
Improving the analysis of dinoflagellate phylogeny based on rDNA
Upload
independentCategory
view
0download
0
Journal of Plankton Research Vol.20 no.12 pp.2291-2312, 1998
Dinoflagellate cyst production at a coastal Mediterranean site
M.Montresor, A.Zingone and D.SarnoStazione Zoologica 'A. Dohrn', Villa Comunale, 80121 Napoli, Italy
Abstract. To assess the diversity and seasonality of dinoflagellate cyst production, surface sedimentand trap samples were studied in the Gulf of Naples (Mediterranean Sea). A total of 59 different cystmorphotypes were recorded. At the stations within the 70 m isobath, sediment assemblages weredominated by calcareous Peridiniales (66-79%), while at the deepest stations non-calcareous Peri-diniales attained the highest percentages (40-49%). The sediment trap sampling, carried out fort-nightly over two annual cycles, revealed high production rates (up to 1.7 x 105 cysts nr2 day1) fromspring to late autumn of both years, with a distinct seasonal production pattern. Although rathersimilar in species composition, the total cyst flux differed markedly between the 2 years (1.26 and 0.55X 10s cysts nr2 year1, respectively). Species-specific production patterns were observed: some speciesformed cysts over several months, others in restricted periods of the year. Cyst-forming species consti-tuted a small part of the planktonic dinoflagellate populations recorded in the area. A couplingbetween the trap material and surface water plankton was observed for calcareous Peridiniales. Thissampling approach allowed the detection of some species never recorded before in the gulf, includingtwo potentially toxic species: Alexandrium andersoni and Gymnodinium catenatum-like species.
Introduction
The production of resting stages is known for ~200 marine dinoflagellates whichinclude species with an extremely wide array of ecological preferences andtrophic habits. Cyst-forming species are not equally distributed between theextant dinoflagellate orders, being much more common for the Peridiniales,Gonyaulacales and Gymnodiniales, and quite rare for the Dinophysiales andProrocentrales (Head, 1996). Cysts are the product of sexual reproduction(Pfiester and Anderson, 1987) and the only diploid phase in most dinoflagellatelife cycles. They have two peculiar characteristics: they are non-motile and aresurrounded by a dense, resistant wall, which cause their settling on the sea bottomand allow them to withstand more unfavourable conditions as compared to motilestages. The formation of resting stages thus notably widens the tolerance rangeof a species and offers the possibility of a better spread of its distribution range.As a general rule, a life cycle including settling stages should mainly be a featureof neritic species, but recent studies also suggest a previously underestimatedimportance of calcareous cyst production in open oceanic waters (Dale, 1992).
Over the last few years, our knowledge of dinoflagellate resting stages hasmainly derived from studies focused on toxic dinoflagellates, where the conse-quences of cyst-mediated species dispersal are particularly important (Hallegraeffand Bolch, 1992; Anderson, 1997). Mapping cyst bed extension (Anderson et al.,1982; Cembella et al., 1988) has become essential for detecting the presence ofnuisance species and tracking their routes of spread. Resting stages also representa valuable tool for paleo-environmental reconstructions (Edwards, 1992; Dale,1996; Versteegh, 1997) due to their ability to fossilize. These applications are basedon an extended knowledge of cyst distribution patterns in modern sediments(Reid, 1975; Wall et al, 1977; Dale, 1983; Harland, 1983) as well as of the biological
© Oxford University Press 2291
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

MJVfontresor, A.Zingone and Djamo
characteristics of single species. Since the majority of dinoflagellate cysts areformed in neritic environments, it is essential to focus on these areas in order toobtain a biogeographic coverage of their distribution.
An understanding of the ecological conditions under which resting stages areproduced is fundamental for the interpretation of the signal left in sediments, aswell as for the comprehension of the different life strategies. A first step in thisdirection is the in situ estimation of cyst production patterns and rates, which canbe approached by sediment trap sampling. Very few such studies are available,the only previous ones being those carried out in the open Atlantic and PacificOceans (Dale, 1992), the Nordic Sea (Dale and Dale, 1992) and, for Scrippsiellatrochoidea, in a Japanese bay (Ishikawa and Taniguchi, 1996).
The Gulf of Naples (Tyrrhenian Sea, Mediterranean Sea) is a deep embayment,which can be broadly subdivided into a coastal subsystem affected by land run-off, and an open-water subsystem, consisting of oligotrophic Tyrrhenian waters(Carrada et al, 1980). The transitional zone between the two subsystems can benarrow and close to the coastline, especially in summer, when it approximatelycorresponds to the 100 m isobath (Marino et al, 1984). Multi-annual informationon plankton distribution is available for this area (Marino et al, 1984; Scotto DiCarlo etal, 1985; Zingone et al, 1990,1995; A.Zingone and D.Sarno, unpublisheddata)
The present study is a 2 year investigation on dinoflagellate cyst production ata coastal station in the Gulf of Naples. The composition of surface sedimentassemblages is also depicted, which provides a more complete coverage of thecoastal system. The aim of this study was to assess the diversity, quantity andseasonality of cysts produced over the annual cycle, and to track the possiblerelationship between cyst production and planktonic dinoflagellates in this area.
Method
Sediment samples were collected at nine stations (Figure 1, Table I). Stations 1-5and MC were located on the shelf (30-70 m depth) at different distances from theNaples harbour, which is one of the main sources of eutrophication in the area.The other three stations were located in the western part of the gulf, at the headof deep underwater valleys, and represent possible accumulation sites.
Table I. Surface sediment sampling stations
Station
1234MC5678
Depth (m)
306040657060
110200300
Sampling device
CorerCorerCorerCorerCorerCorerGrabGrabGrab
Sampling date
20.03.199125.07.199020.03.199105.03.199105.03.199125.07.199015.11.199015.11.199015.11.1990
Sediment type
Sand-siltSand-siltSand-siltSand-siltSand-siltSand-siltSiltSilt-claySilt-clay
2292
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

DinoflageUate cyst production at a coastal site
i <
/ *
\ _.'''.•
/,-. >'' *< ; , 1
'500 ii
/ i \- \ t/
200 y /* *'* *''•/ , 500 / '
/ / / :- ' -- .
j 2 Naples
,%57 MC 3 \ \
' v \ V.«' l v
_ / ;'
• 8 . \
Y*•
200 _>-'
k |
N
i—t1nm
foo * )
Fig. 1. Map of the Gulf of Naples with the location of the sediment sampling stations (1-8 and MC)and the sediment trap station (T).
Surface sediments were collected by means of either a gravity corer or a Shipekgrab. The gravity corer was equipped with Plexiglas cylinders of 4 cm diameter.In the laboratory, the first 2 cm of sediment were collected in a plastic test tubeand stored in the dark at 5-6°C. The undisturbed portion of sediments collectedwith the Shipek grab was subsampled on board with a Plexiglas cylinder of 4 cmdiameter and the uppermost 2 cm were collected and stored as above.
For cyst identification and enumeration, small amounts of the sediment weregently sonicated for 45 s, pre-filtered on a 130-um-mesh metallic sieve, and thenfiltered on 36- and 20-um-mesh metallic sieves. Both the sediment fractions >36and >20 um were examined. Identification and counting were performed using aninverted Zeiss IM35 microscope. About 200 cysts were counted for each sample.Counts included both empty and apparently live cysts, and were expressed asrelative abundance (%) of the examined samples. Cyst diversity was estimatedusing the Shannon Weaver index, H' (Shannon and Weaver, 1949). A hierarch-ical clustering was performed using the complete linkage algorithm on a dissim-ilarity matrix between the sampling stations. Dissimilarity was calculated as 1 -PSI (percentage similarity index; Whittaker, 1952).
A sediment trap (Knauer et ai, 1979; modified) was used for cyst flux estimates.It consisted of two cylindrical Plexiglas collectors (height 52 cm, diameter 7 cm,aspect ratio 7.4) fastened to a gimballed metallic array that ensured their verticalposition. Both collectors were filled with filtered sea water, adjusted to 40 PSUby the addition of reagent grade NaCl. Neutralized formol was added to one of
2293
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

M.Montresor, A.Zingone and Djaino
the collectors to a final concentration of 2% formaldehyde. The trap wasdeployed at 25 m depth at station T (Figure 1). This station (40 m depth) waslocated <1 mile from the long-term monitoring station MC, which was theminimal safety distance from the routes of the marine traffic of the gulf. The trapwas operating for a total of 682 days, from 29 August 1994 until 10 October 1996,and the collectors were substituted by a scuba diver, generally every 15 days.Traps were lost on two occasions, resulting in sampling gaps from 26 October to30 November 1994 (35 days) and from 19 April to 15 May 1995 (57 days).
In the laboratory, fixed trap samples were mixed and two subsamples wereplaced in the refrigerator until examination, usually within 2 weeks from theircollection. For cyst identification and enumeration, 20 ml of each subsample weregently sonicated (60 W) for 15 s and 4 ml subsamples were examined in an Uter-mohl counting chamber using a Zeiss IM35 inverted microscope. Cyst fluxes wereexpressed as daily flux per square metre. Empty cysts, presumably deriving fromresuspension, were not included in flux estimates and graphs. A total of 200 mlof the unpreserved trap material were placed in the refrigerator in the dark andused for germination experiments aimed at identifying the corresponding vege-tative stages. For this purpose, selected cysts were isolated singly and cultureswere established as indicated in Montresor (1995).
Cysts were identified to the species level whenever possible. The biologicaltaxonomy system is used throughout this paper, with the exception of morpho-types for which the corresponding vegetative stage is unknown; these are namedfollowing the paleontological taxonomy system. Unidentified morphotypes wereclassified on the basis of their most relevant morphological features (colour,shape, size, etc.).
Surface phytoplankton samples were collected weekly with Niskin bottles fromFebruary 1995 at station MC. Samples were fixed with neutralized formol to 1.6%formaldehyde concentration and kept in the dark at 4°C until examination,usually within a few weeks. For cell enumeration, subsamples of 1-100 ml wereallowed to settle for 1-3 days in combined chambers and observed under a ZeissAXIOVERT optical microscope equipped with phase contrast. The most abun-dant species were enumerated at X400 magnification on two transepts of thesettling chamber bottom (1/33 of the total chamber bottom). Rarer species werecounted over a fixed subsample of 100 ml at X200 magnification on two transepts(1/16 of the total chamber bottom).
Phytoplankton were identified at the species level whenever possible. Uniden-tified specimens were assigned to suprageneric groups as indicated in Zingone etal. (1995). For biomass estimation, cell biovolumes were calculated frommeasurements taken on the samples, occasionally integrated with literature data,and converted into cell carbon (Strathmann, 1967).
Results
Species composition
A total of 59 different cyst morphotypes were recorded during this study, 52 ofwhich were identified at least to the genus level (Table II). Of these, 24 were
2294
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

Dinoflagellate cyst prodnction at a coastal she
Table n. Check-list of the cysts recorded in the Gulf of Naples
Biological taxonomy system Paleontological taxonomy system
Calcareous Peridiniales
Calciodinellum operosum (Deflandre)Montresor, Janofske and Willems
Pentapharsodiniutn tyrrhenicum (Balech)Montresor, Zingone and Marino
Scrippsiella crystallina LewisS.lachrymosa LewisS.precaria Montresor and ZingoneS.ramonii MontresorS. rotunda LewisS.trochoidea (Stein) Loeblich III (big)S.trochoidea (Stein) Loeblich III (medium)S.trochoidea (Stein) Loeblich III (small)calcareous cyst type2Organic (non-calcareous) Peridinalescf. Dissodium parvum Abe (=Diplopelta parva (Abe) Matsuoka)Diplopsalis group spp.cf. Ensiculifera imariense Kobayashi
and MatsuokaProtoperidinium americanum (Gran and
Braarud) BalechP.avellanum (Meunier) Balech
P.daudicans (Paulsen) BalechP.compressum (Abe) BalechP.conicum (Gran) Balech
P.divaricatum (Meunier) Parke and DodgeP.leone (Pavillard) BalechP.nudum (Meunier) Balech
P.oblongum (Aurivillius) Parke andDodge
P.pentagonum (Gran) Balech
P.subinerme (Paulsen) Loeblich III
Protoperidinium sp. # 6Protoperidinium sp. # 9Protoperidinium sp. # 16Protoperidinium sp. # 15Protoperidinium sp. # 17Protoperidinium sp. # 18 (round grey)Protoperidinium sp. # 19(cf. P.monospinum)Protoperidinium sp. # 20Preperidinium meunierii (Pavillard)Elbrachter (= Zygabikodiniumlenticulatum Loeblich and Loeblich)Round brown cysts
Bicarinellum tricarinelloides VersteeghCalcigonellum infula DeflandreCalciodinellum operosum Deflandre
Calciperidinium asymmetricum VersteeghFollisdinellum splendidum VersteeghCalcicarpinum bivalvum Versteegh
Praecalcigonellum schizosaeptumVersteegh
? Rhabdothorax Kamptner? Rhabdothorax Kamptner? Rhabdothorax Kamptner
Brigantedinium cariacoense (Wall)Lentin and WilliamsVotadinium spinbsum ReidStelladinium reidii BradfordSelenopemphix quanta (Bradford)MatsuokaXandarodinium xanthum ReidQuinquecuspis concreta (Reid) HeadSelenopemphix quanta (Bradford)Matsuoka
Trinovantedinium applanatum (Bradford)Bujak and DaviesSelenopemphix alticincta (Bradford)MatsuokaBrigantedinium sp.Brigantedinium sp.
2295
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

MJVf ontresor, A.Zingone and Djamo
Table n. Continued
Biological taxonomy system Paleontological taxonomy system
GonyaulacalesAlexandrium andersoni BalechGonyaulax scrippsae Kofoid
Gonyaulax sp.Gonyaulax sp.
Lingulodinium polyedrum (Stein) Dodge
Protoceratium reticulalum (Claparedeand Lachmann) BiitschliGymnodinialescf. Gymnodinium catenatum GrahamPolykrikos kofoidii ChattonP.schwartzii BiitschliGymnodiniales type aGymnodiniales type bUnidentified morphotypesOval brown type aRound beigeRound brown, smallRound green type aRound smooth reddishRound spiny reddishRound reticulatedUnknown cysts organic
Spiniferites bulloideus(Deflandre and Cookson)Sarjeant sensu Wall and Dale 1968Spiniferites mirabilis (Rossignol) SarjeantSpiniferites membranaceum (Rossignol)SarjeantUnidentified Spiniferites spp.Lingulodinium machaerophorum(Deflandre and Cookson) WallOperculodiniwn centrocarpum(Deflandre and Cookson) Wall
S, surface sediments; T, sediment trap.
found in both sediment and trap samples, 21 only in sediment and 14 only in trapsamples.
The large majority of cyst types found in the gulf belong to the order Peri-diniales, which includes both phototrophic and heterotrophic taxa. Among them,calcareous cysts were particularly important in terms of abundance and speciesdiversity. We found 12 different calcareous morphotypes, attributable to twogenera, Scrippsiella and Pentapharsodinium, which are presently grouped in thesubfamily Calciodinelloideae (Fensome et ai, 1993) along with Ensiculifera andother fossil calcareous cysts. In both sediment and trap samples, the mostcommon calcareous cysts were oval in shape and had short, pyramidal spineswhich are typical of S.trochoidea (Figure 2a). These exhibited a rather wide rangeof pigmentation and size, and were tentatively grouped into three differentclasses: greenish cysts with a mean length of 20 um and brown cysts with a meanlength of either 30 or 50 um, respectively. Vegetative cells obtained from thegermination of greenish cysts were small (mean length 20 um), lightbrown-greenish in colour, and had the same plate pattern of S.trochoidea.Medium-sized cysts produced larger (mean length 25 um), yellowish-brown cells,in some cases differing from typical S.trochoidea by having a spine on the sulcal
2296
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

Dinoflagellate cyst production at a coastal she
Fig. 2. Selected calcareous cysts recorded in the Gulf of Naples, light micrographs: (a) Scrippsiellatrochoidea, (b) S.precaria, (c) Sxrystallina, (d) Pentapharsodinium tyrrhenicum, (e) Bicarinellumbicarinelloides, (I) Calciodinellum operosum. Scale bars 10 um.
anterior platelet (MMontresor, unpublished data). Attempts to germinate thelargest cysts were unsuccessful.
Calcareous cysts also included other spiny morphotypes {Scrippsiella precaria,Figure 2b; and S.ramonii) as well as cysts surrounded by more or less irregularlyshaped polygonal crystals (e.g. Scrippsiella crystallina, Figure 2c; and S.lachry-mosa). A peculiar group of non-spiny calcareous cysts, with rather complexcoverings formed by minute crystals, was also well represented in the gulf (e.g.Pentapharsodinium tyrrhenicum, Figure 2d; Bicarinellum tricarinelloid.es, Figure2e; Calciodinellum operosum, Figure 2f). These cysts are mainly reported fromfossil records (Versteegh, 1993) and the vegetative stage is known only for a fewof them (Montresor et al., 1993,1994,1997). For others, only the micropaleonto-logical name and description are available.
Non-calcareous peridinioid cysts very similar to those of Ensiculifera imariensewere found which did not germinate. All the other non-calcareous Peridinialescysts were attributable to heterotrophic species, mainly belonging to the genusProtoperidinium. In some cases, identification to the species level was possiblefrom their morphology (Figure 3a and b); others were attributed to this genusafter germination experiments (Figure 3c) or based on their affinity with thepaleontological genus Brigantedinium. Round, brown, smooth-walled cysts(Figure 3d), which cannot be identified with certainty unless their archeopyles arevisible, were all ascribed to Protoperidinium. A few morphotypes belonging tothe heterotrophic "Diplopsalis group' were also occasionally found.
2297
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

M-Montresor, A-Zingone and D-Sarno
Fig. 3. Selected non-calcareous cysts recorded in the Gulf of Naples, light micrographs: (a) Protoperi-dinium claudicans, (b) P.subinerme, (c) P. cf. monospinum, (d) round, brown cyst, (e) Gymnodinialestype a, (I) Gymnodiniales type b, (g) Cymnodinium catenarum-\ike, (h) Protoceratium reticulatum, (i)Alexandrium andersoni. Scale bars 10 nm.
Other organic cyst types, generally produced by species included in the ordersGymnodiniales and Gonyaulacales, were represented in the area with -10species. The first order mainly included small-sized (15-20 um), either smooth(type a; Figure 3e) or 'hairy', cysts (type b; Figure 3f) which produced smallphototrophic cells. We also found empty cysts with surface microreticulation andarcheopyle shape similar to those of Gymnodinium catenatum (Figure 3g).However, it has recently been found that this cyst morphotype can be producedby several distinct species (Bravo, 1997; C.Bolch, personal communication).
Among Gonyaulacales, cysts of Protoceratium reticulatum (Figure 3h) andLingulodinium polyedrum were common but never abundant, the rest beingmainly constituted by Spiniferites cysts. In trap samples, spherical, smooth-walledcysts were also found which germinated into Alexandrium andersoni (Figure 3i).
2298
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

Dinoflagellate cyst production at a coastal site
This species has been described from the eastern US coast and reported as non-toxic (Anderson et al., 1990; Balech, 1995), but a preliminary HPLC analysis onour material has revealed the presence of paralytic shellfish poisoning (PSP)toxins (P.Ciminiello, personal communication).
Spatial distribution of cyst assemblages
Sediment analysis showed that calcareous and organic Peridiniales cysts consti-tuted the large majority of the assemblages (from 81 to 94% of the total number)at all of the stations sampled (Figure 4).
Stations closer to the coast (stations 1-5 and MC) were characterized by themarked dominance of calcareous cysts. Among these, S.trochoidea was the mostcommon and dominated the whole assemblage at the two stations near themouths of the harbour (station 1 and 2). Moving from the harbour (stations 3-5and MC), a more diversified calcareous assemblage was found, with S.trochoideaco-occurring with S.crystallina, Pentapharsodiniwn tyrrhenicum and B.tricarinel-loides. Non-calcareous Peridiniales constituted on average 25% of the total,whereas Gonyaulacales, mainly constituted by L.polyedrum, were proportionallymore abundant only at station 6. Gymnodiniales, mainly represented byPolykrikos spp., never attained >5.6% (station 5) of total numbers. A few cysts
s t 2 s t 1
s t 3st5 stMC
s t 6
s t 8
s t 7
I Scrippsiella trochoidea
| other calcareous Peridinales
|Q] non-calcareous Peridiniales
• Gonyaulacales
• Gymnodiniales
S undet cysts
Fig. 4. Percentage composition of the major groups of dinoflagellate cysts in surface sediments. Thesediment trap station (T) is marked with an asterisk.
2299
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

M-Montresor, A.Zingone and D-Sarno
with microreticulate markings like those of G.catenatum were found at stations3,4,5 and MC.
At the deepest stations (stations 6-8), organic non-calcareous Peridinialesshowed the highest species diversity and were more important as a percentage,dominating or co-dominating the assemblages along with calcareous cysts. Theywere mainly constituted by Protoperidinium spp. (up to 44.7% at station 8),among which P.leonis, P.subinerme, P.avellana and P.conicum often attained rela-tively high percentages. A significant fraction of the Protoperidinium cysts(16-55%) consisted of round, brown cysts. Among the Gonyaulacales, L.poly-edrum reached a maximum of 5.3% of total numbers at station 6, while otherGonyaulacales (Spiniferites spp.) were scarce at all of the sampling sites.
The diversity index (//') ranged from 2.8 to 4.2. Highest values (3.9-4.2) wererecorded at western stations (stations 6-8). Lowest diversity, due to the strongdominance of S.trochoidea, was recorded at stations 1 (//' = 2.8) and 2 (//' = 3.1).The hierarchical clustering confirmed the distinct separation of the stationslocated closer to the mouths of the harbour (Figure 5). Two other clustersincluded the other coastal stations and those located in the western part of thegulf, respectively.
Cyst flux seasonality
Table III shows total flux values for the different groups of cysts integrated overthe first and second year of sampling, respectively. Despite gaps in the sampling,flux values for the first year exceeded those of the second by a factor close to threeor more for most groups. The percentage composition of the cyst flux integratedover, the whole sampling period is similar to that found in the sediments at themost coastal stations (Figure 4; stations 1, 2 and 4). Calcareous Peridiniales,mainly represented by S.trochoidea, dominated the integrated percentagecomposition (57.5%). Non-calcareous Peridiniales were slightly less common inthe traps (13.4%), whereas Gonyaulacales (9.1%) and especially Gymnodiniales
ons
<3
ng
ssa
mpi
i
8
7
6
bMC
3
2
4
1
'
11
0.15 0.2 0.25 0.3 0.35 0.4 0.45 0.5 0.55 0.6 0.65
linkage distance
Fig. 5. Hierarchical clustering of sediment sampling stations, based on cyst assemblage dissimilarity.
2300
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

Dinoflagellate cyst production at a coastal site
Table UL Cyst fluxes (cysts X 106 nr2) measured over two annual cycles
Sept. 1994-Aug. 1995 Sept. 1995-Aug. 1996
Days of deploymentScrippsiella trochoideaOther calcareous PeridinialesNon-calcareous PeridinialesGonyaulacalesGymnodinialesUnidentified cysts
Total
26857.4819.8518.6113.3714.891.63
125.82
37220.936.235.612.86
16.672.31
54.60
(17.8%) were relatively more abundant in trap samples as compared tosediments.
Cyst fluxes showed a marked seasonal variability (Figure 6) with periods ofhigher fluxes from early spring throughout late autumn (up to 17 X 10* cysts m~2
day1 in September 1994 and 11 X 105 cysts nr2 day1 in March 1995). FromDecember onwards, cyst fluxes decreased markedly in both years, for a periodlasting until the end of February and April in the 2 years, respectively. A contri-bution of resuspension events to the trap content cannot be excluded inwinter-early spring, when the water column is mixed and storms can occur in thearea. However, the peak in early spring 1995 coincided with a period of relativelycalm weather.
Species composition in the traps was quite variable throughout the year (Figure6, Table IV). In late summer-early autumn of both years, when daily fluxesranged from 0.9 to 15.5 X 105 cysts m~2 day1, calcareous resting stages dominated.These were mainly represented by small- and medium-sized S.trochoidea andPentapharsodinium tyrrhenicum cysts, followed by Scrippsiella rotunda and largeS.trochoidea, accompanied by a set of other different calcareous morphotypes.Several Protoperidiniwn species, including P.conicum, P.claudicans and bothsmooth and spiny, round, unidentified Protoperidinium cysts, were also numer-ous in the trap samples of this period, dominating the whole trap content at theend of November-beginning of December. From this period onwards, trapcontent was scarce and mainly constituted by empty cysts, most probably deriv-ing from resuspension of sediments. The few apparently viable cysts includedelements of all groups. In February-March 1995, a peak of 11 X 105 cysts nr2
day1 was recorded which was almost entirely (up to 96%) due to calcareousmorphotypes. These mainly consisted of large S.trochoidea (up to 8.8 X 105 nr2
day1 in early March) followed by the smaller-sized S.trochoidea (up to 2.5 X 105
nr2 day1 in late March) and S.crystallina (1.6-1.3 X 10s nr2 day1 in March). Inspring 1996, cyst fluxes were almost undetectable until April. Late spring andsummer samples of both years were characterized by diversified assemblages,with the conspicuous presence of Gymnodiniales, which at times dominated,attaining daily fluxes of 4.7 X 105 cysts irr2 day1 in July 1995. In these periods,calcareous cysts were still abundant, dominating in August and in September, and
2301
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

MJVIontresor, A^Zingone and Diaroo
Table IV. Monthly distribution of dinoflagellate cysts in sediment trap samples. For each month, thehighest mean daily flux value recorded over 2 years was considered
Calcareous PeridinialesBicarinellum tricarinelloides*Calcigonellum infula*Calciodinellum operosumCalciperidinium asymmetricum*Follisdinellum splendidum*Pentapharsodiniwn tyrrhenicumPraecalcigonellum schizosaeptum*Scrippsiella crystallinaS.lachrymosaS.precariaS.ramoniiS. rotundaS.trochoidea (big)S.trochoidea (medium)ytrocnoiaea (small)Calcareous cyst type 2Organic (non-calcareous) PeridinialesDiplopsalis group spp.cf. Ensiculifera imarienseProtoperidinium claudicansP.conicumProtoperidinium sp. # 18(round grey)Protoperidinium sp. # 19(cf. P.monospinum)Protoperidinium sp. # 20Round brown cystsGonyaulacalesAlexandrium andersoniGonyaulax scrippsae(S.bulloideus*)Gonyaulax sp.(cf. S.membranaceum*)Protoceratium reticulatumSpiniferites spp.GymnodinialesPolykrikos schwartziiGymnodiniales type aGymnodiniales type bUnidentified morphotypesOval brown type aRound brown smallRound green type aRound smooth reddishRound spiny reddishRound reticulatedUnknown organic cysts
J
X
X
X
F M A M
X
X X X
X X
m m xX
X
x x am • a xx a a
X
X
a aX X X
X X
X X X X
X X
am m
X
X
ax a
Ju
X
aaa
X
mmmm
u
m•
aa
JI
X
X
X
X
X
mm
X
m
m
mmX
a
A
aX
m
X
mmX
X
X
X
mmX
23•="3
JfJ
X
a
s
X
X
X
X
aX
ama
m
*3
mX
am
mX
X
X
X
X
aX
O N D
X X
X X
X
X
a x aX
X
X
X X
aa x sia x x
X
x
am x mm x
m xa a sX
X X
X
M x
aXX
a xX
X X
'x', flux values 5 0.1 x 105 cysts nr2 day-1; 'grey', 0.1 < flux values < 2.5 x 10s cysts nr2 day1; 'black',flux values > 2.5 x 105 cysts nr2 day1.'Paleontologic name.
2302
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

Dinoflagellate cyst production at a coastal site
were mainly represented by small and medium-sized S.trochoidea. The presenceof Gonyaulacales cysts was restricted to the warmest months, from July toSeptember, and was mainly due to Protoceratium reticulatum (September 1994and July 1995) and A.andersoni (<0.5 X 105 nr2 day"1 in August and September1996).
Species-specific seasonal patterns for cyst production were detected (Table TV)in both years. Small and medium-sized S.trochoidea and the two Gymnodinialescyst types showed a high production rate almost uninterruptedly over severalmonths. Other species had a more restricted cyst production period in latesummer-autumn, as in the case of Pentapharsodinium tyrrhenicum and other lessabundant calcareous cysts, or confined to a few weeks in summer as in the caseof A.andersoni and Protoceratium reticulatum. It was not possible to unravel anyspecies-specific pattern for the heterotrophic dinoflagellates, due to the pro-nounced morphological convergence of their resting stages, probably hidingseveral different species.
| calcareous Peridiniales| Gonyaulacalesnon calcareous Peridiniales
( U Gymnodiniales
s o1995
Fig. 6. Mean daily cyst flux at station T over the sampling period; missing trap samples are markedby a horizontal bar.
2303
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

M.Montresor, A~Zingone and D.Samo
Cysts and phytoplankton
Phytoplankton sampling at station MC was resumed 6 months after the sedimenttrap had been deployed, following a period of interruption. Dinoflagellates,consisting of -90 identified species or supraspecific taxa, constituted as usual aquite limited part of the phytoplankton population in terms of cell numbers,although they often exceeded the other phytoflagellates in terms of biomass.Relatively high concentrations were estimated as of late February in the first year,and peak values were reached in late spring and summer of both years (Figure7). Small (<15 urn), unidentified, unarmoured species represented the majority ofthe population, attaining maximal values from June until late July. Amongthecate species, Prorocentrum triestinum and Heterocapsa niei reached relativelyhigh concentrations (<6 X 105 and 6 X 104 cells I"1, respectively), whereas otherperidinioids rarely exceeded 1.5 X 104 cells H.
Of the dinoflagellates identified to species level, 18 are known to form cystsand, of these, 13 were found in this study. These species did not contribute signifi-cantly to the motile population, only once exceeding 10% of total dinoflagellatecell numbers. This estimation increases, in terms of both species number andcontribution to the motile dinoflagellate cell numbers, if small, cyst-producinggymnodinioids are included. Figure 8 shows concentrations for small gymnodin-ioids at station MC, which presumably included a number of non-cyst formingspecies, and the flux of gymnodinioid cysts at station T. Cell numbers were aver-aged over periods comparable to the corresponding sampling intervals of thetrap. For both resting and vegetative stages, a common pattern was evident, withlow autumn and winter abundances, and an increase from mid-spring throughoutsummer. The low number of these cysts as compared to vegetative cells indicatesthat either a very limited number of species form cysts or that encystment ratesare extremely low for these species.
• other dinoflagellatesa gymnodinioids• Scrippsiella-Wke
0.0 7 >iTi11111111 i 11111 n
F M A M M J J A A S O O N O J F F M A M M J J A A S
1995
Fig. 7. Dinoflagellate abundance at station MC.
2304
1996
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

Dinoflagellate cyst production at a coastal site
I gymnodinioids
-gymnodinioid cysts
F M A M J J A S O N D J F M A M J J A S
Fig. 8. Mean daily flux for gymnodiniod cysts at station T and small-sized gymnodinioid abundanceat station MC, the latter integrated over the corresponding trap sampling intervals. Missing phyto-plankton samples are marked by an asterisk.
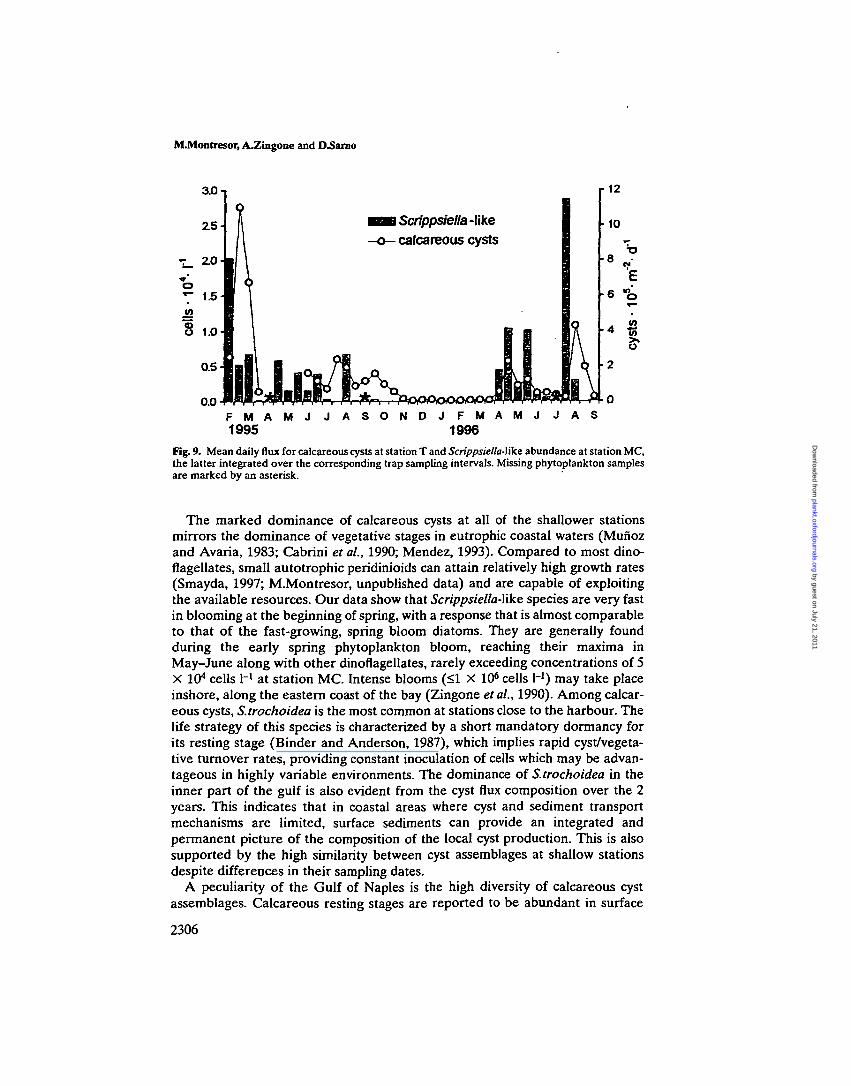
Our findings of cyst-producing dinoflagellates in plankton samples were toosporadic to allow for comparisons with trap data. The co-occurrence of cysts andrelative planktonic stages can be traced only for calcareous peridinioid species(Figure 9). Within this group, only five species were occasionally identified inplankton samples, due to the remarkable similarity of the motile stages. Mostoften, all of the different calcareous cyst-producing species were grouped underScrippsiella-\ike spp. In spring 1995, these species showed an increase in surfacewaters which was followed by the above-mentioned peak in calcareous cyst flux.Cyst/vegetative stage ratios appeared to be higher in autumn of both years and,generally, following most productive periods for surface populations.
Several species found in either sediment or trap samples were not recorded insurface waters over the sampled period. These also included some occasionallyabundant cysts such as A.andersoni and Protoceratium reticulatum. Some species,mainly belonging to Gonyaulax, Protoperidinium and Diplopsalis group, have attimes been observed during previous investigations in the area, but in very lowconcentrations (A.Zingone, unpublished data). Others, such as A.andersoni,several Protoperidinium spp. and several peridinioids producing calcareous cysts,have never been recorded in the area before.
Discussion
Dinoflagellate cysts in the Gulf of Naples form a quite diversified assemblage, withwell-defined patterns of spatial and temporal occurrence. The cyst assemblageincludes 59 morphotypes belonging to widely distributed neritic species, encom-passing cold-temperate to tropical taxa (Wall et al., 1977; Dale, 1983,1996; Dodgeand Harland, 1991).
2305
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from
M.Montresor, A.Zingone and D-Sarno
Scnppsiella -likecalcareous cysts
F M A M J J A S O N D J F M A M J J A S
Fig. 9. Mean daily flux for calcareous cysts at station T and Scrippsiella-like abundance at station MC,the latter integrated over the corresponding trap sampling intervals. Missing phytoplankton samplesare marked by an asterisk.
The marked dominance of calcareous cysts at all of the shallower stationsmirrors the dominance of vegetative stages in eutrophic coastal waters (Mufiozand Avaria, 1983; Cabrini et ah, 1990; Mendez, 1993). Compared to most dino-flagellates, small autotrophic peridinioids can attain relatively high growth rates(Smayda, 1997; M.Montresor, unpublished data) and are capable of exploitingthe available resources. Our data show that Scrippsiella-like species are very fastin blooming at the beginning of spring, with a response that is almost comparableto that of the fast-growing, spring bloom diatoms. They are generally foundduring the early spring phytoplankton bloom, reaching their maxima inMay-June along with other dinoflagellates, rarely exceeding concentrations of 5X 104 cells I"1 at station MC. Intense blooms (<1 X 106 cells H) may take placeinshore, along the eastern coast of the bay (Zingone etal., 1990). Among calcar-eous cysts, S.trochoidea is the most common at stations close to the harbour. Thelife strategy of this species is characterized by a short mandatory dormancy forits resting stage (Binder and Anderson, 1987), which implies rapid cyst/vegeta-tive turnover rates, providing constant inoculation of cells which may be advan-tageous in highly variable environments. The dominance of S.trochoidea in theinner part of the gulf is also evident from the cyst flux composition over the 2years. This indicates that in coastal areas where cyst and sediment transportmechanisms are limited, surface sediments can provide an integrated andpermanent picture of the composition of the local cyst production. This is alsosupported by the high similarity between cyst assemblages at shallow stationsdespite differences in their sampling dates.
A peculiarity of the Gulf of Naples is the high diversity of calcareous cystassemblages. Calcareous resting stages are reported to be abundant in surface
2306
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

Dinoflagellate cyst production at a coastal site
sediments of several other coastal areas, such as the Danish coast (EUegaard etal., 1994), the German Bight (Nehring, 1995), some Spanish rias (Blanco, 1995),and the Japanese (Ishikawa and Taniguchi, 1996) and Australian (Bolch andHallegraeff, 1990; Sonnemann and Hill, 1997) coasts. However, the records ofmorphotypes other than S.trochoidea are very scattered in these areas. Theunusual and distinctive cysts of Pentapharsodinium tyrrhenicum and B.tricarinel-loides, which are rather common in the Gulf of Naples, have only sporadicallybeen recorded in warmer/temperate waters where workers have used non-palynological methods (Wall and Dale, 1968a; Dale 1992; Sonnemann and Hill,1997). Interestingly, many of the calcareous cysts recorded in the gulf are alsocommon in Mediterranean fossil sediments dating back to the Pliocene andPleistocene (Versteegh, 1993), which attests that this high diversity has beenpreserved in the basin for a long time. It can be hypothesized that the rather highalkalinity of Mediterranean waters (Delgado and Estrada, 1994) can favourbiomineralization and maintain a great diversity of species with calcareous cover-ings. However, information on the biogeography of calcareous cysts is extremelylimited, since the widely used acid treatment of sediment samples dissolves thecalcium carbonate, leaving only featureless cysts of difficult classification.
The close association between coastal conditions and calcareous cysts,although repeatedly reported, has some exceptions. In our area, a marked quali-tative shift in cyst assemblages is observed slightly offshore, at stations in thewestern part of the gulf, where non-calcareous peridinioids are dominant due tothe decline of calcareous cysts. This spatial pattern marks the coast-offshore tran-sition towards the oligotrophic, open Tyrrhenian waters, that can penetratedeeply in the gulf for a large part of the year (Marino et al., 1984). Similarly, differ-ent Spanish Atlantic rias show a marked heterogeneity (Blanco, 1995) for cystassemblages, some of which are dominated by S.trochoidea, others by non-calcareous cysts such as Alexandrium spp. On the other hand, calcareous restingstages are not exclusively found in coastal waters, but can also constitute animportant fraction of cyst assemblages in open oceanic waters (Dale, 1992). Here,the most abundant species are Thoracosphaera albatrosiana, a cyst of stillunknown biological affinity, followed by Calciodinellum operosum, while spinyS.trochoidea-\ike morphotypes are almost absent. The production of heavilymineralized resting stages over the deep sea is puzzling, and the life strategy ofthese species deserves investigation to understand how they cope with a presum-ably fast and no-return sinking of their cysts in the ocean depth.
We have estimated that 16% of the species recorded as vegetative stages in theplankton were present as resting stages in either sediment samples or in the traps.If all the 250 species ever recorded at the fixed station MC are considered, 20%can form resting stages and 10% of the total dinoflagellates were recorded as cystsin the present study. At the same time, a high number of dinoflagellate specieswere found through this sampling approach which had never been found in theplankton of the area before. In fact, the benthic sampling efficiently integratestime and space scales which are not appropriately sampled for planktonic stages.The present case demonstrates that species undetected in surface waters can alsoattain high cyst production rates, as is the case for Protoceratium reticulatum and
2307
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

M.Montresor, A-Zingone and Djamo
A.andersoni. Luckily, many toxic dinoflagellates produce resting cysts, and sedi-ment surveys or trap sampling represent an alternative, valuable tool for theirdetection. In this case, the presence of two potentially toxic dinoflagellates,A.andersoni and G.catenatum-like species, has first been detected in the gulfthrough cyst sampling.
For some planktonic organisms, such as neritic copepods, a latitudinal gradi-ent in reproductive strategies has been reported, with a higher proportion ofspecies producing resting eggs at high latitudes (Dahms, 1995). A higher numberof dinoflagellate species with a cyst-based strategy might be expected forenvironments that experience extremely short day lengths and lower tempera-ture minima, if dinoflagellate cysts are to be interpreted as a means for increasedsurvival in unfavourable periods. However, estimates in the Oslofjord (Dale,1976) and in the German and Kiel Bights (Nehring, 1997) resulted in comparablepercentages of cyst-forming species (15, 25 and 20%, respectively), despitemarked differences in seasonal constraints between these geographic areas.
Resting stage production has at times been considered an important traitaccounting for the marked seasonality observed for phytoplankton populationsin temperate waters (Boero, 1994). We present data on high cyst production ratesin dinoflagellates, but also show that only a limited fraction of the species has acyst-based life history. The other species must have alternative overwinteringstrategies. This could include the production of resistant and quiescent life stagesnot distinguishable from vegetative cells, remaining in the water column at unde-tectable concentrations, a strategy known for some diatoms (Peters, 1996).Apparently, the formation of resting stages is not necessarily related to the capa-bility to attain higher concentrations, nor to a timely seasonal occurrence of aspecies in the plankton. In fact, many abundant and also seasonally recurrentdinoflagellates do not produce resting cysts. This is the case for Prorocentrumtriestinum, H.niei and Protoperidinium diabolus in the Gulf of Naples, and forother dinoflagellates in different geographic areas (e.g. Smetacek, 1985; Hesse etal., 1995).
Sediment trap data have shed light on the seasonal trend of cyst productionand on different species-specific patterns. In both years, cyst production ismaximal from late spring to early autumn for the majority of the species, butdifferences in temporal patterns of production are evident. Some species{S.trochoidea and gymnodinioids) show a high cyst production rate almost unin-terruptedly over several months (multivoltine reproduction), while others(Pentapharsodinium tyrrhenicum, A.andersoni and Protoceratium reticulatwri)have a more restricted cyst production period (univoltine reproduction). It hasbeen postulated that coastal, highly variable environments can favour or selectpopulations with multivoltine strategies, while offshore, more stable environ-ments can select populations with a univoltine habit (Wyatt and Jenkinson,1997). Our data on cyst production, as well as the different excystment patternsobserved for a number of species in a Japanese bay (Ishikawa and Taniguchi,1997), demonstrate that the two strategies can co-exist in the same coastalenvironment. In fact, despite their marked unpredictability, coastal environ-ments in temperate or subtropical seas are subject to seasonal constraints which
2308
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

Dinoflagellate cyst production at a coastal site
can account for univoltine strategies in species with restricted ecologicalrequirements.
In some cases, we have observed a coupling between cyst flux and the presenceof motile stages in overlying waters. The simultaneous presence of small gymno-dinioid motile cells and cysts is recorded from early spring until September, buta more precise relationship cannot be tracked due to difficulties in distinguishingboth vegetative and cyst stages in this group of species. A more defined match isobserved in the early appearance and relatively high concentrations of Scripp-siella-Uke cells and respective calcareous cysts in spring 1995, which coincidedwith the anticipated phytoplankton spring bloom. A good relationship for thisgroup of species is also evident for the rest of the sampled period, with peaks ofcalcareous cysts following those of motile peridinioids by some weeks. In bothyears, cyst production and the cyst/vegetative ratio are higher towards the end ofsummer-beginning of autumn, as also observed for 5. trochoidea in a Japanese bay(Ishikawa and Taniguchi, 1996). Cyst production in temperate and cold-temper-ate environments has been shown to be either concomitant to the peaks of themotile stages (Wall and Dale, 1968b; Dale, 1977) or slightly delayed (Nordli, 1951;Wall and Dale, 1968b; Heiskanen, 1993). There are also cases with two seasonalpeaks of abundance, followed by cyst production (Anderson and Morel, 1979)and, as shown by our data, still other cases where both vegetative and restingstages are present over a large part of the year. These annual patterns can bemarkedly altered by latitudinal and/or environmental differences, as demon-strated by the different germination strategies for coastal and offshore popu-lations of Alexandriurn tamarense along the eastern US coast (Anderson andMorel, 1979; Anderson and Keafer, 1987).
These cases show that, for each species, the fraction of the population whichforms cysts is not constant over time, demonstrating the impossibility of inferringphytoplankton concentrations from cyst abundances. Several environmental andbiological factors have been advocated to control encystment rates, and henceaffect cyst/vegetative ratios in natural environments, including nutrient depletionor imbalance, temperature and light signals, and density-dependent factors(Pfiester and Anderson, 1989). In the Gulf of Naples, cysts are formed over arather wide temperature range (15-25°C) and highly fluctuating nutrient concen-trations (from 0.05 to 5.6 umol I"1 of inorganic nitrogen and 0.01-0.26 umol H ofphosphorus). Experimental data obtained for different species producing calcar-eous cysts have shown that shorter day lengths can notably augment cyst produc-tion rates (M.Montresor, unpublished data) which is in agreement with trap datashowing more frequent calcareous cyst peaks in autumn. To account for theseasonal and interannual variability of cyst production, a better understanding ofenvironmental factors driving cyst production is required. This is also a key pointfor a more effective exploitation of cyst data to interpret environmental andclimatic changes over long temporal scales.
Cyst production in dinoflagellates is a complex phenomenon, where species-specific characteristics interplay with environmental variables, producing anarray of diverse strategies. The present study combining trap data and sedimentassemblage composition provides a number of insights on this process and, as
2309
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

M.Montresor, A.Zingone and D-Samo
usual, poses further questions. It confirms that calcareous cysts can attain highproduction rates in coastal areas, and shows that diverse species-specificseasonal and spatial patterns are recognizable within this group as well as forother species. We have shown that signals recorded in the trap data reflect plank-ton variations in some cases, as for the anticipated spring in the first year ofsampling. This supports the possibility of recording plankton variations throughsediment trap sampling, possibly integrating the analysis with resting stages ofother planktonic organisms, which can be particularly useful in the monitoringof remote field stations where long-term plankton sampling is not applicable. Wehave provided evidence that the 'benthic' sampling of planktonic organismsoffers an opportunity for the detection of species which are rare in the watercolumn or scarcely identifiable as vegetative cells. The number of species thatcan be followed from their benthic stages is limited, but this approach may bevery useful for tracking the presence of cyst-producing toxic species. Studyingcyst distribution and in situ production has offered a tool to unveil hidden diver-sity in species composition, ecological performances and life strategies in thecyst-forming dinoflagellates.
Acknowledgements
The authors wish to thank Donato Marino for helpful discussion, Raffaele DeMaio for his professional assistance in modifying the sediment trap, and PasqualeSansone and Ciro Zazo for their help at sea in the deployment and periodicsampling of the sediment trap.
ReferencesAnderson.D.M. (1997) Diversity of harmful algal blooms in coastal waters. Limnol. Oceanogr, Ail,
1009-1022.Anderson.D.M. and Keafer,B.A. (1987) An endogenous annual clock in the toxic marine dinoflagel-
late Gonyaulax tamarensis. Nature, 325, 616-617.Anderson,D.M. and Morel,M.M. (1979) The seeding of two red tide blooms by the germination of
benthic Gonyaulax tamarensis hypnocysts. Estuarine Coastal Mar. Sci., 8,279—293.Anderson,D.M., Kulis,D.M., OrphanosJ.A. and Ceurvels,A.R. (1982) Distribution of the toxic dino-
flagellate Gonyaulax tamarensis in the southern New England region. Estuarine Coastal Shelf ScL,14, 447-458.
Anderson,D.M., KulisJD.M., SullivanJJ., Hall.S. and Lee.C (1990) Dynamics and physiology ofsaxitoxin production by the dinoflagellates Alexandrium spp. Mar. Biol, 104,511-524.
Balech.E. (1995) The Genus Alexandrium Halim (Dinoflagellata). Sherkin Island Marine Station,Sherkin Island, Co. Cork, Ireland.
Binder,BJ- and Anderson,D.M. (1987) Physiological and environmental control of germination inScrippsiella trochoidea (Dinophyceae) resting cysts. /. PhycoL, 23, 99-107.
BlancoJ. (1995) The distribution of dinoflagellate cysts along the Galician (NW Spain) coast. J.Plankton Res., 17,283-302.
BravoJ. (1997) The unique microreticulate cyst? Harmful Algae News, IOC NewsL, 16,4-5.Boero,F. (1994) Fluctuations and variations in coastal marine environments. PSZN1: Mar. EcoL, 15,
3-25.BoIch.CJ. and Hallegraeff,G.M. (1990) Dinoflagellate cysts in recent marine sediments from
Tasmania, Australia. Bot Mar., 33,173-192.Cabrini,M., MilaniJL and Honsell.G. (1990) Fioritura di Scrippsiella trochoidea (Stein) Loeblich III
nel Golfo di Trieste. Oebalia, 16,599-601.Carrada.G.C, Hopkins.T.S., Bonaduce.G., Ianora,A., MarinoJJ., Modigh,M., Ribera D'Alcal5,M.
2310
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

Dinoflagellate cyst production at a coastal site
and Scotto Di CarIo,B. (1980) Variability in the hydrographic and biological features of the Gulfof Naples. PSZNI: Mar. EcoL, 1,105-120.
Cembella,A.D., TurgeonJ., TherriaultJ.-C. and Berland,P. (1988) Spatial distribution of Proto-gonyaulax tamarensis resting cysts in nearshore sediments along the north coast of the lower St.Lawrence estuary. / Shellfish Res., 7, 597-609.
DahmsJH.-U. (1995) Dormancy in the Copepoda—an overview. Hydrobiologia, 306,199-211.Dale.B- (1976) Cyst formation, sedimentation, and preservation: factors affecting dinoflagellate
assemblages in recent sediments from Trondheimsfjord, Norway. Rev. Palaeobot. PalynoL, 22,39-60.
Dale3- (1977) New observations on Peridinium faeroense Paulsen (1905), and classification of smallorthoperidinioid dinoflagellates. Br. PhycoL J., 12, 241-253.
Dale3- (1983) Dinoflagellate resting cysts: 'benthic plankton'. In Fryxell,G.A. (ed.), Survival Strat-egies of the Algae. Cambridge University Press, Cambridge, pp. 69—136.
Dale3- (1992) Dinoflagellate contributions to the open ocean sediment flux. In Dale.B- andDale,A.L. (eds), Dinoflagellate Contributions to the Deep Sea. Ocean Biocenosis Series 5. WoodsHole Oceanographic Institute, Woods Hole, MA, pp. 1-43.
Dale.B- (1996) Dinoflagellate cyst ecology: modelling and geological applications. In JansoniusJ. andMcGregor j).C. (eds), Palynology: Principles and Applications. American Association of Strati-graphic Palynologists Foundation, Dallas, TX, Vol. 3, pp. 1249-1275.
Dale,B. and Dale,A.L. (1992) Dinoflagellate contributions to the sediment flux of the Nordic Seas. InDale.B- and Dale,A.L. (eds), Dinoflagellate Contributions to the Deep Sea. Ocean Biocenosis Series5. Woods Hole Oceanographic Institute, Woods Hole, MA, pp. 45-75.
Delgado.O. and Estrada,M. (1994) CO2 system in a Mediterranean frontal zone. Sci. Mar., 58,237-250.
Dodge JX). and Harland.R. (1991) The distribution of planktonic dinoflagellates and their cysts in theeastern and northeastern Atlantic Ocean. New Phytoi, 118, 593-603.
Edwards.L.E. (1992) New semiquantitative (paleo)temperature estimates using dinoflagellate cysts,an example from the North Atlantic Ocean. In Head,M.J. and WrennJ.H. (eds), Neogene andQuaternary Dinoflagellate Cysts and Acritarchs. American Association of Stratigraphic Palynolo-gists Foundation, Dallas, TX, pp. 69-87.
Ellegaard,M., Christensen,N.F. and Moestrup,0. (1994) Dinoflagellate cysts from recent Danishsediments. Eur. J. Phycoi, 29,183-194.
Fensome,R.A., Taylor,EJ.R., Norris.G., Sarjeant,W.A.S., Wharton,D.I. and Williams.G.I. (1993) AClassification of Living and Fossil Dinoflagellates. Micropaleontology Special Publication Number 7.Sheridan Press, Hanover, PA.
Hallegraeff.G.M. and Bolch.CJ. (1992) Transport of diatom and dinoflagellate resting cysts in ships'ballast water: implications for plankton biogeography and aquaculture. J. Plankton Res., 14,1067-1084.
Harland.R. (1983) Distribution maps of recent dinoflagellate cysts in bottom sediments from the northAtlantic Ocean and adjacent seas. Palaeontology, 26, 321-387.
Head,MJ. (1996) Modem dinoflagellate cysts and their biological affinities. In JansoniusJ. andMcGregor.D.C. (eds), Palynology: Principles and Applications. American Association of Strati-graphic Palynologists Foundation, Dallas, TX, Vol. 3, pp. 1197-1248.
Heiskanen,A-S. (1993) Mass encystment and sinking of dinoflagellates during a spring bloom. Mar.Bioi, 116,161-167.
Hesse,K.-J., Tillmann.U., Nehring,S. and Brockmann.U. (1995) Factors controlling phytoplanktondistribution in coastal waters of the German Bight (North Sea). In EleftheriouA-, Ansell,A.D. andSmith.CJ. (eds), Biology and Ecology of Shallow Coastal Waters. Olsen and Olsen, Fredensborg,pp. 11-22.
Ishikawa,A. and Taniguchi.A. (1996) Contribution of benthic cysts to the population dynamics ofScrippsiella spp. (Dinophyceae) in Onagawa Bay, northeast Japan. Mar. EcoL Prog. Sen, 140,169-178.
Ishikawa.A. and Taniguchi^A. (1997) In situ germination patterns, and bloom formation of somearmoured dinoflagellates in Onagawa Bay, north-east Japan. J. Plankton Res., 14,1783-1791.
Knauer.G.A., MartinJ.H. and BrulandJC.W. (1979) Fluxes of paniculate carbon, nitrogen, and phos-phorous in the upper water column of the northeast Pacific. Deep-Sea Res., 26A, 97-108.
MarinoJ)., Modigh,M. and Zingone,A. (1984) General features of phytoplankton communities andprimary production in the Gulf of Naples and adjacent waters. In Holm-Hansen.O., Bolis.S. andGilles,R. (eds). Marine Phytoplankton Productivity. Lecture Notes on Coastal and Estuarine Studies,Springer-Verlag, Berlin, 8, pp. 89-100.
Mendez,S.M. (1993) Uruguayan red tide monitoring programme: preliminary results (1990-1991). In
2311
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from

M.Montresor, A.Zingone and D.Sarno
Smayda,TJ. and Shimizu.Y. (eds), Toxic Phytoplankton Blooms in the Sea. Elsevier, Amsterdam,pp. 287-291.
Montresor,M. (1995) Scrippsiella ramonii sp. nov. (Peridiniales, Dinophyceae), a marine dinoflagel-late producing a calcareous resting cyst. Phycologia, 34,87-91.
Montresor,M., Zingone A- and Marino^D. (1993) The calcareous resting cyst of Pentapharsodiniumtyrrhenicwn comb. nov. (Dinophyceae). /. PhycoL, 29, 223-230.
Montresor,M., Montesarchio.E., Zingone,A. and MarinoJ). (1994) Calcareous dinoflagellate cysts inmarine sediments of the Gulf of Naples (Mediterranean Sea). Rev. PalaeoboL Palynol, 84, 45-56.
Montresor,M., JanofskeJ). and Willems,H. (1997) The cyst-theca relationship in Calciodinellumoperosum emend. (Peridiniales, Dinophyceae) and a new approach for the study of calcareouscysts. / PhycoL, 33,122-131.
MufiozJ?. and Avaria.S.P. (1983) Scrippsiella trochoidea (Stein) Loeblich III, nuevo organismocausante de marea roja en la Bahia de Valparaiso, Chile. Rev. BioL Mar. Valparaiso, 19, 63-78.
Nehring.S. (1995) Dinoflagellate resting cysts as factors in phytoplankton ecology of the North Sea.Helgol. Meeresunters., 49,375-392.
Nehring.S. (1997) Dinoflagellate resting cysts from recent German coastal sediments. Bot. Mar., 40,307-324.
Nordli,E. (1951) Resting spores in Gonyaulax polyedra Stein. Nytt Mag. Naturvidensk. B, 88,207-212.Peters,E. (1996) Prolonged darkness and diatom mortality: II. Marine temperate species. /. Exp. Mar.
BioL EcoL, 207, 43-58.Pfiester,L.A. and Anderson,D.M. (1987) Dinoflagellate reproduction. In Taylor,RJ.R. (ed.), The
Biology of Dinoflagellates. Blackwell Scientific, Oxford, pp. 611-648.Reid,P.C. (1975) A regional sub-division of dinoflagellate cysts around the British Isles. New Phytol.,
75,589-4503.Scotto Di Carlo3- et "I. (1985) Uno studio integrate) dell'ecosistema costiero del Golfo di Napoli.
Nova Thalassia, 7, 99-128.Shannon.C.E. and Weaver.G. (1949) The Mathematical Theory of Communication. University of
Illinois Press, Urbana, Chicago, IL, pp. 1-125.Smayda.TJ. (1997) Harmful algal blooms: Their ecophysiology and general relevance to phyto-
plankton blooms in the sea. Limnol. Oceanogr., 42,1137-1153.Smetacek.V. (1985) The annual cycle of Kiel Bight plankton: a long-term analysis. Estuaries, 6,
145-157.SonnemannJ.A. and Hill,D.R.A. (1997) A taxonomic survey of cyst-producing dinoflagellates from
recent sediments of Victorian coastal waters, Australia. Bot. Mar., 40,149-177.Strathmann,R.R. (1967) Estimating the organic carbon content of phytoplankton from cell volume
or plasma volume. LimnoL Oceanogr., 12,411-418.Versteegh.GJ.M. (1993) New Pliocene and Pleistocene calcareous dinoflagellate cysts from southern
Italy and Crete. Rev. Palaeobot Palynol., 78, 353-380.Versteegh.GJ.M. (1997) The onset of major Northern Hemisphere glaciations and their impact on
dinoflagellate cysts and acritarchs from the Singa section, Calabria (southern Italy) and DSDPHoles 607/607A (North Atlantic). Mar. MicropaleontoL, 30, 319-343.
Wall.D. and Dale.B. (1968a) Quaternary calcareous dinoflagellates (Calciodinellideae) and theirnatural affinities. /. Paleontol., 42,1395-1408.
WallJ). and Dale.B. (1968b) Modern dinoflagellate cysts and evolution of the Peridiniales. Micro-paleontology, 14, 265-304.
WallJX, Dale3., Lohmann.G.P. and Smith.W.K. (1977) The environmental and climatic distributionof dinoflagellate cysts in modem marine sediments from regions in the north and south AtlanticOceans and adjacent seas. Mar. MicropaleontoL, 2,121-200.
Whittaker,R.H. (1952) A study of summer foliage insect communities in the Great SmokeyMountains. EcoL Monogr., 22,1-44.
Wyatt,T. and JenkinsonJ.R. (1997) Notes on Alexandrium population dynamics. J. Plankton Res., 19,551-575.
ZingoneA-, Montresor,M. and MarinoJD. (1990) Summer phytoplankton physiognomy in coastalwaters of the Gulf of Naples. PSZNI: Mar. EcoL, 11,157-172.
ZingoneA-, Casotti,R., Ribera D'AlcalaJvI., Scardi,M. and MarinoJ). (1995) 'St. Martin's Summer':the case of an autumn phytoplankton bloom in the Gulf of Naples (Mediterranean Sea). /. PlanktonRes., 17, 575-593.
Received on February 3, 1998; accepted on July 7,1998
2312
by guest on July 21, 2011plankt.oxfordjournals.org
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















