
Access to Medical Oxygen Virtual Roundtable - Stop Pneumonia
Upload
independentCategory
view
2download
0
Journal of Medical Virology 2:4S-59 (1978)
Experimental Respiratory Syncytial Virus Pneumonia in Cebus Monkeys Linda S. Richardson, Robert B. Belshe, D. Lewis Sly, William T. London, David A. Prevar, Ena Camargo, and Robert M. Chanock National Institute o f Allergy and Infectious Diseases (Linda Richardson, Robert Belshe, David Prevar, Ena Camargo, and Robert Chanock) and National Institute o f Neurological and Communicative Disorders and Stroke (William London), National Institutes of Health, Bethesda, Maryland; and Meloy Laboratories (Lewis Sly), Rockville, Maryland.
Into 14 juvenile cebus monkeys that lacked serum antibodies for RS virus LO8 plaque-forming units (pfu) of wild-type respiratory syncytial (RS) virus were inoculated transtracheally. Roentgenographic evidence of pneumonia developed in 13 of 14 infected animals. Gross pathologic changes occurred in each of the 13 monkeys that were sacrificed. Patchy areas of red consolida- tion were seen in the lower lobes 24 hours after inoculation, and there was progression t o gray consolidation seven days later. Each of the infected animals had histologic evidence of interstitial pneumonia. Changes were de- tected in thc lung as early as 24 hours after inoculation; they consisted pri- marily of infiltration of the alveolar wall. By the fourth t o sixth day after inoculation there was marked interstitial thickening, pulmonary consolida- tion, formation of multinucleated gianl cells and development of eosinophilic cytoplasmic inclusion bodies within alveolar cells. RS viral antigens, detected by indirect immunofluorcscence, were distributed throughout cells of the alveolar wall and the bronchiolar epithelium. The virus grew t o highest titer in the lungs on the fourth to sixth day after inoculation: up to 10' pfu/gram of tissue were detected. The cebus monkey represents the first experimental host to develop extensive pulmonary lesions during infection with respiratory syncytial virus.
Key words: respiratory syncytial virus, viral pneumonia, cebus monkeys
INTRODUCTION
Until now a suitable experimental animal for the study of RS virus disease has not been available. A number of nvnprimates can be infected in the laboratory, bu t none of these species develop extensive pulmonary pathology [Coates and Chanock, 1962; Dreizin et al, 197 1 ;Prince and Porter, 19761 . In an effort t o identify a species in which t o study the pathogenesis of RS virus disease, we had previously administered lo3
Received for publication September 30, 1977.
Address reprint requests to Dr. Linda S. Richardson, National Institutcs of Health, Bldg 7, Rm 301, Bethesda, MD 20014.
to
0146-661 S/78/0201-004SS02.90 0 1978 Alan R . Liss. Inc

46 Richardson et a1
lo3 .7 plaque-forming units (pfu) of wild-type virus intranasally to RS virus antibody-free chimpanzees, cebus monkeys, rhesus monkeys, and squirrel monkeys [Belshe et al, 19771 . Each species of primates became infected but only the chimpanzees became ill. Each of four infected chimpanzees developed an illness that was limited to the upper respiratory tract and that resembled the “common cold.” Viral replication was most extensive in the chimpanzee (10’ -lo6 pfu/ml of nasopharyngeal swab fluid), while the lower primates shed less virus (102-103.8 pfu/ml).
serious lower respiratory tract disease. Therefore,,we sought a more readily available laboratory animal for further study of RS virus disease. Cebus monkeys had previously been found to be partially permissive for growth of RS virus [Belshe et al, 19771 , and it was considered possible that a larger dose of virus than used in the initial study might overconie host resistance and cause disease in the lungs. We therefore extended our studies of RS virus infection in cebus monkeys by inoculating a large quantity of RS virus (10’ pfu) directly into the lower respiratory tract. In this report, we describe the produc- tion of extensive pulmonary disease in cebus monkeys infected in this manner.
The scarcity of chimpauees precluded more extensive studies designed to produce
MATERIALS AND METHODS
Monkeys
Cebus monkeys (Cebus apella and Cebus albifrons) that lacked RS virus-neutralidng antibodies as detected by the plaque reduction technique [Mills et al, 197 11 were selected from those housed at Meloy Laboratories, Inc., Rockville, Maryland. The monkeys were maintained in facilities accredited by the American Association for the Accreditation of Laboratory Animal Care (AAALAC). Laboratory-raised juveniles that ranged in age from 6 to 20 months and weighed 900-2,150 gm were housed in pairs in air-flow-controlled isolation cages [London et al, 19721 . Their diet consisted of 24% crude protein biscuits supplemented with apples, bread, and vitamin D3 once a week in the water.
Virus
The history of the A2 strain of RS virus has been previously described [Mills et al, 19711 . The virus had undergone many passages in HEp-2 cells. The two suspensions used to inoculate cebus monkeys contained virus that had undergone 2 or 3 additional passages in HeLa spinner culture (Flow Laboratories, Rockville, Maryland). We sedimented lo8 cells that had been infected in spinner culture a t 1,500 rpm for 10 minutes in the cold. The resulting cell pellet (containing about 90% of the total amount of infectious virus) was resuspended in 10 ml of growth medium (representing about a 30-fold concentration of virus), quick-frozen, and stored at -70°C.
Cell Cultures
HEp-2 cells were obtained from Flow Laboratories and maintained as previously described [Gharpure et al, 19691 . HeLa spinner cultures were prepared from cells ob- tained from Flow Laboratories that had been grown in Eagle’s minimum essential medium (Flow Laboratories) supplemented with 5% agamma calf serum, 0.002 M gluta- mine, penicillin (200 units/ml), streptomycin (200 pg/ml), and amphotericin (10 pg/ml).

RS Virus Pneumonia in Cebus Monkeys 47
Experimental Protocol
A total of 18 monkeys were studied in two separate experiments. The monkeys were lightly anesthetized with ketamine hydrochloride administered intramuscularly prior to administration of virus or uninfected cell suspension. With a 23-gauge needle 0.5 ml of virus or control uninfected cell suspension was inoculated transtracheally through sur- gically prepared skin immediately below the cricoid cartilage. Animals were examined daily for evidence of respiratory disease and rectal temperature was recorded. Nasal and throat swab specimens were taken daily and combined in 1 ml of veal infusion broth as described previously [Belshe et al, 19771. These were quick-frozen in a dry ice-alcohol bath and stored at -7OOC until the end of the experiment, at which time they were assayed for RS virus by the plaque technique. Chest x-rays were taken every two days and at the time the animals were sacrificed. On Days 1, 2,4, 6, and 8 after inocula- tion 2-4 monkeys were selected randomly and sacrificed - a total of 13 virus-infected and 3 control monkeys (Table I). Tissue samples from the respiratory tract were taken for gross and histologic examination and for viral assay.
Viral Assays
The quantity of RS virus in each nasopharyngeal swab specimen was determined by the plaque technique using HEp-2 cultures as previously described [Gharpure et al, 19691 I The amount of virus present in the various respiratory tissues was quantitated in the following manner. Animals were sacrificed and nasal turbinates, trachea, lungs, and hilar lymph nodes were removed and weighed. Samples of each tissue were quick-frozen in L-15 medium (Flow Laboratories) supplemented wtih 10% agamma calf serum and glutamine and antibiotics as described above, and stored at -70°C. At the end of the experiment, each tissue sample was thawed and ground in a mortar with 2 ml of L-15 niediuni and processed as previously described [Wright et al, 19701 . The content of Rs virus was determined by plaque assay using HEp-2 monolayers and was expressed as pfu of virus per gram of tissue [Gharpure et al, 19691 .
TABLE I. Experimental Infection of Cebus Monkeys With Respiratory Syncytial Virus
No. of animals tested on indicated Dostinoculation dav
Inoculum Procedure 0 1 2 3 4 5 6 1 8
lO*pfu of Nasopharyngeal 14 14 12 10 10 7 7 4 4 RS virus swab specimen
taken Chest x-ray 14 2 9 0 1 0 0 7 0 4 Sacrificed 0 2 2 0 3 0 3 0 3
Uninfected Nasophaiyngeal 4 4 4 3 3 2 2 1 1 HeLa cells swab specimen
taken Chest x-ray 4 2 3 0 3 0 2 0 0 Sacrificed 0 0 1 0 1 0 1 0 0

48 Richardson et a1
Histology
Tissues from the respiratory tract were removed from each animal and were fixed in formalin, washed in ethanol, and embedded in paraffin. Lungs were inflated at the time of sacrifice with buffered formalin in order to preserve lung tissue in an expanded state [Prince and Porter, 19761.
lmmunofluorescence Technique
Tissue samples from the respiratory tract were removed from each animal, quick- frozen, and stored at -70°C in Cryoforma (International Equipment Co.. Needhain Heights, Massachusetts). In order to preserve lung tissue in an expanded state, lung tissue was inflated with Cryofoim diluted 1:2 in phosphate buffered saline. [Prince and Porter, 19761. Ciyostat sections (12 p in thickness) were stained by the indirect immuno- fluorescence technique as previously described [Schnitzer et al, 1976J . Anti-RS virus bovine serum was applied to the sections followed by fluorescein-labeled rabbit anti- bovine immunoglobulin (Wellcome Research Laboratories, Beckenham, England).
RESULTS
Clinical Response of Cebus Monkeys to Transtracheally Administered RS Virus
Six of the 14 monkeys given 10' pfu of RS virus transtracheally developed mild upper respiratory tract disease characterized by rhinorrliea and conjunctivitis. Only one infected animal developed signs of lower respiratory tract disease, which consisted of coughing, dyspnea, anorexia, and lethargy. The infected animals did not develop a febrile response; their temperature pattern did not differ from that of the control monkeys.
Roentgenographic Examination
Although 13 of the 14 monkeys inoculated with virus had no lower respiratory tract signs, each of the monkeys except one showed roentgenographic evidence of pul- monary disease. The only animal that failed to develop roentgenographic changes was sacrificed 24 hours after infection; however, the other monkey sacrificed at 24 hours did exhibit changes. On the fourth day after inoculation each of the ten monkeys x-rayed had extensive infiltrates in the lower lobes, as did the majority of animals studied on the sixth and eighth day after inoculation (Tabie 11). In contrast, animals inoculated with win - fected cell suspension did not develop abnormalities discernible by x-ray.
Pathology
Gross pathologic changes in the lungs were detected as early as 24 hours after inoculation of virus and were seen in both of the infected animals sacrificed at that time (Table 111). By 48 hours therc were areas of red consolidation throughout both lower lobes. In general, the abnormalities were more prominent in the right lower lobe and more patchy in the left lower lobe. This may reflect the site to which the inoculated virus was aspirated. On the sixth day after inoculation extensive consolidation was evident in both lower lobes as well as in the intermediate and cardiac lobe (Fig 1B). Gray consoli- dation of the entire right lower lobe, cardiac, intermediate, and part of the apical lobe

R S Virus Pneumonia in Cebus Monkeys 49
TABLE 11. Roentgenographic Evidence of Pneumonia in Cebus Monkeys Inoculated With Respiratory Syncytial Virus*
Po s tiiio Lulation day -~ - __ Inoculum 0 1 2 4 6 8
~~~ ~~~
lo8 pfu of KSvi ru r 0/14 1/2 3/9 lO/l0 6/7 3/4
HeLa ccils 0 /4 012 013 0/3 0 / 2 NTa UninfeLted
*Number of monkeys with roentgenographic evidence of pneumonia/ number of monkeys exdrnined on the lndlcated postmocrilation day a h T = not tested
TABLE 111. Gross Pathologic Evidence of Pneumonia in Cebus Monkeys Inoculated With Respiratory Syncytial Virus*
Postinoculation day ~~ ~~
- Inoculum 1 2 4 6 8 ~ - ______ ~
lo8 pfu of RS virus 212 2 /2 3/3 313 313
Uninfectcd HeLacells NT" O i l 0/1 0 / l N1'
"Number of monkcys with gross pathologic evidence of pneumonia/number of monkeys examined on the indicated postinoculation day.
"NT = not tested
was observed on the eighth day. Abnormalities were not noted in the lungs of any of the control animals (Fig 1A). By gross inspection, the nasal turbinates and trachea of the in- fected and control animals appeared normal.
ings. No abnornialities were noted in the nasal turbinates or trachea from control or virus- infected animals. The lungs of control animals were uniformly negative (Table 1V and Fig 2). In contrast, the lungs of each virus-inoculated animal showed signs of interstitial pneumonia. Microscopically, there was thickening of the alveolar wall with infiltration by polyniorphonuclear leukocytes (PMLs) and mononuclear inflammatory cells as early as 24 hours after inoculation (Fig 3). This was accompanied by an inflammatory response within the alveoli that also consisted of PMLs and mononuclear cells. By the fourth day, extensive interstitial infiltration with mononuclear cells and PMLs was seen in the alveoli, and occasional multinucleated giant cells (syncytia) were evident. Syncytia became more prominent by day six, when extensive interstitial pneumonia, areas of consolidation, and mononuclear cell infiltrates were also seen (Fig 4). Eosinophilic cytoplasmic inclusion bodies were additionally noted a t that time (Fig 5). On the eighth day there were exten- sive areas of interstitial thickening, pulmonary consolidation, and alveoli filled with proteinaceous fluid and inflaminatory cells (Fig 6).
Histologic examination of respiratory tissue confirnied the gross pathologic find-

50 Richardson et a1
c
w
'c 0

RS Virus Pneumonia in Cebus Monkeys 51
TABLE 1%'. Histologic Evidence of Pneumonia in Cebus Monkeys Inoculated With Respiratory Syncytial Virus*
Postinoculation day ~ -~
l n o u l u m 1 2 4 6 8 ~ ~ ~
lo8 pfu of RS birus 212 212 313 313 313
HeLa ~ c l l h NT" Oil 011 011 NT linlnfec ted
'Number of monkeys with histologic evidence of pneumonia/ number of monkeys examined on the indicated postinocula- tion day.
"NT = tlot tested
Fig. 2 . Section of control cebus lung. No histologic lesions were identified in monkeys sacrificed o n days two, four, or six after inoculation of uninfected HeLa cells (H and E, original rnagnification x 800).

5 2 Richardson e t a1
Fig. 3 . Thickening of the alveolar walls with intraalveolar inflammatory response in the lung of a cebus monkey sacrificed 24 hours after inoculation of 108pfu of RS virus (H and L, original magnification x 800).
Fig. 4 . Interstitial pneumonia with giant cell formation in the lung of a cebus monkey sacrificed on day six after inoculation of 10'pfu of KS virus. (H and E , original magnification x 800).

RS Virus Pneumonia in Cebus Monkeys 53
Fig. 5 . Two eosinophilic cytoplasmic inclusion bodies (arrows) in an arca of consolidation from the lung o f a cebus monkey sacrificed on day six after inoculation of 10'pfu of KS virus (H and E, original magnification X 2 . O W ) .
Fig. 6 . Consolidation in the lung of a cebus monkey sacrificed on day eight after inoculation of lO'pfu of RS virus (I1 and E, original niagnification x 500).
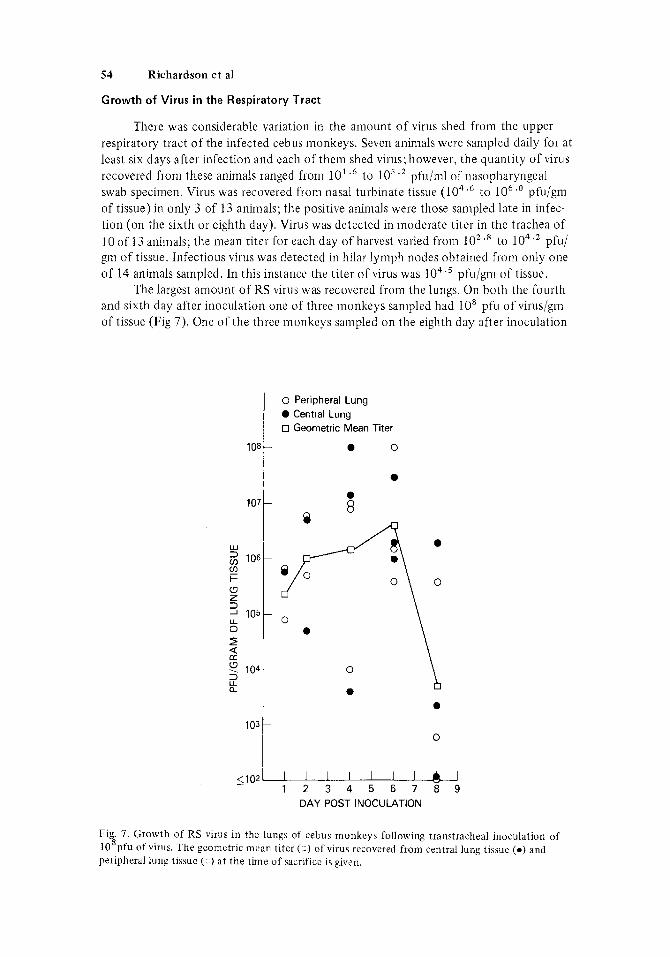
54 Richardson et al
Growth of Virus in the Respiratory Tract
There was considerable variation in the amount o f virus shed from the upper respiratory tract of the infected cebiis monkeys. Seven animals were sampled daily for at lcast six days after infection and each of thcni shed virus; however, the quantity of virus recovered from these animals ranged from 10' .' to los .* pfu/ml of riasopharyngeal swab specimen. Virus was recovered from nasal turbinate tissue (104.6 to 1 O h . ' pfu/gm of tissue) in only 3 of 13 animals; the positive animals were those sampled late in infec- tion (on the sixth or eighth day). Virus was detected in moderate titer in the trachea of 10 of 13 animals: the mean titer for each day of harvest varied from lo2 .* to 104'2 pfu/ grn of tissue. Infectious virus w-as detected in hilar lymph nodes obtained from only one of 14 animals sampled. In this instance the titer of virus was lo4.' pfu/gni of tissue.
The largest amount of RS virus was recovered from the lungs. On both the fourth and sixth day after inoculation one of three monkeys sampled had lo8 pfu o f virus/gm of tissue (Fig 7). One of the three monkeys sampled o n the eighth day after inoculation
108
107
Lu
2 106 v, I- U z
105 U 0 2 2 ? 104 i? a
103
2102
0 Peripheral Lung Central Lung
0 Geometric Mean Titer
0
0
0 \
0
0 - 1 2 3 4 5 6 7 8 9
DAY POST INOCULATION
Fig. 7. Growth of RS virus in thc lungs of cebus monkeys following transtracheal inoculation of 10 pfu of virus. The geometric mean titer ( 1) of virus recovered from central lung tissue ( 0 ) and peripheral lung tissue ( -1 ) at the time of sacrifice is given.
8

RS Virus Pneumonia in Cebus Monkeys 5 5
still had a moderately large quantity of virus in the lungs (lo6 . 2 pfu/gin). There were no significant differences in the amount of virus recovered from central or peripheral areas of the lungs.
Localization of Virus Antigens
RS viral antigens were detected by the indirect immunofluorescence technique in the nasal turbinates, trachea, and lungs of infected animals. In the nasal turbinates, antigen-positive cells were abundant in the surface epithelium. In contrast, only occa- sional discrete cells of the tracheal epithelium contained viral antigens (Fig 8). In lung tissue, antigens were present in alveolar cells dispersed throughout the lung parenchyma (Fig 9), as well as in cells of the bronchiolar epithelium (Fig 10). In general, antigens were detected only in those lungs from which more than lo4.' pfu were isolated per gm of tissue (data not shown).
were stained consecutively by indirect immunofluorescence (to identify the RS viral antigen-positive cells) and periodic acid-Schiff stain ( to identify goblet cells). Infection involved both goblet and ciliated epithelial cells. Howcver, only a srnall proportion of each cell type appeared to be infected.
The unusual staining pattern seen in the trachea was investigated further. Sections
Fig. 8 Section of trachea stained by the Lndirect iinmunofluorescence technique for the presence of RS viral antigens from a ccbus rnonkcy sdcrificed on day six dfter inoculation of 108pfu of RS virus (original magnification x 250)

56 Richardson et a1
Fig. 9 . RS virus antigcn-positive cells as detected by the indirect itnInunofluorescence technique in the alveolar walls of the lung from a cebus monkey sacrificed on day six after inoculation of 108pfu of R S virus (original magnification X 500).
b'ig. 10. RS virus antigen-positive cells as dctected by the indirect iiiitiiunofluorescence technique in the bronchiolar cpitheliuni o f thc lung from a cebus monkey sacrificed on day six after inoculation of 10*pfu of RS virus (original magnification x 150).

RS Virus Pneumonia in Cebus Monkeys 57
Response of Cebus Monkeys to Lower Dose of Virus
One of two animals inociilated with a 10-fold smaller quantity of virus (107pfu) became infected and developed pneumonia similar to that seen in the 14 animals that re- ceived 10spfu of virus. The other monkey that received 107pfu did not become infected and failed t o develop roentgenographic or pathologic changes. Pneumonia did not develop in either of two monkeys inoculated with 106pfu of virus although one animal became in- fected, as evidenced by the recovery of lo4.' pfu of virus per gram of lung tissue.
DISCUSS ION
The development of pneumonia in juvenile cebus monkeys following inoculation of a large quantity of RS virus provides the first experimental aninial in which t o study lower respiratory disease caused by this virus. The presence of large amounts of RS virus in the lungs and the abundance of RS viral antigens in alveolar cells and bronchioles, as well as the complete absence of pathologic changes in control animals, suggest that RS virus was responsible for the observed pneumonia. The histologic finding of multi- nucleated giant cells and eosinophilic cytoplasmic inclusion bodies in the infected cebus lungs are reminiscent of the characteristic syncytia formation [Chanock et al, 19571 and inclusion bodies [Peacock and Clarke, 1961 ; Holzel et al, 19631 seen in cells infected with RS virus in vitro. Similar giant cell formation has also been reported in the nasal epithelium of RS virus-infected ferrets [Coates and Chanock, 19621, as well as the bronchial epithelium of RS virus-infected cotton rats [Dreizen et al, 19711. The cyto- plasmic inclusion bodies seen in the cebus monkey resemble those described by A d a m in fatal inclusion body pneumonia of the newborn as well as in patients who died of acute RS virus bronchiolitis [Adams, 1941; Aherne et al, 1970; Holzel e t al, 196.51. Similar inclusions have also been reported in the epithelium of the nasal turbinates of RS virus-infected ferrets [Coates and Chanock, 1Y621 .
Although it might appear that lo8 pfu is an unnaturally large inoculum, in fact comparable quantities of virus are present in the respiratory tract of RS virus-infected chimpanzees and man. Chimpanzees shed up t o lo6.' pfu of virus per nil o f naso- pharyngeal swab specimen [Belshe et al, 19771 , and this sampling procedure represented at least a lo-' .' dilution of the quantity of virus present in nasopharyngeal secretions [Richardson and Belshe. unpublished observations]. In children, Hall et a1 [1975] have reported quantities of virus 2 106.2 TCID,, /nil (tissue culture infective dose) in nasal wash samples. No estimate was given in this study of the dilution factor; however, 5-9 ml of medium was used per nasal wash. Thus, it is possible that the infants might on occasion be exposed t o a large amount of virus by aspirating their own infectious nasal secretions.
fected. Using double-staining techniques, we observed that RS virus replicated in both the goblet cells and ciliated epithelial cells of the trachea. However, only a small proportion of either cell type became infected. Thus, it is unlikely that RS virus produced a specific effect upon cells that produce niucous in the monkey.
Severe RS virus infection in the human infant is often manifested as either bron- chiolitis or pneumonia. Histologically, necrosis of the bronchiolar epithelium with peri- bronchiolar inflammatory response as well as interstitial pneumonia are seen in infants with fatal RS virus disease [Aherne et al, 1970; Holzel e t al, 1965; Gardner et al, 19701 . Extensive pulmonary changes in RS virus-infected cebus monkeys were limited to inter- stitial pneumonia without significant broncliiolar pathology. None of the 13 infected
In the cebus monkey only occasional cells of the tracheal epithelium became in-

58 Richardson e t a1
monkeys displayed a histopatho!ogic pattern similar to that seen in acute bronchiolitis of infants. However, viral replication occurred in both the alveolar cell wall and bron- chiolar epithelium, as indicated by the indirect immunofluorescence technique. This is in contrast to the response of other animals to RS virus. For example, infection in the ferret is limited to alveolar cells [Prince and Porter, 19761 , while in the cotton rat the bronchiolar epithelium is the primary site of viral replication [Dreizen et d, 19711.
A dose of 108'Opfu appeared to be the pneumonia doseloo, since 107'*pfu of RS virus infected only one of two cebus monkeys; this animal developed gross pneumonia. A dose of lo6 pfu also infected one of two animals, but pneumonia did not develop. Identification of the cebus monkey as an animal that responds to extensive infection by developing definite pulmonary lesions means that an experimental primate system is now available that can be manipulated to convert an inapparent infection (such as that induced by lo6 pfu) into a fulminant infection with development of extensive pulmon- ary pathology. In this way the important determinants of the pathogenesis of RS virus disease may be amenable to experimental study.
ACKNOWLEDGMENTS
This study was supported in part by the National Institutes of Health, NINCDS,
The authors wish to thank Mr. James Edwards, Mr. Ivan Lantz, and Mr. Henry contract NOI-NS-4-2325.
Asbury of Meloy Laboratories for their excellent technical assistance.
REFERENCES
Adams JM (1941): Primary virus pneumonitis with cytoplasmic inclusion bodies. Journal of thc
Aherne W, Bird T, Court SDM, Gardner PS, McQuillin J (1970): Pathological changes in virus infec-
Belshe RB, Richardson LS, London WT, Sly DL, Lorfeld JH, Camargo E, Prevar DA, Chanock RM
American Medical Association 116 :925.
tions of the lower respiratory tract in children. Journal of Clincial Pathology 23:7.
(1977) : Experimental respiratory syncytial virus infection of 4 species of primates. Journal of Medical Virology 1 : 157.
related to chimpanzee coryza agent (CCA). 1. Isolation, properties and characterization. Ameri- can Journal of Hygiene 66:281.
Coates HV, Chanock RM (1962) : Experimental infection with respiratory syncytial virus in several species of animals. American Journal of Hygiene 76:302.
Dreizin RS, Vyshnevetskaia LO, BagdamIan EE, Y ankevich OD, Tarasova LB, Klenova AV (197 1) : [Study of experimental respiratory syncytial virus infection in cotton rats: Virologic and immunofluorescent studies.] Voprosy Virusologii 16 :670.
Gardner PS, McQuillin J ? Court SDM (1970): Speculation on pathogenesis in death from respiratory syncytial virus infection. British Medical Journal 1:327.
Gharpure MA, Wright PF, Chanock RM (1969) : Temperature-sensitive mutants of respiratory syn- cytial virus. Journal of Virology 3:414.
Hall CB, Douglas RG, Geiman JM (1975) : Quantitative shedding patterns of respiratory syncytial virus in infants. Journal of Infectious Diseases 132:151.
Holzel A, Parker L, Patterson WH, Cartmel D, White LLR, Purdy R, Thompson KM, Tobin JOH (1965): Virus isqlations from throats of children admitted to hospital with respiratory and other diseases. Manchester 1962-4. British Medical Journal 1 :6 14.
Chanock RM, Roizman B, Myers R (1957): Recovery from infants with respiratory illness of a virus

RS Virus Pneumonia in Cebus Monkeys 59
Holzel A, Parker L, Patterson WH, White LLR, Thompson KM, Tobin JOH (1963): The isolation of respiratory syncytial virus from children with acute respiratory disease. Lancet 1 :295.
London WT, Alter HJ, Lander J , Purcell RH (1972): Serial transmission in rhesus monkeys of an . agent related to hepatitis-associated antigen. Journal of Infectious Diseases 125 :382.
Mills J , VanKirk JE, Wright PF, Chanock RM (197 1): Experimental respiratory syncytial virus infec- tion of adults. Journal of Immunology 107:123.
Peacock DB, Clarke SKR (1961): Respiratory syncytial virus in Britain. Lancet 1 :446. Prince GA, Porter DD (1976): The pathogenesis of respiratory syncytial virus infection in infant
ferrets. American Journal of Pathology 82:339. Schnitier TJ, Richardson LS, Chanock RM (1976): Growth and genetic stability of the ts-1 mutant
of respiratory syncytial virus a t restrictive temperatures. Journal of Virology 17 :43 1. Wright PF, Woodend WG, Chanock RM (1970) : Temperature-sensitive mutants of respiratory syn-
cytial virus: In vivo studies in hamsters. Journal of Infectious Diseases 122:501.
Copyright © 2022 FDOKUMEN




















