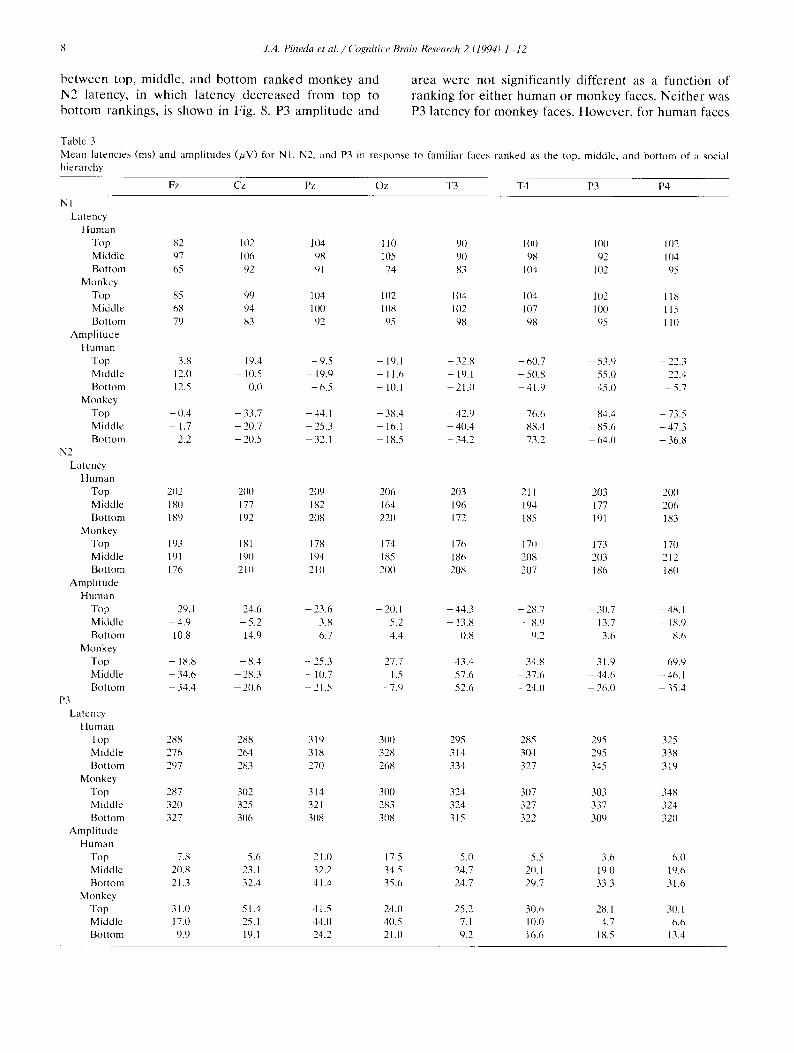
shown in Fig. 7, N2 amplitude responses along midline placement F,, 42 = 2.36, P < 0.05). N2 latency did not electrode sites decreased in amplitude for bottom and show any significant differences to monkey rankings middle ranked monkey faces, but increased in ampli- but did exhibit a linear relationship for human rank- tude for top ranked monkeys (ranking x electrode ings, primarily over temporal sites. This relationship
Monkey Human
5yv _ ,
200 400 600 800
msec
TOP . . . . . . . . . . . . . . . . . . . . . . (f/f,&+
..~~~.._... Bottom
Fig. 6. Grand average ERPs from four monkey subjects to top, middle and bottom ranked monkey (left column) and human (right column) faces, Wa;,eforms from three midline (Fz, Cz, Pz) electrode sites are shown: ‘Note the sustained difference in response to human faces.
x J.A. Pitwdu et ~1. / Cogniriw Bruin Rexwrch 2 (19941 I-12
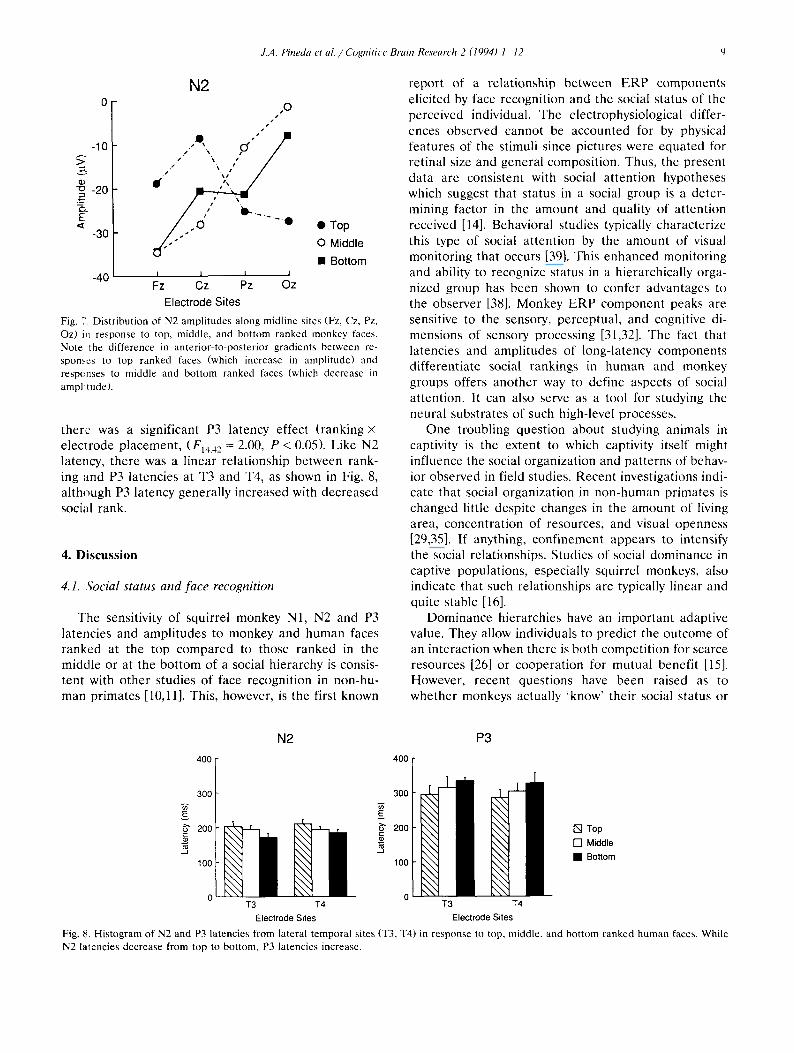
between top, middle, and bottom ranked monkey and area were not significantly different as a function of N2 latency, in which latency decreased from top to ranking for either human or monkey faces. Neither was bottom rankings, is shown in Fig. 8. P3 amplitude and P3 latency for monkey faces. However, for human faces
Table 3 Mean latencies (ms) and amplitudes (PV) for Ni, N2. and I’3 in response to familiar faces ranked as the top, middle. and bottom of B social hierarchy
FL cz PZ OZ
N l
Latency FIuman
Top Middle Bottom
Monkey Top Middle Bottom
Amplitude Human
Top Middle Bottom
Monkey Top Middle Bottom
N2 Latencv
Human Top Middle Bottom
Monkey Top Middle Bottom
Amplitudr Human
Top Middle Bottom
Monkey Top Middle Bottom
P3 Latrncy
Human Top Middle Bottom
Monkey Top Middle Bottom
Amplitude Human
Top Middle Bottom
Monkey Top Middle Bottom
X2 97 hS
I02 IO6 92
I04 9x 91
x5 6X 79
YY 94 x3
104 100 92
3.X 12.0 I2.S
~ 19.4 ~ IO.5
0.0
- 9.5 ~ 19.0
- 6.5
- 0.4 -33.7 - 44.1 - I.7 - 20.7 ~ 25.3
2.2 - 20,s -32.1
202 I80 189
200 177 192
20’) 182 208
193 191 176
1x1 190 210
17x I 04 210
~ 2’). I ~ 24.6 _ 23.6 ~ 4.9 - 5.2 3.X
IO.8 14.9 6.7
-X.4 -2X.3 ~ 20.6
_ 25.3 - 10.7 _ 21.5
288 288 319 276 264 31x 297 2X3 270
2X7 320 327
302 32s 306
S.h 23.1 32.4
51.4 25.1 19.1
314 321 30x
7.x 20.X 21.3
31.0 17.0 Y.9
21 .o 32.2 41.4
41.5 44.0 24.2
I10 1 OS 74
102 I08 YS
- 19.1 - II.6 - IO. I
-3x.4 - 16.1 ~ 18.5
206 I64 220
174 IX.5 200
- 20. I 5. 2 4.4
~ 27.7 ~ 1.5 - 7.9
300 32x 268
300 283 30x
17.5 34.5 35.6
24.0 40.5 21.0
T3 T4 P3 P4
90 90 X3
IO0 ox
IO4
100 92
I02
IO2 I04 YS
104 I02 ox
104 107 ox
IO2 100 YS
II8 IIS I IO
~ 32.x - 19.1 -21.0
- 60.7 - SO.8 -41.9
~ 53.0 ~ 55.0 ~ 45.0
_ 22.3 _ 22.3
-5.7
~ 42.9 - 40.4 - 34.2
- 76.6 ~ XX.4 ~ 73.2
~ x4.4 - X.5.h - 64.0
- 73.5 - 47.3 -36.X
203 211 203 200 I96 lY4 177 206 I72 IX5 191 1x3
I76 IX6 20x
I70 208 207
I70 212 1x0
~ 44.3 - 13.x
0.8
_ 2x.7 - KY ~ 0.7
- 4x. 1 - IX.9
H.h
-33.4 -34.X -57.6 - 37.6 ~ 52.6 - 24.0
I73 203 I Xh
~ 30.7 ~ 13.7
~ 3.6
-31.‘) ~ 44.6 ~ 26.0
20s 29s 34s
303 337 309
3.6 1 Y.0 33.3
28.1 4.7
IX.5
~ 69.9 -46.1 ~ 35.4
2YS 2X5 314 304 334 327
32s 33x 3 I 0
324 324 31.5
34x 324 320
5.0 24.7 24.7
25.2 7.1 0.2
307 327 322
5.5 20. I 29.7
30.6 10.0 16.6
6.0 19.6 31.h
30. I 6.6
13.4
J.A. Pirwdu et ul. / Cognitiw Bruin Resrurch 2 (1994) I-12 9
O-
-10 - _. > 2
8 -20 - 2.
‘La
I -30 -
N2
,’ 0
,’ ,’
.I ,, .I ‘\ d’ .J ‘\ #’
‘\ ’
/’ ‘\ I’ #”
/
#’ .\
d’ .L_.__.
.’ .’
.’
0 Top
0 Middle
W Bottom -40 ’ I I 1 I
Fz Cz Pz Oz Electrode Sites
Fig. _. Distribution of N2 amplitudes along midline sites (Fz, Cz, Pz. Oz) in response to top, middle, and bottom ranked monkey faces. Note the difference in anterior-to-posterior gradients between re- sponses to top ranked faces (which increase in amplitude) and responses to middle and bottom ranked faces (which decrease in amplitude).
there was a significant P3 latency effect (ranking X
electrode placement, (F,,,,, = 2.00, P < 0.05). Like N2 latency, there was a linear relationship between rank- ing ;md P3 latencies at T3 and T4, as shown in Fig. 8, although P3 latency generally increased with decreased social rank.
4. Discussion
4.1. Social status and face recognition
One troubling question about studying animals in captivity is the extent to which captivity itself might influence the social organization and patterns of behav- ior observed in field studies. Recent investigations indi- cate that social organization in non-human primates is changed little despite changes in the amount of living area, concentration of resources, and visual openness [29,351. If anything, confinement appears to intensify the social relationships. Studies of social dominance in captive populations, especially squirrel monkeys, also indicate that such relationships are typically linear and quite stable [16].
The sensitivity of squirrel monkey Nl, N2 and P3 Dominance hierarchies have an important adaptive latencies and amplitudes to monkey and human faces value. They allow individuals to predict the outcome of ranked at the top compared to those ranked in the an interaction when there is both competition for scarce middle or at the bottom of a social hierarchy is consis- resources [261 or cooperation for mutual benefit [15]. tent with other studies of face recognition in non-hu- However, recent questions have been raised as to man primates [lO,ll]. This, however, is the first known whether monkeys actually ‘know’ their social status or
400
300
E
0” 200 5 2
100
0 I--
N2 P3
3
report of a relationship between ERP components elicited by face recognition and the social status of the perceived individual. The electrophysiological differ- ences observed cannot be accounted for by physical features of the stimuli since pictures were equated for retinal size and general composition. Thus, the present data are consistent with social attention hypotheses which suggest that status in a social group is a deter- mining factor in the amount and quality of attention received [14]. Behavioral studies typically characterize this type of social attention by the amount of visual monitoring that occurs [39]. This enhanced monitoring and ability to recognize status in a hierarchically orga- nized group has been shown to confer advantages to the observer [38]. Monkey ERP component peaks are sensitive to the sensory, perceptual, and cognitive di- mensions of sensory processing 131,321. The fact that latencies and amplitudes of long-latency components differentiate social rankings in human and monkey groups offers another way to define aspects of social attention. It can also serve as a tool for studying the neural substrates of such high-level processes.
0 T3 T4
Electrode Sites Electrode Sites
L9 TOP 0 Middle n Bottom
T3 T4
Fig. 8. Histogram of N2 and P3 latencies from lateral temporal sites (T3, T4) in response to top, middle, and bottom ranked human faces. While N2 latencies decrease from top to bottom, P3 latencies increase.

those of their partners [7,8,15]. It is certainly the case that many non-human primates show hierarchically organized relationships in terms of signs of aggression or submission and displays of authority, and these relationships tend to be stable [I(,]. Therefore, some form of hierarchical discrimination might be expected. The differences in ERPs observed in the present study, particularly long-latency components such as the N2. to a top versus lower ranked monkeys supports the hy- pothesis that differences, whether at an individual level or at a categorical level (i.e. very dominant, dominant, subordinate, very subordinate) are perceived, although not whether an animal is cognizant of it. Investigators have argued that the perception of differences in social rank means that some type of mental representation must be involved, allowing monkeys to make compar- isons and draw inferences about social relationships. Furthermore. this type of knowledge in humans is typically declarative rather than procedural. However, the lack of distinct vocalizations for these relationships in most non-human primates argues that monkeys may not be aware of such knowledge [ 15.37].
Squirrel monkeys’ visual perceptual skills are funda- mentally similar to those of Old World monkeys, apes and humans [27]. However, evidence from field re- search suggests that they live in a troop structure where the dispersion of adult males seems to bc associ- ated with a predominantly cohesive female nucleus [2]. To that extent. their social organization differs from that of Old World. terrestrial species. Nonetheless, the prcscnt elcctrophysiological data are consistent with the behavioral findings that many monkeys respond to perceived conspccifics based on their social status [ 11,20.34]. The electrophysiological effects are evident at least as early as 100 ms following stimulus onset, at least in the perception of human faces. and are widely distributed throughout the cortical mantle. The widespread topography of these effects, as well as the apparent enhancement of both positive and ncgativc ERP components, suggests a generalized enhancement in the processing of faces throughout a variety of cortical areas. This is consistent with the idea of an attcntional bias in a distributed network involved in the perception and recognition of meaningful faces.
The ERP responses to human faces as a function of their ranking is surprising because the premise for this hierarchy was not considered analogous to that used to rank monkey faces. Because human rankings were based primarily on the threatening nature of the pic- tured individuals. the systematic change from top to bottom. would be consistent with this categorization. It thus may be that threat. or the possibility of threatcn- ing behavior, is one of several factors involved in establishing monkey social dominance. The sustained difference in response to human faces ranked as top, middle. and bottom of the hierarchy, which occurs
immediately after stimulus presentation, argues for a generalized, non-specific arousal effect. What is sur- prising is that these differences begin so early. A type of priming effect would explain such an early onset. However. the random nature of stimulus presentation would not prime subjects to respond one way or the other to the faces. It is possible, therefore, that catego- rization of the stimuli (i.e. very threatening, somewhat threatening, not threatening) occurs early on in the stream of processing, perhaps at some subcortical level. It is fair to say that monkey Nl, N2, and P.3 compo- nents were all sensitive to the rankings of human t’accs, whereas only N2 was sensitive to the rankings of mon- key faces. The additional effects to the human faces may bc related to the degree of threat perceived. This is an issue that needs further exploration.
The present results indicate that monkeys elicit larger Nl and N2 responses, and tend to elicit larger P3 components, to conspecific compared to non-con- specific faces. Both human and monkey faces elicit waveforms with similar morphologics and peak laten- ties. The main diffcrencc is the cnhanccd response to monkey faces. This additional gain or modulation is similar to the cnhanccd processing of the top ranked monkey compared to the bottom ranked one and may retlcct enhanced attentional processing to conspecifics. The similarities in spatial distribution and in peak latencics but differcnccs in the magnitude of the re- sponse would suggest that the same neural substrates arc being engaged in response to the two categories of faces. Such an interpretation is consistent with the study by Overman and Doty [28] in which monkeys differcntiatcd monkey and human faces from other classes of visual stimuli, suggesting a unique processing mechanism for the stimulus category. Results from single cell studies also show that STS cells respond to faces as a general category rather than to specific individuals (i.e. the ‘grandmother’ cell) [ 171.
A number of explanations for the cnhancemcnt or bias can be postulated. One possibility is that innate mechanisms are hardwired to give special processing to monkey conspecific facts [XI]. Altcrnativcly, the social context and lifetime cxpericnce monkeys have in pro- cessing monkey and human faces would. in itself, be enough to give additional significance or meaning to those facts. One way to test whether prior cxpcriencc is sufficient to produce the type of generalized en- hanccments observed in this study is to cxaminc re- sponses to familiar versus unfamiliar facts.
The present results suggest that familiar and unfa- miliar faces arc indeed processed differently within a

J.A. Pineda et ul. / Cognitiw Bruin Rewurch 2 (I 994) I-12 II
single category (i.e. monkey faces) as well as between categories (i.e. human vs. monkey faces). Monkey ERPs appear sensitive to the familiarity dimension but only for conspecific faces. Within this category, the differ- enccs in processing appear to be component specific. That is, familiar monkey faces produce larger Nls but smaller P3s relative to unfamiliar monkey faces. These effects could result from an additional, overlapping negative wave elicited by familiar stimuli. Thus, famil- iarity per se would not explain the generalized en- hanced negative and positive peaks in response to monkey faces. The lack of statistical differences in response to familiar and unfamiliar human faces sug- gesls that monkey ERPs are unable to distinguish stimuli along this dimension. This implies that there exist distinct face processing mechanisms, or at least distinct processing strategies, for human and monkey facts. Why these differences in processing occur re- mains to be examined.
4.4. Relelwce to human studies
Recognition of faces is an important component of non-verbal human and monkey social communication. Because of the evolutionary link with non-human pri- mates, it is reasonable to assume that the mechanisms by which social status is discriminated may still exist in humans. Such systems may have either retained their function or been adapted for more ethologically rele- vant human behavior, such as allocating attention, via fact recognition, to socially meaningful faces. Human relationships are typically not cast within the context of rigid social hierarchies of the type seen in non-human primate social groups. Some children studies report thar the most aggressive individuals in a group do not necessarily acquire the dominant status in the group. Other factors such as intelligence, skill in interpersonal relations, self confidence, and problem-solving abilities play just as important a role [41]. Nevertheless, some similarities remain. One issue to be explored by a human study is whether, and to what extent, humans are biased to process human faces. Furthermore, ERP stutlies in young children could determine whether sucv a bias is hardwired or simply a function of learn- ing and of tuning a face processing neural network. The phenomenon in which individuals of a racial group, other than one’s own, are difficult to discriminate suggests that learning is critical in human face recogni- tion.
Acknowledgements
This research was supported by a grant from the Air Force Office of Scientific Research (AFOSR).
References
[I] Baldwin, J.D.. The behavior of squirrel monkeys (Srrimiri) in natural environments. In L.A. Rosenblum and C.L. C‘oe (Eda.), Humlhook of‘ Sqrtirrd Monkey Rmwrch Plenum, New York. 19x5, pp. 3.5-53.
[2] Baldwin. J.D. and Baldwin. J.I.. Squirrel monkeys in natural habitats in Panama, Colombia, Brazil. and Peru. Prirnutcs. I2 (lY71) 45-61.
[31 Barrett. S.E. and Rugg. M.D.. Event-related potentials and the semantic matching of faces, N~ctropv?~ho/o~~. 27 (10%)) Y l3-Y27.
[4] Barrett. S.E. and Rug. M.D., Event-related potentials and the semantic matching of pictures. Bruin C‘op~., I3 (IYYO) 2OI~212.
[S] Barrett, SE., Rugg, M.D. and Perrett. D.1.. Event-related po- tentials and the matching of familiar and unfamiliar facts. N~uro/~s~cho/o~~, 26 (198X) 105-I 17.
[6] Baylis. G.C.. Rolls, E.T. and Leonard, C.M., Selectivity hetwren faces in the responses of a population of neurons in the cortex in the superior temporal sulcu~ of the monkey. Bruit! Rc.v.. 34’7 (1985) Y~IOZ.
[71
[Xl
[Yl
[ 101
[Ill
1121
[I31
[I41
[ISI
[If)1
[I71
[IX1
[I’)1
[N
[211
Bernstein, IS.. Dominate: a theoretical perspective for etholo- gist. In D.R. Omark, F.F. Strayer and D.<‘. Freedman (Eds.). Dorninur~c Relatior~s. Gartland Fresh. New York. IYXO. Bernstein. 1.5, Dominance: the baby and the hathwntcr, Uelrrrt,. Bruin Sci., 4 (IYXI) 410-457. Botzel, K. and Grusser. O.J.. Electric brain potentials evoked hy pictures of faces and non-faces: a search for ‘face-specific’ EEG potentials. Exp. Brcm Rex, 77 (19x9) 340%360. Boysen. S.T. and Bernthon. G.G.. C‘ardiac correlate\ of recopn- tion in the chimpanzee (Pun troglotlytec), J. C‘omp PvLY~., IO0 (1986) 312-324. Boyscn. S.T. and Berntson, G.G.. (‘onspecific recognition in the chimpanzee (Purr trog/or/yt~,s ): cardiac response\ to significant others. J. C‘on7p. Psydz., 103 (IYXY) 215%220. Bruce, C., Face recognition by monkeys: absence of an inversion effect. N~rrr~~~~.s~c,llol~~,~i[~. 20 ( IYX2) 5 15-527. Bruce. C.. Drsimone, R. and Gross. C.G.. Visual propertie\ ol neurons in polysensory area in superior temporal suIcu\ of the macaque, J. N~wroph~sio/. . 4h ( I YX I) 369-3X4. Chance, M.R.A. and Larsen, R.R., Tire Socrd Struc~turc o/’ Attention, John Wiley, New York. lY7h. Cheney, D.L. and Seyfarth. R.M.. The representation of social relations hy monkeys, C‘ognitiorr, 37 (IYYO) lh7-IYh. Clark. D.L.. Kessler. K.L. and Dillon. J.E.. Long-term stability of pairwise social dominance in squirrel monkeys. Bull. /~cyc~/wu. sot., 2 (lY73) 203-20s. Desimone, R., Face selective cells in the temporal cortex of monkeys. J. C‘ogr~. Nwro.wi.. 3 (IYYI) I-X. Desimone, R.. Albright. T.D.. Gross. C.G. and Bruce. C.. Stimu- lus-selective responses of inferior temporal neuron, in the macaque, J. Nwrosci.. 4 (lY84) 205 l-2062. Ellis. A.W., Cognitive mechanism\ of face processing. Phil. Trum R. Sot. London Ser. B.. 335 (lW2) I l3- I IY. Fragasry, D.M.. Cognition in squirrel monkeys: a contemporary perspective. In L.A. Rohenhlum and C.L. C‘oe (Eda.), Hrmtlhook of‘ Squirrel Marl key Rrseu rch . Plenum, New York. IYXS. pp. 5%YX. Gauthier, R. and Strayer, F.F.. Empirical techniques fol- the identification of dominance class. In D.M. Tauh and F.A. King (Eds.), C‘lrrretlr Prrsprc~tiws m Primute Socrc~l l~wwrnic~.s. Van Nostrand Reinhold Company, New York. 1086, pp. 120- 133.
[22] Hasselmon, M.E., Rolls. ET. and Baylis, G.C.. The role of expression and identity in the face-selective responses of neu- rons in the temporal visual cortex of the monkey. Wehtr/.. Brui~I Rex. 32 (IYXY) 203-218.

[2.3] liaude. R.H.. Ciraher, J.G. and Farres. A.G.. Visual observing 111 I-hews monkeys: some relationship\ with social dominance rank. A,fi,lln/ [.ctrr,rir1!: B&trr . 4 (iY76) lh3- Ihh.
[24] Jeff!-cys. D.A., A face-responsive potential recorded from the human scalp. /$. Rrcrifr Res., 7X (19X9) 103-202.
[251 Kendrick. K.M. and Bald win. B.A.. Cells in temporal cortex of conscious sheep can respond preferentially to the sight of faces. SC 1(‘11(‘(‘. 2.36 ( 19X7) 44X-150.
[26] Lancaster. J.B.. t’rirutrrc, UC/~CI/ rw rrrrti t/w Etm,r~,~ewc~ of ffw~~rrtl (‘~rlruw. Holt. Rinehart and Winston. New York. lY7.5.
[77] Marriott. B.M., /Qrure pc’tw[~rcorr 11, .\cllrrtwl tuo~r/\c~~.s (Sarrurrt wrrrwr.r). Ph.D. Dissertation University of Aberdeen (University Microfilma International No. 7X102-17). 1Y7h.
[2X] Overman. W.El. and Doty. R.W.. IIemispheric specialization displayed by man but not macaques for analysis of faces, ,\‘cw w/~\w/Io/oh’itr. 20 ( I ‘)X2) I I3- 12X.
[2Yl Prrloe. S.I.. Conflict. afflllation. mating. and the effects of spatial confinement in a captive group of squirrel monkeys (Srr/urrr-; .sc~iur-cws). In D.M. Tauh and F.A. King (Ed\.). C’wrcwt ~cr-;lwc’trt 6’s 111 f%rr~trre Socrtrl @w,nic.s. Van Nostrand Reinhold <‘o.. NC\\ York. IYXh. pp. X%YX.
[3(1] Prrr-ett. D.I.. Rolls. ET. and Caan. W., Visual neuroneb rehpon- sivc to face5 in the monkey temporal cortex, tg,. Urrri/r Rex, 47 ( 1’1x7) 3’%312.
[.?I] Pincda. J.A. and Nau. c‘.. Event-related potentials in macaque monks) during paa5ive and attentional processing of faces in a pl-iminp paradigm. &/ztri Brrrirt Res.. 5.1 (1903) l77- 1X7.
1311 Pincda. J.A. and Sticknry, G.. Human and monkey N400-like component\ during the prowssing of familiar and unfamiliar fact\. in preparation.
[331 Rolls. E.T. and Baylis. G.C., Size and contrast have only small effects on the responses to faces of neurons in the cortex of the superior temporal sulcus of the monkey, ,%/I. Brrritr Rex, 6.5 (1986) 3X-48.
[341 Rosenfeld. S.A. and van Hoesen. G.W.. Face recognition in the rhesus monkey, N~u~ol,.s~cho/ofiicl. I7 (1979) 503-509.
[351 Rowell, T.E., A quantitative comparison of the behavior of a wild and a caged baboon group. Awnul Brlrcrl,., IS (1967) 4”)9%5OY.
[%I Sackctt. G.. Monkeys reared in isolation with pictures as visual stimuli: evidence for an innate releasing mechanism. Scrcnw, 153 (lYh6) 146X-1473.
[37] Seyfarth. R.M. and Cheney. D.L. Meaning and mind in mon- keys. Sci. &I.. 267 (1892) 172-128.
[3X] Smuts. B.B.. Cheney. D.W.. Wrangham. R.W. and Struhsaker. T.T.. Pwmrtr Socierics, University of Chicago Press. Chicago, IYX7.
[39] Strayer, F.F. and Gariepy. J.L.. The structure of social attention and its coordination with cohesive and dispersive activities in captive group5 of squirrel monkeys. In D.M. Taub and F.A. King (Eds.). (‘ut-twrt Pcrcpcc’tir e in Prirmte So&l Dyrumics. Van Nostrand Reinhold Company, New York. lY86. pp. 9’1-1 IO.
[40] Vaughn. B. and Waters. E., Attention structure. sociometric status. and dominance: interrelationa. behavioral correlates. and relationships to social competence. Del, Psq’ch.. 17 (19X1) 27S- ‘XX.
[II] Willhoite Jr.. F.H., Reciprocity. political origins. and legitimacy. Paper presented at the annual meeting of the American Politi- cal Science Association. Washington. DC, 1980.




















