Exploring the backbone dynamics of native spider silk proteins in Black Widow silk glands with...
7
Exploring the backbone dynamics of native spider silk proteins in Black Widow silk glands with solution-state NMR spectroscopy Dian Xu, Jeffery L. Yarger * , Gregory P. Holland * Department of Chemistry and Biochemistry, Magnetic Resonance Research Center, Arizona State University, Tempe, AZ 85287-1604, United States article info Article history: Received 14 March 2014 Received in revised form 21 May 2014 Accepted 8 June 2014 Available online 13 June 2014 Keywords: Spider silk protein Intrinsically disordered proteins NMR abstract Spider dragline silk is an outstanding biopolymer with a strength that exceeds steel by weight and a toughness greater than high-performance fibers like Kevlar. For this reason, understanding how a spider converts the gel-like, aqueous protein spinning dope within the major ampullate (MA) gland into a super fiber is of great importance for developing future biomaterials based on spider silk. In this work, the initial state of the silk proteins within Black Widow MA glands was probed with solution-state NMR spectroscopy. 15 N relaxation parameters, T 1 , T 2 and 15 N-{ 1 H} steady-state NOE were measured for twelve backbone environments at two spectrometer frequencies, 500 and 800 MHz. The NMR relaxation pa- rameters extracted for all twelve environments are consistent with MA silk protein backbone dynamics on the fast sub-nanosecond timescale. Therefore, it is concluded that the repetitive core of spider MA proteins are in an unfolded, highly flexible state in the MA gland. © 2014 Elsevier Ltd. All rights reserved. 1. Introduction Spiders have envolved over three hundred millions years the ability to spin up to six different silks from their spinnerets that have a diverse range of mechanical properties [1e3]. Major ampullate (MA) or dragline spider silk is a proteinaceous fiber with a combination of high tensile strength and extensibility that gives it a toughness that exceeds Kevlar [1e5]. Dragline spider silk is one of the strongest fibers found in nature and serves as the spider's safety line and is used in web construction [1,3]. Understanding the pri- mary and secondary structure of the silk proteins comprising this insoluble super fiber has been intensively studied over the past few decades [6e23]. The two proteins, major ampullate spidroin 1 and 2 (MaSp1 and MaSp2), are the primary components of spider dragline silk [6,7]. The proteins are synthesized in the tail region of the spider's MA glands and then stored within the sac-like region at high concen- tration (~25e50% w/v) [3]. The only full length primary amino acid sequence for MaSp1 and MaSp2 has been reported for the Black Widow (Latrodectus hesperus) spider [17]. The molecular weights of the MaSp1 and MaSp2 proteins are 250 and 312 kDa, respectively. Each of the two proteins has a long repetitive core region flanked by two highly conserved nonrepetitive terminal domains. The sec- ondary structure of the repetitive core of the dragline silk proteins has been extensively studied in the solid fiber [8e16,18e23]. Crystalline b-sheet conformations for the alanine-rich domains, poly(Ala) and poly(GlyeAla), were identified by solid-state NMR [8,16,19e22] and X-ray diffraction [10e14,18]. In addition, solid- state NMR experiments indicated 3 1 -helical structures for the Gly-Gly-X repeats [9,15,18,21] and elastin-like type II b-turns for the Gly-Pro-Gly-X-X motifs [23] in spider dragline silk. Despite the increasing information about the structure of solid dragline silk fibers, producing synthetic fibers with comparable mechnical properties to native spider silk still eludes researchers [24e26]. Spider silk research has been shifting towards understanding the proteins within the MA glands prior to fiber formation and the biochemical processes responsible for spider silk assembly [27e34]. Due to the difficulties associated with extracting and purifying native spider silk proteins, there have only been a few NMR studies to structurally characterize the spider silk proteins in the MA gland (spinning dope) [27e31]. In previous studies, NMR has been applied to characterize silk proteins within intact MA glands from three spider species, Nephila clavipes, Nephila edulis and Argiope aurantia. The silk proteins within the MA glands are in a hydrogel- like state and solution-state [27], solid-state magic angle spinning (MAS) [28] and high resolution (HR)-MAS [31] NMR spectroscopy Abbreviations: HSQC, heteronuclear single quantum correlation; INEPT, insen- sitive nuclei enhanced by polarization transfer; NOE, nuclear overhauser effect; HNCACB, amide proton to nitrogen to a/b carbon correlation; CP-MAS, cross po- larization magic angle spinning; TPPM, two pulse phase modulation. * Corresponding authors. E-mail addresses: [email protected], [email protected] (G.P. Holland). Contents lists available at ScienceDirect Polymer journal homepage: www.elsevier.com/locate/polymer http://dx.doi.org/10.1016/j.polymer.2014.06.018 0032-3861/© 2014 Elsevier Ltd. All rights reserved. Polymer 55 (2014) 3879e3885
Transcript of Exploring the backbone dynamics of native spider silk proteins in Black Widow silk glands with...

lable at ScienceDirect
Polymer 55 (2014) 3879e3885
Contents lists avai
Polymer
journal homepage: www.elsevier .com/locate/polymer
Exploring the backbone dynamics of native spider silk proteins inBlack Widow silk glands with solution-state NMR spectroscopy
Dian Xu, Jeffery L. Yarger*, Gregory P. Holland*
Department of Chemistry and Biochemistry, Magnetic Resonance Research Center, Arizona State University, Tempe, AZ 85287-1604, United States
a r t i c l e i n f o
Article history:Received 14 March 2014Received in revised form21 May 2014Accepted 8 June 2014Available online 13 June 2014
Keywords:Spider silk proteinIntrinsically disordered proteinsNMR
Abbreviations: HSQC, heteronuclear single quantusitive nuclei enhanced by polarization transfer; NOHNCACB, amide proton to nitrogen to a/b carbon colarization magic angle spinning; TPPM, two pulse ph* Corresponding authors.
E-mail addresses: [email protected], gpholla@
http://dx.doi.org/10.1016/j.polymer.2014.06.0180032-3861/© 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
Spider dragline silk is an outstanding biopolymer with a strength that exceeds steel by weight and atoughness greater than high-performance fibers like Kevlar. For this reason, understanding how a spiderconverts the gel-like, aqueous protein spinning dope within the major ampullate (MA) gland into a superfiber is of great importance for developing future biomaterials based on spider silk. In this work, theinitial state of the silk proteins within Black Widow MA glands was probed with solution-state NMRspectroscopy. 15N relaxation parameters, T1, T2 and 15N-{1H} steady-state NOE were measured for twelvebackbone environments at two spectrometer frequencies, 500 and 800 MHz. The NMR relaxation pa-rameters extracted for all twelve environments are consistent with MA silk protein backbone dynamicson the fast sub-nanosecond timescale. Therefore, it is concluded that the repetitive core of spider MAproteins are in an unfolded, highly flexible state in the MA gland.
© 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Spiders have envolved over three hundred millions years theability to spin up to six different silks from their spinnerets thathave a diverse range of mechanical properties [1e3]. Majorampullate (MA) or dragline spider silk is a proteinaceous fiber witha combination of high tensile strength and extensibility that gives ita toughness that exceeds Kevlar [1e5]. Dragline spider silk is one ofthe strongest fibers found in nature and serves as the spider's safetyline and is used in web construction [1,3]. Understanding the pri-mary and secondary structure of the silk proteins comprising thisinsoluble super fiber has been intensively studied over the past fewdecades [6e23].
The two proteins, major ampullate spidroin 1 and 2 (MaSp1 andMaSp2), are the primary components of spider dragline silk [6,7].The proteins are synthesized in the tail region of the spider's MAglands and then stored within the sac-like region at high concen-tration (~25e50% w/v) [3]. The only full length primary amino acidsequence for MaSp1 and MaSp2 has been reported for the BlackWidow (Latrodectus hesperus) spider [17]. The molecular weights of
m correlation; INEPT, insen-E, nuclear overhauser effect;rrelation; CP-MAS, cross po-ase modulation.
gmail.com (G.P. Holland).
the MaSp1 and MaSp2 proteins are 250 and 312 kDa, respectively.Each of the two proteins has a long repetitive core region flanked bytwo highly conserved nonrepetitive terminal domains. The sec-ondary structure of the repetitive core of the dragline silk proteinshas been extensively studied in the solid fiber [8e16,18e23].Crystalline b-sheet conformations for the alanine-rich domains,poly(Ala) and poly(GlyeAla), were identified by solid-state NMR[8,16,19e22] and X-ray diffraction [10e14,18]. In addition, solid-state NMR experiments indicated 31-helical structures for theGly-Gly-X repeats [9,15,18,21] and elastin-like type II b-turns for theGly-Pro-Gly-X-X motifs [23] in spider dragline silk. Despite theincreasing information about the structure of solid dragline silkfibers, producing synthetic fibers with comparable mechnicalproperties to native spider silk still eludes researchers [24e26].Spider silk research has been shifting towards understanding theproteins within the MA glands prior to fiber formation and thebiochemical processes responsible for spider silk assembly[27e34].
Due to the difficulties associated with extracting and purifyingnative spider silk proteins, there have only been a few NMR studiesto structurally characterize the spider silk proteins in the MA gland(spinning dope) [27e31]. In previous studies, NMR has beenapplied to characterize silk proteins within intact MA glands fromthree spider species, Nephila clavipes, Nephila edulis and Argiopeaurantia. The silk proteins within the MA glands are in a hydrogel-like state and solution-state [27], solid-state magic angle spinning(MAS) [28] and high resolution (HR)-MAS [31] NMR spectroscopy

D. Xu et al. / Polymer 55 (2014) 3879e38853880
have been applied to collect isotropic chemical shifts to analyzespider silk protein conformation. The results from conformation-dependent 1H and 13C isotropic chemical shifts indicate that therepetitive core of the silk proteins are primarily in a random coilconformation [27,30,31]. However, it should be noted that the silkproteins may be present in the metastable “molten-globule” stateand the possibility of partial backbone folding where random coilchemical shifts are still observed can not be excluded [27,31,35].Recent Raman spectroscopy studies [29,30] indicate possible pol-yproline II (PPII) and a-helical fragments in addition to random coilstructures for native spider silk proteins.
In order to determine if the MA silk proteins are truly randomcoil or molten-globule, futher studies on the native spider silkprotein dynamics within the MA glands is required. In this work,solution-state NMR spectroscopy was used to characterize thebackbone structure and dynamics of Black Widow silk proteinswithin intact MA glands. Residue-specific 1H, 13C, and 15N isotropicchemical shift measurements were used to characterize proteinconformations on 13C/15N-labeled MA silk proteins. The backbonedynamics of the silk proteins were extracted from 15N NMR relax-ation parameters T1, T2 and 15N-{1H} steady-state NOE measured attwo spectrometer frequencies (500 and 800 MHz) for 15N-labeledsilk proteins. The measured subnanosecond backbone dynamicsobserved for the spider silk proteins within the MA gland provideconvincing evidence that the repetitive core of the silk proteins arehighly flexibile and completely unfolded in agreement with theobserved random coil chemical shifts.
2. Experimental
2.1. Sample preparation
Mature female BlackWidow (Latrodectus hesperus) spiders werefed a 15% (w/v) aqueous solution of 15N-L-alanine for 15N NMRrelaxation measurements or uniformly labeled U-[13C,15N]-L-alanine for HNCACB experiments and solid-state NMR experimentsevery other day while they were forcibly silked at a speed of 2 cm/s[36]. Spiders were typically dissected following 2 weeks of isotopeenrichment. For solution-state NMR experiments, 5 spiders weredissected under a microscope, and 10 intact MA glands werecarefully transferred from the spider's abdomen into 90:10H2O:D2O (v/v) where both gland ducts and tails were thenremoved. The remaining sac-like MA glands were placed into a5 mm Shigemi solution-state NMR tube filled with a solution of90:10 H2O:D2O (v/v), and 5% (w/v) sodium azide at neutral pH(Supporting Information, Fig. S1). For each set of 15N NMR relaxa-tion measurements at one frequency or a single HNCACB experi-ment, a set of gland samples was prepared with the methoddescribed above. Sample conditions were shown to remain stableafter 24 h of sample preparation (Supporting Information, Fig. S2).For solid-state NMR experiments, 2 spiders were dissected in thesame method described above and 4 intact MA glands werecollected and carefully transferred into a Bruker Kel-F 4 mm rotorinsert filled with a solution of 90:10 H2O:D2O (v/v), 5% (w/v) so-dium azide at neutral pH or pH 4.0. Isotopes were purchased fromCambridge Isotope Laboratories, Inc., sodium azide was from Sigmaand all chemicals were used as received.
2.2. NMR experiments
Solution-state NMR data was collected on a Varian VNMRS 500or 800 MHz spectrometer equipped with a HCN triple resonanceprobe with triple-axis gradients at 298 K. All pulse sequences werefrom the Varian Biopack library. 2D 1H-15N HSQC [37,38] spectrawere collected at 800 MHz with 32 scans per t1 point, 50 � 2560
complex points for t1 � t2 dimensions with a sweep width of35 � 16 ppm, and a 2.7 ms 1H-15N INEPT transfer time (1/4JNH).Gradient enhanced triple resonance 3D HNCACB [39e41] experi-ments were acquired at 500 MHz with 1024 � 58 � 28 complexpoints for t3(NH)� t1(13Ca/13Cb)� t2(15N) dimensionswith a sweepwidth of 16 � 80 � 35 ppm. Longitudinal relaxation times (T1),transverse relaxation times (T2), and steady-state 15N-{1H} NOEwere measured with sequences described by Kay et al. [42] andFarrow et al. [43] where pulse field gradients are utilized toenhance sensitivity and select coherence transfer pathway with 1H180� pulse inserted to remove the cross-relaxation effect in T1 andT2 measurements and water flip-back pulses applied to minimizethe effect of water magnetization on fast exchanging amide protonsin NOEmeasurements [44]. Waltz-16 15N decoupling with a 2.7 kHzradio frequency field was applied during acquisitions for all NMRexperiments. The 1H-15N HSQC spectra for measuring relaxationdata were acquired with 50 � 2560 complex points for t1 � t2 di-mensions, 2 s recycle delay and 8 scans per t1 point for T1, T2measurements and 3 s recycle delay and 32 scans per t1 point forNOE measurements. To measure T1 and T2 times, a set of 10 1H-15NHSQC spectra were recorded with delay periods of 0, 50, 100, 300,400, 500, 600, 750,1000,1400ms and 10, 50,110,150, 210, 290, 350,410, 450, 490 ms, respectively. In the NOE experiment, a pair of1H-15N HSQC spectra was collected with and without a 3 s protonsaturation pulse prior to the experiments. All relaxation measure-ments were repeated twice on two different sets of samples withaverage values reported. To minimize systematic errors, all relax-ation measurements were recorded in an interleaved fashion.
Solution-state spectra were processed in NMRPipe [45]. Forprocessing 1H-15N HSQC spectra, a cosine-bell apodization wasapplied to the t2 dimension and a combination of cosine bell and20 Hz exponential line broadening was used for the t1 dimensionwith zero filling to 512 points. For the HNCACB spectrum, cosine-bellapodization functions and zero filling to 1024 � 64 � 32 fort3� t1 � t2 dimensions were used. Processed datawas then analyzedin NMRViewj [46] and peak intensities were used to determinerelaxation times. T1 and T2 parameters were determined by fittingthe peak heights from each spectrumwith a three-parameter single-exponential-decay function with the uncertainty estimated through500 timesMonte Carlo simulations. NOE valueswere calculated fromthe ratios of peak heights from spectra with and without protonsaturation. Uncertainties for NOE values were estimated from theroot-mean-square baseline noise of the spectra.
Additional sets of solution- and solid-state NMR data were ac-quired at 298 K on a Bruker Avance III 400 MHz spectrometerequipped with a 5 mm solution-state broad band observe (BBO)probe optimized for X-detection and a 4 mm Bruker HX solid-stateCP-MAS probe, respectively. For the solution-state NMR experi-ments, a 1D 15N-detected refocused 1H-15N INEPT experiment [47]was collected and compared to a 15N one pulse experiment withgated proton decoupling. Experimental parameters for the solutionexperiments were a sweep width of 200 ppm, 8400 scans, 5 srecycle delay, 0.252 s acquisition time and 2.7 ms refocused INEPTtransfer delays with ~2 kHz waltz-16 1H decoupling during acqui-sition. Spectra were further processed with 5 Hz line broadening.Experimental parameters for solid-state NMR experiments were asweep width of 375 ppm, 34 ms acquisition time with ~2 kHzwaltz-16 1H decoupling, 1024 scans, 5 s recycle delay and 2.7 msrefocused INEPT transfer delays. 1H-15N CP-MAS experiments wereacquired with a sweep width of 50 kHz, 10.2 ms acquisition timewith ~55 kHz TPPM-15 1H decoupling, a 5 s recycle delay, 1 mscontact time, 6400 scans for the neutral pH sample, and 1024 scansfor the sample at pH 4.0 sample. All solid-state spectra were pro-cessed using Bruker Topspin 3.2 with 10 Hz exponential linebroadening applied prior to Fourier transformation.

Fig. 1. 2D 1H-15N HSQC spectrum of (A) 15N-labeled Black Widow spider MA glands in90:10 H2O:D2O, pH 7.0 collected at 298 K, 800 MHz; (B) strip plots of the 3D tripleresonance HNCACB spectrum of U-[13C, 15N]-L-alanine labeled Black Widow spider MAglands in 90:10 H2O:D2O, pH 7.0 collected at 298 K, 500 MHz. The 12 resolved back-bone cross peaks (A) were labeled with one-letter amino acid codes (bold) next to thesymbols of their previous residues. Assignment is based on the (B) HNCACB experi-ment. (B) Selected 15N planes with unique 1H-13C correlations were displayed withantiphased 13Ca (black), 13Cb (red) cross peaks from the corresponding amino acidswith relative higher intensities and their previous amino acids with lower intensities.The (C) consensus amino acid sequence for Black Widow MA silk proteins. (Forinterpretation of the references to color in this figure legend, the reader is referred tothe web version of this article.)
D. Xu et al. / Polymer 55 (2014) 3879e3885 3881
3. Results
In the spider silk spinning process, four factors are believed to beresponsible for silk protein folding including salt concentrationchanges [32,48], a decrease in pH [32e34,48], dehydration [48,49]and mechanical shearing [50]. Thus, when preparing a sample ofsilk proteins withinMA glands for solution-state NMR experiments,the intact MA glands were dissected from the spider's abdomenwith the intact membrane containing the proteins in their nativestate and particular care was taken to not shear or allow the sampleto dehydrate. The solvent used was neutral-pH aqueous solutionwith 5% (w/v) sodium azide added to prevent bacteria growth andextend the life time of the protein sample. If the silk proteinsaggregated and precipitated from the solution phase, an obvioussignal loss in the solution-state NMR spectrumwill be observed. Toconfirm that the silk proteins within the MA glands maintain anear-native state with no protein aggregation within a 24 h timeperiod, 1H projections from 2D 1H-15N HSQC experiments werecollected on a freshly prepared sample (within 30 min) and 24 hlater (Supporting Information, Fig. S2). Essentially, no signal loss orchange in lineshape was observed. However, it should be noted,that the sample does change after multiple days to a week(Supporting Information, Fig. S2) and therefore, all NMR mea-surements were accomplished within a 24 h period for a givensample.
The structure and dynamics of silk proteins within the MAglands of Black Widow spiders was probed with solution-stateNMR spectroscopy. A 2D 1H-15N HSQC spectrum (see Fig. 1A) wasacquired and 12 backbone amide resonances were resolved. Toassign these backbone resonances, a triple resonance 3D HNCACBexperiment was utilized. Fig. 1B shows 15N planes selected for thecross peaks of interest in the 2D 1H-15N HSQC spectrum, and givesdistinguishable 13Ca resonances (black contour) with antiphased13Cb (red contour) for each residue. In addition, the spectrum alsodisplays relatively weaker contours for the previous residue 13Caand/or 13Cb resonance(s). Therefore, an unambiguous backboneresonance assignment can be made for the repetitive amino acidpairs prevalent in the core of the MaSp1 and MaSp2 spider silkproteins.
In Fig. 1A three major residues, alanine, glycine and glutamineare observed for the Black Widow silk proteins within the MAglands. Each amino acid has multiple chemical environmentsincluding 8 Gly, 2 Gln and 4 Ala resonances. Slight differences inchemical shifts for a given amino acid are attributed to theirsequential positions. The backbone resonance assignment providesresidue sequential patterns such as Ala-Ala, Gly-Ala, Ala-Gly, Gly-Gly or Gly-Gln, which agrees with the poly(Ala), poly(Gly-Ala) andGly-Gly-Gln motifs of the repetitive core region of the MaSp1 andMaSp2 spider silk proteins primary amino acid sequence (seeFig. 1C) [17]. From our previous results [51], where Nephila clavipesspiders were fed a 13C-labeled alanine solution for solid-state NMRexperiments on MA silk fibers, 13C enrichment in spider silk pro-teins was achieved for four amino acids, Ala, Gly, Gln and Ser. Thesefour amino acids constitute greater than 85% of the repetitive corefor Black Widow MA silk proteins [52]. Compared to the presentassignment, 15N enrichment in spider silk proteins appears to besimilar to 13C where we observe high 15N enrichment for Ala, Gly,and Gln. The reason Ser is not included is because of its low per-centage (1.1%) in the amino acid composition of Black Widowdragline silk proteins [52]. Although Ser appears to be labeled with15N, its low percentage results in a low S/N ratio for Ser resonancesin 2 and 3D NMR experiments.
The amide proton region in Fig. 1A displays a narrow chemicalshift dispersion (~0.6 ppm) characteristic of an unstructured pro-tein [53,54]. An additional H/D exchange experimentwas applied at
neutral pH and shows that the amide protons were rapidlyexchanged by deuterium within 5 min (Supporting Information,Fig. S3). This also indicates that the silk proteins within the MAglands are unstructured in the repetitive core region. The timescalefor amide proton/deuterium exchange is typically hours to days for

D. Xu et al. / Polymer 55 (2014) 3879e38853882
folded proteins exhibiting defined secondary and tertiary structure[55,56].
In order to further characterize the protein structure within MAglands, isotropic chemical shift analysis was utilized. 13Ca,b, amideproton and 15N chemical shifts of all 12 backbone environmentswere measured from the HNCACB spectrum and 2D 1H-15N HSQCspectrum, respectively. A 2D 1H-13C HSQC spectrum (SupportingInformation, Fig. S4) was also collected to provide unambiguous1Ha,b assignment and allow the extraction of 1Ha,b chemical shiftsfor the Black Widow spider silk proteins within the MA glands. Allmeasured chemical shifts are listed in Table 1 along with theaverage chemical shifts observed for random coil, a-helical and b-sheet structures [57,58]. All 1Ha,b and 13Ca,b chemical shifts of silkproteins within the MA glands closely match random coil chemicalshifts. This indicates the repetitive core region of the silk proteins isunstructured with no evidence for secondary structure such as a-helical, b-sheet, or b-turn in the MA gland prior to silk fiber for-mation. This agrees with previous 1H, 13C chemical shift data onnative silk proteins within the MA glands from Nephila clavipes,Nephila edulis and Argiope aurantia [27,28,31]. The amide 1Hchemical shifts for Ala, Gln and 15N chemical shifts for Gln showreasonable agreement with their corresponding random coilchemical shifts. However, discrepancies such as the one for the Ala15N chemical shifts were also observed. This can be explained bythe lower reliability in predicting protein secondary structuresfrom amide 1H and 15N chemical shifts as the chemical shifts forboth nuclei were found to be inconsistent among different modelpeptides with random coil structures [58]. The latter discrepancymainly originates from the amide 1H and 15N chemical shifts beingsensitive to not only conformation but, other influences such ashydrogen bonding. When utilizing the more reliable 13Ca,b and 1Hachemical shifts that are not impacted to the same extent by otherinfluences, the repetitive silk protein core appears to be present in arandom coil conformation.
To probe the dynamics of spider silk proteins within the MAglands, the relaxation parameters T1, T2 and NOE of the backboneamide 15N nuclei were measured. Fig. 2 displays the 15N relaxationdata recorded at two spectrometer frequencies (500 and 800 MHz).The corresponding average errors for T1 are estimated to be 6.6%and 6.5%, and the estimated average errors for T2 are 2.8% and 2.3%.T1 has the average values of 360 ± 24 ms and 390 ± 29 ms at 500and 800MHz, respectively, while T2 hasmean values of 203 ± 16ms
Table 11Ha, 1Hb,13Ca,b, 1HN and 15N isotropic chemical shiftsa (ppm) of Black Widow spider silkHNCACB (Fig. 1) spectra. Averaged 1H, 13C and 15N chemical shifts for random coil (coil), a-[57,58].
Gland Coil Helix Strand
1Ha
Ala 4.23,4.30 4.25 4.03 4.87Gln 4.35 4.34 4.03 4.97Gly 3.95 3.95 3.84 4.09
13CaGA 52.93, 52.95 52.67 54.86 50.86AA 52.99, 53.03GQ 56.04, 56.09 55.94 58.61 54.33GG 45.41 e 45.50 45.34 47.02 45.08QG 45.50AG 45.37, 45.41
NHGA 8.10, 8.27 8.11 7.99 8.59AA 8.10, 8.25GQ 8.26, 8.35 8.25 8.11 8.51GG 8.19 e 8.40 8.34 8.23 8.27QG 8.51AG 8.24, 8.44
a All chemical shifts are in ppm, referenced to DSS.
at 500 MHz and 171 ± 12 ms at 800 MHz. For all 15N-labeled resi-dues, T1 and T2 remain relatively uniform with consistent trendsbetween the two fields when the error is considered. This indicatesthe core region of the silk proteins in the MA glands exhibit similardynamics. Large negative NOE values (�0.557 ± 0.196) at 500 MHzare detected for all residues indicating that the double quantumrelaxation pathway is more favored than zero quantum relaxation.This is typically observed for highly flexible, unfolded peptidechains or terminal regions of folded proteins where motions ofunrestricted NeH bond vectors are primarily in the high frequencyregime [54,59e61]. Combined with positive NOE values(0.338 ± 0.082) at 800 MHz, the frequencies of gland proteinbackbone motions are constrained within a range of~500e800 MHz (sub-nanosecond timescale). Additionally, the 15Nrelaxation data was analyzed with the reduced spectral densitymapping approach [62,63] (supporting information, Fig. S6 and S7).The spectral density mapping analysis was consistent with sub-nanosecond backbone dynamics for the silk proteins.
4. Discussion
The applicability of conventional solution-state NMR spectros-copy for characterizing the structure and dynamics of spider silkproteins with molecular weights in the ~250e300 kDa range needsto be considered. For a typical folded proteinwith similar molecularweight, the transverse relaxation times could be on the order of theINEPT transfer delays in the HSQC pulse sequence due to proteinrigidity or conformational exchange processes on the ms-ms time-scale and could render some of the NMR signals “invisible” byconventional, INEPT-based solution NMR. However, the spectradisplayed in the present work have relatively narrow linewidths forall three nuclei and the possibility may exist that solution-stateNMR is only sampling a fraction of the silk proteins from a subsetof flexible regions and not the whole protein. To address that theentire proteins are detected by solution-state NMR spectroscopy, aseparate set of solution- and solid-state NMR experiments wereconducted.
A 15N-L-alanine labeled Black Widow spider MA gland samplewas freshly prepared at neutral pH and 15N NMR data was collectedon a BBO solution NMR probe optimized for X-detection with a 15Ndirect (one pulse) experiment with gated proton decoupling and a1H-15N refocused INEPT pulse sequence. The results of this
proteins within the MA gland from solution-state NMR 1H-13C HSQC (Fig. S4) andhelix (helix) and b-strand (Strand) frommodel proteins are also listed for comparison
Gland Coil Helix Strand
1Hb
1.38 1.39 e e
1.97, 2.14 1.99, 2.12 e e
e e e e13Cb18.77, 19.36 19.03 18.27 21.7219.11, 19.1229.50, 29.51 28.67 28.33 31.92e e e e
e
e15N123.72, 123.99 132.52 121.65 125.57122.62, 122.75119.68, 119.72 119.73 118.59 123.14108.49 e 110.68 109.94 107.34 110.19109.85107.62, 108.08

Fig. 2. Bar graph of backbone amide 15N relaxation parameters at 298K, 500 MHz(black) and 800 MHz (red) for different residues labeled with one-letter amino acidcodes. The amino acid residue is indicated in bold along with the preceding aminoacid. (A) longitudinal relaxation times T1. (B) transverse relaxation times T2. (C) steady-state 15N-{1H} NOE values for different residues. (For interpretation of the references tocolor in this figure legend, the reader is referred to the web version of this article.)
D. Xu et al. / Polymer 55 (2014) 3879e3885 3883
experiment are presented in Fig. 3A. When comparing the two 15Nspectra collected with the same number of scans and a long recycledelay for full relaxation, it is determined that the refocused INEPTspectrum is 9-fold more intense compared to the direct spectrumand the spectra are nearly superimposable for all amide environ-ments. The experimentally observed INEPT enhancement is veryclose to the predicted theoretical signal enhancement of 10 (gH/gN)for 1H-15N INEPT. NMR signals with long (�50 ms) to intermediate(5e20 ms) 15N transverse relaxation timescale should becompletely detectable in the direct spectrum and the near
theoretical 9-fold signal enhancement in the INEPT experimentindicates that there is no appreciable signal lost due to interme-diate 15N transverse relaxation on the timescale of the INEPTtransfers. Likewise, if a considerable fraction of the 1H transverserelaxation times were on the intermediate timescale (1e5 ms) asmaller signal enhancement would be expected for the INEPTspectrum in contrast with the observations in Fig. 3A. In addition,relaxation dispersion experiments [64] conducted with a fast andslow Carr-Purcell Meiboom-Gill (CPMG) frequency confirmed thatthere was no appreciable contribution from conformational ex-change on the msems timescale (Supporting information, Fig. S5).
A second series of data was collected with a HX solid-state CP-MAS probe to determine if there was any appreciable signal fromsilk protein components with short (<1 ms) 1H and 15N transverserelaxation times. A static 1D 15N INEPT spectrum (Fig. 3B, red) wascollected serving as a direct comparison to the 15N dimension of the1H-15NHSQC solution-state NMR spectrum (Fig.1). Then 5 kHzMASwas applied to the sample followed by the acquisition of another15N INEPT spectrum (Fig. 3, green) with identical parameters to thestatic 15N INEPT experiment. Comparing the two INEPT spectra,only a slight resolution enhancement from MAS is observed withno obvious signal increase. Another 15N-L-alanine labeled spidergland sample was prepared and treated with pH 4.0 buffer. 12 hlater, strong 1H-15N CP signal was observed in the 1H-15N CP MASspectrum (Fig. 3, black). This indicates protein aggregation wastriggered by the low pH that is readily probed by the appearance ofCP signal via the 1H-15N dipolar interactions. In contrast, in the pHneutral sample, no 1H-15N CP signal was detected from an identicalCP-MAS experiment collected with six times more scans (Fig. 3,blue). The combination of X-detected solution and solid-state NMRdata presented in Fig. 3 indicates that there is no appreciable rigid/aggregated (short transverse relaxation) or solution-state NMRunobservable regions (intermediate transverse relaxation) for theisotopically labeled regions. It is concluded that solution-state NMRis capable of detecting�90% of the 15N-labeled silk protein residuesfor the repetitive core at neutral pH.
In addition to the large molecular weights, the spider silk pro-teins also exhibit highly repetitive primary amino acid sequences.This makes a full sequential assignment of the spider silk proteinswithin the MA glands by solution-state NMR spectroscopyincredibly challenging. However, the combination of a 2D 1H-15NHSQC and a 3D triple resonance HNCACB experiment permits apartial sequential assignment for the spider silk proteins. Specif-ically, this combination of NMR experiments permits the structuraland dynamic characterization of amino acid couplets in the primaryamino acid sequence including Ala-Ala, Aly-Gly, Gly-Gln, Gly-Glyand Gly-Ala. Isotropic chemical shift analysis confirmed that allthese backbone environments in the core region of the spider silkproteins within the MA glands are unstructured as previously re-ported [27,28,31]. As the spider silk proteins are rapidly assembledinto a solid fiber rich in secondary structure, there is a possibilitythat the spider silk proteins are experiencing an intermediate statein the MA gland environment where the proteins are partiallyfolded or in the “molten-globule state” [35]. A simple isotropicchemical shift analysis where random coil chemical shift areobserved does not necessarily rule out this possibility [35]. How-ever, when considering the fast sub-nanosecond dynamics of thespider silk proteins extracted from the 15N NMR relaxation data thesilk protein backbone within the core region is highly flexible.Therefore, no appreciable intermediate states of the spider silkproteins is present within the MA glands and the proteins can becategorized as intrinsically disordered proteins that are truerandom coils and not present as an intermediate molten globulestate [65]. It should be noted that the current isotopic labelingscheme does not provide any information regarding the terminal
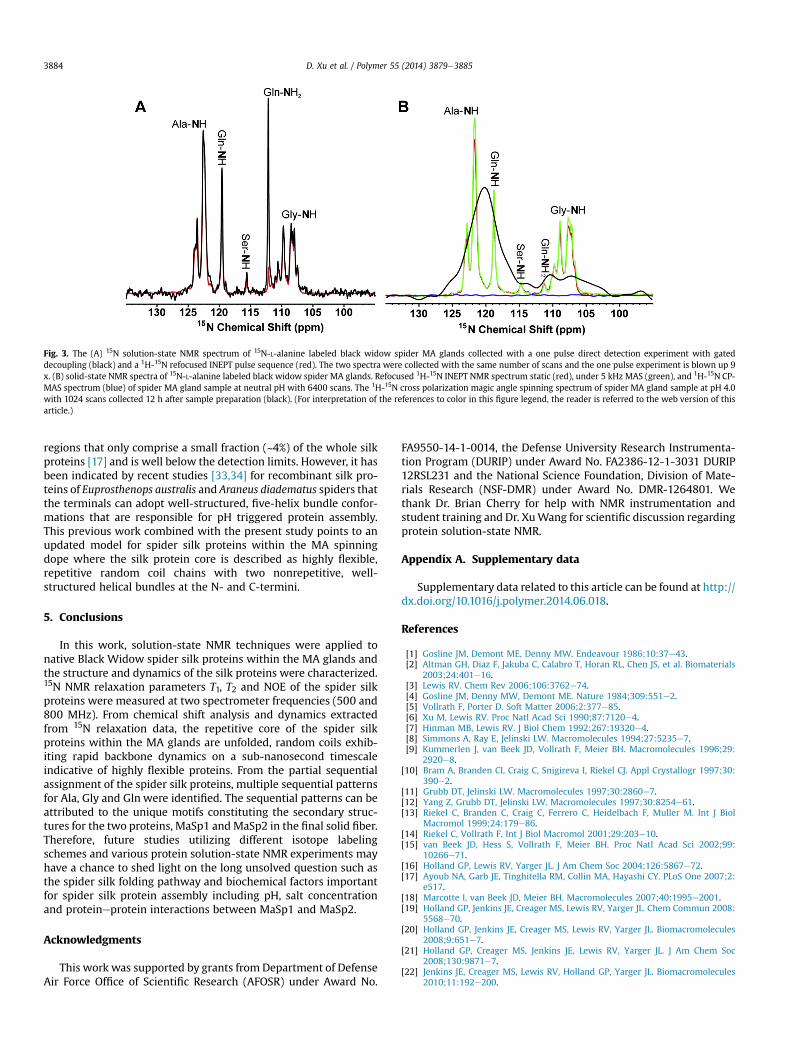
Fig. 3. The (A) 15N solution-state NMR spectrum of 15N-L-alanine labeled black widow spider MA glands collected with a one pulse direct detection experiment with gateddecoupling (black) and a 1H-15N refocused INEPT pulse sequence (red). The two spectra were collected with the same number of scans and the one pulse experiment is blown up 9x. (B) solid-state NMR spectra of 15N-L-alanine labeled black widow spider MA glands. Refocused 1H-15N INEPT NMR spectrum static (red), under 5 kHz MAS (green), and 1H-15N CP-MAS spectrum (blue) of spider MA gland sample at neutral pH with 6400 scans. The 1H-15N cross polarization magic angle spinning spectrum of spider MA gland sample at pH 4.0with 1024 scans collected 12 h after sample preparation (black). (For interpretation of the references to color in this figure legend, the reader is referred to the web version of thisarticle.)
D. Xu et al. / Polymer 55 (2014) 3879e38853884
regions that only comprise a small fraction (~4%) of the whole silkproteins [17] and is well below the detection limits. However, it hasbeen indicated by recent studies [33,34] for recombinant silk pro-teins of Euprosthenops australis and Araneus diadematus spiders thatthe terminals can adopt well-structured, five-helix bundle confor-mations that are responsible for pH triggered protein assembly.This previous work combined with the present study points to anupdated model for spider silk proteins within the MA spinningdope where the silk protein core is described as highly flexible,repetitive random coil chains with two nonrepetitive, well-structured helical bundles at the N- and C-termini.
5. Conclusions
In this work, solution-state NMR techniques were applied tonative Black Widow spider silk proteins within the MA glands andthe structure and dynamics of the silk proteins were characterized.15N NMR relaxation parameters T1, T2 and NOE of the spider silkproteins were measured at two spectrometer frequencies (500 and800 MHz). From chemical shift analysis and dynamics extractedfrom 15N relaxation data, the repetitive core of the spider silkproteins within the MA glands are unfolded, random coils exhib-iting rapid backbone dynamics on a sub-nanosecond timescaleindicative of highly flexible proteins. From the partial sequentialassignment of the spider silk proteins, multiple sequential patternsfor Ala, Gly and Gln were identified. The sequential patterns can beattributed to the unique motifs constituting the secondary struc-tures for the two proteins, MaSp1 andMaSp2 in the final solid fiber.Therefore, future studies utilizing different isotope labelingschemes and various protein solution-state NMR experiments mayhave a chance to shed light on the long unsolved question such asthe spider silk folding pathway and biochemical factors importantfor spider silk protein assembly including pH, salt concentrationand proteineprotein interactions between MaSp1 and MaSp2.
Acknowledgments
This work was supported by grants from Department of DefenseAir Force Office of Scientific Research (AFOSR) under Award No.
FA9550-14-1-0014, the Defense University Research Instrumenta-tion Program (DURIP) under Award No. FA2386-12-1-3031 DURIP12RSL231 and the National Science Foundation, Division of Mate-rials Research (NSF-DMR) under Award No. DMR-1264801. Wethank Dr. Brian Cherry for help with NMR instrumentation andstudent training and Dr. XuWang for scientific discussion regardingprotein solution-state NMR.
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.polymer.2014.06.018.
References
[1] Gosline JM, Demont ME, Denny MW. Endeavour 1986;10:37e43.[2] Altman GH, Diaz F, Jakuba C, Calabro T, Horan RL, Chen JS, et al. Biomaterials
2003;24:401e16.[3] Lewis RV. Chem Rev 2006;106:3762e74.[4] Gosline JM, Denny MW, Demont ME. Nature 1984;309:551e2.[5] Vollrath F, Porter D. Soft Matter 2006;2:377e85.[6] Xu M, Lewis RV. Proc Natl Acad Sci 1990;87:7120e4.[7] Hinman MB, Lewis RV. J Biol Chem 1992;267:19320e4.[8] Simmons A, Ray E, Jelinski LW. Macromolecules 1994;27:5235e7.[9] Kummerlen J, van Beek JD, Vollrath F, Meier BH. Macromolecules 1996;29:
2920e8.[10] Bram A, Branden CI, Craig C, Snigireva I, Riekel CJ. Appl Crystallogr 1997;30:
390e2.[11] Grubb DT, Jelinski LW. Macromolecules 1997;30:2860e7.[12] Yang Z, Grubb DT, Jelinski LW. Macromolecules 1997;30:8254e61.[13] Riekel C, Branden C, Craig C, Ferrero C, Heidelbach F, Muller M. Int J Biol
Macromol 1999;24:179e86.[14] Riekel C, Vollrath F. Int J Biol Macromol 2001;29:203e10.[15] van Beek JD, Hess S, Vollrath F, Meier BH. Proc Natl Acad Sci 2002;99:
10266e71.[16] Holland GP, Lewis RV, Yarger JL. J Am Chem Soc 2004;126:5867e72.[17] Ayoub NA, Garb JE, Tinghitella RM, Collin MA, Hayashi CY. PLoS One 2007;2:
e517.[18] Marcotte I, van Beek JD, Meier BH. Macromolecules 2007;40:1995e2001.[19] Holland GP, Jenkins JE, Creager MS, Lewis RV, Yarger JL. Chem Commun 2008:
5568e70.[20] Holland GP, Jenkins JE, Creager MS, Lewis RV, Yarger JL. Biomacromolecules
2008;9:651e7.[21] Holland GP, Creager MS, Jenkins JE, Lewis RV, Yarger JL. J Am Chem Soc
2008;130:9871e7.[22] Jenkins JE, Creager MS, Lewis RV, Holland GP, Yarger JL. Biomacromolecules
2010;11:192e200.

D. Xu et al. / Polymer 55 (2014) 3879e3885 3885
[23] Jenkins JE, Creager MS, Butler EB, Lewis RV, Yarger JL, Holland GP. ChemCommun 2010;46:6714e6.
[24] Lazaris A, Arcidiacono S, Huang Y, Zhou JF, Duguay F, Chretien N, et al. Science2002;295:472e6.
[25] Brooks AE, Stricker SM, Joshi SB, Kamerzell TJ, Middaugh CR, Lewis RV. Bio-macromolecules 2008;9:1506e10.
[26] Zhang YS, Hu JH, Miao YG, Zhao AC, Zhao TF, Wu DY, et al. Mol Biol Rep2008;35:329e35.
[27] Hijirida DH, Do KG, Michal C, Wong S, Zax D, Jelinski LW. Biophys J 1996;71:3442e7.
[28] Hronska M, van Beek JD, Williamson PTF, Vollrath F, Meier BH. Bio-macromolecules 2004;5:834e9.
[29] Lefevre T, Leclerc J, Rioux-Dube JF, Buffeteau T, Paquin MC, Rousseau ME, et al.Biomacromolecules 2007;8:2342e4.
[30] Lefevre T, Boudreault S, Cloutier C, Pezolet M. Biomacromolecules 2008;9:2399e407.
[31] Jenkins JE, Holland GP, Yarger JL. Soft Matter 2012;8:1947e54.[32] Dicko C, Vollrath F, Kenney JM. Biomacromolecules 2004;5:704e10.[33] Hagn F, Eisoldt L, Hardy JG, Vendrely C, Coles M, Scheibel T, et al. Nature
2010;465:239e42.[34] Askarieh G, Hedhammar M, Nordling K, Saenz A, Casals C, Rising A, et al.
Nature 2010;465:236e8.[35] Ohgushi M, Wada A. FEBS Lett 1983;164:21e4.[36] Work RW, Emerson PD. J Arachnol 1982;10:1e10.[37] Bodenhausen G, Ruben DJ. Chem Phys Lett 1980;69:185e9.[38] Kay LE, Keifer P, Saarinen T. J Am Chem Soc 1992;114:10663e5.[39] Wittekind M, Mueller L. J Magn Reson Ser B 1993;101:201e5.[40] Kay LE, Xu GY, Yamazaki T. J Magn Reson Ser A 1994;109:129e33.[41] Muhandiram DR, Kay LE. J Magn Reson Ser B 1994;103:203e16.[42] Kay LE, Nicholson LK, Delaglio F, Bax A, Torchia DA. J Magn Reson 1992;97:
359e75.
[43] Farrow NA, Muhandiram R, Singer AU, Pascal SM, Kay CM, Gish G, et al.Biochemistry 1994;33:5984e6003.
[44] Grzesiek S, Bax A. J Am Chem Soc 1993;115:12593e4.[45] Delaglio F, Grzesiek S, Vuister GW, Zhu G, Pfeifer J, Bax A. J Biomol NMR
1995;6:277e93.[46] Johnson BA, Blevins RA. J Biomol NMR 1994;4:603e14.[47] Burum DP, Ernst RR. J Magn Reson 1980;39:163e8.[48] Knight DP, Vollrath F. Naturwissenschaften 2001;88:179e82.[49] Tillinghast EK, Chase SF, Townley MA. J Insect Physiol 1984;30:591e6.[50] Iizuka E. Experientia 1983;39:449e54.[51] Shi XY, Yarger JL, Holland GP. Anal Bioanal Chem 2013;405:3997e4008.[52] Creager MS, Jenkins JE, Thagard-Yeaman LA, Brooks AE, Jones JA, Lewis RV,
et al. Biomacromolecules 2010;11:2039e43.[53] Frank MK, Clore GM, Gronenborn AM. Protein Sci 1995;4:2605e15.[54] Bai YW, Chung J, Dyson HJ, Wright PE. Protein Sci 2001;10:1056e66.[55] Englander SW, Kallenbach NR. Q Rev Biophys 1983;16:521e655.[56] Tadeo X, Casta~no D, Millet O. Protein Sci 2007;16:2733e40.[57] Wishart DS, Bigam CG, Holm A, Hodges RS, Sykes BD. J Biomol NMR 1995;5:
67e81.[58] Wang YJ, Jardetzky O. Protein Sci 2002;11:852e61.[59] Kay LE, Torchia DA, Bax A. Biochemistry 1989;28:8972e9.[60] Clore GM, Driscoll PC, Wingfield PT, Gronenborn AM. Biochemistry 1990;29:
7387e401.[61] Eliezer D, Yao J, Dyson HJ, Wright PE. Nat Struct Biol 1998;5:148e55.[62] Farrow NA, Zhang OW, Szabo A, Torchia DA, Kay LE. J Biomol NMR 1995;6:
153e62.[63] Peng JW, Wagner G. Biochemistry 1995;34:16733e52.[64] Loria JP, Rance M, Palmer AG. J Am Chem Soc 1999;121:2331e2.[65] Dunker AK, Lawson JD, Brown CJ, Williams RM, Romero P, Oh JS, et al. J Mol
Graph Model 2001;19:26e59.