
Environmental drivers of temporal succession in recent dinoflagellate cyst assemblages from a...
23
Environmental drivers of temporal succession in recent dinoflagellate cyst assemblages from a coastal site in the North-East Atlantic (Lisbon Bay, Portugal) Sofia Ribeiro 1 , Ana Amorim ⁎ Instituto de Oceanografia, Faculdade de Ciências da Universidade de Lisboa, Campo Grande 1749-016 Lisboa, Portugal Received 21 December 2007; accepted 4 January 2008 Abstract Temporal changes in the community structure of recent dinoflagellate cyst assemblages of Lisbon Bay (Iberian upwelling system) were investigated between 2000 and 2005. The assemblages were diverse and characterized by high inter-annual variability, rather than a clear seasonal pattern. In order to identify the main environmental drivers of community changes, several regional (river runoff, rainfall, upwelling, radiation, daylength) and in situ (sea surface temperature, salinity, bottom and surface chlorophyll a concentration) environmental parameters were tested. Multivariate statistical analysis allowed the identification of water stability as the main environmental gradient influencing the community composition, with river runoff in the preceding rain season and upwelling being the two drivers of stratification and turbulence, respectively. Both these processes can be described as nutrient enrichment processes, but the cyst signal indicates that the two mechanisms select for different functional groups. The main upwelling cyst signal is characterised by the dominance of heterotrophic species (Protoperidinioid species) and presence of the autotrophic chain-forming Gymnodinium catenatum, while the river runoff cyst signal is characterised by dominance of autotrophs forming calcareous cysts, mainly Scrippsiella spp. Lingulodinium polyedrum is suggested to be indicative of upwelling conditions in the region but reflecting an ecological niche different from the more classical heterotrophic assemblage and G. catenatum. Our results reinforce the applicability of dinoflagellate cysts as environmental tracers in the warm-temperate region of the NE Atlantic, and contribute to the development of palaeoenvironmental cyst-based signals. © 2008 Elsevier B.V. All rights reserved. Keywords: Gymnodinium catenatum; Lingulodinium polyedrum; Dinoflagellate cyst signals; Runoff; Upwelling; Water stability 1. Introduction Coastal upwelling regions are within the most productive in the world (Mann, 2000). Phytoplankton succession patterns in these systems are strongly dependent on the physical processes that characterize these regions, namely the upwelling and downwelling cycles, which determine space and time alternation Available online at www.sciencedirect.com Marine Micropaleontology 68 (2008) 156 – 178 www.elsevier.com/locate/marmicro ⁎ Corresponding author. Tel.: +351 217500000x20332; fax: +351 217500009. E-mail addresses: [email protected] (S. Ribeiro), [email protected] (A. Amorim). 1 Currently also at: Department of Biology, Aquatic Biology Section, Faculty of Science, University of Copenhagen, Øster Farimagsgade 2D, DK-1353 Copenhagen K, Denmark; and LNEG - Departamento de Geologia Marinha, Estrada da Portela, Zambujal, 2721–866 Amadora, Portugal. 0377-8398/$ - see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.marmicro.2008.01.013
Transcript of Environmental drivers of temporal succession in recent dinoflagellate cyst assemblages from a...

Available online at www.sciencedirect.com
68 (2008) 156–178www.elsevier.com/locate/marmicro
Marine Micropaleontology
Environmental drivers of temporal succession in recent dinoflagellatecyst assemblages from a coastal site in the North-East Atlantic
(Lisbon Bay, Portugal)
Sofia Ribeiro 1, Ana Amorim ⁎
Instituto de Oceanografia, Faculdade de Ciências da Universidade de Lisboa, Campo Grande 1749-016 Lisboa, Portugal
Received 21 December 2007; accepted 4 January 2008
Abstract
Temporal changes in the community structure of recent dinoflagellate cyst assemblages of Lisbon Bay (Iberian upwellingsystem) were investigated between 2000 and 2005. The assemblages were diverse and characterized by high inter-annualvariability, rather than a clear seasonal pattern. In order to identify the main environmental drivers of community changes, severalregional (river runoff, rainfall, upwelling, radiation, daylength) and in situ (sea surface temperature, salinity, bottom and surfacechlorophyll a concentration) environmental parameters were tested. Multivariate statistical analysis allowed the identification ofwater stability as the main environmental gradient influencing the community composition, with river runoff in the preceding rainseason and upwelling being the two drivers of stratification and turbulence, respectively. Both these processes can be described asnutrient enrichment processes, but the cyst signal indicates that the two mechanisms select for different functional groups. Themain upwelling cyst signal is characterised by the dominance of heterotrophic species (Protoperidinioid species) and presence ofthe autotrophic chain-forming Gymnodinium catenatum, while the river runoff cyst signal is characterised by dominance ofautotrophs forming calcareous cysts, mainly Scrippsiella spp. Lingulodinium polyedrum is suggested to be indicative of upwellingconditions in the region but reflecting an ecological niche different from the more classical heterotrophic assemblage andG. catenatum. Our results reinforce the applicability of dinoflagellate cysts as environmental tracers in the warm-temperate regionof the NE Atlantic, and contribute to the development of palaeoenvironmental cyst-based signals.© 2008 Elsevier B.V. All rights reserved.
Keywords: Gymnodinium catenatum; Lingulodinium polyedrum; Dinoflagellate cyst signals; Runoff; Upwelling; Water stability
⁎ Corresponding author. Tel.: +351 217500000x20332; fax: +351217500009.
E-mail addresses: [email protected] (S. Ribeiro),[email protected] (A. Amorim).1 Currently also at: Department of Biology, Aquatic Biology Section,
Faculty of Science, University of Copenhagen, Øster Farimagsgade2D, DK-1353 Copenhagen K, Denmark; and LNEG - Departamento deGeologia Marinha, Estrada da Portela, Zambujal, 2721–866 Amadora,Portugal.
0377-8398/$ - see front matter © 2008 Elsevier B.V. All rights reserved.doi:10.1016/j.marmicro.2008.01.013
1. Introduction
Coastal upwelling regions are within the mostproductive in the world (Mann, 2000). Phytoplanktonsuccession patterns in these systems are stronglydependent on the physical processes that characterizethese regions, namely the upwelling and downwellingcycles, which determine space and time alternation

157S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
between well-mixed nutrient replete waters and strati-fied relatively poor waters (Margalef, 1978).
Dinoflagellates are a diverse group of protists thatalong with diatoms and coccolithophors are one of themost important and abundant groups of the marinemicrophytoplankton. It is generally assumed that abouthalf of the species are phototrophic, the other half beingheterotrophs (Schnepf and Elbrächter, 1992). However,both these groups may show alternative diverse andcomplex nutritional strategies (e.g. Stoecker, 1999).Manydinoflagellates are meroplanktonic, producing a benthicresting stage or cyst as part of their life cycle (Dale, 1983).This structure is the only known fossilizable stage indinoflagellates, and studies on their present-day occur-rence and distribution may provide valuable informationto micropalaeontology on the ecology, biogeography, andenvironmental optima of species (Dale and Dale, 2002).
Several authors have studied the spatial distributionand abundance of dinoflagellate cysts in relation topresent-day sea surface conditions (e.g. Wall et al., 1977,Dale, 1996, Rochon et al., 1999) and identifiedenvironmental cyst signals that can be used to interpretthe fossil record. The main environmental cyst signalsrecognized so far include the coastal/oceanic transition,the climatic signal, the salinity signal, and the produc-tivity signal (Dale, 1996; de Vernal et al., 2001).
Sediment traps have been used since approximatelythe last decade to study the timing of in situ cystproduction and quantify cyst fluxes at short andecologically relevant time-scales (e.g. Ishikawa andTaniguchi, 1996, 1997; Montresor et al., 1998; Joyceand Pitcher, 2004, Morquecho and Lechuga-Devéze,2004). In general, sediment trap work has highlightedthe existence of species-specific strategies of cystproduction, with some species producing cysts overseveral months (e.g. Scrippsiella trochoidea in the Gulfof Naples, Montresor et al., 1998) and others only overrestricted periods of time (e.g. Zygabikodinium lenticu-latum in the Benguela upwelling system, Joyce andPitcher, 2004). Regarding cyst germination and bloomformation, highly variable species-specific strategies,largely determined by the temporal and spatial fluctua-tions of environmental conditions have also beenidentified. Ishikawa and Taniguchi (1997) proposed“sporadic” and “synchronous” as the two main types ofcyst germination strategies found in Onagawa Bay,Japan, although several sub-types were also identified.
Regarding environmental drivers of change at thecommunity level, Morquecho and Lechuga-Devéze(2004) have identified temperature as the main environ-mental driver of species occurrence in the Gulf ofCalifornia, where transitional hydrographic conditions
in spring and early fall largely determine the seasonalabundance of meroplanktonic dinoflagellates.
Temporal changes in cyst assemblages have alsobeen investigated through the analysis of surfacesediment samples by Harland et al. (2004) and Wanget al. (2004). Both authors identified temporal variationson the composition of cyst assemblages and highlightedcomplex interactions with the environmental conditions.Harland et al. (2004) stressed out the importance ofwater stability and nutrient availability in determiningseasonal cyst production patterns. These results high-light the complexity and unpredictability of ecologicalprocesses associated with cyst production patterns andstress the need for long-term surveys in order to includethe representative temporal scales of variation at theindividual, population and community levels.
Here, we present results from a long-term cyst surveyconducted between 2000 and 2005, at a coastal site inthe NE Atlantic under the influence of seasonalupwelling and river runoff (Lisbon Bay). The temporalvariations on the composition of recently formed cystassemblages is described and discussed in relation to theregional environmental context. Results are expected tocontribute to a better understanding of present-daydinoflagellate cyst ecology and to improve the power ofactualistic methods, allowing more realistic assumptionsto be made from the fossil record.
2. Regional setting
The west coast of Portugal is located at the easternmargin of the North Atlantic Ocean. Seasonal upwellingoccurs when the intensity and steadiness of northerlywinds are favourable, reaching its peak from July toSeptember (Fiúza et al., 1982). During upwellingevents, surface waters are transported offshore andreplaced by deeper, colder and more nutrient-richwaters, enhancing biological productivity (Woosteret al., 1976). Upwelling has been identified as themajor source of seasonal and spatial phytoplanktonvariability along the coast of Portugal (Moita, 2001).
At major topographic features, in particular capes andsubmarine canyons, cold-water filaments can extendoffshore, for hundreds of kilometres, as the upwellingseason progresses (e.g. Sousa and Bricaud, 1992).
Lisbon Bay (Fig. 1) is located south of a majorcoastal discontinuity, the Estremadura Promontory. Itshydrography is highly influenced by the coastalmorphology, bottom topography (Lisbon and Setúbalcanyons) and the input of freshwater from the TagusRiver. The latter is the longest river in the IberianPeninsula with an estimated annual runoff of 14 km3

Fig. 1. Map of the study area showing sampling site at Cascais marina (✳). Bathymetric lines from 100 to 1000 m depth are also represented.
158 S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
(OSPAR, 2000). Water resources in the Tagus basin arehighly variable, with drought periods and floodsoccurring frequently (Benito et al., 2003). More than2.5 million people live in the area of Lisbon, andpartially treated effluents still drain into the river estuary,contributing largely to the amount of dissolved andparticulate material that reaches the coastal zone.
During the upwelling season, a recurrent upwellingplume, extending southward or westward, can be rootedat Cape Roca in the north side of the Bay (Sousa andBricaud, 1992). Phytoplankton studies coupled withstudies on the physical mechanisms associated with thisplume have shown that the leeward side of the plumemay function as a retention area for the development ofphytoplankton, particularly of the chain-forming dino-flagellateGymnodinium catenatum (Moita and Amorim,2001, Moita et al., 2003).
The selected sampling site is an artificial harbourbuilt in 1999 at the northern limit of Lisbon Bay (38°42′N; 09°25′W), located close to the root of the Cape Rocaupwelling plume. This site represents the main sedi-mentary basin in the study region, and it is expected towork as a large sediment trap in an otherwise highlyhydrodynamic open-coast.
3. Materials and methods
3.1. Sample collection and preparation
Twenty-three surface sediment samples were col-lected (Fig. 1) between January 2000 and November2005, using a manual vacuum pump sampler thatallows retrieval of the surface sediment and adjacentflocculent layer (ca. 0–2 cm) (Dale and Dale, 2002,Fig. 10.4).
To assure no cyst germination occurred, samples werekept in the dark at 4 °C and processed within 24 h aftercollection. No chemical treatment was applied to thesediment samples, in order to obtain the whole cystassemblage. Sample preparation involved sonication(1 min) in an ultra-sonic bath, and wet sieving through150 µm onto a 25 µm-mesh calibrated sieve (Ende-cotts™) using alkaline tap water to avoid dissolution ofcalcareous cyst walls. The fraction remaining on the25 μm sieve was washed into a 15 ml centrifuge tube andcentrifuged (2500 rpm, 5 min). To further concentratedinoflagellate cysts, the pellet was re-suspended in a high-density solution of sodium metatungstate (2.016 g cm−3)(Alfa Aesar® ref. 022534) and centrifuged for 15 min at

Table 1Dinoflagellate cyst taxa (theca and cyst-based names) and codes used for abbreviation in data analysis
Theca-based name (biological) Cyst-based name (palaeontological) Code
Order GonyaulacalesAlexandrium spp. rGonyaulax cf. spinifera Spiniferites ramosus SpinN40 µmGonyaulax membranacea Spiniferites membranaceus SpinN40 µmGonyaulax scrippsae Spiniferites bulloideus SbulGonyaulax sp. Spiniferites delicatus SdelGonyaulax sp. indet. Impagidinium spp. rGonyaulax spinifera Nematosphaeropsis labirinthus rGonyaulax spinifera Spiniferites mirabilis/hyperacanthus Smir/hypGonyaulax spinifera complex Achomosphaera spp. AsppGonyaulax spinifera complex Achomosphaera ramulifera AsppGonyaulax spinifera complex Spiniferites sppb40 µm Spinb40 µmGonyaulax spinifera complex Spiniferites sppN40 µm SpinN40 µmGonyaulax verior rLingulodinium polyedrum Lingulodinium machaerophorum LpolProtoceratium reticulatum Operculodinium centrocarpum Pretcf. Protoceratium reticulatum cf. Operculodinium centrocarpum Pretcf. Protoceratium reticulatum Operculodinium cf. israelianum rPyrodinium bahamense Polysphaeridium zoharii r
Order GymnodinialesGymnodinium catenatum Gcatcf Gymnodinium microreticulatum rGymnodinium cf. impudicum rGymnodinioid sp.1 Gym sp1Polykrikos schwartzii PschPolykrikos cf. kofoidii rcf Warnowia rosea r
Order PeridinialesCT196 CT196CTA CTA
cf Ensiculifera imarinensis rDiplopelta parva DparDiplopelta symmetrica rDiplopsalids group Dubridinium ulesterum DuleDiplopsalids group Dubridinium spp. DsppDiplopsalis lenticula Dlen
Echinidinium cf. aculeatum EchsppEchinidinium cf. granulatum rEchinidinium spp. EchsppIslandinium sp.1 r
Protoperidinium americanum PameProtoperidinium avell/punct Brigatedinium cf. cariacoense Pav/puProtoperidinium cf. nudum Selenopemphix quanta PconProtoperidinium claudicans Votadinium spinosum rProtoperidinium conicum b55 µm Selenopemphix quanta PconProtoperidinium conicum N55 µm Selenopemphix quanta PconProtoperidinium divaricatum Xandarodinium xanthum PdivProtoperidinium leonis L. sabrina/Q. concreta PleoProtoperidinium oblongum Votadinium calvum PoblProtoperidinium pentagonum Trinovantedinium applanatum Ppen
Islandinium spp. IsppProtoperidinium spp. Lejeunecysta/Quinquecuspis/Votadinium PsppProtoperidinium spp. Bringatedinium spp. RBProtoperidinium subinerme Selenopemphix nephroides PsubScrippsiella spp. Scrspp
(continued on next page)
159S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178

Table 1 (continued)
Theca-based name (biological) Cyst-based name (palaeontological) Code
Order PeridinialesScrippsiella trochoidea Stro
RBS spp. RBSRBS sp1 RBSsp1
Zygabikodinium lenticulatum Dubridinium caperatum Dcap
Some morphotypes were grouped together under the same code. The code “r” refers to rare cysts excluded from the statistical analysis.
160 S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
1000 rpm (Amorim and Dale, 1998, modified fromMunsterman and Kerstholt, 1996 and Bolch, 1997). Theorganic fraction, floated off by the heavy liquid, wascarefully pipetted, rinsed with tap water through sieving(25 μm) into a 15 ml centrifuge tube and centrifuged(2500 rpm, 5min). Aliquots of the pellet weremounted onmicroscope slides with glycerine jelly, and sealed withparaffin wax.
3.2. Composition of assemblages
Whenever possible, more than 300 cysts werecounted, to assure good statistical power. Slides wereexamined under the light microscope, with differentialinterference contrast (DIC) (Zeiss Axioskop and Olym-pus BX50). Microphotographs were taken with a ZeissAxioVision camera. Counting included both empty cystsand cysts with cell contents. Results are expressed asrelative abundances (%) of the whole cyst assemblages.Identification of dinoflagellate cysts followed Wall andDale (1968), Zonneveld (1997), Rochon et al. (1999),and Head et al. (2001), among others. In accordance tothe nature of this study, nomenclature based on themotile stage was preferred, except when cyst-basednames could better distinguish the different cystmorphotypes encountered. Cyst-theca relationships con-form to Head (1996), and Ellegaard et al. (2003).
Several descriptors were used to characterize the cystassemblages, and their temporal variations: (1) Composi-tion of the assemblages— cyst morphotypes; (2) Speciesrichness; (3) Species diversity and evenness; (4) Ratioof autotrophic to heterotrophic taxa; and (5) Proportionof empty cysts to cysts with cell contents. SpeciesRichness, Diversity and Evenness were estimated basedon all different cyst morphotypes. Shannon–Wiener'sDiversity index (H') and Shannon's Equitability (E)were used (Shannon and Weaver, 1949).
3.3. Excystment experiments
Excystment experiments were conducted, when-ever possible, to establish unknown cyst-motile stagerelationships. Samples were sieved, centrifuged in a high-
density solution of sodiummetatungstate (2.016 g cm−3),and screened for viable cysts belonging to the target groupor species. The cysts were then isolated with a micropip-ette into tissue culture plates (Nunc™ 176740) filled withsterilised seawater from the sampling site. Plates wereincubated at 19 °C under 14 h: 10 h, L:D cycle at 54 µmolphotons m−2 s−1. When germination occurred, motilecells and cysts were described, photographed, andeventually transferred to growth medium (GSe, salinity35) (Doblin et al., 1999).
3.4. Environmental data
Between May 2001 and December 2005, weeklymeasurements of in situ sea surface temperature (SST)and sea surface salinity (SSS) (portable sonar YSI), anddetermination of bottom (bChla) and surface (sChla)chlorophyll a concentrations (Nansen bottle) wereperformed at a fixed point in the main entrance channelof the marina, one hour before high-tide, to assure astrong oceanic influence (Palma, 2003) (data kindlyprovided by the Portuguese Fisheries Institute, IPI-MAR). In addition, the following regional environ-mental parameters were available for the whole studyperiod (2000–2005): upwelling, Tagus river runoff,photosynthetic active radiation (PAR), rainfall anddaylength. The upwelling index (UI) was calculatedaccording to Bakun (1973), based on the N–Scomponent of the wind tension at the closest meteor-ological station (Cape Carvoeiro) (data kindly providedby IPIMAR). Tagus river runoff, PAR and rainfall wereobtained from the Portuguese Water Institute (INAG),and daylength data from the Astronomical Observatoryof the University of Lisbon (OAL).
3.5. Statistical approach
As a first approach, cluster analyses were carried outboth for samples and main cyst morphotypes, using thehierarchical agglomerative Unweighted Pair GroupMethod with Arithmetic Mean (UPGMA) (Sneath andSokal, 1973). UPGMA is a straightforward methodof tree construction. It uses a sequential clustering

161S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
algorithm, in which local homology between units(species or samples) is identified in order of similarity,and the tree is built in a stepwise manner. Clusteranalyses were performed using STATISTICA 7.0. Theschematic representation of these results followed Daleand Dale (2002).
3.5.1. Multivariate analysisOrdinationmethods were performed using CANOCO
for Windows 4.5 (Ter Braak and Smilauer, 2002). Somecyst types were grouped together based on theirmorphological similarities (Table 1). Cyst types withlow representation (b1%) in all samples were excludedfrom the multivariate analysis, to avoid the effect of veryrare species. No logarithmic transformation of data waschosen, because this study originated compositional datacontaining many zeros (Ter Braak and Smilauer, 2002).
As a first approach to determine the nature of therelationship (unimodal or linear) between dinocysts andthe environmental parameters measured, DetrendedCorrespondence Analysis (DCA) was used. CCA waschosen afterwards, in order to identify and test thesignificance of environmental gradients influencing theassemblages. CCA was carried for samples collectedafter November 2001 (n=17), for which data on allenvironmental variables was available.
For multivariate analysis, averages of the environ-mental parameters were calculated for different time-periods (1, 2, 3 and 6 months prior to the samplingdate). A time-lag of 1 month relative to the samplingday was also tested. This approach was chosen basedon the known temporal discrepancy between cystproduction in the water column and cyst deposition inthe sediments (e.g. Montresor et al., 1998; Amorimet al., 2001a), and as an attempt to find the mostrelevant time-frame of influence for each explanatoryenvironmental variable.
The statistical significance of all environmentalvariables was tested by unrestricted Monte Carlopermutation tests (forward selection), based on 499unrestricted permutations. A global permutation test wasused to determine the significance of the first ordinationaxis and of all canonical axes together. Correlationbetween all the statistically significant environmentalvariables was also tested using STATISTICA 7.0.
4. Results
4.1. Taxonomic notes
The designation RB (Round Brown) was created toinclude spherical brown protoperidinioid cysts that
could not be identified due to unsuitable orientation orfragmentation. Protoperidinioids with apical and anta-pical horns not identifiable to the species level, weregrouped as Protoperidinium spp. Cysts with unknownaffinity, characterised by having an organic colourlesswall, highly refractive when cell contents were present,with no signs of paratabulation, and frequently envel-oped by a mucous layer were grouped as UMC(Unidentified Mucous Cysts). These cysts often pre-sented a red-orange pigment body. Successful excyst-ment experiments frequently resulted in motile stagesbelonging to the genus Scrippsiella. UMC was, there-fore, considered an autotrophic group. Round brownspiny cysts, which could not be identified, were groupedas RBS. These cysts are likely to belong to the cyst-based genera Islandinium and Echinidinium. All thesuccessful excystment experiments gave heterotrophiccells with biological affinities to diplopsalids and thegenus Protoperidinium. RBS were, therefore, consid-ered a heterotrophic group.
4.2. Dinoflagellate cyst assemblages
Rich dinoflagellate cyst assemblages were recoveredfrom all samples. A minimum of 212 and a maximum of574 cysts were counted (mean=356). Fifty-eight cysttypes were identified and the majority could be assignedto 21 genera belonging to three orders (Gonyaulacales,Gymnodiniales and Peridiniales) (Table 1).
Species richness was high in all samples, with aminimum of 24 cyst types (September 2000) and amaximum of 39 (September 2003), averaging 31(Table 2). Shannon–Wiener's diversity index (H')ranged from 3.0 to 4.5, indicating high complexitywithin the assemblages. Values between 3.0 and 3.6could be associated with dominance by particular taxa.Between March and September 2003 the low H' wasrelated to high numbers of Scrippsiella spp., whilebetween July and November 2005 diplopsalids were thedominant group. This was also evidenced by lowervalues in the Shannon's equitability index (Evennessindex) (Table 2).
The ratio between cysts produced by autotrophic andheterotrophic dinoflagellate species (A:H ratio) showsthat heterotrophs were dominant with the exceptionof 2003 and January 2000 (Table 2). The fractionof cysts with cell contents in the assemblage variedbetween 17 and 69% (Table 2, Fig. 2). The changes inthe relative abundance of cysts with cell contents wereused as indicative of timing of cyst production. Severalirregular pulses of cyst production could be identifiedwhich reflected different species-specific strategies of
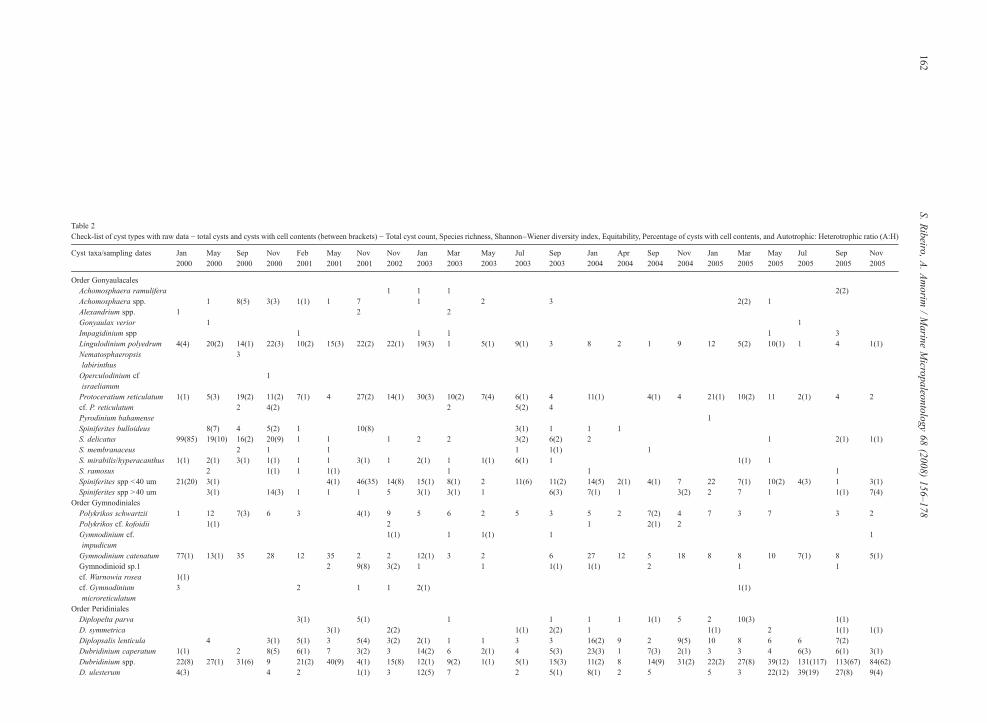
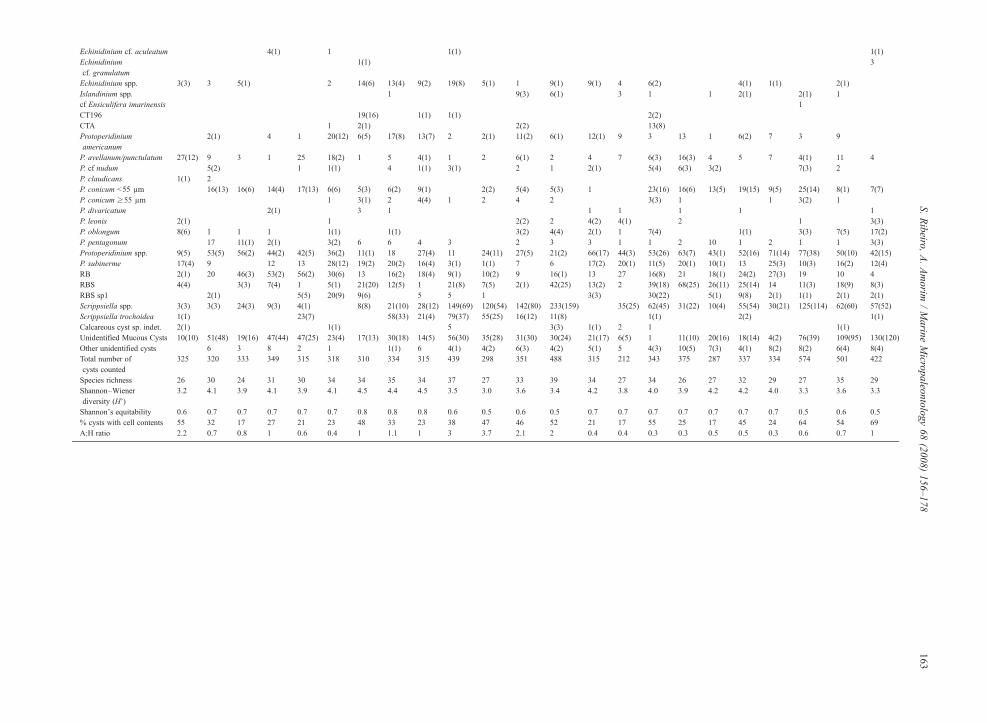
Table 2Check-list of cyst types with raw data − total cysts and cysts with cell contents (between brackets) − Total cyst count, Species richness, Shannon–Wiener diversity index, Equitability, Percentage of cysts with cell contents, and Autotrophic: Heterotrophic ratio (A:H)
Cyst taxa/sampling dates Jan2000
May2000
Sep2000
Nov2000
Feb2001
May2001
Nov2001
Nov2002
Jan2003
Mar2003
May2003
Jul2003
Sep2003
Jan2004
Apr2004
Sep2004
Nov2004
Jan2005
Mar2005
May2005
Jul2005
Sep2005
Nov2005
Order GonyaulacalesAchomosphaera ramulifera 1 1 1 2(2)Achomosphaera spp. 1 8(5) 3(3) 1(1) 1 7 1 2 3 2(2) 1Alexandrium spp. 1 2 2Gonyaulax verior 1 1Impagidinium spp 1 1 1 1 3Lingulodinium polyedrum 4(4) 20(2) 14(1) 22(3) 10(2) 15(3) 22(2) 22(1) 19(3) 1 5(1) 9(1) 3 8 2 1 9 12 5(2) 10(1) 1 4 1(1)Nematosphaeropsislabirinthus
3
Operculodinium cfisraelianum
1
Protoceratium reticulatum 1(1) 5(3) 19(2) 11(2) 7(1) 4 27(2) 14(1) 30(3) 10(2) 7(4) 6(1) 4 11(1) 4(1) 4 21(1) 10(2) 11 2(1) 4 2cf. P. reticulatum 2 4(2) 2 5(2) 4Pyrodinium bahamense 1Spiniferites bulloideus 8(7) 4 5(2) 1 10(8) 3(1) 1 1 1S. delicatus 99(85) 19(10) 16(2) 20(9) 1 1 1 2 2 3(2) 6(2) 2 1 2(1) 1(1)S. membranaceus 2 1 1 1 1(1) 1S. mirabilis/hyperacanthus 1(1) 2(1) 3(1) 1(1) 1 1 3(1) 1 2(1) 1 1(1) 6(1) 1 1(1) 1S. ramosus 2 1(1) 1 1(1) 1 1 1Spiniferites spp b40 um 21(20) 3(1) 4(1) 46(35) 14(8) 15(1) 8(1) 2 11(6) 11(2) 14(5) 2(1) 4(1) 7 22 7(1) 10(2) 4(3) 1 3(1)Spiniferites spp N40 um 3(1) 14(3) 1 1 1 5 3(1) 3(1) 1 6(3) 7(1) 1 3(2) 2 7 1 1(1) 7(4)
Order GymnodinialesPolykrikos schwartzii 1 12 7(3) 6 3 4(1) 9 5 6 2 5 3 5 2 7(2) 4 7 3 7 3 2Polykrikos cf. kofoidii 1(1) 2 1 2(1) 2Gymnodinium cf.impudicum
1(1) 1 1(1) 1 1
Gymnodinium catenatum 77(1) 13(1) 35 28 12 35 2 2 12(1) 3 2 6 27 12 5 18 8 8 10 7(1) 8 5(1)Gymnodinioid sp.1 2 9(8) 3(2) 1 1 1(1) 1(1) 2 1 1cf. Warnowia rosea 1(1)cf. Gymnodiniummicroreticulatum
3 2 1 1 2(1) 1(1)
Order PeridinialesDiplopelta parva 3(1) 5(1) 1 1 1 1 1(1) 5 2 10(3) 1(1)D. symmetrica 3(1) 2(2) 1(1) 2(2) 1 1(1) 2 1(1) 1(1)Diplopsalis lenticula 4 3(1) 5(1) 3 5(4) 3(2) 2(1) 1 1 3 3 16(2) 9 2 9(5) 10 8 6 6 7(2)Dubridinium caperatum 1(1) 2 8(5) 6(1) 7 3(2) 3 14(2) 6 2(1) 4 5(3) 23(3) 1 7(3) 2(1) 3 3 4 6(3) 6(1) 3(1)Dubridinium spp. 22(8) 27(1) 31(6) 9 21(2) 40(9) 4(1) 15(8) 12(1) 9(2) 1(1) 5(1) 15(3) 11(2) 8 14(9) 31(2) 22(2) 27(8) 39(12) 131(117) 113(67) 84(62)D. ulesterum 4(3) 4 2 1(1) 3 12(5) 7 2 5(1) 8(1) 2 5 5 3 22(12) 39(19) 27(8) 9(4)
162S.
Ribeiro,
A.Amorim
/Marine
Micropaleontology
68(2008)
156–178
Echinidinium cf. aculeatum 4(1) 1 1(1) 1(1)Echinidiniumcf. granulatum
1(1) 3
Echinidinium spp. 3(3) 3 5(1) 2 14(6) 13(4) 9(2) 19(8) 5(1) 1 9(1) 9(1) 4 6(2) 4(1) 1(1) 2(1)Islandinium spp. 1 9(3) 6(1) 3 1 1 2(1) 2(1) 1cf Ensiculifera imarinensis 1CT196 19(16) 1(1) 1(1) 2(2)CTA 1 2(1) 2(2) 13(8)Protoperidiniumamericanum
2(1) 4 1 20(12) 6(5) 17(8) 13(7) 2 2(1) 11(2) 6(1) 12(1) 9 3 13 1 6(2) 7 3 9
P. avellanum/punctulatum 27(12) 9 3 1 25 18(2) 1 5 4(1) 1 2 6(1) 2 4 7 6(3) 16(3) 4 5 7 4(1) 11 4P. cf nudum 5(2) 1 1(1) 4 1(1) 3(1) 2 1 2(1) 5(4) 6(3) 3(2) 7(3) 2P. claudicans 1(1) 2P. conicum b55 µm 16(13) 16(6) 14(4) 17(13) 6(6) 5(3) 6(2) 9(1) 2(2) 5(4) 5(3) 1 23(16) 16(6) 13(5) 19(15) 9(5) 25(14) 8(1) 7(7)P. conicum ≥55 µm 1 3(1) 2 4(4) 1 2 4 2 3(3) 1 1 3(2) 1P. divaricatum 2(1) 3 1 1 1 1 1 1P. leonis 2(1) 1 2(2) 2 4(2) 4(1) 2 1 3(3)P. oblongum 8(6) 1 1 1 1(1) 1(1) 3(2) 4(4) 2(1) 1 7(4) 1(1) 3(3) 7(5) 17(2)P. pentagonum 17 11(1) 2(1) 3(2) 6 6 4 3 2 3 3 1 1 2 10 1 2 1 1 3(3)Protoperidinium spp. 9(5) 53(5) 56(2) 44(2) 42(5) 36(2) 11(1) 18 27(4) 11 24(11) 27(5) 21(2) 66(17) 44(3) 53(26) 63(7) 43(1) 52(16) 71(14) 77(38) 50(10) 42(15)P. subinerme 17(4) 9 12 13 28(12) 19(2) 20(2) 16(4) 3(1) 1(1) 7 6 17(2) 20(1) 11(5) 20(1) 10(1) 13 25(3) 10(3) 16(2) 12(4)RB 2(1) 20 46(3) 53(2) 56(2) 30(6) 13 16(2) 18(4) 9(1) 10(2) 9 16(1) 13 27 16(8) 21 18(1) 24(2) 27(3) 19 10 4RBS 4(4) 3(3) 7(4) 1 5(1) 21(20) 12(5) 1 21(8) 7(5) 2(1) 42(25) 13(2) 2 39(18) 68(25) 26(11) 25(14) 14 11(3) 18(9) 8(3)RBS sp1 2(1) 5(5) 20(9) 9(6) 5 5 1 3(3) 30(22) 5(1) 9(8) 2(1) 1(1) 2(1) 2(1)Scrippsiella spp. 3(3) 3(3) 24(3) 9(3) 4(1) 8(8) 21(10) 28(12) 149(69) 120(54) 142(80) 233(159) 35(25) 62(45) 31(22) 10(4) 55(54) 30(21) 125(114) 62(60) 57(52)Scrippsiella trochoidea 1(1) 23(7) 58(33) 21(4) 79(37) 55(25) 16(12) 11(8) 1(1) 2(2) 1(1)Calcareous cyst sp. indet. 2(1) 1(1) 5 3(3) 1(1) 2 1 1(1)Unidentified Mucous Cysts 10(10) 51(48) 19(16) 47(44) 47(25) 23(4) 17(13) 30(18) 14(5) 56(30) 35(28) 31(30) 30(24) 21(17) 6(5) 1 11(10) 20(16) 18(14) 4(2) 76(39) 109(95) 130(120)Other unidentified cysts 6 3 8 2 1 1(1) 6 4(1) 4(2) 6(3) 4(2) 5(1) 5 4(3) 10(5) 7(3) 4(1) 8(2) 8(2) 6(4) 8(4)Total number ofcysts counted
325 320 333 349 315 318 310 334 315 439 298 351 488 315 212 343 375 287 337 334 574 501 422
Species richness 26 30 24 31 30 34 34 35 34 37 27 33 39 34 27 34 26 27 32 29 27 35 29Shannon–Wienerdiversity (H')
3.2 4.1 3.9 4.1 3.9 4.1 4.5 4.4 4.5 3.5 3.0 3.6 3.4 4.2 3.8 4.0 3.9 4.2 4.2 4.0 3.3 3.6 3.3
Shannon's equitability 0.6 0.7 0.7 0.7 0.7 0.7 0.8 0.8 0.8 0.6 0.5 0.6 0.5 0.7 0.7 0.7 0.7 0.7 0.7 0.7 0.5 0.6 0.5% cysts with cell contents 55 32 17 27 21 23 48 33 23 38 47 46 52 21 17 55 25 17 45 24 64 54 69A:H ratio 2.2 0.7 0.8 1 0.6 0.4 1 1.1 1 3 3.7 2.1 2 0.4 0.4 0.3 0.3 0.5 0.5 0.3 0.6 0.7 1
163S.
Ribeiro,
A.Amorim
/Marine
Micropaleontology
68(2008)
156–178
164 S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
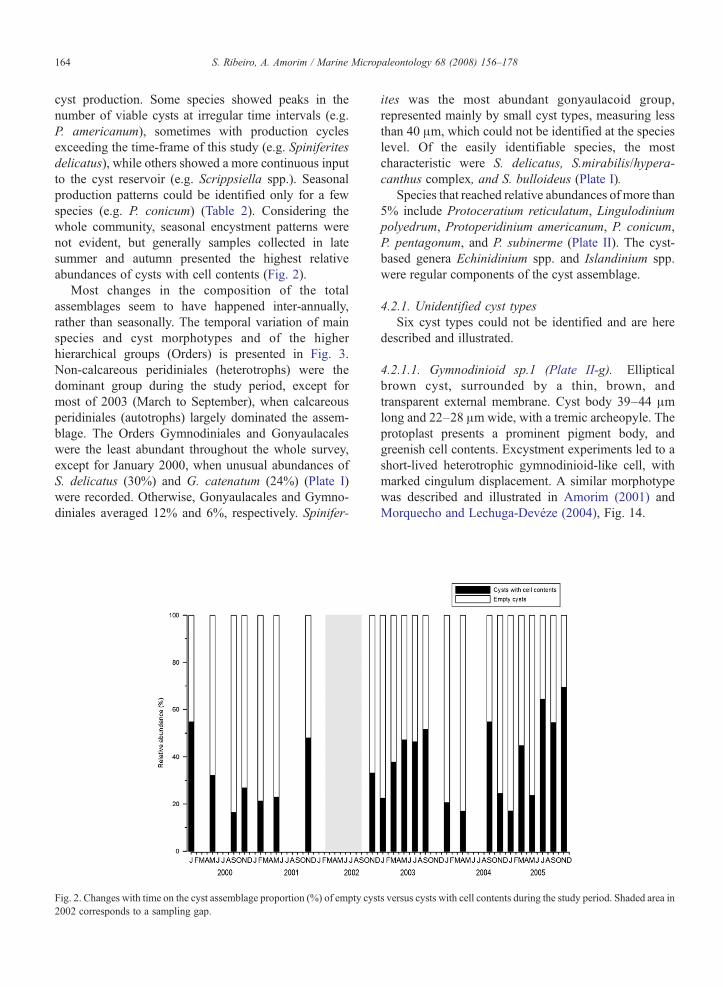
cyst production. Some species showed peaks in thenumber of viable cysts at irregular time intervals (e.g.P. americanum), sometimes with production cyclesexceeding the time-frame of this study (e.g. Spiniferitesdelicatus), while others showed a more continuous inputto the cyst reservoir (e.g. Scrippsiella spp.). Seasonalproduction patterns could be identified only for a fewspecies (e.g. P. conicum) (Table 2). Considering thewhole community, seasonal encystment patterns werenot evident, but generally samples collected in latesummer and autumn presented the highest relativeabundances of cysts with cell contents (Fig. 2).
Most changes in the composition of the totalassemblages seem to have happened inter-annually,rather than seasonally. The temporal variation of mainspecies and cyst morphotypes and of the higherhierarchical groups (Orders) is presented in Fig. 3.Non-calcareous peridiniales (heterotrophs) were thedominant group during the study period, except formost of 2003 (March to September), when calcareousperidiniales (autotrophs) largely dominated the assem-blage. The Orders Gymnodiniales and Gonyaulacaleswere the least abundant throughout the whole survey,except for January 2000, when unusual abundances ofS. delicatus (30%) and G. catenatum (24%) (Plate I)were recorded. Otherwise, Gonyaulacales and Gymno-diniales averaged 12% and 6%, respectively. Spinifer-
Fig. 2. Changes with time on the cyst assemblage proportion (%) of empty cys2002 corresponds to a sampling gap.
ites was the most abundant gonyaulacoid group,represented mainly by small cyst types, measuring lessthan 40 μm, which could not be identified at the specieslevel. Of the easily identifiable species, the mostcharacteristic were S. delicatus, S.mirabilis/hypera-canthus complex, and S. bulloideus (Plate I).
Species that reached relative abundances of more than5% include Protoceratium reticulatum, Lingulodiniumpolyedrum, Protoperidinium americanum, P. conicum,P. pentagonum, and P. subinerme (Plate II). The cyst-based genera Echinidinium spp. and Islandinium spp.were regular components of the cyst assemblage.
4.2.1. Unidentified cyst typesSix cyst types could not be identified and are here
described and illustrated.
4.2.1.1. Gymnodinioid sp.1 (Plate II-g). Ellipticalbrown cyst, surrounded by a thin, brown, andtransparent external membrane. Cyst body 39–44 μmlong and 22–28 μm wide, with a tremic archeopyle. Theprotoplast presents a prominent pigment body, andgreenish cell contents. Excystment experiments led to ashort-lived heterotrophic gymnodinioid-like cell, withmarked cingulum displacement. A similar morphotypewas described and illustrated in Amorim (2001) andMorquecho and Lechuga-Devéze (2004), Fig. 14.
ts versus cysts with cell contents during the study period. Shaded area in
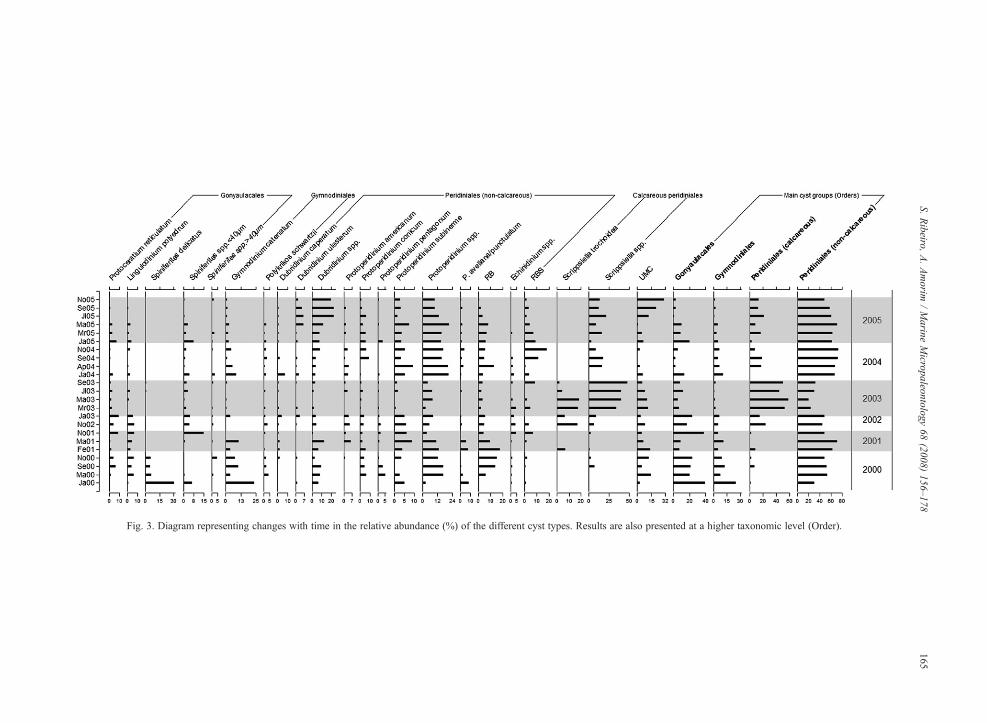
Fig. 3. Diagram representing changes with time in the relative abundance (%) of the different cyst types. Results are also presented at a higher taxonomic level (Order).
165S.
Ribeiro,
A.Amorim
/Marine
Micropaleontology
68(2008)
156–178
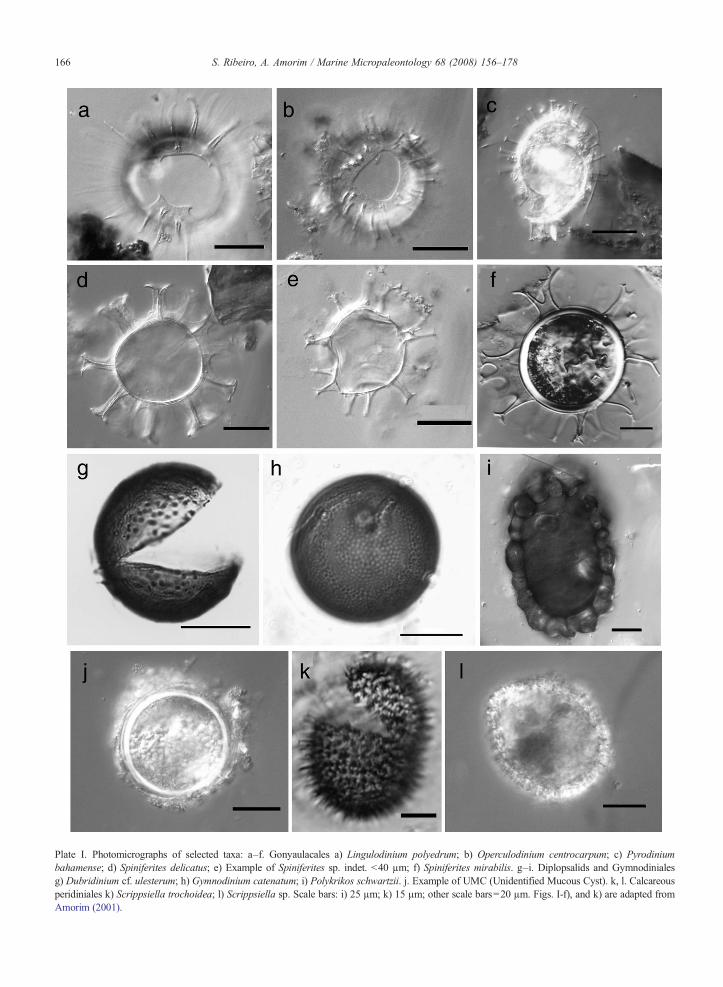
Plate I. Photomicrographs of selected taxa: a–f. Gonyaulacales a) Lingulodinium polyedrum; b) Operculodinium centrocarpum; c) Pyrodiniumbahamense; d) Spiniferites delicatus; e) Example of Spiniferites sp. indet. b40 µm; f) Spiniferites mirabilis. g–i. Diplopsalids and Gymnodinialesg) Dubridinium cf. ulesterum; h) Gymnodinium catenatum; i) Polykrikos schwartzii. j. Example of UMC (Unidentified Mucous Cyst). k, l. Calcareousperidiniales k) Scrippsiella trochoidea; l) Scrippsiella sp. Scale bars: i) 25 µm; k) 15 µm; other scale bars=20 µm. Figs. I-f), and k) are adapted fromAmorim (2001).
166 S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
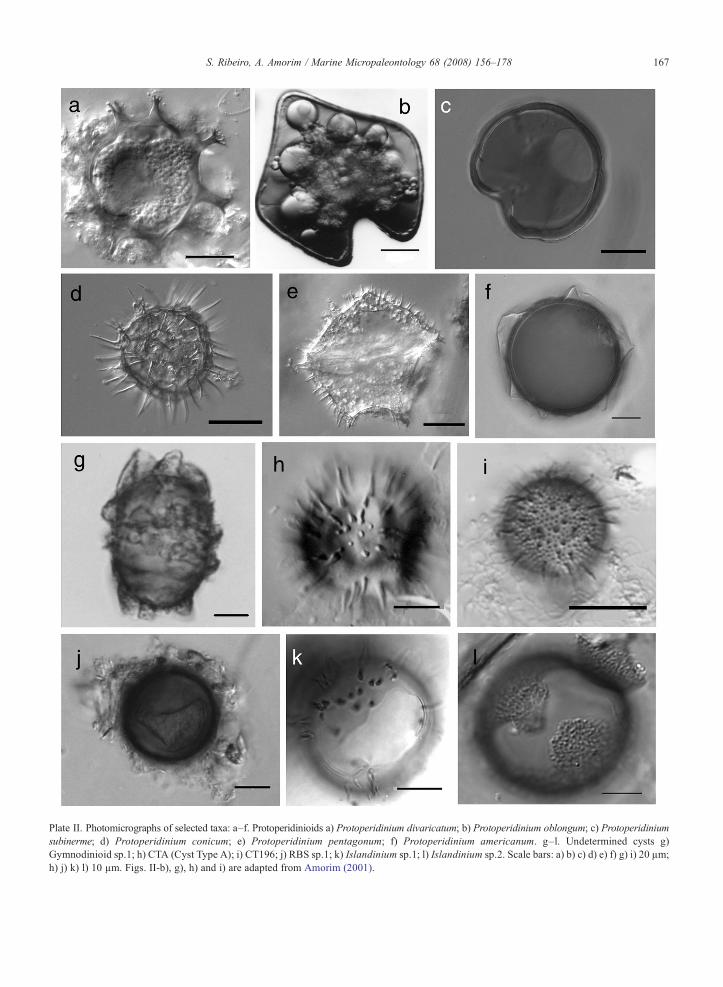
Plate II. Photomicrographs of selected taxa: a–f. Protoperidinioids a) Protoperidinium divaricatum; b) Protoperidinium oblongum; c) Protoperidiniumsubinerme; d) Protoperidinium conicum; e) Protoperidinium pentagonum; f) Protoperidinium americanum. g–l. Undetermined cysts g)Gymnodinioid sp.1; h) CTA (Cyst Type A); i) CT196; j) RBS sp.1; k) Islandinium sp.1; l) Islandinium sp.2. Scale bars: a) b) c) d) e) f) g) i) 20 µm;h) j) k) l) 10 µm. Figs. II-b), g), h) and i) are adapted from Amorim (2001).
167S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
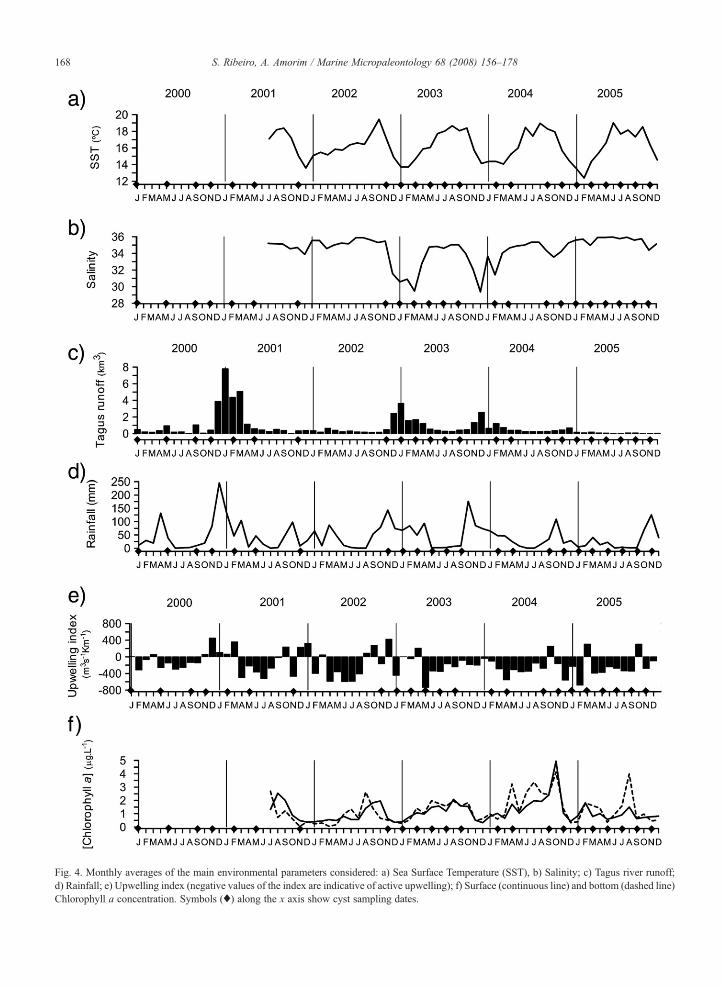
Fig. 4. Monthly averages of the main environmental parameters considered: a) Sea Surface Temperature (SST), b) Salinity; c) Tagus river runoff;d) Rainfall; e) Upwelling index (negative values of the index are indicative of active upwelling); f) Surface (continuous line) and bottom (dashed line)Chlorophyll a concentration. Symbols (♦) along the x axis show cyst sampling dates.
168 S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178

169S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
4.2.1.2. CTA (Cyst type A) (Plate II-h). Spherical lightbrown cyst, ca. 30 μm in body diameter, with numerousrigid conical spines (5–7 μm in length). Spines are notevenly distributed, usually occurring in small groups(2–3). The cyst wall shows no ornamentation. No cystswith visible archeopyle were found. This cyst type wasfirst recorded along the Portuguese coast by Amorim(2001).
4.2.1.3. CT196 (Cyst type 196) (Plate II-i). Smallspherical brown cyst, with long and short spinesmeasuring ca. 7.5 μm and 2.5 μm, respectively. Thecyst body diameter is ca. 26 μm. The shorter pro-cesses may resemble verrucae at high focus. The nameCT196 is used in agreement with the designationused by B. Dale for similar specimens (personalcommunication).
Fig. 5. Cluster diagrams of main cyst types and samples, plotted together with a sthe code presented in Table 1, except for: Dip: Diplopsalids group; Cal: CalcaSpin: Spiniferites spp. The scale indicates 1-Pearson's product moment correla
4.2.1.4. RBS sp.1 (Plate II-j). Spherical brown cystwith very short processes (≈1 μm), hardly distinguish-able at magnifications lower than 1000×. Cyst bodydiameter ca. 27 μm. The archeopyle was neverobserved.
4.2.1.5. Islandinium sp.1 (Plate II-k). Sphericalbrown cyst with numerous processes, 4–6 μm in length,and a plate-like archeopyle. Cyst body diameter ca.29 μm. Smooth wall surface with no visible ornamenta-tion. Processes conical and hollow, not evenly dis-tributed, forming groups/aggregates. During this survey,only one specimen was observed, in November 2002.
4.2.1.6. Islandinium sp.2 (Plate II-l). Spherical browncyst with very short processes and a plate-like arche-opyle. The short processes resemble verrucae at lower
chematic representation of the raw percentage data. Abbreviations followreous peridiniales; Ech: Echinidinium spp.; Ach: Achomosphaera spp.;tion coefficient (0=shortest distance, strongest possible correlation).
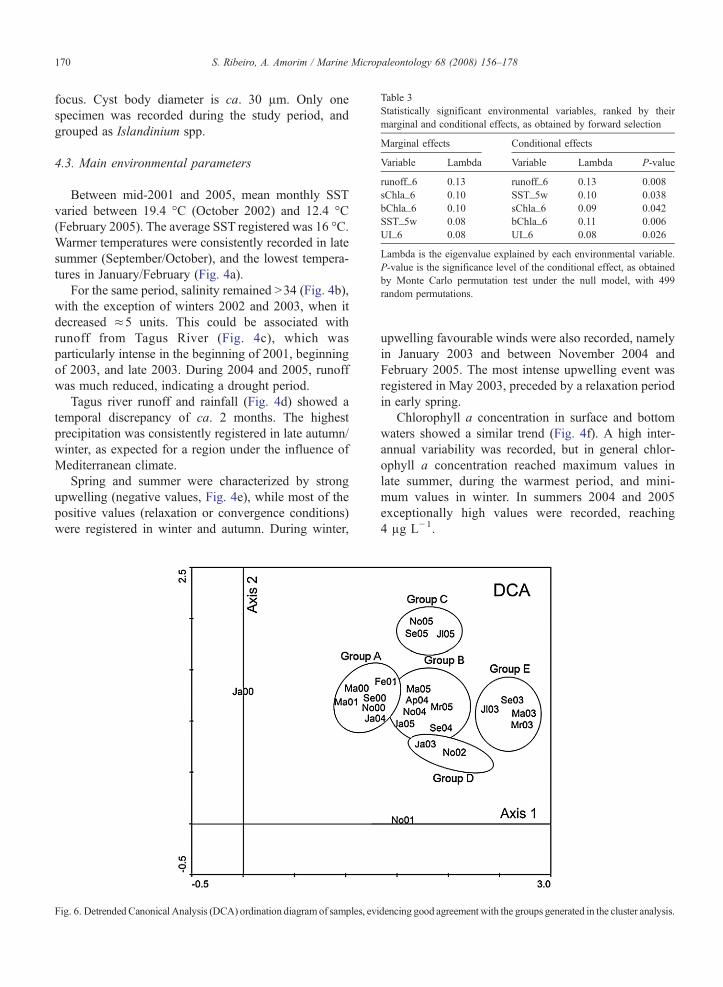
Table 3Statistically significant environmental variables, ranked by theirmarginal and conditional effects, as obtained by forward selection
Marginal effects Conditional effects
Variable Lambda Variable Lambda P-value
runoff_6 0.13 runoff_6 0.13 0.008sChla_6 0.10 SST_5w 0.10 0.038bChla_6 0.10 sChla_6 0.09 0.042SST_5w 0.08 bChla_6 0.11 0.006UI_6 0.08 UI_6 0.08 0.026
Lambda is the eigenvalue explained by each environmental variable.P-value is the significance level of the conditional effect, as obtainedby Monte Carlo permutation test under the null model, with 499random permutations.
170 S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
focus. Cyst body diameter is ca. 30 µm. Only onespecimen was recorded during the study period, andgrouped as Islandinium spp.
4.3. Main environmental parameters
Between mid-2001 and 2005, mean monthly SSTvaried between 19.4 °C (October 2002) and 12.4 °C(February 2005). The average SST registered was 16 °C.Warmer temperatures were consistently recorded in latesummer (September/October), and the lowest tempera-tures in January/February (Fig. 4a).
For the same period, salinity remained N34 (Fig. 4b),with the exception of winters 2002 and 2003, when itdecreased ≈5 units. This could be associated withrunoff from Tagus River (Fig. 4c), which wasparticularly intense in the beginning of 2001, beginningof 2003, and late 2003. During 2004 and 2005, runoffwas much reduced, indicating a drought period.
Tagus river runoff and rainfall (Fig. 4d) showed atemporal discrepancy of ca. 2 months. The highestprecipitation was consistently registered in late autumn/winter, as expected for a region under the influence ofMediterranean climate.
Spring and summer were characterized by strongupwelling (negative values, Fig. 4e), while most of thepositive values (relaxation or convergence conditions)were registered in winter and autumn. During winter,
Fig. 6. Detrended Canonical Analysis (DCA) ordination diagramof samples, ev
upwelling favourable winds were also recorded, namelyin January 2003 and between November 2004 andFebruary 2005. The most intense upwelling event wasregistered in May 2003, preceded by a relaxation periodin early spring.
Chlorophyll a concentration in surface and bottomwaters showed a similar trend (Fig. 4f). A high inter-annual variability was recorded, but in general chlor-ophyll a concentration reached maximum values inlate summer, during the warmest period, and mini-mum values in winter. In summers 2004 and 2005exceptionally high values were recorded, reaching4 µg L−1.
idencing good agreement with the groups generated in the cluster analysis.
171S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
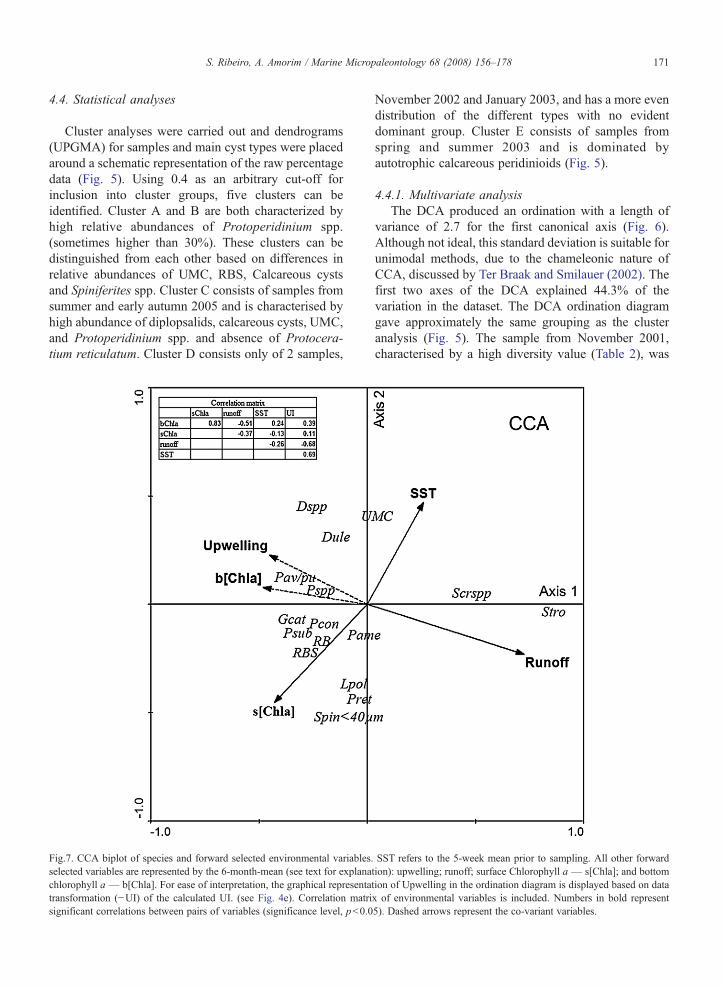
4.4. Statistical analyses
Cluster analyses were carried out and dendrograms(UPGMA) for samples and main cyst types were placedaround a schematic representation of the raw percentagedata (Fig. 5). Using 0.4 as an arbitrary cut-off forinclusion into cluster groups, five clusters can beidentified. Cluster A and B are both characterized byhigh relative abundances of Protoperidinium spp.(sometimes higher than 30%). These clusters can bedistinguished from each other based on differences inrelative abundances of UMC, RBS, Calcareous cystsand Spiniferites spp. Cluster C consists of samples fromsummer and early autumn 2005 and is characterised byhigh abundance of diplopsalids, calcareous cysts, UMC,and Protoperidinium spp. and absence of Protocera-tium reticulatum. Cluster D consists only of 2 samples,
Fig.7. CCA biplot of species and forward selected environmental variables.selected variables are represented by the 6-month-mean (see text for explanachlorophyll a — b[Chla]. For ease of interpretation, the graphical representatransformation (−UI) of the calculated UI. (see Fig. 4e). Correlation matrisignificant correlations between pairs of variables (significance level, pb0.0
November 2002 and January 2003, and has a more evendistribution of the different types with no evidentdominant group. Cluster E consists of samples fromspring and summer 2003 and is dominated byautotrophic calcareous peridinioids (Fig. 5).
4.4.1. Multivariate analysisThe DCA produced an ordination with a length of
variance of 2.7 for the first canonical axis (Fig. 6).Although not ideal, this standard deviation is suitable forunimodal methods, due to the chameleonic nature ofCCA, discussed by Ter Braak and Smilauer (2002). Thefirst two axes of the DCA explained 44.3% of thevariation in the dataset. The DCA ordination diagramgave approximately the same grouping as the clusteranalysis (Fig. 5). The sample from November 2001,characterised by a high diversity value (Table 2), was
SST refers to the 5-week mean prior to sampling. All other forwardtion): upwelling; runoff; surface Chlorophyll a — s[Chla]; and bottomtion of Upwelling in the ordination diagram is displayed based on datax of environmental variables is included. Numbers in bold represent5). Dashed arrows represent the co-variant variables.
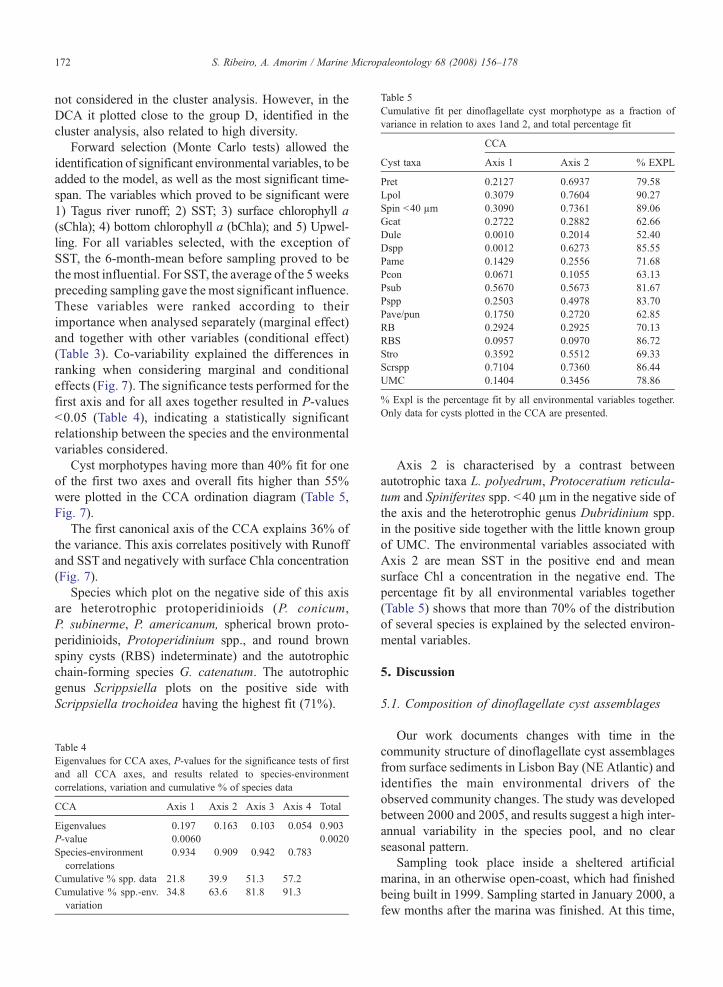
Table 5Cumulative fit per dinoflagellate cyst morphotype as a fraction ofvariance in relation to axes 1and 2, and total percentage fit
CCA
Cyst taxa Axis 1 Axis 2 % EXPL
Pret 0.2127 0.6937 79.58Lpol 0.3079 0.7604 90.27Spin b40 µm 0.3090 0.7361 89.06Gcat 0.2722 0.2882 62.66Dule 0.0010 0.2014 52.40Dspp 0.0012 0.6273 85.55Pame 0.1429 0.2556 71.68Pcon 0.0671 0.1055 63.13Psub 0.5670 0.5673 81.67Pspp 0.2503 0.4978 83.70Pave/pun 0.1750 0.2720 62.85RB 0.2924 0.2925 70.13RBS 0.0957 0.0970 86.72Stro 0.3592 0.5512 69.33Scrspp 0.7104 0.7360 86.44UMC 0.1404 0.3456 78.86
% Expl is the percentage fit by all environmental variables together.Only data for cysts plotted in the CCA are presented.
172 S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
not considered in the cluster analysis. However, in theDCA it plotted close to the group D, identified in thecluster analysis, also related to high diversity.
Forward selection (Monte Carlo tests) allowed theidentification of significant environmental variables, to beadded to the model, as well as the most significant time-span. The variables which proved to be significant were1) Tagus river runoff; 2) SST; 3) surface chlorophyll a(sChla); 4) bottom chlorophyll a (bChla); and 5) Upwel-ling. For all variables selected, with the exception ofSST, the 6-month-mean before sampling proved to bethe most influential. For SST, the average of the 5 weekspreceding sampling gave the most significant influence.These variables were ranked according to theirimportance when analysed separately (marginal effect)and together with other variables (conditional effect)(Table 3). Co-variability explained the differences inranking when considering marginal and conditionaleffects (Fig. 7). The significance tests performed for thefirst axis and for all axes together resulted in P-valuesb0.05 (Table 4), indicating a statistically significantrelationship between the species and the environmentalvariables considered.
Cyst morphotypes having more than 40% fit for oneof the first two axes and overall fits higher than 55%were plotted in the CCA ordination diagram (Table 5,Fig. 7).
The first canonical axis of the CCA explains 36% ofthe variance. This axis correlates positively with Runoffand SST and negatively with surface Chla concentration(Fig. 7).
Species which plot on the negative side of this axisare heterotrophic protoperidinioids (P. conicum,P. subinerme, P. americanum, spherical brown proto-peridinioids, Protoperidinium spp., and round brownspiny cysts (RBS) indeterminate) and the autotrophicchain-forming species G. catenatum. The autotrophicgenus Scrippsiella plots on the positive side withScrippsiella trochoidea having the highest fit (71%).
Table 4Eigenvalues for CCA axes, P-values for the significance tests of firstand all CCA axes, and results related to species-environmentcorrelations, variation and cumulative % of species data
CCA Axis 1 Axis 2 Axis 3 Axis 4 Total
Eigenvalues 0.197 0.163 0.103 0.054 0.903P-value 0.0060 0.0020Species-environmentcorrelations
0.934 0.909 0.942 0.783
Cumulative % spp. data 21.8 39.9 51.3 57.2Cumulative % spp.-env.variation
34.8 63.6 81.8 91.3
Axis 2 is characterised by a contrast betweenautotrophic taxa L. polyedrum, Protoceratium reticula-tum and Spiniferites spp. b40 µm in the negative side ofthe axis and the heterotrophic genus Dubridinium spp.in the positive side together with the little known groupof UMC. The environmental variables associated withAxis 2 are mean SST in the positive end and meansurface Chl a concentration in the negative end. Thepercentage fit by all environmental variables together(Table 5) shows that more than 70% of the distributionof several species is explained by the selected environ-mental variables.
5. Discussion
5.1. Composition of dinoflagellate cyst assemblages
Our work documents changes with time in thecommunity structure of dinoflagellate cyst assemblagesfrom surface sediments in Lisbon Bay (NE Atlantic) andidentifies the main environmental drivers of theobserved community changes. The study was developedbetween 2000 and 2005, and results suggest a high inter-annual variability in the species pool, and no clearseasonal pattern.
Sampling took place inside a sheltered artificialmarina, in an otherwise open-coast, which had finishedbeing built in 1999. Sampling started in January 2000, afew months after the marina was finished. At this time,

173S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
two autotrophic species were dominant, G. catenatum(N20%) and Spiniferites delicatus (ca. 30%) (Fig. 3).Between 1986 and 1995, recurrent blooms ofG. catenatum were observed along the whole coast ofPortugal (e.g. Moita et al., 1998). Earlier reports on thedistribution of G. catenatum cysts in this region haveshown that relative abundances above 20% reflectbloom periods, while values close to 7%, as thoserecorded during most of this study period, represent thebackground values characterizing regions subject torecurrent blooms (Amorim et al., 2001b). In a four yearcyst survey, conducted approximately 1° latitude southof Lisbon Bay (37°57′N, 08°53′W), Amorim (2001)recorded in September 1999 a major increase in therelative abundance of S. delicatus from backgroundlevels of 1% to 42%. The recorded cysts were all withcell contents suggesting recent production. Based on theabove evidence, the exceptional high values found forG. catenatum and S. delicatus in January 2000, atLisbon Bay, seem to reflect a combination of twofactors: (1) the Marina represented a new sedimentarybasin in the system, initially with no true autochtonouscyst assemblage, and (2) these two species were themain bloomers prior to sampling. Thus, the initial cystassemblage was highly influenced by the generalhydrographical processes determining the advectionand sedimentation of cysts recently produced in areasadjoining Lisbon Bay. In later succession stages,integration in time of overall cyst production andtransportation allowed for the build up of a higherdiversity species pool, representative of cyst assem-blages in Lisbon Bay.
From a biogeographical perspective, dinoflagellatecyst assemblages in Lisbon Bay may be classified aswarm-temperate estuarine-neritic. This is evidenced bythe common occurrence of indicator species such asL. polyedrum, S. mirabilis, P. leonis, P. pentagonum andP. conicum (Table 2) and the consistence absence ofseveral cold-water species, namely P. conicoides, Pen-tapharsodinium dalei and Bitectatodinium tepikense(Wall et al., 1977; Edwards and Andrle, 1992; Dale,1996). In general, assemblages were characterised byhigh diversity values (Table 2) in good agreement withresults reported from similar environments in theAtlantic (Wall et al., 1977).
From a trophic perspective, the assemblages weregenerally dominated by heterotrophs, with the exceptionof January 2000 (discussed above) and most of 2003(March to September), when autotrophic calcareouscysts were the dominant group. Heterotrophic dom-inance is usually associated with high-productivityareas, namely the upwelling systems of Peru, Chile,
Mauritania, south-west Iceland and the Faeroe Islands(Dale, 1996), and West-Central Africa (Dale et al.,2002).
About 30 modern dinoflagellate species have so farbeen found to incorporate calcite into their cyst wall.They contribute considerably to the total carbonate fluxto the ocean floor at times (Dale, 1992) and, due to theirextensive fossil record, can be used as paleoenviron-mental indicators (e.g. Zonneveld et al., 1999; Vink,2004). Although calcareous cysts seem to be a commonelement in surface sediments from coastal waters aroundthe world, these can sometimes be overlooked due to thewidely used acid treatment in cyst assemblage studies(Montresor et al., 1998 and references therein). In thissurvey, calcareous cysts (mostly belonging to the genusScrippsiella) were very abundant and were responsiblefor the major shift in the cyst community composition.They were also identified as a major cyst signal (seediscussion below) highlighting their usefulness in neriticzones as palaeoecological proxies.
There are still few published works on cystassemblages from the coastal warm-temperate regionof the NE Atlantic. Our work has allowed the identifi-cation of several morphotypes, which may prove usefulin identifying or strengthening the interpretation ofecological signals. These include round brown spiny(RBS) cysts which most likely belong to the cyst-basedgenera Echinidium (Zonneveld, 1997) and Islandinium(Head et al., 2001), considered to have affinity withthe Protoperidiniaceae and thus contributing to theheterotrophic pool. Echinidiniumwas first reported fromequatorial and subtropical oceanic zones, while Islan-dinium species were first described as cold-marineindicators, although the distribution of these cyst-basedgenera has latter been extended to other biogeographicalzones (Zonneveld, 1997; Head et al., 2001; Pospelovaand Head, 2002). Before the description of these generaseveral authors noted ‘spiny forms’ orMultispinula spp.as common components of high-productivity areas suchas upwelling regions (e.g. Wall et al., 1977; Lewis et al.,1990; Dale and Fjellså, 1994; Dale et al., 2002). Athorough examination of these cyst morphotypes,coupled with germination experiments for the establish-ment of cyst-theca relationships is essential to clarify theecological boundaries, diversity, and taxonomy of thisgroup.
During the study period, two major blooms involvingcyst-producing dinoflagellates (G. catenatum andL. polyedrum) were detected in waters North andSouth of Lisbon Bay. However, these reports were notfollowed by an increase in cysts of the blooming speciesinside the Bay. By contrast, after detection of the G.

174 S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
catenatum bloom, sediment samples collected in theMondego estuary (40°09′N, 8°52′W), North of LisbonBay, had relative abundance values of G. catenatumcysts close to 70% (Artigas et al., in press). L.polyedrum bloomed in August 2005, to the south ofthe sampling site (Moita, personal communication),when southward currents prevail (Ambar and Fiúza,1994). G. catenatum bloomed in the autumn of the sameyear, north of Lisbon Bay (Moita et al., in press) whencurrents are predominantly northwards. These resultshighlight how the sedimentary regime at the time of cystproduction, ultimately determined by the prevailinghydrographic conditions, may influence the composi-tion of cyst assemblages in surface sediments.
5.2. Environmental parameters: drivers of communitychange
In phytoplankton ecology the interaction betweengradients of physical disturbance, irradiance andnutrients is considered to be a major driving force indetermining species succession and community organi-sation (Margalef, 1978; Smayda and Reynolds, 2003).Understanding the environmental factors driving tem-poral succession in dinoflagellate cyst assemblages innature is further complicated by the time-lag and spatialuncertainty existing between the environmental triggeron the encystment processes and the cyst record insediments. Adding to these uncertainties, species-specific physiological properties of cysts, such asmandatory dormancy period and temperature windowfor germination, also contribute for determination of thecyst pool and succession patterns (e.g. Dale, 1976;Montresor et al., 1998; Ishikawa and Taniguchi, 1996,1997; Anderson and Rengefors, 2006). Thus, no attempthas been made to establish direct correlations betweenmeasured environmental variables and cyst occurrences.Instead, statistical methods were used to try to identifythe most significant abiotic parameters driving commu-nity changes (Dale et al., 2002).The short length ofvariance obtained for the DCA and the way samplesplotted in the same quadrant reflect the nature of thisstudy (Fig. 6). Samples were separated in time but camefrom the same site. Cyst assemblages in surfacesediments are known to represent integration in timeof several months to years, thus it is expected that thesesamples will always share common species.
The CCA allowed identification of Tagus river runoffas a major environmental factor affecting dinoflagellatecyst assemblages in Lisbon Bay. This parameter showeda negative correlation with upwelling intensity. Bothvariables were most influential when considering an
average of 6 months prior to the sampling date, probablyreflecting the marked seasonal character of these 2variables, and the integrative nature of the cyst signal.The Tagus river runoff is usually higher in winter, whileseasonal upwelling prevails in spring and summer. Bothupwelling and river runoff are major processes asso-ciated with nutrient enrichment in marine coastalregions. Thus Axis 1 cannot be interpreted as a nutrientgradient.
Most of the heterotrophic cyst types with highestweights on the negative end of Axis 1 are welldocumented in the literature in association with highnutrient upwelling regions (e.g. P. conicum, RB, RBS)(Wall et al., 1977; Lewis et al., 1990; Dale and Fjellså,1994; Dale et al., 2002). G. catenatum, which plotted inassociation with this group, is also well documented asan upwelling relaxation taxa based on the ecology of themotile stage (Smayda, 2002; GEOHAB 2005). Bycontrast, the genus Scrippsiella, with strong affinity tothe positive side of Axis 1, is typical of near-shorehighly stratified nutrient-rich waters (Smayda, 2002;Smayda and Reynolds, 2003). This clearly suggests thatthe strongest ecological gradient affecting speciessuccession in Lisbon Bay is related to water stability.River runoff in the positive domain of Axis 1 reflectingnutrient-rich stratified waters and upwelling in thenegative domain reflecting nutrient-rich verticallymixed waters. Given these two major nutrient sources,Lisbon Bay is not expected to be nutrient limited. Theobservation, all year round, of a highly diverseassemblage of cyst-producing dinoflagellates (mero-planktonic) supports the evidence from previous workthat, in nature, cyst formation cannot generally beexplained by nutrient depletion (e.g. Kremp andHeiskanen, 1999; Anderson and Rengefors, 2006).However, it is interesting to note that the two differentenrichment processes prevalent in Lisbon Bay select fordifferent trophic and functional groups, as expected bythe proposed Assembly Rules of Smayda and Reynolds(2003). The upwelling signal is characterized bydominance of heterotrophs and presence of chain-forming dinoflagellates, tolerant to physically disturbedwaters (Ruderal-strategists of Smayda and Reynolds2003), while water stability induced by river runoffselects for autotrophic, small, fast-growing species(Colonist-strategists of Smayda and Reynolds 2003).These species have cysts with short mandatory dor-mancy periods (e.g. S.trochoidea, Binder and Anderson,1987) that, combined with high vegetative growth rates,result in fast cyst-motile stage turnover rates providingconstant inocula to the water column and deposition ofnewly formed cysts in the sediments. This strategy may

175S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
be an advantage for the exploration of resources inhighly variable environments (Montresor et al., 1998).
As reported from other marine systems influenced byriver plumes (Dale et al., 2002), there is no evidencefrom the cyst record suggesting a salinity signalassociated with Tagus river runoff, similar to thosedescribed for low salinity brackish water environments(Dale, 1996).
Since 1976, blooms of G. catenatum have beenregularly recorded along the west Iberian coast. Massiveblooms were recorded predominantly in late summer–early autumn in the transition from the upwelling to thedownwelling season. However, for several years inter-mittent blooms were also recorded associated withrelaxation episodes during the upwelling season (for areview see GEOHAB, 2005). The fact thatG. catenatumin Lisbon Bay plotted with heterotrophs, well docu-mented as part of the cyst upwelling signal elsewhere,supports previous suggestions of G. catenatum beingpart of this signal in the NE Atlantic (Tarragona et al.,1999; Amorim, 2001). Recently, results from a dino-flagellate cyst study in sediment cores from the Taguspro-Delta covering ca 2000 years have shown that G.catenatum was most likely introduced in the NEAtlantic at the beginning of the XX century (Amorimand Dale, 2006). From a paleoceanographic perspective,this limits the use of this cyst signal in the N. Atlantic toa very narrow period in recent times. However, itsupports the use of G. catenatum cysts as part of theupwelling signal in the upwelling system off California,in the Pacific west coast of North America, presumablythe biogeographical distribution center for this species(Mudie et al., 2002; Amorim and Dale, 2006).
The gradient associated with Axis2 in the CCA ismore difficult to interpret, and further work is needed tobetter resolve its ecological significance. The environ-mental variables most significantly associated with thisAxis were SST in the positive domain and surfacechlorophyll a in the negative domain. The end-membergroups comprising the highest scores on Axis 2 are theheterotrophic diplopsalids and the autotrophic UMC onthe positive end, and the cosmopolitan autotrophicspecies L. polyedrum, Protoceratium reticulatum andsmall Spiniferites spp. in the negative end. The lattergroup is well documented as indicative of coastalassemblages (e.g. Dale et al., 2002). In Lisbon Bay theposition of this group in the CCA diagram, with highscores for Axis 2, and an intermediate position on Axis 1suggests a preference for intermediate turbulence con-ditions. On the other hand, the position on the negativeside of Axis 1, and the close relationship to high surfaceproductivity and low SST, suggests an association with
upwelling events. Along the Portuguese coast, recentblooms of L. polyedrum have been reported in southLisbon Bay and along the south coast in the vicinity ofCape S. Vicente, associated with particular oceano-graphic conditions whereby blooms occur in warmstratified waters compressed by active upwellingfilaments (Amorim et al., 2001a, 2004). In the Cape S.Vicente region, cysts of L. polyedrum may representmore than 50% of the assemblage. In Lisbon Bay valuesnever exceeded 7% of the cyst assemblage, suggestingthe environmental conditions favouring development ofL. polyedrum in the Bay are not so persistent or frequentas in the vicinity of Cape S. Vicente. L. polyedrum haspreviously been documented as part of particularupwelling conditions in other regions of the world,namely South California and Northwest Africa (Dale,1996). This supports our interpretation of L. polyedrumas part of the upwelling signal along the Portuguesecoast. Smayda and Reynolds (2003) group bothL. polyedrum and G. catenatum as upwelling relaxationtaxa. The distinct position of these two species in theCCA suggests the existence of different optima within aprevailing upwelling ecological gradient.
Blooms of L. poyledrum were never observed to berecurrent in the same region and season. By contrast,blooms of G. catenatum may be recurrent during theupwelling season, responding to the intermittent natureof upwelling episodes. Once initiated, blooms are oftentransported along the coast by dominant surface currents(Moita et al., 1998). To our knowledge, these twospecies have never bloomed at the same time and in thesame region along the Iberian coast. Smayda andReynolds (2003) point out that within each strategythere are species-specific differences in the responserates to environmental changes. Several propertiesdistinguishing L. polyedrum and G. catenatum maycontribute to the suggested differences in niche selec-tion. These differences may be due to the classicaldistinction between r-species and k-species proposed bythese authors, combined with differences in functionalmorphology (chain-formation vs. single cells) andphysiology of the dormant cyst. In fact, several studieshave shown that cysts of L. polyedrum need hightemperatures (N19 °C) for germination and have a longmandatory dormancy period (N55 days) (Figueroa andBravo 2005; Anderson and Rengefors, 2006). Thisprevents germination of newly formed cysts andprobably inhibits the occurrence of recurrent bloomsin the same upwelling season. G. catenatum, on theother hand, has a wide temperature window forgermination and one of the shortest known mandatorydormancy periods (Hallegraeff and Fraga, 1998). This

176 S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
probably allows this species to explore the stochasticupwelling/relaxation cycles occurring in a single season.
6. Conclusions
The study of time-series changes in the communitystructure of recent dinoflagellate cyst assemblages inLisbon Bay allowed the identification of waterstability as the major ecological gradient drivingchanges in the assemblages. Statistical analysis ofspecies' relative abundances and environmental datasupport previous interpretations that cysts in sedimentsdo not reflect a direct relation to instant, locallymeasured hydrographic parameters, but a time integra-tion of the abiotic factors affecting their production,transportation and sedimentation.
Most of the cyst signals identified herein areconsistent with results obtained for other areas influ-enced both by upwelling and river runoff, reinforcingthe use of dinoflagellate cysts as environmental tracers.The two processes select for different trophic groups andlife-forms which is reflected on the cyst assemblage.
The main cyst signals recognised in Lisbon Bay are:
1. High water stability induced by Tagus river runoff,characterised by dominance of autotrophic neriticcalcareous cysts (mainly Scrippsiella spp.).
2. Upwelling, characterized by the well describeddominance of heterotrophic species, particularlyProtoperidinioids, and cysts of the autotrophicchain-forming species G. catenatum. L. polyedrumis confirmed as an upwelling indicator, but exploringan ecological niche different from the former group.
Acknowledgements
This work has been developed within projectsPROFIT — “Interdisciplinary study of processesunderlying the phytoplankton dynamics in the Portu-guese upwelling system” — (PDCTE/CTA/50386/2003) and PORTCOAST — “Present and futurePortuguese coastal climate and its impacts on thebiological communities” — (POCTI/CLI/58348/2004),funded by the Portuguese Foundation for Science andTechnology. The authors would like to thank TeresaMoita and Sofia Palma, from IPIMAR, for kindlymaking available environmental data used in this study.Lino Costa is greatly acknowledged for advicesregarding statistical analyses, and Vera Veloso for thehelp with sampling and slide preparation. Thanks arealso due to two anonymous reviewers, for constructivecomments that helped improve this manuscript. Sofia
Ribeiro holds a PhD scholarship from the PortugueseFoundation for Science and Technology (SFRH/BD/30847/2006).
References
Ambar, I., Fiúza, A., 1994. Some features of the Portugal currentsystem: a poleward slope undercurrent, an upwelling-relatedsummer southward flow and an autumn–winter poleward coastalsurface current. In: Katasaros, K.B., Fiúza, A.F.G., Ambar, I.(Eds.), Proceedings of the International Conference on Air–seainteraction and on Meteorology and Oceanography of the CoastalZone. American Meteorological Society, Boston, pp. 286–287.
Amorim, A., 2001. Dinoflagellate cyst distribution along the coast ofPortugal. PhD Thesis. Faculty of Sciences, University of Lisbon,Portugal.
Amorim, A., Dale, B., 1998. Distribution of cysts from toxic orpotentially toxic dinoflagellates along the Portuguese coast. In:Reguera, B., Blanco, J., Fernandez, M.L., Wyatt, T. (Eds.),Harmful Algae. Xunta de Galicia and IOC of UNESCO, Santiagode Compostela, pp. 64–65.
Amorim, A., Dale, B., 2006. Historical cyst record as evidence for therecent introduction of the dinoflagellate Gymnodinium catenatumin the North Eastern Atlantic. African Journal of Marine Science28 (2), 193–197.
Amorim, A., Palma, A.S., Sampayo, M.A., Moita, M.T., 2001a. On aLingulodinium polyedrum bloom in Setúbal bay, Portugal. In:Hallegraeff, G.M., Blackburn, S.I., Bolch, C.J., Lewis, R.J. (Eds.),Harmful Algal Blooms 2000. Intergovernmental OceanographicCommission of UNESCO, pp. 133–136.
Amorim, A., Dale, B., Godinho, R., Brotas, V., 2001b. Gymnodiniumcatenatum-like (Dinophyceae) cysts in recent sediments from thecoast of Portugal. Phycologia 40 (6), 572–582.
Amorim, A., Moita, M.T., Oliveira, P., 2004. Dinoflagellate bloomsrelated to coastal upwelling plumes off Portugal. In: Steidinger, K.A.,Landsberg, J.H., Tomas, C.R., Vargo, C.A. (Eds.). Harmful Algae2002. Florida Fish and Wildlife Conservation Commission, FloridaInstitute of Oceanography, and Intergovernmental OceanographicComission of UNESCO, St. Petersburg, Florida, USA. pp. 89–91.
Anderson, D., Rengefors, K., 2006. Community assembly andseasonal succession of marine dinoflagellates in a temperateestuary: the importance of life cycle events. Limnology andOceanography 5, 860–873.
Artigas, M., Amorim, A., Vale, P., Gomes, S.S., Botelho, M.J.,Rodrigues, S.M., in press. Prolonged toxicity of Scrobiculariaplana after a PSP event and its relation toGymnodinium catenatumcyst consumption and toxin depuration. Proceedings of the XIIInternational Conference on Harmful Algae.
Bakun, A., 1973. Coastal upwelling indices, west coast of NorthAmerica, 1976–71. NOAATechnical Report: 103.
Benito, G., Diez-Herrero, A., de Villalta, M.F., 2003. Magnitude andfrequency of flooding in the Tagus basin (Central Spain) over thelast millennium. Climatic Change 58, 171–192.
Binder, B.J., Anderson, D.M., 1987. Physiological and environmentalcontrol of germination in Scrippsiella trochoidea (Dinophyceae)resting cysts. Journal of Phycology 23 (1), 99–107.
Bolch, C.J., 1997. The use of sodium polytungstate for the separationand concentration of living dinoflagellate cysts from marinesediments. Phycologia 36, 472–478.
Dale, B., 1976. Cyst formation, sedimentation, and preservation:factors affecting dinoflagellate assemblages in recent sediments

177S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
from Trondheimsfjord, Norway. Review of Palaeobotany andPalynology 22, 39–60.
Dale, B., 1983. Dinoflagellate resting cysts: “benthic plankton”. In:Fryxell, G.A. (Ed.), Survival Strategies of the Algae. CambridgeUniversity Press, Cambridge, pp. 69–136.
Dale, B., 1992. Dinoflagellate contributions to the open oceansediment flux. In: Honjo, S. (Ed.), Ocean Biocenosis Series, 5.Woods Hole Oceanographic Institute, Woods Hole, MA, pp. 1–23.
Dale, B., 1996. Dinoflagellate cyst ecology: modelling and geologicalapplications. In: Jansonius, J., McGregory, D.C. (Eds.), Palynol-ogy: principles and applications. . The American Association ofStratigraphic Palynologists Foundation, vol. 3. Publishers Press,Salt Lake City, pp. 1249–1276.
Dale, B., Fjellså, A., 1994. Dinoflagellate cysts as paleoproductivityindicators: state of the art, potential, and limits. In: Zahn, R.,Pedersen, T.F., Kaminski, M.A., Labeyrie, L. (Eds.), CarbonCycling in the Glacial Ocean: Constrains on the ocean's role inGlobal Change. Springer–Verlag, Berlin, pp. 521–537.
Dale, B., Dale, A., 2002. Environmental applications of dinoflagellatecysts and acritarchs. In: Haslett, S. (Ed.), Quaternary EnvironmentalMicropaleontology. Edward Arnold Lda, London, pp. 207–240.
Dale, B., Dale, A.L., Jansen, J.H.F., 2002. Dinoflagellate cysts asenvironmental indicators in surface sediments from the Congodeep-sea fan and adjacent regions. Palaeogeography, Palaeocli-matology, Palaeoecology 185 (3–4), 309–338.
de Vernal, A., Henry, M., Matthiessen, J., Mudie, P., Rochon, A.,Boessenkool, K., Eynaud, F., Grøsfjeld, K., Guiot, J., Hamel, D.,Harland, R., Head, M.J., Kunz-Pirrung, M., Levac, E., Loucheur, V.,Peyron, O., Pospelova, V., Radi, T., Turon, J.-L., Voronina, E., 2001.Dinoflagellate cyst assemblages as tracers of sea-surface conditionsin the northern North Atlantic, Arctic and sub-Arctic seas: the new‘n=677’ data base and its application for quantitative palaeoceano-graphic reconstruction. Journal of Quaternary Science 16 (7),681–698.
Doblin, M., Blackburn, S., Hallegraeff, G., 1999. Comparative studyof selenium requirements of three phytoplankton species: Gym-nodinium catenatum, Alexandrium minutum (Dinophyta) andChaetoceros cf. tenuissimus (Bacillariophyta). Journal of PlanktonResearch 21, 1153–1169.
Edwards, L.E., Andrle, A.S., 1992. Distribution of selected dino-flagellate cysts in modern marine sediments. In: Head, M.J.,Wrenn, L.H. (Eds.), Neogene and Quaternary Dinoflagellate Cystsand Acritarchs. American Association of Stratigraphic Palynolo-gists Foundation, Dallas, TX, pp. 259–288.
Ellegaard, M., Daugbjerg, N., Rochon, A., 2003. Morphological andLSU rDNA sequence variation within the Gonyaulax spinifera–Spiniferites group (Dinophyceae) and proposal of G-elongatacomb. nov and G-membranacea comb. nov. Phycologia 42 (2),151–164.
Figueroa, R.I., Bravo, I., 2005. Sexual reproduction and two differentencystment strategies of Lingulodinium polyedrum (Dinophyceae)in culture. Journal of Phycology 41, 74–83.
Fiúza, A.F., de Macedo, M.E., Guerreiro, M.R., 1982. Climatologicalspace and time variation of the Portuguese coastal upwelling.Oceanologica Acta 5 (1), 31–40.
GEOHAB, 2005. Global Ecology and Oceanography of Harmful AlgalBlooms, GEOHAB Core Research Project: HABs in UpwellingSystems. G. Pitcher, T.Moita, V. Trainer, R. Kudela, P. Figueiras, T.Probyn (Eds.) IOC and SCOR, Paris and Baltimore. 82 pp.
Hallegraeff, G.M., Fraga, S., 1998. Bloom dynamics of the toxicdinoflagellateGymnodinium catenatum, with emphasis on Tasmaniaand Spanish coastal waters. In: Anderson, D.M., Cembella, A.D.,
Hallegraeff, G.M. (Eds.), Physiological ecology of harmful algalblooms. NATO ASI Series, vol. G 41. Springer–Verlag, Heidelberg,pp. 59–80.
Harland, R., Nordberg, K., Filipsson, H., 2004. The seasonaloccurrence of dinoflagellate cysts in surface sediments fromKoljö Fjord, west coast of Sweden — a note. Review ofPalaeobotany and Palynology 128, 107–117.
Head, M.J., 1996. Modern dinoflagellates cysts and their biologicalaffinities. In: Jansonius, J., McGregor, D.C. (Eds.), Palynology:Principles and Applications. American Association of Strati-graphic Palynologists, Dallas, pp. 1197–1248.
Head, M.J., Harland, R., Matthiessen, J., 2001. Cold marine indicatorsof the late quaternary: the new dinoflagellate cyst genus Islandi-nium and related morphotypes. Journal of Quaternary Science 16,621–636.
Ishikawa, A., Taniguchi, A., 1996. Contribution of benthic cysts to thepopulation dynamics of Scrippsiella spp. (Dinophyceae) inOnagawa Bay, northeast Japan. Marine Ecology Progress Series140, 169–178.
Ishikawa, A., Taniguchi, A., 1997. In situ germination patterns of cysts,and bloom formation of some armored dinoflagellates in OnagawaBay, north-east Japan. Journal of Plankton Research 19 (11),1783–1791.
Joyce, L.B., Pitcher, G.C., 2004. Encystment of Zygabikodiniumlenticulatum (Dinophyceae) during a summer bloom of dino-flagellates in the southern Benguela upwelling system Estuarine.Coastal and Shelf Science 59 (1), 1–11.
Kremp, A., Heiskanen, A.-S., 1999. Sexuality and cyst formation ofthe spring-bloom dinoflagellate Scrippsiella hangoei in the coastalnorthern Baltic Sea. Marine Biology 134, 771–777.
Lewis, J., Dodge, J.D., Powell, A.J., 1990. Quaternary dinoflagellate cystsfrom the upwelling system offshore Peru, Hole 686B, ODP LEG 112.In: Suess, E., von Huene, R., et al. (Eds.), Proceedings of the OceanDrilling Program. Scientific Results, vol. 112, pp. 323–327.
Mann, K.H., 2000. Ecology of coastal waters: with implications formanagement. Blackwell Science. 406 pp.
Margalef, R., 1978. Life-forms of phytoplankton as survival alternativesin an unstable environment. Oceanologica Acta 1, 493–509.
Moita, M.T., 2001. Estrutura, Variabilidade e Dinâmica do Fitoplânc-ton na Costa de Portugal Continental (in Portuguese with EnglishAbstract). PhD Thesis. Faculdade de Ciências da Universidade deLisboa. Portugal.
Moita, M.T., Amorim, A., 2001. The relevance of Gymnodiniumcatenatum (Dinophyceae) over-wintering planktonic population vs.cysts as seedbanks for the local development of toxic blooms offWestern Iberia. In: Garcés, E., Zingone, A., Montresor, M., Reguera,B., Dale, B. (Eds.), LIFEHAB— life histories of microalgal speciescausing harmful blooms. European Commission Directorate GeneralScience, Research and Development, pp. 87–89.
Moita, M.T., Vilarinho, M.G., Palma, A.S., 1998. On the variability ofGymnodinium catenatum Graham blooms in Portuguese waters.In: Reguera, B., Blanco, J., Fernandez, M.L., Wyatt, T. (Eds.),Harmful Algae. Xunta de Galicia and IOC of UNESCO, Santiagode Compostela, pp. 118–121.
Moita, M.T., Oliveira, P.B., Mendes, J.C., Palma, A.S., 2003.Distribution of chlorophyll a and Gymnodinium catenatumassociated with coastal upwelling plumes off central Portugal.Acta Oecologica 24, 125–132.
Moita, M.T., Palma, A.S., Oliveira, P.B., Vidal, T., Silva, A., Vilarinho,M.G., in press. The return of Gymnodinium catenatum after10 years: bloom initiation and transport off the Portuguese coast.Proceedings of the XII International Conference on Harmful Algae.

178 S. Ribeiro, A. Amorim / Marine Micropaleontology 68 (2008) 156–178
Montresor, M., Zingone, A., Sarno, D., 1998. Dinoflagellate cystproduction at a coastal Mediterranean site. Journal of PlanktonResearch 20 (12), 2291–2312.
Morquecho, L., Lechuga-Devéze, C.H., 2004. Seasonal occurrence ofplanktonic dinoflagellates and cyst production in relationship toenvironmental variables in subtropical Bahía Concepción, Gulf ofCalifornia. Botanica Marina 47, 313–322.
Mudie, P.J., Rochon, A., Levac, E., 2002. Palynological records of redtide-producing species in Canada: past trends and implications forthe future. Palaeogeography, Palaeoclimatology, Palaeoecology180, 159–186.
Munsterman, D., Kerstholt, S., 1996. Sodium polytungstate, a newnon-toxic alternative to bromoform in heavy liquid separation.Review of Paleobotany and Palynology 91, 417–422.
OSPAR, 2000. Quality status report 2000. Region IV— Bay of Biscayand Iberian Coast. Oslo-Paris Commission, London. 134 pp.
Palma, A.S., 2003. Estudo de um ciclo anual de Sucessão Fitoplanctó-nica na Baía de Cascais. MSc Thesis (in Portuguese with EnglishAbstract). Faculdade deCiências daUniversidade do Porto, Portugal.
Pospelova, V., Head, M.J., 2002. Islandinium brevispinosum sp. nov.(Dinoflagellata), a new organic-walled dinoflagellate cyst frommodern estuarine sediments of New England (USA). Journal ofPhycology 38 (3), 593–601.
Rochon, A., de Vernal, A., Turon, J.-L., Mattiessen, J., Head, M.,1999. Distribution of dinoflagellate cyst assemblages in surfacesediments from the North Atlantic Ocean and adjacent basins andquantitative reconstruction of seasurface parameters. AmericanAssociation of Stratigraphic Palynologists Special ContributionsSer. 35. 146 pp.
Schnepf, E., Elbrächter, M., 1992. Nutritional strategies in dino-flagellates. A review with emphasis on cell biological aspects.European Journal of Protistology 28, 3–24.
Shannon, C.E., Weaver, W., 1949. The mathematical theory ofcommunication. University of Illinois Press, Urbana IL.
Smayda, T.J., 2002. Turbulence, watermass stratification and harmfulalgal blooms: an alternative view and frontal zones as “pelagicseed banks”. Harmful Algae 1, 95–112.
Smayda, T.J., Reynolds, C.S., 2003. Strategies of marine dinoflagellatesurvival and some rules of assembly. Journal of Sea Research 49,95–106.
Sneath, P.H.A., Sokal, R.R., 1973. Numerical Taxonomy: theprinciples and practice of numerical classification. W.H. Freemanand Company, San Francisco. 573 pp.
Sousa, F.M., Bricaud, A., 1992. Satellite-derived phytoplanktonpigment structures in the Portuguese upwelling area. Journal ofGeophysical Research-Oceans 97 (C7), 11343–11356.
Stoecker, D., 1999. Mixotrophy among dinoflagellates. Journal ofEukaryotic Microbiology 46 (4), 397–401.
Tarragona, J., Warnaar, J., Boessenkool, K.P., Brinkhuis, H., Canals,M., 1999. Recent dinoflagellate cyst distribution in the NorthCanary Basin, NWAfrica. Grana 38, 170–178.
Ter Braak, C.J.F., Smilauer, P., 2002. CANOCOReference manual andCanoDraw for Window's User's guide: Software for CanonicalCommunity Ordination (version 4.5). Microcomputor Power,Ithaca, NY, USA. 500 pp.
Vink, A., 2004. Calcareous dinoflagellate cysts in South and equatorialAtlantic surface sediments: diversity, distribution, ecology andpotential for palaeoenvironmental reconstruction. Marine Micro-paleontology 50, 43–88.
Wang, Z.H., Qi, Y.Z., Lu, S.H., Wang, Y., Matsuoka, K., 2004.Seasonal distribution of dinoflagellate resting cysts in surfacesediments from Changjiang River Estuary. Phycological Research52 (4), 387–395.
Wall, D., Dale, B., 1968. Modern dinoflagellate cysts and evolution ofthe Peridiniales. Micropaleontology 14 (3), 265–304.
Wall, D., Dale, B., Lohman, G.P., Smith, W.K., 1977. Theenvironmental and climatic distribution of dinoflagellate cysts inmodern sediments from regions in the North and South Atlanticoceans and adjacent seas. Marine Micropaleontology 2, 121–200.
Wooster, W.S., Bakun, A., McLain, D.R., 1976. The seasonalupwelling cycle along the eastern boundary of the North Atlantic.Journal of Marine Research 34 (2), 131–141.
Zonneveld, K.A.F., 1997. New species of organic walled dinoflagel-late cysts from modern sediments of the Arabian Sea (IndianOcean). Review of Palaeobotany and Palynology 97, 319–337.
Zonneveld, K.A.F., Höll, C., Janofske, D., Karwath, B., Kerntopf, B.,Rühlemann, C., Willems, H., 1999. Calcareous dinoflagellates aspalaeo-environmental tools. Use of proxies in Paleoceanology:Examples from the South Atlantic. by Fischer, G. and Wefer, G.Springer Verlag, Berlin Heidelberg, pp. 145–164.




















