
Difflugia mulanensis nov. spec., a freshwater testate amoeba from Lake Mulan, China
Upload
independentCategory
view
0download
0
Es(
A
a
Bb
RA
A
astdcbmsasv©
K
I
pecd(
0h
Available online at www.sciencedirect.com
European Journal of Protistology 49 (2013) 238–248
nvironmental factors influencing soil testate amoebae in herbaceous andhrubby vegetation along an altitudinal gradient in subarctic tundraAbisko, Sweden)
ndrey N. Tsyganova,∗, Ann Milbaub, Louis Beyensa
Ecosystem Management Research Group, Department of Biology, University of Antwerp, Universiteitsplein 1, B-2610 Antwerp (Wilrijk),elgiumClimate Impacts Research Centre, Department of Ecology and Environmental Sciences, Umeå University, SE-98107 Abisko, Sweden
eceived 26 March 2012; received in revised form 20 July 2012; accepted 11 August 2012vailable online 28 September 2012
bstract
Shifts in community composition of soil protozoa in response to climate change may substantially influence microbial activitynd thereby decomposition processes. However, effects of climate and vegetation on soil protozoa remain poorly understood. Wetudied the distribution of soil testate amoebae in herbaceous and shrubby vegetation along an altitudinal gradient (from belowhe treeline at 500 m to the mid-alpine region at 900 m a.s.l.) in subarctic tundra. To explain patterns in abundance, speciesiversity and assemblage composition of testate amoebae, a data set of microclimate and soil chemical characteristics wasollected. Both elevation and vegetation influenced the assemblage composition of testate amoebae. The variation was regulatedy interactive effects of summer soil moisture, winter soil temperature, soil pH and nitrate ion concentrations. Besides, soiloisture regulated non-linear patterns in species richness across the gradient. This is the first study showing the effects of winter
oil temperatures on species composition of soil protozoa. The effects could be explained by specific adaptations of testatemoebae such as frost-resistant cysts allowing them to survive low winter temperatures. We conclude that the microclimate and
oil chemical characteristics are the main drivers of changes in protozoan assemblage composition in response to elevation andegetation.2012 Elsevier GmbH. All rights reserved.
oil; Tes
tbptt
eywords: Altitudinal gradient; Subarctic tundra; Microclimate; S
ntroduction
Understanding how environmental variables regulate soilrotozoa is useful for effective management of terrestrialcosystems and estimation of their responses to global
hange. Soil protozoa represent a common and abun-ant component of terrestrial ecosystems almost worldwideFoissner and Hawksworth 2009), considerably contributing∗Corresponding author. Fax: +32 3 265 22 71.E-mail address: [email protected] (A.N. Tsyganov).
aBrb(ca
932-4739/$ – see front matter © 2012 Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.ejop.2012.08.004
tate amoebae; Vegetation
o nutrient cycling (Coûteaux and Darbyshire 1998) and localiodiversity (Esteban et al. 2006; Finlay et al. 2000). Theylay an essential role in the functioning of terrestrial ecosys-ems as a part of soil decomposer food webs by exertingop-down control on microbial community composition andctivity (Clarholm 1985; Griffiths 1994; Rønn et al. 2002).y grazing on bacteria and other microorganisms, they indi-
ectly influence vegetation by regulating nutrient release and
acterial production of plant growth-promoting hormonesBonkowski 2004). Thus, changes in protozoan communityomposition in soils can influence below-ground processesnd, potentially, above-ground biota.
urnal o
emClpeceteqedwcctaecaior
eiseeoamTpttotW2eqrz(aesosasar
d2ata2artgaiab
oacs(tSimcstwtd
M
S
Msct31lbaiB
E
A.N. Tsyganov et al. / European Jo
Among the numerous environmental variables which influ-nce soil protozoa, the effects of climatic factors and theechanisms through which they operate are least understood.limate regulates soil microbial communities by determining
ocal meteorological characteristics such as air temperature,recipitation level and their variation patterns. However, theffects of climate-related environmental factors on soil biotaan be considerably mediated by the vegetation (Wookeyt al. 2009), which influences soil microclimate conditionshrough shading, precipitation interception, snow deposition,tc. (Graae et al. 2012). Besides, vegetation determines theuality, quantity, and timing of litter and root exudates whichnter the soil. Climate-dependent variation in primary pro-uction can therefore cause changes in decomposer foodebs (Bardgett and Wardle 2010). In order to understand the
limate regulation of soil protozoan assemblages, studies onlimate effects should account for the effects of vegetationhat could be achieved in observational field studies alongltitudinal gradients. Altitudinal gradients represent ‘naturalxperiments’ with a non-random variation in many climaticomponents including temperature, precipitation and season-lity (Dunne et al. 2004; Lomolino 2001). The data obtainedn such studies can considerably improve our understandingf the effects and the mechanisms of climate- and vegetation-elated factors on soil protozoa.
One of the most interesting groups of soil protozoa forcological studies is testate amoebae, a group of free liv-ng shelled protozoa which constitute a considerable part ofoil biota in terms of biomass and biodiversity (Ehrmannt al. 2012; Esteban et al. 2006; Finlay et al. 2000; Schrötert al. 2003). Testate amoebae prey on a wide range ofrganisms, including bacteria, protozoa, microalgae, fungi,nd micrometazoa, and may also consume dead organicatter (Gilbert et al. 2000; Wilkinson and Mitchell 2010).hanks to the shells, they can be enumerated in soil sam-les by ‘direct’ microscope counting that helps to avoidhe difficulties associated with labour-intensive cultivationechniques which are necessary for studies of other groupsf protozoa (Foissner 1987). Although the ecology of tes-ate amoebae has been intensively studied (Charman and
arner 1992; Lamentowicz et al. 2010; Mitchell et al.000b; Qin et al. 2011), data on their relationships withlevation- and vegetation-related environmental variables areuite scanty and inconsistent. Todorov (1998) observed aeduction in diversity of soil testate amoebae in the sub-alpineone (2000–2500 m a.s.l.) as compared to lower elevations400–2000 m a.s.l.) in samples taken from various soil typeslong a gradient located in south-western Bulgaria. Mitchellt al. (2004) did not detect any overall significant relation-hip between elevation and diversity or community structuref testate amoebae in the moss Hylocomium splendens in theouth-eastern Alps. Krashevska et al. (2007, 2008) reported
unimodal response of species richness and abundance ofoil testate amoebae to elevation with a peak at intermedi-te elevations along a slope located in a tropical mountainain forest in southern Ecuador. These relationships were
aAs
f Protistology 49 (2013) 238–248 239
riven by the availability of food resources (Krashevska et al.008). Studies on the effects of vegetation on soil testatemoebae demonstrated positive relationships between func-ional diversity of vascular plant communities and abundancend/or species diversity of testate amoebae (Carlson et al.010; Ledeganck et al. 2003). Sutton and Wilkinson (2007)lso demonstrated that the presence of Rhododendron specieseduced total abundance and species diversity and changedhe assemblage composition of soil testate amoebae. The sug-ested mechanism of the relationships between vegetationnd testate amoebae is the influence of vegetation on qual-ty and quantity of organic material which enters the soilnd regulates soil chemistry and activity and composition ofacterial communities.
The aim of the present study is to investigate the patternsf soil testate amoeba assemblages in relation to elevationnd vegetation type and to determine how these patterns areorrelated with environmental variables. For that purpose, wetudied the distribution of soil testate amoebae in herbaceousmeadow) and shrubby (willow) vegetation along an eleva-ional gradient (500–900 m a.s.l.) in subarctic tundra (Abisko,weden). To develop a model for explaining the variation
n soil testate amoeba assemblages we collected microcli-atic and chemical soil parameters. We focused on univariate
haracteristics (abundance, species diversity) and speciestructure of the testate amoeba assemblage and hypothesisedhat (i) the assemblage characteristics of soil testate amoebaeill vary with local environmental conditions along the alti-
udinal gradient and (ii) testate amoeba assemblages will beifferent in herbaceous and shrubby vegetation types.
aterial and Methods
tudy site
The study was performed on the north-east facing slope ofount Suorooaivi (1193 m), located approximately 20 km
outh of Abisko, Sweden (68◦17′N, 19◦15′E). The site isharacterised by a subarctic climate with a mean annual airemperature of −0.6 ◦C and a mean annual precipitation of05 mm (measured at Abisko Scientific Research Station,913–2010) (Callaghan et al. 2010). The growing seasonasts for about 130 days (Karlsson and Callaghan 1996). Theedrock at the site consists of salic igneous rocks and quarticnd phyllitic hard schists (Sundqvist et al. 2011). The treelines situated at an elevation of 500–600 m a.s.l. and formed byetula pubescenss sp. czerepanovii (mountain birch).
xperimental design and sampling
Soil sampling for this study was performed in meadownd willow vegetation types along an elevation transect onugust 18 and 19, 2009. The transect was established in the
ummer of 2007 for studying effects of microclimate on plant

2 urnal o
cBettsGwSSGgsowsmmcftspces3
S
mwfgUssvi(Bmmwpc
mUJlttt
ct�
4tuassT
T
abscalcadwfiUmAfworotReoacLHtfsttass
40 A.N. Tsyganov et al. / European Jo
olonisation and was fully described by Graae et al. (2012).riefly, three replicate rectangular plots (1.8 m × 0.8 m) werestablished for each vegetation type at every 100 m alonghe slope ranging from below the treeline at 500 m a.s.l. tohe mid-alpine region at 900 m a.s.l. In total, 30 plots wereet up. Altitudes were checked with a GPS device (Etrex,armin, USA) with an accuracy of ±15 m. The meadow plotsere dominated by herbs (Viola biflora, Carex bigelowii,aussurea alpina, Anthoxanthum alpinum, Bistorta vivipara,olidago virgaurea, Trollius europaeus, Vaccinium myrtillus,eranium sylvaticum, Epilobium angustifolium, Calama-rostis purpurea). The willow plots were dominated by Salixpp. (Salix lanata, Salix glauca, etc.) but also had manyf the same herb species in the understory, except for theillow plots at the 800 m, which were placed on dwarf
hrub dominated patches and were rather dry. In most plots,osses formed 10–20% of the vegetation cover (grid quadratethod). The vegetation types differed with respect to the
omposition of functional groups whereas the number ofunctional groups was equal. Overall, the meadow vegeta-ion was characterised by greater mean species diversity (17.5pecies vs. 13.9 species in the shrubby vegetation). At eachlot, two samples of 5 cm surface soil layer were randomlyollected under the vegetation with a cork corer (1.5 cm diam-ter). This sampling strategy resulted in 60 samples. Theamples were stored in screw-top plastic vials and fixed with% neutralised formaldehyde.
oil characteristics
Both microclimatic and soil chemical characteristics wereeasured in the plots. Soil temperature and soil moistureere continuously recorded in one of the three replicate plots
or each vegetation type at every elevation using data log-ers (Em50 Decagon Devices Inc., Pullman, Washington,SA) from October 2007. Soil temperature (◦C) was mea-
ured in the top 2 cm of soil layer with three temperatureensors (EST, Decagon Devices, Washington, USA). Soilolumetric moisture content (VWC, m3/m3) was measuredn the top 5 cm soil layer using two soil moisture sensorsEC-5, Decagon Devices Inc., Pullman, Washington, USA).oth soil temperature and soil moisture were measured everyinute and mean values per hour were stored. We calculatedean summer (June–September, 2008 and 2009) and meaninter (October–April, 2008/2009 and 2009/2010) soil tem-eratures. For soil moisture, only mean summer values werealculated.
Bio-available amounts of PO43−, NO3
−, NH4+ were deter-
ined in each plot using PST-1 resin capsules (Unibest, MT,SA) which were placed into the soil for two months (end
une till end August 2008). The capsules were then col-
ected and extracted by gently shaking them on a shakerable for 30 min in 10 mL of 2 M KCl. This was repeatedhree times and after each rinse KCl extracts were decantedo create a total extract volume of 30 mL. The KCl extractableS
s
f Protistology 49 (2013) 238–248
oncentrations of the nutrients were determined by colorime-ry. Values are given as “resin absorption quantity” (RAQ) inmol/cm2.For determination of soil pH, three soil cores (diameter of
.5 cm) of the top 5 cm soil layer were collected per plot onhe 29 and 30 July, 2008. The samples were stored at −15 ◦Cntil analyses. Prior to use, the samples were merged togethernd sieved through a 2-mm sieve. Soil pH was measured in theoil suspension (6 g of soil in 50 mL deionised water) afterhaking for about 18 h using a MP220 pH meter (Mettleroledo, Sweden).
estate amoeba analysis
The soil samples were prepared for counting of testatemoebae following a slightly modified version of the waterased procedure (Hendon and Charman 1997). First, the soilamples were air-dried at 40 ◦C for 48 h to normalise the waterontent. After that, the samples were carefully sieved through
2-mm sieve to disaggregate soil particles and removearge-size material, such as gravel and plant roots, whichould disproportionally influence concentrations of testatemoebae increasing sample weight. The sieving residue wasiscarded and 1–2 g of the remaining fine fraction were mixedith an arbitrary amount of deionised water. At the same time,ve tablets of Lycopodium spores (Batch No. 483216, Lundniversity, Sweden) were added to each sample as exoticarker spores for quantitative analysis (Stockmarr 1971).fter 12 h of soaking, the samples were thoroughly shaken
or 5 min to extract testate amoebae from soil. The suspensionas sequentially passed through sieves with mesh openingsf 300 and 10 �m, and the fraction between the two sieves wasetained. We used a finer sieve mesh for back sieving insteadf the 15 �m suggested by the original technique in ordero reduce the possible loss of small species (Payne 2009).ose Bengal was added to the samples in order to distinguishmpty shells from amoebae which were alive at the momentf sampling. Testate amoebae were identified and counted indrop of the concentrate mixed with glycerol at a magnifi-
ation of 400× using a light microscope (Olympus BX-50).iving individuals and empty shells were counted separately.owever, living amoebae were not abundant (2.5% of the
otal counts) therefore only empty shell counts were used forurther analyses. Although empty shell assemblages can toome extent be influenced by decomposition and by passiveransfer of the shells, they can still be used for studying rela-ionships between testate amoebae and environmental factorss they accumulate the effects of long-term trends in the corre-ponding living assemblages. A minimum total of 150 emptyhells was counted in each sample.
tatistical analyses
The statistical analysis of the data was performed in threeteps. First, we tested general patterns in testate amoeba
A.N. Tsyganov et al. / European Journal of Protistology 49 (2013) 238–248 241
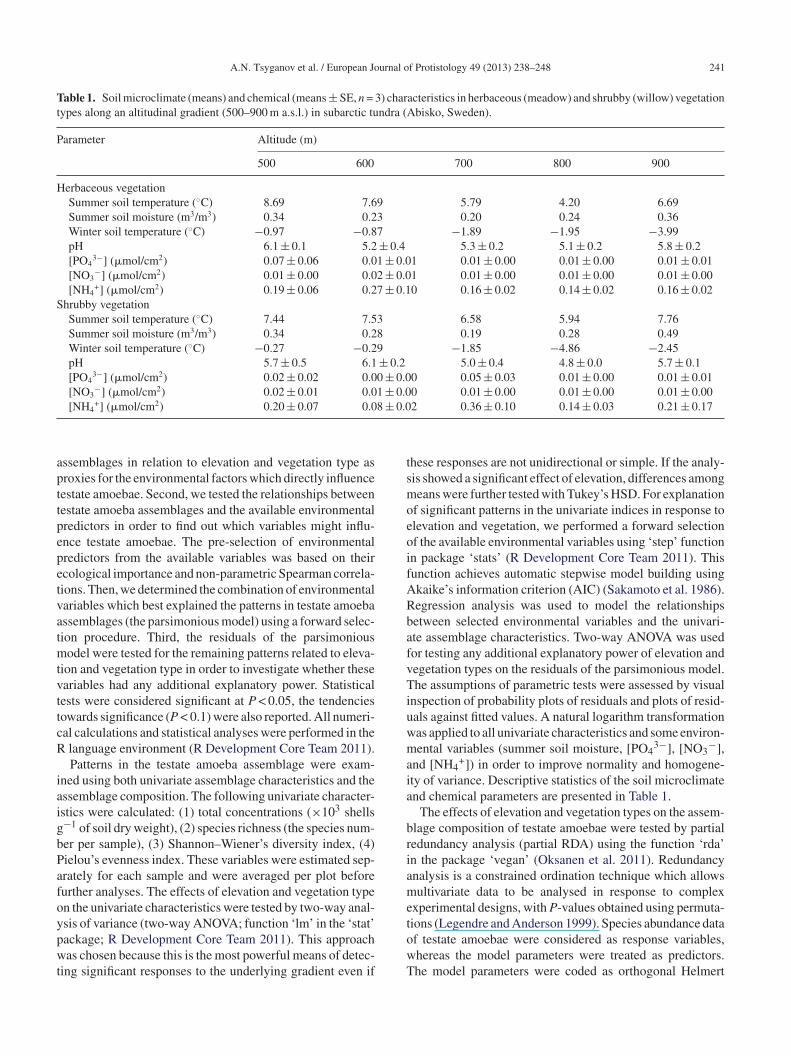
Table 1. Soil microclimate (means) and chemical (means ± SE, n = 3) characteristics in herbaceous (meadow) and shrubby (willow) vegetationtypes along an altitudinal gradient (500–900 m a.s.l.) in subarctic tundra (Abisko, Sweden).
Parameter Altitude (m)
500 600 700 800 900
Herbaceous vegetationSummer soil temperature (◦C) 8.69 7.69 5.79 4.20 6.69Summer soil moisture (m3/m3) 0.34 0.23 0.20 0.24 0.36Winter soil temperature (◦C) −0.97 −0.87 −1.89 −1.95 −3.99pH 6.1 ± 0.1 5.2 ± 0.4 5.3 ± 0.2 5.1 ± 0.2 5.8 ± 0.2[PO4
3−] (�mol/cm2) 0.07 ± 0.06 0.01 ± 0.01 0.01 ± 0.00 0.01 ± 0.00 0.01 ± 0.01[NO3
−] (�mol/cm2) 0.01 ± 0.00 0.02 ± 0.01 0.01 ± 0.00 0.01 ± 0.00 0.01 ± 0.00[NH4
+] (�mol/cm2) 0.19 ± 0.06 0.27 ± 0.10 0.16 ± 0.02 0.14 ± 0.02 0.16 ± 0.02Shrubby vegetation
Summer soil temperature (◦C) 7.44 7.53 6.58 5.94 7.76Summer soil moisture (m3/m3) 0.34 0.28 0.19 0.28 0.49Winter soil temperature (◦C) −0.27 −0.29 −1.85 −4.86 −2.45pH 5.7 ± 0.5 6.1 ± 0.2 5.0 ± 0.4 4.8 ± 0.0 5.7 ± 0.1[PO4
3−] (�mol/cm2) 0.02 ± 0.02 0.00 ± 0.00 0.05 ± 0.03 0.01 ± 0.00 0.01 ± 0.01[NO3
−] (�mol/cm2) 0.02 ± 0.01 0.01 ± 0.00 0.01 ± 0.00 0.01 ± 0.00 0.01 ± 0.00+ 2 8 ± 0.0
apttpepetvatmtvttcR
iaigbPafoypwt
tsmoeoifARbafvTiuwmaia
briamet
[NH4 ] (�mol/cm ) 0.20 ± 0.07 0.0
ssemblages in relation to elevation and vegetation type asroxies for the environmental factors which directly influenceestate amoebae. Second, we tested the relationships betweenestate amoeba assemblages and the available environmentalredictors in order to find out which variables might influ-nce testate amoebae. The pre-selection of environmentalredictors from the available variables was based on theircological importance and non-parametric Spearman correla-ions. Then, we determined the combination of environmentalariables which best explained the patterns in testate amoebassemblages (the parsimonious model) using a forward selec-ion procedure. Third, the residuals of the parsimonious
odel were tested for the remaining patterns related to eleva-ion and vegetation type in order to investigate whether theseariables had any additional explanatory power. Statisticalests were considered significant at P < 0.05, the tendenciesowards significance (P < 0.1) were also reported. All numeri-al calculations and statistical analyses were performed in thelanguage environment (R Development Core Team 2011).Patterns in the testate amoeba assemblage were exam-
ned using both univariate assemblage characteristics and thessemblage composition. The following univariate character-stics were calculated: (1) total concentrations (×103 shells−1 of soil dry weight), (2) species richness (the species num-er per sample), (3) Shannon–Wiener’s diversity index, (4)ielou’s evenness index. These variables were estimated sep-rately for each sample and were averaged per plot beforeurther analyses. The effects of elevation and vegetation typen the univariate characteristics were tested by two-way anal-sis of variance (two-way ANOVA; function ‘lm’ in the ‘stat’
ackage; R Development Core Team 2011). This approachas chosen because this is the most powerful means of detec-ing significant responses to the underlying gradient even if
owT
2 0.36 ± 0.10 0.14 ± 0.03 0.21 ± 0.17
hese responses are not unidirectional or simple. If the analy-is showed a significant effect of elevation, differences amongeans were further tested with Tukey’s HSD. For explanation
f significant patterns in the univariate indices in response tolevation and vegetation, we performed a forward selectionf the available environmental variables using ‘step’ functionn package ‘stats’ (R Development Core Team 2011). Thisunction achieves automatic stepwise model building usingkaike’s information criterion (AIC) (Sakamoto et al. 1986).egression analysis was used to model the relationshipsetween selected environmental variables and the univari-te assemblage characteristics. Two-way ANOVA was usedor testing any additional explanatory power of elevation andegetation types on the residuals of the parsimonious model.he assumptions of parametric tests were assessed by visual
nspection of probability plots of residuals and plots of resid-als against fitted values. A natural logarithm transformationas applied to all univariate characteristics and some environ-ental variables (summer soil moisture, [PO4
3−], [NO3−],
nd [NH4+]) in order to improve normality and homogene-
ty of variance. Descriptive statistics of the soil microclimatend chemical parameters are presented in Table 1.
The effects of elevation and vegetation types on the assem-lage composition of testate amoebae were tested by partialedundancy analysis (partial RDA) using the function ‘rda’n the package ‘vegan’ (Oksanen et al. 2011). Redundancynalysis is a constrained ordination technique which allowsultivariate data to be analysed in response to complex
xperimental designs, with P-values obtained using permuta-ions (Legendre and Anderson 1999). Species abundance data
f testate amoebae were considered as response variables,hereas the model parameters were treated as predictors.he model parameters were coded as orthogonal Helmert
2 urnal o
cwPiorattotinsetoOutasrp
‘etovcttmwtsbtivwtst
R
G
taag
accsaalpt4
Ua
tiowsUeabbRetrtnnFrtnot‘P
Sa
o2sPtP
42 A.N. Tsyganov et al. / European Jo
ontrasts to allow testing the factors and interaction in aay that provides the correct F values (Borcard et al. 2011).rior to the analysis, the species which were encountered
n four samples or less were removed from the data set inrder to eliminate unwarranted effects of rare taxa on theesults of ordination. The final data set for the multivariatenalysis contained 47 testate amoeba taxa (Table A1). Afterhat, species abundances were averaged per plot and Hellingerransformed (function ‘decostand’ in the package ‘vegan’) inrder to preserve Hellinger distance in RDA. The Hellingerransformation helps to avoid problems arising from Euclid-an distance, where the distance between two sites sharingo species in common can be smaller than between two sitesharing common species (Legendre and Gallagher 2001). Theffects of each model parameter and the interaction betweenhem were assessed by partialling out the effects of all thether factors in the model (Legendre and Anderson 1999).rdination diagrams based on the results of the RDA weresed to visualise the relationships between testate amoebaaxa and significant parameters. In order to explain the vari-tion represented by canonical axes we regressed the fittedite scores on the environmental data. Forward selection wasun with ‘step’ function (package ‘stats’) in order to build aarsimonious model.
Variation partitioning (function ‘varpart’ in the packagevegan’) was performed to quantify the unique and combinedffects of elevation together with vegetation and environmen-al variables on the variation in the assemblage compositionf testate amoebae (Borcard et al. 2011). Environmentalariables which explained the variation in the assemblageomposition were selected using the enhanced forward selec-ion procedure suggested by Blanchet et al. (2008). Beforehe forward selection, we tested the significance of the full
odel, which contained all explanatory variables. If the testas significant, the forward selection procedure was run with
he function ‘ordiR2step’ (package ‘vegan’) which performstepwise model selection using a double stopping criterionased on both adjusted R2 and P-values. The variation parti-ioning was repeated for the selected environmental variablesn order to estimate the contribution of each variable to theariation in assemblage composition. Unadjusted R2 valuesere corrected following Peres-Neto et al. (2006) (the func-
ion ‘RsquareAdj’ in the package ‘vegan’). Statistical tests ofignificance were carried out by 999 Monte Carlo permuta-ions under the reduced model.
esults
eneral observations
The analysis of the samples revealed 69 testate amoeba
axa belonging to 19 genera (Table A1). The testate amoebassemblage was dominated by six species: Centropyxiserophila sphagnicola (19% of the total counts), Difflugialobulus (15%), Plagiopyxis declivis (7.8%), Centropyxisnt(B
f Protistology 49 (2013) 238–248
erophila (7.6%), Centropyxis elongata (6.7%), and Tra-heleuglypha dentata (5.3%). All these species were alsoharacterised by high occurrence, in more than 80% of theamples, and thus constituted the core of the testate amoebassemblage at the site. A considerable number of testatemoeba taxa (24 taxa) were rare and were encountered iness than 10% of the samples. The mean species numberer sample was 19.9 ± SE 0.7 (n = 30). The mean concen-ration of testate amoebae at the site amounted to 36.98 ± SE.19 × 103 shells g−1 of dry soil weight (n = 30).
nivariate characteristics of the testate amoebassemblage
Two-way ANOVA showed a significant effect of eleva-ion on the species number and Shannon–Wiener’s diversityndex of the testate amoeba assemblage, whereas the variationf total abundance and Pielou’s evenness along the gradientas only marginally significant (Table 2 and Fig. 1). Both the
pecies number and Shannon–Wiener’s diversity index had-shaped relationships with elevation, being minimum at the
levations of 600 and 700 m, and maximum at 900 m (Fig. 1Bnd C). The patterns in the diversity index were mostly driveny changes in the species number (Spearman’s correlationetween the species number and Shannon–Wiener’s index:= 0.94, P < 0.001, n = 30). Therefore, we statistically mod-
lled the relationships between environmental variables andhe species number only. The forward selection procedureetained mean summer soil moisture as the only explana-ory variable which explained the variation in the speciesumber. The regression analysis showed that the speciesumber increased with mean summer soil moisture (Fig. 2;1,28 = 11.05, P < 0.002, R2 = 0.28). The two-way ANOVA of
esiduals of the regression model showed no extra explana-ory power of elevation and vegetation type on the speciesumber (P > 0.05 for all terms). Vegetation did not influencer mediate the effect of elevation on the univariate charac-eristics of the testate amoeba assemblage (non-significantvegetation type’ and ‘elevation × vegetation type’ terms;> 0.05).
pecies composition of the testate amoebassemblage
Redundancy analysis with elevation and vegetation as thenly explanatory variables showed that the model explained8.7% (R2
adj) of the total variation in the species compo-ition of the testate amoeba assemblage (pseudo-F = 2.30,< 0.001). Partial RDA indicated a significant interac-
ive effect of elevation and vegetation (pseudo-F = 1.64,< 0.001). The model could be decomposed into four sig-
ificant canonical axes (the results are not shown); howeverhe first two axes explained the most of the total variance15.2%), and therefore only these are shown (Fig. 3A and). The distribution of the sampling locations on the diagram

A.N. Tsyganov et al. / European Journal of Protistology 49 (2013) 238–248 243
Fig. 1. The variation of total abundance (A); the species number (B), Shannon–Wiener’s diversity index (C), and Pielou’s evenness index (D)of the soil testate amoeba assemblage in herbaceous (meadow) and shrubby (willow) vegetation types along an altitudinal gradient (500–900 ma ificant( ata are
dbiita
gva
Fbslb
a(wstac
.s.l.) in subarctic tundra (Abisko, Sweden). The letters indicate signbars topped with the same letters are not significantly different). D
id not show any well defined patterns which could be unam-iguously related to the effects of elevation, vegetation or thenteraction between them (Fig. 3A). In other words, the samenteractive environmental gradients influence the patterns ofhe species composition both across the altitudinal gradientnd in the vegetation types.
The first canonical axis accounted for the environmental
radient which was non-linearly related to elevation (the ele-ations in the order of increasing effects: 800, 700, 500, 600nd 900 m) and reflected the differences between meadowig. 2. Linear regression of the loge-transformed species num-er (loge(species number)) dependent on loge-transformed meanummer soil moisture (loge(summer soil moisture)). The solidine is the least square regression, equation: loge(species num-er) = 3.43 + 0.36 × loge(mean summer soil moisture).
pbaw7P
catiTmrt(yaawth
effects of the elevation on the univariate assemblage characteristicsmeans ± 1 SE (n = 3).
nd willow vegetation at the elevations of 600 and 800Fig. 3A). Forward selection showed that this canonical axisas positively related to mean summer soil moisture and
oil pH and negatively related to [NO3−] (Table 3). All
ogether, these factors explained 59.6% (R2adj) of the variance
long the axis (F3,26 = 27.22, P < 0.001). The second canoni-al axis accounted for the environmental gradient which wasositively related to elevation and reflected the differencesetween the vegetation types at all elevations (Fig. 3A). Thexis was negatively related to soil pH, [NO3
−] and meaninter soil temperatures (Table 3) which explained together3.06% (R2
adj) of the variation along the axis (F3,26 = 27.22,< 0.001).Combinations of the factors were associated with the spe-
ific assemblage composition of testate amoebae (Fig. 3And B). The biotopes clustered in the upper left quarter ofhe diagram were characterised by harsh conditions: dry soiln summer, low winter temperatures and acidic environment.he species composition of testate amoebae in these biotopesostly consisted of ubiquitous species: Assulina musco-
um, Corythion dubium, Trinema complanatum, Euglyphauberculata minor, etc. The second group of the biotopesupper right quarter) was associated with the species Arg-nnia (Nebela) dentata, Nebela collaris, Nebela tubulosand Padaungiella wailesi which preferred humid, moderatelycidic conditions and low concentrations of nitrate ions and
ere resistant to low winter temperatures. The biotopes inhe lower right quarter of the diagram were characterised byumid and warm conditions with the least acidic environment

244 A.N. Tsyganov et al. / European Journal of Protistology 49 (2013) 238–248
Table 2. F-Statistics for the two-way analysis of variance (ANOVA) testing the effects of elevation and vegetation types on the univariatecharacteristics of soil testate amoeba assemblages in herbaceous (meadow) and shrubby (willow) vegetation types along an altitudinal gradient(500–900 m a.s.l.) in subarctic tundra (Abisko, Sweden). All assemblage level characteristics were loge-transformed.
dfa Abundance Species number Shannon’s index Pielou’s index
Elevation 4 2.75+ 3.31* 3.15* 2.42+
Vegetation 1 1.40 0.60 1.50 2.43Elevation × vegetation 4 1.29 0.76 0.93 0.96Residuals 20
a
aihawa
tstvsowt(tbv
vacettF
D
G
a
Fabcswo
Degrees of freedom.*P < 0.05.+P < 0.1.
nd medium concentrations of nitrate ions. The biotopes withntermediate moisture, warm winter soil temperatures andigh [NO3
−] (lower middle part of the diagram) were char-cterised by Centropyxis plagiostoma and Cyclopyxis kahlihich seemed to be least tolerant to low winter soil temper-
tures.The test of the full model including all the environmen-
al variables except for elevation and vegetation type wasignificant (pseudo-F = 2.37, P < 0.001) so that forward selec-ion was performed. The procedure retained four explanatoryariables: pH, [NO3
−], mean winter temperature and meanummer soil moisture. The unique and combined effectsf the environmental variables and the elevation togetherith vegetation on the assemblage composition of soil
estate amoebae were quantified by variation partitioningFig. 4). The results indicate that 17% of the total varia-
ion in the assemblage composition was jointly explainedy the elevation- and vegetation-dependent environmentalariables. Apart from that, elevation- and vegetation-relatedpdna
ig. 3. Ordination diagrams illustrating the results of the redundancy anas the only explanatory variables. (A) Distance biplot of site scores (scalinelonging to the same class (white diamonds – herbaceous vegetation; blaorresponding elevations. Dashed lines are the Hellinger distance betweenpecies scores (scaling 2). The ordination was performed on the correlationhich were encountered in more than four samples (see Table A1 for the sf the total variation explained by each axis (both axes are significant; P <
ariables, which were not included in the study, stillccounted for 12% of the total variation in the assemblageomposition (Fig. 4; pseudo-F = 1.65, P < 0.001). The pureffect of elevation- and vegetation-independent variation ofhe selected environmental variables explained 6% of theotal variation in the assemblage composition (Fig. 4; pseudo-= 1.87, P < 0.003).
iscussion
eneral observations
The soil testate amoeba assemblage at the site was char-cterised by high taxonomic diversity and a considerable
roportion of taxa with low occurrence. The assemblage wasominated by typical soil taxa (Centropyxis aerophila sphag-icola, Difflugia globulus, Plagiopyxis declivis, Centropyxiserophila, Centropyxis elongata, Tracheleuglypha dentata),lysis of species abundance data with elevation and vegetation typeg 1). Diamonds are centroids (means ± SE, n = 3) of the site pointsck diamonds – shrubby vegetation). Numbers are centroids of thethe vegetation types at the same elevation. (B) Correlation plot ofmatrix of the Hellinger-transformed concentrations of the species
pecies codes). The numbers in the parentheses are the proportions0.001).

A.N. Tsyganov et al. / European Journal o
Table 3. Results of the multiple regressions showing the environ-mental variables which explained the variation in the assemblagecomposition of testate amoebae in response to elevation and vegeta-tion (the site scores along the two canonical axis: RDA1 and RDA2;see Fig. 3A). Estimate refers to the estimated partial regressionslope.
Parameter Estimate Standarderror
t-Value
RDA1Intercept −2.38 1.03 −2.30loge(summer soil moisture) 0.68 0.24 2.81**
pH 0.34 0.11 3.06**
loge[NO3−] −0.29 0.11 −2.75*
RDA2Intercept 0.80 0.51 1.56Winter soil temperature −0.13 0.03 −4.49***
pH −0.35 0.07 −4.92***
loge[NO3−] −0.19 0.07 −2.78*
*P < 0.1.**P < 0.05.
ydt(eessoo
Fptavpe
oawco
Ua
tSspmtcdsta2nicrlg
d
***P < 0.001.
et moss-dwelling species were regularly encountered. Theominant species have been previously reported to be theypical component of soil ecosystems in the Arctic regionBeyens and Chardez 1995; Beyens et al. 2009; Carlsont al. 2010; Tsyganov et al. 2011) or elsewhere (Schrötert al. 2003; Wanner 1991). The occurrence of moss-dwellingpecies (Heleopera spp., Hyalosphenia spp., Trygonopyxis
pp.) in the soil samples can be explained by the presencef mosses in the understorey of the vegetation. The majorityf the other taxa in the assemblage were either soil-dwellingig. 4. Venn diagram of the variation partitioning representing pro-ortions of unique and shared contributions of the model describinghe variation related to elevation together with the vegetation typend the parsimonious model which includes four environmentalariables (mean summer soil moisture, mean winter temperature,H, and [NO3
−]). The fractions of the circles represent the variationxplained by the models or by their combinations (the intersections).
TbbaatcpStvto2tv
Sa
vvbl
f Protistology 49 (2013) 238–248 245
r ubiquitous. Overall, taxonomic composition of the testatemoeba assemblage at the site was typical for soil biotopesith understory moss vegetation suggesting that our findings
an have bearing for other soil ecosystems beyond the limitsf the studied location.
nivariate characteristics of the testate amoebassemblage
Among all the univariate characteristics of the tes-ate amoeba assemblage, only the species number andhannon–Wiener’s diversity index had an U-shaped relation-hip with elevation. The relationship was governed by theositive link between the species number and mean sum-er soil moisture. These results support our hypothesis about
he greater importance of local environmental conditions inomparison to large-scale gradients in regulation of speciesiversity of soil testate amoebae. These results are also con-istent with the previously reported effects of soil moisture onhe species number and abundance of soil protozoa (Finlaynd Fenchel 2001) and ecological processes in soil (Anderson011). Moist soil biotopes can normally harbour a greaterumber of protozoan species because of the higher availabil-ty of large pores filled with water for large species. Thus,limate-independent variation in soil moisture, which can beelated to topography and/or geology, can considerably alterocal patterns in species diversity of soil biota along climaticradients.
Univariate characteristics of soil testate amoebae did notiffer between the herbaceous and shrubby vegetation types.he vegetation types in our study differed in the species num-er of vascular plants and composition of functional groupsut were similar in the number of functional groups. Thebsence of any effects of the vegetation on the univariatessemblage characteristics indirectly supports the observa-ions of Ledeganck et al. (2003), who found a positiveorrelation between soil testate amoeba diversity and vascularlant functional diversity but not with plant species identity.urprisingly, although the univariate characteristics of the
estate amoebae were similar in the herbaceous and shrubbyegetation, the assemblage composition did differ betweenhe vegetation types. These results are in line with the previ-us observations (Mitchell et al. 2000a; Sutton and Wilkinson007) which indicated that assemblage composition of tes-ate amoebae is more sensitive to variation in environmentalariables than the univariate characteristics.
pecies composition of the testate amoebassemblage
The species composition of testate amoeba assemblages
aried in response to elevation and vegetation. Both theariation across the altitudinal gradient and the differencesetween the herbaceous and shrubby vegetation were regu-ated by the same environmental variables which were soil
2 urnal o
sctbaetsfsas(2awaccibs
eoatoawbwt(msvpiip
tefiscdcsvcsec
ts
A
coFPtaMf
A
fj
R
A
B
B
B
B
B
B
C
C
46 A.N. Tsyganov et al. / European Jo
ummer moisture, winter soil temperature, soil pH and con-entration of nitrate ions. The effects of the variables onhe assemblage composition were interactive so that com-inations of environmental variables resulted in the specificssemblage composition of testate amoebae. To our knowl-dge, this is the first study showing effects of winter soilemperature on the species composition of soil protozoa inubarctic ecosystems. The effects could be explained by theact that low winter temperatures require specific adaptationsuch as cysts which are resistant to freezing. The other vari-bles have previously been recognised as major drivers ofpecies composition of testate amoeba assemblage in soilsKrashevska et al. 2010; Lousier 1974a,b; Schröter et al.003). Moreover, our results demonstrate that these vari-bles also accounted for a considerable part of the variancehich was elevation- and vegetation-independent. This vari-
tion could be structured by geological or topographicalharacteristics at the site. Thus, basic microclimatic and soilhemical characteristics are good predictors of the variationn the species composition of soil testate amoeba assemblagesoth across altitudinal gradients and between herbaceous andhrubby vegetation.
The results of the variation partitioning demonstrate thatlevation and vegetation type explained a considerable partf the variation in the species composition of testate amoebassemblage even after removing the effects of the above men-ioned environmental variables. This means that there werether elevation- and vegetation-related environmental vari-bles which regulated the species composition but whichere not included in the present study. These variables mighte concentrations of minerals (e.g. zinc, calcium, iron, etc.),hich have been shown to influence species composition of
estate amoebae in soils (Carlson et al. 2010) and in mossesMitchell et al. 2004) across altitudinal gradients. It alsoight be the number of freeze–thaw cycles, variation in
easonality and disturbance regimes. These characteristicsary with elevation (Lomolino 2001), but their effects on soilrotozoa have not received much attention yet. Thus, exam-nation of the effects of these variables in future studies maymprove our knowledge of the environmental regulation ofrotozoan assemblages in soils.
Overall, our results demonstrate that the species composi-ion of soil testate amoeba assemblages is very sensitive tolevation- and vegetation-related variation in environmentalactors. Soil moisture is the most important factor regulat-ng not only the composition of soil protozoa but also thepecies number. Therefore, climate effects on soil protozoaould be expected to be mostly driven by variation in climate-ependent soil moisture. The other predictors of speciesomposition of testate amoeba are winter soil temperature,oil pH and concentrations of nitrate ions. The effects of theariables on species composition are highly interactive so
ombinations of factors are associated with specific speciestructure. Besides, other elevation- and vegetation-relatednvironmental factors, such as the number of freeze–thawycles, variation in seasonality and biotic interactions, seemC
f Protistology 49 (2013) 238–248
o influence soil protozoan communities and their effectshould be examined in further studies.
cknowledgements
The fieldwork at the Abisko Research Station was finan-ially supported by a grant from the Royal Swedish Academyf Science (ANS Scholarship 2009). We thank the Kempeoundation for funding the data loggers; Till Jochum, Timflügner, and Ive Van Krunkelsven for setting-up and main-
aining the data loggers; Maja Sundqvist for the nutrientnalyses and Bente Graae, Anna Shevtsova, Nora Osler,aria Mooshammer, Ida-Maria Blåhed and Andrea Claes
or assistance in the field.
ppendix A. Supplementary data
Supplementary data associated with this article can beound, in the online version, at http://dx.doi.org/10.1016/.ejop.2012.08.004.
eferences
nderson, O.R., 2011. Soil respiration, climate change and the roleof microbial communities. Protist 162, 679–690.
ardgett, R.D., Wardle, D.A., 2010. Aboveground–BelowgroundLinkages: Biotic Interactions, Ecosystem Processes, and GlobalChange. Oxford University Press, Oxford.
eyens, L., Chardez, D., 1995. An annotated list of testate amoebaeobserved in the Arctic between the longitudes 27 degrees E and168 degrees W. Arch. Protistenkd. 146, 219–233.
eyens, L., Ledeganck, P., Graae, B.J., Nijs, I., 2009. Are soil biotabuffered against climatic extremes? An experimental test on tes-tate amoebae in arctic tundra (Qeqertarsuaq, West Greenland).Polar Biol. 32, 453–462.
lanchet, F.G., Legendre, P., Borcard, D., 2008. Forward selectionof explanatory variables. Ecology 89, 2623–2632.
onkowski, M., 2004. Protozoa and plant growth: the microbialloop in soil revisited. New Phytol. 162, 617–631.
orcard, D., Gillet, F., Legendre, P., 2011. Numerical Ecology withR. Springer, New York.
allaghan, T.V., Bergholm, F., Christensen, T.R., Jonasson, C.,Kokfelt, U., Johansson, M., 2010. A new climate era in thesub-Arctic: accelerating climate changes and multiple impacts.Geophys. Res. Lett. 37, L14705.
arlson, M.L., Flagstad, L.A., Gillet, F., Mitchell, E.A.D., 2010.Community development along a proglacial chronosequence:are above-ground and below-ground community structure con-trolled more by biotic than abiotic factors? J. Ecol. 98,1084–1095.
harman, D.J., Warner, B.G., 1992. Relationship between testateamebas (Protozoa, Rhizopoda) and microenvironmental param-eters on a forested peatland in northeastern Ontario. Can. J. Zool.70, 2474–2482.

urnal o
C
C
D
E
E
F
F
F
F
G
G
G
H
K
K
K
K
L
L
L
L
L
L
L
M
M
M
O
P
P
Q
R
R
S
S
S
S
A.N. Tsyganov et al. / European Jo
larholm, M., 1985. Interactions of bacteria, protozoa and plantsleading to mineralization of soil nitrogen. Soil Biol. Biochem.17, 181–187.
oûteaux, M.-M., Darbyshire, J.F., 1998. Functional diversityamongst soil protozoa. Appl. Soil Ecol. 10, 229–237.
unne, J.A., Saleska, S.R., Fischer, M.L., Harte, J., 2004. Inte-grating experimental and gradient methods in ecological climatechange research. Ecology 85, 904–916.
hrmann, O., Puppe, D., Wanner, M., Kaczorek, D., Sommer, M.,2012. Testate amoebae in 31 mature forest ecosystems – densitiesand micro-distribution in soils. Eur. J. Protistol. 48, 161–168.
steban, G.F., Clarke, K.J., Olmo, J.L., Finlay, B.J., 2006. Soilprotozoa – an intensive study of population dynamics and com-munity structure in an upland grassland. Appl. Soil Ecol. 33,137–151.
inlay, B.J., Black, H.I.J., Brown, S., Clarke, K.J., Esteban, G.F.,Hindle, R.M., Olmo, J.L., Rollett, A., Vickerman, K., 2000. Esti-mating the growth potential of the soil protozoan community.Protist 151, 69–80.
inlay, B.J., Fenchel, T., 2001. Protozoan community structure in afractal soil environment. Protist 152, 203–218.
oissner, W., 1987. Soil protozoa: fundamental problems, ecologicalsignificance, adaptations in Ciliates and Testateceans, bioindica-tors and guide to the literature. Prog. Protistol. 2, 69–212.
oissner, W., Hawksworth, D.L., 2009. Protist Diversity and Geo-graphical Distribution. Springer, Dordrecht.
ilbert, D., Amblard, C., Bourdier, G., Francez, A.-J., Mitchell,E.A.D., 2000. Le régime alimentaire des thécamoebiens. AnnéeBiol. 39, 57–68.
raae, B.J., De Frenne, P., Kolb, A., Brunet, J., Chabrerie, O., Ver-heyen, K., Pepin, N., Heinken, T., Zobel, M., Shevtsova, A., Nijs,I., Milbau, A., 2012. On the use of weather data in ecologicalstudies along altitudinal and latitudinal gradients. Oikos 121,3–19.
riffiths, B.S., 1994. Microbial-feeding nematodes and protozoa insoil: their effects on microbial activity and nitrogen mineraliza-tion in decomposition hotspots and the rhizosphere. Plant Soil164, 25–33.
endon, D., Charman, D.J., 1997. The preparation of testate amoe-bae (Protozoa: Rhizopoda) samples from peat. Holocene 7,199–205.
arlsson, P.S., Callaghan, T.V., 1996. Plant ecology in the subarcticSwedish Lapland. Ecol. Bull. 45, 220–227.
rashevska, V., Bonkowski, M., Maraun, M., Ruess, L., Kandeler,E., Scheu, S., 2008. Microorganisms as driving factors for thecommunity structure of testate amoebae along an altitudinal tran-sect in tropical mountain rain forests. Soil Biol. Biochem. 40,2427–2433.
rashevska, V., Bonkowski, M., Maraun, M., Scheu, S., 2007. Tes-tate amoebae (protista) of an elevational gradient in the tropicalmountain rain forest of Ecuador. Pedobiologia 51, 319–331.
rashevska, V., Maraun, M., Ruess, L., Scheu, S., 2010. Carbon andnutrient limitation of soil microorganisms and microbial grazersin a tropical montane rain forest. Oikos 119, 1020–1028.
amentowicz, M., Van der Knaap, W., Lamentowicz, L., VanLeeuwen, J.F.N., Mitchell, E.A.D., Goslar, T., Kamenik, C.,2010. A near-annual palaeohydrological study based on testate
amoebae from a sub-alpine mire: surface wetness and the role ofclimate during the instrumental period. J. Quat. Sci. 25, 190–202.edeganck, P., Nijs, I., Beyens, L., 2003. Plant functional groupdiversity promotes soil protist diversity. Protist 154, 239–249.
S
f Protistology 49 (2013) 238–248 247
egendre, P., Anderson, M.J., 1999. Distance-based redundancyanalysis: testing multispecies responses in multifactorial eco-logical experiments. Ecol. Monogr. 69, 1–24.
egendre, P., Gallagher, E.D., 2001. Ecologically meaningfultransformations for ordination of species data. Oecologia 129,271–280.
omolino, M.V., 2001. Elevation gradients of species-density:historical and prospective views. Glob. Ecol. Biogeogr. 10,3–13.
ousier, J.D., 1974a. Effects of experimental soil moisture fluc-tuations on turnover rates of Testacea. Soil Biol. Biochem. 6,19–26.
ousier, J.D., 1974b. Response of soil Testacea to soil moisturefluctuations. Soil Biol. Biochem. 6, 235–239.
itchell, E.A.D., Borcard, D., Buttler, A.J., Grosvernier, P., Gilbert,D., Gobat, J.M., 2000a. Horizontal distribution patterns of tes-tate amoebae (Protozoa) in a Sphagnum magellanicum carpet.Microb. Ecol. 39, 290–300.
itchell, E.A.D., Bragazza, L., Gerdol, R., 2004. Testate amoe-bae (Protista) communities in Hylocomium splendens (Hedw.)B.S.G. (Bryophyta): relationships with altitude, and moss ele-mental chemistry. Protist 155, 423–436.
itchell, E.A.D., Buttler, A., Grosvernier, P., Rydin, H., Albinsson,C., Greenup, A.L., Heijmans, M., Hoosbeek, M.R., Saarinen, T.,2000b. Relationships among testate amoebae (Protozoa), vegeta-tion and water chemistry in five Sphagnum-dominated peatlandsin Europe. New Phytol. 145, 95–106.
ksanen, J., Blanchet, F.G., Kindt, R., Legendre, P., Minchin, P.R.,O’Hara, R.B., Simpson, G.L., Solymos, P., Stevens, M.H.H.,Wagner, H., 2011. vegan: Community Ecology Package.
ayne, R., 2009. The standard preparation method for testate amoe-bae leads to selective loss of the smallest taxa. Quat. Newslett.119, 16–20.
eres-Neto, P.R., Legendre, P., Dray, S., Borcard, D., 2006. Variationpartitioning of species data matrices: estimation and comparisonof fractions. Ecology 87, 2614–2625.
in, Y., Xie, S., Smith, H.G., Swindles, G.T., Gu, Y., 2011. Diver-sity, distribution and biogeography of testate amoebae in China:implications for ecological studies in Asia. Eur. J. Protistol. 47,1–9.
Development Core Team, 2011. R: A Language and Environmentfor Statistical Computing. R Foundation for Statistical Comput-ing, Vienna.
ønn, R., McCaig, A.E., Griffiths, B.S., Prosser, J.I., 2002.Impact of protozoan grazing on bacterial community struc-ture in soil microcosms. Appl. Environ. Microbiol. 68,6094–6105.
akamoto, Y., Ishiguro, M., Kitagawa, G., 1986. Akaike InformationCriterion Statistics. Reidel Publishing Company, Dordrecht.
chröter, D., Wolters, V., De Ruiter, P.C., 2003. C and N mineralisa-tion in the decomposer food webs of a European forest transect.Oikos 102, 294–308.
tockmarr, J., 1971. Tablets with spores used in absolute pollenanalysis. Pollen et Spores 13, 615–621.
undqvist, M.K., Giesler, R., Graae, B.J., Wallander, H., Fogelberg,E., Wardle, D.A., 2011. Interactive effects of vegetation typeand elevation on aboveground and belowground properties in a
subarctic tundra. Oikos 120, 128–142.utton, C.A., Wilkinson, D.M., 2007. The effects of Rhododendronon testate amoebae communities in woodland soils in north westEngland. Acta Protozool. 46, 333–338.

2 urnal o
T
T
W
W
W
Lavorel, S., Shaver, G.R., 2009. Ecosystem feedbacks and cas-
48 A.N. Tsyganov et al. / European Jo
odorov, M., 1998. Observation on the soil and moss testate amoe-bae (Protozoa: Rhizopoda) from Pirin Mountain (Bulgaria). ActaZool. Bulg. 50, 19–29.
syganov, A.N., Nijs, I., Beyens, L., 2011. Does climate warmingstimulate or inhibit soil protist communities? A test on tes-tate amoebae in high-arctic tundra with Free-Air Temperature
Increase. Protist 162, 237–248.anner, M., 1991. Zur Morphologie von Thekamöben (Protozoa:Rhizopoda) in süddeutschen Wäldern. Arch. Protistenkd. 140,236–288.
f Protistology 49 (2013) 238–248
ilkinson, D.M., Mitchell, E.A.D., 2010. Testate amoebae andnutrient cycling with particular reference to soils. Geomicrobiol.J. 27, 520–533.
ookey, P.A., Aerts, R., Bardgett, R.D., Baptist, F., Bråthen, K.A.,Cornelissen, J.H.C., Gough, L., Hartley, I.P., Hopkins, D.W.,
cade processes: understanding their role in the responses ofArctic and alpine ecosystems to environmental change. Glob.Change Biol. 15, 1153–1172.
Copyright © 2022 FDOKUMEN




















