
Evidence for non-traditional activation of complement factor C3 during murine liver regeneration
15
Evidence for Nontraditional Activation of Complement Factor C3 during Murine Liver Regeneration Amelia Clark 1,6 , Alexander Weymann 1,6 , Eric Hartman 1 , Yumirle Turmelle 1 , Michael Carroll 4 , Joshua M. Thurman 5 , V. Michael Holers 5 , Dennis E. Hourcade 2 , and David A. Rudnick *,1,3 1Department of Pediatrics, Washington University School of Medicine, St. Louis, MO 63110 2Department of Medicine, Washington University School of Medicine, St. Louis, MO 63110 3Department of Developmental Biology, Washington University School of Medicine, St. Louis, MO 63110 4CBR Institute for Biomedical Research and Department of Pathology, Harvard Medical School, Boston, MA 02115 5Department of Medicine, University of Colorado Health Sciences Center, Denver CO 80262 Abstract Complement signaling has been implicated as important for normal hepatic regeneration. However, the specific mechanism by which complement is activated during liver regeneration remains undefined. To address this question, we investigated the hepatic regenerative response to partial hepatectomy in wildtype mice, C3-, C4-, and factor B-null mice, and C4-null mice treated with a factor B neutralizing antibody (mAb 1379). The results showed that following partial hepatectomy, C3-null mice exhibit reduced hepatic regeneration compared to wildtype mice as assessed by quantification of hepatic cyclin D1 expression and hepatocellular DNA synthesis and mitosis. In contrast, C4-null mice and factor B-null mice demonstrated normal liver regeneration. Moreover, animals in which all of the traditional upstream C3 activation pathways were disrupted, i.e. C4-null mice treated with mAb 1379, exhibited normal C3 activation and hepatocellular proliferation following partial hepatectomy. In order to define candidate non-traditional mechanisms of C3 activation during liver regeneration, plasmin and thrombin were investigated for their abilities to activate C3 in mouse plasma in vitro. The results showed that both proteases are capable of initiating C3 activation, and that plasmin can do so independent of the classical and alternative pathways. Conclusions—These results show that C3 is required for a normal hepatic regenerative response, but that disruption of the classical- or lectin-dependent pathways (C4-dependent), the alternative pathway (factor B-dependent), or all of these pathways does not impair the hepatic regenerative response, and indicate that non-traditional mechanisms by which C3 is activated during hepatic regeneration must exist. In vitro analysis raises the possibility that plasmin may contribute to non- traditional complement activation during liver regeneration in vivo. *Corresponding Author: David A. Rudnick, M.D.-Ph.D., Department of Pediatrics, Washington University School of Medicine, 660 S Euclid Ave, Box 8208, St. Louis, MO 63110, Phone:314-286-2832, Fax: 314-286-2892, [email protected]. 6 These authors contributed equally to this work. Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. NIH Public Access Author Manuscript Mol Immunol. Author manuscript; available in PMC 2009 June 1. Published in final edited form as: Mol Immunol. 2008 June ; 45(11): 3125–3132. doi:10.1016/j.molimm.2008.03.008. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
uni-heidelberg -
Category
Documents
-
view
1 -
download
0
Transcript of Evidence for non-traditional activation of complement factor C3 during murine liver regeneration

Evidence for Nontraditional Activation of Complement Factor C3during Murine Liver Regeneration
Amelia Clark1,6, Alexander Weymann1,6, Eric Hartman1, Yumirle Turmelle1, MichaelCarroll4, Joshua M. Thurman5, V. Michael Holers5, Dennis E. Hourcade2, and David A.Rudnick*,1,3
1Department of Pediatrics, Washington University School of Medicine, St. Louis, MO 63110
2Department of Medicine, Washington University School of Medicine, St. Louis, MO 63110
3Department of Developmental Biology, Washington University School of Medicine, St. Louis, MO 63110
4CBR Institute for Biomedical Research and Department of Pathology, Harvard Medical School, Boston, MA02115
5Department of Medicine, University of Colorado Health Sciences Center, Denver CO 80262
AbstractComplement signaling has been implicated as important for normal hepatic regeneration. However,the specific mechanism by which complement is activated during liver regeneration remainsundefined. To address this question, we investigated the hepatic regenerative response to partialhepatectomy in wildtype mice, C3-, C4-, and factor B-null mice, and C4-null mice treated with afactor B neutralizing antibody (mAb 1379). The results showed that following partial hepatectomy,C3-null mice exhibit reduced hepatic regeneration compared to wildtype mice as assessed byquantification of hepatic cyclin D1 expression and hepatocellular DNA synthesis and mitosis. Incontrast, C4-null mice and factor B-null mice demonstrated normal liver regeneration. Moreover,animals in which all of the traditional upstream C3 activation pathways were disrupted, i.e. C4-nullmice treated with mAb 1379, exhibited normal C3 activation and hepatocellular proliferationfollowing partial hepatectomy. In order to define candidate non-traditional mechanisms of C3activation during liver regeneration, plasmin and thrombin were investigated for their abilities toactivate C3 in mouse plasma in vitro. The results showed that both proteases are capable of initiatingC3 activation, and that plasmin can do so independent of the classical and alternative pathways.
Conclusions—These results show that C3 is required for a normal hepatic regenerative response,but that disruption of the classical- or lectin-dependent pathways (C4-dependent), the alternativepathway (factor B-dependent), or all of these pathways does not impair the hepatic regenerativeresponse, and indicate that non-traditional mechanisms by which C3 is activated during hepaticregeneration must exist. In vitro analysis raises the possibility that plasmin may contribute to non-traditional complement activation during liver regeneration in vivo.
*Corresponding Author: David A. Rudnick, M.D.-Ph.D., Department of Pediatrics, Washington University School of Medicine, 660 SEuclid Ave, Box 8208, St. Louis, MO 63110, Phone:314-286-2832, Fax: 314-286-2892, [email protected] authors contributed equally to this work.Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptMol Immunol. Author manuscript; available in PMC 2009 June 1.
Published in final edited form as:Mol Immunol. 2008 June ; 45(11): 3125–3132. doi:10.1016/j.molimm.2008.03.008.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

KeywordsPartial Hepatectomy; Alternative Pathway; Classical Pathway; Factor B; C4
IntroductionThe regenerative ability of the liver is important for recovery from hepatic injury and disease.Analyses using the rodent partial hepatectomy model (Higgins and Anderson 1931) show thatfollowing partial hepatectomy, the proliferation of normally quiescent hepatocytes is rapidlyinduced, proceeds until the original liver mass is restored, and is then precisely terminated(Fausto 2000; Diehl and Rai 1999; Michalopoulos and DeFrances 1997). The molecularmechanisms that control this carefully orchestrated response include activation of TNFα-IL6signaling (Akerman et al. 1992; Yamada et al. 1997; Cressman et al. 1996), generation ofmitochondrial reactive oxygen species (Lee et al. 1999) and prostaglandins (Rudnick et al.2001), and activation of stress- and mitogen-activated-protein kinase cascades (Talarmin et al.1999). These events promote activation of NFκB, STAT3, AP1 and other transcription factors(Cressman et al. 1994; Cressman et al. 1995; Taub 1996), which direct an immediate-earlygene expression program (Haber et al. 1993) culminating in growth factor-dependent,hepatocellular re-entry into and progression through the cell cycle, and restoration of normalhepatic mass. Despite the knowledge gained from experimental analyses of liver regeneration,an integrated understanding of the specific mechanisms responsible for initiation, propagation,and termination of the hepatic regenerative response remains elusive.
The complement pathway, well known for its role in host defense, as an effector arm of innateimmunity, and as a regulatory element of acquired immunity (Walport 2001a, 2001b;Volanakis 1998b; Lambris et al. 2008), has recently been implicated as important in liverregeneration. This link was established by studies demonstrating that liver regeneration isimpaired in complement factor C3-null mice, and that such impairment is rescued by exogenousC3 or C3a supplementation (Strey et al. 2003; Markiewski et al. 2004). Similarly, complementfactor C5-deficient mice exhibit abnormal hepatic regeneration which can be rescued bysupplementation with exogenous C5 or C5a (Mastellos et al. 2001; Strey et al. 2003). Theseanalyses also show that pharmacological C5 neutralization and C5a receptor blockade disruptthe hepatic regenerative response and that disruption of complement cascade activation isassociated with suppression of TNFα-IL6, NFκB, and STAT3 activation (Strey et al. 2003).Together, these data provide convincing evidence that activation of complement factor C3 andsubsequent C5-dependent downstream signaling events are required for normal liverregeneration.
Although the important role of C3 and C5 signaling during liver regeneration has beenestablished by the studies described above, the mechanisms by which such signaling isactivated have not yet been defined. Complement is traditionally activated by the classical,lectin-dependent, or alternative pathway (reviewed in Lambris et al. 2008; Volanakis 1998b).The classical pathway is initiated by antibody:antigen complexes, the lectin pathway bymannan-binding lectin:carbohydrate complexes, and the alternative pathway by a variety ofmicrobial surfaces. The alternative pathway is also activated by and can amplify the signalfrom the classical and lectin pathways. The classical and lectin-dependent pathways requirecomplement factors C2 and C4 while the alternative pathway depends on complement factorsB and D. Each activation pathway results in the assembly of the C3 convertases, which are thecentral enzymes of the complement cascade, and which catalyze the proteolytic cleavage ofC3 to form C3a and C3b. C3b is an opsonin that mediates target clearance, and it can alsoassociate with the C3 convertase to form a C5 convertase. The C5 convertase cleaves C5,forming C5a and C5b. C5b initiates the assembly of the membrane attack complex. C3a and
Clark et al. Page 2
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

C5a are anaphylatoxins which trigger various cellular responses via the C3a or C5a receptor,respectively.
In the studies reported here, we describe the results of our analyses of hepatic regeneration inmice disrupted for classical and lectin dependent complement activation pathways (C4-nullmice), for alternative pathway activation (factor B null mice), or for both pathways (C4-nullmice treated with a factor B neutralizing antibody, mAb 1379; Leinhase et al. 2007; Thurmanet al. 2005, 2006; Taube et al. 2006).
Materials and MethodsAnimal Husbandry and Surgery
Wildtype C57Bl/6J mice were obtained from the Jackson Laboratory (Bar Harbor, ME).Complement factor 3-, factor B-, and factor C4-null mice were maintained on C57Bl/6Jbackgrounds as previously described (Circolo et al. 1999; Matsumoto et al. 1997; Wessels etal. 1995). Absence of circulating C3, C4, or factor B in each of the respective null mice wasverified by protein immunoblot performed on plasma recovered from wildtype, C3-null, C4-null, and factor B-null mice (Figure 1). One mg of the inhibitory monoclonal anti-factor Bantibody mAb 1379 was administered by intraperitoneal injection once daily beginning 12hours prior to partial hepatectomy. This is within the dosing range reported to neutralize factorB activity in models of murine inflammation (Leinhase et al. 2007; Taube et al. 2006; Thurmanet al. 2005, 2006). Suppression of the complement alternative pathway in mice treated withmAb 1379 was verified using a modification of the alternative pathway zymosan assay(Thurman et al. 2005; Foley et al. 1993) as described below.
Two to three month old male mice, maintained on 12 h dark-light cycles and standard mousechow, were subjected to partial hepatectomy, allowed to recover, and then sacrificed for plasmaand tissue harvest as previously described (Rudnick et al. 2001; Shteyer et al. 2004; Liao et al.2004). Briefly, mice were sedated with inhaled Isoflurane (VEDCO, Inc., St. Joseph, MO)delivered via an anesthesia cart, then subjected to mid-ventral laparotomy with exposure of theleft and median hepatic lobes, which was followed by sequential ligation and resection of themedian and left lobes and closure of the peritoneal and skin wounds. Hepatectomized animalswere allowed to recover until the time of sacrifice by inhaled carbon dioxide and harvest ofplasma and liver tissue. Three to six animals were examined at each time point and for eachgenotype. All experiments were approved by the Animal Studies Committee of WashingtonUniversity and conducted in accordance with institutional guidelines and the criteria outlinedin the “Guide for Care and Use of Laboratory Animals” (NIH publication 86-23).
Alternative Pathway Zymosan AssayAlternative pathway integrity was measured using a modification of the zymosan assay(Thurman et al. 2005; Foley et al. 1993). Activated zymosan particles (CompTech, Tyler, TX,1×106 per reaction) were added to 100 µl veronal buffered saline containing either 2 mMMgCl2 plus 10 mM EGTA (experimental sample), or 10 mM EDTA (negative control). Mouseplasma (10 µl) was added and the reactions were incubated at 37 degrees for 30 min. Particleswere then washed twice in PBS/1% BSA, resuspended in 100 µl PBS/BSA and treated withFITC-conjugated goat anti-mouse C3 antibody (MP Biomedicals, Solon, OH; 1 µl of 1:10dilution) at 4 degrees for 20 min. Samples were washed once and surface C3 was analyzed byFACS with a FACScaliber instrument (Becton Dickinson). A common size gate was used forall experiments. Two Mg-EGTA control reactions were performed, one without plasma andone with an unrelated FITC-conjugated antibody. Alternative pathway activity was calculatedas the (Mean Particle Fluorescence of the Mg-EGTA reaction) - (Mean Particle Fluorescenceof the EDTA reaction).
Clark et al. Page 3
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Histology and ImmunohistochemistryLiver histology and hepatocellular bromodeoxyuridine (BrdU) incorporation were assessed aspreviously described (Rudnick et al. 2001; Shteyer et al. 2004; Liao et al. 2004). Briefly,animals were injected with 100 mg/kg BrdU 1 hour prior to sacrifice. After harvesting, a portionof the right hepatic lobe was fixed in formalin, paraffin-embedded, and stained either withhematoxylin and eosin or for nuclear BrdU incorporation. The frequency of nuclear BrdUlabeling was determined by two different investigators and by examination of at least threerandom 400x fields and at least 300 cells and nuclei in each tissue section.
Protein Expression AnalysisWhole cell lysates were made from snap frozen liver and their protein concentration determinedas previously described (Rudnick et al. 2001). Twenty-five µg aliquots of protein lysate or 1µl aliquots of plasma were subjected to SDS-PAGE, followed by electrophoretic transfer tonitrocellulose. Filters were probed with primary antibody (Cyclin D1, Upstate, Lake Placid,NY; glyceraldehyde phosphate dehydrogenase, Chemicon International, Temecula, CA;complement factor C4 and complement factor B, CompTech; complement factor C3, MPBiomedicals) followed by a horseradish peroxidase-conjugated secondary antibody, and thendeveloped using the ECL system (Amersham, Piscataway, NJ). Densitometric analysis wasperformed with Scion Image data analysis software (Scion Corporation, Frederick, MD).
In Vivo and In Vitro Analysis of C3 ActivationActivation of C3 during liver regeneration was evaluated by protein immunoblot determinationof the activated ~40 kDa C3α proteolytic fragment in plasma as previously described (Mastelloset al. 2004; Miwa et al. 2007). In vitro activation of C3 was determined using this samemethodology on mouse plasma incubated with thrombin (Sigma Chemical Company, St.Louis; 18 units to 10 µl plasma) or plasmin (Sigma Chemical Company; 10 µg, ≥0.03 units,added to 10 µl of plasma), at 25° C for 20 minutes.
Statistical AnalysisData were analyzed using SigmaPlot and SigmaStat software (SPSS, Chicago, IL). UnpairedStudent’s t-test for pair-wise comparisons and ANOVA for multiple groups (with the Holm-Sidak method for post-hoc comparison) were used to determine statistical significance fordifferences in hepatocellular BrdU incorporation, mitotic body frequency, and proteinexpression. Kaplan-Meier survival analysis was performed using the LogRank statistic tocompare outcomes between groups. Data are reported as mean ± standard error.
ResultsLiver Regeneration is Delayed in C3-Null Mice
Determination of hepatocellular BrdU incorporation following partial hepatectomy in wildtypeand C3-null mice showed that hepatocellular proliferation was inhibited and the time to peakproliferation delayed in C3-null mice (Figure 2A, *p<0.05). Similarly, hepatocellular mitoticbody frequency was reduced in C3-null mice at 48 hours after surgery (Figure 2B, *p<0.02),which is the timepoint corresponding to peak hepatocellular mitosis in wildtype mice (Liao etal. 2004). Finally, induction of hepatic cyclin D1 protein expression, which regulateshepatocellular G1-S phase cell cycle progression during normal regeneration, was suppressedin C3-null mice (Figure 2C: Immunoblot representative of n=6 animals for each timepointshown on left; densitometric analysis of individual data from all animals summarized on right;*p<0.03). Together these data indicate that liver regeneration is delayed by genetic disruptionof complement factor C3 expression. This delay was not associated with histological evidence
Clark et al. Page 4
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

of increased liver tissue injury (Figure 2B) or with increased animal mortality from 0–72 hoursafter surgery.
Liver Regeneration Occurs Normally in C4-Null MiceIn order to determine whether either the complement classical or lectin activation pathwaysare required for normal liver regeneration, the hepatic regenerative response was evaluated incomplement factor C4-null mice. C4 is a necessary component of the classical and lectin-dependent pathway C3- and C5-convertases (Volanakis 1998a). Analysis of the regenerativeresponse to partial hepatectomy in C4-null mice showed that hepatocellular BrdUincorporation, mitotic body frequency, and cyclin D expression were not inhibited or delayedby genetic disruption of C4 expression (Figure 3A–C; Immunoblot shown in Figure 3C isrepresentative of n=6 animals for each timepoint shown on left; densitometric analysis ofindividual data from all animals summarized on right). In addition, there was no evidence forincreased liver tissue injury (Figure 3B) or increased animal mortality in C4-null micesubjected to partial hepatectomy. These data indicate that neither the classical nor the lectin-dependent pathways of complement cascade activation are absolutely required for normalhepatic regeneration.
Liver Regeneration Occurs Normally in Factor B-Null MiceNext, in order to determine whether the complement alternative activation pathway is requiredduring liver regeneration, the hepatic regenerative response to partial hepatectomy wasquantified in complement factor B-null mice. A major fragment of factor B is a component ofthe alternative pathway C3- and C5-convertases (Volanakis 1998a). Moreover, the alternativepathway has been shown, in most instances, to play an obligate role in generating complementactivation products (Thurman and Holers 2006). Analysis of the regenerative response topartial hepatectomy in factor B-null mice showed that hepatocellular BrdU incorporation,cyclin D expression, and mitotic body frequency were not inhibited or delayed by geneticdisruption of factor B expression (Figure 3A–C; Immunoblot shown in Figure 3C isrepresentative of n=6 animals for each timepoint shown on left; densitometric analysis ofindividual data from all animals summarized on right). As was true for C3- and C4-null mice,there was no evidence of increased liver tissue injury or increased mortality in factor B nullmice subjected to partial hepatectomy. These data show that the alternative pathway ofcomplement cascade activation is not required for normal hepatic regeneration.
Liver Regeneration and C3 Activation Occur Normally in C4-Null Mice Treated with mAb 1379Factor B Neutralizing Antibody
The data described above indicate that neither the classical, lectin-dependent, nor alternativepathways of complement activation are absolutely required for normal liver regeneration.However, these analyses do not distinguish between the possibilities that any of thesetraditional upstream pathways may be sufficient to activate C3 and promote normal liverregeneration or, alternatively, that non-traditional mechanisms activate complement signalingduring liver regeneration. To investigate these considerations, the hepatic regenerativeresponse was evaluated in C4-null mice treated with a well characterized anti-factor Bneutralizing monoclonal antibody (mAb 1379, (Leinhase et al. 2007; Taube et al. 2006;Thurman et al. 2005, 2006)). In these animals, the classical and lectin-dependent pathways areblocked by genetic disruption of C4 expression (Figure 1) while the alternative pathway isentirely suppressed by the neutralizing antibody (Figure 4B). The results showed thathepatocellular proliferation 36 hours after partial hepatectomy, the time corresponding to peakproliferation in wildtype mice, is comparable in wildtype untreated-, wildtype mAb 1379-treated-, and C4 null mAb 1379-treated mice (Figure 4A), indicating that disruption of all ofthe traditional upstream activation pathways of complement signaling does not prevent normal
Clark et al. Page 5
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

liver regeneration. Taken together with the observation that complement factor C3 is requiredfor normal liver regeneration (Mastellos et al. 2001;Strey et al. 2003;Markiewski et al. 2004),these data raise the possibility that C3 is activated independently of the classical-, lectin-dependent-, and alternative-pathways during the hepatic regenerative response. To test thispossibility, C3 activation during liver regeneration was quantified in wildtype and mAb1379-treated C4 null mice subjected to partial hepatectomy by protein immunoblot analysis ofmouse plasma for the proteolytic fragment corresponding to activated C3α (Miwa et al.2007). The results showed comparable C3 activation in each case (Figure 4C; Immunoblotshown on left is representative of n=3 animals for each timepoint and treatment group;densitometric analysis of individual data from all animals summarized on right), indicatingthat C3 must be activated by non-traditional mechanisms during liver regeneration.
In Vitro Proteolytic Activation of C3Based on the data described above and a recent report demonstrating non-traditional (C3-independent) proteolytic activation of complement factor C5 by thrombin (Huber-Lang et al.2006), the effect of thrombin on in vitro C3 activation was assessed by incubating purifiedprotease with wildtype mouse plasma. The results showed thrombin-dependent appearance ofactivated C3α (Figure 5, lanes a–c; The immunoblot shown is representative of an experimentperformed in duplicate). The protease plasmin, which is an important regulator of liverregeneration (Drixler et al. 2003), was also investigated, and similar results were seen (Figure5, lanes a–b,d). Further analysis showed that plasmin dependent C3 activation was comparablein wildtype, factor B null-, C4 null-, and mAb 1379 treated C4 null-mice (Figure 5, lanes d,h,l,p)whereas thrombin dependent activation was reduced in plasma from factor B null- and mAb1379 treated C4 null-mice (Figure 5, lanes c,g,k,o). These data show that under theseexperimental conditions plasmin can promote C3 activation independent of the alternative,classical, and lectin dependent pathways in vitro. This observation raises the possibility thatplasmin may contribute to non-traditional complement activation during liver regeneration invivo.
Discussion and ConclusionsThe complement system is well known for its immunological role in defense against invadingpathogens, and has also been implicated in the pathogenesis of autoimmune and other diseasesin which tissue injury results from dysregulated complement-dependent release ofinflammatory mediators (reviewed in Volanakis 1998b; Lambris et al. 2008). More recentevidence indicates that complement may also be an important regulator of variousdevelopmental processes including bone and cartilage growth, reproduction, hematopoiesis,vascular development, and tissue and organ regeneration (Mastellos and Lambris 2002).Complement activation during host defense traditionally occurs through one of three pathways:the classical-, lectin-dependent, or alternative pathways (reviewed in Walport 2001a, 2001b;Volanakis 1998b). The classical and lectin-dependent pathways require complement factorsC2 and C4 while the alternative pathway depends on complement factors B and D. Theregulation of complement activation during various developmental processes is not as wellestablished. For example, although complement signaling is known to be required forregeneration of the newt limb (Kimura et al. 2003; Rio-Tsonis et al. 1998) and the mammalianliver (Mastellos et al. 2001; Strey et al. 2003; Markiewski et al. 2004), the events that regulatecomplement cascade activation in these settings have not yet been defined. Therefore, in theanalyses reported here, we verified the role of C3 and assessed the functional requirement foreach of the known upstream complement activation pathways during liver regeneration.
Our data show that C3-null mice exhibit impaired hepatic regeneration characterized bysignificantly delay in hepatocellular proliferation, mitotic progression, and cyclin D expression
Clark et al. Page 6
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

following partial hepatectomy (Figure 1). Although these observations are consistent with theprevious report showing that C3 is required for normal liver regeneration (Strey et al. 2003),the phenotype in C3-null mice reported here is not as severe, with decreased tissue injury andmortality, compared to that described in the prior study. The basis for this difference is notknown, but is suspected to reflect technical differences in application of the surgical model:the partial hepatectomy methodology used here has been modified from that reported byHiggins and Anderson (Higgins and Anderson 1931) to incorporate use of controlled vaporizer-based delivery of inhaled Isoflurane anesthesia, a change which has resulted in diminishedmorbidity and mortality in this model system (personal observations of Clark, Weymann, andRudnick). Nevertheless, our data support the conclusion that complement factor C3 is requiredfor normal liver regeneration.
The data described here also show that disruption of specific factors required for the classicaland lectin activation pathways (C4 null mice) or for the alternative pathway (factor B null mice)does not impair hepatocellular proliferation following partial hepatectomy. With respect to C4disruption, two complement activation mechanisms involving classical or lectin pathwaycomponents that bypass the standard C4 requirement have been reported: Antibody-antigencomplexes, which normally activate the classical pathway, and mannose-binding-lectin:lectincomplexes, which normally activate the lectin pathway are both capable of activating C3 inthe absence of C4 (Atkinson and Frank 2006; Selander et al. 2006). However, in each case thealternative pathway plays a critical role. In contrast, our data demonstrate that liver regenerationand C3 activation are also normal in C4 null mice treated with the factor B-neutralizingantibody (Figure 4).
Taken together, the data reported here indicate that despite the established requirement forcomplement factors C3 and C5 during the hepatic regenerative response (Mastellos et al.2001; Strey et al. 2003; Markiewski et al. 2004), none of the traditional upstream complementactivation pathways are absolutely necessary for normal liver regeneration. The moststraightforward interpretation of these data is that proteolytic C3 activation during liverregeneration may occur via non-traditional mechanisms. This conclusion is particularlyintriguing in light of a recently published report demonstrating non-traditional (C3-independent) activation of C5 by thrombin both in vivo in a murine model of lung injury andin vitro (Huber-Lang et al. 2006), and raise the possibility that thrombin or other factors mayalso promote C3 activation during liver regeneration. Based on such analyses and recentlyreported observations highlighting the existence of crosstalk between the complement andcoagulation proteolytic cascades (Markiewski et al. 2007), the effects of thrombin and plasminon in vitro C3 activation were determined. The results showed that both of these proteases canpromote the generation of activated C3α, and furthermore that under the experimentalconditions employed here plasmin can promote such activation in the absence of factor B(Figure 5). Together, our data raise the possibility that non-traditional mechanisms involvingplasmin or other proteases may promote complement factor C3 activation during normal liverregeneration in vivo. Future studies should investigate the extent to which such mechanismscontribute to complement function in liver regeneration and other settings.
AcknowledgementsWe are grateful to Drs. Harvey Colten and Hector Molina for providing the C3-null and factor B-null mice, to LynnMitchell for technical assistance with the zymosan assay, and to Dr. Xiaobo Wu for assistance with the complementfactor C3 activation immunoblot analysis. The studies reported here were supported by grants to DAR from NIH(DK068219) and March of Dimes (Basil O’Connor Award), to DH from NIH (AI05143), and by the Digestive DiseaseResearch Core Center (NIH grant # P30 DK52574). AW was supported in part by a post-doctoral fellowship awardfrom the American Liver Foundation. YPT was supported in part by Institutional Training Grant T32-HD07409. JMTis supported by NIH DK064790.
Clark et al. Page 7
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Reference List1. Akerman P, Cote P, Yang SQ, McClain C, Nelson S, Bagby GJ, Diehl AM. Antibodies to tumor
necrosis factor-alpha inhibit liver regeneration after partial hepatectomy. American Journal ofPhysiology 1992;263:G579–G585. [PubMed: 1415718]
2. Atkinson JP, Frank MM. Bypassing complement: evolutionary lessons and future implications.J.Clin.Invest 2006;116:1215–1218. [PubMed: 16670764]
3. Circolo A, Garnier G, Fukuda W, Wang X, Hidvegi T, Szalai AJ, Briles DE, Volanakis JE, WetselRA, Colten HR. Genetic disruption of the murine complement C3 promoter region generates deficientmice with extrahepatic expression of C3 mRNA. Immunopharmacology 1999;42:135–149. [PubMed:10408374]
4. Cressman DE, Diamond RH, Taub R. Rapid activation of the STAT3 transcription complex in liverregeneration. Hepatology 1995;21:1443–1449. [PubMed: 7737651]
5. Cressman DE, Greenbaum LE, DeAngelis RA, Ciliberto G, Furth EE, Poli V, Taub R. Liver failureand defective hepatocyte regeneration in interleukin-6-deficient mice. Science 1996;274:1379–1383.[PubMed: 8910279]
6. Cressman DE, Greenbaum LE, Haber BA, Taub R. Rapid activation of post-hepatectomy factor/nuclearfactor kappa B in hepatocytes, a primary response in the regenerating liver. J.Biol.Chem1994;269:30429–30435. [PubMed: 7982957]
7. Diehl, AM.; Rai, R. Liver Regeneration. In: Schiff, R.; Sorrell, MF.; Maddrey, WC., editors. Schiff'sDiseases of the Liver. Philadelphia: Lippincott-Raven; 1999. p. 39-54.
8. Drixler TA, Vogten JM, Gebbink MF, Carmeliet P, Voest EE, Borel R, I. Plasminogen mediates liverregeneration and angiogenesis after experimental partial hepatectomy. British Journal of Surgery2003;90:1384–1390. [PubMed: 14598419]
9. Fausto N. Liver regeneration. J Hepatol 2000;32:19–31. [PubMed: 10728791]10. Foley S, Li B, Dehoff M, Molina H, Holers VM. Mouse Crry/p65 is a regulator of the alternative
pathway of complement activation. European Journal of Immunology 1993;23:1381–1384.[PubMed: 8500531]
11. Haber BA, Mohn KL, Diamond RH, Taub R. Induction patterns of 70 genes during nine days afterhepatectomy define the temporal course of liver regeneration. J.Clin.Invest 1993;91:1319–1326.[PubMed: 8473485]
12. Higgins GM, Anderson RM. Experimental Pathology of the Liver. 1. Restoration of the liver of thewhite rat following partial surgical removal. Arch.Pathol 1931;12:186–202.
13. Huber-Lang M, Sarma JV, Zetoune FS, Rittirsch D, Neff TA, McGuire SR, Lambris JD, Warner RL,Flierl MA, Hoesel LM, Gebhard F, Younger JG, Drouin SM, Wetsel RA, Ward PA. Generation ofC5a in the absence of C3: a new complement activation pathway. Nat.Med 2006;12:682–687.[PubMed: 16715088]
14. Kimura Y, Madhavan M, Call MK, Santiago W, Tsonis PA, Lambris JD, Rio-Tsonis K. Expressionof complement 3 and complement 5 in newt limb and lens regeneration. J.Immunol 2003;170:2331–2339. [PubMed: 12594255]
15. Lambris JD, Ricklin D, Geisbrecht BV. Complement evasion by human pathogens.Nat.Rev.Microbiol 2008;6:132–142. [PubMed: 18197169]
16. Lee FY, Li Y, Zhu H, Yang S, Lin HZ, Trush M, Diehl AM. Tumor necrosis factor increasesmitochondrial oxidant production and induces expression of uncoupling protein-2 in the regeneratingmice [correction of rat] liver. Hepatology 1999;29:677–687. [PubMed: 10051468]
17. Leinhase I, Rozanski M, Harhausen D, Thurman JM, Schmidt OI, Hossini AM, Taha ME, RittirschD, Ward PA, Holers VM, Ertel W, Stahel PF. Inhibition of the alternative complement activationpathway in traumatic brain injury by a monoclonal anti-factor B antibody: a randomized placebo-controlled study in mice. J.Neuroinflammation 2007;4:13. [PubMed: 17474994]
18. Liao Y, Shikapwashya ON, Shteyer E, Dieckgraefe BK, Hruz PW, Rudnick DA. Delayedhepatocellular mitotic progression and impaired liver regeneration in early growth response-1-deficient mice. J.Biol.Chem 2004;279:43107–43116. [PubMed: 15265859]
Clark et al. Page 8
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

19. Markiewski MM, Mastellos D, Tudoran R, DeAngelis RA, Strey CW, Franchini S, Wetsel RA, ErdeiA, Lambris JD. C3a and C3b activation products of the third component of complement (C3) arecritical for normal liver recovery after toxic injury. Journal of Immunology 2004;173:747–754.
20. Markiewski MM, Nilsson B, Ekdahl KN, Mollnes TE, Lambris JD. Complement and coagulation:strangers or partners in crime? Trends Immunol 2007;28:184–192. [PubMed: 17336159]
21. Mastellos D, Lambris JD. Complement: more than a 'guard' against invading pathogens? Trends inImmunology 2002;23:485–491. [PubMed: 12297420][Review] [36 refs]
22. Mastellos D, Papadimitriou JC, Franchini S, Tsonis PA, Lambris JD. A novel role of complement:mice deficient in the fifth component of complement (C5) exhibit impaired liver regeneration.J.Immunol 2001;166:2479–2486. [PubMed: 11160308]
23. Mastellos D, Prechl J, Laszlo G, Papp K, Olah E, Argyropoulos E, Franchini S, Tudoran R,Markiewski M, Lambris JD, Erdei A. Novel monoclonal antibodies against mouse C3 interferingwith complement activation: description of fine specificity and applications to variousimmunoassays. Mol.Immunol 2004;40:1213–1221. [PubMed: 15104126]
24. Matsumoto M, Fukuda W, Circolo A, Goellner J, Strauss-Schoenberger J, Wang X, Fujita S, HidvegiT, Chaplin DD, Colten HR. Abrogation of the alternative complement pathway by targeted deletionof murine factor B. Proceedings of the National Academy of Sciences of the United States of America1997;94:8720–8725. [PubMed: 9238044]
25. Michalopoulos GK, DeFrances MC. Liver regeneration. Science 1997;276:60–66. [PubMed:9082986][Review]
26. Miwa T, Zhou L, Tudoran R, Lambris JD, Madaio MP, Nangaku M, Molina H, Song WC. DAF/Crrydouble deficiency in mice exacerbates nephrotoxic serum-induced proteinuria despite markedlyreduced systemic complement activity. Mol.Immunol 2007;44:139–146. [PubMed: 16887189]
27. Rio-Tsonis K, Tsonis PA, Zarkadis IK, Tsagas AG, Lambris JD. Expression of the third componentof complement, C3, in regenerating limb blastema cells of urodeles. Journal of Immunology1998;161:6819–6824.
28. Rudnick DA, Perlmutter DH, Muglia LJ. Prostaglandins are required for CREB activation and cellularproliferation during liver regeneration. Proceedings of the National Academy of Sciences of theUnited States of America 2001;98:8885–8890. [PubMed: 11447268]
29. Selander B, Martensson U, Weintraub A, Holmstrom E, Matsushita M, Thiel S, Jensenius JC,Truedsson L, Sjoholm AG. Mannan-binding lectin activates C3 and the alternative complementpathway without involvement of C2. J.Clin.Invest 2006;116:1425–1434. [PubMed: 16670774]
30. Shteyer E, Liao Y, Muglia LJ, Hruz PW, Rudnick DA. Disruption of hepatic adipogenesis is associatedwith impaired liver regeneration in mice. Hepatology 2004;40:1322–1332. [PubMed: 15565660]
31. Strey CW, Markiewski M, Mastellos D, Tudoran R, Spruce LA, Greenbaum LE, Lambris JD. Theproinflammatory mediators C3a and C5a are essential for liver regeneration. Journal of ExperimentalMedicine 2003;198:913–923. [PubMed: 12975457]
32. Talarmin H, Rescan C, Cariou S, Glaise D, Zanninelli G, Bilodeau M, Loyer P, Guguen-GuillouzoC, Baffet G. The mitogen-activated protein kinase kinase/extracellular signal-regulated kinasecascade activation is a key signalling pathway involved in the regulation of G(1) phase progressionin proliferating hepatocytes. Molecular & Cellular Biology 1999;19:6003–6011. [PubMed:10454547]
33. Taub R. LIVER REGENERATION .4. Transcriptional control of liver regeneration. FASEB Journal1996;10:413–427. [PubMed: 8647340]
34. Taube C, Thurman JM, Takeda K, Joetham A, Miyahara N, Carroll MC, Dakhama A, Giclas PC,Holers VM, Gelfand EW. Factor B of the alternative complement pathway regulates developmentof airway hyperresponsiveness and inflammation. Proc.Natl.Acad.Sci.U.S.A 2006;103:8084–8089.[PubMed: 16702544]
35. Thurman JM, Holers VM. The central role of the alternative complement pathway in human disease.J.Immunol 2006;176:1305–1310. [PubMed: 16424154]
36. Thurman JM, Kraus DM, Girardi G, Hourcade D, Kang HJ, Royer PA, Mitchell LM, Giclas PC,Salmon J, Gilkeson G, Holers VM. A novel inhibitor of the alternative complement pathway preventsantiphospholipid antibody-induced pregnancy loss in mice. Mol.Immunol 2005;42:87–97. [PubMed:15488947]
Clark et al. Page 9
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

37. Thurman JM, Royer PA, Ljubanovic D, Dursun B, Lenderink AM, Edelstein CL, Holers VM.Treatment with an inhibitory monoclonal antibody to mouse factor B protects mice from inductionof apoptosis and renal ischemia/reperfusion injury. J.Am.Soc.Nephrol 2006;17:707–715. [PubMed:16467447]
38. Volanakis, JE. The Human Complement System in Health and Disease. Volanakis, JE.; Frank, MM.,editors. New York: Marcel Dekker, Inc; 1998a. p. 149-166.
39. Volanakis, JE. Overview of the Complement System. In: Volanakis, JE.; Frank, MM., editors. TheHuman Complement System in Health and Disease. New York: Marcel Dekker, Inc; 1998b. p. 9-32.
40. Walport MJ. Complement. First of two parts. N.Engl.J.Med 2001a;344:1058–1066. [PubMed:11287977]
41. Walport MJ. Complement. Second of two parts. N.Engl.J.Med 2001b;344:1140–1144. [PubMed:11297706]
42. Wessels MR, Butko P, Ma M, Warren HB, Lage AL, Carroll MC. Studies of group B streptococcalinfection in mice deficient in complement component C3 or C4 demonstrate an essential role forcomplement in both innate and acquired immunity. Proc.Natl.Acad.Sci.U.S.A 1995;92:11490–11494. [PubMed: 8524789]
43. Yamada Y, Kirillova I, Peschon JJ, Fausto N. Initiation of liver growth by tumor necrosis factor:deficient liver regeneration in mice lacking type I tumor necrosis factor receptor. Proceedings of theNational Academy of Sciences of the United States of America 1997;94:1441–1446. [PubMed:9037072]
Clark et al. Page 10
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
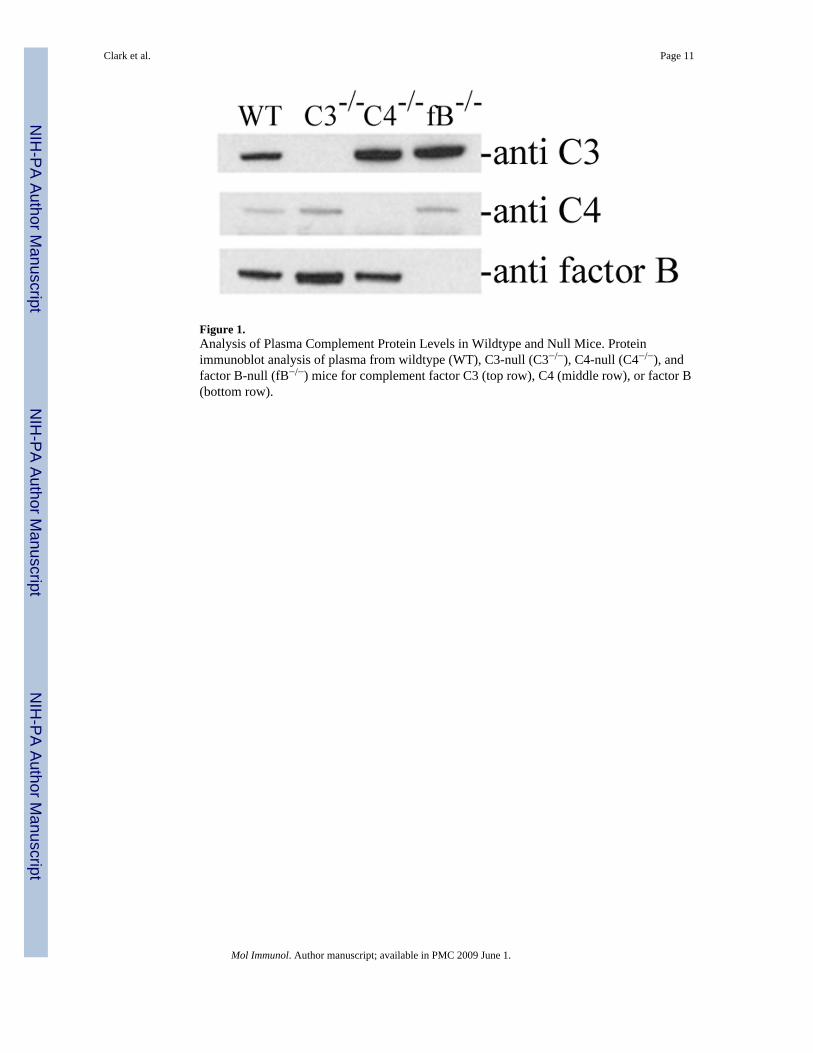
Figure 1.Analysis of Plasma Complement Protein Levels in Wildtype and Null Mice. Proteinimmunoblot analysis of plasma from wildtype (WT), C3-null (C3−/−), C4-null (C4−/−), andfactor B-null (fB−/−) mice for complement factor C3 (top row), C4 (middle row), or factor B(bottom row).
Clark et al. Page 11
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
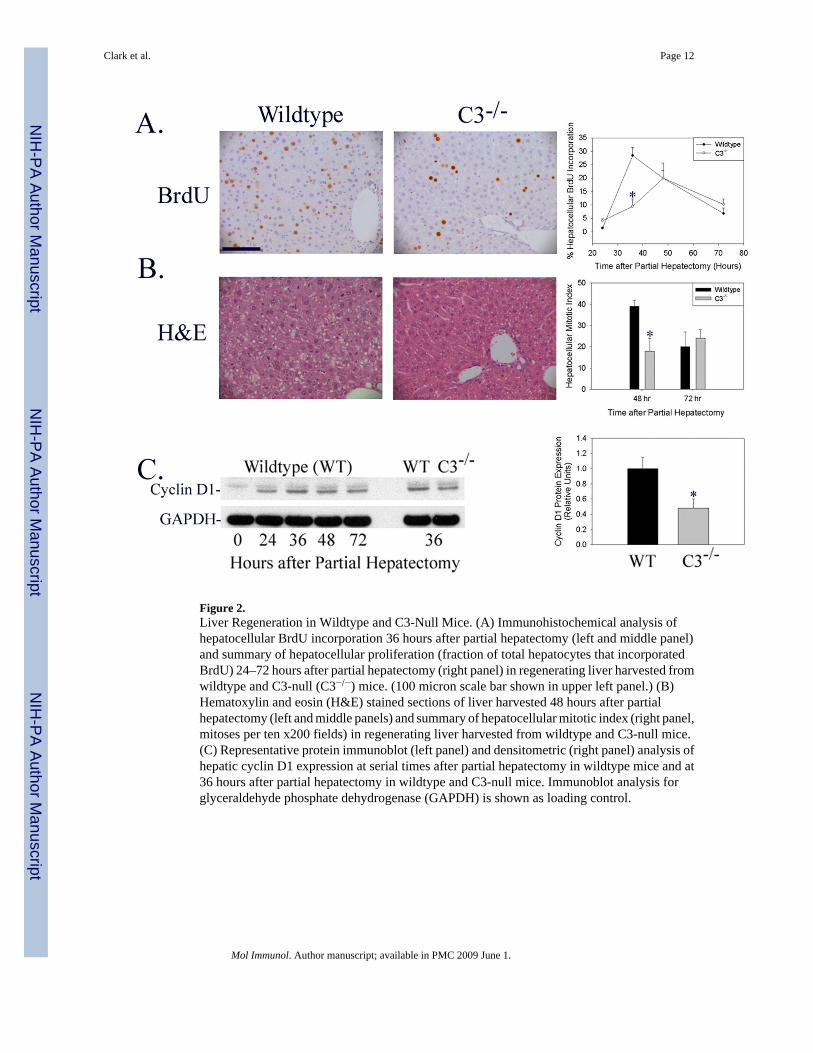
Figure 2.Liver Regeneration in Wildtype and C3-Null Mice. (A) Immunohistochemical analysis ofhepatocellular BrdU incorporation 36 hours after partial hepatectomy (left and middle panel)and summary of hepatocellular proliferation (fraction of total hepatocytes that incorporatedBrdU) 24–72 hours after partial hepatectomy (right panel) in regenerating liver harvested fromwildtype and C3-null (C3−/−) mice. (100 micron scale bar shown in upper left panel.) (B)Hematoxylin and eosin (H&E) stained sections of liver harvested 48 hours after partialhepatectomy (left and middle panels) and summary of hepatocellular mitotic index (right panel,mitoses per ten x200 fields) in regenerating liver harvested from wildtype and C3-null mice.(C) Representative protein immunoblot (left panel) and densitometric (right panel) analysis ofhepatic cyclin D1 expression at serial times after partial hepatectomy in wildtype mice and at36 hours after partial hepatectomy in wildtype and C3-null mice. Immunoblot analysis forglyceraldehyde phosphate dehydrogenase (GAPDH) is shown as loading control.
Clark et al. Page 12
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
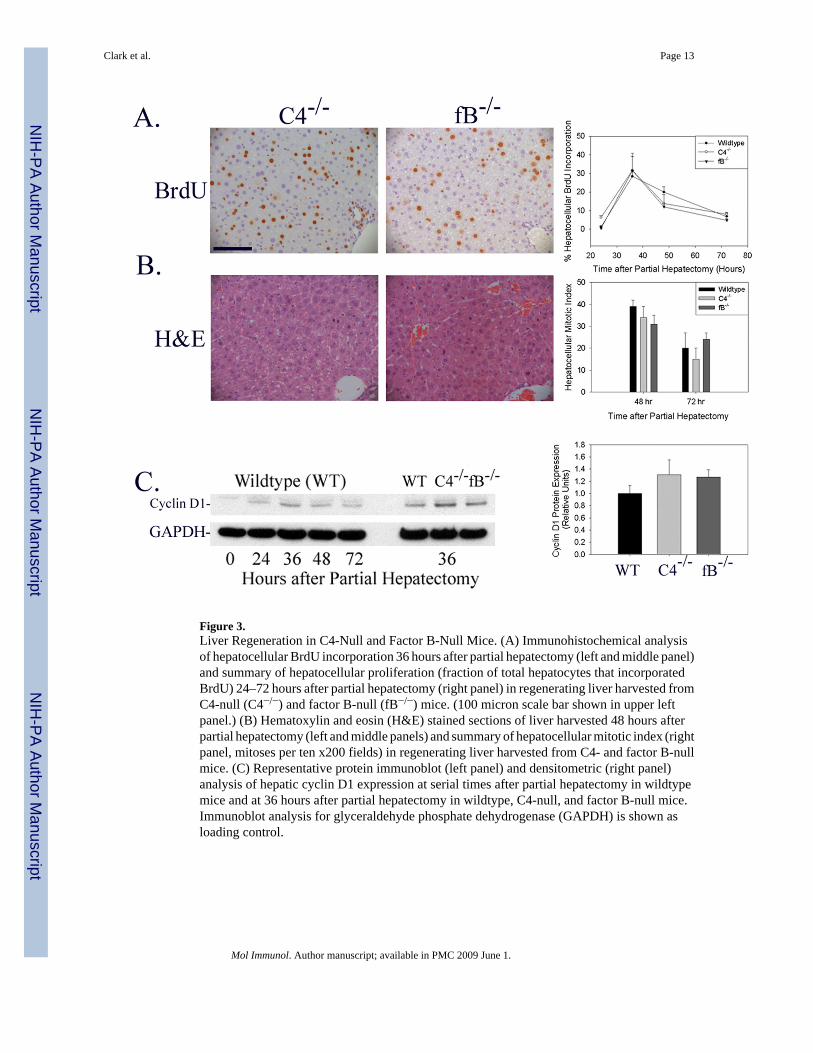
Figure 3.Liver Regeneration in C4-Null and Factor B-Null Mice. (A) Immunohistochemical analysisof hepatocellular BrdU incorporation 36 hours after partial hepatectomy (left and middle panel)and summary of hepatocellular proliferation (fraction of total hepatocytes that incorporatedBrdU) 24–72 hours after partial hepatectomy (right panel) in regenerating liver harvested fromC4-null (C4−/−) and factor B-null (fB−/−) mice. (100 micron scale bar shown in upper leftpanel.) (B) Hematoxylin and eosin (H&E) stained sections of liver harvested 48 hours afterpartial hepatectomy (left and middle panels) and summary of hepatocellular mitotic index (rightpanel, mitoses per ten x200 fields) in regenerating liver harvested from C4- and factor B-nullmice. (C) Representative protein immunoblot (left panel) and densitometric (right panel)analysis of hepatic cyclin D1 expression at serial times after partial hepatectomy in wildtypemice and at 36 hours after partial hepatectomy in wildtype, C4-null, and factor B-null mice.Immunoblot analysis for glyceraldehyde phosphate dehydrogenase (GAPDH) is shown asloading control.
Clark et al. Page 13
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
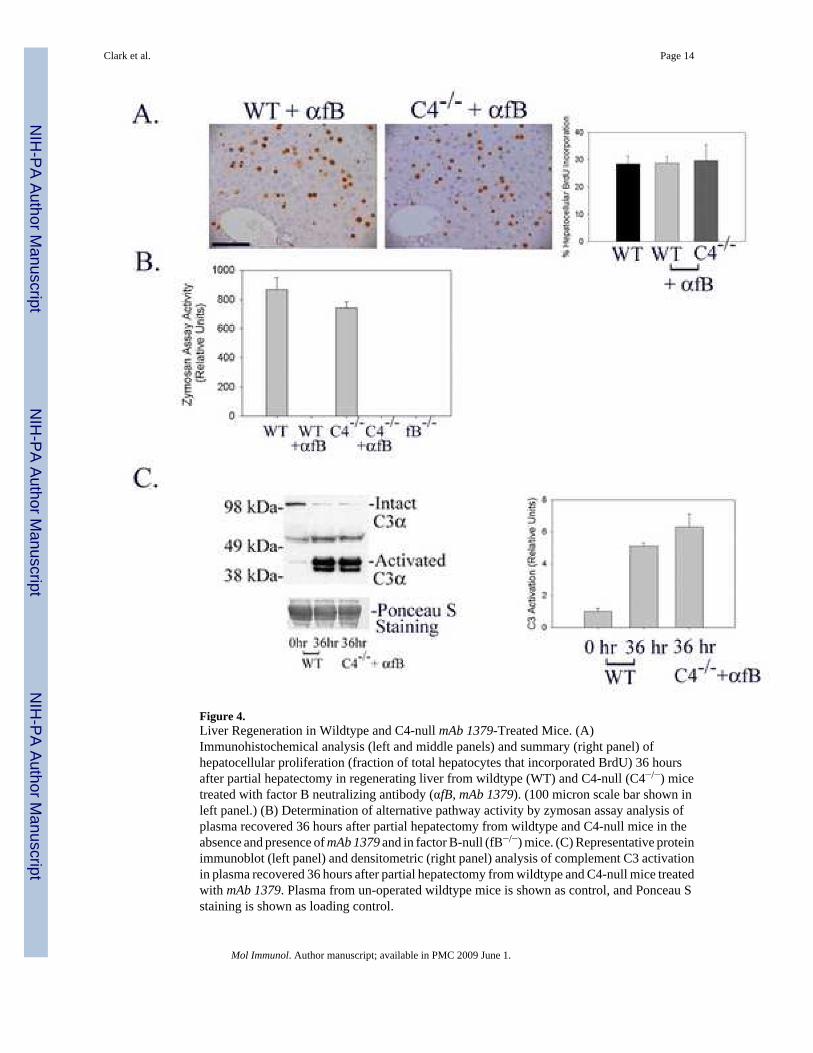
Figure 4.Liver Regeneration in Wildtype and C4-null mAb 1379-Treated Mice. (A)Immunohistochemical analysis (left and middle panels) and summary (right panel) ofhepatocellular proliferation (fraction of total hepatocytes that incorporated BrdU) 36 hoursafter partial hepatectomy in regenerating liver from wildtype (WT) and C4-null (C4−/−) micetreated with factor B neutralizing antibody (αfB, mAb 1379). (100 micron scale bar shown inleft panel.) (B) Determination of alternative pathway activity by zymosan assay analysis ofplasma recovered 36 hours after partial hepatectomy from wildtype and C4-null mice in theabsence and presence of mAb 1379 and in factor B-null (fB−/−) mice. (C) Representative proteinimmunoblot (left panel) and densitometric (right panel) analysis of complement C3 activationin plasma recovered 36 hours after partial hepatectomy from wildtype and C4-null mice treatedwith mAb 1379. Plasma from un-operated wildtype mice is shown as control, and Ponceau Sstaining is shown as loading control.
Clark et al. Page 14
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5.Representative protein immunoblot analysis of complement C3 activation in vitro in plasmafrom wildtype (WT), factor B null (fB−/−), C4 null (C4−/−), and mAb 1379 treated C4 null(C4−/−+αfB) mice unincubated (lanes a,e,i,m), or incubated in the absence (lanes b,f,j,n) orpresence of thrombin (lanes c,g,k,o) or plasmin (lanes d,h,l,p). Ponceau S staining is shown asloading control.
Clark et al. Page 15
Mol Immunol. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript




















