
Expression patterns of FHL/SLIM family members suggest important functional roles in skeletal muscle...
7
Gene expression pattern Expression patterns of FHL/SLIM family members suggest important functional roles in skeletal muscle and cardiovascular system Po-Hsien Chu a,b , Pilar Ruiz-Lozano a,b , Qiang Zhou a,b , Chenleng Cai a,b , Ju Chen a,b, * a UCSD-Salk Program in Molecular Medicine, University of California at San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0613, USA b Department of Medicine, University of California at San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0613, USA Received 4 February 2000; received in revised form 14 April 2000; accepted 14 April 2000 Abstract LIM domain containing proteins play critical roles in animal development and cellular differentiation. Here, we describe the cloning and expression patterns of three members of the four and a half LIM domain-only protein family, FHL1, 2, and 3, from mouse. A comparison of embryonic expression patterns of these three highly-related genes indicates that they are expressed in an overlapping pattern in the developing cardiovascular system, and skeletal muscle. In adult tissues, the three genes are expressed in a predominant and overlapping manner in cardiac and skeletal muscle. Of the three genes, FHL2 appears to have the most restricted expression pattern during development, in heart, blood vessels, and skeletal muscle. Expression in heart is highest in cardiac septa and in the region adjacent to the atrio-ventricular ring, suggesting a potential role in septation or conduction system development. In the heart, FHL1expression was observed strongly in developing outflow tract, and to a lesser extent in myocardium. FHL3 displays low and ubiquitous expression during mouse development. Cardiac ventricular expression of FHL1, but not FHL2 or FHL3, was upregulated in two mouse models of cardiac hypertrophic and dilated cardiomyopathy. Taken together, these data indicate the potential importance of this FHL family in the development and maintenance of the cardiovascular system and striated muscle, and suggest that FHL1 may play a role in the development of heart disease. q 2000 Elsevier Science Ireland Ltd. All rights reserved. Keywords: Four and half LIM domains; SLIM; MLP; Cardiovasculature; Skeletal muscle; Cardiac hypertrophy 1. Results and discussion LIM domains are protein-protein interaction domains that are receiving increasing attention from cellular and mole- cular biologists. Named after their initial discovery in two Caenorhabdits elegans gene products, lin-11 and mec-3, and the rat insulin enhancer binding protein, Islet-1 (Way and Chalfie, 1988; Freyd et al., 1990; Karlsson et al., 1990), LIM domains have since been identified in many proteins (Dawid et al., 1998). The LIM domain is a cysteine rich motif having the consensus sequence CX 2 CX 16– 23 HX 2 CX 2 CX 2 CX 16–21 CX 2 (C, H, D) that coordinately binds two zinc atoms and mediates protein-protein interactions (Arber and Caroni, 1996). Previous studies have revealed that some LIM domain containing proteins are nuclear proteins involved in cell lineage determination and pattern formation during development, whereas others encode cyto- solic proteins associated with the cytoskeleton, and have been found to play a role in adhesion plaque and actin microfilament organization (Zhou et al., 1980; Lumsden, 1995; Beckerle, 1997; Dawid et al., 1998). A newly identified group of LIM-only proteins with four and half LIM domains (FHL) (Chan et al., 1998; Lee et al., 1998) are enriched in striated muscle. This family consists of four members, three of which are expressed in adult striated muscle: FHL1/SLIM1, FHL2/SLIM3, and FHL3/ SLIM2 (SLIM is an acronym for skeletal muscle LIM, Morgan and Madgwick, 1996). The fourth member of this family, FHL4 (accession number: AA144955), may be expressed exclusively in testes, as inferred from electronic ‘Northern blot’ analyses, utilizing the full-length FHL4 cDNA sequence to search the EST data base. This analysis revealed that matching sequences were found only in ESTs from testis cDNA libraries. In confirmation of our search, a recent publication has also shown that FHL4 is expressed in testes (Morgan and Madgwick, 1999). An accumulating body of evidence indicates that FHL proteins may play important roles in both cardiac and striated muscle (Genini et al., 1997; Hwang et al., 1997; Takahashi et al., 1998). A search of the mouse EST database with the LIM domain of MLP revealed multiple EST clones correspond- Mechanisms of Development 95 (2000) 259–265 0925-4773/00/$ - see front matter q 2000 Elsevier Science Ireland Ltd. All rights reserved. PII: S0925-4773(00)00341-5 www.elsevier.com/locate/modo * Corresponding author. Tel.: 11-858-822-2452; fax: 11-858-534-2069. E-mail address: [email protected] (J. Chen).
Transcript of Expression patterns of FHL/SLIM family members suggest important functional roles in skeletal muscle...

Gene expression pattern
Expression patterns of FHL/SLIM family members suggest importantfunctional roles in skeletal muscle and cardiovascular system
Po-Hsien Chua,b, Pilar Ruiz-Lozanoa,b, Qiang Zhoua,b, Chenleng Caia,b, Ju Chena,b,*
aUCSD-Salk Program in Molecular Medicine, University of California at San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0613, USAbDepartment of Medicine, University of California at San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0613, USA
Received 4 February 2000; received in revised form 14 April 2000; accepted 14 April 2000
Abstract
LIM domain containing proteins play critical roles in animal development and cellular differentiation. Here, we describe the cloning and
expression patterns of three members of the four and a half LIM domain-only protein family, FHL1, 2, and 3, from mouse. A comparison of
embryonic expression patterns of these three highly-related genes indicates that they are expressed in an overlapping pattern in the
developing cardiovascular system, and skeletal muscle. In adult tissues, the three genes are expressed in a predominant and overlapping
manner in cardiac and skeletal muscle. Of the three genes, FHL2 appears to have the most restricted expression pattern during development,
in heart, blood vessels, and skeletal muscle. Expression in heart is highest in cardiac septa and in the region adjacent to the atrio-ventricular
ring, suggesting a potential role in septation or conduction system development. In the heart, FHL1expression was observed strongly in
developing out¯ow tract, and to a lesser extent in myocardium. FHL3 displays low and ubiquitous expression during mouse development.
Cardiac ventricular expression of FHL1, but not FHL2 or FHL3, was upregulated in two mouse models of cardiac hypertrophic and dilated
cardiomyopathy. Taken together, these data indicate the potential importance of this FHL family in the development and maintenance of the
cardiovascular system and striated muscle, and suggest that FHL1 may play a role in the development of heart disease. q 2000 Elsevier
Science Ireland Ltd. All rights reserved.
Keywords: Four and half LIM domains; SLIM; MLP; Cardiovasculature; Skeletal muscle; Cardiac hypertrophy
1. Results and discussion
LIM domains are protein-protein interaction domains that
are receiving increasing attention from cellular and mole-
cular biologists. Named after their initial discovery in two
Caenorhabdits elegans gene products, lin-11 and mec-3,
and the rat insulin enhancer binding protein, Islet-1 (Way
and Chal®e, 1988; Freyd et al., 1990; Karlsson et al., 1990),
LIM domains have since been identi®ed in many proteins
(Dawid et al., 1998). The LIM domain is a cysteine rich
motif having the consensus sequence CX2CX16±
23HX2CX2CX2CX16±21CX2(C, H, D) that coordinately binds
two zinc atoms and mediates protein-protein interactions
(Arber and Caroni, 1996). Previous studies have revealed
that some LIM domain containing proteins are nuclear
proteins involved in cell lineage determination and pattern
formation during development, whereas others encode cyto-
solic proteins associated with the cytoskeleton, and have
been found to play a role in adhesion plaque and actin
micro®lament organization (Zhou et al., 1980; Lumsden,
1995; Beckerle, 1997; Dawid et al., 1998).
A newly identi®ed group of LIM-only proteins with four
and half LIM domains (FHL) (Chan et al., 1998; Lee et al.,
1998) are enriched in striated muscle. This family consists
of four members, three of which are expressed in adult
striated muscle: FHL1/SLIM1, FHL2/SLIM3, and FHL3/
SLIM2 (SLIM is an acronym for skeletal muscle LIM,
Morgan and Madgwick, 1996). The fourth member of this
family, FHL4 (accession number: AA144955), may be
expressed exclusively in testes, as inferred from electronic
`Northern blot' analyses, utilizing the full-length FHL4
cDNA sequence to search the EST data base. This analysis
revealed that matching sequences were found only in ESTs
from testis cDNA libraries. In con®rmation of our search, a
recent publication has also shown that FHL4 is expressed in
testes (Morgan and Madgwick, 1999). An accumulating
body of evidence indicates that FHL proteins may play
important roles in both cardiac and striated muscle (Genini
et al., 1997; Hwang et al., 1997; Takahashi et al., 1998).
A search of the mouse EST database with the LIM
domain of MLP revealed multiple EST clones correspond-
Mechanisms of Development 95 (2000) 259±265
0925-4773/00/$ - see front matter q 2000 Elsevier Science Ireland Ltd. All rights reserved.
PII: S0925-4773(00)00341-5
www.elsevier.com/locate/modo
* Corresponding author. Tel.: 11-858-822-2452; fax: 11-858-534-2069.
E-mail address: [email protected] (J. Chen).

ing to cDNAs encoding FHL1, FHL2, and FHL3. EST
clones AA220340 (FHL1), AA023645 and AA419967
(FHL2) and AA016340 (FHL3) were purchased from
Genome Systems Inc. and fully sequenced. Alignment of
deduced amino acid sequences and their human orthologues
were performed utilizing the CLUSTAL program (http://
www.clustalw.genome.ad.jp) (Fig. 1).
All FHL family members contain four and a half LIM
domains and share high homology throughout their amino
acid sequences. Lengths of the LIM1, LIM2, and LIM3
domains (53, 53, 51 amino acids, respectively) are similar
among all FHL family members, as is the length of the half-
LIM domain (25 amino acids). However, the length of the
LIM4 domain varies among different FHL family members.
LIM4 consists of 56 amino acids in both human and mouse
FHL1, and 55 amino acids in human and mouse FHL2, and
human FHL3. Interestingly, when compared to human
FHL3, mouse FHL3 LIM4 has a 9 amino acid insertion
resulting in a 64 amino acid domain.
To ensure that the 9 amino acid insertion in mouse FHL3
was not a cloning artifact, we performed reverse-transcrip-
tion polymerase chain reaction (RT-PCR) analysis of adult
heart RNA utilizing two primers ¯anking the insertion. Gel
electrophoresis revealed one PCR product and sequencing
P.-H. Chu et al. / Mechanisms of Development 95 (2000) 259±265260
Fig. 1. Alignment of mouse and human FHL1, FHL2, and FHL3 amino acid sequences. Identical residues are indicated with an asterisk. Each LIM domain is
indicated in bold. The cDNA sequences of mouse FHL1, FHL2, and FHL3 were deposited in Genbank (accession numbers AF114380, AF114381, and
AF114382, respectively).
of the subcloned PCR product con®rmed the 9 amino acid
insertion. The functional consequences of this insertion
remain to be explored.
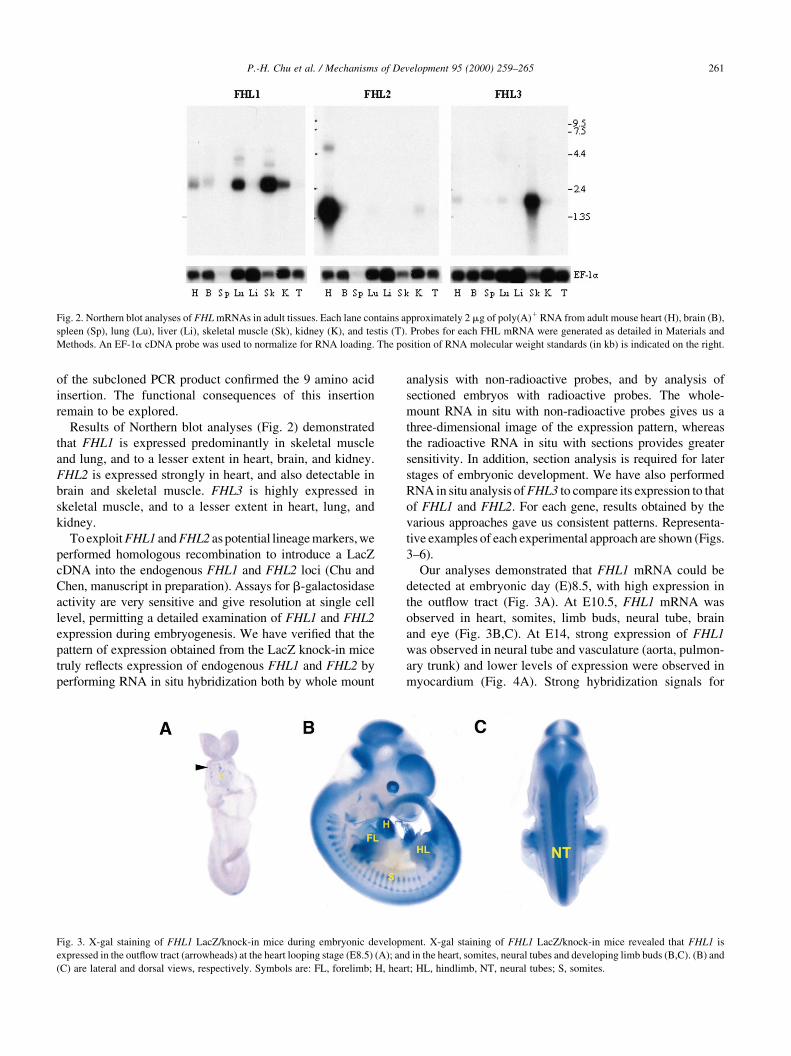
Results of Northern blot analyses (Fig. 2) demonstrated
that FHL1 is expressed predominantly in skeletal muscle
and lung, and to a lesser extent in heart, brain, and kidney.
FHL2 is expressed strongly in heart, and also detectable in
brain and skeletal muscle. FHL3 is highly expressed in
skeletal muscle, and to a lesser extent in heart, lung, and
kidney.
To exploit FHL1 and FHL2 as potential lineage markers, we
performed homologous recombination to introduce a LacZ
cDNA into the endogenous FHL1 and FHL2 loci (Chu and
Chen, manuscript in preparation). Assays for b-galactosidase
activity are very sensitive and give resolution at single cell
level, permitting a detailed examination of FHL1 and FHL2
expression during embryogenesis. We have veri®ed that the
pattern of expression obtained from the LacZ knock-in mice
truly re¯ects expression of endogenous FHL1 and FHL2 by
performing RNA in situ hybridization both by whole mount
analysis with non-radioactive probes, and by analysis of
sectioned embryos with radioactive probes. The whole-
mount RNA in situ with non-radioactive probes gives us a
three-dimensional image of the expression pattern, whereas
the radioactive RNA in situ with sections provides greater
sensitivity. In addition, section analysis is required for later
stages of embryonic development. We have also performed
RNA in situ analysis of FHL3 to compare its expression to that
of FHL1 and FHL2. For each gene, results obtained by the
various approaches gave us consistent patterns. Representa-
tive examples of each experimental approach are shown (Figs.
3±6).
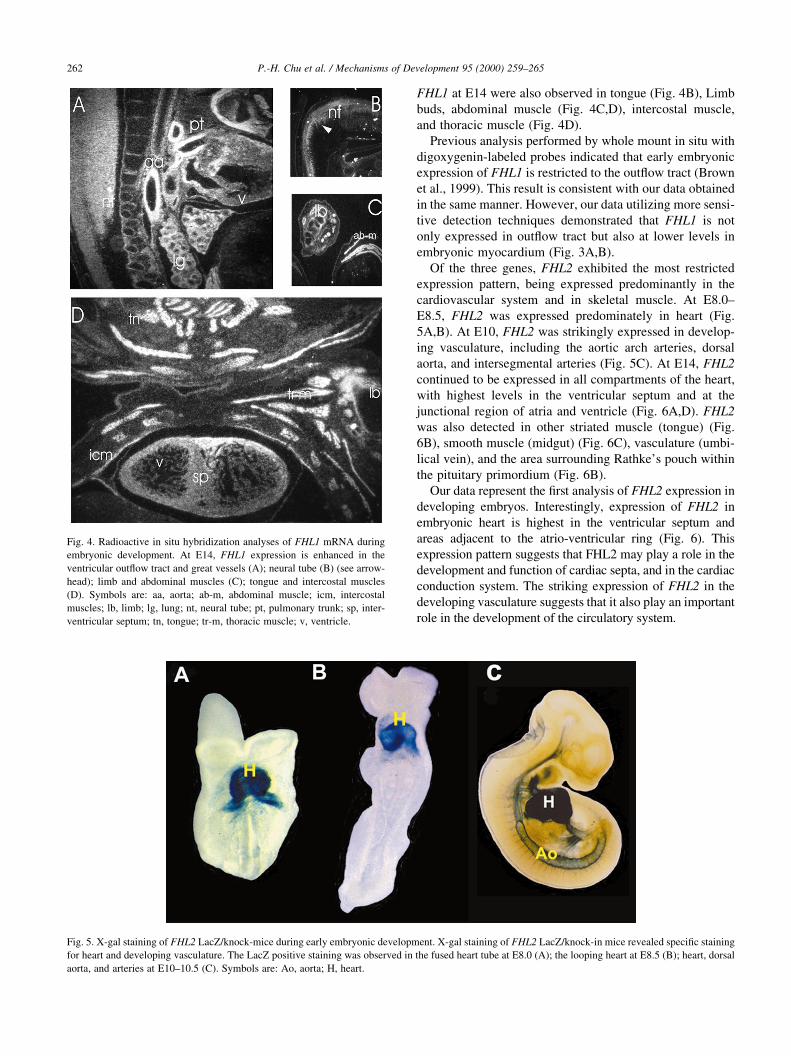
Our analyses demonstrated that FHL1 mRNA could be
detected at embryonic day (E)8.5, with high expression in
the out¯ow tract (Fig. 3A). At E10.5, FHL1 mRNA was
observed in heart, somites, limb buds, neural tube, brain
and eye (Fig. 3B,C). At E14, strong expression of FHL1
was observed in neural tube and vasculature (aorta, pulmon-
ary trunk) and lower levels of expression were observed in
myocardium (Fig. 4A). Strong hybridization signals for
P.-H. Chu et al. / Mechanisms of Development 95 (2000) 259±265 261
Fig. 2. Northern blot analyses of FHL mRNAs in adult tissues. Each lane contains approximately 2 mg of poly(A)1 RNA from adult mouse heart (H), brain (B),
spleen (Sp), lung (Lu), liver (Li), skeletal muscle (Sk), kidney (K), and testis (T). Probes for each FHL mRNA were generated as detailed in Materials and
Methods. An EF-1a cDNA probe was used to normalize for RNA loading. The position of RNA molecular weight standards (in kb) is indicated on the right.
Fig. 3. X-gal staining of FHL1 LacZ/knock-in mice during embryonic development. X-gal staining of FHL1 LacZ/knock-in mice revealed that FHL1 is
expressed in the out¯ow tract (arrowheads) at the heart looping stage (E8.5) (A); and in the heart, somites, neural tubes and developing limb buds (B,C). (B) and
(C) are lateral and dorsal views, respectively. Symbols are: FL, forelimb; H, heart; HL, hindlimb, NT, neural tubes; S, somites.
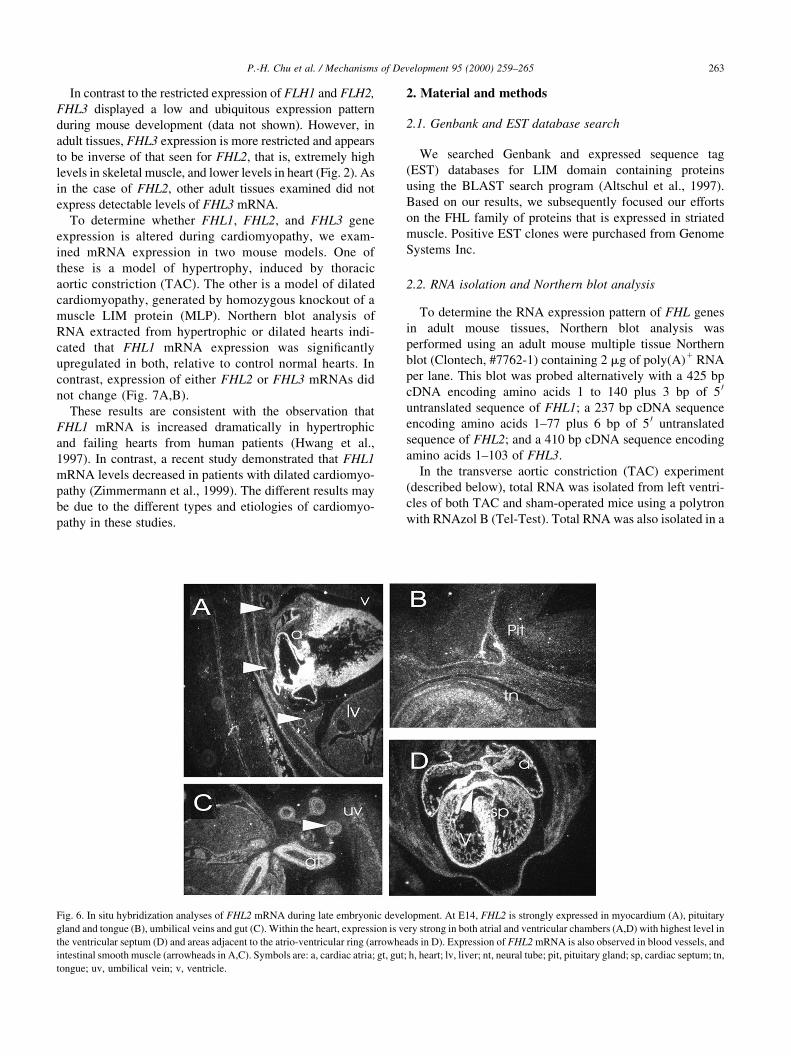
FHL1 at E14 were also observed in tongue (Fig. 4B), Limb
buds, abdominal muscle (Fig. 4C,D), intercostal muscle,
and thoracic muscle (Fig. 4D).
Previous analysis performed by whole mount in situ with
digoxygenin-labeled probes indicated that early embryonic
expression of FHL1 is restricted to the out¯ow tract (Brown
et al., 1999). This result is consistent with our data obtained
in the same manner. However, our data utilizing more sensi-
tive detection techniques demonstrated that FHL1 is not
only expressed in out¯ow tract but also at lower levels in
embryonic myocardium (Fig. 3A,B).
Of the three genes, FHL2 exhibited the most restricted
expression pattern, being expressed predominantly in the
cardiovascular system and in skeletal muscle. At E8.0±
E8.5, FHL2 was expressed predominately in heart (Fig.
5A,B). At E10, FHL2 was strikingly expressed in develop-
ing vasculature, including the aortic arch arteries, dorsal
aorta, and intersegmental arteries (Fig. 5C). At E14, FHL2
continued to be expressed in all compartments of the heart,
with highest levels in the ventricular septum and at the
junctional region of atria and ventricle (Fig. 6A,D). FHL2
was also detected in other striated muscle (tongue) (Fig.
6B), smooth muscle (midgut) (Fig. 6C), vasculature (umbi-
lical vein), and the area surrounding Rathke's pouch within
the pituitary primordium (Fig. 6B).
Our data represent the ®rst analysis of FHL2 expression in
developing embryos. Interestingly, expression of FHL2 in
embryonic heart is highest in the ventricular septum and
areas adjacent to the atrio-ventricular ring (Fig. 6). This
expression pattern suggests that FHL2 may play a role in the
development and function of cardiac septa, and in the cardiac
conduction system. The striking expression of FHL2 in the
developing vasculature suggests that it also play an important
role in the development of the circulatory system.
P.-H. Chu et al. / Mechanisms of Development 95 (2000) 259±265262
Fig. 4. Radioactive in situ hybridization analyses of FHL1 mRNA during
embryonic development. At E14, FHL1 expression is enhanced in the
ventricular out¯ow tract and great vessels (A); neural tube (B) (see arrow-
head); limb and abdominal muscles (C); tongue and intercostal muscles
(D). Symbols are: aa, aorta; ab-m, abdominal muscle; icm, intercostal
muscles; lb, limb; lg, lung; nt, neural tube; pt, pulmonary trunk; sp, inter-
ventricular septum; tn, tongue; tr-m, thoracic muscle; v, ventricle.
Fig. 5. X-gal staining of FHL2 LacZ/knock-mice during early embryonic development. X-gal staining of FHL2 LacZ/knock-in mice revealed speci®c staining
for heart and developing vasculature. The LacZ positive staining was observed in the fused heart tube at E8.0 (A); the looping heart at E8.5 (B); heart, dorsal
aorta, and arteries at E10±10.5 (C). Symbols are: Ao, aorta; H, heart.
In contrast to the restricted expression of FLH1 and FLH2,
FHL3 displayed a low and ubiquitous expression pattern
during mouse development (data not shown). However, in
adult tissues, FHL3 expression is more restricted and appears
to be inverse of that seen for FHL2, that is, extremely high
levels in skeletal muscle, and lower levels in heart (Fig. 2). As
in the case of FHL2, other adult tissues examined did not
express detectable levels of FHL3 mRNA.
To determine whether FHL1, FHL2, and FHL3 gene
expression is altered during cardiomyopathy, we exam-
ined mRNA expression in two mouse models. One of
these is a model of hypertrophy, induced by thoracic
aortic constriction (TAC). The other is a model of dilated
cardiomyopathy, generated by homozygous knockout of a
muscle LIM protein (MLP). Northern blot analysis of
RNA extracted from hypertrophic or dilated hearts indi-
cated that FHL1 mRNA expression was signi®cantly
upregulated in both, relative to control normal hearts. In
contrast, expression of either FHL2 or FHL3 mRNAs did
not change (Fig. 7A,B).
These results are consistent with the observation that
FHL1 mRNA is increased dramatically in hypertrophic
and failing hearts from human patients (Hwang et al.,
1997). In contrast, a recent study demonstrated that FHL1
mRNA levels decreased in patients with dilated cardiomyo-
pathy (Zimmermann et al., 1999). The different results may
be due to the different types and etiologies of cardiomyo-
pathy in these studies.
2. Material and methods
2.1. Genbank and EST database search
We searched Genbank and expressed sequence tag
(EST) databases for LIM domain containing proteins
using the BLAST search program (Altschul et al., 1997).
Based on our results, we subsequently focused our efforts
on the FHL family of proteins that is expressed in striated
muscle. Positive EST clones were purchased from Genome
Systems Inc.
2.2. RNA isolation and Northern blot analysis
To determine the RNA expression pattern of FHL genes
in adult mouse tissues, Northern blot analysis was
performed using an adult mouse multiple tissue Northern
blot (Clontech, #7762-1) containing 2 mg of poly(A)1 RNA
per lane. This blot was probed alternatively with a 425 bp
cDNA encoding amino acids 1 to 140 plus 3 bp of 5 0
untranslated sequence of FHL1; a 237 bp cDNA sequence
encoding amino acids 1±77 plus 6 bp of 5 0 untranslated
sequence of FHL2; and a 410 bp cDNA sequence encoding
amino acids 1±103 of FHL3.
In the transverse aortic constriction (TAC) experiment
(described below), total RNA was isolated from left ventri-
cles of both TAC and sham-operated mice using a polytron
with RNAzol B (Tel-Test). Total RNA was also isolated in a
P.-H. Chu et al. / Mechanisms of Development 95 (2000) 259±265 263
Fig. 6. In situ hybridization analyses of FHL2 mRNA during late embryonic development. At E14, FHL2 is strongly expressed in myocardium (A), pituitary
gland and tongue (B), umbilical veins and gut (C). Within the heart, expression is very strong in both atrial and ventricular chambers (A,D) with highest level in
the ventricular septum (D) and areas adjacent to the atrio-ventricular ring (arrowheads in D). Expression of FHL2 mRNA is also observed in blood vessels, and
intestinal smooth muscle (arrowheads in A,C). Symbols are: a, cardiac atria; gt, gut; h, heart; lv, liver; nt, neural tube; pit, pituitary gland; sp, cardiac septum; tn,
tongue; uv, umbilical vein; v, ventricle.

similar manner from ventricles of wild type and homozy-
gous null MLP mutant mice. Total RNA (10 mg) was elec-
trophoresed on a 1% agarose gel and transferred to a nylon
membrane, which was hybridized subsequently with
[a-32P]dATP-labeled FHL1, 2, 3, or GAPDH cDNA probes
utilizing QuickHyb solution (Stratagene) as described by
the company.
2.3. In situ hybridization
Radioactive in situ hybridization of mouse tissues was
performed on 7±10 mm paraf®n sections as described
(Ruiz-Lozano et al., 1997). Probes were designed as
follows: a 470 bp BamH I-Hind III fragment of FHL1
cDNA (encoding AA 1±140 plus 50 bp of 5 0 untranslated
region), a 490 bp EcoR I- Pst I fragment of FHL2 cDNA
(encoding AA 1±160 plus 10 bp of untranslated region), and
a 362 bp Pst I fragment of FHL3 cDNA (encoding AA 165±
285) were each subcloned into pBluescript II KS (Strata-
gene) to generate templates for sense and antisense 35S-
UTP- or digoxigenin-labeled probes. To evaluate expression
patterns of FHL mRNAs during mouse development, both
radioactive and non-radioactive RNA in situ hybridization
were performed. Non-radioactive in situ hybridization of
whole-mount embryos was performed using digoxigenin-
labeled synthetic oligo-DNA probes following standard
techniques (Faerman and Shani, 1997).
2.4. Whole-mount X-gal staining of FHL1 and FHL2
transgenic embryos
For whole-mount LacZ expression analysis, transgenic
embryos were ®xed and stained using standard methods
(Ross et al., 1996). The duration of staining was dependent
on color development and ranged from 4 to 12 h at room
temperature or 308C. Whole embryos or tissues were
analyzed and photographed on a Zeiss SV-6 dissecting
microscope with a Nikon C-mount 35mm camera.
2.5. Transverse aortic constriction (TAC)
Pressure overload hypertrophy was induced in mice by
using a microsurgical technique involving transverse aortic
constriction (TAC) as previously described (Rockman et al.,
1991). Sham-operated animals underwent the same opera-
tion with no aortic constriction. Transthoracic echocardio-
graphy and histological studies were utilized to ensure that
ventricular hypertrophic had occurred (data not shown).
Acknowledgements
We thank Dr Kenneth R. Chien for his enthusiastic
support throughout the project, and Drs Sylvia Evans,
Tara R. St Amand and Raymond Clark for critical reading
of the manuscript. The authors would like to thank Julie
Anderson and Wendy Bardwell for expert technical assis-
tance. Dr Chu was supported by a grant from the Chung
Gang Memorial Hospital at Taiwan.
References
Altschul, S.F., Madden, T.L., SchaÈffer, A.A., Zhang, J., Zhang, Z., Miller,
W., Lipman, D.J., 1997. Gapped BLAST and PSI-BLAST: a new
generation of protein database search programs. Nucl. Acids Res. 25,
3389±3402.
Arber, S., Caroni, P., 1996. Speci®city of single LIM motifs in targeting and
LIM/LIM interactions in situ. Genes Dev. 10, 289±300.
Beckerle, M.C., 1997. Zyxin: zinc ®ngers at sites of cell adhesion. Bioes-
says 19, 949±957.
Brown, S., McGrath, M.J., Ooms, L.M., Gurung, R., Maimone, M.M.,
Mitchell, C.A., 1999. Characterization of two isoforms of the skeletal
muscle LIM protein 1, SLIM1 ± localization of SLIM1 at focal adhesions
and the isoform slimmer in the nucleus of myoblasts and cytoplasm of
myotubes suggests distinct roles in the cytoskeleton and in nuclear-cyto-
plasmic communication. J. Biol. Chem. 274, 27083±27091.
Chan, K.K., Tsui, S.K., Lee, S.M., Luk, S.C., Liew, C.C., Fung, K.P.,
Waye, M.M., Lee, C.Y., 1998. Molecular cloning and characterization
of FHL2, a novel LIM domain protein preferentially expressed in
human heart. Gene 210, 345±350.
Dawid, I.B., Breen, J.J., Toyama, R., 1998. LIM domains: multiple roles as
adapters and functional modi®ers in protein interactions. Trends Genet.
14, 156±162.
Faerman, A., Shani, M., 1997. Transgenic mice: production and analysis of
expression. Methods Cell Biol. 52, 373±403.
Freyd, G., Kim, S.K., Horvitz, H.R., 1990. Novel cysteine-rich motif and
homeodomain in the product of the Caenorhabditis elegans cell lineage
gene lin-11. Nature 344, 876±879.
Genini, M., Schwalbe, P., Scholl, F.A., Remppis, A., Mattei, M.G., SchaÈfer,
P.-H. Chu et al. / Mechanisms of Development 95 (2000) 259±265264
Fig. 7. Northern blot analysis of FHL mRNAs in normal and cardiomyo-
pathic hearts. Probes for FHL1, FHL2, and FHL3 RNAs were generated as
described in Section 2. Approximately 10 mg total RNA were loaded to
each lane. Normalization for loading was performed by hybridization to a
probe for GAPDH. (A). RNA from 6-month old MLP knockout mice (lanes
3 and 4) and wildtype littermate controls (lanes 1 and 2). (B). RNA from
hypertrophic left ventricles of TAC mice (T) and sham-operated (S)
controls. Note greatly increased expression of FHL1 mRNA in both dilated
hearts from MLP mice, and hypertrophic hearts following TAC. In contrast,
expressions of FHL2 and FHL3 mRNAs exhibit no change.

B.W., 1997. Subtractive cloning and characterization of DRAL, a novel
LIM-domain protein down-regulated in rhabdomyosarcoma. DNA Cell
Biol. 16, 433±442.
Hwang, D.M., Dempsey, A.A., Wang, R.X., Rezvani, M., Barrans, J.D.,
Dai, K.S., Wang, H.Y., Ma, H., Cukerman, E., Liu, Y.Q., Gu, J.R.,
Zhang, J.H., Tsui, S.K., Waye, M.M., Fung, K.P., Lee, C.Y., Liew,
C.C., 1997. A genome-based resource for molecular cardiovascular
medicine: toward a compendium of cardiovascular genes. Circulation
96, 4146±4203.
Karlsson, O., Thor, S., Norberg, T., Ohlsson, H., Edlund, T., 1990. Insulin
gene enhancer binding protein Isl-1 is a member of a novel class of
proteins containing both a homeo- and a Cys-His domain. Nature 344,
879±882.
Lee, S.M., Tsui, S.K., Chan, K.K., Garcia-Barcelo, M., Waye, M.M., Fung,
K.P., Liew, C.C., Lee, C.Y., 1998. Chromosomal mapping, tissue distri-
bution and cDNA sequence of four-and-a-half LIM domain protein 1
(FHL1). Gene 216, 163±170.
Lumsden, A., 1995. Neural development. A `LIM code' for motor neurons?
Curr Biol. 5, 491±495.
Morgan, M.J., Madgwick, A.J., 1996. Slim de®nes a novel family of LIM-
proteins expressed in skeletal muscle. Biochem. Biophys. Res.
Commun. 225, 632±638.
Morgan, M.J., Madgwick, A.J., 1999. The fourth member of the FHL
family of LIM proteins is expressed exclusively in the testis. Biochem.
Biophys. Res. Commun. 255, 251±255.
Rockman, H.A., Ross, R.S., Harris, A.N., Knowlton, K.U., Steinhelper,
M.E., Field, L.J., Ross Jr., J., Chien, K.R., 1991. Segregation of
atrial-speci®c and inducible expression of an atrial natriuretic factor
transgene in an in vivo murine model of cardiac hypertrophy. Proc.
Natl. Acad. Sci. USA 88, 8277±8281.
Ross, R.S., Navankasattusas, S., Harvey, R.P., Chien, K.R., 1996. An HF-
1a/HF-1b/MEF-2 combinatorial element confers cardiac ventricular
speci®city and established an anterior-posterior gradient of expression.
Development 122, 1799±1809.
Ruiz-Lozano, P., Doevendans, P., Brown, A., Gruber, P.J., Chien, K.R.,
1997. Developmental expression of the murine spliceosome-associated
protein mSAP49. Dev. Dyn. 208, 482±490.
Takahashi, H., Koshimizu, U., Nakamura, T., 1998. A novel transcript
encoding truncated LIM kinase 2 is speci®cally expressed in male
germ cells undergoing meiosis. Biochem. Biophys. Res. Commun.
249, 138±145.
Way, J.C., Chal®e, M., 1988. mec-3, a homeobox-containing gene that
speci®es differentiation of the touch receptor neurons in C. elegans.
Cell 54, 5±16.
Zhou, Q., Ruiz-Lozano, P., Martone, M.E., Chen, J., 1999. Cypher, a
striated muscle-restricted PDZ and LIM domain-containing protein,
binds to a-actin-2 and protein kinase C. J.Biol. Chem. 274, 7±
19813.
Zimmermann, R., Kostin, S., Kotaka, M., Waye, M.M., Tsui, S.K., Lee,
S.M., Chen, K.K., Lee, C.Y., Fung, K.P., Klsverkorn, W.P., Schaper, J.,
1999. A possible role for novel LIM domain proteins in dilated cardi-
omyopathy. Circulation 100 (Suppl. 1), 565.
P.-H. Chu et al. / Mechanisms of Development 95 (2000) 259±265 265




















