
Evolutionary history of mammalian sucking lice (Phthiraptera: Anoplura)

733
Part Opener TitleText to come.
Part XPlant Form and Function
Part opener figure 1 title. Figure legend.

734 Part X Plant Form and Function
Part opener figure 2 title. Figure legend.

735
37Evolutionary History
of Plants
Concept Outline
37.1 Plants have multicellular haploid and diploidstages in their life cycles.
The Evolutionary Origins of Plants. Plants evolvedfrom freshwater green algae and eventually developedcuticles, stomata, conducting systems, and reproductivestrategies that adapt them well for life on land.Plant Life Cycles. Plants have haplodiplontic life cycles.Diploid sporophytes produce haploid spores which developinto haploid gametophytes that produce haploid gametes.
37.2 Nonvascular plants are relatively unspecialized,but successful in many terrestrial environments.
Mosses, Liverworts, and Hornworts. The mostconspicuous part of a nonvascular plant is the greenphotosynthetic gametophyte, which supports the smallersporophyte nutritionally.
37.3 Seedless vascular plants have well-developedconducting tissues in their sporophytes.
Features of Vascular Plants. In vascular plants,specialized tissue called xylem conducts water and dissolvedminerals within the plant, and tissue called phloem conductssucrose and plant growth regulators within the plant.Seedless Vascular Plants. Seedless vascular plants have amuch more conspicuous sporophyte than nonvascularplants do, and many have well-developed conductingsystems in stem, roots, and leaves.
37.4 Seeds protect and aid in the dispersal of plantembryos.
Seed Plants. In seed plants, the sporophyte is dominant.Male and female gametophytes develop within thesporophyte and depend on it for food. Seeds allow embryosto germinate when conditions are favorable.Gymnosperms. In gymnosperms, the femalegametophyte (ovule) is not completely enclosed bysporophyte tissue at the time of pollination by malegametophytes (pollen).Angiosperms. In angiosperms, the ovule is completelyenclosed by sporophyte tissue at the time of pollination.Angiosperms, by far the most successful plant group,produce flowers.
Plant evolution is the story of the conquest of land bygreen algal ancestors. For about 500 million years,
algae were confined to a watery domain, limited by theneed for water to reproduce, provide structural support,prevent water loss, and provide some protection from thesun’s ultraviolet irradiation. Numerous evolutionary solu-tions to these challenges have resulted in over 300,000species of plants dominating all terrestrial communitiestoday, from forests to alpine tundra, from agriculturalfields to deserts (figure 37.1). Most plants are photosyn-thetic, converting light energy into chemical-bond energyand providing oxygen for all aerobic organisms. We relyon plants for food, clothing, wood for shelter and fuel,chemicals, and many medicines. This chapter explores theevolutionary history and strategies that have allowedplants to inhabit most terrestrial environments over mil-lions of years.
FIGURE 37.1 An arctic tundra. This is one of the harshest environments onearth, yet a diversity of plants have made this home. Theseecosystems are fragile and particularly susceptible to globalchange.

Adaptations to Land
Plants and fungi are the only major groups of organismsthat are primarily terrestrial. Most plants are protectedfrom desiccation—the tendency of organisms to lose waterto the air—by a waxy cuticle that is secreted onto their ex-posed surfaces. The cuticle is relatively impermeable andprovides an effective barrier to water loss.
This solution creates another problem by limiting gasexchange essential for respiration and photosynthesis.Water and gas diffusion into and out of a plant occursthrough tiny mouth-shaped openings called stomata (sin-gular, stoma).
The evolution of leaves resulted in increased photo-synthetic surface area. The shift to a dominant diploidgeneration, accompanied by the structural support of vas-cular tissue, allowed plants to take advantage of the verti-cal dimension of the terrestrial environment, resulting intrees.
Plants evolved from freshwater green algae andeventually developed cuticles, stomata, conductingsystems, and reproductive strategies that adapt themwell for life on land.
736 Part X Plant Form and Function
The Evolutionary Origins of PlantsBiologists have long suspected that plants are the evolu-tionary descendants of green algae. Now we are sure. Theevolutionary history of plants was laid bare at the 1999 In-ternational Botanical Congress held in St. Louis, Mis-souri, by a team of 200 biologists from 12 countries thathad been working together for five years with U.S. federalfunding. Their project, Deep Green (more formallyknown as The Green Plant Phylogeny Research Coordi-nation Group), coordinated the efforts of laboratoriesusing molecular, morphological, and anatomical traits tocreate a new “Tree of Life.” Deep Green confirmed thelong-standing claim that green algae were ancestral toplants. More surprising was the finding that just a singlespecies of green algae gave rise to the entire terrestrialplant lineage from moss through the flowering plants (an-giosperms). Exactly what this ancestral alga was is still amystery, but close relatives are believed to exist in fresh-water lakes today. DNA sequence data is consistent withthe claim that a single “Eve” gave rise to the entire king-dom Plantae 450 million years ago. At each subsequentstep in evolution, the evidence suggests that only a singlefamily of plants made the transition. The fungi appear tohave branched later than the plants and are more closelyrelated to us.
There are 12 plant phyla, all of which afford some pro-tection to their embryos. All plants also have a haploid anda diploid stage that is multicellular. The trend over timehas been toward increasing embryo protection and asmaller haploid stage in the life cycle. The plants are di-vided into two groups based on the presence or absence ofvascular tissues which facilitate the transport of water andnutrients in plants. Three phyla (mosses, liverworts, andhornworts) lack vascular tissue and are referred to as thenonvascular plants. Members of 9 of the 12 plant phylaare collectively called vascular plants, and include, amongothers, the ferns, conifers, and flowering plants. Vascularplants have water-conducting xylem and food-conductingphloem strands of tissues in their stems, roots, and leaves.Vascular plants can be further grouped based on how muchprotection embryos have. The seedless vascular plants(ferns) provide less protection than the seeds of the gym-nosperms (conifers) and angiosperms (flowering plants)(figure 37.2). About 150 million years ago the angiospermsarose with further innovations—flowers to attract pollina-tors and fruit surrounding the seed to protect the embryoand aid in seed dispersal. Many of these lineages have per-sisted. If you could travel back 65 million years to the di-nosaur era, you would encounter oak, walnut, andsycamore trees!
37.1 Plants have multicellular haploid and diploid stages in their life cycles.
Non
vasc
ular
pla
nts
See
dles
s va
scul
ar p
lant
s
Gym
nosp
erm
s
Ang
iosp
erm
s
Plants
FIGURE 37.2The four major groups of plants. In this chapter we will discussfour major groups of plants. The green algae, discussed in chapter35, are the protists from which plants are thought to have evolved.

Plant Life CyclesAll plants undergo mitosis after both gametefusion and meiosis. The result is a multicellu-lar haploid and a multicellular diploid individ-ual, unlike us where gamete fusion directly fol-lows meiosis. We have a diplontic life cycle(only the diploid stage is multicellular), but theplant life cycle is haplodiplontic (with multi-cellular haploid and diploid stages). The basichaplodiplontic cycle is summarized in figure37.3. Brown, red, and green algae are also hap-lodiplontic (see chapter 35). While we producegametes via meiosis, plants actually producegametes by mitosis in a multicellular, haploidindividual. The diploid generation, or sporo-phyte, alternates with the haploid generation,or gametophyte. Sporophyte means “sporeplant,” and gametophyte means “gameteplant.” These terms indicate the kinds of re-productive cells the respective generationsproduce.
The diploid sporophyte undergoes meiosisto produce haploid spores (not gametes).Meiosis takes place in structures called spo-rangia, where diploid spore mother cells(sporocytes) undergo meiosis, each producingfour haploid spores. Spores divide by mitosis,producing a multicellular, haploid gameto-phyte. Spores are the first cells of the gameto-phyte generation.
In turn, the haploid gametophyte produces haploid ga-metes by mitosis. When the gametes fuse, the zygotethey form is diploid and is the first cell of the next sporo-phyte generation. The zygote grows into a diploid sporo-phyte that produces sporangia in which meiosis ulti-mately occurs.
While all plants are haplodiplontic, the haploid genera-tion consumes a much larger chunk of the life cycle inmosses than in gymnosperms and angiosperms. In mosses,liverworts, and ferns, the gametophyte is photosyntheticand free-living; in other plants it is either nutritionally de-pendent on the sporophyte, or saprobic (deriving its en-ergy directly from nonliving organic matter). When youlook at moss, what you see is largely gametophyte tissue;their sporophytes are usually smaller, brownish or yellow-ish structures attached to or enclosed within tissues of thegametophyte. In most vascular plants the gametophytes aremuch smaller than the sporophytes. In seed plants, the ga-metophytes are nutritionally dependent on the sporophytesand are enclosed within their tissues. When you look at agymnosperm or angiosperm, what you see, with rare excep-tions, is a sporophyte.
The difference between dominant gametophytes andsporophytes is key to understanding why there are nomoss trees. What we identify as a moss plant is a gameto-
phyte and it produces gametes at its tip. The egg is sta-tionery and sperm lands near the egg in a droplet ofwater. If the moss were the height of a sequoia, not onlywould it need vascular tissue for conduction and support,the sperm would have to swim up the tree! In contrast,the fern gametophyte develops on the forest floor wheregametes can meet. Fern trees abound in Australia and thehaploid spores fall to the ground and develop into game-tophytes.
Having completed our overview of plant life cycles, wewill consider the major plant groups. As we do, we willsee a progressive reduction of the gametophyte fromgroup to group, a loss of multicellular gametangia(structures in which gametes are produced), and increas-ing specialization for life on the land, culminating withthe remarkable structural adaptations of the floweringplants, the dominant plants today. Similar trends musthave characterized the progression to seed plants over thehundreds of millions of years since a freshwater algamade the move onto land.
Plants have haplodiplontic life cycles. Diploidsporophytes produce haploid spores which develop intohaploid gametophytes that produce haploid gametes.
Chapter 37 Evolutionary History of Plants 737
2n2n
Egg
Mitosis
Spore
2n
Sporophyte(2n)
Sporangia
Spore mothercell
Sporesn
nn
n
Gametophyte(n)
Gamete fusion
Sperm
Embryo
Zygote
Meiosis
Haploid
Diploid
FIGURE 37.3A generalized plant life cycle. Anything yellow is haploid and anything blue isdiploid. Note that both haploid and diploid individuals can be multicellular. Also,spores are produced by meiosis while gametes are produced by mitosis.

Mosses, Liverworts, andHornwortsThere are about 24,700 bryophytes—mosses, liverworts, and hornworts—that are simply but highly adapted to adiversity of terrestrial environments(even deserts!). Scientists now agreethat bryophytes consist of three quitedistinct phyla of relatively unspecial-ized plants. Their gametophytes arephotosynthetic. Sporophytes are at-tached to the gametophytes and de-pend on them nutritionally to varyingdegrees. Bryophytes, like ferns and cer-tain other vascular plants, requirewater (for example, rainwater) to re-produce sexually. It is not surprisingthat they are especially common inmoist places, both in the tropics and tem-perate regions.
Most bryophytes are small; few exceed 7centimeters in height. The gametophytesare more conspicuous than the sporo-phytes. Some of the sporophytes are com-pletely enclosed within gametophyte tissue;others are not and usually turn brownish orstraw-colored at maturity.
Mosses (Bryophyta)
The gametophytes of mosses typicallyconsist of small leafl ike structuresarranged spirally or alternately around astemlike axis (figure 37.4); the axis is an-chored to its substrate by means of rhi-zoids. Each rhizoid consists of severalcells that absorb water, but nothing likethe volume of water absorbed by a vascu-lar plant root. Moss “leaves” have little incommon with true leaves, except for thesuperficial appearance of the green, flat-tened blade and slightly thickened midribthat runs lengthwise down the middle.They are only one cell thick (except at themidrib), lack vascular strands and stomata, and all thecells are haploid.
Water may rise up a strand of specialized cells in thecenter of a moss gametophyte axis, but most water used bythe plant travels up the outside of the plant. Some mosses
also have specialized food-conductingcells surrounding those that conductwater.
Multicellular gametangia are formedat the tips of the leafy gametophytes(figure 37.5). Female gametangia(archegonia) may develop either on thesame gametophyte as the male ga-metangia (antheridia) or on separateplants. A single egg is produced in theswollen lower part of an archegoniumwhile numerous sperm are produced inan antheridium. When sperm are re-leased from an antheridium, they swimwith the aid of flagella through a film ofdew or rainwater to the archegonia.One sperm (which is haploid) uniteswith an egg (also haploid), forming adiploid zygote. The zygote divides bymitosis and develops into the sporo-
phyte, a slender, basal stalk with a swollencapsule, the sporangium, at its tip. As thesporophyte develops, its base becomes em-bedded in gametophyte tissue, its nutri-tional source. The sporangium is oftencylindrical or club-shaped. Spore mothercells within the sporangium undergo meio-sis, each becoming four haploid spores. Atmaturity, the top of the sporangium popsoff, and the spores are released. A sporethat lands in a suitable damp location maygerminate and grow into a threadlike struc-ture that branches to form rhizoids and“buds” that grow upright. Each bud devel-ops into a new gametophyte plant consist-ing of a leafy axis.
In the Arctic and the Antarctic, mossesare the most abundant plants, boasting notonly the largest number of individuals inthese harsh regions, but also the largestnumber of species. Many mosses are able towithstand prolonged periods of drought, al-though they are not common in deserts.Most are remarkably sensitive to air pollu-tion and are rarely found in abundance in
or near cities or other areas with high levels of air pollu-tion. Some mosses, such as the peat mosses (Sphagnum),can absorb up to 25 times their weight in water and arevaluable commercially as a soil conditioner, or as a fuelwhen dry.
738 Part X Plant Form and Function
37.2 Nonvascular plants are relatively unspecialized, but successful in manyterrestrial environments.
Non
vasc
ular
pla
nts
See
dles
s va
scul
ar p
lant
s
Gym
nosp
erm
s
Ang
iosp
erm
s
FIGURE 37.4A hair-cup moss, Polytrichum(phylum Bryophyta). The“leaves” belong to thegametophyte. Each of theyellowish-brown stalks, with thecapsule, or sporangium, at itssummit, is a sporophyte.

Liverworts (Hepaticophyta)
The old English word wyrt means“plant” or “herb.” Some common liver-worts have flattened gametophytes withlobes resembling those of liver—hencethe combination “liverwort.” Althoughthe lobed liverworts are the best-knownrepresentatives of this phylum, theyconstitute only about 20% of thespecies (figure 37.6). The other 80%are leafy and superficially resemblemosses. Liverworts are less complexthan mosses. Gametophytes are pros-trate instead of erect, and the rhizoidsare one-celled.
Some liverworts have air chamberscontaining upright, branching rows ofphotosynthetic cells, each chamberhaving a pore at the top to facilitate gasexchange. Unlike stomata, the poresare fixed open and cannot close.
Sexual reproduction in liverworts issimilar to that in mosses. Lobed liver-worts form gametangia in umbrella-like structures. Asexual reproductionoccurs when lens-shaped pieces oftissue that are released from thegametophyte grow to form newgametophytes.
Hornworts (Anthocerotophyta)
The origins of hornworts are a puzzle.They are most likely among the earliestland plants, yet the earliest fossil sporesdate from the Cretaceous period, 65 to145 million years ago, when an-giosperms were emerging.
The small hornwort sporophytes re-semble tiny green broom handles risingfrom filmy gametophytes usually lessthan 2 centimeters in diameter (figure37.7). The sporophyte base is embed-ded in gametophyte tissue, from whichit derives some of its nutrition. How-ever, the sporophyte has stomata, isphotosynthetic, and provides much ofthe energy needed for growth and re-production. Hornwort cells usuallyhave a single chloroplast.
The three major phyla ofnonvascular plants are all relativelyunspecialized, but well suited fordiverse terrestrial environments.
Chapter 37 Evolutionary History of Plants 739
Sperm
FERTILIZATION
Zygote
Developingsporophyte inarchegonium
Maturesporophyte
Parentgametophyte
Sporangium
Antheridium
Egg
Archegonium
Male Female
Gametophytes
Bud
Rhizoid
Germinatingspores Spores
MEIOSIS
n 2n
Mitosis
FIGURE 37.5Life cycle of a typical moss. The majority of the life cycle of a moss is in the haploidstate. The leafy gametophyte is photosynthetic, while the smaller sporophyte is not, andis nutritionally dependent on the gametophyte. Water is required to carry sperm to the egg.
FIGURE 37.6A common liverwort, Marchantia(phylum Marchantiophyta). Thesporophytes are borne within the tissues ofthe umbrella-shaped structures that arisefrom the surface of the flat, green, creepinggametophyte.
FIGURE 37.7Hornworts (phylum Anthocerotophyta).Hornwort sporophytes are seen in thisphoto. Unlike the sporophytes of otherbryophytes, these are photosynthetic, butthey also depend on the gametophyte fornutrition.

Features of Vascular PlantsThe first vascular plants for which we have a relativelycomplete record belonged to the phylum Rhyniophyta;they flourished some 410 million years ago but are now ex-tinct. We are not certain what the very earliest of these vas-cular plants looked like, but fossils of Cooksonia providesome insight into their characteristics (figure 37.8). Cookso-nia, the first known vascular land plant, appeared in thelate Silurian period about 420 million years ago. It was suc-cessful partly because it encountered little competition as itspread out over vast tracts of land. The plants were only afew centimeters tall and had no roots or leaves. They con-sisted of little more than a branching axis, the branchesforking evenly and expanding slightly toward the tips. Theywere homosporous (producing only one type of spore).Sporangia formed at branch tips. Other ancient vascularplants that followed evolved more complex arrangementsof sporangia. Leaves began to appear as protuberancesfrom stems.
Cooksonia and the other early plants that followed it be-came successful colonizers of the land through the develop-ment of efficient water- and food-conducting systemsknown as vascular tissues. The word vascular comes fromthe Latin vasculum, meaning a “vessel or duct.” These tis-sues consist of strands of specialized cylindrical or elon-gated cells that form a network throughout a plant, extend-ing from near the tips of the roots, through the stems, andinto true leaves. One type of vascular tissue, the xylem, con-ducts water and dissolved minerals upward from the roots;another type of tissue, phloem, conducts sucrose and hor-monal signals throughout the plant. It is important to notethat vascular tissue developed in the sporophyte, but (withfew exceptions) not the gametophyte. (See the discussion ofvascular tissue structure in chapter 38.) The presence of acuticle and stomata are also characteristic of vascularplants.
The nine phyla of vascular plants (table 37.1) dominateterrestrial habitats everywhere, except for the highestmountains and the tundra. The haplodiplontic life cyclepersists, but the gametophyte has been reduced during evo-lution of vascular plants. A similar reduction in multicellu-lar gametangia has occurred.
Accompanying this reduction in size and complexity ofthe gametophytes has been the appearance of the seed.Seeds are highly resistant structures well suited to protecta plant embryo from drought and to some extent frompredators. In addition, most seeds contain a supply offood for the young plant. Seeds occur only in het-erosporous plants (plants that produce two types ofspores). Heterospory is believed to have arisen multiple
times in the plants. Fruits in the flowering plants add alayer of protection to seeds and attract animals that assistin seed dispersal, expanding the potential range of thespecies. Flowers, which evolved among the angiosperms,attract pollinators. Flowers allow plants to overcome lim-itations of their rooted, immobile nature and secure thebenefits of wide outcrossing in promoting geneticdiversity.
Most vascular plants have well-developed conductingtissues, specialized stems, leaves, roots, cuticles, andstomata. Many have seeds which protect embryos untilconditions are suitable for further development.
740 Part X Plant Form and Function
37.3 Seedless vascular plants have well-developed conducting tissues in theirsporophytes.
Sporangia
FIGURE 37.8Cooksonia, the first known vascular land plant. Its upright,branched stems, which were no more than a few centimeters tall,terminated in sporangia, as seen here. It probably lived in moistenvironments such as mudflats, had a resistant cuticle, andproduced spores typical of vascular plants. This fossil represents aplant that lived some 410 million years ago. Cooksonia belongs tophylum Rhyniophyta, consisting entirely of extinct plants.

Chapter 37 Evolutionary History of Plants 741
Table 37.1 The Nine Phyla of Extant Vascular Plants
ApproximateNumber of
Phylum Examples Key Characteristics Living Species
Heterosporous seed plants. Sperm not motile; conducted 250,000to egg by a pollen tube. Seeds enclosed within a fruit. Leaves greatly varied in size and form. Herbs, vines, shrubs, trees. About 14,000 genera.
Primarily homosporous (a few heterosporous) vascular 11,000plants. Sperm motile. External water necessary for fertilization. Leaves are megaphylls that uncoil as they mature. Sporophytes and virtually all gametophytes photosynthetic. About 365 genera.
Homosporous or heterosporous vascular plants. Sperm 1,150motile. External water necessary for fertilization. Leaves are microphylls. About 12–13 genera.
Heterosporous seed plants. Sperm not motile; conducted 601to egg by a pollen tube. Leaves mostly needlelike or scalelike. Trees, shrubs. About 50 genera.
Heterosporous vascular seed plants. Sperm flagellated and 206motile but confined within a pollen tube that grows to the vicinity of the egg. Palmlike plants with pinnate leaves. Secondary growth slow compared with that of the conifers. Ten genera.
Heterosporous vascular seed plants. Sperm not motile; 65conducted to egg by a pollen tube. The only gymnosperms with vessels. Trees, shrubs, vines. Three very diverse genera (Ephedra, Gnetum, Welwitschia).
Homosporous vascular plants. Sperm motile. External 15water necessary for fertilization. Stems ribbed, jointed, either photosynthetic or nonphotosynthetic. Leaves scalelike, in whorls, nonphotosynthetic at maturity. One genus.
Homosporous vascular plants. Sperm motile. External 6water necessary for fertilization. No differentiation between root and shoot. No leaves; one of the two genera has scalelike enations and the other leaflike appendages.
Heterosporous vascular seed plants. Sperm flagellated and 1motile but conducted to the vicinity of the egg by a pollen tube. Deciduous tree with fan-shaped leaves that have evenly forking veins. Seeds resemble a small plum with fleshy, ill-scented outer covering.
Anthophyta
Pterophyta
Lycophyta
Coniferophyta
Cycadophyta
Gnetophyta
Arthrophyta
Psilophyta
Ginkgophyta
Flowering plants(angiosperms)
Ferns
Club mosses
Conifers (including pines, spruces, firs, yews,redwoods, and others)
Cycads
Gnetophytes
Horsetails
Whisk ferns
Ginkgo

Seedless Vascular PlantsThe earliest vascular plants lackedseeds. Members of four phyla of livingvascular plants lack seeds, as do at leastthree other phyla known only from fos-sils. As we explore the adaptations of thevascular plants, we focus on both repro-ductive strategies and the advantages ofincreasingly complex transport systems.We will begin with the most familiarphylum of seedless vascular plants, theferns.
Ferns (Pterophyta)
Ferns are the most abundant group ofseedless vascular plants, with about12,000 living species. The fossil recordindicates that they originated during theDevonian period about 350 million yearsago and became abundant and varied in form during thenext 50 million years. Their apparent ancestors had nobroad leaves and were established on land as much as 375million years ago.
Today, ferns flourish in a wide range of habitatsthroughout the world; about 75% of the species, however,occur in the tropics. The conspicuous sporophytes may beless than a centimeter in diameter—as seen in small aquaticferns such as Azolla—or more than 24 meters tall and withleaves up to 5 meters or more long in the tree ferns (figure37.9). The sporophytes and the smaller gametophytes,which rarely reach 6 millimeters in diameter, are both pho-tosynthetic. The fern life cycle differs from that of a mossprimarily in the much greater development, independence,and dominance of the fern’s sporophyte. The fern’s sporo-phyte is much more complex than that of the moss’s; thefern sporophyte has vascular tissue and well-differentiatedroots, stems, and leaves. The gametophyte, however, lacksvascular tissue.
Fern sporophytes typically have a horizontal under-ground stem called a rhizome, with roots emerging fromthe sides. The leaves, referred to as fronds, usually de-velop at the tip of the rhizome as tightly rolled-up coils(“fiddleheads”) that unroll and expand. Many fronds arehighly dissected and feathery, making the ferns that pro-duce them prized as ornamentals. Some ferns, such asMarsilea, have fronds that resemble a four-leaf clover,but Marsilea fronds still begin as coiled fiddleheads.Other ferns produce a mixture of photosynthetic frondsand nonphotosynthetic reproductive fronds that tend tobe brownish in color.
Most ferns are homosporous, producing distinctive, spo-rangia, usually in clusters called sori, typically on the backsof the fronds. Sori are often protected during their devel-opment by a transparent, umbrella-like covering. At first
glance, one might mistake the sori for aninfection on the plant. Diploid sporemother cells in each sporangium un-dergo meiosis, producing haploid spores.At maturity, the spores are catapultedfrom the sporangium by a snapping ac-tion, and those that land in suitabledamp locations may germinate, produc-ing gametophytes which are often heart-shaped, are only one cell thick (except inthe center) and have rhizoids that anchorthem to their substrate. These rhizoidsare not true roots as they lack vasculartissue, but as with many of the nonvascu-lar plants they do aid in transportingwater and nutrients from the soil. Flask-shaped archegonia and globular an-theridia are produced on either the sameor different gametophyte.
742 Part X Plant Form and Function
Non
vasc
ular
pla
nts
Gym
nosp
erm
s
Ang
iosp
erm
s
See
dles
s va
scul
ar p
lant
s
FIGURE 37.9A tree fern (phylum Pterophyta) in the forests of Malaysia.The ferns are by far the largest group of seedless vascular plants.Trees in other phyla are now extinct.
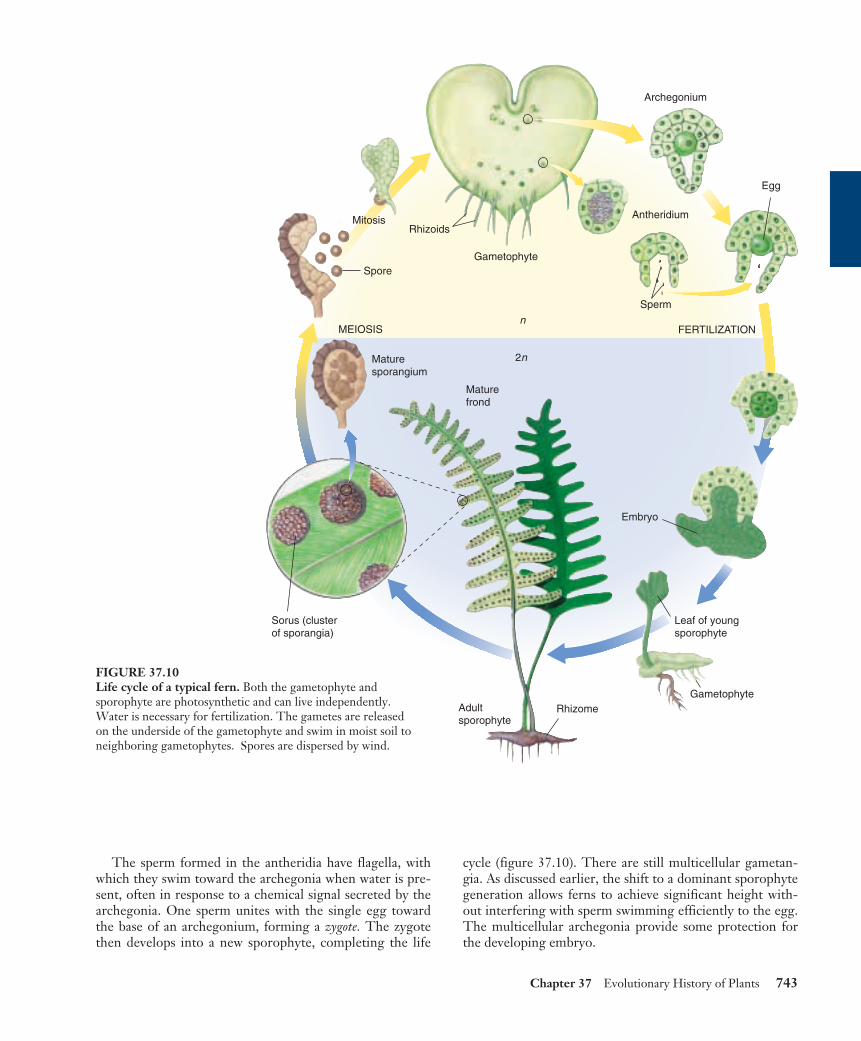
The sperm formed in the antheridia have flagella, withwhich they swim toward the archegonia when water is pre-sent, often in response to a chemical signal secreted by thearchegonia. One sperm unites with the single egg towardthe base of an archegonium, forming a zygote. The zygotethen develops into a new sporophyte, completing the life
cycle (figure 37.10). There are still multicellular gametan-gia. As discussed earlier, the shift to a dominant sporophytegeneration allows ferns to achieve significant height with-out interfering with sperm swimming efficiently to the egg.The multicellular archegonia provide some protection forthe developing embryo.
Chapter 37 Evolutionary History of Plants 743
Archegonium
Egg
Antheridium
Sperm
FERTILIZATION
Embryo
Leaf of youngsporophyte
GametophyteRhizome
Maturefrond
Maturesporangium
MEIOSIS
Spore
Rhizoids
Gametophyte
n
2n
Adultsporophyte
Mitosis
Sorus (clusterof sporangia)
FIGURE 37.10Life cycle of a typical fern. Both the gametophyte andsporophyte are photosynthetic and can live independently.Water is necessary for fertilization. The gametes are releasedon the underside of the gametophyte and swim in moist soil toneighboring gametophytes. Spores are dispersed by wind.

Whisk Ferns (Psilophyta)
The three other phyla of seedless vascular plants, the Psilo-phyta, (whisk ferns), Lycophyta (club mosses), and Arthro-phyta (horsetails), have many features in common withferns. For example, they all form antheridia and archego-nia. Free water is required for the process of fertilization,during which the sperm, which have flagella, swim to andunite with the eggs. In contrast, most seed plants have non-flagellated sperm; none form antheridia, although a fewform archegonia.
The origins of the two genera of whisk ferns, whichoccur in the tropics and subtropics, are not clear, butthey are considered to be living remnants of the very ear-liest vascular plants. Certainly they are the simplest of allextant vascular plants, consisting merely of evenly forkinggreen stems without roots or leaves. The two or threespecies of the genus Psilotum do, however, have tiny,green, spirally arranged, flaps of tissue lacking veins andstomata. Another genus, Tmespiteris , has leaflikeappendages.
The gametophytes of whisk ferns were unknown formany years until their discovery in the soil beneath thesporophytes. They are essentially colorless and are less than2 millimeters in diameter, but they can be up to 18 mil-limeters long. They form saprobic or parasitic associationswith fungi, which furnish their nutrients. Some develop el-ements of vascular tissue and have the distinction of beingthe only gametophytes known to do so.
Club Mosses (Lycophyta)
The club mosses are worldwide in distribution but aremost abundant in the tropics and moist temperate re-gions. Several genera of club mosses, some of them tree-like, became extinct about 270 million years ago. Mem-bers of the four genera and nearly 1000 living species ofclub mosses superficially resemble true mosses, but oncetheir internal structure and reproductive processes be-came known it was clear that these vascular plants arequite unrelated to mosses. Modern club mosses are eitherhomosporous or heterosporous. The sporophytes haveleafy stems that are seldom more than 30 centimeterslong.
Horsetails (Arthrophyta)
The 15 living species of horsetails, also called scouringrushes, are all heterosporous and herbaceous. They con-stitute a single genus, Equisetum. Fossil forms of Equise-tum extend back 300 million years to an era when some oftheir relatives were treelike. Today, they are widely scat-tered around the world, mostly in damp places. Some thatgrow among the coastal redwoods of California may reacha height of 3 meters, but most are less than a meter tall(figure 37.11).
Horsetail sporophytes consist of ribbed, jointed, photo-synthetic stems that arise from branching underground rhi-zomes with roots at their nodes. A whorl of nonphotosyn-thetic, scalelike leaves emerges at each node. The stems,which are hollow in the center, have silica deposits in theepidermal cells of the ribs, and the interior parts of thestems have two sets of vertical, somewhat tubular canals.The larger outer canals, which alternate with the ribs, con-tain air, while the smaller inner canals opposite the ribscontain water.
Ferns and other seedless vascular plants have a muchlarger and more conspicuous sporophyte, with vasculartissue. Many have well-differentiated roots, stem, andleaves. The shift to a dominant sporophyte lead to theevolution of trees.
744 Part X Plant Form and Function
FIGURE 37.11A horsetail, Equisetum telmateia, a representative of the onlyliving genus of the phylum Arthrophyta. This species formstwo kinds of erect stems; one is green and photosynthetic, and theother, which terminates in a spore-producing “cone,” is mostlylight brown.

Seed PlantsSeed plants first appeared about 425 million years ago.Their ancestors appear to have been spore-bearing plantsknown as progymnosperms. Progymnosperms shared sev-eral features with modern gymnosperms, including sec-ondary xylem and phloem (which allows for an increase ingirth later in development). Some progymnosperms hadleaves. Their reproduction was very simple, and it is notcertain which particular group of progymnosperms gaverise to seed plants.
From an evolutionary and ecological perspective, theseed represents an important advance. The embryo is pro-tected by an extra layer of sporophyte tissue creating theovule. During development this tissue hardens to producethe seed coat. In addition to protection from drought, dis-persal is enhanced. Perhaps even more significantly, a dor-mant phase is introduced into the life cycle that allows theembryo to survive until environmental conditions are fa-vorable for further growth.
Seed plants produce two kinds of gametophytes—maleand female, each of which consists of just a few cells. Pollengrains, multicellular male gametophytes, are conveyed tothe egg in the female gametophyte by wind or a pollinator.The sperm move toward the egg through a growing pollentube. This eliminates the need for water. In contrast to theseedless plants, the whole male gametophyte rather thanjust the sperm moves to the female gametophyte. A femalegametophyte develops within an ovule. In flowering plants(angiosperms), the ovules are completely enclosed withindiploid sporophyte tissue (ovaries which develop into thefruit). In gymnosperms (mostly cone-bearing seed plants),the ovules are not completely enclosed by sporophyte tissueat the time of pollination.
A common ancestor with seeds gave rise to thegymnosperms and the angiosperms. Seeds can allow fora pause in the life cycle until environmental conditionsare more optimal.
Chapter 37 Evolutionary History of Plants 745
37.4 Seeds protect and aid in the dispersal of plant embryos.
filament The stalklike structure that sup-ports the anther of a stamen.gametophyte The multicellular, haploidphase of a plant life cycle in which gametesare produced by mitosis.gynoecium The carpel(s) of a flower.heterosporous Refers to a plant that pro-duces two types of spores: microspores andmegaspores.homosporous Refers to a plant that pro-duces only one type of spore.integument The outer layer(s) of anovule; integuments become the seed coat ofa seed.micropyle The opening in the ovule in-tegument through which the pollen tubegrows.nucellus The tissue of an ovule in whichan embryo sac develops.ovary The basal, swollen portion of acarpel (gynoecium); it contains the ovulesand develops into the fruit.ovule A seed plant structure within an ovary;it contains a female gametophyte surroundedby the nucellus and one or two integuments.At maturity, an ovule becomes a seed.pollen grain A binucleate or trinucleateseed plant structure produced from a mi-
crospore in a microsporangium.pollination The transfer of a pollen grainfrom an anther to a stigma in angiosperms,or to the vicinity of the ovule in gym-nosperms.primary endosperm nucleus Thetriploid nucleus resulting from the fusion ofa single sperm with the two polar nuclei.seed A reproductive structure that devel-ops from an ovule in seed plants. It consistsof an embryo and a food supply surroundedby a seed coat.seed coat The protective layer of a seed; itdevelops from the integument or integu-ments.spore A haploid reproductive cell, pro-duced when a diploid spore mother cell un-dergoes meiosis; it gives rise by mitosis to agametophyte.sporophyte The multicellular, diploidphase of a plant life cycle; it is the genera-tion that ultimately produces spores.stamen A unit of an androecium; it con-sists of a pollen-bearing anther and usually astalklike filament.stigma The uppermost pollen-receptiveportion of a gynoecium.
androecium The stamens of a flower.anther The pollen-producing portion of astamen. This is a sporophyte structurewhere male gametophytes are produced bymeiosis.antheridium The male sperm-producingstructure found in the gametophytes ofseedless plants and certain fungi.archegonium The multicellular egg-producing structure in the gametophytes ofseedless plants and gymnosperms.carpel A leaflike organ in angiospermsthat encloses one or more ovules; a unit of agynoecium.double fertilization The process, uniqueto angiosperms, in which one sperm fuseswith the egg, forming a zygote, and theother sperm fuses with the two polar nuclei,forming the primary endosperm nucleus.endosperm The usually triploid (althoughit can have a much higher ploidy level) foodsupply of some angiosperm seeds.
A Vocabulary ofPlant Terms

GymnospermsThere are several groups of living gym-nosperms (conifers, cycads, gneto-phytes, and Ginkgo), none of which aredirectly related to one another, but allof which lack the flowers and fruits ofangiosperms. In all of them the ovule,which becomes a seed, rests exposed ona scale (modified leaf) and is not com-pletely enclosed by sporophyte tissues atthe time of pollination. The name gym-nosperm combines the Greek root gym-nos, or “naked,” with sperma, or “seed.”In other words, gymnosperms arenaked-seeded plants. However, al-though the ovules are naked at the timeof pollination, the seeds of gym-nosperms are sometimes enclosed byother sporophyte tissues by the timethey are mature.
Details of reproduction vary somewhat in gym-nosperms, and their forms vary greatly. For example, cy-cads and Ginkgo have motile sperm, even though thesperm are borne within a pollen tube, while many othershave sperm with no flagella. The female cones range fromtiny woody structures weighing less than 25 grams with adiameter of a few millimeters, to massive structuresweighing more than 45 kilograms growing to lengthsmore than a meter.
Conifers (Coniferophyta)
The most familiar gymnosperms are conifers (phylumConiferophyta), which include pines (figure 37.12),spruces, firs, cedars, hemlocks, yews, larches, cypresses, andothers. The coastal redwood (Sequoia sempervirens), aconifer native to northwestern California and southwesternOregon, is the tallest living vascular plant; it may attainnearly 100 meters (300 feet) in height. Another conifer, thebristlecone pine (Pinus longaeva) of the White Mountains ofCalifornia is the oldest living tree; one is 4900 years of age.Conifers are found in the colder temperate and sometimesdrier regions of the world, especially in the northern hemi-sphere. They are sources of timber, paper, resin, turpen-tine, taxol (used to treat cancer) and other economicallyimportant products.
Pines. More than 100 species of pines exist today, allnative to the northern hemisphere, although the range ofone species does extend a little south of the equator. Pinesand spruces are members of the vast coniferous foreststhat lie between the arctic tundra and the temperate de-ciduous forests and prairies to their south. During thepast century, pines have been extensively planted in thesouthern hemisphere.
Pines have tough, needlelike leavesproduced mostly in clusters of two tofive. The leaves, which have a thick cuti-cle and recessed stomata, represent anevolutionary adaptation for retardingwater loss. This is important becausemany of the trees grow in areas wherethe topsoil is frozen for part of the year,making it difficult for the roots to obtainwater. The leaves and other parts of thesporophyte have canals into which sur-rounding cells secrete resin. The resin,apparently secreted in response towounding, deters insect and fungal at-tacks. The resin of certain pines is har-vested commercially for its volatile liquidportion, called turpentine, and for thesolid rosin, which is used on stringed in-struments. The wood of pines consistsprimarily of xylem tissue that lacks someof the more rigid cell types found in
other trees. Thus it is considered a “soft” rather than a“hard” wood. The thick bark of pines represents anotheradaptation for surviving fires and subzero temperatures.Some cones actually depend on fire to open, releasing seedto reforest burnt areas.
746 Part X Plant Form and Function
Non
vasc
ular
pla
nts
See
dles
s va
scul
ar p
lant
s
Ang
iosp
erm
s
Gym
nosp
erm
s
FIGURE 37.12Conifers. Slash pines, Pinus palustris, in Florida, arerepresentative of the Coniferophyta, the largest phylum ofgymnosperms.

As mentioned earlier, all seed plantsare heterosporous, so the spores giverise to two types of gametophytes (fig-ure 37.13). The male gametophytes ofpines develop from pollen grains, whichare produced in male cones that developin clusters of 30 to 70, typically at thetips of the lower branches; there may behundreds of such clusters on any singletree. The male cones generally are 1 to4 centimeters long and consist of small,papery scales arranged in a spiral or inwhorls. A pair of microsporangia formas sacs within each scale. Numerous mi-crospore mother cells in the microspo-rangia undergo meiosis, each becomingfour microspores. The microspores de-velop into four-celled pollen grains witha pair of air sacs that give them addedbuoyancy when released into the air. Asingle cluster of male pine cones mayproduce more than 1 million pollengrains.
Female cones typically are producedon the upper branches of the same treethat produces male cones. Femalecones are larger than male cones, andtheir scales become woody. Twoovules develop toward the base of eachscale. Each ovule contains a megaspo-rangium called the nucellus. The nu-cellus itself is completely surroundedby a thick layer of cells called the in-tegument that has a small opening (themicropyle) toward one end. One ofthe layers of the integument later be-comes the seed coat. A single mega-spore mother cell within each mega-sporangium undergoes meiosis,becoming a row of four megaspores. Three of the mega-spores break down, but the remaining one, over the betterpart of a year, slowly develops into a female gametophyte.The female gametophyte at maturity may consist of thou-sands of cells, with two to six archegonia formed at themicropylar end. Each archegonium contains an egg solarge it can be seen without a microscope.
Female cones usually take two or more seasons to ma-ture. At first they may be reddish or purplish in color,but they soon turn green, and during the first spring, thescales spread apart. While the scales are open, pollengrains carried by the wind drift down between them,some catching in sticky fluid oozing out of the mi-cropyle. As the sticky fluid evaporates, the pollen grainsare slowly drawn down through the micropyle to the topof the nucellus, and the scales close shortly thereafter.The archegonia and the rest of the female gametophyte
are not mature until about a year later. While the femalegametophyte is developing, a pollen tube emerges from apollen grain at the bottom of the micropyle and slowlydigests its way through the nucellus to the archegonia.While the pollen tube is growing, one of the pollengrain’s four cells, the generative cell, divides by mitosis,with one of the resulting two cells dividing once more.These last two cells function as sperm. The germinatedpollen grain with its two sperm is the mature malegametophyte.
About 15 months after pollination, the pollen tubereaches an archegonium, and discharges its contents into it.One sperm unites with the egg, forming a zygote. Theother sperm and cells of the pollen grain degenerate. Thezygote develops into an embryo within a seed. After disper-sal and germination of the seed, the young sporophyte ofthe next generation grows into a tree.
Chapter 37 Evolutionary History of Plants 747
Longisection ofseed, showingembryo
FERTILIZATION(15 months afterpollination)
Ovulate (seed-bearing)cone
Megasporemother cell
Microsporemother cell
Microspores
Pollen
Pollen-bearingcone
PollentubePollination
n
2n
Megaspore
MEIOSIS
Sporophyte
Seedling
Pineseed
Mature seedcone (2nd year)
Scale
Embryo
Scale
Mitosis
Mitosis
FIGURE 37.13Life cycle of a typical pine. The male and female gametophytes have been dramaticallyreduced in size. Wind generally disperses sperm that is within the male gametophyte(pollen). Pollen tube growth delivers the sperm to the egg on the female cone. Additionalprotection for the embryo is provided by the ovule which develops into the seed coat.

Cycads (Cycadophyta)
Cycads are slow-growing gymnosperms of tropical andsubtropical regions. The sporophytes of most of the 100known species resemble palm trees (figure 37.14a) withtrunks that can attain heights of 15 meters or more. Unlikepalm trees—which are flowering plants—cycads producecones and have a life cycle similar to that of pines. The fe-male cones, which develop upright among the leaf bases,are huge in some species and can weigh up to 45 kilograms.The sperm of cycads, although conveyed to an archego-nium by a pollen tube, have thousands of spirally arrangedflagella. Several species are facing extinction in the wild andsoon may exist only in botanical gardens.
Gnetophytes (Gnetophyta)
There are three genera and about 70 living species ofGnetophyta. Gnetophytes are the closest living relatives ofangiosperms and probably share a common ancestor withthat group. They are the only gymnosperms with vessels (aparticularly efficient conducting cell type) in their xylem—a common feature in angiosperms. The members of thethree genera differ greatly from one another in form. Oneof the most bizarre of all plants is Welwitschia, which oc-curs in the Namib and Mossamedes deserts of southwest-ern Africa (figure 37.14b). The stem is shaped like a large,shallow cup that tapers into a taproot below the surface. Ithas two strap-shaped, leathery leaves that grow continu-ously from their base, splitting as they flap in the wind.The reproductive structures of Welwitschia are conelike,appear toward the bases of the leaves around the rims ofthe stems, and are produced on separate male and femaleplants.
More than half of the gnetophyte species are in thegenus Ephedra, which is common in arid regions of thewestern United States and Mexico. The plants are shrubby,with stems that superficially resemble those of horsetails asthey are jointed and have tiny, scalelike leaves at each node.Male and female reproductive structures may be produced
on the same or different plants. The drug ephedrine,widely used in the treatment of respiratory problems, wasin the past extracted from Chinese species of Ephedra, but ithas now been largely replaced with synthetic preparations.Mormon tea is brewed from Ephedra stems in the south-western United States.
The best known species of Gnetum is a tropical tree, butmost species are vinelike. All species have broad leaves sim-ilar to those of angiosperms. One Gnetum species is culti-vated in Java for its tender shoots, which are cooked as avegetable.
Ginkgo (Ginkgophyta)
The fossil record indicates that members of the Ginkgofamily were once widely distributed, particularly in thenorthern hemisphere; today only one living species, themaidenhair tree (Ginkgo biloba), remains. The tree, whichsheds its leaves in the fall, was first encountered by Euro-peans in cultivation in Japan and China; it apparently nolonger exists in the wild (figure 37.14c). The commonname comes from the resemblance of its fan-shaped leavesto the leaflets of maidenhair ferns. Like the sperm of cy-cads, those of Ginkgo have flagella. The Ginkgo isdiecious, that is, the male and female reproductive struc-tures of Ginkgo are produced on separate trees. The fleshyouter coverings of the seeds of female Ginkgo plants exudethe foul smell of rancid butter caused by butyric andisobutyric acids. In the Orient, however, the seeds areconsidered a delicacy. In Western countries, because ofthe seed odor, male plants vegetatively propagated fromshoots are preferred for cultivation. Because it is resistantto air pollution, Ginkgo is commonly planted along citystreets.
Gymnosperms are mostly cone-bearing seed plants. Ingymnosperms, the ovules are not completely enclosedby sporophyte tissue at pollination.
748 Part X Plant Form and Function
FIGURE 37.14Three phyla ofgymnosperms.(a) An African cycad,Encephalartostransvenosus. (b) Welwitschiamirabilis, one of thethree genera ofgnetophytes. (c) Maidenhair tree,Ginkgo biloba, the onlyliving representative ofthe phylumGinkgophyta.
(a) (b) (c)

AngiospermsThe 250,000 known species of flower-ing plants are called angiosperms be-cause their ovules, unlike those ofgymnosperms, are enclosed withindiploid tissues at the time of pollina-tion. The name angiosperm derivesfrom the Greek words angeion, “ves-sel,” and sperma, “seed.” The “vessel”in this instance refers to the carpel,which is a modified leaf that encapsu-lates seeds. The carpel develops intothe fruit, a unique angiosperm feature.While some gymnosperms, includingyew, have fleshlike tissue around theirseeds, it is of a different origin and nota true fruit.
The origins of the angiospermspuzzled even Darwin (his “abominablemystery”). Recently, consensus hasbeen reached on the most basal, living angiosperm—Amborella trichopoda (figure 37.15). This has ended the de-bate between the supporters of magnolias and those ofwater lilies as the closest relatives of the original an-giosperm. Amborella, with small, cream-colored flowers, iseven more primitive than either the magnolias or waterlilies. This small shrub found only on the island of NewCaledonia in the South Pacific is the last remaining speciesof the earliest extant lineage of the angiosperms. About 135million years ago a close relative of Amborella developedfloral parts and branched off from the gymnosperms.While Amborella is not the original angiosperm, it is suffi-ciently close that much will be learned from studying its re-productive biology that will help us understand the earlyradiation of the angiosperms.
Flowering plants (phylum Anthophyta) exhibit an almostinfinite variety of shapes, sizes, and textures. They vary, forexample, from the huge Tasmanian Eucalyptus trees, whichhave nearly as much mass as the giant redwoods, to thetiniest duckweeds, which are less than 1 millimeter long. Inaddition to the typical flattened green leaves with whicheveryone is familiar, flowering plant leaves may be succu-lent, floating, submerged, cup-shaped, spinelike, scalelike,feathery, papery, hairy, or insect-trapping, and of almostany color. Some are so tiny one needs a microscope to ex-amine them, while others, such as those of the SeychellesIsland palm, can be up to 6 meters long. Their flowers varyfrom the simple blossoms of buttercups to the extraordi-narily complex flowers of some orchids, which may luretheir pollinators with drugs, forcibly attach bags of pollento their bodies, or dunk them in fluid they secrete. Theflowers may weigh less than 1 gram and remain functionalfor only a few minutes, or they can weigh up to 9 kilogramsand be functional for months. Plants of several families areparasitic or partially parasitic (for example, dodder, or
mistletoe) on other plants, or mycotrophic (deriving their nu-trients from fungi that form a mutualism with plant roots).Others, such as many orchids, are epiphytic (attached toother plants, with no roots in the ground, and not in anyway parasitic).
The Structure of Flowers
Flowers are considered to be modified stems bearing modi-fied leaves. Regardless of their size and shape, they all sharecertain features (see figure 37.16). Each flower originates asa primordium that develops into a bud at the end of a stalkcalled a pedicel. The pedicel expands slightly at the tipinto a base, the receptacle, to which the remaining flowerparts are attached. The other flower parts typically are at-tached in circles called whorls. The outermost whorl iscomposed of sepals. In most flowers there are three to fivesepals, which are green and somewhat leaflike; they oftenfunction in protecting the immature flower and in somespecies may drop off as the flower opens. The next whorlconsists of petals that are often colored and attract pollina-tors such as insects and birds. The petals, which commonlynumber three to five, may be separate, fused together, ormissing altogether in wind-pollinated flowers.
The third whorl consists of stamens, collectively calledthe androecium, a term derived from the Greek words an-dros, “male,” and oikos, “house.” Each stamen consists of apollen-bearing anther and a stalk called a filament, whichmay be missing in some flowers. The gynoecium, consist-ing of one or more carpels, is at the center of the flower.The term gynoecium derives from the Greek words gynos,which means “female,” and oikos, or “house.” The firstcarpel is believed to have been formed from a leaflike struc-ture with ovules along its margins. The edges of the bladethen rolled inward and fused together, forming a carpel.
Chapter 37 Evolutionary History of Plants 749
Non
vasc
ular
pla
nts
See
dles
s va
scul
ar p
lant
s
Gym
nosp
erm
s
Ang
iosp
erm
s
FIGURE 37.15A flowering plant. Amborella trichopoda.This plant is believed to be the closestliving relative to the original angiosperm.

Primitive flowers can have several to many separate carpels,but in most flowers, two to several carpels are fused to-gether. Such fusion can be seen in an orange sliced in half;each segment represents one carpel. A carpel has threemajor regions (figure 37.16). The ovary is the swollen base,which contains from one to hundreds of ovules; the ovarylater develops into a fruit. The tip of the pistil is called astigma. Most stigmas are sticky or feathery, causing pollengrains that land on them to adhere. Typically there is a neckor stalk called a style connecting the stigma and the ovary;in some flowers, the style may be very short or even miss-ing. Many flowers have nectar-secreting glands called nec-taries, often located toward the base of the ovary. Nectar is afluid containing sugars, amino acids, and other moleculesused to attract insects, birds, and other animals to flowers.
The Angiosperm Life Cycle
While a flower bud is developing, a single megasporemother cell in the ovule undergoes meiosis, producing fourmegaspores (figure 37.17). In most flowering plants, threeof the megaspores soon disappear while the nucleus of theremaining one divides mitotically, and the cell slowly ex-pands until it becomes many times its original size. Whilethis expansion is occurring, each of the daughter nuclei di-vide twice, resulting in eight haploid nuclei arranged in twogroups of four. At the same time, two layers of the ovule,the integuments, differentiate and become the seed coat ofa seed. The integuments, as they develop, leave a small gapor pore at one end—the micropyle (see figure 37.16). Onenucleus from each group of four migrates toward the cen-ter, where they function as polar nuclei. Polar nuclei mayfuse together, forming a single diploid nucleus, or they mayform a single cell with two haploid nuclei. Cell walls alsoform around the remaining nuclei. In the group closest tothe micropyle, one cell functions as the egg; the other twonuclei are called synergids. At the other end, the threecells are now called antipodals; they have no apparentfunction and eventually break down and disappear. Thelarge sac with eight nuclei in seven cells is called an em-bryo sac; it constitutes the female gametophyte. Althoughit is completely dependent on the sporophyte for nutrition,it is a multicellular, haploid individual.
While the female gametophyte is developing, a similarbut less complex process takes place in the anthers. Mostanthers have patches of tissue (usually four) that eventu-ally become chambers lined with nutritive cells. The tis-sue in each patch is composed of many diploid mi-crospore mother cells that undergo meiosis more or lesssimultaneously, each producing four microspores. Thefour microspores at first remain together as a quartet ortetrad, and the nucleus of each microspore divides once;in most species the microspores of each quartet then sep-arate. At the same time, a two-layered wall developsaround each microspore. As the anther matures, the wallbetween adjacent pairs of chambers breaks down, leavingtwo larger sacs. At this point, the binucleate microsporeshave become pollen grains. The outer pollen grain walllayer often becomes beautifully sculptured, and it con-tains chemicals that may react with others in a stigma tosignal whether or not development of the male gameto-phyte should proceed to completion. The pollen grainhas areas called apertures, through which a pollen tubemay later emerge.
Pollination is simply the mechanical transfer of pollenfrom its source (an anther) to a receptive area (the stigma ofa flowering plant). Most pollination takes place betweenflowers of different plants and is brought about by insects,wind, water, gravity, bats, and other animals. In as many asa quarter of all angiosperms, however, a pollen grain maybe deposited directly on the stigma of its own flower, andself-pollination occurs. Pollination may or may not be fol-lowed by fertilization, depending on the genetic compatibil-ity of the pollen grain and the flower on whose stigma ithas landed. (In some species, complex, genetically con-trolled mechanisms prevent self-fertilization to enhance ge-netic diversity in the progeny.) If the stigma is receptive,the pollen grain’s dense cytoplasm absorbs substances fromthe stigma and bulges through an aperture. The bulge de-velops into a pollen tube that responds to chemicals releasedby the embryo sac. It follows a diffusion gradient of thechemicals and grows down through the style and into themicropyle. The pollen tube usually takes several hours totwo days to reach the micropyle, but in a few instances, itmay take up to a year. One of the pollen grain’s two nuclei,the generative nucleus, lags behind. This nucleus divides, ei-
750 Part X Plant Form and Function
Stigma
Style
Ovule
Ovary
Pistil
Anther
FilamentStamen
Petal
Sepal
Receptacle
Pedicel
Megasporemother cell
Nucellus
Integuments
Micropyle
Stalk of ovule (funiculus)(a) (b)
FIGURE 37.16Diagram of an angiosperm flower.(a) The main structures of the flower arelabeled. (b) Details of an ovule. The ovaryas it matures will become a fruit; as theovule’s outer layers (integuments) mature,they will become a seed coat.

ther in the pollen grain or in the pollen tube, producingtwo sperm nuclei. Unlike sperm in mosses, ferns, and somegymnosperms, the sperm of flowering plants have no fla-gella. At this point, the pollen grain with its tube and spermhas become a mature male gametophyte.
As the pollen tube enters the embryo sac, it destroys asynergid in the process and then discharges its contents.Both sperm are functional, and an event called doublefertilization, unique to angiosperms, follows. One spermunites with the egg and forms a zygote, which developsinto an embryo sporophyte plant. The other sperm andthe two polar nuclei unite, forming a triploid primary en-dosperm nucleus. The primary endosperm nucleus beginsdividing rapidly and repeatedly, becoming triploid en-dosperm tissue that may soon consist of thousands ofcells. Endosperm tissue can become an extensive part ofthe seed in grasses such as corn (see figure 41.7). But inmost flowering plants, it provides nutrients for the embryothat develops from the zygote; in many species, such as
peas and beans, it disappears completely by the time theseed is mature. Following double fertilization, the integu-ments harden and become the seed coat of a seed. Thehaploid cells remaining in the embryo sac (antipodals, syn-ergid, tube nucleus) degenerate. There is some evidencefor a type of double fertilization in gymnosperms believedto be closely related to the angiosperms. Further studies ofthis and of fertilization in Amborella, the most basal, extantangiosperm, may provide clues to the evolution of thisdouble fertilization event.
Angiosperms are characterized by ovules that atpollination are enclosed within an ovary at the base of acarpel—a structure unique to the phylum; a fruitdevelops from the ovary. Evolutionary innovationsincluding flowers to attract pollinators, fruits to protectand aid in embryo dispersal, and double fertilizationproviding additional nutrients for the embryo all havecontributed to the widespread success of this phylum.
Chapter 37 Evolutionary History of Plants 751
FIGURE 37.17Life cycle of a typical angiosperm. As in pines, water is no longer required for fertilization. In most species of angiosperms, animalscarry pollen to the carpel. The outer wall of the carpel forms the fruit that entices animals to disperse seed.
Rhizome
n2n
Generative cell
8-nucleateembryo sac
(megagametophyte ) (n)Formation ofpollination tube (n)
MEIOSIS
MEIOSIS Tube nucleus
Tubenucleus
Sperm
Style
Sperm
Egg
Polarnuclei
Seed coat
Embryo (2n)
Seed (2n)
Endosperm (3n)
Youngsporophyte
(2n)
Adultsporophytewith flowers
(2n)
Anther
Ovary
Stigma
Anther (2n)
Microsporemother cells (2n)
Microspores (n) Pollen grains(microgametophytes) (n)
Megaspore (n)
Megasporemother cell (2n) Pollen tube
Ovule
DOUBLE FERTILIZATION
Germination
Egg
Mitosis
Mitosis

752 Part X Plant Form and Function
Chapter 37 Summary Questions Media Resources
37.1 Plants have multicellular haploid and diploid stages in their life cycles.
• Plants evolved from a multicellular, freshwater greenalgae 450 million years ago. The evolution of theirconducting tissues, cuticle, stomata, and seeds hasmade them progressively less dependent on externalwater for reproduction.
• All plants have a haplodiplontic life cycle in whichhaploid gametophytes alternate with diploidsporophytes.
1. Where did the most recentancestors of land plants live?What were they like? Whatadaptations were necessary forthe “move” onto land?2. What does it mean for a plantto alternate generations?Distinguish between sporophyteand gametophyte.
• Three phyla of plants lack well-developed vasculartissue, are the simplest in structure, and have beengrouped as bryophytes. This grouping does notreflect a common ancestry or close relationship.
• Sporophytes of mosses, liverworts, and hornworts areusually nutritionally dependent on the gametophytes,which are more conspicuous and photosynthetic.
3. Distinguish between malegametophytes and femalegametophytes. Which specifichaploid spores give rise to eachof these? 4. What reproductive limitationswould a moss tree (if oneexisted) face?
37.2 Nonvascular plants are relatively unspecialized, but successful in many terrestrial environments.
• Nine of the 12 plant phyla contain vascular plants,which have two kinds of well-defined conductingtissues: xylem, which is specialized to conduct waterand dissolved minerals; and phloem, which isspecialized to conduct the sugars produced byphotosynthesis and plant growth regulators.
• In ferns and other seedless vascular plants, thesporophyte generation is dominant. The fernsporophyte has vascular tissue and well-differentiatedroots, stems, and leaves.
5. In what ways are thegametophytes of seedless plantsdifferent from the gametophytesof seed plants?6. Which generation(s) of thefern are nutritionallyindependent?
37.3 Seedless vascular plants have well-developed conducting tissues in their sporophytes.
• Seeds were an important evolutionary advanceproviding for a dormant stage in development.
• In gymnosperms, ovules are exposed directly topollen at the time of pollination; in angiosperms,ovules are enclosed within an ovary, and a pollen tubegrows from the stigma to the ovule.
• The pollen of gymnosperms is usually disseminatedby wind. In most angiosperms the pollen istransported by insects and other animals. Bothflowers and fruits are found only in angiosperms andmay account for the extensive colonization ofterrestrial environments by the flowering plants.
7. What is a seed? Why is theseed a crucial adaptation toterrestrial life? 8. What is the principaldifference betweengymnosperms and angiosperms? 9. If all the offspring of a plantwere to develop in a small area,they would suffer from limitedresources. Compare dispersalstrategies in moss, pine, andangiosperms.
37.4 Seeds protect and aid in the dispersal of plant embryos.
www.mhhe.com/raven6e www.biocourse.com
• Life Cycles of Plants
• Student Research:Plant Biodiversity inNew Hampshire
• Book Review: A RumAffair by Sabbagh
• Book Review: TheOrchid Thief byOrlean
• Non-Vascular Plants
• Seedless VascularPlants
• Gymnosperms• Angiosperms
Copyright © 2022 FDOKUMEN



















