
Enhanced reporter gene expression in the rat brain from helper virus-free HSV1 vectors packaged in...
16
Molecular Brain Research 90 (2001) 1–16 www.elsevier.com / locate / bres Research report Enhanced reporter gene expression in the rat brain from helper virus-free HSV-1 vectors packaged in the presence of specific mutated HSV-1 proteins that affect the virion a a a a a Tianzhong Yang , Guo-rong Zhang , Wei Zhang , Mei Sun , Xiaodan Wang , a,b, * Alfred I. Geller a Division of Endocrinology, Children’ s Hospital, 300 Longwood Avenue, Boston, MA 02115, USA b Program in Neuroscience, Harvard Medical School, Boston, USA Accepted 6 February 2001 Abstract Herpes simplex virus (HSV-1) gene expression is hypothesized to shut off promoters in HSV-1 vectors, but in a helper virus-free HSV-1 vector system, a number of promoters support only short-term expression. Thus, recombinant gene expression remains short-term in the absence of |99% of the HSV-1 genome. To resolve this paradox, we hypothesized that specific HSV-1 proteins that affect the virion can shut off recombinant gene expression. This study evaluated expression from HSV-1 vectors, containing neuronal-specific promoters, that were packaged in the presence of specific mutated HSV-1 proteins that affect the virion. The mutated HSV-1 proteins that were examined included two protein kinases (U 13 and U 3), the virion host shut-off factor ( vhs), the transactivator of immediate early promoters L S (VP16), and a virion protein that affects RNA metabolism (U 11). Helper virus-free packaging could occur in the presence of each S mutated protein alone or specific combinations of two or three mutated proteins. In BHK and PC12 cells, vectors packaged in the presence of each mutated protein increased ( |2-fold) the level of expression per cell, and vectors packaged in the presence of specific combinations of mutated proteins supported larger (4–7-fold) increases. In the rat striatum, vectors packaged in the presence of a mutated U 3 S displayed enhanced gene transfer (13–18-fold increases in the number of cells at 4 days), and vectors packaged in the presence of mutated U 13 or VP16 enhanced long-term expression (2 months). Vectors packaged in the presence of mutated vhs or U 11 displayed minimal L S changes in expression. 2001 Elsevier Science B.V. All rights reserved. Theme: Cellular and molecular biology Topic: Gene structure and function: general Keywords: Long-term expression; VP16; Protein kinase; Neurofilament promoter; Herpes simplex virus vector; Gene transfer 1. Introduction of cells that support expression within the first few weeks after gene transfer into the rat brain [56]. Interestingly, in Herpes simplex virus type one (HSV-1) vectors have HSV-1 plasmid vectors, two neuronal type-specific pro- potential for gene transfer into neurons, but the lack of moters (preproenkephalin and tyrosine hydroxylase (TH)), long-term expression remains a primary problem. Using and a chimeric promoter that contains 59 upstream se- either plasmid (amplicon) or recombinant HSV-1 vectors quences from the TH promoter, support some long-term that contain a wide range of viral or neuronal-specific expression (2–6 months). Although there is an initial large cellular promoters, there is a large decrease in the numbers decrease in expression, the remaining expression is rela- tively stable [22,27,49,53,56]. Thus, if the causes of this initial decrease in expression could be addressed, HSV-1 *Corresponding author. Tel.: 11-617-355-6185; fax: 11-617-734- plasmid vectors that contain specific promoters might 0062. E-mail address: geller [email protected] (A.I. Geller). support long-term expression. ] 0169-328X / 01 / $ – see front matter 2001 Elsevier Science B.V. All rights reserved. PII: S0169-328X(01)00059-6
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Enhanced reporter gene expression in the rat brain from helper virus-free HSV1 vectors packaged in...

Molecular Brain Research 90 (2001) 1–16www.elsevier.com/ locate /bres
Research report
Enhanced reporter gene expression in the rat brain from helpervirus-free HSV-1 vectors packaged in the presence of specific mutated
HSV-1 proteins that affect the viriona a a a aTianzhong Yang , Guo-rong Zhang , Wei Zhang , Mei Sun , Xiaodan Wang ,
a,b ,*Alfred I. GelleraDivision of Endocrinology, Children’s Hospital, 300 Longwood Avenue, Boston, MA 02115, USA
bProgram in Neuroscience, Harvard Medical School, Boston, USA
Accepted 6 February 2001
Abstract
Herpes simplex virus (HSV-1) gene expression is hypothesized to shut off promoters in HSV-1 vectors, but in a helper virus-free HSV-1vector system, a number of promoters support only short-term expression. Thus, recombinant gene expression remains short-term in theabsence of |99% of the HSV-1 genome. To resolve this paradox, we hypothesized that specific HSV-1 proteins that affect the virion canshut off recombinant gene expression. This study evaluated expression from HSV-1 vectors, containing neuronal-specific promoters, thatwere packaged in the presence of specific mutated HSV-1 proteins that affect the virion. The mutated HSV-1 proteins that were examinedincluded two protein kinases (U 13 and U 3), the virion host shut-off factor (vhs), the transactivator of immediate early promotersL S
(VP16), and a virion protein that affects RNA metabolism (U 11). Helper virus-free packaging could occur in the presence of eachS
mutated protein alone or specific combinations of two or three mutated proteins. In BHK and PC12 cells, vectors packaged in the presenceof each mutated protein increased (|2-fold) the level of expression per cell, and vectors packaged in the presence of specific combinationsof mutated proteins supported larger (4–7-fold) increases. In the rat striatum, vectors packaged in the presence of a mutated U 3S
displayed enhanced gene transfer (13–18-fold increases in the number of cells at 4 days), and vectors packaged in the presence of mutatedU 13 or VP16 enhanced long-term expression (2 months). Vectors packaged in the presence of mutated vhs or U 11 displayed minimalL S
changes in expression. 2001 Elsevier Science B.V. All rights reserved.
Theme: Cellular and molecular biology
Topic: Gene structure and function: general
Keywords: Long-term expression; VP16; Protein kinase; Neurofilament promoter; Herpes simplex virus vector; Gene transfer
1. Introduction of cells that support expression within the first few weeksafter gene transfer into the rat brain [56]. Interestingly, in
Herpes simplex virus type one (HSV-1) vectors have HSV-1 plasmid vectors, two neuronal type-specific pro-potential for gene transfer into neurons, but the lack of moters (preproenkephalin and tyrosine hydroxylase (TH)),long-term expression remains a primary problem. Using and a chimeric promoter that contains 59 upstream se-either plasmid (amplicon) or recombinant HSV-1 vectors quences from the TH promoter, support some long-termthat contain a wide range of viral or neuronal-specific expression (2–6 months). Although there is an initial largecellular promoters, there is a large decrease in the numbers decrease in expression, the remaining expression is rela-
tively stable [22,27,49,53,56]. Thus, if the causes of thisinitial decrease in expression could be addressed, HSV-1*Corresponding author. Tel.: 11-617-355-6185; fax: 11-617-734-plasmid vectors that contain specific promoters might0062.
E-mail address: geller [email protected] (A.I. Geller). support long-term expression.]
0169-328X/01/$ – see front matter 2001 Elsevier Science B.V. All rights reserved.PI I : S0169-328X( 01 )00059-6

2 T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16
The shut-off of recombinant gene expression may be 2. Materials and methodsrelated to the repression of viral gene expression that is ahallmark of latent HSV-1. This shut-off might be caused 2.1. Materialsdirectly by HSV-1 gene expression or indirectly by thecytopathic effects and inflammatory response due to HSV- Restriction endonucleases and T4 DNA ligase were1 gene expression [23,24,54]. Of note, gene transfer with obtained from New England Biolabs and Boehringerhelper virus-free HSV-1 vectors produces substantially less Mannheim. Dulbecco’s modified minimal essentialcytopathic effects and inflammatory response compared to medium, fetal bovine serum, G418, lipofectamine, andthose observed using helper virus systems [13,57]. How- OPTI-MEM I were obtained from GIBCO/BRL. 5-ever, helper virus-free vectors support modest, if any, Bromo-4-chloro-3-indoyl-b-D-galactopyranoside (X-gal)enhancements of long-term expression from a number of was obtained from Sigma. Rabbit anti-Escherichia coliviral or cellular promoters [13,53]. Thus, recombinant gene b-galactosidase antibody was obtained from ICN, andexpression remains short-term, even in the absence of mouse monoclonal anti-NeuN antibody was obtained from|99% of the HSV-1 genome, the LAT gene, and HSV-1 Chemicon. Fluorescein isothiocyanate-conjugated goat an-gene expression. ti-rabbit IgG and rhodamine isothiocyanate-conjugated
To resolve this paradox, we hypothesized that specific goat anti-mouse IgG were obtained from Jackson Immuno-HSV-1 proteins that affect the virion can shut off recombi- Research Laboratories.nant gene expression. Many studies on changes in geneexpression following HSV-1 infection have focused on the 2.2. Cells and virusesactivity of specific HSV-1 promoters in fibroblast cells(reviewed in Ref. [43]) rather than cellular promoters in The growth of baby hamster kidney fibroblast (BHK21)neurons. Thus, it is difficult to postulate detailed mechanis- cells, 2-2 cells [48], and PC12 cells [17] has beentic pathways by which specific HSV-1 proteins might described. KOS strain 5dl1.2 [33] was grown in 2-2 cellsaffect the activity of specific cellular promoters in neurons. and titered by plaque assay.Nonetheless, specific HSV-1 proteins that assist the virusin taking over a cell’s biosynthetic machinery have ac- 2.3. Plasmidstivities that might be capable of shutting off recombinantgene expression from HSV-1 vectors. Two HSV-1 encoded vhs pSc243 [25], VP16 in14 [1], U 11 MutB/421, andS
protein kinases (U 13 and U 3) phosphorylate specific U 11 MutG/421 [44] have been described.L S S
virion proteins located in either the tegument or the We isolated point mutations in U 13 and U 3 designedL S
envelope [6,8,9,36,38–41]. U 13 also phosphorylates at to lack protein kinase activity. A conserved Lys residue,L
least one cellular protein, elongation factor 1d [28], and present in subdomain II of all protein kinases, is requiredU 3 has anti-apoptotic functions [30]. U 13, but not U 3, for efficient phosphoryl transfer [19]. Mutation of this LysS L S
is present in virions. The virion host shut-off protein (vhs, is a standard method for inactivating protein kinases [19].U 41) degrades cellular mRNAs in fibroblast cell lines Two fragments that contain the U 13 mutation (ul13g,L L
[25,37,42] and may inhibit antigen presentation [20]. VP16 generates a Kpn I site) and extend 59 or 39 were produced(U 48), a tegument protein, is a powerful enhancer that using PCR (template, cos28 [7]; primers: 59 fragment,L
increases transcription from HSV-1 immediate early (IE) 59 GCGGGAT CCGT GTGTGTGCTGGTAATTTATACA -promoters and interacts with a number of cellular proteins CCG39 (nucleotides 27,056–27,082) [34] andand transcription factors [16]. U 11, another tegument 59CGCGGTACCATAAAGGAAAAGGAGTGGTTTGCC-S
protein, affects cellular RNA metabolism and the transla- 39 (complementary to nucleotides 27,950–27,979 excepttional machinery in fibroblast cell lines [4,11,44,45]. for ACC replacing CTT starting at nucleotide 27,977); 39
In this study, we examined expression from HSV-1 fragment, 59CCCGGTACCAACGGCAAGCTTATGTTC-vectors that contain neuronal-specific promoters and were GCGAATCAGTTGG39 (nucleotides 27,974–28,010 ex-packaged in the presence of specific mutated HSV-1 cept for ACC replacing CTT) and 59CGCGAATTCAAG-proteins. In cultured cells, vectors packaged in the pres- CAGCGACCAACGCTTCTCTGTGG39 (complementaryence of each mutated protein, or specific combinations of to nucleotides 28,528–28,553)). After digestion (59 frag-two or three mutated proteins, exhibited increases in the ment, BamH I and Kpn I; 39 fragment Kpn I and EcoR I),level of expression per cell. In the rat striatum, vectors the fragments were inserted into pUC19 (BamH I andpackaged in the presence of mutated U 3 displayed EcoR I) to yield pUC19/ul13g. Two fragments thatS
enhanced gene transfer (number of cells at 4 days), and contain the U 3 mutation (us3g, generates a Pvu II site)S
vectors packaged in the presence of mutated U 13 or VP16 and extend 59 or 39 were produced using PCR (template,L
displayed enhanced long-term expression (2 months). cos48Da [13]; primers: 59 fragment, 59CGGGAATT-Vectors packaged in the presence of mutated vhs or U 11 CTTCTGGTCAAGGAGGGTCATTACG39 (nucleotidesS
exhibited minimal changes. 134,885–134,908) and 59TCGTGTACCACCCAGCTGC-

T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16 3
CACGATTACCCGTTGG39 (complementary to nucleo- 103,379); 2.7 kb (VP16 in14, 102,921–105,530); andtides 135,863–135,897 except for GCAGCT replacing 8.0 kb (104,512–112,492).AAGGCG starting at nucleotide 135,879); 39 fragment, • cos48Da /us3g: vector fragment, 16.8 kb (135,985–59AACGGGTAATCGTGGCAGCTGGGTGGTACACGA- 108,670 (includes SuperCos I)); 28.4 kb (3,375 ofGC39 (nucleotides 135,865–135,899 except for GCAGCT SuperCos I-133,466); 20.0 kb (115,702–135,880); andreplacing AAGGCG) and 59GCGAAGCTTCACCGACA- 2.0 kb (pUC19/us3g, 134,885–136,807).AGAACCAAAAGGAATGG39 (complementary to • cos6Da /us11B, cos6Da /us11G: vector fragment, 11.8nucleotides 136,783–136,807)). After digestion (59 frag- kb (28,630–144,897 (includes SuperCos I)); either 3.0ment, EcoR I and Pvu II; 39 fragment, Pvu II and Hind kb (U 11B, 230–3,204) or 3.7 kb (U 11G, 230–S S
III), the fragments were inserted into pUC19 (EcoR I and 3,999); either 10.4 kb (with U 11B, 145,246–3,338)S
Hind III) to yield pUC19/us3g. or 10.2 kb (with U 11G, 145,442–3,381); 11.8 kbS
(1,308–13,089); and 19.1 kb (10,636–29,733).• cos6Da /ul13g,us11B, cos6Da /ul13g,us11G: vector
2.4. Cosmids fragment, 15.7 kb (cos6Da /ul13g, 22,872–143,028(includes SuperCos I)); and 34.1 kb (cos6Da /us11B or
Cosmid set C (cos6, cos14, cos28, cos48, cos56) [7] cos6Da /us11G, 141,221–23,161).represents the HSV-1 genome; the a sequence was deleted • cos56/vhsSc243,vp16in14: vector fragment, 29.2 kbfrom the two cosmids that contain it (cos6Da, cos48Da) (cos56/vp16in14, 97,669–84,252 (includes SuperCos[13]. I)); and 24.5 kb (cos56/vhsSc243, 71,016–103,379).
Mutations were introduced into the cosmids by homolo-gous recombination cloning in E. coli. Recombinationcloning in E. coli BJ5183 has been used to manipulate 2.5. Vectors35-kb adenovirus vectors [5], and we modified this method[29] to allow the use of up to six DNA fragments to pHSVlac [15] contains the HSV-1 IE 4/5 promoter.reconstruct an HSV-1 cosmid. Each transformation con- pNFHlac [53] contains the mouse NF-H promoter (0.6-kbtained 30 ml E. coli BJ5183, 3-ng vector, and the other fragment, from pH-615) [47]. pTH-NFHlac [56] contains afragments (1:1 molar ratio relative to vector). Candidates 6.3-kb BamH I fragment from the rat TH promoter (0.5–were analyzed by digestion with several enzymes, includ- 6.8 kb) [3] inserted at the 59 end of the NF-H promoter.ing BamH I and Dra I, and the presence of a mutation wasverified by assaying for an additional site or fragment due
2.6. Viruses that harbor ul13g or us3gto a mutation (ul13g, Kpn I; vhs Sc243, Xho I; VP16 in14,BamH I; us3g, Pvu II; U 11B, Kpn I (|100-bp smallerS Viruses were derived [7] by cotransfecting 2-2 cellsfragment due to the deletion); and U 11G, Sac I (|500-bpS 6(1310 cells, 60-mm dish; lipofectamine procedure) withlarger fragment due to the insertion)). Correct cosmids
cosmid DNAs (0.5 mg of each) digested with Pac I.were transferred into SURE cells.HSV-1/ul13g: cos6Da /ul13g, cos28/ul13g, cos14, cos56,Specific cosmids were constructed using the followingand cos48. HSV-1/us3g: cos6, cos28, cos14, cos56, andfragments:cos48Da /us3g. One a sequence in a cosmid set is suffi-cient to produce virus [13]. Single plaques were isolatedand passaged on 2-2 cells.
• cos6Da /ul13g: vector fragment, 6.0 kb (28,166–5,548SuperCos I (Stratagene; nucleotides refer to HSV-1
2.7. Protein kinase assayssequences [34] except where SuperCos I is specified));9.3 kb (4,727 SuperCos I-147,352); 25.4 kb (144,513–
U 13 was assayed [6] by incubating disrupted virions in17,669); 13.2 kb (14,074–27,265); and 1.5 kb L32(pUC19/ul13g, 27,014–28,586). the presence of [g- P]ATP (New England Nuclear).
• cos28/ul13g: vector fragment, 25.2 kb (48,599– Reaction products were separated by SDS-polyacrylamide27,014 (includes SuperCos I)); 2.7 kb (24,982– (9%) gel electrophoresis; each lane contained 0.7 mg27,673); 1.5 kb (pUC19/ul13g, 27,014–28,586); 7.6 protein (Bio-Rad Protein Assay). The gel was dried andkb (28,166–35,757); and 30.9 kb (30,535–61,393). opposed to film for 30 min.
• cos56/vhsSc243: vector fragment, 24.5 kb (110,660– U 3 was assayed [39] by fractionating extracts ofS
91,257 (includes SuperCos I)); 1.8 kb (pSC243, infected BHK cells (multiplicity of infection (MOI), 10;90,940–92,760); 8.6 kb (91,910–100,561); and 18.0 harvested at 19 h after infection) on DEAE-cellulose (DE-
32kb (97,669–115,152). 52, Whatman) and incubating fractions with [g- P]ATP• cos56/vp16in14: vector fragment, 24.5 kb (110,660– and protamine sulfate. Reaction products were spotted on
91,257 (includes SuperCos I)); 14.0 kb (89,364– P81 paper (Whatman), washed with trichloroacetic acid,

4 T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16
rinsed in ethanol, dried, and subjected to liquid scintillation (Ig) G, and a mouse monoclonal anti-NeuN antibody [35]spectrometry. (Chemicon) was visualized using a rhodamine isothio-
cyanate-conjugated goat anti-mouse IgG.2.8. Vector packaging
2.12. Cell countsHelper virus-free packaging [13] was performed using a
modified protocol [52] that improves the efficiency. In A number of 25-mm coronal sections that contained thesome packagings, the cells were infected with pre-viral striatum were prepared. Every fourth section was analyzedDNA replication enveloped particles (PREPs) [10] prior to for expression of b-galactosidase and |12 of these sectionsthe transfection [52]. Vector stocks were purified [31] and contained either the X-gal positive cells or the b-galacto-titered by counting the numbers of 5-bromo-4-chloro-3- sidase-IR positive cells. Positive cells were identifiedindoyl-b-D-galactopyranoside (X-gal) positive cells ob- under low power (103), and cell counts were performedtained at 1 day after infection of BHK or PC12 cells. under 403 magnification. To verify the accuracy of the
cell counts, each section was counted at least twice, on2.9. Titering vector genomes different days; the two values differed by ,10% for each
section. The statistical significance of the differences in theDNA was isolated from purified vector stocks. Lac Z numbers of X-gal positive cells were analyzed using
sequences were detected using PCR (30 cycles; 948C, 1 Student’s unpaired t-test.min; 558C, 1 min; 728C, 4 min. Primers:59GTTGATTGAACTGCCTGAACTACC39 (nucleotides2,034–2,057) [26], 59CACTTCAACATCAACGGTAAT- 3. ResultsCG39 (complementary to nucleotides 2,594–2,616)). PCRwas performed on 3-fold serial dilutions of each sample to 3.1. Packaging systems that contain mutated HSV-1identify a concentration range that produced a linear proteinsincrease in signal with increasing sample. The standardDNA was pNFHlac plasmid digested with BamH I (single Because multiple HSV-1 proteins that affect the virionsite) and added to a sample obtained from a stock of may reduce expression, enhanced expression may requirepTH-NFHpkcD (expresses a protein kinase C). Reaction packaging vectors in the presence of multiple mutatedproducts were displayed on 0.6% agarose gels. The amount HSV-1 proteins. To increase the probability that specificof DNA in a sample was determined by comparing the combinations of mutated tegument proteins might besignals produced using dilutions of the sample to those assembled into biologically active vector particles, weproduced using dilutions of the standard. The titers of preferred mutations that are likely to cause only minimalvector genomes were calculated from the DNA concen- changes in the structure of a protein. Linker insertiontrations and corrected for packaging a vector as a con- mutations in vhs [25] and VP16 [1] display either reducedcatamer similar in size to the HSV-1 genome (pNFHlac mRNA degradation activity or reduced IE promoter trans-(10.2 kb, 15 copies), 153 kb; pTH-NFHlac (16.5 kb, nine activation activity, respectively. We used both a partialcopies), 148.5 kb). deletion and a null mutation in U 11 [44] because thereS
appears to be insufficient information to support the2.10. Expression experiments in cell lines rational isolation of point mutations; U 11B contains anS
internal deletion that removes the ribosome association and5BHK or PC12 cells (5310 cells /ml, 60-mm dish) were RNA binding activities, and U 11G does not expressS
infected with vector stocks (MOI, 0.1), and 1 day later U 11. Only deletion mutations in U 13 or U 3 have beenS L S
b-galactosidase was assayed using either o-nitrophenyl-D- reported [6,39]; we isolated point mutations in U 13 andL
galactopyranoside (ONPG) or X-gal [12,32]. U 3 that were designed to lack protein kinase activityS
(ul13g and us3g). These mutations were introduced into2.11. Expression experiments in the brain the cosmids used for helper virus-free packaging.
The established assays for U 13 [6] or U 3 [39] proteinL S
Vector stocks were delivered by stereotactic injection kinase activity use either disrupted virions or infected cell(two sites, 3 ml / site) into the striatum of male Sprague– lysates, respectively. We derived viruses that contain eitherDawley rats (150–175 gm; injection coordinates are ul13g or us3g. U 13 activity was assayed by incubatingL
described in Ref. [56]). These studies were approved by disrupted virions with radiolabeled ATP; reaction productsthe Children’s Hospital IACUC. Rats were perfused, brains were displayed on a polyacrylamide gel. The results (Fig.were sectioned, and X-gal staining was performed [56]. 1A) show that disrupted HSV-1/ul13g virions incorporatedAlternatively [56], a rabbit anti-E. coli b-galactosidase less radioactivity into an |57-kDa band, and other bands,antibody (ICN) was visualized using a fluorescein iso- compared to virions that contain wild type (wt) U 13.L
thiocyanate-conjugated goat anti-rabbit immunoglobulin U 13 is a |57-kDa protein with an autophosphorylationL

T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16 5
Fig. 1. Protein kinase assays on disrupted virions (A) or infected cell extracts (B). (A) HSV-1/ul13g or 5dl1.2 (U 13 wt) virions were disrupted with NP40L
and then incubated with radiolabeled ATP and varying concentrations of NaCl. Aliquots of the reaction products were subjected to SDS-polyacrylamide gelelectrophoresis. The resulting autoradiogram shows that disrupted HSV-1/ul13g, but not 5dl1.2, virions lack a radiolabeled|57-kDa band (curved arrow);57 kDa is the predicted size of U 13 [6]. Disrupted HSV-1/ul13g virions also show reduced levels of a number of other bands (straight arrows) comparedL
to disrupted 5dl1.2 virions. Molecular weight sizes are from the Rainbow Molecular Weight Marker kit (Amersham). (B) Protein kinase activity followingcolumn chromatography of lysates prepared from cells infected with either HSV-1/us3g or 5dl1.2 (U 3 wt), or mock infected. Cell lysates were prepared atS
19 h after infection of BHK cells and fractionated by column chromatography on DEAE-cellulose. Protein kinase assays were performed using radiolabeledATP and exogenous protamine sulfate as the substrate. 5dl1.2 infected cell lysates contain a late eluting protein kinase activity that is absent from lysatesprepared from either mock infected cells or HSV-1/us3g infected cells. The late eluting protein kinase activity in lysates from 5dl1.2 infected cells elutes ata KCl concentration consistent with U 3 protein kinase [39].S
activity [6]. U 3 activity was detected by subjecting to pHSVlac /wt. Packaging could occur in the presence ofS
infected cell lysates to column chromatography followed specific combinations of two or three mutated proteins, butby a protein kinase assay. The results (Fig. 1B) demon- the titers were reduced (Table 1). pHSVlac packaged instrate that both an HSV-1/us3g-infected cell lysate and a the presence of specific combinations of four mutatedmock-infected cell lysate lack a late-eluting activity pres- proteins, or all five of these mutated proteins, yielded veryent in a 5dl1.2 (U 3 wt) infected cell lysate. These results low, or no detectable, titers (Table 1).S
are similar to those reported using the deletion mutants We tried one approach to increase the titers. Treating the[6,39]. packaging cells with PREPs [10] results in a modest
(3–5-fold) increase in the titers [52]. Because PREPs3.2. Packaging in the presence of mutated HSV-1 supply tegument proteins, we reasoned that the PREPsproteins might support larger increases in titers from packaging in
the presence of specific mutated proteins. Packaging usingpHSVlac was packaged in the presence of each mutated PREPs, in the presence of either two or all five mutated
HSV-1 protein under study (nomenclature, vector followed proteins, resulted in modest or minimal, respectively,by the mutated protein(s) present during packaging), and increases in the titers (Table 1).the vector stocks were titered. The results (Table 1) show Because we are interested in obtaining long-term, neuro-that pHSVlac /vhsSc243, pHSVlac /us3g, and pHSVlac / nal-specific expression in the brain, we packaged pNFHlacus11(B or G) supported titers similar to that of pHSVlac / in the presence of specific mutated proteins. The titerswt. The titers of pHSVlac /ul13g and pHSVlac /vp16in14 obtained using BHK cells were similar to, or slightlywere reduced |20- and |100-fold, respectively, compared higher than, those obtained using PC12 cells (Table 1),

6 T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16
Table 1The titers obtained from packaging pHSVlac or pNFHlac in the presence of specific mutated HSV-1 proteins
aPackaging Titer, IVP/mlcondition
pHSVlac pNFHlacbBHK PREPs , BHK BHK PC12
6 6 5 5wt 2.561.1310 6.062.4310 3.662.0310 1.160.53105 4 3ul13g 1.260.6310 2.561.0310 7.562.53105 4 4vhs Sc243 8.062.0310 4.861.6310 1.260.53104 3 2VP16 in14 3.062.0310 4.062.0310 5.562.03105 5 4us3g 8.563.5310 1.560.5310 4.262.03106 5 4U 11B 1.560.5310 1.560.5310 2.561.0310S6U 11G 1.560.6310S
3ul13g, vhs Sc243 3.661.83104 5 3 2ul13g, us3g 4.562.0310 1.160.6310 4.261.0310 4.562.03104ul13g, U 11B 2.562.0310S3vhs Sc243, VP16 in14 4.061.63102ul13g, vhs Sc243, VP16 in14 2.061.03102ul13g, vhs Sc243, U 11B 2.061.2310S4 3ul13g, vhs Sc243, us3g 1.561.0310 2.061.03103ul13g, VP16 in14, us3g 1.560.53104ul13g, us3g, U 11B 1.860.6310S4 3ul13g, us3g, U 11G 2.060.5310 2.561.5310S2vhs Sc243, VP16 in14, us3g 5.062.03102vhs Sc243, VP16 in14, U 11B 3.062.0310S4 3 2vhs Sc243, us3g, U 11B 3.061.0310 4.561.0310 4.362.2310S3VP16 in14, us3g, U 11B 4.062.0310S
ul13g, vhs Sc243, VP16 in14, us3g ,1ul13g, vhs Sc243, us3g, U 11B 20620S
ul13g, us3g, VP16 in14, U 11B ,1S2vhs Sc243, VP16 in14, us3g, U 11B 2.061.1310S
ul13g, vhs Sc243,VP16 in14, us3g, U 11B ,1 ,1S
pHSVlac or pNFHlac were packaged in the presence of specific mutated HSV-1 proteins, or none of the mutations (wt). BHK or PC12 cells were infectedwith the resulting vector stocks. Then 1 day later, the number of cells expressing b-galactosidase was determined using X-gal. Each titer is the averagefrom two to four packagings and the standard deviations are shown.a The units of the titers are infectious vector particles (IVP) /ml.b 2-2 Cells were infected with PREPs prior to the transfection of the vector and cosmid DNAs [52].
even though neurofilament genes are expressed in PC12 fully active in BHK cells, the biological titers of thecells but not in BHK cells. In attempts to increase the titers pNFHlac stocks may underestimate the titer by |10-fold.on PC12 cells, we either plated PC12 cells at high densities Vector particles produced in the presence of a specific(to obtain a monolayer) or infected PC12 cells in suspen- mutated HSV-1 protein might have a reduced capability tosion (at high density), but we observed only decreases in bind to, or enter, BHK cells and subsequently expressthe titers (not shown). Expression from pNFHlac in b-galactosidase (transducing efficiency). A reduced trans-fibroblast cells represents ectopic expression, and this ducing efficiency might be detected as an increase in theectopic expression declines at longer times after gene ratio of vector genomes to biological titer for a specifictransfer (not shown). Use of this ectopic expression may packaging condition. In contrast, the ratio of vectorresult in underestimating the titers, and the titers of genomes to biological titer was similar for pNFHlac /wtpNFHlac were 5–10-fold lower than the titers of pHSVlac and each packaging condition that used specific mutatedfor specific packaging conditions (Table 1). We determined proteins (Table 2). Thus, the vector particles produced bythe titers of vector genomes. PCR was performed on serial these different packaging conditions appear to have similardilutions of DNAs isolated from these stocks or a standard transducing efficiencies.(pNFHlac plasmid). A linear response range (Fig. 2) wasused to determine the titer of vector genomes (Table 2). 3.3. Enhanced expression in cultured cellsThe ratio of vector genomes to biological titer was 1.5 fora pHSVlac stock. Thus, the physical and biological titers The amount of b-galactosidase activity was quantitated,were similar, and the IE 4/5 promoter in pHSVlac is active 1 day after infection of BHK or PC12 cells, using ONPG,in BHK cells. In contrast, the ratio of vector genomes to and the number of cells expressing b-galactosidase wasbiological titer was 8.5–11.5 for the pNFHlac stocks. This determined using X-gal. The b-galactosidase activity / in-result suggests that because the NFH promoter may not be fected cell was calculated from these data. The results

T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16 7
We investigated the expression supported by pNFHlacpackaged in the presence of specific combinations of themutated proteins. Compensatory crosstalk between proteinkinase signaling pathways is well known; to increase theprobability of enhancing expression, we examinedpNFHlac /ul13g,us3g. Both vhs and U 11 affect RNAS
metabolism; to address possible redundancy in the func-tions of these proteins, we examined pNFHlac /vhs-Sc243,us3g,us11B (us3g was included because it affectsthe efficiency of gene transfer, see below). These twocombinations were designed to detect synergistic effectsbetween four of the five proteins under study. VP16 in14
Fig. 2. PCR analysis of Lac Z sequences in samples prepared from was omitted from these combinations because of the largepNFHlac stocks. To identify the linear range for PCR, 3-fold serial
reduction in the titer caused by this mutation (Table 1).dilutions of each DNA sample and the standard DNA were analyzed. TheThe results (Fig. 3) showed that pNFHlac /ul13g,us3g andstandard was pNFHlac plasmid DNA digested with BamH I (single site)pNFHlac /vhsSc243,us3g,us11B support 4–7-fold in-and added to a sample obtained from a stock of pTH-NFHpkcD. The
amount of DNA in a sample was determined by comparing the signals creases in the levels of expression.produced using the dilutions to those produced using the dilutions of thestandard. The predicted size of the PCR products is 583 bp. Lanes: 3.4. Enhanced expression in the brainplasmid, pNFHlac plasmid DNA standard; N, no DNA; S, bacteriophagel BstE II size standards; VP16in14, pNFHlac /vp16in14; ul13g,us3g,
pNFHlac stocks were microinjected into the striatum,pNFHlac /ul13g,us3g; U, undiluted; 1,2,3,4,5 indicate 3, 9, 27, 81, and243-fold dilutions, respectively. The lane of standard undiluted (plasmid, the rats were sacrificed 4 days, 2 weeks, 1 month, or 2
28U) represents the signal from 7.3310 mg of pNFHlac, which corre- months later, and X-gal staining was performed. Rats that3sponds to 6.7310 molecules of pNFHlac (10.2 kb).received either PBS or a vector that expresses a proteinkinase C lacked X-gal positive striatal cells, but faintly
(Fig. 3) showed that pNFHlac packaged in the presence of positive cells were occasionally observed in brain vascula-each mutated protein supported |2-fold, statistically sig- ture endothelium (not shown). At 4 days after gene transfernificant, increases in the levels of expression /cell com- with these pNFHlac stocks, we observed numerous X-galpared to those obtained using pNFHlac /wt. positive cells proximal to the injection sites (Fig. 4A, B
Table 2The numbers of X-gal positive striatal cells from rats sacrificed at various times after microinjection of vectors that were packaged in the presence ofspecific mutated HSV-1 proteins
aVector Packaging Purified titers Average X-gal positive cells per rat Relative efficiencybcondition of gene transfer
VG/ml IVP/ml VG/IVP Time after gene transfer
4 Days 2 Weeks 1 Month 2 months7 6 1pNFHlac ul13g 2.2310 2.4310 9.3 278633 170611 1.27 6pNFHlac vhs Sc243 1.1310 1.1310 10.0 141636 1.57 6 3 3 1,4pNFHlac VP16 in14 4.5310 6.1310 7.3 10136560 2826110 127645 4665 1.86 6pNFHlac us3g 4.5310 1.9310 8.6 24776532 12.9
6pNFHlac U 11B 1.1310 193656 2.0S
pNFHlac vhs Sc243, us3g,7 6 2U 11B 1.3310 1.1310 11.5 16766290 283646 17.4S6 5 1 1,4 2,4pNFHlac ul13g, us3g 4.5310 5.2310 8.6 8236150 477657 251693 140653 17.17 6,c c c,2 cpNFHlac wt 1.8310 2.1310 8.5 192630 25613 060 1.0
8 8pHSVlac wt 1.5310 1.0310 1.5
6 5 3 1,4 2,5,6pTH-NFHlac ul13g, us3g 5.5310 4.5310 12.2 8646182 718697 492618 219639 17.87 7,c c c,2 c,2 c,2pTH-NFHlac wt 5.0310 1.2310 4.1 17196384 315644 260695 230635 1.0
The rats were perfused 4 days to 2 months after gene transfer into the striatum, and expression of b-galactosidase was detected using X-gal [12,49]. Three1 2 3rats were analyzed with each packaging condition at each time point, and the means6S.D. are shown. P,0.05, P,0.01, or P.0.05, compared to 4 days;
4 5 6P,0.05 or P,0.01 compared to 2 weeks; P,0.001 compared to 1 month; Student’s unpaired t-test.a The titers of each vector stock following purification and concentration. VG/ml is the vector genomes /ml.b The efficiency of gene transfer is the number of X-gal positive cells at 4 days after gene transfer divided by the amount of vector injected. The relativeefficiency of gene transfer is the efficiency of gene transfer with a specific vector and packaging condition divided by the efficiency of gene transfer withthat vector and wt packaging conditions.c These data are reported in Ref. [56], but were collected concurrently with the other data in this table, and are included only to enable comparisons.

8 T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16
Fig. 3. The levels of b-galactosidase activity in BHK or PC12 cells at 1 day after infection with pNFHlac packaged in the presence, or absence, of specificmutated HSV-1 proteins. The amount of b-galactosidase activity was quantitated using ONPG (a standard curve was generated using purified E. colib-galactosidase (Sigma)), and the number of cells expressing b-galactosidase was determined using X-gal. These data were used to calculate theb-galactosidase activity per 1000 infected cells. The experiment was repeated twice, and a total of four plates of each cell line were used for eachmeasurement (ONPG or X-gal) with each vector stock. The values shown are the means6S.D. [P,0.05; 1P,0.01; *P,0.001 compared to the same celltype infected with pNFHlac /wt using Student’s unpaired t-test.
and E). Low power views show the locations and sizes of trix, an inflammatory response, and physiological differ-the areas of positive striatal cells. Higher power views ences between striatal cells and BHK cells.reveal some individual cell bodies in the densely stained Long-term expression was examined by comparing theareas proximal to the injection sites as well as outlying numbers of positive cells at 2 weeks, 1 month, or 2 monthspositive cells. Due to retrograde transport of HSV-1 [51], after gene transfer to the number of positive cells at 4 days.small numbers of positive cells were also observed at Because this comparison uses the number of positive cellsdistant sites, such as specific areas of neocortex, that at 4 days as the initial value, this comparison is designed tocontain neurons that project to the striatum (not shown). be independent of any variability in either the gene transferSubsequent analyses focused on the positive striatal cells. process or the titering. Using pNFHlac /wt, at 2 weeks, the
The efficiency of gene transfer was quantitated as the number of positive cells had declined to |15% of that at 4number of positive cells at 4 days after gene transfer days; and by 1 month, no positive cells were observed.divided by the amount of vector that was injected. To This decline in expression is similar to that previouslymeasure any changes in the efficiency of gene transfer reported [53]. Although pNFHlac /vhsSc243,us3g,us11Bcaused by packaging in the presence of specific mutated increased the efficiency of gene transfer, by 2 weeks,proteins, we compared the efficiencies of gene transfer pNFHlac /vhsSc243,us3g,us11B displayed a large declinesupported by pNFHlac packaged in the presence of specific in expression similar in magnitude to that observed usingmutated proteins to that supported by pNFHlac /wt. This pNFHlac /wt (Table 2 and Fig. 5A; photomicrographs, Fig.ratio is also designed to correct for any underestimation of 4C and D). Thus, longer time points were not examinedthe titers of these pNFHlac stocks due to the use of BHK using pNFHlac /vhsSc243,us3g,us11B or vectors packagedcells in the titering. The results (Table 2) showed that in the presence of any of these three mutated proteinspNFHlac /us3g supported a 12.9-fold increase in the individually. pNFHlac /vp16in14 supported increases inrelative efficiency of gene transfer. In contrast, pNFHlac / long-term expression, and some positive cells were stillul13g, pNFHlac /vhsSc243, pNFHlac /vp16in14, and observed at 2 months (Table 2 and Fig. 5A). BothpNFHlac /us11B had minimal effects on the efficiency of pNFHlac /ul13g and pNFHlac /ul13g,us3g supported largegene transfer. pNFHlac /ul13g,us3g and pNFHlac /vhs- increases in long-term expression; at 2 weeks, we observedSc243,us3g,us11B supported 17-fold increases in the rela- |60% of the numbers of positive cells obtained at 4 daystive efficiency of gene transfer. For a specific vector and (Table 2 and Fig. 5A; pNFHlac /ul13g,us3g photomicrog-packaging condition, the efficiency of gene transfer into raph, Fig. 4F). More positive cells were observed usingthe brain may differ from the transducing efficiency of pNFHlac /ul13g,us3g than pNFHlac /ul13g, presumablycultured BHK cells because of specific aspects of striatal because packaging in the presence of us3g enhanced genestructure and physiology, including the extracellular ma- transfer. Using pNFHlac /ul13g,us3g, significant levels of
T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16 9
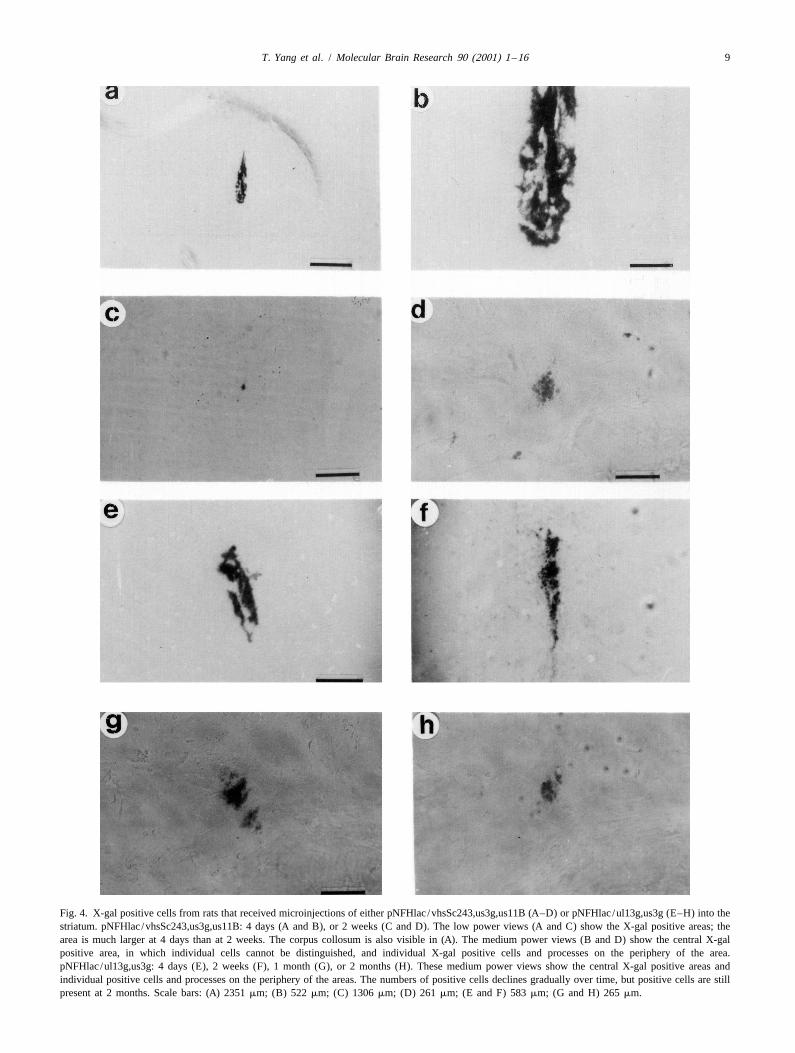
Fig. 4. X-gal positive cells from rats that received microinjections of either pNFHlac /vhsSc243,us3g,us11B (A–D) or pNFHlac /ul13g,us3g (E–H) into thestriatum. pNFHlac /vhsSc243,us3g,us11B: 4 days (A and B), or 2 weeks (C and D). The low power views (A and C) show the X-gal positive areas; thearea is much larger at 4 days than at 2 weeks. The corpus collosum is also visible in (A). The medium power views (B and D) show the central X-galpositive area, in which individual cells cannot be distinguished, and individual X-gal positive cells and processes on the periphery of the area.pNFHlac /ul13g,us3g: 4 days (E), 2 weeks (F), 1 month (G), or 2 months (H). These medium power views show the central X-gal positive areas andindividual positive cells and processes on the periphery of the areas. The numbers of positive cells declines gradually over time, but positive cells are stillpresent at 2 months. Scale bars: (A) 2351 mm; (B) 522 mm; (C) 1306 mm; (D) 261 mm; (E and F) 583 mm; (G and H) 265 mm.

10 T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16
Fig. 5. The stabilities of expression in the striatum supported by pNFHlac (A) or pTH-NFHlac (B) packaged in the presence, or absence, of specificmutated HSV-1 proteins. For each vector, packaging condition, and time point, the % of X-gal positive cells at 4 days is shown (the average number ofX-gal positive cells at 2 weeks, 1 month, or 2 months after gene transfer divided by the average number of X-gal positive cells at 4 days (3100); calculatedusing the data in Table 2). The data with pNFHlac /wt and pTH-NFHlac /wt are reported in Ref. [56], but were collected concurrently with the other data inthis study, and are included in this figure only to enable comparisons.
expression were observed at both 1 and 2 months (Table 2 3.5. Packaging in the presence of the mutated proteinand Fig. 5A; photomicrographs, Fig. 4G and H), although kinases acts cooperatively with a TH-NFH promoter tothe amount of expression exhibited a statistically signifi- enhance expressioncant decline during this period. To confirm that pNFHlac /ul13g,us3g targets expression to neurons (ul13g or us3g pTH-NFHlac /wt supports more stable expression thanmight potentially affect the cell type specificity of expres- pNFHlac /wt; although a large decrease in expression ission), we performed double staining using antibodies observed between 4 days and 2 weeks, the remainingdirected against either E. coli b-galactosidase or a neuronal expression is relatively stable for 6 months [56]. Becausemarker, NeuN. The results (Fig. 6A and B) revealed this initial decrease might be caused by specific proteinsb-galactosidase-IR in striatal cells that contain NeuN-IR. that affect the vector particle, we examined expression

T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16 11
Fig. 6. b-Galactosidase-IR positive cells that also contain NeuN-IR from rats sacrificed at 4 days after microinjection of either pNFHlac /ul13g,us3g intothe striatum (A and B) or pTH-NFHlac /ul13g,us3g into either the striatum (C and D) or the hippocampus (E and F). b-Galactosidase-IR (A, C, and E) wasdetected with a rabbit anti-E. coli b-galactosidase antibody that was visualized using a fluorescein isothiocyanate-conjugated goat anti-rabbit IgG. NeuN-IR(B, D, and F) was detected in the same sections with mouse monoclonal anti-NeuN antibody that was visualized using a rhodamine isothiocyanate-conjugated goat anti-mouse IgG. NeuN is a neuronal marker found in the nucleus. (A and B) Using pNFHlac /ul13g,us3g, a striatal section contains a largeb-galactosidase-IR positive cell body with a process, and this cell is also positive for NeuN (arrow). (C and D) Using pTH-NFHlac /ul13g,us3g in thestriatum, a section contains several b-galactosidase-IR positive cell bodies with processes, and these cells also contain NeuN-IR (arrows). (E and F) UsingpTH-NFHlac /ul13g,us3g in the hippocampus, a section contains numerous b-galactosidase-IR positive cells in both the dentate granule cell layer (many ofthese positive cells also display processes) and the hilar region. Many of the b-galactosidase-IR cells also contain NeuN-IR. Scale bars: (A and B) 250 mm;(C–F) 550 mm.
from pTH-NFHlac that was packaged in the presence of 5B): at 2 weeks, the stability of expression was 83%; at 1the preferred combination of mutated proteins for enhanc- month, 57%; and at 2 months, 25%. Thus, the stability ofing expression (ul13g and us3g). The results (Table 2; Fig. expression supported by pTH-NFHlac /ul13g,us3g ex-7) showed that pTH-NFHlac /ul13g,us3g increased the hibited statistically significant, gradual decreases from 2efficiency of gene transfer 18-fold compared to that weeks to 2 months; in contrast, the expression supportedobserved using pTH-NFHlac /wt. This increase was similar by pTH-NFHlac /wt remained essentially unchanged dur-in magnitude to that observed between pNFHlac /ul- ing this period [56]. Nonetheless, the stability of expres-13g,us3g and pNFHlac /wt. Of note, pTH-NFHlac /ul- sion obtained using pTH-NFHlac /ul13g,us3g is higher13g,us3g enhanced long-term expression (Table 2; Fig. than those observed using either pTH-NFHlac /wt or

12 T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16
Fig. 7. X-gal positive cells from rats sacrificed at 4 days (A and B), 2 weeks (C and D), 1 month (E and F), or 2 months (G and H) after microinjection ofpTH-NFHlac /ul13g,us3g into the striatum. Low (A, C, E, and G) and medium (B, D, F, and H) power views of X-gal positive cells are shown at each timepoint. In (A) and (C), most of the striatum and a portion of the corpus collosum are visible. The medium power views reveal a central X-gal positive areasurrounded by individual X-gal positive cells. Although the numbers of X-gal positive cells declines gradually with time, many X-gal positive cells arepresent at 2 months after gene transfer. Scale bars: (A and C) 2570 mm; (B and D) 510 mm; (E and G) 1290 mm; (F and H) 260 mm.

T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16 13
pNFHlac /ul13g,us3g. To confirm that pTH-NFHlac /ul- activities might be capable of shutting off recombinant13g,us3g targeted expression to neurons, we performed gene expression.double staining using antibodies directed against either Vector packaged in the presence of us3g exhibitedb-galactosidase or NeuN. At 4 days after microinjection enhanced levels of expression in cultured cells and en-into either the striatum or hippocampus, we observed hanced gene transfer in the striatum. Thus, U 3 appears toS
doubly labeled cells in either brain area (Fig. 6C–F). either inhibit the gene transfer process or mediate a rapiddecline in expression that occurs within 4 days after genetransfer. U 3 can protect cells from the apoptosis inducedS
4. Discussion by HSV-1 infection [30], and this anti-apoptotic functionmight be responsible for the observed effects on expres-
Helper virus-free HSV-1 vectors reduce the cytopathic sion. Of note, U 3 is not present in HSV-1 virions; unlessS
effects and inflammatory response associated with gene U 3 is present at a low level in vector particles, the effectsS
transfer, but do not enhance long-term expression. Thus, on expression are likely due to a substrate(s) of U 3 that isS
expression from many promoters remains short-term, even incorporated into virions. U 34, the best characterizedL
in the absence of |99% of the HSV-1 genome. To resolve substrate of U 3, is a non-glycosylated membrane proteinS
this paradox, we hypothesized that specific HSV-1 proteins present at low levels in virions, and, in the absence ofthat affect the virion can shut off recombinant gene phosphorylation by U 3, U 34 associates with a number ofS L
expression. The data in this study support this hypothesis. other, genetically unrelated, phosphoproteins [41]. Addi-In particular, we show that U 3 affects the efficiency of tionally, HSV-2 U 3 phosphorylates another virion protein,S S
gene transfer, and both U 13 and VP16 affect long-term U 9 [9].L S
expression. Vectors packaged in the presence of ul13g exhibitedenhanced levels of expression in cultured cells and en-
4.1. Titers hanced long-term expression in the striatum. Thus, U 13L
appears to destabilize expression. Substrates for U 13L
Although packaging could occur in the presence of each include a number of HSV-1 proteins (ICP0, ICP22, gE, andmutated protein and specific combinations of two or three VP22 (U 49)) [6,36,38,40] and elongation factor 1d [28].L
mutated proteins, the titers were reduced by specific ICP0 and ICP22 are not present in virions and thus aremutated proteins and by most combinations of mutated unlikely to reduce expression. Following infection, theproteins. The |20- or 100-fold reductions in titer caused by viral Fc receptor (gE and gI) is displayed on the cellul13g or VP16 in14, respectively, are not surprising surface where it might influence the inflammatory responsebecause viruses that harbor a deletion in U 13 [6] or VP16 and any consequent effects on expression. U 49 is presentL L
in14 [2] display reduced growth in fibroblast cells. In- at low levels in virions, but little is known about itsdividually, vhs Sc243, us3g, U 11B, or U 11G had function. Changes in the activity of elongation factor 1d, inS S
minimal effects on the titer, consistent with the relatively response to phosphorylation by U 13, might affect theL
normal growth in fibroblast cells of viruses that harbor level of expression, congruent with the results in culturedthese mutations [25,39,44]. However, packaging with cells, but even low levels of b-galactosidase are sufficientspecific combinations of these mutated proteins reduced to support X-gal staining. More generally, U 13 mayL
the titers. phosphorylate additional, unidentified, cellular proteins,Enhanced titers might be obtained by developing alter- and protein kinase signaling pathways are known to
native mutations; for example, a mutation in U 13 that regulate gene expression.L
preserves protein kinase activity but prevents its incorpora- pNFHlac packaged in the presence of VP16 in14tion into vector particles might support higher titers than exhibited enhanced levels of expression in cultured cellsul13g. Recently, bacterial artificial chromosomes (BACs) and enhanced expression in the striatum, although thisthat harbor the HSV-1 genome have been developed for enhancement was less than that observed using pNFHlac /helper virus-free packaging [21,46,50]. Although the pres- ul13g. Thus, these results suggest that VP16 may affect,ent HSV/BACs produce some wt HSV-1 [21], HSV/BACs perhaps indirectly, the activity of a cellular promoter in amay eventually improve vector packaging, and mutations vector, and this effect is dependent upon the transactiva-in specific HSV-1 proteins could be introduced into HSV/ tion activity of VP16 (VP16 in14 has minimal transactiva-BACs. tion activity on IE promoters) [1]. The HSV-1 ori frag-s
ment in pNFHlac contains the IE 4/5 promoter and4.2. Pathways that might affect expression adjacent TAATGARAT elements [53]. Enhanced tran-
scription from the IE 4/5 promoter might interfere withDuring an HSV-1 infection, each of the five HSV-1 expression from the NF-H promoter, even though pNFHlac
proteins under study exhibits an activity that assists the contains three polyadenylation sites between these promot-virus in taking over the cell biosynthetic machinery [43]. ers. Alternatively, wt HSV-1, but not in1814, activates theThus, in the context of HSV-1 vectors, each of these c-jun N-terminal kinase /stress-activated protein kinase

14 T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16
[55]. Perhaps this signaling pathway can modulate the 4.4. Implications for physiological studiesactivity of the NF-H promoter in an HSV-1 vector.
pNFHlac packaged in the presence of mutations in vhs Although most of the published physiological studiesor U 11 displayed enhanced expression in cultured cells with HSV-1 vectors use only short-term expression, a fewS
but minimal changes in the striatum. vhs causes the studies rely upon long-term expression [14]. The levels ofdegradation of cellular mRNAs in many cell lines long-term expression reported here equal or exceed those[25,37,42]; vhs might degrade Lac Z RNA in BHK or previously achieved. Thus, this system (pTH-NFHlac /PC12 cells. However, vhs does not appear to inhibit ul13,us3) may support some physiological studies thatprotein synthesis in cultured neurons [37], consistent with require long-term expression.the minimal changes observed in the striatum. U 11 has aS
wide range of functions that might decrease the levels ofb-galactosidase: U 11 binds to specific mRNAs and to 60S AcknowledgementsS ribosome subunits [44,45], U 11 has functions similar toS
HIV Rev and Tat proteins [11], and U 11 can bind toS We gratefully thank Dr J. Smiley for vhs pSc243, Dr C.protein kinase R and inhibit phosphorylation of eIF2a and Preston for VP16 in14, Drs R. Roller and B. Roizman forthe resulting shut-off of protein synthesis [4]. Effects of U 11 MutB/421 and U 11 MutG/421, Dr A. Davison forS Svhs or U 11 on the level of expression may not beS HSV-1 cosmid set C, Dr R. Sandri-Goldin for 2-2 cells, Drapparent in the in vivo experiments which measured only P. Schaffer for 5dl1.2 virus, Dr K. O’Malley for the THnumbers of positive cells. promoter, and Dr W. Schlaepfer for the NF-H promoter.
This work was supported by F32NS10805 (M.S.),R01AG16777, R01NS42016, and R01NS34025 (A.G.).
4.3. Multiple HSV-1 proteins, and additional factors,may reduce expression
ReferencesThe enhancements in expression are larger at 2 weekscompared to longer times, and at least 3 HSV-1 proteins
[1] C.I. Ace, M.A. Dalrymple, F.H. Ramsay, V.G. Preston, C.M.(U 3, U 13, VP16) affect expression in vivo. AdditionalS L Preston, Mutational analysis of the herpes simplex virus type 1HSV-1 proteins, not examined in this study, may also trans-inducing factor Vmw65, J. Gen. Virol. 69 (1988) 2595–2605.reduce expression. Thus, one explanation for the remaining [2] C.I. Ace, T.A. McKee, J.M. Ryan, J.M. Cameron, C.M. Preston,
Construction and characterization of a herpes simplex virus type 1gradual declines in expression is that we mutated onlymutant unable to transinduce immediate-early gene expression, J.some of the critical proteins, and other proteins can stillVirol. 63 (1989) 2260–2269.reduce expression, but at a slower rate. Additional en-
[3] E.R. Brown, G.T.D. Coker, K.L. O’Malley, Organization andhancements in expression might be obtained by packaging evolution of the rat tyrosine hydroxylase gene, Biochemistry 26vectors in the presence of more complex combinations of (1987) 5208–5212.
[4] K.A. Cassady, M. Gross, B. Roizman, The herpes simplex virusmutations.US11 protein effectively compensates for the gamma1(34.5) gene ifOther factors may contribute to the gradual decline inpresent before activation of protein kinase R by precluding itsexpression. Although the inflammatory response to helperphosphorylation and that of the alpha subunit of eukaryotic transla-
virus-free HSV-1 vectors declines rapidly and to a low tion initiation factor 2, J. Virol. 72 (1998) 8620–8626.level by 1 month, the numbers of infiltrating cells remains [5] C. Chartier, E. Degryse, M. Gantzer, A. Dieterle, A. Pavirani, M.
Mehtali, Efficient generation of recombinant adenovirus vectors bygreater than that evoked by vehicle alone [57]. Similarly,homologous recombination in Escherichia coli, J. Virol. 70 (1996)lymphokines are produced in the trigeminal ganglia for at4805–4810.least 2 months after ocular inoculation of wt HSV-1, even
[6] L.J. Coulter, H.W. Moss, J. Lang, D.J. McGeoch, A mutant of herpesthough the latent state is achieved [18]. The inflammatory simplex virus type 1 in which the UL13 protein kinase gene isresponse might reduce expression. Alternatively, although disrupted, J. Gen. Virol. 74 (1993) 387–395.
[7] C. Cunningham, A.J. Davison, A cosmid-based system for con-the HSV-1 genome can persist indefinitely in the latentstructing mutants of herpes simplex virus type 1, Virology 197state, these vector DNAs might be degraded at a specific(1993) 116–124.rate. Another possibility is that the structure of the vector,
[8] C. Cunningham, A.J. Davison, A. Dolan, M.C. Frame, D.J.including the promoter, may contribute to reduced expres- McGeoch, D.M. Meredith, H.W. Moss, A.C. Orr, The UL13 virionsion. protein of herpes simplex virus type 1 is phosphorylated by a novel
It seems unlikely that cell death is a major contributor to virus-induced protein kinase, J. Gen. Virol. 73 (1992) 303–311.[9] T. Daikoku, R. Kurachi, T. Tsurumi, Y. Nishiyama, Identification ofthe observed declines in expression because of the absence
a target protein of US3 protein kinase of herpes simplex virus typeof virus replication and HSV-1 genes and because pTH-2, J. Gen. Virol. 75 (1994) 2065–2068.
NFHlac /ul13g,us3g exhibited less than a 20% reduction in [10] D.J. Dargan, A.H. Patel, J.H. Subak-Sharpe, PREPs: herpes simplexthe number of X-gal positive cells between 4 days and 2 virus type 1-specific particles produced by infected cells when viralweeks. DNA replication is blocked, J. Virol. 69 (1995) 4924–4932.

T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16 15
[11] J.J. Diaz, M.D. Dodon, N. Schaerer-Uthurralt, D. Simonin, K. [30] R. Leopardi, C. Van Sant, B. Roizman, The herpes simplex virus 1Kindbeiter, L. Gazzolo, J.J. Madjar, Post-transcriptional transactiva- protein kinase US3 is required for protection from apoptosis inducedtion of human retroviral envelope glycoprotein expression by herpes by the virus, Proc. Natl. Acad. Sci. USA 94 (1997) 7891–7896.simplex virus Us11 protein, Nature 379 (1996) 273–277. [31] F. Lim, D. Hartley, P. Starr, P. Lang, S. Song, L. Yu, Y. Wang, A.I.
[12] P.C. Emson, S. Shoham, C. Feler, T. Buss, J. Price, C.J. Wilson, The Geller, Generation of high-titer defective HSV-1 vectors using an IEuse of a retroviral vector to identify foetal striatal neurones 2 deletion mutant and quantitative study of expression in culturedtransplanted into the adult striatum, Exp. Brain Res. 79 (1990) cortical cells, Biotechniques 20 (1996) 460–469.427–430. [32] K. Lim, C.B. Chae, A simple assay for DNA transfection by
[13] C. Fraefel, S. Song, F. Lim, P. Lang, L. Yu, Y. Wang, P. Wild, A.I. incubation of the cells in culture dishes with substrates for beta-Geller, Helper virus-free transfer of herpes simplex virus type 1 galactosidase, Biotechniques 7 (1989) 576–579.plasmid vectors into neural cells, J. Virol. 70 (1996) 7190–7197. [33] A.M. McCarthy, L. McMahan, P.A. Schaffer, Herpes simplex virus
[14] A.I. Geller, Genetic analysis of the role of protein kinase C signaling type 1 ICP27 deletion mutants exhibit altered patterns of transcrip-pathways in behaviors by direct gene transfer with HSV-1 vectors, tion and are DNA deficient, J. Virol. 63 (1989) 18–27.Rev. Neurosci. 10 (1999) 1–13. [34] D.J. McGeoch, M.A. Dalrymple, A.J. Davison, A. Dolan, M.C.
[15] A.I. Geller, X.O. Breakefield, A defective HSV-1 vector expresses Frame, D. McNab, L.J. Perry, J.E. Scott, P. Taylor, The completeEscherichia coli beta-galactosidase in cultured peripheral neurons, DNA sequence of the long unique region in the genome of herpesScience 241 (1988) 1667–1669. simplex virus type 1, J. Gen. Virol. 69 (1988) 1531–1574.
[16] C.R. Goding, P. O’Hare, Herpes simplex virus Vmw65-octamer [35] R.J. Mullen, C.R. Buck, A.M. Smith, NeuN, a neuronal specificbinding protein interaction: a paradigm for combinatorial control of nuclear protein in vertebrates, Development 116 (1992) 201–211.transcription, Virology 173 (1989) 363–367. [36] T.I. Ng, W.O. Ogle, B. Roizman, UL13 protein kinase of herpes
[17] L.A. Greene, A.S. Tischler, Establishment of a noradrenergic clonal simplex virus 1 complexes with glycoprotein E and mediates theline of rat adrenal pheochromocytoma cells which respond to nerve phosphorylation of the viral Fc receptor: glycoproteins E and I,growth factor, Proc. Natl. Acad. Sci. USA 73 (1976) 2424–2428. Virology 241 (1998) 37–48.
[18] W.P. Halford, B.M. Gebhardt, D.J. Carr, Persistent cytokine expres- [37] P.F. Nichol, J.Y. Chang, E.M. Johnson Jr., P.D. Olivo, Infection ofsion in trigeminal ganglion latently infected with herpes simplex sympathetic and sensory neurones with herpes simplex virus doesvirus type 1, J. Immunol. 157 (1996) 3542–3549. not elicit a shut-off of cellular protein synthesis: implications for
[19] S.K. Hanks, A.M. Quinn, T. Hunter, The protein kinase family: viral latency and herpes vectors, Neurobiol. Dis. 1 (1994) 83–94.conserved features and deduced phylogeny of the catalytic domains, [38] W.O. Ogle, T.I. Ng, K.L. Carter, B. Roizman, The UL13 proteinScience 241 (1988) 42–52. kinase and the infected cell type are determinants of posttranslation-
[20] A.B. Hill, B.C. Barnett, A.J. McMichael, D.J. McGeoch, HLA class al modification of ICP0, Virology 235 (1997) 406–413.I molecules are not transported to the cell surface in cells infected [39] F.C. Purves, R.M. Longnecker, D.P. Leader, B. Roizman, Herpeswith herpes simplex virus types 1 and 2, J. Immunol. 152 (1994) simplex virus 1 protein kinase is encoded by open reading frame2736–2741. US3 which is not essential for virus growth in cell culture, J. Virol.
[21] B.C. Horsburgh, M.M. Hubinette, D. Qiang, M.L. MacDonald, F. 61 (1987) 2896–2901.Tufaro, Allele replacement: an application that permits rapid ma- [40] F.C. Purves, B. Roizman, The UL13 gene of herpes simplex virus 1nipulation of herpes simplex virus type 1 genomes, Gene Ther. 6 encodes the functions for posttranslational processing associated(1999) 922–930. with phosphorylation of the regulatory protein alpha 22, Proc. Natl.
[22] B.K. Jin, M. Belloni, B. Conti, H.J. Federoff, R. Starr, J.H. Son, H. Acad. Sci. USA 89 (1992) 7310–7314.Baker, T.H. Joh, Prolonged in vivo gene expression driven by a [41] F.C. Purves, D. Spector, B. Roizman, The herpes simplex virus 1tyrosine hydroxylase promoter in a defective herpes simplex virus protein kinase encoded by the US3 gene mediates posttranslationalamplicon vector, Hum. Gene Ther. 7 (1996) 2015–2024. modification of the phosphoprotein encoded by the UL34 gene, J.
[23] P.A. Johnson, A. Miyanohara, F. Levine, T. Cahill, T. Friedmann, Virol. 65 (1991) 5757–5764.Cytotoxicity of a replication-defective mutant of herpes simplex [42] G.S. Read, N. Frenkel, Herpes simplex virus mutants defective invirus type 1, J. Virol. 66 (1992) 2952–2965. the virion-associated shutoff of host polypeptide synthesis and
[24] P.A. Johnson, M.J. Wang, T. Friedmann, Improved cell survival by exhibiting abnormal synthesis of alpha (immediate early) viralthe reduction of immediate-early gene expression in replication- polypeptides, J. Virol. 46 (1983) 498–512.defective mutants of herpes simplex virus type 1 but not by mutation [43] B. Roizman, A.E. Sears, Herpes simplex viruses and their replica-of the virion host shutoff function, J. Virol. 68 (1994) 6347–6362. tion, in: B. Roizman, R.J. Whitley, C. Lopez (Eds.), The Human
[25] F.E. Jones, C.A. Smibert, J.R. Smiley, Mutational analysis of the Herpesviruses, Raven Press, New York, 1993, pp. 11–68.herpes simplex virus virion host shutoff protein: evidence that vhs [44] R.J. Roller, L.L. Monk, D. Stuart, B. Roizman, Structure andfunctions in the absence of other viral proteins, J. Virol. 69 (1995) function in the herpes simplex virus 1 RNA-binding protein U(s)11:4863–4871. mapping of the domain required for ribosomal and nucleolar
[26] A. Kalnins, K. Otto, U. Ruther, B. Muller-Hill, Sequence of the lacZ association and RNA binding in vitro, J. Virol. 70 (1996) 2842–gene of Escherichia coli, EMBO J. 2 (1983) 593–597. 2851.
[27] M.G. Kaplitt, A.D. Kwong, S.P. Kleopoulos, C.V. Mobbs, S.D. [45] R.J. Roller, B. Roizman, The herpes simplex virus 1 RNA bindingRabkin, D.W. Pfaff, Preproenkephalin promoter yields region-spe- protein US11 is a virion component and associates with ribosomalcific and long-term expression in adult brain after direct in vivo gene 60S subunits, J. Virol. 66 (1992) 3624–3632.transfer via a defective herpes simplex viral vector, Proc. Natl. [46] Y. Saeki, T. Ichikawa, A. Saeki, E.A. Chiocca, K. Tobler, M.Acad. Sci. USA 91 (1994) 8979–8983. Ackermann, X.O. Breakefield, C. Fraefel, Herpes simplex virus type
[28] Y. Kawaguchi, C. Van Sant, B. Roizman, Eukaryotic elongation 1 DNA amplified as bacterial artificial chromosome in Escherichiafactor 1delta is hyperphosphorylated by the protein kinase encoded coli: rescue of replication-competent virus progeny and packagingby the U(L)13 gene of herpes simplex virus 1, J. Virol. 72 (1998) of amplicon vectors, Hum. Gene Ther. 9 (1998) 2787–2794.1731–1736. [47] M.L. Schwartz, C. Katagi, J. Bruce, W.W. Schlaepfer, Brain-specific
[29] Y. Kong, T. Yang, A.I. Geller, An efficient in vivo recombination enhancement of the mouse neurofilament heavy gene promoter incloning procedure for modifying and combining HSV-1 cosmids, J. vitro, J. Biol. Chem. 269 (1994) 13444–13450.Virol. Methods 80 (1999) 129–136. [48] I.L. Smith, M.A. Hardwicke, R.M. Sandri-Goldin, Evidence that the

16 T. Yang et al. / Molecular Brain Research 90 (2001) 1 –16
herpes simplex virus immediate early protein ICP27 acts post- [53] Y. Wang, L. Yu, A.I. Geller, Diverse stabilities of expression in thetranscriptionally during infection to regulate gene expression, Virol- rat brain from different cellular promoters in a helper virus-freeogy 186 (1992) 74–86. herpes simplex virus type 1 vector system, Hum. Gene Ther. 10
[49] S. Song, Y. Wang, S.Y. Bak, P. Lang, D. Ullrey, R.L. Neve, K.L. (1999) 1763–1771.O’Malley, A.I. Geller, An HSV-1 vector containing the rat tyrosine [54] M.J. Wood, A.P. Byrnes, D.W. Pfaff, S.D. Rabkin, H.M. Charlton,hydroxylase promoter enhances both long-term and cell type-spe- Inflammatory effects of gene transfer into the CNS with defectivecific expression in the midbrain, J. Neurochem. 68 (1997) 1792– HSV-1 vectors, Gene Ther. 1 (1994) 283–291.1803. [55] G. Zachos, B. Clements, J. Conner, Herpes simplex virus type 1
[50] T.A. Stavropoulos, C.A. Strathdee, An enhanced packaging system infection stimulates p38/c-Jun N-terminal mitogen-activated proteinfor helper-dependent herpes simplex virus vectors, J. Virol. 72 kinase pathways and activates transcription factor AP-1, J. Biol.(1998) 7137–7143. Chem. 274 (1999) 5097–5103.
[51] J.G. Stevens, Latent herpes simplex virus and the nervous system, [56] G. Zhang, X. Wang, T. Yang, M. Sun, W. Zhang, Y. Wang, A.I.Curr. Top. Microbiol. Immunol. 70 (1975) 31–50. Geller, A tyrosine hydroxylase–neurofilament chimeric promoter
[52] M. Sun, G.R. Zhang, T. Yang, L. Yu, A.I. Geller, Improved titers for enhances long-term expression in rat forebrain neurons from helperhelper virus-free herpes simplex virus type 1 plasmid vectors by virus-free HSV-1 vectors, Mol. Brain Res. 84 (2000) 17–31.optimization of the packaging protocol and addition of non-infecti- [57] W. Zhang, C. Schott, G. Zhang, Y. Wang, J. Guerriere, G. Lewan-ous herpes simplex virus-related particles (previral DNA replication dowski, L.A. Lampson, A.I. Geller, Comparison of the inflammatoryenveloped particles) to the packaging procedure, Hum. Gene Ther. response to gene transfer into the rat brain with helper-containing or10 (1999) 2005–2011. helper virus-free HSV-1 vectors, (2000) (submitted for publication).




















