
Extremely high MHC class I variation in a population of a long-distance migrant, the Scarlet...
11
ORIGINAL PAPER Extremely high MHC class I variation in a population of a long-distance migrant, the Scarlet Rosefinch (Carpodacus erythrinus) M. Promerová & T. Albrecht & J. Bryja Received: 15 October 2008 / Accepted: 20 April 2009 / Published online: 19 May 2009 # Springer-Verlag 2009 Abstract Although the number of studies focusing on the major histocompatibility complex (MHC) in non-model vertebrates is increasing, results are often contradictory, and the structure of MHC is still poorly understood in wild species. Here, we describe the structure and diversity of exon 3 of MHC class I in a passerine bird, the Scarlet Rosefinch (Carpodacus erythrinus). Using capillary elec- trophoresis single-strand conformation polymorphism, we identified 82 different MHC class I variants in one Rose- finch population nesting at one site in the Czech Republic. Thus far, this is the highest intra-populational MHC class I variation observed in birds. We have not found support for ‘minimal essential’ MHC in this species since individuals exhibited between three and nine different exon 3 sequen- ces, indicating that there may be at least five amplified MHC class I genes. By cloning, we obtained and analysed 29 exon sequences and found that all of them could be translated into potentially functional proteins. We also show that strong positive selection appears to be acting mainly, but not only, on previously described antigen-binding sites in MHC class I genes. Furthermore, our results indicate that recombination has played an important role in generating genetic diversity of these genes in the Scarlet Rosefinch; we discuss the significance of this extremely high genetic diversity in light of the life history traits of this species, such as long-distance migration. Keywords MHC class I . Scarlet Rosefinch . Carpodacus erythrinus . Positive selection . Recombination Introduction The genes of the major histocompatibility complex (MHC) represent an important component of the vertebrate ac- quired immune system, since they encode transmembrane proteins which present antigen peptides to T cells for recognition, thus initiating an immune response (e.g. Klein 1986; Abbas et al. 1994; Hughes and Yeager 1998). MHC genes are by far the most polymorphic coding regions of the vertebrate genome (Klein 1986). In birds, the most complex studies of MHC structure involve galliform species, mainly the domestic chicken (Gallus gallus), in which MHC is called the ‘B-complex’. This region contains two class I and two class II B genes together with 15 other closely linked genes (Kaufman et al. 1999). In addition to the B-complex, MHC genes in the domestic chicken have been found in another genomic region called Rfp-Y (Miller et al. 1994). The term ‘minimal essential’ is used to characterise the chicken MHC because this genomic region is much smaller than that of mammals and very gene-rich, as the genes are flanked by very short introns (Kaufman et al. 1995; Kaufman 1999). Some evidence indicates that this pattern is also similar in other galliforms, e.g. ring-necked pheasants (Phasianus colchi- cus; Jarvi et al. 1999; Wittzell et al. 1999a) and the Black grouse (Tetrao tetrix; Strand et al. 2007), as well as in other Immunogenetics (2009) 61:451–461 DOI 10.1007/s00251-009-0375-x The sequence data described in this paper are accessible in GenBank data library under accession numbers FJ392762–FJ392790. M. Promerová (*) : T. Albrecht : J. Bryja Department of Population Biology, Institute of Vertebrate Biology, Academy of Sciences of the Czech Republic, Studenec 122, 675 02 Koněšín, Czech Republic e-mail: [email protected] T. Albrecht Department of Zoology, Faculty of Science, Charles University in Prague, Viničná 7, 128 44 Prague, Czech Republic
Transcript of Extremely high MHC class I variation in a population of a long-distance migrant, the Scarlet...

ORIGINAL PAPER
Extremely high MHC class I variation in a populationof a long-distance migrant the Scarlet Rosefinch(Carpodacus erythrinus)
M Promerovaacute amp T Albrecht amp J Bryja
Received 15 October 2008 Accepted 20 April 2009 Published online 19 May 2009 Springer-Verlag 2009
Abstract Although the number of studies focusing on themajor histocompatibility complex (MHC) in non-modelvertebrates is increasing results are often contradictory andthe structure of MHC is still poorly understood in wildspecies Here we describe the structure and diversity ofexon 3 of MHC class I in a passerine bird the ScarletRosefinch (Carpodacus erythrinus) Using capillary elec-trophoresis single-strand conformation polymorphism weidentified 82 different MHC class I variants in one Rose-finch population nesting at one site in the Czech RepublicThus far this is the highest intra-populational MHC class Ivariation observed in birds We have not found support forlsquominimal essentialrsquo MHC in this species since individualsexhibited between three and nine different exon 3 sequen-ces indicating that there may be at least five amplifiedMHC class I genes By cloning we obtained and analysed29 exon sequences and found that all of them could betranslated into potentially functional proteins We also showthat strong positive selection appears to be acting mainlybut not only on previously described antigen-binding sitesin MHC class I genes Furthermore our results indicate thatrecombination has played an important role in generating
genetic diversity of these genes in the Scarlet Rosefinch wediscuss the significance of this extremely high geneticdiversity in light of the life history traits of this speciessuch as long-distance migration
Keywords MHC class I Scarlet Rosefinch
Carpodacus erythrinus Positive selection Recombination
Introduction
The genes of the major histocompatibility complex (MHC)represent an important component of the vertebrate ac-quired immune system since they encode transmembraneproteins which present antigen peptides to T cells forrecognition thus initiating an immune response (eg Klein1986 Abbas et al 1994 Hughes and Yeager 1998) MHCgenes are by far the most polymorphic coding regions ofthe vertebrate genome (Klein 1986)
In birds the most complex studies of MHC structureinvolve galliform species mainly the domestic chicken(Gallus gallus) in which MHC is called the lsquoB-complexrsquoThis region contains two class I and two class II B genestogether with 15 other closely linked genes (Kaufman et al1999) In addition to the B-complex MHC genes in thedomestic chicken have been found in another genomicregion called Rfp-Y (Miller et al 1994) The term lsquominimalessentialrsquo is used to characterise the chicken MHC becausethis genomic region is much smaller than that of mammalsand very gene-rich as the genes are flanked by very shortintrons (Kaufman et al 1995 Kaufman 1999) Someevidence indicates that this pattern is also similar in othergalliforms eg ring-necked pheasants (Phasianus colchi-cus Jarvi et al 1999 Wittzell et al 1999a) and the Blackgrouse (Tetrao tetrix Strand et al 2007) as well as in other
Immunogenetics (2009) 61451ndash461DOI 101007s00251-009-0375-x
The sequence data described in this paper are accessible in GenBankdata library under accession numbers FJ392762ndashFJ392790
M Promerovaacute () T Albrecht J BryjaDepartment of Population Biology Institute of Vertebrate BiologyAcademy of Sciences of the Czech RepublicStudenec 122675 02 Koněšiacuten Czech Republice-mail promerovaseznamcz
T AlbrechtDepartment of Zoology Faculty of ScienceCharles University in PragueViničnaacute 7128 44 Prague Czech Republic
groups of birds such as parrots (Psittaciformes Hughes etal 2008) and owls (Strigiformes Burri et al 2008)However the MHC of some other galliform species suchas the quail (Coturnix japonica) has a larger number ofduplicated genes and the overall structure is much morecomplex (Hosomichi et al 2006 Shiina et al 2004) Aneven more complex MHC structure has been reported forseveral passerine species with many gene copies andabundant pseudogenes (eg Westerdahl et al 1999 2000Hess et al 2000 Freeman-Gallant et al 2002 Miller andLambert 2004 Bonneaud et al 2006a)
Most ecological studies of MHC in birds are traditionallybased on MHC class II while MHC class I genes are oftendisregarded There are only very limited data on MHC classI structure of passerine birds in fact they have only beendescribed in two species In the great reed warbler(Acrocephalus arundinaceus) Westerdahl et al (1999)found four transcribed MHC class I loci in one individualand in the house sparrow (Passer domesticus) there seemto be at least three (Bonneaud et al 2004) to six (Bonneaudet al 2006a) MHC class I loci per individual Despite thefact that MHC class I genes are used only rarely asimmunogenetic markers they might be very important forunderstanding the pathogenndashhost relationships in wildpopulations For example recent studies in wild-livingsongbird species have shown that there may be significantassociations of certain MHC class I variants with theresistance or susceptibility to some parasites (Westerdahl etal 2005 Bonneaud et al 2006b Loiseau et al 2008)
The peptides derived from pathogen fragments arebound to MHC proteins in the so-called peptide-bindingregion (PBR) which is encoded by codons of exon 2 andexon 3 of the MHC class I genes (Wallny et al 2006 Kochet al 2007) These sites are believed to be under strongpositive selection and very variable in order to pursue thearms race with fast-evolving pathogens On the other handthere are sites that are conserved and lack variation sincethey are essential for anchoring the peptide-terminal main-chain atoms (Kaufman et al 1994 Shum et al 1999 Mesaet al 2004) Because many studies use MHC genes asgenetic markers it is important to understand theirevolutionary history revealing the mechanisms maintaininghigh polymorphism may help to achieve this goal Forexample previous molecular studies have disclosed theexistence of trans-species polymorphism (occurrence ofsimilar alleles across different species originating from acommon ancestor) at the MHC (Klein et al 1998) whichsupports the hypothesis of balancing selection Anotherexplanation of the extraordinarily high variation at theMHC may be gene conversion which can produce newsequences or cause concerted evolution when duplicatedgenes evolve in parallel this particular mechanism seems tobe common in birds (eg Wittzell et al 1999b)
Although knowledge regarding the structure and func-tion of the avian MHC has increased significantly in the lastdecade there are still questions that remain unresolved Acomparative approach could help to understand the evolu-tionary mechanisms and ecological importance of MHC inwild populations (see eg Gouumly de Bellocq et al 2008)however before specific hypotheses can be tested theMHC structure in a wide array of species still needs to bestudied because genetic variation especially in MHC class Igenes has only been described in a few wild birds (Jarvi etal 1999 Westerdahl et al 1999 Bonneaud et al 2004Mesa et al 2004 Xia et al 2005) Here we describe thestructure and extremely high intra-population diversity ofexon 3 of MHC class I genes in a long-distance migratorysongbird with pronounced sexual dimorphism the ScarletRosefinch (Carpodacus erythrinus) The study populationnests in central Europe and winters in southern Asiaparticularly in India (Cramp et al 1994) thus these birdstravel thousands of kilometres and are potentially exposedto a wide range of pathogens and parasites Furthermoreusing state-of-the-art methods of sequence analyses weexamine the role of selection and recombination in formingthe diversity of MHC class I in rosefinches and otherpasserine species where data on MHC class I structure areavailable
Materials and methods
Study population sampling and DNA extraction
The Scarlet Rosefinch is a socially monogamous migratorypasserine with a distribution that has been spreadingwestward from Russia in recent years (Cramp et al 1994)The population occupying the Šumava Mountains NationalPark Czech Republic (48deg49prime N 13deg56prime E sim750 m asl)has been studied since 1995 (Albrecht 2004 Polaacutekovaacute et al2007 Albrecht et al 2007) The present study uses samplesfrom 66 adult males and 54 adult females captured at thestudy site during the breeding season in the years 2000ndash2007 Most individuals were captured soon after theirarrival at the study site using mist nets although some birdswere caught later during the chick provisioning stage(Albrecht et al 2007) Blood (approximately 30 μl) wassampled by wing venipuncture and stored in ethanol atminus20degC until DNA extraction Genomic DNA was extractedfrom blood using the Dneasy Blood amp Tissue Kit (Qiagen)according to the manufacturers instructions
Primer design for amplifying genomic exon 3 sequences
We amplified the complete exon 3 of MHC class Itogether with portions of adjacent introns (550 bp) using
452 Immunogenetics (2009) 61451ndash461
the primers HN34 (5prime-CCATGGGTCTCTGTGGGTA-3prime)and HN45 (5prime-CCATGGAATTCCCACAGGAA-3prime) takenfrom Westerdahl et al (2004) using the polymerase chainreaction (PCR) with the following conditions 15 mMMgCl2 04 μM of each primer 02 mM dNTPs 1 UμlTaq polymerase (Fermentas) in appropriate 1times PCR bufferand 1 μl of extracted DNA Deionised water was added toa 10-μl reaction volume The amplification consisted of aninitial denaturation at 94degC (3 min) followed by 30 cyclesof denaturation at 94degC (40 s) annealing at 60degC (30 s)and extension at 72degC (1 min) with a final extension stepat 72degC (10 min) The reactions were run on a Master-cycler ep (Eppendorf) The PCR products were purifiedusing the MinElute PCR purification kit (Qiagen) ligatedto the vector and transformed to bacteria using the pGEM-T Easy Cloning Kit (Promega) according to manufac-turers protocols Positive transformants containing insertsof appropriate length were identified by PCR screeningusing primers on the vector and agarose gel electrophore-sis We randomly chose eight clones extracted plasmidDNA using the QIAprep Spin Miniprep Kit (Qiagen) andsequenced the inserts using the BigDye Terminators Se-quencing Kit v11 and an ABI PRISM 3130 GeneticAnalyzer (Applied Biosystems) Based on these eightsequences a new reverse primer specific for the ScarletRosefinch was designed (CE-MHCREX3 5prime-ACAGGAATTCTGCTCCCACC-3prime) that together with forwardprimer HN34 amplifies the complete exon 3 of the MHCclass I genes of the Scarlet Rosefinch under almost the samePCR conditions as described above (the only difference isthe annealing step which changes to 30 s at 59degC)
Capillary electrophoresis single-strand conformationpolymorphism
Exon 3 of MHC class I was amplified by PCR usingfluorescently labelled primers (HN34 by 6-FAM and CE-MHCREX3 by NED) and conditions described abovePositive PCR amplifications were checked by agarose gelelectrophoresis and then single-strand conformation poly-morphism (SSCP) analysis (a method able to detect even asingle nucleotide substitution) was performed using capil-lary electrophoresis on an ABI PRISM 3130 GeneticAnalyzer (Applied Biosystems) We mixed 2 μl of PCRproduct with 05 μl of GeneScan-500 LIZtrade Size standard(Applied Biosystems) and 12 μl Hi-Ditrade Formamide anddenaturated the mix at 95degC for 3 min After denaturationsamples were immediately put on ice for at least 2 min andelectrophoresis was run at 22degC in 5 non-denaturingconformation analysis polymer (prepared from 9 CAPpolymer Applied Biosystems according to manufacturersinstructions) on an ABI PRISM 3130 Data were analysedusing the software GeneMapper v37 (Applied Biosystems)
and user-defined size standard files for the alignment ofdifferent runs The alleles were identified by both theposition and the shape of peaks For each allele one 6-FAMand one NED peak was identified It is important to notethat neither SSCP peaks nor sequences can be assigned tospecific loci but for simplification in this study they aretermed lsquoallelesrsquo To obtain the sequences of particularalleles (ie SSCP peaks) we cloned the PCR products(obtained by non-labelled primers HN34 and CE-MHCREX3) using the techniques as described aboveSSCP was subsequently used for the identification ofinserted sequences after cloning This procedure allowedus to (1) minimise cost by avoiding the sequencing of thesame sequence in many clones and (2) compare SSCPpatterns of clones with those obtained from individuals toidentify PCR artefacts which are common when analysingMHC genes with a cloning and sequencing approach (Bryjaet al 2005) Inserts providing the SSCP patterns thatmatched the SSCP profile of an individual bird ie theinserts contained one of the exon 3 variants of theindividual were re-amplified by specific PCRs and se-quenced using Big Dye kits as described above
Analyses of MHC class I sequence variation
The sequences of MHC class I derived from C erythrinuswere edited in SeqScape v25 (Applied Biosystems) andaligned in BioEdit Sequence Alignment Editor (Hall 1999)To determine whether all sequences represent functionalclassical class I alleles the sequences were examined (1)for the presence of insertions or deletions (lsquoindelsrsquo) causingshift of the reading frame andor stop codons and (2) forthe conservation of residues that anchor the peptide-terminal main-chain atoms Possible codons encoding theamino acid residues included in the peptide binding wereidentified on the basis of homology between chicken MHCand human leukocyte antigen (HLA)-A (eg Wallny et al2006) and by analysing the detailed structural models ofpeptide-binding grooves of chicken MHC class I proteins(Koch et al 2007) Some of the residues are generallyhighly conserved throughout evolution in classical class Imolecules (Kaufman et al 1994 Mesa et al 2004) whileothers are thought to be under strong positive selection andallow interaction with a wide variety of antigens Sequencesof chicken (GenBank acc no X12780) Japanese quail(D29813) and duck (AY294416) were used to compareMHC sequences of Scarlet Rosefinches with those ofdistantly related bird species
Detecting amino acid sites under selection
To detect selection acting on MHC class I gene variationsequences (partial or complete) of exon 3 MHC class I for
Immunogenetics (2009) 61451ndash461 453
three additional genera of passerines were downloadedfrom GenBank (accession nos in parentheses) theseinclude two species of warbler of the genus Acrocephalus(AF449688ndashAF449701 AJ005503 AJ005504 AJ005506AJ005508ndashAJ005510 AJ005512 AJ005514ndashAJ005517AY306008 AY306009 AJ557874ndashAJ557883) the housesparrow (AY284994ndashAY285013 EF429132) and threespecies of tit of the genus Parus (AF346821 AF346823AF346825 AF346827 AF346829ndashAF346832 AM232707 AM232708 AM232710 AM232712ndashAM232715 EF446975ndashEF446978 EF446980ndashEF446982EF446985ndashEF446987) pseudogenes and identical sequen-ces were removed from the analyses (Fig 1) Theserepresent all the accessible sequences of exon 3 class IMHC available to date for songbirds Alignment gaps weretreated as missing data in all analyses Pairwise distancesbetween sequences of different genera were calculated inBioEdit Sequence Alignment Editor (Hall 1999) using theKimura 2-parameter model The length of the sequencesvaried between species and for the analysis of sequencedistances the sequences were cut to a uniform lengthaccording to the shortest available sequence
Many methods for identifying sites under positive ornegative selection are available and we chose three ofthem First we examined the presence of selection using awidely used likelihood ratio modelling approach inCodeML which is included in the PAML 314 programsuite (Yang 1997) Maximum-likelihood models were usedwhich account for heterogeneity among site partitions usingdifferent ω (=dNdS) parameters for the partitions Themodels applied for the analysis of four alignments (fourpasserine genera) in this study are M7 and M8 (Yang et al2000) Models were compared using likelihood ratio teststatistics calculated as follows 2(LbminusLa) was comparedwith χ2 distribution with PbminusPa degrees of freedom where
La and Lb are log-likelihood values and Pa and Pb are thenumbers of parameters for each of the compared nestedmodels (M8 and M7 respectively) If the alternative modelie M8 suggests the presence of sites with ωgt1 the testcan be considered a test of positive selection (Yang et al2000) Posterior probabilities for site classes were calculat-ed by the Bayes Empirical Bayes (BEB) method in modelM8 If the posterior means of ω for some sites are gt1 thosesites are likely to be under positive selection (Yang et al2005)
The likelihood methods for quantifying selection pres-sure on codon alignments may have high rates of falsepositives if the sequences have undergone recombination(Anisimova et al 2003) therefore nonrecombinant frag-ments were identified using the Genetic Algorithm Recom-bination Detection (GARD Kosakovsky Pond et al 2006)Subsequently the presence of selection on particularcodons was analysed separately for each of them usingtwo methods First the random effects likelihood (REL)approach which involves fitting a distribution of substitu-tion rates across sites and then inferring the rate at whichindividual sites evolve was used Like CodeML thismethod makes inference on selection using an empiricalBayes approach Second the fixed effects likelihood (FEL)method was also applied this approach directly estimatesthe rates of nonsynonymous and synonymous substitutionsat each site (Kosakovsky Pond and Frost 2005) GARDREL and FEL analyses were implemented in the HyPhysoftware package (Kosakovsky Pond et al 2005) and aweb-based interface running on a cluster of computers athttpwwwdatamonkeyorg was employed (KosakovskyPond and Frost 2005)
Analysis of recombination in the Scarlet Rosefinch MHCclass I
Recombination events were recently shown to be frequentinside the MHC domain (either intra-locus or inter-locus)We used three approaches to detect if recombination wasimportant in generating recent diversity of MHC class Isequences in the Scarlet Rosefinch We analysed threecommonly used statistics (r2 Dprime and G4) by permutation inthe program PERMUTE (included in the package OME-GAMAP Wilson and McVean 2006) to test the correlationbetween linkage disequilibrium and physical distance of thesingle-nucleotide polymorphism sites In the absence ofrecombination all sites are linked and there should be zerocorrelation but if recombination has occurred the correla-tion should be significantly negative
The level of recombination rate in the history of the setof 29 aligned sequences was estimated using the Bayesianapproach incorporated in OMEGAMAP (Wilson andMcVean 2006) which is primarily employed to co-
3 4 5 6 7 8
No of exon 3 variants
0
10
20
30
40
50
60
No
of in
divi
dual
s
Fig 1 Distribution of numbers of different exon 3 MHC class Ialleles per individual in a Scarlet Rosefinch population (N=120)
454 Immunogenetics (2009) 61451ndash461
estimate the selection parameter ω and the recombinationrate (ρ=4Nec) along the sequence
Results
MHC class I variation in the Scarlet Rosefinch
Amplification of genomic DNA with primers HN34 andCE-MHCREX3 gave a single PCR product of about 325 bpin all individuals including the whole exon 3 of 270 bp(273 bp in some alleles see below) and encoding thealpha2 domain (corresponding to part of the PBR and smallparts of adjacent introns) Among 120 adult Scarlet Rose-finches we identified 82 different variants of amplifiedMHC class I sequences using high-resolution capillarySSCP The SSCP profiles were highly repeatable andindividuals exhibited between three and nine variants (oneof these variants was impossible to distinguish by SSCPalthough it was repeatedly sequenced [see below]) Theseresults suggest that as many as five MHC class I genes arepresent in the Scarlet Rosefinch and the number ofamplified genes is variable between individuals
Due to the extremely high diversity of MHC class I inthe studied population not all variants were analysed indetail In total we obtained 29 sequences of exon 3 ofMHC class I genes by cloning and sequencing PCRproducts from 18 highly heterozygous individuals (identi-fied by SSCP) Each sequence was provided with thespecies abbreviation and a number (ie CaerU01-CaerU29) In this abbreviation lsquoUrsquo stands for theunknown locus designation of the gene (Mesa et al2004) In three individuals (ID 541 667 and 692) wemanaged to sequence all the variants identified by SSCP ofthese individuals (six four and eight alleles respectively)and all these sequences correspond to exon 3 of MHC classI when BLASTed (httpblastncbinlmnihgovBlastcgi)In total 597 clones were analysed by SSCP 156 of whichcontained either PCR artefacts or were contaminated byPCR product from the individual bird 55 contained noinsert and 122 contained the most frequent sequenceCaerU02 This variant of exon 3 occurred in all analysedadults except one female However the true number ofalleles is probably underestimated because some sequencevariants were difficult to distinguish by SSCP alleleCaerU11 was poorly visualised on SSCP because the6-FAM peak was very low and the NED peak wasoverlapped by CaerU02 (for this reason this allele mighthave been overlooked in some individuals and thereforewe do not have the precise number of individuals in whichthis allele occurred (Fig 2) even if it was repeatedlysequenced in four individuals) Most SSCP patterns werehighly specific in fact we found only one case in which
two different MHC sequences repeatedly confirmed bycloning and sequencing gave identical SSCP peaks incapillary electrophoresis (CaerU18 and CaerU19)
All 29 sequences were obtained from randomly chosenclones whose SSCP pattern matched exactly the SSCPpattern found in the profile of the individual from which theclone was isolated (to avoid sequencing PCR artefacts)Seventeen of these sequences were obtained from oneindividual each (seven of them were only present in oneindividual in the population) the remaining sequences wereidentified in clones originating from at least two differentbirds (two PCRs) These sequences translate into 28 uniqueamino acid sequences (CaerU02 and CaerU19 translateto identical alpha2 domains) and none of them containdeletions insertions or stop codons in the reading frameHowever four sequences have an insertion of 3 bp codingfor isoleucine (Fig 2) The pairwise distances between bothnucleotide and aminoacid sequences of the Scarlet Rose-finch and three other passerine genera (AcrocephalusPasser and Parus) chicken G gallus the Japanese quailC japonica and duck Anas platyrhynchos are shown inTable 1 (see also Fig 2)
Seven amino acid residues in the alpha2 domain offunctional MHC class I molecules are reported to be highlyconserved [Y123 T143 K146 W147 Y159 L160 Y171(Kaufman et al 1994 Shum et al 1999 Mesa et al 2004)]In all our Scarlet Rosefinch sequences four out of seven ofthese residues are conserved (W144W147 Y157Y159L158L160 Y169Y171 Fig 2) Y123 is not conserved inany avian sequence but the phenylalanine present in allRosefinch sequences (F123) is also reported in other non-mammalian species (Shum et al 1999 Mesa et al 2004)Threonine at position T140T143 is replaced by serine in theCaerU23 allele (S140T143) but both these residues belongto the same biochemical group of amino acids (ie theyshould substitute each other in peptide binding) The sameis true for K146 where lysine is replaced by arginine(R143K146) in all alleles (Fig 2)
Positive selection on MHC class I sequences in passerinebirds
Likelihood ratio tests comparing models M8 and M7 in thePAML package identified significant variability in selectivepressure among sites in exon 3 and the presence of two to15 sites under positive selection in four studied passerinespecies (Table 2) GARD analyses identified one significantglobal recombination in alignments of MHC sequencesfrom Carpodacus although no recombination point wasfound in Acrocephalus Parus or Passer REL and FELanalyses which incorporate information of recombinationprovided the evidence that two to 14 and one to eight sitesare under positive selection respectively (Table 2) The
Immunogenetics (2009) 61451ndash461 455
positively selected sites were often identical between thefour compared species as well as between the three methodsof analysis (Fig 3)
Recombination in MHC class I of the Scarlet Rosefinch
Significant negative correlation between physical distanceand linkage disequilibrium was detected using all threeanalyses in the program PERMUTE (plt0001) suggestingthat recombination regularly occurs in exon 3 of MHC classI genes The mean population recombination per codon (ρ=0233) identified in the program OMEGAMAP is approxi-mately half of the mean total population mutation (θ=0534)
again suggesting that recombination is important for gener-ating MHC class I variants in the Scarlet Rosefinch
Discussion
Structure of classical MHC class I genes in passerine birds
In this study the structure and diversity of MHC class I wasstudied in a wild-living songbird species Up to ninedifferent MHC class I sequences were present in oneScarlet Rosefinch individual which means that at least fiveamplified MHC class I genes could occur in a single bird
98 108 118 128 138 148 158 168 178 |||||||||||||||||| CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEG-EAEGWTNYLKHECPEWLQKYVGYGQKELER 1 CaerU12 R-VENVR 2 CaerU29 Y-VENV 1 CaerU05 RLRYP-PRRH 4 CaerU02 LWRLPKKPNSTGKIVRNVSRR 119CaerU17 LWLPKKPNSTGKIVRNVSRRERH 4 CaerU18 LWLPKKPNSTGKIVRNVSRR 32 CaerU08 FIQWLSVRRAFIED-FRR 27 CaerU16 FIQWLSVRRAFIEDK-FRR 1 CaerU22 FIQWLSVRRAFIEDK-FIRE 7 CaerU20 FIQWLSVDARFIP-VKR 19 CaerU06 WRSSEVYYRFIPDVKE-VFRE 51 CaerU07 WRSSEVYYWRFIPDVKE-VFRE 111CaerU11 WRSSEVYYRFIPDVKE-VFRER CaerU04 LWVRHNRFID-FIC 9 CaerU13 LRVRRARFIRDK-MRDV 1 CaerU14 LRVRRARFIRD-MRDV 1 CaerU27 WRVRRARFIDG-VRRDV 1 CaerU28 LRVRRARFIRDKE-VF 1 CaerU03 LNVRYNRFIPD-PRW 4 CaerU09 RNVRYNRFIELDKE-AF 11CaerU10 WRNVRYNRFIPDKE-VFIH 5 CaerU15 WRNVRYNRFIEPD-PRVI 6 CaerU23 LWNVRYNRFIPDSKE-VLH 11CaerU24 WRNVRYRFIPD-PRRVH 4 CaerU25 WRSNVRFYNRFIPDKE-VFDQH 3 CaerU21 LWNVRDARFIDNGKE-VF 7 CaerU26 LRNVRDARFIDNGK-VEMRE 10Gallus gallus SVQWMIEGPRYYQMARFTAKTMTTVPEVPKKES-PRKEETVRREKAG Coturnix japonica SVQWMIDTTRYNQYARFIVKDTMTTVPEVPKKEGD-YRQKHEETVQRRHENKAG Anas platyrhynchos SVQRMKRFEQYERFIALKDTLTTDAQKKETYRTKYENTIRSKDV + + ++ ++ +
Fig 2 Amino acid sequences encoded by exon 3 in the ScarletRosefinch corresponding to alpha2 domain of the protein comparedwith GenBank sequences from chicken (X12780) quail (D29813) andduck (AY294416) Identity with the Scarlet Rosefinch allele CaerU1is indicated by dots gaps which are added to get the best alignment bydashes Highly conserved residues that anchor the peptide-terminalmain-chain atoms are marked by crosses other residues forming
variable part of peptide-binding region in HLA-A and chicken(Wallny et al 2006 Koch et al 2007) are marked by asterisks Thenumbering of amino acid residues is based on the complete MHCclass I protein in the duck (Mesa et al 2004) The numbers on theright show the number of individuals with the particular sequence (N=120) question mark the precise number of individuals bearing theallele CaerU11 is not known (see the text for more details)
Genus Numbera Amino acid sequence distance plusmn SD Nucleotide sequence distance plusmn SD
Acrocephalus 37 0514plusmn0189 0185plusmn0053
Parus 24 0734plusmn0167 0260plusmn00483
Passer 21 0538plusmn0154 0187plusmn0040
Gallus 1 1390plusmn0102 0689plusmn0040
Coturnix 1 1618plusmn0149 0739plusmn0050
Anas 1 1247plusmn0122 0607plusmn0037
Table 1 Average distancesbetween the 29 Scarlet Rose-finch sequences and sequencesof six other bird species
a Number of sequences in theanalysed alignment
456 Immunogenetics (2009) 61451ndash461
Although gene expression analysis was not performedstrong correlation between signatures for balancing selec-tion and expression is suggested Furthermore the presenceof conserved sites that anchor the peptide-terminal main-chain atoms and a reading frame with no terminal codons orframeshift mutations suggest that possibly all sequences ofMHC class I described in this study might be functionalalthough further study at the level of cDNA is required toconfirm this There is an increasing number of studiesshowing that even nearly intact MHC sequences are notexpressed (eg Indovina et al 2001 Bonneaud et al 2004Shiina et al 2006) Hence MHC class I genes seem to behighly variable in the Scarlet Rosefinch and this compli-cated and complex structure is very different from non-
passerines For example the best described MHC in birdsthe chicken lsquominimal essentialrsquo MHC has very few buthighly condensed genes (eg there are two classical MHCclass I genes but only a single dominantly expressedKaufman et al 1999) it appears that this is the ancestralstate for birds since it occurs in various groups such asother gallinaceous birds (Jarvi et al 1999 Wittzell et al1999a Worley et al 2008) owls (Burri et al 2008) andparrots (Hughes et al 2008) Up to now the mostcomplicated MHC class I gene set has been described intwo species of passerines in the great reed warbler sevengenomic class I sequences were found in one individualone of which contained a frameshift mutation and washence nonfunctional (Westerdahl et al 1999) and eight
Table 2 Results of the likelihood ratio tests of positive selection on exon 3 of MHC class I in four genera of passerine birds
Genus Length Numbera M8 vs M7 Likelihood ratio test Number of positively selected sites
df Test statistic Significance PAML REL FEL
Carpodacus 91 29 2 578 plt0001 15 7 8
Acrocephalus 68 37 2 712 plt0001 8 14 4
Parus 69 24 2 472 plt0001 13 14 5
Passer 82 21 2 232 plt0001 2 2 1
Test statistic was computed as 2(LbminusLa) where La and Lb are log-likelihood values for each of the nested models being compared (M8 and M7respectively)
Length length of the aminoacid sequencesa Number of sequences in the analysed alignment
A - PAML 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ + B ndash REL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
C ndash FEL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
Fig 3 Comparison of results of three methods for detecting aminoacid sites under selection in four passerine species The analyses arebased on the alignments of 29 Rosefinch 37 warbler 24 tit and 21sparrow complete or partial sequences of exon 3 encoding alpha2domain of the protein Amino acid residues found to be under positiveselection by three methods (A PAML B REL C FEL) are bold (fordetails on the methods see the text) If the same residue was found tobe positively selected in at least two genera it is marked by rectangle
Regions excluded from analysis are marked by a squiggle Highlyconserved residues that anchor the peptide-terminal main-chain atomsare marked by crosses other residues forming variable part of peptide-binding region in HLA-A and chicken (Wallny et al 2006 Koch et al2007) are marked by asterisks The numbering of amino acid residuesis based on the complete MHC class I protein in the duck (Mesa et al2004) (Acrocephalus-AF449690) (Parus-AM232710) and (Passer-AY285012)
Immunogenetics (2009) 61451ndash461 457
different cDNA sequences were also found in an individualIn a separate study of the same species 20 genomicsequences of exon 3 of class I were found in fiveindividuals five of which were pseudogenes (Westerdahlet al 2004) Similar results were found in house sparrowswhere 20 sequences were verified in seven individualsthree of which were proved to be transcribed althoughthere were no pseudogenes (Bonneaud et al 2004) Ourresults concerning individual exon 3 variation are similar tothe ones mentioned above suggesting that complex MHCstructure could be a general pattern in songbirds
Evolution of MHC class I sequences in passerines
Several mechanisms of molecular evolution have beenalready documented in the avian MHC sequences includ-ing positive selection (Alcaide et al 2007 Hughes et al2008) concerted evolution (Wittzell et al 1999b) andtrans-species polymorphism (Richardson and Westerdahl2003 Alcaide et al 2007) In the present study model-based tests of selection (used here for the first time inpasserine species) have demonstrated the presence ofpositively selected sites in exon 3 of MHC class I in fourpasserine genera This part of the gene encodes the alpha 2domain of the protein which is included in forming thepeptide-binding groove Positive selection acting on somecodons of exon 3 is in accordance with published data onMHC class I in other birds where the ratio of non-synonymous to synonymous substitutions (dNdS) wasusually slightly higher at PBR sites than at non-PBR ones(Westerdahl et al 1999 Richardson and Westerdahl 2003Bonneaud et al 2004) The differences however were notsignificant and this could be caused either (1) by in-correctly identified PBR sites since the MHC proteinstructure in the majority of species studied is not well-understood In fact the PBR sites have usually beenidentified on the basis of the three-dimensional structureof human HLA-A protein (Bjorkman et al 1987) A modelof the three-dimensional structure of a passerine MHC classI molecule is highly desirable Or (2) another reason mightbe the fact that some of the studied sequences come fromloci that lost their importance and they are not underselection that is why it is crucial to identify sequences toparticular loci in the future studies (see Worley et al 2008)
Our results indicate extreme variability in selectivepressure among sites in the exon 3 and the presence of anumber of sites under balancing selection Approximatelyhalf of the positively selected sites in passerines (Fig 3)have been previously assumed to play a role in antigenbinding based on the structural models (Wallny et al 2006Koch et al 2007) However we found at least twoadditional sites (amino acid residues 126 and 136) understrong positive selection in several genera which have not
been previously reported for PBR (either in human HLA orchicken BF) The maintenance of considerable nonsynon-ymous variation at these specific codons by positiveselection suggests that such polymorphism may havefunctional consequences for peptide binding and presenta-tion to T cells (see similar recent results concerning teleostfish MHC molecules Aguilar and Garza 2007 or inmammals Amills et al 2008) The predictive power ofcodon-based models has not yet been tested in MHC andthe structural model describing peptide-binding groove inpasserine birds is still required to confirm the functionalrole of amino acid residues under different types ofselection
It appears that mammalian and avian MHCs may differin frequency of genetic exchange between different genesof multigene families Concerted evolution where the inter-locus gene conversion events are frequent in highlyduplicated MHC genes may be the predominant evolu-tionary mechanism in birds (Edwards et al 1995 Alcaide etal 2007 Garrigan and Edwards 1999 Wittzell et al 1999bHess and Edwards 2002) and probably also in fish (Reuschet al 2004 van Oosterhout et al 2006 Aguilar and Garza2007) and reptiles (Miller et al 2006) but not in mammals(Edwards et al 1997) This process creates high homologyamong MHC sequences derived from various loci and itbecomes impossible to assign different MHC alleles tospecific loci In birds so far most of the studies evidencinginter-locus recombination were conducted on MHC class IIConcerning MHC class I it has only been studied in thechicken where no evidence was found for concertedevolution to occur (Shaw et al 2007) Thus our resultson recombination analyses in the finches (ie the totalamount of recombination per codon was half of that ofsubstitution and we failed to assign the sequences tospecific loci) are in agreement with previous reports andfurthermore they suggest that concerted evolution may befrequent also in MHC class I genes in passerine birds Toconfirm this hypothesis the more detailed description ofMHC structure (especially the affiliation of particularalleles to individual loci in a population and the geneticvariation outside exon 3) is necessary
Extreme MHC class I variation in a single population
Previous studies on songbirds using the method ofdenaturing gradient gel electrophoresis or restriction frag-ment length polymorphism (RFLP) have also indicatedhigh sequence variation at exon 3 of MHC class I forexample in an outbred population of house sparrows 46sequences were found in 101 individuals or between oneand 11 alleles per individual (Bonneaud et al 2006a) in thegreat reed warbler 49 different RFLP genotypes werefound in 55 birds (Westerdahl et al 2004) Even so in the
458 Immunogenetics (2009) 61451ndash461
Scarlet Rosefinch 82 alleles were found in a singlepopulation and most of the birds displayed uniquegenotypes which is the highest exon 3 MHC class Ivariability observed thus far This result may still be anunderestimate considering that in at least one case theSSCP pattern was identical for two different allelesAlthough we were not able to sequence all the allelesdisplayed on SSCP it is very probable that all the peaksidentified by SSCP are true alleles and that there could beadditional ones (due to peak overlapping) Another reasonwe believe this variation has not been overestimated is thatan extraordinarily high variability of selectively neutralmarkers has been found in this species at two microsatelliteloci more than 70 alleles were found (Albrecht et al 2007)
Interestingly one of the alleles (CaerU02) was possessedby all individuals but one female (no 515) investigated inthis study From parentndashoffspring genotypes (Promerovaacute etal unpublished data) we know that also some of thisfemales offspring missed this allele thus this absence issurely heritable We conclude that CaerU02 is the mostfrequent allele on a locus with very low variability Thefemale no 515 is either homo- or heterozygote for other veryrare allele(s) or the locus (at least the copy with alleleCaerU02) is completely missing in her MHC haplotype
Our optimization of a genotyping method for analyses ofMHC class I variation will allow us to search for answers tofundamental questions concerning recent immunoecologysuch as the role of MHC in disease resistance expression ofsecondary ornaments and mating success (eg Bonneaud etal 2006a b Loiseau et al 2008 Richardson et al 2004Westerdahl et al 2005) Furthermore our data could beused for future comparative studies of the correlationbetween MHC diversity and migratory strategies in birdsWe presume that migratory birds such as the ScarletRosefinch (which migrates from the Czech Republic toIndia ca 5000 km) might encounter a larger variety ofpathogens during their lifetime than sedentary birds (forexample there is evidence that migrating individualsencounter more malarial parasites Peacuterez-Tris and Bensch2005) In that case more diverse immune mechanismscould help them fight these pathogens Moreover it hasalready been suggested that migratory species have highergenetic variability than sedentary species (Fitzpatrick 1994)and also that the higher genetic variability of migrants hasan impact on higher occurrence of extra-pair paternity(Spottiswoode and Moslashller 2004) The hypothesis thatmigration in birds might elicit a higher variability onMHC was first evaluated by Shiina et al (2004) whocompared the MHC of migratory quail with that ofsedentary chicken The quail has a more complicatedMHC than the chicken the class I region in the quail istwice as long as that of the chicken (Shiina et al 2004) andthere are five expressed class I genes although not all of
them are classical ones (Shiina et al 2006) SimilarlyRichardson and Westerdahl (2003) reported a difference inMHC variability between the migratory Great reed warblerand the sedentary inbred Seychelles warbler In this casehowever the low MHC variation in the latter could be dueto a severe bottleneck leading to strong decrease of overallgenetic variation Furthermore the endemic Basra reedwarbler (Acrocephalus griseldis) is as variable at MHC asthe Great reed warbler even though it is sedentary (Hanssonand Richardson 2005) High level of MHC polymorphismwas found in the house sparrow which is a non-migratoryspecies (Bonneaud et al 2004) which also contradicts thepredictions of the above hypothesis Unfortunately formost species (including the Scarlet Rosefinch) there is noprecise information about the functional variation ienumber of expressed MHC variants It is evident that dataon real MHC variation in more species are inevitablyrequired to perform a rigorous comparative meta-analysisand to test the predictions of migration-MHC hypothesis
Acknowledgements We thank Pavel Munclinger Michal VinklerJan Schnitzer and Radka Polaacutekovaacute for their help in the field Thanksto Anna Bryjovaacute and Dagmar Čiacutežkovaacute for their valuable advice inthe laboratory We are very grateful to Heidi C Hauffe for improvingthe English and two anonymous referees for critical comments on themanuscript This study was funded by the Grant Agency of theAcademy of Sciences of the Czech Republic (project noIAA600930608) the Grant Agency of the Czech Rep (project no206060851) and by the Ministry of Education of the Czech Repwhose Research Centre no LC06073 and Long-term Research Plan(no MSM 0021620828) to the Charles University in Prague formed aframework for a part of this study
References
Abbas AK Lichtman AH Pober JS (1994) Cellular and molecularimmunology W B Saunders Company Philadelphia
Aguilar A Garza JC (2007) Patterns of historical balancing selectionon the salmonid major histocompatibility complex class II betagene J Mol Evol 6534ndash43 doi101007s00239-006-0222-8
Albrecht T (2004) Edge effect in wetland-arable land boundarydetermines nesting success of scarlet rosefinches Carpodacuserythrinus in the Czech Republic Auk 121361ndash371doi1016420004-8038(2004)121[0361EEIWLB]20CO2
Albrecht T Schnitzer J Kreisinger J Exnerovaacute A Bryja JMunclinger P (2007) Extrapair paternity and the opportunity forsexual selection in long-distant migratory passerines Behav Ecol18477ndash486 doi101093behecoarm001
Alcaide M Edwards SV Negro JJ (2007) Characterization polymor-phism and evolution ofMHCClass II B genes in birds of prey JMolEvol 65541ndash554 doi101007s00239-007-9033-9
Amills M Ramiacuterez O Tomaacutes A Obexer-Ruff G Vidal O (2008)Positive selection on mammalian MHC-DQ genes revisited from amultispecies perspective Genes Immun 9651ndash658 doi101038gene200862
Anisimova M Nielsen R Yang Z (2003) Effect of recombination onthe accuracy of the likelihood method for detecting positiveselection at amino acid sites Genetics 1641229ndash1236
Immunogenetics (2009) 61451ndash461 459
Bjorkman PJ Saper MA Samraoui B Bennett WS Strominger JLWiley DC (1987) Structure of human class I histocompatibil-ity antigen HLA-A2 Nature 329506ndash512 doi101038329506a0
Bonneaud C Sorci G Morin V Westerdahl H Zoorob R Wittzell H(2004) Diversity of MHC class I and IIB genes in house sparrows(Passer domesticus) Immunogenetics 55855ndash865 doi101007s00251-004-0648-3
Bonneaud C Chastel O Federici P Westerdahl H Sorci G (2006a)Complex MHC-based mate choice in a wild passerine Proc RSoc 2731111ndash1116 doi101098rspb20053325
Bonneaud C Peacuterez-Tris J Federici P Chastel O Sorci G (2006b) Majorhistocompatibility alleles associated with local resistance to malariain a passerine Evolution Int J Org Evolution 60383ndash389
Bryja J Galan M Charbonnel N Cosson J-F (2005) Analysis ofmajor histocompatibility complex class II gene in water volesusing capillary electrophoresis-single stranded conformationpolymorphism Mol Ecol Notes 5173ndash176 doi101111j1471-8286200400855x
Burri R Niculita-Hirzel H Roulin A Fumagalli L (2008) Isolation andcharacterization of major histocompatibility complex (MHC) class IIB genes in teh Barn owl (Aves Tyto alba) Immunogenetics 60543ndash550 doi101007s00251-008-0308-0
Cramp S Simmons KEL Perrins CM (eds) (1977-1994) The birds ofthe Western Palearctic Oxford University Press Oxford
Edwards SV Wakeland EK Potts WK (1995) Contrasting histories ofavian and mammalian MHC genes revealed by class II B genesof songbirds Proc Natl Acad Sci U S A 9212200ndash12204doi101073pnas922612200
Edwards SV Chesnut K Satta Y Wakeland EK (1997) Ancestralpolymorphism of Mhc Class II genes in mice implications forbalancing selection and the mammalian molecular clock Genetics146655ndash668
Fitzpatrick S (1994) Colourful migratory birds evidence for amechanism other than parasite resistance for the maintenance oflsquogood genesrsquo sexual selection Proc R Soc Lond B Biol Sci257155ndash166 doi101098rspb19940109
Freeman-Gallant CR Johnson EM Saponara F Stanger M (2002)Variation at the major histocompatibility complex in Savannahsparrows Mol Ecol 111125ndash1130 doi101046j1365-294X200201508x
Garrigan D Edwards SV (1999) Polymorphism across an exon-intronboundary in an avian Mhc class II B gene Mol Biol Evol 161599ndash1606
Gouumly de Bellocq J Charbonnel N Morand S (2008) Coevolutionaryrelationship between helminth diversity and Mhc class II polymor-phism in rodents J Evol Biol 211144ndash1150 doi101111j1420-9101200801538x
Hall TA (1999) BioEdit a user-friendly biological sequence alignmenteditor and analisis program for Windows 9598NT NucleicAcids Symp Ser 4195ndash98
Hansson B Richardson DS (2005) Genetic variation in twoendangered Acrocephalus species compared to a widespreadcongener estimates based on functional and random loci AnimConserv 883ndash90 doi101017S1367943004001878
Hess CM Edwards SV (2002) The evolution of major histocompat-ibility genes in birds Bioscience 52423ndash431 doi1016410006-3568(2002)052[0423TEOTMH]20CO2
Hess CM Gasper J Hoekstra H Hill C Edwards SV (2000) MHCclass II pseudogene and genomic signature of a 32-kb cosmid inthe House Finch (Carpodacus mexicanus) Genome Res 1013ndash23 doi101101gr105613
Hosomichi K Shiina T Suzuki S Tanaka M Shimizu S Iwamoto SHara H Yoshida Y Kulski JK Inoko H Hanzawa K (2006) Themajor histocompatibility complex (Mhc) class IIB region hasgreater genomic structural flexibility and diversity in the quail
than the chicken BMC Genomics 7322ndash335 doi1011861471-2164-7-322
Hughes AL Yeager M (1998) Natural selection at major histocom-patibility complex loci of vertebrates Annu Rev Genet 32415ndash435 doi101146annurevgenet321415
Hughes CR Miles S Walbroehl JM (2008) Support for the minimalessential MHC hypothesis a parrot with a single highlypolymorphic MHC class II B gene Immunogenetics 60219ndash231 doi101007s00251-008-0287-1
Indovina P Megiorni F Fontemaggi G Coni P Mora B Mazzilli MC(2001) Absence of in vivo DNA-Protein interactions in theDQA2 and DQB2 promoter regions Hum Immunol 62504ndash508doi101016S0198-8859(01)00236-1
Jarvi SI Goto RM Gee GF Briles WE Miller MM (1999)Identification inheritance and linkage of B-G-like and MHCclass I genes in cranes Am Genet Assoc 90152ndash159
Kaufman J (1999) Co-evolving genes in MHC haplotypes the lsquorulersquofor nonmammalian vertebrates Immunogenetics 50228ndash236doi101007s002510050597
Kaufman J Salamonsen J Flajnik M (1994) Evolutionary conserva-tion of MHC class I and class II molecules-different yet the sameSemin Immunol 6411ndash424 doi101006smim19941050
Kaufman J Voumllk H Wallny H (1995) A bdquominimal essential MHCldquoand an bdquounrecognized MHCldquo two extremes in selection forpolymorphism Immunol Rev 14363ndash88 doi101111j1600-065X1995tb00670x
Kaufman J Jacob J Shaw I Walker B Milne S Beck S SalomonsenJ (1999) Gene organisation determines evolution of function inthe chicken MHC Immunol Rev 167101ndash117 doi101111j1600-065X1999tb01385x
Klein J (1986) Natural history of the major histocompatibilitycomplex Wiley New York
Klein J Sato A Nagl S Oh Uigin C (1998) Molecular trans-speciespolymorphism Annu Rev Ecol Syst 291ndash21 doi101146annurevecolsys2911
Koch M Camp S Collen T Avila D Salomonsen J Wallny HJ vanHateren A Hunt L Jacob JP Johnston F Marston DA Shaw IDunbar PR Cerundolo V Jones EY Kaufman J (2007)Structures of an MHC class I molecule from B21 chickensillustrate promiscuous peptide binding Immunity 27885ndash899doi101016jimmuni200711007
Kosakovsky Pond SL Frost SDW (2005) Not so different after all acomparison of methods for detecting amino acid sites under selectionMol Biol Evol 221208ndash1222 doi101093molbevmsi105
Kosakovsky Pond SL Frost SDW Muse SV (2005) HyPhyhypothesis testing using phylogenies Bioinformatics 21676ndash679 doi101093bioinformaticsbti079
Kosakovsky Pond SL Posada D Gravenor MB Woelk CH FrostSDW (2006) Automated phylogenetic detection of recombinationusing a genetic algorithm Mol Biol Evol 231891ndash1901doi101093molbevmsl051
Loiseau C Zoorob R Garnier S Birard J Federici P Julliard R SorciG (2008) Antagonistic effects of a MHC class I allele on malaria-infected house sparrows Ecol Lett 11258ndash265 doi101111j1461-0248200701141x
Mesa CM Thulien KJ Moon DA Veniamin SM Magor KE (2004)The dominant MHC class I gene is adjacent to the polymorphicTAP2 gene in the duck Anas platyrhynchos Immunogenetics56192ndash203 doi101007s00251-004-0672-3
Miller HC Lambert DM (2004) Gene duplication and geneconversion in class II MHC genes of New Zealand robins(Petroicidae) Immunogenetics 56178ndash191
Miller MM Goto RM Bernot A Zoorob R Auffray C Bumstead NBriles WE (1994) Two Mhc class I and two Mhc class II genes mapto the chicken Rfp-Y system outside the B complex Proc Natl AcadSci U S A 914397ndash4401 doi101073pnas91104397
460 Immunogenetics (2009) 61451ndash461
Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461

groups of birds such as parrots (Psittaciformes Hughes etal 2008) and owls (Strigiformes Burri et al 2008)However the MHC of some other galliform species suchas the quail (Coturnix japonica) has a larger number ofduplicated genes and the overall structure is much morecomplex (Hosomichi et al 2006 Shiina et al 2004) Aneven more complex MHC structure has been reported forseveral passerine species with many gene copies andabundant pseudogenes (eg Westerdahl et al 1999 2000Hess et al 2000 Freeman-Gallant et al 2002 Miller andLambert 2004 Bonneaud et al 2006a)
Most ecological studies of MHC in birds are traditionallybased on MHC class II while MHC class I genes are oftendisregarded There are only very limited data on MHC classI structure of passerine birds in fact they have only beendescribed in two species In the great reed warbler(Acrocephalus arundinaceus) Westerdahl et al (1999)found four transcribed MHC class I loci in one individualand in the house sparrow (Passer domesticus) there seemto be at least three (Bonneaud et al 2004) to six (Bonneaudet al 2006a) MHC class I loci per individual Despite thefact that MHC class I genes are used only rarely asimmunogenetic markers they might be very important forunderstanding the pathogenndashhost relationships in wildpopulations For example recent studies in wild-livingsongbird species have shown that there may be significantassociations of certain MHC class I variants with theresistance or susceptibility to some parasites (Westerdahl etal 2005 Bonneaud et al 2006b Loiseau et al 2008)
The peptides derived from pathogen fragments arebound to MHC proteins in the so-called peptide-bindingregion (PBR) which is encoded by codons of exon 2 andexon 3 of the MHC class I genes (Wallny et al 2006 Kochet al 2007) These sites are believed to be under strongpositive selection and very variable in order to pursue thearms race with fast-evolving pathogens On the other handthere are sites that are conserved and lack variation sincethey are essential for anchoring the peptide-terminal main-chain atoms (Kaufman et al 1994 Shum et al 1999 Mesaet al 2004) Because many studies use MHC genes asgenetic markers it is important to understand theirevolutionary history revealing the mechanisms maintaininghigh polymorphism may help to achieve this goal Forexample previous molecular studies have disclosed theexistence of trans-species polymorphism (occurrence ofsimilar alleles across different species originating from acommon ancestor) at the MHC (Klein et al 1998) whichsupports the hypothesis of balancing selection Anotherexplanation of the extraordinarily high variation at theMHC may be gene conversion which can produce newsequences or cause concerted evolution when duplicatedgenes evolve in parallel this particular mechanism seems tobe common in birds (eg Wittzell et al 1999b)
Although knowledge regarding the structure and func-tion of the avian MHC has increased significantly in the lastdecade there are still questions that remain unresolved Acomparative approach could help to understand the evolu-tionary mechanisms and ecological importance of MHC inwild populations (see eg Gouumly de Bellocq et al 2008)however before specific hypotheses can be tested theMHC structure in a wide array of species still needs to bestudied because genetic variation especially in MHC class Igenes has only been described in a few wild birds (Jarvi etal 1999 Westerdahl et al 1999 Bonneaud et al 2004Mesa et al 2004 Xia et al 2005) Here we describe thestructure and extremely high intra-population diversity ofexon 3 of MHC class I genes in a long-distance migratorysongbird with pronounced sexual dimorphism the ScarletRosefinch (Carpodacus erythrinus) The study populationnests in central Europe and winters in southern Asiaparticularly in India (Cramp et al 1994) thus these birdstravel thousands of kilometres and are potentially exposedto a wide range of pathogens and parasites Furthermoreusing state-of-the-art methods of sequence analyses weexamine the role of selection and recombination in formingthe diversity of MHC class I in rosefinches and otherpasserine species where data on MHC class I structure areavailable
Materials and methods
Study population sampling and DNA extraction
The Scarlet Rosefinch is a socially monogamous migratorypasserine with a distribution that has been spreadingwestward from Russia in recent years (Cramp et al 1994)The population occupying the Šumava Mountains NationalPark Czech Republic (48deg49prime N 13deg56prime E sim750 m asl)has been studied since 1995 (Albrecht 2004 Polaacutekovaacute et al2007 Albrecht et al 2007) The present study uses samplesfrom 66 adult males and 54 adult females captured at thestudy site during the breeding season in the years 2000ndash2007 Most individuals were captured soon after theirarrival at the study site using mist nets although some birdswere caught later during the chick provisioning stage(Albrecht et al 2007) Blood (approximately 30 μl) wassampled by wing venipuncture and stored in ethanol atminus20degC until DNA extraction Genomic DNA was extractedfrom blood using the Dneasy Blood amp Tissue Kit (Qiagen)according to the manufacturers instructions
Primer design for amplifying genomic exon 3 sequences
We amplified the complete exon 3 of MHC class Itogether with portions of adjacent introns (550 bp) using
452 Immunogenetics (2009) 61451ndash461
the primers HN34 (5prime-CCATGGGTCTCTGTGGGTA-3prime)and HN45 (5prime-CCATGGAATTCCCACAGGAA-3prime) takenfrom Westerdahl et al (2004) using the polymerase chainreaction (PCR) with the following conditions 15 mMMgCl2 04 μM of each primer 02 mM dNTPs 1 UμlTaq polymerase (Fermentas) in appropriate 1times PCR bufferand 1 μl of extracted DNA Deionised water was added toa 10-μl reaction volume The amplification consisted of aninitial denaturation at 94degC (3 min) followed by 30 cyclesof denaturation at 94degC (40 s) annealing at 60degC (30 s)and extension at 72degC (1 min) with a final extension stepat 72degC (10 min) The reactions were run on a Master-cycler ep (Eppendorf) The PCR products were purifiedusing the MinElute PCR purification kit (Qiagen) ligatedto the vector and transformed to bacteria using the pGEM-T Easy Cloning Kit (Promega) according to manufac-turers protocols Positive transformants containing insertsof appropriate length were identified by PCR screeningusing primers on the vector and agarose gel electrophore-sis We randomly chose eight clones extracted plasmidDNA using the QIAprep Spin Miniprep Kit (Qiagen) andsequenced the inserts using the BigDye Terminators Se-quencing Kit v11 and an ABI PRISM 3130 GeneticAnalyzer (Applied Biosystems) Based on these eightsequences a new reverse primer specific for the ScarletRosefinch was designed (CE-MHCREX3 5prime-ACAGGAATTCTGCTCCCACC-3prime) that together with forwardprimer HN34 amplifies the complete exon 3 of the MHCclass I genes of the Scarlet Rosefinch under almost the samePCR conditions as described above (the only difference isthe annealing step which changes to 30 s at 59degC)
Capillary electrophoresis single-strand conformationpolymorphism
Exon 3 of MHC class I was amplified by PCR usingfluorescently labelled primers (HN34 by 6-FAM and CE-MHCREX3 by NED) and conditions described abovePositive PCR amplifications were checked by agarose gelelectrophoresis and then single-strand conformation poly-morphism (SSCP) analysis (a method able to detect even asingle nucleotide substitution) was performed using capil-lary electrophoresis on an ABI PRISM 3130 GeneticAnalyzer (Applied Biosystems) We mixed 2 μl of PCRproduct with 05 μl of GeneScan-500 LIZtrade Size standard(Applied Biosystems) and 12 μl Hi-Ditrade Formamide anddenaturated the mix at 95degC for 3 min After denaturationsamples were immediately put on ice for at least 2 min andelectrophoresis was run at 22degC in 5 non-denaturingconformation analysis polymer (prepared from 9 CAPpolymer Applied Biosystems according to manufacturersinstructions) on an ABI PRISM 3130 Data were analysedusing the software GeneMapper v37 (Applied Biosystems)
and user-defined size standard files for the alignment ofdifferent runs The alleles were identified by both theposition and the shape of peaks For each allele one 6-FAMand one NED peak was identified It is important to notethat neither SSCP peaks nor sequences can be assigned tospecific loci but for simplification in this study they aretermed lsquoallelesrsquo To obtain the sequences of particularalleles (ie SSCP peaks) we cloned the PCR products(obtained by non-labelled primers HN34 and CE-MHCREX3) using the techniques as described aboveSSCP was subsequently used for the identification ofinserted sequences after cloning This procedure allowedus to (1) minimise cost by avoiding the sequencing of thesame sequence in many clones and (2) compare SSCPpatterns of clones with those obtained from individuals toidentify PCR artefacts which are common when analysingMHC genes with a cloning and sequencing approach (Bryjaet al 2005) Inserts providing the SSCP patterns thatmatched the SSCP profile of an individual bird ie theinserts contained one of the exon 3 variants of theindividual were re-amplified by specific PCRs and se-quenced using Big Dye kits as described above
Analyses of MHC class I sequence variation
The sequences of MHC class I derived from C erythrinuswere edited in SeqScape v25 (Applied Biosystems) andaligned in BioEdit Sequence Alignment Editor (Hall 1999)To determine whether all sequences represent functionalclassical class I alleles the sequences were examined (1)for the presence of insertions or deletions (lsquoindelsrsquo) causingshift of the reading frame andor stop codons and (2) forthe conservation of residues that anchor the peptide-terminal main-chain atoms Possible codons encoding theamino acid residues included in the peptide binding wereidentified on the basis of homology between chicken MHCand human leukocyte antigen (HLA)-A (eg Wallny et al2006) and by analysing the detailed structural models ofpeptide-binding grooves of chicken MHC class I proteins(Koch et al 2007) Some of the residues are generallyhighly conserved throughout evolution in classical class Imolecules (Kaufman et al 1994 Mesa et al 2004) whileothers are thought to be under strong positive selection andallow interaction with a wide variety of antigens Sequencesof chicken (GenBank acc no X12780) Japanese quail(D29813) and duck (AY294416) were used to compareMHC sequences of Scarlet Rosefinches with those ofdistantly related bird species
Detecting amino acid sites under selection
To detect selection acting on MHC class I gene variationsequences (partial or complete) of exon 3 MHC class I for
Immunogenetics (2009) 61451ndash461 453
three additional genera of passerines were downloadedfrom GenBank (accession nos in parentheses) theseinclude two species of warbler of the genus Acrocephalus(AF449688ndashAF449701 AJ005503 AJ005504 AJ005506AJ005508ndashAJ005510 AJ005512 AJ005514ndashAJ005517AY306008 AY306009 AJ557874ndashAJ557883) the housesparrow (AY284994ndashAY285013 EF429132) and threespecies of tit of the genus Parus (AF346821 AF346823AF346825 AF346827 AF346829ndashAF346832 AM232707 AM232708 AM232710 AM232712ndashAM232715 EF446975ndashEF446978 EF446980ndashEF446982EF446985ndashEF446987) pseudogenes and identical sequen-ces were removed from the analyses (Fig 1) Theserepresent all the accessible sequences of exon 3 class IMHC available to date for songbirds Alignment gaps weretreated as missing data in all analyses Pairwise distancesbetween sequences of different genera were calculated inBioEdit Sequence Alignment Editor (Hall 1999) using theKimura 2-parameter model The length of the sequencesvaried between species and for the analysis of sequencedistances the sequences were cut to a uniform lengthaccording to the shortest available sequence
Many methods for identifying sites under positive ornegative selection are available and we chose three ofthem First we examined the presence of selection using awidely used likelihood ratio modelling approach inCodeML which is included in the PAML 314 programsuite (Yang 1997) Maximum-likelihood models were usedwhich account for heterogeneity among site partitions usingdifferent ω (=dNdS) parameters for the partitions Themodels applied for the analysis of four alignments (fourpasserine genera) in this study are M7 and M8 (Yang et al2000) Models were compared using likelihood ratio teststatistics calculated as follows 2(LbminusLa) was comparedwith χ2 distribution with PbminusPa degrees of freedom where
La and Lb are log-likelihood values and Pa and Pb are thenumbers of parameters for each of the compared nestedmodels (M8 and M7 respectively) If the alternative modelie M8 suggests the presence of sites with ωgt1 the testcan be considered a test of positive selection (Yang et al2000) Posterior probabilities for site classes were calculat-ed by the Bayes Empirical Bayes (BEB) method in modelM8 If the posterior means of ω for some sites are gt1 thosesites are likely to be under positive selection (Yang et al2005)
The likelihood methods for quantifying selection pres-sure on codon alignments may have high rates of falsepositives if the sequences have undergone recombination(Anisimova et al 2003) therefore nonrecombinant frag-ments were identified using the Genetic Algorithm Recom-bination Detection (GARD Kosakovsky Pond et al 2006)Subsequently the presence of selection on particularcodons was analysed separately for each of them usingtwo methods First the random effects likelihood (REL)approach which involves fitting a distribution of substitu-tion rates across sites and then inferring the rate at whichindividual sites evolve was used Like CodeML thismethod makes inference on selection using an empiricalBayes approach Second the fixed effects likelihood (FEL)method was also applied this approach directly estimatesthe rates of nonsynonymous and synonymous substitutionsat each site (Kosakovsky Pond and Frost 2005) GARDREL and FEL analyses were implemented in the HyPhysoftware package (Kosakovsky Pond et al 2005) and aweb-based interface running on a cluster of computers athttpwwwdatamonkeyorg was employed (KosakovskyPond and Frost 2005)
Analysis of recombination in the Scarlet Rosefinch MHCclass I
Recombination events were recently shown to be frequentinside the MHC domain (either intra-locus or inter-locus)We used three approaches to detect if recombination wasimportant in generating recent diversity of MHC class Isequences in the Scarlet Rosefinch We analysed threecommonly used statistics (r2 Dprime and G4) by permutation inthe program PERMUTE (included in the package OME-GAMAP Wilson and McVean 2006) to test the correlationbetween linkage disequilibrium and physical distance of thesingle-nucleotide polymorphism sites In the absence ofrecombination all sites are linked and there should be zerocorrelation but if recombination has occurred the correla-tion should be significantly negative
The level of recombination rate in the history of the setof 29 aligned sequences was estimated using the Bayesianapproach incorporated in OMEGAMAP (Wilson andMcVean 2006) which is primarily employed to co-
3 4 5 6 7 8
No of exon 3 variants
0
10
20
30
40
50
60
No
of in
divi
dual
s
Fig 1 Distribution of numbers of different exon 3 MHC class Ialleles per individual in a Scarlet Rosefinch population (N=120)
454 Immunogenetics (2009) 61451ndash461
estimate the selection parameter ω and the recombinationrate (ρ=4Nec) along the sequence
Results
MHC class I variation in the Scarlet Rosefinch
Amplification of genomic DNA with primers HN34 andCE-MHCREX3 gave a single PCR product of about 325 bpin all individuals including the whole exon 3 of 270 bp(273 bp in some alleles see below) and encoding thealpha2 domain (corresponding to part of the PBR and smallparts of adjacent introns) Among 120 adult Scarlet Rose-finches we identified 82 different variants of amplifiedMHC class I sequences using high-resolution capillarySSCP The SSCP profiles were highly repeatable andindividuals exhibited between three and nine variants (oneof these variants was impossible to distinguish by SSCPalthough it was repeatedly sequenced [see below]) Theseresults suggest that as many as five MHC class I genes arepresent in the Scarlet Rosefinch and the number ofamplified genes is variable between individuals
Due to the extremely high diversity of MHC class I inthe studied population not all variants were analysed indetail In total we obtained 29 sequences of exon 3 ofMHC class I genes by cloning and sequencing PCRproducts from 18 highly heterozygous individuals (identi-fied by SSCP) Each sequence was provided with thespecies abbreviation and a number (ie CaerU01-CaerU29) In this abbreviation lsquoUrsquo stands for theunknown locus designation of the gene (Mesa et al2004) In three individuals (ID 541 667 and 692) wemanaged to sequence all the variants identified by SSCP ofthese individuals (six four and eight alleles respectively)and all these sequences correspond to exon 3 of MHC classI when BLASTed (httpblastncbinlmnihgovBlastcgi)In total 597 clones were analysed by SSCP 156 of whichcontained either PCR artefacts or were contaminated byPCR product from the individual bird 55 contained noinsert and 122 contained the most frequent sequenceCaerU02 This variant of exon 3 occurred in all analysedadults except one female However the true number ofalleles is probably underestimated because some sequencevariants were difficult to distinguish by SSCP alleleCaerU11 was poorly visualised on SSCP because the6-FAM peak was very low and the NED peak wasoverlapped by CaerU02 (for this reason this allele mighthave been overlooked in some individuals and thereforewe do not have the precise number of individuals in whichthis allele occurred (Fig 2) even if it was repeatedlysequenced in four individuals) Most SSCP patterns werehighly specific in fact we found only one case in which
two different MHC sequences repeatedly confirmed bycloning and sequencing gave identical SSCP peaks incapillary electrophoresis (CaerU18 and CaerU19)
All 29 sequences were obtained from randomly chosenclones whose SSCP pattern matched exactly the SSCPpattern found in the profile of the individual from which theclone was isolated (to avoid sequencing PCR artefacts)Seventeen of these sequences were obtained from oneindividual each (seven of them were only present in oneindividual in the population) the remaining sequences wereidentified in clones originating from at least two differentbirds (two PCRs) These sequences translate into 28 uniqueamino acid sequences (CaerU02 and CaerU19 translateto identical alpha2 domains) and none of them containdeletions insertions or stop codons in the reading frameHowever four sequences have an insertion of 3 bp codingfor isoleucine (Fig 2) The pairwise distances between bothnucleotide and aminoacid sequences of the Scarlet Rose-finch and three other passerine genera (AcrocephalusPasser and Parus) chicken G gallus the Japanese quailC japonica and duck Anas platyrhynchos are shown inTable 1 (see also Fig 2)
Seven amino acid residues in the alpha2 domain offunctional MHC class I molecules are reported to be highlyconserved [Y123 T143 K146 W147 Y159 L160 Y171(Kaufman et al 1994 Shum et al 1999 Mesa et al 2004)]In all our Scarlet Rosefinch sequences four out of seven ofthese residues are conserved (W144W147 Y157Y159L158L160 Y169Y171 Fig 2) Y123 is not conserved inany avian sequence but the phenylalanine present in allRosefinch sequences (F123) is also reported in other non-mammalian species (Shum et al 1999 Mesa et al 2004)Threonine at position T140T143 is replaced by serine in theCaerU23 allele (S140T143) but both these residues belongto the same biochemical group of amino acids (ie theyshould substitute each other in peptide binding) The sameis true for K146 where lysine is replaced by arginine(R143K146) in all alleles (Fig 2)
Positive selection on MHC class I sequences in passerinebirds
Likelihood ratio tests comparing models M8 and M7 in thePAML package identified significant variability in selectivepressure among sites in exon 3 and the presence of two to15 sites under positive selection in four studied passerinespecies (Table 2) GARD analyses identified one significantglobal recombination in alignments of MHC sequencesfrom Carpodacus although no recombination point wasfound in Acrocephalus Parus or Passer REL and FELanalyses which incorporate information of recombinationprovided the evidence that two to 14 and one to eight sitesare under positive selection respectively (Table 2) The
Immunogenetics (2009) 61451ndash461 455
positively selected sites were often identical between thefour compared species as well as between the three methodsof analysis (Fig 3)
Recombination in MHC class I of the Scarlet Rosefinch
Significant negative correlation between physical distanceand linkage disequilibrium was detected using all threeanalyses in the program PERMUTE (plt0001) suggestingthat recombination regularly occurs in exon 3 of MHC classI genes The mean population recombination per codon (ρ=0233) identified in the program OMEGAMAP is approxi-mately half of the mean total population mutation (θ=0534)
again suggesting that recombination is important for gener-ating MHC class I variants in the Scarlet Rosefinch
Discussion
Structure of classical MHC class I genes in passerine birds
In this study the structure and diversity of MHC class I wasstudied in a wild-living songbird species Up to ninedifferent MHC class I sequences were present in oneScarlet Rosefinch individual which means that at least fiveamplified MHC class I genes could occur in a single bird
98 108 118 128 138 148 158 168 178 |||||||||||||||||| CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEG-EAEGWTNYLKHECPEWLQKYVGYGQKELER 1 CaerU12 R-VENVR 2 CaerU29 Y-VENV 1 CaerU05 RLRYP-PRRH 4 CaerU02 LWRLPKKPNSTGKIVRNVSRR 119CaerU17 LWLPKKPNSTGKIVRNVSRRERH 4 CaerU18 LWLPKKPNSTGKIVRNVSRR 32 CaerU08 FIQWLSVRRAFIED-FRR 27 CaerU16 FIQWLSVRRAFIEDK-FRR 1 CaerU22 FIQWLSVRRAFIEDK-FIRE 7 CaerU20 FIQWLSVDARFIP-VKR 19 CaerU06 WRSSEVYYRFIPDVKE-VFRE 51 CaerU07 WRSSEVYYWRFIPDVKE-VFRE 111CaerU11 WRSSEVYYRFIPDVKE-VFRER CaerU04 LWVRHNRFID-FIC 9 CaerU13 LRVRRARFIRDK-MRDV 1 CaerU14 LRVRRARFIRD-MRDV 1 CaerU27 WRVRRARFIDG-VRRDV 1 CaerU28 LRVRRARFIRDKE-VF 1 CaerU03 LNVRYNRFIPD-PRW 4 CaerU09 RNVRYNRFIELDKE-AF 11CaerU10 WRNVRYNRFIPDKE-VFIH 5 CaerU15 WRNVRYNRFIEPD-PRVI 6 CaerU23 LWNVRYNRFIPDSKE-VLH 11CaerU24 WRNVRYRFIPD-PRRVH 4 CaerU25 WRSNVRFYNRFIPDKE-VFDQH 3 CaerU21 LWNVRDARFIDNGKE-VF 7 CaerU26 LRNVRDARFIDNGK-VEMRE 10Gallus gallus SVQWMIEGPRYYQMARFTAKTMTTVPEVPKKES-PRKEETVRREKAG Coturnix japonica SVQWMIDTTRYNQYARFIVKDTMTTVPEVPKKEGD-YRQKHEETVQRRHENKAG Anas platyrhynchos SVQRMKRFEQYERFIALKDTLTTDAQKKETYRTKYENTIRSKDV + + ++ ++ +
Fig 2 Amino acid sequences encoded by exon 3 in the ScarletRosefinch corresponding to alpha2 domain of the protein comparedwith GenBank sequences from chicken (X12780) quail (D29813) andduck (AY294416) Identity with the Scarlet Rosefinch allele CaerU1is indicated by dots gaps which are added to get the best alignment bydashes Highly conserved residues that anchor the peptide-terminalmain-chain atoms are marked by crosses other residues forming
variable part of peptide-binding region in HLA-A and chicken(Wallny et al 2006 Koch et al 2007) are marked by asterisks Thenumbering of amino acid residues is based on the complete MHCclass I protein in the duck (Mesa et al 2004) The numbers on theright show the number of individuals with the particular sequence (N=120) question mark the precise number of individuals bearing theallele CaerU11 is not known (see the text for more details)
Genus Numbera Amino acid sequence distance plusmn SD Nucleotide sequence distance plusmn SD
Acrocephalus 37 0514plusmn0189 0185plusmn0053
Parus 24 0734plusmn0167 0260plusmn00483
Passer 21 0538plusmn0154 0187plusmn0040
Gallus 1 1390plusmn0102 0689plusmn0040
Coturnix 1 1618plusmn0149 0739plusmn0050
Anas 1 1247plusmn0122 0607plusmn0037
Table 1 Average distancesbetween the 29 Scarlet Rose-finch sequences and sequencesof six other bird species
a Number of sequences in theanalysed alignment
456 Immunogenetics (2009) 61451ndash461
Although gene expression analysis was not performedstrong correlation between signatures for balancing selec-tion and expression is suggested Furthermore the presenceof conserved sites that anchor the peptide-terminal main-chain atoms and a reading frame with no terminal codons orframeshift mutations suggest that possibly all sequences ofMHC class I described in this study might be functionalalthough further study at the level of cDNA is required toconfirm this There is an increasing number of studiesshowing that even nearly intact MHC sequences are notexpressed (eg Indovina et al 2001 Bonneaud et al 2004Shiina et al 2006) Hence MHC class I genes seem to behighly variable in the Scarlet Rosefinch and this compli-cated and complex structure is very different from non-
passerines For example the best described MHC in birdsthe chicken lsquominimal essentialrsquo MHC has very few buthighly condensed genes (eg there are two classical MHCclass I genes but only a single dominantly expressedKaufman et al 1999) it appears that this is the ancestralstate for birds since it occurs in various groups such asother gallinaceous birds (Jarvi et al 1999 Wittzell et al1999a Worley et al 2008) owls (Burri et al 2008) andparrots (Hughes et al 2008) Up to now the mostcomplicated MHC class I gene set has been described intwo species of passerines in the great reed warbler sevengenomic class I sequences were found in one individualone of which contained a frameshift mutation and washence nonfunctional (Westerdahl et al 1999) and eight
Table 2 Results of the likelihood ratio tests of positive selection on exon 3 of MHC class I in four genera of passerine birds
Genus Length Numbera M8 vs M7 Likelihood ratio test Number of positively selected sites
df Test statistic Significance PAML REL FEL
Carpodacus 91 29 2 578 plt0001 15 7 8
Acrocephalus 68 37 2 712 plt0001 8 14 4
Parus 69 24 2 472 plt0001 13 14 5
Passer 82 21 2 232 plt0001 2 2 1
Test statistic was computed as 2(LbminusLa) where La and Lb are log-likelihood values for each of the nested models being compared (M8 and M7respectively)
Length length of the aminoacid sequencesa Number of sequences in the analysed alignment
A - PAML 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ + B ndash REL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
C ndash FEL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
Fig 3 Comparison of results of three methods for detecting aminoacid sites under selection in four passerine species The analyses arebased on the alignments of 29 Rosefinch 37 warbler 24 tit and 21sparrow complete or partial sequences of exon 3 encoding alpha2domain of the protein Amino acid residues found to be under positiveselection by three methods (A PAML B REL C FEL) are bold (fordetails on the methods see the text) If the same residue was found tobe positively selected in at least two genera it is marked by rectangle
Regions excluded from analysis are marked by a squiggle Highlyconserved residues that anchor the peptide-terminal main-chain atomsare marked by crosses other residues forming variable part of peptide-binding region in HLA-A and chicken (Wallny et al 2006 Koch et al2007) are marked by asterisks The numbering of amino acid residuesis based on the complete MHC class I protein in the duck (Mesa et al2004) (Acrocephalus-AF449690) (Parus-AM232710) and (Passer-AY285012)
Immunogenetics (2009) 61451ndash461 457
different cDNA sequences were also found in an individualIn a separate study of the same species 20 genomicsequences of exon 3 of class I were found in fiveindividuals five of which were pseudogenes (Westerdahlet al 2004) Similar results were found in house sparrowswhere 20 sequences were verified in seven individualsthree of which were proved to be transcribed althoughthere were no pseudogenes (Bonneaud et al 2004) Ourresults concerning individual exon 3 variation are similar tothe ones mentioned above suggesting that complex MHCstructure could be a general pattern in songbirds
Evolution of MHC class I sequences in passerines
Several mechanisms of molecular evolution have beenalready documented in the avian MHC sequences includ-ing positive selection (Alcaide et al 2007 Hughes et al2008) concerted evolution (Wittzell et al 1999b) andtrans-species polymorphism (Richardson and Westerdahl2003 Alcaide et al 2007) In the present study model-based tests of selection (used here for the first time inpasserine species) have demonstrated the presence ofpositively selected sites in exon 3 of MHC class I in fourpasserine genera This part of the gene encodes the alpha 2domain of the protein which is included in forming thepeptide-binding groove Positive selection acting on somecodons of exon 3 is in accordance with published data onMHC class I in other birds where the ratio of non-synonymous to synonymous substitutions (dNdS) wasusually slightly higher at PBR sites than at non-PBR ones(Westerdahl et al 1999 Richardson and Westerdahl 2003Bonneaud et al 2004) The differences however were notsignificant and this could be caused either (1) by in-correctly identified PBR sites since the MHC proteinstructure in the majority of species studied is not well-understood In fact the PBR sites have usually beenidentified on the basis of the three-dimensional structureof human HLA-A protein (Bjorkman et al 1987) A modelof the three-dimensional structure of a passerine MHC classI molecule is highly desirable Or (2) another reason mightbe the fact that some of the studied sequences come fromloci that lost their importance and they are not underselection that is why it is crucial to identify sequences toparticular loci in the future studies (see Worley et al 2008)
Our results indicate extreme variability in selectivepressure among sites in the exon 3 and the presence of anumber of sites under balancing selection Approximatelyhalf of the positively selected sites in passerines (Fig 3)have been previously assumed to play a role in antigenbinding based on the structural models (Wallny et al 2006Koch et al 2007) However we found at least twoadditional sites (amino acid residues 126 and 136) understrong positive selection in several genera which have not
been previously reported for PBR (either in human HLA orchicken BF) The maintenance of considerable nonsynon-ymous variation at these specific codons by positiveselection suggests that such polymorphism may havefunctional consequences for peptide binding and presenta-tion to T cells (see similar recent results concerning teleostfish MHC molecules Aguilar and Garza 2007 or inmammals Amills et al 2008) The predictive power ofcodon-based models has not yet been tested in MHC andthe structural model describing peptide-binding groove inpasserine birds is still required to confirm the functionalrole of amino acid residues under different types ofselection
It appears that mammalian and avian MHCs may differin frequency of genetic exchange between different genesof multigene families Concerted evolution where the inter-locus gene conversion events are frequent in highlyduplicated MHC genes may be the predominant evolu-tionary mechanism in birds (Edwards et al 1995 Alcaide etal 2007 Garrigan and Edwards 1999 Wittzell et al 1999bHess and Edwards 2002) and probably also in fish (Reuschet al 2004 van Oosterhout et al 2006 Aguilar and Garza2007) and reptiles (Miller et al 2006) but not in mammals(Edwards et al 1997) This process creates high homologyamong MHC sequences derived from various loci and itbecomes impossible to assign different MHC alleles tospecific loci In birds so far most of the studies evidencinginter-locus recombination were conducted on MHC class IIConcerning MHC class I it has only been studied in thechicken where no evidence was found for concertedevolution to occur (Shaw et al 2007) Thus our resultson recombination analyses in the finches (ie the totalamount of recombination per codon was half of that ofsubstitution and we failed to assign the sequences tospecific loci) are in agreement with previous reports andfurthermore they suggest that concerted evolution may befrequent also in MHC class I genes in passerine birds Toconfirm this hypothesis the more detailed description ofMHC structure (especially the affiliation of particularalleles to individual loci in a population and the geneticvariation outside exon 3) is necessary
Extreme MHC class I variation in a single population
Previous studies on songbirds using the method ofdenaturing gradient gel electrophoresis or restriction frag-ment length polymorphism (RFLP) have also indicatedhigh sequence variation at exon 3 of MHC class I forexample in an outbred population of house sparrows 46sequences were found in 101 individuals or between oneand 11 alleles per individual (Bonneaud et al 2006a) in thegreat reed warbler 49 different RFLP genotypes werefound in 55 birds (Westerdahl et al 2004) Even so in the
458 Immunogenetics (2009) 61451ndash461
Scarlet Rosefinch 82 alleles were found in a singlepopulation and most of the birds displayed uniquegenotypes which is the highest exon 3 MHC class Ivariability observed thus far This result may still be anunderestimate considering that in at least one case theSSCP pattern was identical for two different allelesAlthough we were not able to sequence all the allelesdisplayed on SSCP it is very probable that all the peaksidentified by SSCP are true alleles and that there could beadditional ones (due to peak overlapping) Another reasonwe believe this variation has not been overestimated is thatan extraordinarily high variability of selectively neutralmarkers has been found in this species at two microsatelliteloci more than 70 alleles were found (Albrecht et al 2007)
Interestingly one of the alleles (CaerU02) was possessedby all individuals but one female (no 515) investigated inthis study From parentndashoffspring genotypes (Promerovaacute etal unpublished data) we know that also some of thisfemales offspring missed this allele thus this absence issurely heritable We conclude that CaerU02 is the mostfrequent allele on a locus with very low variability Thefemale no 515 is either homo- or heterozygote for other veryrare allele(s) or the locus (at least the copy with alleleCaerU02) is completely missing in her MHC haplotype
Our optimization of a genotyping method for analyses ofMHC class I variation will allow us to search for answers tofundamental questions concerning recent immunoecologysuch as the role of MHC in disease resistance expression ofsecondary ornaments and mating success (eg Bonneaud etal 2006a b Loiseau et al 2008 Richardson et al 2004Westerdahl et al 2005) Furthermore our data could beused for future comparative studies of the correlationbetween MHC diversity and migratory strategies in birdsWe presume that migratory birds such as the ScarletRosefinch (which migrates from the Czech Republic toIndia ca 5000 km) might encounter a larger variety ofpathogens during their lifetime than sedentary birds (forexample there is evidence that migrating individualsencounter more malarial parasites Peacuterez-Tris and Bensch2005) In that case more diverse immune mechanismscould help them fight these pathogens Moreover it hasalready been suggested that migratory species have highergenetic variability than sedentary species (Fitzpatrick 1994)and also that the higher genetic variability of migrants hasan impact on higher occurrence of extra-pair paternity(Spottiswoode and Moslashller 2004) The hypothesis thatmigration in birds might elicit a higher variability onMHC was first evaluated by Shiina et al (2004) whocompared the MHC of migratory quail with that ofsedentary chicken The quail has a more complicatedMHC than the chicken the class I region in the quail istwice as long as that of the chicken (Shiina et al 2004) andthere are five expressed class I genes although not all of
them are classical ones (Shiina et al 2006) SimilarlyRichardson and Westerdahl (2003) reported a difference inMHC variability between the migratory Great reed warblerand the sedentary inbred Seychelles warbler In this casehowever the low MHC variation in the latter could be dueto a severe bottleneck leading to strong decrease of overallgenetic variation Furthermore the endemic Basra reedwarbler (Acrocephalus griseldis) is as variable at MHC asthe Great reed warbler even though it is sedentary (Hanssonand Richardson 2005) High level of MHC polymorphismwas found in the house sparrow which is a non-migratoryspecies (Bonneaud et al 2004) which also contradicts thepredictions of the above hypothesis Unfortunately formost species (including the Scarlet Rosefinch) there is noprecise information about the functional variation ienumber of expressed MHC variants It is evident that dataon real MHC variation in more species are inevitablyrequired to perform a rigorous comparative meta-analysisand to test the predictions of migration-MHC hypothesis
Acknowledgements We thank Pavel Munclinger Michal VinklerJan Schnitzer and Radka Polaacutekovaacute for their help in the field Thanksto Anna Bryjovaacute and Dagmar Čiacutežkovaacute for their valuable advice inthe laboratory We are very grateful to Heidi C Hauffe for improvingthe English and two anonymous referees for critical comments on themanuscript This study was funded by the Grant Agency of theAcademy of Sciences of the Czech Republic (project noIAA600930608) the Grant Agency of the Czech Rep (project no206060851) and by the Ministry of Education of the Czech Repwhose Research Centre no LC06073 and Long-term Research Plan(no MSM 0021620828) to the Charles University in Prague formed aframework for a part of this study
References
Abbas AK Lichtman AH Pober JS (1994) Cellular and molecularimmunology W B Saunders Company Philadelphia
Aguilar A Garza JC (2007) Patterns of historical balancing selectionon the salmonid major histocompatibility complex class II betagene J Mol Evol 6534ndash43 doi101007s00239-006-0222-8
Albrecht T (2004) Edge effect in wetland-arable land boundarydetermines nesting success of scarlet rosefinches Carpodacuserythrinus in the Czech Republic Auk 121361ndash371doi1016420004-8038(2004)121[0361EEIWLB]20CO2
Albrecht T Schnitzer J Kreisinger J Exnerovaacute A Bryja JMunclinger P (2007) Extrapair paternity and the opportunity forsexual selection in long-distant migratory passerines Behav Ecol18477ndash486 doi101093behecoarm001
Alcaide M Edwards SV Negro JJ (2007) Characterization polymor-phism and evolution ofMHCClass II B genes in birds of prey JMolEvol 65541ndash554 doi101007s00239-007-9033-9
Amills M Ramiacuterez O Tomaacutes A Obexer-Ruff G Vidal O (2008)Positive selection on mammalian MHC-DQ genes revisited from amultispecies perspective Genes Immun 9651ndash658 doi101038gene200862
Anisimova M Nielsen R Yang Z (2003) Effect of recombination onthe accuracy of the likelihood method for detecting positiveselection at amino acid sites Genetics 1641229ndash1236
Immunogenetics (2009) 61451ndash461 459
Bjorkman PJ Saper MA Samraoui B Bennett WS Strominger JLWiley DC (1987) Structure of human class I histocompatibil-ity antigen HLA-A2 Nature 329506ndash512 doi101038329506a0
Bonneaud C Sorci G Morin V Westerdahl H Zoorob R Wittzell H(2004) Diversity of MHC class I and IIB genes in house sparrows(Passer domesticus) Immunogenetics 55855ndash865 doi101007s00251-004-0648-3
Bonneaud C Chastel O Federici P Westerdahl H Sorci G (2006a)Complex MHC-based mate choice in a wild passerine Proc RSoc 2731111ndash1116 doi101098rspb20053325
Bonneaud C Peacuterez-Tris J Federici P Chastel O Sorci G (2006b) Majorhistocompatibility alleles associated with local resistance to malariain a passerine Evolution Int J Org Evolution 60383ndash389
Bryja J Galan M Charbonnel N Cosson J-F (2005) Analysis ofmajor histocompatibility complex class II gene in water volesusing capillary electrophoresis-single stranded conformationpolymorphism Mol Ecol Notes 5173ndash176 doi101111j1471-8286200400855x
Burri R Niculita-Hirzel H Roulin A Fumagalli L (2008) Isolation andcharacterization of major histocompatibility complex (MHC) class IIB genes in teh Barn owl (Aves Tyto alba) Immunogenetics 60543ndash550 doi101007s00251-008-0308-0
Cramp S Simmons KEL Perrins CM (eds) (1977-1994) The birds ofthe Western Palearctic Oxford University Press Oxford
Edwards SV Wakeland EK Potts WK (1995) Contrasting histories ofavian and mammalian MHC genes revealed by class II B genesof songbirds Proc Natl Acad Sci U S A 9212200ndash12204doi101073pnas922612200
Edwards SV Chesnut K Satta Y Wakeland EK (1997) Ancestralpolymorphism of Mhc Class II genes in mice implications forbalancing selection and the mammalian molecular clock Genetics146655ndash668
Fitzpatrick S (1994) Colourful migratory birds evidence for amechanism other than parasite resistance for the maintenance oflsquogood genesrsquo sexual selection Proc R Soc Lond B Biol Sci257155ndash166 doi101098rspb19940109
Freeman-Gallant CR Johnson EM Saponara F Stanger M (2002)Variation at the major histocompatibility complex in Savannahsparrows Mol Ecol 111125ndash1130 doi101046j1365-294X200201508x
Garrigan D Edwards SV (1999) Polymorphism across an exon-intronboundary in an avian Mhc class II B gene Mol Biol Evol 161599ndash1606
Gouumly de Bellocq J Charbonnel N Morand S (2008) Coevolutionaryrelationship between helminth diversity and Mhc class II polymor-phism in rodents J Evol Biol 211144ndash1150 doi101111j1420-9101200801538x
Hall TA (1999) BioEdit a user-friendly biological sequence alignmenteditor and analisis program for Windows 9598NT NucleicAcids Symp Ser 4195ndash98
Hansson B Richardson DS (2005) Genetic variation in twoendangered Acrocephalus species compared to a widespreadcongener estimates based on functional and random loci AnimConserv 883ndash90 doi101017S1367943004001878
Hess CM Edwards SV (2002) The evolution of major histocompat-ibility genes in birds Bioscience 52423ndash431 doi1016410006-3568(2002)052[0423TEOTMH]20CO2
Hess CM Gasper J Hoekstra H Hill C Edwards SV (2000) MHCclass II pseudogene and genomic signature of a 32-kb cosmid inthe House Finch (Carpodacus mexicanus) Genome Res 1013ndash23 doi101101gr105613
Hosomichi K Shiina T Suzuki S Tanaka M Shimizu S Iwamoto SHara H Yoshida Y Kulski JK Inoko H Hanzawa K (2006) Themajor histocompatibility complex (Mhc) class IIB region hasgreater genomic structural flexibility and diversity in the quail
than the chicken BMC Genomics 7322ndash335 doi1011861471-2164-7-322
Hughes AL Yeager M (1998) Natural selection at major histocom-patibility complex loci of vertebrates Annu Rev Genet 32415ndash435 doi101146annurevgenet321415
Hughes CR Miles S Walbroehl JM (2008) Support for the minimalessential MHC hypothesis a parrot with a single highlypolymorphic MHC class II B gene Immunogenetics 60219ndash231 doi101007s00251-008-0287-1
Indovina P Megiorni F Fontemaggi G Coni P Mora B Mazzilli MC(2001) Absence of in vivo DNA-Protein interactions in theDQA2 and DQB2 promoter regions Hum Immunol 62504ndash508doi101016S0198-8859(01)00236-1
Jarvi SI Goto RM Gee GF Briles WE Miller MM (1999)Identification inheritance and linkage of B-G-like and MHCclass I genes in cranes Am Genet Assoc 90152ndash159
Kaufman J (1999) Co-evolving genes in MHC haplotypes the lsquorulersquofor nonmammalian vertebrates Immunogenetics 50228ndash236doi101007s002510050597
Kaufman J Salamonsen J Flajnik M (1994) Evolutionary conserva-tion of MHC class I and class II molecules-different yet the sameSemin Immunol 6411ndash424 doi101006smim19941050
Kaufman J Voumllk H Wallny H (1995) A bdquominimal essential MHCldquoand an bdquounrecognized MHCldquo two extremes in selection forpolymorphism Immunol Rev 14363ndash88 doi101111j1600-065X1995tb00670x
Kaufman J Jacob J Shaw I Walker B Milne S Beck S SalomonsenJ (1999) Gene organisation determines evolution of function inthe chicken MHC Immunol Rev 167101ndash117 doi101111j1600-065X1999tb01385x
Klein J (1986) Natural history of the major histocompatibilitycomplex Wiley New York
Klein J Sato A Nagl S Oh Uigin C (1998) Molecular trans-speciespolymorphism Annu Rev Ecol Syst 291ndash21 doi101146annurevecolsys2911
Koch M Camp S Collen T Avila D Salomonsen J Wallny HJ vanHateren A Hunt L Jacob JP Johnston F Marston DA Shaw IDunbar PR Cerundolo V Jones EY Kaufman J (2007)Structures of an MHC class I molecule from B21 chickensillustrate promiscuous peptide binding Immunity 27885ndash899doi101016jimmuni200711007
Kosakovsky Pond SL Frost SDW (2005) Not so different after all acomparison of methods for detecting amino acid sites under selectionMol Biol Evol 221208ndash1222 doi101093molbevmsi105
Kosakovsky Pond SL Frost SDW Muse SV (2005) HyPhyhypothesis testing using phylogenies Bioinformatics 21676ndash679 doi101093bioinformaticsbti079
Kosakovsky Pond SL Posada D Gravenor MB Woelk CH FrostSDW (2006) Automated phylogenetic detection of recombinationusing a genetic algorithm Mol Biol Evol 231891ndash1901doi101093molbevmsl051
Loiseau C Zoorob R Garnier S Birard J Federici P Julliard R SorciG (2008) Antagonistic effects of a MHC class I allele on malaria-infected house sparrows Ecol Lett 11258ndash265 doi101111j1461-0248200701141x
Mesa CM Thulien KJ Moon DA Veniamin SM Magor KE (2004)The dominant MHC class I gene is adjacent to the polymorphicTAP2 gene in the duck Anas platyrhynchos Immunogenetics56192ndash203 doi101007s00251-004-0672-3
Miller HC Lambert DM (2004) Gene duplication and geneconversion in class II MHC genes of New Zealand robins(Petroicidae) Immunogenetics 56178ndash191
Miller MM Goto RM Bernot A Zoorob R Auffray C Bumstead NBriles WE (1994) Two Mhc class I and two Mhc class II genes mapto the chicken Rfp-Y system outside the B complex Proc Natl AcadSci U S A 914397ndash4401 doi101073pnas91104397
460 Immunogenetics (2009) 61451ndash461
Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461

the primers HN34 (5prime-CCATGGGTCTCTGTGGGTA-3prime)and HN45 (5prime-CCATGGAATTCCCACAGGAA-3prime) takenfrom Westerdahl et al (2004) using the polymerase chainreaction (PCR) with the following conditions 15 mMMgCl2 04 μM of each primer 02 mM dNTPs 1 UμlTaq polymerase (Fermentas) in appropriate 1times PCR bufferand 1 μl of extracted DNA Deionised water was added toa 10-μl reaction volume The amplification consisted of aninitial denaturation at 94degC (3 min) followed by 30 cyclesof denaturation at 94degC (40 s) annealing at 60degC (30 s)and extension at 72degC (1 min) with a final extension stepat 72degC (10 min) The reactions were run on a Master-cycler ep (Eppendorf) The PCR products were purifiedusing the MinElute PCR purification kit (Qiagen) ligatedto the vector and transformed to bacteria using the pGEM-T Easy Cloning Kit (Promega) according to manufac-turers protocols Positive transformants containing insertsof appropriate length were identified by PCR screeningusing primers on the vector and agarose gel electrophore-sis We randomly chose eight clones extracted plasmidDNA using the QIAprep Spin Miniprep Kit (Qiagen) andsequenced the inserts using the BigDye Terminators Se-quencing Kit v11 and an ABI PRISM 3130 GeneticAnalyzer (Applied Biosystems) Based on these eightsequences a new reverse primer specific for the ScarletRosefinch was designed (CE-MHCREX3 5prime-ACAGGAATTCTGCTCCCACC-3prime) that together with forwardprimer HN34 amplifies the complete exon 3 of the MHCclass I genes of the Scarlet Rosefinch under almost the samePCR conditions as described above (the only difference isthe annealing step which changes to 30 s at 59degC)
Capillary electrophoresis single-strand conformationpolymorphism
Exon 3 of MHC class I was amplified by PCR usingfluorescently labelled primers (HN34 by 6-FAM and CE-MHCREX3 by NED) and conditions described abovePositive PCR amplifications were checked by agarose gelelectrophoresis and then single-strand conformation poly-morphism (SSCP) analysis (a method able to detect even asingle nucleotide substitution) was performed using capil-lary electrophoresis on an ABI PRISM 3130 GeneticAnalyzer (Applied Biosystems) We mixed 2 μl of PCRproduct with 05 μl of GeneScan-500 LIZtrade Size standard(Applied Biosystems) and 12 μl Hi-Ditrade Formamide anddenaturated the mix at 95degC for 3 min After denaturationsamples were immediately put on ice for at least 2 min andelectrophoresis was run at 22degC in 5 non-denaturingconformation analysis polymer (prepared from 9 CAPpolymer Applied Biosystems according to manufacturersinstructions) on an ABI PRISM 3130 Data were analysedusing the software GeneMapper v37 (Applied Biosystems)
and user-defined size standard files for the alignment ofdifferent runs The alleles were identified by both theposition and the shape of peaks For each allele one 6-FAMand one NED peak was identified It is important to notethat neither SSCP peaks nor sequences can be assigned tospecific loci but for simplification in this study they aretermed lsquoallelesrsquo To obtain the sequences of particularalleles (ie SSCP peaks) we cloned the PCR products(obtained by non-labelled primers HN34 and CE-MHCREX3) using the techniques as described aboveSSCP was subsequently used for the identification ofinserted sequences after cloning This procedure allowedus to (1) minimise cost by avoiding the sequencing of thesame sequence in many clones and (2) compare SSCPpatterns of clones with those obtained from individuals toidentify PCR artefacts which are common when analysingMHC genes with a cloning and sequencing approach (Bryjaet al 2005) Inserts providing the SSCP patterns thatmatched the SSCP profile of an individual bird ie theinserts contained one of the exon 3 variants of theindividual were re-amplified by specific PCRs and se-quenced using Big Dye kits as described above
Analyses of MHC class I sequence variation
The sequences of MHC class I derived from C erythrinuswere edited in SeqScape v25 (Applied Biosystems) andaligned in BioEdit Sequence Alignment Editor (Hall 1999)To determine whether all sequences represent functionalclassical class I alleles the sequences were examined (1)for the presence of insertions or deletions (lsquoindelsrsquo) causingshift of the reading frame andor stop codons and (2) forthe conservation of residues that anchor the peptide-terminal main-chain atoms Possible codons encoding theamino acid residues included in the peptide binding wereidentified on the basis of homology between chicken MHCand human leukocyte antigen (HLA)-A (eg Wallny et al2006) and by analysing the detailed structural models ofpeptide-binding grooves of chicken MHC class I proteins(Koch et al 2007) Some of the residues are generallyhighly conserved throughout evolution in classical class Imolecules (Kaufman et al 1994 Mesa et al 2004) whileothers are thought to be under strong positive selection andallow interaction with a wide variety of antigens Sequencesof chicken (GenBank acc no X12780) Japanese quail(D29813) and duck (AY294416) were used to compareMHC sequences of Scarlet Rosefinches with those ofdistantly related bird species
Detecting amino acid sites under selection
To detect selection acting on MHC class I gene variationsequences (partial or complete) of exon 3 MHC class I for
Immunogenetics (2009) 61451ndash461 453
three additional genera of passerines were downloadedfrom GenBank (accession nos in parentheses) theseinclude two species of warbler of the genus Acrocephalus(AF449688ndashAF449701 AJ005503 AJ005504 AJ005506AJ005508ndashAJ005510 AJ005512 AJ005514ndashAJ005517AY306008 AY306009 AJ557874ndashAJ557883) the housesparrow (AY284994ndashAY285013 EF429132) and threespecies of tit of the genus Parus (AF346821 AF346823AF346825 AF346827 AF346829ndashAF346832 AM232707 AM232708 AM232710 AM232712ndashAM232715 EF446975ndashEF446978 EF446980ndashEF446982EF446985ndashEF446987) pseudogenes and identical sequen-ces were removed from the analyses (Fig 1) Theserepresent all the accessible sequences of exon 3 class IMHC available to date for songbirds Alignment gaps weretreated as missing data in all analyses Pairwise distancesbetween sequences of different genera were calculated inBioEdit Sequence Alignment Editor (Hall 1999) using theKimura 2-parameter model The length of the sequencesvaried between species and for the analysis of sequencedistances the sequences were cut to a uniform lengthaccording to the shortest available sequence
Many methods for identifying sites under positive ornegative selection are available and we chose three ofthem First we examined the presence of selection using awidely used likelihood ratio modelling approach inCodeML which is included in the PAML 314 programsuite (Yang 1997) Maximum-likelihood models were usedwhich account for heterogeneity among site partitions usingdifferent ω (=dNdS) parameters for the partitions Themodels applied for the analysis of four alignments (fourpasserine genera) in this study are M7 and M8 (Yang et al2000) Models were compared using likelihood ratio teststatistics calculated as follows 2(LbminusLa) was comparedwith χ2 distribution with PbminusPa degrees of freedom where
La and Lb are log-likelihood values and Pa and Pb are thenumbers of parameters for each of the compared nestedmodels (M8 and M7 respectively) If the alternative modelie M8 suggests the presence of sites with ωgt1 the testcan be considered a test of positive selection (Yang et al2000) Posterior probabilities for site classes were calculat-ed by the Bayes Empirical Bayes (BEB) method in modelM8 If the posterior means of ω for some sites are gt1 thosesites are likely to be under positive selection (Yang et al2005)
The likelihood methods for quantifying selection pres-sure on codon alignments may have high rates of falsepositives if the sequences have undergone recombination(Anisimova et al 2003) therefore nonrecombinant frag-ments were identified using the Genetic Algorithm Recom-bination Detection (GARD Kosakovsky Pond et al 2006)Subsequently the presence of selection on particularcodons was analysed separately for each of them usingtwo methods First the random effects likelihood (REL)approach which involves fitting a distribution of substitu-tion rates across sites and then inferring the rate at whichindividual sites evolve was used Like CodeML thismethod makes inference on selection using an empiricalBayes approach Second the fixed effects likelihood (FEL)method was also applied this approach directly estimatesthe rates of nonsynonymous and synonymous substitutionsat each site (Kosakovsky Pond and Frost 2005) GARDREL and FEL analyses were implemented in the HyPhysoftware package (Kosakovsky Pond et al 2005) and aweb-based interface running on a cluster of computers athttpwwwdatamonkeyorg was employed (KosakovskyPond and Frost 2005)
Analysis of recombination in the Scarlet Rosefinch MHCclass I
Recombination events were recently shown to be frequentinside the MHC domain (either intra-locus or inter-locus)We used three approaches to detect if recombination wasimportant in generating recent diversity of MHC class Isequences in the Scarlet Rosefinch We analysed threecommonly used statistics (r2 Dprime and G4) by permutation inthe program PERMUTE (included in the package OME-GAMAP Wilson and McVean 2006) to test the correlationbetween linkage disequilibrium and physical distance of thesingle-nucleotide polymorphism sites In the absence ofrecombination all sites are linked and there should be zerocorrelation but if recombination has occurred the correla-tion should be significantly negative
The level of recombination rate in the history of the setof 29 aligned sequences was estimated using the Bayesianapproach incorporated in OMEGAMAP (Wilson andMcVean 2006) which is primarily employed to co-
3 4 5 6 7 8
No of exon 3 variants
0
10
20
30
40
50
60
No
of in
divi
dual
s
Fig 1 Distribution of numbers of different exon 3 MHC class Ialleles per individual in a Scarlet Rosefinch population (N=120)
454 Immunogenetics (2009) 61451ndash461
estimate the selection parameter ω and the recombinationrate (ρ=4Nec) along the sequence
Results
MHC class I variation in the Scarlet Rosefinch
Amplification of genomic DNA with primers HN34 andCE-MHCREX3 gave a single PCR product of about 325 bpin all individuals including the whole exon 3 of 270 bp(273 bp in some alleles see below) and encoding thealpha2 domain (corresponding to part of the PBR and smallparts of adjacent introns) Among 120 adult Scarlet Rose-finches we identified 82 different variants of amplifiedMHC class I sequences using high-resolution capillarySSCP The SSCP profiles were highly repeatable andindividuals exhibited between three and nine variants (oneof these variants was impossible to distinguish by SSCPalthough it was repeatedly sequenced [see below]) Theseresults suggest that as many as five MHC class I genes arepresent in the Scarlet Rosefinch and the number ofamplified genes is variable between individuals
Due to the extremely high diversity of MHC class I inthe studied population not all variants were analysed indetail In total we obtained 29 sequences of exon 3 ofMHC class I genes by cloning and sequencing PCRproducts from 18 highly heterozygous individuals (identi-fied by SSCP) Each sequence was provided with thespecies abbreviation and a number (ie CaerU01-CaerU29) In this abbreviation lsquoUrsquo stands for theunknown locus designation of the gene (Mesa et al2004) In three individuals (ID 541 667 and 692) wemanaged to sequence all the variants identified by SSCP ofthese individuals (six four and eight alleles respectively)and all these sequences correspond to exon 3 of MHC classI when BLASTed (httpblastncbinlmnihgovBlastcgi)In total 597 clones were analysed by SSCP 156 of whichcontained either PCR artefacts or were contaminated byPCR product from the individual bird 55 contained noinsert and 122 contained the most frequent sequenceCaerU02 This variant of exon 3 occurred in all analysedadults except one female However the true number ofalleles is probably underestimated because some sequencevariants were difficult to distinguish by SSCP alleleCaerU11 was poorly visualised on SSCP because the6-FAM peak was very low and the NED peak wasoverlapped by CaerU02 (for this reason this allele mighthave been overlooked in some individuals and thereforewe do not have the precise number of individuals in whichthis allele occurred (Fig 2) even if it was repeatedlysequenced in four individuals) Most SSCP patterns werehighly specific in fact we found only one case in which
two different MHC sequences repeatedly confirmed bycloning and sequencing gave identical SSCP peaks incapillary electrophoresis (CaerU18 and CaerU19)
All 29 sequences were obtained from randomly chosenclones whose SSCP pattern matched exactly the SSCPpattern found in the profile of the individual from which theclone was isolated (to avoid sequencing PCR artefacts)Seventeen of these sequences were obtained from oneindividual each (seven of them were only present in oneindividual in the population) the remaining sequences wereidentified in clones originating from at least two differentbirds (two PCRs) These sequences translate into 28 uniqueamino acid sequences (CaerU02 and CaerU19 translateto identical alpha2 domains) and none of them containdeletions insertions or stop codons in the reading frameHowever four sequences have an insertion of 3 bp codingfor isoleucine (Fig 2) The pairwise distances between bothnucleotide and aminoacid sequences of the Scarlet Rose-finch and three other passerine genera (AcrocephalusPasser and Parus) chicken G gallus the Japanese quailC japonica and duck Anas platyrhynchos are shown inTable 1 (see also Fig 2)
Seven amino acid residues in the alpha2 domain offunctional MHC class I molecules are reported to be highlyconserved [Y123 T143 K146 W147 Y159 L160 Y171(Kaufman et al 1994 Shum et al 1999 Mesa et al 2004)]In all our Scarlet Rosefinch sequences four out of seven ofthese residues are conserved (W144W147 Y157Y159L158L160 Y169Y171 Fig 2) Y123 is not conserved inany avian sequence but the phenylalanine present in allRosefinch sequences (F123) is also reported in other non-mammalian species (Shum et al 1999 Mesa et al 2004)Threonine at position T140T143 is replaced by serine in theCaerU23 allele (S140T143) but both these residues belongto the same biochemical group of amino acids (ie theyshould substitute each other in peptide binding) The sameis true for K146 where lysine is replaced by arginine(R143K146) in all alleles (Fig 2)
Positive selection on MHC class I sequences in passerinebirds
Likelihood ratio tests comparing models M8 and M7 in thePAML package identified significant variability in selectivepressure among sites in exon 3 and the presence of two to15 sites under positive selection in four studied passerinespecies (Table 2) GARD analyses identified one significantglobal recombination in alignments of MHC sequencesfrom Carpodacus although no recombination point wasfound in Acrocephalus Parus or Passer REL and FELanalyses which incorporate information of recombinationprovided the evidence that two to 14 and one to eight sitesare under positive selection respectively (Table 2) The
Immunogenetics (2009) 61451ndash461 455
positively selected sites were often identical between thefour compared species as well as between the three methodsof analysis (Fig 3)
Recombination in MHC class I of the Scarlet Rosefinch
Significant negative correlation between physical distanceand linkage disequilibrium was detected using all threeanalyses in the program PERMUTE (plt0001) suggestingthat recombination regularly occurs in exon 3 of MHC classI genes The mean population recombination per codon (ρ=0233) identified in the program OMEGAMAP is approxi-mately half of the mean total population mutation (θ=0534)
again suggesting that recombination is important for gener-ating MHC class I variants in the Scarlet Rosefinch
Discussion
Structure of classical MHC class I genes in passerine birds
In this study the structure and diversity of MHC class I wasstudied in a wild-living songbird species Up to ninedifferent MHC class I sequences were present in oneScarlet Rosefinch individual which means that at least fiveamplified MHC class I genes could occur in a single bird
98 108 118 128 138 148 158 168 178 |||||||||||||||||| CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEG-EAEGWTNYLKHECPEWLQKYVGYGQKELER 1 CaerU12 R-VENVR 2 CaerU29 Y-VENV 1 CaerU05 RLRYP-PRRH 4 CaerU02 LWRLPKKPNSTGKIVRNVSRR 119CaerU17 LWLPKKPNSTGKIVRNVSRRERH 4 CaerU18 LWLPKKPNSTGKIVRNVSRR 32 CaerU08 FIQWLSVRRAFIED-FRR 27 CaerU16 FIQWLSVRRAFIEDK-FRR 1 CaerU22 FIQWLSVRRAFIEDK-FIRE 7 CaerU20 FIQWLSVDARFIP-VKR 19 CaerU06 WRSSEVYYRFIPDVKE-VFRE 51 CaerU07 WRSSEVYYWRFIPDVKE-VFRE 111CaerU11 WRSSEVYYRFIPDVKE-VFRER CaerU04 LWVRHNRFID-FIC 9 CaerU13 LRVRRARFIRDK-MRDV 1 CaerU14 LRVRRARFIRD-MRDV 1 CaerU27 WRVRRARFIDG-VRRDV 1 CaerU28 LRVRRARFIRDKE-VF 1 CaerU03 LNVRYNRFIPD-PRW 4 CaerU09 RNVRYNRFIELDKE-AF 11CaerU10 WRNVRYNRFIPDKE-VFIH 5 CaerU15 WRNVRYNRFIEPD-PRVI 6 CaerU23 LWNVRYNRFIPDSKE-VLH 11CaerU24 WRNVRYRFIPD-PRRVH 4 CaerU25 WRSNVRFYNRFIPDKE-VFDQH 3 CaerU21 LWNVRDARFIDNGKE-VF 7 CaerU26 LRNVRDARFIDNGK-VEMRE 10Gallus gallus SVQWMIEGPRYYQMARFTAKTMTTVPEVPKKES-PRKEETVRREKAG Coturnix japonica SVQWMIDTTRYNQYARFIVKDTMTTVPEVPKKEGD-YRQKHEETVQRRHENKAG Anas platyrhynchos SVQRMKRFEQYERFIALKDTLTTDAQKKETYRTKYENTIRSKDV + + ++ ++ +
Fig 2 Amino acid sequences encoded by exon 3 in the ScarletRosefinch corresponding to alpha2 domain of the protein comparedwith GenBank sequences from chicken (X12780) quail (D29813) andduck (AY294416) Identity with the Scarlet Rosefinch allele CaerU1is indicated by dots gaps which are added to get the best alignment bydashes Highly conserved residues that anchor the peptide-terminalmain-chain atoms are marked by crosses other residues forming
variable part of peptide-binding region in HLA-A and chicken(Wallny et al 2006 Koch et al 2007) are marked by asterisks Thenumbering of amino acid residues is based on the complete MHCclass I protein in the duck (Mesa et al 2004) The numbers on theright show the number of individuals with the particular sequence (N=120) question mark the precise number of individuals bearing theallele CaerU11 is not known (see the text for more details)
Genus Numbera Amino acid sequence distance plusmn SD Nucleotide sequence distance plusmn SD
Acrocephalus 37 0514plusmn0189 0185plusmn0053
Parus 24 0734plusmn0167 0260plusmn00483
Passer 21 0538plusmn0154 0187plusmn0040
Gallus 1 1390plusmn0102 0689plusmn0040
Coturnix 1 1618plusmn0149 0739plusmn0050
Anas 1 1247plusmn0122 0607plusmn0037
Table 1 Average distancesbetween the 29 Scarlet Rose-finch sequences and sequencesof six other bird species
a Number of sequences in theanalysed alignment
456 Immunogenetics (2009) 61451ndash461
Although gene expression analysis was not performedstrong correlation between signatures for balancing selec-tion and expression is suggested Furthermore the presenceof conserved sites that anchor the peptide-terminal main-chain atoms and a reading frame with no terminal codons orframeshift mutations suggest that possibly all sequences ofMHC class I described in this study might be functionalalthough further study at the level of cDNA is required toconfirm this There is an increasing number of studiesshowing that even nearly intact MHC sequences are notexpressed (eg Indovina et al 2001 Bonneaud et al 2004Shiina et al 2006) Hence MHC class I genes seem to behighly variable in the Scarlet Rosefinch and this compli-cated and complex structure is very different from non-
passerines For example the best described MHC in birdsthe chicken lsquominimal essentialrsquo MHC has very few buthighly condensed genes (eg there are two classical MHCclass I genes but only a single dominantly expressedKaufman et al 1999) it appears that this is the ancestralstate for birds since it occurs in various groups such asother gallinaceous birds (Jarvi et al 1999 Wittzell et al1999a Worley et al 2008) owls (Burri et al 2008) andparrots (Hughes et al 2008) Up to now the mostcomplicated MHC class I gene set has been described intwo species of passerines in the great reed warbler sevengenomic class I sequences were found in one individualone of which contained a frameshift mutation and washence nonfunctional (Westerdahl et al 1999) and eight
Table 2 Results of the likelihood ratio tests of positive selection on exon 3 of MHC class I in four genera of passerine birds
Genus Length Numbera M8 vs M7 Likelihood ratio test Number of positively selected sites
df Test statistic Significance PAML REL FEL
Carpodacus 91 29 2 578 plt0001 15 7 8
Acrocephalus 68 37 2 712 plt0001 8 14 4
Parus 69 24 2 472 plt0001 13 14 5
Passer 82 21 2 232 plt0001 2 2 1
Test statistic was computed as 2(LbminusLa) where La and Lb are log-likelihood values for each of the nested models being compared (M8 and M7respectively)
Length length of the aminoacid sequencesa Number of sequences in the analysed alignment
A - PAML 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ + B ndash REL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
C ndash FEL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
Fig 3 Comparison of results of three methods for detecting aminoacid sites under selection in four passerine species The analyses arebased on the alignments of 29 Rosefinch 37 warbler 24 tit and 21sparrow complete or partial sequences of exon 3 encoding alpha2domain of the protein Amino acid residues found to be under positiveselection by three methods (A PAML B REL C FEL) are bold (fordetails on the methods see the text) If the same residue was found tobe positively selected in at least two genera it is marked by rectangle
Regions excluded from analysis are marked by a squiggle Highlyconserved residues that anchor the peptide-terminal main-chain atomsare marked by crosses other residues forming variable part of peptide-binding region in HLA-A and chicken (Wallny et al 2006 Koch et al2007) are marked by asterisks The numbering of amino acid residuesis based on the complete MHC class I protein in the duck (Mesa et al2004) (Acrocephalus-AF449690) (Parus-AM232710) and (Passer-AY285012)
Immunogenetics (2009) 61451ndash461 457
different cDNA sequences were also found in an individualIn a separate study of the same species 20 genomicsequences of exon 3 of class I were found in fiveindividuals five of which were pseudogenes (Westerdahlet al 2004) Similar results were found in house sparrowswhere 20 sequences were verified in seven individualsthree of which were proved to be transcribed althoughthere were no pseudogenes (Bonneaud et al 2004) Ourresults concerning individual exon 3 variation are similar tothe ones mentioned above suggesting that complex MHCstructure could be a general pattern in songbirds
Evolution of MHC class I sequences in passerines
Several mechanisms of molecular evolution have beenalready documented in the avian MHC sequences includ-ing positive selection (Alcaide et al 2007 Hughes et al2008) concerted evolution (Wittzell et al 1999b) andtrans-species polymorphism (Richardson and Westerdahl2003 Alcaide et al 2007) In the present study model-based tests of selection (used here for the first time inpasserine species) have demonstrated the presence ofpositively selected sites in exon 3 of MHC class I in fourpasserine genera This part of the gene encodes the alpha 2domain of the protein which is included in forming thepeptide-binding groove Positive selection acting on somecodons of exon 3 is in accordance with published data onMHC class I in other birds where the ratio of non-synonymous to synonymous substitutions (dNdS) wasusually slightly higher at PBR sites than at non-PBR ones(Westerdahl et al 1999 Richardson and Westerdahl 2003Bonneaud et al 2004) The differences however were notsignificant and this could be caused either (1) by in-correctly identified PBR sites since the MHC proteinstructure in the majority of species studied is not well-understood In fact the PBR sites have usually beenidentified on the basis of the three-dimensional structureof human HLA-A protein (Bjorkman et al 1987) A modelof the three-dimensional structure of a passerine MHC classI molecule is highly desirable Or (2) another reason mightbe the fact that some of the studied sequences come fromloci that lost their importance and they are not underselection that is why it is crucial to identify sequences toparticular loci in the future studies (see Worley et al 2008)
Our results indicate extreme variability in selectivepressure among sites in the exon 3 and the presence of anumber of sites under balancing selection Approximatelyhalf of the positively selected sites in passerines (Fig 3)have been previously assumed to play a role in antigenbinding based on the structural models (Wallny et al 2006Koch et al 2007) However we found at least twoadditional sites (amino acid residues 126 and 136) understrong positive selection in several genera which have not
been previously reported for PBR (either in human HLA orchicken BF) The maintenance of considerable nonsynon-ymous variation at these specific codons by positiveselection suggests that such polymorphism may havefunctional consequences for peptide binding and presenta-tion to T cells (see similar recent results concerning teleostfish MHC molecules Aguilar and Garza 2007 or inmammals Amills et al 2008) The predictive power ofcodon-based models has not yet been tested in MHC andthe structural model describing peptide-binding groove inpasserine birds is still required to confirm the functionalrole of amino acid residues under different types ofselection
It appears that mammalian and avian MHCs may differin frequency of genetic exchange between different genesof multigene families Concerted evolution where the inter-locus gene conversion events are frequent in highlyduplicated MHC genes may be the predominant evolu-tionary mechanism in birds (Edwards et al 1995 Alcaide etal 2007 Garrigan and Edwards 1999 Wittzell et al 1999bHess and Edwards 2002) and probably also in fish (Reuschet al 2004 van Oosterhout et al 2006 Aguilar and Garza2007) and reptiles (Miller et al 2006) but not in mammals(Edwards et al 1997) This process creates high homologyamong MHC sequences derived from various loci and itbecomes impossible to assign different MHC alleles tospecific loci In birds so far most of the studies evidencinginter-locus recombination were conducted on MHC class IIConcerning MHC class I it has only been studied in thechicken where no evidence was found for concertedevolution to occur (Shaw et al 2007) Thus our resultson recombination analyses in the finches (ie the totalamount of recombination per codon was half of that ofsubstitution and we failed to assign the sequences tospecific loci) are in agreement with previous reports andfurthermore they suggest that concerted evolution may befrequent also in MHC class I genes in passerine birds Toconfirm this hypothesis the more detailed description ofMHC structure (especially the affiliation of particularalleles to individual loci in a population and the geneticvariation outside exon 3) is necessary
Extreme MHC class I variation in a single population
Previous studies on songbirds using the method ofdenaturing gradient gel electrophoresis or restriction frag-ment length polymorphism (RFLP) have also indicatedhigh sequence variation at exon 3 of MHC class I forexample in an outbred population of house sparrows 46sequences were found in 101 individuals or between oneand 11 alleles per individual (Bonneaud et al 2006a) in thegreat reed warbler 49 different RFLP genotypes werefound in 55 birds (Westerdahl et al 2004) Even so in the
458 Immunogenetics (2009) 61451ndash461
Scarlet Rosefinch 82 alleles were found in a singlepopulation and most of the birds displayed uniquegenotypes which is the highest exon 3 MHC class Ivariability observed thus far This result may still be anunderestimate considering that in at least one case theSSCP pattern was identical for two different allelesAlthough we were not able to sequence all the allelesdisplayed on SSCP it is very probable that all the peaksidentified by SSCP are true alleles and that there could beadditional ones (due to peak overlapping) Another reasonwe believe this variation has not been overestimated is thatan extraordinarily high variability of selectively neutralmarkers has been found in this species at two microsatelliteloci more than 70 alleles were found (Albrecht et al 2007)
Interestingly one of the alleles (CaerU02) was possessedby all individuals but one female (no 515) investigated inthis study From parentndashoffspring genotypes (Promerovaacute etal unpublished data) we know that also some of thisfemales offspring missed this allele thus this absence issurely heritable We conclude that CaerU02 is the mostfrequent allele on a locus with very low variability Thefemale no 515 is either homo- or heterozygote for other veryrare allele(s) or the locus (at least the copy with alleleCaerU02) is completely missing in her MHC haplotype
Our optimization of a genotyping method for analyses ofMHC class I variation will allow us to search for answers tofundamental questions concerning recent immunoecologysuch as the role of MHC in disease resistance expression ofsecondary ornaments and mating success (eg Bonneaud etal 2006a b Loiseau et al 2008 Richardson et al 2004Westerdahl et al 2005) Furthermore our data could beused for future comparative studies of the correlationbetween MHC diversity and migratory strategies in birdsWe presume that migratory birds such as the ScarletRosefinch (which migrates from the Czech Republic toIndia ca 5000 km) might encounter a larger variety ofpathogens during their lifetime than sedentary birds (forexample there is evidence that migrating individualsencounter more malarial parasites Peacuterez-Tris and Bensch2005) In that case more diverse immune mechanismscould help them fight these pathogens Moreover it hasalready been suggested that migratory species have highergenetic variability than sedentary species (Fitzpatrick 1994)and also that the higher genetic variability of migrants hasan impact on higher occurrence of extra-pair paternity(Spottiswoode and Moslashller 2004) The hypothesis thatmigration in birds might elicit a higher variability onMHC was first evaluated by Shiina et al (2004) whocompared the MHC of migratory quail with that ofsedentary chicken The quail has a more complicatedMHC than the chicken the class I region in the quail istwice as long as that of the chicken (Shiina et al 2004) andthere are five expressed class I genes although not all of
them are classical ones (Shiina et al 2006) SimilarlyRichardson and Westerdahl (2003) reported a difference inMHC variability between the migratory Great reed warblerand the sedentary inbred Seychelles warbler In this casehowever the low MHC variation in the latter could be dueto a severe bottleneck leading to strong decrease of overallgenetic variation Furthermore the endemic Basra reedwarbler (Acrocephalus griseldis) is as variable at MHC asthe Great reed warbler even though it is sedentary (Hanssonand Richardson 2005) High level of MHC polymorphismwas found in the house sparrow which is a non-migratoryspecies (Bonneaud et al 2004) which also contradicts thepredictions of the above hypothesis Unfortunately formost species (including the Scarlet Rosefinch) there is noprecise information about the functional variation ienumber of expressed MHC variants It is evident that dataon real MHC variation in more species are inevitablyrequired to perform a rigorous comparative meta-analysisand to test the predictions of migration-MHC hypothesis
Acknowledgements We thank Pavel Munclinger Michal VinklerJan Schnitzer and Radka Polaacutekovaacute for their help in the field Thanksto Anna Bryjovaacute and Dagmar Čiacutežkovaacute for their valuable advice inthe laboratory We are very grateful to Heidi C Hauffe for improvingthe English and two anonymous referees for critical comments on themanuscript This study was funded by the Grant Agency of theAcademy of Sciences of the Czech Republic (project noIAA600930608) the Grant Agency of the Czech Rep (project no206060851) and by the Ministry of Education of the Czech Repwhose Research Centre no LC06073 and Long-term Research Plan(no MSM 0021620828) to the Charles University in Prague formed aframework for a part of this study
References
Abbas AK Lichtman AH Pober JS (1994) Cellular and molecularimmunology W B Saunders Company Philadelphia
Aguilar A Garza JC (2007) Patterns of historical balancing selectionon the salmonid major histocompatibility complex class II betagene J Mol Evol 6534ndash43 doi101007s00239-006-0222-8
Albrecht T (2004) Edge effect in wetland-arable land boundarydetermines nesting success of scarlet rosefinches Carpodacuserythrinus in the Czech Republic Auk 121361ndash371doi1016420004-8038(2004)121[0361EEIWLB]20CO2
Albrecht T Schnitzer J Kreisinger J Exnerovaacute A Bryja JMunclinger P (2007) Extrapair paternity and the opportunity forsexual selection in long-distant migratory passerines Behav Ecol18477ndash486 doi101093behecoarm001
Alcaide M Edwards SV Negro JJ (2007) Characterization polymor-phism and evolution ofMHCClass II B genes in birds of prey JMolEvol 65541ndash554 doi101007s00239-007-9033-9
Amills M Ramiacuterez O Tomaacutes A Obexer-Ruff G Vidal O (2008)Positive selection on mammalian MHC-DQ genes revisited from amultispecies perspective Genes Immun 9651ndash658 doi101038gene200862
Anisimova M Nielsen R Yang Z (2003) Effect of recombination onthe accuracy of the likelihood method for detecting positiveselection at amino acid sites Genetics 1641229ndash1236
Immunogenetics (2009) 61451ndash461 459
Bjorkman PJ Saper MA Samraoui B Bennett WS Strominger JLWiley DC (1987) Structure of human class I histocompatibil-ity antigen HLA-A2 Nature 329506ndash512 doi101038329506a0
Bonneaud C Sorci G Morin V Westerdahl H Zoorob R Wittzell H(2004) Diversity of MHC class I and IIB genes in house sparrows(Passer domesticus) Immunogenetics 55855ndash865 doi101007s00251-004-0648-3
Bonneaud C Chastel O Federici P Westerdahl H Sorci G (2006a)Complex MHC-based mate choice in a wild passerine Proc RSoc 2731111ndash1116 doi101098rspb20053325
Bonneaud C Peacuterez-Tris J Federici P Chastel O Sorci G (2006b) Majorhistocompatibility alleles associated with local resistance to malariain a passerine Evolution Int J Org Evolution 60383ndash389
Bryja J Galan M Charbonnel N Cosson J-F (2005) Analysis ofmajor histocompatibility complex class II gene in water volesusing capillary electrophoresis-single stranded conformationpolymorphism Mol Ecol Notes 5173ndash176 doi101111j1471-8286200400855x
Burri R Niculita-Hirzel H Roulin A Fumagalli L (2008) Isolation andcharacterization of major histocompatibility complex (MHC) class IIB genes in teh Barn owl (Aves Tyto alba) Immunogenetics 60543ndash550 doi101007s00251-008-0308-0
Cramp S Simmons KEL Perrins CM (eds) (1977-1994) The birds ofthe Western Palearctic Oxford University Press Oxford
Edwards SV Wakeland EK Potts WK (1995) Contrasting histories ofavian and mammalian MHC genes revealed by class II B genesof songbirds Proc Natl Acad Sci U S A 9212200ndash12204doi101073pnas922612200
Edwards SV Chesnut K Satta Y Wakeland EK (1997) Ancestralpolymorphism of Mhc Class II genes in mice implications forbalancing selection and the mammalian molecular clock Genetics146655ndash668
Fitzpatrick S (1994) Colourful migratory birds evidence for amechanism other than parasite resistance for the maintenance oflsquogood genesrsquo sexual selection Proc R Soc Lond B Biol Sci257155ndash166 doi101098rspb19940109
Freeman-Gallant CR Johnson EM Saponara F Stanger M (2002)Variation at the major histocompatibility complex in Savannahsparrows Mol Ecol 111125ndash1130 doi101046j1365-294X200201508x
Garrigan D Edwards SV (1999) Polymorphism across an exon-intronboundary in an avian Mhc class II B gene Mol Biol Evol 161599ndash1606
Gouumly de Bellocq J Charbonnel N Morand S (2008) Coevolutionaryrelationship between helminth diversity and Mhc class II polymor-phism in rodents J Evol Biol 211144ndash1150 doi101111j1420-9101200801538x
Hall TA (1999) BioEdit a user-friendly biological sequence alignmenteditor and analisis program for Windows 9598NT NucleicAcids Symp Ser 4195ndash98
Hansson B Richardson DS (2005) Genetic variation in twoendangered Acrocephalus species compared to a widespreadcongener estimates based on functional and random loci AnimConserv 883ndash90 doi101017S1367943004001878
Hess CM Edwards SV (2002) The evolution of major histocompat-ibility genes in birds Bioscience 52423ndash431 doi1016410006-3568(2002)052[0423TEOTMH]20CO2
Hess CM Gasper J Hoekstra H Hill C Edwards SV (2000) MHCclass II pseudogene and genomic signature of a 32-kb cosmid inthe House Finch (Carpodacus mexicanus) Genome Res 1013ndash23 doi101101gr105613
Hosomichi K Shiina T Suzuki S Tanaka M Shimizu S Iwamoto SHara H Yoshida Y Kulski JK Inoko H Hanzawa K (2006) Themajor histocompatibility complex (Mhc) class IIB region hasgreater genomic structural flexibility and diversity in the quail
than the chicken BMC Genomics 7322ndash335 doi1011861471-2164-7-322
Hughes AL Yeager M (1998) Natural selection at major histocom-patibility complex loci of vertebrates Annu Rev Genet 32415ndash435 doi101146annurevgenet321415
Hughes CR Miles S Walbroehl JM (2008) Support for the minimalessential MHC hypothesis a parrot with a single highlypolymorphic MHC class II B gene Immunogenetics 60219ndash231 doi101007s00251-008-0287-1
Indovina P Megiorni F Fontemaggi G Coni P Mora B Mazzilli MC(2001) Absence of in vivo DNA-Protein interactions in theDQA2 and DQB2 promoter regions Hum Immunol 62504ndash508doi101016S0198-8859(01)00236-1
Jarvi SI Goto RM Gee GF Briles WE Miller MM (1999)Identification inheritance and linkage of B-G-like and MHCclass I genes in cranes Am Genet Assoc 90152ndash159
Kaufman J (1999) Co-evolving genes in MHC haplotypes the lsquorulersquofor nonmammalian vertebrates Immunogenetics 50228ndash236doi101007s002510050597
Kaufman J Salamonsen J Flajnik M (1994) Evolutionary conserva-tion of MHC class I and class II molecules-different yet the sameSemin Immunol 6411ndash424 doi101006smim19941050
Kaufman J Voumllk H Wallny H (1995) A bdquominimal essential MHCldquoand an bdquounrecognized MHCldquo two extremes in selection forpolymorphism Immunol Rev 14363ndash88 doi101111j1600-065X1995tb00670x
Kaufman J Jacob J Shaw I Walker B Milne S Beck S SalomonsenJ (1999) Gene organisation determines evolution of function inthe chicken MHC Immunol Rev 167101ndash117 doi101111j1600-065X1999tb01385x
Klein J (1986) Natural history of the major histocompatibilitycomplex Wiley New York
Klein J Sato A Nagl S Oh Uigin C (1998) Molecular trans-speciespolymorphism Annu Rev Ecol Syst 291ndash21 doi101146annurevecolsys2911
Koch M Camp S Collen T Avila D Salomonsen J Wallny HJ vanHateren A Hunt L Jacob JP Johnston F Marston DA Shaw IDunbar PR Cerundolo V Jones EY Kaufman J (2007)Structures of an MHC class I molecule from B21 chickensillustrate promiscuous peptide binding Immunity 27885ndash899doi101016jimmuni200711007
Kosakovsky Pond SL Frost SDW (2005) Not so different after all acomparison of methods for detecting amino acid sites under selectionMol Biol Evol 221208ndash1222 doi101093molbevmsi105
Kosakovsky Pond SL Frost SDW Muse SV (2005) HyPhyhypothesis testing using phylogenies Bioinformatics 21676ndash679 doi101093bioinformaticsbti079
Kosakovsky Pond SL Posada D Gravenor MB Woelk CH FrostSDW (2006) Automated phylogenetic detection of recombinationusing a genetic algorithm Mol Biol Evol 231891ndash1901doi101093molbevmsl051
Loiseau C Zoorob R Garnier S Birard J Federici P Julliard R SorciG (2008) Antagonistic effects of a MHC class I allele on malaria-infected house sparrows Ecol Lett 11258ndash265 doi101111j1461-0248200701141x
Mesa CM Thulien KJ Moon DA Veniamin SM Magor KE (2004)The dominant MHC class I gene is adjacent to the polymorphicTAP2 gene in the duck Anas platyrhynchos Immunogenetics56192ndash203 doi101007s00251-004-0672-3
Miller HC Lambert DM (2004) Gene duplication and geneconversion in class II MHC genes of New Zealand robins(Petroicidae) Immunogenetics 56178ndash191
Miller MM Goto RM Bernot A Zoorob R Auffray C Bumstead NBriles WE (1994) Two Mhc class I and two Mhc class II genes mapto the chicken Rfp-Y system outside the B complex Proc Natl AcadSci U S A 914397ndash4401 doi101073pnas91104397
460 Immunogenetics (2009) 61451ndash461
Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461
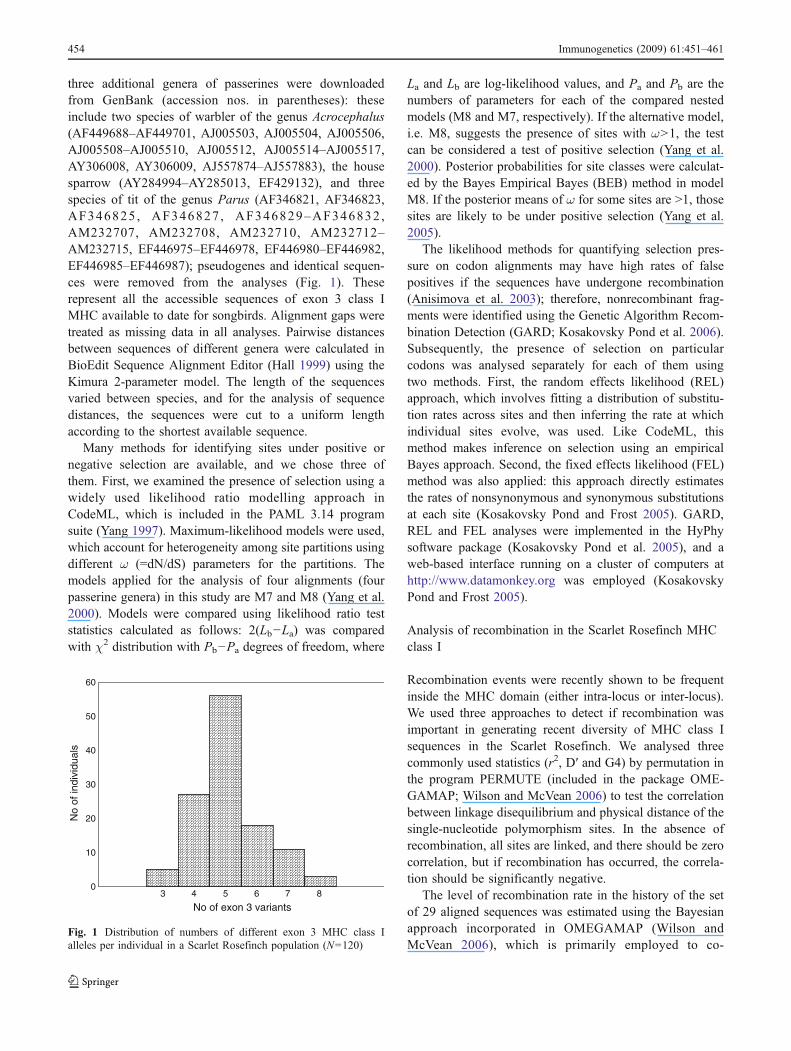
three additional genera of passerines were downloadedfrom GenBank (accession nos in parentheses) theseinclude two species of warbler of the genus Acrocephalus(AF449688ndashAF449701 AJ005503 AJ005504 AJ005506AJ005508ndashAJ005510 AJ005512 AJ005514ndashAJ005517AY306008 AY306009 AJ557874ndashAJ557883) the housesparrow (AY284994ndashAY285013 EF429132) and threespecies of tit of the genus Parus (AF346821 AF346823AF346825 AF346827 AF346829ndashAF346832 AM232707 AM232708 AM232710 AM232712ndashAM232715 EF446975ndashEF446978 EF446980ndashEF446982EF446985ndashEF446987) pseudogenes and identical sequen-ces were removed from the analyses (Fig 1) Theserepresent all the accessible sequences of exon 3 class IMHC available to date for songbirds Alignment gaps weretreated as missing data in all analyses Pairwise distancesbetween sequences of different genera were calculated inBioEdit Sequence Alignment Editor (Hall 1999) using theKimura 2-parameter model The length of the sequencesvaried between species and for the analysis of sequencedistances the sequences were cut to a uniform lengthaccording to the shortest available sequence
Many methods for identifying sites under positive ornegative selection are available and we chose three ofthem First we examined the presence of selection using awidely used likelihood ratio modelling approach inCodeML which is included in the PAML 314 programsuite (Yang 1997) Maximum-likelihood models were usedwhich account for heterogeneity among site partitions usingdifferent ω (=dNdS) parameters for the partitions Themodels applied for the analysis of four alignments (fourpasserine genera) in this study are M7 and M8 (Yang et al2000) Models were compared using likelihood ratio teststatistics calculated as follows 2(LbminusLa) was comparedwith χ2 distribution with PbminusPa degrees of freedom where
La and Lb are log-likelihood values and Pa and Pb are thenumbers of parameters for each of the compared nestedmodels (M8 and M7 respectively) If the alternative modelie M8 suggests the presence of sites with ωgt1 the testcan be considered a test of positive selection (Yang et al2000) Posterior probabilities for site classes were calculat-ed by the Bayes Empirical Bayes (BEB) method in modelM8 If the posterior means of ω for some sites are gt1 thosesites are likely to be under positive selection (Yang et al2005)
The likelihood methods for quantifying selection pres-sure on codon alignments may have high rates of falsepositives if the sequences have undergone recombination(Anisimova et al 2003) therefore nonrecombinant frag-ments were identified using the Genetic Algorithm Recom-bination Detection (GARD Kosakovsky Pond et al 2006)Subsequently the presence of selection on particularcodons was analysed separately for each of them usingtwo methods First the random effects likelihood (REL)approach which involves fitting a distribution of substitu-tion rates across sites and then inferring the rate at whichindividual sites evolve was used Like CodeML thismethod makes inference on selection using an empiricalBayes approach Second the fixed effects likelihood (FEL)method was also applied this approach directly estimatesthe rates of nonsynonymous and synonymous substitutionsat each site (Kosakovsky Pond and Frost 2005) GARDREL and FEL analyses were implemented in the HyPhysoftware package (Kosakovsky Pond et al 2005) and aweb-based interface running on a cluster of computers athttpwwwdatamonkeyorg was employed (KosakovskyPond and Frost 2005)
Analysis of recombination in the Scarlet Rosefinch MHCclass I
Recombination events were recently shown to be frequentinside the MHC domain (either intra-locus or inter-locus)We used three approaches to detect if recombination wasimportant in generating recent diversity of MHC class Isequences in the Scarlet Rosefinch We analysed threecommonly used statistics (r2 Dprime and G4) by permutation inthe program PERMUTE (included in the package OME-GAMAP Wilson and McVean 2006) to test the correlationbetween linkage disequilibrium and physical distance of thesingle-nucleotide polymorphism sites In the absence ofrecombination all sites are linked and there should be zerocorrelation but if recombination has occurred the correla-tion should be significantly negative
The level of recombination rate in the history of the setof 29 aligned sequences was estimated using the Bayesianapproach incorporated in OMEGAMAP (Wilson andMcVean 2006) which is primarily employed to co-
3 4 5 6 7 8
No of exon 3 variants
0
10
20
30
40
50
60
No
of in
divi
dual
s
Fig 1 Distribution of numbers of different exon 3 MHC class Ialleles per individual in a Scarlet Rosefinch population (N=120)
454 Immunogenetics (2009) 61451ndash461
estimate the selection parameter ω and the recombinationrate (ρ=4Nec) along the sequence
Results
MHC class I variation in the Scarlet Rosefinch
Amplification of genomic DNA with primers HN34 andCE-MHCREX3 gave a single PCR product of about 325 bpin all individuals including the whole exon 3 of 270 bp(273 bp in some alleles see below) and encoding thealpha2 domain (corresponding to part of the PBR and smallparts of adjacent introns) Among 120 adult Scarlet Rose-finches we identified 82 different variants of amplifiedMHC class I sequences using high-resolution capillarySSCP The SSCP profiles were highly repeatable andindividuals exhibited between three and nine variants (oneof these variants was impossible to distinguish by SSCPalthough it was repeatedly sequenced [see below]) Theseresults suggest that as many as five MHC class I genes arepresent in the Scarlet Rosefinch and the number ofamplified genes is variable between individuals
Due to the extremely high diversity of MHC class I inthe studied population not all variants were analysed indetail In total we obtained 29 sequences of exon 3 ofMHC class I genes by cloning and sequencing PCRproducts from 18 highly heterozygous individuals (identi-fied by SSCP) Each sequence was provided with thespecies abbreviation and a number (ie CaerU01-CaerU29) In this abbreviation lsquoUrsquo stands for theunknown locus designation of the gene (Mesa et al2004) In three individuals (ID 541 667 and 692) wemanaged to sequence all the variants identified by SSCP ofthese individuals (six four and eight alleles respectively)and all these sequences correspond to exon 3 of MHC classI when BLASTed (httpblastncbinlmnihgovBlastcgi)In total 597 clones were analysed by SSCP 156 of whichcontained either PCR artefacts or were contaminated byPCR product from the individual bird 55 contained noinsert and 122 contained the most frequent sequenceCaerU02 This variant of exon 3 occurred in all analysedadults except one female However the true number ofalleles is probably underestimated because some sequencevariants were difficult to distinguish by SSCP alleleCaerU11 was poorly visualised on SSCP because the6-FAM peak was very low and the NED peak wasoverlapped by CaerU02 (for this reason this allele mighthave been overlooked in some individuals and thereforewe do not have the precise number of individuals in whichthis allele occurred (Fig 2) even if it was repeatedlysequenced in four individuals) Most SSCP patterns werehighly specific in fact we found only one case in which
two different MHC sequences repeatedly confirmed bycloning and sequencing gave identical SSCP peaks incapillary electrophoresis (CaerU18 and CaerU19)
All 29 sequences were obtained from randomly chosenclones whose SSCP pattern matched exactly the SSCPpattern found in the profile of the individual from which theclone was isolated (to avoid sequencing PCR artefacts)Seventeen of these sequences were obtained from oneindividual each (seven of them were only present in oneindividual in the population) the remaining sequences wereidentified in clones originating from at least two differentbirds (two PCRs) These sequences translate into 28 uniqueamino acid sequences (CaerU02 and CaerU19 translateto identical alpha2 domains) and none of them containdeletions insertions or stop codons in the reading frameHowever four sequences have an insertion of 3 bp codingfor isoleucine (Fig 2) The pairwise distances between bothnucleotide and aminoacid sequences of the Scarlet Rose-finch and three other passerine genera (AcrocephalusPasser and Parus) chicken G gallus the Japanese quailC japonica and duck Anas platyrhynchos are shown inTable 1 (see also Fig 2)
Seven amino acid residues in the alpha2 domain offunctional MHC class I molecules are reported to be highlyconserved [Y123 T143 K146 W147 Y159 L160 Y171(Kaufman et al 1994 Shum et al 1999 Mesa et al 2004)]In all our Scarlet Rosefinch sequences four out of seven ofthese residues are conserved (W144W147 Y157Y159L158L160 Y169Y171 Fig 2) Y123 is not conserved inany avian sequence but the phenylalanine present in allRosefinch sequences (F123) is also reported in other non-mammalian species (Shum et al 1999 Mesa et al 2004)Threonine at position T140T143 is replaced by serine in theCaerU23 allele (S140T143) but both these residues belongto the same biochemical group of amino acids (ie theyshould substitute each other in peptide binding) The sameis true for K146 where lysine is replaced by arginine(R143K146) in all alleles (Fig 2)
Positive selection on MHC class I sequences in passerinebirds
Likelihood ratio tests comparing models M8 and M7 in thePAML package identified significant variability in selectivepressure among sites in exon 3 and the presence of two to15 sites under positive selection in four studied passerinespecies (Table 2) GARD analyses identified one significantglobal recombination in alignments of MHC sequencesfrom Carpodacus although no recombination point wasfound in Acrocephalus Parus or Passer REL and FELanalyses which incorporate information of recombinationprovided the evidence that two to 14 and one to eight sitesare under positive selection respectively (Table 2) The
Immunogenetics (2009) 61451ndash461 455
positively selected sites were often identical between thefour compared species as well as between the three methodsof analysis (Fig 3)
Recombination in MHC class I of the Scarlet Rosefinch
Significant negative correlation between physical distanceand linkage disequilibrium was detected using all threeanalyses in the program PERMUTE (plt0001) suggestingthat recombination regularly occurs in exon 3 of MHC classI genes The mean population recombination per codon (ρ=0233) identified in the program OMEGAMAP is approxi-mately half of the mean total population mutation (θ=0534)
again suggesting that recombination is important for gener-ating MHC class I variants in the Scarlet Rosefinch
Discussion
Structure of classical MHC class I genes in passerine birds
In this study the structure and diversity of MHC class I wasstudied in a wild-living songbird species Up to ninedifferent MHC class I sequences were present in oneScarlet Rosefinch individual which means that at least fiveamplified MHC class I genes could occur in a single bird
98 108 118 128 138 148 158 168 178 |||||||||||||||||| CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEG-EAEGWTNYLKHECPEWLQKYVGYGQKELER 1 CaerU12 R-VENVR 2 CaerU29 Y-VENV 1 CaerU05 RLRYP-PRRH 4 CaerU02 LWRLPKKPNSTGKIVRNVSRR 119CaerU17 LWLPKKPNSTGKIVRNVSRRERH 4 CaerU18 LWLPKKPNSTGKIVRNVSRR 32 CaerU08 FIQWLSVRRAFIED-FRR 27 CaerU16 FIQWLSVRRAFIEDK-FRR 1 CaerU22 FIQWLSVRRAFIEDK-FIRE 7 CaerU20 FIQWLSVDARFIP-VKR 19 CaerU06 WRSSEVYYRFIPDVKE-VFRE 51 CaerU07 WRSSEVYYWRFIPDVKE-VFRE 111CaerU11 WRSSEVYYRFIPDVKE-VFRER CaerU04 LWVRHNRFID-FIC 9 CaerU13 LRVRRARFIRDK-MRDV 1 CaerU14 LRVRRARFIRD-MRDV 1 CaerU27 WRVRRARFIDG-VRRDV 1 CaerU28 LRVRRARFIRDKE-VF 1 CaerU03 LNVRYNRFIPD-PRW 4 CaerU09 RNVRYNRFIELDKE-AF 11CaerU10 WRNVRYNRFIPDKE-VFIH 5 CaerU15 WRNVRYNRFIEPD-PRVI 6 CaerU23 LWNVRYNRFIPDSKE-VLH 11CaerU24 WRNVRYRFIPD-PRRVH 4 CaerU25 WRSNVRFYNRFIPDKE-VFDQH 3 CaerU21 LWNVRDARFIDNGKE-VF 7 CaerU26 LRNVRDARFIDNGK-VEMRE 10Gallus gallus SVQWMIEGPRYYQMARFTAKTMTTVPEVPKKES-PRKEETVRREKAG Coturnix japonica SVQWMIDTTRYNQYARFIVKDTMTTVPEVPKKEGD-YRQKHEETVQRRHENKAG Anas platyrhynchos SVQRMKRFEQYERFIALKDTLTTDAQKKETYRTKYENTIRSKDV + + ++ ++ +
Fig 2 Amino acid sequences encoded by exon 3 in the ScarletRosefinch corresponding to alpha2 domain of the protein comparedwith GenBank sequences from chicken (X12780) quail (D29813) andduck (AY294416) Identity with the Scarlet Rosefinch allele CaerU1is indicated by dots gaps which are added to get the best alignment bydashes Highly conserved residues that anchor the peptide-terminalmain-chain atoms are marked by crosses other residues forming
variable part of peptide-binding region in HLA-A and chicken(Wallny et al 2006 Koch et al 2007) are marked by asterisks Thenumbering of amino acid residues is based on the complete MHCclass I protein in the duck (Mesa et al 2004) The numbers on theright show the number of individuals with the particular sequence (N=120) question mark the precise number of individuals bearing theallele CaerU11 is not known (see the text for more details)
Genus Numbera Amino acid sequence distance plusmn SD Nucleotide sequence distance plusmn SD
Acrocephalus 37 0514plusmn0189 0185plusmn0053
Parus 24 0734plusmn0167 0260plusmn00483
Passer 21 0538plusmn0154 0187plusmn0040
Gallus 1 1390plusmn0102 0689plusmn0040
Coturnix 1 1618plusmn0149 0739plusmn0050
Anas 1 1247plusmn0122 0607plusmn0037
Table 1 Average distancesbetween the 29 Scarlet Rose-finch sequences and sequencesof six other bird species
a Number of sequences in theanalysed alignment
456 Immunogenetics (2009) 61451ndash461
Although gene expression analysis was not performedstrong correlation between signatures for balancing selec-tion and expression is suggested Furthermore the presenceof conserved sites that anchor the peptide-terminal main-chain atoms and a reading frame with no terminal codons orframeshift mutations suggest that possibly all sequences ofMHC class I described in this study might be functionalalthough further study at the level of cDNA is required toconfirm this There is an increasing number of studiesshowing that even nearly intact MHC sequences are notexpressed (eg Indovina et al 2001 Bonneaud et al 2004Shiina et al 2006) Hence MHC class I genes seem to behighly variable in the Scarlet Rosefinch and this compli-cated and complex structure is very different from non-
passerines For example the best described MHC in birdsthe chicken lsquominimal essentialrsquo MHC has very few buthighly condensed genes (eg there are two classical MHCclass I genes but only a single dominantly expressedKaufman et al 1999) it appears that this is the ancestralstate for birds since it occurs in various groups such asother gallinaceous birds (Jarvi et al 1999 Wittzell et al1999a Worley et al 2008) owls (Burri et al 2008) andparrots (Hughes et al 2008) Up to now the mostcomplicated MHC class I gene set has been described intwo species of passerines in the great reed warbler sevengenomic class I sequences were found in one individualone of which contained a frameshift mutation and washence nonfunctional (Westerdahl et al 1999) and eight
Table 2 Results of the likelihood ratio tests of positive selection on exon 3 of MHC class I in four genera of passerine birds
Genus Length Numbera M8 vs M7 Likelihood ratio test Number of positively selected sites
df Test statistic Significance PAML REL FEL
Carpodacus 91 29 2 578 plt0001 15 7 8
Acrocephalus 68 37 2 712 plt0001 8 14 4
Parus 69 24 2 472 plt0001 13 14 5
Passer 82 21 2 232 plt0001 2 2 1
Test statistic was computed as 2(LbminusLa) where La and Lb are log-likelihood values for each of the nested models being compared (M8 and M7respectively)
Length length of the aminoacid sequencesa Number of sequences in the analysed alignment
A - PAML 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ + B ndash REL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
C ndash FEL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
Fig 3 Comparison of results of three methods for detecting aminoacid sites under selection in four passerine species The analyses arebased on the alignments of 29 Rosefinch 37 warbler 24 tit and 21sparrow complete or partial sequences of exon 3 encoding alpha2domain of the protein Amino acid residues found to be under positiveselection by three methods (A PAML B REL C FEL) are bold (fordetails on the methods see the text) If the same residue was found tobe positively selected in at least two genera it is marked by rectangle
Regions excluded from analysis are marked by a squiggle Highlyconserved residues that anchor the peptide-terminal main-chain atomsare marked by crosses other residues forming variable part of peptide-binding region in HLA-A and chicken (Wallny et al 2006 Koch et al2007) are marked by asterisks The numbering of amino acid residuesis based on the complete MHC class I protein in the duck (Mesa et al2004) (Acrocephalus-AF449690) (Parus-AM232710) and (Passer-AY285012)
Immunogenetics (2009) 61451ndash461 457
different cDNA sequences were also found in an individualIn a separate study of the same species 20 genomicsequences of exon 3 of class I were found in fiveindividuals five of which were pseudogenes (Westerdahlet al 2004) Similar results were found in house sparrowswhere 20 sequences were verified in seven individualsthree of which were proved to be transcribed althoughthere were no pseudogenes (Bonneaud et al 2004) Ourresults concerning individual exon 3 variation are similar tothe ones mentioned above suggesting that complex MHCstructure could be a general pattern in songbirds
Evolution of MHC class I sequences in passerines
Several mechanisms of molecular evolution have beenalready documented in the avian MHC sequences includ-ing positive selection (Alcaide et al 2007 Hughes et al2008) concerted evolution (Wittzell et al 1999b) andtrans-species polymorphism (Richardson and Westerdahl2003 Alcaide et al 2007) In the present study model-based tests of selection (used here for the first time inpasserine species) have demonstrated the presence ofpositively selected sites in exon 3 of MHC class I in fourpasserine genera This part of the gene encodes the alpha 2domain of the protein which is included in forming thepeptide-binding groove Positive selection acting on somecodons of exon 3 is in accordance with published data onMHC class I in other birds where the ratio of non-synonymous to synonymous substitutions (dNdS) wasusually slightly higher at PBR sites than at non-PBR ones(Westerdahl et al 1999 Richardson and Westerdahl 2003Bonneaud et al 2004) The differences however were notsignificant and this could be caused either (1) by in-correctly identified PBR sites since the MHC proteinstructure in the majority of species studied is not well-understood In fact the PBR sites have usually beenidentified on the basis of the three-dimensional structureof human HLA-A protein (Bjorkman et al 1987) A modelof the three-dimensional structure of a passerine MHC classI molecule is highly desirable Or (2) another reason mightbe the fact that some of the studied sequences come fromloci that lost their importance and they are not underselection that is why it is crucial to identify sequences toparticular loci in the future studies (see Worley et al 2008)
Our results indicate extreme variability in selectivepressure among sites in the exon 3 and the presence of anumber of sites under balancing selection Approximatelyhalf of the positively selected sites in passerines (Fig 3)have been previously assumed to play a role in antigenbinding based on the structural models (Wallny et al 2006Koch et al 2007) However we found at least twoadditional sites (amino acid residues 126 and 136) understrong positive selection in several genera which have not
been previously reported for PBR (either in human HLA orchicken BF) The maintenance of considerable nonsynon-ymous variation at these specific codons by positiveselection suggests that such polymorphism may havefunctional consequences for peptide binding and presenta-tion to T cells (see similar recent results concerning teleostfish MHC molecules Aguilar and Garza 2007 or inmammals Amills et al 2008) The predictive power ofcodon-based models has not yet been tested in MHC andthe structural model describing peptide-binding groove inpasserine birds is still required to confirm the functionalrole of amino acid residues under different types ofselection
It appears that mammalian and avian MHCs may differin frequency of genetic exchange between different genesof multigene families Concerted evolution where the inter-locus gene conversion events are frequent in highlyduplicated MHC genes may be the predominant evolu-tionary mechanism in birds (Edwards et al 1995 Alcaide etal 2007 Garrigan and Edwards 1999 Wittzell et al 1999bHess and Edwards 2002) and probably also in fish (Reuschet al 2004 van Oosterhout et al 2006 Aguilar and Garza2007) and reptiles (Miller et al 2006) but not in mammals(Edwards et al 1997) This process creates high homologyamong MHC sequences derived from various loci and itbecomes impossible to assign different MHC alleles tospecific loci In birds so far most of the studies evidencinginter-locus recombination were conducted on MHC class IIConcerning MHC class I it has only been studied in thechicken where no evidence was found for concertedevolution to occur (Shaw et al 2007) Thus our resultson recombination analyses in the finches (ie the totalamount of recombination per codon was half of that ofsubstitution and we failed to assign the sequences tospecific loci) are in agreement with previous reports andfurthermore they suggest that concerted evolution may befrequent also in MHC class I genes in passerine birds Toconfirm this hypothesis the more detailed description ofMHC structure (especially the affiliation of particularalleles to individual loci in a population and the geneticvariation outside exon 3) is necessary
Extreme MHC class I variation in a single population
Previous studies on songbirds using the method ofdenaturing gradient gel electrophoresis or restriction frag-ment length polymorphism (RFLP) have also indicatedhigh sequence variation at exon 3 of MHC class I forexample in an outbred population of house sparrows 46sequences were found in 101 individuals or between oneand 11 alleles per individual (Bonneaud et al 2006a) in thegreat reed warbler 49 different RFLP genotypes werefound in 55 birds (Westerdahl et al 2004) Even so in the
458 Immunogenetics (2009) 61451ndash461
Scarlet Rosefinch 82 alleles were found in a singlepopulation and most of the birds displayed uniquegenotypes which is the highest exon 3 MHC class Ivariability observed thus far This result may still be anunderestimate considering that in at least one case theSSCP pattern was identical for two different allelesAlthough we were not able to sequence all the allelesdisplayed on SSCP it is very probable that all the peaksidentified by SSCP are true alleles and that there could beadditional ones (due to peak overlapping) Another reasonwe believe this variation has not been overestimated is thatan extraordinarily high variability of selectively neutralmarkers has been found in this species at two microsatelliteloci more than 70 alleles were found (Albrecht et al 2007)
Interestingly one of the alleles (CaerU02) was possessedby all individuals but one female (no 515) investigated inthis study From parentndashoffspring genotypes (Promerovaacute etal unpublished data) we know that also some of thisfemales offspring missed this allele thus this absence issurely heritable We conclude that CaerU02 is the mostfrequent allele on a locus with very low variability Thefemale no 515 is either homo- or heterozygote for other veryrare allele(s) or the locus (at least the copy with alleleCaerU02) is completely missing in her MHC haplotype
Our optimization of a genotyping method for analyses ofMHC class I variation will allow us to search for answers tofundamental questions concerning recent immunoecologysuch as the role of MHC in disease resistance expression ofsecondary ornaments and mating success (eg Bonneaud etal 2006a b Loiseau et al 2008 Richardson et al 2004Westerdahl et al 2005) Furthermore our data could beused for future comparative studies of the correlationbetween MHC diversity and migratory strategies in birdsWe presume that migratory birds such as the ScarletRosefinch (which migrates from the Czech Republic toIndia ca 5000 km) might encounter a larger variety ofpathogens during their lifetime than sedentary birds (forexample there is evidence that migrating individualsencounter more malarial parasites Peacuterez-Tris and Bensch2005) In that case more diverse immune mechanismscould help them fight these pathogens Moreover it hasalready been suggested that migratory species have highergenetic variability than sedentary species (Fitzpatrick 1994)and also that the higher genetic variability of migrants hasan impact on higher occurrence of extra-pair paternity(Spottiswoode and Moslashller 2004) The hypothesis thatmigration in birds might elicit a higher variability onMHC was first evaluated by Shiina et al (2004) whocompared the MHC of migratory quail with that ofsedentary chicken The quail has a more complicatedMHC than the chicken the class I region in the quail istwice as long as that of the chicken (Shiina et al 2004) andthere are five expressed class I genes although not all of
them are classical ones (Shiina et al 2006) SimilarlyRichardson and Westerdahl (2003) reported a difference inMHC variability between the migratory Great reed warblerand the sedentary inbred Seychelles warbler In this casehowever the low MHC variation in the latter could be dueto a severe bottleneck leading to strong decrease of overallgenetic variation Furthermore the endemic Basra reedwarbler (Acrocephalus griseldis) is as variable at MHC asthe Great reed warbler even though it is sedentary (Hanssonand Richardson 2005) High level of MHC polymorphismwas found in the house sparrow which is a non-migratoryspecies (Bonneaud et al 2004) which also contradicts thepredictions of the above hypothesis Unfortunately formost species (including the Scarlet Rosefinch) there is noprecise information about the functional variation ienumber of expressed MHC variants It is evident that dataon real MHC variation in more species are inevitablyrequired to perform a rigorous comparative meta-analysisand to test the predictions of migration-MHC hypothesis
Acknowledgements We thank Pavel Munclinger Michal VinklerJan Schnitzer and Radka Polaacutekovaacute for their help in the field Thanksto Anna Bryjovaacute and Dagmar Čiacutežkovaacute for their valuable advice inthe laboratory We are very grateful to Heidi C Hauffe for improvingthe English and two anonymous referees for critical comments on themanuscript This study was funded by the Grant Agency of theAcademy of Sciences of the Czech Republic (project noIAA600930608) the Grant Agency of the Czech Rep (project no206060851) and by the Ministry of Education of the Czech Repwhose Research Centre no LC06073 and Long-term Research Plan(no MSM 0021620828) to the Charles University in Prague formed aframework for a part of this study
References
Abbas AK Lichtman AH Pober JS (1994) Cellular and molecularimmunology W B Saunders Company Philadelphia
Aguilar A Garza JC (2007) Patterns of historical balancing selectionon the salmonid major histocompatibility complex class II betagene J Mol Evol 6534ndash43 doi101007s00239-006-0222-8
Albrecht T (2004) Edge effect in wetland-arable land boundarydetermines nesting success of scarlet rosefinches Carpodacuserythrinus in the Czech Republic Auk 121361ndash371doi1016420004-8038(2004)121[0361EEIWLB]20CO2
Albrecht T Schnitzer J Kreisinger J Exnerovaacute A Bryja JMunclinger P (2007) Extrapair paternity and the opportunity forsexual selection in long-distant migratory passerines Behav Ecol18477ndash486 doi101093behecoarm001
Alcaide M Edwards SV Negro JJ (2007) Characterization polymor-phism and evolution ofMHCClass II B genes in birds of prey JMolEvol 65541ndash554 doi101007s00239-007-9033-9
Amills M Ramiacuterez O Tomaacutes A Obexer-Ruff G Vidal O (2008)Positive selection on mammalian MHC-DQ genes revisited from amultispecies perspective Genes Immun 9651ndash658 doi101038gene200862
Anisimova M Nielsen R Yang Z (2003) Effect of recombination onthe accuracy of the likelihood method for detecting positiveselection at amino acid sites Genetics 1641229ndash1236
Immunogenetics (2009) 61451ndash461 459
Bjorkman PJ Saper MA Samraoui B Bennett WS Strominger JLWiley DC (1987) Structure of human class I histocompatibil-ity antigen HLA-A2 Nature 329506ndash512 doi101038329506a0
Bonneaud C Sorci G Morin V Westerdahl H Zoorob R Wittzell H(2004) Diversity of MHC class I and IIB genes in house sparrows(Passer domesticus) Immunogenetics 55855ndash865 doi101007s00251-004-0648-3
Bonneaud C Chastel O Federici P Westerdahl H Sorci G (2006a)Complex MHC-based mate choice in a wild passerine Proc RSoc 2731111ndash1116 doi101098rspb20053325
Bonneaud C Peacuterez-Tris J Federici P Chastel O Sorci G (2006b) Majorhistocompatibility alleles associated with local resistance to malariain a passerine Evolution Int J Org Evolution 60383ndash389
Bryja J Galan M Charbonnel N Cosson J-F (2005) Analysis ofmajor histocompatibility complex class II gene in water volesusing capillary electrophoresis-single stranded conformationpolymorphism Mol Ecol Notes 5173ndash176 doi101111j1471-8286200400855x
Burri R Niculita-Hirzel H Roulin A Fumagalli L (2008) Isolation andcharacterization of major histocompatibility complex (MHC) class IIB genes in teh Barn owl (Aves Tyto alba) Immunogenetics 60543ndash550 doi101007s00251-008-0308-0
Cramp S Simmons KEL Perrins CM (eds) (1977-1994) The birds ofthe Western Palearctic Oxford University Press Oxford
Edwards SV Wakeland EK Potts WK (1995) Contrasting histories ofavian and mammalian MHC genes revealed by class II B genesof songbirds Proc Natl Acad Sci U S A 9212200ndash12204doi101073pnas922612200
Edwards SV Chesnut K Satta Y Wakeland EK (1997) Ancestralpolymorphism of Mhc Class II genes in mice implications forbalancing selection and the mammalian molecular clock Genetics146655ndash668
Fitzpatrick S (1994) Colourful migratory birds evidence for amechanism other than parasite resistance for the maintenance oflsquogood genesrsquo sexual selection Proc R Soc Lond B Biol Sci257155ndash166 doi101098rspb19940109
Freeman-Gallant CR Johnson EM Saponara F Stanger M (2002)Variation at the major histocompatibility complex in Savannahsparrows Mol Ecol 111125ndash1130 doi101046j1365-294X200201508x
Garrigan D Edwards SV (1999) Polymorphism across an exon-intronboundary in an avian Mhc class II B gene Mol Biol Evol 161599ndash1606
Gouumly de Bellocq J Charbonnel N Morand S (2008) Coevolutionaryrelationship between helminth diversity and Mhc class II polymor-phism in rodents J Evol Biol 211144ndash1150 doi101111j1420-9101200801538x
Hall TA (1999) BioEdit a user-friendly biological sequence alignmenteditor and analisis program for Windows 9598NT NucleicAcids Symp Ser 4195ndash98
Hansson B Richardson DS (2005) Genetic variation in twoendangered Acrocephalus species compared to a widespreadcongener estimates based on functional and random loci AnimConserv 883ndash90 doi101017S1367943004001878
Hess CM Edwards SV (2002) The evolution of major histocompat-ibility genes in birds Bioscience 52423ndash431 doi1016410006-3568(2002)052[0423TEOTMH]20CO2
Hess CM Gasper J Hoekstra H Hill C Edwards SV (2000) MHCclass II pseudogene and genomic signature of a 32-kb cosmid inthe House Finch (Carpodacus mexicanus) Genome Res 1013ndash23 doi101101gr105613
Hosomichi K Shiina T Suzuki S Tanaka M Shimizu S Iwamoto SHara H Yoshida Y Kulski JK Inoko H Hanzawa K (2006) Themajor histocompatibility complex (Mhc) class IIB region hasgreater genomic structural flexibility and diversity in the quail
than the chicken BMC Genomics 7322ndash335 doi1011861471-2164-7-322
Hughes AL Yeager M (1998) Natural selection at major histocom-patibility complex loci of vertebrates Annu Rev Genet 32415ndash435 doi101146annurevgenet321415
Hughes CR Miles S Walbroehl JM (2008) Support for the minimalessential MHC hypothesis a parrot with a single highlypolymorphic MHC class II B gene Immunogenetics 60219ndash231 doi101007s00251-008-0287-1
Indovina P Megiorni F Fontemaggi G Coni P Mora B Mazzilli MC(2001) Absence of in vivo DNA-Protein interactions in theDQA2 and DQB2 promoter regions Hum Immunol 62504ndash508doi101016S0198-8859(01)00236-1
Jarvi SI Goto RM Gee GF Briles WE Miller MM (1999)Identification inheritance and linkage of B-G-like and MHCclass I genes in cranes Am Genet Assoc 90152ndash159
Kaufman J (1999) Co-evolving genes in MHC haplotypes the lsquorulersquofor nonmammalian vertebrates Immunogenetics 50228ndash236doi101007s002510050597
Kaufman J Salamonsen J Flajnik M (1994) Evolutionary conserva-tion of MHC class I and class II molecules-different yet the sameSemin Immunol 6411ndash424 doi101006smim19941050
Kaufman J Voumllk H Wallny H (1995) A bdquominimal essential MHCldquoand an bdquounrecognized MHCldquo two extremes in selection forpolymorphism Immunol Rev 14363ndash88 doi101111j1600-065X1995tb00670x
Kaufman J Jacob J Shaw I Walker B Milne S Beck S SalomonsenJ (1999) Gene organisation determines evolution of function inthe chicken MHC Immunol Rev 167101ndash117 doi101111j1600-065X1999tb01385x
Klein J (1986) Natural history of the major histocompatibilitycomplex Wiley New York
Klein J Sato A Nagl S Oh Uigin C (1998) Molecular trans-speciespolymorphism Annu Rev Ecol Syst 291ndash21 doi101146annurevecolsys2911
Koch M Camp S Collen T Avila D Salomonsen J Wallny HJ vanHateren A Hunt L Jacob JP Johnston F Marston DA Shaw IDunbar PR Cerundolo V Jones EY Kaufman J (2007)Structures of an MHC class I molecule from B21 chickensillustrate promiscuous peptide binding Immunity 27885ndash899doi101016jimmuni200711007
Kosakovsky Pond SL Frost SDW (2005) Not so different after all acomparison of methods for detecting amino acid sites under selectionMol Biol Evol 221208ndash1222 doi101093molbevmsi105
Kosakovsky Pond SL Frost SDW Muse SV (2005) HyPhyhypothesis testing using phylogenies Bioinformatics 21676ndash679 doi101093bioinformaticsbti079
Kosakovsky Pond SL Posada D Gravenor MB Woelk CH FrostSDW (2006) Automated phylogenetic detection of recombinationusing a genetic algorithm Mol Biol Evol 231891ndash1901doi101093molbevmsl051
Loiseau C Zoorob R Garnier S Birard J Federici P Julliard R SorciG (2008) Antagonistic effects of a MHC class I allele on malaria-infected house sparrows Ecol Lett 11258ndash265 doi101111j1461-0248200701141x
Mesa CM Thulien KJ Moon DA Veniamin SM Magor KE (2004)The dominant MHC class I gene is adjacent to the polymorphicTAP2 gene in the duck Anas platyrhynchos Immunogenetics56192ndash203 doi101007s00251-004-0672-3
Miller HC Lambert DM (2004) Gene duplication and geneconversion in class II MHC genes of New Zealand robins(Petroicidae) Immunogenetics 56178ndash191
Miller MM Goto RM Bernot A Zoorob R Auffray C Bumstead NBriles WE (1994) Two Mhc class I and two Mhc class II genes mapto the chicken Rfp-Y system outside the B complex Proc Natl AcadSci U S A 914397ndash4401 doi101073pnas91104397
460 Immunogenetics (2009) 61451ndash461
Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461

estimate the selection parameter ω and the recombinationrate (ρ=4Nec) along the sequence
Results
MHC class I variation in the Scarlet Rosefinch
Amplification of genomic DNA with primers HN34 andCE-MHCREX3 gave a single PCR product of about 325 bpin all individuals including the whole exon 3 of 270 bp(273 bp in some alleles see below) and encoding thealpha2 domain (corresponding to part of the PBR and smallparts of adjacent introns) Among 120 adult Scarlet Rose-finches we identified 82 different variants of amplifiedMHC class I sequences using high-resolution capillarySSCP The SSCP profiles were highly repeatable andindividuals exhibited between three and nine variants (oneof these variants was impossible to distinguish by SSCPalthough it was repeatedly sequenced [see below]) Theseresults suggest that as many as five MHC class I genes arepresent in the Scarlet Rosefinch and the number ofamplified genes is variable between individuals
Due to the extremely high diversity of MHC class I inthe studied population not all variants were analysed indetail In total we obtained 29 sequences of exon 3 ofMHC class I genes by cloning and sequencing PCRproducts from 18 highly heterozygous individuals (identi-fied by SSCP) Each sequence was provided with thespecies abbreviation and a number (ie CaerU01-CaerU29) In this abbreviation lsquoUrsquo stands for theunknown locus designation of the gene (Mesa et al2004) In three individuals (ID 541 667 and 692) wemanaged to sequence all the variants identified by SSCP ofthese individuals (six four and eight alleles respectively)and all these sequences correspond to exon 3 of MHC classI when BLASTed (httpblastncbinlmnihgovBlastcgi)In total 597 clones were analysed by SSCP 156 of whichcontained either PCR artefacts or were contaminated byPCR product from the individual bird 55 contained noinsert and 122 contained the most frequent sequenceCaerU02 This variant of exon 3 occurred in all analysedadults except one female However the true number ofalleles is probably underestimated because some sequencevariants were difficult to distinguish by SSCP alleleCaerU11 was poorly visualised on SSCP because the6-FAM peak was very low and the NED peak wasoverlapped by CaerU02 (for this reason this allele mighthave been overlooked in some individuals and thereforewe do not have the precise number of individuals in whichthis allele occurred (Fig 2) even if it was repeatedlysequenced in four individuals) Most SSCP patterns werehighly specific in fact we found only one case in which
two different MHC sequences repeatedly confirmed bycloning and sequencing gave identical SSCP peaks incapillary electrophoresis (CaerU18 and CaerU19)
All 29 sequences were obtained from randomly chosenclones whose SSCP pattern matched exactly the SSCPpattern found in the profile of the individual from which theclone was isolated (to avoid sequencing PCR artefacts)Seventeen of these sequences were obtained from oneindividual each (seven of them were only present in oneindividual in the population) the remaining sequences wereidentified in clones originating from at least two differentbirds (two PCRs) These sequences translate into 28 uniqueamino acid sequences (CaerU02 and CaerU19 translateto identical alpha2 domains) and none of them containdeletions insertions or stop codons in the reading frameHowever four sequences have an insertion of 3 bp codingfor isoleucine (Fig 2) The pairwise distances between bothnucleotide and aminoacid sequences of the Scarlet Rose-finch and three other passerine genera (AcrocephalusPasser and Parus) chicken G gallus the Japanese quailC japonica and duck Anas platyrhynchos are shown inTable 1 (see also Fig 2)
Seven amino acid residues in the alpha2 domain offunctional MHC class I molecules are reported to be highlyconserved [Y123 T143 K146 W147 Y159 L160 Y171(Kaufman et al 1994 Shum et al 1999 Mesa et al 2004)]In all our Scarlet Rosefinch sequences four out of seven ofthese residues are conserved (W144W147 Y157Y159L158L160 Y169Y171 Fig 2) Y123 is not conserved inany avian sequence but the phenylalanine present in allRosefinch sequences (F123) is also reported in other non-mammalian species (Shum et al 1999 Mesa et al 2004)Threonine at position T140T143 is replaced by serine in theCaerU23 allele (S140T143) but both these residues belongto the same biochemical group of amino acids (ie theyshould substitute each other in peptide binding) The sameis true for K146 where lysine is replaced by arginine(R143K146) in all alleles (Fig 2)
Positive selection on MHC class I sequences in passerinebirds
Likelihood ratio tests comparing models M8 and M7 in thePAML package identified significant variability in selectivepressure among sites in exon 3 and the presence of two to15 sites under positive selection in four studied passerinespecies (Table 2) GARD analyses identified one significantglobal recombination in alignments of MHC sequencesfrom Carpodacus although no recombination point wasfound in Acrocephalus Parus or Passer REL and FELanalyses which incorporate information of recombinationprovided the evidence that two to 14 and one to eight sitesare under positive selection respectively (Table 2) The
Immunogenetics (2009) 61451ndash461 455
positively selected sites were often identical between thefour compared species as well as between the three methodsof analysis (Fig 3)
Recombination in MHC class I of the Scarlet Rosefinch
Significant negative correlation between physical distanceand linkage disequilibrium was detected using all threeanalyses in the program PERMUTE (plt0001) suggestingthat recombination regularly occurs in exon 3 of MHC classI genes The mean population recombination per codon (ρ=0233) identified in the program OMEGAMAP is approxi-mately half of the mean total population mutation (θ=0534)
again suggesting that recombination is important for gener-ating MHC class I variants in the Scarlet Rosefinch
Discussion
Structure of classical MHC class I genes in passerine birds
In this study the structure and diversity of MHC class I wasstudied in a wild-living songbird species Up to ninedifferent MHC class I sequences were present in oneScarlet Rosefinch individual which means that at least fiveamplified MHC class I genes could occur in a single bird
98 108 118 128 138 148 158 168 178 |||||||||||||||||| CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEG-EAEGWTNYLKHECPEWLQKYVGYGQKELER 1 CaerU12 R-VENVR 2 CaerU29 Y-VENV 1 CaerU05 RLRYP-PRRH 4 CaerU02 LWRLPKKPNSTGKIVRNVSRR 119CaerU17 LWLPKKPNSTGKIVRNVSRRERH 4 CaerU18 LWLPKKPNSTGKIVRNVSRR 32 CaerU08 FIQWLSVRRAFIED-FRR 27 CaerU16 FIQWLSVRRAFIEDK-FRR 1 CaerU22 FIQWLSVRRAFIEDK-FIRE 7 CaerU20 FIQWLSVDARFIP-VKR 19 CaerU06 WRSSEVYYRFIPDVKE-VFRE 51 CaerU07 WRSSEVYYWRFIPDVKE-VFRE 111CaerU11 WRSSEVYYRFIPDVKE-VFRER CaerU04 LWVRHNRFID-FIC 9 CaerU13 LRVRRARFIRDK-MRDV 1 CaerU14 LRVRRARFIRD-MRDV 1 CaerU27 WRVRRARFIDG-VRRDV 1 CaerU28 LRVRRARFIRDKE-VF 1 CaerU03 LNVRYNRFIPD-PRW 4 CaerU09 RNVRYNRFIELDKE-AF 11CaerU10 WRNVRYNRFIPDKE-VFIH 5 CaerU15 WRNVRYNRFIEPD-PRVI 6 CaerU23 LWNVRYNRFIPDSKE-VLH 11CaerU24 WRNVRYRFIPD-PRRVH 4 CaerU25 WRSNVRFYNRFIPDKE-VFDQH 3 CaerU21 LWNVRDARFIDNGKE-VF 7 CaerU26 LRNVRDARFIDNGK-VEMRE 10Gallus gallus SVQWMIEGPRYYQMARFTAKTMTTVPEVPKKES-PRKEETVRREKAG Coturnix japonica SVQWMIDTTRYNQYARFIVKDTMTTVPEVPKKEGD-YRQKHEETVQRRHENKAG Anas platyrhynchos SVQRMKRFEQYERFIALKDTLTTDAQKKETYRTKYENTIRSKDV + + ++ ++ +
Fig 2 Amino acid sequences encoded by exon 3 in the ScarletRosefinch corresponding to alpha2 domain of the protein comparedwith GenBank sequences from chicken (X12780) quail (D29813) andduck (AY294416) Identity with the Scarlet Rosefinch allele CaerU1is indicated by dots gaps which are added to get the best alignment bydashes Highly conserved residues that anchor the peptide-terminalmain-chain atoms are marked by crosses other residues forming
variable part of peptide-binding region in HLA-A and chicken(Wallny et al 2006 Koch et al 2007) are marked by asterisks Thenumbering of amino acid residues is based on the complete MHCclass I protein in the duck (Mesa et al 2004) The numbers on theright show the number of individuals with the particular sequence (N=120) question mark the precise number of individuals bearing theallele CaerU11 is not known (see the text for more details)
Genus Numbera Amino acid sequence distance plusmn SD Nucleotide sequence distance plusmn SD
Acrocephalus 37 0514plusmn0189 0185plusmn0053
Parus 24 0734plusmn0167 0260plusmn00483
Passer 21 0538plusmn0154 0187plusmn0040
Gallus 1 1390plusmn0102 0689plusmn0040
Coturnix 1 1618plusmn0149 0739plusmn0050
Anas 1 1247plusmn0122 0607plusmn0037
Table 1 Average distancesbetween the 29 Scarlet Rose-finch sequences and sequencesof six other bird species
a Number of sequences in theanalysed alignment
456 Immunogenetics (2009) 61451ndash461
Although gene expression analysis was not performedstrong correlation between signatures for balancing selec-tion and expression is suggested Furthermore the presenceof conserved sites that anchor the peptide-terminal main-chain atoms and a reading frame with no terminal codons orframeshift mutations suggest that possibly all sequences ofMHC class I described in this study might be functionalalthough further study at the level of cDNA is required toconfirm this There is an increasing number of studiesshowing that even nearly intact MHC sequences are notexpressed (eg Indovina et al 2001 Bonneaud et al 2004Shiina et al 2006) Hence MHC class I genes seem to behighly variable in the Scarlet Rosefinch and this compli-cated and complex structure is very different from non-
passerines For example the best described MHC in birdsthe chicken lsquominimal essentialrsquo MHC has very few buthighly condensed genes (eg there are two classical MHCclass I genes but only a single dominantly expressedKaufman et al 1999) it appears that this is the ancestralstate for birds since it occurs in various groups such asother gallinaceous birds (Jarvi et al 1999 Wittzell et al1999a Worley et al 2008) owls (Burri et al 2008) andparrots (Hughes et al 2008) Up to now the mostcomplicated MHC class I gene set has been described intwo species of passerines in the great reed warbler sevengenomic class I sequences were found in one individualone of which contained a frameshift mutation and washence nonfunctional (Westerdahl et al 1999) and eight
Table 2 Results of the likelihood ratio tests of positive selection on exon 3 of MHC class I in four genera of passerine birds
Genus Length Numbera M8 vs M7 Likelihood ratio test Number of positively selected sites
df Test statistic Significance PAML REL FEL
Carpodacus 91 29 2 578 plt0001 15 7 8
Acrocephalus 68 37 2 712 plt0001 8 14 4
Parus 69 24 2 472 plt0001 13 14 5
Passer 82 21 2 232 plt0001 2 2 1
Test statistic was computed as 2(LbminusLa) where La and Lb are log-likelihood values for each of the nested models being compared (M8 and M7respectively)
Length length of the aminoacid sequencesa Number of sequences in the analysed alignment
A - PAML 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ + B ndash REL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
C ndash FEL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
Fig 3 Comparison of results of three methods for detecting aminoacid sites under selection in four passerine species The analyses arebased on the alignments of 29 Rosefinch 37 warbler 24 tit and 21sparrow complete or partial sequences of exon 3 encoding alpha2domain of the protein Amino acid residues found to be under positiveselection by three methods (A PAML B REL C FEL) are bold (fordetails on the methods see the text) If the same residue was found tobe positively selected in at least two genera it is marked by rectangle
Regions excluded from analysis are marked by a squiggle Highlyconserved residues that anchor the peptide-terminal main-chain atomsare marked by crosses other residues forming variable part of peptide-binding region in HLA-A and chicken (Wallny et al 2006 Koch et al2007) are marked by asterisks The numbering of amino acid residuesis based on the complete MHC class I protein in the duck (Mesa et al2004) (Acrocephalus-AF449690) (Parus-AM232710) and (Passer-AY285012)
Immunogenetics (2009) 61451ndash461 457
different cDNA sequences were also found in an individualIn a separate study of the same species 20 genomicsequences of exon 3 of class I were found in fiveindividuals five of which were pseudogenes (Westerdahlet al 2004) Similar results were found in house sparrowswhere 20 sequences were verified in seven individualsthree of which were proved to be transcribed althoughthere were no pseudogenes (Bonneaud et al 2004) Ourresults concerning individual exon 3 variation are similar tothe ones mentioned above suggesting that complex MHCstructure could be a general pattern in songbirds
Evolution of MHC class I sequences in passerines
Several mechanisms of molecular evolution have beenalready documented in the avian MHC sequences includ-ing positive selection (Alcaide et al 2007 Hughes et al2008) concerted evolution (Wittzell et al 1999b) andtrans-species polymorphism (Richardson and Westerdahl2003 Alcaide et al 2007) In the present study model-based tests of selection (used here for the first time inpasserine species) have demonstrated the presence ofpositively selected sites in exon 3 of MHC class I in fourpasserine genera This part of the gene encodes the alpha 2domain of the protein which is included in forming thepeptide-binding groove Positive selection acting on somecodons of exon 3 is in accordance with published data onMHC class I in other birds where the ratio of non-synonymous to synonymous substitutions (dNdS) wasusually slightly higher at PBR sites than at non-PBR ones(Westerdahl et al 1999 Richardson and Westerdahl 2003Bonneaud et al 2004) The differences however were notsignificant and this could be caused either (1) by in-correctly identified PBR sites since the MHC proteinstructure in the majority of species studied is not well-understood In fact the PBR sites have usually beenidentified on the basis of the three-dimensional structureof human HLA-A protein (Bjorkman et al 1987) A modelof the three-dimensional structure of a passerine MHC classI molecule is highly desirable Or (2) another reason mightbe the fact that some of the studied sequences come fromloci that lost their importance and they are not underselection that is why it is crucial to identify sequences toparticular loci in the future studies (see Worley et al 2008)
Our results indicate extreme variability in selectivepressure among sites in the exon 3 and the presence of anumber of sites under balancing selection Approximatelyhalf of the positively selected sites in passerines (Fig 3)have been previously assumed to play a role in antigenbinding based on the structural models (Wallny et al 2006Koch et al 2007) However we found at least twoadditional sites (amino acid residues 126 and 136) understrong positive selection in several genera which have not
been previously reported for PBR (either in human HLA orchicken BF) The maintenance of considerable nonsynon-ymous variation at these specific codons by positiveselection suggests that such polymorphism may havefunctional consequences for peptide binding and presenta-tion to T cells (see similar recent results concerning teleostfish MHC molecules Aguilar and Garza 2007 or inmammals Amills et al 2008) The predictive power ofcodon-based models has not yet been tested in MHC andthe structural model describing peptide-binding groove inpasserine birds is still required to confirm the functionalrole of amino acid residues under different types ofselection
It appears that mammalian and avian MHCs may differin frequency of genetic exchange between different genesof multigene families Concerted evolution where the inter-locus gene conversion events are frequent in highlyduplicated MHC genes may be the predominant evolu-tionary mechanism in birds (Edwards et al 1995 Alcaide etal 2007 Garrigan and Edwards 1999 Wittzell et al 1999bHess and Edwards 2002) and probably also in fish (Reuschet al 2004 van Oosterhout et al 2006 Aguilar and Garza2007) and reptiles (Miller et al 2006) but not in mammals(Edwards et al 1997) This process creates high homologyamong MHC sequences derived from various loci and itbecomes impossible to assign different MHC alleles tospecific loci In birds so far most of the studies evidencinginter-locus recombination were conducted on MHC class IIConcerning MHC class I it has only been studied in thechicken where no evidence was found for concertedevolution to occur (Shaw et al 2007) Thus our resultson recombination analyses in the finches (ie the totalamount of recombination per codon was half of that ofsubstitution and we failed to assign the sequences tospecific loci) are in agreement with previous reports andfurthermore they suggest that concerted evolution may befrequent also in MHC class I genes in passerine birds Toconfirm this hypothesis the more detailed description ofMHC structure (especially the affiliation of particularalleles to individual loci in a population and the geneticvariation outside exon 3) is necessary
Extreme MHC class I variation in a single population
Previous studies on songbirds using the method ofdenaturing gradient gel electrophoresis or restriction frag-ment length polymorphism (RFLP) have also indicatedhigh sequence variation at exon 3 of MHC class I forexample in an outbred population of house sparrows 46sequences were found in 101 individuals or between oneand 11 alleles per individual (Bonneaud et al 2006a) in thegreat reed warbler 49 different RFLP genotypes werefound in 55 birds (Westerdahl et al 2004) Even so in the
458 Immunogenetics (2009) 61451ndash461
Scarlet Rosefinch 82 alleles were found in a singlepopulation and most of the birds displayed uniquegenotypes which is the highest exon 3 MHC class Ivariability observed thus far This result may still be anunderestimate considering that in at least one case theSSCP pattern was identical for two different allelesAlthough we were not able to sequence all the allelesdisplayed on SSCP it is very probable that all the peaksidentified by SSCP are true alleles and that there could beadditional ones (due to peak overlapping) Another reasonwe believe this variation has not been overestimated is thatan extraordinarily high variability of selectively neutralmarkers has been found in this species at two microsatelliteloci more than 70 alleles were found (Albrecht et al 2007)
Interestingly one of the alleles (CaerU02) was possessedby all individuals but one female (no 515) investigated inthis study From parentndashoffspring genotypes (Promerovaacute etal unpublished data) we know that also some of thisfemales offspring missed this allele thus this absence issurely heritable We conclude that CaerU02 is the mostfrequent allele on a locus with very low variability Thefemale no 515 is either homo- or heterozygote for other veryrare allele(s) or the locus (at least the copy with alleleCaerU02) is completely missing in her MHC haplotype
Our optimization of a genotyping method for analyses ofMHC class I variation will allow us to search for answers tofundamental questions concerning recent immunoecologysuch as the role of MHC in disease resistance expression ofsecondary ornaments and mating success (eg Bonneaud etal 2006a b Loiseau et al 2008 Richardson et al 2004Westerdahl et al 2005) Furthermore our data could beused for future comparative studies of the correlationbetween MHC diversity and migratory strategies in birdsWe presume that migratory birds such as the ScarletRosefinch (which migrates from the Czech Republic toIndia ca 5000 km) might encounter a larger variety ofpathogens during their lifetime than sedentary birds (forexample there is evidence that migrating individualsencounter more malarial parasites Peacuterez-Tris and Bensch2005) In that case more diverse immune mechanismscould help them fight these pathogens Moreover it hasalready been suggested that migratory species have highergenetic variability than sedentary species (Fitzpatrick 1994)and also that the higher genetic variability of migrants hasan impact on higher occurrence of extra-pair paternity(Spottiswoode and Moslashller 2004) The hypothesis thatmigration in birds might elicit a higher variability onMHC was first evaluated by Shiina et al (2004) whocompared the MHC of migratory quail with that ofsedentary chicken The quail has a more complicatedMHC than the chicken the class I region in the quail istwice as long as that of the chicken (Shiina et al 2004) andthere are five expressed class I genes although not all of
them are classical ones (Shiina et al 2006) SimilarlyRichardson and Westerdahl (2003) reported a difference inMHC variability between the migratory Great reed warblerand the sedentary inbred Seychelles warbler In this casehowever the low MHC variation in the latter could be dueto a severe bottleneck leading to strong decrease of overallgenetic variation Furthermore the endemic Basra reedwarbler (Acrocephalus griseldis) is as variable at MHC asthe Great reed warbler even though it is sedentary (Hanssonand Richardson 2005) High level of MHC polymorphismwas found in the house sparrow which is a non-migratoryspecies (Bonneaud et al 2004) which also contradicts thepredictions of the above hypothesis Unfortunately formost species (including the Scarlet Rosefinch) there is noprecise information about the functional variation ienumber of expressed MHC variants It is evident that dataon real MHC variation in more species are inevitablyrequired to perform a rigorous comparative meta-analysisand to test the predictions of migration-MHC hypothesis
Acknowledgements We thank Pavel Munclinger Michal VinklerJan Schnitzer and Radka Polaacutekovaacute for their help in the field Thanksto Anna Bryjovaacute and Dagmar Čiacutežkovaacute for their valuable advice inthe laboratory We are very grateful to Heidi C Hauffe for improvingthe English and two anonymous referees for critical comments on themanuscript This study was funded by the Grant Agency of theAcademy of Sciences of the Czech Republic (project noIAA600930608) the Grant Agency of the Czech Rep (project no206060851) and by the Ministry of Education of the Czech Repwhose Research Centre no LC06073 and Long-term Research Plan(no MSM 0021620828) to the Charles University in Prague formed aframework for a part of this study
References
Abbas AK Lichtman AH Pober JS (1994) Cellular and molecularimmunology W B Saunders Company Philadelphia
Aguilar A Garza JC (2007) Patterns of historical balancing selectionon the salmonid major histocompatibility complex class II betagene J Mol Evol 6534ndash43 doi101007s00239-006-0222-8
Albrecht T (2004) Edge effect in wetland-arable land boundarydetermines nesting success of scarlet rosefinches Carpodacuserythrinus in the Czech Republic Auk 121361ndash371doi1016420004-8038(2004)121[0361EEIWLB]20CO2
Albrecht T Schnitzer J Kreisinger J Exnerovaacute A Bryja JMunclinger P (2007) Extrapair paternity and the opportunity forsexual selection in long-distant migratory passerines Behav Ecol18477ndash486 doi101093behecoarm001
Alcaide M Edwards SV Negro JJ (2007) Characterization polymor-phism and evolution ofMHCClass II B genes in birds of prey JMolEvol 65541ndash554 doi101007s00239-007-9033-9
Amills M Ramiacuterez O Tomaacutes A Obexer-Ruff G Vidal O (2008)Positive selection on mammalian MHC-DQ genes revisited from amultispecies perspective Genes Immun 9651ndash658 doi101038gene200862
Anisimova M Nielsen R Yang Z (2003) Effect of recombination onthe accuracy of the likelihood method for detecting positiveselection at amino acid sites Genetics 1641229ndash1236
Immunogenetics (2009) 61451ndash461 459
Bjorkman PJ Saper MA Samraoui B Bennett WS Strominger JLWiley DC (1987) Structure of human class I histocompatibil-ity antigen HLA-A2 Nature 329506ndash512 doi101038329506a0
Bonneaud C Sorci G Morin V Westerdahl H Zoorob R Wittzell H(2004) Diversity of MHC class I and IIB genes in house sparrows(Passer domesticus) Immunogenetics 55855ndash865 doi101007s00251-004-0648-3
Bonneaud C Chastel O Federici P Westerdahl H Sorci G (2006a)Complex MHC-based mate choice in a wild passerine Proc RSoc 2731111ndash1116 doi101098rspb20053325
Bonneaud C Peacuterez-Tris J Federici P Chastel O Sorci G (2006b) Majorhistocompatibility alleles associated with local resistance to malariain a passerine Evolution Int J Org Evolution 60383ndash389
Bryja J Galan M Charbonnel N Cosson J-F (2005) Analysis ofmajor histocompatibility complex class II gene in water volesusing capillary electrophoresis-single stranded conformationpolymorphism Mol Ecol Notes 5173ndash176 doi101111j1471-8286200400855x
Burri R Niculita-Hirzel H Roulin A Fumagalli L (2008) Isolation andcharacterization of major histocompatibility complex (MHC) class IIB genes in teh Barn owl (Aves Tyto alba) Immunogenetics 60543ndash550 doi101007s00251-008-0308-0
Cramp S Simmons KEL Perrins CM (eds) (1977-1994) The birds ofthe Western Palearctic Oxford University Press Oxford
Edwards SV Wakeland EK Potts WK (1995) Contrasting histories ofavian and mammalian MHC genes revealed by class II B genesof songbirds Proc Natl Acad Sci U S A 9212200ndash12204doi101073pnas922612200
Edwards SV Chesnut K Satta Y Wakeland EK (1997) Ancestralpolymorphism of Mhc Class II genes in mice implications forbalancing selection and the mammalian molecular clock Genetics146655ndash668
Fitzpatrick S (1994) Colourful migratory birds evidence for amechanism other than parasite resistance for the maintenance oflsquogood genesrsquo sexual selection Proc R Soc Lond B Biol Sci257155ndash166 doi101098rspb19940109
Freeman-Gallant CR Johnson EM Saponara F Stanger M (2002)Variation at the major histocompatibility complex in Savannahsparrows Mol Ecol 111125ndash1130 doi101046j1365-294X200201508x
Garrigan D Edwards SV (1999) Polymorphism across an exon-intronboundary in an avian Mhc class II B gene Mol Biol Evol 161599ndash1606
Gouumly de Bellocq J Charbonnel N Morand S (2008) Coevolutionaryrelationship between helminth diversity and Mhc class II polymor-phism in rodents J Evol Biol 211144ndash1150 doi101111j1420-9101200801538x
Hall TA (1999) BioEdit a user-friendly biological sequence alignmenteditor and analisis program for Windows 9598NT NucleicAcids Symp Ser 4195ndash98
Hansson B Richardson DS (2005) Genetic variation in twoendangered Acrocephalus species compared to a widespreadcongener estimates based on functional and random loci AnimConserv 883ndash90 doi101017S1367943004001878
Hess CM Edwards SV (2002) The evolution of major histocompat-ibility genes in birds Bioscience 52423ndash431 doi1016410006-3568(2002)052[0423TEOTMH]20CO2
Hess CM Gasper J Hoekstra H Hill C Edwards SV (2000) MHCclass II pseudogene and genomic signature of a 32-kb cosmid inthe House Finch (Carpodacus mexicanus) Genome Res 1013ndash23 doi101101gr105613
Hosomichi K Shiina T Suzuki S Tanaka M Shimizu S Iwamoto SHara H Yoshida Y Kulski JK Inoko H Hanzawa K (2006) Themajor histocompatibility complex (Mhc) class IIB region hasgreater genomic structural flexibility and diversity in the quail
than the chicken BMC Genomics 7322ndash335 doi1011861471-2164-7-322
Hughes AL Yeager M (1998) Natural selection at major histocom-patibility complex loci of vertebrates Annu Rev Genet 32415ndash435 doi101146annurevgenet321415
Hughes CR Miles S Walbroehl JM (2008) Support for the minimalessential MHC hypothesis a parrot with a single highlypolymorphic MHC class II B gene Immunogenetics 60219ndash231 doi101007s00251-008-0287-1
Indovina P Megiorni F Fontemaggi G Coni P Mora B Mazzilli MC(2001) Absence of in vivo DNA-Protein interactions in theDQA2 and DQB2 promoter regions Hum Immunol 62504ndash508doi101016S0198-8859(01)00236-1
Jarvi SI Goto RM Gee GF Briles WE Miller MM (1999)Identification inheritance and linkage of B-G-like and MHCclass I genes in cranes Am Genet Assoc 90152ndash159
Kaufman J (1999) Co-evolving genes in MHC haplotypes the lsquorulersquofor nonmammalian vertebrates Immunogenetics 50228ndash236doi101007s002510050597
Kaufman J Salamonsen J Flajnik M (1994) Evolutionary conserva-tion of MHC class I and class II molecules-different yet the sameSemin Immunol 6411ndash424 doi101006smim19941050
Kaufman J Voumllk H Wallny H (1995) A bdquominimal essential MHCldquoand an bdquounrecognized MHCldquo two extremes in selection forpolymorphism Immunol Rev 14363ndash88 doi101111j1600-065X1995tb00670x
Kaufman J Jacob J Shaw I Walker B Milne S Beck S SalomonsenJ (1999) Gene organisation determines evolution of function inthe chicken MHC Immunol Rev 167101ndash117 doi101111j1600-065X1999tb01385x
Klein J (1986) Natural history of the major histocompatibilitycomplex Wiley New York
Klein J Sato A Nagl S Oh Uigin C (1998) Molecular trans-speciespolymorphism Annu Rev Ecol Syst 291ndash21 doi101146annurevecolsys2911
Koch M Camp S Collen T Avila D Salomonsen J Wallny HJ vanHateren A Hunt L Jacob JP Johnston F Marston DA Shaw IDunbar PR Cerundolo V Jones EY Kaufman J (2007)Structures of an MHC class I molecule from B21 chickensillustrate promiscuous peptide binding Immunity 27885ndash899doi101016jimmuni200711007
Kosakovsky Pond SL Frost SDW (2005) Not so different after all acomparison of methods for detecting amino acid sites under selectionMol Biol Evol 221208ndash1222 doi101093molbevmsi105
Kosakovsky Pond SL Frost SDW Muse SV (2005) HyPhyhypothesis testing using phylogenies Bioinformatics 21676ndash679 doi101093bioinformaticsbti079
Kosakovsky Pond SL Posada D Gravenor MB Woelk CH FrostSDW (2006) Automated phylogenetic detection of recombinationusing a genetic algorithm Mol Biol Evol 231891ndash1901doi101093molbevmsl051
Loiseau C Zoorob R Garnier S Birard J Federici P Julliard R SorciG (2008) Antagonistic effects of a MHC class I allele on malaria-infected house sparrows Ecol Lett 11258ndash265 doi101111j1461-0248200701141x
Mesa CM Thulien KJ Moon DA Veniamin SM Magor KE (2004)The dominant MHC class I gene is adjacent to the polymorphicTAP2 gene in the duck Anas platyrhynchos Immunogenetics56192ndash203 doi101007s00251-004-0672-3
Miller HC Lambert DM (2004) Gene duplication and geneconversion in class II MHC genes of New Zealand robins(Petroicidae) Immunogenetics 56178ndash191
Miller MM Goto RM Bernot A Zoorob R Auffray C Bumstead NBriles WE (1994) Two Mhc class I and two Mhc class II genes mapto the chicken Rfp-Y system outside the B complex Proc Natl AcadSci U S A 914397ndash4401 doi101073pnas91104397
460 Immunogenetics (2009) 61451ndash461
Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461

positively selected sites were often identical between thefour compared species as well as between the three methodsof analysis (Fig 3)
Recombination in MHC class I of the Scarlet Rosefinch
Significant negative correlation between physical distanceand linkage disequilibrium was detected using all threeanalyses in the program PERMUTE (plt0001) suggestingthat recombination regularly occurs in exon 3 of MHC classI genes The mean population recombination per codon (ρ=0233) identified in the program OMEGAMAP is approxi-mately half of the mean total population mutation (θ=0534)
again suggesting that recombination is important for gener-ating MHC class I variants in the Scarlet Rosefinch
Discussion
Structure of classical MHC class I genes in passerine birds
In this study the structure and diversity of MHC class I wasstudied in a wild-living songbird species Up to ninedifferent MHC class I sequences were present in oneScarlet Rosefinch individual which means that at least fiveamplified MHC class I genes could occur in a single bird
98 108 118 128 138 148 158 168 178 |||||||||||||||||| CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEG-EAEGWTNYLKHECPEWLQKYVGYGQKELER 1 CaerU12 R-VENVR 2 CaerU29 Y-VENV 1 CaerU05 RLRYP-PRRH 4 CaerU02 LWRLPKKPNSTGKIVRNVSRR 119CaerU17 LWLPKKPNSTGKIVRNVSRRERH 4 CaerU18 LWLPKKPNSTGKIVRNVSRR 32 CaerU08 FIQWLSVRRAFIED-FRR 27 CaerU16 FIQWLSVRRAFIEDK-FRR 1 CaerU22 FIQWLSVRRAFIEDK-FIRE 7 CaerU20 FIQWLSVDARFIP-VKR 19 CaerU06 WRSSEVYYRFIPDVKE-VFRE 51 CaerU07 WRSSEVYYWRFIPDVKE-VFRE 111CaerU11 WRSSEVYYRFIPDVKE-VFRER CaerU04 LWVRHNRFID-FIC 9 CaerU13 LRVRRARFIRDK-MRDV 1 CaerU14 LRVRRARFIRD-MRDV 1 CaerU27 WRVRRARFIDG-VRRDV 1 CaerU28 LRVRRARFIRDKE-VF 1 CaerU03 LNVRYNRFIPD-PRW 4 CaerU09 RNVRYNRFIELDKE-AF 11CaerU10 WRNVRYNRFIPDKE-VFIH 5 CaerU15 WRNVRYNRFIEPD-PRVI 6 CaerU23 LWNVRYNRFIPDSKE-VLH 11CaerU24 WRNVRYRFIPD-PRRVH 4 CaerU25 WRSNVRFYNRFIPDKE-VFDQH 3 CaerU21 LWNVRDARFIDNGKE-VF 7 CaerU26 LRNVRDARFIDNGK-VEMRE 10Gallus gallus SVQWMIEGPRYYQMARFTAKTMTTVPEVPKKES-PRKEETVRREKAG Coturnix japonica SVQWMIDTTRYNQYARFIVKDTMTTVPEVPKKEGD-YRQKHEETVQRRHENKAG Anas platyrhynchos SVQRMKRFEQYERFIALKDTLTTDAQKKETYRTKYENTIRSKDV + + ++ ++ +
Fig 2 Amino acid sequences encoded by exon 3 in the ScarletRosefinch corresponding to alpha2 domain of the protein comparedwith GenBank sequences from chicken (X12780) quail (D29813) andduck (AY294416) Identity with the Scarlet Rosefinch allele CaerU1is indicated by dots gaps which are added to get the best alignment bydashes Highly conserved residues that anchor the peptide-terminalmain-chain atoms are marked by crosses other residues forming
variable part of peptide-binding region in HLA-A and chicken(Wallny et al 2006 Koch et al 2007) are marked by asterisks Thenumbering of amino acid residues is based on the complete MHCclass I protein in the duck (Mesa et al 2004) The numbers on theright show the number of individuals with the particular sequence (N=120) question mark the precise number of individuals bearing theallele CaerU11 is not known (see the text for more details)
Genus Numbera Amino acid sequence distance plusmn SD Nucleotide sequence distance plusmn SD
Acrocephalus 37 0514plusmn0189 0185plusmn0053
Parus 24 0734plusmn0167 0260plusmn00483
Passer 21 0538plusmn0154 0187plusmn0040
Gallus 1 1390plusmn0102 0689plusmn0040
Coturnix 1 1618plusmn0149 0739plusmn0050
Anas 1 1247plusmn0122 0607plusmn0037
Table 1 Average distancesbetween the 29 Scarlet Rose-finch sequences and sequencesof six other bird species
a Number of sequences in theanalysed alignment
456 Immunogenetics (2009) 61451ndash461
Although gene expression analysis was not performedstrong correlation between signatures for balancing selec-tion and expression is suggested Furthermore the presenceof conserved sites that anchor the peptide-terminal main-chain atoms and a reading frame with no terminal codons orframeshift mutations suggest that possibly all sequences ofMHC class I described in this study might be functionalalthough further study at the level of cDNA is required toconfirm this There is an increasing number of studiesshowing that even nearly intact MHC sequences are notexpressed (eg Indovina et al 2001 Bonneaud et al 2004Shiina et al 2006) Hence MHC class I genes seem to behighly variable in the Scarlet Rosefinch and this compli-cated and complex structure is very different from non-
passerines For example the best described MHC in birdsthe chicken lsquominimal essentialrsquo MHC has very few buthighly condensed genes (eg there are two classical MHCclass I genes but only a single dominantly expressedKaufman et al 1999) it appears that this is the ancestralstate for birds since it occurs in various groups such asother gallinaceous birds (Jarvi et al 1999 Wittzell et al1999a Worley et al 2008) owls (Burri et al 2008) andparrots (Hughes et al 2008) Up to now the mostcomplicated MHC class I gene set has been described intwo species of passerines in the great reed warbler sevengenomic class I sequences were found in one individualone of which contained a frameshift mutation and washence nonfunctional (Westerdahl et al 1999) and eight
Table 2 Results of the likelihood ratio tests of positive selection on exon 3 of MHC class I in four genera of passerine birds
Genus Length Numbera M8 vs M7 Likelihood ratio test Number of positively selected sites
df Test statistic Significance PAML REL FEL
Carpodacus 91 29 2 578 plt0001 15 7 8
Acrocephalus 68 37 2 712 plt0001 8 14 4
Parus 69 24 2 472 plt0001 13 14 5
Passer 82 21 2 232 plt0001 2 2 1
Test statistic was computed as 2(LbminusLa) where La and Lb are log-likelihood values for each of the nested models being compared (M8 and M7respectively)
Length length of the aminoacid sequencesa Number of sequences in the analysed alignment
A - PAML 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ + B ndash REL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
C ndash FEL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
Fig 3 Comparison of results of three methods for detecting aminoacid sites under selection in four passerine species The analyses arebased on the alignments of 29 Rosefinch 37 warbler 24 tit and 21sparrow complete or partial sequences of exon 3 encoding alpha2domain of the protein Amino acid residues found to be under positiveselection by three methods (A PAML B REL C FEL) are bold (fordetails on the methods see the text) If the same residue was found tobe positively selected in at least two genera it is marked by rectangle
Regions excluded from analysis are marked by a squiggle Highlyconserved residues that anchor the peptide-terminal main-chain atomsare marked by crosses other residues forming variable part of peptide-binding region in HLA-A and chicken (Wallny et al 2006 Koch et al2007) are marked by asterisks The numbering of amino acid residuesis based on the complete MHC class I protein in the duck (Mesa et al2004) (Acrocephalus-AF449690) (Parus-AM232710) and (Passer-AY285012)
Immunogenetics (2009) 61451ndash461 457
different cDNA sequences were also found in an individualIn a separate study of the same species 20 genomicsequences of exon 3 of class I were found in fiveindividuals five of which were pseudogenes (Westerdahlet al 2004) Similar results were found in house sparrowswhere 20 sequences were verified in seven individualsthree of which were proved to be transcribed althoughthere were no pseudogenes (Bonneaud et al 2004) Ourresults concerning individual exon 3 variation are similar tothe ones mentioned above suggesting that complex MHCstructure could be a general pattern in songbirds
Evolution of MHC class I sequences in passerines
Several mechanisms of molecular evolution have beenalready documented in the avian MHC sequences includ-ing positive selection (Alcaide et al 2007 Hughes et al2008) concerted evolution (Wittzell et al 1999b) andtrans-species polymorphism (Richardson and Westerdahl2003 Alcaide et al 2007) In the present study model-based tests of selection (used here for the first time inpasserine species) have demonstrated the presence ofpositively selected sites in exon 3 of MHC class I in fourpasserine genera This part of the gene encodes the alpha 2domain of the protein which is included in forming thepeptide-binding groove Positive selection acting on somecodons of exon 3 is in accordance with published data onMHC class I in other birds where the ratio of non-synonymous to synonymous substitutions (dNdS) wasusually slightly higher at PBR sites than at non-PBR ones(Westerdahl et al 1999 Richardson and Westerdahl 2003Bonneaud et al 2004) The differences however were notsignificant and this could be caused either (1) by in-correctly identified PBR sites since the MHC proteinstructure in the majority of species studied is not well-understood In fact the PBR sites have usually beenidentified on the basis of the three-dimensional structureof human HLA-A protein (Bjorkman et al 1987) A modelof the three-dimensional structure of a passerine MHC classI molecule is highly desirable Or (2) another reason mightbe the fact that some of the studied sequences come fromloci that lost their importance and they are not underselection that is why it is crucial to identify sequences toparticular loci in the future studies (see Worley et al 2008)
Our results indicate extreme variability in selectivepressure among sites in the exon 3 and the presence of anumber of sites under balancing selection Approximatelyhalf of the positively selected sites in passerines (Fig 3)have been previously assumed to play a role in antigenbinding based on the structural models (Wallny et al 2006Koch et al 2007) However we found at least twoadditional sites (amino acid residues 126 and 136) understrong positive selection in several genera which have not
been previously reported for PBR (either in human HLA orchicken BF) The maintenance of considerable nonsynon-ymous variation at these specific codons by positiveselection suggests that such polymorphism may havefunctional consequences for peptide binding and presenta-tion to T cells (see similar recent results concerning teleostfish MHC molecules Aguilar and Garza 2007 or inmammals Amills et al 2008) The predictive power ofcodon-based models has not yet been tested in MHC andthe structural model describing peptide-binding groove inpasserine birds is still required to confirm the functionalrole of amino acid residues under different types ofselection
It appears that mammalian and avian MHCs may differin frequency of genetic exchange between different genesof multigene families Concerted evolution where the inter-locus gene conversion events are frequent in highlyduplicated MHC genes may be the predominant evolu-tionary mechanism in birds (Edwards et al 1995 Alcaide etal 2007 Garrigan and Edwards 1999 Wittzell et al 1999bHess and Edwards 2002) and probably also in fish (Reuschet al 2004 van Oosterhout et al 2006 Aguilar and Garza2007) and reptiles (Miller et al 2006) but not in mammals(Edwards et al 1997) This process creates high homologyamong MHC sequences derived from various loci and itbecomes impossible to assign different MHC alleles tospecific loci In birds so far most of the studies evidencinginter-locus recombination were conducted on MHC class IIConcerning MHC class I it has only been studied in thechicken where no evidence was found for concertedevolution to occur (Shaw et al 2007) Thus our resultson recombination analyses in the finches (ie the totalamount of recombination per codon was half of that ofsubstitution and we failed to assign the sequences tospecific loci) are in agreement with previous reports andfurthermore they suggest that concerted evolution may befrequent also in MHC class I genes in passerine birds Toconfirm this hypothesis the more detailed description ofMHC structure (especially the affiliation of particularalleles to individual loci in a population and the geneticvariation outside exon 3) is necessary
Extreme MHC class I variation in a single population
Previous studies on songbirds using the method ofdenaturing gradient gel electrophoresis or restriction frag-ment length polymorphism (RFLP) have also indicatedhigh sequence variation at exon 3 of MHC class I forexample in an outbred population of house sparrows 46sequences were found in 101 individuals or between oneand 11 alleles per individual (Bonneaud et al 2006a) in thegreat reed warbler 49 different RFLP genotypes werefound in 55 birds (Westerdahl et al 2004) Even so in the
458 Immunogenetics (2009) 61451ndash461
Scarlet Rosefinch 82 alleles were found in a singlepopulation and most of the birds displayed uniquegenotypes which is the highest exon 3 MHC class Ivariability observed thus far This result may still be anunderestimate considering that in at least one case theSSCP pattern was identical for two different allelesAlthough we were not able to sequence all the allelesdisplayed on SSCP it is very probable that all the peaksidentified by SSCP are true alleles and that there could beadditional ones (due to peak overlapping) Another reasonwe believe this variation has not been overestimated is thatan extraordinarily high variability of selectively neutralmarkers has been found in this species at two microsatelliteloci more than 70 alleles were found (Albrecht et al 2007)
Interestingly one of the alleles (CaerU02) was possessedby all individuals but one female (no 515) investigated inthis study From parentndashoffspring genotypes (Promerovaacute etal unpublished data) we know that also some of thisfemales offspring missed this allele thus this absence issurely heritable We conclude that CaerU02 is the mostfrequent allele on a locus with very low variability Thefemale no 515 is either homo- or heterozygote for other veryrare allele(s) or the locus (at least the copy with alleleCaerU02) is completely missing in her MHC haplotype
Our optimization of a genotyping method for analyses ofMHC class I variation will allow us to search for answers tofundamental questions concerning recent immunoecologysuch as the role of MHC in disease resistance expression ofsecondary ornaments and mating success (eg Bonneaud etal 2006a b Loiseau et al 2008 Richardson et al 2004Westerdahl et al 2005) Furthermore our data could beused for future comparative studies of the correlationbetween MHC diversity and migratory strategies in birdsWe presume that migratory birds such as the ScarletRosefinch (which migrates from the Czech Republic toIndia ca 5000 km) might encounter a larger variety ofpathogens during their lifetime than sedentary birds (forexample there is evidence that migrating individualsencounter more malarial parasites Peacuterez-Tris and Bensch2005) In that case more diverse immune mechanismscould help them fight these pathogens Moreover it hasalready been suggested that migratory species have highergenetic variability than sedentary species (Fitzpatrick 1994)and also that the higher genetic variability of migrants hasan impact on higher occurrence of extra-pair paternity(Spottiswoode and Moslashller 2004) The hypothesis thatmigration in birds might elicit a higher variability onMHC was first evaluated by Shiina et al (2004) whocompared the MHC of migratory quail with that ofsedentary chicken The quail has a more complicatedMHC than the chicken the class I region in the quail istwice as long as that of the chicken (Shiina et al 2004) andthere are five expressed class I genes although not all of
them are classical ones (Shiina et al 2006) SimilarlyRichardson and Westerdahl (2003) reported a difference inMHC variability between the migratory Great reed warblerand the sedentary inbred Seychelles warbler In this casehowever the low MHC variation in the latter could be dueto a severe bottleneck leading to strong decrease of overallgenetic variation Furthermore the endemic Basra reedwarbler (Acrocephalus griseldis) is as variable at MHC asthe Great reed warbler even though it is sedentary (Hanssonand Richardson 2005) High level of MHC polymorphismwas found in the house sparrow which is a non-migratoryspecies (Bonneaud et al 2004) which also contradicts thepredictions of the above hypothesis Unfortunately formost species (including the Scarlet Rosefinch) there is noprecise information about the functional variation ienumber of expressed MHC variants It is evident that dataon real MHC variation in more species are inevitablyrequired to perform a rigorous comparative meta-analysisand to test the predictions of migration-MHC hypothesis
Acknowledgements We thank Pavel Munclinger Michal VinklerJan Schnitzer and Radka Polaacutekovaacute for their help in the field Thanksto Anna Bryjovaacute and Dagmar Čiacutežkovaacute for their valuable advice inthe laboratory We are very grateful to Heidi C Hauffe for improvingthe English and two anonymous referees for critical comments on themanuscript This study was funded by the Grant Agency of theAcademy of Sciences of the Czech Republic (project noIAA600930608) the Grant Agency of the Czech Rep (project no206060851) and by the Ministry of Education of the Czech Repwhose Research Centre no LC06073 and Long-term Research Plan(no MSM 0021620828) to the Charles University in Prague formed aframework for a part of this study
References
Abbas AK Lichtman AH Pober JS (1994) Cellular and molecularimmunology W B Saunders Company Philadelphia
Aguilar A Garza JC (2007) Patterns of historical balancing selectionon the salmonid major histocompatibility complex class II betagene J Mol Evol 6534ndash43 doi101007s00239-006-0222-8
Albrecht T (2004) Edge effect in wetland-arable land boundarydetermines nesting success of scarlet rosefinches Carpodacuserythrinus in the Czech Republic Auk 121361ndash371doi1016420004-8038(2004)121[0361EEIWLB]20CO2
Albrecht T Schnitzer J Kreisinger J Exnerovaacute A Bryja JMunclinger P (2007) Extrapair paternity and the opportunity forsexual selection in long-distant migratory passerines Behav Ecol18477ndash486 doi101093behecoarm001
Alcaide M Edwards SV Negro JJ (2007) Characterization polymor-phism and evolution ofMHCClass II B genes in birds of prey JMolEvol 65541ndash554 doi101007s00239-007-9033-9
Amills M Ramiacuterez O Tomaacutes A Obexer-Ruff G Vidal O (2008)Positive selection on mammalian MHC-DQ genes revisited from amultispecies perspective Genes Immun 9651ndash658 doi101038gene200862
Anisimova M Nielsen R Yang Z (2003) Effect of recombination onthe accuracy of the likelihood method for detecting positiveselection at amino acid sites Genetics 1641229ndash1236
Immunogenetics (2009) 61451ndash461 459
Bjorkman PJ Saper MA Samraoui B Bennett WS Strominger JLWiley DC (1987) Structure of human class I histocompatibil-ity antigen HLA-A2 Nature 329506ndash512 doi101038329506a0
Bonneaud C Sorci G Morin V Westerdahl H Zoorob R Wittzell H(2004) Diversity of MHC class I and IIB genes in house sparrows(Passer domesticus) Immunogenetics 55855ndash865 doi101007s00251-004-0648-3
Bonneaud C Chastel O Federici P Westerdahl H Sorci G (2006a)Complex MHC-based mate choice in a wild passerine Proc RSoc 2731111ndash1116 doi101098rspb20053325
Bonneaud C Peacuterez-Tris J Federici P Chastel O Sorci G (2006b) Majorhistocompatibility alleles associated with local resistance to malariain a passerine Evolution Int J Org Evolution 60383ndash389
Bryja J Galan M Charbonnel N Cosson J-F (2005) Analysis ofmajor histocompatibility complex class II gene in water volesusing capillary electrophoresis-single stranded conformationpolymorphism Mol Ecol Notes 5173ndash176 doi101111j1471-8286200400855x
Burri R Niculita-Hirzel H Roulin A Fumagalli L (2008) Isolation andcharacterization of major histocompatibility complex (MHC) class IIB genes in teh Barn owl (Aves Tyto alba) Immunogenetics 60543ndash550 doi101007s00251-008-0308-0
Cramp S Simmons KEL Perrins CM (eds) (1977-1994) The birds ofthe Western Palearctic Oxford University Press Oxford
Edwards SV Wakeland EK Potts WK (1995) Contrasting histories ofavian and mammalian MHC genes revealed by class II B genesof songbirds Proc Natl Acad Sci U S A 9212200ndash12204doi101073pnas922612200
Edwards SV Chesnut K Satta Y Wakeland EK (1997) Ancestralpolymorphism of Mhc Class II genes in mice implications forbalancing selection and the mammalian molecular clock Genetics146655ndash668
Fitzpatrick S (1994) Colourful migratory birds evidence for amechanism other than parasite resistance for the maintenance oflsquogood genesrsquo sexual selection Proc R Soc Lond B Biol Sci257155ndash166 doi101098rspb19940109
Freeman-Gallant CR Johnson EM Saponara F Stanger M (2002)Variation at the major histocompatibility complex in Savannahsparrows Mol Ecol 111125ndash1130 doi101046j1365-294X200201508x
Garrigan D Edwards SV (1999) Polymorphism across an exon-intronboundary in an avian Mhc class II B gene Mol Biol Evol 161599ndash1606
Gouumly de Bellocq J Charbonnel N Morand S (2008) Coevolutionaryrelationship between helminth diversity and Mhc class II polymor-phism in rodents J Evol Biol 211144ndash1150 doi101111j1420-9101200801538x
Hall TA (1999) BioEdit a user-friendly biological sequence alignmenteditor and analisis program for Windows 9598NT NucleicAcids Symp Ser 4195ndash98
Hansson B Richardson DS (2005) Genetic variation in twoendangered Acrocephalus species compared to a widespreadcongener estimates based on functional and random loci AnimConserv 883ndash90 doi101017S1367943004001878
Hess CM Edwards SV (2002) The evolution of major histocompat-ibility genes in birds Bioscience 52423ndash431 doi1016410006-3568(2002)052[0423TEOTMH]20CO2
Hess CM Gasper J Hoekstra H Hill C Edwards SV (2000) MHCclass II pseudogene and genomic signature of a 32-kb cosmid inthe House Finch (Carpodacus mexicanus) Genome Res 1013ndash23 doi101101gr105613
Hosomichi K Shiina T Suzuki S Tanaka M Shimizu S Iwamoto SHara H Yoshida Y Kulski JK Inoko H Hanzawa K (2006) Themajor histocompatibility complex (Mhc) class IIB region hasgreater genomic structural flexibility and diversity in the quail
than the chicken BMC Genomics 7322ndash335 doi1011861471-2164-7-322
Hughes AL Yeager M (1998) Natural selection at major histocom-patibility complex loci of vertebrates Annu Rev Genet 32415ndash435 doi101146annurevgenet321415
Hughes CR Miles S Walbroehl JM (2008) Support for the minimalessential MHC hypothesis a parrot with a single highlypolymorphic MHC class II B gene Immunogenetics 60219ndash231 doi101007s00251-008-0287-1
Indovina P Megiorni F Fontemaggi G Coni P Mora B Mazzilli MC(2001) Absence of in vivo DNA-Protein interactions in theDQA2 and DQB2 promoter regions Hum Immunol 62504ndash508doi101016S0198-8859(01)00236-1
Jarvi SI Goto RM Gee GF Briles WE Miller MM (1999)Identification inheritance and linkage of B-G-like and MHCclass I genes in cranes Am Genet Assoc 90152ndash159
Kaufman J (1999) Co-evolving genes in MHC haplotypes the lsquorulersquofor nonmammalian vertebrates Immunogenetics 50228ndash236doi101007s002510050597
Kaufman J Salamonsen J Flajnik M (1994) Evolutionary conserva-tion of MHC class I and class II molecules-different yet the sameSemin Immunol 6411ndash424 doi101006smim19941050
Kaufman J Voumllk H Wallny H (1995) A bdquominimal essential MHCldquoand an bdquounrecognized MHCldquo two extremes in selection forpolymorphism Immunol Rev 14363ndash88 doi101111j1600-065X1995tb00670x
Kaufman J Jacob J Shaw I Walker B Milne S Beck S SalomonsenJ (1999) Gene organisation determines evolution of function inthe chicken MHC Immunol Rev 167101ndash117 doi101111j1600-065X1999tb01385x
Klein J (1986) Natural history of the major histocompatibilitycomplex Wiley New York
Klein J Sato A Nagl S Oh Uigin C (1998) Molecular trans-speciespolymorphism Annu Rev Ecol Syst 291ndash21 doi101146annurevecolsys2911
Koch M Camp S Collen T Avila D Salomonsen J Wallny HJ vanHateren A Hunt L Jacob JP Johnston F Marston DA Shaw IDunbar PR Cerundolo V Jones EY Kaufman J (2007)Structures of an MHC class I molecule from B21 chickensillustrate promiscuous peptide binding Immunity 27885ndash899doi101016jimmuni200711007
Kosakovsky Pond SL Frost SDW (2005) Not so different after all acomparison of methods for detecting amino acid sites under selectionMol Biol Evol 221208ndash1222 doi101093molbevmsi105
Kosakovsky Pond SL Frost SDW Muse SV (2005) HyPhyhypothesis testing using phylogenies Bioinformatics 21676ndash679 doi101093bioinformaticsbti079
Kosakovsky Pond SL Posada D Gravenor MB Woelk CH FrostSDW (2006) Automated phylogenetic detection of recombinationusing a genetic algorithm Mol Biol Evol 231891ndash1901doi101093molbevmsl051
Loiseau C Zoorob R Garnier S Birard J Federici P Julliard R SorciG (2008) Antagonistic effects of a MHC class I allele on malaria-infected house sparrows Ecol Lett 11258ndash265 doi101111j1461-0248200701141x
Mesa CM Thulien KJ Moon DA Veniamin SM Magor KE (2004)The dominant MHC class I gene is adjacent to the polymorphicTAP2 gene in the duck Anas platyrhynchos Immunogenetics56192ndash203 doi101007s00251-004-0672-3
Miller HC Lambert DM (2004) Gene duplication and geneconversion in class II MHC genes of New Zealand robins(Petroicidae) Immunogenetics 56178ndash191
Miller MM Goto RM Bernot A Zoorob R Auffray C Bumstead NBriles WE (1994) Two Mhc class I and two Mhc class II genes mapto the chicken Rfp-Y system outside the B complex Proc Natl AcadSci U S A 914397ndash4401 doi101073pnas91104397
460 Immunogenetics (2009) 61451ndash461
Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461
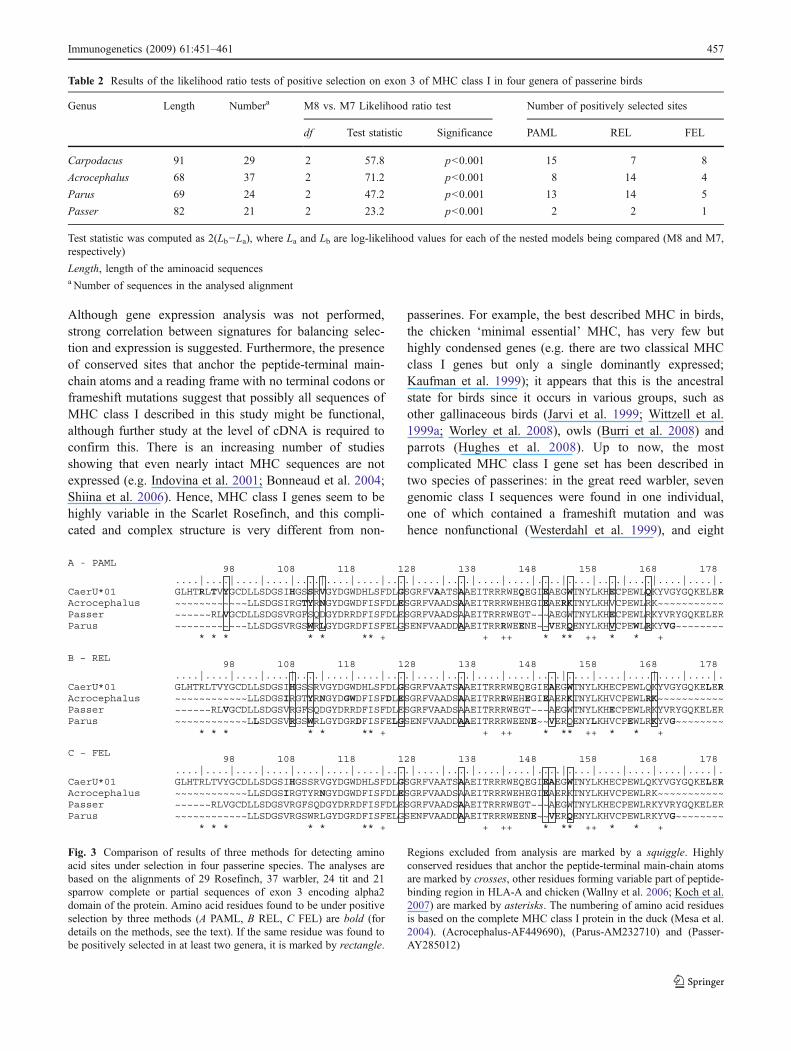
Although gene expression analysis was not performedstrong correlation between signatures for balancing selec-tion and expression is suggested Furthermore the presenceof conserved sites that anchor the peptide-terminal main-chain atoms and a reading frame with no terminal codons orframeshift mutations suggest that possibly all sequences ofMHC class I described in this study might be functionalalthough further study at the level of cDNA is required toconfirm this There is an increasing number of studiesshowing that even nearly intact MHC sequences are notexpressed (eg Indovina et al 2001 Bonneaud et al 2004Shiina et al 2006) Hence MHC class I genes seem to behighly variable in the Scarlet Rosefinch and this compli-cated and complex structure is very different from non-
passerines For example the best described MHC in birdsthe chicken lsquominimal essentialrsquo MHC has very few buthighly condensed genes (eg there are two classical MHCclass I genes but only a single dominantly expressedKaufman et al 1999) it appears that this is the ancestralstate for birds since it occurs in various groups such asother gallinaceous birds (Jarvi et al 1999 Wittzell et al1999a Worley et al 2008) owls (Burri et al 2008) andparrots (Hughes et al 2008) Up to now the mostcomplicated MHC class I gene set has been described intwo species of passerines in the great reed warbler sevengenomic class I sequences were found in one individualone of which contained a frameshift mutation and washence nonfunctional (Westerdahl et al 1999) and eight
Table 2 Results of the likelihood ratio tests of positive selection on exon 3 of MHC class I in four genera of passerine birds
Genus Length Numbera M8 vs M7 Likelihood ratio test Number of positively selected sites
df Test statistic Significance PAML REL FEL
Carpodacus 91 29 2 578 plt0001 15 7 8
Acrocephalus 68 37 2 712 plt0001 8 14 4
Parus 69 24 2 472 plt0001 13 14 5
Passer 82 21 2 232 plt0001 2 2 1
Test statistic was computed as 2(LbminusLa) where La and Lb are log-likelihood values for each of the nested models being compared (M8 and M7respectively)
Length length of the aminoacid sequencesa Number of sequences in the analysed alignment
A - PAML 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ + B ndash REL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
C ndash FEL 98 108 118 128 138 148 158 168 178 ||||||||||||||||||CaerU01 GLHTRLTVYGCDLLSDGSIHGSSRVGYDGWDHLSFDLGSGRFVAATSAAEITRRRWEQEGIEAEGWTNYLKHECPEWLQKYVGYGQKELERAcrocephalus ~~~~~~~~~~~~LLSDGSIRGTYRNGYDGWDFISFDLESGRFVAADSAAEITRRRWEHEGIEAERKTNYLKHVCPEWLRK~~~~~~~~~~~Passer ~~~~~~RLVGCDLLSDGSVRGFSQDGYDRRDFISFDLESGRFVAADSAAEITRRRWEGT~~~AEGWTNYLKHECPEWLRKYVRYGQKELERParus ~~~~~~~~~~~~LLSDGSVRGSWRLGYDGRDFISFELGSENFVAADDAAEITRRRWEENE~~VERQENYLKHVCPEWLRKYVG~~~~~~~~ + + ++ ++ +
Fig 3 Comparison of results of three methods for detecting aminoacid sites under selection in four passerine species The analyses arebased on the alignments of 29 Rosefinch 37 warbler 24 tit and 21sparrow complete or partial sequences of exon 3 encoding alpha2domain of the protein Amino acid residues found to be under positiveselection by three methods (A PAML B REL C FEL) are bold (fordetails on the methods see the text) If the same residue was found tobe positively selected in at least two genera it is marked by rectangle
Regions excluded from analysis are marked by a squiggle Highlyconserved residues that anchor the peptide-terminal main-chain atomsare marked by crosses other residues forming variable part of peptide-binding region in HLA-A and chicken (Wallny et al 2006 Koch et al2007) are marked by asterisks The numbering of amino acid residuesis based on the complete MHC class I protein in the duck (Mesa et al2004) (Acrocephalus-AF449690) (Parus-AM232710) and (Passer-AY285012)
Immunogenetics (2009) 61451ndash461 457
different cDNA sequences were also found in an individualIn a separate study of the same species 20 genomicsequences of exon 3 of class I were found in fiveindividuals five of which were pseudogenes (Westerdahlet al 2004) Similar results were found in house sparrowswhere 20 sequences were verified in seven individualsthree of which were proved to be transcribed althoughthere were no pseudogenes (Bonneaud et al 2004) Ourresults concerning individual exon 3 variation are similar tothe ones mentioned above suggesting that complex MHCstructure could be a general pattern in songbirds
Evolution of MHC class I sequences in passerines
Several mechanisms of molecular evolution have beenalready documented in the avian MHC sequences includ-ing positive selection (Alcaide et al 2007 Hughes et al2008) concerted evolution (Wittzell et al 1999b) andtrans-species polymorphism (Richardson and Westerdahl2003 Alcaide et al 2007) In the present study model-based tests of selection (used here for the first time inpasserine species) have demonstrated the presence ofpositively selected sites in exon 3 of MHC class I in fourpasserine genera This part of the gene encodes the alpha 2domain of the protein which is included in forming thepeptide-binding groove Positive selection acting on somecodons of exon 3 is in accordance with published data onMHC class I in other birds where the ratio of non-synonymous to synonymous substitutions (dNdS) wasusually slightly higher at PBR sites than at non-PBR ones(Westerdahl et al 1999 Richardson and Westerdahl 2003Bonneaud et al 2004) The differences however were notsignificant and this could be caused either (1) by in-correctly identified PBR sites since the MHC proteinstructure in the majority of species studied is not well-understood In fact the PBR sites have usually beenidentified on the basis of the three-dimensional structureof human HLA-A protein (Bjorkman et al 1987) A modelof the three-dimensional structure of a passerine MHC classI molecule is highly desirable Or (2) another reason mightbe the fact that some of the studied sequences come fromloci that lost their importance and they are not underselection that is why it is crucial to identify sequences toparticular loci in the future studies (see Worley et al 2008)
Our results indicate extreme variability in selectivepressure among sites in the exon 3 and the presence of anumber of sites under balancing selection Approximatelyhalf of the positively selected sites in passerines (Fig 3)have been previously assumed to play a role in antigenbinding based on the structural models (Wallny et al 2006Koch et al 2007) However we found at least twoadditional sites (amino acid residues 126 and 136) understrong positive selection in several genera which have not
been previously reported for PBR (either in human HLA orchicken BF) The maintenance of considerable nonsynon-ymous variation at these specific codons by positiveselection suggests that such polymorphism may havefunctional consequences for peptide binding and presenta-tion to T cells (see similar recent results concerning teleostfish MHC molecules Aguilar and Garza 2007 or inmammals Amills et al 2008) The predictive power ofcodon-based models has not yet been tested in MHC andthe structural model describing peptide-binding groove inpasserine birds is still required to confirm the functionalrole of amino acid residues under different types ofselection
It appears that mammalian and avian MHCs may differin frequency of genetic exchange between different genesof multigene families Concerted evolution where the inter-locus gene conversion events are frequent in highlyduplicated MHC genes may be the predominant evolu-tionary mechanism in birds (Edwards et al 1995 Alcaide etal 2007 Garrigan and Edwards 1999 Wittzell et al 1999bHess and Edwards 2002) and probably also in fish (Reuschet al 2004 van Oosterhout et al 2006 Aguilar and Garza2007) and reptiles (Miller et al 2006) but not in mammals(Edwards et al 1997) This process creates high homologyamong MHC sequences derived from various loci and itbecomes impossible to assign different MHC alleles tospecific loci In birds so far most of the studies evidencinginter-locus recombination were conducted on MHC class IIConcerning MHC class I it has only been studied in thechicken where no evidence was found for concertedevolution to occur (Shaw et al 2007) Thus our resultson recombination analyses in the finches (ie the totalamount of recombination per codon was half of that ofsubstitution and we failed to assign the sequences tospecific loci) are in agreement with previous reports andfurthermore they suggest that concerted evolution may befrequent also in MHC class I genes in passerine birds Toconfirm this hypothesis the more detailed description ofMHC structure (especially the affiliation of particularalleles to individual loci in a population and the geneticvariation outside exon 3) is necessary
Extreme MHC class I variation in a single population
Previous studies on songbirds using the method ofdenaturing gradient gel electrophoresis or restriction frag-ment length polymorphism (RFLP) have also indicatedhigh sequence variation at exon 3 of MHC class I forexample in an outbred population of house sparrows 46sequences were found in 101 individuals or between oneand 11 alleles per individual (Bonneaud et al 2006a) in thegreat reed warbler 49 different RFLP genotypes werefound in 55 birds (Westerdahl et al 2004) Even so in the
458 Immunogenetics (2009) 61451ndash461
Scarlet Rosefinch 82 alleles were found in a singlepopulation and most of the birds displayed uniquegenotypes which is the highest exon 3 MHC class Ivariability observed thus far This result may still be anunderestimate considering that in at least one case theSSCP pattern was identical for two different allelesAlthough we were not able to sequence all the allelesdisplayed on SSCP it is very probable that all the peaksidentified by SSCP are true alleles and that there could beadditional ones (due to peak overlapping) Another reasonwe believe this variation has not been overestimated is thatan extraordinarily high variability of selectively neutralmarkers has been found in this species at two microsatelliteloci more than 70 alleles were found (Albrecht et al 2007)
Interestingly one of the alleles (CaerU02) was possessedby all individuals but one female (no 515) investigated inthis study From parentndashoffspring genotypes (Promerovaacute etal unpublished data) we know that also some of thisfemales offspring missed this allele thus this absence issurely heritable We conclude that CaerU02 is the mostfrequent allele on a locus with very low variability Thefemale no 515 is either homo- or heterozygote for other veryrare allele(s) or the locus (at least the copy with alleleCaerU02) is completely missing in her MHC haplotype
Our optimization of a genotyping method for analyses ofMHC class I variation will allow us to search for answers tofundamental questions concerning recent immunoecologysuch as the role of MHC in disease resistance expression ofsecondary ornaments and mating success (eg Bonneaud etal 2006a b Loiseau et al 2008 Richardson et al 2004Westerdahl et al 2005) Furthermore our data could beused for future comparative studies of the correlationbetween MHC diversity and migratory strategies in birdsWe presume that migratory birds such as the ScarletRosefinch (which migrates from the Czech Republic toIndia ca 5000 km) might encounter a larger variety ofpathogens during their lifetime than sedentary birds (forexample there is evidence that migrating individualsencounter more malarial parasites Peacuterez-Tris and Bensch2005) In that case more diverse immune mechanismscould help them fight these pathogens Moreover it hasalready been suggested that migratory species have highergenetic variability than sedentary species (Fitzpatrick 1994)and also that the higher genetic variability of migrants hasan impact on higher occurrence of extra-pair paternity(Spottiswoode and Moslashller 2004) The hypothesis thatmigration in birds might elicit a higher variability onMHC was first evaluated by Shiina et al (2004) whocompared the MHC of migratory quail with that ofsedentary chicken The quail has a more complicatedMHC than the chicken the class I region in the quail istwice as long as that of the chicken (Shiina et al 2004) andthere are five expressed class I genes although not all of
them are classical ones (Shiina et al 2006) SimilarlyRichardson and Westerdahl (2003) reported a difference inMHC variability between the migratory Great reed warblerand the sedentary inbred Seychelles warbler In this casehowever the low MHC variation in the latter could be dueto a severe bottleneck leading to strong decrease of overallgenetic variation Furthermore the endemic Basra reedwarbler (Acrocephalus griseldis) is as variable at MHC asthe Great reed warbler even though it is sedentary (Hanssonand Richardson 2005) High level of MHC polymorphismwas found in the house sparrow which is a non-migratoryspecies (Bonneaud et al 2004) which also contradicts thepredictions of the above hypothesis Unfortunately formost species (including the Scarlet Rosefinch) there is noprecise information about the functional variation ienumber of expressed MHC variants It is evident that dataon real MHC variation in more species are inevitablyrequired to perform a rigorous comparative meta-analysisand to test the predictions of migration-MHC hypothesis
Acknowledgements We thank Pavel Munclinger Michal VinklerJan Schnitzer and Radka Polaacutekovaacute for their help in the field Thanksto Anna Bryjovaacute and Dagmar Čiacutežkovaacute for their valuable advice inthe laboratory We are very grateful to Heidi C Hauffe for improvingthe English and two anonymous referees for critical comments on themanuscript This study was funded by the Grant Agency of theAcademy of Sciences of the Czech Republic (project noIAA600930608) the Grant Agency of the Czech Rep (project no206060851) and by the Ministry of Education of the Czech Repwhose Research Centre no LC06073 and Long-term Research Plan(no MSM 0021620828) to the Charles University in Prague formed aframework for a part of this study
References
Abbas AK Lichtman AH Pober JS (1994) Cellular and molecularimmunology W B Saunders Company Philadelphia
Aguilar A Garza JC (2007) Patterns of historical balancing selectionon the salmonid major histocompatibility complex class II betagene J Mol Evol 6534ndash43 doi101007s00239-006-0222-8
Albrecht T (2004) Edge effect in wetland-arable land boundarydetermines nesting success of scarlet rosefinches Carpodacuserythrinus in the Czech Republic Auk 121361ndash371doi1016420004-8038(2004)121[0361EEIWLB]20CO2
Albrecht T Schnitzer J Kreisinger J Exnerovaacute A Bryja JMunclinger P (2007) Extrapair paternity and the opportunity forsexual selection in long-distant migratory passerines Behav Ecol18477ndash486 doi101093behecoarm001
Alcaide M Edwards SV Negro JJ (2007) Characterization polymor-phism and evolution ofMHCClass II B genes in birds of prey JMolEvol 65541ndash554 doi101007s00239-007-9033-9
Amills M Ramiacuterez O Tomaacutes A Obexer-Ruff G Vidal O (2008)Positive selection on mammalian MHC-DQ genes revisited from amultispecies perspective Genes Immun 9651ndash658 doi101038gene200862
Anisimova M Nielsen R Yang Z (2003) Effect of recombination onthe accuracy of the likelihood method for detecting positiveselection at amino acid sites Genetics 1641229ndash1236
Immunogenetics (2009) 61451ndash461 459
Bjorkman PJ Saper MA Samraoui B Bennett WS Strominger JLWiley DC (1987) Structure of human class I histocompatibil-ity antigen HLA-A2 Nature 329506ndash512 doi101038329506a0
Bonneaud C Sorci G Morin V Westerdahl H Zoorob R Wittzell H(2004) Diversity of MHC class I and IIB genes in house sparrows(Passer domesticus) Immunogenetics 55855ndash865 doi101007s00251-004-0648-3
Bonneaud C Chastel O Federici P Westerdahl H Sorci G (2006a)Complex MHC-based mate choice in a wild passerine Proc RSoc 2731111ndash1116 doi101098rspb20053325
Bonneaud C Peacuterez-Tris J Federici P Chastel O Sorci G (2006b) Majorhistocompatibility alleles associated with local resistance to malariain a passerine Evolution Int J Org Evolution 60383ndash389
Bryja J Galan M Charbonnel N Cosson J-F (2005) Analysis ofmajor histocompatibility complex class II gene in water volesusing capillary electrophoresis-single stranded conformationpolymorphism Mol Ecol Notes 5173ndash176 doi101111j1471-8286200400855x
Burri R Niculita-Hirzel H Roulin A Fumagalli L (2008) Isolation andcharacterization of major histocompatibility complex (MHC) class IIB genes in teh Barn owl (Aves Tyto alba) Immunogenetics 60543ndash550 doi101007s00251-008-0308-0
Cramp S Simmons KEL Perrins CM (eds) (1977-1994) The birds ofthe Western Palearctic Oxford University Press Oxford
Edwards SV Wakeland EK Potts WK (1995) Contrasting histories ofavian and mammalian MHC genes revealed by class II B genesof songbirds Proc Natl Acad Sci U S A 9212200ndash12204doi101073pnas922612200
Edwards SV Chesnut K Satta Y Wakeland EK (1997) Ancestralpolymorphism of Mhc Class II genes in mice implications forbalancing selection and the mammalian molecular clock Genetics146655ndash668
Fitzpatrick S (1994) Colourful migratory birds evidence for amechanism other than parasite resistance for the maintenance oflsquogood genesrsquo sexual selection Proc R Soc Lond B Biol Sci257155ndash166 doi101098rspb19940109
Freeman-Gallant CR Johnson EM Saponara F Stanger M (2002)Variation at the major histocompatibility complex in Savannahsparrows Mol Ecol 111125ndash1130 doi101046j1365-294X200201508x
Garrigan D Edwards SV (1999) Polymorphism across an exon-intronboundary in an avian Mhc class II B gene Mol Biol Evol 161599ndash1606
Gouumly de Bellocq J Charbonnel N Morand S (2008) Coevolutionaryrelationship between helminth diversity and Mhc class II polymor-phism in rodents J Evol Biol 211144ndash1150 doi101111j1420-9101200801538x
Hall TA (1999) BioEdit a user-friendly biological sequence alignmenteditor and analisis program for Windows 9598NT NucleicAcids Symp Ser 4195ndash98
Hansson B Richardson DS (2005) Genetic variation in twoendangered Acrocephalus species compared to a widespreadcongener estimates based on functional and random loci AnimConserv 883ndash90 doi101017S1367943004001878
Hess CM Edwards SV (2002) The evolution of major histocompat-ibility genes in birds Bioscience 52423ndash431 doi1016410006-3568(2002)052[0423TEOTMH]20CO2
Hess CM Gasper J Hoekstra H Hill C Edwards SV (2000) MHCclass II pseudogene and genomic signature of a 32-kb cosmid inthe House Finch (Carpodacus mexicanus) Genome Res 1013ndash23 doi101101gr105613
Hosomichi K Shiina T Suzuki S Tanaka M Shimizu S Iwamoto SHara H Yoshida Y Kulski JK Inoko H Hanzawa K (2006) Themajor histocompatibility complex (Mhc) class IIB region hasgreater genomic structural flexibility and diversity in the quail
than the chicken BMC Genomics 7322ndash335 doi1011861471-2164-7-322
Hughes AL Yeager M (1998) Natural selection at major histocom-patibility complex loci of vertebrates Annu Rev Genet 32415ndash435 doi101146annurevgenet321415
Hughes CR Miles S Walbroehl JM (2008) Support for the minimalessential MHC hypothesis a parrot with a single highlypolymorphic MHC class II B gene Immunogenetics 60219ndash231 doi101007s00251-008-0287-1
Indovina P Megiorni F Fontemaggi G Coni P Mora B Mazzilli MC(2001) Absence of in vivo DNA-Protein interactions in theDQA2 and DQB2 promoter regions Hum Immunol 62504ndash508doi101016S0198-8859(01)00236-1
Jarvi SI Goto RM Gee GF Briles WE Miller MM (1999)Identification inheritance and linkage of B-G-like and MHCclass I genes in cranes Am Genet Assoc 90152ndash159
Kaufman J (1999) Co-evolving genes in MHC haplotypes the lsquorulersquofor nonmammalian vertebrates Immunogenetics 50228ndash236doi101007s002510050597
Kaufman J Salamonsen J Flajnik M (1994) Evolutionary conserva-tion of MHC class I and class II molecules-different yet the sameSemin Immunol 6411ndash424 doi101006smim19941050
Kaufman J Voumllk H Wallny H (1995) A bdquominimal essential MHCldquoand an bdquounrecognized MHCldquo two extremes in selection forpolymorphism Immunol Rev 14363ndash88 doi101111j1600-065X1995tb00670x
Kaufman J Jacob J Shaw I Walker B Milne S Beck S SalomonsenJ (1999) Gene organisation determines evolution of function inthe chicken MHC Immunol Rev 167101ndash117 doi101111j1600-065X1999tb01385x
Klein J (1986) Natural history of the major histocompatibilitycomplex Wiley New York
Klein J Sato A Nagl S Oh Uigin C (1998) Molecular trans-speciespolymorphism Annu Rev Ecol Syst 291ndash21 doi101146annurevecolsys2911
Koch M Camp S Collen T Avila D Salomonsen J Wallny HJ vanHateren A Hunt L Jacob JP Johnston F Marston DA Shaw IDunbar PR Cerundolo V Jones EY Kaufman J (2007)Structures of an MHC class I molecule from B21 chickensillustrate promiscuous peptide binding Immunity 27885ndash899doi101016jimmuni200711007
Kosakovsky Pond SL Frost SDW (2005) Not so different after all acomparison of methods for detecting amino acid sites under selectionMol Biol Evol 221208ndash1222 doi101093molbevmsi105
Kosakovsky Pond SL Frost SDW Muse SV (2005) HyPhyhypothesis testing using phylogenies Bioinformatics 21676ndash679 doi101093bioinformaticsbti079
Kosakovsky Pond SL Posada D Gravenor MB Woelk CH FrostSDW (2006) Automated phylogenetic detection of recombinationusing a genetic algorithm Mol Biol Evol 231891ndash1901doi101093molbevmsl051
Loiseau C Zoorob R Garnier S Birard J Federici P Julliard R SorciG (2008) Antagonistic effects of a MHC class I allele on malaria-infected house sparrows Ecol Lett 11258ndash265 doi101111j1461-0248200701141x
Mesa CM Thulien KJ Moon DA Veniamin SM Magor KE (2004)The dominant MHC class I gene is adjacent to the polymorphicTAP2 gene in the duck Anas platyrhynchos Immunogenetics56192ndash203 doi101007s00251-004-0672-3
Miller HC Lambert DM (2004) Gene duplication and geneconversion in class II MHC genes of New Zealand robins(Petroicidae) Immunogenetics 56178ndash191
Miller MM Goto RM Bernot A Zoorob R Auffray C Bumstead NBriles WE (1994) Two Mhc class I and two Mhc class II genes mapto the chicken Rfp-Y system outside the B complex Proc Natl AcadSci U S A 914397ndash4401 doi101073pnas91104397
460 Immunogenetics (2009) 61451ndash461
Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461

different cDNA sequences were also found in an individualIn a separate study of the same species 20 genomicsequences of exon 3 of class I were found in fiveindividuals five of which were pseudogenes (Westerdahlet al 2004) Similar results were found in house sparrowswhere 20 sequences were verified in seven individualsthree of which were proved to be transcribed althoughthere were no pseudogenes (Bonneaud et al 2004) Ourresults concerning individual exon 3 variation are similar tothe ones mentioned above suggesting that complex MHCstructure could be a general pattern in songbirds
Evolution of MHC class I sequences in passerines
Several mechanisms of molecular evolution have beenalready documented in the avian MHC sequences includ-ing positive selection (Alcaide et al 2007 Hughes et al2008) concerted evolution (Wittzell et al 1999b) andtrans-species polymorphism (Richardson and Westerdahl2003 Alcaide et al 2007) In the present study model-based tests of selection (used here for the first time inpasserine species) have demonstrated the presence ofpositively selected sites in exon 3 of MHC class I in fourpasserine genera This part of the gene encodes the alpha 2domain of the protein which is included in forming thepeptide-binding groove Positive selection acting on somecodons of exon 3 is in accordance with published data onMHC class I in other birds where the ratio of non-synonymous to synonymous substitutions (dNdS) wasusually slightly higher at PBR sites than at non-PBR ones(Westerdahl et al 1999 Richardson and Westerdahl 2003Bonneaud et al 2004) The differences however were notsignificant and this could be caused either (1) by in-correctly identified PBR sites since the MHC proteinstructure in the majority of species studied is not well-understood In fact the PBR sites have usually beenidentified on the basis of the three-dimensional structureof human HLA-A protein (Bjorkman et al 1987) A modelof the three-dimensional structure of a passerine MHC classI molecule is highly desirable Or (2) another reason mightbe the fact that some of the studied sequences come fromloci that lost their importance and they are not underselection that is why it is crucial to identify sequences toparticular loci in the future studies (see Worley et al 2008)
Our results indicate extreme variability in selectivepressure among sites in the exon 3 and the presence of anumber of sites under balancing selection Approximatelyhalf of the positively selected sites in passerines (Fig 3)have been previously assumed to play a role in antigenbinding based on the structural models (Wallny et al 2006Koch et al 2007) However we found at least twoadditional sites (amino acid residues 126 and 136) understrong positive selection in several genera which have not
been previously reported for PBR (either in human HLA orchicken BF) The maintenance of considerable nonsynon-ymous variation at these specific codons by positiveselection suggests that such polymorphism may havefunctional consequences for peptide binding and presenta-tion to T cells (see similar recent results concerning teleostfish MHC molecules Aguilar and Garza 2007 or inmammals Amills et al 2008) The predictive power ofcodon-based models has not yet been tested in MHC andthe structural model describing peptide-binding groove inpasserine birds is still required to confirm the functionalrole of amino acid residues under different types ofselection
It appears that mammalian and avian MHCs may differin frequency of genetic exchange between different genesof multigene families Concerted evolution where the inter-locus gene conversion events are frequent in highlyduplicated MHC genes may be the predominant evolu-tionary mechanism in birds (Edwards et al 1995 Alcaide etal 2007 Garrigan and Edwards 1999 Wittzell et al 1999bHess and Edwards 2002) and probably also in fish (Reuschet al 2004 van Oosterhout et al 2006 Aguilar and Garza2007) and reptiles (Miller et al 2006) but not in mammals(Edwards et al 1997) This process creates high homologyamong MHC sequences derived from various loci and itbecomes impossible to assign different MHC alleles tospecific loci In birds so far most of the studies evidencinginter-locus recombination were conducted on MHC class IIConcerning MHC class I it has only been studied in thechicken where no evidence was found for concertedevolution to occur (Shaw et al 2007) Thus our resultson recombination analyses in the finches (ie the totalamount of recombination per codon was half of that ofsubstitution and we failed to assign the sequences tospecific loci) are in agreement with previous reports andfurthermore they suggest that concerted evolution may befrequent also in MHC class I genes in passerine birds Toconfirm this hypothesis the more detailed description ofMHC structure (especially the affiliation of particularalleles to individual loci in a population and the geneticvariation outside exon 3) is necessary
Extreme MHC class I variation in a single population
Previous studies on songbirds using the method ofdenaturing gradient gel electrophoresis or restriction frag-ment length polymorphism (RFLP) have also indicatedhigh sequence variation at exon 3 of MHC class I forexample in an outbred population of house sparrows 46sequences were found in 101 individuals or between oneand 11 alleles per individual (Bonneaud et al 2006a) in thegreat reed warbler 49 different RFLP genotypes werefound in 55 birds (Westerdahl et al 2004) Even so in the
458 Immunogenetics (2009) 61451ndash461
Scarlet Rosefinch 82 alleles were found in a singlepopulation and most of the birds displayed uniquegenotypes which is the highest exon 3 MHC class Ivariability observed thus far This result may still be anunderestimate considering that in at least one case theSSCP pattern was identical for two different allelesAlthough we were not able to sequence all the allelesdisplayed on SSCP it is very probable that all the peaksidentified by SSCP are true alleles and that there could beadditional ones (due to peak overlapping) Another reasonwe believe this variation has not been overestimated is thatan extraordinarily high variability of selectively neutralmarkers has been found in this species at two microsatelliteloci more than 70 alleles were found (Albrecht et al 2007)
Interestingly one of the alleles (CaerU02) was possessedby all individuals but one female (no 515) investigated inthis study From parentndashoffspring genotypes (Promerovaacute etal unpublished data) we know that also some of thisfemales offspring missed this allele thus this absence issurely heritable We conclude that CaerU02 is the mostfrequent allele on a locus with very low variability Thefemale no 515 is either homo- or heterozygote for other veryrare allele(s) or the locus (at least the copy with alleleCaerU02) is completely missing in her MHC haplotype
Our optimization of a genotyping method for analyses ofMHC class I variation will allow us to search for answers tofundamental questions concerning recent immunoecologysuch as the role of MHC in disease resistance expression ofsecondary ornaments and mating success (eg Bonneaud etal 2006a b Loiseau et al 2008 Richardson et al 2004Westerdahl et al 2005) Furthermore our data could beused for future comparative studies of the correlationbetween MHC diversity and migratory strategies in birdsWe presume that migratory birds such as the ScarletRosefinch (which migrates from the Czech Republic toIndia ca 5000 km) might encounter a larger variety ofpathogens during their lifetime than sedentary birds (forexample there is evidence that migrating individualsencounter more malarial parasites Peacuterez-Tris and Bensch2005) In that case more diverse immune mechanismscould help them fight these pathogens Moreover it hasalready been suggested that migratory species have highergenetic variability than sedentary species (Fitzpatrick 1994)and also that the higher genetic variability of migrants hasan impact on higher occurrence of extra-pair paternity(Spottiswoode and Moslashller 2004) The hypothesis thatmigration in birds might elicit a higher variability onMHC was first evaluated by Shiina et al (2004) whocompared the MHC of migratory quail with that ofsedentary chicken The quail has a more complicatedMHC than the chicken the class I region in the quail istwice as long as that of the chicken (Shiina et al 2004) andthere are five expressed class I genes although not all of
them are classical ones (Shiina et al 2006) SimilarlyRichardson and Westerdahl (2003) reported a difference inMHC variability between the migratory Great reed warblerand the sedentary inbred Seychelles warbler In this casehowever the low MHC variation in the latter could be dueto a severe bottleneck leading to strong decrease of overallgenetic variation Furthermore the endemic Basra reedwarbler (Acrocephalus griseldis) is as variable at MHC asthe Great reed warbler even though it is sedentary (Hanssonand Richardson 2005) High level of MHC polymorphismwas found in the house sparrow which is a non-migratoryspecies (Bonneaud et al 2004) which also contradicts thepredictions of the above hypothesis Unfortunately formost species (including the Scarlet Rosefinch) there is noprecise information about the functional variation ienumber of expressed MHC variants It is evident that dataon real MHC variation in more species are inevitablyrequired to perform a rigorous comparative meta-analysisand to test the predictions of migration-MHC hypothesis
Acknowledgements We thank Pavel Munclinger Michal VinklerJan Schnitzer and Radka Polaacutekovaacute for their help in the field Thanksto Anna Bryjovaacute and Dagmar Čiacutežkovaacute for their valuable advice inthe laboratory We are very grateful to Heidi C Hauffe for improvingthe English and two anonymous referees for critical comments on themanuscript This study was funded by the Grant Agency of theAcademy of Sciences of the Czech Republic (project noIAA600930608) the Grant Agency of the Czech Rep (project no206060851) and by the Ministry of Education of the Czech Repwhose Research Centre no LC06073 and Long-term Research Plan(no MSM 0021620828) to the Charles University in Prague formed aframework for a part of this study
References
Abbas AK Lichtman AH Pober JS (1994) Cellular and molecularimmunology W B Saunders Company Philadelphia
Aguilar A Garza JC (2007) Patterns of historical balancing selectionon the salmonid major histocompatibility complex class II betagene J Mol Evol 6534ndash43 doi101007s00239-006-0222-8
Albrecht T (2004) Edge effect in wetland-arable land boundarydetermines nesting success of scarlet rosefinches Carpodacuserythrinus in the Czech Republic Auk 121361ndash371doi1016420004-8038(2004)121[0361EEIWLB]20CO2
Albrecht T Schnitzer J Kreisinger J Exnerovaacute A Bryja JMunclinger P (2007) Extrapair paternity and the opportunity forsexual selection in long-distant migratory passerines Behav Ecol18477ndash486 doi101093behecoarm001
Alcaide M Edwards SV Negro JJ (2007) Characterization polymor-phism and evolution ofMHCClass II B genes in birds of prey JMolEvol 65541ndash554 doi101007s00239-007-9033-9
Amills M Ramiacuterez O Tomaacutes A Obexer-Ruff G Vidal O (2008)Positive selection on mammalian MHC-DQ genes revisited from amultispecies perspective Genes Immun 9651ndash658 doi101038gene200862
Anisimova M Nielsen R Yang Z (2003) Effect of recombination onthe accuracy of the likelihood method for detecting positiveselection at amino acid sites Genetics 1641229ndash1236
Immunogenetics (2009) 61451ndash461 459
Bjorkman PJ Saper MA Samraoui B Bennett WS Strominger JLWiley DC (1987) Structure of human class I histocompatibil-ity antigen HLA-A2 Nature 329506ndash512 doi101038329506a0
Bonneaud C Sorci G Morin V Westerdahl H Zoorob R Wittzell H(2004) Diversity of MHC class I and IIB genes in house sparrows(Passer domesticus) Immunogenetics 55855ndash865 doi101007s00251-004-0648-3
Bonneaud C Chastel O Federici P Westerdahl H Sorci G (2006a)Complex MHC-based mate choice in a wild passerine Proc RSoc 2731111ndash1116 doi101098rspb20053325
Bonneaud C Peacuterez-Tris J Federici P Chastel O Sorci G (2006b) Majorhistocompatibility alleles associated with local resistance to malariain a passerine Evolution Int J Org Evolution 60383ndash389
Bryja J Galan M Charbonnel N Cosson J-F (2005) Analysis ofmajor histocompatibility complex class II gene in water volesusing capillary electrophoresis-single stranded conformationpolymorphism Mol Ecol Notes 5173ndash176 doi101111j1471-8286200400855x
Burri R Niculita-Hirzel H Roulin A Fumagalli L (2008) Isolation andcharacterization of major histocompatibility complex (MHC) class IIB genes in teh Barn owl (Aves Tyto alba) Immunogenetics 60543ndash550 doi101007s00251-008-0308-0
Cramp S Simmons KEL Perrins CM (eds) (1977-1994) The birds ofthe Western Palearctic Oxford University Press Oxford
Edwards SV Wakeland EK Potts WK (1995) Contrasting histories ofavian and mammalian MHC genes revealed by class II B genesof songbirds Proc Natl Acad Sci U S A 9212200ndash12204doi101073pnas922612200
Edwards SV Chesnut K Satta Y Wakeland EK (1997) Ancestralpolymorphism of Mhc Class II genes in mice implications forbalancing selection and the mammalian molecular clock Genetics146655ndash668
Fitzpatrick S (1994) Colourful migratory birds evidence for amechanism other than parasite resistance for the maintenance oflsquogood genesrsquo sexual selection Proc R Soc Lond B Biol Sci257155ndash166 doi101098rspb19940109
Freeman-Gallant CR Johnson EM Saponara F Stanger M (2002)Variation at the major histocompatibility complex in Savannahsparrows Mol Ecol 111125ndash1130 doi101046j1365-294X200201508x
Garrigan D Edwards SV (1999) Polymorphism across an exon-intronboundary in an avian Mhc class II B gene Mol Biol Evol 161599ndash1606
Gouumly de Bellocq J Charbonnel N Morand S (2008) Coevolutionaryrelationship between helminth diversity and Mhc class II polymor-phism in rodents J Evol Biol 211144ndash1150 doi101111j1420-9101200801538x
Hall TA (1999) BioEdit a user-friendly biological sequence alignmenteditor and analisis program for Windows 9598NT NucleicAcids Symp Ser 4195ndash98
Hansson B Richardson DS (2005) Genetic variation in twoendangered Acrocephalus species compared to a widespreadcongener estimates based on functional and random loci AnimConserv 883ndash90 doi101017S1367943004001878
Hess CM Edwards SV (2002) The evolution of major histocompat-ibility genes in birds Bioscience 52423ndash431 doi1016410006-3568(2002)052[0423TEOTMH]20CO2
Hess CM Gasper J Hoekstra H Hill C Edwards SV (2000) MHCclass II pseudogene and genomic signature of a 32-kb cosmid inthe House Finch (Carpodacus mexicanus) Genome Res 1013ndash23 doi101101gr105613
Hosomichi K Shiina T Suzuki S Tanaka M Shimizu S Iwamoto SHara H Yoshida Y Kulski JK Inoko H Hanzawa K (2006) Themajor histocompatibility complex (Mhc) class IIB region hasgreater genomic structural flexibility and diversity in the quail
than the chicken BMC Genomics 7322ndash335 doi1011861471-2164-7-322
Hughes AL Yeager M (1998) Natural selection at major histocom-patibility complex loci of vertebrates Annu Rev Genet 32415ndash435 doi101146annurevgenet321415
Hughes CR Miles S Walbroehl JM (2008) Support for the minimalessential MHC hypothesis a parrot with a single highlypolymorphic MHC class II B gene Immunogenetics 60219ndash231 doi101007s00251-008-0287-1
Indovina P Megiorni F Fontemaggi G Coni P Mora B Mazzilli MC(2001) Absence of in vivo DNA-Protein interactions in theDQA2 and DQB2 promoter regions Hum Immunol 62504ndash508doi101016S0198-8859(01)00236-1
Jarvi SI Goto RM Gee GF Briles WE Miller MM (1999)Identification inheritance and linkage of B-G-like and MHCclass I genes in cranes Am Genet Assoc 90152ndash159
Kaufman J (1999) Co-evolving genes in MHC haplotypes the lsquorulersquofor nonmammalian vertebrates Immunogenetics 50228ndash236doi101007s002510050597
Kaufman J Salamonsen J Flajnik M (1994) Evolutionary conserva-tion of MHC class I and class II molecules-different yet the sameSemin Immunol 6411ndash424 doi101006smim19941050
Kaufman J Voumllk H Wallny H (1995) A bdquominimal essential MHCldquoand an bdquounrecognized MHCldquo two extremes in selection forpolymorphism Immunol Rev 14363ndash88 doi101111j1600-065X1995tb00670x
Kaufman J Jacob J Shaw I Walker B Milne S Beck S SalomonsenJ (1999) Gene organisation determines evolution of function inthe chicken MHC Immunol Rev 167101ndash117 doi101111j1600-065X1999tb01385x
Klein J (1986) Natural history of the major histocompatibilitycomplex Wiley New York
Klein J Sato A Nagl S Oh Uigin C (1998) Molecular trans-speciespolymorphism Annu Rev Ecol Syst 291ndash21 doi101146annurevecolsys2911
Koch M Camp S Collen T Avila D Salomonsen J Wallny HJ vanHateren A Hunt L Jacob JP Johnston F Marston DA Shaw IDunbar PR Cerundolo V Jones EY Kaufman J (2007)Structures of an MHC class I molecule from B21 chickensillustrate promiscuous peptide binding Immunity 27885ndash899doi101016jimmuni200711007
Kosakovsky Pond SL Frost SDW (2005) Not so different after all acomparison of methods for detecting amino acid sites under selectionMol Biol Evol 221208ndash1222 doi101093molbevmsi105
Kosakovsky Pond SL Frost SDW Muse SV (2005) HyPhyhypothesis testing using phylogenies Bioinformatics 21676ndash679 doi101093bioinformaticsbti079
Kosakovsky Pond SL Posada D Gravenor MB Woelk CH FrostSDW (2006) Automated phylogenetic detection of recombinationusing a genetic algorithm Mol Biol Evol 231891ndash1901doi101093molbevmsl051
Loiseau C Zoorob R Garnier S Birard J Federici P Julliard R SorciG (2008) Antagonistic effects of a MHC class I allele on malaria-infected house sparrows Ecol Lett 11258ndash265 doi101111j1461-0248200701141x
Mesa CM Thulien KJ Moon DA Veniamin SM Magor KE (2004)The dominant MHC class I gene is adjacent to the polymorphicTAP2 gene in the duck Anas platyrhynchos Immunogenetics56192ndash203 doi101007s00251-004-0672-3
Miller HC Lambert DM (2004) Gene duplication and geneconversion in class II MHC genes of New Zealand robins(Petroicidae) Immunogenetics 56178ndash191
Miller MM Goto RM Bernot A Zoorob R Auffray C Bumstead NBriles WE (1994) Two Mhc class I and two Mhc class II genes mapto the chicken Rfp-Y system outside the B complex Proc Natl AcadSci U S A 914397ndash4401 doi101073pnas91104397
460 Immunogenetics (2009) 61451ndash461
Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461

Scarlet Rosefinch 82 alleles were found in a singlepopulation and most of the birds displayed uniquegenotypes which is the highest exon 3 MHC class Ivariability observed thus far This result may still be anunderestimate considering that in at least one case theSSCP pattern was identical for two different allelesAlthough we were not able to sequence all the allelesdisplayed on SSCP it is very probable that all the peaksidentified by SSCP are true alleles and that there could beadditional ones (due to peak overlapping) Another reasonwe believe this variation has not been overestimated is thatan extraordinarily high variability of selectively neutralmarkers has been found in this species at two microsatelliteloci more than 70 alleles were found (Albrecht et al 2007)
Interestingly one of the alleles (CaerU02) was possessedby all individuals but one female (no 515) investigated inthis study From parentndashoffspring genotypes (Promerovaacute etal unpublished data) we know that also some of thisfemales offspring missed this allele thus this absence issurely heritable We conclude that CaerU02 is the mostfrequent allele on a locus with very low variability Thefemale no 515 is either homo- or heterozygote for other veryrare allele(s) or the locus (at least the copy with alleleCaerU02) is completely missing in her MHC haplotype
Our optimization of a genotyping method for analyses ofMHC class I variation will allow us to search for answers tofundamental questions concerning recent immunoecologysuch as the role of MHC in disease resistance expression ofsecondary ornaments and mating success (eg Bonneaud etal 2006a b Loiseau et al 2008 Richardson et al 2004Westerdahl et al 2005) Furthermore our data could beused for future comparative studies of the correlationbetween MHC diversity and migratory strategies in birdsWe presume that migratory birds such as the ScarletRosefinch (which migrates from the Czech Republic toIndia ca 5000 km) might encounter a larger variety ofpathogens during their lifetime than sedentary birds (forexample there is evidence that migrating individualsencounter more malarial parasites Peacuterez-Tris and Bensch2005) In that case more diverse immune mechanismscould help them fight these pathogens Moreover it hasalready been suggested that migratory species have highergenetic variability than sedentary species (Fitzpatrick 1994)and also that the higher genetic variability of migrants hasan impact on higher occurrence of extra-pair paternity(Spottiswoode and Moslashller 2004) The hypothesis thatmigration in birds might elicit a higher variability onMHC was first evaluated by Shiina et al (2004) whocompared the MHC of migratory quail with that ofsedentary chicken The quail has a more complicatedMHC than the chicken the class I region in the quail istwice as long as that of the chicken (Shiina et al 2004) andthere are five expressed class I genes although not all of
them are classical ones (Shiina et al 2006) SimilarlyRichardson and Westerdahl (2003) reported a difference inMHC variability between the migratory Great reed warblerand the sedentary inbred Seychelles warbler In this casehowever the low MHC variation in the latter could be dueto a severe bottleneck leading to strong decrease of overallgenetic variation Furthermore the endemic Basra reedwarbler (Acrocephalus griseldis) is as variable at MHC asthe Great reed warbler even though it is sedentary (Hanssonand Richardson 2005) High level of MHC polymorphismwas found in the house sparrow which is a non-migratoryspecies (Bonneaud et al 2004) which also contradicts thepredictions of the above hypothesis Unfortunately formost species (including the Scarlet Rosefinch) there is noprecise information about the functional variation ienumber of expressed MHC variants It is evident that dataon real MHC variation in more species are inevitablyrequired to perform a rigorous comparative meta-analysisand to test the predictions of migration-MHC hypothesis
Acknowledgements We thank Pavel Munclinger Michal VinklerJan Schnitzer and Radka Polaacutekovaacute for their help in the field Thanksto Anna Bryjovaacute and Dagmar Čiacutežkovaacute for their valuable advice inthe laboratory We are very grateful to Heidi C Hauffe for improvingthe English and two anonymous referees for critical comments on themanuscript This study was funded by the Grant Agency of theAcademy of Sciences of the Czech Republic (project noIAA600930608) the Grant Agency of the Czech Rep (project no206060851) and by the Ministry of Education of the Czech Repwhose Research Centre no LC06073 and Long-term Research Plan(no MSM 0021620828) to the Charles University in Prague formed aframework for a part of this study
References
Abbas AK Lichtman AH Pober JS (1994) Cellular and molecularimmunology W B Saunders Company Philadelphia
Aguilar A Garza JC (2007) Patterns of historical balancing selectionon the salmonid major histocompatibility complex class II betagene J Mol Evol 6534ndash43 doi101007s00239-006-0222-8
Albrecht T (2004) Edge effect in wetland-arable land boundarydetermines nesting success of scarlet rosefinches Carpodacuserythrinus in the Czech Republic Auk 121361ndash371doi1016420004-8038(2004)121[0361EEIWLB]20CO2
Albrecht T Schnitzer J Kreisinger J Exnerovaacute A Bryja JMunclinger P (2007) Extrapair paternity and the opportunity forsexual selection in long-distant migratory passerines Behav Ecol18477ndash486 doi101093behecoarm001
Alcaide M Edwards SV Negro JJ (2007) Characterization polymor-phism and evolution ofMHCClass II B genes in birds of prey JMolEvol 65541ndash554 doi101007s00239-007-9033-9
Amills M Ramiacuterez O Tomaacutes A Obexer-Ruff G Vidal O (2008)Positive selection on mammalian MHC-DQ genes revisited from amultispecies perspective Genes Immun 9651ndash658 doi101038gene200862
Anisimova M Nielsen R Yang Z (2003) Effect of recombination onthe accuracy of the likelihood method for detecting positiveselection at amino acid sites Genetics 1641229ndash1236
Immunogenetics (2009) 61451ndash461 459
Bjorkman PJ Saper MA Samraoui B Bennett WS Strominger JLWiley DC (1987) Structure of human class I histocompatibil-ity antigen HLA-A2 Nature 329506ndash512 doi101038329506a0
Bonneaud C Sorci G Morin V Westerdahl H Zoorob R Wittzell H(2004) Diversity of MHC class I and IIB genes in house sparrows(Passer domesticus) Immunogenetics 55855ndash865 doi101007s00251-004-0648-3
Bonneaud C Chastel O Federici P Westerdahl H Sorci G (2006a)Complex MHC-based mate choice in a wild passerine Proc RSoc 2731111ndash1116 doi101098rspb20053325
Bonneaud C Peacuterez-Tris J Federici P Chastel O Sorci G (2006b) Majorhistocompatibility alleles associated with local resistance to malariain a passerine Evolution Int J Org Evolution 60383ndash389
Bryja J Galan M Charbonnel N Cosson J-F (2005) Analysis ofmajor histocompatibility complex class II gene in water volesusing capillary electrophoresis-single stranded conformationpolymorphism Mol Ecol Notes 5173ndash176 doi101111j1471-8286200400855x
Burri R Niculita-Hirzel H Roulin A Fumagalli L (2008) Isolation andcharacterization of major histocompatibility complex (MHC) class IIB genes in teh Barn owl (Aves Tyto alba) Immunogenetics 60543ndash550 doi101007s00251-008-0308-0
Cramp S Simmons KEL Perrins CM (eds) (1977-1994) The birds ofthe Western Palearctic Oxford University Press Oxford
Edwards SV Wakeland EK Potts WK (1995) Contrasting histories ofavian and mammalian MHC genes revealed by class II B genesof songbirds Proc Natl Acad Sci U S A 9212200ndash12204doi101073pnas922612200
Edwards SV Chesnut K Satta Y Wakeland EK (1997) Ancestralpolymorphism of Mhc Class II genes in mice implications forbalancing selection and the mammalian molecular clock Genetics146655ndash668
Fitzpatrick S (1994) Colourful migratory birds evidence for amechanism other than parasite resistance for the maintenance oflsquogood genesrsquo sexual selection Proc R Soc Lond B Biol Sci257155ndash166 doi101098rspb19940109
Freeman-Gallant CR Johnson EM Saponara F Stanger M (2002)Variation at the major histocompatibility complex in Savannahsparrows Mol Ecol 111125ndash1130 doi101046j1365-294X200201508x
Garrigan D Edwards SV (1999) Polymorphism across an exon-intronboundary in an avian Mhc class II B gene Mol Biol Evol 161599ndash1606
Gouumly de Bellocq J Charbonnel N Morand S (2008) Coevolutionaryrelationship between helminth diversity and Mhc class II polymor-phism in rodents J Evol Biol 211144ndash1150 doi101111j1420-9101200801538x
Hall TA (1999) BioEdit a user-friendly biological sequence alignmenteditor and analisis program for Windows 9598NT NucleicAcids Symp Ser 4195ndash98
Hansson B Richardson DS (2005) Genetic variation in twoendangered Acrocephalus species compared to a widespreadcongener estimates based on functional and random loci AnimConserv 883ndash90 doi101017S1367943004001878
Hess CM Edwards SV (2002) The evolution of major histocompat-ibility genes in birds Bioscience 52423ndash431 doi1016410006-3568(2002)052[0423TEOTMH]20CO2
Hess CM Gasper J Hoekstra H Hill C Edwards SV (2000) MHCclass II pseudogene and genomic signature of a 32-kb cosmid inthe House Finch (Carpodacus mexicanus) Genome Res 1013ndash23 doi101101gr105613
Hosomichi K Shiina T Suzuki S Tanaka M Shimizu S Iwamoto SHara H Yoshida Y Kulski JK Inoko H Hanzawa K (2006) Themajor histocompatibility complex (Mhc) class IIB region hasgreater genomic structural flexibility and diversity in the quail
than the chicken BMC Genomics 7322ndash335 doi1011861471-2164-7-322
Hughes AL Yeager M (1998) Natural selection at major histocom-patibility complex loci of vertebrates Annu Rev Genet 32415ndash435 doi101146annurevgenet321415
Hughes CR Miles S Walbroehl JM (2008) Support for the minimalessential MHC hypothesis a parrot with a single highlypolymorphic MHC class II B gene Immunogenetics 60219ndash231 doi101007s00251-008-0287-1
Indovina P Megiorni F Fontemaggi G Coni P Mora B Mazzilli MC(2001) Absence of in vivo DNA-Protein interactions in theDQA2 and DQB2 promoter regions Hum Immunol 62504ndash508doi101016S0198-8859(01)00236-1
Jarvi SI Goto RM Gee GF Briles WE Miller MM (1999)Identification inheritance and linkage of B-G-like and MHCclass I genes in cranes Am Genet Assoc 90152ndash159
Kaufman J (1999) Co-evolving genes in MHC haplotypes the lsquorulersquofor nonmammalian vertebrates Immunogenetics 50228ndash236doi101007s002510050597
Kaufman J Salamonsen J Flajnik M (1994) Evolutionary conserva-tion of MHC class I and class II molecules-different yet the sameSemin Immunol 6411ndash424 doi101006smim19941050
Kaufman J Voumllk H Wallny H (1995) A bdquominimal essential MHCldquoand an bdquounrecognized MHCldquo two extremes in selection forpolymorphism Immunol Rev 14363ndash88 doi101111j1600-065X1995tb00670x
Kaufman J Jacob J Shaw I Walker B Milne S Beck S SalomonsenJ (1999) Gene organisation determines evolution of function inthe chicken MHC Immunol Rev 167101ndash117 doi101111j1600-065X1999tb01385x
Klein J (1986) Natural history of the major histocompatibilitycomplex Wiley New York
Klein J Sato A Nagl S Oh Uigin C (1998) Molecular trans-speciespolymorphism Annu Rev Ecol Syst 291ndash21 doi101146annurevecolsys2911
Koch M Camp S Collen T Avila D Salomonsen J Wallny HJ vanHateren A Hunt L Jacob JP Johnston F Marston DA Shaw IDunbar PR Cerundolo V Jones EY Kaufman J (2007)Structures of an MHC class I molecule from B21 chickensillustrate promiscuous peptide binding Immunity 27885ndash899doi101016jimmuni200711007
Kosakovsky Pond SL Frost SDW (2005) Not so different after all acomparison of methods for detecting amino acid sites under selectionMol Biol Evol 221208ndash1222 doi101093molbevmsi105
Kosakovsky Pond SL Frost SDW Muse SV (2005) HyPhyhypothesis testing using phylogenies Bioinformatics 21676ndash679 doi101093bioinformaticsbti079
Kosakovsky Pond SL Posada D Gravenor MB Woelk CH FrostSDW (2006) Automated phylogenetic detection of recombinationusing a genetic algorithm Mol Biol Evol 231891ndash1901doi101093molbevmsl051
Loiseau C Zoorob R Garnier S Birard J Federici P Julliard R SorciG (2008) Antagonistic effects of a MHC class I allele on malaria-infected house sparrows Ecol Lett 11258ndash265 doi101111j1461-0248200701141x
Mesa CM Thulien KJ Moon DA Veniamin SM Magor KE (2004)The dominant MHC class I gene is adjacent to the polymorphicTAP2 gene in the duck Anas platyrhynchos Immunogenetics56192ndash203 doi101007s00251-004-0672-3
Miller HC Lambert DM (2004) Gene duplication and geneconversion in class II MHC genes of New Zealand robins(Petroicidae) Immunogenetics 56178ndash191
Miller MM Goto RM Bernot A Zoorob R Auffray C Bumstead NBriles WE (1994) Two Mhc class I and two Mhc class II genes mapto the chicken Rfp-Y system outside the B complex Proc Natl AcadSci U S A 914397ndash4401 doi101073pnas91104397
460 Immunogenetics (2009) 61451ndash461
Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461

Bjorkman PJ Saper MA Samraoui B Bennett WS Strominger JLWiley DC (1987) Structure of human class I histocompatibil-ity antigen HLA-A2 Nature 329506ndash512 doi101038329506a0
Bonneaud C Sorci G Morin V Westerdahl H Zoorob R Wittzell H(2004) Diversity of MHC class I and IIB genes in house sparrows(Passer domesticus) Immunogenetics 55855ndash865 doi101007s00251-004-0648-3
Bonneaud C Chastel O Federici P Westerdahl H Sorci G (2006a)Complex MHC-based mate choice in a wild passerine Proc RSoc 2731111ndash1116 doi101098rspb20053325
Bonneaud C Peacuterez-Tris J Federici P Chastel O Sorci G (2006b) Majorhistocompatibility alleles associated with local resistance to malariain a passerine Evolution Int J Org Evolution 60383ndash389
Bryja J Galan M Charbonnel N Cosson J-F (2005) Analysis ofmajor histocompatibility complex class II gene in water volesusing capillary electrophoresis-single stranded conformationpolymorphism Mol Ecol Notes 5173ndash176 doi101111j1471-8286200400855x
Burri R Niculita-Hirzel H Roulin A Fumagalli L (2008) Isolation andcharacterization of major histocompatibility complex (MHC) class IIB genes in teh Barn owl (Aves Tyto alba) Immunogenetics 60543ndash550 doi101007s00251-008-0308-0
Cramp S Simmons KEL Perrins CM (eds) (1977-1994) The birds ofthe Western Palearctic Oxford University Press Oxford
Edwards SV Wakeland EK Potts WK (1995) Contrasting histories ofavian and mammalian MHC genes revealed by class II B genesof songbirds Proc Natl Acad Sci U S A 9212200ndash12204doi101073pnas922612200
Edwards SV Chesnut K Satta Y Wakeland EK (1997) Ancestralpolymorphism of Mhc Class II genes in mice implications forbalancing selection and the mammalian molecular clock Genetics146655ndash668
Fitzpatrick S (1994) Colourful migratory birds evidence for amechanism other than parasite resistance for the maintenance oflsquogood genesrsquo sexual selection Proc R Soc Lond B Biol Sci257155ndash166 doi101098rspb19940109
Freeman-Gallant CR Johnson EM Saponara F Stanger M (2002)Variation at the major histocompatibility complex in Savannahsparrows Mol Ecol 111125ndash1130 doi101046j1365-294X200201508x
Garrigan D Edwards SV (1999) Polymorphism across an exon-intronboundary in an avian Mhc class II B gene Mol Biol Evol 161599ndash1606
Gouumly de Bellocq J Charbonnel N Morand S (2008) Coevolutionaryrelationship between helminth diversity and Mhc class II polymor-phism in rodents J Evol Biol 211144ndash1150 doi101111j1420-9101200801538x
Hall TA (1999) BioEdit a user-friendly biological sequence alignmenteditor and analisis program for Windows 9598NT NucleicAcids Symp Ser 4195ndash98
Hansson B Richardson DS (2005) Genetic variation in twoendangered Acrocephalus species compared to a widespreadcongener estimates based on functional and random loci AnimConserv 883ndash90 doi101017S1367943004001878
Hess CM Edwards SV (2002) The evolution of major histocompat-ibility genes in birds Bioscience 52423ndash431 doi1016410006-3568(2002)052[0423TEOTMH]20CO2
Hess CM Gasper J Hoekstra H Hill C Edwards SV (2000) MHCclass II pseudogene and genomic signature of a 32-kb cosmid inthe House Finch (Carpodacus mexicanus) Genome Res 1013ndash23 doi101101gr105613
Hosomichi K Shiina T Suzuki S Tanaka M Shimizu S Iwamoto SHara H Yoshida Y Kulski JK Inoko H Hanzawa K (2006) Themajor histocompatibility complex (Mhc) class IIB region hasgreater genomic structural flexibility and diversity in the quail
than the chicken BMC Genomics 7322ndash335 doi1011861471-2164-7-322
Hughes AL Yeager M (1998) Natural selection at major histocom-patibility complex loci of vertebrates Annu Rev Genet 32415ndash435 doi101146annurevgenet321415
Hughes CR Miles S Walbroehl JM (2008) Support for the minimalessential MHC hypothesis a parrot with a single highlypolymorphic MHC class II B gene Immunogenetics 60219ndash231 doi101007s00251-008-0287-1
Indovina P Megiorni F Fontemaggi G Coni P Mora B Mazzilli MC(2001) Absence of in vivo DNA-Protein interactions in theDQA2 and DQB2 promoter regions Hum Immunol 62504ndash508doi101016S0198-8859(01)00236-1
Jarvi SI Goto RM Gee GF Briles WE Miller MM (1999)Identification inheritance and linkage of B-G-like and MHCclass I genes in cranes Am Genet Assoc 90152ndash159
Kaufman J (1999) Co-evolving genes in MHC haplotypes the lsquorulersquofor nonmammalian vertebrates Immunogenetics 50228ndash236doi101007s002510050597
Kaufman J Salamonsen J Flajnik M (1994) Evolutionary conserva-tion of MHC class I and class II molecules-different yet the sameSemin Immunol 6411ndash424 doi101006smim19941050
Kaufman J Voumllk H Wallny H (1995) A bdquominimal essential MHCldquoand an bdquounrecognized MHCldquo two extremes in selection forpolymorphism Immunol Rev 14363ndash88 doi101111j1600-065X1995tb00670x
Kaufman J Jacob J Shaw I Walker B Milne S Beck S SalomonsenJ (1999) Gene organisation determines evolution of function inthe chicken MHC Immunol Rev 167101ndash117 doi101111j1600-065X1999tb01385x
Klein J (1986) Natural history of the major histocompatibilitycomplex Wiley New York
Klein J Sato A Nagl S Oh Uigin C (1998) Molecular trans-speciespolymorphism Annu Rev Ecol Syst 291ndash21 doi101146annurevecolsys2911
Koch M Camp S Collen T Avila D Salomonsen J Wallny HJ vanHateren A Hunt L Jacob JP Johnston F Marston DA Shaw IDunbar PR Cerundolo V Jones EY Kaufman J (2007)Structures of an MHC class I molecule from B21 chickensillustrate promiscuous peptide binding Immunity 27885ndash899doi101016jimmuni200711007
Kosakovsky Pond SL Frost SDW (2005) Not so different after all acomparison of methods for detecting amino acid sites under selectionMol Biol Evol 221208ndash1222 doi101093molbevmsi105
Kosakovsky Pond SL Frost SDW Muse SV (2005) HyPhyhypothesis testing using phylogenies Bioinformatics 21676ndash679 doi101093bioinformaticsbti079
Kosakovsky Pond SL Posada D Gravenor MB Woelk CH FrostSDW (2006) Automated phylogenetic detection of recombinationusing a genetic algorithm Mol Biol Evol 231891ndash1901doi101093molbevmsl051
Loiseau C Zoorob R Garnier S Birard J Federici P Julliard R SorciG (2008) Antagonistic effects of a MHC class I allele on malaria-infected house sparrows Ecol Lett 11258ndash265 doi101111j1461-0248200701141x
Mesa CM Thulien KJ Moon DA Veniamin SM Magor KE (2004)The dominant MHC class I gene is adjacent to the polymorphicTAP2 gene in the duck Anas platyrhynchos Immunogenetics56192ndash203 doi101007s00251-004-0672-3
Miller HC Lambert DM (2004) Gene duplication and geneconversion in class II MHC genes of New Zealand robins(Petroicidae) Immunogenetics 56178ndash191
Miller MM Goto RM Bernot A Zoorob R Auffray C Bumstead NBriles WE (1994) Two Mhc class I and two Mhc class II genes mapto the chicken Rfp-Y system outside the B complex Proc Natl AcadSci U S A 914397ndash4401 doi101073pnas91104397
460 Immunogenetics (2009) 61451ndash461
Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461

Miller HC Belov K Daugherty CH (2006) MHC Class I genes in theTuatara (Sphenodon spp) evolution of the MHC in an ancientreptilian order Mol Biol Evol 23949ndash956 doi101093molbevmsj099
Peacuterez-Tris J Bensch S (2005) Dispersal increases local transmissionof avian malarial parasites Ecol Lett 8838ndash845 doi101111j1461-0248200500788x
Polaacutekovaacute R Vyskočilovaacute M Martin JF Mays HL Jr Hill GE Bryja JSchnitzer J Albrecht T (2007) A multiplex set of microsatellitemarkers for the Scarlet Rosefinch (Carpodacus erythrinus) MolEcol Notes 71375ndash1378 doi101111j1471-8286200701892x
Reusch TBH Schaschl H Wegner KM (2004) Recent duplication andinter-locus gene conversion in major histocompatibility class II-genes in a teleost the three-spined stickleback Immunogenetics56427ndash437 doi101007s00251-004-0704-z
Richardson DS Westerdahl H (2003) MHC diversity in twoAcrocephalus sepcies the outbred Great reed warbler and theinbred Seychelles warbler Mol Ecol 123523ndash3529 doi101046j1365-294X200302005x
Richardson DS Komdeur J Burke T von Schantz T (2004) MHC-based patterns of social and extra-pair mate choice in theSeychelles warbler Proc R Soc Lond B Biol Sci 272759ndash767doi101098rspb20043028
Shaw I Powell TJ Marston DA Baker K van Hateren A Riegert PWiles MV Milne S Beck S Kaufman J (2007) Differentevolutionary histories of the two classical class I genes BF1and BF2 illustrate drift and selection within the stable MHChaplotypes of chickens J Immunol 1785744ndash5752
Shiina T Shimizu S Hosomichi K Kohara S Watanabe S HanzawaK Beck S Kulski JK Inoko H (2004) Comparative genomicanalysis of two avian (Quail and Chicken) MHC regions JImmunol 1726751ndash6763
Shiina T Hosomichi K Hanzawa K (2006) Comparative genomics ofthe poultry major histocompatibility complex Anim Sci J77151ndash162 doi101111j1740-0929200600333x
Shum BP Rajalingam R Magor KE Azumi K Carr WH Dixon BStet RJ Adkison MA Hedrick RP Parham P (1999) A divergentnon-classical class I gene conserved in salmonids Immunoge-netics 49479ndash490 doi101007s002510050524
Spottiswoode C Moslashller AP (2004) Extra-pair paternity migration andbreeding synchrony in birds Behav Ecol 1541ndash57
Strand T Westerdahl H Houmlglund J Alatalo RV Siitari H (2007) The Mhcclass II of the Black grouse (Tetrao tetrix) consists of low numbersof B and Y genes with variable diversity and expressionImmunogenetics 59725ndash734 doi101007s00251-007-0234-6
van Oosterhout C Joyce DA Cummings SM (2006) Evolution of MHCclass IIB in the genome of wild and ornamental guppies Poeciliareticulata Heredity 97111ndash118 doi101038sjhdy6800843
Wallny H Avila D Hunt L Powell T Riegert P Salomonsen J SkjodtK Vainio O Vilbois F Wiles M Kaufman J (2006) Peptidemotifs of the single dominantly expressed class I moleculeexplain the striking MHC-determined response to Rous sarcomavirus in chicken Proc Natl Acad Sci U S A 1031434ndash1439doi101073pnas0507386103
Westerdahl H Wittzell H von Schantz T (1999) Polymorphism andtranscription of MHC class I genes in a passerine bird the greatreed warbler Immunogenetics 49158ndash170 doi101007s002510050477
Westerdahl H Witzel H von Schantz T (2000) MHC diversity intwo passerine birds no evidence for a minimal essentialMHC Immunogenetics 5292ndash100 doi101007s002510000256
Westerdahl H Wittzell H von Schantz T Bensch S (2004) MHC classI typing in a songbird with numerous loci and high polymor-phism using motif-specific PCR and DGGE Heredity 92534ndash542 doi101038sjhdy6800450
Westerdahl H Waldenstroumlm J Hansson B Hasselquist D von Schantz TBensch S (2005) Associations between malaria andMHC genes in amigratory songbird Proc R Soc Lond B Biol Sci 2721511ndash1518doi101098rspb20053113
Wilson DJ McVean G (2006) Estimating diversifying selection andfunctional constraint in the presence of recombination Genetics1721411ndash1425 doi101534genetics105044917
Wittzell H Bernot A Auffray C Zoorob R (1999a) Concertedevolution of two Mhc class II B loci in pheasants and domesticchicken Mol Biol Evol 16479ndash490
Wittzell H Madsen T Westerdahl H Shine R von Schantz T (1999b)MHC variation in birds and reptiles Genetica 104301ndash309doi101023A1026421607089
Worley K Gillingham M Jensen P Kennedy LJ Pizzari T KaufmanJ Richardson DS (2008) Single locus typing of MHC class I andclass II B loci in a population of red jungle fowl Immunogenet-ics 60233ndash247 doi101007s00251-008-0288-0
Xia C Hu T Yang T Wang L Xu G Lin C (2005) cDNA cloninggenomic structure and expression analysis of the goose (Ansercygnoides) MHC class I gene Vet Immunol Immunopathol107291ndash302 doi101016jvetimm200505005
Yang Z (1997) PAML a program package for phylogenetic analysisby maximum likelihood Comput Appl Biosci 13555ndash556
Yang Z Swanson WJ Vacquier VD (2000) Maximum-likelihoodanalysis of molecular adaptation in abalone sperm lysin revealsvariable selective pressures among lineages and sites Mol BiolEvol 171446ndash1455
Yang Z Wong WSW Nielsen R (2005) Bayes empirical Bayesinference of amino acid sites under positive selection Mol BiolEvol 221107ndash1118 doi101093molbevmsi097
Immunogenetics (2009) 61451ndash461 461




















